
1. Биология как наука. Методы биологии | |
| 1.1. Роль биологии в формировании современной естественнонаучной картины мира, в практической деятельности людей. Методы изучения живых объектов. Биологический эксперимент. Наблюдение, описание, измерение биологических объектов | Конспект 1, Конспект 2 |
2. Признаки живых организмов | ТЕСТЫ |
| 2.1. Клеточное строение организмов как доказательство их родства, единства живой природы. Гены и хромосомы. Нарушения в строении и функционировании клеток — одна из причин заболеваний организмов. Вирусы — неклеточные формы жизни | Конспект 1, Конспект 2, Конспект 3 |
| 2.2. Признаки организмов. Наследственность и изменчивость — свойства организмов. Одноклеточные и многоклеточные организмы. Ткани, органы, системы органов растений и животных, выявление изменчивости организмов. Приемы выращивания и размножения растений и домашних животных, ухода за ними | Конспект 1, Конспект 2, Конспект 3, Конспект 4, Конспект 5, Конспект 6. |
3. Система, многообразие и эволюция живой природы | ТЕСТЫ |
| 3.1. Царство Бактерии. Роль бактерий в природе, жизни человека и собственной деятельности. Бактерии — возбудители заболеваний растений, животных, человека | Конспект |
| 3.2. Царство Грибы. Роль грибов в природе, жизни человека и собственной деятельности. Роль лишайников в природе, жизни человека и собственной деятельности | Конспект 1, Конспект 2 |
| 3.3. Царство Растения. Роль растений в природе, жизни человека и собственной деятельности | Конспект |
| 3.4. Царство Животные. Роль животных в природе, жизни человека и собственной деятельности | Конспект |
| 3.5. Учение об эволюции органического мира. Ч. Дарвин — основоположник учения об эволюции. Усложнение растений и животных в процессе эволюции. Биологическое разнообразие как основа устойчивости биосферы и результата эволюции | Конспект 1, Конспект 2, Конспект 3 |
4. Человек и его здоровье | ТЕСТЫ |
| 4.1. Сходство человека с животными и отличие от них. Общий план строения и процессы жизнедеятельности человека | Конспект 1, Конспект 2, Конспект 3 |
| 4.2. Нейро-гуморальная регуляция процессов жизнедеятельности организма. Нервная система. Рефлекс. Рефлекторная дуга. Железы внутренней секреции. Гормоны | |
| 4.3. Питание. Система пищеварения. Роль ферментов в пищеварении | Конспект |
| 4.4. Дыхание. Система дыхания | Конспект |
| 4.5. Внутренняя среда организма: кровь, лимфа, тканевая жидкость. Группы крови. Иммунитет | Конспект 1, Конспект 2 |
| 4.6. Транспорт веществ. Кровеносная и лимфатическая системы | Конспект 1, Конспект 2 |
| 4.7. Обмен веществ и превращение энергии в организме человека. Витамины | Конспект 1, Конспект 2 |
| 4.8. Выделение продуктов жизнедеятельности. Система выделения | Конспект |
| 4.9. Покровы тела и их функции | Конспект |
| 4.10. Размножение и развитие организма человека. Наследование признаков у человека. Наследственные болезни, их причины и предупреждение | Конспект 1, Конспект 2. |
| 4.11. Опора и движение. Опорно-двигательный аппарат | Конспект 1, Конспект 2 |
| 4.12. Органы чувств, их роль в жизни человека | Конспект |
| 4.13. Психология и поведение человека. Высшая нервная деятельность. Условные и безусловные рефлексы, их биологическое значение. Познавательная деятельность мозга. Сон, его значение. Биологическая природа и социальная сущность человека. Сознание человека. Память, эмоции, речь, мышление. Особенности психики человека: осмысленность восприятия, словесно-логическое мышление, способность к накоплению и передаче из поколения в поколение информации. Значение интеллектуальных, творческих и эстетических потребностей. Цели и мотивы деятельности. Индивидуальные особенности личности: способности, темперамент, характер. Роль обучения и воспитания в развитии психики и поведения человека | Конспект |
| 4.14. Соблюдение санитарно-гигиенических норм и правил здорового образа жизни. Переливание крови. Профилактические прививки. Уход за кожей, волосами, ногтями. Укрепление здоровья: закаливание, двигательная активность, сбалансированное питание, рациональная организация труда и отдыха, чистый воздух. Факторы риска: несбалансированное питание, гиподинамия, курение, употребление алкоголя и наркотиков, стресс, вредные условия труда, и др. Инфекционные заболевания: грипп, гепатит, ВИЧ-инфекция и другие инфекционные заболевания (кишечные, мочеполовые, органов дыхания). Предупреждение инфекционных заболеваний. Профилактика: отравлений, вызываемых ядовитыми растениями и грибами; заболеваний, вызываемых паразитическими животными и животными-переносчиками возбудителей болезней; травматизма; ожогов; обморожений; нарушения зрения и слуха | Конспект |
| 4.15. Приемы оказания первой доврачебной помощи: при отравлении некачественными продуктами, ядовитыми грибами и растениями, угарным газом; спасении утопающего; кровотечениях; травмах опорно-двигательного аппарата; ожогах; обморожениях; повреждении зрения | Конспект |
5. Взаимосвязи организмов и окружающей среды | ТЕСТЫ |
| 5.1. Влияние экологических факторов на организмы. Приспособления организмов к различным экологическим факторам. Популяция. Взаимодействия разных видов (конкуренция, хищничество, симбиоз, паразитизм). Сезонные изменения в живой природе | Конспект |
| 5.2. Экосистемная организация живой природы. Роль производителей, потребителей и разрушителей органических веществ в экосистемах и круговороте веществ в природе. Пищевые связи в экосистеме. Цепи питания. Особенности агроэкосистем | |
| 5.3. Биосфера — глобальная экосистема. Роль человека в биосфере. Экологические проблемы, их влияние на собственную жизнь и жизнь других людей. Последствия деятельности человека в экосистемах, влияние собственных поступков на живые организмы и экосистемы | Конспект |
Подготовка к ОГЭ по биологии. Материалы по каждому заданию
Подготовка к ОГЭ по биологии 2020.
Материалы по каждому заданию
На выполнение ОГЭ по биологии отводится 2,5 часа (150 минут). Работа состоит из 2 частей, включающих 30 заданий. Часть I включает задания с кратким ответом: 19 заданий с выбором одного ответа из четырёх (в том числе на работу с графиками и таблицами, умение определять правильность биологических суждений), а также 6 заданий с кратким ответом в виде последовательности цифр. Часть II содержит задания с развёрнутым ответом. Подготовка к ОГЭ по биологии 2020 может проводиться путем разбора материала по каждому заданию (комментарии к заданиям ОГЭ).
Задание 01 Задание 02 Задание 03 Задание 04
Задание 05 Задание 06 Задание 07 Задание 08
Задание 09 Задание 10 Задание 11 Задание 12
Задание 13 Задание 14 Задание 15 Задание 16
Задание 17 Задание 18 Задание 19
Задание 21 Задание 22 Задание 23 Задание 24
Задание 25 Задание 26 Задание 27 Задание 28
Задание 29 Задание 30
Шкала перевода баллов
ОГЭ 2020 Биология. Максимальное количество первичных баллов, которое может получить участник ОГЭ за выполнение всей экзаменационной работы — 45 баллов.
0—12 баллов — отметка «2»
13—24 баллов — отметка «3»
25—35 баллов — отметка «4»
36—45 баллов — отметка «5»
Рекомендуемый минимальный первичный балл для отбора обучающихся в профильные классы для обучения по образовательным программам среднего общего образования — 33 балла.
Государственная итоговая аттестация (ГИА) выпускников 9-х классов в настоящее время является распространённой практикой выпускной аттестации школьников как государственного механизма контроля качества образования. ОГЭ по биологии проводится с использованием современных тестовых технологий, являющихся одним из направлений модернизации российского образования.
ГИА направлена на проверку реализации требований, предъявляемых к учащимся на современном этапе обучения, проверку как общебиологических знаний, так и метапредметных умений. Контрольные измерительные материалы (КИМ) ГИА включают задания по
Данное пособие предназначено учащимся 9-х классов для подготовки к ОГЭ по биологии. В то же время оно полезно и учителям, желающим найти оптимальные формы организации подготовки учеников к этому экзамену. Предлагаемое пособие составлено с учётом характерных особенностей различных учебных (рабочих) программ по биологии для общеобразовательных учреждений. Подобраны материалы по каждому заданию ОГЭ по биологии 2020 года.
Подготовка к ОГЭ по биологии.
Материалы по каждому заданию
Подготовка к огэ по биологии. Биология как наука. Методы биологии: теория и практика
ТЕОРЕТИЧЕСКИЙ МАТЕРИАЛ
БИОЛОГИЯ КАК НАУКА. МЕТОДЫ БИОЛОГИИ
Биология – наука о жизни, ее закономерностях и формах проявления, о существовании и распространении ее во времени и пространстве. Она исследует происхождение жизни и ее сущность, развитие, взаимосвязи и многообразие. Биология относится к естественным наукам.
Впервые термин «биология» употребил немецкий профессор анатомии Т. Руз в 1779г. Однако общепринятым он стал в 1802 г., после того как его стал употреблять в своих работах французский натуралист Ж.-Б. Ламарк.
Современная биология представляет собой комплексную науку, состоящую из ряда самостоятельных научных дисциплин со своими объектами исследования.
БИОЛОГИЧЕСКИЕ ДИСЦИПЛИНЫ
Ботаника — наука о растениях,
Зоология – наука о животных,
Микология — о грибах,
Вирусология — о вирусах,
Микробиология — о бактериях.
Анатомия — наука, изучающая внутреннее строение организмов (отдельных органов, тканей). Анатомия растений изучает строение растений, анатомия животных — строение животных.
Морфология — наука, изучающая внешнее строение организмов
Физиология — наука, изучающая процессы жизнедеятельности организма, функции отдельных органов.
Гигиена – наука о сохранении и укреплении здоровья человека.
Цитология — наука о клетке.
Гистология – наука о тканях.
Систематика — наука, о классификации живых организмов. Классификация — разделение организмов на группы (виды, рода, семейства и т. д.) на основании особенностей строения, происхождения, развития и др.
Палеонтология — наука, изучающая ископаемые останки (отпечатки, окаменелости и др.) организмов.
Эмбриология – наука, изучающая индивидуальное (зародышевое) развитие организмов.
Экология — наука, изучающая взаимоотношения организмов друг с другом и с окружающей средой.
Этология – наука о поведении животных.
Генетика — наука о закономерностях наследственности и изменчивости.
Селекция — наука, о выведении новых и улучшением существующих пород домашних животных, сортов культурных растений и штаммов бактерий и грибов.
Эволюционное учение – изучает вопросы возникновения и законы исторического развития жизни на Земле.
Антропология – наука о возникновении и развитии человека.
Клеточная инженерия — направление науки, занимающееся получением гибридных клеток. Примером может служить гибридизация раковых клеток и лимфоцитов, слияние протопластов разных растительных клеток, а также клонирование.
Генная инженерия — направление науки, занимающееся получением гибридных молекул ДНК или РНК. Если клеточная инженерия работает на уровне клетки, то генная работает на молекулярном уровне. В данном случае специалисты «пересаживают» гены одного организма другому. Одним из результатов генной инженерии является получение генетически модифицированных организмов (ГМО).
Бионика — направление в науке, занимающееся поиском возможностей применения принципов организации, свойств и структур живой природы в технических устройствах.
Биотехнология — дисциплина, изучающая возможности использования организмов или биологических процессов для получения веществ, нужных человеку. Обычно в биотехнологических процессах используются бактерии и грибы.
ОБЩИЕ МЕТОДЫ БИОЛОГИИ
Метод – это способ познания действительности.
1. Наблюдение и описание.
2. Измерение
3. Сравнение
4. Эксперимент или опыт
5. Моделирование
6. Исторический.
ЭТАПЫ научного исследования
Проводится наблюдение над объектом или явлением
на основе полученных данных выдвигается гипотеза
проводится научный эксперимент (с контрольным опытом)
проверенная в ходе эксперимента гипотеза может быть названа
теорией или законом
СВОЙСТВА ЖИВОГО
Обмен веществ (метаболизм) и поток энергии — важнейшее свойство живого. Все живые организмы поглощают необходимые им вещества из внешней среды и выделяют в нее продукты жизнедеятельности.
Единство химического состава. Среди химических элементов в живых организмах преобладают углерод, кислород, водород и азот. Кроме того, важнейшим признаком живых организмов является наличие органических веществ: жиров, углеводов, белков и нуклеиновых кислот.
Клеточное строение. Все организмы состоят из клеток. Неклеточное строение имеют только вирусы, но и они проявляют признаки живого, только попав в клетку-хозяина.
Раздражимость — способность организма реагировать на внешние или внутренние воздействия.
Самовоспроизведение. Все живые организмы способны к размножению, т. е. воспроизведению себе подобных. Воспроизведение организмов происходит в соответствии с генетической программой, записанной в молекулах ДНК.
Наследственность и изменчивость.
Наследственность — свойство организмов, передавать свои признаки потомкам. Наследственность обеспечивает преемственность жизни. Изменчивость — способность организмов приобретать новые признаки в процессе своего развития. Наследственная изменчивость является важным фактором эволюции.
Рост и развитие.
Рост — количественные изменения (например, увеличение массы).
Развитие — качественные изменения (например, формирование систем органов, цветение и плодоношение).
Саморегуляция — способность организмов поддерживать постоянство своего химического состава и процессов жизнедеятельности — гомеостаз.
Приспособленность (адаптация)
Ритмичность – периодические изменения интенсивности физиологических функций с различными периодами колебаний (суточные, сезонные ритмы). (Например, фотопериодизм – реакция организма на длину светового дня).
Уровни организации жизни
Номер | Название | Чем представлен | Примеры |
VI | Биосферный | Совокупность всех экосистем | |
V | Экосистемный (биогеоценотический) | Система популяций разных | Саванна, тундра |
IV | Популяционно- | Совокупность популяций, | Медведи белые, |
III | Организменный | Организм как целостная система | Бактерия, обезьяна |
II | Клеточный | Клетка и её структурные компоненты | Эритроциты, митохондрии, хлоропласты |
I | Молекулярный | Органические и неорганические вещества | – белки, углеводы; – вода, ионы солей |
Тестовые задания в формате ОГЭ
Какая наука изучает сортовое разнообразие растений?
1)физиология 2)систематика 3)экология 4)селекция
2. Выяснить, необходим ли свет для образования крахмала в листьях, можно с помощью
1) описания органов растений 2) сравнения растений разных природных зон
3) наблюдения за ростом растения 4) эксперимента по фотосинтезу
3. В какой области биологии была разработана клеточная теория?
1)вирусологии 2) цитологии 3)анатомии 4) эмбриологии
4. Для разделения органоидов клетки по плотности Вы выберете метод
1) наблюдения 2) хроматографии 3) центрифугирования 4)выпаривания
5.На фотографии изображена модель фрагмента ДНК. Какой метод позволил учёным создать такое трехмерное изображение молекулы?

1) классификации 2) эксперимента 3) наблюдения 4) моделирования
6. На фотографии изображен шаростержневой фрагмент ДНК. Какой метод позволил ученым создать такое трехмерное изображение молекулы?

классификации 2) эксперимента 3) наблюдения 4) моделирования
7. Применение какого научного метода иллюстрирует сюжет картины голландского художника Я. Стена «Пульс», написанной в середине XVII в.?

1) моделирование 2) измерение 3) эксперимент 4) наблюдение
8. Изучите график, отражающий процесс роста и развития насекомого.
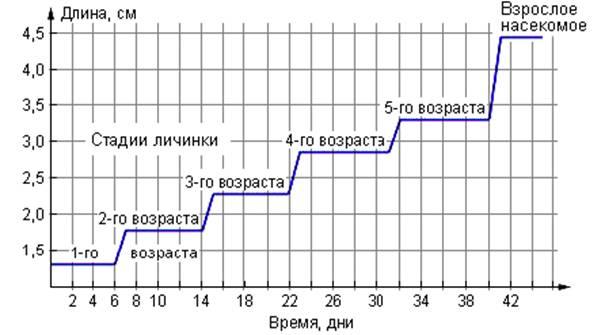 Определите длину насекомого на 30-й день его развития.
Определите длину насекомого на 30-й день его развития.
1) 3,4 2) 2,8 3) 2,5 4) 2,0
9. Кого из перечисленных ученых считают создателем эволюционного учения?
1) И.И. Мечникова 2) Л. Пастера 3) Ч. Дарвина 4) И.П. Павлова
10. Какая наука изучает сортовое разнообразие растений?
1) физиология 2) систематика 3) экология 4) селекция
11. Выберите пару животных, в экспериментах с которыми были сделаны основные открытия в области физиологии животных и человека.
1) лошадь и корова 2) пчела и бабочка 3) собака и лягушка 4) ящерица и голубь
12. В какой области биологии была разработана клеточная теория?
1) вирусологии 2) цитологии 3) анатомии 4) эмбриологии
13. Точно установить степень влияния удобрений на рост растений можно методом
1) эксперимента 2) моделирования 3) анализа 4) наблюдения
14. Примером применения экспериментального метода исследования является
1) описание строения нового растительного организма
2)сравнение двух микропрепаратов с различными тканями
3)подсчёт пульса у человека до и после нагрузки
4) формулирование положения на основе полученных фактов
15. Микробиолог хотел узнать, насколько быстро размножается один из видов бактерий в разных питательных средах. Он взял две колбы, заполнил их до половины разными питательными средами и поместил туда примерно одинаковое количество бактерий. Каждые 20 минут он извлекал пробы и подсчитывал в них количество бактерий. Данные его исследования отражены в таблице.
Изучите таблицу «Изменение скорости размножения бактерий за определённое время» и ответьте на вопросы.
Изменение скорости размножения бактерий за определённое время
Время после введения бактерий в культуру, мин. | Число бактерий в колбе 1 | Число бактерий в колбе 2 |
20 | 18 | 20 |
40 | 36 | 40 |
60 | 72 | 80 |
80 | 140 | 160 |
100 | 262 | 314 |
120 | 402 | 620 |
140 | 600 | 1228 |
1) Сколько бактерий поместил учёный в каждую колбу в самом начале эксперимента?
2) Как изменялась скорость размножения бактерий на протяжении эксперимента в каждой колбе?
3) Чем можно объяснить полученные результаты?
Литература
Каменский А.А., Криксунов Е.А., Пасечник В.В. Биология. Общая биология 9 класс: учеб. для общеобразовательных учреждение. М.: Дрофа, 2013.
Заяц Р.Г., Рачковская И.В., Бутиловский В.Э., Давыдов В.В. Биология для абитуриентов: вопросы, ответы, тесты, задачи.- Минск: Юнипресс, 2011.-768 с.
«Решу ОГЭ»: биология. Обучающая система Дмитрия Гущина [Электронный ресурс] – URL:http:// oge.sdamgia.ru
Методы познания живой природы Метод – это путь исследования, который проходит ученый, решая какую-либо научную задачу или проблему | |||||
Метод (название) | Описание / Суть метода | Пример | |||
Классические методы исследования в биологии (в науке вообще). Эмпирические – основаны на чувственном познании | |||||
Описание | В основе его лежит наблюдение. Последующая запись в полевой дневник наблюдений или на другой носитель. | Он широко применялся еще учёными древности, занимавшихся сбором фактического материала и его описанием (изучение и описание животных и растений), а также применяется в настоящее время (например, при открытии/описании новых видов). | |||
Наблюдение | Наблюдение — метод, с помощью которого исследователь собирает информацию об объекте (восприятие природных объектов с помощью органов чувств). | Наблюдать можно визуально, например, за поведением животных. Можно наблюдать с помощью приборов за изменениями, происходящими в живых объектах: например, при снятии кардиограммы в течение суток, при замерах веса телёнка в течение месяца. Наблюдать можно за сезонными изменениями в природе, за линькой животных и т.д. Выводы, сделанные наблюдателем, проверяются либо повторными наблюдениями, либо экспериментально. | |||
Измерение | Измерение различных показателей биологического объекта или явления с помощью измерительных приборов. | Измерение ЧСС человека, измерение веса, роста, объема грудной клетки и т.п. В палеонтологии измерение веса и линейных размеров находок/ископаемых. | |||
Эксперимент (опыт) | Метод, с помощью которого проверяют результаты наблюдений, выдвинутые предположения (гипотезы). Это всегда получение нового знания с помощью поставленного опыта | Скрещивания животных или растений с целью получения новой породы или сорта, проверка нового лекарства, выявление функций/роли органоида клетки. | |||
Классические методы исследования в биологии. Теоретические умозаключения (и не только) на основе фактов, полученных эмпирическими методами. Из них чаще всего используются метод исторических аналогий, анализ, моделирование, экстраполяция, абстрагирование и конкретизация, индукция и дедукция, сравнение, обобщение, систематизация и интерпретация фактов. Одной практикой не обойтись, хороший исследователь должен опираться на практику, но потом хорошенько всё обдумать и систематизировать. | |||||
Сравнение | Сравнение полученных в ходе исследования данных с предыдущими исследованиями или литературными данными. | Для наглядности в дальнейшем составление графиков, таблиц, диаграмм и т.п. Например, сравнение среднего количества птенцов в гнездах в разные годы. | |||
Классификация | Объединение нескольких объектов в группы на основании общих признаков, заданных исследователем. | Основной метод таксономистов (людей, занимающихся биоразнообразием). Классификация растений и животных, грибов и микроорганизмов. | |||
Анализ | Процесс мысленного разделения предмета на части (признаки, свойства, отношения) с последующим обдумыванием составляющих по отдельности и в целом (а это уже синтез). | Тут довольно трудно с примерами, но чисто интуитивно каждый понимает, что это такое. Анализ результатов проведенного исследования в соответствии с поставленными задачами. Или теоретическое обобщение на основе литературных данных изучение отдельных свойств и структуры органоидов клетки. | |||
Синтез | Соединение выделенных в ходе анализа сторон предмета в единое целое. | И дальнейшее объединение этих умозаключений в единую систему, описывающую взаимосвязь и необходимость отдельных структур клетки. | |||
Обобщение | Метод построения гипотез и, в дальнейшем, теорий на основе полученных эмпирических и литературных данных. Установление родства биологических объектов. Выявление черт сходства и различия и их важности для конкретного исследования. | Синтетическая теория эволюции обобщила факты и результаты экспериментов из различных областей биологической науки. | |||
Моделирование | Метод, при котором создаётся некий образ объекта (явления, процесса), модель, с помощью которой ученые получают необходимые сведения об изучаемом объекте. | При установлении структуры молекулы ДНК Джеймс Уотсон и Френсис Крик создали из пластмассовых элементов модель молекулы ДНК (двойную спираль), отвечающую данным рентгенологических и биохимических исследований. В настоящее время компьютерное моделирование процессов и явлений встречается в научных исследованиях всё чаще. | |||
Исторический | Применяется для установления взаимосвязей между фактами, процессами, явлениями, происходившими на исторически длительном промежутке времени. | Эволюционное учение развивалось во многом благодаря этому методу. | |||
Палеонтологический | Сбор данных во время археологических/палеонтологических раскопок. Входит во все исследования по эволюции и применим для изучения практически всех эволюционных процессов. | Позволяет выявить родство между древними организмами, останки которых находятся в земной коре, в разных геологический слоях. Возможно определение относительного возраста осадочных толщ земной коры по сохранившимся в них ископаемым остаткам организмов. Позволяет составить филогенетические ряды современных лошади и кита. | |||
Абстрагирование | Отвлечение в процессе познания от некоторых свойств объекта с целью углубленного исследования одной его стороны. Позволяет не учитывать ряд свойств объекта в ходе конкретного исследования. Помогает выделить то, что важно. | Для классификации организмов важны как свойства в совокупности, как и по отдельности. Или, в генетике при анализе наследования конкретного признака, к примеру, окраска венчика цветка, нужно анализировать именно его, абстрагируясь при этом от формы и цвета семени, размера куста и прочих. | |||
Современные методы изучения биологических объектов. | |||||
Цитологический/ цитогенетический | Исследование строения клетки, её структур, с помощью различных методов микроскопирования (далее). | Метод обнаружения болезнетворных микроорганизмов в пробах, мазках и др. Часто используется в мед.диагностике заболеваний. Цитогенетический исследует кариотип человека (хромосомы ядра), выявляет наследственные заболевания, связанные с изменением структуры и количества хромосом. | |||
Световая микроскопия | Довольно дешевый и эффективный метод исследования. Актуален до сих пор и без него не обойтись практически ни одному биологу. используют световой микроскоп. Строение его довольно простое, обязательно повторите. | Позволяет наблюдать живые объекты. Можно рассматривать клетку целиком, срез органа, ткань, но не органоиды. Иногда видно ядро и хлоропласты, клеточную стенку. | |||
Метод микроскопирования в темном поле | Мелкие структуры, невидимые при обычном микроскопировании, становятся заметны в отраженных лучах. | Рассматривание мелких структур / объектов (менее 0,2 мкм). Используется в микробиологической диагностике патогенных микроорганизмов. | |||
Флуоресцентная микроскопия | Некоторые вещества/структуры способны светиться при поглощении световой энергии. Существуют флуорохромы (см. картинку), способные избирательно связываться с липидами, полисахаридами и некоторыми белками. | Позволяет изучить локализацию различных химических веществ в живой и фиксированной клетке. | |||
Фазово-контрастная микроскопия | Основана на том, что отдельные структуры, прозрачной, в целом, клетки отличаются друг от друга по светопреломлению и плотности. Проходя через эти структуры, свет изменяет свою фазу, но наш глаз не воспринимает это изменение. Специальный объектив на микроскопе создаёт черно-белое контрастное изображение. | Микробиологическая диагностика патогенных микроорганизмов. | |||
Электронная микроскопия | Требует длительной и сложной подготовки объекта к микроскопированию, дорогостоящий метод, однако позволяющий рассматривать самые мелкие клеточные структуры. | Изучение повехностных структур клетки, её органоидов, отдельных элементов, ультраструктуры, всё это возможно только благодаря электронному микроскопу. Структура вирусов исследуется и была открыта только таким методом. | |||
Биохимический | Исследование химических процессов, происходящих в организме. | Исследование биохимического анализа крови человека. Может быть частным методом генетики как науки. Был использован для выявления частных болезней обмена веществ, связанных с наследственностью. | |||
Центрифугирование | Разделение смесей на составляющие под действием центробежной силы. Изучение состава и свойств смесей. | Применяется для разделения органоидов клетки, легких и тяжелых фракций органических соединений. | |||
Хроматография | Метод разделения компонентов смесей, основа на распределении компонентов между двумя фазами: неподвижной (нанесенной на колонку) и подвижной, протекающей через неподвижную. | Метод разделения пигментов растительной клетки. Метод определения беременности (по наличию определенного гормона в моче или в крови) | |||
Электрофорез | Близкий к хроматографии метод, разделению веществ в геле способствует электрический ток. проводится в хроматографической камере с электродами. | Основной метод ДНК-диагностики. Выделяет ДНК-фрагмент определенного размера/веса/длины из смеси. Фрагменты видны в УФ-излучении, благодаря предварительному окрашиванию. | |||
Метод меченых атомов | Чтобы проследить за превращением какого-либо вещества в него вводят радиоактивную метку (изотоп какого-либо элемента) | Применяется для изучения процессов, происходящих в живых клетках. Позволяет проследить круговорот элемента в природе или осаждение вещества в каком-либо органе или ткани. | |||
Спектральный анализ | Известно, что каждый элемент поглощает/излучает характерный только для него световой спектр Метод изучает химический состав веществ на основании их способности к поглощению и испусканию света. | Современные методы позволяют определить до миллиардных долей грамма определенного вещества в пробе. | |||
Метод витального (прижизненного) окрашивания | В низких концентрациях красители малотоксичны для живых клеток. | Этот метод позволяет судить о жизнедеятельности клетки при различных внешних воздействиях. | |||
Метод культивирования клеток и тканей | Основан на выращивании отдельных клеток, тканей и органов вне организма. Отдельные клетки или кусочки тканей выращивают обычно погруженными в питательную среду. | Таким образом можно получить стерильные материалы для посадки растений. Можно вырастить кусочки тканей человека для трансплантации его собственных тканей или даже органов. Таким методом выращивают редкие орхидеи, продающиеся в каждом цветочном магазине, наращивают биомассу женьшеня и т.п. | |||
Клеточная инженерия | Совокупность методов, используемых для конструирования новых клеток. Включает культивирование и клонирование клеток на специально подобранных средах, гибридизацию клеток, пересадку клеточных ядер и другие микрохирургические операции по «разборке» и «сборке» (реконструкции) жизнеспособных клеток из отдельных фрагментов. Клеточная инженерия основана на культивировании растительных и животных клеток и тканей, способных вне организма производить нужные для человека вещества. | Соматическую гибридизацию, т. е. получение гибридов без участия полового процесса, проводят, культивируя совместно клетки различных линий одного вида или клетки различных видов. При определённых условиях происходит слияние двух разных клеток в одну гибридную, содержащую оба генома объединившихся клеток. Удалось получить гибриды между клетками животных, далёких по систематическому положению, напр. мыши и курицы. Соматические гибриды нашли широкое применение как в научных исследованиях, так и в биотехнологии. | |||
Генная инженерия | Совокупность приёмов, методов и технологий получения рекомбинантных РНК и ДНК, выделения генов из организма (клеток), осуществления манипуляций с генами, введения их в другие организмы и выращивания искусственных организмов после удаления выбранных генов из ДНК. Генетическая инженерия не является наукой в широком смысле, но является инструментом биотехнологии | Все методы генетической инженерии применяются для осуществления одного из следующих этапов решения генно-инженерной задачи: Получение изолированного гена. Введение гена в вектор (плазмиду или бактериофаг) для переноса в организм. Перенос вектора с геном в модифицируемый организм. Преобразование клеток организма. Отбор генетически модифицированных организмов (ГМО) и устранение тех, которые не были успешно модифицированы. | |||
Методы генетики как науки. | |||||
Гибридологический | Основной метод исследования в Менделевской генетике. Им же и был введен в науку: подбирают родительские пары по паре альтернативных признаков и анализируют фенотипическое расщепление в потомстве. | Берем чистые линии родительских особей, отличающихся по какому-либо исследуемому признаку, скрещиваем, наблюдаем в потомстве единообразие, делаем вывод относительно доминирования. | |||
Цитогенетический | Был описан выше в общих методах. Применяется для исследования кариотипа человека. | С помощью цитогенетических методов можно выявить геномные мутации у человека (синдром Дауна, синдром Клайнфельтера и т.д.). | |||
Генеалогический | Применяется при составлении родословных людей, выявлении характера наследования некоторых признаков. | С помощью этого метода выявили сцепленное с полом наследование гемофилии. | |||
Близнецовый | Применяется для выявления степени влияния окружающей среды, воспитания и условий жизни на генотип. Изучаются генотипические и фенотипические особенности одно- и двуяйцовых близнецов. | Можно попробовать разделить влияние наслественности и среды на организм. Близнецов иногда даже воспитывают в разных семьях. | |||
Популяционно-статистический | Это метод изучения распространения наследственных признаков (наследственных заболеваний) в популяциях. Существенным моментом при использовании этого метода является статистическая обработка получаемых данных. | Изучением генетической структуры популяций занимается особый раздел генетики — популяционная генетика. Для выяснения частот встречаемости тех или иных генов и генотипов используется закон Харди-Вайнберга. | |||
Биохимический | Был описан выше, применяется и как частный метод в генетике. | Позволяет обнаружить нарушения в обмене веществ, вызванные изменением генов и, как следствие, изменением активности различных ферментов. Наследственные болезни обмена веществ подразделяются на болезни углеводного обмена (сахарный диабет), обмена аминокислот, липидов, минералов и др. | |||
Методы селекции как науки. | Теоретической основой селекции является генетика, так как именно знание законов генетики позволяет целенаправленно управлять закреплением мутаций, предсказывать результаты скрещивания, правильно проводить отбор гибридов. В результате применения знаний по генетике удалось создать более 10 тысяч сортов пшеницы на основе нескольких исходных диких сортов, получить новые штаммы микроорганизмов, выделяющих пищевые белки, лекарственные вещества, витамины и т. п. | ||||
Скрещивание | |||||
Родственное (инбридинг) | Применяется с целью сохранения/закрепления необходимых признаков у сорта/породы. Для закрепления полезных наследственных свойств необходимо повысить гомозиготность нового сорта. Иногда для этого применяют самоопыление перекрёстноопыляемых растений. | ||||
Неродственное (аутбридинг) | -внутрисортовое (внутрипородное) -межсортовое (межпородное) | Метод межлинейной гибридизации, при котором часто наблюдается эффект гетерозиса: гибриды первого поколения обладают высокой урожайностью и устойчивостью к неблагоприятным воздействиям. Гетерозис характерен для гибридов первого поколения, которые получаются при скрещивании не только разных линий, но и разных сортов и даже видов. Эффект гетерозиготной (или гибридной) мощности бывает сильным только в первом гибридном поколении, а в следующих поколениях постепенно снижается. Основная причина гетерозиса заключается в устранении в гибридах вредного проявления накопившихся рецессивных генов. Другая причина — объединение в гибридах доминантных генов родительских особей и взаимное усиление их эффектов. | |||
Отдаленная гибридизация | Отдаленная гибридизация домашних животных менее эффективна, чем растений. Межвидовые гибриды животных часто бывают бесплодными. При этом восстановление плодовитости у животных представляет более сложную задачу, поскольку получение полиплоидов на основе умножения числа хромосом у них невозможно. Правда, в некоторых случаях отдаленная гибридизация сопровождается нормальным слиянием гамет, обычным мейозом и дальнейшим развитием зародыша, что позволило получить некоторые породы, сочетающие ценные признаки обоих использованных в гибридизации видов. | ||||
Искусственный отбор | Первоначально в основе селекции лежал искусственный отбор, когда человек отбирает растения или животных с интересующими его признаками. До XVI—XVII веков отбор происходил нерегулярно и неметодично: для посева отбирали лучшие плоды (на посадку) или особи (для воспроизводства) просто рассчитывая на повторение результата; забавно, что это соседствовало с теологической убеждённостью неизменности «божьих созданий». Только в последние столетия, ещё не зная законов генетики, стали использовать отбор сознательно и целенаправленно, скрещивая экземпляры с ярко выраженными полезными свойствами. | ||||
Массовый отбор | Отбор особей по фенотипу без проверки генотипа. Для перекрёстноопыляемых растений применяют массовый отбор особей с желаемыми свойствами. В противном случае невозможно получить материал для дальнейшего скрещивания. Таким образом получают, например, новые сорта ржи. Эти сорта не являются генетически однородными. | ||||
Индивидуальный отбор | Прием искусственного отбора, который проводится на основе индивидуальной наследственной изменчивости особи. В селекции животных применяют жесткий индивидуальный отбор по хозяйственно ценным признакам, выносливости, экстерьеру. В селекции растений индивидуальный отбор используют при работе с самоопыляющимися растениями, при этом выделяются чистые линии — потомство одной самоопыляющейся особи. Благодаря индивидуальному отбору от одного вида дикого сизого голубя выведено около 150 пород домашних голубей; от одного вида собаки получено все разнообразие пород. Большинство сортов пшеницы, ячменя, овса были получено методом индивидуального отбора. | ||||
Полиплоидизация | Получают искусственные полиплоиды при помощи химических веществ, которые разрушают веретено деления, в результате чего удвоившиеся хромосомы не могут разойтись, оставаясь в одном ядре. Одно из таких веществ — колхицин. | Полиплоиды отличаются быстрым ростом, крупными размерами и высокой урожайностью. В сельскохозяйственной практике широко используются триплоидная сахарная свекла, четырёхплоидный клевер, рожь и твердая пшеница, а также шестиплоидная мягкая пшеница. Один из путей преодоления стерильности межвидовых гибридов. | |||
Искусственный мутагенез | Путём искусственного мутагенеза и последующего отбора мутантов были получены новые высокоурожайные сорта ячменя и пшеницы. Этими же методами удалось получить новые штаммы грибов, выделяющие в 20 раз больше антибиотиков, чем исходные формы. За последние 70 лет выведено более 2250 сортов сельскохозяйственных растений, созданных при помощи физического и химического мутагенеза. | ||||
Методы исследования эволюции | |||||
Палеонтологические методы | Практически все методы палеонтологии применимы для изучения эволюционных процессов. Важнейшие из этих методов: выявление ископаемых промежуточных форм, восстановление филогенетических рядов и обнаружение последовательности ископаемых форм. | Наибольшую информацию палеонтологические методы дают о состоянии биосферы на различных этапах развития органического мира вплоть до современности, о последовательности смен флор и фаун. | |||
Биогеографические методы | Биогеографические методы основаны на анализе распространения ныне существующих видов. Особое значение имеет изучение распространения реликтовых форм. | Даёт информацию о местонахождении очагов происхождения таксонов, путях их расселения, влиянии климатических условий и изоляции на развитие видов. | |||
Эмбриологический | Данный метод основан на проявлении закономерностей эмбрионального развития, таких как закон зародышевого сходства и биогенетический закон | Выявление зародышевого сходства. В первой половине XIX в. выдающийся натуралист К. Бэр сформулировал «закон зародышевого сходства: чем более ранние стадии индивидуального развития исследуются, тем больше сходства обнаруживается между различными организмами. Например, на ранних стадиях развития эмбрионы позвоночных не отличаются друг от друга. Лишь на средних стадиях развития в сравниваемом ряду у зародышей появляются особенности, характерные для рыб и амфибий; на ее более поздних стадиях – особенности рептилий, птиц и млекопитающих. | |||
Морфологические методы | Морфологические (сравнительно-анатомические, гистологические и др.) методы позволяют на основе сравнения сходств и различий в строении организмов судить о степени их родства. | Изучение строения современных организмов, относящихся к группам с различным эволюционным возрастом с целью выявления общих и различных черт их организации. Изучают гомологичные и аналогичные органы, рудименты и атавизмы. | |||
Молекулярно-генетические методы | Сравнения последовательности ДНК разных генов у разных организмов могут сказать учёному много нового об эволюционных взаимоотношениях организмов, которые не могут иначе быть обнаружены на основе на морфологии, или внешней форме организмов, и их внутренней структуре. Поскольку геномы эволюционируют через постепенное накопление мутаций, количество отличий последовательности нуклеотидов между парой геномов разных организмов должно указать, как давно эти два генома разделили общего предка. Два генома, которые разделились в недавнем прошлом, должны иметь меньшие отличий, чем два генома, чей общий предок очень давний. Потому, сравнивая разные геномы друг с другом, возможно получить сведения об эволюционном взаимоотношения между ними. Это является главной задачей молекулярной филогенетики. | Большая и разнообразная группа методов, предназначенная для выявления вариаций (повреждений) в структуре участка ДНК (аллеля, гена, региона хромосомы) вплоть до расшифровки первичной последовательности оснований. В основе этих методов лежат генно-инженерные манипуляции с ДНК и РНК. | |||
что нужно знать, чтобы подготовиться к экзамену
Биология не похожа на физику. Чем
больше мы узнаем, тем очевиднее
становится, что одного объяснения, до
которого нужно докопаться, не будет.
Стив Джонс
Экзамен по биологии в 9 классе входит в список необязательных предметов, доступных к сдаче по выбору. Предмет пользуется популярностью и высокий процент учащихся сдаёт его на максимальный бал.
Почему выбирают ОГЭ по биологии?
- На уровне 9 класса материал достаточно прост.
- Объёмы информации невелики (биологию изучают с 5 по 9 класс, если образовательное учреждение или направление обучения – непрофильное).
- После 9 класса с экзаменом по биологии можно поступать/переводиться в медицинские колледжи и университеты.
- Это отличная тренировка, если в планах – сдавать ЕГЭ по биологии в 11 классе.
Таким образом, экзамен чаще всего выбирают в двух случаях:
- либо у школьника ещё нет чётких планов на обучение, тогда экзамен «выигрывает» за счёт своей простоты;
- либо же – напротив, уже определена дальнейшая программа и в планах – профильное направление в старшей школе или в профессиональном училище.
Ещё один неоспоримый плюс: вопросы и задания ОГЭ по биологии из года в год остаются фактически неизменными. При подготовке можно пользоваться не только материалами 2018-2019 года, но даже периода 2015-2017 года. Также, в сборники тестов входят дополнительные упражнения, которые не встретятся на самом испытании, но будут крайне полезны для оттачивания навыков сдачи и прохождения теста – рекомендуем уделить им внимание.
Структура экзамена по биологии
- Длительность: 180 минут (3 часа).
- Объём: 32 вопроса.
- Формат испытания: 2 части (с коротким ответом и с развёрнутым комментарием).
- Использование материалов во время сдачи – запрещено.
Сгруппировать задания можно:
— по уровням сложности
- базовый – 22 задания (1-22)
- повышенный – 7 заданий (23-29)
- высокий – 3 задания (30-32)
— по формату выполнения
Задания 1-22: ответ записывается с помощью одной цифры (бланк №1).
Пример
Плесневые грибы человек использует при:
- выпечке хлеба
- силосовании кормов
- получении сыров
- приготовлении столового вина
Задания 23-28: ответ записывается в виде последовательности цифр (бланк №1).
Пример
Вставьте в текст «Типы клеток» пропущенные термины из предложенного перечня, используя для этого цифровые обозначения. Запишите в текст цифры выбранных ответов, а затем получившуюся последовательность цифр (по тексту) запишите в таблицу.
ТИПЫ КЛЕТОК
Первыми на пути исторического развития появились организмы, имеющие мелкие клетки с простой организацией, – _________(А). Эти доядерные клетки не имеют оформленного _________(Б). В них выделяется лишь ядерная зона, содержащая _________(В) ДНК. Такие клетки есть у современных _________(Г) и синезелёных.
Перечень терминов:
- хромосома
- прокариоты
- цитоплазма
- кольцевая молекула
- ядро
- одноклеточное животное
- бактерия
- эукариоты
Задания 29-32: даётся развёрнутый ответ (бланк №2).
Используя содержание текста «Происхождение живых существ», ответьте на следующие вопросы.
- Какое оборудование использовал в своём эксперименте Ф. Реди?
- Что было объектом исследования в опытах Л. Пастера?
- Как на мясе в открытых банках могли появиться черви?
| Ответ: |
| Банки и марля / 8 банок, марля |
| Мясной бульон |
| Черви — червеобразные личинки насекомых образуются из яиц, отложенных комнатными мухами |
Основные темы при сдаче ОГЭ по биологии
Чтобы разнообразить подготовку, задания ОГЭ по биологии 2019 года, как и задания прошлых лет, можно сгруппировать по темам. Количественно они тоже отличаются: где-то встретится всего 1-2 вопроса, а обширные темы охватывают большой объём материала и насчитывают до 50-60.
- Роль биологии в практической деятельности людей
- Клеточное строение организмов
- Одноклеточные и многоклеточные организмы. Царство грибы
- Царство Растения
- Царство Животные
- Сходство человека с животными и отличие от них
- Нейрогуморальная регуляция процессов жизнедеятельности
- Опора и движение
- Внутренняя среда
- Транспорт веществ
- Питание. Дыхание
- Обмен веществ. Выделение. Покровы тела
- Органы чувств
- Психология и поведение человека
- Соблюдение санитарно-гигиенических норм
- Влияние экологических факторов на организмы
- Экосистемная организация живой природы. Биосфера
- Интерпретирование результатов научных исследований
- Определение структуры объекта
- Умение оценивать правильность биологических суждений
- Умение проводить множественный выбор
- Умение устанавливать соответствие
- Биологические процессы, явления, объекты
- Пропущенные термины и понятия из числа предложенных
- Соотнесение морфологических признаков организма
- Работа с текстом биологического содержания
- Статистические данные, представленные в табличной форме
- Определение энерготрат и составление рациона
- Обосновывание рационального и здорового питания
Обратите внимание на последние четыре темы. Их знание потребуется, если вы планируете сдать вторую часть теста ОГЭ по биологии (29-32/С1-С4) на высокие баллы.
Оценки и критерии
Оценивание ОГЭ по биологии строится следующим образом.
| Задание | Максимальный балл |
|---|---|
| 1-22 | 1 |
| 23-27; 32 | 2 |
| 28-31 | 3 |
Максимальный балл | 46 |
Для перевода оценок в пятибалльную шкалу воспользоваться можно этой таблицей:
Пятибалльная шкала
Балл за ОГЭ по биологии
Минимальный проходной балл – 13. Для его достижения необходимо без ошибок решить минимум 13 заданий базового уровня.
Советы по подготовке к экзамену
Когда предмет для сдачи выбран, возникает логичный вопрос: как готовиться к ОГЭ по биологии?

Даём советы, следуя которым, вы сможете подготовиться и сдать на высокий балл.
- Начинайте подготовку заранее. Бывает такое, что всё внимание «перетягивают» на себя основные экзамены (русский язык и математика), а на биологию и другие дополнительные экзамены совершенно не хватает времени. Не нервничайте: подготовиться, чтобы хорошо сдать, получится и за короткий срок (хотя, конечно, лучше не затягивать). Главное: выявить сложности и слабые места, грамотно распланировать подготовку и чётко следовать намеченной структуре.
- Определите уровень знаний по предмету. Сделать это можно как с помощью репетитора, так и самому – достаточно без подготовки решить несколько вариантов теста и оценить результат. Это поможет выявить слабые места и «западающие» темы. После того, как вы поймёте все эти моменты – можно приступать к планированию подготовки.
- Осуществляйте постоянный контроль. Выработайте у себя привычку отслеживания и проверки результатов вашей подготовки. Если вы занимаетесь на курсах или с репетитором, то ваши действия регулирует преподаватель, и корректирует, при надобности. Но это не освобождает вас от необходимости самостоятельно отслеживать собственные шаги, успехи и, напротив, слабые места.
- Помните про возможность подачи апелляции. Этот совет – на самый крайний случай. Но если вдруг случится ситуация, где нужно будет оспорить справедливость оценки, почему бы не воспользоваться им.
Если вы сознательно выбрали сдачу биологии, а результаты пригодятся в будущем, то мы желаем вам успехов! Напоминаем, что, если у вас возникли трудности при подготовке к сдаче ОГЭ по биологии, вы всегда можете записаться на курсы в Ruseducenter.
Занятия в учебном центре Ruseducenter проходят в 2 филиалах по адресам:
Москва, Старопименовский переулок, 18
Москва, ул.Мясницкая, 22
Телефон: 8 (495) 189-69-80 Записаться
E-mail: [email protected]
Читать похожие статьи
ОГЭ по биологии: материалы для подготовки
Главная » 2016 » Июнь » 5 » Материалы для подготовки к ОГЭ по биологии14:15 Материалы для подготовки к ОГЭ по биологии | |
 На сайте появился новый комплект материалов к ОГЭ по биологии: На сайте появился новый комплект материалов к ОГЭ по биологии: ● краткая теория ● курс биологии в таблицах ● подробный словарь терминов ● комплект материалов по следующим разделам: 1. Биология как наука. Методы научного познания 2. Клетка как биологическая система 3. Организм как биологическая система 4. Система и многообразие органического мира 5. Организм человека и его здоровье 6. Эволюция живой природы 7. Экосистемы и присущие им закономерности ● Таблицы по курсу школьной программы: • 6 класс. Живой организм • 6 класс. Ботаника • 7 класс. Зоология • 7, 9-11 классы. Растительные сообщества • 8 класс. Анатомия и физиология • 9-11 классы. Генетика • 9-11 классы. Общая биология. Экология • 9-11 классы. Эволюция систем органов • 8-9 классы. Человек • 10-11 классы. Эволюционное учение • Введение в Экологию • Вещества растений, клеточное строение • Общее знакомство с цветковыми растениями • Растение — живой организм • Химия клетки + бонус: Для того, чтобы сдать ОГЭ 2020 по биологии на пятёрку, мы рекомендуем: ● Онлайн-курсы Фоксфорда ● Очные курсы в твоём городе ● Репетиторы Фоксфорда (онлайн) ● Репетиторы (вживую и онлайн) | |
| Просмотров: 49766 | | Теги: теория, шпаргалки, словарь, ОГЭ, биология, материалы, шпоры, скачать, 9 класс, таблицы, бесплатно | |

Подготовка с нуля к ОГЭ по биологии
Теперь ты знаешь, что такое ОГЭ по биологии. Но какой же уровень знаний у тебя сегодня? Узнать это нужно как можно скорее.
Есть несколько вариантов решения поставленной задачи.
Вариант первый – самый простой. Подойди к учителю биологии и спроси вариант экзаменационной работы прошлых лет. Реши его, избегая соблазна посмотреть ответы или решение в интернете. Помни, что на выполнение работы дается 3 часа. Попроси учителя биологии проверить выполненный вариант в твоем присутствии. Это очень важно! Ты узнаешь, как оцениваются задания экзаменационной работы. Полученные баллы переведи в отметку – это и есть твой результат сегодняшнего дня.
Вариант второй – самостоятельный. Найди вариант экзаменационной работы по биологии в интернете или приобрети готовые варианты для подготовки к ОГЭ по биологии. Важно, чтобы задания найденного варианта соответствовали структуре ОГЭ в этом году. (смотри статью «Что такое ОГЭ по биологии» ССЫЛКА). Сравни количество заданий первой части, сравни количество заданий второй части. Посмотри, есть ли в найденном варианте ответы для проверки. Выполни и проверь, используя ответы и рекомендации. Полученные баллы переведи в отметку.
Вариант третий – распространенный. Обратиться за помощью к репетитору, который даст пробную работу для выполнения, проверит ее, определит темы для изучения и поможет подготовиться к ОГЭ по биологии.
Используя любой из предложенных вариантов, ты получаешь результат сегодняшнего дня и первую практику самостоятельного выполнения экзаменационной работы ОГЭ по биологии.
Этот небольшой тест поможет тебе понять, какие у тебя сильные и слабые стороны, где пробелы в знаниях по биологии. И действительно ли у тебя «подготовка с нуля». По первым 16 тестовым заданиям можно определить, какой блок материала по биологии ты знаешь, а какой надо учить. Весь материал ОГЭ по биологии разделен на пять блоков:
1. Биология как наука;
2. Признаки живых организмов;
3. Система, многообразия и эволюция живой природы;
4. Человек и его здоровье;
5. Взаимосвязь организмов и окружающей среды
В варианте ОГЭ:
Задания 1 и 2 — по блоку «Признаки живых организмов»
Задания 3-5 — по блоку «Система, многообразия и эволюция живой природы»
Задания 6-15 — по блоку «Человек и его здоровье»
Задание 16 — по блоку «Взаимосвязь организмов и окружающей среды»
Начинай с этих заданий. Основной материал для изучения этих блоков ты найдешь в учебниках по биологии за 7, 8, 9 классы.
Автор Некипелова
Определение теории и примеры — Биологический онлайн-словарь
Определение
существительное, множественное число: теории
(общее)
(1) Предположение, мнение, предположение или предположение, основанное на ограниченной информации или опыте, необязательно по фактам.
(2) Ожидание того, что может произойти, за исключением непредвиденных обстоятельств. Например, теория о том, что преступники обычно возвращаются на место преступления.
(наука) Хорошо обоснованное объяснение некоторых аспектов естественного мира, основанное на совокупности фактов, неоднократно подтвержденных посредством наблюдений и экспериментов.
(математика) Раздел математики, пытающийся описать определенный класс конструкций и включающий аксиомы, теоремы, примеры, и т. Д. .
Дополнение
Общее использование термина теория подразумевает домыслы или предположения, которые не были проверены или имеют ограниченные доказательства. В науке недоказанная идея или простое теоретическое предположение рассматривается как гипотеза, а не как научная теория. Однако в науке теория — это хорошо обоснованное объяснение или набор утверждений, подтвержденных в ходе множества независимых экспериментов.Для сравнения: законы объясняют вещи, но не описывают их, тогда как теории объясняют, почему мы видим набор наблюдений. Например, закон всемирного тяготения Ньютона описывает, как работает гравитация, но не объясняет, почему она существует. Теория эволюции путем естественного отбора Дарвина объясняет нам, почему наблюдения за жизнью на этой планете такие, какими мы их видим. И научные теории, и законы основаны на фактах и признаны научным сообществом истинными. И оба используются для предсказания будущих событий.
Слово происхождение: L theria
Производные термины: теорема, теоретический ( прилагательное ), теоретизирующий ( глагол ).
Связанные термины: теория вероятностей, теория темперамента, теория игр, теория питекоида.
Сравните: гипотеза, закон.
Теория графов и анализ биологических данных в вычислительной биологии
1. Введение
Теория сложных сетей играет важную роль в широком спектре дисциплин, от коммуникаций до молекулярной и популяционной биологии. Основное внимание в этой статье уделяется методам теории графов для вычислительной биологии. Мы рассмотрим методы и подходы в теории графов, а также текущие приложения в биомедицинской информатике. В области биологии и медицины потенциальные применения сетевого анализа с использованием теории графов включают идентификацию мишеней для лекарств, определение роли белков или генов с неизвестной функцией.Существует несколько биологических областей, в которых методы теории графов применяются для извлечения знаний из данных. Мы разделили эти проблемы на несколько различных областей, которые описаны ниже.
Моделирование биомолекулярных сетей. В нем представлены методы моделирования биомолекулярных сетей, таких как сети взаимодействия белков, метаболические сети, а также сети регуляции транскрипции.
Измерение центральности и важности в биомолекулярных сетях.Идентификация наиболее важных узлов в большой сложной сети имеет фундаментальное значение в вычислительной биологии. Мы представим несколько исследований, в которых применялись меры центральности для идентификации структурно важных генов или белков в сетях взаимодействия и изучалась биологическая значимость генов или белков, идентифицированных таким образом.
Идентификация мотивов или функциональных модулей в биологических сетях. В наиболее важных биологических процессах, таких как передача сигналов, регуляция судьбы клеток, транскрипция и трансляция, задействовано более четырех, но гораздо меньше, чем сотни белков или генов.Наиболее важные процессы в биологических сетях соответствуют мотивам или функциональным модулям. Это предполагает, что определенные функциональные модули очень часто встречаются в биологических сетях и используются для их категоризации.
Новые пути добычи из биомолекулярных сетей. Биологические пути позволяют лучше понять механизмы взаимодействия молекул. Экспериментальная проверка идентификации путей у различных организмов в условиях влажной лаборатории требует огромных затрат времени и усилий.Таким образом, существует потребность в инструментах теории графов, которые помогают ученым предсказывать пути в биомолекулярных сетях.
Наша основная цель в данной статье — дать как можно более широкий обзор основных достижений в этой области. Кроме того, мы также выделяем то, что было достигнуто, а также некоторые из наиболее важных открытых проблем, которые необходимо решить. Наконец, мы надеемся, что эта глава послужит полезным введением в эту область для тех, кто не знаком с литературой.
2. Определения и математические предварительные сведения
2.1. Понятие графа
Понятие графа является фундаментальным для материала, который будет обсуждаться в этой главе. Граф G состоит из набора вершин V ( G ) и набора ребер E ( G ). В простом графе две вершины в G связаны, если существует ребро ( vi , vj ) E ( G ), соединяющее вершины vi и vj в графе G так, что vi V ( G ) и vj V ( G ).Количество вершин будет обозначаться как V ( G ), а набор вершин, смежных с вершиной vi , называется соседями vi , N ( vi ). Степень вершины vi — это количество ребер, с которыми она инцидентна, что обозначено как d ( vi ). Два графика, G1 и G2 , называются изоморфными ( G1 G2 ), если однозначное преобразование V1 в V2 приводит к однозначному преобразованию E1 на E2 .Подграф G ´ графа G — это граф, набор вершин и набор ребер которого удовлетворяют соотношениям: V ( G ´) V ( G ) и E ( G ´) E ( G ), и если G ´ является подграфом G , то G называется надграфом G ´. Линейный граф L ( G ) неориентированного графа G — это такой граф, что каждая вершина в L ( G ) указывает ребро в G и любые пары вершин в L ( G ) являются смежными тогда и только тогда, когда их соответствующие ребра имеют общую конечную точку в G .
2.2. Направленные и неориентированные графы
Граф может быть неориентированным , что означает, что нет различия между двумя вершинами, связанными с каждым ребром, или его ребра могут быть направлены от одной вершины к другой. Формально конечный ориентированный граф G состоит из набора вершин или узлов V ( G ) = { v1 , …, vn } вместе с набором ребер E ( G ) V ( G ) V ( G ).Интуитивно каждое ребро ( u , v ) E ( G ) можно рассматривать как соединение начального узла u с конечным узлом v . Ненаправленный граф G также состоит из набора вершин V ( G ) и набора ребер E ( G ). Однако в этом случае с краями не связано никакого направления. Следовательно, элементы E ( G ) представляют собой просто два подмножества элементов V ( G ), а не упорядоченные пары как ориентированные графы.Как и в случае с ориентированными графами, мы будем использовать обозначение uv (или vu , поскольку направление неважно) для обозначения ребра { u , v } в неориентированном графе. Для двух вершин, u , v , неориентированного графа, uv является ребром тогда и только тогда, когда vu также является ребром. Мы не имеем дело с мультиграфами, поэтому между любой парой вершин в неориентированном графе может быть не более одного ребра. То есть мы обсуждаем простой график.Простой граф — это неориентированный граф, в котором нет петель и не более одного ребра между любыми двумя разными вершинами. В простом графе ребра графа образуют набор, и каждое ребро представляет собой пару из различных вершин. Количество вершин n в ориентированном или неориентированном графе — это размер или порядок графа.
2.3. Степень узла и матрица смежности
Для неориентированного графа G , мы будем писать d ( u ) для степени узла u в V ( G ).Это просто общее количество ребер и . Для графов, которые мы будем рассматривать, это равно количеству соседей u , d ( u ) = | N ( u ) |. В ориентированном графе G внутренняя степень , d + ( u ) (внешняя степень, d — ( u )) вершины u задается количеством ребер, которые заканчиваются (или начинаются) на и .Предположим, что вершины графа (ориентированного или неориентированного) G упорядочены как v 1 , …, v n . Тогда матрица смежности A для G имеет вид
aij = {1ifvivj∈E (G) 0ifvivj∉E (G) E1
Таким образом, матрица смежности неориентированного графа симметрична, в то время как это необходимо не в случае ориентированного графа.
2.4 Путь, длина пути и связанный граф
Пусть u , v — две вершины в графе G .Тогда последовательность вершин u = v1 , v2 , …, vk = v , такая, что для i = 1, …, k -1, быть путем длиной k -1 от u до v . Геодезическое расстояние, или просто расстояние, d ( u , v ), от u до v — это длина кратчайшего пути от u до v в G .Если такого пути не существует, то мы устанавливаем d ( u , v ) = 1. Если для каждой пары вершин ( u, v ) в графе G есть путь от u к v , то мы говорим, что G подключен.
3. Моделирование биомолекулярных сетей
3.1. Введение
Было изучено несколько классов биомолекулярных сетей: транскрипционные регуляторные сети, сети взаимодействия белков и метаболические сети.В биологии сети регуляции транскрипции и метаболические сети обычно моделируются в виде ориентированных графов. Например, в сети регуляции транскрипции узлы будут представлять гены с краями, обозначающими транскрипционные отношения между ними. Это был бы направленный граф, потому что, если ген A регулирует ген B, тогда существует естественное направление, связанное с границей между соответствующими узлами, начиная с A и заканчивая B. В последние годы внимание было сосредоточено на протеине — сети взаимодействия белков различных простых организмов (Itzkovitz & Alon, 2005).Эти сети описывают прямые физические взаимодействия между белками в протеоме организма, и нет направления, связанного с взаимодействиями в таких сетях. Следовательно, сети PPI обычно моделируются как неориентированные графы, в которых узлы представляют белки, а ребра представляют взаимодействия. В следующих разделах мы индивидуально представим эти биомолекулярные сети.
3.2. Сети регуляции транскрипции
Сети регуляции транскрипции описывают регуляторные взаимодействия между генами.Здесь узлы соответствуют отдельным генам, и направленный край проводится от гена A к гену B, если A положительно или отрицательно регулирует ген B. Созданы сети для сетей регуляции транскрипции E. coli и S. cerevisiae ( Salgado et al., 2006; Lee et al., 2002; Salgado et al., 2006; Keseler et al., 2005) и поддерживаются в таких базах данных, как RegulonDB (Salgado et al., 2006) и EcoCyc (Keseler et al. ., 2005). Такие сети обычно создаются путем сочетания высокопроизводительных экспериментов по определению местоположения генома и поиска литературы.В прошлом сообщалось о многих типах подходов, связанных с регуляцией транскрипции генов. Их природа и состав подразделяются на несколько факторов: с учетом значений экспрессии генов (Keedwell & Narayanan, 2005; Shmulevich et al., 2002), причинно-следственная связь между генами, например с байесовским анализом или динамическими байесовскими сетями (Zou & Conzen, 2005; Husmeier, 2003), а также во временной области, например дискретное или непрерывное время (Li et al., 2006; He & Zeng, 2006; Filkov et al., 2002; Qian et al., 2001). Одним из ограничений приложений теории графов при анализе биохимических сетей является статическое качество графов. Биохимические сети динамичны, и абстракция до графов может маскировать временные аспекты информационного потока. Узлы и звенья биохимических сетей меняются со временем. Однако представление системы в виде статического графа является предпосылкой для построения подробных динамических моделей (Zou & Conzen, 2005). Большинство подходов к динамическому моделированию можно использовать для имитации сетевой динамики, используя графическое представление в качестве скелета модели.Моделирование динамики биохимических сетей обеспечивает более близкое к реальности повторение поведения системы in silico , что может быть полезно для разработки более количественных гипотез.
3.3. Сети взаимодействия белков
Понимание взаимодействий белков — одна из важных проблем вычислительной биологии. Эти сети белок-белковых взаимодействий (PPI) обычно представлены в формате неориентированного графа с узлами, соответствующими белкам, и ребрами, соответствующими взаимодействиям белок-белок.Объем экспериментальных данных о белок-белковых взаимодействиях быстро увеличивается за счет усовершенствования высокопроизводительных методов, позволяющих производить большие партии ИПП. Например, дрожжи содержат более 6000 белков, и в настоящее время между дрожжевыми белками идентифицировано более 78000 ИПП, и сотни лабораторий по всему миру постоянно добавляют в этот список. Ожидается, что у человека будет около 120000 белков и около 10 6 PPI. Ожидается, что отношения между структурой сети PPI и функцией сотовой связи будут изучены.Крупномасштабные сети PPI (Rain et al., 2001; Giot et al., 2003; Li et al., 2004; Von Mering et al., 2004; Mewes et al., 2002) были построены недавно с использованием высокой пропускной способности. такие подходы, как дрожжевой-2-гибридный скрининг (Ito et al., 2001) или методы масс-спектрометрии (Gavin et al., 2002) для выявления взаимодействий с белками.
Огромные объемы данных, связанных с PPI, которые постоянно генерируются по всему миру, хранятся в многочисленных базах данных. Данные о взаимодействии белков также хранятся в базах данных, таких как база данных взаимодействующих белков (DIP) (Xenarios et al., 2000). Мы кратко упомянем основные базы данных, включая базы данных нуклеотидных последовательностей, белковых последовательностей и PPI. Самыми крупными базами данных нуклеотидных последовательностей являются EMBL (Stoesser et al., 2002), DDBJ (Tateno et al., 2002) и GenBank (Benson et al., 2002). Они содержат последовательности из литературы, а также те, которые предоставлены непосредственно отдельными лабораториями. Эти базы данных хранят информацию в общем для всех организмов. Для многих организмов существуют специфические базы данных. Например, полный геном дрожжей и родственных штаммов дрожжей можно найти в базе данных генома Saccharomyces (SGD) (Dwight et al., 2002). FlyBase (Ashburner, 1993) содержит полный геном плодовой мушки Drosophila melanogaster . Это одна из самых ранних баз данных моделей организмов. Ensembl (Hubbard et al., 2002) содержит черновой вариант последовательности генома человека вместе с предсказанием генов и крупномасштабной аннотацией. SwissProt (Bairoch & Apweiler, 2000) и Protein Information Resource (PIR) (McGarvey et al., 2000) — две основные базы данных последовательностей белков. SwissProt поддерживает высокий уровень аннотаций для каждого белка, включая его функцию, доменную структуру и информацию о посттрансляционной модификации.
Понимание взаимодействий между белками в клетке может быть полезно с помощью модели сети PPI. Полное описание сетей взаимодействия белков требует сложной модели, которая охватывала бы ненаправленные физические взаимодействия белок-белок, другие типы взаимодействий, уровень достоверности взаимодействия или метод и множественность взаимодействия, информацию о направленном пути, временную информацию о наличии или отсутствии. ИЦП, а также информацию о силе взаимодействия.Это может быть достигнуто путем разработки функции оценки и присвоения весов узлам и краям сети PPI.
3.4. Метаболические сети
Метаболические сети описывают биохимические взаимодействия внутри клетки, посредством которых субстраты превращаются в продукты посредством реакций, катализируемых ферментами. Метаболические сети обычно требуют более сложных представлений, таких как гиперграфы, поскольку реакции в метаболических сетях обычно преобразуют несколько входов в и несколько выходов с помощью других компонентов.Альтернативой является взвешенный двудольный граф для уменьшения представления метаболической сети. В таких графах используются два типа узлов для представления реакций и соединений соответственно. Ребра взвешенного двудольного графа соединяют узлы разных типов, представляя отношения либо субстрата, либо продукта. Эти сети могут представлять собой полный набор метаболических и физических процессов, которые определяют физиологические и биохимические свойства клетки. Метаболические сети сложны.В таких сетях существует множество видов узлов (белки, частицы, молекулы) и множество соединений (взаимодействий). Даже если можно определить подсети, которые можно осмысленно описать в относительной изоляции, от нее всегда есть соединения с другими сетями. Как и в случае сетей взаимодействия с белками, метаболические сети в масштабе генома были сконструированы для множества простых организмов, включая S. cerevisiae и E. coli (Jeong et al., 2000; Overbeek et al., 2000; Karp et al. ., 2002; Edwards et al., 2000) и хранятся в базах данных, таких как базы данных KEGG (Kanehisa & Goto, 2000) или BioCyc (Karp et al., 2005). Обычный подход к построению таких сетей состоит в том, чтобы сначала использовать аннотированный геном организма для идентификации ферментов в сети, а затем объединить биохимическую и генетическую информацию для получения связанных с ними реакций (Kauffman et al., 2000; Edwards и др., 2001). Несмотря на то, что были предприняты усилия для автоматизации определенных аспектов этого процесса, все еще существует потребность в проверке сетей, созданных автоматически вручную, по экспериментальным биохимическим результатам (Segre et al., 2003). Для метаболических сетей также были достигнуты значительные успехи в моделировании реакций, которые происходят в таких сетях. Общая структура сети может быть описана несколькими различными параметрами. Например, среднее количество соединений, которые узел имеет в сети, или вероятность того, что узел имеет заданное число соединений. Теоретическая работа показала, что разные модели создания сети будут давать разные значения для этих параметров. Классическая теория случайных сетей (Erdös & Renyi, 1960) утверждает, что при заданном наборе узлов соединения между узлами устанавливаются случайным образом.Это дает сеть, в которой у большинства узлов одинаковое количество подключений. Недавние исследования показали, что эта модель не соответствует структуре нескольких важных сетей. Вместо этого эти сложные сети лучше описывать с помощью так называемой безмасштабной модели, в которой у большинства узлов есть только несколько соединений, но несколько узлов (называемых концентраторами) имеют очень большое количество соединений. Недавняя работа показывает, что метаболические сети являются примерами таких безмасштабных сетей (Jeong et al., 2000). Этот результат важен и, вероятно, приведет к новому пониманию функции метаболических и сигнальных сетей, а также к эволюционной истории сетей.Устойчивость — еще одно важное свойство метаболических сетей. Это способность сети обеспечивать по существу одно и то же поведение, даже когда различные параметры, управляющие ее компонентами, варьируются в значительных пределах. Напр., Недавняя работа указывает на то, что сеть полярности сегментов у эмбриона Drosophila может удовлетворительно функционировать с удивительно большим количеством случайно выбранных наборов параметров (von Dassow et al. , 2000). Параметры не нужно тщательно настраивать или оптимизировать.Это имеет биологический смысл, а это означает, что метаболическая сеть должна быть толерантной к мутациям или большим изменениям окружающей среды.
Другой важной новой темой исследования является понимание метаболических сетей с точки зрения их функции в организме и с учетом уже имеющихся данных. Это требует объединения информации из большого количества источников, таких как классическая биохимия, геномика, функциональная геномика, эксперименты с микрочипами, сетевой анализ и моделирование.Теория клетки должна сочетать описание структур в ней с теоретическим и вычислительным описанием динамики жизненных процессов. Одна из важнейших задач будущего — как сделать всю эту информацию понятной с биологической точки зрения. Это необходимо для облегчения использования информации в целях прогнозирования, чтобы предсказать, что произойдет после определенного набора обстоятельств. Такого рода предсказательная сила будет достигнута только в том случае, если сложность биологических процессов можно будет обрабатывать с помощью вычислений.
4. Измерение центральности и важности в биомолекулярных сетях
Биологическая функция — чрезвычайно сложное следствие действия большого количества различных молекул, которые взаимодействуют по-разному. Геномные ассоциации между генами отражают функциональные ассоциации между их продуктами (белками) (Huynen et al., 2000; Yanai et al., 2001). Кроме того, сила геномных ассоциаций коррелирует с силой функциональных ассоциаций.Гены, которые часто встречаются в одном опероне у разных видов, с большей вероятностью будут физически взаимодействовать, чем гены, которые встречаются вместе в опероне только у двух видов ((Huynen et al., 2000), и белки, связанные слиянием генов или сохранение порядка генов, скорее всего, будут субъединицами комплекса, чем белки, которые просто кодируются в тех же геномах (Enright et al., 1999). Для сетевых исследований использовались другие типы ассоциаций, но они сосредоточены на определенных определенные типы функциональных взаимодействий, такие как последующие ферментативные этапы метаболических путей или физические взаимодействия.Выяснение вклада каждой молекулы в определенную функцию казалось бы безнадежным, если бы эволюция не сформировала взаимодействие молекул таким образом, чтобы они участвовали в функциональных единицах или строительных блоках функции организма (Callebaut et al., 2005). Эти строительные блоки можно назвать модулями, чьи взаимодействия, взаимосвязи и отказоустойчивость можно исследовать с точки зрения более высокого уровня, что позволяет получить синтетический, а не аналитический взгляд на биологические системы (Sprinzak et al., 2005). Признание модулей как отдельных объектов, функции которых отделимы от функций других модулей (Hartwell et al., 1999), вводит критический уровень биологической организации, который позволяет проводить исследования in silico и .
Интуитивно понятно, что модульность должна быть следствием эволюционного процесса. Модульность подразумевает возможность изменения с минимальным нарушением функции, функция, которая напрямую выбирается для (Wilke et al., 2003). Однако, если модуль необходим, его независимость от других модулей не имеет значения, если только в случае нарушения его функция не может быть восстановлена либо избыточным геном, либо альтернативным путем или модулем.Более того, модульность должна влиять на сами эволюционные механизмы, поэтому устойчивость и эволюционируемость можно оптимизировать одновременно (Lenski et al., 2006). Анализ этих концепций требует как понимания того, что составляет модуль в биологических системах, так и инструментов для распознавания модулей среди групп генов. В частности, системный взгляд на биологическую функцию требует разработки словаря, который не только классифицирует модули в соответствии с ролью, которую они играют в сети модулей и мотивов, но также и тем, как эти модули и их взаимосвязи изменяются в процессе эволюции, например, как они составляют единицы эволюции, на которые нацелен непосредственно процесс отбора (Schlosser et al., 2004). Идентификация биологических модулей обычно основана либо на функциональных, либо на топологических критериях. Например, гены, которые коэкспрессируются или корегулируются, можно классифицировать по модулям, идентифицируя их общие факторы транскрипции (Segal et al., 2004), в то время как гены, которые сильно связаны ребрами в сети, образуют кластеры, которые только слабо связаны с другие кластеры (Rives et al., 2003). С точки зрения эволюции гены, которые наследуются вместе, но не вместе с другими, часто образуют модули (Snel et al., 2004; Слоним и др., 2006). Однако концепция модульности вообще не определена. Например, доля белков, составляющих ядро модуля и наследуемых вместе, мала (Snel et al., 2004), что подразумевает, что модули нечеткие, но также гибкие, поэтому их можно быстро перепрограммировать, что позволяет организму адаптироваться к новым обстоятельствам (Campillos et al., 2006).
Набор данных обеспечивается генетическими взаимодействиями (Reguly et al., 2006), такими как синтетические летальные пары генов или пары спасения дозы, в которых нокаут или мутация гена подавляется чрезмерной экспрессией другого гена.Такие пары интересны, потому что они открывают окно для клеточной устойчивости и модульности, вызванных условной экспрессией генов. В самом деле, взаимодействие между генами эпистаза (Wolf et al., 2000) было использовано для успешной идентификации модулей в метаболических генах дрожжей (Segre et al., 2005). Однако часто взаимодействующие пары генов лежат в альтернативных путях, а не в функциональных модулях. Эти гены не взаимодействуют напрямую, и поэтому ожидается, что они будут перемещаться между модулями чаще, чем лежать в одном (Jeong et al., 2000).
In silico evolution — мощный инструмент, если могут быть созданы сложные сети, которые разделяют всеобъемлющие характеристики биологических сетей, такие как устойчивость к ошибкам, возможность соединения в малых масштабах и безмасштабное распределение степеней (Jeong et al., 2000). Если, кроме того, каждый узел в сети представляет смоделированные химические или белковые реакции, катализирующие эти молекулы, то можно провести подробный функциональный анализ сети, моделируя эксперименты по нокдауну или сверхэкспрессии.Затем эти функциональные данные могут быть объединены с эволюционной и топологической информацией, чтобы прийти к более четкой концепции модульности, которая может быть протестирована in vitro, когда станет доступно больше генетических данных. Предыдущая работа по in silico эволюции метаболизма (Pfeiffer et al., 2005), передачи сигналов (Soyer & Bonhoeffer, 2006; Soyer et al., 2006), биохимического (Francois et al., 2004; Paladugu et al., 2006) , регуляторный (Ciliberti et al., 2007), а также булевский (Ma’ayan et al., 2006), электронный (Kashtan et al., 2005), а нейронные (Hampton et al., 2004) сети начали обнаруживать, как свойства сети, такие как концентрированность, масштабирование, мутационная устойчивость, а также короткая длина пути, могут проявляться в чисто дарвиновских условиях. В частности, in silico эксперименты , проверяющие эволюцию модульности как в абстрактных (Lipson et al., 2002), так и в смоделированных электронных сетях, предполагают, что изменение окружающей среды является ключом к модульной организации функций. Эти сети сложны, топологически интересны (Adami, 2002) и функционируют в смоделированных средах с различной изменчивостью, которыми можно произвольно управлять.
5. Идентификация мотивов или функциональных модулей в биологических сетях
Биологические системы, рассматриваемые как сети, можно легко сравнить с инженерными системами, которые традиционно описываются сетями, такими как блок-схемы. Примечательно, что при таком сравнении биологические сети и инженерные сети, как видно, разделяют структурные принципы, такие как модульность и повторяемость элементов схемы (Alon, 2003). Функционирование и инженерия биологических систем организованы по модульному принципу.Инженерные системы можно разделить на функциональные модули на разных уровнях (Hansen et al., 1999), подпрограммы в программном обеспечении (Myers, 2003) и сменные части в машинах. В случае биологических сетей, хотя нет единого мнения о точных группах генов и взаимодействиях, которые образуют модули, ясно, что они обладают модульной структурой (Babu et al., 2004). Алон предложил рабочее определение модуля, основанное на сравнении с инженерным. Модуль в сети — это набор узлов, которые имеют сильные взаимодействия и общую функцию (Alon, 2003).В модуле определены входные и выходные узлы, которые управляют взаимодействием с остальной частью сети.
Различные базовые функциональные модули часто повторно используются в инженерных и биологических системах. Например, цифровая схема может включать в себя множество базовых функциональных модулей, таких как мультиплексоры и так далее (Hansen et al., 1999). Биология демонстрирует тот же принцип, снова и снова используя ключевые схемы разводки по всей сети. Например, метаболические сети используют регуляторные цепи, такие как подавление обратной связи во многих различных путях (Alon, 2003).Помимо базовых функциональных модулей, недавно был обнаружен небольшой набор повторяющихся элементов схемы, названный мотивом , , в широком диапазоне биологических и инженерных сетей (Milo et al., 2002). Мотивы — это небольшие (примерно 3 или 4 узла) подграфы, которые встречаются в реальных сетях значительно чаще, чем ожидалось случайно, и обнаруживаются исключительно с помощью топологического анализа. Это открытие вызвало большой интерес к организации и функции мотивов, и в последние годы было опубликовано множество статей по теме.Наблюдаемое чрезмерное представление мотивов было интерпретировано как проявление функциональных ограничений и принципов проектирования, которые сформировали архитектуру сети на локальном уровне (Milo et al., 2002). Некоторые исследователи считают, что мотивы — это базовые строительные блоки, которые могут выполнять определенные функции в качестве элементарных вычислительных схем (Milo et al., 2002). Хотя мотивы кажутся тесно связанными с традиционными строительными блоками, их взаимосвязь не имеет адекватного и точного анализа, а их метод интеграции в полные сети не был полностью исследован.Кроме того, неясно, что определяет частоту всех возможных сетевых мотивов в конкретной сети.
6. Поиск новых путей из биомолекулярных сетей
При изучении организмов на системном уровне биологи недавно упомянули (Kelley et al., 2003) следующие вопросы: (1) Существует ли минимальный набор необходимых путей? всеми организмами? (2) В какой степени сохраняются геномные пути у разных видов? (3) Как организмы связаны скорее по расстоянию между путями, чем по уровню сходства последовательностей ДНК? В основе таких вопросов лежит определение путей в различных организмах.Однако экспериментальная проверка огромного количества возможных кандидатов в условиях влажной лаборатории требует огромного количества времени и усилий. Таким образом, существует потребность в инструментах сравнительной геномики, которые помогают ученым предсказывать пути в биологической сети организма. Из-за сложной и неполной природы биологических данных в настоящее время полностью автоматизированное прогнозирование вычислительных путей является чрезмерно амбициозным. Метаболический путь — это набор биологических реакций, каждая из которых потребляет набор метаболитов, называемых субстратами, и производит другой набор метаболитов, называемых продуктами.Реакция катализируется ферментом (или белком) или набором ферментов. Существует множество веб-ресурсов, которые обеспечивают доступ к тщательно подобранным, а также прогнозируемым коллекциям путей, например, KEGG (Kanehisa et al. 2004), EcoCyc (Keseler et al. 2005), Reactome (Joshi-Tope et al. 2005) и PathCase (Озсойоглу и др., 2006). На сегодняшний день работа по обнаружению биологических сетей может быть организована под двумя основными названиями: (i) Определение пути (Yamanishi et al., 2007; Shlomi et al., 2006) и (ii) Whole-Network Detection (Tu et al., 2006; Яманиши и др. 2005). Даже при наличии геномной схемы живой системы и функциональных аннотаций ее предполагаемых генов экспериментальное выяснение ее биохимических процессов по-прежнему остается сложной задачей. Хотя возможно организовать гены по широким функциональным ролям, объединение их вручную в последовательные биохимические пути быстро становится трудноразрешимым. Ряд инструментов реконструкции метаболических путей был разработан с момента появления первого микробного генома, Haemophilus influenza (Fleischmann et al., 1995). К ним относятся PathoLogic (Karp & Riley, 1994), MAGPIE (Gaasterland & Sensen, 1996) и WIT (Overbeek et al., 2000) и PathFinder (Goesmann et al., 2002). Целью большинства методов определения путей обычно было сопоставление предположительно идентифицированных ферментов с известными или эталонными путями. Хотя реконструкция является важной отправной точкой для выяснения метаболических возможностей организма на основе предшествующих знаний о путях, реконструированные пути часто имеют много отсутствующих ферментов, даже в основных путях.Проблема переопределения микробных биохимических путей на основе отсутствующих белков важна, так как существует множество примеров альтернатив стандартным путям у различных организмов (Cordwell, 1999). Более того, создание нового пути проникновения в организм с помощью гетерологичных ферментов также требует способности вывести новые биохимические пути. В связи с тем, что все больше проектов по геномному секвенированию находится в стадии реализации, и отсутствуют надежные функциональные характеристики многих генов, автоматизированные стратегии прогнозирования биохимических путей могут помочь биологам в разгадывании сложных процессов в живых системах.В то же время подходы к выводу о путях также могут помочь в разработке синтетических процессов с использованием репертуарных биокатализаторов, доступных в природе.
7. Заключение
Крупномасштабные данные о биомолекулярных взаимодействиях, которые становятся все более доступными, позволяют заглянуть в сложные сотовые сети. Математическая теория графов — это простой способ представления этой информации, а модели на основе графов могут использовать глобальные и локальные характеристики этих сетей, относящиеся к клеточной биологии.Более того, потребность в более систематическом подходе к анализу живых организмов, наряду с доступностью беспрецедентных объемов данных, привела к значительному росту активности в теории и анализе сложных биологических сетей в последние годы. Сети повсеместны в биологии, происходящие на всех уровнях от биохимических реакций внутри клетки до сложных сетей социальных и сексуальных взаимодействий, которые управляют динамикой распространения болезней среди людей. Сетевые графы имеют то преимущество, что они очень просты для размышлений и в целом соответствуют информации, которая сегодня глобально доступна на сетевом уровне.Однако, хотя информация о бинарных отношениях действительно представляет собой важный аспект сетей взаимодействия, многие биологические процессы, по-видимому, требуют более подробных моделей. Для разработки более сложных и эффективных стратегий лечения таких заболеваний, как рак, необходимо всестороннее понимание этих сетей. В конечном итоге это может доказать, что математические модели крупномасштабных наборов данных ценны для медицинских проблем, таких как определение ключевых игроков и их взаимоотношений, ответственных за многофакторное поведение в сетях болезней человека.В заключение можно сказать, что анализ биологических сетей необходим в области исследований в области биоинформатики, и это волнующие проблемы. Надеемся, что эта глава будет полезна исследователям, освещая последние достижения в этой области.
.Телеологических понятий в биологии (Стэнфордская энциклопедия философии)
1. Обстановка дебатов
Дискуссия о биологической телеологии имеет древние корни. это особенно заметно в описании Платоном божественного Ремесленник или «Демиург» в Тимей и обсуждение Аристотелем конечных причин в Физика (см. Раздел о телеологии в статье о Аристотель). Однако понимание Платоном и Аристотелем телеологии, а также их аргументы в пользу телеологии в естественном мир, различны (Lennox 1992; Ariew 2002; Johnson 2005).В то время как Платон телеология антропоцентрична и креационистка, Аристотель — натуралистический и функциональный. С платонической точки зрения, Демиург — это источник всего движения как на небесах, так и на земле, и Вселенная и все живые существа в ней — артефакты, смоделированные на Формы (см. Соответствующие разделы записей на Платон и Платоновская метафизика). Цель, к которой стремятся все, включая живые существа. направлено внешнее и вечное благо Форм. Напротив, с точки зрения Аристотеля, телеология, которая управляет поведением живые существа имманентны.Например, в развитии организма толчком для этого целенаправленного процесса является принцип изменения внутри организма, и телос , или цель, развития также неотъемлемое свойство. Хотя часто взгляды Платон и Аристотель по телеологии оказали влияние на историческую науку. дебаты по биологической телеологии, и все еще можно найти платонические и Аристотелевские идеи в нынешних дебатах о биологических функциях.
Помимо своей роли в античной философии и космологии, телеология долгое время была важной темой в физиологии и лекарство. Галена об использовании деталей ( De usu partium ) является ранним примером телеологического рассуждения, применяемого к физиология (см. раздел о телеологии в статье о Гален). В этом тексте Гален представляет функциональный анализ различных части живых организмов, в которых «существование, структура и атрибуты всех частей должны быть объяснены со ссылкой на их функции в содействии деятельности всего организма » (Шифски 2007: 371). Для Галена телеологическое описание частей превосходит чисто причинно-механический, поскольку функция или цель роли играет неотъемлемую роль в объяснении часть и ее деятельность.Этот галеновский взгляд на анатомию с ее явно аристотелевская опора на конечные причины в значительной степени преобладала медицинская мысль до семнадцатого века. Уильям Харви О движении сердца и крови ( De motu cordis ) видели многие его современники (например, Гоббс, Декарт) как поворотный момент в сторону от галеновского или аристотелевского подходов к анатомии, с их апелляцией к конечным причинам, к новая механистическая и экспериментальная наука семнадцатого века (Французский 1994).Его попытка эмпирически установить структуру и движение сердца, даже не утверждая, что определил последний причина обращения, а также использование им механических аналогий, таких как как аналогия от расширения артерий к раздуванию перчатка, поддержите эту оценку. Роберт Бойл, однако предложил иное толкование Харви работай. Бойль считал это приемлемым для своего «компатибилиста» подход, который пытался показать, что «механический и телеологические объяснения биологических явлений совместимы »(Lennox 1983: 38).Недавние комментаторы также предположил, что Харви находился под сильным влиянием аристотелевской телеологической мышление, и поэтому является лиминальной фигурой перехода от виталистической к чисто механистическому объяснению в физиологии и медицине (Distelzweig 2014, 2016; Lennox 2017), хотя Леннокс отмечает, что «Харви, похоже, проводит аналогию между искусством и природой способами, которые не аристотелевские, а более платонические в дух »(Леннокс 2017: 191).
Анализ телеологии Иммануила Канта в книге Critique of Решение (Kant 1790 [2000]) также сыграло важную роль в биология.Согласно Канту, люди неизбежно понимают живое. как если бы они были телеологическими системами (Zammito 2006). Однако на По мнению Канта, телеология, которую мы видим в мире природы, очевидный; это продукт наших ограниченных когнитивных способностей (см. раздел 3 статьи о Канта эстетика). Но кроме того, по Канту, есть определенная немашинный характер организмов, проявляющийся в их способности к расти и воспроизводиться, что приводит к механическому необъяснимость.Ханна Гинсборг (2004) утверждает, что для Канта это невозможность объяснения организмов исключительно механистическими терминами делает не отличить их от сложных артефактов; но, утверждает она, Кант считал, что регенеративные и репродуктивные аспекты организмов заставляет нас приписывать своего рода естественную целесообразность, которая отсутствует в артефактах, что соответствует оправданию Аристотелем естественная, имманентная телеология. Кантовский анализ такого рода обнаруживается в исследования начала девятнадцатого века в том, что впоследствии назовут органическая химия.Ученые того времени пытались определить, живые системы были не более чем сложными химическими системами, полностью поддается анализу с точки зрения физических и химических процессов. Те исследователи, принявшие кантовский подход, выступали за телеомеханиста стратегия осмысления целенаправленной природы живых систем, которые стремились рассматривать организм как средство и как цель, и таким образом включены элементы как телеологического, так и механистического объяснения (Ленуар 1982).
В восемнадцатом и девятнадцатом веках статус телеологии в биологии был также оспаривается как часть дебатов виталистов и механистов в физиологии и медицина (обзор см. в первом разделе статьи о жизнь).В то время как механики стремились описать все живое в чисто механические термины, виталисты утверждали, что только физические свойства не мог объяснить целенаправленную организацию живых существ. Они утверждали, что «жизненные силы» также необходимы для объясните разницу между физическими и живыми системами. Хотя они попали в немилость, некоторые виталистические учения сохранялись в двадцатого века, например, в книге философа Анри Бергсона ‘ élan vital ’ (см. Раздел креатив эволюция в статье о Бергсон) и в концепции биолога и философа Ганса Дриша о «Энтелехия» (Дриш 1908).
Много споров о роли телеологии в биологии двадцатого века. века, особенно среди архитекторов «модерна» синтез », восходит к теории Чарльза Дарвина эволюция путем естественного отбора. Биолог и философ Майкл Гизелин, выражая общую интерпретацию роли Дарвина в эти дебаты, утверждает в своем предисловии к работе Дарвина об орхидеях что теория Дарвина смогла «избавиться от телеологии и заменив ее новым взглядом на адаптация »(Darwin 1862/1877 [1984: xiii], цит. по: Lennox 1993: 409).С этой точки зрения теория естественного отбора объясняет, как виды «были изменены, чтобы обрести это совершенство структура и соадаптация »без обращения к доброжелательному Создатель (Дарвин 1859: 3). До Дарвина лучшее объяснение биологическая адаптация была аргументом от дизайна, наиболее влиятельным представлено в книге Уильяма Пэли Natural Theology (Paley 1802): живые существа имеют ту же структуру и поведение, что и потому что они были созданы для определенных целей доброжелательной Создатель (см. Запись по естественному богословию и естественной религии).Теория Дарвина предусматривает биологии с ресурсами, чтобы противостоять этому аргументу, предлагая полностью натурализованное объяснение адаптации. Хотя большинство согласны с тем, что Теория Дарвина действительно очищает эволюционную биологию от любого незаконное обращение к внешней, платонической телеологии, есть разногласия относительно того, являются ли эволюционные объяснения Дарвина телеологический (см. соответствующий раздел статьи о Дарвинизм). Даже современники Дарвина расходились во мнениях относительно того, теория естественного отбора очистила телеологические объяснения от биологии или возродили их (Lennox 2010).В любом случае ясно, что Дарвин использовал язык «конечных причин» для описания функция биологических частей в его «Записках о видах» и во всем его жизнь; он также часто размышлял об отношениях между естественный отбор и телеология (Леннокс, 1993).
2. Объяснительный телеонатурализм
Философский натурализм обозначает широкий диапазон отношения к онтологическим вопросам. Мы использовать «телеонатурализм» для обозначения столь же широкого диапазона натуралистические объяснения телеологии в биологии, которые объединены в отказ от любой зависимости от ментальных или преднамеренных представлений в объяснение использования телеологических терминов в биологических контекстах.Таким образом, те, кто отвергает телеоментализм, обычно ищут условия истины. для телеологических утверждений в биологии, основанных на нементальных факты об организмах и их свойствах.
Некоторые телеонатуралисты анализируют телеологический язык как прежде всего описательный, а не объяснительный, поддерживая эту телеологию в биология подходит для биологических систем, которые демонстрируют целенаправленное, целенаправленные модели поведения (для которых Питтендрай (1958) ввел термин «телеономический»). Для таких представлений основной научная задача состоит в том, чтобы объяснить телеономию, а не использовать телеономию как объясняется (см. Thompson 1987).В то время как кибернетика потеряла свою привлекательность в во второй половине двадцатого века, более современные подходы к живые системы, которые рассматривают их как самоорганизующиеся или «Аутопоэтический» (Maturana & Varela 1980) несомненен близость к описательному отношению к телеонатурализм — хотя большинство сторонников утверждают, что концепции разработанные в рамках этих подходов носят пояснительный характер.
Следовательно, большинство телеонатуралистов отдают предпочтение объяснению биологической функции. что делает объяснительную роль этого понятия желаемой.Натуралистические отчеты обычно стремятся удовлетворить два дополнительных Desiderata. Они должны отличать настоящие биологические функции от случайное использование (например, носы, поддерживающие очки), и они должны зафиксировать нормативное измерение функции, чтобы сохранить различие между функцией и неисправностью. Хотя эти три Desiderata ни общепризнанных, ни условий адекватности в в строгом смысле, они, тем не менее, достигли канонического статуса в современные дебаты о биологических функциях.
В следующих разделах мы разделим различные способы, которыми Телеонатуралистические описания функций можно выделить. Наш первый различие между (а) взглядами, которые ассимилируют функциональные объяснения в биологии к шаблонам объяснения в небиологические науки, и (б) точки зрения рассматривают функциональное объяснение как отчетливо биологический.
3. Ассимиляция к небиологическим объяснениям
Эрнест Нагель (1961) и Карл Хемпель (1965) предоставляют ранние попытки философы науки непосредственно усваивают функциональное объяснение в биологии к более общим схемам объяснения.В частности, они оба рассматривают функциональное объяснение в рамках Дедуктивно-номологический учет научного объяснения. Они считают, что функциональные утверждения связаны с объяснениями наличие в организме признака. Их счета различаются в первую очередь от того, следует ли говорить, что признак T имеет функцию F в организме O когда T достаточно для производства F в O (версия Hempel) или когда T необходимо производить F в O (версия Нагеля).
Ларри Райт (1973, 1976) также предлагает пояснительный отчет, ориентированный на на наличие признака, но он критикует предыдущие отчеты за неспособность уловить очевидную целенаправленность функциональных черты. Его так называемый «этиологический» анализ утверждает, что функция X — Z означает: (a) X есть потому что он выполняет Z , и (б) Z является следствием (или результат) наличия X . Из-за предполагаемого широты анализа Райта критиковались концептуальные основания (e.g., Boorse 1976), но общая направленность этиологического счет выживает в счетах функций, основанных на естественном отборе, обсуждается в разделе 4.
Камминс (1975) критиковал Хемпеля и Нагеля на том основании, что правильная цель объяснения заявлений о биологической функции не наличие признака, но возможности биологических органов и организмы. Сложные возможности можно проанализировать с точки зрения вклад, который их компоненты вносят в эти возможности.Для Например, сердце двустворчатого животного перекачивает кровь, которая в этом способ способствует способности организма доставлять кислород и питательные вещества для его тканей. Само сердце можно еще разложить на части (камеры, клапаны и т. д.), каждая из которых играет разные функциональные роли в содействии способности этого органа качать кровь. Среди философов такой подход к функциональному анализу больше всего ассоциируется с Cummins (1975), хотя биологи выдвигал аналогичные идеи, иногда независимо (Hinde 1975; Lauder 1982), и этот подход можно проследить гораздо раньше, поскольку цитата из Уильяма Харви во введении подсказывает.Идеи Cummins о функциональном анализе были учтены в недавней дискуссии о механизмах в биологических науках (см. вход на механизмы в науке). Например, Craver (2007) явно опирается на Cummins (1975) в своей счет и важен для переноса телеологических описаний на на молекулярном уровне (см. также Craver 2001, 2013).
Согласно Cummins, хотя биологические системы, безусловно, имеют возможности, которые являются однозначно биологическими, нет ничего конкретного биологические о модели объяснения, предлагаемой функциональными анализ; он одинаково хорошо применим, например, к взносам сделаны компонентами артефактов (Lewens 2004), например вклад поршней двигателя в качестве автомобиль для перевозки людей.Из-за общности рамки, также можно дать функциональный анализ того, как какая-то часть биологической системы обычно способствует результатам рассматривается как отрицательный, например болезнь или смерть. Некоторые комментаторы считают это достоинством подхода, в то время как другие считают это слишком отдельно от стандартной биологической практики. Соответственно, функционал аналитический подход не соответствует общепринятому желанию адекватный отчет должен обеспечивать анализ неисправности — desideratum, который Cummins явно отвергает (см. также Wouters 1999 и Дэвис 2001).Сердце с дырой в перегородке может не циркулировать кровь на уровне, достаточном для поддержания жизни, но тем самым просто не хватает этой функции.
4. Счета естественного отбора
Многие философы биологии считают, что функциональное объяснение уникально подходит для биологии, обращаясь к теории Дарвина спуск с модификацией, чтобы обосновать практику приписывания функции. Как Райт, Хемпель и Нагель, естественный отбор телеонатуралисты считают, что основной целью объяснения является наличие у организмов различных признаков.
Здесь мы различаем два способа использования естественного отбора для наземная биологическая телеология.
- Косвенные подходы рассматривают адаптивную, самоорганизующуюся природу живые клетки и организмы как естественная основа телеологических свойства их черт, но отдать должное силе естественный отбор, чтобы произвести такую самоорганизационную сложность, как есть найдено в живых системах.
- Прямые подходы явно вызывают естественный отбор, когда объяснение функциональных утверждений, либо в этиологическом смысле на основе история выбора или в диспозиционном смысле на основе приспособленность организмов, обладающих признаками.
4.1 Косвенный
Основная мотивация самых ранних косвенных, кибернетических отчетов биологической телеологии должны были объяснить очевидную целесообразность биологические организмы, например, поддержание постоянного тела температура в эндотермах. Эти учетные записи были нацелены на натурализованное объяснение целенаправленного поведения биологических системы через ссылку на их организацию. Во влиятельном В ранней статье Норберт Винер и его коллеги пытались объяснить целенаправленное поведение биологических организмов и машин как возникающее от использования ими механизмов отрицательной обратной связи (Rosenblueth et al. al.1943; для дальнейшего развития см. также Braithwaite (1953), Зоммерхофф (1950) и Нагель (1953)). Атрибуции телеологических или они утверждали, что целенаправленное поведение по отношению к животным или машинам означает не более чем «цель, управляемая обратной связью» (Rosenblueth et al. 1943, 23).
Это кибернетическое описание телеологии вдохновило биолога Колина. Питтендрай, чтобы ввести термин «телеономия» в литературе (Питтендрай, 1958). Питтендрай надеялся, что этот неологизм очистить биологию от любых остатков аристотелевских конечных причин, пока обеспечение биологии приемлемым термином для описания адаптированных, целенаправленные системы.Этот термин был использован в 1960-х гг. эволюционные биологи, такие как Эрнст Майр (1974) и Джордж Уильямс (1966), а также учеными, изучающими клеточный метаболизм и регулирования, которые только начинали выяснять структурные и молекулярная основа механизмов клеточной обратной связи (Monod & Jacob 1961; Дэвис 1961). По мнению сторонников, принятие кибернетического учет целенаправленного поведения в биологических системах разделяет объяснительная проблема в двух. С одной стороны, телеологическая активность в биологический мир можно объяснить наличием телеономических системы с механизмами отрицательной обратной связи, тогда как само наличие этих телеономных систем в живых организмах, с другой стороны, можно объяснить действием естественного отбора (Monod 1970 [1971]).
Хотя явные кибернетические описания биологической телеологии вышли из моды, другие организационные подходы к биологическому Функция недавно получила возрождение в литературе о функциях. Эти организационные или теоретико-системные подходы часто создают на ранних кибернетических счетах или стремятся расширить Maturana и Влиятельное понятие автопоэзиса Варелы (1980), относящееся к к самоорганизующейся, самоподдерживающейся характеристике жизни системы (см. запись о Воплощенное познание для дальнейшего описания).Эти учетные записи определяют функцию биологическая черта посредством анализа роли, которую эта черта играет внутри организованной системы, способствуя как своей собственной устойчивости и устойчивость системы в целом (Schlosser 1998; McLaughlin 2001; Mossio et al. 2009; Saborido et al. 2011; Moreno И Mossio 2015). Хотя они различаются деталями, организационные подходы к биологической функции обычно соглашаются с маркер признака T имеет функцию F , когда выполнение F по T участвует в обслуживании комплекса организация системы, что, в свою очередь, приводит к T продолжение существования.Например, у сердца есть функция качать кровь, согласно этим отчетам, потому что она способствует поддержание всего организма за счет циркуляции крови, который облегчает циркуляцию кислорода и питательных веществ. На В то же время этот оборот также частично отвечает за настойчивость самого сердца, так как сердце тоже приносит пользу непосредственно из этой функции (то есть сердечные клетки получают кислород и питательные вещества, необходимые для их выживания).
Подобно учетным записям прямого естественного отбора, учетные записи организаций может быть направленным вперед или назад: функция признака может определить его диспозиционный вклад в сложную организацию система, которая приводит к ее собственному постоянству или воспроизводству в будущее (перспективный; Schlosser 1998) или функциональная атрибуция может определить прошлый вклад характеристики (этиологический или ретроспективный; Маклафлин 2001).Группа Альваро Морено перенимает третья позиция. Они заявляют о своей организационной концепции функции объединяет эти две точки зрения (Mossio et al 2009; ср. Artiga & Мартинес 2016). Все эти организационные аккаунты отличаются от прямых естественного отбора, однако, в том, что они не обращаются к история выбора признака. Вместо этого функция признака можно вывести из настоящей или прошлой роли черты в поддерживая себя в сложной организованной системе без далее считая, что черта была выбрана для этой роли.На этом точки зрения, функциональные атрибуции в биологии объясняют не потому, что отбора, а скорее из-за причинной роли, которую черты характера играют в способствуя поддержанию организации системы, которая в свою очередь позволяет самим чертам сохраняться.
4.2 Подходы прямого естественного отбора
Учет биологической функции, относящейся к естественному отбору обычно имеют форму, которую функции черты каузально объясняют наличие или поддержание этой черты в данной популяции через механизм естественного отбора.Уильям Вимсатт (1972), Рут Милликен (1984) и Карен Неандер (1991a) рассматривают прошлую историю. естественного отбора как процесса отбора, узаконивающего понятие биологической функции. В рамках таких подходов есть спор о точной роли естественного отбора, как источника вариации (иногда называемой «творческой» ролью естественного отбора, например, Neander 1988; также Ayala 1970, 1977), или только как фильтр для вариаций, возникающих независимо (Sober 1984).
Позиции, лежащие в основе функциональных требований естественного отбора много общего с этиологическим описанием Райта. Тем не мение, поскольку заземление специфично для биологии, они могут избегать контрпримеров к счету Райта, представленных критиками таких как Кристофер Бурс, основанный на идее Райта счет предназначен для обеспечения более общего концептуального анализа. А Связанная с этим проблема проистекает из утверждения, что додарвиновские мыслители например, Харви правильно определил функциональные свойства биологических органов, и поэтому естественный отбор не может быть требование правильного концептуального анализа функции.Defenders объяснений функций прямого естественного отбора ответили в различные пути. Один из способов, проиллюстрированный Милликеном (1989), — это аргументировать этот концептуальный анализ не играет роли в формулировании того, что по сути, теоретический термин в современной эволюционной биологии. Другой способ, приведенный Неандером (1991b), состоит в том, чтобы сказать, что задача концептуального анализа подходит, но ограничивается концепциями соответствующего научного сообщества.
Пол Дэвис (2001) и Арно Воутерс (2005) утверждают, что и Милликен и Неандер неправильно рассматривают неисправность как важную теоретический или концептуальный аспект практики атрибуции функции биологами.Воутерс заявляет о желании, чтобы изучение биологическая функция должна быть освобождена «от ига философия разума »(2005: 148). Однако Эма Салливан-Биссетт (2017) утверждает что, хотя задача объяснения биологической практики философами биологии полезно отличить от более широких целей философы, стремящиеся к натуралистическим объяснениям разума и языка (см. запись на телеологические теории ментального содержания), последний служит законным целям. Она считает, что неисправность должна быть неотъемлемой частью последнего проекта, даже если не бывший.Дэвис (2001) утверждает, что счета естественного отбора не в состоянии предоставить отчет о неисправности, поскольку они функционально индивидуализировать черты, в результате чего предположительно неисправный признак не является экземпляром функционально определенного Добрый. Салливан-Биссет рассматривает возражение Дэвиса следующим образом: включение структурного условия на индивидуацию черт. (См. Также Гарсон 2016: 48–49, дополнительное обсуждение и критика точки зрения Дэвиса.)
Возвращаясь к чертам характера, изучаемым биологами, некоторые теоретики различать первоначальное распространение нового фенотипа черта в популяции и недавнее поддержание черт в населения.Возьмите такую черту, как перья, возникающую в популяции что бы ни значило. Первоначально эта черта могла распространяться из-за роли в дисплеях сопряжения. Позже перья, возможно, способствовали улучшению терморегуляция. А еще позже черта могла стать более широко распространены, потому что перья являются хорошими поверхностями управления полетом. Если дисплейная или терморегуляторная функция перьев становится меньше важно в некоторых нишах, тем не менее, эта черта может сохраняться в популяция из-за отбора для выполнения функции управления полетом.В смещение функционального профиля также может быть связано с различие по форме, например, между пушистыми перьями и полетом перья.
Некоторые биологи использовали термин «предварительная адаптация», чтобы выразить идею о том, что черта, выбранная для одной функции, может оказаться очень полезной для что-то другое. Однако Гулд и Врба (1982) ввели термин «Exaptation», чтобы зафиксировать такие переходы и избежать того, что они считали чрезмерно телеологическими последствиями преадаптации, поскольку а также признать, что невыбранные черты организмов также могут использоваться для выполнения определенной функции, повышая физическую форму без каких-либо дальнейшая модификация естественным отбором (Lloyd & Gould 2017).Критики этиологического естественного отбора иногда подходят к утверждают, что ретроспективные подходы слишком расплывчаты в отношении вопросы о том, в какой момент черты приобретают или теряют функции, и что, следовательно, они не поддаются эмпирической проверке (Amundson & Лаудер 1994). Годфри-Смит (1994) независимо предложил Теория функций «современной истории» для решения этих проблемы. Точно так же Гриффитс (1993: 417) использует понятие «Последний эволюционно значимый период времени» для обработки эти проблемы, но многие критики по-прежнему не убеждены (например,g., Wouters 1999; Дэвис 2001).
Еще одна проблема, с которой сталкиваются методы прямого естественного отбора, — это очевидная полезность приписывания функций новым признакам организмов развиваются в течение одной жизни, например, способность мозга приобретать новые представления о вещах, ранее не испытанных в эволюционная линия или иммунная система для развития антитела к новым возбудителям инфекции. Ранее Милликен (1984) имел предложил понятие «производной собственной функции» для захвата вот такой пример.Совсем недавно Бушар (2013) и Гарсон (2017) разработали более подробные отчеты о производной функции, соответственно с использованием «дифференциальной стойкости» и «Дифференциальное удержание» в организме продолжительность жизни, чтобы играть роль, которую играет дифференциальное воспроизводство счета прямого естественного отбора.
Некоторые биологи и философы биологии руководствовались проблемы с ретроспективным этиологическим подходом или видением примеры из биологии, которые стремятся идентифицировать настоящие функции черта.Для решения этих вопросов они предлагают диспозиционные или дальновидный подход, который анализирует функции с точки зрения тех эффекты, которые он имеет тенденцию вызывать, настоящее или будущее сохранение признака в популяции организмы. Различные способы разъяснения этого включают Hinde’s (1975) описание сильной функции, Boorse (1976, 2002) биостатистическая теория, склонность Бигелоу и Парджеттера (1987) теория и теория отношений Уолша (1996) (см. также и Уолш & Ariew 1996).
5. Объединение и плюрализм
Некоторые теоретики отстаивают плюралистическую идею о том, что биология может включить (по крайней мере) два понятия функции, одно для объяснения наличие черт и другое, чтобы объяснить, как эти черты вносят свой вклад в комплексные способности организмов (Милликен 1989; Sober 1993; Годфри-Смит 1994). Рон Амундсон и Джордж Лаудер (Amundson & Lauder 1994) утверждают, что палеонтология является частью биологии, которая не может использовать этиологический учет, потому что режим отбора для вымерших организмов, как правило, недоступен, и поэтому он должен зависеть от функционального анализа в стиле Cummins окаменелые останки.Однако защитники плюрализма могут ответить, что даже если правильно сказать, что не все разделы биологии могут использовать оба понятия функции, это согласуется с обоими учетными записями, имеющими роль в биологии.
Некоторые теоретики утверждали, что эти два явно разных понятия функции можно объединить, рассматривая цель объяснения как биологическая пригодность всего организма (например, Griffiths 1993; Kitcher 1994; и, возможно, Тинберген 1963 года по Питеру Годфри-Смит 1994).Морено и его коллеги (Mossio et al., 2009; Moreno & Mossio 2015) также заявили, что их организационные подход объединен для ретроспективных и перспективных счетов описывая действия, которые временно объясняют продолжающийся стойкость черт. Жизнеспособность этой учетной записи как в отличие от этиологических отчетов оспаривается Марком Артига и Маноло Мартинес (Artiga & Martínez 2016), который утверждают, что непременная характеристика разных поколений организационное закрытие, необходимое для выполнения биологических функций для родительские дары потомству (передаются через гаметы или поведение) влечет за собой стандартную этиологическую структуру, обнаруженную в Счет Райта.
Внимание к актуальным объяснительным практикам современных биологов занимает центральное место в этих дискуссиях. Этот фокус на научных практика отражает более широкие тенденции в философии науки, выходя за рамки общих и абстрактных вопросов о научных эпистемология и метафизика, которые доминировали в двадцатом веке. В разнообразие самой жизни отражается в разнообразии научных пытается понять это, и как философы биологии преследуют дальнейшее взаимодействие с этим разнообразием, новые взгляды на роль и уместность функциональных и телеологических понятий кажется вероятной к результату.
.Биология
Биология, также называемая биологическими науками, — это изучение живых организмов с использованием научных методов.
Биология изучает структуру, функции, рост, происхождение, эволюцию и распространение живых существ.
Он классифицирует и описывает организмы, их функции, способы возникновения видов и их взаимодействия друг с другом и с природной средой.
Четыре объединяющих принципа составляют основу современной биологии: клеточная теория, эволюция, генетика и гомеостаз.
Биология как отдельная наука была разработана в девятнадцатом веке, когда ученые обнаружили, что организмы имеют общие фундаментальные характеристики.
Биология в настоящее время является стандартным предметом преподавания в школах и университетах по всему миру, и более миллиона статей публикуются ежегодно в широком спектре журналов по биологии и медицине.
Большинство биологических наук являются специализированными дисциплинами.
Традиционно их группируют по типу изучаемых организмов: ботаника, изучение растений; зоология, изучение животных; и микробиология, изучение микроорганизмов.
Области биологии далее подразделяются в зависимости от масштаба изучаемых организмов и методов, используемых для их изучения: биохимия изучает фундаментальную химию жизни; молекулярная биология изучает сложные взаимодействия систем биологических молекул; клеточная биология исследует основной строительный блок всего живого — клетку; физиология изучает физические и химические функции тканей и систем органов организма; а экология изучает, как взаимосвязаны различные организмы.
Прикладные области биологии, такие как медицина и генетические исследования, включают множество специализированных дисциплин.
Центральная организационная концепция в биологии состоит в том, что жизнь изменяется и развивается в процессе эволюции и что все известные формы жизни имеют общее происхождение.
Чарльз Дарвин установил эволюцию как жизнеспособную теорию, сформулировав ее движущую силу, естественный отбор (Альфред Рассел Уоллес признан соавтором этой концепции).
Дарвин предположил, что виды и породы возникли в результате процессов естественного отбора, а также путем искусственного отбора или селекции.
Генетический дрейф стал дополнительным механизмом эволюционного развития в современном синтезе теории.
Биологическая форма и функция создаются и передаются следующему поколению генами, которые являются первичными единицами наследования.
Физиологическая адаптация организма к окружающей среде не может быть закодирована в его генах и не может быть унаследована его потомством.
Примечательно, что очень разные организмы, включая бактерии, растения, животных и грибы, имеют один и тот же базовый механизм, который копирует и транскрибирует ДНК в белки.
Например, бактерии со встроенной ДНК человека будут правильно давать соответствующий белок человека.
.
Leave A Comment