
Устойчивость растений к переувлажнению
Переувлажнение наблюдается при заболачивании почв, затяжных дождях, в результате весеннего таяния снега, при нарушении поливных норм на орошаемых территориях.
Избыток влаги оказывает крайне неблагоприятное влияние на растения. В этих условиях появление всходов затруднено; растения угнетаются из-за образования корки на поверхности почвы; оголяется узел кущения у злаков; опыление из-за туманов и дождей плохое, поэтому мало образуется завязей; а нередко наблюдается осыпание завязей и незрелых плодов; происходит загнивание корнеклубнеплодов, плодов овощных культур; переувлажнение валков при раздельной уборке вызывает прорастание зерна в валках, а у культур с коротким периодом покоя или его отсутствием — прорастание семян на корню. После затяжных дождей наблюдается растрескивание плодов у томатов, ягодных культур, корнеплодов; у подсолнечника загнивает тыльная часть корзинки. Наблюдается полегание посевов, особенно высокорослых сортов или на участках с усиленным азотным питанием.
Если сильные дожди сопровождаются градом, то растения повреждаются травматически, иногда до полного уничтожения посевов.
Одна из основных причин угнетения растений при избыточном увлажнении почвы состоит в нарушении воздушного режима в зоне корней. При оптимальной обеспеченности водой почва содержит достаточное количество кислорода. Но при затоплении воздух из пор вытесняется водой.
Растения страдают от недостатка кислорода (гипоксия) или его полного отсутствия (аноксия). В условиях кислородной недостаточности часто оказываются озимые, ячмень, соя, рис, хлопчатник.
Кислород необходим как для дыхания корней, так и для жизнедеятельности аэробных микроорганизмов. При длительном затоплении в почве развиваются анаэробные процессы: спиртовое, маслянокислое и другие виды брожения. Происходит подкисление почвы в ризосфере, накапливаются углекислый газ, метан, органические кислоты, спирты и другие соединения. Многие из этих соединений ядовиты для корней растений, их называют болотными токсинами. В таких условиях корни начинают загнивать и отмирать, что приводит к гибели растений.
Многие из этих соединений ядовиты для корней растений, их называют болотными токсинами. В таких условиях корни начинают загнивать и отмирать, что приводит к гибели растений.
Нарушение аэробного дыхания в клетках корней сопряжено с прекращением транспорта электронов и окислительного фосфорилирования, прерываются реакции цикла трикарбоновых кислот. В этих условиях окисление углеводов в клетках переключается на путь спиртового или молочнокислого брожения. В результате в условиях аноксии образование АТФ происходит только на этапе гликолиза, а цикл трикарбоновых кислот полностью выключается. Поэтому из каждой молекулы глюкозы образуется только 2 молекулы АТФ (при нормальном снабжении кислородом и аэробном дыхании образуется 36 молекул АТФ). В итоге растения испытывают недостаток энергии для сохранения процессов метаболизма, роста и развития.
В процессе брожения в клетках накапливаются продукты брожения — молочная кислота и этиловый спирт. Спирт ядовит для клеток, а органические кислоты вызывают подкисление цитоплазмы и увеличение проницаемости клеточных мембран.
В связи с угнетением дыхания резко снижается поглотительная активность корней, так как поглощение питательных веществ требует затраты энергии, поставляемой в форме АТФ процессом дыхания. Наблюдается задержка роста растений. Недостаточно эффективное поглощение корнями минеральных элементов и их слабый транспорт в побеги сопровождается усиленной реутилизацией азота, фосфора и других элементов из более старых листьев в более молодые. Следствием этого является быстрое старение и гибель закончивших рост листьев.
Надземные органы растений не испытывают недостатка кислорода, а влияние затопления на них осуществляется опосредовано через нарушение трофических связей с корневой системой. Это определяется нарушением поступления из корневой системы в надземные органы элементов минерального питания и физиологически активных веществ.
Отрицательное влияние высокой влажности почвы и воздуха на процесс формирования урожайности сельскохозяйственных растений проявляется также в ухудшении условий для созревания семян. На завершающем этапе онтогенеза происходит обезвоживание семян. Затяжные дожди в предуборочный и уборочный периоды ухудшают условия для высыхания зерна и вызывают прорастание его в валках или на корню. Это происходит в результате биохимического распада веществ зерна и, в первую очередь, гидролиза крахмала до более простых соединений — сахаров под влиянием амилолитических ферментов. При сильном прорастании может начаться также распад клейковинных белков и липидов. Прорастание семян на корню или в валках нередко отмечается во влажные годы у ржи, пшеницы, тритикале, гороха, гречихи и других культур. Прорастание зерна резко ухудшает хлебопекарные свойства ржи и пшеницы: хлеб получается малого объема, мякиш плотный, липкий, легко заминается, цвет корки белесый с сильными подрывами.
Сельскохозяйственные растения в большинстве плохо переносят затопление. К наиболее чувствительным к кислородному дефициту относятся горох, томаты, соя; к относительно устойчивым — кукуруза, пшеница, овес, картофель. Самыми устойчивыми являются растения водные и влажных (заболоченных) местообитаний. Из растений сельскохозяйственного назначения к ним относятся рис и сахарный тростник.
К наиболее чувствительным к кислородному дефициту относятся горох, томаты, соя; к относительно устойчивым — кукуруза, пшеница, овес, картофель. Самыми устойчивыми являются растения водные и влажных (заболоченных) местообитаний. Из растений сельскохозяйственного назначения к ним относятся рис и сахарный тростник.
У устойчивых растений в процессе эволюции сформированы разнообразные морфологические, анатомические приспособления и биохимические механизмы, которые позволяют им выживать при дефиците кислорода. Основная особенность таких растений — это наличие аэренхимы — основной ткани, содержащей очень крупные межклетники. Межклетники и воздушные полости в корнях, стеблях и черешках листьев сообщаются между собой и служат резервуаром для кислорода, необходимого для дыхания всех тканей.
У риса затопление вызывает удлинение клеток междоузлий, поэтому рост всего стебля ускоряется и верхняя часть его всегда находится над водой. Первичная кора корня и стебля дифференцируется в аэренхиму.
Выживаемость растений сахарного тростника при затоплении определяется также способностью образовывать придаточные корни на узлах, расположенных выше уровня воды.
У растений-мезофитов аэренхима при нормальных условиях обеспеченности кислородом отсутствует, но может формироваться в ответ на недостаток кислорода в основании стебля и в корнях. Формирование этой ткани регулируется этиленом. В образовании этилена участвуют ферменты аминоциклопропанкарбосинтаза (АЦК-синтаза) и АЦК-оксидаза. Эти ферменты синтезируется в кончиках корней в ответ на дефицит кислорода. Синтезируются ферменты, связанные с лизисом клеточных стенок и образованием аэренхимы. В результате происходит гибель клеток первичной коры корня и стебля, а в местах гибели остаются полости, которые используются как резервуары для кислорода, поступающего в них из надземных органов. В образовании этой ткани участвуют также ионы кальция.
Формирование этой ткани регулируется этиленом. В образовании этилена участвуют ферменты аминоциклопропанкарбосинтаза (АЦК-синтаза) и АЦК-оксидаза. Эти ферменты синтезируется в кончиках корней в ответ на дефицит кислорода. Синтезируются ферменты, связанные с лизисом клеточных стенок и образованием аэренхимы. В результате происходит гибель клеток первичной коры корня и стебля, а в местах гибели остаются полости, которые используются как резервуары для кислорода, поступающего в них из надземных органов. В образовании этой ткани участвуют также ионы кальция.
Формирование аэренхимы часто бывает недостаточным для нормальной жизнедеятельности растений в условиях затопления, поэтому большое значение имеют физиолого-биохимические приспособления, которые обеспечивают достаточную активность обмена веществ в условиях пониженной концентрации кислорода. Эти приспособления связаны в основном с процессом дыхания — его путями.
В условиях дефицита кислорода повышается активность пентозофосфатного пути дыхания с последующим окислением в митохондриях образующегося НАДФН и сопряженного образования АТФ. Увеличивается значение гликолитического пути распада глюкозы. Возрастает значение системы детоксикации продуктов анаэробного распада (этилового спирта, молочной кислоты) путем удаления этих веществ или включения в обмен веществ. Нередко устойчивые к кислородному дефициту растения не накапливают продукты брожения. В этом случае образующиеся продукты брожения могут выходить из корней в почву или подниматься с транспирационным током в надземную часть, а затем удаляться через листья или чечевички побегов.
Увеличивается значение гликолитического пути распада глюкозы. Возрастает значение системы детоксикации продуктов анаэробного распада (этилового спирта, молочной кислоты) путем удаления этих веществ или включения в обмен веществ. Нередко устойчивые к кислородному дефициту растения не накапливают продукты брожения. В этом случае образующиеся продукты брожения могут выходить из корней в почву или подниматься с транспирационным током в надземную часть, а затем удаляться через листья или чечевички побегов.
Образующийся этиловый спирт может разрушаться с участием фермента алкогольдегидрогеназы. Опытами установлено, что активность этого фермента в анаэробных условиях повышается. Нейтрализация конечных продуктов брожения вызывает усиление процесса гликолиза.
Аноксия вызывает образование новых белков и ферментов. В частности обнаруживаются новые изоферменты, участвующие в гликолизе. Они поддерживают более надежное функционирование гликолиза в изменившихся условиях и снабжение растений энергией при подавлении аэробного дыхания. Изоферменты различаются между собой по физико-химическим свойствам, но катализируют одну и ту же реакцию.
Изоферменты различаются между собой по физико-химическим свойствам, но катализируют одну и ту же реакцию.
К физиолого-биохимическим приспособлениям у растений к недостатку кислорода относится способность их использовать в качестве конечного акцептора электронов не кислорода (как при аэробном дыхании), а других соединений, например нитратов или соединений, имеющих двойные связи — жирных кислот, каротиноидов. Процесс переноса электронов и протонов на NO3- получил названия нитратного дыхания.
Таким образом, у растений, произрастающих в условиях переувлажнения, устойчивость к гипоксии и аноксии достигается широким комплексом приспособлений.
Под влиянием избыточного увлажнения у полевых культур особенно высокорослых сортов нередко наблюдается полегание растений. Это вызывает большие неудобства для возделывания растений и при их уборке. Особенно большими потерями сопровождается полегание злаковых культур. При избыточном увлажнении рост растений ускоряется, междоузлия вытягиваются, а образование механической ткани отстает, поэтому прочность стебля снижается.
Интенсивность полегания определяется в баллах по результатам визуального наблюдения за посевами.
Повышение устойчивости растений к переувлажнению.
В условиях затопления при отсутствии в почве кислорода растения некоторых видов и сортов способны переносить электроны на другие акцепторы (нитраты, а также соединения, имеющие двойные связи — жирные кислоты, каротиноиды).
Благодаря этому при недостатке или отсутствии кислорода поддерживается жизнедеятельность растений. Подкормки таких культур нитратными удобрениями продлевают их жизнедеятельность в условиях переувлажнения и затопления. Устойчивость хлебных злаков к избытку воды в почве повышает также замачивание семян в растворе сульфата марганца (0,1 %), что способствует развитию генеративных органов, формированию зерновок в колосьях, а также в растворах хлорхолинхлорида или никотиновой кислоты.
Механизмы адаптации растений к недостатку кислорода (из Т.В. Чирковой, 2002)
Оценка интенсивности полегания растений
| Интенсивность полегания | Оценка, балл |
Очень сильная. Стебли практически лежат на земле. Механизированная уборка даже при проходе комбайна в одном направлении без значительных потерь урожая невозможна. Стебли практически лежат на земле. Механизированная уборка даже при проходе комбайна в одном направлении без значительных потерь урожая невозможна. | 1 |
| Сильная. Стебли сильно наклонены. Механизированная уборка возможна только в одном направлении (против направления полегания) с приспособлением для уборки полеглых хлебов. | 2 |
| Средняя. Стебли сильно наклонены. Механизированная уборка возможна при наличии специальных приспособлений для уборки полеглых посевов, но при этом возможны потери урожая. | 3 |
| Слабая. Стебли наклонены в слабой степени, как правило, местами. При механизированной уборке затруднения невелики, и потери урожая несрезанными колосьями не бывает. | 4 |
| Полегания нет. Посевы неполеглые. | 5 |
Для предупреждения полегания растений необходимо предпринимать следующие меры: соблюдение оптимальных норм высева семян и недопущение загущения посевов, соблюдение норм внесения азотных удобрений и недопущение перекорма азотом, соблюдение норм полива для культур, возделываемых в условиях орошения.
Для предотвращения полегания растений высокорослых сортов зерновых культур рекомендуется использовать ретарданты, например, хлорхолинхлорид (ССС) и его аналоги. Обычно обработку зерновых культур проводят в начале выхода в трубку. Эти соединения замедляют рост стебля, способствуют накоплению целлюлозы и лигнина, утолщению клеточной стенки. В результате утолщается стебель и повышается его механическая прочность.
Реферат на тему Приспособление растений к водному режиму
Содержание:
- Характеристика основных групп растений по отношению к воде
- Анатомо-морфологические приспособления растений к водному режиму
- Физиологические адаптации растений, приуроченных к местообитаниям разной увлажненности
- Заключение
| Предмет: | Ботаника |
| Тип работы: | Реферат |
| Язык: | Русский |
| Дата добавления: | 04. 01.2019 01.2019 |
- Данный тип работы не является научным трудом, не является готовой работой!
- Данный тип работы представляет собой готовый результат обработки, структурирования и форматирования собранной информации, предназначенной для использования в качестве источника материала для самостоятельной подготовки учебной работы.
Если вам тяжело разобраться в данной теме напишите мне в whatsapp разберём вашу тему, согласуем сроки и я вам помогу!
По этой ссылке вы сможете найти рефераты по биологии на любые темы и посмотреть как они написаны:
Посмотрите похожие темы возможно они вам могут быть полезны:
Введение:
Вода является самым важным экологическим фактором для всей жизни на Земле. Для осуществления обменных процессов с окружающей средой, составляющей основу жизни, необходимо участие воды как растворителя и продукта обмена веществ. Так, в растениях вода участвует в реакции фотосинтеза, а неорганические соли поступают в растение только в виде раствора воды из почвы.
Так, в растениях вода участвует в реакции фотосинтеза, а неорганические соли поступают в растение только в виде раствора воды из почвы.
Вода является основным компонентом растительного тела. Даже в состоянии анабиоза растения содержат воду. Особую роль воды наземных растений играет постоянное пополнение больших затрат на испарение за счет развития больших фотосинтетических поверхностей. Вода, вызывающая необходимый тургор, определенным образом участвует в поддержании формы наземных растений как живых организмов, не имеющих опорного скелета. А для большой группы растений, населяющих водоемы, моря и океаны, вода является непосредственной средой обитания.
Характеристика основных групп растений по отношению к воде
Среди наземных растений различают три основных экологических типа по различным условиям увлажнения и развитию соответствующих приспособлений, а также по близости к месту обитания: гигрофиты, мезофиты и ксерофиты.
Растения, вода которых является не только необходимым экологическим фактором, но и непосредственным местом обитания, относятся к водным растениям и называются гидрофобными.
Гигрофиты. Это растения в чрезмерно влажных местообитаниях с повышенной влажностью воздуха и почвы. Благодаря достаточно широкому разнообразию местообитаний, особенностям водоемов, анатомо-морфологическим особенностям, все гигрофиты являются травянистыми растениями и эпифитами влажных тропических лесов, которые не переносят заметного снижения влажности воздуха, а также отсутствия устройств, ограничивающих водопотребление.
Даже в разгар сезона дождей мелкие эпифиты на стволах деревьев теряют свой тургор и высыхают, когда на них в течение 2-3 часов падают солнечные лучи. Отличительной особенностью Гигрофитов являются травянистые хвойные леса (щавель, майника двулопастный, двулопастный Альпийский). К гигрофитам относятся также виды, произрастающие в открытых и светлых местообитаниях, но в местах, где грунтовые воды выходят наружу, вблизи водоема, в дельте реки наступает состояние избыточной влажности почвы и в наших широтах примером могут служить прибрежные виды: Caltha palaris, Плакучая трава-Lyustrum salikaria, Ясима, в жарком климате–папирус, яшии. Из культурных растений можно включить рис, выращенный на полях, заполненных водой.
Из культурных растений можно включить рис, выращенный на полях, заполненных водой.
Ксерофиты. Это растения засушливой среды обитания, способные выдержать значительный дефицит воды в почве и атмосферную засуху. Они распространены, многочисленны и разнообразны в районах с жарким и сухим климатом. К этой группе относятся пустыни, засушливые степи, саванны, колючие леса и сухие субтропические виды. В более влажных районах ксерофиты участвуют в растительном покрове только в наиболее нагретых и наименее влажных местообитаниях (например, на склонах южной экспозиции).
Неблагоприятный водный режим растения в засушливой среде обитания, во-первых, при недостатке влаги в почве, а во-вторых, при высокой сухости воздуха и транспирации при высоких температурах, поэтому для преодоления недостатка влаги возможны различные способы: увеличить ее поглощение, уменьшить расход и, кроме того, способность переносить большие потери воды. Все это используется ксерофитами при адаптации к сухости, но разные растения имеют разную степень, поэтому некоторые авторы предполагают, что засуха может быть вызвана ксерофитами.
В зависимости от особенностей строения и способов регулирования водного режима ксерофиты (Хенкель П. А. выделяют несколько типов): эвксениты, ремиксирующие, Поэсилотерии. В группу ксерофитов также входят суккуленты с сочными листьями и стеблями. Есть листовые суккуленты (агавы, алоэ) и стеблевые, листья редуцированы,а наземная часть представлена мясистыми стеблями (кактус, некоторые молочаи).
Вид с наиболее выраженными гетероморфными особенностями строения листьев имел уникальный внешний вид, за что и получил название склеродермии. Появление типичной склеродермии легко представить на примере чертополоха-Carduus crispus и пустынной полыни, ковыля, саксаула.
К этой группе относятся растения, произрастающие в условиях умеренной влажности, в том числе луговые растения, лесные травы, лиственные и кустарниковые виды из районов с умеренной влажностью климата, а также большинство культурных растений.
Мезофиты очень разнообразны не только с точки зрения видового состава, но и с точки зрения различных экологических оттенков, обусловленных различным сочетанием факторов в их естественной среде обитания. есть виды с повышенной влажностью, предпочитающие влажные или временно затопляемые участки (степной лисохвост-alopecurus pratensis, бекмания обыкновенная-beckmannia eruciformis).
есть виды с повышенной влажностью, предпочитающие влажные или временно затопляемые участки (степной лисохвост-alopecurus pratensis, бекмания обыкновенная-beckmannia eruciformis).
Они объединяются в переходную группу гигромезофитов вместе с некоторыми влаголюбивыми лесными травами, которые предпочитают наиболее сырые леса, лесные ущелья (Impatiens nolitangere). С другой стороны, в местообитаниях с перемежающимся или стойким (малым) недостатком влаги многие группы мезодермы со специфической атипией с повышенной физиологической устойчивостью к засухе мигрировали между промежуточными остистыми клетками и пигментными колюче-пластиформными остистыми. Примерами могут служить многие виды северных степей, сухих сосновых лесов и песчаных местообитаний: Клевер белоголовый-Trifolium montanum, подмаренник желтый-Galium verum и др.
Особое место среди Мезофитов занимают эфемериды и эфемероиды весной в степях и пустынях. К этой группе относятся растения, покрывающие Луга и пустыни красочными цветочными коврами (многолетники-тюльпаны, гуси-Луки, однолетники — маки, вероники) ранней весной. Это виды с очень короткими вегетационными периодами и длительными периодами покоя, имеющие опыт однолетних эфемероидов, таких как семена, и многолетних эфемероидов — в виде спящих луковиц, клубней, кроме весенних есть еще и осенние эфемероиды, произрастающие в регионах со средиземноморским климатическим ритмом. Это включает в себя род Крокус, Сцилла, и других видов.
Это виды с очень короткими вегетационными периодами и длительными периодами покоя, имеющие опыт однолетних эфемероидов, таких как семена, и многолетних эфемероидов — в виде спящих луковиц, клубней, кроме весенних есть еще и осенние эфемероиды, произрастающие в регионах со средиземноморским климатическим ритмом. Это включает в себя род Крокус, Сцилла, и других видов.
По многим особенностям строения и физиологии растения испытывают недостаток влаги, который близок к нулевому Фитоценозу и связан по каким-то причинам с действием низких температур. Иногда такие виды включаются как особое подразделение ксерофитной группы, а иногда их выделяют в самостоятельные экологические типы (сапрофиты и мерзлые грибы), также Психрофиты-в прохладную среду обитания, влажные, холодные почвы растений высокогорий и северных широт. Несмотря на достаточное увлажнение почвы, им часто не хватает влаги (из-за физиологической сухости, вызванной низкими температурами, или из-за землистых травянистых растений, таких как торфяные почвы (например, северные луговые травы: Белоус-Nardus strikta; высокогорные травы белые), высокогорные, Фестовые и тундровые кустарники и вечнозеленые кустарники (call-evergreen). к психрофилам относятся хвойные породы умеренных и северных широт.
к психрофилам относятся хвойные породы умеренных и северных широт.
Экологически очень близок к психотическим грибам, связан с ними переходной формой. Это растения сухой и холодной среды обитания, сухие участки тундры, скалы и чешуи. Как обычно и предполагают, он характеризуется психрофитом не из-за его многочисленных сходных морфологических и физиологических особенностей. Однако некоторые холодные растения также имеют очень своеобразную форму.
Это гидрофильное вещество. Это водные растения. Согласно их образу жизни и структуре, они содержат погруженные в воду растения и растения с плавающими листьями. Погруженные растения делятся на укореняющиеся в донной почве и подвешенные в толще воды. Из высших растений первым является телорез — Stratiotes aloides, а водным шильником — Subularia aquatika. В эту группу также входят водоросли, которые прилипают к Земле. Среди растений, подвешенных в толще воды, можно упомянуть погруженные в воду роговые водоросли — ceratophyllum demelthum, обыкновенную пузырчатку — Utrikularia vulgaris и многочисленные виды плавающих водорослей.
Растения с плавающими листьями частично используют воду, частично — воздух. Из них в землю пускают корни водяные лилии рода Nymphaea, стручки рода Nuphar, rdesta и водяные орехи Trapa natans.
Многие виды, наряду с плавающими листьями на поверхности воды, также встречаются в воде. Ряска и водокрас плавают на поверхности воды, не приживаясь.
Гелофиты или амфибии — группа растений земноводных, которые очень тесно связаны с реальными водными растениями, обычно рассматриваются вместе с ними. Это типы прибрежных и прибрежных местообитаний с избыточным или изменчивым увлажнением. Они могут расти как в воздухе, так и частично в воде, выдерживая полное временное наводнение. Как и в природе, здесь нет резкой границы между водными и сухопутными местообитаниями для растений, поэтому группа герофитов, с одной стороны, имеет реальные гидрофильные, а с другой-примеры наземных Гелофитов-растения прибрежной зоны пресноводных водоемов и рек: Стрелец тонтифолия, ежеголовка-сагоголовка-Sparganium ramosum.
Анатомо-морфологические приспособления растений к водному режиму
Растения, ограниченные ареалом обитания в различных увлажнительных условиях, выработали соответствующую адаптацию к водному режиму.
Характерной структурной особенностью Гигрофитов является тонкая и мягкая листовая пластинка с небольшим количеством пор, не имеющих толстой кутикулы, и рыхлый слой листовой ткани с большим межклеточным пространством.
Для ксерофитов большое значение имеют различные структурные приспособления к состоянию недостатка влаги.
Корневая система обычно сильно развита, что помогает растению увеличить поступление почвенной влаги. По общей массе корневая система ксерофитов часто превышает надземную часть, иногда очень крупную. Например, многие виды трав и кустарников в пустынях Средней Азии имеют подземные массы в 9-10 раз больше, чем наземные массы, в то время как корневая система кеслофитов в холодных пустынях Памирского нагорья часто является типом, то есть растениями с длинными корнями, которые распространяются по большим массам почвы, но относительно далеко расходятся. При более глубоком проникновении таких корней сухие грибы могут использовать влагу в глубоких слоях почвы, а в некоторых случаях и грунтовые воды.
При более глубоком проникновении таких корней сухие грибы могут использовать влагу в глубоких слоях почвы, а в некоторых случаях и грунтовые воды.
У других видов корневая система имеет интенсивный тип: они покрывают сравнительно небольшое количество почвы, но за счет очень плотного ветвления максимизируют влажность почвы. Корни многих аэробных видов имеют специальные приспособления для хранения влаги.
Наземные органы ксерофитов также отличаются уникальным (так называемым гетеротипическим признаком), который несет на себе отпечаток сложных условий водоснабжения. Они имеют высокоразвитую систему водоснабжения, что хорошо видно по плотности венозной сети листьев, которые снабжают водой ткани. Эта особенность позволяет легко пополнять запасы влаги, которые ксерофиты используют для испарения.
Различные конструктивные устройства защитного характера, направленные на снижение водопотребления, сводятся в основном к следующему:
- Общее уменьшение поверхности транспирации.
 Многие ксерофиты имеют небольшие, узкие и очень редуцированные листовые пластинки. В особо сухих пустынных местообитаниях листья некоторых древесных и кустарниковых пород редуцируются до едва заметных чешуек. У таких видов фотосинтез осуществляется зелеными ветвями.
Многие ксерофиты имеют небольшие, узкие и очень редуцированные листовые пластинки. В особо сухих пустынных местообитаниях листья некоторых древесных и кустарниковых пород редуцируются до едва заметных чешуек. У таких видов фотосинтез осуществляется зелеными ветвями. - Уменьшение листовой поверхности в самые жаркие и засушливые периоды вегетации. Для многих кустарников Средней Азии, Северной Африки и других пустынь, а также для некоторых видов средиземноморских сухих субтропиков характерны сезонные варианты листьев.
- Защита листьев от больших потерь влаги при транспирации. Это достигается за счет развития мощной покровной ткани (толстостенного, иногда многослойного эпидермиса), а в различных выростах и других видах, которые часто образуют толстые «войлочные» удары по поверхности листа, поверхность кутикулы покрывается толстым слоем или восковым гидроизоляционным покрытием.
 Многие ксерофиты имеют небольшие, узкие и очень редуцированные листовые пластинки. В особо сухих пустынных местообитаниях листья некоторых древесных и кустарниковых пород редуцируются до едва заметных чешуек. У таких видов фотосинтез осуществляется зелеными ветвями.
Многие ксерофиты имеют небольшие, узкие и очень редуцированные листовые пластинки. В особо сухих пустынных местообитаниях листья некоторых древесных и кустарниковых пород редуцируются до едва заметных чешуек. У таких видов фотосинтез осуществляется зелеными ветвями.Развитие защитного покрова листьев является причиной того, что луговая трава имеет тусклый сероватый оттенок, который значительно отличается от ярко-зеленого луга. Поры ксерофитов обычно защищены от чрезмерной потери влаги, например, расположены в специальном углублении ткани листа, иногда с помощью волос и других дополнительных защитных приспособлений.
Поры ксерофитов обычно защищены от чрезмерной потери влаги, например, расположены в специальном углублении ткани листа, иногда с помощью волос и других дополнительных защитных приспособлений.
Ковили и другие степные травы обладают интересным механизмом защиты устьиц в самое жаркое и сухое время суток:при большой потере воды, во влажную погоду эпидермиса клетки эпидермиса восстанавливают выпуклость, и лопасти листьев снова разворачиваются.
Повышенное развитие механической ткани. Клетки ткани листа ксерофитов имеют небольшие размеры и очень плотную упаковку, то есть малую генерацию межклеточных клеток, что значительно уменьшает внутреннее испарение поверхности листа. Ксерофиты обычно живут в открытых, светлых местообитаниях, поэтому многие особенности ксероморфного строения листьев являются также особенностями светлого строения. Так, у многих видов листья часто имеют мощную, иногда многорядную частоколообразную паренхиму, расположенную с обеих сторон.
Основными морфологическими и эктопическими признаками Мезофитов были средние значения между гигрофитами и ксофитами. Мезофиты умеренно развивают широкий и интенсивный тип корневой системы и имеют все переходы между ними. Листья характеризуются дифференцировкой тканей на рыхлую губчатую паренхиму с системой более или менее плотных палисадных паренхим и просветов. Сеть вен относительно разрежена. Покровная ткань имеет признаки отдельной атипичной формы, но не выражена, как при сухом дерматите.
Мезофиты умеренно развивают широкий и интенсивный тип корневой системы и имеют все переходы между ними. Листья характеризуются дифференцировкой тканей на рыхлую губчатую паренхиму с системой более или менее плотных палисадных паренхим и просветов. Сеть вен относительно разрежена. Покровная ткань имеет признаки отдельной атипичной формы, но не выражена, как при сухом дерматите.
Ментальная зародышевая клетка имеет сильную гетерогенную структуру листа. Таким образом, кишечнополостные являются узколистными, хорошо развитыми проводящими и механическими тканями. Вечнозеленые кустарники имеют плотные кожистые листья, иногда с очень сильной кутикулой, плотный частокол паренхимы. Нижняя сторона, которая поддерживает поры, часто покрыта слоем полового созревания или воска. Однако некоторые влажные особенности отчетливо прослеживаются в строении листьев, главным образом в развитии больших межклеточных пространств в больших клеточных размерах и губчатых тканях.
Анатомо-морфологические особенности водных растений существенно отличают их от наземных растений. Уменьшение механической ткани заметно. Проводящие системы уменьшаются так же сильно. Если земля мезодермы имеет длину жилок на 1 кв. из см листьев около 100 мм и более, у сухих грибов она достигает 300, А у водных и прибрежных растений в несколько раз меньше. Вот некоторые примеры.
Уменьшение механической ткани заметно. Проводящие системы уменьшаются так же сильно. Если земля мезодермы имеет длину жилок на 1 кв. из см листьев около 100 мм и более, у сухих грибов она достигает 300, А у водных и прибрежных растений в несколько раз меньше. Вот некоторые примеры.
У некоторых погруженных растений, не прикрепленных к земле, корни полностью редуцируются, остальные корни сохраняются, но отдельные плавающие части растения можно обойтись и без них. Однако многие виды имеют толстые и крепкие корневища, которые играют роль якорей, резервов и вегетативных органов.
Листья погруженных в воду водных растений очень тонкие и нежные и имеют упрощенную мезофильную структуру без выраженной дифференциации на частокол и губчатую паренхиму. Листья в воде без устьиц. В некоторых местах имеется группа клеток эпидермиса с тонкими стенками. Считается, что они играют большую роль в усвоении воды и растворенных минеральных солей.
У растений, которые лишь частично погружены в воду, гетерофилл хорошо выражен разница в строении листьев на поверхности одна и та же особь и в воде. Первый имеет общую черту в листьях второго наземного растения — очень тонкую или рассеченную листовую пластинку. Гетерофилия наблюдалась у водяных кимбана, водяных лилий и стручков, стреловидных и других видов. Интересным примером является поручейник, где можно увидеть несколько форм листьев, представляющих все переходы от типичной суши к воде.
Первый имеет общую черту в листьях второго наземного растения — очень тонкую или рассеченную листовую пластинку. Гетерофилия наблюдалась у водяных кимбана, водяных лилий и стручков, стреловидных и других видов. Интересным примером является поручейник, где можно увидеть несколько форм листьев, представляющих все переходы от типичной суши к воде.
Физиологические адаптации растений, приуроченных к местообитаниям разной увлажненности
Помимо морфологических особенностей растения, оно ограничено, развито и физиологично в местах с повышенной влажностью в различных условиях.
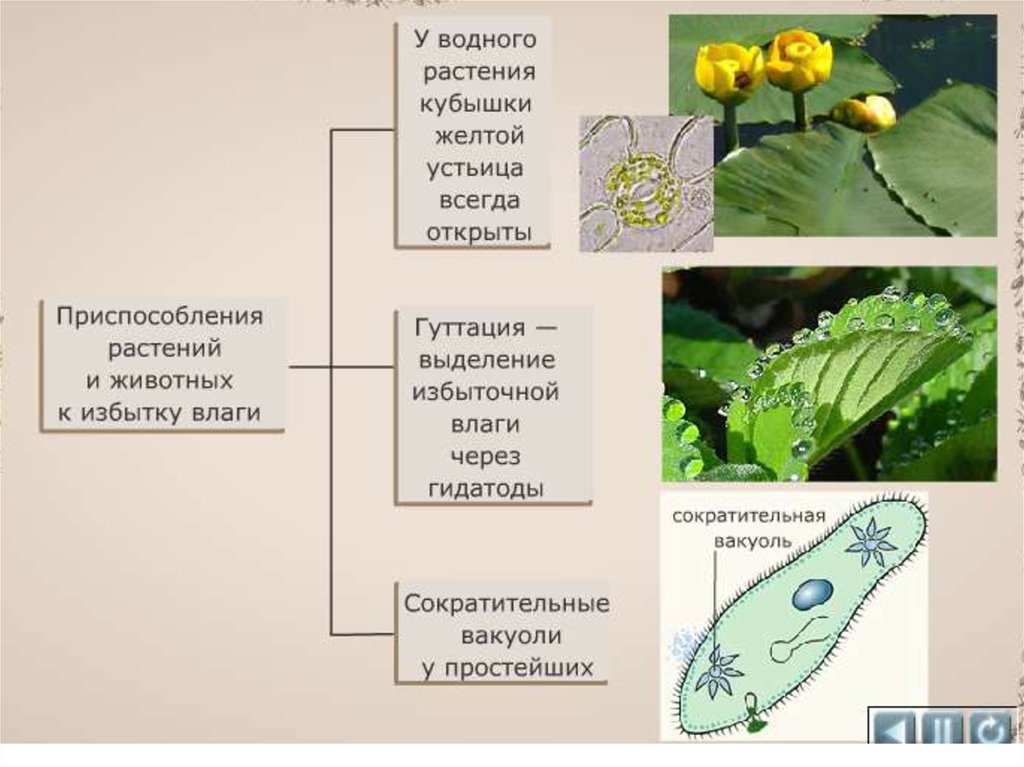
Способность гигроскопичности регулировать режим воды ограничена:устьица в основном широко открыты, поэтому транспирация мало отличается от физического испарения. Благодаря беспрепятственному поступлению воды и отсутствию защитных приспособлений интенсивность транспирации очень высока:у легких гигрофитов в дневное время листья могут терять в весе в 4-5 раз больше, чем за время полива, а высокое содержание воды в тканях Гигрофитов поддерживается главным образом за счет постоянного поступления влаги из окружающей среды.
Другой характерной физиологической особенностью гигрофитов, из-за ее легкого использования влаги, является низкое осмотическое давление в клеточном соке, что приводит к быстрой потере водоподготовки, низкой водоудерживающей способности. Например, в случае кислицы и майника потеря от 15% до 20% подачи воды уже необратима и приводит к гибели людей. В некоторых случаях растения в очень влажной среде обитания нуждаются в удалении лишней влаги. Обычно это происходит, когда почва теплая,корни активно впитывают воду,а транспирации нет (например, утро или туман с влажностью 100%). Избыток влаги удаляется путем выделения воды через специальные выделительные ячейки, расположенные по краям или на кончиках листьев.
Ксерофиты имеют много различных физиологических приспособлений, которые могут успешно противостоять недостатку влаги.
Ксерофиты обычно повышают осмотическое давление клеточных соков, что позволяет им поглощать воду даже при большом водоотводящем усилии в почве.
 Она измеряется в тысячах кПа, а некоторые пустынные кустарники регистрируют регистрационный номер, который достигает от 10000 до 30000 кПа.
Она измеряется в тысячах кПа, а некоторые пустынные кустарники регистрируют регистрационный номер, который достигает от 10000 до 30000 кПа.В течение длительного времени внимание было привлечено к вопросу потребления воды сухими батареями для испарения. Представляется, что значительное уменьшение транспирации должно способствовать большому количеству анатомических приспособлений, надежно защищающих наземную часть ксерофитов от сильного испарения. Но оказывается, что это не совсем так. При достаточном запасе воды большинство сухих грибов обладают достаточно высокой транспирацией, но с наступлением сухих условий они значительно ее снижают. При этом играет роль и закрытие пор, и начало сильного обезвоживания подводнии листа. Несомненно, анатомо-морфологические приспособления имеют определенное значение, но главная роль в устойчивости к высыханию сухих грибов в настоящее время отводится физиологическим механизмам.
Эти механизмы включают высокую водоудерживающую способность тканей и клеток, обусловленную многими физиологическими и биохимическими особенностями.
Большое значение для выживания ксерофитов при недостатке влаги имеет способность переносить глубокое обезвоживание тканей без потери жизнеспособности, а при возобновлении благоприятных условий растения — ксерофиты могут потерять 75% общего запаса воды, но все же жить.
Другая адаптивная система, обеспечивающая выживание ксерофитов в засушливых условиях, заключается в использовании растениями наиболее благоприятных периодов года для вегетации, а в районах с ярко выраженным средиземноморским климатом с сухим периодом лета за счет жизнедеятельности во время засух многие виды имеют «двухтактный» ритм аэробного сезонного развития: во время весенних дождей; в течение лета происходит регенерация растительности; в течение лета следует зимний отдых. Аналогичное явление наблюдается у сухих луговых растений в середине и конце лета: потеря части листовой поверхности, приостановка развития, сильное обезвоживание тканей и др.
Физиологическая адаптация суккулентов очень своеобразна и требует отдельного рассмотрения.
Основным способом преодоления условий высыхания суккулентов является накопление большого количества воды в ткани и использование ее очень экономно. В жарком и сухом климате весь запас воды может быть быстро израсходован, но растение имеет защитное устройство для уменьшения испарения. Одна из них — своеобразная форма надземной части суккулентов. Кроме того, многие суккуленты имеют поверхность, защищенную восковым покрытием витилиго, но суккуленты с тонким незащищенным эпидермисом также имеют очень мало пор и часто погружаются в ткани листьев или стеблей. В течение дня устьица обычно закрыты, и потеря воды проходит главным образом через покровную ткань.
Транспирация суккулентов очень низкая. Его трудно поймать за короткий промежуток времени и необходимо определить расход воды. Водоудерживающая способность мясистых тканей была значительно выше, чем у других растений экологической группы, за счет содержания в клетках гидрофильных веществ. Поэтому, даже не имея доступа к влаге, суккуленты потребляют воду очень медленно и продолжают жить в течение длительного времени даже в растительных образцах.
Ограничения, обусловленные особенностями водного режима суккулентов, вызывают и другие трудности для жизни этих растений в сухих условиях. Слабая транспирация сводит к минимуму возможность терморегуляции, которая связана с сильным нагреванием огромных наземных органов суккулентов. Поскольку устьица обычно закрыты днем и открыты ночью, близость углекислого газа и света не совпадает во времени, возникают также трудности для фотосинтеза. Поэтому суккуленты разработали особый метод фотосинтеза, при котором продукты дыхания частично используются в качестве источника углекислого газа. Иными словами, в экстремальных ситуациях растения частично используют принцип замкнутой системы с повторным использованием метаболических отходов. Из-за всех этих ограничений интенсивность фотосинтеза у суккулентов низкая, их рост и накопление массы очень медленные,поэтому они не обладают высокой биологической продуктивностью и могут быть использованы для закрытых растений.
Физиологические показатели подтверждают их промежуточное положение: они характеризуются умеренным значением осмотического давления, количеством воды в листьях и маргинальным дефицитом воды. Что касается транспирации, то ее величина в большей степени зависит от условий освещения и других элементов микроклимата.
Что касается транспирации, то ее величина в большей степени зависит от условий освещения и других элементов микроклимата.
Одни и те же среднетемпературные виды, попадая в различные условия водоснабжения, проявляют определенную пластичность, приобретают более глубокие свойства в сухих условиях, обладают более влажными свойствами во влажных условиях.
Пластичность листьев проявляется не только в разных местообитаниях, но даже у одних и тех же особей. Например, у дерева на опушке леса листья на стороне, обращенной к лесу, имеют более умеренный и затененный характер, по сравнению со слегка деформированными листьями на внешней стороне дерева. Разные высотные слои листьев одного и того же растения находятся в разных условиях полива, так как приток воды к вершине связан с преодолением большого сопротивления. Кроме того, верхние листья дерева обычно находятся в другом микроклимате.
Различие анатомо-физиологических показателей листьев на разных сторонах кроны дерева, растущего на опушке леса.
Поскольку водная среда существенно отличается от воздушной, водные растения обладают многими уникальными физиологическими адаптивными функциями. Интенсивность света в воде сильно ослаблена, так как часть падающего излучения отражается от поверхности воды, а другие поглощаются ее толщей. Из-за ослабления света фотосинтез у растений в воде значительно уменьшается с увеличением глубины. Выживание глубоководного фитопланктона в зоне, где освещенность ниже точки коррекции, обеспечивается периодическим вертикальным перемещением в Верхнюю зону, где происходит интенсивный фотосинтез и пополнение органического вещества.
В воде, помимо недостатка света, растения могут испытывать еще одну трудность, необходимую для фотосинтеза-недостаток доступного CO2. Углекислый газ поступает в воду в результате растворения СО2, содержащегося в воздухе,дыхания водных организмов, разложения органических остатков и выделения из них карбонатов. Вот почему при интенсивном фотосинтезе растений потребление CO2 увеличивается, и его легко не хватает.
Гидрофиты реагируют на увеличение содержания СО2 в воде значительным увеличением фотосинтеза.
В погруженном растении нет испарения, поэтому нет «верхнего двигателя», который поддерживает поток воды растения. Однако этот поток, снабжающий ткани питательными веществами, присутствует с ежедневной частотой. Активная роль в его поддержании принадлежит корневому давлению и активности особых клеток, выделяющих воду-водяных устьиц.
Заключение
Плавающие или выступающие листья обычно имеют сильную транспирацию, но они непосредственно соприкасаются с водой и располагаются в воздушном слое с высокой влажностью. Поры широко открыты и полностью закрываются только ночью.
Транспирация одинаково высока у прибрежных растений, у которых значительное количество воды потребляется не только листьями, но и стеблями.
Осмотическое давление в водных и прибрежных растениях очень низкое, так как нет необходимости преодолевать водоудерживающую способность почвы при поглощении воды.
Примеры осмотического давления (в кПа) у водных растений и гелофитов (по Гесснеру Ф.,1959).
Адаптация растений к засухе
1. Брей ЕА: Реакция растений на дефицит воды. Trends Plant Sci. 1997;2(2):48–54. 10.1016/С1360-1385(97)82562-9 [CrossRef] [Google Scholar]
2. Дай А: Усиление засухи при глобальном потеплении в наблюдениях и моделях. Нат Клим Чанг. 2012;3:52–8. 10.1038/nclimate1633 [CrossRef] [Google Scholar]
3. Бонерт Х.Дж., Нельсон Д.Э., Дженсен Р.Г.: Адаптации к экологическим стрессам. Растительная клетка. 1995;7(7):1099–111. 10.1105/тпк.7.7.1099 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
4. Левитт Дж.: Реакции растений на стрессы окружающей среды. Том 2. Вода, радиация, соль и другие стрессы. (Академическая пресса), 1980. Справочный источник [Google Scholar]
5.
Джонс М.М., Тернер Н.К., Осмонд С.Б.:
Механизмы засухоустойчивости. Физиология и биохимия засухоустойчивости растений. 1981; 1:15–37. [Google Scholar]
1981; 1:15–37. [Google Scholar]
6. Морган Дж. М.: Осморегуляция и водный стресс у высших растений. Annu Rev Plant Physiol. 1984; 35: 299–319. 10.1146/аннурев.стр.35.060184.001503 [CrossRef] [Google Scholar]
7. Гафф Д.Ф.: Устойчивые к засухе растения Южной Америки. Экология. 1987;74(1):133–6. 10.1007/BF00377357 [PubMed] [CrossRef] [Google Scholar]
8. Гафф Д.Ф.: Устойчивые к засухе цветковые растения на юге Африки. Наука. 1971; 174 (4013): 1033–4. 10.1126/наука.174.4013.1033 [PubMed] [CrossRef] [Академия Google]
9. Порембски С: Обнажения гранитов и гнейсов (инзельберги) как центры разнообразия устойчивых к высыханию сосудистых растений. Завод Экол. 2000;151(1):19–28. 10.1023/А:1026565817218 [CrossRef] [Google Scholar]
10.
Чонди Б., Фукай С., Купер М.:
Водный потенциал листьев и осмотическая адаптация как физиологические признаки для улучшения засухоустойчивости риса. Полевые культуры Res. 2002;76(2–3):153–63.
10.1016/С0378-4290(02)00036-9
[CrossRef] [Google Scholar]
2002;76(2–3):153–63.
10.1016/С0378-4290(02)00036-9
[CrossRef] [Google Scholar]
11. Пантуван Г., Фукай С., Купер М. и др.: Урожайность риса ( Oryza sativa L.) генотипы к засухе на богарных низинах: 3. Растительные факторы, способствующие засухоустойчивости. Полевые культуры Res. 2002;73(2–3):181–200. 10.1016/С0378-4290(01)00194-0 [CrossRef] [Google Scholar]
12. Венупрасад Р., Лафит Х.Р., Атлин Г.Н.: Ответ на прямой отбор по урожайности зерна при стрессе от засухи у риса. Растениеводство. 2007;47(1):285–293. 10.2135/cropsci2006.03.0181 [CrossRef] [Google Scholar]
13. Гуан Ю.С., Серрадж Р., Лю С.Х. и др.: Одновременное повышение урожайности в условиях засухи и в нестрессовых условиях: пример риса ( Oryza sativa L.). J Опытный бот. 2010;61(15):4145–56. 10.1093/jxb/erq212 [PubMed] [CrossRef] [Google Scholar]
14.
Чон Дж.С., Ким Ю.С., Редиллас М.С. и др.: Сверхэкспрессия OsNAC5 увеличивает диаметр корня у растений риса, что приводит к повышенной засухоустойчивости и повышению урожайности зерна в поле. Завод Биотехнолог J. 2013;11(1):101–14.
10.1111/пби.12011
[PubMed] [CrossRef] [Google Scholar]
Завод Биотехнолог J. 2013;11(1):101–14.
10.1111/пби.12011
[PubMed] [CrossRef] [Google Scholar]
15. Redillas MC, Jeong JS, Kim YS и др.: Гиперэкспрессия OsNAC9 изменяет строение корней растений риса, повышая засухоустойчивость и урожайность зерна в полевых условиях. Завод Биотехнолог J. 2012;10(7):792–805. 10.1111/j.1467-7652.2012.00697.x [PubMed] [CrossRef] [Google Scholar]
16. Чон Дж.С., Ким Й.С., Пэк К.Х. и др.: Специфическое для корня выражение OsNAC10 повышает засухоустойчивость и урожайность риса в условиях полевой засухи. Физиол растений. 2010;153(1):185–97. 10.1104/стр.110.154773 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
17. Амбаварам М.М., Басу С., Кришнан А. и др.: Скоординированная регуляция фотосинтеза у риса повышает урожайность и устойчивость к стрессам окружающей среды. Нац.коммун. 2014;5:5302. 10.1038/ncomms6302 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
18. Рамеговда В., Басу С., Кришнан А. и др.:
Рис РОСТ В УСЛОВИЯХ ЗАСУХИ КИНАЗА необходима для обеспечения засухоустойчивости и урожайности зерна в нормальных условиях и в условиях засушливого стресса. Физиол растений. 2014;166(3):1634–45.
10.1104/стр.114.248203
[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Рамеговда В., Басу С., Кришнан А. и др.:
Рис РОСТ В УСЛОВИЯХ ЗАСУХИ КИНАЗА необходима для обеспечения засухоустойчивости и урожайности зерна в нормальных условиях и в условиях засушливого стресса. Физиол растений. 2014;166(3):1634–45.
10.1104/стр.114.248203
[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
19. Йорданов И, Великова В, Цонев Т: Реакция растений на засуху, акклиматизация и стрессоустойчивость. Фотосинтетика. 2000;38(2):171–86. 10.1023/А:1007201411474 [CrossRef] [Google Scholar]
20. Брей ЕА: Молекулярные реакции на дефицит воды. Физиол растений. 1993;103(4):1035–40. [Бесплатная статья PMC] [PubMed] [Google Scholar]
21. Осакабе Ю., Осакабе К., Шинозаки К. и др.: Реакция растений на водный стресс. Front Plant Sci. 2014;5:86. 10.3389/fpls.2014.00086 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
22.
Рейнольдс М., Тубероза Р.:
Трансляционные исследования, влияющие на продуктивность сельскохозяйственных культур в засушливых условиях. Curr Opin Plant Biol. 2008;11(2):171–9.
10.1016/j.pbi.2008.02.005
[PubMed] [CrossRef] [Google Scholar]
Curr Opin Plant Biol. 2008;11(2):171–9.
10.1016/j.pbi.2008.02.005
[PubMed] [CrossRef] [Google Scholar]
23. Тезара В., Митчелл В.Дж., Дрисколл С.Д. и др.: Водный стресс ингибирует фотосинтез растений за счет уменьшения фактора связи и АТФ. Природа. 1999; 401:914–7. 10.1038/44842 [CrossRef] [Google Scholar]
24. Лоулор Д.У., Корник Г.: Фотосинтетическая ассимиляция углерода и связанный с ней метаболизм в связи с дефицитом воды у высших растений. Окружающая среда растительных клеток. 2002;25(2):275–94. 10.1046/j.0016-8025.2001.00814.x [PubMed] [CrossRef] [Google Scholar]
25. Нийоги К.К.: ПЕРЕСМОТР ФОТОЗАЩИТЫ: Генетические и молекулярные подходы . Annu Rev Plant Physiol Plant Mol Biol. 1999;50:333–59. 10.1146/аннурев.арзавод.50.1.333 [PubMed] [CrossRef] [Google Scholar]
26.
Деммиг-Адамс Б., Адамс В.В., III:
Роль каротиноидов ксантофиллового цикла в защите фотосинтеза. Trends Plant Sci. 1996;1(1):21–6.
10.1016/С1360-1385(96)80019-7
[CrossRef] [Google Scholar]
1996;1(1):21–6.
10.1016/С1360-1385(96)80019-7
[CrossRef] [Google Scholar]
27. Деммиг-Адамс Б., Адамс В.В., 3-й: Фотозащита в экологическом контексте: замечательная сложность рассеивания тепловой энергии. Новый Фитол. 2006;172(1):11–21. 10.1111/j.1469-8137.2006.01835.х [PubMed] [CrossRef] [Google Scholar]
28. Лоулор Д.У.: Ограничение фотосинтеза в листьях, подверженных водному стрессу: устьица против . Метаболизм и роль АТФ. Энн Бот. 2002; 89 (спец. №): 871–85. 10.1093/аоб/mcf110 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
29. Medrano H, Parry MAJ, Socias X и др.: Длительный водный стресс инактивирует Rubisco в подземном клевере. Энн Appl Biol. 1997;131(3):491–501. 10.1111/j.1744-7348.1997.tb05176.x [CrossRef] [Google Scholar]
30.
Chaves MM, Flexas J, Pinheiro C:
Фотосинтез в условиях засухи и солевого стресса: механизмы регуляции от всего растения к клетке. Энн Бот. 2009;103(4):551–60. 10.1093/аоб/mcn125
[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
10.1093/аоб/mcn125
[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
31. Эдвардс Г., Уокер Д.: C3, C4: механизмы, клеточная и экологическая регуляция фотосинтеза. Калифорнийский университет Press, 1983. Справочный источник [PubMed] [Академия Google]
32. Говик У, Вестхофф П.: Путь от С. 3 до С 4 фотосинтез. Физиол растений. 2011;155(1):56–63. 10.1104/стр.110.165308 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
33. Пинейро С, Чавиш М.М.: Фотосинтез и засуха: можем ли мы установить метаболические связи на основе имеющихся данных? J Опытный бот. 2011;62(3):869–82. 10.1093/jxb/erq340 [PubMed] [CrossRef] [Google Scholar]
34. Уилкинсон С., Кудоярова Г.Р., Веселов Д.С. и др.: Взаимодействие гормонов растений: инновационные цели для селекции и управления сельскохозяйственными культурами. J Опытный бот. 2012;63(9):3499–509. 10.1093/jxb/ers148 [PubMed] [CrossRef] [Google Scholar]
35. Уилкинсон С., Дэвис В.Дж.:
Засуха, озон, ABA и этилен: новые идеи от клетки к растению и сообществу. Окружающая среда растительных клеток. 2010;33(4):510–25.
10.1111/j.1365-3040.2009.02052.x
[PubMed] [CrossRef] [Google Scholar]
Уилкинсон С., Дэвис В.Дж.:
Засуха, озон, ABA и этилен: новые идеи от клетки к растению и сообществу. Окружающая среда растительных клеток. 2010;33(4):510–25.
10.1111/j.1365-3040.2009.02052.x
[PubMed] [CrossRef] [Google Scholar]
36. Ji X, Dong B, Shiran B и др.: Контроль катаболизма абсцизовой кислоты и гомеостаза абсцизовой кислоты важен для стрессоустойчивости репродуктивной стадии у злаков. Физиол растений. 2011;156(2):647–62. 10.1104/стр.111.176164 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
37. Сперотто Р.А., Рикаченевский Ф.К., Дуарте Г.Л. и др.: Идентификация генов с повышенной регуляцией во флаговых листьях во время наполнения рисового зерна и характеристика Os NAC5, новый АБК-зависимый фактор транскрипции. Планта. 2009;230(5):985–1002. 10.1007/s00425-009-1000-9 [PubMed] [CrossRef] [Google Scholar]
38.
Лян С, Ван И, Чжу И и др.:
OsNAP связывает абсцизовую кислоту и старение листьев путем точной настройки биосинтеза абсцизовой кислоты и прямого воздействия на гены, связанные со старением, у риса. Proc Natl Acad Sci U S A. 2014;111(27):10013–8.
10.1073/пнас.1321568111
[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Proc Natl Acad Sci U S A. 2014;111(27):10013–8.
10.1073/пнас.1321568111
[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
39. Chen X, Wang Y, Lv B и др.: Фактор транскрипции семейства NAC OsNAP обеспечивает реакцию на абиотический стресс через путь ABA. Физиол клеток растений. 2014;55(3):604–19. 10.1093/pcp/pct204 [PubMed] [CrossRef] [Google Scholar]
40. Ду Х, Ван Н, Цуй Ф и др.: Характеристика гена бета-каротингидроксилазы DSM2 , придающий рису устойчивость к засухе и окислительному стрессу за счет увеличения синтеза ксантофиллов и абсцизовой кислоты. Физиол растений. 2010;154(3):1304–18. 10.1104/стр.110.163741 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
41.
Пелег З., Регера М., Тумимбанг Э. и др.:
Цитокинин-опосредованные модификации источника/поглотителя улучшают устойчивость к засухе и повышают урожайность зерна риса в условиях водного стресса. Завод Биотехнолог J. 2011;9(7):747–58. 10.1111/j.1467-7652.2010.00584.x
[PubMed] [CrossRef] [Академия Google]
10.1111/j.1467-7652.2010.00584.x
[PubMed] [CrossRef] [Академия Google]
42. Пелег З., Блюмвальд Э.: Гормональный баланс и устойчивость к абиотическим стрессам у сельскохозяйственных культур. Curr Opin Plant Biol. 2011;14(3):290–5. 10.1016/j.pbi.2011.02.001 [PubMed] [CrossRef] [Google Scholar]
43. Се Зи, Цзян Д, Цао В и др.: Взаимосвязь эндогенных фитогормонов с накоплением белка и крахмала в зерне озимой пшеницы при различном постантезисном водном статусе почвы. Регулятор роста растений. 2003;41(2):117–27. 10.1023/А:1027371
9 [Перекрестная ссылка] [Академия Google]44. Чжан С., Ли С., Цао Дж. и др.: Измененная архитектура и повышенная засухоустойчивость риса за счет подавления индол-3-уксусной кислоты с помощью TLD1/OsGh4.13 активация. Физиол растений. 2009; 151(4):1889–901. 10.1104/стр.109.146803 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
45.
Уга Ю., Сугимото К., Огава С. и др. :
Управление архитектурой корневой системы с помощью ГЛУБОКОЕ Укоренение 1 повышает урожайность риса в засушливых условиях. Нат Жене. 2013;45(9):1097–102.
10.1038/нг.2725
[PubMed] [CrossRef] [Google Scholar]
:
Управление архитектурой корневой системы с помощью ГЛУБОКОЕ Укоренение 1 повышает урожайность риса в засушливых условиях. Нат Жене. 2013;45(9):1097–102.
10.1038/нг.2725
[PubMed] [CrossRef] [Google Scholar]
46. Ван С, Ян А, Инь Х и др.: Влияние водного стресса на содержание эндогенных гормонов и повреждение клеток проростков кукурузы. J Integr Plant Biol. 2008;50(4):427–34. 10.1111/j.1774-7909.2008.00638.x [PubMed] [CrossRef] [Google Scholar]
47. Мунне-Бош С., Алегре Л.: Умри и дай жить: старение листьев способствует выживанию растений в условиях засухи. Функц. биол. растений. 2004;31(3):203–216. 10.1071/ФП03236 [CrossRef] [Google Scholar]
48. Фукао Т., Сюй К., Рональд П.С. и др.: Вариабельный кластер генов, подобных фактору отклика на этилен, регулирует реакции метаболизма и развития на погружение риса в воду. Растительная клетка. 2006;18(8):2021–34. 10.1105/тпк.106.043000 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
49. Перата П., Воесенек Л.А.:
Устойчивость к погружению в рис требует Sub1A , ген, подобный фактору реакции на этилен. Trends Plant Sci. 2007;12(2):43–6.
10.1016/j.tрастения.2006.12.005
[PubMed] [CrossRef] [Google Scholar]
Перата П., Воесенек Л.А.:
Устойчивость к погружению в рис требует Sub1A , ген, подобный фактору реакции на этилен. Trends Plant Sci. 2007;12(2):43–6.
10.1016/j.tрастения.2006.12.005
[PubMed] [CrossRef] [Google Scholar]
50. Раджала А., Пелтонен-Сайнио П.: Влияние регуляторов роста растений на рост корней и побегов яровых зерновых. Агрон Дж. 2001;93(4):936–943. 10.2134/агронж2001.934936х [CrossRef] [Google Scholar]
51. Резкий RE: Взаимодействие с этиленом: изменение взглядов на роль абсцизовой кислоты в реакциях роста корней и побегов на водный стресс. Окружающая среда растительных клеток. 2002;25(2):211–22. 10.1046/j.1365-3040.2002.00798.x [PubMed] [CrossRef] [Google Scholar]
52. Yang JC, Zhang JH, Ye YX и др.: Участие абсцизовой кислоты и этилена в реакциях зерен риса на водный стресс при наполнении. Окружающая среда растительных клеток. 2004;27(8):1055–64. 10.1111/j.1365-3040.2004.01210.x [CrossRef] [Google Scholar]
53. Гомес-Ролдан В., Фермас С., Брюэр П.Б. и др.:
Ингибирование стриголактоном ветвления побегов. Природа. 2008;455(7210):189–94.
10.1038/природа07271
[PubMed] [CrossRef] [Google Scholar]
Гомес-Ролдан В., Фермас С., Брюэр П.Б. и др.:
Ингибирование стриголактоном ветвления побегов. Природа. 2008;455(7210):189–94.
10.1038/природа07271
[PubMed] [CrossRef] [Google Scholar]
54. Син Ю, Чжан Цюй: Генетические и молекулярные основы урожайности риса. Annu Rev Plant Biol. 2010;61:421–42. 10.1146/annurev-arplant-042809-112209 [PubMed] [CrossRef] [Google Scholar]
55. Сюн Л, Чжу Дж: Молекулярно-генетические аспекты реакции растений на осмотический стресс. Окружающая среда растительных клеток. 2002;25(2):131–9. 10.1046/j.1365-3040.2002.00782.x [PubMed] [CrossRef] [Академия Google]
56. Микко В.Д., Аронн Г.: Реакция растений на стресс от засухи. (изд. Арока. Рикардо). Спрингер, 2002. [Google Scholar]
57.
Миядзава С., Йошимура С., Шинзаки Ю. и др.:
Деактивация аквапоринов снижает внутреннюю проводимость к CO. 2 диффузия в листьях табака, выращенных в условиях длительной засухи. Функц. биол. растений. 2008;35(7):553–564.
10.1071/ФП08117
[CrossRef] [Google Scholar]
растений. 2008;35(7):553–564.
10.1071/ФП08117
[CrossRef] [Google Scholar]
58. Тосенс Т., Ниинеметс У., Вислап В. и др.: Онтогенетические изменения диффузионной проводимости мезофилла и фотосинтетической способности в условиях разной освещенности и водообеспеченности у Populus tremula : как структура ограничивает функцию. Окружающая среда растительных клеток. 2012;35(5):839–56. 10.1111/ж.1365-3040.2011.02457.х [PubMed] [CrossRef] [Google Scholar]
59. Сюй Цзи, Чжоу Г: Реакция плотности устьиц листьев на водный статус и ее связь с фотосинтезом в траве. J Опытный бот. 2008;59(12):3317–25. 10.1093/jxb/ern185 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
60. Блюм А: Засухоустойчивость, эффективность использования воды и потенциал урожайности — совместимы ли они, противоречат друг другу или исключают друг друга? Aust J Agric Res. 2005;56(11):1159–1168. 10.1071/AR05069 [CrossRef] [Google Scholar]
61. Караба А., Диксит С., Греко Р. и др.:
Повышение эффективности использования воды при выращивании риса за счет ХАРДИ , Ген засухо- и солеустойчивости Arabidopsis . Proc Natl Acad Sci U S A. 2007;104(39):15270–5.
10.1073/пнас.0707294104
[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Караба А., Диксит С., Греко Р. и др.:
Повышение эффективности использования воды при выращивании риса за счет ХАРДИ , Ген засухо- и солеустойчивости Arabidopsis . Proc Natl Acad Sci U S A. 2007;104(39):15270–5.
10.1073/пнас.0707294104
[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
62. Spollen WG, Sharp RE: Пространственное распределение тургора и роста корней при низком водном потенциале. Физиол растений. 1991;96(2):438–43. 10.1104/стр.96.2.438 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
63. Дик К.И., Малами Дж.: Осмотическая регуляция архитектуры корневой системы. Завод Ж. 2005;43(1):17–28. 10.1111/j.1365-313X.2005.02425.x [PubMed] [CrossRef] [Google Scholar]
64. Со ПиДжей, Пак Си: Гомеостаз ауксинов при развитии боковых корней в условиях засухи. Поведение сигнала установки. 2009;4(10):1002–4. 10.4161/псб.4.10.9716 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
65. Чен Х, Ли Зи, Сюн Л:
Растительная микроРНК регулирует адаптацию корней к засушливому стрессу. ФЭБС Письмо. 2012;586(12):1742–1747.
10.1016/j.febslet.2012.05.013
[PubMed] [CrossRef] [Google Scholar]
Чен Х, Ли Зи, Сюн Л:
Растительная микроРНК регулирует адаптацию корней к засушливому стрессу. ФЭБС Письмо. 2012;586(12):1742–1747.
10.1016/j.febslet.2012.05.013
[PubMed] [CrossRef] [Google Scholar]
66. Джаффе М.Дж., Такахаши Х., Биро Р.Л.: Мутант гороха для изучения гидротропизма корней. Наука. 1985; 230(4724):445–7. 10.1126/наука.230.4724.445 [PubMed] [CrossRef] [Академия Google]
67. Такахаси Н., Ямадзаки Ю., Кобаяши А. и др.: Гидротропизм взаимодействует с гравитропизмом, разрушая амилопласты в корнях проростков арабидопсиса и редьки. Физиол растений. 2003;132(2):805–10. 10.1104/стр.102.018853 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
68. Блилоу И., Сюй Дж., Уайлдуотер М. и др.: Сеть посредников оттока ауксинов PIN контролирует рост и формирование паттерна в Корни арабидопсиса . Природа. 2005;433(7021):39–44. 10.1038/природа03184 [PubMed] [CrossRef] [Google Scholar]
69. Сенгупта Д., Редди А.Р.:
Дефицит воды как регуляторный переключатель реакции корней бобовых культур. Поведение сигнала установки. 2011;6(6):914–7.
10.4161/псб.6.6.15340
[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Сенгупта Д., Редди А.Р.:
Дефицит воды как регуляторный переключатель реакции корней бобовых культур. Поведение сигнала установки. 2011;6(6):914–7.
10.4161/псб.6.6.15340
[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
70. Диксит С., Кумар Бисвал А., Мин А. и др.: Действие нескольких генов внутри QTL, объединенных вокруг совместно локализованного транскрипционного фактора, лежит в основе QTL с большим эффектом. Научный представитель 2015;5: 15183. 10.1038/srep15183 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
71. Генри А., Кэл А.Дж., Батото Т.С. и др.: Корневые атрибуты, влияющие на водопоглощение риса ( Oryza sativa ) в условиях засухи. J Опытный бот. 2012;63(13):4751–63. 10.1093/jxb/ers150 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
72.
Хуанг Б., Фрай Д.Д.:
Анатомические, физиологические и морфологические реакции корней на стресс от засухи для сортов высокорослых овсяниц. Растениеводство. 1998;38(4):1017–1022.
10.2135/cropsci1998.0011183X003800040022x
[Перекрестная ссылка] [Академия Google]
Растениеводство. 1998;38(4):1017–1022.
10.2135/cropsci1998.0011183X003800040022x
[Перекрестная ссылка] [Академия Google]
73. Штойдл Э: Поглощение воды корнями: последствия водного дефицита. J Опытный бот. 2000;51(350):1531–42. 10.1093/jexbot/51.350.1531 [PubMed] [CrossRef] [Google Scholar]
74. Чавиш М.М., Оливейра М.М.: Механизмы, лежащие в основе устойчивости растений к дефициту воды: перспективы водосберегающего сельского хозяйства. J Опытный бот. 2004;55(407):2365–84. 10.1093/jxb/erh369 [PubMed] [CrossRef] [Google Scholar]
75. Мейер Р.Ф., Бойер Дж.С.: Чувствительность клеточного деления и удлинения клеток к низкому водному потенциалу в гипокотилях сои. Планта. 1972;108(1):77–87. 10.1007/BF00386508 [PubMed] [CrossRef] [Google Scholar]
76.
Серрадж Р., Синклер Т.Р.:
Накопление осмолитов: действительно ли это может помочь увеличить урожай в условиях засухи? Окружающая среда растительных клеток. 2002;25(2):333–41.
10.1046/j.1365-3040.2002.00754.x
[PubMed] [CrossRef] [Google Scholar]
2002;25(2):333–41.
10.1046/j.1365-3040.2002.00754.x
[PubMed] [CrossRef] [Google Scholar]
77. Oosterhuis DM, Wullschleger SD: Осмотическая регулировка хлопка ( Gossypium hirsutum L.) Листья и корни в ответ на водный стресс. Физиол растений. 1987;84(4):1154–1157. 10.1104/стр.84.4.1154 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
78. Заутер А., Дэвис В.Дж., Хартунг В.: Дальний сигнал абсцизовой кислоты в засушливом растении: судьба гормона на пути от корня к побегу. J Опытный бот. 2001; 52(363):1991–7. 10.1093/jexbot/52.363.1991 [PubMed] [CrossRef] [Google Scholar]
79. Ашраф М., Фулад М.Р.: Роль глицина бетаина и пролина в повышении устойчивости растений к абиотическим стрессам. Бот Environment Exp. 2007;59(2):206–16. 10.1016/j.envexpbot.2005.12.006 [CrossRef] [Google Scholar]
80.
Ю Л.Х., Ву С.Дж., Пэн Ю.С. и др.: Арабидопсис EDT1 / HDG11 улучшает засухо- и солеустойчивость хлопка и тополя и повышает урожайность хлопка в поле. Завод Биотехнолог J. 2016;14(1):72–84.
10.1111/pbi.12358
[PubMed] [CrossRef] [Google Scholar]
Завод Биотехнолог J. 2016;14(1):72–84.
10.1111/pbi.12358
[PubMed] [CrossRef] [Google Scholar]
81. He X, Chen Z, Wang J и др.: Сахароза: ген фруктан-6-фруктозилтрансферазы (6-SFT) из Psathyrostachys huashanica придает табаку устойчивость к абиотическому стрессу. Ген. 2015;570(2):239–47. 10.1016/j.gene.2015.06.023 [PubMed] [CrossRef] [Google Scholar]
82. Гэ Л, Чао Д, Ши М и др.: Сверхэкспрессия гена трегалозо-6-фосфатфосфатазы OsTPP1 придает рису устойчивость к стрессу и приводит к активации генов, чувствительных к стрессу. Планта. 2008;228(1):191–201. 10.1007/s00425-008-0729-х [PubMed] [CrossRef] [Академия Google]
83. Ан С., Андерсон Дж. А., Сорреллс М. Е. и др.: Гомеологические отношения хромосом риса, пшеницы и кукурузы. Мол Ген Генет. 1993;241(5–6):483–90. [PubMed] [Google Scholar]
84.
Хирота О, Ока М, Такеда Т:
Оценка активности стока по размеру стока и увеличению сухого вещества на стадии созревания ячменя ( Hordeum vulgare ) и рис ( Oryza sativa ). Энн Бот. 1990;65(4):349–353.
Справочный источник
[Академия Google]
Энн Бот. 1990;65(4):349–353.
Справочный источник
[Академия Google]
85. Гладун И.В., Карпов Е.А.: Образование и распределение ассимилятов между метелкой и вегетативными органами риса после цветения. Русс Ж Завод Физиол. 1993;40(5):629-633. Справочный источник [Google Scholar]
86. Li X, Lawas LMF, Malo R и др.: Метаболические и транскриптомные сигнатуры цветочных органов риса показывают, что сахарное голодание является фактором репродуктивной недостаточности в условиях стресса от жары и засухи. Окружающая среда растительных клеток. 2015;38(10):2171–92. 10.1111/шт.12545 [PubMed] [CrossRef] [Академия Google]
87. Нгуен Г.Н., Хейлстоунс Д.Л., Уилкс М. и др.: СТРЕСС ЗАСУХИ: роль метаболизма углеводов в вызванной засухой мужской стерильности в пыльниках риса. J Agron Crop Sci. 2010;196(5):346–57. 10.1111/j.1439-037X.2010.00423.x [CrossRef] [Google Scholar]
88.
Шеоран И.С., Сайни Х.С.:
Вызванное засухой мужское бесплодие у риса: изменения уровня углеводов и активности ферментов, связанные с ингибированием накопления крахмала в пыльце. Репродукция полового растения. 1996;9(3):161–9.
10.1007/BF02221396
[CrossRef] [Google Scholar]
Репродукция полового растения. 1996;9(3):161–9.
10.1007/BF02221396
[CrossRef] [Google Scholar]
89. Джин И, Ян Х, Вэй Зи и др.: Развитие самцов риса в условиях засушливого стресса: фенотипические изменения и стадийно-зависимое транскриптомное перепрограммирование. Мол Завод. 2013;6(5):1630–45. 10.1093/мп/сст067 [PubMed] [CrossRef] [Google Scholar]
90. Тубероза Р: Фенотипирование засухоустойчивости сельскохозяйственных культур в эпоху геномики. Фронт Физиол. 2012;3:347. 10.3389/ффиз.2012.00347 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
91. Нгуен Т.Т., Клюева Н., Чамарек В. и др.: Картирование насыщения областей QTL и идентификация предполагаемых генов-кандидатов на устойчивость к засухе у риса. Мол Генет Геномикс. 2004;272(1):35–46. 10.1007/s00438-004-1025-5 [PubMed] [CrossRef] [Google Scholar]
92.
Ду Х, Ву Н, Фу Дж и др.:
Член семейства Gh4, OsGh4-2, модулирует уровни ауксина и абсцизовой кислоты и по-разному влияет на устойчивость риса к засухе и холоду. J Опытный бот. 2012;63(18):6467–80.
10.1093/jxb/ers300
[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
J Опытный бот. 2012;63(18):6467–80.
10.1093/jxb/ers300
[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
93. Oh SJ, Kim YS, Kwon CW и др.: Сверхэкспрессия фактора транскрипции AP37 в рисе повышает урожайность зерна в условиях засухи. Физиол растений. 2009;150(3):1368–79. 10.1104/стр.109.137554 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
94. He Y, Hao Q, Li W и др.: Идентификация и характеристика рецепторов АБК в Oryza sativa . PLoS Один. 2014;9(4):e95246. 10.1371/journal.pone.0095246 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
95. Guo C, Luo C, Guo L и др.: OsSIDP366 , ген DUF1644, позитивно регулирует реакцию риса на засуху и солевой стресс. J Integr Plant Biol. 2016;58(5):492–502. 10.1111/jipb.12376 [PubMed] [CrossRef] [Google Scholar]
96.
Сюн Х., Ли Дж., Лю П. и др.:
Гиперэкспрессия OsMYB48-1 , новый фактор транскрипции, связанный с MYB, повышает устойчивость риса к засухе и засолению. PLoS Один. 2014;9(3):e92913.
10.1371/journal.pone.0092913
[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
PLoS Один. 2014;9(3):e92913.
10.1371/journal.pone.0092913
[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
97. Че М., Ли Дж. С., Нам М. Х. и др.: Протеинкиназа 2, родственная SNF1, индуцируемая дегидратацией риса, фосфорилирует фактор связывания элемента, чувствительного к абсцизовой кислоте, и связывается с передачей сигналов ABA. Растительный Мол Биол. 2007;63(2):151–69. 10.1007/s11103-006-9079-х [PubMed] [CrossRef] [Google Scholar]
98. Чжан С., Хайдер И., Колен В. и др.: Функция гена HD-Zip I Oshox22 в опосредованной АБК засухе и солеустойчивости риса. Растительный Мол Биол. 2012;80(6):571–85. 10.1007/s11103-012-9967-1 [PubMed] [CrossRef] [Google Scholar]
99. Ху Х, Дай М, Яо Дж и др.: Сверхэкспрессия факторов транскрипции NAM, ATAF и CUC (NAC) повышает засухоустойчивость и солеустойчивость риса. Proc Natl Acad Sci U S A. 2006;103(35):12987–92. 10.1073/пнас.0604882103 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
100. Hu H, You J, Fang Y и др.:
Характеристика гена фактора транскрипции SNAC2 , придающий рису устойчивость к холоду и соли. Растительный Мол Биол. 2008;67(1–2):169–81.
10.1007/с11103-008-9309-5
[PubMed] [CrossRef] [Google Scholar]
Hu H, You J, Fang Y и др.:
Характеристика гена фактора транскрипции SNAC2 , придающий рису устойчивость к холоду и соли. Растительный Мол Биол. 2008;67(1–2):169–81.
10.1007/с11103-008-9309-5
[PubMed] [CrossRef] [Google Scholar]
101. Ю Дж, Ху Х, Сюн Л: Орнитин δ -ген аминотрансферазы OsOAT придает рису устойчивость к засухе и окислительному стрессу. Науки о растениях. 2012;197:59–69. 10.1016/j.plantsci.2012.090,002 [PubMed] [CrossRef] [Google Scholar]
102. Сян И, Тан Н, Ду Х и др.: Характеристика OsbZIP23 как ключевого игрока семейства основных факторов транскрипции лейциновой молнии для придания рису чувствительности к абсцизовой кислоте, солености и засухоустойчивости. Физиол растений. 2008; 148 (4): 1938–52. 10.1104/стр.108.128199 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
103.
Джиша В., Дампанабойна Л., Вадассери Дж. и др.:
Сверхэкспрессия фактора транскрипции типа AP2/ERF OsEREBP1 Придает рису устойчивость к биотическим и абиотическим стрессам. PLoS Один. 2015;10(6):e0127831.
10.1371/журн.pone.0127831
[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
PLoS Один. 2015;10(6):e0127831.
10.1371/журн.pone.0127831
[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
104. Лю С., Мао Б., Оу С. и др.: OsbZIP71, фактор транскрипции bZIP, придает рису устойчивость к засолению и засухе. Растительный Мол Биол. 2014;84(1–2):19–36. 10.1007/с11103-013-0115-3 [PubMed] [CrossRef] [Академия Google] F1000 Рекомендация
105. Тан Н, Чжан Х, Ли Х и др.: Конститутивная активация фактора транскрипции OsbZIP46 улучшает устойчивость риса к засухе. Физиол растений. 2012;158(4):1755–68. 10.1104/стр.111.1
[Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]106. Цзоу М., Гуан И, Рен Х и др.: Фактор транскрипции bZIP, OsABI5, участвует в фертильности и стрессоустойчивости риса. Растительный Мол Биол. 2008;66(6):675–83. 10.1007/s11103-008-9298-4 [PubMed] [CrossRef] [Google Scholar]
107.
Йошида Т., Фудзита Ю., Саяма Х. и др.:
AREB1, AREB2 и ABF3 являются основными факторами транскрипции, которые совместно регулируют ABRE-зависимую передачу сигналов ABA, участвующую в устойчивости к засухе, и требуют ABA для полной активации..jpg) Завод Ж. 2010;61(4):672–85.
10.1111/j.1365-313X.2009.04092.x
[PubMed] [CrossRef] [Google Scholar]
Завод Ж. 2010;61(4):672–85.
10.1111/j.1365-313X.2009.04092.x
[PubMed] [CrossRef] [Google Scholar]
108. You J, Zong W, Li X и др.: Ген, нацеленный на SNAC1 OsSRO1c модулирует закрытие устьиц и устойчивость к окислительному стрессу, регулируя перекись водорода в рисе. J Опытный бот. 2013;64(2):569–83. 10.1093/jxb/ers349 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
109. Цай С., Цзян Г., Е Н. и др.: Ключевой катаболический ген АБК, OsABA8ox3 участвует в обеспечении устойчивости риса к засухе. PLoS Один. 2015;10(2):e0116646. 10.1371/journal.pone.0116646 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Адаптация растений к избытку или дефициту воды. – Живая природа
Растениям требуется вода почти на всех этапах их жизненного цикла:
- Семенам требуется влага для прорастания и они останутся в состоянии покоя, если упадут на сухую почву. Эта потребность во влаге может быть вызвана необходимостью выщелачивания ингибиторов роста из семенной кожуры (особенно у пустынных растений, где ингибиторы препятствуют прорастанию после небольшого ливня и задерживают его до сильных зимних дождей), чтобы смягчить твердую оболочку семени, чтобы корень корня мог появляться.
 Эта потребность во влаге может быть вызвана необходимостью выщелачивания ингибиторов роста из семенной кожуры (особенно у пустынных растений, где ингибиторы препятствуют прорастанию после небольшого ливня и задерживают его до сильных зимних дождей), чтобы смягчить твердую оболочку семени, чтобы корень корня мог появляться.
Эта потребность во влаге может быть вызвана необходимостью выщелачивания ингибиторов роста из семенной кожуры (особенно у пустынных растений, где ингибиторы препятствуют прорастанию после небольшого ливня и задерживают его до сильных зимних дождей), чтобы смягчить твердую оболочку семени, чтобы корень корня мог появляться.- Рост растений требует влаги, так как при недостаточном увлажнении газообмен в листве резко снижается. Растения регулируют потерю воды листьями за счет образования воскового поверхностного слоя (кутикулы), который почти непроницаем для воды. Однако кутикула также довольно непроницаема для СО2 и О2, таким образом, проблемы были решены с развитием устьиц (крохотных отверстий или пор, используемых для газообмена), которые соединяют атмосферу листа с внешней атмосферой.
- С другой стороны, активное поглощение влаги необходимо, так как транспирационный поток транспортирует питательные вещества от корней к листьям. Растения забирают воду из почвы до тех пор, пока не исчезнет градиент натяжения влаги между растением и почвой. Транспирация создает водный стресс в растении, что приводит к напряжению влаги внутри корней.
 Растения забирают воду из почвы до тех пор, пока не исчезнет градиент натяжения влаги между растением и почвой. Транспирация создает водный стресс в растении, что приводит к напряжению влаги внутри корней.
Растения забирают воду из почвы до тех пор, пока не исчезнет градиент натяжения влаги между растением и почвой. Транспирация создает водный стресс в растении, что приводит к напряжению влаги внутри корней.- Когда почва влажная, вода поступает в корни в ответ на это напряжение. По мере высыхания почвы градиент между почвой и растением уменьшается, и поглощается меньше воды. Когда почва высыхает до такой степени, что натяжение влаги в почве приближается к напряжению влаги в растениях, поглощение внезапно резко снижается, растения увядают, устьица закрываются и транспирация прекращается. Потенциал влажности почвы, при котором это происходит, известен как точка временного увядания . Если водный потенциал почвы еще больше уменьшится, растения увядают до точки, при которой они не восстанавливают тургор (постоянная точка увядания).
- Количество воды в почве (% по весу) в точке постоянного увядания называется «% постоянного увядания (PW%)». Это сильно зависит от текстуры почвы, потому что текстура определяет натяжение, с которым оставшаяся вода удерживается в капиллярах по мере высыхания почвы.
 Это сильно зависит от текстуры почвы, потому что текстура определяет натяжение, с которым оставшаяся вода удерживается в капиллярах по мере высыхания почвы.
Это сильно зависит от текстуры почвы, потому что текстура определяет натяжение, с которым оставшаяся вода удерживается в капиллярах по мере высыхания почвы.- Хотя PW% практически одинаков для всех растений (таким образом, его можно рассматривать как параметр почвы, а не параметр растения), могут быть небольшие различия , которые могут обеспечить растение достаточным количеством влаги, чтобы поддерживать его жизнь в течение 6 лет. полных дней, что может иметь значение между выживанием и гибелью растения.
Растения можно классифицировать в соответствии с их адаптацией к водному стрессу.
- Гидрофиты : приспособлены для частичного или полного погружения в свободную воду. Они не могут расти при напряжении почвенной влаги более 500-1000 кПа.
- Мезофиты: наземные растения, приспособленные к умеренному водоснабжению. Они могут расти при напряжении почвенной влаги до 2000 кПа. У них отсутствуют специализированные приспособления к избытку или недостатку влаги.
 У них отсутствуют специализированные приспособления к избытку или недостатку влаги.
У них отсутствуют специализированные приспособления к избытку или недостатку влаги.- Ксерофиты : адаптированы к засушливым зонам, могут расти при давлении влаги до 4000 кПа. Для них характерны разнообразные ксероморфные и физиологические приспособления.
A) ИЗБЫТОЧНАЯ ВЛАГА
Термин гидрофит включает водные растения, произрастающие на болотах, болотах и топях. Негидрофитные растения обычно исключаются из этих насыщенных полуземных местообитаний из-за дефицита кислорода в почве. Они развивают некоторые из следующих приспособлений:
- Внутренние полости и губчатые ткани , которые облегчают перемещение кислорода от листьев и стеблей к корням, что позволяет им функционировать в среде с недостатком кислорода. Хорошим примером этого является рисовое растение 9.0003 (Oryza sativa) , у которого погруженные корни могут содержать до 18% O2, тогда как илистый субстрат не содержит его.

- Ограничение роста корней лучше аэрируемой поверхностью почвы.
- Развитие пневматофоров: особые корневые ответвления, растущие вертикально вверх до выхода в воздух и обычно снабженные устьицами и хорошо развитой межклеточной системой воздушных пространств.
- Способность дышать анаэробно в течение ограниченного периода времени.
- В целом низкая потребность в кислороде.
B) ДЕФИЦИТ ВЛАГИ
Ксерофиты можно классифицировать в зависимости от того, как они реагируют на недостаток влаги:
- Засухоустойчивые растения
- Засухоустойчивые: Предотвращение стресса от засухи (экономия и расход воды) и устойчивость к стрессу от засухи (предотвращение обезвоживания, устойчивость к обезвоживанию).
1) Засухокрылые: эфемеры
Они устраняют большую часть или все свои функции воды и приостанавливают метаболические функции, которые зависят от воды. Они могут сделать это, войдя в стадию семян или спор. На самом деле, в засушливых регионах обычно произрастает большое разнообразие небольших однолетних видов растений, которые цветут только в дождливые сезоны. У них отсутствуют ксероморфные признаки, поэтому они не переносят почвенную засуху. Их основными характеристиками являются небольшой размер, высокое соотношение корней и побегов и способность быстро завершать свой цикл (несколько недель после периода дождей).
Они могут сделать это, войдя в стадию семян или спор. На самом деле, в засушливых регионах обычно произрастает большое разнообразие небольших однолетних видов растений, которые цветут только в дождливые сезоны. У них отсутствуют ксероморфные признаки, поэтому они не переносят почвенную засуху. Их основными характеристиками являются небольшой размер, высокое соотношение корней и побегов и способность быстро завершать свой цикл (несколько недель после периода дождей).
Следует отметить, что семена большинства растений устойчивы к засухе, что не все однолетники могут выжить в засушливых условиях и что большинство пустынных эфемеров не растут за пределами засушливых регионов.
2) Засухоустойчивые.
2.1.) Предотвращение стресса от засухи
Это включает поддержание высокого водного потенциала, когда растение подвергается воздействию внешнего водного стресса. Существуют две основные адаптации: водосбережение (водосберегающие) и использование воды (водорасходатели).
Те, кто экономит воду, могут терять всего лишь 1/4300 своего веса воды в день, в то время как те, кто тратит воду, могут терять в пять раз больше своего веса воды в час. Поэтому те, кто тратит воду, могут терять воду в 500 000 раз быстрее, чем те, кто экономит воду.
2.1.1) Водосберегающие : лучшим примером являются суккуленты (кактусы…) и некоторые склерофиллы. Оба способны ограничивать транспирацию задолго до того, как происходит увядание.
Суккуленты
- Суккуленты избегают засухи, поглощая большое количество воды и накапливая ее в набухших парехиматозных тканях и вакуолях.
- Их корни имеют тенденцию быть очень поверхностными, чтобы использовать даже небольшие осадки.
- Корни часто чувствительны к засухе, отмирают вскоре после дождя и вновь отрастают, когда появляется влага.
- Потеря воды исключена, так как устьица открываются только ночью и ненадолго утром. CO2, необходимый в течение дня для фотосинтеза, получается из органических кислот, накапливаемых ночью.
- Потери воды также снижаются благодаря очень малому отношению площади поверхности к объему.
- Листья редуцируются или даже исчезают, растение представляет собой просто вздутый стебель.
- Потеря воды в результате повреждения кутикулы травоядными животными предотвращается за счет развития острых шипов и химической защиты.

Несуккулентные водосберегающие растения: включают засухоустойчивые хвойные и вечнозеленые покрытосеменные растения.
- Непроницаемая кутикула. Кутикулярная транспирация составляет гораздо меньшую долю (от 1/5 до 1/50 устьичной транспирации), чем у мезофитов (от 1/2 до 1/5 устьичной транспирации). По мере потери воды транспирация кутикулы снижается до тех пор, пока не станет незначительной, прежде чем произойдет увядание.
- Отложение дополнительных липидов в кутикуле способствует замедлению транспирации кутикулы.
2.1.2) Водопоглотители : ускоренное водопоглощение позволяет поддерживать высокое внутреннее содержание воды, несмотря на высокую интенсивность транспирации ксерофитными водопоглотителями. Это позволяет им держать устьица открытыми в течение дня, и они связаны с более высокими темпами фотосинтеза и роста, чем сохраняет вода. У них есть следующие приспособления, которые позволяют этому явно расточительному расходованию воды:
Это позволяет им держать устьица открытыми в течение дня, и они связаны с более высокими темпами фотосинтеза и роста, чем сохраняет вода. У них есть следующие приспособления, которые позволяют этому явно расточительному расходованию воды:
- Жилки листа расположены ближе друг к другу, и плотность жилок выше, чем у мезофитов, что приводит к более быстрой доставке воды к поверхности листа и более высокой скорости транспирации.
- Меньшие листья имеют меньшее сопротивление трению потоку воды, чем большие листья, и могут испаряться быстрее.
- Очень высокое соотношение корней и побегов обеспечивает растения большим запасом влаги в почве, что помогает поддерживать транспирацию.
- Более крупная корневая система может проникать глубоко в почву к источникам воды.
- Пониженная точка постоянного увядания.
Pinus densiflora, , имеющая самое высокое соотношение корней и побегов, является одним из самых засухоустойчивых хвойных деревьев.
Корни деревьев Джарра (Eucalyptus marginata) также были измерены на глубине 15 м.
Eucalyptus marginata встречается в юго-западной части Западной Австралии, как правило, там, где изогиета осадков превышает 600 мм.
Расходующий воду человек может частично смягчить стресс от засухи, поглощая атмосферную влагу или ночную росу непосредственно через листву. Это может быть важно для растений в туманных местах или районах, характеризующихся жаркими, сухими днями и холодными ночами.
Расточители воды также могут экономить воду, когда запасы воды становятся крайне ограниченными.
2.2.) Устойчивость к засухе
Растения либо избегают обезвоживания, либо переносят обезвоживание посредством различных механизмов:
- Высокий уровень растворенных веществ в клеточном соке придает клеткам низкий осмотический потенциал (высокое осмотическое давление), который сохраняет их набухшими даже в условиях засухи, тем самым избегая обезвоживания.
- Смерть от засухи может быть следствием «голодания», сопровождающего ограниченный обмен СО2. Таким образом, устойчивость к засухе может быть достигнута путем поддержания устьиц открытыми, несмотря на пониженное содержание воды. В этих условиях транспирация снижается из-за низкого осмотического потенциала клеток, но обмен СО2 продолжается.
- В качестве альтернативы голодания можно избежать, если снизить скорость метаболизма. Это снижает потребность в поглощении CO2 и продлевает доступный фотосинтез.
- Некоторые растения просто не повреждаются при высыхании. Мхи, лишайники и некоторые папоротники могут оставаться в воздушно-сухих условиях в течение длительного времени без повреждений. Было обнаружено, что папоротник Polypodium polypochoides выжил без повреждений после потери 97% своего нормального содержания воды.
- На засухоустойчивость, как и на устойчивость к низким температурам, влияет предварительное кондиционирование или «закаливание»: воздействие сублетального стресса, которое приводит к устойчивости к смертельному стрессу.
 Мхи, лишайники и некоторые папоротники могут оставаться в воздушно-сухих условиях в течение длительного времени без повреждений. Было обнаружено, что папоротник Polypodium polypochoides выжил без повреждений после потери 97% своего нормального содержания воды.
Мхи, лишайники и некоторые папоротники могут оставаться в воздушно-сухих условиях в течение длительного времени без повреждений. Было обнаружено, что папоротник Polypodium polypochoides выжил без повреждений после потери 97% своего нормального содержания воды.
2.3.) Морфологические и физиологические приспособления к засухе
Общие ксероморфные признаки:
2.3.1) Морфологические признаки
25 9013
Leave A Comment