
Рибонуклеиновая кислота — это… Что такое Рибонуклеиновая кислота?
Рибонуклеи́новая кисло́та (РНК) — одна из трёх основных макромолекул (две другие — ДНК и белки), которые содержатся в клетках всех живых организмов.
Так же, как ДНК (дезоксирибонуклеиновая кислота), РНК состоит из длинной цепи, в которой каждое звено называется нуклеотидом. Каждый нуклеотид состоит из азотистого основания, сахара рибозы и фосфатной группы. Последовательность нуклеотидов позволяет РНК кодировать генетическую информацию. Все клеточные организмы используют РНК (мРНК) для программирования синтеза белков.
Клеточные РНК образуются в ходе процесса, называемого транскрипцией, то есть синтеза РНК на матрице ДНК, осуществляемого специальными ферментами — РНК-полимеразами. Затем матричные РНК (мРНК) принимают участие в процессе, называемом трансляцией. Трансляция — это синтез белка на матрице мРНК при участии рибосом. Другие РНК после транскрипции подвергаются химическим модификациям, и после образования вторичной и третичной структур выполняют функции, зависящие от типа РНК.
Для одноцепочечных РНК характерны разнообразные пространственные структуры, в которых часть нуклеотидов одной и той же цепи спарены между собой. Некоторые высокоструктурированные РНК принимают участие в синтезе белка клетки, например, транспортные РНК служат для узнавания кодонов и доставки соответствующих аминокислот к месту синтеза белка, а рибосомные РНК служат структурной и каталитической основой рибосом.
Однако функции РНК в современных клетках не ограничиваются их ролью в трансляции. Так, малые ядерные РНК принимают участие в сплайсинге эукариотических матричных РНК и других процессах.
Помимо того, что молекулы РНК входят в состав некоторых ферментов (например, теломеразы), у отдельных РНК обнаружена собственная ферментативная активность: способность вносить разрывы в другие молекулы РНК или, наоборот, «склеивать» два РНК-фрагмента. Такие РНК называются рибозимами.
Геномы ряда вирусов состоят из РНК, то есть у них она играет роль, которую у высших организмов выполняет ДНК. На основании разнообразия функций РНК в клетке была выдвинута гипотеза, согласно которой РНК — первая молекула, которая была способна к самовоспроизведению в добиологических системах.
История изучения
Нуклеиновые кислоты были открыты в 1868 году швейцарским учёным Иоганном Фридрихом Мишером, который назвал эти вещества «нуклеин», поскольку они были обнаружены в ядре (лат. nucleus)
Химический состав и модификации мономеров
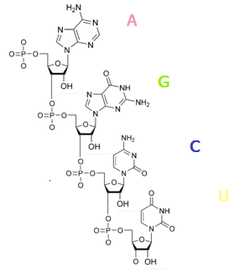 Химическое строение полинуклеотида РНК
Химическое строение полинуклеотида РНКНуклеотиды РНК состоят из сахара — рибозы, к которой в положении 1′ присоединено одно из оснований: аденин, гуанин, цитозин или урацил. Фосфатная группа соединяет рибозы в цепочку, образуя связи с 3′ атомом углерода одной рибозы и в 5′ положении другой. Фосфатные группы при физиологическом рН отрицательно заряжены, поэтому РНК — полианион. РНК транскрибируется как полимер четырёх оснований (аденина (A), гуанина (G), урацила (U) и цитозина (C), но в «зрелой» РНК есть много модифицированных оснований и сахаров
У псевдоуридина (Ψ) связь между урацилом и рибозой не C — N, а C — C, этот нуклеотид встречается в разных положениях в молекулах РНК. В частности, псевдоуридин важен для функционирования тРНК
Роль многих других модификаций не до конца изучена, но в рибосомальной РНК многие пост-транскрипционные модификации находятся в важных для функционирования рибосомы участках. Например, на одном из рибонуклеотидов, участвующем в образовании пептидной связи[14].
Структура
Азотистые основания в составе РНК могут образовывать водородные связи между цитозином и гуанином, аденином и урацилом, а также между гуанином и урацилом
Важная структурная особенность РНК, отличающая её от ДНК — наличие гидроксильной группы в 2′ положении рибозы, которая позволяет молекуле РНК существовать в А, а не В-конформации, наиболее часто наблюдаемой у ДНК[17]. У А-формы глубокая и узкая большая бороздка и неглубокая и широкая малая бороздка
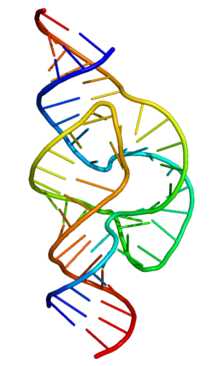
«Рабочая» форма одноцепочечной молекулы РНК, как и у белков, часто обладает третичной структурой. Третичная структура образуется на основе элементов вторичной структуры, образуемой с помощью водородных связей внутри одной молекулы. Различают несколько типов элементов вторичной структуры — стебель-петли, петли и псевдоузлы[20]. В силу большого числа возможных вариантов спаривания оснований предсказание вторичной структуры РНК — гораздо более сложная задача, чем предсказание вторичной структуры белков, но в настоящее время есть эффективные программы, например, mfold
Примером зависимости функции молекул РНК от их вторичной структуры являются участки внутренней посадки рибосомы (IRES). IRES — структура на 5′ конце информационной РНК, которая обеспечивает присоединение рибосомы в обход обычного механизма инициации синтеза белка, требующего наличия особого модифицированного основания (кэпа) на 5′ конце и белковых факторов инициации. Первоначально IRES были обнаружены в вирусных РНК, но сейчас накапливается всё больше данных о том, что клеточные мРНК также используют IRES-зависимый механизм инициации в условиях стресса
Многие типы РНК, например, рРНК и мяРНК в клетке функционируют в виде комплексов с белками, которые ассоциируют с молекулами РНК после их синтеза или (у эукариот) экспорта из ядра в цитоплазму. Такие РНК-белковые комплексы называются рибонуклеопротеиновыми комплексами или рибонуклеопротеидами.
Сравнение с ДНК
Между ДНК и РНК есть три основных отличия:
- ДНК содержит сахар дезоксирибозу, РНК — рибозу, у которой есть дополнительная, по сравнению с дезоксирибозой, гидроксильная группа. Эта группа увеличивает вероятность гидролиза молекулы, то есть уменьшает стабильность молекулы РНК.
- Нуклеотид, комплементарный аденину, в РНК не тимин, как в ДНК, а урацил — неметилированная форма тимина.
- ДНК существует в форме двойной спирали, состоящей из двух отдельных молекул. Молекулы РНК, в среднем, гораздо короче и преимущественно одноцепочечные.
Структурный анализ биологически активных молекул РНК, включая тРНК, рРНК, мяРНК и другие молекулы, которые не кодируют белков, показал, что они состоят не из одной длинной спирали, а из многочисленных коротких спиралей, расположенных близко друг к другу и образующих нечто, похожее на третичную структуру белка. В результате этого РНК может катализировать химические реакции, например, пептидил-трансферазный центр рибосомы, участвующий в образовании пептидной связи белков, полностью состоит из РНК[23][24].
Синтез
Синтез РНК в живой клетке проводится ферментом — РНК-полимеразой. У эукариот разные типы РНК синтезируются разными, специализированными РНК-полимеразами. В целом матрицей синтеза РНК может выступать как ДНК, так и другая молекула РНК. Например, полиовирусы используют РНК-зависимую РНК-полимеразу для репликации своего генетического материала, состоящего из РНК[25]. Но РНК-зависимый синтез РНК, который раньше считался характерным только для вирусов, происходит и в клеточных организмах, в процессе так называемой РНК-интерференции[26].
Как в случае ДНК-зависимой РНК-полимеразы, так и в случае РНК-зависимой РНК-полимеразы фермент присоединяется к промоторной последовательности. Вторичная структура молекулы матрицы расплетается с помощью хеликазной активности полимеразы, которая при движении субстрата в направлении от 3′ к 5′ концу молекулы синтезирует РНК в направлении 5′ → 3′. Терминатор транскрипции в исходной молекуле определяет окончание синтеза. Многие молекулы РНК синтезируются в качестве молекул-предшественников, которые подвергаются «редактированию» — удалению ненужных частей с помощью РНК-белковых комплексов[27].
Например, у кишечной палочки гены рРНК расположены в составе одного оперона (в rrnB порядок расположения такой: 16S — tRNAGlu2 — 23S —5S) считываются в виде одной длинной молекулы, которая затем подвергается расщеплению в нескольких участках с образованием сначала пре-рРНК, а затем зрелых молекул рРНК[28]. Процесс изменения нуклеотидной последовательности РНК после синтеза носит название процессинга или редактирования РНК.
После завершения транскрипции РНК часто подвергается модификациям (см. выше), которые зависят от функции, выполняемой данной молекулой. У эукариот процесс «созревания» РНК, то есть её подготовки к синтезу белка, часто включает сплайсинг: удаление некодирующих белок последовательностей (интронов) с помощью рибонуклеопротеида сплайсосомы. Затем к 5′ концу молекулы пре-мРНК эукариот добавляется особый модифицированный нуклеотид (кэп), а к 3′ концу несколько аденинов, так назваемый «полиА-хвост»[27].
Типы РНК
 Структура молоточкового (hammerhead) рибозима, который расщепляет РНК
Структура молоточкового (hammerhead) рибозима, который расщепляет РНКМатричная (информационная) РНК — РНК, которая служит посредником при передаче информации, закодированной в ДНК к рибосомам, молекулярным машинам, синтезирующим белки живого организма. Кодирующая последовательность мРНК определяет последовательность аминокислот полипептидной цепи белка[29]. Однако подавляющее большинство РНК не кодируют белок. Эти некодирующие РНК могут транскрибироваться с отдельных генов (например, рибосомальные РНК) или быть производными интронов[30]. Классические, хорошо изученные типы некодирующих РНК — это транспортные РНК (тРНК) и рРНК, которые участвуют в процессе трансляции[31]. Существуют также классы РНК, ответственные за регуляцию генов, процессинг мРНК и другие роли. Кроме того, есть и молекулы некодирующих РНК, способные катализировать химические реакции, такие, как разрезание и лигирование молекул РНК[32]. По аналогии с белками, способными катализировать химические реакции — энзимами (ферментами), каталитические молекулы РНК называются рибозимами.
Участвующие в трансляции
Роль разных типов РНК в синтезе белка (по Уотсону)Информация о последовательности аминокислот белка содержится в мРНК. Три последовательных нуклеотида (кодон) соответствуют одной аминокислоте. В эукариотических клетках транскирибированный предшественник мРНК или пре-мРНК процессируется с образованием зрелой мРНК. Процессинг включает удаление некодирующих белок последовательностей (интронов). После этого мРНК экспортируется из ядра в цитоплазму, где к ней присоединяются рибосомы, транслирующие мРНК с помощью соединённых с аминокислотами тРНК.
В безъядерных клетках (бактерии и археи) рибосомы могут присоединяться к мРНК сразу после транскрипции участка РНК. И у эукариот, и у прокариот цикл жизни мРНК завершается её контролируемым разрушением ферментами рибонуклеазами[29].
Транспортные (тРНК) — малые, состоящие из приблизительно 80 нуклеотидов, молекулы с консервативной третичной структурой. Они переносят специфические аминокислоты в место синтеза пептидной связи в рибосоме. Каждая тРНК содержит участок для присоединения аминокислоты и антикодон для узнавания и присоединения к кодонам мРНК. Антикодон образует водородные связи с кодоном, что помещает тРНК в положение, способствующее образованию пептидной связи между последней аминокислотой образованного пептида и аминокислотой, присоединённой к тРНК[30].
Рибосомальные РНК (рРНК) — каталитическая составляющая рибосом. Эукариотические рибосомы содержат четыре типа молекул рРНК: 18S, 5.8S, 28S и 5S. Три из четырёх типов рРНК синтезируются в ядрышке. В цитоплазме рибосомальные РНК соединяются с рибосомальными белками и формируют нуклеопротеин, называемый рибосомой[29]. Рибосома присоединяется к мРНК и синтезирует белок. рРНК составляет до 80 % РНК, обнаруживаемой в цитоплазме эукариотической клетки[33].
Необычный тип РНК, который действует в качестве тРНК и мРНК (тмРНК) обнаружен во многих бактериях и пластидах. При остановке рибосомы на дефектных мРНК без стоп-кодонов тмРНК присоединяет небольшой пептид, направляющий белок на деградацию[34].
Участвующие в регуляции генов
В живых клетках обнаружено несколько типов РНК, которые могут уменьшать степень выражения гена при комплементарности мРНК или самому гену. Микро-РНК (21-22 нуклеотида в длину) найдены у эукариот и оказывают воздействие через механизм РНК-интерференции. При этом комплекс микро-РНК и ферментов может приводить к метилированию нуклеотидов в ДНК промотора гена, что служит сигналом для уменьшения активности гена. При использовании другого типа регуляции мРНК, комплементарная микро-РНК, деградируется[35]. Однако есть и миРНК, которые увеличивают, а не уменьшают экспрессию генов[36]. Малые интерферирующие РНК (миРНК, 20-25 нуклеотидов) часто образуются в результате расщепления вирусных РНК, но существуют и эндогенные клеточные миРНК[37]. Малые интерферирующие РНК также действуют через РНК-интерференцию по сходным с микро-РНК механизмам[38]. У животных найдены так называемыме РНК, взаимодействующие с Piwi (piRNA, 29-30 нуклеотидов), действующие в половых клетках против транспозиции и играющие роль в образовании гамет[39][40]. Кроме того, piRNA могут эпигенетически наследоваться по материнской линии, передавая потомству своё свойство ингибировать экспрессию транспозонов[41].
Антисмысловые РНК широко распространены у бактерий, многие из них подавляют выражение генов, но некоторые активируют экспрессию[42]. Действуют антисмысловые РНК, присоединяясь к мРНК, что приводит к образованию двуцепочечных молекул РНК, которые деградируются ферментами.[43]. У эукариот обнаружены высокомолекулярные, мРНК-подобные молекулы РНК. Эти молекулы также регулируют выражение генов[44]. В качестве примера можно привести Xist, присоединяющуюся и инактивирующую одну из двух Х-хромосом у самок млекопитающих[45].
Кроме роли отдельных молекул в регуляции генов, регуляторные элементы могут формироваться в 5′ и 3′ нетранслируемых участках мРНК. Эти элементы могут действовать самостоятельно, предотвращая инициацию трансляции, либо присоединять белки, например, ферритин или малые молекулы, например, биотин[46].
В процессинге РНК
Многие РНК принимают участие в модификации других РНК. Интроны вырезаются из пре-мРНК сплайсосомами, которые, кроме белков, содержат несколько малых ядерных РНК (мяРНК)[31]. Кроме того, интроны могут катализировать собственное вырезание[47]. Синтезированая в результате транскрипции РНК также может быть химически модифицирована. У эукариот химические модификации нуклеотидов РНК, например, их метилирование, выполняется малыми ядерными РНК (мяРНК, 60-300 нуклеотидов). Этот тип РНК локализуется в ядрышко и тельцах Кахаля[30]. После ассоциации мяРНК с ферментами, мяРНК связываются с РНК-мишенью путём образования пар между основаниями двух молекул, а ферменты модифицируют нуклеотиды РНК-мишени. Рибосомальные и транспортные РНК содержат много подобных модификаций, конкретное положение которых часто сохраняется в процессе эволюции. Также могут быть модифицированы мяРНК и сами мяРНК[48][49]. Гидовые РНК осуществляют процесс редактирования РНК в кинетопласте — особом участке митохондрии протистов-кинетопластид (например, трипаносом).
Геномы, состоящие из РНК
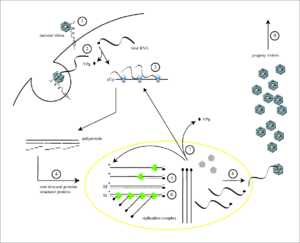 Жизненный цикл вируса с РНК геномом на примере полиовируса: 1 — присоединение исходного вириона к рецептору; 2 — вирион попадает в клетку; 3 — трансляция белков вируса с его РНК с образованием полипетида; 4 — полимеразы вируса размножают его РНК
Жизненный цикл вируса с РНК геномом на примере полиовируса: 1 — присоединение исходного вириона к рецептору; 2 — вирион попадает в клетку; 3 — трансляция белков вируса с его РНК с образованием полипетида; 4 — полимеразы вируса размножают его РНККак и ДНК, РНК может хранить информацию о биологических процессах. РНК может использоваться в качестве генома вирусов и вирусоподобных частиц. РНК-геномы можно разделить на те, которые не имеют промежуточной стадии ДНК и те, которые для размножения копируются в ДНК-копию и обратно в РНК (ретровирусы).
РНК-содержащие вирусы
Многие вирусы, например, вирус гриппа, на всех стадиях содержат геном, состоящий исключительно из РНК. РНК содержится внутри обычно белковой оболочки и реплицируется с помощью закодированных в ней РНК-зависимых РНК-полимераз. Вирусные геномы, состоящие из РНК разделяются на
- содержащие «плюс-цепь РНК», которая используется в качестве и мРНК, и генома;
- «минус-цепь РНК», которая служит только геномом, а в качестве мРНК используется комплементарная ей молекула;
- двухцепоченые вирусы.
Вироиды — другая группа патогенов, содержащих РНК-геном и не содержащих белок. Они реплицируются РНК-полимеразами организма хозяина[50].
Ретровирусы и ретротранспозоны
У других вирусов РНК-геном есть в течение только одной из фаз жизненного цикла. Вирионы так называемых ретровирусов содержат молекулы РНК, которые при попадании в клетки хозяина служат матрицей для синтеза ДНК-копии. В свою очередь, с матрицы ДНК считывается РНК-геном. Кроме вирусов обратную транскрипции применяют и класс мобильных элементов генома — ретротранспозоны[51].
Гипотеза РНК-мира
Способность молекул РНК одновременно служить как в качестве носителя информации, так и в качестве катализатора химических реакций, позволила выдвинуть гипотезу о том, что РНК была первым сложным полимером, появившимся в процессе добиологической эволюции. Эта гипотеза названа «гипотеза РНК-мира»[52][53]. Согласно ей, РНК на первых этапах эволюции автокатализировала синтез других молекул РНК, а затем и ДНК. На втором этапе эволюции синтезированные молекулы ДНК, как более стабильные, стали хранилищем генетической информации. Синтез белка на матрице РНК с помощью пра-рибосом, полностью состоящих из РНК, расширил свойства добиологических систем, постепенно белок заменил РНК в структурных аспектах. Из этой гипотезы делается вывод, что многие РНК, принимающие участие в синтезе белка в современных клетках, в особенности рРНК и тРНК — это реликты РНК-мира.
См. также
Примечания
- ↑ Dahm R (2005). «Friedrich Miescher and the discovery of DNA». Developmental Biology 278 (2): 274–88. PMID 15680349.
- ↑ Nierhaus KH, Wilson DN. Protein Synthesis and Ribosome Structure. — Wiley-VCH, 2004. — С. 3. — ISBN 3-527-30638-2
- ↑ Carlier M L’ADN, cette «simple» molécule. Esprit libre (июнь 2003). Архивировано из первоисточника 23 августа 2011. Проверено ???.
- ↑ А. С. Спирин. Биоорганическая химия. — М.: Высшая школа, 1986. — С. 10.
- ↑ Ochoa S. Enzymatic synthesis of ribonucleic acid. Nobel Lecture (1959). Архивировано из первоисточника 23 августа 2011. Проверено ???.
- ↑ Holley RW et al. Structure of a ribonucleic acid // Science. — 1965. — Vol. 147. — № 1664. — P. 1462–65. — DOI:10.1126/science.147.3664.1462
- ↑ Szathmáry E. The origin of the genetic code: amino acids as cofactors in an RNA world // Trends Genet.. — 1999. — Vol. 15. — № 6. — P. 223–9. — DOI:10.1016/S0168-9525(99)01730-8
- ↑ Fiers W et al. Complete nucleotide-sequence of bacteriophage MS2-RNA: primary and secondary structure of replicase gene // Nature. — 1976. — Vol. 260. — P. 500–7. — PMID 1264203.
- ↑ Napoli C, Lemieux C, Jorgensen R. Introduction of a chimeric chalcone synthase gene into petunia results in reversible co-suppression of homologous genes in trans. // Plant Cell. — 1990. — Vol. 2. — № 4. — P. 279–89. — PMID 12354959.
- ↑ Ruvkun G. Glimpses of a tiny RNA world // Science. — 2001. — Vol. 294. — № 5543. — P. 797–99. — DOI:10.1126/science.1066315
- ↑ Jankowski JAZ, Polak JM Clinical gene analysis and manipulation: tools, techniques and troubleshooting. — Cambridge University Press. — P. 14. — ISBN 0521478960
- ↑ Kiss T (2001). «Small nucleolar RNA-guided post-transcriptional modification of cellular RNAs». The EMBO Journal 20: 3617–22. DOI:10.1093/emboj/20.14.3617.
- ↑ Yu Q, Morrow CD (2001). «Identification of critical elements in the tRNA acceptor stem and TΨC loop necessary for human immunodeficiency virus type 1 infectivity». J Virol. 75 (10): 4902–6. DOI:10.1128/JVI.75.10.4902-4906.2001.
- ↑ King TH, Liu B, McCully RR, Fournier MJ (2002). «Ribosome structure and activity are altered in cells lacking snoRNPs that form pseudouridines in the peptidyl transferase center». Molecular Cell 11 (2): 425–35. DOI:10.1016/S1097-2765(03)00040-6.
- ↑ Barciszewski J, Frederic B, Clark C RNA biochemistry and biotechnology. — Springer. — P. 73–87. — ISBN 0792358627
- ↑ Lee JC, Gutell RR (2004). «Diversity of base-pair conformations and their occurrence in rRNA structure and RNA structural motifs». J. Mol. Biol. 344 (5): 1225–49. DOI:10.1016/j.jmb.2004.09.072. PMID 15561141.
- ↑ Salazar M, Fedoroff OY, Miller JM, Ribeiro NS, Reid BR (1992). «The DNA strand in DNAoRNA hybrid duplexes is neither B-form nor A-form in solution». Biochemistry 1993 (32): 4207–15. PMID 7682844.
- ↑ Hermann T, Patel DJ (2000). «RNA bulges as architectural and recognition motifs». Structure 8 (3): R47–R54. DOI:10.1016/S0969-2126(00)00110-6.
- ↑ Mikkola S, Nurmi K, Yousefi-Salakdeh E, Strömberg R, Lönnberg H (1999). «The mechanism of the metal ion promoted cleavage of RNA phosphodiester bonds involves a general acid catalysis by the metal aquo ion on the departure of the leaving group». Perkin transactions 2: 1619–26. DOI:10.1039/a903691a.
- ↑ Mathews DH, Disney MD, Childs JL, Schroeder SJ, Zuker M, Turner DH (2004). «Incorporating chemical modification constraints into a dynamic programming algorithm for prediction of RNA secondary structure». Proc. Natl. Acad. Sci. USA 101 (19): 7287–92. DOI:10.1073/pnas.0401799101.
- ↑ Redirect
- ↑ Spriggs KA, Stoneley M, Bushell M, Willis AE. (2008). «Re-programming of translation following cell stress allows IRES-mediated translation to predominate». Biol Cell. 100 (1): 27–38.
- ↑ Higgs PG (2000). «RNA secondary structure: physical and computational aspects». Quarterly Reviews of Biophysics 33: 199–253. DOI:10.1017/S0033583500003620.
- ↑ Nissen P, Hansen J, Ban N, Moore PB, Steitz TA (2000). «The structural basis of ribosome activity in peptide bond synthesis». Science 289 (5481): 920–30. DOI:10.1126/science.289.5481.920.
- ↑ Jeffrey L Hansen, Alexander M Long, Steve C Schultz (1997). «Structure of the RNA-dependent RNA polymerase of poliovirus». Structure 5 (8): 1109–22. DOI:10.1016/S0969-2126(97)00261-X.
- ↑ Ahlquist P (2002). «RNA-Dependent RNA Polymerases, Viruses, and RNA Silencing». Science 296 (5571): 1270–73. DOI:10.1126/science.1069132.
- ↑ 1 2 Alberts Bruce Molecular Biology of the Cell; Fourth Edition. — New York and London: Garland Science. — P. 302–303. — ISBN ISBN 0-8153-3218-1
- ↑ Wagner R., Theissen G., Zacharias Regulation of Ribosomal RNA synthesis and Control of ribosome Formation in E.coli. — 1993. — P. 119–129.
- ↑ 1 2 3 Cooper GC, Hausman RE The Cell: A Molecular Approach. — 3rd edition. — Sinauer. — P. 261–76, 297, 339–44. — ISBN 0-87893-214-3
- ↑ 1 2 3 Wirta W Mining the transcriptome – methods and applications. — ISBN 91-7178-436-5
- ↑ 1 2 Berg JM, Tymoczko JL, Stryer L Biochemistry. — 5th edition. — WH Freeman and Company, 2002. — P. 118–19, 781–808. — ISBN 0-7167-4684-0
- ↑ Rossi JJ (2004). «Ribozyme diagnostics comes of age». Chemistry & Biology 11 (7): 894–95. DOI:10.1016/j.chembiol.2004.07.002.
- ↑ Kampers T, Friedhoff P, Biernat J, Mandelkow E-M, Mandelkow E (1996). «RNA stimulates aggregation of microtubule-associated protein tau into Alzheimer-like paired helical filaments». FEBS Letters 399: 98–100, 344–49. PMID 8985176.
- ↑ Gueneau de Novoa P, Williams KP (2004). «The tmRNA website: reductive evolution of tmRNA in plastids and other endosymbionts». Nucleic Acids Res. 32 (Database issue): D104-8. DOI:10.1093/nar/gkh202. PMID 14681369.
- ↑ Matzke MA, Matzke AJM (2004). «Planting the seeds of a new paradigm». PLoS Biology 2 (5): e133. DOI:10.1371/journal.pbio.0020133. PMID 15138502.
- ↑ Check E (2007). «RNA interference: hitting the on switch». Nature 448 (7156): 855–58. DOI:10.1038/448855a. PMID 17713502.
- ↑ Vazquez F, Vaucheret H, Rajagopalan R, Lepers C, Gasciolli V, Mallory AC, Hilbert J, Bartel DP, Crété P (2004). «Endogenous trans-acting siRNAs regulate the accumulation of Arabidopsis mRNAs». Molecular Cell 16 (1): 69–79. DOI:10.1016/j.molcel.2004.09.028. PMID 15469823.
- ↑ Doran G (2007). «RNAi – Is one suffix sufficient?». Journal of RNAi and Gene Silencing 3 (1): 217–19.
- ↑ name=fruitfly_piRNA>Horwich MD, Li C Matranga C, Vagin V, Farley G, Wang P, Zamore PD (2007). «The Drosophila RNA methyltransferase, DmHen1, modifies germline piRNAs and single-stranded siRNAs in RISC». Current Biology 17: 1265–72. DOI:10.1016/j.cub.2007.06.030. PMID 17604629.
- ↑ Girard A, Sachidanandam R, Hannon GJ, Carmell MA (2006). «A germline-specific class of small RNAs binds mammalian Piwi proteins». Nature 442: 199–202. DOI:10.1038/nature04917. PMID 16751776.
- ↑ Brennecke J, Malone CD, Aravin AA, Sachidanandam R, Stark A, Hannon GJ (November 2008). «An epigenetic role for maternally inherited piRNAs in transposon silencing». Science (journal) 322 (5906): 1387–92. DOI:10.1126/science.1165171. PMID 19039138.
- ↑ Wagner EG, Altuvia S, Romby P (2002). «Antisense RNAs in bacteria and their genetic elements». Adv Genet. 46: 361–98. PMID 11931231.
- ↑ Gilbert SF Developmental Biology. — 7th ed. — Sinauer, 2003. — P. 101–3. — ISBN 0878932585
- ↑ Hüttenhofer A, Schattner P, Polacek N (2005). «Non-coding RNAs: hope or hype?». Trends Genet. 21 (5): 289–97. DOI:10.1016/j.tig.2005.03.007. PMID 15851066.
- ↑ Heard E, Mongelard F, Arnaud D, Chureau C, Vourc’h C, Avner P (1999). «Human XIST yeast artificial chromosome transgenes show partial X inactivation center function in mouse embryonic stem cells». Proc. Natl. Acad. Sci. USA 96 (12): 6841–46. DOI:10.1073/pnas.96.12.6841. PMID 10359800.
- ↑ Batey RT (2006). «Structures of regulatory elements in mRNAs». Curr. Opin. Struct. Biol. 16 (3): 299–306. DOI:10.1016/j.sbi.2006.05.001. PMID 16707260.
- ↑ Steitz TA, Steitz JA (1993). «A general two-metal-ion mechanism for catalytic RNA». Proc. Natl. Acad. Sci. U.S.A. 90 (14): 6498–502. DOI:10.1073/pnas.90.14.6498. PMID 8341661.
- ↑ Covello PS, Gray MW (1989). «RNA editing in plant mitochondria». Nature 341: 662–66. DOI:10.1038/341662a0. PMID 2552326.
- ↑ Omer AD, Ziesche S, Decatur WA, Fournier MJ, Dennis PP (2003). «RNA-modifying machines in archaea». Molecular Microbiology 48 (3): 617–29. DOI:10.1046/j.1365-2958.2003.03483.x. PMID 12694609.
- ↑ Daròs JA, Elena SF, Flores R (2006). «Viroids: an Ariadne’s thread into the RNA labyrinth». EMBO Rep. 7 (6): 593–8. DOI:10.1038/sj.embor.7400706. PMID 16741503.
- ↑ Kalendar R, Vicient CM, Peleg O, Anamthawat-Jonsson K, Bolshoy A, Schulman AH (2004). «Large retrotransposon derivatives: abundant, conserved but nonautonomous retroelements of barley and related genomes». Genetics 166 (3): D339. DOI:10.1534/genetics.166.3.1437. PMID 15082561.
- ↑ Gilbert, Walter (Feb 1986). «The RNA World». Nature 319: 618. DOI:10.1038/319618a0.
- ↑ Woese Carl The Genetic Code. — Harper & Row, 1968. — ISBN 978-0060471767
Литература
Ссылки
dic.academic.ru
Рибонуклеиновые кислоты — это… Что такое Рибонуклеиновые кислоты?
Рибонуклеи́новые кисло́ты (РНК) — нуклеиновые кислоты, полимеры нуклеотидов, в состав которых входят остаток ортофосфорной кислоты, рибоза (в отличие от ДНК, содержащей дезоксирибозу) и азотистые основания — аденин, цитозин, гуанин и урацил (в отличие от ДНК, содержащей вместо урацила тимин). Эти молекулы содержатся в клетках всех живых организмов, а также в некоторых вирусах.
Клеточные РНК образуются в ходе процесса, называемого транскрипцией, то есть синтеза РНК на матрице ДНК, осуществляемого специальными ферментами — РНК-полимеразами. Затем матричные РНК (мРНК) подвергаются сплайсингу и принимают участие в процессе, называемом трансляцией. Трансляция — это синтез белка на матрице мРНК при участии рибосом. Другие РНК после транскрипции подвергаются химическим модификациям, и после образования вторичной и третичной структур выполняют функции, зависящие от типа РНК.
Для одноцепочечных РНК характерны разнообразные пространственные структуры, в которых часть нуклеотидов одной и той же цепи спарены между собой. Некоторые высокоструктурированные РНК принимают участие в синтезе белка клетки, например, транспортные РНК служат для узнавания кодонов и доставки соответствующих аминокислот к месту синтеза белка, а рибосомные РНК служат структурной и каталитической основой рибосом.
Однако функции РНК в современных клетках не ограничиваются их ролью в трансляции. Так малые ядерные РНК принимают участие в сплайсинге эукариотических матричных РНК и других процессах.
Помимо того, что молекулы РНК входят в состав некоторых ферментов (например, теломеразы) у отдельных РНК обнаружена собственная энзиматическая активность, способность вносить разрывы в другие молекулы РНК или, наоборот, «склеивать» два РНК-фрагмента. Такие РНК называются рибозимами.
Геномы ряда вирусов состоят из РНК, то есть у них она играет роль, которую у высших организмов выполняет ДНК. На основании разнообразия функций РНК в клетке была выдвинута гипотеза, согласно которой РНК — первая молекула, которая была способна к самовоспроизведению в добиологических системах.
История изучения
Нуклеиновые кислоты были открыты в 1868 году швейцарским учёным Иоганном Фридрихом Мишером, который назвал эти вещества «нуклеин», поскольку они были обнаружены в ядре (лат. nucleus)[1]. Позже было обнаружено, что бактериальные клетки, в которых нет ядра, тоже содержат нуклеиновые кислоты. Значение РНК в синтезе белков было предположено в 1939 году в работе Торбьёрна Оскара Касперссона, Жана Брачета и Джека Шульца [2]. Джерард Маирбакс выделил первую матричную РНК, кодирующую гемоглобин кролика и показал, что при её введении в ооциты образуется тот же самый белок[3]. В Советском Союзе в 1956-57 годах проводились работы (А. Белозёрский, А. Спирин, Э. Волкин, Ф. Астрахан) по определению состава РНК клеток, которые привели к выводу, что основную массу РНК в клетке составляет рибосомальная РНК.[4] Северо Очоа получил Нобелевскую премию по медицине в 1959 году за открытие механизма синтеза РНК[5]. Последовательность 77 нуклеотидов одной из тРНК дрожжей S. cerevisiae была определена в 1965 году в лаборатории Роберта Холея, за что в 1968 году он получил Нобелевскую премию по медицине [6]. В 1967 Карл Вёзе предположил, что РНК обладают каталитическими свойствами. Он выдвинул так называемую Гипотезу РНК-мира, в котором РНК прото-организмов служила и в качестве молекулы хранения информации (сейчас эта роль выполняется в основном ДНК) и молекулы, которая катализировала метаболические реакции (сейчас это делают в основном ферменты)[7]. В 1976 Уолтер Фаэрс и его группа в Гентском Университете (Голландия) определили первую последовательность генома РНК-содержащего вируса, бактериофага MS2[8]. В начале 1990-х было обнаружено, что введение чужеродных генов в геном растений приводит к подавлению выражения аналогичных генов растения[9]. Приблизительно в это же время было показано, что РНК длиной около 22 оснований, которые сейчас называются микроРНК, играют регуляторную роль в онтогенезе нематод C.elegans [10].
Химический состав и модификации мономеров
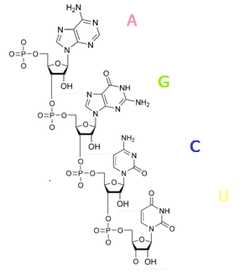 Химическое строение полинуклеотида РНК
Химическое строение полинуклеотида РНКНуклеотиды РНК состоят из сахара — рибозы, к которой в положении 1′ присоединено одно из оснований: аденин, гуанин, цитозин или урацил. Фосфатная группа соединяет рибозы в цепочку, образуя связи с 3′ атомом углерода одной рибозы и в 5′ положении другой. Фосфатные группы при физиологическом рН отрицательно заряжены, поэтому РНК — полианион. РНК транскрибируется как полимер четырёх оснований (аденина (A), гуанина (G), урацила (U) и цитозина (C), но в «зрелой» РНК есть много модифицированных оснований и сахаров[11]. Всего в РНК насчитывается около 100 разных видов модифицированных нуклеозидов, из которых 2′-О-метилрибоза наиболее частая модификация сахара, а псевдоуридин — наиболее часто встречающееся модифицированное основание [12]. У псевдоуридина (Ψ) связь между урацилом и рибозой не C — N, а C — C, этот нуклеотид встречается в разных положениях в молекулах РНК. В частности, псевдоуридин важен для функционирования тРНК[13]. Другое заслуживающее внимания модифицированное основание — гипоксантин, деаминированный гуанин, нуклеозид которого носит название инозина. Инозин играет важную роль в обеспечении вырожденности генетического кода. Роль многих других модификаций не до конца изучена, но в рибосомальной РНК многие пост-транскрипционные модификации находятся в важных для функционирования рибосомы участках. Например, на одном из рибонуклеотидов, участвующим в образовании пептидной связи[14].
Структура
Азотистые основания в составе РНК могут образовывать водородные связи между цитозином и гуанином, аденином и урацилом, а также между гуанином и урацилом [15]. Однако возможны и другие взаимодействия, например, несколько аденинов могут образовывать петлю, или петля, состоящая из четырёх нуклеотидов, в которой есть пара оснований аденин — гуанин[16].
Разные формы нуклеиновых кислот. На рисунке (слева направо) представлены A (типична для РНК), B (ДНК) и Z (редкая форма ДНК)Важная структурная особенность РНК, отличающая её от ДНК — наличие гидроксильной группы в 2′ положении рибозы, которая позволяет молекуле РНК существовать в А, а не В-конформации, наиболее часто наблюдаемой у ДНК[17]. У А-формы глубокая и узкая большая бороздка и неглубокая и широкая малая бороздка[18]. Второе последствие наличия 2′ гидроксильной группы состоит в том, что конформационно пластичные, то есть не принимающие участие в образовании двойной спирали, участки молекулы РНК могут химически атаковать другие фосфатные связи и их расщеплять [19].
«Рабочая» форма одноцепочечной молекулы РНК, как и у белков, часто обладает третичной структурой. Третичная структура образуется на основе элементов вторичной структуры, образуемой с помощью водородных связей внутри одной молекулы. Различают несколько типов элементов вторичной структуры — стебель-петли, петли и псевдоузлы[20]. В силу большого числа возможных вариантов спаривания оснований предсказание вторичной структуры РНК — гораздо более сложная задача, чем предсказание вторичной структуры белков, но в настоящее время есть эффективные программы, например, mfold [21].
Примером зависимости функции молекул РНК от их вторичной структуры являются участки внутренней посадки рибосомы (IRES). IRES — структура на 5′ конце информационной РНК, которая обеспечивает присоединение рибосомы в обход обычного механизма инициации синтеза белка, требующего наличия особого модифицированного основания (кэпа) на 5′ конце и белковых факторов инициации. Первоначально IRES были обнаружены в вирусных РНК, но сейчас накапливается всё больше данных о том, что клеточные мРНК также используют IRES-зависимый механизм инициации в условиях стресса[22].
Многие типы РНК, например, рРНК и мяРНК в клетке функционируют в виде комплексов с белками, которые ассоциируют с молекулами РНК после их синтеза или (у эукариот) экспорта из ядра в цитоплазму. Такие РНК-белковые комплексы называются рибонуклеопротеиновыми комплексами или рибонуклеопротеидами.
Сравнение с ДНК
Между ДНК и РНК есть три основных отличия:
- ДНК содержит сахар дезоксирибозу, РНК — рибозу, у которой есть дополнительная, по сравнению с дезоксирибозой, гидроксильная группа. Эта группа увеличивает вероятность гидролиза молекулы, то есть уменьшает стабильность молекулы РНК.
- Нуклеотид, комплементарный аденину, в РНК не тимин, как в ДНК, а урацил — неметилированная форма тимина.
- ДНК существует в форме двойной спирали, состоящей из двух отдельных молекул. Молекулы РНК, в среднем, гораздо короче и преимущественно одноцепочечные.
Структурный анализ биологически активных молекул РНК, включая тРНК, рРНК, мяРНК и другие молекулы, которые не кодируют белков, показал, что они состоят не из одной длинной спирали, а из многочисленных коротких спиралей, расположенных близко друг к другу и образующих нечто, похожее на третичную структуру белка. В результате этого РНК может катализировать химические реакции, например, пептидил-трансферазный центр рибосомы, участвующий в образовании пептидной связи белков, полностью состоит из РНК [23][24].
Синтез
Синтез РНК в живой клетке проводится ферментом — РНК-полимеразой. У эукариот разные типы РНК синтезируются разными, специализированными РНК-полимеразами. В целом матрицей синтеза РНК может выступать как ДНК, так и другая молекула РНК. Например, полиовирусы используют РНК-зависимую РНК-полимеразу для репликации своего генетического материала, состоящего из РНК[25]. Но РНК-зависимый синтез РНК, который раньше считался характерным только для вирусов, происходит и в клеточных организмах, в процессе так называемой РНК-интерференции[26].
Как в случае ДНК-зависимой РНК-полимеразы, так и в случае РНК-зависимой РНК-полимеразы фермент присоединяется к промоторной последовательности. Вторичная структура молекулы матрицы расплетается с помощью хеликазной активности полимеразы, которая при движении субстрата в направлении от 5′ к 3′ концу молекулы синтезирует РНК в 3′ —> 5′ направлении. Терминатор транскрипции в исходной молекуле определяет окончание синтеза. Многие молекулы РНК синтезируются в качестве молекул-предшественников, которые подвергаются «редактированию» — удалению ненужных частей с помощью РНК-белковых комплексов[27].
Например, у кишечной палочки гены рРНК расположены в составе одного оперона (в rrnB порядок расположения такой: 16S — tRNAGlu2 — 23S —5S) считываются в виде одной длинной молекулы, которая затем подвергается расщеплению в нескольких участках с образованием сначала пре-рРНК, а затем зрелых молекул рРНК[28]. Процесс изменения нуклеотидной последовательности РНК после синтеза носит название процессинга или редактирования РНК.
После завершения транскрипции РНК часто подвергается модификациям (см. выше), которые зависят от функции, выполняемой данной молекулой. У эукариот процесс «созревания» РНК, то есть её подготовки к синтезу белка, часто включает сплайсинг: удаление некодирующих белок последовательностей (интронов) с помощью рибонуклеопротеида сплайсосомы. Затем к 5′ концу молекулы пре-мРНК эукариот добавляется особый модифицированный нуклеотид (кэп), а к 3′ концу несколько аденинов, так назваемый «полиА-хвост»[27].
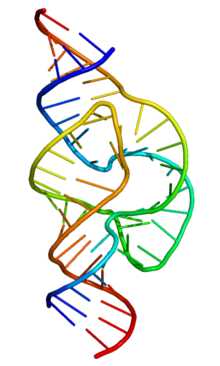 Структура молоточкового (hammerhead) рибозима, который расщепляет РНК
Структура молоточкового (hammerhead) рибозима, который расщепляет РНК.
Типы РНК
Матричная (информационная) РНК — РНК, которая служит посредником при передаче информации, закодированной в ДНК к рибосомам, молекулярным машинам, синтезирующим белки живого организма. Кодирующая последовательность мРНК определяет последовательность аминокислот полипептидной цепи белка [29]. Однако подавляющее большинство РНК не кодируют белок. Эти некодирующие РНК могут транскрибироваться с отдельных генов (например, рибосомальные РНК) или быть производными интронов [30]. Классические, хорошо изученные типы некодирующих РНК — это транспортные РНК (тРНК) и рРНК, которые участвуют в процессе трансляции [31]. Существуют также классы РНК, ответственные за регуляцию генов, процессинг мРНК и другие роли. Кроме того, есть и молекулы некодирующих РНК, способные катализировать химические реакции, такие, как разрезание и лигирование молекул РНК[32]. По аналогии с белками, способными катализировать химические реакции — энзимами (ферментами), каталитические молекулы РНК называются рибозимами.
Участвующие в трансляции
Информация о последовательности аминокислот белка содержится в мРНК. Три последовательных нуклеотида (кодон) соответствуют одной аминокислоте. В эукариотических клетках транскирибированный предшественник мРНК или пре-мРНК процессируется с образованием зрелой мРНК. Процессинг включает удаление некодирующих белок последовательностей (интронов). После этого мРНК экспортируется из ядра в цитоплазму, где к ней присоединяются рибосомы, транслирующие мРНК с помощью соединённых с аминокислотами тРНК.
В безъядерных клетках (бактерии и археи) рибосомы могут присоединяться к мРНК сразу после транскрипции участка РНК. И у эукариот, и у прокариот цикл жизни мРНК завершается её контролируемым разрушением ферментами рибонуклеазами [29].
Транспортные (тРНК) — малые, состоящие из приблизительно 80 нуклеотидов, молекулы с консервативной третичной структурой. Они переносят специфические аминокислоты в место синтеза пептидной связи в рибосоме. Каждая тРНК содержит участок для присоединения аминокислоты и антикодон для узнавания и присоединения к кодонам мРНК. Антикодон образует водородные связи с кодоном, что помещает тРНК в положение, способствующее образованию пептидной связи между последней аминокислотой образованного пептида и аминокислотой, присоединённой к тРНК [30].
Рибосомальные РНК (рРНК) — каталитическая составляющая рибосом. Эукариотические рибосомы содержат четыре типа молекул рРНК: 18S, 5.8S, 28S и 5S. Три из четырёх типов рРНК синтезируются в ядрышке. В цитоплазме рибосомальные РНК соединяются с рибосомальными белками и формируют нуклеопротеин, называемый рибосомой[29]. Рибосома присоединяется к мРНК и синтезирует белок. рРНК составляет до 80 % РНК, обнаруживаемой в цитоплазме эукариотической клетки [33].
Необычный тип РНК, который действует в качестве тРНК и мРНК (тмРНК) обнаружен во многих бактериях и пластидах. При остановке рибосомы на дефектных мРНК без стоп-кодонов тмРНК присоединяет небольшой пептид, направляющий белок на деградацию [34].
Участвующие в регуляции генов
В живых клетках обнаружено несколько типов РНК, которые могут уменьшать степень выражения гена при комплементарности мРНК или самому гену. Микро-РНК (21-22 нуклеотида в длину) найдены у эукариот и оказывают воздействие через механизм РНК-интерференции. При этом комплекс микро-РНК и ферментов может приводить к метилированию нуклеотидов в ДНК промотора гена, что служит сигналом для уменьшения активности гена. При использовании другого типа регуляции мРНК, комплементарная микро-РНК, деградируется.[35]. Однако есть и миРНК, которые увеличивают, а не уменьшают экспрессию генов.[36]. Малые интерферирующие РНК (миРНК, 20-25 нуклеотидов) часто образуются в результате расщепления вирусных РНК, но существуют и эндогенные клеточные миРНК[37]. Малые интерферирующие РНК также действуют через РНК-интерференцию по сходным с микро-РНК механизмам[38]. У животных найдены так называемыме РНК, взаимодействующие с Piwi (piRNA, 29-30 нуклеотидов), действующие в половых клетках против транспозиции и играющие роль в образовании гамет[39][40]. Кроме того, piRNA могут эпигенетически наследоваться по материнской линии, передавая потомству своё свойство ингибировать экспрессию транспозонов[41].
Антисмысловые РНК широко распространены у бактерий, многие из них подавляют выражение генов, но некоторые активируют экспрессию[42]. Действуют антисмысловые РНК, присоединяясь к мРНК, что приводит к образованию двуцепочечных молекул РНК, которые деградируются ферментами.[43]. У эукариот обнаружены высокомолекулярные, мРНК-подобные молекулы РНК. Эти молекулы также регулируют выражение генов,[44]. В качестве примера можно привести Xist, присоединяющуюся и инактивирующую одну из двух Х-хромосом у самок млекопитающих.[45].
Кроме роли отдельных молекул в регуляции генов, регуляторные элементы могут формироваться в 5′ и 3′ нетранслируемых участках мРНК. Эти элементы могут действовать самостоятельно, предотвращая инициацию трансляции, либо присоединять белки, например, ферритин или малые молекулы, например, биотин.[46].
В процессинге РНК
Многие РНК принимают участие в модификации других РНК. Интроны вырезаются из пре-мРНК сплайсосомами, которые, кроме белков, содержат несколько малых ядерных РНК (мяРНК)[31]. Кроме того, интроны могут катализировать собственное вырезание[47]. Синтезированая в результате транскрипции РНК также может быть химически модифицирована. У эукариот химические модификации нуклеотидов РНК, например, их метилирование, выполняется малыми ядерными РНК (мяРНК, 60-300 нуклеотидов). Этот тип РНК локализуется в ядрышко и тельцах Кахаля [30]. После ассоциации мяРНК с ферментами, мяРНК связываются с РНК-мишенью путём образования пар между основаниями двух молекул, а ферменты модифицируют нуклеотиды РНК-мишени. Рибосомальные и транспортные РНК содержат много подобных модификаций, конкретное положение которых часто сохраняется в процессе эволюции. Также могут быть модифицированы мяРНК и сами мяРНК[48][49]Гидовые РНК осуществляют процесс редактирования РНК в кинетопласте — особом участке митохондрии протистов-кинетопластид (например, трипаносом).
Геномы, состоящие из РНК
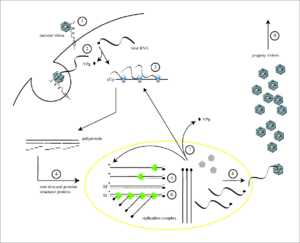 Жизненный цикл вируса с РНК геномом на примере полиовируса: 1 — присоединение исходного вириона к рецептору; 2 — вирион попадает в клетку; 3 — трансляция белков вируса с его РНК с образованием полипетида; 4 — полимеразы вируса размножают его РНК
Жизненный цикл вируса с РНК геномом на примере полиовируса: 1 — присоединение исходного вириона к рецептору; 2 — вирион попадает в клетку; 3 — трансляция белков вируса с его РНК с образованием полипетида; 4 — полимеразы вируса размножают его РНККак и ДНК, РНК может хранить информацию о биологических процессах. РНК может использоваться в качестве генома вирусов и вирусоподобных частиц. РНК-геномы можно разделить на те, которые не имеют промежуточной стадии ДНК и те, которые для размножения копируются в ДНК-копию и обратно в РНК (ретровирусы).
РНК-содержащие вирусы
Многие вирусы, например, вирус гриппа, на всех стадиях содержат геном, состоящий исключительно из РНК. РНК содержится внутри обычно белковой оболочки и реплицируется с помощью закодированных в ней РНК-зависимых РНК-полимераз. Вирусные геномы, состоящие из РНК разделяются на
- содержащие «плюс-цепь РНК», которая используется в качестве и мРНК, и генома;
- «минус-цепь РНК», которая служит только геномом, а в качестве мРНК используется комплементарная ей молекула;
- двухцепоченые вирусы.
Вироиды — другая группа патогенов, содержащих РНК-геном и не содержащих белок. Они реплицируются РНК-полимеразами организма хозяина[50].
Ретровирусы и ретротранспозоны
У других вирусов РНК-геном есть в течение только одной из фаз жизненного цикла. Вирионы так называемых ретровирусов содержат молекулы РНК, которые при попадании в клетки хозяина служат матрицей для синтеза ДНК-копии. В свою очередь, с матрицы ДНК считывается РНК-геном. Кроме вирусов обратную транскрипции применяют и класс мобильных элементов генома — ретротранспозоны [51]
Гипотеза РНК-мира
Способность молекул РНК одновременно служить как в качестве носителя информации, так и в качестве катализатора химических реакций, позволила выдвинуть гипотезу о том, что РНК была первым сложным полимером, появившимся в процессе добиологической эволюции. Эта гипотеза названа «гипотеза РНК-мира» [52][53]. Согласно ей, РНК на первых этапах эволюции автокатализировала синтез других молекул РНК, а затем и ДНК. На втором этапе эволюции синтезированные молекулы ДНК, как более стабильные, стали хранилищем генетической информации. Синтез белка на матрице РНК с помощью пра-рибосом, полностью состоящих из РНК, расширил свойства добиологических систем, постепенно белок заменил РНК в структурных аспектах. Из этой гипотезы делается вывод, что многие РНК, принимающие участие в синтезе белка в современных клетках, в особенности рРНК и тРНК — это реликты РНК-мира.
См. также
Примечания
Рекомендуемая литература
Ссылки
| Типы нуклеиновых кислот | |
|---|---|
| Азотистые основания | Пурины (Аденин, Гуанин) | Пиримидины (Урацил, Тимин, Цитозин) |
| Нуклеозиды | Аденозин | Гуанозин | Уридин | Тимидин | Цитидин |
| Нуклеотиды | монофосфаты (АМФ, ГМФ, UMP, ЦМФ) | дифосфаты (АДФ, ГДФ, УДФ, ЦДФ) | трифосфаты (АТФ, ГТФ, УТФ, ЦТФ) | циклические (цАМФ, цГМФ, cADPR) |
| Рибонуклеиновые кислоты | РНК | мРНК | тРНК | рРНК | антисмысловые РНК | gRNA | микроРНК | некодирующие РНК | piwi-interacting RNA | shRNA | малые интерферирующие РНК | малые ядерные РНК | малые ядрышковые РНК | тмРНК |
| Дезоксирибонуклеиновые кислоты | ДНК | кДНК | Геном | msDNA | Митохондриальная ДНК |
| Аналоги нуклеиновых кислот | en:glycerol nucleic acid | en:locked nucleic acid | ПНК | ТНК | Морфолино |
| Типы векторов | en:phagemid | Плазмиды | Фаг лямбда | en:cosmid | en:P1 phage | en:fosmid | en:Bacterial artificial chromosome | en:Yeast artificial chromosome | en:Human artificial chromosome |
dic.academic.ru
Рибонуклеиновая кислота : Р — Словарь : Все про гены!
РНК содержатся главным образом в цитоплазме клеток. Эти молекулы синтезируются в клетках всех клеточных живых организмов, а также содержатся в вироидах и некоторых вирусах.Основные функции РНК
в клеточных организмах — это шаблон для трансляции генетической информации в белки и поставка соответствующих аминокислот к рибосомам. В вирусах является носителем генетической информации (кодирует белки оболочки и ферменты вирусов). Вироиды состоят из кольцевой молекулы РНК и не содержат в себе других молекул. Существует гипотеза мира РНК, согласно которой, РНК возникли перед белками и были первыми формами жизни.Клеточные РНК образуются в ходе процесса, называемого транскрипцией, то есть синтеза РНК на матрице ДНК, осуществляемого специальными ферментами — РНК-полимеразы. Затем матричные РНК (мРНК) участвуют в процессе, называемом трансляцией. Трансляция — это синтез белка на матрице мРНК при участии рибосом. Другие РНК после транскрипции подвергаются химическим модификациям, и после образования вторичной и третичной структур выполняют функции, зависящие от типа РНК.
Для одноцепочечной РНК характерны разнообразные пространственные структуры, в которых часть нуклеотидов одной и той же цепи спарены между собой. Некоторые высокоструктурированные РНК принимают участие в синтезе белка клетки, например, транспортные РНК служат для узнавания кодонов и доставки соответствующих аминокислот к месту синтеза белка, а матричные РНК служат структурной и каталитической основой рибосом.
Однако функции РНК в современных клетках не ограничиваются их ролью в трансляции. Так мРНК участвуют в сплайсинге эукариотических матричных РНК и других процессах.
Кроме того, что молекулы РНК входят в состав некоторых ферментов (например, теломеразы) у отдельных РНК обнаружена собственная энзиматическая активность, способность вносить разрывы в другие молекулы РНК или, наоборот, «склеивать» два РНК-фрагмента. Такие РНК называются рибозимами.
Геномы ряда вирусов состоят из РНК, то есть у них она играет роль, которую в высших организмах выполняет ДНК. На основании разнообразия функций РНК в клетке была выдвинута гипотеза, согласно которой РНК — первая молекула, способная к самовоспроизведению в добиологических системах.
История изучения РНК
Нуклеиновые кислоты были открыты в 1868 году швейцарским ученым Иоганном Фридрихом Мишером, который назвал эти вещества «нуклеин», поскольку они были обнаружены в ядре (лат. nucleus). Позже было обнаружено, что бактериальные клетки, в которых нет ядра, тоже содержат нуклеиновые кислоты.
Значение РНК в синтезе белков было предположено в 1939 году в работе Торберна Оскара Касперссона, Жана Брачета и Джека Шульца. Джерард Маирбакс выделил первую матричную РНК, кодирующую гемоглобин кролика и показал, что при ее введении в ооциты образуется тот же самый белок.
В Советском Союзе в 1956-57 годах проводились работы (А. Белозерский, А. Спирин, Э. Волкин, Ф. Астрахан) по определению состава РНК клеток, которые привели к выводу, что основную массу РНК в клетке составляют рибосомные РНК.
В 1959 году Северо Очоа получил Нобелевскую премию по медицине за открытие механизма синтеза РНК. Последовательность из 77 нуклеотидов одной из тРНК дрожжей S. cerevisiae была определена в 1965 году в лаборатории Роберта Холле, за что в 1968 году он получил Нобелевскую премию по медицине.
В 1967 Карл Везе предположил, что РНК имеют каталитические свойства. Он выдвинул так называемую Гипотезу РНК-мира, в котором РНК прото-организмов служили и как молекулы хранения информации (сейчас эта роль выполняется ДНК) и как молекулы, которые катализировали метаболические реакции (сейчас это делают ферменты).
В 1976 Уолтер Фаерс и его группа из Гентского университета (Голландия) впервые определили последовательность генома РНК — содержащегося в вирусе, бактериофага MS2.
В начале 1990-х было обнаружено, что введение чужеродных генов в геном растений приводит к подавлению выражения аналогичных генов растения. Примерно в это же время было показано, что РНК длиной около 22 оснований, которые сейчас называются микро-РНК, играют регуляторную роль в онтогенезе круглых червей.
Гипотеза о значении РНК в синтезе белков была высказана Торбьерном Касперссоном (Torbjörn Caspersson) на основе исследований 1937-1939 гг., в результате которых было показано, что клетки, активно синтезирующие белок, содержат большое количество РНК. Подтверждение гипотезы было получено Юбером Шантренном (Hubert Chantrenne).
Особенности строения РНК
Нуклеотиды РНК состоят из сахара — рибозы, к которой в положении 1 ‘присоединена одна из основ: аденин, гуанин, цитозин или урацил. Фосфатная группа объединяет рибозы в цепочку, образуя связи с 3 ‘атомом углерода одной рибозы и в 5’ положении другого. Фосфатные группы при физиологическом рН отрицательно заряжены, поэтому РНК — можно назвать полианионом.
РНК транскрибируется как полимер четырех оснований (аденина (A), гуанина (G), урацила (U) и цитозина (C)), но в «зрелой» РНК есть много модифицированных оснований и сахаров. Всего в РНК насчитывается около 100 различных видов модифицированных нуклеозидов, из которых:
— 2′-О-метилрибоза наиболее частая модификация сахара;
— Псевдоуридин — наиболее часто модифицированная основа, которая встречается чаще всего. В псевдоуридине (Ψ) связь между урацилом и рибозой не C — N, а C — C, этот нуклеотид встречается в разных положениях в молекулах РНК. В частности, псевдоуридин важен для функционирования тРНК.
Еще одной модифицированной основой, о которой стоит сказать является — гипоксантин, деаминованний гуанин, нуклеозид которого носит название инозин. Инозин играет важную роль в обеспечении вырожденности генетического кода.
Роль многих других модификаций не до конца изучена, но в рибосомальной РНК много пост-транскрипционных модификаций находятся в важных для функционирования рибосомы участках. Например, на одном из рибонуклеотидов, участвующих в образовании пептидной связи. Азотистые основания в составе РНК могут образовывать водородные связи между цитозином и гуанином, аденином и урацилом, а также между гуанином и урацилом. Однако возможны и другие взаимодействия, например, несколько аденинов могут образовывать петлю, или петля, состоящая из четырех нуклеотидов, в которой есть пара оснований аденин — гуанин.
Важная структурная особенность РНК, отличающая ее от ДНК — наличие гидроксильной группы в 2 ‘положении рибозы, которая позволяет молекуле РНК существовать в А, а не В-конформации, которая наиболее часто наблюдается в ДНК. В А-форме глубокая и узкая большая бороздка и неглубокая и широкая малая бороздка. Второе последствие наличия 2 ‘гидроксильной группы состоит в том, что конформационно пластичные, то есть, не принимающие участие в образовании двойной спирали, участки молекулы РНК могут химически атаковать другие фосфатные связи и их расщеплять.
«Рабочая» форма одноцепочечной молекулы РНК, как и у белков, часто обладает третичной структурой. Третичная структура образуется на основе элементов вторичной структуры, образуется посредством водородных связей внутри одной молекулы. Различают несколько типов элементов вторичной структуры — стебель-петли, петли и псевдоузлы. В силу большого количества возможных вариантов спаривания оснований, предсказания вторичной структуры РНК — гораздо более сложная задача, чем структуры белков, но в настоящее время есть эффективные программы, например, mfold.
Примером зависимости функций молекул РНК от их вторичной структуры являются участки внутренней посадки рибосомы (IRES). IRES — структура на 5 ‘конце информационной РНК, которая обеспечивает присоединение рибосомы в обход обычного механизма инициации синтеза белка, требует наличия особого модифицированного основания (кэпа) на 5’ конце и белковых факторов инициации. Сначала IRES были обнаружены в вирусных РНК, но сейчас накапливается все больше данных о том, что клеточные мРНК также используют IRES-зависимый механизм инициации в условиях стресса. Многие типы РНК, например, рРНК и мяРНК (мяРНК) в клетке функционируют в виде комплексов с белками, которые ассоциируют с молекулами РНК после их синтеза или (у эукариот) экспорта из ядра в цитоплазму. Такие РНК-белковые комплексы называются рибонуклеопротеиновыми комплексами или рибонуклеопротеидами.
Матричная рибонуклеиновая кислота (мРНК, синоним — информационная РНК, иРНК) — РНК, отвечающая за перенос информации о первичной структуре белков от ДНК к местам синтеза белков. мРНК синтезируется на основе ДНК в ходе транскрипции, после чего, в свою очередь, используется при трансляции как матрица для синтеза белков. Тем самым мРНК играет важную роль в «проявлении» (экспрессии) генов.
Длина типичной зрелой мРНК составляет от нескольких сотен до нескольких тысяч нуклеотидов. Самые длинные мРНК отмечены у (+) оц РНК-содержащих вирусов, например пикорнавирусов, однако следует помнить, что у этих вирусов мРНК образует весь их геном.
Подавляющее большинство РНК не кодируют белок. Эти некодирующие РНК могут транскрибировать из отдельных генов (например, рибосомальные РНК) или быть производными интронов. Классические, хорошо изученные типы некодирующих РНК — это транспортные РНК (тРНК) и рРНК, участвующие в процессе трансляции. Существуют также классы РНК, ответственные за регуляцию генов, процессинг мРНК и другие роли. Кроме того, есть и молекулы некодирующих РНК, способные катализировать химические реакции, такие, как разрезание и лигирование молекул РНК. По аналогии с белками, способными катализировать химические реакции — энзимами (ферментами), каталитические молекулы РНК называются рибозимами.
Транспортные (тРНК) — малые, состоящие из примерно 80 нуклеотидов, молекулы с консервативной третичной структурой. Они переносят специфические аминокислоты к месту синтеза пептидной связи в рибосоме. Каждая тРНК содержит участок для присоединения аминокислоты и антикодон для узнавания и присоединения к кодону мРНК. Антикодон образует водородные связи с кодоном, что помещает тРНК в положение, способствующее образованию пептидной связи между последней аминокислотой образованного пептида и аминокислотой, присоединенной к тРНК.
Рибосомальные РНК (рРНК) — каталитическая составляющая рибосом. Эукариотические рибосомы содержат четыре типа молекул рРНК: 18S, 5.8S, 28S и 5S. Три из четырех типов рРНК синтезируются на полисомах. В цитоплазме рибосомальные РНК соединяются с рибосомальными белками и формируют нуклеопротеины, называемые рибосомами. Рибосома присоединяется к мРНК и синтезирует белок. рРНК составляет до 80% РНК, обнаруживается в цитоплазме эукариотической клетки.
Необычный тип РНК, который действует в качестве тРНК и мРНК (тмРНК) обнаружен во многих бактериях и пластидах. При остановке рибосомы на дефектных мРНК без стоп-кодонов тмРНК присоединяет небольшой пептид, направляющий белок на деградацию.
Микро-РНК (21-22 нуклеотида в длину) найдены у эукариот и влияют через механизм РНК-интерференции. При этом комплекс микро-РНК и ферментов может приводит к метилированию нуклеотидов в ДНК промотора гена, что служит сигналом для уменьшения активности гена. При использовании другого типа регуляции мРНК, комплементарная микро-РНК, деградирует. Однако есть и миРНК, которые увеличивают, а не уменьшают экспрессию генов.
Малые интерферирующие РНК (миРНК, 20-25 нуклеотидов) часто образуются в результате расщепления вирусных РНК, но существуют и эндогенные клеточные миРНК. Малые интерферирующие РНК также действуют через РНК-интерференцию по сходным с микро-РНК механизмам.
Сравнение с ДНК
Между ДНК и РНК есть три основных отличия:
1. ДНК содержит сахар дезоксирибозу, РНК — рибозу, у которой есть дополнительная, по сравнению с дезоксирибозой, гидроксильная группа. Эта группа увеличивает вероятность гидролиза молекулы, то есть уменьшает стабильность молекулы РНК.
2. Нуклеотид, комплементарный аденину, в РНК не тимин, как в ДНК, а урацил — неметилированная форма тимина.
3. ДНК существует в форме двойной спирали, состоящей из двух отдельных молекул. Молекулы РНК, в среднем, намного короче и преимущественно одноцепочечные. Структурный анализ биологически активных молекул РНК, включая тРНК, рРНК мяРНК и другие молекулы, которые не кодируют белков, показал, что они состоят не из одной длинной спирали, а из многочисленных коротких спиралей, расположенных близко друг к другу и образуют нечто, похожее на третичную структуру белка. В результате этого РНК может катализировать химические реакции, например, пептид-трансферазный центр рибосомы, участвующий в образовании пептидной связи белков, полностью состоит из РНК.
Особенности функций:
1. Процессинг
Многие РНК принимают участие в модификации других РНК. Интроны вырезают из про-мРНК сплайсосомы, которые, кроме белков, содержат несколько малых ядерных РНК (мяРНК). Кроме того, интроны могут катализировать собственное вырезание. Синтезированная в результате транскрипции РНК также может быть химически модифицирована. У эукариот химические модификации нуклеотидов РНК, например, их метилирование, выполняется малыми ядерными РНК (мяРНК, 60-300 нуклеотидов). Этот тип РНК локализуется в ядрышке и тельцах Кахаля. После ассоциации мяРНК с ферментами, мяРНК связывается с РНК-мишенью путем образования пар между основаниями двух молекул, а ферменты модифицируют нуклеотиды РНК-мишени. Рибосомальные и транспортные РНК содержат много подобных модификаций, конкретное положение которых часто сохраняется в процессе эволюции. Также могут быть модифицированы мяРНК и сами мяРНК.
2. Трансляция
тРНК присоединяют определенные аминокислоты в цитоплазме и направляется к месту синтеза белка на иРНК где связывается с кодоном и отдает аминокислоту которая используется для синтеза белка.
3. Информационная функция
У некоторых вирусов РНК выполняет те функции которые ДНК выполняет у эукариот. Также информационную функцию выполняет иРНК которая переносит информацию о белках и является местом его синтеза.
4. Регуляция генов
Некоторые типы РНК участвуют в регуляции генов увеличивая или уменьшая его активность. Это так называемые миРНК (малые интерферирующие РНК) и микро-РНК.
5. Каталитическая функция
Есть так называемые ферменты которые относятся к РНК они называются рибозимы. Эти ферменты выполняют различные функции и имеют своеобразное строение
vse-pro-geny.ru
Рибонуклеиновые кислоты функция — Справочник химика 21
Рибонуклеиновые кислоты — полимерные молекулы, которые по своей структуре подобны ДНК. Отличительной особенностью РНК является то, что углеводной компонентой в них является О-рибофураноза, а место тимина занимает урацил. Последовательность оснований в скелете природных РНК еще не известна причем в противоположность ДНК, РНК состоят из простых поли-нуклеотидных цепей, в структуре которых последовательность пуриновых и пиримидиновых оснований варьируется в значительно меньшей степени, чем в нуклеотидном составе ДНК. В зависимости от характера выполняемых функций РНК делятся на три группы. Это прежде всего рибосомальные РНК, являющиеся основным компонентом клетки. Полагают, что рибосомальные РНК участвуют в создании клеточных образований — рибосом, однако их функция окончательно не выяснена. Информационные РНК являются как бы шаблонами в синтезе белка и составляют активную часть полирибосом. Так, характер синтезируемого белка зависит от последовательности оснований (А, Ц, У и Г) в полинуклеотидной цепи информационной РНК. Наконец, третья форма — растворимые РНК, являются как бы адаптором аминокислот, направляющим аминокислоты к специальным участкам (шаблонам) информационной РНК, осуществляющей синтез белка. Более детально биологическая роль ДНК и РНК обсуждается в специальных обзорах [21, 24]. [c.335]Биохимические исследования давно привели к заключению, что синтезы таких специфических белков, как ферменты и т.д., контролируются шаблонами или матрицами, называемыми генами. Гены выполняют двойную функцию — воспроизведение собственной копии и обеспечение специфической структуры молекулы белка. Приведенные выше новые исследования, а также работы, направленные на изз ение размножения вирусов (см. Вирусы ), являются важным началом в познании химической природы генов. Большинство белков синтезируется в клеточной плазме в определенных полимеризационных центрах, называемых микросомами. Последние содержат только рибонуклеиновую кислоту и белки. Были открыты ферменты, связывающие аминокислоты с аденозинмонофосфорной кислотой с образованием смешанных ангидридов. Оказалось также, что эти ангидриды соединяются далее до входа в микросомы с рибонуклеиновой кислотой небольшого молекулярного веса, служащей, вероятно, переносчиком (М. Б. Хогланд 1956 г.). Таким образом, время выяснения механизма синтеза белков теперь кажется не очень далеким. [c.779]
Тот же самый принцип активации карбоксильной группы используется н в синтезе белков in vivo. Карбоксильная группа аминокислоты активируется, реагируя с АТР с промежуточным образованием ангидрида. Однако следующая стадия не сводится просто к атаке такого ангидрида второй аминокислотой, поскольку синтез белков включает строго определенное последовательное присоединение многих (до нескольких сотен) аминокислот. Матрица, или организующая поверхность , должна участвовать в этом процессе для того, чтобы обеспечить правильную последовательность белковой молекулы. Макромолекулой, выполняющей функцию такой матрицы, является полинуклеотидтранс-портная рибонуклеиновая кислота (тРНК) строение полинуклеотидов описано в следующей главе. [c.56]
Нуклеиновые кислоты, содержащие в молекулах остатки рибозы, называются рибонуклеиновыми кислотами (РНК), й содержащие остатки дезоксирибозы — дезоксирибонуклеиновыми кислотами (ДНК). ДНК содержится, по-видимому, только в ядрах клеток, а РНК — в протоплазме и таких органоидах, как рибосомы, митохондрии и др. Функции ДНК связаны с передачей наследственных признаков, а функции РНК — с биосинтезом белков, т. е. существует важнейшая [c.410]
Наряду с ДНК в сложном, катализируемом ферментами процессе синтеза белка участвует три типа рибонуклеиновых кислот. РНК синтезируются в ядре и благодаря сравнительно низкой молекулярной массе проникают сквозь оболочку ядра в цитоплазму клетки, где и выполняют свои функции. [c.333]
Структура и функции рибонуклеиновых кислот [c.184]
Молекулы рибонуклеиновых кислот (РНК) синтезируются в ядре клетки, однако свои функции они осуществляют в цитоплазме. Имеются три вида РНК, отличающиеся друг от друга молекулярным весом и вторичной структурой. Все они имеют значительно более низкий молекулярный вес, чем ДНК, и поэтому могут проникать через оболочку ядра клетки. [c.453]
Во всех живых клетках белки синтезируются рибосомами. Рибосома представляет собой крупную макромолекулу со сложной асимметричной четвертичной структурой, построенной из рибонуклеиновых кислот (рибосомных РНК) и белков. Для того чтобы синтезировать белок, рибосома должна быть снабжена а) программой, задающей порядок чередования аминокислотных остатков в полипептидной цепи белка б) аминокислотным материалом, из которого надлежит строить белок в) энергией. Сама рибосома обладает каталитической (энзиматической) функцией, ответственной за образование пептидных связей и, соответственно, полимеризацию аминокислотных остатков в полипептидную цепь белка. [c.7]
Известны нуклеиновые кислоты двух типов дезоксирибонуклеиновые кислоты (ДНК) и рибонуклеиновые кислоты (РНК). Функция РНК состоит в том, чтобы запустить в работу информацию, которая хранится в ДНК. [c.533]
Пусть теперь читатель будет готов к тому, что при рассмотрении связи между структурой и функцией (см. гл. 22.5) предстоит обсуждать еще более сложные проблемы. Пока что мы обратимся к более простым вопросам, касающимся природы рибонуклеиновых кислот. [c.53]
Нуклеиновые кислоты — информационные макромолекулы, состоящие из мононуклеотидов. В клетках содержится дезоксирибонуклеиновая кислота (ДНК) и рибонуклеиновые кислоты (РНК). ДНК — самая большая макромолекула в живых системах. Она состоит из многих тысяч пар нуклеотидов, соединенных друг с другом в определенной последовательности. Молекулы РНК по размеру много меньше, чем ДНК, однако их общее количество превышает ДНК. Для нуклеиновых кислот несвойственно многообразие функций, зато хранение и передача генетической информации является основой размножения и функционирования клеток. [c.9]
В зависимости от строения моносахарида, входящего в состав нуклеиновых кислот (НК), различают дезоксирибонуклеиновые кислоты (ДНК) и рибонуклеиновые кислоты (РНК). Необходимость подобной классификации определяется не только различным химическим строением ДНК и РНК, но и различием выполняемых ими биологических функций. Дезоксирибонуклеиновые кислоты ответственны за передачу наследственных признаков в ряду поколений живых организмов, поэтому конкретное строение ДНК каждого вида животных организмов будет строго специфично, однако общая структура ДНК одинакова для многих типов клеток. Рибонуклеиновые кислоты участвуют в процессе биосинтеза белка. [c.613]
С признанием по крайней мере некоторых биологических функций рибонуклеиновых кислот возникло представление о том, что порядок расположения различных нуклеотидных звеньев имеет особое значение, как и в случае белков и полипептидов. Экспериментальное определение этой последовательности представляет собой главную проблему сегодняшнего дня пока же наибольший фрагмент из природной рибонуклеиновой кислоты, структура которого установлена с определенностью, имеет длину меньше десяти нуклеотидов . Вследствие относительно небольшого числа разновидностей мономеров, участвующих в образовании молекулы нуклеиновой кислоты, методы, вкл
www.chem21.info
Рибонуклеиновые кислоты — Мегаэнциклопедия Кирилла и Мефодия — статья
Рибонуклеи́новые кисло́ты (РНК), семейство нуклеиновых кислот, содержащих в качестве углеводного компонента остаток рибозы. PНK присутствуют во всех живых клетках, участвуя в процессах, связанных с передачей генетической информации от дезоксирибонуклеиновой кислоты(ДНК) к белку. Из РНК образованы геномы многих вирусов. За редким исключением все PНK состоят из одиночных полинуклеотидных цепей. Их многомерные единицы — монорибонуклеотиды — содержат пуриновые— аденин и гуанин и пиримидиновыеоснования — цитозин и урацил. Обычно нуклеотиды обозначают начальными буквами названий входящих в их состав оснований на английском или русском (в русскоязычной научной литературе) языках: соответственно А, G (Г), С (Ц) и U (У). Как и в молекулах ДНК, отдельные нуклеотиды связаны между собой 3′-, 5′- фосфодиэфирными связями: остаток фосфорной кислоты служит связующим звеном между 3′-атомом углерода рибозы одного нуклеотида и 5′-атомом углерода рибозы другого (исходя из этого различают 3′-; и 5′-конец молекулы).Молекулы PНK содержат от нескольких десятков до нескольких десятков тысяч нуклеотидов. Все РНК способны к формированию вторичной структуры, основным элементом которой являются сравнительно короткие двуцелочечные тяжи, образованные комплементарными основаниями одной и той же молекулы, и связывающие их однотяжевые участки.
В живой клетке синтез РНК происходит с помощью фермента РНК-полимеразы в процессе транскрипции — считывания (переписывания) информации заложенной в молекуле ДНК, или в гене (часто группе генов).
Молекулы РНК формируются на матрице, которой служит одна из цепей ДНК, последовательность нуклеотидов в которой определяет порядок включения рибонуклеотидов по принципу комплементарности (гуанин против цитозина, урацил против аденина и т. д.). РНК-полимераза, продвигаясь вдоль одной из цепей ДНК, соединяет нуклеотиды в том порядке, который определен матрицей. Образовавшиеся молекулы РНК называют транскриптами.
Впервые РНК была выделена в 1889 году немецким биохимиком Р. Альтманом из клеток дрожжей. Но только в 1950-х гг., после доказательства генетической роли ДНК и установления ее двуцепочечной структуры, были описаны основные типы РНК и определены их функции. Было показано, что основная масса РНК в клетке (до 80%) приходится на рибосомные рибонуклеиновые кислоты (рРНК), которые в комплексе с белками образуют клеточную органеллу — рибосому, и непосредственно участвуют в биосинтезе белка (см. Трансляция).В начале 19б0-х гг. на основании того, что у эукариотпочти вся ДНК сосредоточена в ядре, а синтез белка протекает главным образом в цитоплазмена рибосомах, была высказана мысль о том, что какой-то вид РНК несет генетическую информацию для синтеза белка, то есть должна существовать какая-то молекула, переносящая информацию от ядра к рибосомам. Вскоре было установлено, что эту роль играют информационные, или матричные, рибонуклеиновые кислоты (иРНК, мРНК) — комплементарные копии генов.Каждому работающему гену (или группе генов) соответствует своя молекула мРНК. Первичный транскрипт (мРНК-предшественник) по размерам намного (в 4-5 раз) превышает зрелую мРНК, которая, в свою очередь, служит матрицей для синтеза белка на рибосомах. Это связано прежде всего с экзон-интронным строением большинства эукариотических генов. В ходе созревания (процессинга) мРНК ее интронные участки удаляются, а экзоны — сшиваются друг с другом, или, как говорят, подвергаются сплайсингу. Кроме того, молекула мРНК претерпевает и некоторые другие изменения (укорачивание со стороны 3′-конца, присоединение от 20 до 200 остатков адениловой кислоты и т. д.). Каждые три последовательно расположенных основания в зрелой мРНК, называемые кодонами, соответствуют определенной аминокислоте в молекуле белка, который синтезируется по мере продвижения вдоль нее рибосомы. У прокариот мРНК не претерпевает существенных изменений после синтеза на ДНК.
В расшифровке же (декодировании) записанной в мРНК информации участвуют транспортные рибонуклеиновые кислоты (тРНК). Они переносят нужную аминокислоту к растущей полипептидной цепи в ходе синтеза белка. Узнавание кодона в мРНК осуществляется с помощью трех оснований в тРНК, называемых антикодонами, а доставляемая аминокислота присоединена к ее 3′-концу. Специфичность такой системы переноса обеспечивается тем, что имеется по крайней мере одна тРНК для каждой аминокислоты (валиновая тРНК переносит валин, аланиновая — аланин и т. д.). Но, как правило, в цитоплазме клеток встречается около 40 видов тРНК, которые доставляют 20 аминокислот, так как одна и та же аминокислота может кодироваться несколькими разными кодонами (см. Генетический код).В дальнейшем были разработаны способы определения последовательности нуклеотидов в РНК, установления ее пространственной структуры. Крупнейшим открытием явилось обнаружение у некоторых РНК, названных рибозимами, способности расщеплять полирибонуклеотидные цепи, то есть играть роль фермента (ранее считалось, что ферментами могут быть только белки). Этот факт дал основания предполагать, что в период зарождения жизни на Земле РНК могла действовать самостоятельно (без белков и ДНК) и все биохимические превращения осуществлялись при ее участии.
Среди всех вышеперечисленных основных трех классов РНК наиболее изучены тРНК. Отличительной особенностью тРНК является наличие в их структуре так называемых минорных (необычных) оснований (описано более 40 таких оснований). Среди них наиболее часто встречаются инозин — производное аденина, псевдоуридин и другие производные урацила. Минорные основания появляются в тРНК уже после ее синтеза на ДНК, причем превращение обычных оснований в модифицированные происходит при участии специальных ферментов. Различные виды тРНК имеют сходную структуру и состоят примерно из 76 нуклеотидов (молекулярная масса 25 тыс.). Укладка молекул тРНК напоминает по форме клеверный лист. В ней выделяют 4 участка (стебли), в которых комплементарные пары оснований образуют водородные связи, и 3 одноцепочечных участка (петли). Значительно сложнее выглядят структуры высокомолекулярных рРНК или геномной РНК вирусов, однако детали пространственной организации их молекул пока неизвестны.
Помимо вышеописанных типов РНК, все эукариотические клетки содержат множество низкомолекулярных (коротких) молекул РНК, которые находятся в виде комплекса с молекулами белка (нуклеопротеидные частицы). Так называемые малые ядерные РНК (мяРНК) играют ключевую роль в процессинге транскриптов. Роль цитоплазматических низкомолекулярных РНК до конца не ясна.
У РНК-содержащих вирусов геномы могут быть представлены или двуцепочечной РНК, или одноцепочечной. Во многих случаях структурная организация геномных РНК сходна с мРНК эукариот. Поэтому при синтезе вирусных белков в клетке вирусная РНК может использоваться непосредственно в качестве матрицы.
Репликация ДНК
megabook.ru
14.Рибонуклеиновые кислоты, их виды, строение, назначение.
РНК. Строение молекул РНК во многом сходно со строением молекул ДНК. Однако имеется и ряд существенных отличий. В молекуле РНК вместо дезоксирибозы в состав нуклеотидов входит рибоза, вместо тимидилового нуклеотида (Т) — уридило-вый (У). Главное отличие от ДНК состоит в том, что молекула РНК представляет собой одну цепь. Однако ее нуклеотиды способны образовывать водородные связи между собой (например, в молекулах тРНК, рРНК), но в этом случае речь идет о внутри-цепочечном соединении комплементарных нуклеотидов. Цепочки РНК значительно короче ДНК.
В клетке существует несколько видов РНК, которые различаются по величине молекул, структуре, расположению в клетке и функциям:
1. Информационная (матричная) РНК(иРНК). Этот вид наиболее разнороден по размерам и структуре. иРНК представляет собой незамкнутую полинуклеотидную цепь. Она синтезируется в ядре при участии фермента РНК-полимеразы, комплементарнаучастку ДНК, на котором происходит ее синтез. Несмотря на относительно низкое содержание (3—5% РНК клетки), она выполняет важнейшую функцию в клетке: служит в качестве матрицы для синтеза белков, передавая информацию об их структуре с молекул ДНК. Каждь|й белок клетки кодируется специфической иРНК, поэтому число их типов в клетке соответствует числу видов белков.
2. Рибосомная РНК (рРНК). Это одноцепочечные нуклеиновые кислоты, образующие в комплексе с белками рибосомы — орга-неллы, на которых происходит синтез белка. Рибосомные РНК синтезируются в ядре. Информация об их структуре закодирована в участках ДНК, которые расположены в области вторичной перетяжки хромосом. Рибосомные РНК составляют 80% всей РНК клетки, поскольку в клетке имеется огромное количество рибосом. Рибосомные РНК обладают сложной вторичной и третичной структурой, образуя петли на комплементарных участках, что приводит к самоорганизации этих молекул в сложное по форме тело. В состав рибосом входит три типа рРНК у прокариот и четыре типа рРНК у эукариот.
3. Транспортная (трансферная) РНК(тРНК).Молекула тРНК состоит в среднем из 80 нуклеотидов. Содержание тРНК в клетке — около 15% всей РНК. Функция тРНК — перенос аминокислот к месту синтеза белка. Число различных типов тРНК в клетке невелико (20—60). Все они имеют сходную пространственную организацию. Благодаря внутрицепочечным водородным связям молекула тРНК приобретает характерную вторичную структуру, называемую клеверным листам. Трехмерная же модель тРНК выглядит несколько иначе. В тРНК выделяют четыре петли: акцепторную (служит местом присоединения аминокислоты), антикодоновую (узнает кодон в иРНК в процессе трансляции) и две боковые.
15.Органические вещества в клетках, их назначение.
В клетке содержится множество разнообразных органических соединений, разнообразных по структуре и выполняемым функциям. Органические вещества могут быть низкомолекулярными (аминокислоты, сахары, органические кислоты, нуклеотиды, липиды и т.д.) и высокомолекулярными. Большинство высокомолекулярных органических соединений в клетке являются биополимерами. Полимерами называются молекулы, состоящие из большого числа повторяющихся единиц –мономеров, соединенных друг с другом ковалентными связями. К биополимерам, т.е. к полимерам, входящим в состав клетки, относятся белки, полисахариды и нуклеиновые кислоты.
Особую группу органических соединений клетки составляют липиды (жиры и жироподобные вещества). Все они являются гидрофобными соединениями, т.е. нерастворимы в воде, но растворимы в неполярных органических растворителях (хлороформе, бензоле, эфире) К липидам относятся нейтральные жиры, фосфолипиды , воски, стероиды и некоторые другие соединения. Функции липидов в живых организмах разнообразны. Фосфолипиды присутствуют во всех клетках, выполняя структурную функцию в качестве основы биологических мембран. Стероид холестерин является важным компонентом мембран у животных. Нейтральные жиры и некоторые другие липиды обеспечивают энергетическую функцию. Они накапливаются в живых организмах в качестве запасных питательных веществ. При окислении 1 г жира высвобождается 38 кДж энергии, что в два раза больше, чем при окислении такого же количества глюкозы. С энергетической функцией жиров связана их запасающая функция. В виде жира хранится значительная часть энергетического запаса организма. Кроме того, жиры служат источником воды, которая выделяется при его окислении. Это особенно важно для пустынных животных, испытывающих дефицит воды. Например, именно жировые отложения находятся в горбе у верблюда. Для ряда липидов свойственна защитная функция. У млекопитающих подкожный жир выступает в качестве термоизолятора. Воск предохраняет перья и шерсть животных от смачивания. Ряд липидов выполняет в организме регуляторную функцию. Например, гормоны коры надпочечников по своей химической природе являются стероидами. Часть липидов принимают активное участие в обмене веществ, например жирорастворимые витамины А, D, E и K .
Углеводы (сахары, сахариды) представляют собой соединения с общей химической формулой Сn(h3O)n. По количеству звеньев в полимерной цепи различают три основных класса углеводов: моносахариды (простые сахары),олигосахариды (состоят из 2-10 молекул простых сахаров)и полисахариды (состоят более чем из 10 молекул простых сахаров). В зависимости от числа атомов углерода, входящих в состав моносахарида, различают триозы, тетрозы, пентозы, гексозы и гептозы. В природе наиболее распространены гексозы (глюкоза и фруктоза) и пентозы (рибоза идезоксирибоза). Глюкоза является основным источником энергии для клетки, при полном окислении 1 г глюкозы выделяется 17,6 кДж энергии. Рибоза и дезоксирибоза входят в состав нуклеиновых кислот. Из олигосахаридов наиболее часто встречаются дисахариды мальтоза (солодовый сахар), лактоза (молочный сахар), сахароза (свекловичный сахар). Моносахарида и дисахариды хорошо растворимы в воде и обладают сладким вкусом. Полисахариды имеют высокую молекулярную массу, не имеют сладкого вкуса и неспособны растворяться в воде. Они являются биополимерами. К наиболее распространенным в природе полисахаридам относятся полимеры глюкозы крахмал, гликоген и целлюлоза, а такжехитин, состоящий из остатков глюкозамина. Крахмал является основным запасным веществом у растений, гликоген – у животных. Целлюлоза и хитин выполняют защитную функцию, обеспечивая прочность покровов растений, животных и грибов. Таким образом, основные функции углеводов в природе — энергетическая, запасающая и структурная.
Белки– это биополимеры, мономерами которых являются аминокислоты. В образовании белков участвует 20 различных аминокислот. Аминокислоты в молекулах белка соединены ковалентными пептидными связями. В молекулу белка может входить до нескольких тысяч аминокислот. Выделяют 4 уровня пространственной организации молекулы белков. Последовательность аминокислот в полипептидной цепочке называется первичной структурой белка. Первичная структура молекулы любого белка уникальна и определяет его пространственную организацию, свойства и функции в клетке. Вторичная структура белка определяется укладкой цепочки аминокислот в определенные структуры, называемые a-спиралью и b-слоем. Вторичная структура белка формируется за счет водородных связей. Третичная структура образуется при сворачивании полипептидной цепи с элементами вторичной структуры в клубок (глобулу) и поддерживается за счет ионных, гидрофильных и ковалентных (дисульфидных) связей между различными остатками аминокислот. Четвертичная структура характерна для белков, состоящих из нескольких полипептидных цепей. Утрата белковой молекулой своей структурной организации, например вследствие нагревания, называется денатурацией. Денатурация может быть обратимой и необратимой. При обратимой денатурации может нарушаться четвертичная, третичная и вторичная структуры белка, но первичная структура не нарушается, и при возвращении нормальных условий за счет этого возможна ренатурация – восстановление нормальной конфигурации. При нарушении первичной структуры денатурация бывает необратимой.
Важнейшей функцией белков является каталитическая. Все ферменты, биологические катализаторы являются белками. Благодаря ферментам скорость химических реакций в клетке возрастает в миллионы раз. Ферменты высокоспецифичны: каждый фермент катализирует определенный тип химической реакции в клетке. Именно благодаря ферментам возможны все реакции обмена веществ, происходящие в живых организмах.
+ нуклеиновые кислоты!(см.выше вопрос 13)
studfiles.net
Нуклеиновые кислоты
Нуклеиновые кислоты — это ДНК (дезоксирибонуклеиновая кислота) и РНК (рибонуклеиновая кислота). Они определяют генетический материал всех живых организмов.
Молекулы ДНК (дезоксирибонуклеиновой кислоты) — это самые крупные линейные нерегулярные биополимеры, их мономером является нуклеотиды. Каждый нуклеотид состоит из остатков трех веществ: азотистого основания, углевода дезоксирибозы и фосфорной кислоты (рис.4). Известны 4 разновидности нуклеотидов: цитидиловый (Ц), тимидиловый (Т), адениловый (А), гуаниловый (Г). В названии каждого нуклеотида отражено название азотистого основания. Они отличаются друг от друга азотистыми основаниями. Цитозин и тимин — производные пиримидина, аденин и гуанин — относят к производным пурина. Соединение нуклеотидов в нити ДНК происходит через углевод одного нуклеотида и остаток фосфорной кислоты соседнего (фосфодиэфирных связей).
Рис. 5. Соединение нуклеотидов в полинуклеотидную цепь.
Согласно модели ДНК, предложенной Дж.Уотсоном и Ф.Криком (1953 г.), молекула ДНК представляет собой две спирально обвивающие друг друга нити. Обе нити вместе закручены вокруг общей оси. Две нити молекулы удерживаются рядом водородными связями, которые возникают между их комплементарными азотистыми основаниями. Аденин комплементарен тимину, а гуанин — цитозину. Между аденином и тимином возникают две водородные связи, между гуанином и цитозином — три.
ДНК находится в ядре, где она вместе с белками образует линейные структуры — хромосомы. Хромосомы хорошо видны при микроскопировании в период деления ядра; в интерфазе они деспирализованы.
ДНК имеется в митохондриях и пластидах (хлоропластах и лейкопластах), где их молекулы образуют кольцевые структуры. В клетках прокариот также присутствует кольцевая ДНК.
ДНК способна к самоудвоению (редупликации). Это имеет место в определенном периоде жизненного цикла клетки (синтетический период интерфазы). Редупликация позволяет сохранить постоянство структуры ДНК. Если под воздействием различных факторов в процессе репликации в молекуле ДНК происходят изменения в числе, порядке следования нуклеотидов, то возникают мутации.
Основная функция ДНК — хранение наследственной информации, заключенной в последовательности нуклеотидов, образующих ее молекулу, ипередача этой информациидочерним клеткам. Возможность передачи наследственной информации от клетки к клетке обеспечивается способностью хромосом к разделению на хроматиды с последующей редупликацией молекулы ДНК.
В ДНК заключена вся информация о структуре и деятельности клеток, о признаках каждой клетки и организма в целом. Эта информация называется генетической.
В молекуле ДНК закодирована генетическая информация о последовательности аминокислот в молекуле белка. Участок ДНК, несущий информацию об одной полипептидной цепи, называется геном. Передача и реализация информации осуществляется в клетке при участии рибонуклеиновых кислот.
Рибонуклеиновые кислоты бывают нескольких видов. Есть рибосомальная, транспортная и информационная РНК. Нуклеотиды РНК включают одно из азотистых оснований (аденин, гуанин, цитозин или урацил), углевод — рибозу и остатк фосфорной кислоты. Молекулы РНК представлены одной цепочкой нуклеотидов, образуя одноцепочечный линейный нерегулярный биополимер.
Рибосомальная РНК (р-РНК) в соединении с белком входит в состав субъединиц рибосом. Рибосомы участвуют в синтезе белка. Рибосомальная РНК составляет 80% от всей РНК в клетке. Количество нуклеотидов в р-РНК от 3 до 5 тыс. Этот вид РНК синтезируется особыми генами, находящимися в нескольких хромосомах в ядрышковом организаторе.
Информационная РНК (и-РНК) составляет от 1 до 10% от всей РНК в клетке. По строению и-РНК комплементарна участку молекулы ДНК, несущему информацию о синтезе определенного белка. Длина и-РНК зависит от длины участка ДНК, с которого считывали информацию. И-РНК переносит информацию о синтезе белка из ядра в цитоплазму. Перед выходом из ядра происходит редактирование и-РНК — сплайсинг.
Транспортная РНК (т-РНК) составляет около 10- 15% всей РНК. Она имеет короткую цепь нуклеотидов, состоящую из 80 нуклеотидов. Имеет модифицированные нуклеотиды, которые защищают т-РНК от разрушения ферментами. Образуется в ядре на ДНК, находится в цитоплазме. Т-РНК имеет форму трилистника. На верхушке центрального изгиба находится триплет нуклеотидов (антикодон), кодирующий определенную аминокислоту. На другом конце имеется триплет нуклеотидов- посадочная площадка, к которому присоединяется аминокислота. Т-РНК присоединяет определенные аминокислоты и подвозит их к месту синтеза белка к рибосомам. Известно 20 видов т-РНК
РНК находится в ядрышке, в цитоплазме, в рибосомах, в митохондриях и пластидах.
В природе есть еще один вид РНК. Это вирусная РНК. У одних вирусов она выполняет функцию хранения и передачи наследственной информации. У других вирусов эту функцию выполняет вирусная ДНК.
studfiles.net
Leave A Comment