
Возрастная анатомия опорно-двигательного аппарата
Рис. 1. Филогенез скелета животных.
У животных выделяют наружный и внутренний скелет.
Наружный скелет у разных животных (рис. 1) имеет разное строение и происхождение. У многих беспозвоночных он является продуктом выделения кожного эпителия: кутикула дождевого червя, хитин членистоногих, известковые раковины молюсков.
Наружный скелет у позвоночных появляется в форме чешуи у рыб. Из чешуй у высших рыб развиваются покровные кости головы и плечевого пояса.
Чешуя рыб и кожные окостенения наземных позвоночных всегда дополняются внутренним скелетом.
Внутренний скелет у низших животных (рис. 1) развит слабо и представляет собой систему соединительнотканных образований, иногда включающих рогоподобные волокна, кремниевые или известковые иглы.
Внутренний скелет у головоногих молюсков представлен хрящом.
У позвоночных животных внутренний скелет всегда хорошо развит.
У бесчерепных он перепончатый, у низших рыб – хрящевой, у высших рыб и наземных позвоночных он построен преимущественно из костной ткани.
Рис. 2. Онтогенез скелета.
Согласно основному биогенетическому закону Геккеля-Мюллера онтогенез есть краткое повторение филогенеза. Онтогенез твердого скелета у человека не является исключением: в развитии костей у человека выделяются три последовательных стадии (рис. 2):
1. Соединительнотканная.
2. Хрящевая.
3. Костная.
Большинство костей в своем развитии последовательно проходят все три стадии – это вторичные кости. Ряд костей при развитии пропускают хрящевую стадию – это
Рис. 3. Классификация костей по развитию.

По развитию кости человека делятся на две группы (рис. 3):
- Первичные – проходят в своем развитии две стадии: соединительнотканная и костная.
- Вторичные кости – проходят в своем развитии три последовательных стадии: соединительнотканную, хрящевую и костную.
Рис. 4.Клетки, формирующие костную ткань.
Для развития костной ткани в костях необходимо наличие популяций двух видов клеток (рис. 4):
- Остеобласты.
- Остеокласты.
Остеобласты представляют собой кубовидной формы клетки (20-30 мкм в диаметре) с одним крупным ядром, располагающиеся близко друг к другу на костном матриксе (межклеточном веществе). Фибробласты продуцируют все компоненты костного матрикса. Они имеют два разных эмбриональных источника:
- нервные гребешковые клетки (выделяются из краев нервного желобка эмбриона при замыкании его в нервную трубку).
- мезенхимальные клетки закладки кости. Они дают начало пластинчатой костной ткани.
Остеокласты — многоядерные (от 2 до 100 ядер в клетке), большие (от 20 до 100 мкм) клетки гемопоэтической природы. Заносятся в соединительнотканные и хрящевые закладки костей по кровеносным сосудам. Функция остеокластов – резорбция кости.
Для формирования кости как органа необходимо совместная работа двух видов клеток: остеобластов и остеокластов.
Рис. 5. Способы окостенения.
Рис. 6. Рост костей в толщину и длину.
В зависимости от того где начинается формирование костной ткани в костях (включая их закладки) выделяют четыре способа окостенения (рис. 5):
- Эндесмальное окостенение.
- Перихондральной окостенение.
- Энхондральное окостенение.
- Периостальной окостенение.

При эндесмальном окостенении (рис. 5) первичная точка окостенения появляется в центре соединительнотканной закладки кости. Затем новообразующаяся костная ткань распространяется от цента органа к периферии. Таким способом окостеневают первичные кости. На месте первичной точки окостенения обычно наблюдается утолщение (например, теменной бугор, наружный затылочный выступ и т.п.).
Периходральное окостенение характерно для вторичных костей. Остеобласты выстраиваются на поверхности хрящевой закладки кости и начинают синтезировать костный матрикс. Это приводит с сдавливанию и нарушению трофика подлежащей хрящевой ткани, изменения которой активирует остеокласты. В результате этого на поверхности хрящевой закладки кости появляется и постепенно нарастает костная ткань (рис. 5). За счет перихондрального окостенения формируется компактное костное вещество. У длинных трубчатых костей так во внутриутробном периоде образуется диафиз.
При энхондральном окостенении точка (первичный очаг) окостенения появляется в центре хрящевой закладки кости. Затем костная ткань разрастается из центра к периферии (рис. 6). В результате этого формируется губчатое костное вещество. Этим способом развиваются вторичные кости: эпифизы и апофизы трубчатых костей, губчатые, плоские (кроме свода черепа) кости.
Периостальное окостенение происходит за счет надкостницы (
Рис. 7. Развитие костей туловища.
Рис. 8. Развитие и аномалии развития позвонков.
Рис. 9. Расщелина дуг позвонков на протяжении всех грудных позвонков.
Кости туловища по развитию относятся к вторичным костям. Они окостеневают энхондрально (рис. 7).
Развитие позвонков:
У зародыша закладывается 38 позвонков: 7 шейных, 13 грудных, 5 поясничных, 12-13 крестцовых и копчиковых (рис. 8).
13-й грудной превращается в 1-й поясничный, последний поясничный – в 1-й крестцовый, Идет редукция большинства копчиковых позвонков.
Каждый позвонок имеет первоначально три ядра окостенения: в теле и по одному в каждой половинке дуги. Они срастаются лишь к третьему году жизни.
Вторичные центры появляются по верхнему и нижнему краям тела позвонка у девочек в 6-8 лет, у мальчиков – в 7-9 лет. Они прирастают к телу позвонка в 20-25 лет.
Самостоятельные ядра окостенения образуются в отростках позвонков.
Аномалии развития позвонков (рис. 8, 9):
— Врожденные расщелины позвонков:
— Spina bifida — расщелина только дуг.
— Рахишизис – полная расщелина (тело и дуга).

— Клиновидные позвонки и полупозвонки.
— Платиспондилия – расширение тела позвонка в поперечнике.
— Брахиспондилия – уменьшение тела позвонка по высоте, уплощение и укорочение.
— Аномалии суставных отростков: аномалии положения, аномалии величины, аномалии сочленения, отсутствие суставных отростков.
— Спондилолиз – дефект в межсуставной части дуги позвонка.
— Врожденные синостозы: полный и частичный.
— Os odontoideum – неслияние зуба с телом осевого позвонка.
— Ассимиляция (окципитализация) атланта – слияние атланта с затылочной костью.
— Шейные ребра.
— Сакрализация – полное или частичное слияние последнего поясничного позвонка с крестцом.
— Люмбализация – наличие шестого поясничного позвонка (за счет мобилизации первого крестцового).
Рис. 10. Развитие и аномалии развития ребер.
Рис.
 11. Развитие и аномалии развития грудины.
11. Развитие и аномалии развития грудины.Развитие ребер (рис. 10):
Закладывается 13 пар ребер. Затем 13-е ребро редуцируется и срастается с поперечным отростком 1-го поясничного позвонка.
Основных точек окостенения в ребре две: точка окостенения на месте будущего угла ребра (окостеневает тело ребра) и в головке ребра (на 15-20 году жизни). У 10 верхних ребер появляется точка окостенения в бугорке ребра.
Передние концы 9 пар верхних ребер образуют грудные полоски – источник развития грудины.
Развитие грудины (рис. 11):
Источником развития грудины являются грудные полоски – расширенные концы хрящевых концов девяти пар верхних ребер. В грудине бывает до 13 точек окостенения.
Аномалии развития ребер (рис. 10):
— Отсутствие ребра
— Отсутствие части ребра
— Дефект ребра
— Раздвоение ребра (вилка Лушки)
— Шейное ребро
— XIII ребро
Аномалии развития грудины (рис. 11):
11):
— Аплазия рукоятки грудины
— Отсутствие отдельных сегментов тела грудины — Расщепление грудину
— Отсутствие тела грудины
— Воронкообразная деформация
— Куриная грудь
Рис. 12. Развитие костей конечностей.
Рис. 13. Развитие костей конечностей.
Рис. 14. Развитие эпифизов трубчатых костей.
Рис. 15. Развитие костей верхней конечности.
Рис. 16. Развитие тазовой и бедренной костей.
Рис. 17. Развитие бедренной кости.
Рис. 18. Развитие костей голени и стопы.
Кости конечностей по развитию относятся к вторичным костям. Исключение представляет собой ключица: ее тело и акромиальный конец окостеневают эндесмально (точка окостенения появляется на 6-7-й неделях внутриутробного развития.
Исключение представляет собой ключица: ее тело и акромиальный конец окостеневают эндесмально (точка окостенения появляется на 6-7-й неделях внутриутробного развития.
Диафизы длинных трубчатых костей окостеневают перихондральными и энходральными способами. В диафизах первичная точка окостенения появляется на 2-м – начале 3-го месяцев внутриутробного развития и растет по направлению к проксимального и дистальному эпифизам.
Эпифизы и апофизы длинных трубчатых костей окостеневают энходральным способом. Они у новорожденных хрящевые. Вторичные точки окостенения появляются в течение первых 5-10 лет жизни. Исключение составляют эпифизы костей, образующих коленный сустав: точка окостенения в дистальном конце бедренной кости появляется на 6 месяце, а в проксимальном конце большеберцовой кости – на 7 месяце внутриутробного развития. Прирастают эпифизы к диафизам после 15-17 лет и позже.
Рис. 19. Аномалии развития костей верхней конечности.
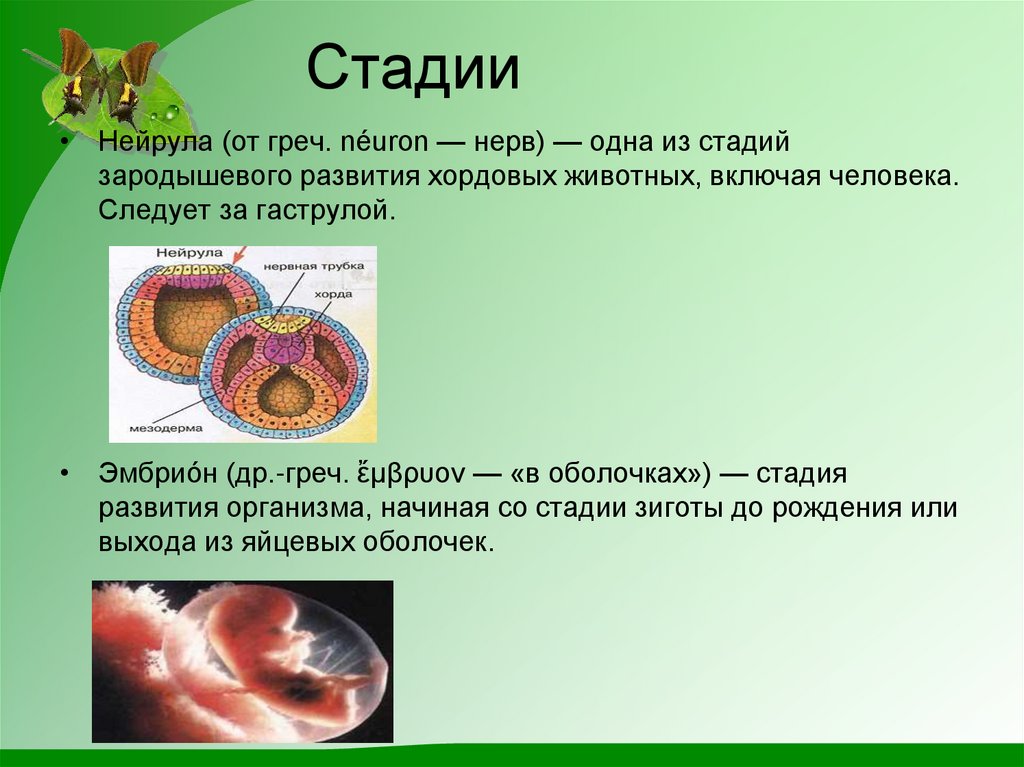
Рис. 20. Аномалии развития костей нижней конечности.
Аномалии развития лопатки:
- Глубокая вырезка, иногда отверстие
- Несращение акромиона (синхондроз)
Аномалии развития ключицы:
- Варьируют изгибы
- Нет конусовидного бугорка и трапециевидной линии.
Варианты и аномалии развития плечевой кости
- Processus supracondylaris – над медиальным надмыщелком.
Аномалии развития костей предплечья:
- Локтевой отросток не срастается с телом локтевой кости
- Отсутствие лучевой кости
Аномалии развития костей кисти:
- Добавочные кости запястья, например, os centrale
- Добавочный палец (полидактилия)
Варианты и аномалии развития тазовой кости:
- Отверстие в центре подвздошной ямки
- Удлинение подвздошных остей
Варианты и аномалии развития бедренной кости:
- Увеличение ягодичной бугристости – третий вертел, trochanter tertius
Варианты и аномалии развития костей голени:
- Уплощенное (не трехгранное) тело большеберцовой кости
Варианты и аномалии развития костей стопы
- Добавочные кости предплюсны
- Добавочные пальцы
Рис.
 21. Развитие костей черепа.
21. Развитие костей черепа.Рис. 22. Источники развития костей лицевого черепа.
Рис. 23. Развитие костей черепа.
Рис. 24. Развитие костей черепа после рождения.
Кости свода и лицевого черепа по развитию относятся к первичным костям, окостеневающим на основе соединительной ткани эндесмальным способом окостенения.
Кости лицевого черепа развиваются на основе жаберных дуг (первой и второй висцеральной дуги).
Из первой висцеральной дуги развиваются следующие кости: верхняя, нижняя челюсти, частично скуловая и небные кости, медиальная пластинка крыловидного отростка клиновидной кости; молоточек, наковальня – слуховые косточки; костное небо и его швы, нижняя часть глазницы.
Из второй висцеральной дуги развиваются: стремечко, шиловидный отросток височной кости, малые рога подъязычной.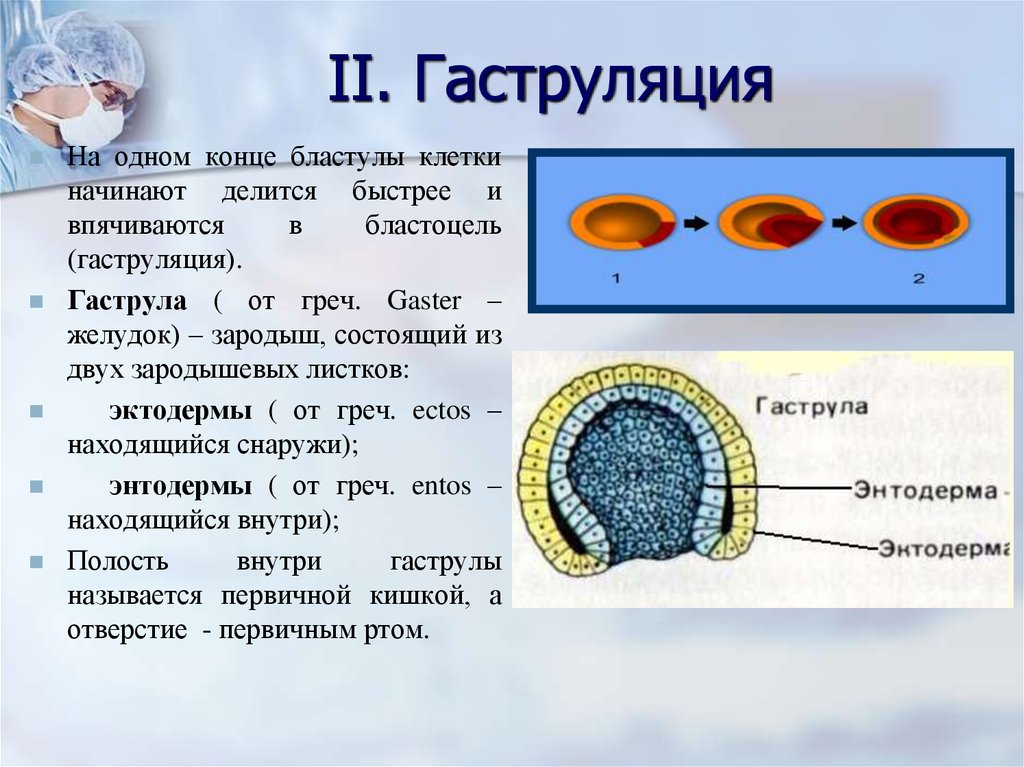
Кости основания черепа проходят три стадии развития: соединительнотканную, хрящевую и костную. Т.е. они являются вторичными. Они окостеневают энхондрально.
Рис. 25. Вставочные кости швов черепа (слева), деформации черепа (справа).
Рис. 26. Аэнцефалия.
Рис. 27. Аномалии развития твердого неба.
Известны следующие аномалии развития черепа
- Непостоянные (вставочные, вормиевы) кости швов
- Кости родничков
- Непостоянные швы
- Большие теменные отверстия
- Тонкая теменная кость
- Дырчатый череп
- Краниосхизис – головной мозг и череп открыты с дорсальной стороны
- Краниостеноз – преждевременный синостоз отдельных или всех швов, ведет к деформациям черепа:
- Башенный
- Ладьевидный
- Клиновидный
- Скошенный
- Микроцефалия
- Макроцефалия
- Гидроцефалия
- Аэнцефалия (отсутствует свод черепа, сочетается с недоразвитием головного мозга).
- Аномалии лицевого черепа:
- Заячья губа
- Волчья пасть
- Колобома
- Др.
Рис. 28. Скелет костистой рыбы.
Рис. 29. Двоякодышашая рыба.
Рис. 30. Скелет лягушки.
Рассмотрим филогенез соединения костей хордовых. У беспозвоночных твердого скелета нет, весь скелет представлен соединтельнотканным тяжем — хордой.
У рыб, обитающих в водной среде, многочисленные кости скелета (рис. 28) соединяются при помощи непрерывных соединений: соеденительнотканных и хрящевых.
Важным биомеханическим фактором, повлиявшим на филогенез соединений костей, является выход животных на сушу. Кратковременное пребываниена твердой поверхности (в том числе перемещение с места на место), которое наблюдается, напрмер, у двоякодышащих рыб, приводит к появлению гемиартрозов между костями конечностей (рис.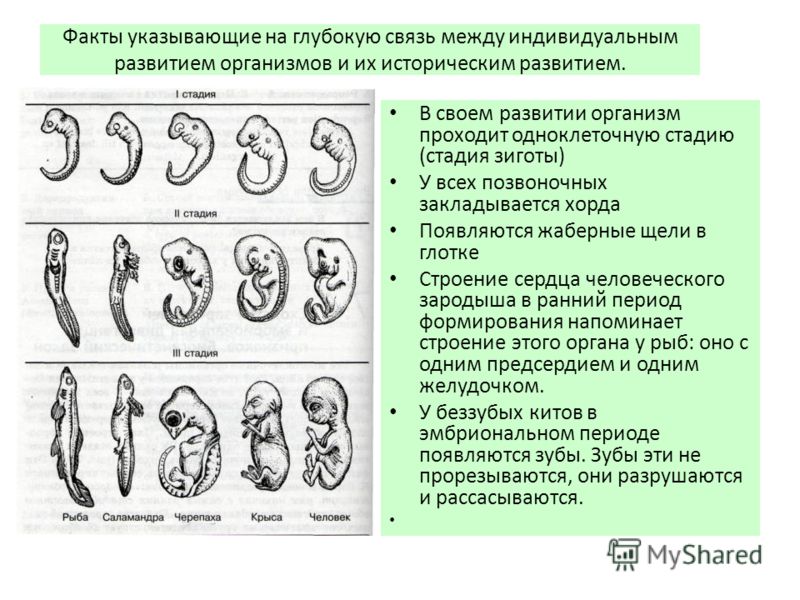 29). Большинство исследователей считают такую форму пререходной от снартроза к диартрозу.
29). Большинство исследователей считают такую форму пререходной от снартроза к диартрозу.
Окончательный выход животных на сушу формирует два направления морфогенеза мягкого остова. Во-первых, формируются суставы со всеми обязательными и вспомогательными элементами и высокой степенью подвижности. Во-вторых, в местах контакта костей с увеличившейся нагрузкой (из-за возросшего действия силы тяжести) формируются синостозы (кости срастаются между собой). Признаки обоих изменений строения соединений костей уже выявляются у земноводных (рис. 30).
Рис. 31. Варианты дисплазии тазобедренного сустава.
В онтегенезе соединения костей наблюдаются сходные с филогенезом тенденции. Первоначально все соединения образованые скоплением мезенхимальных клеток (эмбриональная соединительная ткань).
В конце первой половины пренатального онтогенеза (16-18-ая недели внутриутробного развития) между костями, которые смещаются (движутся) друг относительно друга, формируются суставы.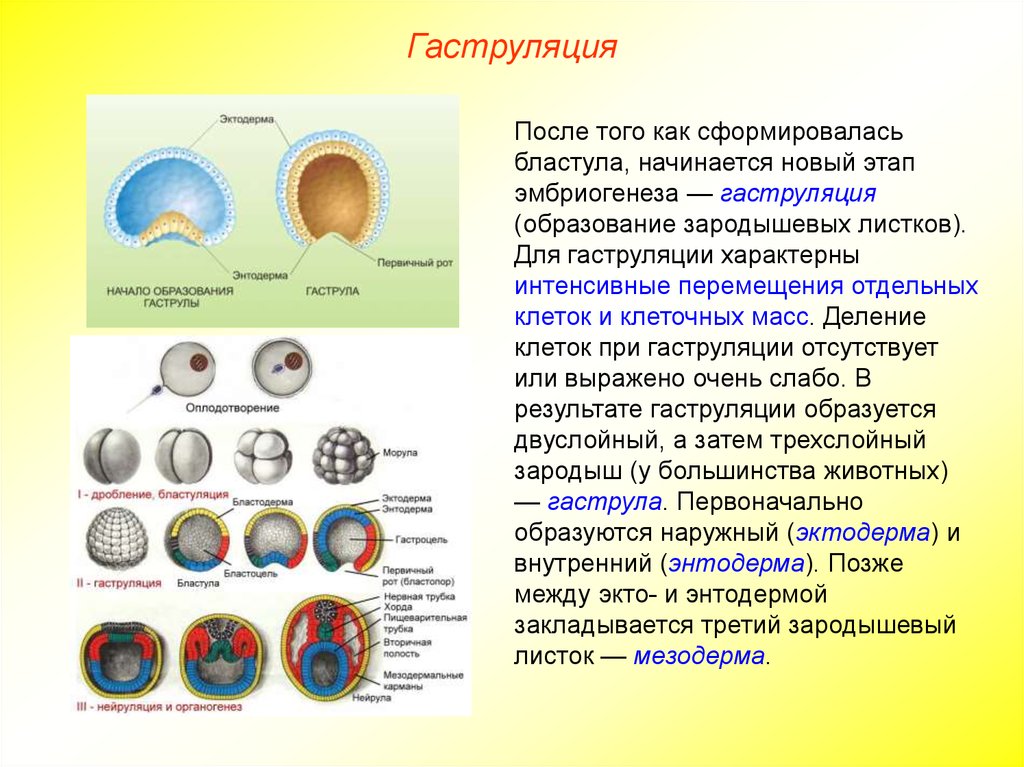 Важным биомеханическим фактором их морфогенеза являются силы мышц, двигающих кости. Между зачатками костей, которые соединяются без смещание, формируются непрерывные соединения.
Важным биомеханическим фактором их морфогенеза являются силы мышц, двигающих кости. Между зачатками костей, которые соединяются без смещание, формируются непрерывные соединения.
У новорожденных имеется закладка всех элементов суставов на нижней конечности. Однако большинство из них достигают функциональной зрелости к юношескому возрасту.
Основной аномалией развития соединения костей является дисплазия суставов. Для данные аномалии характерно изменение формы одной из суставных поверхностей, сопровождающееся изменениями строения расположенных рядом элементов сустава (рис. 31).
Автор лекции доцент Стрижков А.Е.
© Первый МГМУ им. И.М. Сеченова, 2017.
Эмбриогенез | Кинезиолог
Краткое описание:
Библиографическая ссылка для цитирования: Сазонов В.Ф. Эмбриогенез [Электронный ресурс] // Кинезиолог, 2009-2022: [сайт]. Дата обновления: 24.11.2022. URL: https://kineziolog.su/content/embriogenez (дата обращения: __.__.20___). _______________Ранние стадии эмбриогенеза.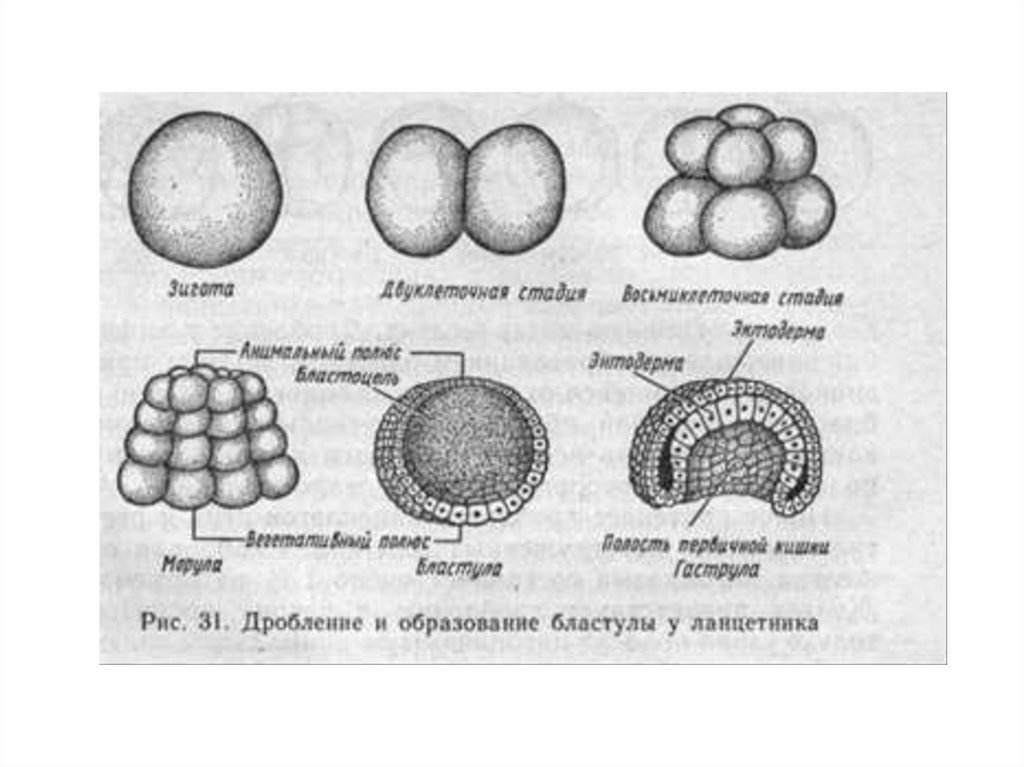
Определение понятий
Онтогенез — это процесс развития организма от момента зачатия (образования зиготы) до смерти.
Эмбриогенез — это период развития организма от образования зиготы до рождения (выхода из яйцевых оболочек).
Однако не надо упускать того, что есть ещё и преэмбриональный (синоним: проэмбриональный) период — это процесс от начала формирования половых клеток (мейоза) до слияния мужской и женской половой клетки (оплодотворения).
Интересно, как следует расценивать статус половой клетки по отношению к индивидуальному развитию человека? Интуиция подсказывает, что человеческую половую клетку ещё нельзя считать человеком. Однако, эта клетка имеет полный набор чисто человеческих генов, достаточный для построения полноценного человеческого организма. Только этот набор одинарный (гаплоидный), а не двойной (диплоидный), как положено для нормального человеческого организма.
Что же это тогда такое: человеческая половая клетка?
Человеческая половая клетка — это человеческое существо на ранней гаплоидной стадии проэмбрионального развития человека.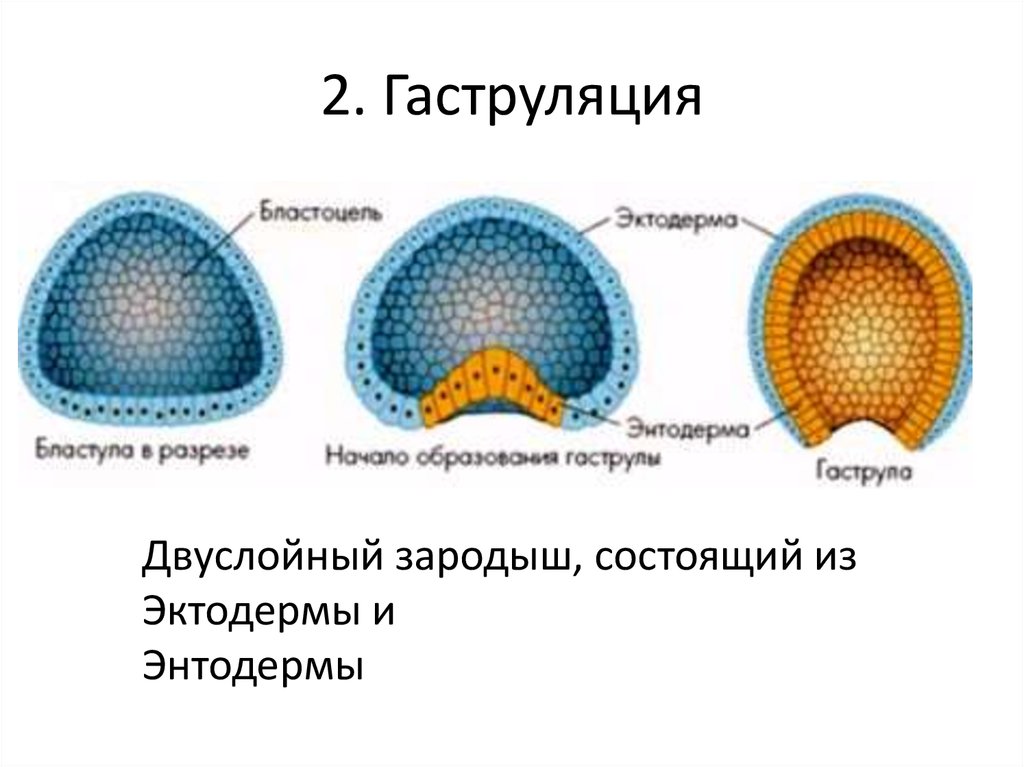 © 2022 Сазонов В.Ф. © 2022 kineziolog.su
© 2022 Сазонов В.Ф. © 2022 kineziolog.su
Это ещё не человек, однако это уже человеческое существо в особом обличии, которое живёт, действует по определённой генетической программе и умирает, если не встретится со своей «второй половинкой», то есть половой клеткой «другого пола». Яйцеклетка и сперматозоид должны встретиться и слиться, чтобы продолжить своё долгое совместное существование уже в виде диплоидного организма.
Онтогенез делится на пренатальное развитие (дородовое — от зачатия до рождения) и постнатальное (послеродовое). Эти же периоды называются эмбриональным и постэмбриональным развитием.
Оплодотворением называют слияние мужской и женской половых клеток, в результате которого возникает зигота (оплодотворённая яйцеклетка) с диплоидным (двойным) набором хромосом.
Оплодотворение
Оплодотворение чаще всего совершается в расширении женского яйцевода (в маточных трубах). Сперматозоиды, излившиеся в составе спермы во влагалище, благодаря своей исключительной подвижности и активности продвигаются в полость матки, проходят через неё до яйцеводов и в одном из них встречаются со зрелой яйцеклеткой.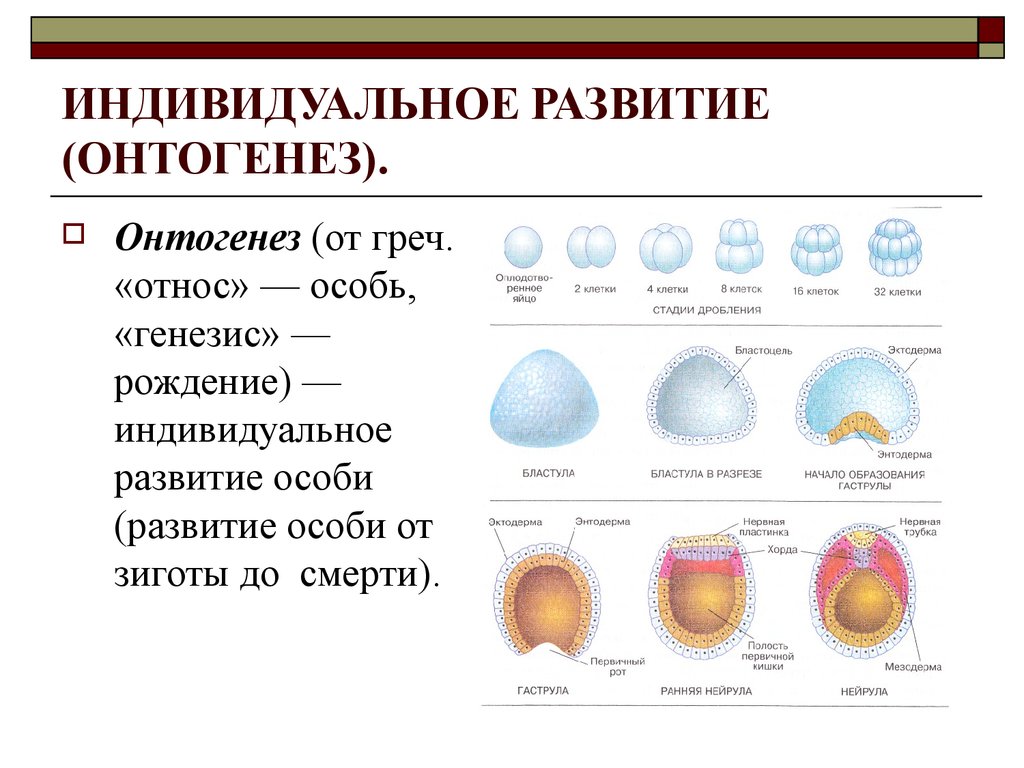 Здесь сперматозоид внедряется в яйцеклетку и оплодотворяет её. Сперматозоид вносит в яйцеклетку наследственные свойства, характерные для отцовского организма, содержащиеся в упакованном виде в хромосомах мужской половой клетки. Образно можно сказать, что зигота и развившийся из неё организм — это результат встречи яйцеклетки, наиболее удачливой среди других яйцеклеток, с самым успешным «сперматозоидом-чемпионом», опередившим десятки миллионов своих конкурентов.
Здесь сперматозоид внедряется в яйцеклетку и оплодотворяет её. Сперматозоид вносит в яйцеклетку наследственные свойства, характерные для отцовского организма, содержащиеся в упакованном виде в хромосомах мужской половой клетки. Образно можно сказать, что зигота и развившийся из неё организм — это результат встречи яйцеклетки, наиболее удачливой среди других яйцеклеток, с самым успешным «сперматозоидом-чемпионом», опередившим десятки миллионов своих конкурентов.
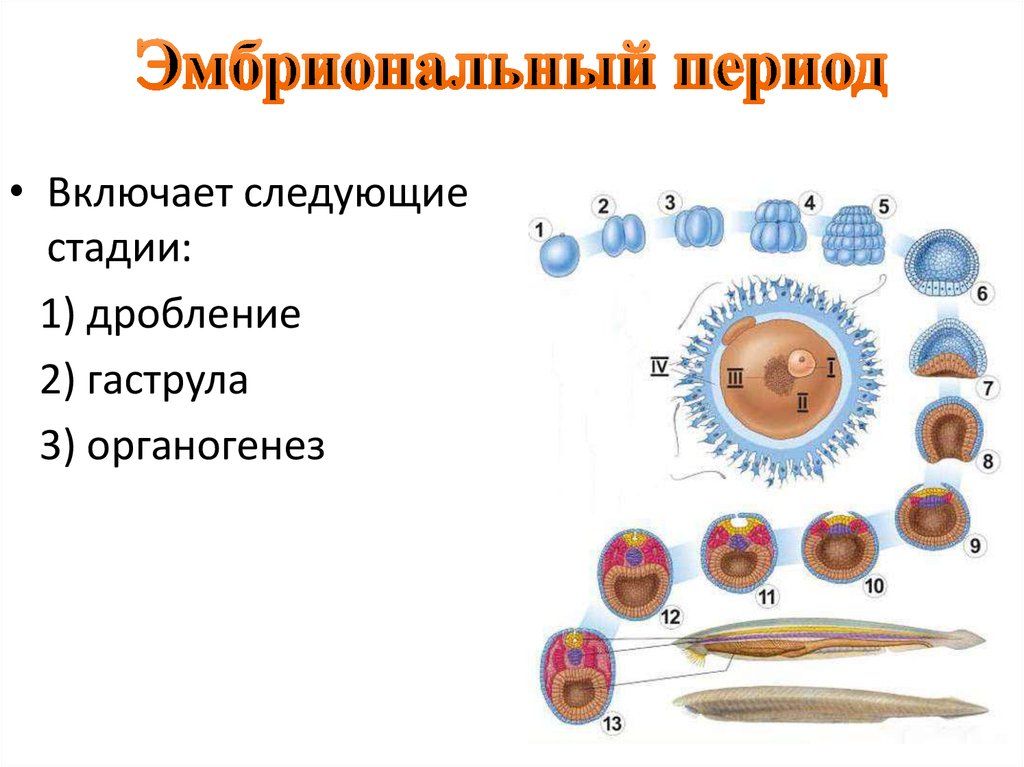
Дробление
Дробление — это процесс клеточного деления, в который вступает зигота. Размеры образующихся клеток при этом не увеличиваются, т.к. они не успевают расти, а только делятся.
После того как оплодотворённое яйцо начинает делиться, его называют эмбрионом. Происходит активация зиготы; начинается её дробление. Дробление идёт медленно. На 4-е сутки зародыш состоит из 8-12 бластомеров (бластомеры — это клетки, образующиеся в результате дробления, они всё более мелкие после очередного деления).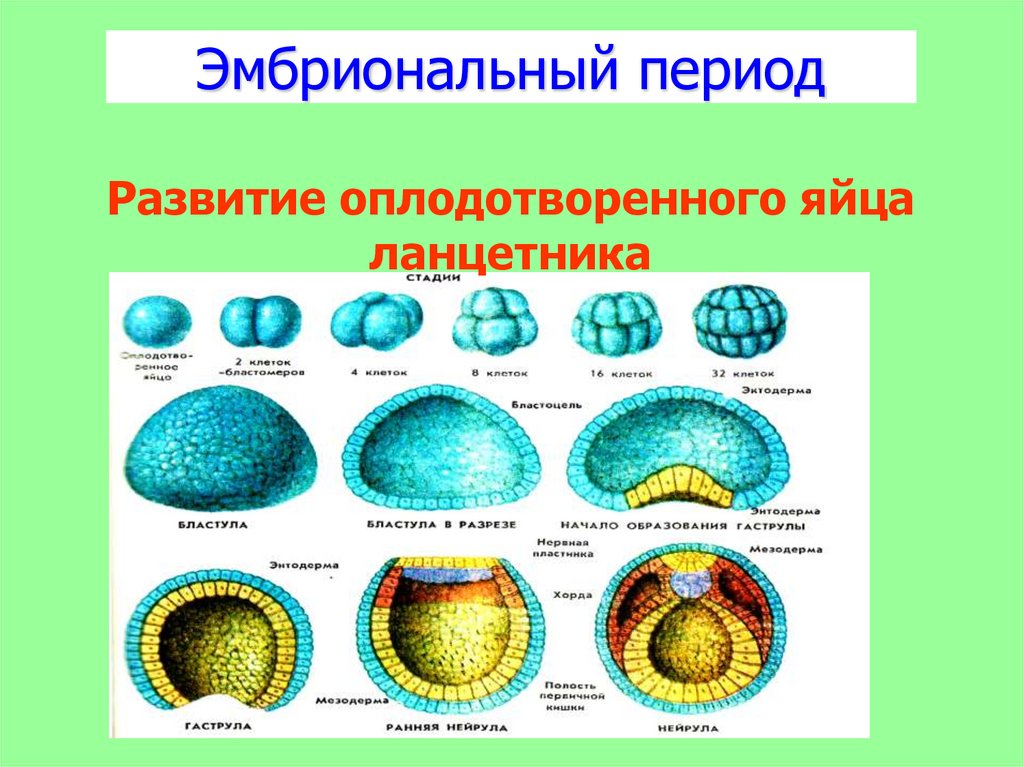
Рисунок: Начальные стадии эмбриогенеза млекопитающих животных
I – стадия 2-х бластомеров; II – стадия 4-х бластомеров; III – морула; IV–V – образование трофобласта; VI – бластоциста и первая фаза гаструляции:
1 – темные бластомеры; 2 – светлые бластомеры; 3 – трофобласт;
4 – эмбриобласт; 5 – эктодерма; 6 – энтодерма.
Рис. __. Эмбрион морской звезды Echinaster brasiliensis. Микроскопия в темном поле, увеличение в 60 раз. Вверху слева стадия двух бластомеров, справа — четырех. Затем, в процессе дробления образуется морула (внизу слева), а после гаструляции — личинка (внизу справа). Автор: Alvaro Esteves Migotto, Universidade de Sâo Paulo.
Рис. Фотографии ранних стадий (дробление) поясняются схемами.
Морула
Морула («тутовая ягода», она же «ягода шелковицы») — это группа бластомеров, образовавшихся в результате дробления зиготы.
Рис. Морула человека, срок развития зародыша — 80 часов.
Фото: Ягоды шелковицы (тутовника): Morus nigra и Morus alba, давшие название данной стадии эмбриогенеза
Рис. Дробление: от двух бластомеров до бластоцисты. Источник изображения: http://higheredbcs.wiley.com/legacy/college/tortora/0470565101/hearthis_…
Бластула
Бластула (пузырёк) — это однослойный зародыш. Клетки расположены в нём в один слой.
Бластула образуется из морулы за счёт того, что в ней появляется полость. Полость называется первичная полость тела. Она содержит жидкость. В дальнейшем полость заполняется внутренними органами и превращается в брюшную и грудную полости.
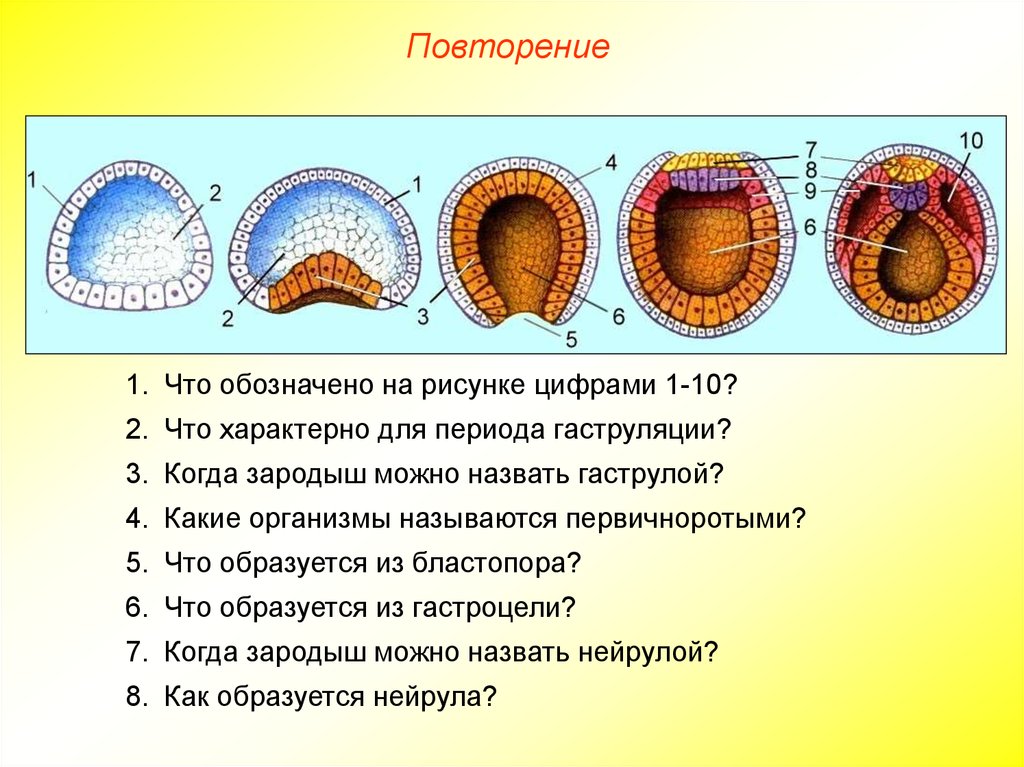
Гаструла
Гаструла — это двухслойный (или трёхслойный) зародыш. Клетки в этом «зародышевом пузырьке» образуют стенки в два или в три слоя.
Гаструляция (образование двухслойного и далее трёхслойного зародыша) — это очередной этап эмбрионального развития. Внешний слой гаструлы называется эктодерма. Он в дальнейшем формирует кожные покровы тела и нервную систему. Очень важно запомнить, что нервная система происходит из эктодермы (наружного зародышевого листка, первого), поэтому она ближе по своим особенностям к коже, чем к таким внутренним органам, как желудок и кишечник. Внутренний слой называется энтодерма (именно энто-, а не эндо-, обратите на это внимание). Энтодерма даёт начало пищеварительной системе и дыхательной. Тоже важно запомнить, что дыхательная и пищеварительная система связаны общим происхождением. Жаберные щели у рыб — это отверстия в кишке, а лёгкие у сухопутных позвоночных животных — это выросты кишки, которые у рыб образуют плавательный пузырь. Между эктодермой и энтодермой несколько позже вклинивается слой мезодермы. Это по времени происхождения третий зародышевый слой, поэтому он называется третьим зародышевым листком, хотя по своему расположению он кажется вроде бы вторым.
Внешний слой гаструлы называется эктодерма. Он в дальнейшем формирует кожные покровы тела и нервную систему. Очень важно запомнить, что нервная система происходит из эктодермы (наружного зародышевого листка, первого), поэтому она ближе по своим особенностям к коже, чем к таким внутренним органам, как желудок и кишечник. Внутренний слой называется энтодерма (именно энто-, а не эндо-, обратите на это внимание). Энтодерма даёт начало пищеварительной системе и дыхательной. Тоже важно запомнить, что дыхательная и пищеварительная система связаны общим происхождением. Жаберные щели у рыб — это отверстия в кишке, а лёгкие у сухопутных позвоночных животных — это выросты кишки, которые у рыб образуют плавательный пузырь. Между эктодермой и энтодермой несколько позже вклинивается слой мезодермы. Это по времени происхождения третий зародышевый слой, поэтому он называется третьим зародышевым листком, хотя по своему расположению он кажется вроде бы вторым.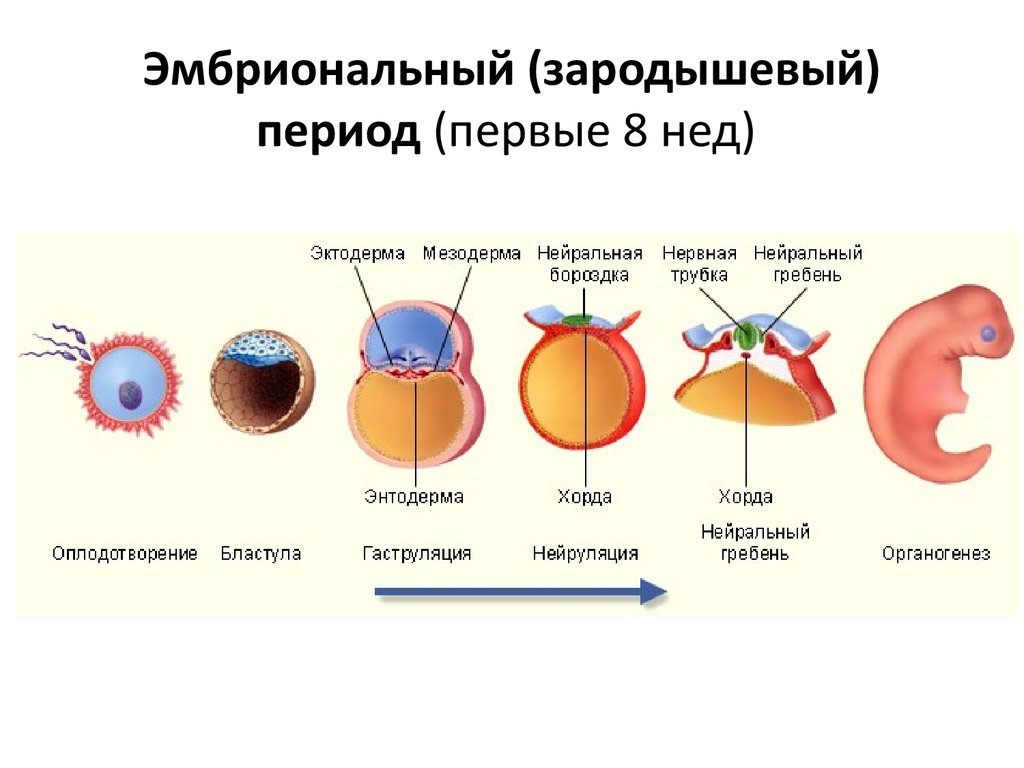
Нейрула
Нейрула — это зародыш на стадии формирования нервной трубки.
Можно назвать нейрулу «поздней гаструлой», но если считать гаструлой только двухслойный зародыш, то получается, что нейрула — это уже не гаструла, а отдельный этап зародышевого развития — трёхслойный зародыш. В процессе нейруляции, т.е. преобразования гаструлы в нейрулу, пузырёк гаструлы вытягивается, а сверху образуется желобок. Этот желобок из вдавленной эктодермы сворачивается в трубку — это нервная трубка. Под ним формируется тяж из выпячивания кишки — это хорда. Так что хорда происходит из энтодермы. Вокруг неё с течением времени уже из мезодермы будет образовываться сначала хрящевая, а потом и костная ткань — и получится позвоночник. Образно можно сказать, что в развитии «хорда не имеет будущего», она ничего не производит. Её «душит» мезодермальная костная ткань. Остаточки хорды можно найти между позвонками рыбы в виде студенистых дисков. Ниже хорды энтодерма вытягивается в кишечную трубку.
Ниже хорды энтодерма вытягивается в кишечную трубку.
Комплекс осевых органов — это нервная трубка, хорда и кишечная трубка.
Гисто- и органогенез
После нейруляции начинается следующий этап в развитии зародыша — гистогенез и органогенез, т.е. формирование тканей («гисто-» — это ткань) и органов. На этом этапе происходит разрастание третьего зародышевого слоя — мезодермы.
Следует обратить внимание на то, что с момента формирования органов и нервной системы, зародыш называют плодом, а период развития — плодным или фетальным.
Плод, развивающийся в матке, находится в особых оболочках, образующих как бы мешок, заполненный околоплодными водами. Эти воды дают возможность плоду свободно передвигаться в мешке, обеспечивают защиту плода от внешних повреждений и инфекций, а также способствуют нормальному течению родового акта.
Рис. __. Эмбриогенез ланцетника для общего представления об этом этапе развития организма.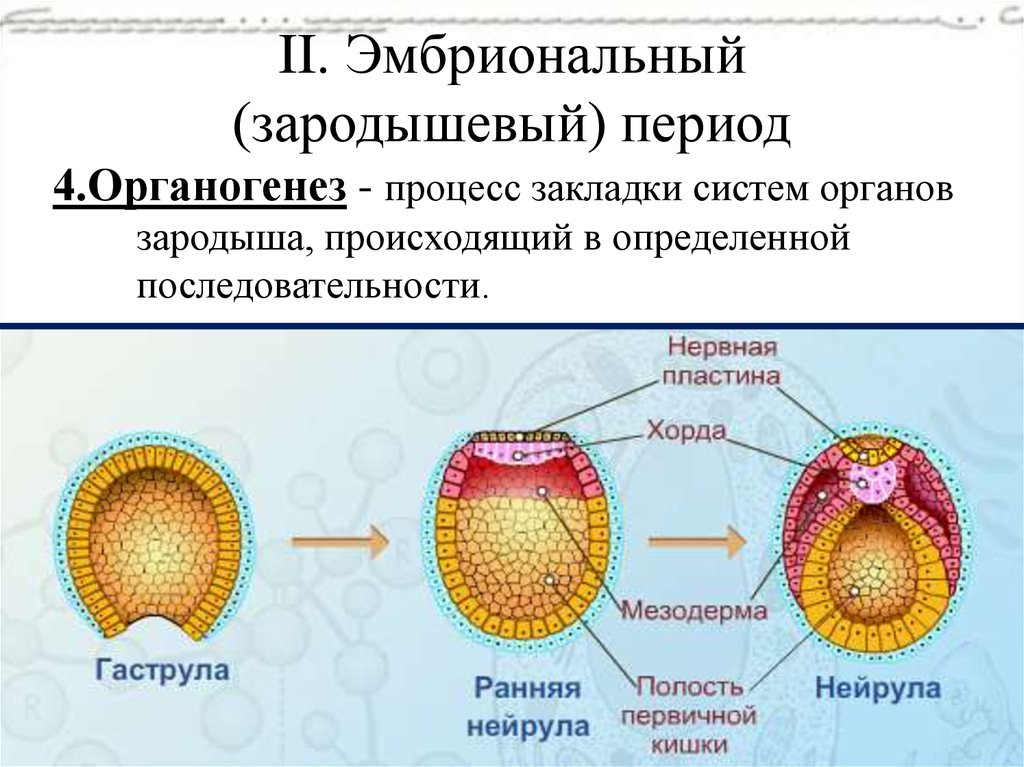 Источник изображения: http://www.studfiles.ru/preview/4333719/
Источник изображения: http://www.studfiles.ru/preview/4333719/
Особенности эмбрионального развития человека
Развитие человеческого эмбриона происходит внутриутробно, как и у других млекопитающих. Поэтому, в отличие от более примитивных животных типа ланцетника, эмбриону нужны дополнительные структуры, позволяющие ему питаться за счёт матери. Это делает процесс эмбрионального развития человека довольно сложным.
Рис. __. Нейруляция у человека: а — вид со спины; б — поперечные срезы. 1 — передний нейропор; 2 — задний ней-ропор; 3 — эктодерма; 4 — нервная пластинка; 5 — нервный желобок; 6 — мезодерма; 7 — хорда; 8 — энтодерма; 9 — нервная трубка; 10 — нервный гребень; 11 — головной мозг; 12 — спинной мозг; 13 — спинномозговой канал Источник изображения: http://vmede.org/sait/?id=Gistologiya_embriol_cit_afanasev_2012&menu=Gis…
Внезародышевые (=провизорные, временные) органы
Источниками развития для тканей внезародышевых органов являются троф-эктодерма и все три зародышевых листка. Общие свойства тканей внезародышевых органов: 1) их развитие является сокращенным и ускоренным; 2) соединительная ткань содержит мало клеточных форм, но много аморфного вещества, богатого гликозаминогликанами; 3) старение этих тканей происходит очень быстро и завершается к концу внутриутробного развития.
Общие свойства тканей внезародышевых органов: 1) их развитие является сокращенным и ускоренным; 2) соединительная ткань содержит мало клеточных форм, но много аморфного вещества, богатого гликозаминогликанами; 3) старение этих тканей происходит очень быстро и завершается к концу внутриутробного развития.
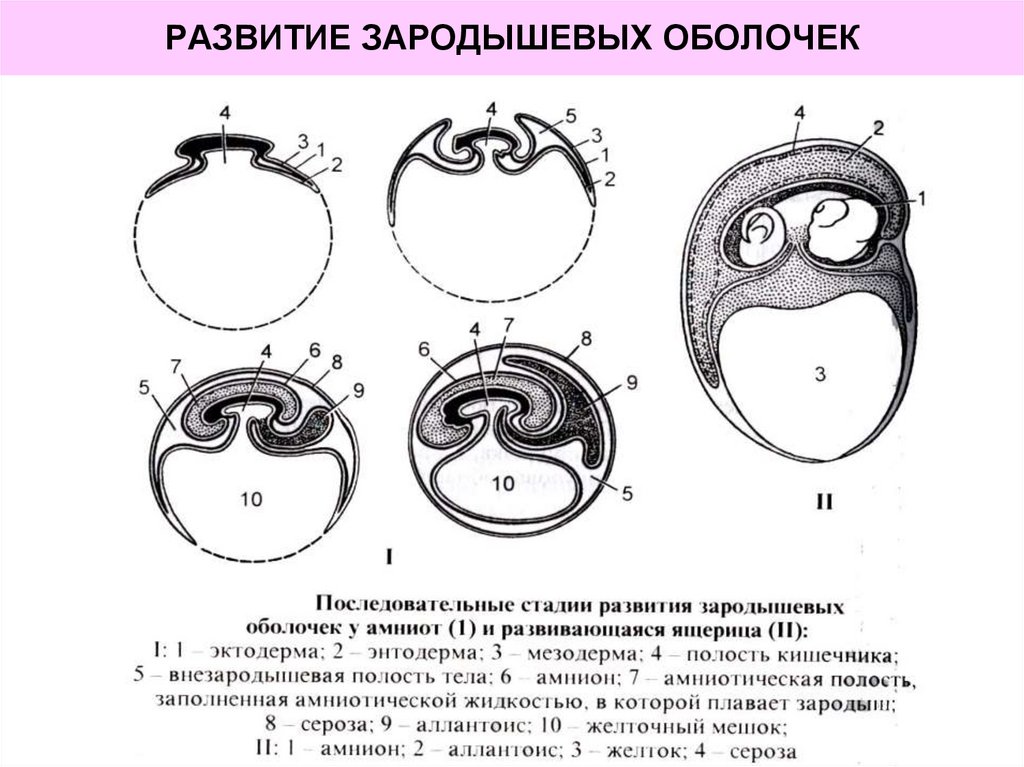
Внезародышевые органы развиваются в процессе эмбриогенеза вне тела зародыша. Они обеспечивают его рост и развитие. Те из них, которые окружают зародыш, называются также зародышевыми оболочками. К ним относятся амнион, желточный мешок, аллантоис, хорион, плацента.
Амнион — временный орган, обеспечивающий водную среду для развития зародыша. Амниотическая оболочка образует стенку резервуара, заполненного амниотической жидкостью, в которой плавает плод. Основная функция амниотической оболочки — выработка околоплодных вод, обеспечивающих среду для развития зародыша и предохраняющих его от механических повреждений.
Желточный мешок — наиболее древний в эволюции внезародышевый орган, возникший для хранения питательных веществ в виде желтка, необходимых для развития зародыша.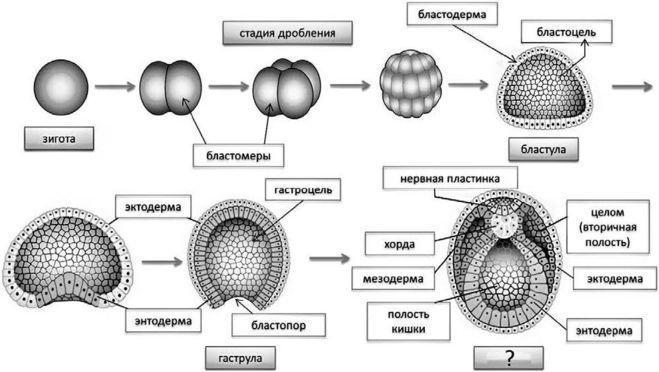 У человека это рудиментарное образование (желточный пузырек), так как с 3-й нед развития устанавливается связь плода с материнским организмом, и происходит переход на гематотрофное («кровяное») питание. В качестве кроветворного органа желточный мешок функционирует у зародыша человека до 7-8-й недели, а затем подвергается обратному развитию и остается в составе пупочного канатика в виде узкой трубочки, служащей проводником кровеносных сосудов к плаценте.
У человека это рудиментарное образование (желточный пузырек), так как с 3-й нед развития устанавливается связь плода с материнским организмом, и происходит переход на гематотрофное («кровяное») питание. В качестве кроветворного органа желточный мешок функционирует у зародыша человека до 7-8-й недели, а затем подвергается обратному развитию и остается в составе пупочного канатика в виде узкой трубочки, служащей проводником кровеносных сосудов к плаценте.
Аллантоис представляет собой небольшой пальцевидный отросток в хвостовом (каудальном) отделе зародыша, врастающий в амниотическую ножку. Он является производным желточного мешка и состоит из внезародышевой энтодермы и висцерального листка мезодермы. По нему к хориону растут сосуды, располагающиеся в пупочном канатике. Аллантоис — орган газообмена и выделения. По сосудам аллантоиса доставляется кислород, а в аллантоис выделяются продукты обмена веществ зародыша. На 2-м месяце эмбриогенеза аллантоис редуцируется и превращается в тяж клеток, который вместе с редуцированным желточным пузырьком входит в состав пупочного канатика.
Пупочный канатик, или пуповина, представляет собой упругий тяж, соединяющий зародыш (плод) с плацентой. Он покрыт амниотической оболочкой, окружающей слизистую соединительную ткань («вартонов студень») с кровеносными сосудами (две пупочные артерии и одна вена) и рудиментами желточного пузырька и аллантоиса.
Хорион, или ворсинчатая оболочка, появляется впервые у млекопитающих, развивается из трофобласта и внезародышевой мезодермы. Именно ворсинки трофобласта при имплантации бластоцисты выделяют протеолитические ферменты, разрушающие слизистую оболочку матки и обеспечивают имплантацию. Затем на 2-3-й неделе развития трофобласт превращается в хорион, или ворсинчатую оболочку, имеющую вторичные эпителиомезенхимальные ворсинки. В начале 3-й недели в ворсинки хориона врастают кровеносные капилляры и формируются третичные ворсинки. Это совпадает с началом гематотрофного питания зародыша. Дальнейшее развитие хориона связано с двумя процессами — разрушением слизистой оболочки матки вследствие протеолитической активности наружного (симпластического) слоя и развитием плаценты.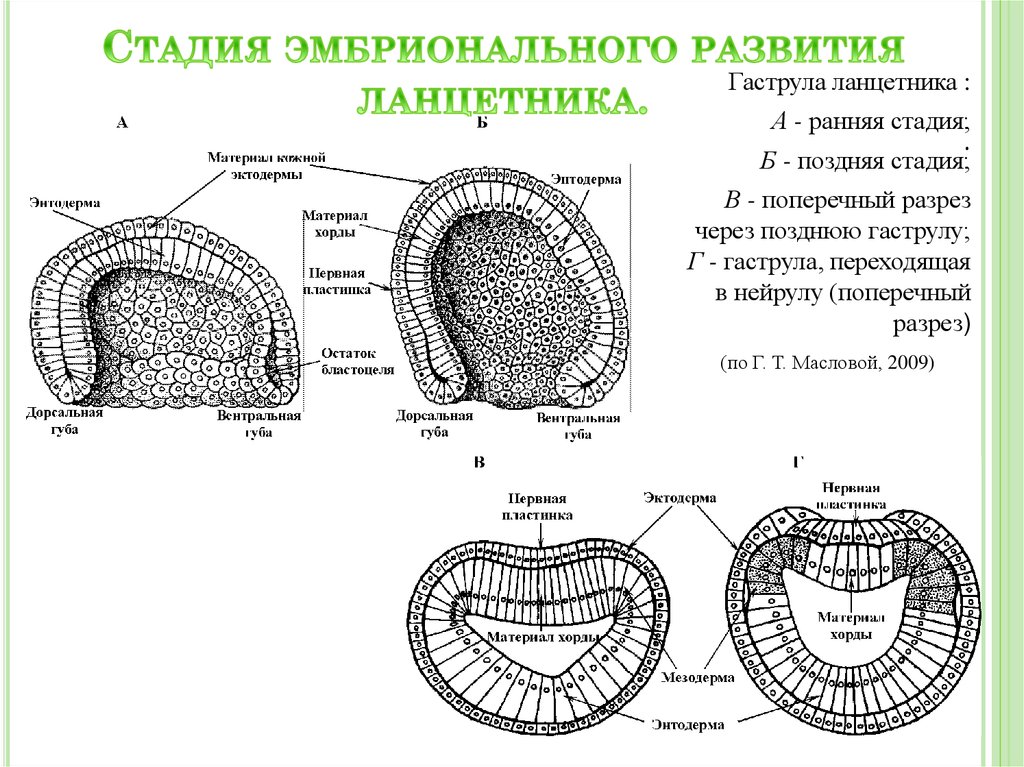
Плацента (детское место) человека относится к типу дискоидальных гемохориальных ворсинчатых плацент. Это важный временный орган с многообразными функциями, которые обеспечивают связь плода с материнским организмом. Вместе с тем плацента создает барьер между кровью матери и плода, за счёт которого их кровь не смешивается. Плацента состоит из двух частей: зародышевой, или плодной (pars fetalis), и материнской (pars materna). Плодная часть представлена ветвистым хорионом и приросшей к нему изнутри амниотической оболочкой, а материнская — видоизмененной слизистой оболочкой матки, отторгающейся при родах (decidua basalis). Развитие плаценты начинается на 3-й неделе, когда во вторичные ворсины начинают врастать сосуды и образовываться третичные ворсины, и заканчивается к концу 3-го месяца беременности. При развитии плаценты происходят разрушение слизистой оболочки матки, обусловленное протеолитической активностью хориона, и смена гистиотрофного питания на гематотрофное.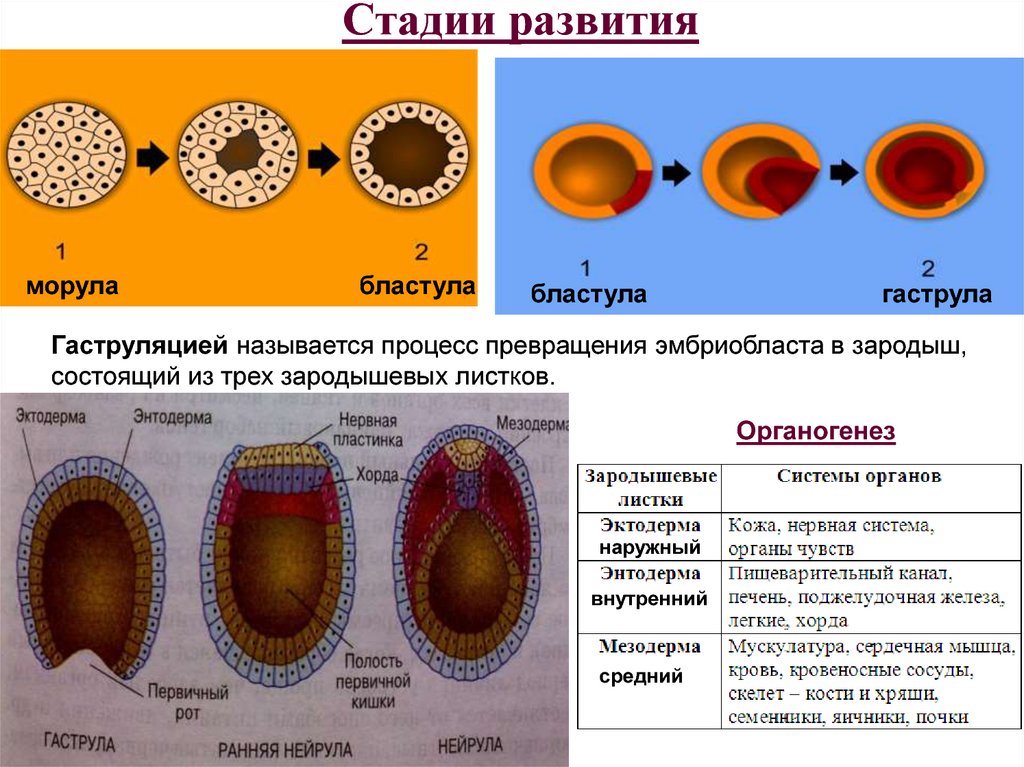 Это означает, что ворсины хориона омываются кровью матери, излившейся из разрушенных сосудов эндометрия в лакуны. Однако кровь матери и плода в нормальных условиях никогда не смешивается. Гематохориальный барьер, разделяющий оба кровотока, состоит из эндотелия сосудов плода, окружающей сосуды соединительной ткани, эпителия хориальных ворсин (цитотрофобласт и симпластотрофобласт), а кроме того, из фибриноида, который местами покрывает ворсины снаружи. Во второй половине беременности и особенно в её конце трофобласт сильно истончается и ворсины покрываются фибриноподобной оксифильной массой, являющейся продуктом свертывания плазмы и распада трофобласта («фибриноид Лангханса»).
Это означает, что ворсины хориона омываются кровью матери, излившейся из разрушенных сосудов эндометрия в лакуны. Однако кровь матери и плода в нормальных условиях никогда не смешивается. Гематохориальный барьер, разделяющий оба кровотока, состоит из эндотелия сосудов плода, окружающей сосуды соединительной ткани, эпителия хориальных ворсин (цитотрофобласт и симпластотрофобласт), а кроме того, из фибриноида, который местами покрывает ворсины снаружи. Во второй половине беременности и особенно в её конце трофобласт сильно истончается и ворсины покрываются фибриноподобной оксифильной массой, являющейся продуктом свертывания плазмы и распада трофобласта («фибриноид Лангханса»).
Структурно-функциональной единицей сформированной плаценты является котиледон, образованный стволовой («якорной») ворсиной и её вторичными и третичными (конечными) разветвлениями. Общее количество котиледонов в плаценте достигает 200.
Материнская часть плаценты представлена базальной пластинкой и соединительнотканными септами, отделяющими котиледоны друг от друга, а также лакунами (полостями), заполненными материнской кровью.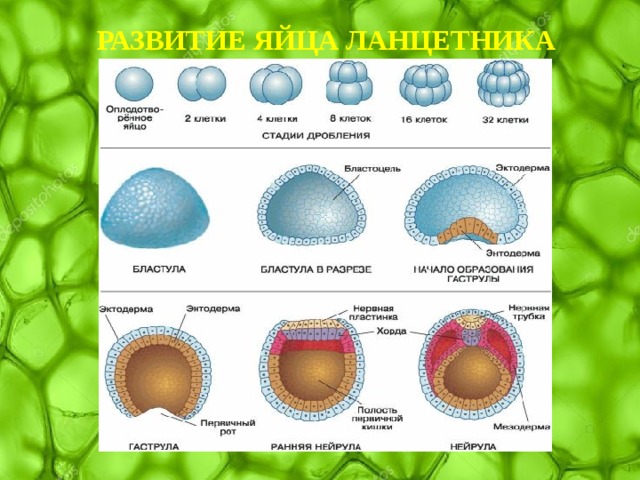 В местах контакта стволовых ворсин с отпадающей оболочкой встречаются также трофобластические клетки (периферический трофобласт).
В местах контакта стволовых ворсин с отпадающей оболочкой встречаются также трофобластические клетки (периферический трофобласт).
Основные функции плаценты: 1) дыхательная; 2) транспорт питательных веществ; воды; электролитов и иммуноглобулинов; 3) выделительная; 4) эндокринная; 5) участие в регуляции сокращения миометрия.
Рис. __. Развитие внезародышевых органов у зародыша человека (схема): 1 — амниотический пузырек; 1а — полость амниона; 2 — тело эмбриона; 3 — желточный мешок; 4 — внеэмбриональный целом; 5 — первичные ворсины хориона; 6 — вторичные ворсины хориона; 7 — стебелек аллантоиса; 8 — третичные ворсины хориона; 9 — аллан-тоис; 10 — пупочный канатик; 11 — гладкий хорион; 12 — котиледоны. Источник изображения: http://vmede.org/sait/?id=Gistologiya_embriol_cit_afanasev_2012&menu=Gis…
Источниками развития тканей внезародышевых органов являются троф-эктодерма и все три зародышевых листка.
Видеолекция: Ранний эмбриогенез
 youtube.com/embed/2ZC2HyEATh5″>
youtube.com/embed/2ZC2HyEATh5″> Видеолекция: Гаструляция
Видео: Имплантация и развитие зародыша
Видеолекция на англ.языке (English): Ранний эмбриогенез
Видео: Анимация гаструляции
Видео: Анимация нейруляции
Основные особенности ранних стадий развития зародыша человека
1) асинхронный тип полного дробления и образование «светлых» и «тёмных» бластомеров;
2) раннее обособление и формирование внезародышевых органов;
3) раннее образование амниотического пузырька и отсутствие амниотических складок;
4) наличие в стадии гаструляции двух механизмов — деламинации и иммиграции, в течение которых происходит также развитие провизорных органов;
5) интерстициальный тип имплантации;
6) сильное развитие амниона, хориона, плаценты и слабое развитие желточного мешка и аллантоиса.
Характеристики хордовых | Биология для специальности II
Результаты обучения
- Описать отличительные характеристики хордовых
Позвоночные относятся к царству Animalia и типу Chordata (рис. 1). Позвоночные демонстрируют два основных нововведения в своей эволюции от хордовых беспозвоночных. Эти нововведения могут быть связаны с дупликациями всего генома, что привело к четырехкратной дупликации основного генома хордовых, включая Hox локусы генов, которые регулируют размещение структур по трем осям тела. Одним из первых крупных шагов было появление четвероногих в форме земноводных. Вторым шагом была эволюция амниотического яйца, которая, подобно эволюции пыльцы и семян растений, освободила наземных животных от их зависимости от воды для оплодотворения и эмбрионального развития. У амниот модификации ороговевших эпидермальных структур привели к появлению чешуи, когтей, волос и перьев. Чешуя рептилий защищала их кожу от потери воды, в то время как волосы и перья обеспечивали изоляцию, поддерживая эволюцию эндотермии, а также выполняли другие функции, такие как маскировка и привлечение партнеров в линиях позвоночных, которые привели к птицам и млекопитающим.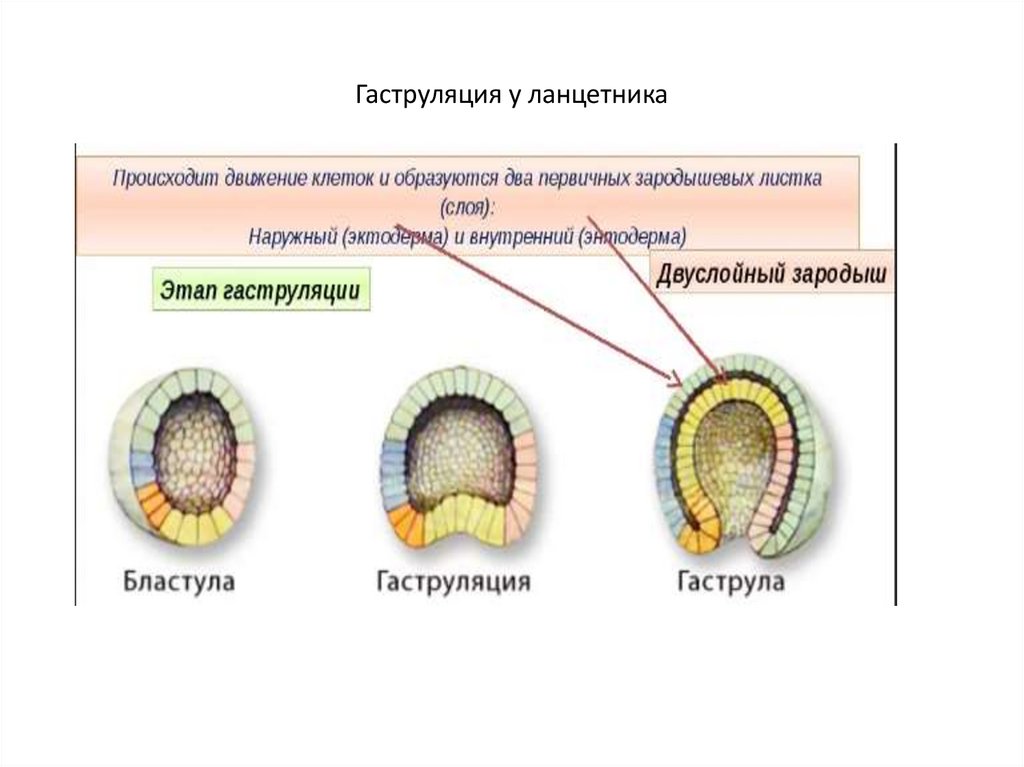
Рисунок 1. Все хордовые относятся к вторичноротым, имеющим хорду.
Характеристики хордовых
Животные типа Хордовые имеют пять общих ключевых характеристик, которые проявляются на определенной стадии их развития: хорда, дорсальный полый (трубчатый) нервный тяж, глоточные жаберные дуги или щели, постанальный хвост и эндостиль/щитовидная железа (рис. 2). В некоторых группах некоторые из этих ключевых характеристик присутствуют только во время эмбрионального развития.
Хордовые названы в честь хорды , которая представляет собой гибкую мезодермальную структуру в форме стержня, которая обнаруживается на эмбриональной стадии у всех хордовых и на взрослой стадии у некоторых видов хордовых. Он укреплен гликопротеинами, подобными хрящам, и покрыт коллагеновой оболочкой. Нотокорд расположен между пищеварительной трубкой и нервным тяжем и обеспечивает жесткую опору скелета, а также гибкое место для прикрепления осевых мышц. У некоторых хордовых хорда служит основной осевой опорой тела на протяжении всей жизни животного. Однако у позвоночных (черепных) хорда присутствует только во время эмбрионального развития, когда она индуцирует развитие нервной трубки и служит опорой для развивающегося эмбрионального тела. Однако хорда не обнаруживается на постэмбриональных стадиях позвоночных; в этот момент его заменил позвоночник (то есть позвоночник).
Однако у позвоночных (черепных) хорда присутствует только во время эмбрионального развития, когда она индуцирует развитие нервной трубки и служит опорой для развивающегося эмбрионального тела. Однако хорда не обнаруживается на постэмбриональных стадиях позвоночных; в этот момент его заменил позвоночник (то есть позвоночник).
Рисунок 2. У хордовых в какой-то момент развития появляются четыре общих признака: хорда, спинной полый нервный тяж, глоточные щели и постанальный хвост.
Практический вопрос
Какое из следующих утверждений об общих чертах хордовых верно?
- Спинной полый нервный тяж является частью центральной нервной системы хордовых.
- У позвоночных рыб глоточные щели становятся жабрами.
- Люди не хордовые, потому что у них нет хвоста.
- Позвоночные не имеют хорды на любом этапе своего развития; вместо этого у них есть позвоночник.
Показать ответ
Спинной полый нервный тяж происходит из эктодермы, которая в процессе развития скручивается в полую трубку. У хордовых он располагается дорсально от хорды. Напротив, нервная система у типов первичноротых животных характеризуется солидными нервными тяжами, которые расположены либо вентрально, либо латерально по отношению к кишечнику. У позвоночных нервная трубка развивается в головной и спинной мозг, которые вместе образуют центральную нервную систему (ЦНС). Периферическая нервная система (ПНС) относится к периферическим нервам (включая черепные нервы), лежащим за пределами головного и спинного мозга.
У хордовых он располагается дорсально от хорды. Напротив, нервная система у типов первичноротых животных характеризуется солидными нервными тяжами, которые расположены либо вентрально, либо латерально по отношению к кишечнику. У позвоночных нервная трубка развивается в головной и спинной мозг, которые вместе образуют центральную нервную систему (ЦНС). Периферическая нервная система (ПНС) относится к периферическим нервам (включая черепные нервы), лежащим за пределами головного и спинного мозга.
Щели глотки являются отверстиями в глотке (область сразу за ртом), которые выходят во внешнюю среду. У организмов, обитающих в водной среде, глоточные щели обеспечивают выход воды, попадающей в рот во время кормления. Некоторые хордовые беспозвоночные используют глоточные щели для фильтрации пищи из воды, попадающей в рот. Эндостиль представляет собой полоску реснитчатой ткани, продуцирующей слизь, на дне глотки. Попавшие в слизь частицы пищи перемещаются по эндостилю в сторону кишечника. Эндостиль также продуцирует вещества, сходные с гормонами щитовидной железы, и гомологичен щитовидной железе позвоночных. У позвоночных рыб глоточные щели преобразованы в жаберные опоры, а у челюстных рыб — в челюстные опоры. У четвероногих (наземных позвоночных) щели сильно видоизменены в компоненты уха, миндалин и вилочковой железы. У других позвоночных дуги глотки, происходящие из всех трех зародышевых листков, дают начало ротовой челюсти от первой дуги глотки, а вторая дуга становится подъязычной и опорой челюсти.
Эндостиль также продуцирует вещества, сходные с гормонами щитовидной железы, и гомологичен щитовидной железе позвоночных. У позвоночных рыб глоточные щели преобразованы в жаберные опоры, а у челюстных рыб — в челюстные опоры. У четвероногих (наземных позвоночных) щели сильно видоизменены в компоненты уха, миндалин и вилочковой железы. У других позвоночных дуги глотки, происходящие из всех трех зародышевых листков, дают начало ротовой челюсти от первой дуги глотки, а вторая дуга становится подъязычной и опорой челюсти.
Постанальный хвост является задним удлинением тела, выходящим за пределы ануса. Хвост содержит скелетные элементы и мышцы, которые обеспечивают передвижение водных видов, таких как рыбы. У некоторых наземных позвоночных хвост также помогает сохранять равновесие, ухаживать и подавать сигналы, когда приближается опасность. У людей и других человекообразных обезьян постанальный хвост уменьшен до рудиментарного копчика («хвостика»), который помогает сохранять равновесие во время сидения.
Посмотрите это видео, в котором обсуждается эволюция хордовых и пять общих для них характеристик.
Попробуйте
Внесите свой вклад!
У вас есть идеи по улучшению этого контента? Мы будем признательны за ваш вклад.
Улучшить эту страницуПодробнее
Хордовые | Биология II
Цели обучения
К концу этого раздела вы должны решить следующие задачи:
- Описать отличительные признаки хордовых
- Определите производный признак черепных, который отличает их от других хордовых
- Опишите развитие хорды у позвоночных
Позвоночные относятся к царству Animalia и типу Chordata (рис. 1). Напомним, что животных, обладающих билатеральной симметрией, можно разделить на две группы — первичноротых и вторичноротых — на основании особенностей их эмбрионального развития. Вторичноротые, чье название переводится как «второй рот», состоят из двух типов: хордовых и иглокожих.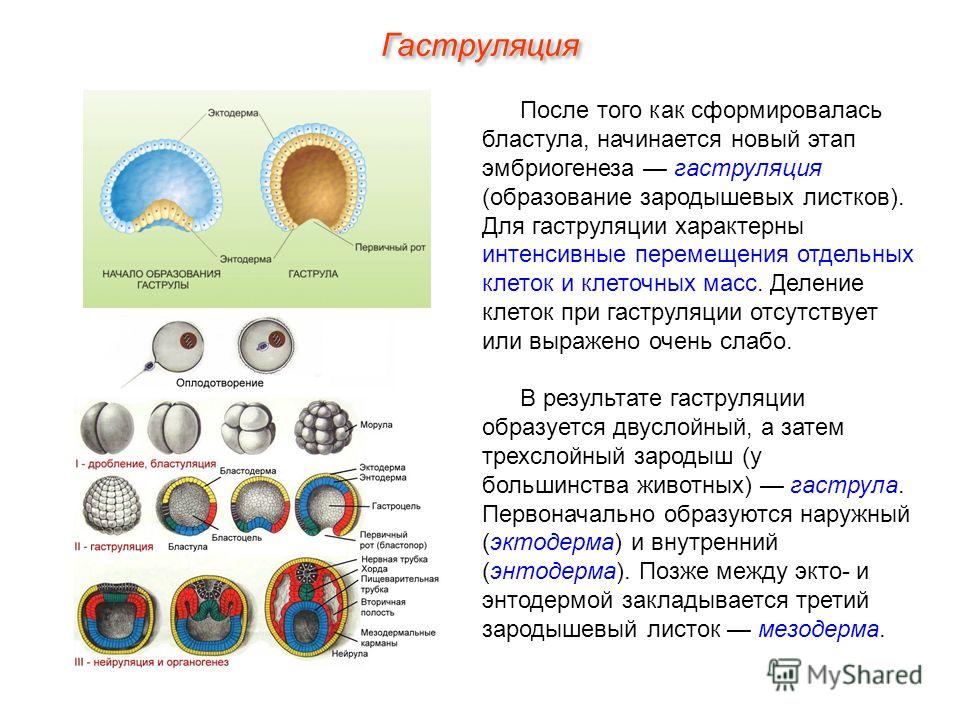 Иглокожие — это беспозвоночные морские животные, имеющие пятилучевую симметрию и колючее покрытие тела, группа, в которую входят морские звезды, морские ежи и морские огурцы. Наиболее заметными и знакомыми представителями хордовых являются позвоночные, но этот тип также включает две группы беспозвоночных хордовых.
Иглокожие — это беспозвоночные морские животные, имеющие пятилучевую симметрию и колючее покрытие тела, группа, в которую входят морские звезды, морские ежи и морские огурцы. Наиболее заметными и знакомыми представителями хордовых являются позвоночные, но этот тип также включает две группы беспозвоночных хордовых.
Рисунок 1. Все хордовые относятся к вторичноротым, имеющим хорду.
Характеристики хордовых
Животные типа Chordata имеют четыре общих ключевых признака, появляющихся на определенной стадии их развития: хорда, дорсальный полый нервный тяж, глоточные щели и постанальный хвост (рис. 2). В некоторых группах некоторые из них присутствуют только во время эмбрионального развития.
Хордовые названы в честь хорды , которая представляет собой гибкую палочковидную структуру, которая обнаруживается на эмбриональной стадии всех хордовых и на взрослой стадии некоторых видов хордовых. Он расположен между пищеварительной трубкой и нервным тяжем и обеспечивает опору скелета по всей длине тела.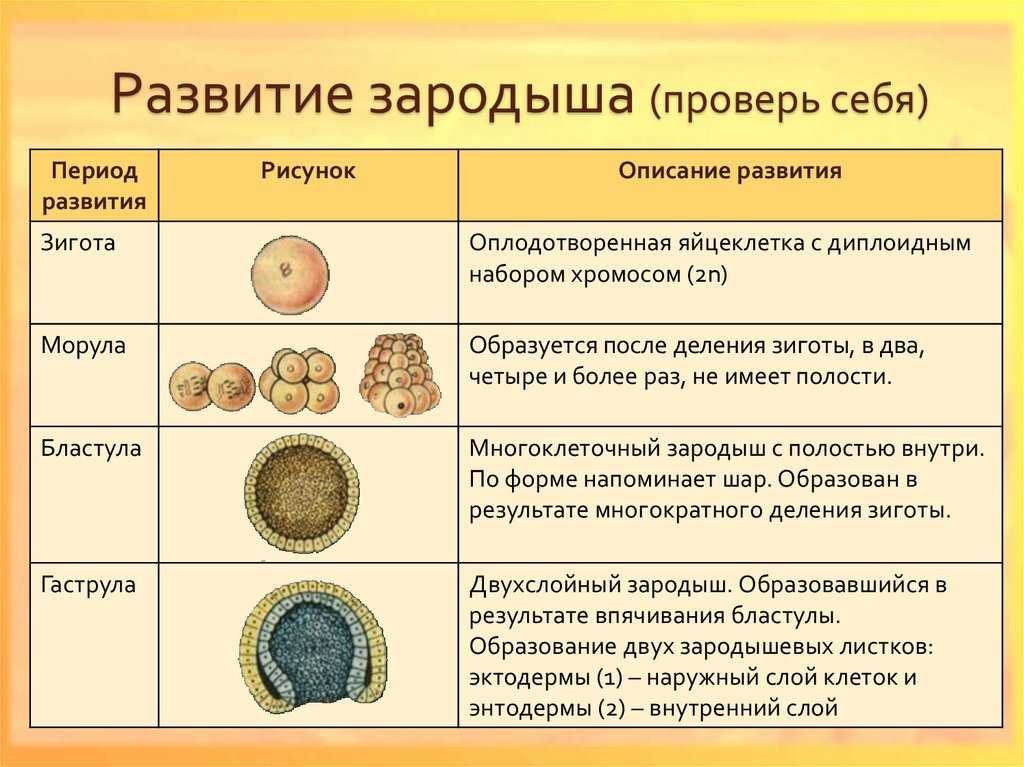 У некоторых хордовых хорда служит основной осевой опорой тела на протяжении всей жизни животного. У позвоночных хорда присутствует во время эмбрионального развития, в это время она индуцирует развитие нервной трубки и служит опорой для развивающегося эмбрионального тела. Однако хорда не обнаруживается на постнатальной стадии позвоночных; в этот момент его заменил позвоночник (то есть позвоночник).
У некоторых хордовых хорда служит основной осевой опорой тела на протяжении всей жизни животного. У позвоночных хорда присутствует во время эмбрионального развития, в это время она индуцирует развитие нервной трубки и служит опорой для развивающегося эмбрионального тела. Однако хорда не обнаруживается на постнатальной стадии позвоночных; в этот момент его заменил позвоночник (то есть позвоночник).
Art Connection
Рисунок 2. У хордовых в какой-то момент развития появляются четыре общих признака: хорда, спинной полый нервный тяж, глоточные щели и постанальный хвост.
Какое из следующих утверждений об общих чертах хордовых верно?
- Спинной полый нервный тяж является частью центральной нервной системы хордовых.
- У позвоночных рыб глоточные щели становятся жабрами.
- Люди не хордовые, потому что у них нет хвоста.
- Позвоночные не имеют хорды на любом этапе своего развития; вместо этого у них есть позвоночник.

Утверждение А верно.
Спинной полый нервный тяж происходит из эктодермы, которая во время развития скручивается в полую трубку. У хордовых она расположена дорсальнее хорды. Напротив, другие типы животных характеризуются солидными нервными тяжами, расположенными либо вентрально, либо латерально. Нервный тяж, обнаруженный у большинства эмбрионов хордовых, развивается в головной и спинной мозг, составляющие центральную нервную систему.
Глоточные щели — это отверстия в глотке (область сразу за ртом), которые выходят во внешнюю среду. У организмов, обитающих в водной среде, глоточные щели обеспечивают выход воды, попадающей в рот во время кормления. Некоторые хордовые беспозвоночные используют глоточные щели для фильтрации пищи из воды, попадающей в рот. У позвоночных рыб глоточные щели преобразованы в жаберные опоры, а у челюстных рыб — в челюстные опоры. У четвероногих щели преобразованы в компоненты уха и миндалин. Тетрапод буквально означает «четырехногий», что относится к филогенетической истории различных групп, которые развивались соответствующим образом, хотя некоторые из них сейчас обладают менее чем двумя парами ходячих придатков.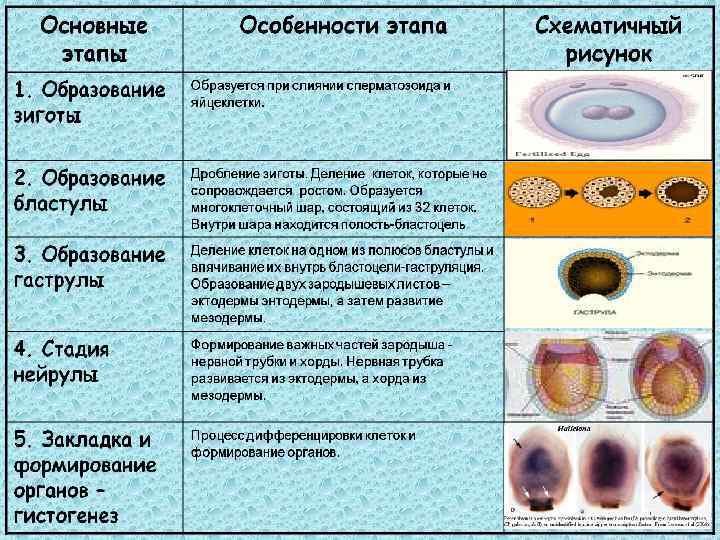 К четвероногим относятся амфибии, рептилии, птицы и млекопитающие.
К четвероногим относятся амфибии, рептилии, птицы и млекопитающие.
Постанальный хвост представляет собой заднее удлинение тела, выходящее за пределы ануса. Хвост содержит скелетные элементы и мышцы, которые обеспечивают передвижение водных видов, таких как рыбы. У некоторых наземных позвоночных хвост также помогает сохранять равновесие, ухаживать и подавать сигналы, когда приближается опасность. У человека постанальный хвост является рудиментарным, то есть уменьшенным в размерах и нефункциональным.
Ссылка на обучение
Посмотрите это видео, в котором обсуждается эволюция хордовых и пять общих для них характеристик.
Хордовые и эволюция позвоночных
Chordata также содержит две группы беспозвоночных: Urochordata и Cephalochordata. Члены этих групп также обладают четырьмя отличительными чертами хордовых в какой-то момент своего развития.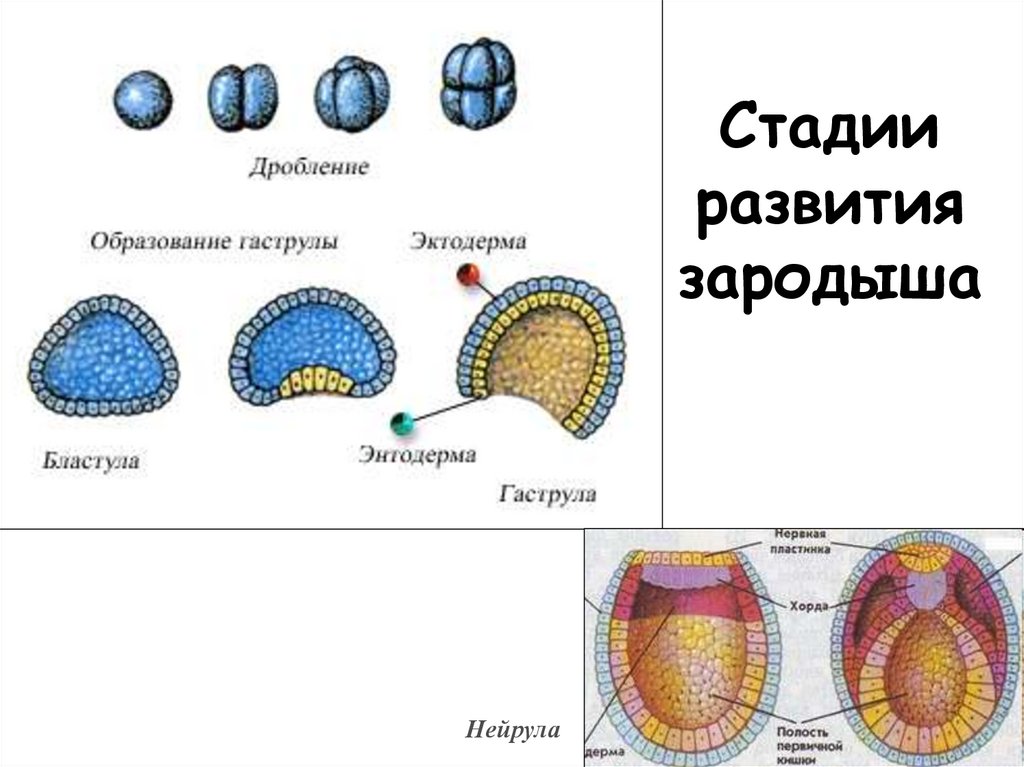
Урохордата
Члены Урохордата также известны как оболочники (рис. 3). Название оболочник происходит от целлюлозоподобного углеводного материала, называемого туникой, которая покрывает внешнее тело оболочников. Хотя взрослые оболочники классифицируются как хордовые, у них нет хорды, спинного полого нервного шнура или постанального хвоста, хотя у них есть глоточные щели. Однако личиночная форма обладает всеми четырьмя структурами. Большинство оболочников — гермафродиты. Личинки оболочников вылупляются из яиц внутри тела взрослых оболочников. После вылупления оболочниковая личинка плавает в течение нескольких дней, пока не найдет подходящую поверхность, на которой она может прикрепиться, обычно в темном или затененном месте. Затем он прикрепляется через голову к поверхности и претерпевает метаморфоз во взрослую форму, после чего хорда, нервный тяж и хвост исчезают.
Рисунок 3. (а) На этой фотографии показана колония оболочников Botrylloides violaceus.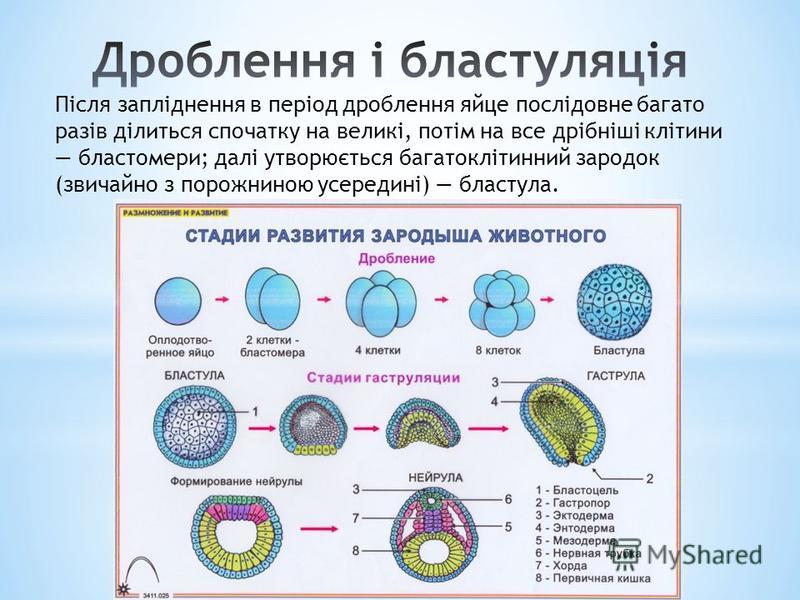 (б) Личиночная стадия оболочника обладает всеми признаками, характерными для хордовых: хордой, дорсальным полым нервным тяжем, глоточными щелями и постанальным хвостом. (c) На взрослой стадии хорда, нервный тяж и хвост исчезают. (кредит: модификация работы Дэнна Блэквуда, Геологическая служба США)
(б) Личиночная стадия оболочника обладает всеми признаками, характерными для хордовых: хордой, дорсальным полым нервным тяжем, глоточными щелями и постанальным хвостом. (c) На взрослой стадии хорда, нервный тяж и хвост исчезают. (кредит: модификация работы Дэнна Блэквуда, Геологическая служба США)
Большинство оболочников ведут сидячий образ жизни на дне океана и питаются суспензией. Основной пищей оболочников являются планктон и детрит. Морская вода поступает в тело оболочника через входной сифон. Взвешенный материал отфильтровывается из этой воды слизистой сеткой (глоточными щелями) и под действием ресничек попадает в кишечник. Анус впадает в отводящий сифон, который выбрасывает отходы и воду. Туникаты встречаются на мелководье в океанских водах по всему миру.
Головохордовые
Члены Cephalochordata обладают хордой, спинным полым нервным тяжем, глоточными щелями и постанальным хвостом на взрослой стадии (рис. 4). Хорда переходит в голову, что дало название подтипу.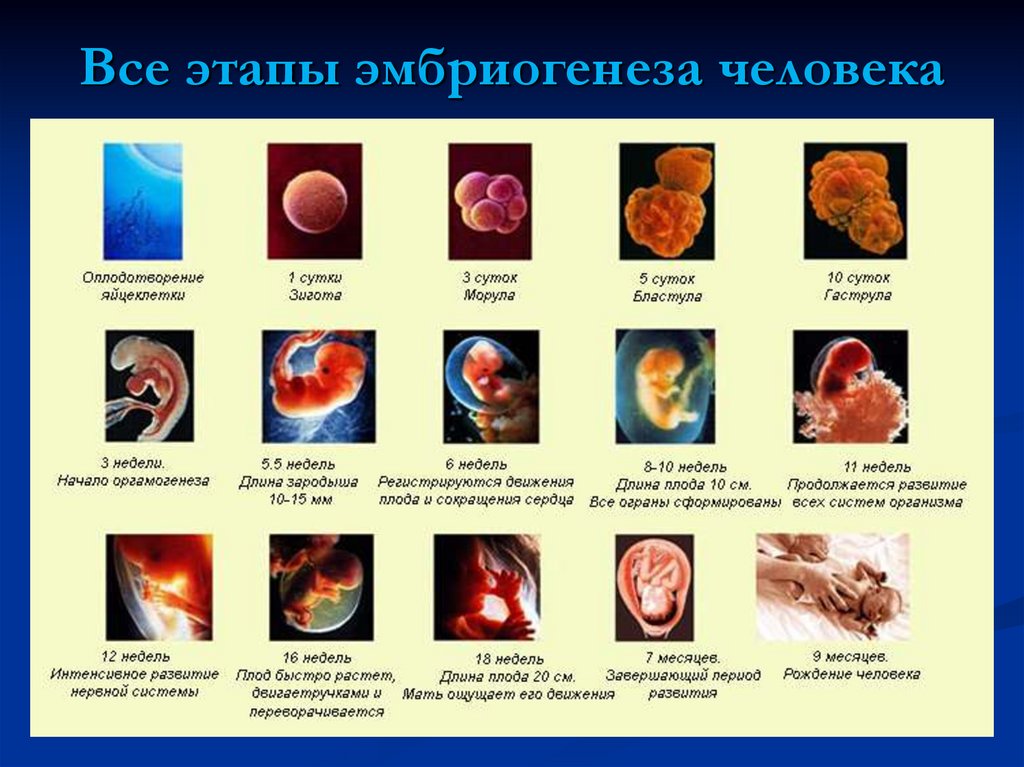 Вымершие представители этого подтипа включают Pikaia , который является старейшим из известных головохордовых. Окаменелости Pikaia были извлечены из сланцев Берджесс в Канаде и датированы серединой кембрийского периода, то есть им более 500 миллионов лет.
Вымершие представители этого подтипа включают Pikaia , который является старейшим из известных головохордовых. Окаменелости Pikaia были извлечены из сланцев Берджесс в Канаде и датированы серединой кембрийского периода, то есть им более 500 миллионов лет.
Существующие представители Cephalochordata — это ланцетников , названных в честь их лезвиеобразной формы. Ланцетники имеют длину всего несколько сантиметров и обычно находятся зарытыми в песок на дне теплых умеренных и тропических морей. Как и оболочники, они питаются подвеской.
Рисунок 4. У ланцетника, как и у всех головохордовых, есть голова. Взрослые ланцетники сохраняют четыре ключевых признака хордовых: хорду, спинной полый нервный тяж, глоточные щели и постанальный хвост. Вода изо рта попадает в глоточные щели, которые отфильтровывают частицы пищи. Затем отфильтрованная вода собирается в атриуме и выходит через атриопору.
Черепные и позвоночные
Рисунок 5.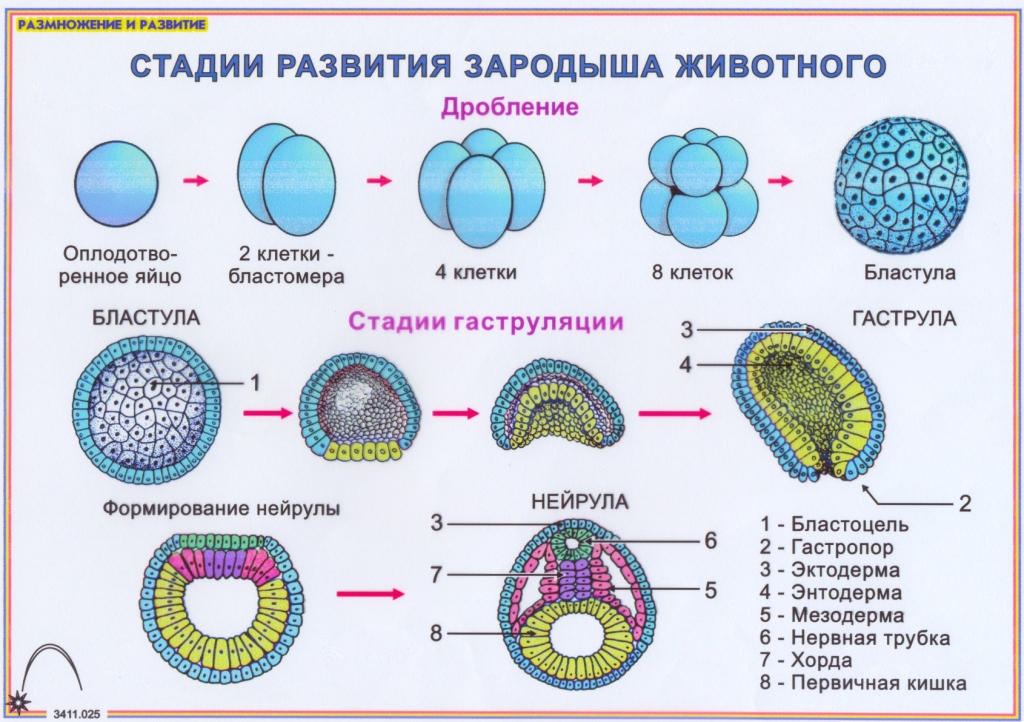 Craniata, включая эту рыбу (Dunkleosteus sp.), характеризуется наличием черепа, нижней челюсти и других лицевых костей. (кредит: «Стевок 86»/Викисклад)
Craniata, включая эту рыбу (Dunkleosteus sp.), характеризуется наличием черепа, нижней челюсти и других лицевых костей. (кредит: «Стевок 86»/Викисклад)
Череп представляет собой костную, хрящевую или волокнистую структуру, окружающую головной мозг, челюсть и лицевые кости (рис. 5). У большинства билатерально-симметричных животных есть голова; из них те, у которых есть череп, составляют кладу Craniata . Craniata включает миксин (Myxini), у которых есть череп, но нет позвоночника, и все организмы, называемые «позвоночными».
Позвоночные относятся к кладе Vertebrata . Позвоночные обладают четырьмя характерными чертами хордовых; однако члены этой группы также имеют общие производные характеристики, которые отличают их от хордовых беспозвоночных. Позвоночные названы в честь позвоночного столба , состоящего из позвонков, ряда отдельных костей, соединенных вместе в виде позвоночника (рис. 6). У взрослых позвоночных позвоночный столб заменяет хорду, которая наблюдается только на эмбриональной стадии.
Рисунок 6. Для позвоночных характерно наличие позвоночника, такого как тот, который проходит через середину этой рыбы. Все позвоночные относятся к кладе Craniata и имеют череп. (Фото: Эрнест В. Мор; сделано в Смитсоновском музее естественной истории, Вашингтон, округ Колумбия)
На основе молекулярного анализа позвоночные, по-видимому, более тесно связаны с ланцетниками (головохордовые), чем с оболочниками (урохордовые) среди хордовых беспозвоночных. Эти данные свидетельствуют о том, что головохордовые отошли от Urochordata, а позвоночные впоследствии отошли от головохордовых. Эта гипотеза также подтверждается открытием в Китае окаменелости рода 9.0009 Хайкоуэла . Этот организм, по-видимому, является промежуточной формой между головохордовыми и позвоночными. Окаменелости Haikouella имеют возраст около 530 миллионов лет и кажутся похожими на современных ланцетников. У этих организмов были мозг и глаза, как у позвоночных, но не было черепа, как у черепных.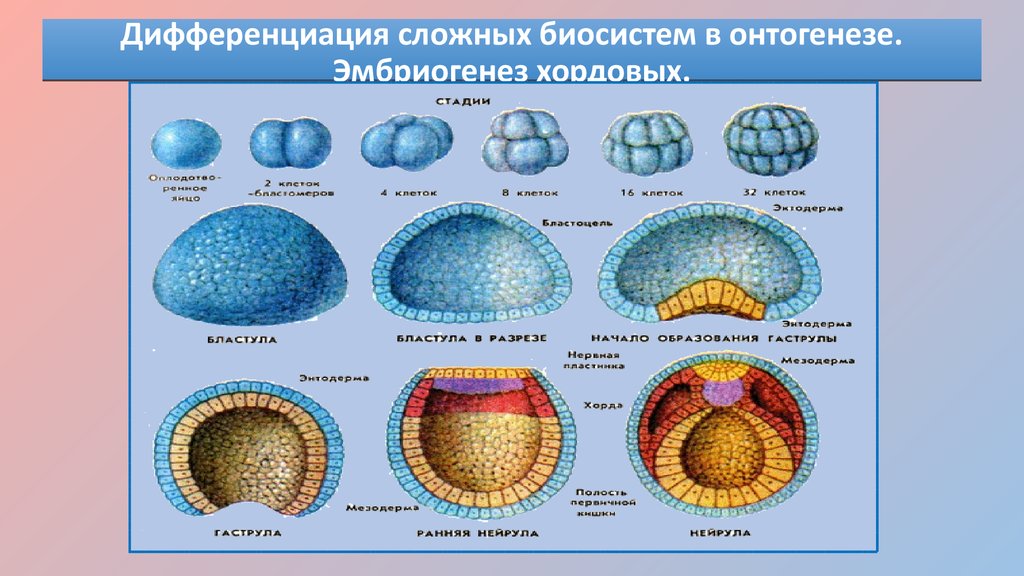 1 Эти данные свидетельствуют о том, что позвоночные возникли во время кембрийского взрыва. Напомним, что «кембрийским взрывом» называют относительно короткий промежуток времени в кембрийском периоде, в течение которого возникло и быстро диверсифицировалось множество групп животных. Большинство современных типов животных возникли во время кембрийского взрыва.
1 Эти данные свидетельствуют о том, что позвоночные возникли во время кембрийского взрыва. Напомним, что «кембрийским взрывом» называют относительно короткий промежуток времени в кембрийском периоде, в течение которого возникло и быстро диверсифицировалось множество групп животных. Большинство современных типов животных возникли во время кембрийского взрыва.
Позвоночные — крупнейшая группа хордовых, насчитывающая более 62 000 живых видов. Позвоночные классифицируются по анатомическим и физиологическим признакам. Для этих животных используется более одной схемы классификации и наименования. Здесь мы рассмотрим традиционные группы Agnatha, Chondrichthyes, Osteichthyes, Amphibia, Reptilia, Aves и Mammalia, составляющие классы подтипа Vertebrata. Многие современные авторы относят птиц к Reptilia, что правильно отражает их эволюционное наследие. Мы рассмотрим их отдельно только для удобства. Далее мы будем рассматривать миксин и миног вместе как бесчелюстных рыб, бесчелюстных, хотя появляющиеся классификационные схемы разделяют их на хордовых бесчелюстных рыб (миноги) и позвоночных бесчелюстных рыб (миноги).
Животные, обладающие челюстями, известны как гнатостомы, что означает «челюстной рот». Гнатостомы включают рыб и четвероногих — земноводных, рептилий, птиц и млекопитающих. Тетраподы можно разделить на две группы: амфибии и амниоты. Амниоты — это животные, яйца которых приспособлены к наземной жизни, и в эту группу входят млекопитающие, рептилии и птицы. Амниотические зародыши, развивающиеся либо во внешне отлитом яйце, либо в яйце, выносимом самкой, обеспечены влагоудерживающей средой и защищены амниотическими мембранами.
Резюме раздела
Характерными признаками хордовых являются хорда, спинной полый нервный тяж, глоточные щели и постанальный хвост. Chordata включает две клады беспозвоночных: Urochordata (оболочечные) и Cephalochordata (ланцетники), вместе с позвоночными позвоночными. Большинство оболочников живут на дне океана и питаются взвесью. Ланцетники питаются фитопланктоном и другими микроорганизмами. Позвоночные названы в честь позвоночного столба, который характерен почти для всех членов этой клады.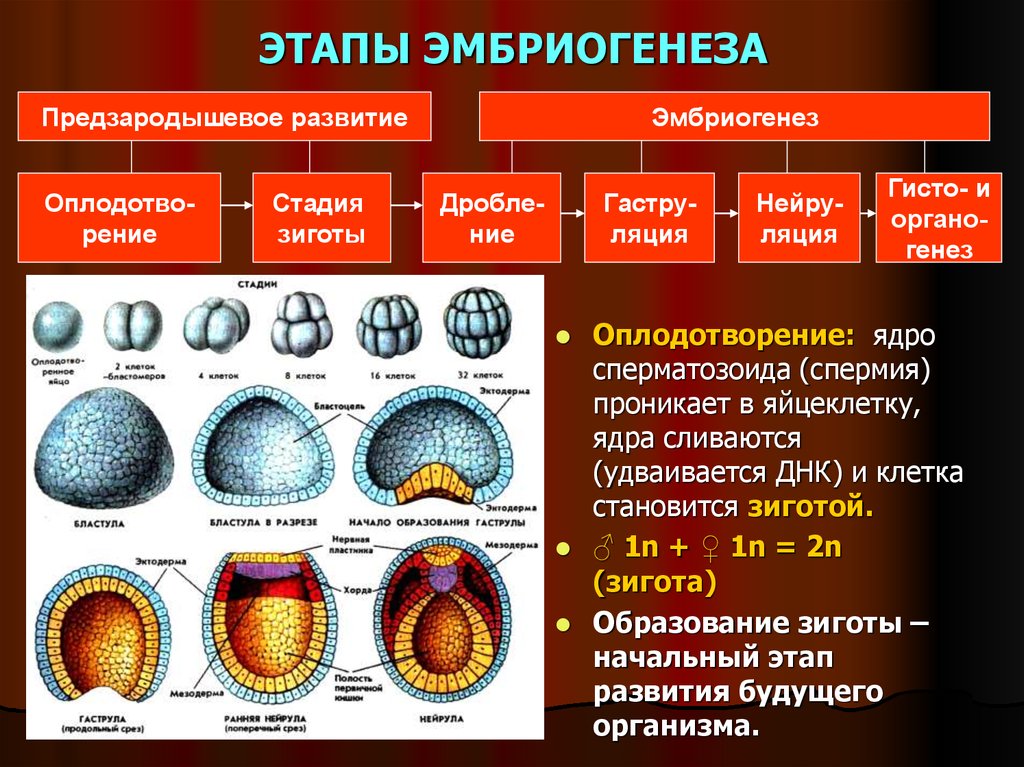
Leave A Comment