
Половые клетки человека, хромосомы, оплодотворение — статья МЦРМ
Половые клетки — гаметы (от греч. gametes — «супруг») можно обнаружить уже у двухнедельного эмбриона человека. Их называют первичными половыми клетками. В это время они совсем не похожи на сперматозоиды или яйцеклетки и выглядят абсолютно одинаковыми. Никаких различий, присущих зрелым гаметам, на этой стадии развития зародыша обнаружить у первичных половых клеток не удается. Это не единственная их особенность. Во-первых, первичные половые клетки появляются у зародыша гораздо раньше собственно половой железы (гонады), а во-вторых, они возникают на значительном удалении от того места, где эти железы сформируются позднее. В определенный момент происходит совершенно удивительный процесс — первичные половые клетки дружно устремляются к половой железе и заселяют, «колонизируют» ее.
После того, как будущие гаметы попали в половые железы, они начинают интенсивно делиться, и количество их увеличивается.
 Теперь в гаплоидной клетке копируются имеющиеся 23 хромосомы и эти копии используются для образования новой клетки. Таким образом, в результате описанных двух делений из одной первичной половой клетки образуется 4 новых.
Теперь в гаплоидной клетке копируются имеющиеся 23 хромосомы и эти копии используются для образования новой клетки. Таким образом, в результате описанных двух делений из одной первичной половой клетки образуется 4 новых.
Причем, в 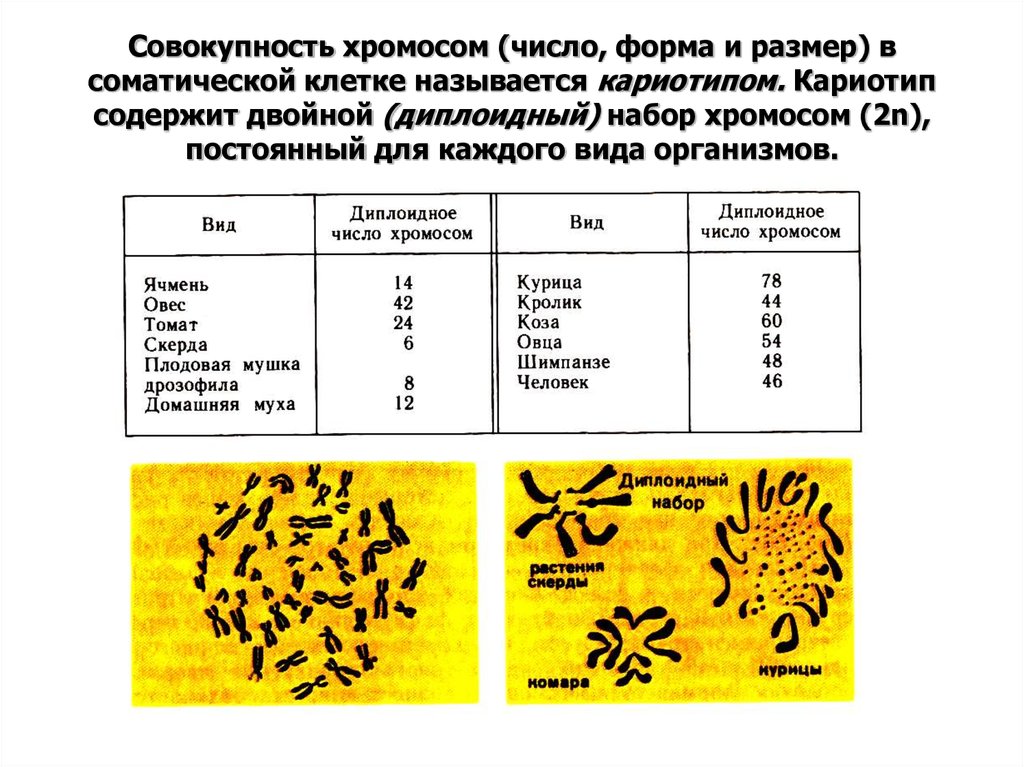
Первичные половые клетки, оказавшиеся в мужской половой железе, до поры до времени не делятся. Их деление начинается только в период полового созревания и приводит к образованию когорты так называемых стволовых диплоидных клеток, из которых и формируются сперматозоиды. Запас стволовых клеток в яичках постоянно пополняется. Здесь уместно напомнить описанную выше особенность сперматогенеза — из одной клетки образуется 4 зрелых сперматозоида. Таким образом, после полового созревания у мужчины в течение всей жизни формируются сотни миллиардов новых сперматозоидов.
Формирование яйцеклеток протекает иначе. Едва заселив половую железу, первичные половые клетки начинают интенсивно делиться. К 5 месяцу внутриутробного развития их количество достигает 6-7 миллионов, но затем происходит массовая гибель этих клеток. В яичниках новорожденной девочки их остается не более 1-2 миллионов, к 7-летнему возрасту — всего лишь около 300 тысяч, а в период полового созревания 30 —50 тысяч. Общее же число яйцеклеток, которые достигнут зрелого состояния за период половой зрелости, будет еще меньше. Хорошо известно, что в течение одного менструального цикла в яичнике обычно созревает лишь один фолликул. Нетрудно подсчитать, что в течение репродуктивного периода, продолжающегося у женщин 30 — 35 лет, образуется около 400 зрелых яйцеклеток.
В яичниках новорожденной девочки их остается не более 1-2 миллионов, к 7-летнему возрасту — всего лишь около 300 тысяч, а в период полового созревания 30 —50 тысяч. Общее же число яйцеклеток, которые достигнут зрелого состояния за период половой зрелости, будет еще меньше. Хорошо известно, что в течение одного менструального цикла в яичнике обычно созревает лишь один фолликул. Нетрудно подсчитать, что в течение репродуктивного периода, продолжающегося у женщин 30 — 35 лет, образуется около 400 зрелых яйцеклеток.
Если мейоз в сперматогенезе начинается в период полового созревания и повторяется миллиарды раз в течение жизни мужчины, в оогенезе формирующиеся женские гаметы вступают в мейоз еще в периоде внутриутробного развития. Причем начинается этот процесс почти одновременно у всех будущих яйцеклеток. Начинается, но не заканчивается! Будущие яйцеклетки доходят только до середины первой фазы мейоза, а дальше процесс деления блокируется на 12 — 50 лет! Лишь с приходом половой зрелости мейоз в оогенезе продолжится, причем не всех клеток сразу, а лишь для 1- 2 яйцеклеток ежемесячно.
Сперматогенез и оогенез
Вынашивание и рождение ребенка является чрезвычайно тяжелой нагрузкой на организм. Врачи говорят, что беременность — это проба на здоровье. Каким родится ребенок — напрямую зависит от состояния здоровья матери. Здоровье, как известно, не вечно. Старость и болезни, к сожалению, неотвратимы. Природа дает женщине строго ограниченное невосполнимое число половых клеток.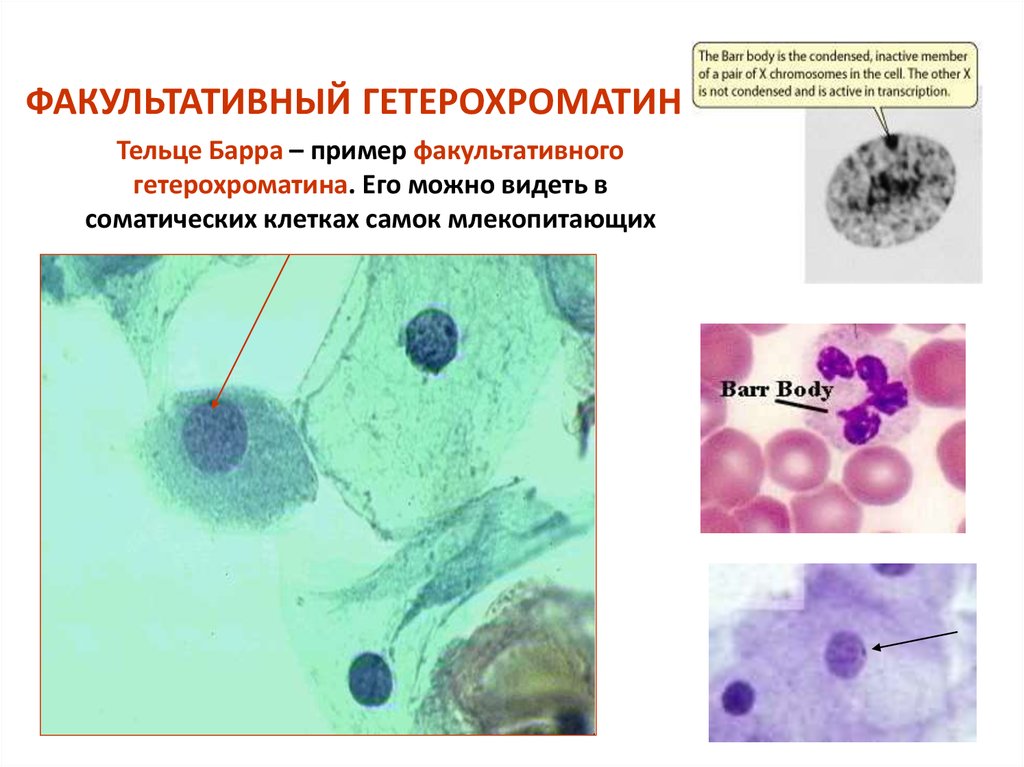
По каким законам происходит деление хромосом? Как передается наследственная информация? Для того чтобы разобраться с этим вопросом, можно привести простую аналогию с картами.
Для того чтобы получить из диплоидной соматической клетки половую клетку, число хромосом должно быть уменьшено вдвое. При этом половая клетка обязательно должна содержать полный одинарный (гаплоидный) набор хромосом. Ни одна не должна потеряться! В случае карт такой набор можно получить следующим образом. Взять наугад из каждой пары карт черной масти по одной и таким образом сформировать два одиночных набора. Каждый набор будет включать все карты черной масти от шестерки до туза, однако, какие именно это будут карты (трефы или пики) определил случай. Например, в одном таком наборе шестерка может быть пиковой, а в другом — трефовой.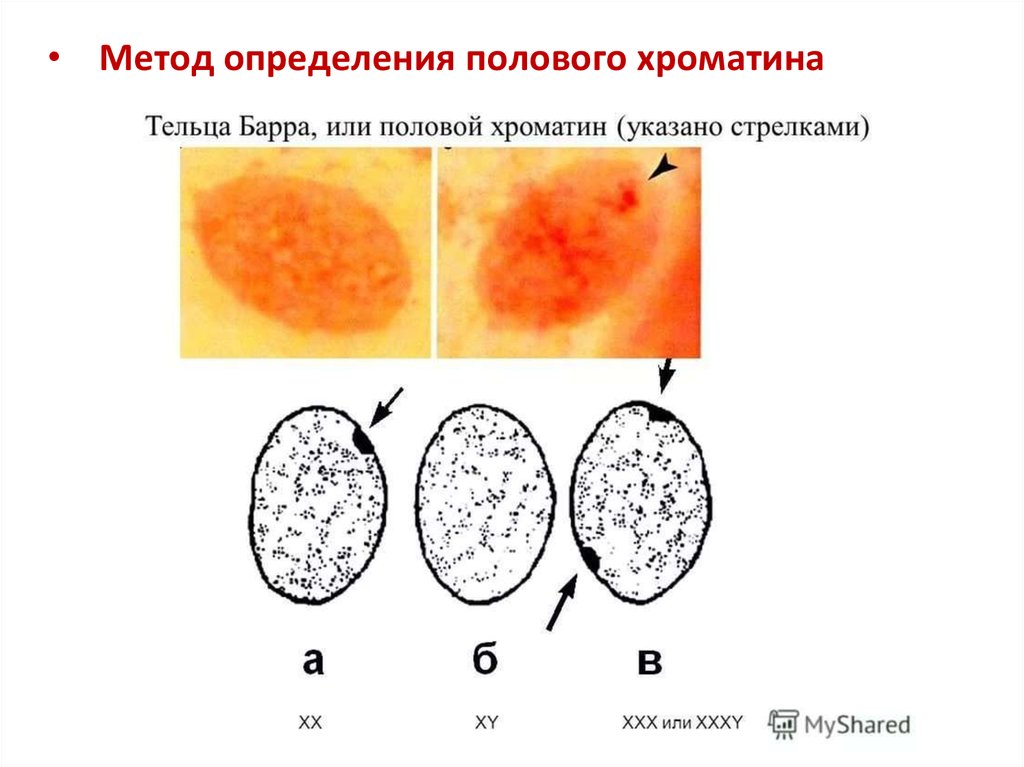
Точно также будем составлять одиночный набор из ее карт красной масти. Получим еще более 500 разных вариантов. Из его одиночного и ее одиночного набора карт составим двойной набор. Он получится мягко сказать «пестреньким»: в каждой паре карт одна будет красной масти, а другая — черной. Общее число таких возможных наборов 500×500, то есть 250 тысяч вариантов.
Примерно также, по закону случайной выборки, поступает и природа с хромосомами в процессе мейоза. В результате из клеток с двойным, диплоидным набором хромосом получаются клетки, каждая из которых содержит одиночный, гаплоидный полный набор хромосом. Предположим, в результате мейоза в вашем теле образовалась половая клетка. Сперматозоид или яйцеклетка — в данном случае не важно. Она обязательно будет содержать гаплоидный набор хромосом — ровно 23 штуки. Что именно это за хромосомы? Рассмотрим для примера хромосому № 7.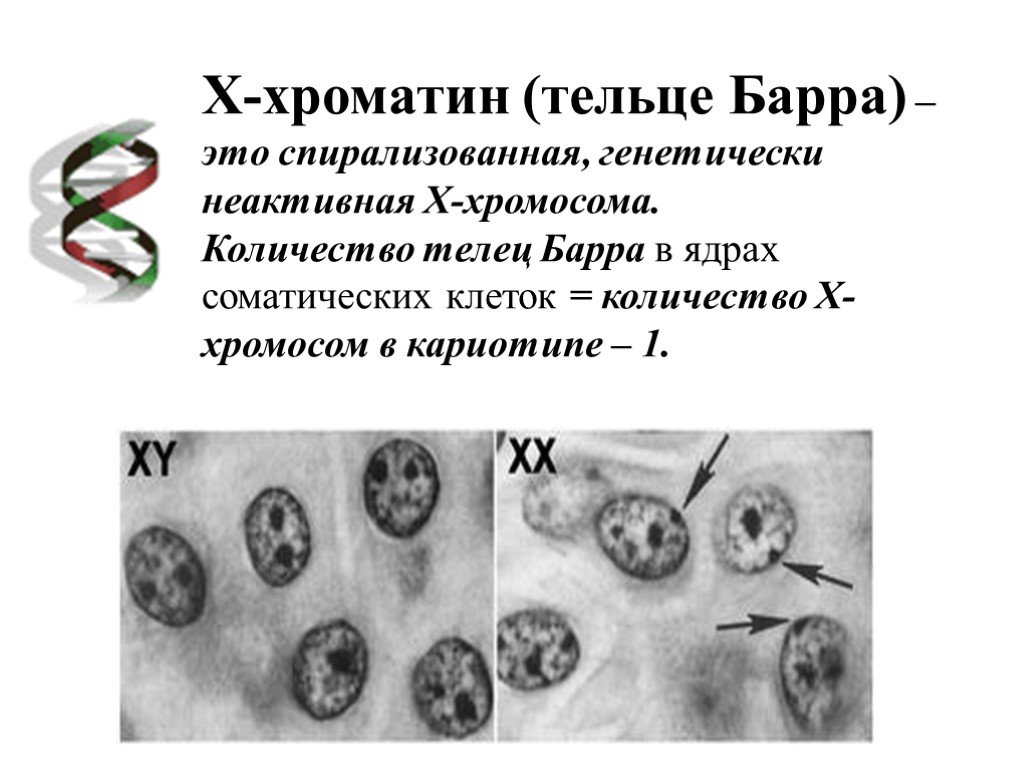 Это может быть хромосома, которую вы получили от отца. С равной вероятностью она может быть хромосомой, которую вы получили от матери. То же самое справедливо для хромосомы № 8, и для любой другой.
Это может быть хромосома, которую вы получили от отца. С равной вероятностью она может быть хромосомой, которую вы получили от матери. То же самое справедливо для хромосомы № 8, и для любой другой.
Поскольку у человека число хромосом гаплоидного набора равно 23, то число возможных вариантов половых гаплоидных клеток, образующихся из диплоидных соматических, равно 2 в степени 23. Получается более 8 миллионов вариантов! В процессе оплодотворения две половые клетки соединяются между собой. Следовательно, общее число таких комбинаций будет равно 8 млн. х 8 млн. = 64000 млрд. вариантов! На уровне пары гомологичных хромосом основа этого разнообразия выглядит так. Возьмем любую пару гомологичных хромосом вашего диплоидного набора. Одну из таких хромосом вы получили от матери, но это может быть хромосома либо вашей бабушки, либо вашего дедушки по материнской линии. Вторую гомологичную хромосому вы получили от отца. Однако она опять-таки может быть независимо от первой либо хромосомой вашей бабушки, либо вашего дедушки уже по отцовской линии.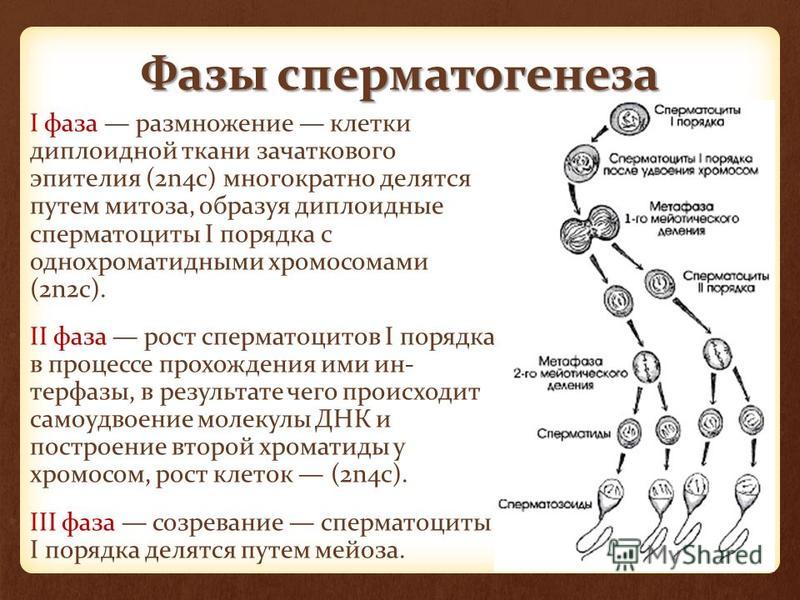 А таких гомологических хромосом у вас 23 пары! Получается невероятное число возможных комбинаций. Неудивительно, что при этом у одной пары родителей, рождаются дети, которые отличаются друг от друга и внешностью, и характером.
А таких гомологических хромосом у вас 23 пары! Получается невероятное число возможных комбинаций. Неудивительно, что при этом у одной пары родителей, рождаются дети, которые отличаются друг от друга и внешностью, и характером.
Кстати, из приведенных выше расчетов следует простой, но важный вывод. Каждый человек, ныне здравствующий, или когда-либо живший в прошлом на Земле, абсолютно уникален. Шансы появления второго такого же практически равны нулю. Поэтому не надо себя ни с кем сравнивать. Каждый из вас неповторим, и тем уже интересен!
Однако вернемся к нашим половым клеткам. Каждая диплоидная клетка человека содержит 23 пары хромосом. Хромосомы с 1 по 22 пару называются соматическим и по форме они одинаковы. Хромосомы же 23-й пары (половые хромосомы) одинаковы только у женщин. Они и обозначаются латинскими буквами ХХ. У мужчин хромосомы этой пары различны и обозначаются ХY. В гаплоидном наборе яйцеклетки половая хромосома всегда только Х, сперматозоид же может нести или Х или Y хромосому. Если яйцеклетку оплодотворит Х сперматозоид, родится девочка, если Y сперматозоид — мальчик. Все просто!
Если яйцеклетку оплодотворит Х сперматозоид, родится девочка, если Y сперматозоид — мальчик. Все просто!
Почему мейоз у яйцеклетки так долго растянут во времени? Каким образом ежемесячно происходит выбор когорты фолликулов, которые начинают свое развитие и как из них выделяется лидирующий, доминантный, овуляторный фолликул, в котором созреет яйцеклетка? На все эти непростые вопросы у биологов нет пока однозначных ответов. Процесс формирования зрелых яйцеклеток у человека ждет новых исследователей!
Образование и созревание сперматозоидов, как уже было сказано, происходит в семенных канальцах мужской половой железы — яичках. Сформированный сперматозоид имеет длину около 50-60 микрон. Ядро сперматозоида находится в его головке. Оно содержит отцовский наследственный материал. За головкой располагается шейка, в которой имеется крупная извитая митохондрия — органоид, обеспечивающий движения хвоста. Иначе говоря, это своеобразная «энергетическая станция». На головке сперматозоида есть «шапочка». Благодаря ей форма головки — овальная. Но, дело не в форме, а в том, что содержится под «шапочкой». «Шапочка» эта на самом деле является контейнером и называется акросомой, а содержатся в ней ферменты, которые способны растворять оболочку яйцеклетки, что необходимо для проникновения сперматозоида внутрь — в цитоплазму яйцеклетки. Если у сперматозоида нет акросомы, головка у него не овальная, а круглая. Эта патология сперматозоидов называется глобулоспермия (круглоголовые сперматозоиды). Но, беда опять не в форме, а в том, что такой сперматозоид не может оплодотворить яйцеклетку, и мужчина с таким нарушением сперматогенеза до начала 90-х прошлого столетия был обречен на бездетность. Сегодня благодаря ВРТбесплодие у этих мужчин может быть преодолено, но об этом мы расскажем позднее в главе, посвященной микроманипуляциям, в частности, ИКСИ.
На головке сперматозоида есть «шапочка». Благодаря ей форма головки — овальная. Но, дело не в форме, а в том, что содержится под «шапочкой». «Шапочка» эта на самом деле является контейнером и называется акросомой, а содержатся в ней ферменты, которые способны растворять оболочку яйцеклетки, что необходимо для проникновения сперматозоида внутрь — в цитоплазму яйцеклетки. Если у сперматозоида нет акросомы, головка у него не овальная, а круглая. Эта патология сперматозоидов называется глобулоспермия (круглоголовые сперматозоиды). Но, беда опять не в форме, а в том, что такой сперматозоид не может оплодотворить яйцеклетку, и мужчина с таким нарушением сперматогенеза до начала 90-х прошлого столетия был обречен на бездетность. Сегодня благодаря ВРТбесплодие у этих мужчин может быть преодолено, но об этом мы расскажем позднее в главе, посвященной микроманипуляциям, в частности, ИКСИ.
Перемещение сперматозоида осуществляется за счет движения его хвостика.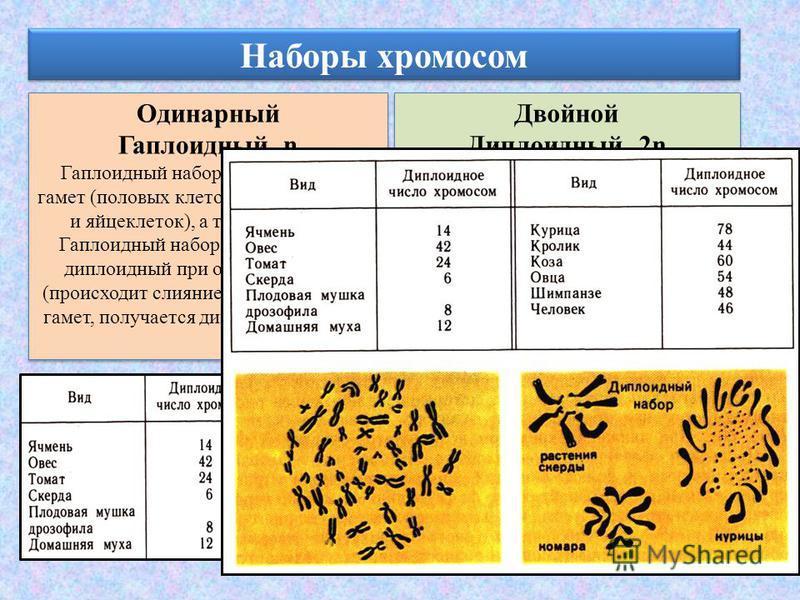 Скорость движения сперматозоида не превышает 2-3 мм в минуту. Казалось бы, немного, однако, за 2-3 часа в женском половом тракте сперматозоиды проходят путь, в 80000 раз превышающий их собственные размеры! Будь на месте сперматозоида в этой ситуации человек, ему пришлось бы двигаться вперед со скоростью 60-70 км/час — то есть со скоростью автомобиля!
Скорость движения сперматозоида не превышает 2-3 мм в минуту. Казалось бы, немного, однако, за 2-3 часа в женском половом тракте сперматозоиды проходят путь, в 80000 раз превышающий их собственные размеры! Будь на месте сперматозоида в этой ситуации человек, ему пришлось бы двигаться вперед со скоростью 60-70 км/час — то есть со скоростью автомобиля!
Сперматозоиды, находящиеся в яичке, неподвижны. Способность к движению они приобретают лишь, проходя по семявыводящим путям под воздействием жидкостей семявыводящих протоков и семенных пузырьков, секрета предстательной железы. В половых путях женщины сперматозоиды сохраняют подвижность в течение 3 — 4 суток, но оплодотворить яйцеклетку они должны в течение 24 часов. Весь процесс развития от стволовой клетки до зрелого сперматозоида длится примерно 72 дня. Однако, поскольку сперматогенез происходит непрерывно и в него одномоментно вступает громадное число клеток, то в яичках всегда есть большое количество спермиев, находящихся на разных этапах сперматогенеза, а запас зрелых сперматозоидов постоянно пополняется. Активность сперматогенеза индивидуальна, но с возрастом снижается.
Активность сперматогенеза индивидуальна, но с возрастом снижается.
Как мы уже говорили, яйцеклетки находятся в фолликулах яичника. В результате овуляции яйцеклетка попадает в брюшную полость, откуда она «вылавливается» фимбриями маточной трубы и переносится в просвет ее ампулярного отдела. Именно здесь происходит встреча яйцеклетки со сперматозоидами.
Какое же строение имеет зрелая яйцеклетка? Она довольно крупная и достигает 0,11-0,14 мм в диаметре. Сразу после овуляции яйцеклетка окружена скоплением мелких клеток и желатинообразной массой (так называемым лучистым венцом). Видимо, в таком виде фимбриям маточной трубы удобнее захватывать яйцеклетку. В просвете маточной трубы с помощью ферментов и механического воздействия (биения ресничек эпителия), происходит «очистка» яйцеклетки от лучистого венца. Окончательно освобождение яйцеклетки от лучистого венца происходит после встречи ее со сперматозоидами, которые буквально облепляют яйцеклетку. Каждый сперматозоид выделяет из акросомы фермент, растворяющий не только лучистый венец, но и действующий на оболочку самой яйцеклетки. Эта оболочка называется блестящей, так она выглядит под микроскопом. Выделяя фермент, все сперматозоиды стремятся оплодотворить яйцеклетку, но блестящая оболочка пропустит лишь один из них. Получается, что устремляясь к яйцеклетки, воздействуя на нее коллективно, сперматозоиды «расчищают дорогу» только для одного счастливчика. Отбором сперматозоида роль блестящей оболочки не ограничивается, на ранних стадиях развития эмбриона она поддерживает упорядоченное расположение его клеток (бластомеров). В какой-то момент блестящая оболочка становится тесной, она разрывается и происходит хетчинг (от анг. hatching — «вылупление») — вылупление эмбриона. Эмбрион готов к имплантации в эндометрий.
46 — норма? Считаем хромосомы: сколько человеку для счастья нужно
В отличие от зубов, хромосом человеку положено иметь строго определенное число — 46 штук. Однако при ближайшем рассмотрении выясняется, что каждый из нас может оказаться носителем лишних хромосом. Откуда они берутся, где прячутся и какой наносят вред (а может, пользу?) — разберемся с участием современной научной литературы
Прожиточный оптимум
Сначала договоримся о терминологии. Окончательно человеческие хромосомы посчитали чуть больше полувека назад — в 1956 году. С тех пор мы знаем, что в соматических, то есть не половых клетках, их обычно 46 штук — 23 пары.
Хромосомы в паре (одна получена от отца, другая — от матери) называют гомологичными. На них расположены гены, выполняющие одинаковые функции, однако нередко различающиеся по строению. Исключение составляют половые хромосомы — Х и Y, генный состав которых совпадает не полностью. Все остальные хромосомы, кроме половых, называют аутосомами.
Количество наборов гомологичных хромосом — плоидность — в половых клетках равно одному, а в соматических, как правило, двум.
Интересно, что не у всех видов млекопитающих число хромосом постоянно. Например, у некоторых представителей грызунов, собак и оленей обнаружили так называемые В-хромосомы. Это небольшие дополнительные хромосомы, в которых практически нет участков, кодирующих белки, а делятся и наследуются они вместе с основным набором и, как правило, не влияют на работу организма. Полагают, что В-хромосомы — это просто удвоенные фрагменты ДНК, «паразитирующие» на основном геноме.
У человека до сих пор В-хромосомы обнаружены не были. Зато иногда в клетках возникает дополнительный набор хромосом — тогда говорят о полиплоидии, а если их число не кратно 23 — об анеуплоидии. Полиплоидия встречается у отдельных типов клеток и способствует их усиленной работе, в то время как анеуплоидия обычно свидетельствует о нарушениях в работе клетки и нередко приводит к ее гибели.
Делиться надо честно
Чаще всего неправильное количество хромосом является следствием неудачного деления клеток. В соматических клетках после удвоения ДНК материнская хромосома и ее копия оказываются сцеплены вместе белками когезинами. Потом на их центральные части садятся белковые комплексы кинетохоры, к которым позже прикрепляются микротрубочки. При делении по микротрубочкам кинетохоры разъезжаются к разным полюсам клетки и тянут за собой хромосомы. Если сшивки между копиями хромосомы разрушатся раньше времени, то к ним могут прикрепиться микротрубочки от одного и того же полюса, и тогда одна из дочерних клеток получит лишнюю хромосому, а вторая останется обделенной.
Деление при образовании половых клеток (мейоз) устроено более сложно. После удвоения ДНК каждая хромосома и ее копия, как обычно, сшиты когезинами. Затем гомологичные хромосомы (полученные от отца и матери), а точнее их пары, тоже сцепляются друг с другом, и получается так называемая тетрада, или четверка. А дальше клетке предстоит поделиться два раза. В ходе первого деления расходятся гомологичные хромосомы, то есть дочерние клетки содержат пары одинаковых хромосом. А во втором делении эти пары расходятся, и в результате половые клетки несут одинарный набор хромосом.
Мейоз тоже нередко проходит с ошибками. Проблема в том, что конструкция из сцепленных двух пар гомологичных хромосом может перекручиваться в пространстве или разделяться в неположенных местах. Результатом снова будет неравномерное распределение хромосом. Иногда половой клетке удается это отследить, чтобы не передавать дефект по наследству. Лишние хромосомы часто неправильно уложены или разорваны, что запускает программу гибели. Например, среди сперматозоидов действует такой отбор по качеству. А вот яйцеклеткам повезло меньше. Все они у человека образуются еще до рождения, готовятся к делению, а потом замирают. Хромосомы уже удвоены, тетрады образованы, а деление отложено. В таком виде они живут до репродуктивного периода. Дальше яйцеклетки по очереди созревают, делятся первый раз и снова замирают. Второе деление происходит уже сразу после оплодотворения. И на этом этапе проконтролировать качество деления уже сложно. А риски больше, ведь четыре хромосомы в яйцеклетке остаются сшитыми в течение десятков лет. За это время в когезинах накапливаются поломки, и хромосомы могут спонтанно разделяться. Поэтому чем старше женщина, тем больше вероятность неправильного расхождения хромосом в яйцеклетке.
Анеуплоидия в половых клетках неизбежно ведет к анеуплоидии зародыша. При оплодотворении здоровой яйцеклетки с 23 хромосомами сперматозоидом с лишней или недостающей хромосомами (или наоборот) число хромосом у зиготы, очевидно, будет отлично от 46. Но даже если половые клетки здоровы, это не дает гарантий здорового развития. В первые дни после оплодотворения клетки зародыша активно делятся, чтобы быстро набрать клеточную массу. Судя по всему, в ходе быстрых делений нет времени проверять корректность расхождения хромосом, поэтому могут возникнуть анеуплоидные клетки. И если произойдет ошибка, то дальнейшая судьба зародыша зависит от того, в каком делении это случилось. Если равновесие нарушено уже в первом делении зиготы, то весь организм вырастет анеуплоидным. Если же проблема возникла позже, то исход определяется соотношением здоровых и аномальных клеток.
Часть последних может дальше погибнуть, и мы никогда не узнаем об их существовании. А может принять участие в развитии организма, и тогда он получится мозаичным — разные клетки будут нести разный генетический материал. Мозаицизм доставляет немало хлопот пренатальным диагностам. Например, при риске рождения ребенка с синдромом Дауна иногда извлекают одну или несколько клеток зародыша (на той стадии, когда это не должно представлять опасности) и считают в них хромосомы. Но если зародыш мозаичен, то такой метод становится не особенно эффективным.
Третий лишний
Все случаи анеуплоидии логично делятся на две группы: недостаток и избыток хромосом. Проблемы, возникающие при недостатке, вполне ожидаемы: минус одна хромосома означает минус сотни генов.
Расположение хромосом в ядре клетки человека (хромосомные территории). Изображение: Bolzer et al. , 2005 / Wikimedia Commons / CC BY 2.5
Если гомологичная хромосома работает нормально, то клетка может отделаться только недостаточным количеством закодированных там белков. Но если среди оставшихся на гомологичной хромосоме генов какие-то не работают, то соответствующих белков в клетке не появится совсем.
В случае избытка хромосом все не так очевидно. Генов становится больше, но здесь — увы — больше не значит лучше.
Во-первых, лишний генетический материал увеличивает нагрузку на ядро: дополнительную нить ДНК нужно разместить в ядре и обслужить системами считывания информации.
Ученые обнаружили, что у людей с синдромом Дауна, чьи клетки несут дополнительную 21-ю хромосому, в основном нарушается работа генов, находящихся на других хромосомах. Видимо, избыток ДНК в ядре приводит к тому, что белков, поддерживающих работу хромосом, не хватает на всех.
Во-вторых, нарушается баланс в количестве клеточных белков. Например, если за какой-то процесс в клетке отвечают белки-активаторы и белки-ингибиторы и их соотношение обычно зависит от внешних сигналов, то дополнительная доза одних или других приведет к тому, что клетка перестанет адекватно реагировать на внешний сигнал. И наконец, у анеуплоидной клетки растут шансы погибнуть. При удвоении ДНК перед делением неизбежно возникают ошибки, и клеточные белки системы репарации их распознают, чинят и запускают удвоение снова. Если хромосом слишком много, то белков не хватает, ошибки накапливаются и запускается апоптоз — программируемая гибель клетки. Но даже если клетка не погибает и делится, то результатом такого деления тоже, скорее всего, станут анеуплоиды.
Жить будете
Если даже в пределах одной клетки анеуплоидия чревата нарушениями работы и гибелью, то неудивительно, что целому анеуплоидному организму выжить непросто. На данный момент известно только три аутосомы — 13, 18 и 21-я, трисомия по которым (то есть лишняя, третья хромосома в клетках) как-то совместима с жизнью. Вероятно, это связано с тем, что они самые маленькие и несут меньше всего генов. При этом дети с трисомией по 13-й (синдром Патау) и 18-й (синдром Эдвардса) хромосомам доживают в лучшем случае до 10 лет, а чаще живут меньше года. И только трисомия по самой маленькой в геноме, 21-й хромосоме, известная как синдром Дауна, позволяет жить до 60 лет.
Совсем редко встречаются люди с общей полиплоидией. В норме полиплоидные клетки (несущие не две, а от четырех до 128 наборов хромосом) можно обнаружить в организме человека, например в печени или красном костном мозге. Это, как правило, большие клетки с усиленным синтезом белка, которым не требуется активное деление.
Дополнительный набор хромосом усложняет задачу их распределения по дочерним клеткам, поэтому полиплоидные зародыши, как правило, не выживают. Тем не менее описано около 10 случаев, когда дети с 92 хромосомами (тетраплоиды) появлялись на свет и жили от нескольких часов до нескольких лет. Впрочем, как и в случае других хромосомных аномалий, они отставали в развитии, в том числе и умственном. Однако многим людям с генетическими аномалиями приходит на помощь мозаицизм. Если аномалия развилась уже в ходе дробления зародыша, то некоторое количество клеток могут остаться здоровыми. В таких случаях тяжесть симптомов снижается, а продолжительность жизни растет.
Гендерные несправедливости
Однако есть и такие хромосомы, увеличение числа которых совместимо с жизнью человека или даже проходит незаметно. И это, как ни удивительно, половые хромосомы. Причиной тому — гендерная несправедливость: примерно у половины людей в нашей популяции (девочек) Х-хромосом в два раза больше, чем у других (мальчиков). При этом Х-хромосомы служат не только для определения пола, но и несут более 800 генов (то есть в два раза больше, чем лишняя 21-я хромосома, доставляющая немало хлопот организму). Но девочкам приходит на помощь естественный механизм устранения неравенства: одна из Х-хромосом инактивируется, скручивается и превращается в тельце Барра. В большинстве случаев выбор происходит случайно, и в ряде клеток в результате активна материнская Х-хромосома, а в других — отцовская. Таким образом, все девочки оказываются мозаичными, потому что в разных клетках работают разные копии генов. Классическим примером такой мозаичности являются черепаховые кошки: на их Х-хромосоме находится ген, отвечающий за меланин (пигмент, определяющий, среди прочего, цвет шерсти). В разных клетках работают разные копии, поэтому окраска получается пятнистой и не передается по наследству, так как инактивация происходит случайным образом.
Кошка черепахового окраса. Фото: Lisa Ann Yount / Flickr / Public domain
В результате инактивации в клетках человека всегда работает только одна Х-хромосома. Этот механизм позволяет избежать серьезных неприятностей при Х-трисомии (девочки ХХХ) и синдромах Шерешевского — Тернера (девочки ХО) или Клайнфельтера (мальчики ХХY). Таким рождается примерно один из 400 детей, но жизненные функции в этих случаях обычно не нарушены существенно, и даже бесплодие возникает не всегда. Сложнее бывает тем, у кого хромосом больше трех. Обычно это значит, что хромосомы не разошлись дважды при образовании половых клеток. Случаи тетрасомии (ХХХХ, ХХYY, ХХХY, XYYY) и пентасомии (XXXXX, XXXXY, XXXYY, XXYYY, XYYYY) встречаются редко, некоторые из них описаны всего несколько раз за всю историю медицины. Все эти варианты совместимы с жизнью, и люди часто доживают до преклонных лет, при этом отклонения проявляются в аномальном развитии скелета, дефектах половых органов и снижении умственных способностей. Что характерно, дополнительная Y-хромосома сама по себе влияет на работу организма несильно. Многие мужчины c генотипом XYY даже не узнают о своей особенности. Это связано с тем, что Y-хромосома сильно меньше Х и почти не несет генов, влияющих на жизнеспособность.
У половых хромосом есть и еще одна интересная особенность. Многие мутации генов, расположенных на аутосомах, приводят к отклонениям в работе многих тканей и органов. В то же время большинство мутаций генов на половых хромосомах проявляется только в нарушении умственной деятельности. Получается, что в существенной степени половые хромосомы контролируют развитие мозга. На основании этого некоторые ученые высказывают гипотезу, что именно на них лежит ответственность за различия (впрочем, не до конца подтвержденные) между умственными способностями мужчин и женщин.
Кому выгодно быть неправильным
Несмотря на то что медицина знакома с хромосомными аномалиями давно, в последнее время анеуплоидия продолжает привлекать внимание ученых. Оказалось, что более 80% клеток опухолей содержат необычное количество хромосом. С одной стороны, причиной этому может служить тот факт, что белки, контролирующие качество деления, способны его затормозить. В опухолевых клетках часто мутируют эти самые белки-контролеры, поэтому снимаются ограничения на деление и не работает проверка хромосом. С другой стороны, ученые полагают, что это может служить фактором отбора опухолей на выживаемость. Согласно такой модели, клетки опухоли сначала становятся полиплоидными, а дальше в результате ошибок деления теряют разные хромосомы или их части. Получается целая популяция клеток с большим разнообразием хромосомных аномалий. Большинство из них нежизнеспособны, но некоторые могут случайно оказаться успешными, например если случайно получат дополнительные копии генов, запускающих деление, или потеряют гены, его подавляющие. Однако если дополнительно стимулировать накопление ошибок при делении, то клетки выживать не будут. На этом принципе основано действие таксола — распространенного лекарства от рака: он вызывает системное нерасхождение хромосом в клетках опухоли, которое должно запускать их программируемую гибель.
Получается, что каждый из нас может оказаться носителем лишних хромосом, по крайней мере в отдельных клетках. Однако современная наука продолжает разрабатывать стратегии борьбы с этими нежеланными пассажирами. Одна из них предлагает использовать белки, отвечающие за Х-хромосому, и натравить, например, на лишнюю 21-ю хромосому людей с синдромом Дауна. Сообщается, что на клеточных культурах этот механизм удалось привести в действие. Так что, возможно, в обозримом будущем опасные лишние хромосомы окажутся укрощены и обезврежены.
Теги
ГенетикаРоссийская наукаМедицина
хромосомных наблюдений на трех культурах эпителиальных клеток, полученных из карциномы шейки матки человека23 | JNCI: Журнал Национального института рака
Фильтр поиска панели навигации JNCI: Journal of the National Cancer InstituteЭтот выпускJNCI PortfolioMedicine and HealthBooksJournalsOxford Academic Мобильный телефон Введите поисковый запрос
Закрыть
Фильтр поиска панели навигации JNCI: Journal of the National Cancer InstituteЭтот выпускJNCI PortfolioMedicine and HealthBooksJournalsOxford Academic Введите поисковый запрос
Расширенный поиск
Журнальная статья
Получить доступ
Нелли Ауэрсперг,
Нелли Ауэрсперг
Ищите другие работы этого автора на:
Оксфордский академический
пабмед
Google Scholar
Аннет П. Гаврилюк
Аннет П. Гаврилюк
Ищите другие работы этого автора на:
Оксфордский академический
пабмед
Google Scholar
JNCI: Журнал Национального института рака , том 28, выпуск 3, март 1962 г., страницы 605–627, https://doi.org/10.1093/jnci/28.3.605
Опубликовано:
01 марта 1962
История статьи
Получено:
17 августа 1961 г.
Опубликовано:
01 марта 1962 г.
- Содержание статьи
- Рисунки и таблицы
- видео
- Аудио
- Дополнительные данные
Цитировать
Cite
Nelly Auersperg, Annette P.
 Hawryluk, Наблюдения за хромосомами трех культур эпителиальных клеток, полученных из карциномы шейки матки человека, JNCI: Журнал Национального института рака , том 28, выпуск 3, март 1962 г., страницы 605–627, https://doi.org/10.1093/jnci/28.3.605
Hawryluk, Наблюдения за хромосомами трех культур эпителиальных клеток, полученных из карциномы шейки матки человека, JNCI: Журнал Национального института рака , том 28, выпуск 3, март 1962 г., страницы 605–627, https://doi.org/10.1093/jnci/28.3.605Выберите формат Выберите format.ris (Mendeley, Papers, Zotero).enw (EndNote).bibtex (BibTex).txt (Medlars, RefWorks)
Закрыть
Разрешения
- Электронная почта
- Твиттер
- Фейсбук
- Подробнее
Фильтр поиска панели навигации JNCI: Journal of the National Cancer InstituteЭтот выпускJNCI PortfolioMedicine and HealthBooksJournalsOxford Academic Мобильный телефон Введите поисковый запрос
Закрыть
Фильтр поиска панели навигации JNCI: Journal of the National Cancer InstituteЭтот выпускJNCI PortfolioMedicine and HealthBooksJournalsOxford Academic Введите поисковый запрос
Advanced Search
Резюме
Эпителиальные культуры были выращены из биопсий 3 инвазивных плоскоклеточных карцином шейки матки и наблюдались in vitro на 6, 9 и 10 месяцев соответственно. Три опухоли различались клинически по характеру роста и радиочувствительности, а также гистологически по степени дифференцировки. In vitro каждый из них имел характерный характер роста, особенно в отношении морфологии клеток, когезии и стратификации. При выращивании в целлюлозной губке гистологические признаки напоминали плоскоклеточный рак. Три культуры имели модальные числа хромосом 45, 51 и 120–130 соответственно. Культура с 45 хромосомами имела дополнительный № 3 и дополнительный аномальный крупный метацентрик; один № 2, один № 16–18 и либо один № 6–12 + X, либо один № 13–15 отсутствовали. В культуре с 51 хромосомой отсутствовала одна №3; было 5 дополнительных № 6–12 + X и 1 дополнительный № 19.–20. В культуре с модальным числом 120–130 было много грубо аномальных хромосом и относительный избыток метацентриков. Эти паттерны хромосом были идентичны в нескольких случаях, а также в разных сублиниях в каждой культуре.
Этот контент доступен только в формате PDF.
Раздел выпуска:
Статьи
В настоящее время у вас нет доступа к этой статье.
Скачать все слайды
Войти
Получить помощь с доступом
Получить помощь с доступом
Доступ для учреждений
Доступ к контенту в Oxford Academic часто предоставляется посредством институциональных подписок и покупок. Если вы являетесь членом учреждения с активной учетной записью, вы можете получить доступ к контенту одним из следующих способов:
Доступ на основе IP
Как правило, доступ предоставляется через институциональную сеть к диапазону IP-адресов. Эта аутентификация происходит автоматически, и невозможно выйти из учетной записи с IP-аутентификацией.
Войдите через свое учреждение
Выберите этот вариант, чтобы получить удаленный доступ за пределами вашего учреждения. Технология Shibboleth/Open Athens используется для обеспечения единого входа между веб-сайтом вашего учебного заведения и Oxford Academic.
- Нажмите Войти через свое учреждение.
- Выберите свое учреждение из предоставленного списка, после чего вы перейдете на веб-сайт вашего учреждения для входа.
- Находясь на сайте учреждения, используйте учетные данные, предоставленные вашим учреждением. Не используйте личную учетную запись Oxford Academic.
- После успешного входа вы вернетесь в Oxford Academic.
Если вашего учреждения нет в списке или вы не можете войти на веб-сайт своего учреждения, обратитесь к своему библиотекарю или администратору.
Войти с помощью читательского билета
Введите номер своего читательского билета, чтобы войти в систему. Если вы не можете войти в систему, обратитесь к своему библиотекарю.
Члены общества
Доступ члена общества к журналу достигается одним из следующих способов:
Войти через сайт сообщества
Многие общества предлагают единый вход между веб-сайтом общества и Oxford Academic. Если вы видите «Войти через сайт сообщества» на панели входа в журнале:
- Щелкните Войти через сайт сообщества.
- При посещении сайта общества используйте учетные данные, предоставленные этим обществом. Не используйте личную учетную запись Oxford Academic.
- После успешного входа вы вернетесь в Oxford Academic.
Если у вас нет учетной записи сообщества или вы забыли свое имя пользователя или пароль, обратитесь в свое общество.
Вход через личный кабинет
Некоторые общества используют личные аккаунты Oxford Academic для предоставления доступа своим членам. См. ниже.
Личный кабинет
Личную учетную запись можно использовать для получения оповещений по электронной почте, сохранения результатов поиска, покупки контента и активации подписок.
Некоторые общества используют личные аккаунты Oxford Academic для предоставления доступа своим членам.
Просмотр учетных записей, вошедших в систему
Щелкните значок учетной записи в правом верхнем углу, чтобы:
- Просмотр вашей личной учетной записи и доступ к функциям управления учетной записью.
- Просмотр институциональных учетных записей, предоставляющих доступ.
Выполнен вход, но нет доступа к содержимому
Oxford Academic предлагает широкий ассортимент продукции. Подписка учреждения может не распространяться на контент, к которому вы пытаетесь получить доступ. Если вы считаете, что у вас должен быть доступ к этому контенту, обратитесь к своему библиотекарю.
Ведение счетов организаций
Для библиотекарей и администраторов ваша личная учетная запись также предоставляет доступ к управлению институциональной учетной записью. Здесь вы найдете параметры для просмотра и активации подписок, управления институциональными настройками и параметрами доступа, доступа к статистике использования и т. д.
Покупка
Стоимость подписки и заказ этого журнала
Варианты покупки книг и журналов в Oxford Academic
Кратковременный доступ
Чтобы приобрести краткосрочный доступ, пожалуйста, войдите в свой личный аккаунт выше.
У вас еще нет личного кабинета? регистр
Хромосомные наблюдения на трех культурах эпителиальных клеток, полученных из карциномы шейки матки человека23 — 24-часовой доступ
ЕВРО €36,00
32 фунта стерлингов
39 долларов США.
Реклама
Цитаты
Альтметрика
Дополнительная информация о метриках
Оповещения по электронной почте
Оповещение об активности статьи
Предварительные уведомления о статьях
Оповещение о новой проблеме
Получайте эксклюзивные предложения и обновления от Oxford Academic
Ссылки на статьи по телефону
Последний
Самые читаемые
Самые цитируемые
Линейная связь между дозой радиоактивного йода и риском второго первичного злокачественного новообразования при раке щитовидной железы
Характеристики программ навигации для пациентов в рамках инициативы ACCSIS по скринингу колоректального рака
Cancer MoonshotИспользование менопаузальной гормональной терапии и риск рака яичников во французском когортном исследовании
Курение как прогностический фактор выживаемости больных диссеминированным герминогенным раком
Гистерэктомия, двусторонняя оофорэктомия и риск рака молочной железы в проспективном когортном исследовании с участием представителей разных рас
Ассистент кафедры инфекционных болезней
Нью-Хейвен, Коннектикут
Ассистент кафедры инфекционных болезней
Нью-Хейвен, Коннектикут
Штатный врач — нейроонколог
Луисвилл, Кентукки
Академический специалист по легочной медицине сна в Scenic Central Pennsylvania
Херши, Пенсильвания
Просмотреть все вакансии
Реклама
Leave A Comment