
1) Наследственную изменчивость Ч. Дарвин называл: а) модификационной; б)групповой; в)
а) модификационной;б)групповой;
в) неопределенной;
г) определенной.
2) Движущая сила эволюции (по Ч. Дарвину) видов — это:
а) искусственный отбор;
б) наследственная изменчивость;
в) популяционные волны;
г) борьба за существование.
3) Материалом для эволюции служит:
а) борьба за существование;
б) естественный отбор;
в) модификационная изменчивость;
г) наследственная изменчивость.
4) Укажите неверное утверждение: «Результат действия естественного отбора — это…»:
а) приспособленность организмов к среде обитания;
б) многообразие органического мира;
в) наследственная изменчивость;
г) образование новых видов.
5)Основная заслуга Ч.Дарвина состоит:
а) в формулировании биогенетического закона;
б) создании первой эволюционной теории;
в) разработка теории естественного отбора;
г) создании закона наследственных рядов.
6) По Ч. Дарвину, движущими силами эволюции являются:
а) борьба за существование;
б) наследственная изменчивость;
в) естественный отбор;
г) все перечисленные.
7) Ведущую роль в эволюции играет следующий вид изменчивости:
а) определенная;
б) модификационная;
в) групповая;
г) мутационная.
8) Способность организмов производить большое количество потомков и ограниченность мест обитания и жизненных ресурсов являются непосредственными причинами:
а) наследственной изменчивости;
б) борьбы за существование;
в) вымирания;
г) видообразования.
9) Наследственная изменчивость в процессе эволюции:
б) доставляет материал для эволюции;
в) закрепляет созданный в процессе эволюции материал;
г) сохраняет наиболее полезные изменения.
10) Результатом действия естественного отбора не является:
а) приспособленность организмов к среде обитания;
б) многообразие органического мира;
в) борьба за существование;
г) совершенствование организации живых существ.
Тест по теме «эволюционное учение»( 11 класс) 1 наследственную изменчивость ч дарвин называл
Тест по теме «Эволюционное учение»( 11 класс)
1. Наследственную изменчивость Ч.Дарвин называл:
а) модификационной
б) групповой
в) неопределённой
г) определенной
2. Движущая сила эволюции (по Дарвину) видов – это:
а) искусственный отбор
б) наследственная изменчивость
в) популяционные волны
г) борьба за существование
3. Материалом для эволюции служит:
а) борьба за существование
б) естественный отбор
в) модификационная изменчивость
г) наследственная изменчивость
4. Укажите неверное утверждение: «Результат действия естественного отбора – это…»
а) приспособленность организмов к среде обитания
б) многообразие органического мира
в) наследственная изменчивость
г) образование новых видов
5. Основная заслуга Дарвина состоит:
а) в формулировании биогенетического закона
б) создании первой эволюционной теории
в) разработке теории естественного отбора
г) создании закона наследственных рядов
6. По Ч.Дарвину, движущими силами эволюции являются:
а) борьба за существование
б) наследственная изменчивость
в) естественный отбор
г) всё перечисленное
7. Ведущую роль в эволюции играет следующий вид изменчивости:
а) определённая
б) модификационная
в) групповая
г) мутационная
8. Способность организмов производить большое количество потомков и ограниченность мест обитания и жизненность ресурсов являются непосредственными причинами:
а) наследственной изменчивости;
б) борьбы за существование;
в) вымирания;
г) видообразования
9. Наследственная изменчивость в процессе эволюции:
а) создает новые виды;
б) доставляет материал для эволюции;
в) закрепляет созданный в процессе эволюции материал;
г) сохраняет наиболее полезные изменения.
10. Результатом действия естественного отбора не является:
а) приспособленность организмов к среде обитания;
б) многообразие органического мира;
в) борьба за существование;
г) совершенствование организации живых существ.
Эволюция Дарвина
Эволюция (лат. evolutio – развертывание, развитие), как принято считать в биологии, это необратимое историческое развитие естественных и искусственных систем. Обычно эволюцию противопоставляли революции – быстрым и значительным по масштабу изменениям. В настоящее время стало ясно, что процесс развития искусственных и естественных систем слагается из изменений как постепенных, так и резких, как быстрых, так и длящихся много поколений.
Основные характерные черты биологической эволюции это: преемственность; возникновение в эволюционном процессе целесообразности; усложнение и совершенствование структур.
Согласно второму началу термодинамики все совершающиеся в природе процессы направлены в сторону разрушения структур, снижению уровня сложности, увеличения доли беспорядка (энтропии) во всех системах. А в процессе эволюции происходит лишь местное усложнение системы, которое достигается ценой лишней затраты энергии на развитие организма.
Впервые термин эволюция был использован в биологии швейцарским ученым Ш. Бонне в 1782 году. Под эволюцией понимают медленные постепенные количественные и качественные изменения объекта. При этом каждое новое состояние объекта должно иметь по сравнению с предыдущим более высокий уровень развития и организации.
Теорию эволюции науку о закономерностях и причинах эволюционного процесса называют эволюционным учением. В настоящее время существует большое число вариантов различных концепций эволюции. Основное их различие в том, какую изменчивость они берут за основу эволюции определенную направленную приспособительную или же неопределенную ненаправленную и оказывающуюся приспособительной только случайно.
В биологии эволюция определяется наследственной изменчивостью, борьбой за существование, естественным и искусственным отбором. Эволюция приводит к формированию адаптации (приспособлений) организмов к условиям их существования, изменению генетического состава от популяции видов, а также отмиранию неприспособленных видов. Под адаптацией понимается процесс приспособления строения и функций организмов и их органов к условиям окружающей среды. В науке под адаптацией понимают процесс накопления и использования информации в системе, направленный на достижение ее (системы) оптимального состояния, при первоначальной неопределенности и изменяющихся внешних условиях. О том, что явление адаптации имеется в живой природе, было известно биологам прошлых веков. В настоящее время генетика или теория генетики утверждает, что адаптация не является какой-то внутренней сущностью, заранее приданной организму, но она всегда возникает и развивается. Такое развитие осуществляется под воздействием четырех основных признаков: наследственности, изменчивости, естественного отбора, искусственного отбора.
С развитием теории эволюции ее идеи все больше используются при моделировании мышления и поведения человека, создании современных компьютеров и т. д. В связи с этим возникают целые новые отрасли знаний и науки. Например,
Важную роль в явлениях наследственности играют хромосомы — нитевидные структуры, находящиеся в клеточном ядре. Каждый вид характеризуется вполне определенным числом хромосом, во всех случаях оно четное, и их можно распределить попарно.
В клетках существует сложный и важный механизм перераспределения генетического материала. Каждая клетка организма имеет одинаковое число хромосом. Потомки имеют то же самое число, причем ровно половина от отца, а половина от матери. При таком обмене передаются наследственные признаки.
Ч. Вильсон в 1900 г. определил, что гены находятся в хромосомах. Две гомологичные хромосомы (одна от отца, а другая от матери) сближаются при созревании половых клеток и обмениваются частям. Это явление назвали кроссинговер. Оно происходит между разными генами случайным образом с разной частотой. Модель хромосомы в настоящее время – это нить, на которую, словно бусины, нанизаны гены.
Ч. Дарвин первый ученый, который определил в живой природе существование общего принципа – естественного отбора. В эволюционном учении различают две стороны: учение о материале для эволюции и учение о ее факторах, ее движущей силе. Движущая сила эволюции естественный отбор. В основе наследования лежат неделимые и несмешиваемые факторы – гены. Именно через отбор и происходит направленное влияние условий жизни на наследственную изменчивость. Сама по себе наследственная изменчивость случайна. Под воздействием окружающей среды отбираются признаки, которые лучше других соответствуют условиям жизни.
Наследственные изменения отдельных генов де Фриз назвал мутациями. Мутации и служат элементарным материалом для эволюционного процесса. Они закономерно возникают в природных условиях.
Изменчивость – разнообразие признаков и свойств у особей и групп особей любой степени родства. Различают изменчивость направленную и ненаправленную.
Направленная, или определенная, изменчивость обычно массовая и приспособительная. В данном случае наследуется не изменение признака, а способность к изменению, но в разной степени. Такие изменения называют модификациями. Определенная изменчивость – продукт эволюции, способность к ней возникает в результате отбора в течение многих поколений. Но само изменение признака под влиянием какого-нибудь фактора внешней среды исчезает с гибелью организма, потомки должны обретать его заново. Только в этом смысле определенная изменчивость ненаследственна. Ее нельзя называть изменчивостью: это наследственность, проявляющаяся в фенотипе не всегда, а лишь при воздействии определенного фактора внешней среды.
Ненаправленная, или неопределенная, изменчивость возникает независимо от природы вызвавшего ее фактора, причем изменяющийся признак может изменяться и в сторону усиления, и в сторону ослабления. При этом она не массовая, а единичная. Различают два типа неопределенной изменчивости комбинативную и мутационную. На основе комбинативной изменчивости при образовании потомства во время мейоза возникают новые сочетания материнских и отцовских хромосом. При этом хромосомы иногда обмениваются частями (кроссинговер), так что число комбинаций генов в каждом новом поколении резко возрастает. Мутационная изменчивость – процесс изменения генетической структуры организма, его генотипа. При этом изменяется число хромосом, или их строение, или же структура слагающих хромосому генов. Как и комбинативная изменчивость, мутационная процесс ненаправленный (признаки могут при ней изменяться случайным образом), немассовый (одновременное возникновение какой-нибудь одной мутации у целого ряда особей в популяции невозможно) и неприспособительный (мутации могут и повышать и понижать жизнеспособность их носителей). Неопределенная изменчивость – материал для процесса эволюции. Изменения организмов, по Ч. Дарвину, определяются факторами внешней среды. При этом с большей вероятностью выживают и оставляют потомство носители полезных в данной среде признаков, возникших в результате мутации или рекомбинации определяющих эти признаки генов.
Понятия ген и генетика ввел датский ученый В. Иогансен. Приведем факторы, которые меняют генетический состав природной популяции: мутационный процесс, изоляция, «волны жизни», отбор.
Особое положение генов состоит в их уникальности. В хромосомном наборе каждый ген представлен только один раз. В 30-е годы было доказано, что генная мутация – это небольшое химическое изменение. Следовательно, ген имеет химическую природу, являясь молекулой или частью большой молекулы.
В 1953 г. с работ М. Уоткена и Ф. Крика началась новая наука – молекулярная генетика. Биохимические особенности живых организмов наследуются по законам, которые открыл Г. Мендель. В генах «записаны» планы строения белков – планы всех наследственных признаков. Генетический код оказался общим для всех естественных систем на нашей планете. Он практически расшифрован. Каждая хромосома уникальна морфологически и генетически и не может быть заменена другой либо восстановлена при утере. При потере хромосомы клетка, как правило, погибает. Каждый биологический вид имеет определенное, постоянное число хромосом. В процессах наследования признаков определяющую роль играет поведение хромосом при делении клеток. Существует два основных типа деления клеток: митоз и мейоз. Митоз – непрямое деление клеток тела, это механизм точного распределения хромосом между двумя образующимися дочерними клетками. Мейоз – механизм редукции (уменьшения числа хромосом вдвое).
Классическая генетика к началу 1940-х годов пришла к пониманию дискретности таких качеств, как наследственность и изменчивость. Это стало возможным в первую очередь благодаря формированию теории гена в работах школы Т. Моргана. Основные положения этой теории можно сформулировать следующим образом:
- все признаки организмов находятся под контролем генов;
- гены – элементарные единицы наследственной информации, они находятся в хромосомах;
- гены могут изменяться – мутировать;
- мутации отдельных генов приводят к изменению отдельных элементарных признаков, или фенов.


Сейчас считают, что ген – реально существующая независимая комбинирующаяся и расщепляющаяся при скрещиваниях единица наследственности, самостоятельно наследующийся наследственный фактор. Ген определяют как структурную единицу наследственной информации, неделимую в функциональном отношении. Его рассматривают как участок молекулы ДНК, кодирующий синтез одной макромолекулы или выполняющий какую-либо другую элементарную функцию. Совокупность генов составляет генотип. Фенотип – совокупность всех внешних и внутренних признаков. Комплекс генов, содержащихся в наборе хромосом одного организма, образует геном.
Основой регулярной (общей) рекомбинации является кроссинговер, т. е. обмен гомологичными участками в различных точках гомологичных хромосом, приводящий к появлению нового сочетания сцепленных генов.
Расщепление при независимом наследовании и при кроссинговере определяет изменчивость организмов вследствие комбинаторики существующих генов (аллелей). Аллелями называют определенное химическое состояние гена. Мутации – это возникновение качественно новых генов (аллелей), хромосом и наборов хромосом. Сочетание обоих типов изменчивости вызывает общую изменчивость генотипа.
Генетический материал обладает такими универсальными свойствами, как дискретность, непрерывность, линейность и относительная стабильность, выявляемыми в ходе генетического анализа. Повышение разрешающей способности генетического анализа возможно с помощью изучения большого числа особей, применения селективных методов, ускорения мутационного процесса.
Все эти методы имеют важное значение в естественных системах и могут найти применение в построении искусственных систем. Увеличение числа особей приводит к росту разнообразия генетического материала, что означает увеличение исходного набора контролируемых генами функций, а это, в свою очередь, позволяет провести более широкий отбор функций. Ускорение мутационного процесса ведет к получению все более разнообразного генетического материала.
Ускорение мутационного процесса ведет к получению все более разнообразного генетического материала.
В естественных и искусственных системах роль мутаций заключается в том, что именно они генерируют новые функции, затем происходит дупликация, закрепляющая обе функции, а после этого начинается отдельная эволюция исходной и новой функции. Эта эволюция и показывает, что новая или возникшая в результате мутации функция обладает более высокими адаптационными качествами либо прежняя функция выполняет эту роль лучше.
В общем же эволюция стремится к усреднению (так как происходит все более однородное смешение разного по качеству с нашей точки зрения генетического материала). Поэтому в качестве одного из методов для получения наилучших результатов развития используется селекция. Селекция представляет собой форму искусственного отбора. Селекция, как наука, создана Ч. Дарвином, который выделял три формы отбора:
- естественный отбор, вызывающий изменения, связанные с приспособлением популяции к новым условиям;
- бессознательный отбор, при котором в популяции сохраняются лучшие экземпляры;
- методический отбор, при котором проводится целенаправленное изменение популяции в сторону установленного идеала.
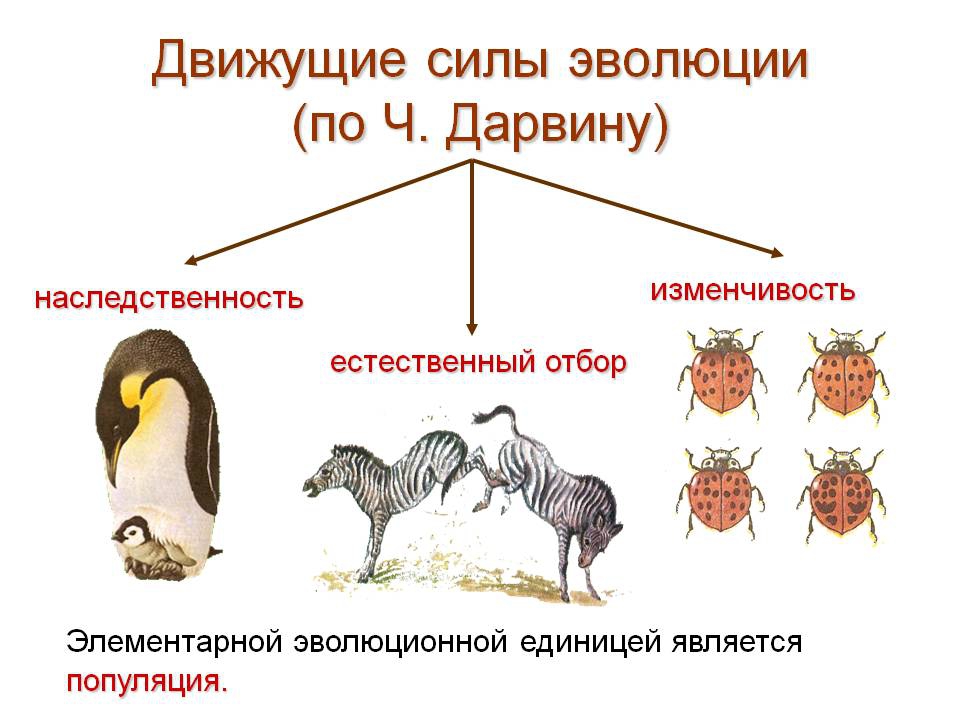
Движущими силами эволюции по Ч. Дарвину являются:
- неопределенная изменчивость, т. е. наследственно обусловленное разнообразие организмов каждой популяции;
- борьба за существование, в ходе которой устраняются от размножения менее приспособленные организмы;
- естественный отбор – выживание более приспособленных особей, в результате которого накапливаются и суммируются полезные наследственные изменения и возникают новые адаптации.
Дарвинизм адаптацию объясняет эволюцией. В результате естественного отбора вновь возникающие мутации комбинируются генами уже прошедших отбор особей, их фенотипическое выражение меняется и на их основе возникают новые адаптации. Следовательно, отбор – основной фактор эволюции, обуславливающий возникновение новых адаптации, преобразование организмов и видообразование. Отбор проявляется в трех основных формах:
Отбор проявляется в трех основных формах:
- движущий (ведущий). Он приводит к выработке новых адаптаций;
- стабилизирующий. Он обеспечивает сохранение в неизменных условиях среды уже сформировавшихся адаптации;
- дизруптивный (разрывающий). Он обуславливает возникновение полиморфизма при разнонаправленных изменениях среды обитания популяции.
Отбор идет по обшей приспособленности организма, а не по какому-нибудь отдельному признаку.
При движущем отборе большую вероятность оставить потомство имеют особи, изменившиеся по каким-нибудь признакам по сравнению со средней дня данного вида величиной (нормой). При этом отбирается один тип отклонений от нормы.
Стабилизирующий отбор сохраняет в популяции среднее значение признаков (норму) и не пропускает в следующее поколение наиболее отклонившихся от этой нормы особей. Это делает сохранения видов неизменными.
При дизруптивном, или разрывающем, отборе отбирается не один тип отклонений от нормы, а два или больше. Это путь дробления предкового вида на дочерние группировки, каждая из которых может стать новым видом.
Эволюционные процессы, протекающие внутри вида и завершающиеся видообразованием, называют микроэволюцией. Макроэволюцией называется развитие групп организмов надвидового ранга. Задачами макроэволюции являются анализ соотношения индивидуального и исторического развития организмов, анализ закономерностей направления эволюционного процесса.
Шмальгаузен разработал и привел концепции целостности организма в индивидуальном и историческом развитии. Он исследовал механизмы эволюционного процесса и индивидуального развития организмов как саморегулирующихся систем, и изложил эволюционную теорию с позиций кибернетики.
В учении об отборе Ч. Дарвин доказал, что главной движущей силой эволюции является отбор наилучших форм, требующий для успеха таких условий: правильный выбор исходного материала, точная постановка цели, проведение селекции в достаточно широких масштабах и возможно более жесткая браковка материала, отбор по одному основному признаку.
Вид – основная структурная единица в естественных системах, качественный этап их эволюции. Новые виды возникают в результате межвидового скрещивания. Вид – это группа популяций, особи которых могут скрещиваться в естественных условиях, но изолированы от других видов. Вид подразделяется на несколько популяций, каждая из которых эволюционирует самостоятельно. Процесс перехода одного вида в другой не скачкообразен и генетическая изоляция между ними может не возникать.
Основой эволюционных процессов в естественных системах служит популяция. Популяция – это многочисленная совокупность особей определенного вида, в течение длительного времени (большого числа поколений) населяющих определенный участок географического пространства, внутри которого осуществляется та или иная степень случайного свободного скрещивания. В популяции нет абсолютно тождественных особей. Каждая особь является носителем уникального генотипа, который управляет формированием фенотипа. Существование каждой особи ограничено некоторым временным интервалом, по истечении которого особь погибает. При этом генотип особи исключается из генофонда популяции, но при жизни особь может передать наследственную информацию.
Известны три механизма передачи наследственной информации при рождении потомства; бесполое размножение, половое размножение, промежуточная (между бесполым и половым) форма размножения. Устойчивая передача генов от родителей к потомкам зависит в первую очередь от способности молекул ДНК к репродукции и авторепродукции. Каждая особь в течение жизни подвергается воздействиям внешней среды. В некоторых случаях эти воздействия могут привести к перестройкам молекул ДНК, переносящих наследственную информацию. Изменение первоначальной последовательности генов в молекулах ДНК приводит к изменению свойств этой молекулы, а следовательно, и наследственной информации.
Рассмотренные выше факторы по степени влияния на эволюцию можно упорядочить по убыванию влияния:
- естественный отбор;
- изоляция популяции;
- колебание численности популяции;
- мутационные процессы.

Пусковой механизм эволюции функционирует в результате совместного действия эволюционных факторов в пределах популяции. В результате действия эволюционных сил в каждой популяции многократно возникают элементарные эволюционные изменения. Со временем некоторые из них суммируются и ведут к возникновению новых приспособлений, что и лежит в основе видообразования.
Направленная молекулярная эволюция подобна искусственному отбору. Если надо создать молекулу, обладающую каким-либо химическим свойством, следует выбирать из большой популяции молекул те, которые в наилучшей степени выражают это свойство, и произвести из них дочерние, в разной степени похожие на родителей. Этот процесс отбора и дупликации повторяется до тех пор, пока не будет достигнут нужный результат.
Согласно теории Ч. Дарвина, эволюция осуществляется во взаимодействии трех повторяющихся вновь процессов: отбора, амплификации, мутации. Амплификация – процесс производства потомков или более точно, копирование особей, действует в природе совместно с отбором. Критерий отбора подобен библейскому: «плодитесь и размножайтесь». Стратегия повторяющейся рандомизации может заставить молекулы эволюционизировать в направлении улучшения функциональных характеристик.
Ведущую роль в эволюции играет следующий вид. Тесты по биологии
Урок 2 ВИД, ЕГО КРИТЕРИИ
Цели: сформировать понятие о виде, структуре вида, критериях; научить описывать особей видов по морфологическому критерию.
Оборудование: комнатные растения для проведения лабораторной работы.
Ход урока
I . Проверка знаний.
1. Тестирование.
1) Наследственную изменчивость Ч. Дарвин называл:
а) модификационной;
б)групповой;
в) неопределенной;
г) определенной.
2) Движущая сила эволюции (по Ч. Дарвину) видов — это:
а) искусственный отбор;
в) популяционные волны;
г) борьба за существование.
3) Материалом для эволюции служит:
а) борьба за существование;
б) естественный отбор;
в) модификационная изменчивость;
г) наследственная изменчивость.
4) Укажите неверное утверждение: «Результат действия ес-тественного отбора — это…»:
в) наследственная изменчивость;
г) образование новых видов.
5)Основная заслуга Ч.Дарвина состоит:
а) в формулировании биогенетического закона;
б) создании первой эволюционной теории;
в) разработка теории естественного отбора;
г) создании закона наследственных рядов.
6) По Ч. Дарвину, движущими силами эволюции являются:
а) борьба за существование;
б) наследственная изменчивость;
в) естественный отбор;
г) все перечисленные.
7) Ведущую роль в эволюции играет следующий вид изменчивости:
а) определенная;
б) модификационная;
в) групповая;
г) мутационная.
8) Способность организмов производить большое количество потомков и ограниченность мест обитания и жизненных ресурсов являются непосредственными причинами:
а) наследственной изменчивости;
б) борьбы за существование;
в) вымирания;
г) видообразования.
9) Наследственная изменчивость в процессе эволюции:
а) создаст новые виды;
б) доставляет материал для эволюции;
в) закрепляет созданный в процессе эволюции материал;
г) сохраняет наиболее полезные изменения.
10) Результатом действия естественного отбора не является:
а) приспособленность организмов к среде обитания;
б) многообразие органического мира;
в) борьба за существование;
г) совершенствование организации живых существ.
2. Задание.
Укажите номера предложений, в которых сделаны ошибки, исправьте их:
1) Не все (все) организмы изменчивы.
2) Все (не все) различия между организмами обязательно пе-редаются по наследству.
3) Организмы размножаются в арифметической (геометри-ческой) прогрессии, и все доживают до половозрелого со-стояния.
4) Жизненные ресурсы не ограничены (ограничены), и в борьбе за существование погибают только больные, ослаблен-ные особи (не только, но и менее приспособленные).
5) В результате борьбы за существование происходит естественный отбор — выживают те особи, которые располагают по-лезными в иных условиях свойствами.
3. Задание.
Установите соответствие между именем ученого и его вкла-дом в биологию.
Имя ученого
1.4. Лайель
2. Аристотель
3. К. Линней
4. К. Бэр 5.4. Дарвин
Заслуги ученых
а) Описал более 500 видов растений и животных, расположив их в опреде-ленном порядке.
б) При классификации использовал принцип соподчиненности система-тических категорий.
в) Сформулировал закон зародыше-вого сходства.
г) Изучая геологическую историю Земли, выдвинул идею движущих факторов эволюции Земли.
д) Вскрыл движущие силы и выявил причины протекания биологической эволюции.
4. Устные ответы у доски по вопросам в конце § 52, №3,4.
II . Изучение нового материала.
1. Вид.
Биологический вид — это совокупность особей, обладающих способностью к скрещиванию с образованием плодовитого по-томства; населяющих определенный ареал; обладающих рядом общих морфологических и физиологических признаков и сходством во взаимоотношениях с биотической и абиотической сре-дой.
Вид характеризуется целостностью и обособленностью. Це-лостность вида проявляется в том, что его особи могут жить и размножаться лишь взаимодействуя друг с другом — благодаря выработанным в процессе эволюции взаимным приспособлени-ям организмов: особенностям скоординированности строения материнского организма и эмбриона, системам сигнализации и восприятия у животных, общности территории и т. д.
Обособленность вида поддерживается репродуктивной изоляцией которая препятствует его смешению с другими видами при размножении. Изоляция обеспечивается различиями в строении половых органов, разобщенностью ар юн. расхожде-нием в сроках или местах размножения, различиями поведения и т. д. Благодаря изоляции виды не смешиваются друг с другом.
Изоляция обеспечивается различиями в строении половых органов, разобщенностью ар юн. расхожде-нием в сроках или местах размножения, различиями поведения и т. д. Благодаря изоляции виды не смешиваются друг с другом.
2. Критерии вида.
Характерные признаки и свойства, по которым одни виды отличаются от других, называют критериями вида.
Морфологический Физиологический Биохимический
Признаки вида
Генетический Экологический Географический Исторический
. Морфологический критерий.
В основе морфологического критерия лежит сходство внеш-него и внутреннего строения особей одного вида.
Но особи в пределах вида иногда настолько изменчивы, что только по морфологическому критерию не всегда удается опре-делить вид. Вместе с тем существуют виды морфологически сходные, однако особи таких видов не скрещиваются между со-бой. Это виды-двойники.
. Генетический критерий.
Имеется в виду набор хромосом, свойственный конкретному виду. Виды обычно отличаются по числу хромосом или по осо-бенностям их строения, поэтому генетический критерий доста-точно надежен. Однако и он не абсолютен. Встречаются случаи, когда виды имеют практически не различимые по строению хромосомы. Кроме того, в пределах вида могут быть широко распространены хромосомные мутации, что затрудняет его точ-ное определение.
. Физиологический критерий.
В основе лежит сходство всех процессов жизнедеятельно-сти особей одного вида, прежде всего сходство размножения. Представители разных видов, как правило, не скрещиваются, или потомство их бесплодно. Нескрещиваемость видов объ-ясняется различиями в строении полового аппарата, сроках размножения и др. Однако в природе есть виды, которые скрещиваются и дают плодовитое потомство (некоторые виды канареек, зябликов, тополей). Следовательно, физиологический критерий недостаточен для определения видовой принадлежности особей.
. Географический критерий.
Это определенный ареал, занимаемый видом в природе.
. Экологический критерий.
Основа экологического критерия — совокупность факторов внешней среды, в которой существует вид. Например, лютик едкий распространен на лугах и полях, в более сырых местах растет лютик ползучий; по берегам рек и прудов, на болотистых местах встречается лютик жгучий.
. Биохимический критерий.
Биохимический критерий позволяет различить виды по био-химическим параметрам (состав и структура определенных бел-ков, нуклеиновых кислот и других веществ).
. Исторический критерий.
Общность предков, единая история возникновения и развития вида.
Ни один из критериев в отдельности не может служить для определения вида. Охарактеризовать вид можно только по их совокупности.
III. Закрепление.
Лабораторная работа
Тема: МОРФОЛОГИЧЕСКИЕ ОСОБЕННОСТИ РАСТЕНИЙ РАЗЛИЧНЫХ ВИДОВ
Цели: обеспечить усвоение учащимися понятия морфоло-гического критерия вида; закрепить умение составлять описа-тельную характеристику растений.
Оборудование: живые растения или гербарные мате-риалы растений разных видов.
Ход работы
1. Рассмотрите растения двух видов, запишите их названия, составьте морфологическую характеристику растений каждого вида, то есть опишите особенности их внешнего строения (осо-бенности листьев, стеблей, корней, цветков, плодов).
2. Сравните растения двух видов, выявите черты сходства и различия. Чем объясняются сходства (различия) растений?
Домашнее задание: § 53. Письменное задание: составить списки известных вам видов растений и животных, сгруппиро-вать известные вам виды по степени сходства: а) морфологиче-ского, б)экологического.
Обобщающий урок по теме: «Основы учения об эволюции»
Уровень А.
1. Ведущую роль в эволюции играет следующий вид изменчивости
А) определенная
Б) модификационная
В) групповая
Г) мутационная
2. Способность живых существ производить большое количество потомков и ограниченность мест обитания и жизненных ресурсов являются непосредственными причинами:
А) наследственной изменчивости
Б) борьбы за существование
В) вымирания
Г) видообразования
3. Наиболее напряженной формой борьбы за существование Ч.Дарвин считал:
Наиболее напряженной формой борьбы за существование Ч.Дарвин считал:
А) борьбу с неблагоприятными условиями
Б) внутривидовую
В) межвидовую
Г) все перечисленные в равной степени
4. Наследственная изменчивость в процессе эволюции:
А) создает новые виды
Б) поставляет материал для эволюции
В) закрепляет созданный в процессе эволюции материал
Г) сохраняет наиболее полезные изменения
5. Естественный отбор действует на уровне:
А) отдельного организма
Б) популяции
Г) биоценоза
6. Движущая форма отбора обычно приводит:
А) к уничтожению особей с отклонениями от прежней нормы реакции
Б) расширению прежней нормы реакции
В) сужению прежней нормы реакции
Г) сдвигу прежней нормы реакции
7. Результатом действия естественного отбора не является:
А) приспособленность организмов к среде обитания
Б) многообразие органического мира
В) борьба за существование
Г) совершенствование организации живых существ
А) ароморфоза
Б) идиоадаптации
В) общей дегенерации
Г) направленной эволюции
11. К идиоадаптациям следует отнести:
А) многоклеточность
Б) четырехкамерное сердце млекопитающих
В) игольчатые листья растений пустынь
Г) все перечисленные эволюционные изменения
12. Примером ароморфоза является:
А) уплощение тела у донных рыб
Б) покровительственная окраска
Г) возникновение полового процесса
13. Естественный отбор – это:
А) сложные отношения между организмами и неживой природой
Б) процесс сохранения особей с полезными им наследственными признаками
В) процесс образования новых видов в природе
Г) процесс роста численности популяции
14. Борьба за существование играет большую роль в эволюции:
А) сохраняет особей преимущественно с полезными изменениями
Б) сохраняет особей с любыми наследственными изменениями
В) поставляет материал для отбора
Г) обостряет взаимоотношения между особями
15. В результате
взаимодействия движущих сил эволюции происходит:
В результате
взаимодействия движущих сил эволюции происходит:
А) размножение организмов
Б) образование новых видов в природе
В) мутационный процесс
Г) изоляция популяций
16. Укажите неверное утверждение: «В процессе эволюции борьба с неблагоприятными условиями приводит…»
А) к повышению сопротивляемости организмов
Б) выживанию наиболее жизнеспособных особей
В) вымиранию вида
Г) совершенствованию вида
17. К движущим силам эволюции относят:
А) многообразие видов
Б) борьбу за существование
В) видообразование
Г) приспособленность
18. Фактор эволюции, основу которого составляет возникновение преград к свободному скрещиванию особей , называют:
А) модификацией
Б) естественным отбором
В) изоляцией
Г) популяционными волнами
19. Среди движущих сил эволюции, ведущих к возникновению приспособлений у особей к среде обитания , направляющий характер имеет:
А) естественный отбор
Б) искусственный отбор
В) изоляция
Г) борьба за существование
20. Вид отбора, действующий в популяциях, обитающих в относительно постоянных условиях среды , — …
А) стихийный
Б) стабилизирующий
В) движущий
Г) половой
21. Каковы последствия стабилизирующего отбора?
А) сохранение старых видов
Б) сохранение нормы реакции
В) появление новых видов
Г) сохранение особей с неизменными признаками
22. Каковы последствия движущего отбора?
А) сохранение старых видов
Б) поддержание нормы реакции
В) появление новых видов
Г) устранение особей с новыми мутациями
23. Образование новых видов в природе происходит в результате
А) методического отбора
Б) искусственного отбора
В) деятельности человека
Г) взаимодействия движущих сил эволюции
24. При географическом видообразовании формирование нового вида происходит в результате:
А) распадения или расширения исходного ареала
Б) искусственного отбора
В) изоляции популяций внутри старого ареала
Г) дрейфа генов
Уровень В.
Установите соответствия между признаками изменчивости и ее видами:
- Обусловлена появлением новых сочетаний генов а. Мутационная
Обусловлена изменение генов и хромосом б. Комбинативная
У потомков появляются новые признаки
У потомков сочетаются родительские признаки
У особей изменяется количество или структура ДНК
У особей не изменяется количество или структура ДНК
Приспособления к жизни в воде, сформировавшиеся в процессе эволюции у китов:
Б) дыхание кислородом , растворенным в воде
В) обтекаемая форма тела
Г) развитый подкожный слой жира
Д) постоянная температура тела
Результатом эволюции является:
Б) появление новых засухоустойчивых сортов растений
В) выведение высокопродуктивных пород крупного рогатого скота
Г) формирование новых приспособлений к жизни в изменившихся условиях
Д) сохранение новых видов в изменившихся условиях
Е) получение высокопродуктивных бройлерных кур
Уровень С.
Какие ароморфозы позволили древним земноводным освоить сушу?
Какие ароморфозы позволили покрытосеменным растениям занять господствующее положение на Земле?
Тест по биологии ВАРИАНТ – 1. 1. Основная заслуга Ч. Дарвина состоит в: А) формулирование биогенетического закона; В) разработка теории естественного отбора; Б) создание первой эволюционной теории; Г) создание закона естественных рядов. 2. Наиболее…
Тест по биологии ВАРИАНТ – 1. 1. Основная заслуга Ч. Дарвина состоит в: А) формулирование биогенетического закона; В) разработка теории естественного отбора; Б) создание первой эволюционной теории; Г) создание закона естественных рядов. 2. Наиболее напряжённой формой борьбы за существование Ч. Дарвин считал: А) борьбу с неблагоприятными условиями; В) межвидовую; Б) внутривидовую; Г) все перечисленные формы в равной степени. 3. Естественный отбор действует на уровне: А) отдельного организма; В) вида; Б) популяции; Г) биоценоза. 4.Гомологичными органами являются: А) лапа кошки и нога мухи; В) чешуя рептилий и перья птицы; Б) глаз человека и глаз паука; Г) крыло бабочки и крыло птицы. 5. К обезьянолюдям относят: А) кроманьонца; В) питекантропа; Б) австралопитека; Г) неандертальца. 6. Экологический фактор, выходящий за пределы выносливости, называют: А) стимулирующим; В) абиотическим; Б) лимитирующим; Г) антропогенным 7. Эукариоты: А) способны к хемосинтезу; В) не имеют многих органоидов;
3. Естественный отбор действует на уровне: А) отдельного организма; В) вида; Б) популяции; Г) биоценоза. 4.Гомологичными органами являются: А) лапа кошки и нога мухи; В) чешуя рептилий и перья птицы; Б) глаз человека и глаз паука; Г) крыло бабочки и крыло птицы. 5. К обезьянолюдям относят: А) кроманьонца; В) питекантропа; Б) австралопитека; Г) неандертальца. 6. Экологический фактор, выходящий за пределы выносливости, называют: А) стимулирующим; В) абиотическим; Б) лимитирующим; Г) антропогенным 7. Эукариоты: А) способны к хемосинтезу; В) не имеют многих органоидов;
Тест по биологии ВАРИАНТ – 1. 1. Основная заслуга Ч. Дарвина состоит в: А) формулирование биогенетического закона; В) разработка теории естественного отбора; Б) создание первой эволюционной теории; Г) создание закона естественных рядов. 2. Наиболее напряжённой формой борьбы за существование Ч. Дарвин считал: А) борьбу с неблагоприятными условиями; В) межвидовую; Б) внутривидовую; Г) все перечисленные формы в равной степени. 3. Естественный отбор действует на уровне: А) отдельного организма; В) вида; Б) популяции; Г) биоценоза. 4.Гомологичными органами являются: А) лапа кошки и нога мухи; В) чешуя рептилий и перья птицы; Б) глаз человека и глаз паука; Г) крыло бабочки и крыло птицы. 5. К обезьянолюдям относят: А) кроманьонца; В) питекантропа; Б) австралопитека; Г) неандертальца. 6. Экологический фактор, выходящий за пределы выносливости, называют: А) стимулирующим; В) абиотическим; Б) лимитирующим; Г) антропогенным 7. Эукариоты: А) способны к хемосинтезу; В) не имеют многих органоидов; Б) имеют ДНК кольцевой формы; Г) имеют ядро с собственной оболочкой. 8. Общим признаком растительной и животной клетки является: А) гетеротрофность; В) наличие хлоропластов; Б) наличие митохондрий; Г) наличие жёсткой клеточной стенки. 9. Биополимерами являются: А) белки; В) нуклеиновые кислоты; Б) полисахариды; Г) всё перечисленное. 10. Урацил образует комплиментарную связь с: А) аденином В) цитозином Б) тимином Г) гуанином. 11. Гликолизом называется: А) совокупность всех процессов энергетического обмена в клетке; Б) бескислородное расщепление глюкозы; В) полное расщепление глюкозы; Г) полимеризация глюкозы с образованием гликогена. 12. Очерёдность стадии митоза следующая: А) метафаза, телофаза, профаза, анафаза; В) профаза, метафаза, телофаза, анафаза; Б) профаза, метафаза, анафаза, телофаза; Г) телофаза, профаза, метафаза, анафаза; 13. Удвоение хромосом происходит в: А) интерфазе В) метафазе Б) профазе Г) телофазе 14. В анафазе митоза происходит расхождение: А) дочерних хромосом В) негомологичных хромосом Б) гомологичных хромосом Г) органоидов клетки. 15. Из перечисленных животных самая крупная яйцеклетка у: А) осетра В) ящерицы Б) лягушки Г) курицы. 16. из эктодермы образуются: А) мышцы В) скелет Б) лёгкие Г) органы чувств. 17. При Менделеевском моногибридном скрещивании доля особей хотя бы с одним рецессивным геном во втором поколении будет равна: А) 25% Б) 50% В) 75% Г) 100%
11. Гликолизом называется: А) совокупность всех процессов энергетического обмена в клетке; Б) бескислородное расщепление глюкозы; В) полное расщепление глюкозы; Г) полимеризация глюкозы с образованием гликогена. 12. Очерёдность стадии митоза следующая: А) метафаза, телофаза, профаза, анафаза; В) профаза, метафаза, телофаза, анафаза; Б) профаза, метафаза, анафаза, телофаза; Г) телофаза, профаза, метафаза, анафаза; 13. Удвоение хромосом происходит в: А) интерфазе В) метафазе Б) профазе Г) телофазе 14. В анафазе митоза происходит расхождение: А) дочерних хромосом В) негомологичных хромосом Б) гомологичных хромосом Г) органоидов клетки. 15. Из перечисленных животных самая крупная яйцеклетка у: А) осетра В) ящерицы Б) лягушки Г) курицы. 16. из эктодермы образуются: А) мышцы В) скелет Б) лёгкие Г) органы чувств. 17. При Менделеевском моногибридном скрещивании доля особей хотя бы с одним рецессивным геном во втором поколении будет равна: А) 25% Б) 50% В) 75% Г) 100%
18. Сцепленными называют гены, находящиеся в: А) одной хромосоме В) половых хромосомах Б) гомологичных хромосомах Г) аутосомах. 19. Мутации проявляются фенотипически: А) всегда В) только в гомозиготном состоянии Б) только в гетерозиготном состоянии Г) никогда. 20. Полиплоидия заключается в: А) изменении числа отдельных хромосом В) изменении структуры хромосом Б) кратном изменении гаплоидного числа хромосом; Г) изменении структуры отдельных генов. ОТВЕТ: 1 – В, 2 – Б, 3 – Б, 4 – В, 5 – В, 6 – Б, 7 – Г, 8 – Б, 9 – Г, 10 – А, 11 – Б, 12 – Б, 13 – А, 14 – А, 15 – Г, 16 – Г, 17 – В, 18 – А, 19 – В, 20 – Б. ВАРИАНТ – 2 1. По Ч. Дарвину, движущими силами эволюции являются: А) борьба за существование; В) естественный отбор; Б) наследственная изменчивость; Г) все перечисленные. 2. Ведущую роль в эволюции играет следующий вид изменчивости: А) определённая; В) групповая; Б) модификационная; Г) мутационная. 3. Движущая форма отбора обычно приводит к: А) уничтожению особей с отклонениями В) расширению прежней нормы реакции; от прежней нормы реакции; Б) сужению прежней нормы реакции; Г) сдвигу прежней нормы реакции. 4. Аналогичными органами являются: А) жабры рака и жабры рыбы; В) листья берёзы и иголки кактуса; Б) лапа собаки и крыло птицы; Г) все перечисленные пары. 5. В эпоху оледенения жили: А) кроманьонцы; В) синантропы; Б) неандертальцы; Г) все перечисленные. 6. Продуктивностью экосистемы называется: А) её суммарная биомасса; В) суммарная биомасса продуцентов; Б) прирост этой биомассы за единицу времени; Г) суммарная биомасса консументов. 7.В клетках прокариот имеются: А) ядра; В) митохондрии; Б) рибосомы; Г) все перечисленные органоиды. 8. Лейкопласты – это органоиды клетки, в которых: А) осуществляется синтез белка; В) находятся пигменты красного и жёлтого цвета; Б) осуществляется процесс фотосинтеза; Г) накапливается крахмал. 9. Нуклеотиды в нити молекулы ДНК соединяются следующей связью: А) ковалентной; В) пептидной; Б) водородной; Г) дисульфидными мостиками. 10. Транскрипция – это: А) синтез молекулы и-РНК В) доставка аминокислот к рибосомам по матрице одной из цепей ДНК; во время синтеза белка; Б) перенос информации с и-РНК на белок Г) процесс сборки белковой молекулы. во время его синтеза; 11. Синтез АТФ в клетке происходит в процессе: А) гликолиза; В) клеточного дыхания; Б) фотосинтеза; Г) всех перечисленных. 12.Самой продолжительной фазой митоза является:
4. Аналогичными органами являются: А) жабры рака и жабры рыбы; В) листья берёзы и иголки кактуса; Б) лапа собаки и крыло птицы; Г) все перечисленные пары. 5. В эпоху оледенения жили: А) кроманьонцы; В) синантропы; Б) неандертальцы; Г) все перечисленные. 6. Продуктивностью экосистемы называется: А) её суммарная биомасса; В) суммарная биомасса продуцентов; Б) прирост этой биомассы за единицу времени; Г) суммарная биомасса консументов. 7.В клетках прокариот имеются: А) ядра; В) митохондрии; Б) рибосомы; Г) все перечисленные органоиды. 8. Лейкопласты – это органоиды клетки, в которых: А) осуществляется синтез белка; В) находятся пигменты красного и жёлтого цвета; Б) осуществляется процесс фотосинтеза; Г) накапливается крахмал. 9. Нуклеотиды в нити молекулы ДНК соединяются следующей связью: А) ковалентной; В) пептидной; Б) водородной; Г) дисульфидными мостиками. 10. Транскрипция – это: А) синтез молекулы и-РНК В) доставка аминокислот к рибосомам по матрице одной из цепей ДНК; во время синтеза белка; Б) перенос информации с и-РНК на белок Г) процесс сборки белковой молекулы. во время его синтеза; 11. Синтез АТФ в клетке происходит в процессе: А) гликолиза; В) клеточного дыхания; Б) фотосинтеза; Г) всех перечисленных. 12.Самой продолжительной фазой митоза является:
А) профаза; В) анафаза; Б) метафаза; Г) телофаза. 13.Редукция числа хромосом происходит во время: А) анафазы митоза; В) II деления мейоза; Б) I деления мейоза; Г) во всех перечисленных случаях. 14. Биологическое значение мейоза заключается в обеспечении: А) генетической стабильности; В) генетической изменчивости; Б) регенерации тканей и увеличения Г) бесполого размножения. числа клеток в организме; 15. Нервная система образуется из: А) эктодермы; В) мезодермы; Б) энтодермы; Г) нет верного ответа. 16. Из мезодермы образуются: А) лёгкие; В) кровеносная система; Б) нервная система; Г) органы чувств. 17. Сколько типов гамет образуют дигетерозиготные особи: А) один; В) четыре; Б) два; Г) нет верного ответа. 18. К мутационной изменчивости относятся: А) изменения в хромосомах; В) изменения, передающиеся по наследству; Б) изменения в генах; Г) все перечисленные. 19. Основным источником комбинативной изменчивости является: А) перекрест хромосом В) независимое расхождение хроматид В профазе I деления мейоза; в анафазе II деления мейоза; Б) независимое расхождение Г) все перечисленные процессы в равной степени. гомологичных хромосом в анафазе I деления мейоза; 20. Межлинейная гибридизация культурных растений приводит к: А) сохранению прежней продуктивности; В) повышению продуктивности; Б) выщеплению новых признаков; Г) закреплению признаков. ОТВЕТ: 1 – Г, 2 – Г, 3 – Г, 4 – А, 5 – Б, 6 – Б, 7 – Б, 8 – Г, 9 – А, 10 – А, 11 – Г, 12 – А, 13 – Б, 14 – В, 15 – А, 16 – В, 17 – В, 18 – Г, 19 – Г, 20 – В.
К мутационной изменчивости относятся: А) изменения в хромосомах; В) изменения, передающиеся по наследству; Б) изменения в генах; Г) все перечисленные. 19. Основным источником комбинативной изменчивости является: А) перекрест хромосом В) независимое расхождение хроматид В профазе I деления мейоза; в анафазе II деления мейоза; Б) независимое расхождение Г) все перечисленные процессы в равной степени. гомологичных хромосом в анафазе I деления мейоза; 20. Межлинейная гибридизация культурных растений приводит к: А) сохранению прежней продуктивности; В) повышению продуктивности; Б) выщеплению новых признаков; Г) закреплению признаков. ОТВЕТ: 1 – Г, 2 – Г, 3 – Г, 4 – А, 5 – Б, 6 – Б, 7 – Б, 8 – Г, 9 – А, 10 – А, 11 – Г, 12 – А, 13 – Б, 14 – В, 15 – А, 16 – В, 17 – В, 18 – Г, 19 – Г, 20 – В.
ВАРИАНТ – 1.
1. Основная заслуга Ч. Дарвина состоит в:
А) формулирование биогенетического закона; В) разработка теории естественного отбора;
Б) создание первой эволюционной теории; Г) создание закона естественных рядов.
2. Наиболее напряжённой формой борьбы за существование Ч. Дарвин считал:
А) борьбу с неблагоприятными условиями; В) межвидовую;
Б) внутривидовую; Г) все перечисленные формы в равной степени.
3. Естественный отбор действует на уровне:
А) отдельного организма; В) вида;
Б) популяции; Г) биоценоза.
4.Гомологичными органами являются:
А) лапа кошки и нога мухи; В) чешуя рептилий и перья птицы;
Б) глаз человека и глаз паука; Г) крыло бабочки и крыло птицы.
5. К обезьянолюдям относят:
А) кроманьонца; В) питекантропа;
Б) австралопитека; Г) неандертальца.
6. Экологический фактор, выходящий за пределы выносливости, называют:
А) стимулирующим; В) абиотическим;
Б) лимитирующим; Г) антропогенным
7. Эукариоты:
А) способны к хемосинтезу; В) не имеют многих органоидов;
Б) имеют ДНК кольцевой формы; Г) имеют ядро с собственной оболочкой.
8. Общим признаком растительной и животной клетки является:
А) гетеротрофность; В) наличие хлоропластов;
Б) наличие митохондрий; Г) наличие жёсткой клеточной стенки.
9. Биополимерами являются:
А) белки; В) нуклеиновые кислоты;
Б) полисахариды; Г) всё перечисленное.
10. Урацил образует комплиментарную связь с:
А) аденином В) цитозином
Б) тимином Г) гуанином.
11. Гликолизом называется:
А) совокупность всех процессов энергетического обмена в клетке;
Б) бескислородное расщепление глюкозы;
В) полное расщепление глюкозы; Г) полимеризация глюкозы с образованием гликогена.
12. Очерёдность стадии митоза следующая:
А) метафаза, телофаза, профаза, анафаза; В) профаза, метафаза, телофаза, анафаза;
Б) профаза, метафаза, анафаза, телофаза; Г) телофаза, профаза, метафаза, анафаза;
13. Удвоение хромосом происходит в:
А) интерфазе В) метафазе
Б) профазе Г) телофазе
14. В анафазе митоза происходит расхождение:
А) дочерних хромосом В) негомологичных хромосом
Б) гомологичных хромосом Г) органоидов клетки.
15. Из перечисленных животных самая крупная яйцеклетка у:
А) осетра В) ящерицы
Б) лягушки Г) курицы.
16. из эктодермы образуются:
А) мышцы В) скелет
Б) лёгкие Г) органы чувств.
17. При Менделеевском моногибридном скрещивании доля особей хотя бы с одним рецессивным геном во втором поколении будет равна:
А) 25% Б) 50% В) 75% Г) 100%
18. Сцепленными называют гены, находящиеся в:
А) одной хромосоме В) половых хромосомах
Б) гомологичных хромосомах Г) аутосомах.
19. Мутации проявляются фенотипически:
А) всегда В) только в гомозиготном состоянии
Б) только в гетерозиготном состоянии Г) никогда.
20. Полиплоидия заключается в:
А) изменении числа отдельных хромосом В) изменении структуры хромосом
Б) кратном изменении гаплоидного числа хромосом; Г) изменении структуры отдельных генов.
ОТВЕТ: 1 – В, 2 – Б, 3 – Б, 4 – В, 5 – В, 6 – Б, 7 – Г, 8 – Б, 9 – Г, 10 – А, 11 – Б, 12 – Б, 13 – А, 14 – А, 15 – Г, 16 – Г, 17 – В, 18 – А, 19 – В, 20 – Б.
Проверочный тест по биологии 11 класс. (1)ВАРИАНТ – 2
1. По Ч. Дарвину, движущими силами эволюции являются:
А) борьба за существование; В) естественный отбор;
Б) наследственная изменчивость; Г) все перечисленные.
2. Ведущую роль в эволюции играет следующий вид изменчивости:
А) определённая; В) групповая;
Б) модификационная; Г) мутационная.
3. Движущая форма отбора обычно приводит к:
А) уничтожению особей с отклонениями В) расширению прежней нормы реакции;
от прежней нормы реакции;
Б) сужению прежней нормы реакции; Г) сдвигу прежней нормы реакции.
4. Аналогичными органами являются:
А) жабры рака и жабры рыбы; В) листья берёзы и иголки кактуса;
Б) лапа собаки и крыло птицы; Г) все перечисленные пары.
5. В эпоху оледенения жили:
А) кроманьонцы; В) синантропы;
Б) неандертальцы; Г) все перечисленные.
6. Продуктивностью экосистемы называется:
А) её суммарная биомасса; В) суммарная биомасса продуцентов;
Б) прирост этой биомассы за единицу времени; Г) суммарная биомасса консументов.
7.В клетках прокариот имеются:
А) ядра; В) митохондрии;
Б) рибосомы; Г) все перечисленные органоиды.
8. Лейкопласты – это органоиды клетки, в которых:
А) осуществляется синтез белка; В) находятся пигменты красного и жёлтого цвета;
Б) осуществляется процесс фотосинтеза; Г) накапливается крахмал.
9. Нуклеотиды в нити молекулы ДНК соединяются следующей связью:
А) ковалентной; В) пептидной;
Б) водородной; Г) дисульфидными мостиками.
10. Транскрипция – это:
А) синтез молекулы и-РНК В) доставка аминокислот к рибосомам
по матрице одной из цепей ДНК; во время синтеза белка;
Б) перенос информации с и-РНК на белок Г) процесс сборки белковой молекулы.
во время его синтеза;
11. Синтез АТФ в клетке происходит в процессе:
А) гликолиза; В) клеточного дыхания;
Б) фотосинтеза; Г) всех перечисленных.
12.Самой продолжительной фазой митоза является:
А) профаза; В) анафаза;
Б) метафаза; Г) телофаза.
13.Редукция числа хромосом происходит во время:
А) анафазы митоза; В) II деления мейоза;
Б) I деления мейоза; Г) во всех перечисленных случаях.
14. Биологическое значение мейоза заключается в обеспечении:
А) генетической стабильности; В) генетической изменчивости;
Б) регенерации тканей и увеличения Г) бесполого размножения.
числа клеток в организме;
15. Нервная система образуется из:
А) эктодермы; В) мезодермы;
Б) энтодермы; Г) нет верного ответа.
16. Из мезодермы образуются:
А) лёгкие; В) кровеносная система;
Б) нервная система; Г) органы чувств.
17. Сколько типов гамет образуют дигетерозиготные особи:
А) один; В) четыре;
Б) два; Г) нет верного ответа.
18. К мутационной изменчивости относятся:
А) изменения в хромосомах; В) изменения, передающиеся по наследству;
Б) изменения в генах; Г) все перечисленные.
19. Основным источником комбинативной изменчивости является:
А) перекрест хромосом В) независимое расхождение хроматид
В профазе I деления мейоза; в анафазе II деления мейоза;
Б) независимое расхождение Г) все перечисленные процессы в равной степени.
гомологичных хромосом
в анафазе I деления мейоза;
20. Межлинейная гибридизация культурных растений приводит к:
А) сохранению прежней продуктивности; В) повышению продуктивности;
Б) выщеплению новых признаков; Г) закреплению признаков.
ОТВЕТ: 1 – Г, 2 – Г, 3 – Г, 4 – А, 5 – Б, 6 – Б, 7 – Б, 8 – Г, 9 – А, 10 – А, 11 – Г, 12 – А, 13 – Б, 14 – В, 15 – А, 16 – В, 17 – В, 18 – Г, 19 – Г, 20 – В.
ВАРИАНТ – 1.
1. Дезоксирибонуклеиновая кислота – это уровень организации живой природы:
Дезоксирибонуклеиновая кислота – это уровень организации живой природы:
А) клеточный; Б) молекулярный;
В) организменный; Г) популяционный
2. Наука цитология изучает:
А) строение клеток одноклеточных и многоклеточных организмов;
Б) строение органов и системы органов многоклеточных организмов;
В) фенотип организмов разных царств;
Г) морфологию растений и особенности их развития.
3. Белки в клетки синтезируются:
А) в цитоплазме; Б) в лизосомах;
В) на рибосомах; Г) в комплексе Гольджи.
4. Белки, способные ускорять химические реакции, выполняют в клетке функцию:
А) гормональную Б) сигнальную
В) ферментативную Г) информационную.
5. Транспортная РНК – это:
А) белок Б) жир
В) фермент Г) нуклеиновая кислота.
6. Конъюгация хромосом характерна для процесса:
А) оплодотворения Б) профазы второго деления мейоза
В) митоза Г) профазы первого деления мейоза
7. Бластула состоит из полости и:
А) двух слоёв клеток Б) соединительной ткани
В) одного слоя клеток Г) эпителиальной ткани.
8. Какие клетки передают потомству мутации при половом размножении:
А) эпителиальные Б) мышечные
В) гаметы Г) нейроны.
9. К древним людям относят:
А) неандертальца Б) питекантропа
В) синантропа Г) кроманьонца.
10. Между лосем и зубром наблюдается конкуренция, так как они:
А) питаются сходной пищей; Б) имеют примерно одинаковые параметры тела
В) имеют немногочисленное потомство; Г) относятся к классу млекопитающих.
11. К агроэкосистемам относят:
А) смешанный лес Б) заливной луг
В) зарастающее озеро Г) пшеничное поле.
12. Приспособленность в процессе эволюции возникает в результате:
А) географической изоляции Б) взаимодействия движущих сил эволюции
В) мутационной изменчивости Г) искусственного отбора.
13. Цитоплазма в клетке не выполняет функцию:
А) транспорта веществ Б) внутренней среды
В) осуществления связи между ядром и органоидами; Г) фотосинтеза.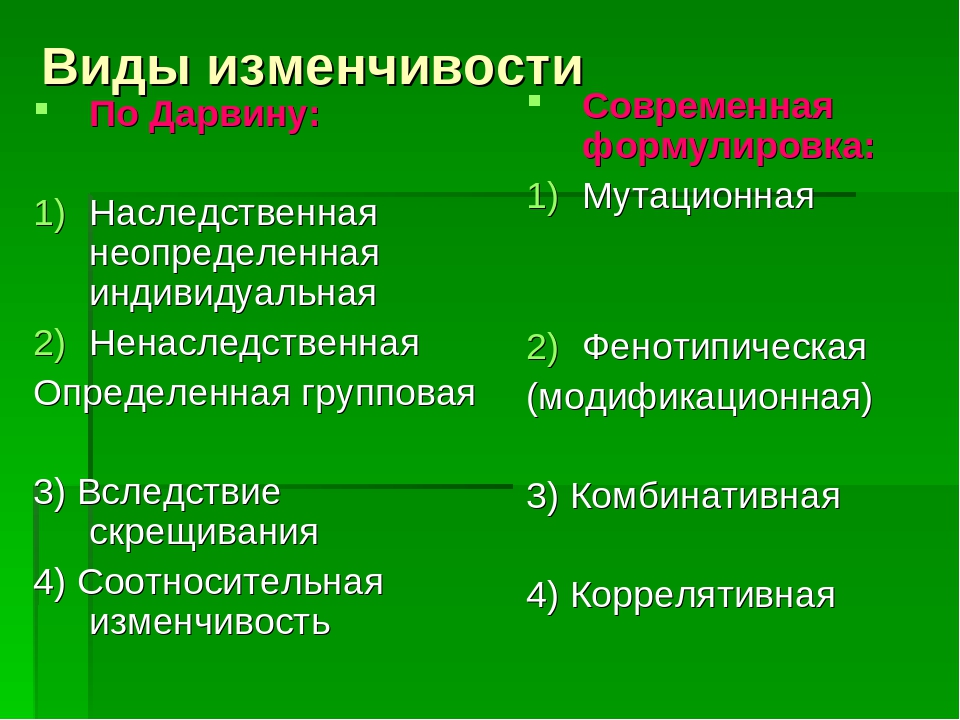
14. Способность плазматической мембраны окружать твёрдую частицу пищи и перемещать её внутрь клетки лежит в основе процесса:
А) диффузии Б) осмоса
В) фагоцитоза Г) пиноцитоза.
15. Где протекает анаэробный этап гликолиза?
А) в митохондриях Б) в лёгких
В) в пищеварительной трубке Г) в цитоплазме.
16. У особи с генотипом Аавв образуются гаметы:
А) Ав, вв Б) Ав, ав
В) Аа, АА Г) Аа, вв.
17. При скрещивании гетерозиготных растений с красными и круглыми плодами с рецессивными по обоим признакам особями (красные А и круглые В – доминантные признаки) появится потомство с генотипами АаВв, ааВв, Аавв, аавв в соотношении:
А) 3:1, Б) 9:3:3:1
В) 1:1:1:1 Г) 1:2:1.
18.Из яйцеклетки развивается девочка, если в процессе оплодотворения в зиготе оказались хромосомы:
А) 44 аутосомы +ХY Б) 23 аутосомы +Х
В) 44 аутосомы +ХХ Г) 23 аутосомы +Y.
19. Значительная часть мутаций не проявляется в фенотипе потомства, так как они6
А) не связанны с изменением генов Б) не связанны с изменением хромосом
В) носят доминантный характер Г) носят рецессивный характер.
20. Водоём, заселённый разнообразными видами растений и животных – это:
А) биогеоценоз Б) ноосфера
В) биосфера Г) агроэкосистема.
Ответы : 1 – Б, 2 – А, 3 – В, 4 – В, 5 – Г, 6 – Г, 7 – В, 8 – В, 9 – А, 10 – А, 11 – Г, 12 – Б, 13 – Г, 14 – В, 15 – Г, 16 – Б, 17 – В, 18 – В, 19 – Г, 20 – А.
Проверочный тест по биологии 11 класс. (2)ВАРИАНТ – 2
1. Для выявления общих анатомических признаков, характерных для царства живой природы, используют метод:
А) микроскопирования Б) прогнозирования
В) сравнения Г) моделирования
2. согласно клеточной теории в эукариотических клетках обязательно есть:
А) клеточная стенка Б) ядро
В) вакуоли Г) пластиды
3. В процессе деления клетки наиболее существенные преобразования претерпевают:
А) рибосомы Б) хромосомы
В) митохондрии Г) лизосомы
4. Структура молекулы ДНК представляет собой:
Структура молекулы ДНК представляет собой:
А) две спирально закрученные одна вокруг другой полинуклеотидные нити
Б) одну спирально закрученную полинуклеотидную нить
В) две спирально закрученные полипептидные нити
Г) одну прямую полипептидную нить.
5. На подготовительном этапе энергетического обмена энергия:
А) поглощается в виде тепла Б) выделяется в виде тепла
В) поглощается цитоплазмой клетки Г) выделяется за счёт расщепления АТФ.
6. «Преобладающий признак одного из родителей» Г. Мендель назвал:
А) рецессивным Б) доминантным
В) гомозиготным Г) гетерозиготным
7. При скрещивании гетерозиготы с гомозиготой доля гомозигот в потомстве составит:
А) 0% Б) 25% В) 50% Г) 100%
8. Изменения, которые не передаются по наследству и возникают как приспособления к внешней среде, называют:
А) неопределёнными Б) индивидуальными
В) мутационными Г) модификационными.
9. Закон гомологических рядов в наследственной изменчивости был установлен:
А) В.И. Вернадским Б) И.В. Мичуриным
В) Н.И. Вавиловым Г) Т. Морганом.
10. Наличие у человека, как и у других млекопитающих животных, живорождения, выкармливания детёнышей молоком свидетельствует:
А) о более высоком уровне развития человека; Б) об их дивергентной эволюции
В) об историческом развитии класса млекопитающих; Г) об их родстве.
11. Обмен веществ в клетке состоит из процессов:
А) возбуждения и торможения Б) пластического и энергетического обмена
В) роста и развития Г) транспорта гормонов и витаминов.
12. К продуцентам в экосистемах относят:
В) сапрофитные бактерии Г) грибы.
13. Новые виды в природе возникают в результате взаимодействия:
А) наследственной изменчивости, борьбы за существование, естественного отбора
Б) ненаследственных изменений и сезонных изменений в природе
В) приспособленности организмов и искусственного отбора
Г) абиотических факторов среды
14. Главной причиной биологического регресса многих видов в настоящее время является:
Главной причиной биологического регресса многих видов в настоящее время является:
А) изменение климата Б) хозяйственная деятельность человека
В) изменение рельефа Г) увеличение численности хищников.
15. Ядро играет большую роль в клетке, так как оно участвует в синтезе:
А) глюкозы Б) клетчатки
В) липидов Г) нуклеиновых кислот.
16. Вещества, способные в живом организме образовывать с водой водородные связи или вступать в электростатическое взаимодействие, являются:
А) гидрофобными Б) гидрофильными
В) нейтральными Г) щелочными.
17. В бескислородной стадии энергетического обмена расщепляются молекулы:
А) глюкозы до пировиноградной кислоты Б) белка до аминокислот
В) крахмала до глюкозы Г) пировиноградной кислоты до углекислого газа и воды
18. При делении клеток животных и растений основным источником энергии являются молекулы:
А) АТФ Б) тРНК В) иРНК Г) ДНК.
19. Разнообразие подводных и надводных листьев стрелолиста – пример:
А) модификационной изменчивости Б) действия мутогенов
В) комбинативной изменчивости Г) различия в генотипах разных клеток
20. В эволюции человека начальные вехи развития искусства обнаружены среди:
А) неандертальцев Б) кроманьонцев
В) австралопитеков питекантропов
Ответы : 1 – В, 2 – Б, 3 – Б, 4 – А, 5 – Б, 6 – Б, 7 – В, 8 – Г, 9 – В, 10 – Г, 11 – Б, 12 – А, 13 – А, 14 – Б, 15 – Г, 16 – Б, 17 – А, 18 – А, 19 – А, 20 – Б.
Чарльз Дарвин – путешественник и исследователь
Самым значимым событием в своей жизни Чарльз Дарвин называл путешествие на корабле «Бигль». 5-летнее странствие по отдалённым уголкам земного шара позволило натуралисту собрать солидный фактологический материал. Он лёг в основу дальнейших исследований и натолкнул учёного на ряд важных научных заключений.
Путешествие на «Бигле»
Дарвин пустился в плавание в 1831 году, в возрасте 23 лет. Он закончил обучение естественным наукам в Кембриджском университете и устроился в команду корабля «Бигль». Судно отправлялось в длительный кругосветный вояж по морям и океанам Южного полушария. В ходе экспедиции «Бигль» обогнул Южную Америку, Африку и Австралию.
Он закончил обучение естественным наукам в Кембриджском университете и устроился в команду корабля «Бигль». Судно отправлялось в длительный кругосветный вояж по морям и океанам Южного полушария. В ходе экспедиции «Бигль» обогнул Южную Америку, Африку и Австралию.
Южная Америка
Дарвин смог сравнить природу разных широт и континентов. За время экспедиции он собрал данные, которые объясняют закономерности географического распространения флоры и фауны в соответствии с природными зонами: от экваториальной до субантарктической. Дарвин также обратил внимание, что растительный и животный мир меняется при подъёме на высоту в горах.
Натуралиста интересовали не только существующие виды живых организмов, но и уже вымершие. Сравнивая скелеты ископаемых и настоящих ленивцев и броненосцев, Дарвин констатировал их сходство. Были заметны и отличия. Учёный высказал мнение, что у современных и вымерших животных общее происхождение. Изменения в них произошли под воздействием факторов внешней среды.
На основании находок, сделанных в Южной Америке, естествоиспытатель сделал заключение, что живые организмы в прошлом вымирали не только в результате глобальных катаклизмов.
Острова – Галапагосы и Огненная земля
По-новому взглянуть на природную среду и её обитателей заставило Дарвина посещение Галапагосских островов. Особенно его заинтересовали птицы из семейства вьюрковых. На архипелаге обитало 13 разных видов. Они отличались друг от друга прежде всего видом клюва. Дарвин предположил, что строение клюва приспособлено к типу пищи.
Попутно он выдвинул гипотезу, что птицы на островах имеют одинаковое происхождение с континентальными видами, а их особенности вызваны обитанием в удалённой от других местности.
Ещё одним знаковым местом стало посещение Огненной Земли. Наблюдение за аборигенами натолкнуло учёного на революционную идею о происхождении homo sapiens от животных.
Исследования строения коралловых рифов привели впоследствии к выдвижению теории формирования коралловых островов.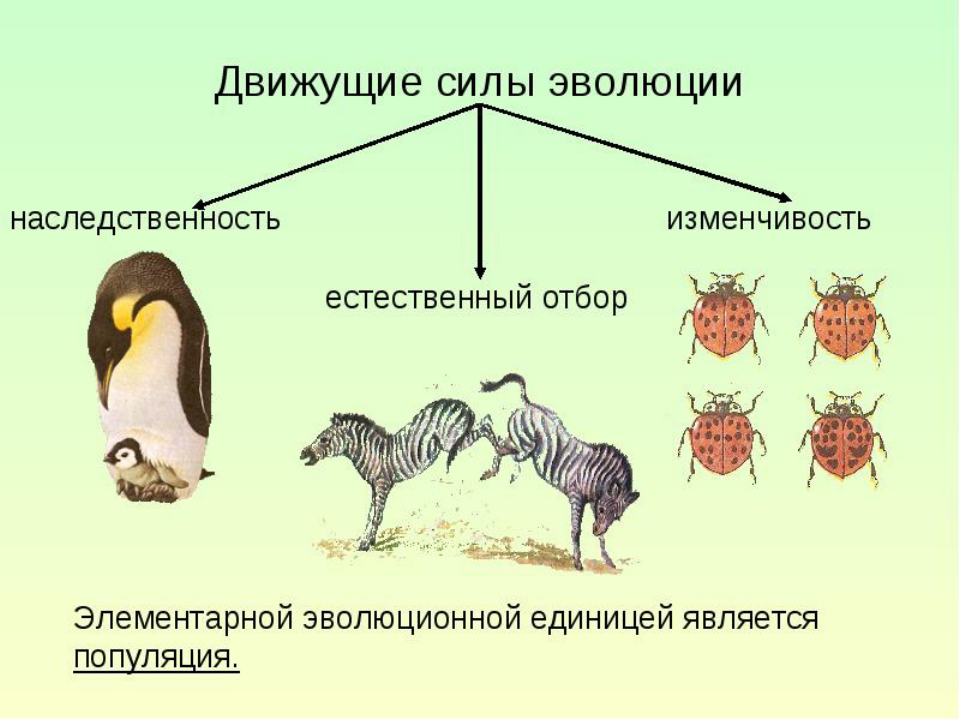
После путешествия
«Бигль» вернулся в Лондон 2 октября 1936 года. С этого времени Дарвин начинает систематизировать, обобщать и анализировать собранные материалы. Свои выводы он обосновывает в фундаментальных трудах по зоологии, биологии, геологии.
«Самую сильную черту отличия человека от животных составляет нравственное чувство, или совесть. И господство его выражается в коротком, но могучем и крайне выразительном слове «должен».
Чарльз Дарвин
Основные труды
В 1859 году Чарльз Дарвин издаёт «Происхождение видов путём естественного отбора», в ней сформулирована теория эволюции, получившая определение «дарвинизм». Основными причинами биологического многообразия и адаптации организмов учёный считает:
- естественный отбор;
- наследственную изменчивость.
В 1868 году опубликовано издание «Изменение домашних животных и культурных растений», где со всех сторон рассматривается влияние изменчивости, генетики, искусственного отбора. Новая работа, содержащая множественные подтверждения антропогенеза – «Происхождение человека и половой отбор», увидела свет в 1871 году.
Научная трилогия Дарвина приводит бесспорные доказательства развития естественной среды в ходе истории, устанавливает факторы, влияющие на эволюцию.
Выставка «Великие учителя человечества» в ЭТНОМИРе
Калужская область, Боровский район, деревня Петрово
Экcпозиция «Великие учителя человечества» расположена на втором и третьем этажах Культурного центра Индии. Она включает в себя свыше 100 экспонатов, это величайшее собрание бюстов мудрецов всех времён и народов, которые оставили миру самое ценное наследие – знания, указали и на собственном примере продемонстрировали пути духовного развития. Изучая труды, научные открытия, философские трактаты этих учителей, мы приходим к пониманию, что в основе базовой системы ценностей лежит единый фундамент: единство религий, единство народов и единство человека и природы. Около каждого бюста на выставке посетитель найдёт информационную табличку с коротким рассказом об основных заслугах Учителя перед человечеством, с указанием знаковых дат и перечнем его трудов. Экспозиция всегда открыта для самостоятельного изучения.
Около каждого бюста на выставке посетитель найдёт информационную табличку с коротким рассказом об основных заслугах Учителя перед человечеством, с указанием знаковых дат и перечнем его трудов. Экспозиция всегда открыта для самостоятельного изучения.
Проверочная работа по биологии 11класс «Теория Ч. Дарвина. Критерии вида»
Проверочная работа по теме: Теория Ч.Дарвина. Критерии вида.
1) Наследственную изменчивость Ч. Дарвин называл:
а) модификационной; б) групповой; в) неопределенной; г) определенной.
2) Движущая сила эволюции (по Ч. Дарвину) видов – это:
а) искусственный отбор б) наследственная изменчивость; в) популяционные волны; г) борьба за существование.
3) Материалом для эволюции служит:
а) борьба за существование б) естественный отбор; в) модификационная изменчивость;
г) наследственная изменчивость.
4) Укажите неверное утверждение: «Результат действия естественного отбора – это…»:
а) приспособленность организмов к среде обитания; б) многообразие органического мира;
в) наследственная изменчивость; г) образование новых видов.
5) Основная заслуга Ч. Дарвина состоит:
а) в формулировании биогенетического закона; б) создании первой эволюционной теории;
в) разработке теории естественного отбора; г) создании закона наследственных рядов.
6) По Ч. Дарвину, движущими силами эволюции являются:
а) борьба за существование; б) наследственная изменчивость ;в) естественный отбор; г) все перечисленные.
7) Ведущую роль в эволюции играет следующий вид изменчивости:
а) определенная; б) модификационная; в) групповая; г) мутационная.
8) Способность организмов производить большое количество потомков и ограниченность мест обитания и жизненных ресурсов являются непосредственными причинами:
а) наследственной изменчивости; б) борьбы за существование в) вымирания; г) видообразования.
9) Наследственная изменчивость в процессе эволюции:
а) создает новые виды; б) доставляет материал для эволюции; в) закрепляет созданный в процессе эволюции материал; г) сохраняет наиболее полезные изменения.
10) Результатом действия естественного отбора не является:
а) приспособленность организмов к среде обитания; б) многообразие органического мира;
в) борьба за существование; г) совершенствование организации живых существ.
Задание2. Прочитайте текст. Найдите предложения , в которых описывается морфологический критерий.
1.Вероника дубравная растет на полянах, лугах, склонах холмов. 2. Растение имеет ползучее корневище и стебель 10-40см высотой. 3. Листья с зубчатыми краями.4. Цветет вероника дубравная с конца мая по август.5. Опыляется вероника дубравная пчелами и мухами. 6. Цветки небольшие, синего цвета, собраны в соцветие кисть.
Прочитайте текст. Найдите предложения , в которых описывается экологический критерий.
1. Африканский страус –крупная птица массой до 90кг. 2. Обитает в открытых саваннах и полупустынях. 3. Клюв прямой, плоский. 4. Ноги мощные, двупалые, оперение рыхлое. 5. Обычной пищей являются растения-побеги, цветы.6. Африканский страус может длительное время обходится без воды, получая влагу их поедаемых растений.
Задание3. Прочитайте текст. Какие критерии вида указаны в данном тексте, назовите его и укажите номер предложения
Лиственница широко распространена в нашей стране, особенно в Сибири. В высоту достигает 30-40м, а ствол в диаметре-2м. Крона ажурная, ветви редкие, листья – мягкие плоские иголки светло-зеленого цвета. На молодых побегах хвоинки располагаются поодиночке, на старых они собраны в пучки. Лиственница очень светолюбива и холодостойка. Она неприхотлива и может расти на разных почвах – сухих песках, каменистых и заболоченных почвах. Каждую осень лиственница сбрасывает свои иголки, а весной у нее вырастает новая хвоя и образуются мужские и женские шишки. После опыления ветром женские шишки темнеют, становятся похожими на сушеные грибы. Крылатые семена разбрасывает лиственница зимой. Древесина у нее тяжелая, смолистая.
В высоту достигает 30-40м, а ствол в диаметре-2м. Крона ажурная, ветви редкие, листья – мягкие плоские иголки светло-зеленого цвета. На молодых побегах хвоинки располагаются поодиночке, на старых они собраны в пучки. Лиственница очень светолюбива и холодостойка. Она неприхотлива и может расти на разных почвах – сухих песках, каменистых и заболоченных почвах. Каждую осень лиственница сбрасывает свои иголки, а весной у нее вырастает новая хвоя и образуются мужские и женские шишки. После опыления ветром женские шишки темнеют, становятся похожими на сушеные грибы. Крылатые семена разбрасывает лиственница зимой. Древесина у нее тяжелая, смолистая.
Биология для студентов — 06. Теория эволюции Ч. Дарвина. Изменчивость организмов в природе. Борьба за существование и естественный отбор
Формы изменчивости по Ч. Дарвину
Ч. Дарвин установил, между особями одного вида всегда существуют как хорошо заметные различия (спорты), так и малозаметные. Ч. Дарвин считал, что именно мелкие различия между особями являются материалом для отбора.
Эти различия Ч. Дарвин назвал изменчивостью и выделил несколько форм изменчивости:
- определенную,
- неопределенную,
- комбинативную,
- коррелятивную.
Ненаследственная изменчивость является определенной, поскольку, изменяя условия развития организмов, можно предвидеть направление изменчивости. В то же время, она является и групповой, поскольку вся группа особей, подвергаемая одинаковому изменению условия развития, изменяется в одном направлении. В XX веке такую изменчивость долгое время называли модификационной.
Изменчивость неопределенная индивидуальная форма изменчивости, под которой Ч. Дарвин (1859) понимал появление разнообразных отличий у особей одного и того же вида, сорта, породы, которыми, существуя в сходных условиях, одна особь отличается от др. Например, из семян одной коробочки вырастают не тождественные растения, потомки одной пары животны не бывают совершенно подобными, хотя и развиваются в сходных условиях. Характер изменчивости, по мнению Дарвина, определяется не только условиями внешней среды, но и особенностью организма, его состоянием
Характер изменчивости, по мнению Дарвина, определяется не только условиями внешней среды, но и особенностью организма, его состоянием
Коррелятивная изменчивость. Пример с изменением целого комплекса признаков у поросят показывает, что при изменении условий развития изменяется не один признак, а целый комплекс признаков. Одновременное изменение нескольких признаков Ч. Дарвин назвал соотносительной, или коррелятивной изменчивостью.
Наследственная изменчивость также может быть коррелятивной: изменение одного признака влечет за собой изменение других признаков. Например, все белые голубоглазые коты – глухие, у бесшерстных собак определенной породы – недоразвиты зубы. В настоящее время установлено, что наследственная коррелятивная изменчивость может быть обусловлена множественным действием генов, а также сцеплением генов – истинным сцеплением или квазисцеплением.
Комбинативная изменчивость. Во времена Дарвина (еще до опытов Менделя) было известно, что разные наследственные признаки могут образовывать различные комбинации (например, разнообразные сочетания окраски и длины шерсти у животных). Независимую изменчивость признаков Ч. Дарвин назвал комбинативной изменчивостью.
Формы борьбы за существование
Под выражением «борьба за существование» понимается все многообразие отношений между организмами и средой их обитания. Для изучения форм борьбы за существование немецкий эволюционист Э. Геккель создал особый раздел биологии – экологию.
Борьба за существование не прекращается даже в оптимальных условиях, однако при изменении действия лимитирующих или элиминирующих факторов одни ее формы могут заменяться на другие. Результатом борьбы за существование является физическая или генетическая элиминация части организмов.
Существует множество классификаций форм борьбы за существование.
Классификация Л. Моргана и Л. Плате (1906–1916).
Эти авторы выделили три основные формы борьбы за существование:
- Конституциональная борьба с климатическими факторами, или борьба с неблагоприятными условиями среды. Именно от конституции организма (то есть от его анатомо-морфологических, физиологических и биохимических особенностей), в первую очередь, зависит его общая жизнеспособность.
- Межвидовая борьба с конкурентами, хищниками и паразитами. Ведущее место в межвидовой борьбе занимает конкуренция с близкими видами.
- Внутривидовая борьба (конкуренция) за пищу, пространство и возможность размножения. Эта форма борьбы, наиболее напряженная.
 Именно от конституции организма (то есть от его анатомо-морфологических, физиологических и биохимических особенностей), в первую очередь, зависит его общая жизнеспособность.
Именно от конституции организма (то есть от его анатомо-морфологических, физиологических и биохимических особенностей), в первую очередь, зависит его общая жизнеспособность.Классификация А.Н. Северцова и И.И. Шмальгаузена
Выдающиеся отечественные эволюционисты Алексей Николаевич Северцов и Иван Иванович Шмальгаузен рассматривали три группы форм борьбы за существование:
- Пассивное соревнование
При пассивном соревновании наблюдается дифференциальная смертность особей при воздействии различных элиминирующих факторов.
1.1. Прямая элиминация абиотическими факторами (экстремальные температуры, недостаток или избыток влаги, недостаток света, ветер, течение воды, прибой, ливни…).
1.2. Прямая элиминация биотическими факторами (хищники, паразиты, патогенные микроорганизмы…).
1.3. Косвенная элиминация при голоде (голод усиливает действие прямых элиминирующих факторов).
- Активная конкуренция
Активная конкуренция наблюдается при наличии у разных групп организмов общих ресурсов, дефицит которых является лимитирующим фактором.
2.1. Прямая, или интерференционная конкуренция. Особи вступают «в схватку» (прямой контакт) за свет, воду, пищу, возможность для размножения.
2.2. Косвенная, или эксплуатационная конкуренция. Особи соревнуются при добывании пищи и других ресурсов, не вступая в непосредственную «схватку».
2.3. Диффузная конкуренция. Возникает при взаимодействии нескольких конкурирующих групп.
Конкуренция может быть межвидовой (при перекрывании экологических ниш) и межвидовой.
- Различные формы биотических взаимодействий
3. 1. Внутривидовой каннибализм, в том числе, и в пределах одной возрастной группы.
1. Внутривидовой каннибализм, в том числе, и в пределах одной возрастной группы.
3.2. Внутривидовой альтруизм («самопожертвование»), например, у рабочих пчел и муравьев.
3.3. Межвидовой альтруизм (протокооперация). Мюллеровская мимикрия (сходство многих защищенных видов перепончатокрылых, например, ос, шмелей, пчел).
3.4. Комменсализм («нахлебничество»).
3.5. Информационный паразитизм: бэтсовская мимикрия, или миметизм (сходство защищенных и незащищенных видов, например, жалящих перепончатокрылых и бабочек–стеклянниц).
3.6. Различные формы симбиоза (мутуализм).
Дарвин и генетика | Генетика
Abstract
Дарвиновской теории естественного отбора не хватало адекватного объяснения наследования, что делало ее логически неполной. Мы рассматриваем взаимодействие между эволюцией и генетикой, показывая, как, в отличие от Менделя, отсутствие у Дарвина модели механизма наследования сделало его неспособным интерпретировать свои собственные данные, показывающие менделевские отношения, даже несмотря на то, что он разделял с Менделем более математические и вероятностные взгляды. чем большинство биологов его времени.Собственная модель «пангенезиса» Дарвина предоставила механизм для создания обширной изменчивости, на которую мог воздействовать отбор. Однако это связано с наследованием признаков, приобретенных в течение жизни организма, что, как знал сам Дарвин, не может объяснить некоторые эволюционные ситуации. Как только были поняты основы генетики в виде частиц, стало ясно, что они позволяют передавать вариации в неизменном виде новым поколениям, и затем эволюцию можно было понять как процесс изменения частот стабильных вариантов.Впоследствии эволюционная генетика стала центральной частью биологии. Принципы Дарвина сейчас играют большую роль в биологии, чем когда-либо прежде, что мы проиллюстрируем некоторыми примерами исследований естественного отбора, в которых используются данные о последовательностях ДНК, и некоторыми недавними достижениями в ответах на вопросы, впервые заданные Дарвином.
Сила отбора, реализуемая человеком или задействованная природой в борьбе за существование и последующее выживание наиболее приспособленных, полностью зависит от изменчивости органических существ.Без изменчивости ничего не получится; однако небольших индивидуальных различий достаточно для работы и, вероятно, они являются главным или единственным средством создания новых видов.
Чарльз Дарвин (1868)
ЧАРЛЬЗ Дарвин был первым, кто четко осознал, что эволюция зависит от существования наследственной изменчивости внутри вида, порождающей различия между предками и потомками. Развитие представлений Дарвина о природе и причинах эволюции четко задокументировано в его записных книжках «трансмутации» 1836–1838 гг. (Barrett et al. 1987). Решив, что виды произошли в результате «происхождения с модификацией», Дарвин быстро осознал необходимость найти механизм для выполнения соответствующих изменений. При формулировании идеи естественного отбора на него большое влияние оказал опыт селекционеров по искусственному отбору популяций домашних животных и растений. Глава 1 из Происхождение видов (Дарвин, 1859) знаменито посвящена документированию существования изменчивости в этих популяциях и эффективности искусственного отбора:
Ключ — это способность человека к кумулятивному отбору: природа дает последовательные вариации; человек складывает их в определенных направлениях, полезных для себя (Darwin 1859, p.30).
Это был лишь небольшой шаг до применения этого наблюдения к отбору в природе:
Можно ли считать это невероятным, учитывая, что полезные для человека вариации, несомненно, имели место, что другие вариации, полезные каким-то образом для каждого существа в великой и сложной жизненной битве, должны иногда происходить в течение тысяч поколений? … Это сохранение благоприятных вариаций и отказ от вредных вариаций я называю естественным отбором (Darwin 1859, стр.
 81–82).
81–82).
Большинство книг и статей, опубликованных Дарвином после Происхождение видов , были посвящены описанию того, как широкий спектр биологических явлений — от половых систем растений до анатомии и поведения человека — может быть интерпретирован с точки зрения эволюции естественный отбор или особая форма естественного отбора, представленная половым отбором. Удивительно (по крайней мере, с сегодняшней точки зрения), но многие биологи долгое время были далеки от убеждений в том, что естественный отбор является преобладающей движущей силой эволюции.Так продолжалось до 1920-х годов. Во введении к тому 1 своего трактата по эволюционной генетике Сьюэлл Райт отметил:
Наряду с всеобщим принятием биологами эволюции как факта, во второй половине девятнадцатого века росло недовольство естественным отбором как основной теорией причинности (Wright 1968, стр.7-8).
СКЕПТИЗМ В ОТНОШЕНИИ ЕСТЕСТВЕННОГО ОТБОРА
Выдающиеся ранние генетики, такие как Уильям Бейтсон, Хьюго де Фриз и Ричард Гольдшмидт, были печально известными скептиками в отношении естественного отбора и эволюционной роли небольших индивидуальных различий, на которые опирался Дарвин, вместо этого подчеркивая роль мутаций с большие и разнообразные эффекты (Provine 1971).Многие натуралисты и палеонтологи придерживались того, что сейчас кажется нам полумистическими теориями, такими как внутренние стремления к улучшению или совершенствованию; многие из них придерживались ламаркистских взглядов вплоть до 1930-х годов (во Франции и в Советском Союзе и его сателлитах ламаркизм сохранялся вплоть до 1960-х годов). В своей классической истории современной науки, The Edge of Objectivity , Чарльз Коулстон Гиллиспи цитирует ведущего историка биологии 1929 года Эрика Норденшельда, который заявил, что утверждение о том, что естественный отбор «не действует в той форме, в которой он был представлен Дарвином, безусловно, должен считаться доказанным »(Gillispie 1960, p. 320). Книга Evolution в свете современных знаний , сборник эссе 13 ведущих британских биологов, опубликованный Blackie and Son в 1925 году, чтобы предоставить (согласно примечанию издателя) «авторитетное заявление» о «доктрине эволюция … после всеобщего переворота фундаментальных теорий за последние 20 лет », не имеет указателя на естественный отбор. Это контрастирует с 3253 статьями, упоминающими естественный отбор и эволюцию в 2008 году в базе данных Web of Science.Подробное обсуждение антидарвиновских эволюционных идей см. В Bowler (1983) и Gayon (1998).
320). Книга Evolution в свете современных знаний , сборник эссе 13 ведущих британских биологов, опубликованный Blackie and Son в 1925 году, чтобы предоставить (согласно примечанию издателя) «авторитетное заявление» о «доктрине эволюция … после всеобщего переворота фундаментальных теорий за последние 20 лет », не имеет указателя на естественный отбор. Это контрастирует с 3253 статьями, упоминающими естественный отбор и эволюцию в 2008 году в базе данных Web of Science.Подробное обсуждение антидарвиновских эволюционных идей см. В Bowler (1983) и Gayon (1998).
Почему был такой скептицизм по отношению к естественному отбору и почему все так сильно изменилось? Одной из причин было отсутствие при жизни Дарвина прямых доказательств естественного отбора. Это начало меняться в конце 19-го и начале 20-го веков благодаря работам Бампуса (1899) в Соединенных Штатах и Велдона (1895, 1901) и его ученика Ди Чеснола (1907) в Европе. Эти ученые положили начало области, ныне известной как экологическая генетика, и теперь у нас есть буквально тысячи примеров, когда полевые натуралисты продемонстрировали действие естественного отбора в дикой природе как на дискретные полиморфизмы, так и на количественные признаки (Kingsolver et al. 2001; Bell 2008; Лейму и Фишер, 2008 г.).
ТРУДНОСТИ ДАРВИНА С НАСЛЕДОВАНИЕМ: МЕНДЕЛ ДЛЯ СПАСЕНИЯ
Другим важным фактором, конечно же, был тот факт, что Дарвин не смог прийти к пониманию механизма наследования, несмотря на то, что осознавал его важность и прилагал огромные усилия для сбора доказательств в его Вариациях животных и растений при одомашнивании (Дарвин 1868). К сожалению, он не знал о работе Менделя, несмотря на то, что она была опубликована двумя годами ранее (Mendel 1866).Работа Менделя теперь, конечно, навсегда изменила наше понимание наследственности, и его трагическая неспособность получить признание при жизни — хорошо известная история. Менее известно, что Мендель хорошо осознавал важность понимания генетики для эволюции:
Это кажется единственным правильным способом окончательного решения вопроса, значение которого для эволюционной истории органических форм нельзя недооценивать (Mendel 1866, p.
 2).
2).
К сожалению, даже если бы Мендель дожил до повторного открытия своей работы, он, вероятно, не был бы удовлетворен, увидев ее вклад в эволюционное понимание, потому что даже после того, как генетика начала свое быстрое развитие в первые десятилетия 20-го века. , биологи-эволюционисты изначально не понимали, как включить генетику в свою работу. Мы обозначим эти неудачи для синтеза позже, но сначала рассмотрим попытки Дарвина понять наследование и то, как его подход не соответствовал подходу Менделя.
Способность Менделя решать самую сложную проблему биологии 19 века после механизма эволюции основывается на использовании им уникального на тот момент подхода: сочетания строгих генетических экспериментов с количественными, вероятностными предсказаниями их ожидаемых результатов: другими словами, использование биологических данные для проверки количественной гипотезы. То, что Мендель предложил свою гипотезу наследования частиц задолго до того, как биологи животных или растений достигли правильного понимания клеточной основы полового размножения, является триумфом продуктивных теоретических рассуждений (Farley 1982).
Это достижение ускользнуло от Дарвина, другого величайшего ума в биологии 19-го века, хотя он был близок к тому, чтобы увидеть те же явления, что и Мендель, и часто рассматривал данные в количественном отношении (Howard 2009). Дарвин неоднократно упоминал феномен возврата к предковым типам в «Вариация животных и растений при одомашнивании » (Дарвин 1868). Он также собрал примеры передачи признаков от нескольких поколений родословных и получил помощь от математического физика сэра Джорджа Стоукса, чтобы показать, что эти случаи вряд ли возникли случайно, — один из первых примеров теста на статистическую значимость в биологии (Дарвин 1868, гл.12).
По иронии судьбы, Дарвин проанализировал данные своих собственных экспериментов по скрещиванию дистилли у видов Primula (суммированных в Darwin 1877, глава 5), которые дали то, что мы теперь можем рассматривать как четкое свидетельство менделевских соотношений (см. Также Bulmer 2003, p.112). , и Ховард 2009). У дистильных видов (Рисунок 1) теперь известно, что морфы с длинным стилем (L) являются гомозиготами ss по аллелям в нескольких локусах в супергене, контролирующем длину стиля, положение тычинок, расположение пыльцы и стигмы, морфологию и несовместимость. , тогда как морф с коротким стилем (S) является гетерозиготным Ss .Единственные спаривания, которые неизменно успешны, — это L × S и S × L (Дарвин назвал это «законное опыление»), и они дают соотношение L и S растений 1: 1 (Таблица 1). Иногда возможно получить семена путем самоопыления, и в этом случае L-растения дают только L-потомство (Таблица 1). Дарвин заявил:
Также Bulmer 2003, p.112). , и Ховард 2009). У дистильных видов (Рисунок 1) теперь известно, что морфы с длинным стилем (L) являются гомозиготами ss по аллелям в нескольких локусах в супергене, контролирующем длину стиля, положение тычинок, расположение пыльцы и стигмы, морфологию и несовместимость. , тогда как морф с коротким стилем (S) является гетерозиготным Ss .Единственные спаривания, которые неизменно успешны, — это L × S и S × L (Дарвин назвал это «законное опыление»), и они дают соотношение L и S растений 1: 1 (Таблица 1). Иногда возможно получить семена путем самоопыления, и в этом случае L-растения дают только L-потомство (Таблица 1). Дарвин заявил:
Дистилли в примулах. (A) Длиннопестистые (иглы) и коротковистые (барабанные) цветки Primula vulgaris . (B) Вертикально разделенные цветы с указанием совместимых видов опыления.(Пыльца с высоких пыльников совместима с рыльцами длиннопестичных растений, а пыльца с низких пыльников совместима с рыльцами короткоствольных растений, в то время как два других типа опыления несовместимы.)
ТАБЛИЦА 1Результаты Дарвина для потомства длинно- и короткоперистых примул, скрещенных с одной и той же морфой
Из длиннопестичной формы, всегда оплодотворенной пыльцой собственной формы, в первом поколении я вырастил три длиннопестичных растения, из их семян 53 длиннопестичных внука, из их семян 4 длиннопестичных правнука, из их семя 20 праправнуков с длинным стилем и, наконец, от их семени 8 праправнуков с длинным стилем и 2 праправнука с коротким стилем.… Всего было выращено 162 растения,… 156 длиннопестичных и 6 короткопестичных (Darwin 1877, стр. 228–229).
Несколько короткоствольных растений в последнем поколении предположительно были контаминантами (эксперименты Дарвина на удивление не содержали их). Самоопыление S-растений должно приводить к соотношению 3: 1, как обнаружил Дарвин (см. Таблицу 1; ни одно из соотношений не отличается значительно от ожидаемого соотношения). Он отметил:
Он отметил:
Сначала я сделал рейз с короткостайловой [ P.sinensis ] растение, удобренное пыльцой собственной формы: один длиннопестичный и семь короткоствольных незаконнорожденных саженцев…. Доктор Хильдебранд вырастил четырнадцать растений, из которых одиннадцать были короткоствольными и три длиннопестистыми (Дарвин 1877, стр. 216).
ТРУДНОСТИ ДАРВИНА С СМЕШИВАНИЕМ НАСЛЕДОВАНИЯ И ЕГО ТЕОРИИ ПАНГЕНЕЗА
Дарвин не понимал значения этих результатов, потому что у него не было модели наследования частиц, которую можно было бы применить к генетическим данным.В самом деле, Дарвин, похоже, сохранял веру в преобладание смешанного наследования, как и почти все его современники. Как указал Фишер в главе 1 книги Генетическая теория естественного отбора (Фишер, 1930), в опубликованных работах Дарвина есть несколько явных заявлений на этот счет, хотя они появляются в некоторых из его неопубликованных заметок и эссе. Кроме того, глава 15 документа «Разнообразие животных и растений при одомашнивании » (Дарвин 1868) начинается с раздела «О скрещивании как причине единообразия признака », в котором неявно предполагается, что скрещивание ведет к смешиванию.Однако неясно, в какой степени он считал потомство продуктом полного слияния генетического вклада его родителей (Bulmer 2003, гл. 4).
Смешивание наследования приводит к затруднению, на которое убедительно указал Флиминг Дженкин (Jenkin 1867), профессор инженерии в Эдинбургском университете (здание рядом с нашим, к сожалению, возможно, названо в его честь). При смешанном наследовании вариабельность быстро затухает, потому что все генотипы потомков скрещивания одинаковы и являются промежуточными между генотипами двух родителей.При случайном спаривании генетическая дисперсия количественного признака затем уменьшается вдвое в каждом поколении (Fisher 1930, p. 4). Принятие смешанного наследования явно вызывает сомнения в способности естественного или искусственного отбора вносить постоянные изменения в популяции. В шестом издании Происхождение видов , опубликованном в 1872 году, Дарвин отреагировал на Дженкина следующим образом:
В шестом издании Происхождение видов , опубликованном в 1872 году, Дарвин отреагировал на Дженкина следующим образом:
Тем не менее, до тех пор, пока я не прочитал способную и ценную статью в «North British Review» (1867 г.), я не осознавал, как редко отдельные вариации, незначительные или сильно выраженные, могут сохраняться (Darwin 1859, стр.111–112).
Поскольку для того, чтобы отбор был эффективным, требуется наследственная изменчивость, а Дарвинский обзор результатов искусственного отбора убедил его в том, что существует достаточно вариаций, чтобы он был эффективным, Дарвин искал способ создать изобилие таких вариаций. Это было обеспечено его теорией пангенезиса, согласно которой вариации, испытываемые человеком в течение его жизни, передаются в половые клетки гипотетическими «геммулами» (Darwin 1868, гл.27). Это гипотеза о наследовании приобретенных признаков, которую Дарвин принял как экспериментально установленный факт (есть обширное обсуждение передачи увечий в Darwin 1868, глава 12).
Однако Дарвин явно не был в этом уверен. Например, он упомянул, что обрезание младенцев мужского пола не привело к потере крайней плоти в еврейской общине (Darwin 1868, Vol. 1, p. 558). Он также отметил, что есть некоторые примеры эволюции, которые не могут быть объяснены этой гипотезой, в частности адаптивные характеристики стерильных каст социальных насекомых:
Ибо никакие упражнения, привычки или воления совершенно бесплодных членов сообщества не могли повлиять на структуру или инстинкты плодовитых членов, которые сами по себе оставляют потомков.Я удивлен, что никто не выдвинул этот показательный случай стерильных насекомых против хорошо известной доктрины Ламарка (Darwin 1859, p. 242).
Использование Дарвином этого естественного случая селекции сибсов для опровержения ламаркизма предвосхищает более позднее использование тех же рассуждений, которые достигли пика совершенства в экспериментах Ледербергов по нанесению покрытия на реплики Escherichia coli (Lederberg and Lederberg 1952).
ЭВОЛЮЦИЯ КАК ПРОЦЕСС ИЗМЕНЕНИЯ ЧАСТОТ МЕНДЕЛЬСКИХ ВАРИАНТОВ В НАСЕЛЕНИЯХ
В отличие от Дарвина, который рассматривал наследование приобретенных признаков в основном как источник вариаций, на которые может действовать отбор, сторонники ламарковского наследования 20-го века рассматривали его как альтернативное объяснение адаптивной эволюции.Как было блестяще изложено Фишером в главе 1 книги «Генетическая теория естественного отбора » и, несомненно, знакомо читателям «Генетики», все трудности, создаваемые смешиванием, исчезают вместе с менделевским наследованием: изменчивость в популяции сохраняется, а не потерянный, когда никакие эволюционные силы не действуют, генетический эквивалент закона инерции Галилея. Следовательно, наследование приобретенных признаков не требуется для восстановления генетической изменчивости.
Конечно, хорошо известно, что наши знания физических основ генов и их поведения в настоящее время в значительной степени исключают наследование по Ламарку.Однако недавние исследования выявили некоторые ситуации, в которых ДНК определенных участков генома модифицируется в течение жизни человека, и эти эпигенетические метки с функциями контроля развития и других процессов могут иногда передаваться через мейотические деления к потомкам (, например, ). , Cubas и др. 1999; Richards 2006; Namekawa et al. 2007; Heijmans et al. 2008; Сидоренко и Чендлер 2008). Если варианты, которые возникают таким образом, стабильно передаются, то их можно рассматривать как менделевские варианты, которые можно использовать в эволюции.Если их наследование нестабильно, как это часто бывает, они не могут вносить значительный вклад в эволюцию.
Прорыв в понимании природы вариации количественных признаков (эквивалент Дарвина «незначительные различия, которые можно назвать« индивидуальными различиями »; см. Эпиграф к этой статье) произошел в первые годы генетики, начиная с экспериментов с чистыми линии, особи которых имеют практически идентичные генотипы. Эти эксперименты показали, что среди таких индивидуумов существует обильная фенотипическая изменчивость, но они не передаются потомству (Johannsen 1909; Wright 1920), что привело к отказу генетического сообщества от наследования по Ламарку.Кроме того, вариабельность количественных признаков (которые часто демонстрируют очевидное смешение в скрещиваниях F 1 между чистыми линиями) увеличивается у F 2 и более поздних поколений (Nilsson-Ehle 1909; East 1910), как и ожидалось с частичным менделевским наследованием. Более того, ответственные факторы могут быть сопоставлены с хромосомными регионами, а иногда (с помощью современных методов) с отдельными генами или вариантами нуклеотидов (Flint and Mackay 2009). Даже изначально озадачивающие случаи очень сложных моделей наследования, таких как бусинки и усеченное крыло у дрозофилы, были связаны с факторами, связанными с хромосомными генами, и виртуальная универсальность менделизма была установлена к началу 1920-х годов (Altenburg and Muller 1920).В отличие от наследования приобретенных признаков, мутации оказались очень редкими, стабильными модификациями генов и возникали независимо от того, придают ли они повышенную приспособленность в данной среде (Muller 1932).
Эти эксперименты показали, что среди таких индивидуумов существует обильная фенотипическая изменчивость, но они не передаются потомству (Johannsen 1909; Wright 1920), что привело к отказу генетического сообщества от наследования по Ламарку.Кроме того, вариабельность количественных признаков (которые часто демонстрируют очевидное смешение в скрещиваниях F 1 между чистыми линиями) увеличивается у F 2 и более поздних поколений (Nilsson-Ehle 1909; East 1910), как и ожидалось с частичным менделевским наследованием. Более того, ответственные факторы могут быть сопоставлены с хромосомными регионами, а иногда (с помощью современных методов) с отдельными генами или вариантами нуклеотидов (Flint and Mackay 2009). Даже изначально озадачивающие случаи очень сложных моделей наследования, таких как бусинки и усеченное крыло у дрозофилы, были связаны с факторами, связанными с хромосомными генами, и виртуальная универсальность менделизма была установлена к началу 1920-х годов (Altenburg and Muller 1920).В отличие от наследования приобретенных признаков, мутации оказались очень редкими, стабильными модификациями генов и возникали независимо от того, придают ли они повышенную приспособленность в данной среде (Muller 1932).
К 1920-м годам стало ясно, что (вопреки убеждениям многих ранних генетиков, которые подчеркивали значительный эффект драматических мутаций и игнорировали доказательства менделевской основы количественной изменчивости признаков) дарвиновская эволюция естественным отбором Фактически, благодаря менделевскому наследованию: мутации в генах обеспечивают источник новых, стабильных вариантов, на которые может воздействовать отбор.Это подготовило почву для понимания того, что эволюция — это, по сути, процесс изменения частот менделевских вариантов внутри популяций и видов, ведущий к развитию классической популяционной и количественной генетики. Увлекательная борьба за достижение этого понимания хорошо описана Provine (1971).
НАСКОЛЬКО НАТУРАЛЬНЫЙ ОТБОР ДЛЯ СОВРЕМЕННОЙ БИОЛОГИИ?
Главный постдарвиновский компонент, имеющий большое значение в современном эволюционном мышлении, — это идея генетического дрейфа и, в частности, возможность того, что значительная часть изменчивости и эволюции вариантов последовательности ДНК вызвана случайными колебаниями в частотах вариантов с мало или совсем не влияет на физическую форму (Kimura 1983). У самого Дарвина была идея избирательного нейтралитета:
У самого Дарвина была идея избирательного нейтралитета:
Вариации, ни полезные, ни вредные, не будут затронуты естественным отбором и останутся колеблющимся элементом, как, возможно, мы видим у видов, называемых полиморфными (Darwin 1859, p. 81).
В неожиданном повороте событий концепция избирательного нейтралитета стала краеугольным камнем современных тестов естественного отбора, предоставив нулевую гипотезу, которая может быть проверена на данных по изменчивости и эволюции последовательностей.Эволюционная биология в настоящее время достаточно созрела, чтобы выплатить долг генетике, и действительно теперь (вместе с генетическими и молекулярно-генетическими подходами) является центральной для работы, инициированной в основном с функциональными генетическими мотивами, включая секвенирование генома.
Учитывая некоторую генетическую изменчивость интересующего фенотипа, экологические генетические подходы могут соотносить приспособленность с различиями между индивидуумами в пределах одной естественной популяции, иногда используя данные о ненарушенных особях (Bell 2008).При большем беспокойстве организмов различия между популяциями также могут быть проверены на их селективную значимость с помощью таких методов, как эксперименты по реципрокной трансплантации. Изменения частот генотипов можно проследить с течением времени в таких экспериментах или после отклонения аллелей от их собственных частот. Эти подходы четко документировали действие отбора, иногда на неожиданные признаки, такие как инверсионные полиморфизмы Drosophila (Wright and Dobzhansky 1946). Однако этот подход может упустить многие случаи отбора, потому что даже самые крупные и наиболее чувствительные эксперименты, такие как эксперименты с конкуренцией между штаммами дрожжей или бактерий, не могут обнаружить селективные различия <∼0.1% по величине (Dykhuizen 1990).
На другом краю эволюционной шкалы времени можно использовать сравнительный подход, чтобы связать различия в экологических условиях, в которых находятся разные развивающиеся линии, с различиями в результатах эволюции путем естественного отбора (Harvey and Pagel 1991). Дарвин был первым биологом, который явно использовал сравнительный подход для этой цели. Этот подход в настоящее время является высоко статистическим (Felsenstein 2004) и часто использует филогении на основе последовательностей, которые имеют то преимущество, что они гораздо менее восприимчивы к действию естественного отбора, вызывая вариации в скорости и направлении изменения признаков, чем ранее использовавшиеся морфологические признаки в филогенетическом анализе.Даже не имея современных методов, Дарвин с успехом использовал сравнительный метод в своей работе по эволюции системы спаривания растений, например, в своем обзоре литературы, чтобы показать, что инбридинговые растения имеют меньшие цветки и, как правило, менее привлекательны для опылителей по сравнению с ауткроссингами (Дарвин, 1876 г.), открытие, подтвержденное более обширными современными исследованиями и говорящее нам о том, что для привлечения опылителей требуются ресурсы (, например, , Ornduff 1969). Однако сравнительный подход неспособен дать оценку интенсивности отбора, вызвавшего наблюдаемые изменения.
Дарвин был первым биологом, который явно использовал сравнительный подход для этой цели. Этот подход в настоящее время является высоко статистическим (Felsenstein 2004) и часто использует филогении на основе последовательностей, которые имеют то преимущество, что они гораздо менее восприимчивы к действию естественного отбора, вызывая вариации в скорости и направлении изменения признаков, чем ранее использовавшиеся морфологические признаки в филогенетическом анализе.Даже не имея современных методов, Дарвин с успехом использовал сравнительный метод в своей работе по эволюции системы спаривания растений, например, в своем обзоре литературы, чтобы показать, что инбридинговые растения имеют меньшие цветки и, как правило, менее привлекательны для опылителей по сравнению с ауткроссингами (Дарвин, 1876 г.), открытие, подтвержденное более обширными современными исследованиями и говорящее нам о том, что для привлечения опылителей требуются ресурсы (, например, , Ornduff 1969). Однако сравнительный подход неспособен дать оценку интенсивности отбора, вызвавшего наблюдаемые изменения.
Современная технология секвенирования ДНК дает популяционным генетикам возможность изучать степень, в которой отбор влияет на варианты в геноме, в отличие от мутаций и случайного генетического дрейфа. После нескольких десятилетий использования экологических генетических и сравнительных подходов для выявления естественного отбора по видимым или физиологическим признакам биологи теперь могут проверять избирательные эффекты конкретных генетических различий между людьми, не зная их фенотипических эффектов.Для этих тестов нейтральность обеспечивает важную нулевую гипотезу. Благодаря нашей недавно приобретенной способности применять методы статистической популяционной генетики для анализа моделей внутривидовой изменчивости и межвидовой дивергенции в больших, полногеномных наборах данных, можно обнаружить чрезвычайно слабое давление отбора, намного ниже разрешающей способности экспериментальных методов. и измерил. Многие из используемых в настоящее время подходов во многом основаны на классической работе Фишера, Кимуры и Райта о поведении вариантов, подверженных мутации, отбору и генетическому дрейфу, которые кратко изложены в книге Кимуры (1983), The Neutral Theory Молекулярной эволюции .Эти методы часто требуют больших вычислительных ресурсов, особенно если принять во внимание такие осложнения, как недавние изменения численности популяции.
и измерил. Многие из используемых в настоящее время подходов во многом основаны на классической работе Фишера, Кимуры и Райта о поведении вариантов, подверженных мутации, отбору и генетическому дрейфу, которые кратко изложены в книге Кимуры (1983), The Neutral Theory Молекулярной эволюции .Эти методы часто требуют больших вычислительных ресурсов, особенно если принять во внимание такие осложнения, как недавние изменения численности популяции.
С увеличением доступности больших наборов данных об изменении последовательности ДНК в геномах человека и Drosophila melanogaster , мы приближаемся к ответам на такие вопросы, как: Каково распределение коэффициентов отбора для вновь возникающих вредных мутаций аминокислот? Какая доля вариантов аминокислот, различающих родственные виды, фиксируется естественным отбором, в отличие от генетического дрейфа, действующего на нейтральные или слегка вредные варианты? В какой степени варианты на сайтах синонимичного кодирования и на сайтах некодирования подлежат отбору и насколько сильна эта отборка?
Иногда результаты просто поразительны.Например, довольно убедительно установлено, что типичный человек является гетерозиготным по нескольким сотням аминокислотных мутаций, большинство из которых оказывают лишь очень небольшое влияние на приспособленность (порядка 10 -3 ) (Boyko et al. 2008), что около 50% вариантов аминокислот, различающих родственные виды дрозофилы, были зафиксированы путем отбора (Sella et al. 2009), и что больше некодирующих сайтов, чем кодирующих сайтов, как у людей, так и у дрозофилы могут мутировать в селективно вредные альтернативы, которые быстро удаляются выбором (Encode Project Consortium 2007; Haag-Liautard et al. 2007).
В дополнение к этим прямым тестам отбора вариантов, мы также можем использовать информацию о нейтральных или почти нейтральных вариантах, которые сами не подвергаются отбору, чтобы делать выводы о выборе на связанных сайтах в геноме. Одним из примеров является обнаружение выборочных зачисток, вызванных недавним распространением селективно благоприятных мутаций. Распространение полезного аллеля может быстро привести к очень низкой вариабельности затронутого гена, а тесно связанные области также могут иметь уменьшенное разнообразие в результате автостопа через популяцию сегмента хромосомы, содержащего исходную полезную мутацию (Мейнард Смит и Хей 1974).Эти эффекты на связанные нейтральные или почти нейтральные варианты могут быть использованы в статистических тестах действия естественного отбора. Это позволило генетикам обнаружить и оценить силу отбора, воздействующего на гены, такие как гены устойчивости к лекарствам у малярийного паразита человека, с помощью вариабельности микросателлитных маркеров (, например, , Nash и др. 2005), чтобы обнаружить многочисленные примеры недавние адаптации в человеческих популяциях из-за их влияния на паттерны изменчивости связанных SNP ( e.грамм. , Currat et al. 2002; Sabeti et al. 2002; Williamson et al. 2007; Akey 2009) и для поиска генов, участвующих в ответах на искусственный отбор (Walsh 2008).
Одним из примеров является обнаружение выборочных зачисток, вызванных недавним распространением селективно благоприятных мутаций. Распространение полезного аллеля может быстро привести к очень низкой вариабельности затронутого гена, а тесно связанные области также могут иметь уменьшенное разнообразие в результате автостопа через популяцию сегмента хромосомы, содержащего исходную полезную мутацию (Мейнард Смит и Хей 1974).Эти эффекты на связанные нейтральные или почти нейтральные варианты могут быть использованы в статистических тестах действия естественного отбора. Это позволило генетикам обнаружить и оценить силу отбора, воздействующего на гены, такие как гены устойчивости к лекарствам у малярийного паразита человека, с помощью вариабельности микросателлитных маркеров (, например, , Nash и др. 2005), чтобы обнаружить многочисленные примеры недавние адаптации в человеческих популяциях из-за их влияния на паттерны изменчивости связанных SNP ( e.грамм. , Currat et al. 2002; Sabeti et al. 2002; Williamson et al. 2007; Akey 2009) и для поиска генов, участвующих в ответах на искусственный отбор (Walsh 2008).
И наоборот, высокая изменчивость в регионе может выдавать действие естественного отбора, действующего таким образом, что различные аллели сохраняются как полиморфизмы в течение длительного периода за счет уравновешивания отбора; это делит популяцию на два или более компартментов, между которыми нейтральные различия могут накапливаться в сайтах, которые тесно связаны с мишенями отбора, где рекомбинация неэффективна для предотвращения дифференциации между компартментами (Hudson 1990; Nordborg 1997).Хорошо известным примером является область MHC человека, в которой не только много полиморфных аминокислот в экзоне 2, который кодирует большую часть пептид-связывающих остатков зрелых белков MHC человека, но и после чрезвычайно многочисленных полиморфных вариантов синонимичные и интронные сайты по сравнению с другими локусами в тех же популяциях (Raymond et al.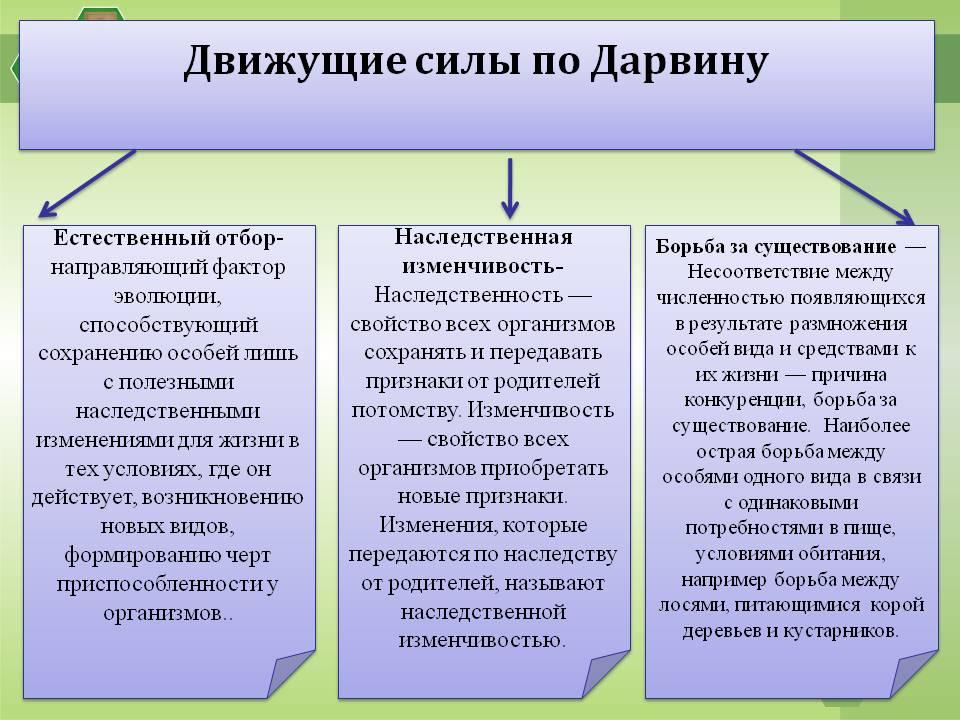 2005). Точно так же частотно-зависимый отбор четко поддерживает полиморфизм последовательностей в течение длительного времени эволюции в локусах самонесовместимости растений и грибов, последовательности которых сильно расходятся ( e.грамм. , Векеманс, Слаткин 1994; Richman et al. 1996; мая и др. 1999).
2005). Точно так же частотно-зависимый отбор четко поддерживает полиморфизм последовательностей в течение длительного времени эволюции в локусах самонесовместимости растений и грибов, последовательности которых сильно расходятся ( e.грамм. , Векеманс, Слаткин 1994; Richman et al. 1996; мая и др. 1999).
С помощью молекулярно-эволюционных подходов можно изучать не только отбор в пределах одной популяции, но и различия между популяциями, обусловленные местной адаптацией, также можно выявить по увеличению дивергенции на участках, связанных с мишенями отбора (Beaumont and Balding 2004). Действительно, сканирование геномов человека и других видов на предмет последовательностей, более дифференцированных, чем большинство генов, является основным способом обнаружения кандидатов на гены, которые в настоящее время проходят отбор (Akey 2009).
СОВРЕМЕННЫЙ ПРОГРЕСС В ОТВЕТЕ НА НЕКОТОРЫЕ СТАРЫЕ ВОПРОСЫ: ИСПОЛЬЗОВАНИЕ ПРИРОДНЫХ ГЕНЕТИЧЕСКИХ МАРКЕРОВ
Другой способ, которым современные эволюционные исследования внесли свой вклад в генетику, в отличие от генетики, способствующей эволюционной биологии, — это интерес к количественной оценке степени генетической изменчивости. [первоначально мотивированные дебатами о том, состоит ли вариация внутри вида в значительной степени из недавних мутаций или включает значительную долю вариантов, поддерживаемых балансирующим отбором (Добжанский, 1955; Левонтин, 1974)] в конечном итоге привели к открытию огромного числа вариантов последовательностей ДНК, которые могут могут использоваться в качестве генетических маркеров для картирования (хотя эти данные сами по себе не разрешают споры о том, поддерживает ли отбор вариативность).Существование обильных маркеров было предсказано давно:
Соответственно, в случае человека было бы желательно провести обширный и тщательный поиск как можно большего числа факторов, которые можно было бы использовать… в качестве идентификаторов.
 Они должны, предпочтительно, включать в себя различия характера, которые: (1) являются обычным явлением, (2) могут быть идентифицированы с уверенностью, (3) наследуются в простой менделевской манере. Кажется разумным предположить, что у такого гетерозиготного вида действительно должно быть бесчисленное множество таких факторов.… Кажется очевидным, что у более податливых организмов, таких как одомашненные и лабораторные расы животных и растений, анализ характера посредством изучения связи с идентифицирующими факторами получит более широкое применение (Altenburg and Muller 1920).
Они должны, предпочтительно, включать в себя различия характера, которые: (1) являются обычным явлением, (2) могут быть идентифицированы с уверенностью, (3) наследуются в простой менделевской манере. Кажется разумным предположить, что у такого гетерозиготного вида действительно должно быть бесчисленное множество таких факторов.… Кажется очевидным, что у более податливых организмов, таких как одомашненные и лабораторные расы животных и растений, анализ характера посредством изучения связи с идентифицирующими факторами получит более широкое применение (Altenburg and Muller 1920).
У некоторых видов естественные маркеры теперь могут быть получены настолько плотно, что необходимы новые подходы для генетического картирования, потому что существует очень низкая вероятность перекрестного события между ближайшими маркерами ( e.грамм. , Черчилль и др. 2004; Ван Ос и др. 2005; Flibotte et al. 2009). Возможность получения большого количества генетических маркеров привела к новому прогрессу в картировании генов, влияющих на количественные признаки, и для таких исследований разрабатываются новые подходы, в том числе ассоциативное картирование, в котором используется популяционная генетическая концепция неравновесия по сцеплению (ассоциации между аллельными состояниями различных локусов или сайтов в последовательности; см. обзор в Slatkin 2008).Изучение популяционной генетики мультилокусных систем когда-то казалось эзотерической областью, далекой от эмпирических данных, что способствовало репутации теоретической популяционной генетики как сухой и не имеющей отношения к биологии. Тем не менее, были разработаны очень важные принципы, которые сейчас широко используются другими генетиками, включая способы измерения неравновесия по сцеплению [также теперь используются для оценки скорости рекомбинации в геномах с использованием выборок последовательностей из популяций (Myers et al. 2005)] и концепция, согласно которой отбор, действующий на данный вариант последовательности или аллель, оказывает влияние на тесно связанные варианты (см. Выше).
Выше).
Виды подходов, о которых только что упоминалось, больше не ограничиваются людьми и генетическими модельными организмами, представляющими наибольший интерес для функциональной молекулярно-генетической работы. Одно из общепринятых способов использования маркеров — это определение систем спаривания популяций в дикой природе (Ritland 1990). Дарвин предвидел это, когда использовал фенотипические различия, в том числе окраску цветков, которые, как он явно предполагал, передавались по наследству, чтобы сделать вывод о происхождении семян:
Всего было выращено 233 растения, из которых 155 были беспородными, а из остальных 78 не половина были абсолютно чистыми.Я повторил эксперимент, посадив рядом две разновидности капусты с пурпурно-зелеными и бело-зелеными лаковыми листьями; и из 325 саженцев пурпурно-зеленого сорта 165 имели бело-зеленые и 160 пурпурно-зеленые листья. Из 466 саженцев бело-зеленого сорта 220 имели пурпурно-зеленые и 246 бело-зеленые листья. Эти случаи показывают, насколько сильно пыльца соседнего сорта капусты нейтрализует действие собственной пыльцы растения (Darwin 1876, p. 393).
В настоящее время становится возможным проводить мелкомасштабные исследования генетического картирования немодельных видов, в том числе представляющих прикладной интерес, таких как одомашненные животные и растения и их патогены, для которых картированию QTL помогает большое количество новых маркеров.Генетическое картирование дает надежду на проверку гипотез, таких как тесное сцепление генов, участвующих в гетеростилии у примулы и других видов растений (Li et al. 2007; Labonne et al. 2009) и мимикрии у бабочек (Baxter et al. 2008), примеры задач, интересовавших Дарвина. Картирование генов также важно в современной работе по генетике видообразования, которая, наконец, выявляет гены, участвующие в репродуктивной изоляции между близкородственными видами, и выявляет доказательства гипотезы Добжанки-Мюллера о том, что естественный отбор важен для создания генетических различий между популяциями, которые снижают выживаемость или фертильность гибридов F 1 или F 2 в результате вредоносных эпистатических взаимодействий между аллелями, происходящими из двух популяций ( e. грамм. , Барбаш и др. 2003; Presgraves et al. 2003). Как известно, сам Дарвин загадывал эволюцию репродуктивной изоляции:
грамм. , Барбаш и др. 2003; Presgraves et al. 2003). Как известно, сам Дарвин загадывал эволюцию репродуктивной изоляции:
Важность того факта, что гибриды в большинстве своем бесплодны, я думаю, сильно недооценивалась некоторыми поздними авторами. В теории естественного отбора этот случай особенно важен, поскольку бесплодие гибридов не могло быть для них никакой пользы и, следовательно, не могло быть достигнуто путем постоянного сохранения последовательных прибыльных степеней стерильности (Darwin 1859, p.245).
Однако название Происхождение видов не относится к этой центральной загадке, касающейся видообразования, а скорее к эволюции приспособлений и различий характеров; до появления генетики было бы практически невозможно разработать правильную интерпретацию репродуктивной изоляции.
Еще одна давно обсуждаемая тема, для понимания которой наличие генетических маркеров должно помочь нашему пониманию, — это вопрос о генетической основе инбридинговой депрессии и гетерозиса.Хотя пагубные последствия инбридинга были известны некоторым более ранним биологам, Дарвин был первым, кто тщательно изучил это явление, потому что он понял, что оно дает объяснение существованию сложных приспособлений растений для предотвращения инбридинга. В книге Дарвина Эффекты скрещивания и самооплодотворения в растительном царстве описываются его собственные эксперименты по сравнению потомства, полученного путем самооплодотворения и перекрестного оплодотворения у 57 видов растений, и его краткое изложение основных результатов предвосхищает будущую работу, которая позволила нам измерить инбридинг. (в современном понимании коэффициенты инбридинга):
Некоторые растения, например, Viola tricolor, Digitalis purpurea, Sarothamnus scoparius, Cyclamen persicum и т. Д., которые подвергались естественному перекрестному оплодотворению в течение многих или всех предыдущих поколений, должны сильно пострадать от одного акта самооплодотворения — это самый удивительный факт.
 У наших домашних животных ничего подобного не наблюдалось; но тогда мы должны помнить, что самое близкое возможное скрещивание с такими животными, то есть между братьями и сестрами, не может рассматриваться как почти столь же тесный союз, как союз между пыльцой и семяпочками одного и того же цветка. Пока не известно, увеличивается ли зло от самооплодотворения в течение следующих друг за другом поколений; но мы можем сделать вывод из моих экспериментов, что это увеличение, если оно вообще есть, далеко не быстрое.После того, как растения размножались путем самоопыления в течение нескольких поколений, единичное скрещивание со свежим подвоем восстанавливает их первозданную силу; и у нас есть строго аналогичный результат с нашими домашними животными (Darwin 1876, p. 438).
У наших домашних животных ничего подобного не наблюдалось; но тогда мы должны помнить, что самое близкое возможное скрещивание с такими животными, то есть между братьями и сестрами, не может рассматриваться как почти столь же тесный союз, как союз между пыльцой и семяпочками одного и того же цветка. Пока не известно, увеличивается ли зло от самооплодотворения в течение следующих друг за другом поколений; но мы можем сделать вывод из моих экспериментов, что это увеличение, если оно вообще есть, далеко не быстрое.После того, как растения размножались путем самоопыления в течение нескольких поколений, единичное скрещивание со свежим подвоем восстанавливает их первозданную силу; и у нас есть строго аналогичный результат с нашими домашними животными (Darwin 1876, p. 438).
Как указано Фишером в его План экспериментов (Fisher 1935, глава 3), Дарвин использовал парные контрасты характеристик инбредного и беспородного растения, выращенного в одном горшке, — метод, который широко используется в современная биологическая статистика.Понимание Дарвином полезности этого подхода было испорчено повторным анализом его данных, проведенным его двоюродным братом Фрэнсисом Гальтоном, предположительно более опытным статистиком.
Так же, как с его теорией полового отбора для объяснения диморфизма между мужчинами и женщинами (Darwin 1871), которой в значительной степени пренебрегали до 1970-х годов, индивидуальное селективное преимущество ауткроссинга, возникающее из инбредной депрессии, постулированное Дарвином, было отвергнуто ведущими мыслителями 20-го века. эволюция растений, таких как C.Д. Дарлингтон и Г. Л. Стеббинс в пользу гипотез группового отбора о преимуществах повышенной изменчивости для популяции или вида. Однако в настоящее время роль инбридинговой депрессии в эволюции систем спаривания хорошо известна (Barrett 2002).
Хотя Дарвин не смог дать удовлетворительной интерпретации своих наблюдений, сейчас хорошо известно, что инбридинговая депрессия является генетическим феноменом, а гибридная энергия (гетерозис) широко используется в сельском хозяйстве. Также хорошо известно, что генетическую основу этих явлений трудно установить и что это может помешать попыткам наилучшим образом использовать гетерозис.Нет сомнений в том, что редкие вредные мутации играют важную роль (Charlesworth and Charlesworth 1999): инбридинг, производя гомозигот для таких мутаций, снижает выживаемость и фертильность, потому что большая часть вредных мутаций рецессивны или частично таковы и вызывают только легкий вред при гетерозиготности, как было впервые ясно предложено DF Jones (Jones 1917). Гетерозис также можно объяснить на этом основании, потому что разные инбредные штаммы будут гомозиготными по разным вредным мутациям, а разные популяции вида в природе будут одинаково различаться по некоторой доле их генов, особенно если популяции сильно изолированы (Ingvarsson et al. 2000; Эскобар и др. 2008 г.). Все еще гораздо менее ясно, вносят ли локусы с избыточными аллелями (аллели, демонстрирующие преимущество гетерозигот) какую-либо основную часть инбридинговой депрессии или гетерозиса, хотя интуитивно легко понять, что, если такие локусы являются общими, эти эффекты будут произведены. Определение генетических факторов, участвующих в инбридинговой депрессии или гетерозисе, с помощью методов мелкомасштабного картирования, упомянутых выше, должно помочь ответить на эти вопросы.
Также хорошо известно, что генетическую основу этих явлений трудно установить и что это может помешать попыткам наилучшим образом использовать гетерозис.Нет сомнений в том, что редкие вредные мутации играют важную роль (Charlesworth and Charlesworth 1999): инбридинг, производя гомозигот для таких мутаций, снижает выживаемость и фертильность, потому что большая часть вредных мутаций рецессивны или частично таковы и вызывают только легкий вред при гетерозиготности, как было впервые ясно предложено DF Jones (Jones 1917). Гетерозис также можно объяснить на этом основании, потому что разные инбредные штаммы будут гомозиготными по разным вредным мутациям, а разные популяции вида в природе будут одинаково различаться по некоторой доле их генов, особенно если популяции сильно изолированы (Ingvarsson et al. 2000; Эскобар и др. 2008 г.). Все еще гораздо менее ясно, вносят ли локусы с избыточными аллелями (аллели, демонстрирующие преимущество гетерозигот) какую-либо основную часть инбридинговой депрессии или гетерозиса, хотя интуитивно легко понять, что, если такие локусы являются общими, эти эффекты будут произведены. Определение генетических факторов, участвующих в инбридинговой депрессии или гетерозисе, с помощью методов мелкомасштабного картирования, упомянутых выше, должно помочь ответить на эти вопросы.
ЯРКОЕ БУДУЩЕЕ!
Приведенные здесь примеры показывают ценность продолжающегося взаимодействия между генетикой и изучением эволюции. Генетика, которая не была главной головной болью для ранних сторонников эволюции, проложила путь для моделей эволюции, основанных на известных свойствах наследования, так что ограничения, испытываемые генами и геномами в процессе эволюции, были правильно включены в количественные модели, и новые возможности, неизвестные Дарвину были открыты.
Эволюционная генетика по своей сути междисциплинарна, плодотворно комбинируя модели (часто математические и часто стохастические, учитывая характер генетики) с эмпирическими данными. Эта интеллектуальная традиция, которой сейчас около 100 лет, заслуживает празднования вместе с годовщинами Дарвина. Мы надеемся, что показали, что эволюция занимает более важное место в современных биологических исследованиях, чем когда-либо прежде, и что это продуктивное сотрудничество с генетикой, как можно предвидеть, принесет много новых чистых и прикладных научных достижений в следующие сто лет.Чтобы это произошло, необходимо удовлетворить потребность в достаточно широком образовании. Биологи и врачи должны будут разбираться в генетике и даже некоторых концепциях популяционной генетики, по крайней мере, в достаточной степени, чтобы сотрудничать с людьми, имеющими опыт применения соответствующих количественных методов. Математические идеи должны быть по возможности демистифицированы, чтобы биологи, использующие филогенетические и генетические маркеры или анализ разнообразия, знали, что скрывается за компьютерными программами, которые они используют, понимание, без которого полученные числа могут привести к неправильным выводам.Нам нужно восстановить уважение к полезности статистики во всей биологии и использовать ее для проверки наших идей, как начал это делать Дарвин. То же самое относится и к теоретическому моделированию, направленному на проверяемые гипотезы, для которых идея естественного отбора по-прежнему является прекрасным примером, даже несмотря на то, что она была распространена на гораздо более широкую сферу биологии, чем первоначально предполагал Дарвин, и дала нам много ценных инструментов в области биологии. граница между генетикой и эволюцией. Сам Дарвин интересовался функционированием организмов, а не только их морфологией, взаимоотношениями и историей жизни, и он, несомненно, был бы рад увидеть, куда его идеи привели нас до сих пор и как они по-прежнему занимают центральное место в биологии. .По знаменитым словам Добжанского:
Эта интеллектуальная традиция, которой сейчас около 100 лет, заслуживает празднования вместе с годовщинами Дарвина. Мы надеемся, что показали, что эволюция занимает более важное место в современных биологических исследованиях, чем когда-либо прежде, и что это продуктивное сотрудничество с генетикой, как можно предвидеть, принесет много новых чистых и прикладных научных достижений в следующие сто лет.Чтобы это произошло, необходимо удовлетворить потребность в достаточно широком образовании. Биологи и врачи должны будут разбираться в генетике и даже некоторых концепциях популяционной генетики, по крайней мере, в достаточной степени, чтобы сотрудничать с людьми, имеющими опыт применения соответствующих количественных методов. Математические идеи должны быть по возможности демистифицированы, чтобы биологи, использующие филогенетические и генетические маркеры или анализ разнообразия, знали, что скрывается за компьютерными программами, которые они используют, понимание, без которого полученные числа могут привести к неправильным выводам.Нам нужно восстановить уважение к полезности статистики во всей биологии и использовать ее для проверки наших идей, как начал это делать Дарвин. То же самое относится и к теоретическому моделированию, направленному на проверяемые гипотезы, для которых идея естественного отбора по-прежнему является прекрасным примером, даже несмотря на то, что она была распространена на гораздо более широкую сферу биологии, чем первоначально предполагал Дарвин, и дала нам много ценных инструментов в области биологии. граница между генетикой и эволюцией. Сам Дарвин интересовался функционированием организмов, а не только их морфологией, взаимоотношениями и историей жизни, и он, несомненно, был бы рад увидеть, куда его идеи привели нас до сих пор и как они по-прежнему занимают центральное место в биологии. .По знаменитым словам Добжанского:
Ничто в биологии не имеет смысла, кроме как в свете эволюции (Добжанский, 1973).
Благодарности
Мы благодарим Адама Уилкинса и двух рецензентов за их полезные предложения по улучшению рукописи.
- Авторские права © 2009 Американского генетического общества
Ссылки
- ↵
Акей, Дж. М., 2009 Построение геномных карт положительного отбора у людей: что мы будем делать дальше? Genome Res.19 : 711–722.
- ↵
Альтенбург Э. и Х. Дж. Мюллер, 1920 Генетическая основа усеченного крыла: непостоянный и изменяемый признак у дрозофилы. Генетика 5 : 1–59.
- ↵
Barbash, D. A., D. F. Siino, A. M. Tarone и J. Roote, 2003 Быстро эволюционирующий белок, родственный MYB, вызывает у дрозофилы изоляцию видов. Proc. Natl. Акад. Sci. США 6 : 5302–5307.
- ↵
Барретт, П.Х., П. Дж. Готри, С. Гербер, Д. Кон и С. Смит, 1987 г. Записные книжки Чарльза Дарвина, 1836–1844 гг. Cornell University Press, Итака, Нью-Йорк.
- ↵
Барретт, С. К. Х., 2002 Эволюция полового разнообразия растений. Nat. Преподобный Жене. 3 : 274–284.
- ↵
Бакстер, С., Р. Папа, Н. Чемберлен, С. Хамфрей, М. Джорон и др. , 2008 Конвергентная эволюция генетической основы мимикрии Мюллера у бабочек Heliconius.Генетика 180 : 1567–1577.
- ↵
Бомонт, М. А., и Д. Дж. Болдинг, 2004 Выявление адаптивного генетического расхождения среди популяций с помощью сканирования генома. Мол. Ecol. 13 : 969–980.
- ↵
Bell, G., 2008 Selection , Ed. 2. Издательство Оксфордского университета, Оксфорд.
- ↵
Боулер П. Дж., 1983 Затмение дарвинизма: антидарвиновские теории эволюции в десятилетие около 1900 года. Johns Hopkins University Press, Балтимор.
- ↵
Бойко А., С. Х. Уильямсон, А. Р. Индап, Дж. Д. Дегенхардт, Р. Д. Эрнандес, и др. , 2008 Оценка эволюционного влияния аминокислотных мутаций в геноме человека.
PLOS Genet. 5 : e1000083. - ↵
Bulmer, M. G., 2003 Francis Galton. Johns Hopkins University Press, Балтимор.
- ↵
Bumpus, H.C., 1899 Устранение непригодных на примере представленного воробья, Passer domesticus. Biol. Лекции Woods Hole Marine Biol. Станция 6 : 209–226.
- ↵
Чарльзуорт, Б. и Д. Чарльзуорт, 1999 Генетическая основа инбридинговой депрессии. Genet. Res. 74 : 329–340.
- ↵
Черчилль, Г. А., Д. К. Эйри, Х. Аллай, Дж. М. Ангел, А. Д. Атти и др. , 2004 Коллаборативный крест, ресурс сообщества по генетическому анализу сложных признаков.Nat. Genet. 36 : 1133–1137.
- ↵
Cubas, P., C. Vincent и E. S. Coen, 1999 Эпигенетическая мутация, ответственная за естественную изменчивость цветочной асимметрии. Природа 401 : 157–161.
- ↵
Currat, M., G. Trabuchet, D. Rees, P. Perrin, R.M. Harding et al. , 2002 Молекулярный анализ кластера генов бета-глобина в популяции Ниохоло Манденка показывает недавнее происхождение мутации бета (S) Сенегала.Являюсь. J. Hum. Genet. 70 : 207–223.
- ↵
Дарвин, К. Р., 1859 Происхождение видов. Джон Мюррей, Лондон.
- ↵
Дарвин, К. Р., 1868 Разнообразие животных и растений при одомашнивании. Джон Мюррей, Лондон.
- ↵
Дарвин, К. Р., 1871 Происхождение человека и отбор в отношении пола. Джон Мюррей, Лондон.
- ↵
Дарвин, К.Р., 1876 г. Эффекты перекрестного и самооплодотворения в растительном мире. Джон Мюррей, Лондон.
- ↵
Дарвин, К. Р., 1877 Различные формы цветов на растениях одного вида. Джон Мюррей, Лондон.
- ↵
Ди Чеснола, А. П., 1907 Первое исследование естественного отбора на Helix arbustorum (Helicogena). Биометрика 5 : 387–399.
- ↵
Добжанский, Т., 1955 Обзор некоторых фундаментальных понятий и проблем популяционной генетики. Колд-Спринг-Харбор Symp. Quant. Биол. 20 : 1–15.
- ↵
Добжанский Т., 1973 Ничто в биологии не имеет смысла, кроме как в свете эволюции. Являюсь. Биол. Учить. 35 : 125–129.
- ↵
Dykhuizen, D. E., 1990 Экспериментальные исследования естественного отбора у бактерий. Анну. Rev. Ecol. Evol. Syst. 21 : 373–398.
- ↵
East, E.М., 1910 Менделирующая интерпретация вариации, которая, по-видимому, непрерывна. Являюсь. Nat. 44 : 65–82.
- ↵
Encode Project Consortium, 2007 Идентификация и анализ функциональных элементов в 1% генома человека в рамках пилотного проекта ENCODE. Природа 447 : 799–816.
- ↵
Эскобар, Дж., А. Никот и П. Дэвид, 2008 г. Различные источники вариабельности инбредной депрессии, гетерозиса и аутбредной депрессии в метапопуляции Physa acuta .Генетика 180 : 1593–1608.
- ↵
Farley, J., 1982 Gametes and Spores. Johns Hopkins Press, Балтимор.
- ↵
Felsenstein, J., 2004 Inferring Phylogenies. Sinauer, Сандерленд, Массачусетс.
- ↵
Фишер Р. А., 1930 Генетическая теория естественного отбора. Clarendon Press, Оксфорд.
- ↵
Фишер Р. А., 1935 Планирование экспериментов. Оливер и Бойд, Эдинбург.
- ↵
Flibotte, S., M. Edgley, J. Maydan, J. Taylor, R. Zapf et al. , 2009 Быстрое картирование гибридизации генома для сравнения однонуклеотидного полиморфизма с высоким разрешением у Caenorhabditis elegans .
Генетика 181 : 33–37. - ↵
Флинт Дж. И Т. Ф. К. Маккей, 2009 Генетическая архитектура количественных признаков у мышей, мух и людей. Genome Res. 19 : 723–733.
- ↵
Гайон, Дж., 1998 Борьба дарвинизма за выживание: наследственность и гипотеза естественного отбора. Cambridge University Press, Кембридж, Великобритания.
- ↵
Гиллиспи, К. К., 1960 На грани объективности. Princeton University Press, Принстон, Нью-Джерси.
- ↵
Haag-Liautard, C., M. Dorris, X. Maside, S. Macaskill, D. L. Halligan et al. , 2007 Прямая оценка частоты нуклеотидных и геномных вредных мутаций у Drosophila .Природа 445 : 82–85.
- ↵
Харви, П. Х. и М. Пагель, 1991 Сравнительный метод в эволюционной биологии. Oxford University Press, Оксфорд.
- ↵
Heijmans, B. T., E. W. Tobi, A. D. Stein, H. Putter, G. J. Blauw et al. , 2008 Устойчивые эпигенетические различия, связанные с пренатальным воздействием голода у людей. Proc. Natl. Акад. Sci. США 6 : 17046–17049.
- ↵
Ховард, Дж., 2009 Почему Дарвин не открыл законы Менделя? J. Biol. 8 : 15.
- ↵
Хадсон Р. Р., 1990 Генеалогии и процесс слияния, стр. 1–44, в Oxford Surveys in Evolutionary Biology, Vol. 7, под редакцией Дугласа Футуймы и Яниса Антоновичса. Издательство Оксфордского университета, Нью-Йорк.
- ↵
Ингварссон, П. К., М. К. Уитлок и Т. Хэтфилд, 2000 Локальная дрейфовая нагрузка и гетерозис взаимосвязанных популяций.Наследственность 84 : 452–457.
- ↵
Дженкин Ф., 1867 Происхождение видов. North British Rev. 46 : 277–318.
- ↵
Johannsen, W.
, 1909 Elemente der exakten Erblichkeitslehre. Фишер, Йена, Германия. - ↵
Джонс, Д. Ф., 1917 г. Преобладание связанных факторов как средство учета гетерозиса. Генетика 2 : 466–479.
- ↵
Кимура, М., 1983 Нейтральная теория молекулярной эволюции. Cambridge University Press, Кембридж, Великобритания.
- ↵
Кингсолвер, Дж. Г., Х. Э. Хокстра, Дж. М. Хокстра, Д. Берриган, С. Н. Виньери и др. , 2001 Сила фенотипического отбора в естественных популяциях. Являюсь. Nat. 157 : 245–261.
- ↵
Labonne, J., A. Goultiaeva и J. Shore, 2009 Картирование с высоким разрешением S-локуса в Turnera привело к открытию трех генов, тесно связанных с S-аллелями.Мол. Genet. Геномика 281 : 673–685.
Ледерберг, Дж. И Э. М. Ледерберг, 1952 Посев на реплики и непрямой отбор бактериальных мутантов. J. Bacteriol. 63 : 399–406.
- ↵
Leimu, R., and M. Fischer, 2008 Метаанализ локальной адаптации растений. PLoS ONE 3 : e4010.
- ↵
Левонтин Р. К., 1974 Генетическая основа эволюционных изменений. Columbia University Press, Нью-Йорк.
- ↵
Li, J., M. Webster, M. Furuya and P. Gilmartin, 2007 Идентификация и характеристика аллелей булавок и головок двух генов, ко-сегрегированных с локусом Primula S . Плант Дж. 51 : 18–31.
- ↵
Мэй, Г., Ф. Шоу, Х. Бадран и Х. Векеманс, 1999 Сигнатура балансирующего отбора: эволюция гена совместимости с грибами. Proc. Natl. Акад. Sci.США 96 : 9172–9177.
- ↵
Мейнард Смит, Дж. И Дж. Хей, 1974 Эффект автостопом благоприятного гена. Genet. Res. 23 : 23–35.
- ↵
Mendel, G.
, 1866 Versuche über Pflanzenhybriden. Вер. Naturforsch. Вер. Брюнн 4 : 3–47. - ↵
Мюллер, Х. Дж., 1932 Дальнейшие исследования природы и причин генных мутаций. Материалы 6-го Международного конгресса по генетике, Vol.1, 213–255.
- ↵
Майерс, С., Л. Боттоло, К. Фриман, Г. Маквин и П. Доннелли, 2005 г. Мелкомасштабная карта скоростей рекомбинации и горячих точек в геноме человека. Наука 310 : 312–324.
- ↵
Намекава, С. Х., Дж. Л. Вандеберг, Дж. Р. МакКерри и Дж. Т. Ли, 2007 Подавление половых хромосом в зародышевой линии сумчатых самцов. Proc. Natl. Акад. Sci. США 104 : 9730–9735.
- ↵
Нэш, Д., С. Наир, М. Майксай, П. Н. Ньютон, Ж.-П. Guthmann et al. , 2005 Сила отбора и автостоп вокруг двух генов антималярийной устойчивости. Proc. Биол. Sci. 272 : 1153–1161.
- ↵
Nilsson-Ehle, H., 1909 Kreuzungsuntersuchungen an Hafer und Weizen. Lunds Univ. Арскрифт НС 5 : 1–12.
- ↵
Nordborg, M., 1997 Структурированные процессы слияния в различных временных масштабах. Генетика 146 : 1501–1514.
- ↵
Орндафф Р., 1969 Репродуктивная биология в отношении систематики. Таксон 18 : 121–133.
- ↵
Прегрейвс, Д. К., Л. Балагопалан, С. М. Абмайр и Х. А. Орр, 2003 г. Адаптивная эволюция приводит к дивергенции гибридного гена неуязвимости между двумя видами дрозофилы. Природа 423 : 715–719.
- ↵
Provine, W. B., 1971 Истоки теоретической популяционной генетики. Чикагский университет Press, Чикаго.
- ↵
Raymond, C. K., A. Kas, M. Paddock, R. Qiu, Y. Zhou et al. , 2005 г. Древние гаплотипы региона HLA класса II. Genome Res. 15 : 1250–1257.
- ↵
Ричардс, Э. Дж., 2006 Унаследованная эпигенетическая изменчивость: возвращаясь к мягкому наследованию. Nat. Преподобный Жене. 7 : 395–401.
- ↵
Ричман, А. Д., М. К. Уенояма и Дж.Р. Кон, 1996 г. Аллельное разнообразие и генеалогия в локусе самонесовместимости у Solanaceae. Наука 273 : 1212–1216.
- ↵
Ritland, K., 1990 Серия компьютерных программ FORTRAN для оценки систем скрещивания растений. J. Hered. 81 : 235–237.
- ↵
Сабети, П. К., Д. Э. Райх, Дж. М. Хиггинс, Х. З. Левин, Д. Дж. Рихтер и др. , 2002 Обнаружение недавнего положительного отбора в геноме человека по гаплотипической структуре.Природа 419 : 832–837.
- ↵
Селла, Г., Д. А. Петров, М. Пржеворски и П. Андольфатто, 2009 Всесторонний естественный отбор в геноме дрозофилы? PLOS Genet. 6 : e1000495.
- ↵
Sidorenko, L., and V. Chandler, 2008 РНК-зависимая РНК-полимераза необходима для опосредованного энхансером подавления транскрипции, связанного с парамутацией в гене p1 кукурузы. Генетика 180 : 1983–1993.
- ↵
Слаткин, М., 2008 Неравновесие по сцеплению: понимание эволюционного прошлого и картирование медицинского будущего. Nat. Преподобный Жене. 9 : 477.
- ↵
Van Os, H., P. Stam, R. G. Visser и H. J. Van Eck, 2005 RECORD: новый метод упорядочивания локусов на карте генетического сцепления. Теор. Прил. Genet. 112 : 30–40.
- ↵
Векеманс, X., и М. Слаткин, 1994 Генная и аллельная генеалогия в локусе гаметофитной самонесовместимости.Генетика 137 : 1157–1165.
- ↵
Уолш Б., 2008 Использование молекулярных маркеров для обнаружения генов одомашнивания, улучшения и адаптации. Euphytica 161 : 1–17.
- ↵
Велдон, W. F. R., 1895 Попытка измерить уровень смертности из-за избирательного уничтожения Carcinus moenas по определенному измерению. Proc. R. Soc. Ser B 58 : 360–379.
- ↵
Weldon, W.F. R., 1901 Первое исследование естественного отбора на Clausilia ламината (Монтегю). Биометрика 1 : 109–124.
- ↵
Williamson, S., M. J. Hubisz, A. G. Clark, B. Payseur, C. Bustamante et al. , 2007 Локализация недавней адаптивной эволюции в геноме человека. PLoS Genet, 3 : 901–915.
- ↵
Райт, С., 1920 Относительная важность наследственности и окружающей среды в определении пегого образа морских свинок.Proc. Natl. Акад. Sci. США 6 : 320–333.
- ↵
Райт, С., 1968 Эволюция и генетика популяций : Генетические и биометрические основы , Vol. 1. Издательство Чикагского университета, Чикаго.
- ↵
Райт С., Добжанские Т., 1946 Генетика природных популяций XII. Экспериментальное воспроизведение некоторых изменений, вызванных естественным отбором, в определенных популяциях Drosophila pseudoobscura .Генетика 31 : 125–156.
 PLOS Genet. 5 : e1000083.
PLOS Genet. 5 : e1000083. 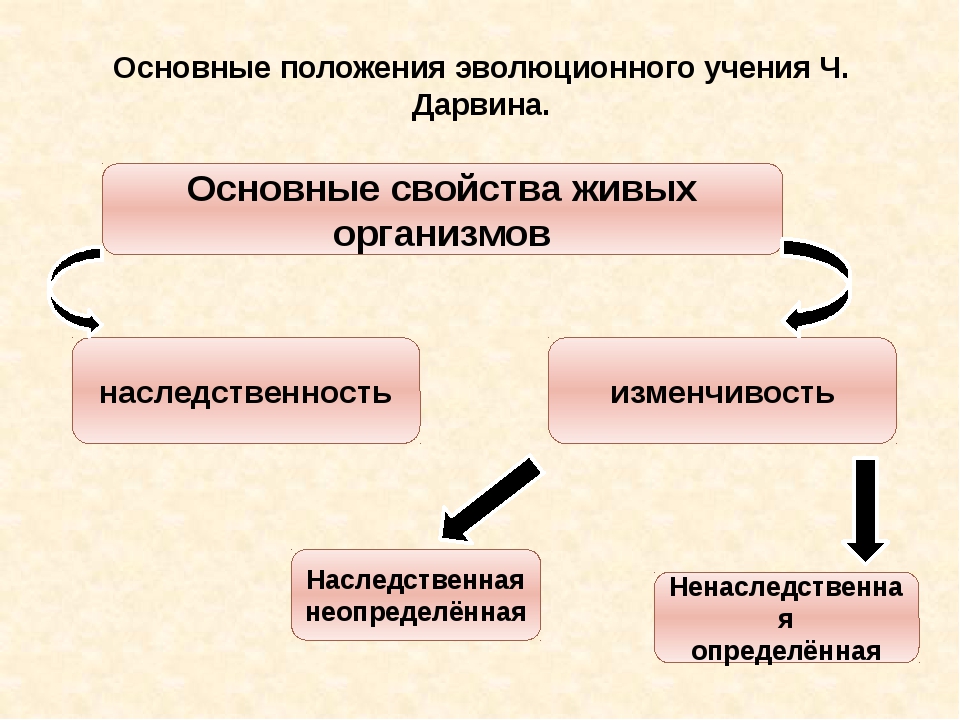
 Генетика 181 : 33–37.
Генетика 181 : 33–37.  , 1909 Elemente der exakten Erblichkeitslehre. Фишер, Йена, Германия.
, 1909 Elemente der exakten Erblichkeitslehre. Фишер, Йена, Германия.  , 1866 Versuche über Pflanzenhybriden. Вер. Naturforsch. Вер. Брюнн 4 : 3–47.
, 1866 Versuche über Pflanzenhybriden. Вер. Naturforsch. Вер. Брюнн 4 : 3–47. Дарвин и теория эволюции
Результаты обучения
- Опишите, как работа Дарвина превратилась в теорию эволюции
Естественный отбор может иметь место только при наличии вариаций или различий между особями в популяции. Важно отметить, что эти различия должны иметь некую генетическую основу; в противном случае выбор не приведет к изменениям в следующем поколении.Это очень важно, потому что различия между людьми могут быть вызваны негенетическими причинами, такими как рост человека из-за лучшего питания, а не из-за разных генов.
Генетическое разнообразие в популяции происходит за счет двух основных механизмов: мутации и полового размножения. Мутация, изменение ДНК, является основным источником новых аллелей или новых генетических вариаций в любой популяции. Генетические изменения, вызванные мутацией, могут иметь один из трех последствий для фенотипа. Мутация влияет на фенотип организма таким образом, что снижает его приспособленность — меньшую вероятность выживания или меньшее количество потомков.Мутация может вызвать фенотип, положительно влияющий на приспособленность. И многие мутации также не будут влиять на приспособленность фенотипа; они называются нейтральными мутациями. Мутации также могут иметь целый ряд размеров воздействия на приспособленность организма, который выражает их в своем фенотипе, от небольшого эффекта до большого эффекта. Половое размножение также ведет к генетическому разнообразию: когда два родителя воспроизводятся, уникальные комбинации аллелей собираются, чтобы произвести уникальные генотипы и, следовательно, фенотипы в каждом из потомков.
Унаследованная черта, которая помогает выживанию и воспроизводству организма в его нынешней среде, называется адаптацией . Ученые описывают группы организмов, которые адаптируются к своей среде, когда с течением времени происходит изменение диапазона генетических вариаций, которое увеличивает или поддерживает «приспособленность» популяции к окружающей среде. Перепончатые лапы утконоса — приспособление для плавания. Густой мех снежного барса — приспособление для жизни на морозе.Быстрая скорость гепардов — приспособление для ловли добычи.
Является ли признак благоприятным, зависит от условий окружающей среды в данный момент. Не всегда выбираются одни и те же черты, потому что условия окружающей среды могут измениться. Например, рассмотрим вид растений, которые росли во влажном климате и не нуждались в экономии воды. Были выбраны крупные листья, потому что они позволяли растению получать больше энергии от солнца. Для содержания больших листьев требуется больше воды, чем для ухода за маленькими листьями, а влажная среда обеспечивает благоприятные условия для поддержки крупных листьев.Спустя тысячи лет климат изменился, и в этом районе больше не было избытка воды. Направление естественного отбора изменилось, и в результате были выбраны растения с мелкими листьями, потому что эти популяции были способны экономить воду, чтобы выжить в новых условиях окружающей среды.
Эволюция видов привела к огромному разнообразию форм и функций. Иногда эволюция порождает группы организмов, которые сильно отличаются друг от друга. Когда два вида эволюционируют в разных направлениях из общей точки, это называется дивергентной эволюцией.Такую дивергентную эволюцию можно увидеть в формах репродуктивных органов цветковых растений, которые имеют одну и ту же базовую анатомию; однако они могут выглядеть совершенно по-разному в результате отбора в разных физических средах и адаптации к разным типам опылителей (рис. 1).
Рис. 1. Цветковые растения произошли от общего предка. Обратите внимание, что (а) плотная пылающая звезда ( Liatrus spicata ) и (b) пурпурный эхинацея ( Echinacea purpurea ) различаются по внешнему виду, но оба имеют схожую базовую морфологию.(кредит А: модификация работы Дрю Эйвери; кредит б: модификация работы Кори Занкера)
В других случаях подобные фенотипы развиваются независимо у отдаленно родственных видов. Например, полет эволюционировал как у летучих мышей, так и у насекомых, и у них обоих есть структуры, которые мы называем крыльями, которые являются адаптацией к полету. Однако крылья летучих мышей и насекомых произошли от очень разных оригинальных структур. Это явление называется конвергентной эволюцией , при которой сходные черты эволюционируют независимо у видов, не имеющих недавнего общего предка.Эти два вида выполняли одну и ту же функцию — летали, но выполняли это отдельно друг от друга.
Эти физические изменения происходят в течение огромных промежутков времени и помогают объяснить, как происходит эволюция. Естественный отбор действует на отдельные организмы, которые, в свою очередь, могут формировать целый вид. Хотя естественный отбор может воздействовать на отдельное поколение в отдельности, эволюция генотипа целого вида может занять тысячи или даже миллионы лет. Именно за эти большие промежутки времени жизнь на Земле изменилась и продолжает меняться.
Вкратце: Дарвин и теория эволюции
Естественный отбор, движущая сила эволюции, может работать только в том случае, если между организмами существуют различия. В конечном итоге вариации возникают из-за генетических мутаций. Разнообразие поощряется половым размножением. По мере изменения среды выборочное давление смещается и способствует разным адаптациям. Таким образом, через тысячи или миллионы лет, виды эволюционируют.
Внесите свой вклад!
У вас была идея улучшить этот контент? Нам очень понравится ваш вклад.
Улучшить эту страницуПодробнее
Мендель и Дарвин: разгадывая непреходящую загадку
Иоганн Мендель родился в 1822 году в Хинчице (Хайзендорф) Сноска 1 в Австрийской Силезии, в настоящее время в Чехии (Чешская Республика). Принеся большую жертву своей семье, он посещал школы-интернаты до 21 года. Он был успешным в учебе и в 1843 году был принят в августинский орден монастыря Св. Фомы в Брно (Брюнн), где принял имя Грегор.
С тех пор он оказался в необычайно интеллектуальном окружении. Он был монахом, а не монахом в том смысле, что монахи-августинцы его монастыря служили обществу, открыто взаимодействуя с местным населением, получая высшее образование по светским предметам, участвуя в научных обществах, выполняя общественные роли, проводя исследования и писательскую деятельность. и обучение своим академическим специальностям в школах. Последствия попытки Менделя служить священником лучше всего описываются словами его настоятеля Сирилла Наппа: «он очень усердно изучает науки; но … он гораздо менее приспособлен для работы приходским священником по той причине, что он охвачен непреодолимой робостью … [поэтому] я счел необходимым освободить его от службы приходским священником »(Iltis, 1966, p. .58). Робость Менделя неоднократно всплывала на поверхность, оказывая положительное и отрицательное влияние на его научные занятия.
Мендель как учитель естествознания
Случайным следствием робости Менделя стало то, что она побудила Наппа поручить ему преподавать естественные науки в Зноймо (Знайм), к юго-западу от Брно. Это задание позволило ему заниматься наукой вместе со студентами, не обремененным обязанностями приходского священника. Он был превосходным и популярным учителем, хотя и не сертифицированным. В ноябре 1849 года австрийское правительство издало указ, согласно которому все учителя должны пройти аттестацию через университетские экзамены (Klein and Klein 2013).Венский университет будет проводить экзамен Менделя, состоящий из письменной части вне офиса (называемой домашним заданием), а также письменных и устных сессий на месте.
Летом 1850 года Мендель получил домашнее задание, в котором мы встречаем его первые слова, намекающие на эволюцию:
С течением времени, когда Земля достигла способности, необходимой для образования и сохранения органической жизни, растения и впервые появились животные низшего вида.Период органического образования нередко прерывался катастрофами, угрожавшими жизни организмов и отчасти приводившими к их упадку. … Растительный и животный мир развивался все более и более богато; его старые формы частично исчезли, чтобы уступить место новым и более совершенным.
(Орёл и др., 1983, с. 237 * , сноска 2 ).
Влияние катастрофизма старой земли на мысли Менделя очевидно, вероятно, из книги Карла Германа Конрада Бурмейстера (1843) Die Geschichte der Schöpfung ( История сотворения мира, ), на которую Мендель цитировал.Учитывая принятие Менделем катастрофизма старой Земли на этом раннем этапе своей карьеры, неясно, рассматривал ли он происхождение от выживших видов, в отличие от создания de novo, как механизм появления новых видов. Примечательно, что Орел и др. (1983) и Кляйн и Кляйн (2013) указали, что Мендель использовал слова «zum Theile», переведенные как «частично», имея в виду угасание жизни, оставляя открытой интерпретацию, что новые виды возникли из тех, которые пережили катастрофы. В любом случае он собирался войти в одну из самых активных и спорных додарвиновских эволюционных сред Европы, где эволюция путем нисхождения на протяжении эонов геологического времени была основной темой.Нет никаких свидетельств того, что он когда-либо вернулся к катастрофе.
Экзаменаторы Менделя по физике Андреас фон Баумгартнер и Кристиан Доплер высоко оценили его эссе по физике. Эксперт по геологии и естественной истории Рудольф Кнер определил, что отчет и источники Менделя неадекватны и устарели. Тем не менее, Мендель был приглашен на письменные и устные занятия в Венском университете. Он преуспел в одних частях и провалил другие, и поэтому не сдал сертификационный экзамен.Тем не менее, его экзаменаторы согласились, что он хорошо подготовлен к учебе в университете, что позволило ему поступить в Венский университет. Согласно Орлу (1996): «Если бы он сдал экзамен учителя, он остался бы в Зноймо гимназии , и несколько поколений учеников средней школы получили бы отличного учителя. С другой стороны, наука почти наверняка потеряла бы одного из своих ведущих первооткрывателей »(стр. 66).
В Венском университете
В ноябре 1851 года Мендель поступил в Венский университет в качестве «экстраординарного» студента, что означало, что он не получал ученую степень, не проверял свои классы и не проходил формальную оценку (Klein and Klein 2013 ).В течение 22 месяцев (с ноября 1851 г. по август 1853 г.) он записался на курсы физики, химии, математики, зоологии, ботаники и палеонтологии, некоторые из которых были посвящены эволюции. Его исследования едва ли можно было лучше приурочить к всплеску эволюционного энтузиазма, который сильно затронул интеллектуальные и религиозные сообщества Вены. Один из профессоров ботаники Менделя, Франц Унгер, был ключевым игроком в этом всплеске. К 1851 году Унгер энергично продвигал то, что Глибофф (1999) назвал «теорией Унгера о всеобщем общем происхождении» (стр.223), рассматривая все новые виды как происходящие от предыдущих видов, и, по словам Унгера, «никоим образом не одностороннее линейное развитие, но излучение, расширяющееся во все стороны» (Unger 1853, с. 107).
С мая по октябрь 1851 года Унгер популяризировал эволюцию посредством серии из семнадцати статей под названием Botanische Briefe ( Botanical Letters ) в Wiener Zeitung ( Vienna News ), которые позже были собраны в книгу ( Унгер 1852 г.).Унгер популяризировал то, как летопись окаменелостей показала эволюцию растений и животных на протяжении эонов геологического времени. 17-я и последняя часть была опубликована 18 октября 1851 года, всего за девять дней до прибытия Менделя в Вену (Gliboff 1998).
Унгер придерживался точки зрения, согласно которой новые виды возникли из предыдущих в отрывке, который примечателен тем, что предсказывает тему генетической рекомбинации в эволюции, к которой Мендель обратится в своем исследовании (как отмечено Olby 1985; Orel 1996; Klein and Klein 2013) : «Кто может отрицать, что новые комбинации элементов возникают из этой перестановки растительности, когда-либо сводимой к определенному закону, — комбинации, которые освобождаются от предшествующих характеристик вида и появляются как новые виды?» (Унгер 1853, стр.92). В том же году Унгер (1851) также опубликовал важную популярную книгу об эволюции Die Urwelt in ihren verschiedenen Bildungsperioden ( Первобытный мир в его различные переходные периоды ), которая была хорошо принята в Вене (Gliboff 1998).
Публикации Унгера вызвали гнев выдающегося римско-католического священника Себастьяна Бруннера, редактора-основателя Wiener Kirchenzeitung ( Венские церковные новости ), который опубликовал несколько редакционных статей, высмеивающих его.Первый появился 25 октября 1851 года, всего за два дня до приезда Менделя. Бруннер утверждал (в отрывках, переведенных Олби 1985 г.), что «язычество преподается в университетах во всех областях науки» (стр. 201), что Унгер был «человеком, который открыто отрицал творение и Создателя» (стр. 202). ), и что «профессора так называемых католических университетов годами читают лекции по действительно чудовищным теориям» (стр. 203). Мендель, несомненно, знал об этом публичном споре, но в 1852 году записался в классы Унгера, что, возможно, поставило его в шаткое положение как члена духовенства.
Возвращение в Брно и церковное осуждение
В августе 1853 года Мендель вернулся в Брно, все еще не получивший диплом учителя естественных наук, но хорошо подготовленный в области научной теории и методологии. Он сразу же столкнулся с атмосферой официального осуждения светских занятий, мало чем отличавшегося от церковного осуждения науки, свидетелем которого он стал в Вене. Епископу Брно, Антону Эрнсту Шаффготчу, было поручено исследовать августинские монастыри в Праге, Кракове и Брно на предмет секуляризма, начиная с 1853 года, когда Мендель еще находился в Вене (Orel 1996; Klein and Klein 2013).В 1854 году, после возвращения Менделя, Шаффготч провел на месте расследование. В своем отчете, рекомендуя распустить монастырь, он написал (в переводе Кляйна и Кляйна, 2013 г.): «Одним словом, в доме, соблюдающем Правило Св. Августина, царит светский дух, который немногочисленны прихожанами августинцев. не в состоянии прикрыть », и особенно Менделя,« он изучает профанные науки в светском учреждении в Вене за счет монастыря, чтобы стать профессором указанных наук в государственном учреждении »(стр.295).
Тем временем нападки Бруннера достигли своего финала в начале 1856 года в редакционной статье под названием «Священник Исиды и филистимлянин», специально адресованной Унгеру и его коллегам: «современных первосвященников Исиды, то есть держателей стульев в различных ветви естественной истории редко или никогда не входят в церковь — согласно их заявлениям, природа — их храм »*. Затем Бруннер высмеивает венских ботаников: «Они делают все, что в их силах, чтобы превратить себя в растения ботанических знаний, которые можно почувствовать издалека, — и добровольно попадают на вечно вонючее навозное ложе пантеистического мировоззрения, которое, тем не менее, порождает новое мировоззрение. определенное богатство цветов »*.
И в Вене, и в Брно среда, противопоставляющая церковь эволюции, стала чрезвычайно едкой. Будучи священнослужителем, терпящим явные упреки епископа Брно и обучавшимся у такого печально известного эволюциониста, как Унгер, Мендель, несомненно, страдал от обременительных конфликтов. К сожалению, ни одно из его сохранившихся произведений не раскрывает ничего о том, как он лично относился к этой дилемме.
Спустя всего несколько недель после того, как спор Унгера с Бруннером достиг апогея, 5 мая 1856 года Мендель поехал в Венский университет, чтобы сдать второй экзамен на получение сертификата учителя.Один из его товарищей рассказал о том, что произошло: «К счастью, он получил простые вопросы, но после первого он заболел, так что писать было невозможно. В любом случае кажется, что он нервничает. … Опасаясь дальнейших подобных нападений, он вернулся домой, ничего не добившись »(Kříženecký 1963, с. 308 *). Можно только догадываться, насколько противоречивые привязанности Менделя к церкви и к своей научной честности способствовали этому нервному срыву.Немедленный результат был ошеломляющим. Мендель был прикован к постели, и известие, отправленное его семье, побудило его больного отца и зятя поехать в Брно, чтобы позаботиться о нем (Iltis 1966, Orel 1996, Klein and Klein 2013). Долгосрочный результат также деморализовал. Несмотря на свои амбиции, он никогда не получит диплом учителя естественных наук.
Классическая статья Менделя и первоначальное влияние Дарвина (1856–66)
Роль гибридизации в эволюции новых видов была одной из наиболее широко исследованных тем при жизни Менделя.За два года до поступления Менделя в Венский университет Карл Фридрих фон Гертнер (1849) опубликовал свой большой труд, Versuche und Beobachtungen über die Bastarderzeugung im Pflanzenreiche ( Эксперименты и наблюдения по гибридному производству в растительном царстве ), который получил Мендель. , наверное, когда учился в университете. Его личный экземпляр содержит его аннотации (Olby 1985), и он часто ссылается на них в своей классической статье. Дарвин (1859) также считал эту тему важной, посвятив целую главу гибридизму в Происхождение видов .
Мендель начал свои эксперименты по гибридизации с горохом посевным ( Pisum sativum ) примерно одновременно со своим вторым экзаменом на получение сертификата учителя, весной 1856 г. Он хорошо спланировал эти эксперименты, проведя два предыдущих лета (1854 г.) и 1855), выращивая и исследуя сорта, которые он выбрал бы в качестве родителей, чтобы убедиться, что они являются истинными. Его классическая статья «Versuche über Pflanzen Hybriden» («Эксперименты с гибридами растений») будет опубликована десять лет спустя (Mendel 1866).
При поддержке Чарльза Лайеля Дарвин начал писать крупный многотомный труд по естественному отбору той же весной, 14 мая 1856 г. (Darwin 1856, Darwin 1887, p. 67). Два года спустя он начал реорганизацию и переписывание материала из этой рукописи, чтобы создать несколько книг, в том числе две, которые больше всего повлияли на Менделя: Происхождение видов (1859) и Разнообразие животных и растений при одомашнивании (1868).
Происхождение видов впервые появилось в ноябре 1859 г. (Дарвин 1859 г.), а первый немецкий перевод — в 1860 г. (Дарвин 1860 г.).Мендель владел вторым изданием немецкого перевода, опубликованным в 1863 году (Darwin 1863; Orel 1971b; Fairbanks and Rytting 2001). Поскольку в том же году Мендель завершил свои эксперименты по гибридизации гороха, он, вероятно, прочитал и аннотировал свою копию Происхождение видов после сбора всех своих данных, но перед подготовкой своих лекций 1865 года по ним. Согласно этой временной шкале, Происхождение видов не могло ни мотивировать, ни направлять его эксперименты, но могло повлиять на его интерпретации.Статья Фишера (1936) «Была ли заново открыта работа Менделя?» больше всего помнят за разжигание полемики Менделя-Фишера, хотя это значительно более обширная статья. Среди основных моментов Фишера — его утверждение, что Дарвин оказал влияние на Менделя. Ссылаясь на параграф, в котором Мендель отверг представление о том, что «условия жизни» вызывают новые наследственные вариации, Фишер заметил: «Отражение мысли Дарвина безошибочно…». (стр.134).
Фэрбенкс и Риттинг (2001) опубликовали аннотации Менделя в его экземпляре Происхождение видов .Они обнаружили, что термин «Lebensbedingungen» (или его альтернативы «Lebens-Bedingungen» и «Lebens Bedingungen») встречается в трех отрывках, аннотированных Менделем, каждый из которых переведен с «условий жизни» Дарвина, которые охарактеризовали Фэрбенкс и Эбботт (2016). как «типично дарвиновская фраза, встречающаяся 107 раз в Происхождение видов » (стр. 404). Мендель дважды использовал «Lebensbedingungen» в своей классической статье, причем оба раза в абзаце, который, как подчеркнул Фишер, отражает мысль Дарвина.Одно это слово дает доказательства, подтверждающие утверждение Фишера, но это только начало таких доказательств.
Исходя из предпосылки, что Мендель читал Происхождение видов во время написания своей рукописи, Эбботт и Фэрбенкс (2016) опубликовали новый дарвинизованный английский перевод классической работы Менделя. Они сравнили немецкий текст Менделя с его немецким переводом Происхождение видов и определили в обоих словах и словосочетаниях один и тот же или один и тот же корень.Затем они использовали оригинальный английский Дарвина в переводе, насколько это было возможно, чтобы придать особый тон девятнадцатого века. Слова из отрывков, помеченных Менделем в его немецком переводе, были заметно чаще в конце его статьи.
По крайней мере две, не исключающие друг друга, возможности объясняют это наблюдение. Во-первых, Мендель мог прочитать Происхождение видов после того, как он подготовил большую часть своей рукописи, и поэтому эти отрывки повлияли на ее заключительную часть.Во-вторых, последние два раздела статьи Менделя посвящены теоретической интерпретации и больше подходят для обсуждения тем, затронутых Дарвином.
Фэрбенкс и Эбботт (2016) обнаружили, что значительное количество этих слов впервые появляется в статье Менделя в том же абзаце, указанном Фишером (1936). В этом абзаце Мендель написал: «Часто высказывается мнение, что стабильность вида была в значительной степени нарушена или полностью нарушена в результате выращивания» (Abbott and Fairbanks, 2016, p.418). Вероятно, это завуалированная ссылка на Дарвина в ответ на первый отрывок, отмеченный Менделем в его копии Происхождение видов (рис. 1). На оригинальном английском языке Дарвина в нем говорится: «Кажется довольно очевидным, что органические существа должны подвергаться в течение нескольких поколений новым условиям жизни, чтобы вызвать какие-либо заметные изменения; и что, когда организация однажды начала меняться, она, как правило, продолжает меняться на протяжении многих поколений »(Darwin 1861, p. 7). «Новые условия жизни» в этом контексте — это условия приручения, которые, по мнению Дарвина, вызывают большую степень наследственной изменчивости по сравнению с теми, которые существуют в природе.
Рис. 1: Аннотация Менделя на первой странице первой главы его немецкого перевода Происхождение видов .В оригинальном английском языке Дарвина вся фраза гласит: «Кажется довольно очевидным, что органические существа должны подвергаться в течение нескольких поколений новым условиям жизни, чтобы вызвать какие-либо заметные изменения; и что, когда организация однажды начала меняться, она, как правило, продолжает меняться на протяжении многих поколений ». Обратите внимание на термин «Lebens-Bedingungen», соответствующий «условиям жизни» Дарвина.
Мендель разъяснил в своей классической статье, как его открытия не соответствовали этой точке зрения: «Типичные вариации должны появиться, если условия жизни для вида изменяются, и он имеет способность адаптироваться к новым условиям. … Если бы изменение условий растительности было единственной причиной изменчивости , то можно было бы ожидать, что те одомашненные растения, которые веками культивировались почти в одних и тех же условиях, приобрели бы стабильность .Как хорошо известно, дело обстоит не так… »(курсив в оригинале, Abbott and Fairbanks, 2016, стр. 418–419).
Здесь Мендель использовал несколько иной подход, чем Дарвин, согласовывая свою точку зрения с одной, предложенной Унгером 14 лет назад, — что изменяющиеся условия жизни не являются причиной новых наследственных вариаций: «Таким образом, попытка проследить разнообразие видов. влияние внешних воздействий, таких как природа почвы, несомненно, упускает из виду истинную причину »(Unger 1853, p.93). Представление о том, что изменение условий жизни может формировать наследственные вариации, было предметом споров задолго до Дарвина, начиная, по крайней мере, с восемнадцатого века (Roberts 1929; Olby 1967). Мендель ясно дал понять, что его наблюдения имеют отношение к этой дискуссии.
Не только Мендель в Брно обратился к этой теме. Общество естествознания в Брно собиралось ежемесячно, а презентацию в январе 1865 года сделал Александр Маковский, ботаник и палеонтолог, который был другом Менделя, соавтором исследования и автором книги, автором которой был Мендель.Мендель и Маковски были учителями в Realschule в Брно, где проводились ежемесячные собрания Общества естественных наук (Орел, 1996). Презентация Маковски «Теория Убера Дарвина» («Теория Дарвина об органическом сотворении») была мощным подтверждением Дарвина Происхождение видов (Маковски 1866; Фэрбенкс и Эбботт 2019). Презентации Менделя следовали сразу же за презентациями Маковского, занимая следующие две встречи, в феврале и марте.В своей речи Маковски обратился к «условиям жизни» Дарвина, но поддержал точку зрения Менделя, преуменьшая роль окружающей среды как причины новых наследственных вариаций.
Исходя из этого, что мы знаем о взглядах Менделя на Дарвина в частности и на эволюцию в целом на момент написания его классической статьи? Во введении к его статье 1866 года содержится часто цитируемая ссылка на эволюцию: «Определенно требуется определенное мужество, чтобы предпринять такую обширную работу; тем не менее, это кажется единственным подходящим средством для окончательного решения вопроса об эволюционной истории органических форм, важность которого нельзя недооценивать »(Abbott and Fairbanks, 2016, p.407). Позже в статье он писал: «Это обстоятельство особенно важно для эволюционной истории растений, потому что постоянные гибриды приобретают статус нового вида . Истинность этого факта была подтверждена наиболее выдающимися наблюдателями и не подлежит сомнению »(курсив в оригинале, Abbott and Fairbanks 2016, p. 420).
В обоих случаях Мендель использовал составной термин «Entwicklungsgeschichte» (в одном случае переносится через дефис), который можно перевести как «эволюционная история» или «история развития».«Эволюция» (пишется одинаково на немецком и английском языках) не было таким распространенным словом ни в одном из языков в то время для обозначения того, что стало биологической эволюцией двадцатого века. На немецком языке «Entwicklung» и его производные объединили развитие и эволюцию в единое целое. Часто утверждается, что Дарвин никогда не использовал слово «эволюция» в Происхождение видов , что позволяет избежать того факта, что он использовал форму причастия прошедшего времени «эволюционировала» как буквально последнее слово книги: «… бесконечные формы, самые красивые и самые прекрасные. чудесные были и развиваются »(Darwin 1859, p.490). Генрих Георг Бронн, переводчик немецкой версии, принадлежащей Менделю, перевел дарвиновское «эволюционировало» как «entwickelt», от того же корня, что и «Entwicklung», слово «Мендель» больше, чем какое-либо другое, связывало бы с биологической эволюцией.
Загадка Менделя как эволюциониста или креациониста
Мендель был ученым и священнослужителем в то время, когда конфликт между эволюцией и религией достиг апогея, что вызывает вопросы относительно его взглядов. Большинство историков, изучавших историю Менделя, приходят к выводу, что он принимал эволюцию и рассматривал свои эксперименты и теорию как совместимые с экспериментами Дарвина (см. Iltis, 1966, Olby, 1985; Orel, 1996; Fairbanks and Rytting, 2001, и Klein and Klein, 2013 для подробностей. обсуждения).Глибофф (1999) предположил, что теория эволюционного происхождения Унгера сильно повлияла на Менделя до того, как он узнал о Дарвине. Однако меньшинство историков утверждает обратное. Каллендер (1988) утверждал, что «Мендель был противником фундаментального принципа самой эволюции, то есть происхождения с модификацией …». (стр. 41), и «менделизм исторически возник как сложная форма доктрины особого творения», которая «лишенная явных ссылок на Творца, находилась в открытом конфликте с дарвиновской концепцией эволюции» (стр.72).
Самая крайняя точка зрения принадлежит Бишопу (1996), который писал: «Единственная цель Менделя при написании его статьи Pisum , опубликованной в 1866 году, заключалась в том, чтобы внести свой вклад в дискуссию об эволюции, которая бушевала с момента публикации книги Дарвина. Происхождение видов в 1859 году »и« Мендель был сторонником ортодоксальной доктрины особого творения »(стр. 212). Бишоп попытался показать, что Мендель начал свои эксперименты в 1861 году (а не в 1856 году, как заявил Мендель) как прямая реакция на изучение Дарвина.Этот аргумент основан на представлении, предложенном Бейтсоном (1909) и дополненном ДиТроккио (1991), о том, что отчет Менделя о его экспериментах был вымышленным. Орел (1971a) и Фэрбенкс и Риттинг (2001) собрали ботанические и исторические свидетельства, окончательно опровергающие как утверждение, что эксперименты Менделя были вымышленными, так и анахроничное утверждение, что Дарвин Происхождение видов побудило Менделя их инициировать. Это свидетельство дополнило хорошо изученный вывод Фишера (1936) о том, что «я считаю, что не может быть никаких сомнений в том, что его отчет [Менделя] следует воспринимать полностью буквально, и что его эксперименты проводились именно так и во многом. порядок их пересчета »(стр.132).
В предпоследнем абзаце своей статьи 1866 года Мендель отметил точку зрения Гертнера (1849) о том, что у каждого вида есть «фиксированные пределы, за которыми он не может измениться», вторя словам Линнея (Aulie, 1983). Авторы ряда статей в современной креационистской литературе и на веб-сайтах опирались на это утверждение, чтобы заявить, что Мендель поддерживал неизменность видов как принцип креационизма. В следующем предложении, однако, делается вывод Менделя о том, что «этой точке зрения нельзя признать безоговорочную действительность».Мендель пришел к выводу, что эксперименты Гертнера подтвердили «сделанное ранее предположение об изменчивости культурных растений», которое не является аргументом в пользу того, что виды являются фиксированными, а скорее, что изменение условий жизни при одомашнивании не «настолько сильно увеличивается, что виды вскоре теряют всякую стабильность. »(Эбботт и Фэрбенкс, 2016 г., стр. 419).
Утверждения, что Мендель был ортодоксальным сторонником особого творения, весьма предположительны. Мендель явно не соответствовал прототипу священнослужителя; тем не менее, имеющиеся свидетельства указывают на то, что он оставался верным своим обетам.Фрагменты недатированных рукописных заметок к двум проповедям показывают, что он использовал ботанические образы для передачи глубоко набожных и искренних посланий. В этих заметках Мендель писал: «Так должны соединиться естественное и сверхъестественное в осознании святости человека» (Zumkeller 1971, p. 250 *). Однако полное отсутствие каких-либо намеков на сверхъестественное или религию в его научных трудах, опубликованных и частных, указывает на то, что он разделял свои религиозные и научные взгляды, последовательно относясь ко всем научным занятиям с максимальной объективностью.
Мендель и Дарвин по классической статье Менделя (1866–1884)
Есть ли какие-либо свидетельства того, что Дарвин читал статью Менделя или читал о нем? Этот вопрос обсуждается более 50 лет (Vorzimmer 1968), и краткий ответ — категоричное «нет». Мендель получил сорок экземпляров своей статьи; Известна судьба лишь немногих (Орел, 1976, 1996). Ходят слухи, что необрезанный оттиск работы Менделя был обнаружен в коллекции Дарвина после его смерти (примеры см. В Hennig 2000; Leonard 2005; Fishman 2018), но при этом отсутствуют убедительные доказательства, подтверждающие это.Вероятно, это произошло из-за того, что книга Фокке (1881 г.), Die Pflanzen-Mischlinge ( Гибриды растений ) была в библиотеке Дарвина с кратким изложением экспериментов Менделя, однако страницы этих сводок остались неразрезанными. Тот факт, что Дарвин владел этой книгой, вероятно, превратился в слух о том, что у него есть неразрезанный оттиск статьи Менделя, хотя на самом деле у него была неразрезанная ссылка на нее, приобретенная немногим более чем за год до его смерти (Fairbanks and Rytting 2001).
Переписка Менделя с Нэгели
Другой вопрос, почему нет доказательств того, что кто-либо, включая Менделя, пытался сообщить Дарвину о его работе.Мендель послал оттиск Карлу Вильгельму фон Нэгели, который ответил, положив начало сотрудничеству с 1867 по 1873 год. Нэгели часто критикуют за то, что он не полностью усвоил статью Менделя 1866 года, но еще одним упущением является его очевидное пренебрежение к информированию Дарвина о Менделе. Нэгели переписывался с Менделем и Дарвином примерно в то же время, отправив свое первое письмо Менделю 24 февраля 1867 г. (Darbishire 1906) и первое письмо Дарвину 31 марта 1867 г. (Burkhardt and Secord 2006). Хотя большая часть переписки Нэгели с Дарвином была утеряна, в том, что осталось, нет никаких указаний на то, что он упоминал Менделя, хотя его переписка с обоими адресована одному и тому же растению: Hieracium (ястребиное растение).
Распространенная критика, что Нэгели непреднамеренно сбил с толку Менделя, поощряя его проводить исследования Hieracium , не имеет под собой оснований. Мендель полностью ожидал результатов, которые он наблюдал в Hieracium , поскольку он подтвердил его описание постоянных гибридов в его классической статье 1866 года (Van Dijk and Ellis 2016). По мнению Менделя, гибридов Pisum были вариабельными, а гибридов Hieracium — постоянными (Mendel 1870). Более того, Мендель и Нэгели были далеко не одиноки в исследовании Hieracium .В 1860-х и 70-х годах этот род стал моделью эволюции растений с точки зрения гибридизации и биогеографии. Другими, которые одновременно активно исследовали Hieracium , были Антон Кернер фон Марилаун, Александр Маковский и Густав Ниссл фон Майендорф, двое последних друзей, сотрудников и соратников Менделя по Обществу естествознания в Брно (Iltis 1966).
Нэгели был известным сторонником эволюции и Дарвина. В статье Менделя (1870 г.) «Über einige aus künstlicher Befruchtung gewonnenen Hieracium -Bastarde» («На Hieracium Hybrids, полученных путем искусственной гибридизации») Нагели дважды упоминается, сначала без его имени, но с его соединением. происхождение многочисленных и постоянных промежуточных форм в последнее время вызывает немалый интерес, поскольку известный специалист по Hieracium в духе дарвиновского учения отстаивал точку зрения, согласно которой эти формы должны рассматриваться как [возникающие] в результате трансмутации потерянные или все еще существующие виды »(Stern and Sherwood, 1966, p.51). Это единственное открытое упоминание Менделем Дарвина в его публикациях. Позже в газете Мендель поблагодарил Нэгели по имени за то, что он прислал ему растительные материалы.
Комментарии Менделя в книге Дарвина «Разнообразие животных и растений при одомашнивании»
Мендель трижды частным образом упоминал Дарвина в письмах к Нэгели, каждый раз в 1870 году, ссылаясь на книгу Дарвина «Разнообразие животных и растений при одомашнивании» (Darwin 1868a, 1868b). У Менделя был немецкий перевод этой книги (Darwin 1868c, 1868d), и он подробно комментировал второй том.Орел (1996) насчитал только пять аннотаций в копии Менделя первого тома (которая не переплетена и в основном не обрезана) по сравнению с 57 во втором (вырезанном и переплетенном). На сегодняшний день аннотации Менделя во 2 томе цитируются лишь в виде нескольких избранных цитат (Орел, 1971b, 1996). Я собрал их полностью в виде пронумерованных записей в файле дополнительной информации 1 (SIF1).
Из первых пяти аннотаций (записи 1–5 SIF1) три находятся в главе 15 («Пересечение»), одна — в главе 22 (Причины изменчивости) и одна — в главе 26 («Законы изменения», продолжение — резюме ).Они рассказывают о наблюдениях, которые Дарвин не может объяснить, но которые полностью объясняются теорией Менделя. Например, второй аннотированный отрывок описывает менделевскую доминантно-рецессивную модель, хотя и в неколичественных терминах (в оригинальном английском Дарвина): «эти цвета никогда не смешивались, но потомство родило чисто белые или чисто желтые цветы; первое — в большей пропорции ». (стр. 124, SIF1, запись 2).
Подавляющее большинство аннотаций сильно сгруппированы (52 из 57) в главе 27 (Предварительная гипотеза пангенезиса) и сосредоточены на трех темах: (1) собственно пангенезис, (2) относительный вклад родителей-мужчин и женщин. во время оплодотворения и (3) влияние изменения условий жизни на наследственную изменчивость.Мендель обратился к этим двум последним темам в своей классической статье (Mendel 1866).
Аннотации Менделя в главе 27, как правило, подчеркивают неуверенность Дарвина в отношении пангенезиса, а также выводы Дарвина, несовместимые с экспериментальными наблюдениями Менделя. Первый отмеченный отрывок в этой главе гласит: «Как отмечает Уэвелл, историк индуктивных наук:« Гипотезы часто могут быть полезны науке, если они включают в себя определенную часть неполноты и даже ошибки »(стр. .357, запись SIF1 6). Позже Мендель отмечал: «Существование свободных геммул является необоснованным предположением …» (стр. 378, запись 21 SIF1). В обоих этих отрывках Дарвин признает, что его гипотеза пангенезиса является в высшей степени спекулятивной и мало подтвержденной доказательствами, а аннотации Менделя, как здесь, так и позже, показывают его согласие.
Часть книги, которая, по-видимому, привлекла внимание Менделя больше, чем какая-либо другая, — это та, которую он отметил двойными вертикальными линиями на полях, рядом с ней большой восклицательный знак, некоторое подчеркивание слов и рукописная заметка внизу страницу (рис.2, SIF1, записи 23 и 24). Предложение Дарвина гласит: «Поскольку каждая единица или группа подобных единиц по всему телу отбрасывает свои геммулы, и поскольку все они содержатся в мельчайшем яйце или семени, а также в каждом сперматозоиде или пыльцевом зерне, их количество и мелкость должны быть чем-то определенным. немыслимо »(стр. 379, запись 23 SIF1). Рукописная заметка Менделя: «sich einem Eindrucke ohne Reflexion hingeben» («наслаждаться впечатлением, не задумываясь» *). Здесь Мендель раскрывает свой скептицизм по поводу необоснованного взгляда Дарвина на бесчисленные геммулы, которые «отбрасываются» единицами внутри тела и накапливаются в репродуктивных клетках.
Рис. 2: Аннотации Менделя на странице 497 его немецкого перевода книги Дарвина «Разнообразие животных и растений при одомашнивании» .В предложении, отмеченном Менделем, говорится (на оригинальном английском Дарвина с подчеркиванием, соответствующим подчеркнутым словам Менделя): «Поскольку каждая единица или группа подобных единиц по всему телу отбрасывает свои геммулы, и поскольку все они содержатся в самом маленьком яйце» или семя, и в каждом сперматозоиде или пыльцевом зерне их количество и мелкость должны быть чем-то невероятным ».Внизу страницы Мендель написал (в английском переводе) «произвести впечатление без размышлений» *.
Отмеченный отрывок в конце главы гласит: «Таким образом, гибридизированные растения могут быть в любой степени размножены почками, но постоянно подвержены реверсии семенами, то есть потере их гибридных или промежуточных растений. персонаж. Я не могу предложить удовлетворительного объяснения этого факта »(стр. 396, запись 36 SIF1). Мендель дал очень удовлетворительное объяснение такой реверсии в своей классической статье (1866), заметив, что доля гомозигот увеличивается при повторном самооплодотворении, феномен, который он продемонстрировал как экспериментально, так и математически, в то время как размножение почками происходит вегетативно, а не половым путем.Объяснение Менделя, конечно, противоречит пангенезису. Краткое прочтение 52 отрывков, отмеченных Менделем в этой главе, большинство из которых несовместимо с его открытиями, совершенно ясно дает понять, почему он написал Нэгели: «Утверждения Дарвина относительно гибридов родов, упомянутых в« Разнообразии животных и растений под их влиянием ». Одомашнивание », согласно другим сообщениям, во многих отношениях нуждается в исправлении» (Stern and Sherwood 1966, p. 93).
Что касается относительного вклада унаследованного материала родителями-мужчинами и женщинами, Дарвин утверждает, что вклады могут быть или не быть равными, что родитель-мужчина должен внести значительное количество геммул в яйцеклетку для успешного оплодотворения и, следовательно, оплодотворения. в растениях требуется более одного пыльцевого зерна.Мендель отметил десять отрывков, в которых Дарвин обращается к этой теме (SIF1, статьи 11, 26, 27, 28, 29, 30, 32, 46, 47 и 53).
Несмотря на то, что Мендель страдал от сильного напряжения глаз, он писал Нэгели: «Но один эксперимент показался мне настолько важным, что я не мог заставить себя отложить его на более поздний срок. Это касается мнения Наудина и Дарвина о том, что одного пыльцевого зерна недостаточно для оплодотворения семяпочки »(Stern and Sherwood 1966, p. 92). Мендель написал Нэгели, что у него были вручную оплодотворенные цветы Mirabilis jalappa с отдельными пыльцевыми зернами, и, в противоречие утверждению Дарвина в этом отрывке, он получил «18 хорошо развитых семян и такое же количество растений, из которых 10 уже всходили. цветение »(Стерн и Шервуд, 1966, с.92). Этот комментарий к Нэгели напрямую соответствует аннотациям Менделя на странице 478 его немецкого перевода, где Дарвин излагает эксперименты, о которых Мендель упоминает в своем письме Нэгели (рис. 3, SIF1, запись 11).
Рис. 3. Аннотации Менделя на странице 478 его немецкого перевода книги Дарвина «Разнообразие животных и растений при одомашнивании» .Отрезок, отмеченный Менделем, утверждает (на оригинальном английском Дарвина с подчеркиванием, соответствующим подчеркнутым словам Менделя): «даже тридцать зерен не оплодотворяли ни одного семени; но когда на рыльце было нанесено сорок зерен, образовалось несколько семян небольшого размера ».Мендель также подчеркнул «Наудин», который, как объяснил Дарвин, заметил, что для оплодотворения Mirabilis требовалось несколько зерен пыльцы (полный текст этого отрывка на английском языке см. В записи 11 SIF1). Эта аннотация особенно важна, потому что она побудила Менделя, несмотря на напряжение глаз, повторить эксперименты Нодина, изложив их в письме Нэгели. Мендель обнаружил, что в повторяющихся случаях одного пыльцевого зерна было достаточно для оплодотворения, что согласуется с его теорией наследственности.
Мендель ранее обращался к этой теме в своей классической статье 1866 года: «Согласно мнению известных физиологов, у фанерогамов с целью воспроизводства одна половая клетка и одна клетка пыльцы объединяются в единую клетку, которая может развиваться в независимый организм »(Abbott, Fairbanks, 2016, p. 420). К «известным физиологам», несомненно, относится Унгер, который был втянут в спор с Эдуардом Фенцлем (другим профессором ботаники Менделя), когда Мендель учился у них.Унгер утверждал, что одна клетка пыльцы оплодотворяет одну яйцеклетку, обеспечивая равный родительский вклад в каждое потомство растения. Фенцль утверждал, что оплодотворение пыльцы не вносит генетический материал, а служит просто для питания яйца (Olby, 1985; Orel, 1996; Klein and Klein, 2013). Наблюдения Менделя полностью соответствовали утверждению Унгера, и Мендель включил длинную сноску, подтверждающую точку зрения Унгера и противоречащую точке зрения Фенцля, не называя ни того, ни другого (Abbott and Fairbanks 2016, p.420).
Мендель снова упомянул Дарвина в своем девятом письме Нэгели: «Дарвин и Вирхов указали на высокую степень независимости, которая характерна для отдельных персонажей и целых групп характеров животных и растений. Поведение растений гибридов, бесспорно, дает важное доказательство правильности этой точки зрения»(Стерны и Шервуд 1966 года, стр. 96). Хотя две предыдущие ссылки Менделя на Дарвина представляют собой критику по конкретным темам, это объективно хвалит точность идей Дарвина.
Дарвиновское заявление Менделя Нэгели
Ближе к концу десятого письма Менделя Нэгели (18 ноября 1873 г.) это, пожалуй, самый дарвиновский отрывок среди его существующих сочинений. Здесь Мендель сделал вывод, что несколько диких видов Hieracium , которые естественным образом самоопыляются, но иногда дают гибриды в природе, в конечном итоге вымрут, потому что, если гибриды «будут повторяться слишком часто или станут постоянными, [они] в конечном итоге приведут к появлению исчезновения вовлеченных видов, в то время как то или иное из более удачно организованных потомков, лучше приспособленных к преобладающим теллурическим и космическим условиям, могло бы успешно начать борьбу за существование и продолжать ее в течение длительного периода времени, пока, наконец, не то же самое. судьба настигла его »(Stern and Sherwood 1966, p.102). Термин «борьба за существование», конечно, является характерным дарвиновским, и слова, которые использовал Мендель, «Kampf ums Dasein» такие же, как и в его немецких переводах Происхождение видов и Варианты животных и растений при одомашнивании. для фразы Дарвина «борьба за существование». Комментируя этот отрывок, Илтис (1966) воскликнул: «Клерикалисты, заклеймившие Менделя-либерала как дарвиниста, не так уж сильно ошибались!» (стр.204).
Разрыв между девятым и десятым письмами Менделя Нэгели превышал три года.В своем десятом письме Мендель отметил, что он отправит оставшиеся гибридов Hieracium в Нэгели. С тех пор Мендель резко сократил свои исследования по гибридизации растений. Нэгели снова дважды писал Менделю, но нет никаких записей о том, что Мендель ответил (Iltis 1966). Менделю удавалось проводить исследования в дополнение к административным обязанностям с того момента, как он был избран настоятелем в 1868 году, но эти обязанности становились все более обременительными. Он все чаще сталкивался с враждебными действиями со стороны своего начальства, отчасти из-за его преданности науке, а отчасти из-за его упорства в игнорировании требований церкви и государственных властей.В ближайшие годы эта враждебность усилится, оставив его поглощенным, ожесточенным и недоверчивым до самой смерти 6 января 1884 года.
Резюме и выводы
Мы можем рассматривать связь Менделя с Дарвином в частности и эволюцию в целом с двух точек зрения: во-первых, интеллектуальная среда, которая контекстуализировала его научные занятия, а во-вторых, его собственная точка зрения через его сочинения и аннотации к книгам. С точки зрения первой точки зрения, Мендель был погружен в интеллектуальную среду, весьма благоприятную для эволюции на протяжении всех его продуктивных с научной точки зрения лет, а также для Дарвина с 1863 г.С точки зрения второй точки зрения, работы Менделя раскрывают влияние этих сред, особенно Дарвина, Унгера и Нэгели.
Ни в одной из его существующих работ нет ни одного упоминания о том, чтобы Мендель отверг дарвиновскую теорию эволюции путем естественного отбора. Фактически, как отмечалось ранее, Мендель предложил дарвиновский сценарий естественного отбора, призывающий к «борьбе за существование». Его опасения по поводу Дарвина не были общими, а были сосредоточены на трех конкретных темах: предварительная гипотеза Дарвина о пангенезисе, относительный вклад родителей-мужчин и женщин в оплодотворение и влияние изменений условий жизни на наследственные вариации.По всем трем темам Мендель основывал свои взгляды на своих экспериментальных наблюдениях.
Те, кто были доверенными лицами Менделя, которые дожили до времени повторного открытия, чтобы поделиться воспоминаниями, вспомнили беспристрастный подход Менделя к Дарвину, совместимый с его существующими работами. Например, Илтис (1966) взял интервью у сотрудника Менделя, Густава Ниссла фон Майендорфа, который, по словам Илтиса, «рассказывает, что Мендель, который очень интересовался идеей эволюции, был далеко не противником дарвиновской теории, но всегда когда всплыло имя Дарвина, он сказал, что теория неадекватна, что чего-то не хватает »(стр.103). Племянник Менделя, Фердинанд Шиндлер, врач, который во время учебы жил недалеко от Менделя и часто навещал его со своими братьями, высказал подобное воспоминание в письме 1902 года Уильяму Бейтсону. Принося извинения за любые грамматические ошибки, Шиндлер написал письмо на английском языке, цитируемое здесь точно так же, как в оригинале:
Умерший аббат Мендель был человеком либеральных принципов…. Он с величайшим интересом читал работы Дарвина в немецком переводе и восхищался его гением, хотя и не соглашался со всеми принципами этого бессмертного натурфилософа.Но может случиться так, что мой дядя в конце своей жизни отказался от вопросов научной эволюции, потому что у него было много клерикальных врагов. Он часто говорил нам, племянникам, что мы найдем в его наследстве документы для публикации, которые он не мог опубликовать при жизни. Но от обители мы ничего не получали, даже на память.
Воспоминания Шиндлера о рукописях Мендель отложил публикацию из страха перед «клерикальными врагами» может пролить свет на вопиющий вопрос: почему Мендель в своих публикациях явно включает завуалированные ссылки на Дарвина, Унгера и Нэгели, не называя их? Все трое были известны в то время своими эволюционными взглядами, Дарвин и Унгер страдали от публичных поношений со стороны видных представителей духовенства, последнее на форуме, свидетелем которого стал Мендель.Контекстуальная формулировка в статьях Менделя такова, что пропуски названий не являются явными. Коллеги-натуралисты, читающие работы Менделя, знали бы, о ком он имел в виду, но безымянные личности не были бы очевидны для тех, кто не знаком с соответствующей наукой. Возможно, это был подход Менделя к признанию экспертов-эволюционистов, который не был бы очевиден для его «клерикальных врагов». Тем не менее, как это часто бывает, мы можем здесь только строить догадки, учитывая отсутствие прямого обоснования.
Когда существующие свидетельства рассматриваются в целом, возникает образ Менделя как дотошного ученого, который занимался вопросами эволюции, хорошо разбирался в эволюционной литературе своего времени, был готов в частном порядке указывать на ошибки в конкретных случаях. Темы в книгах Дарвина все же хвалят его по сравнению с другими, и чьи общие исследования гибридизации растений имели эволюционное значение, совпадающее с таковыми его предшественников и современников. Слова и аннотации Менделя не отражают ни пылкой защиты Дарвина, которую поддерживали некоторые из его ближайших соратников, ни язвительного опровержения теории эволюции, которым выставляли напоказ некоторые из его сослуживцев.Как отмечают Хартл и Фэрбенкс (2007), «прежде всего, работа Менделя, кажется, отражает простоту, скромность и бесхитростность автора» (стр. 975). Эти атрибуты выходят далеко за рамки классической работы Менделя; они пронизывают все его сочинения и согласуются с воспоминаниями тех, кто его знал.
Мутации — сырье для эволюции
Мутация — это изменение последовательности ДНК организма. Что вызывает мутацию? Мутации могут быть вызваны источниками высокой энергии, такими как радиация, или химическими веществами в окружающей среде.Они также могут появляться спонтанно во время репликации ДНК.
Мутации обычно делятся на два типа: точечные мутации и хромосомные аберрации. При точечных мутациях изменяется одна пара оснований. Например, геном человека содержит более 3,1 миллиарда оснований ДНК, и каждое основание должно быть точно воспроизведено, чтобы произошло деление клетки. Ошибки случаются, хотя и на удивление редко. Приблизительно одна из каждых 10 10 (10 000 000 000) пар оснований изменяется. Самый распространенный тип ошибки — это подмена очков.Более редким явлением является невозможность скопировать одну из баз (удаление), создание двух копий для одной базы (точечное дублирование) или добавление новой базы или даже нескольких баз (вставка). Хромосомные аберрации — это крупномасштабные мутации, которые могут возникать во время мейоза в результате неравных событий кроссинговера, проскальзывания во время рекомбинации ДНК или из-за активности мобильных событий. Гены и даже целые хромосомы могут быть заменены, дублированы или удалены из-за этих ошибок (рис. 1).
Рисунок 1: Мутации в последовательности ДНК семи родственных видов тропических рыб (данные взяты из интрона 6 гена LDH-A , секвенированного автором из эпинефелиновых серранид)
Вверху — исходные последовательности, внизу — последовательности, скорректированные для вертикального выравнивания аналогичных оснований ДНК.Замены точек выделены красным цветом, а желтая рамка с пунктиром указывает на удаление 12 оснований.
Мутации могут иметь ряд эффектов. Часто они могут быть вредными. Другие имеют незначительный вред или не оказывают никакого вредного воздействия. А иногда, хотя и очень редко, изменение последовательности ДНК может даже оказаться полезным для организма.
Мутация, которая возникает в клетках организма и не передается последующим поколениям, является соматической мутацией. Мутации, происходящие в гамете или в клетке, которые дают начало гаметам, являются особенными, потому что они влияют на следующее поколение и могут вообще не влиять на взрослых.Такие изменения называются мутациями зародышевой линии, потому что они происходят в клетке, используемой для размножения (зародышевой клетке), что дает возможность изменениям со временем стать более многочисленными. Если мутация пагубно влияет на фенотип потомства, мутация называется генетическим заболеванием. С другой стороны, если мутация положительно влияет на приспособленность потомства, это называется адаптацией. Таким образом, все мутации, влияющие на приспособленность будущих поколений, являются агентами эволюции.
Мутации необходимы для эволюции.Изначально каждая генетическая особенность каждого организма была результатом мутации. Новый генетический вариант (аллель) распространяется посредством воспроизводства, и дифференциальное воспроизводство является определяющим аспектом эволюции. Легко понять, как мутация, которая позволяет организму питаться, расти или воспроизводиться более эффективно, может привести к тому, что мутантный аллель со временем станет более распространенным. Вскоре популяция может весьма экологически и / или физиологически отличаться от исходной популяции, которой не хватало адаптации.Даже вредные мутации могут вызывать эволюционные изменения, особенно в небольших популяциях, путем удаления особей, которые могут нести адаптивные аллели других генов.
Рисунок 2: История серой древесной лягушки, Hyla versicolor , является примером мутации и ее потенциальных последствий. Когда серая древесная лягушка Hyla chrysocelis не смогла отсортировать свои 24 хромосомы во время мейоза, в результате получилось H. versicolor . Эта древесная лягушка идентична по размеру, форме и цвету H.chrysocelis , но имеет 48 хромосом и призыв к спариванию, который отличается от исходного H. chrysocelis .
Большинство мутаций происходят в отдельных точках гена, изменяя, возможно, один белок, и поэтому могут казаться несущественными. Например, гены контролируют структуру и эффективность пищеварительных ферментов в слюнных железах (и всех других позвоночных). На первый взгляд может показаться, что мутации ферментов слюны не могут повлиять на выживаемость. Тем не менее, именно накопление небольших мутаций в слюне является причиной змеиного яда и, следовательно, большей части эволюции змеи.Естественный отбор у некоторых предковых змей благоприятствовал ферментам со все более агрессивными свойствами, но сами мутации были случайными, создавая разные яды в разных группах змей. Змеиный яд на самом деле представляет собой коктейль из разных белков с разными эффектами, поэтому генетически родственные виды имеют смесь, отличную от смеси других ядовитых семейств змей. Предки морских змей, коралловых змей и кобр (семейство Elapidae) развили яд, который атакует нервную систему, в то время как яд гадюк (семейство Viperidae; включая гремучих змей и бушмейстеров) действует на сердечно-сосудистую систему.Оба семейства имеют много разных видов, которые унаследовали небольшое преимущество в силе яда от своих предков, и по мере накопления мутаций разнообразие ядов и разнообразие видов со временем увеличивалось. Хотя на историю многих видов повлияло постепенное накопление крошечных точечных мутаций, иногда эволюция протекает намного быстрее. У некоторых типов организмов есть предок, который не смог правильно пройти мейоз до начала полового размножения, что привело к полному дублированию каждой пары хромосом.Такой процесс создал событие «мгновенного видообразования» у серой древесной лягушки Северной Америки (рис. 2).
Следствием удвоения размера генома у растений часто являются ненормально большие семена или плоды, что может иметь явное преимущество, если вы цветущее растение! Большинство злаков, которые едят люди, имеют огромные семена по сравнению с другими травами, и это часто связано с геномными дупликациями, которые произошли у предков современного риса и пшеницы, и, поскольку ошибка произошла в репродуктивных органах, была успешно передана будущим поколениям.Сами люди подражали этому процессу, скрещивая отдельные растения с самыми крупными плодами и семенами в процессе искусственного отбора, создавая многие из наших современных сортов сельскохозяйственных культур. Идея эволюции путем естественного отбора, впервые описанная Чарльзом Дарвином и Альфредом Расселом Уоллесом, требует дифференцированного выживания из-за того, что некоторые люди обладают большей эволюционной приспособленностью. Независимо от того, влияют ли на эту приспособленность генетические нарушения, ядовитая слюна или увеличенное потомство, наследственные вариации могут возникнуть только в результате мутации.Эволюция просто невозможна без случайных генетических изменений исходного материала.
Смешанное наследование — обзор
Мы надеемся, что анализ, проведенный в предыдущих главах, приведет читателей к пониманию того, что даже несмотря на то, что основные эволюционные ограничения не были идентифицированы на уровне генов, нельзя исключать идею о том, что генетический вклад на уровне генома может быть самым важным эволюционным препятствием. Согласно теории генома, единицей эволюционного отбора является геном, а не отдельные гены (Heng, 2007b, 2009).Таким образом, поиск главного геномного вклада эволюционных ограничений должен быть сосредоточен на геноме, а не на генах. Основываясь на этой точке зрения, особенно после осознания того, что изменения генома приводят к макроэволюции, была выдвинута гипотеза, что давно игнорируемые ограничения на уровне генома обеспечат ключевой генетический механизм застоя организма.
6.5.2.1 Почему важно обсуждать ограничения на уровне генома?
Учитывая сложность как биологической системы, так и эволюционного процесса, необходимо изучить вопрос о нескольких уровнях ограничений и их динамических взаимодействиях, включая гены / эпигены, белки, генетические и белковые сети, геном, индивидуум , популяции, общество и экология (Heng, 2009).Поскольку Футуйма представил всестороннее резюме и проницательный анализ для многих из этих уровней (Futuyma, 2010), уровень генома будет в центре внимания в этом разделе не только потому, что он более или менее игнорировался в традиционных эволюционных исследованиях, но и потому, что он представляет собой основной уровень ограничения / контроля и определяет потенциальные взаимодействия, которые могут происходить на различных нижних и верхних уровнях. Для сравнения, другие типы эволюционных ограничений важны, но не являются основными определяющими факторами по сравнению с ограничениями генома.Например, снижение ограничений на экологическом уровне может повысить нестабильность системы. В условиях такой нестабильности возрастают уровни краткосрочной адаптации и изменений генома, которые обеспечивают повышенный потенциал для образования новых видов. Однако до тех пор, пока не будет сформирован новый кластер генома, нет возможности передать эти измененные геномы. Кластер генома относится к нескольким людям, которые имеют одинаковый геном и могут спариваться. Появление кластеров генома обычно происходит в переходный период, когда появляются и утверждаются новые виды.
Поскольку важность генома обсуждалась в предыдущих главах, будут кратко изложены некоторые выводы, которые имеют непосредственное отношение к этой теме.
а. Геном кодирует пакет всей системы.
Традиционное генетическое кодирование конкретно относится к комбинациям трех нуклеотидов, которые определяют аминокислоты в белках или РНК. 4D геномика, напротив, подчеркивает всестороннее кодирование как системное наследование, возникающее в результате геномных отношений между всеми генами и другими структурными / регуляторными элементами одной и той же системы.Примечательно, что элементы управления информацией на уровне генома могут не храниться непосредственно в отдельной молекуле ДНК, а вместо этого могут существовать в топологической взаимосвязи между генетическими локусами в ядрах.
Это объяснение также имеет смысл и согласуется с различными биологическими наблюдениями. Базовая кодировка для изготовления материалов универсальна для видов, имеющих общего предка. Однако похожие гены (материалы или инструменты) могут создавать множество геномных архитектур с определенными наборами хромосом, которые определяют разные виды.Каждый геном представляет собой уникальный код взаимодействия или сборки, который не является общим для других видов, за исключением сходства между видами, принадлежащими к одному роду и / или семейству. «Материальное» кодирование ДНК по существу сохраняется среди видов из-за его базового уровня генетической организации. Напротив, архитектурное кодирование очень динамично и менее предсказуемо, поскольку этот новый тип информации не кодируется в последовательности ДНК, а существует на хромосомном уровне и достигается за счет принципа самоорганизации, основанного на генах и топологических отношениях генома.Такие свойства на уровне генома наследуются для каждого вида и гарантируются консервацией генома.
г. Секс защищает целостность генома.
Поддержание целостности генома было горячей темой в молекулярной биологии. Предлагаемые механизмы включают, помимо прочего, различные системы репарации ДНК, контрольные точки клеточного цикла, запрограммированную гибель клеток, надежность сети, разделение между клетками зародышевой линии и соматическими клетками и эпигенетический сброс. Недавно было признано, что половое размножение играет ключевую роль в поддержании целостности генома у большинства эукариот, размножающихся половым путем (глава 5).
Контекст генома можно сохранить, поддерживая как состав хромосом, так и общий порядок генетических локусов вдоль каждой хромосомы. Таким образом, пол служит фильтром для устранения наиболее значительных изменений на уровне хромосом, одновременно допуская изменения на уровне генов. Таким образом, мейоз выполняет две основные функции: он способствует изменениям генов, но ограничивает изменения генома.
г. Целостность генома, опосредованная полом, обеспечивает долгосрочную эволюционную стабильность.
Поскольку секс может сохранять границы системы, он обеспечивает идеальный баланс между геномной динамикой и ограничениями в размножающихся половым путем организмах.В краткосрочной перспективе динамика генов, возникающая в результате мутации, генетической рекомбинации и сплайсинга, может эффективно обеспечивать эволюционную адаптацию путем передачи генома, обеспечивая при этом в долгосрочной перспективе, что геном сохраняет свою структуру. Во время каждого нормального репродуктивного цикла большинство накопленных генетических изменений на уровне генома будет устранено, возвращая систему в исходное состояние. Несмотря на некоторые мелкие вариации числа копий и некоторые интеграции ретровирусов, которые могут накапливаться, для большинства видов, размножающихся половым путем, структура генома в основном остается одним и тем же поколением за поколением.Напротив, бесполые организмы постоянно проходят как микро-, так и макроэволюцию, поскольку нет эффективных механизмов, таких как секс, для очистки их геномов и сохранения четко определенных границ генома.
г. Геном, а не ген — это макроэволюционная единица отбора.
Возникла обширная дискуссия о том, почему гены или особи должны или не должны быть основным объектом естественного отбора (Mayr, 1997). Предполагается, что геном является основным объектом отбора, особенно на макроэволюционной фазе (Heng, 2009, 2015).Следующие пункты дополнительно подтверждают эту концепцию. (1) Геном и ген представляют разные уровни генетической организации, где геном определяет наследование системы, важный компонент макроэволюции (см. Главу 4). (2) Существует разделение между индивидуумами и геномами (некоторые люди с измененными геномами могут передавать нормальный геном следующему поколению с более высокой частотой из-за полового фильтра (например, XXX, XYY и люди с синдромом Дауна) ( 3) Существует различие между клетками зародышевой линии (передаваемый геном) и конечной функцией соматических клеток.Обратите внимание, что эта разница может вносить вклад в индивидуальные вариации, происходящие за пределами генома. Таким образом, существует разница между успехом фенотипа и типами генома, которые размножаются (Heng, 2010).
Здесь стоит обсудить два момента. В микроэволюции избирательный вклад отдельных генов можно измерить, если они демонстрируют относительно простые черты. Однако большинство генов не являются независимыми информационными единицами, а скорее зависят от определенной геномной сети в определенных условиях окружающей среды.Отбор действует на пакет генома, а не на конкретные гены. Конечно, геном определяет границу генетического потенциала вида, который включает большое количество комбинаций генов, которые проявляются разными людьми.
В макроэволюции отбор на уровне генома имеет решающее значение во время видообразования, поскольку вновь сформированный геном должен выжить в условиях конкуренции. (Традиционная концепция настаивает на том, что видообразование часто вызывается полиплоидией или географической изоляцией, что приводит к большому генетическому дрейфу, не требующему какого-либо отбора.Однако в эволюции рака очевиден отбор на уровне генома). Как только вид устойчиво установлен, геном служит ограничением для поддержания системы, в которой половое размножение предотвращает резкие изменения на уровне генома в зародышевой линии, в то время как динамика генов / геномов на уровне соматических клеток способствует индивидуальному выживанию. Эффект при краткосрочной адаптации заключается в том, что динамика генов и эпигенов высока, тогда как при долгосрочном отборе, где силы отбора постоянно меняются, мутации генов / изменения эпигена приходят и уходят.До тех пор, пока размер популяции является здоровым и разные люди демонстрируют различные характеристики (интерактивные результаты окружающей среды и различные генетические профили, определяемые геномом), данный вид может передаваться путем передачи основного генома, который также определяет различные потенциальные фенотипы при различных условиях. среды.
Неудивительно, что Майр заявил:
Перечитывая свой анализ, я был весьма удивлен, насколько редко мне приходилось упоминать генетические аспекты, ответственные за фенотип.По-видимому, не имеет большого значения, как скомбинированы гены или насколько генотип должен быть изменен, при условии, что результирующий фенотип будет одобрен отбором. Важна адаптированность конечного продукта.
Mayr, 1997
Mayr частично прав, потому что все фенотипы являются продуктами взаимодействия генотипа и окружающей среды. Все отобранные особи также передают основной геном (репрезентативный кариотип), который содержит постоянно изменяющиеся генные и эпигенные ландшафты.
6.5.2.2 Различные факторы способствуют ограничению генома
Очень важно осознание того, что большинство эукариот достигли эволюционных ограничений благодаря целостности генома, сохраненной половым размножением. В нем рассматривается взаимосвязь между сохранением генома и динамикой генных мутаций и эпигенетических изменений. Следует отметить, что многие другие факторы, такие как экологические и даже социальные взаимодействия, могут либо способствовать, либо уменьшать ограничения генома. Важен баланс между динамикой и ограничениями разных уровней.Например, повышенное генетическое разнообразие на уровне генов полезно для индивидуального выживания, но сохранение генома гарантирует, что выжившая система может поддерживаться в течение длительного времени без превращения в новую систему. Интересно, что более высокий уровень ограничений системы уменьшит влияние динамики более низкого уровня в долгосрочной перспективе, несмотря на его важность для адаптации организма в краткосрочной перспективе. Это можно объяснить как новую версию истории Красной Королевы: попробуйте все, чтобы поддерживать жизнь, включая генные мутации, но не изменяйте свой геном, чтобы изменить то, кем вы являетесь.В конечном итоге имеет значение только существование генома.
Кроме того, то, как система ограничивает свою геномную систему, также определяет модель эволюции. У эукариот, которые все содержат хромосомы, половые фильтры затрудняют изменение генетической идентичности данного вида, поскольку большинство хромосомных модификаций устраняются полом. Однако, когда происходит видообразование, реорганизация генома может привести к очень разным геномам, и некоторые из них могут иметь потенциал для больших эволюционных скачков.Напротив, в то время как прокариоты могут развиваться как за счет микро-, так и макроэволюции, их макроэволюция часто менее радикальна по сравнению с организмами, способными к реорганизации генома на основе хромосом (например, количество и степень создания новой и сложной геномной информации). Таким образом, формирование типичных хромосом и установление пола представляют собой ключевые эволюционные события, разделяющие эти две основные группы. Соответственно, эволюционные паттерны у них различаются. Вероятно, что у прокариот экологические ограничения эквивалентны мощным ограничениям генома, опосредованным полом.
(PDF) Дарвин об изменениях и наследственности
Дарвин о вариациях и наследственности 455
Годфри-Смит, Питер. 1996. Сложность и функции разума в природе. Кембридж, Великобритания:
Cambridge University Press.
Гулд, Стивен Дж. 1977 г. «Вечные метафоры палеэнтологии». In Patterns of Evolution as
Illustrated by Fossil Record, ed. Энтони Халлам, стр. 1-26. Амстердам: Эльзевир.
Гриземер, Джеймс Р. 1993. «Информационный ген и материальное тело: обобщение
эволюционной теории с помощью абстракции.»Философский факультет Калифорнийского университета
, Дэвис.
—— В процессе подготовки. Воспроизведение в эволюционном процессе.
Геккель, Эрнст. 1866. Generelle Morphologie, 2 vols. Берлин: Реймер.
Ходж, М. Дж. С. 1985. «Дарвин как теоретик пожизненного поколения». В дарвиновском
Наследие, изд. Дэвид Кон, стр. 207–243. Принстон: Издательство Принстонского университета.
Халл, Дэвид Л. 1973. Дарвин и его критики. Чикаго: Издательство Чикагского университета.
Кон, Дэвид. 1980. «Теории для работы: отвергнутые теории, воспроизведение и путь Дарвина
к естественному отбору». В исследованиях по истории биологии, под ред. Уильям Коулман и
Камилла Лимож, т. 4. С. 67–170. Балтимор: Издательство Университета Джона Хопкинса.
Кун, Томас С. 1970. Структура научных революций, 2-е изд. Чикаго: Университет
, издательство Chicago Press.
Левинс, Ричард и Ричард С. Левонтин. 1985. Диалектический биолог.Кембридж, Массачусетс:
Издательство Гарвардского университета.
Майеншайн, Джейн. 1987. «Наследственность / развитие в Соединенных Штатах, около 1900 года». История
и философия наук о жизни 9: 79–93.
Олби, Роберт К. 1963. «Рукопись Пангенезиса Чарльза Дарвина». Британский журнал
History of Science 1: 250–63.
—— 1966. Истоки менделизма. Нью-Йорк: Schocken Books.
Осповат, дов. 1981. Развитие теории Дарвина: естественная история, естественная теология и естественный отбор, 1838–1859 гг.Кембридж, Великобритания: Издательство Кембриджского университета.
Ояма, Сьюзан. 1985. Онтогенез информации: системы развития и эволюция.
Кембридж, Великобритания: Издательство Кембриджского университета.
Пекхэм, Морс (ред.). 1959. Чарльз Дарвин «Происхождение видов»: текст Variorum.
Филадельфия: Издательство Пенсильванского университета.
Ричардс, Роберт Дж. 1992. Значение эволюции. Чикаго: Издательство Чикагского университета.
Робинсон, Глория. 1979. Прелюдия к генетике.Теории материальной субстанции наследственности:
Дарвин Вейсманну. Лоуренс, KS: Coronado Press.
Ворзиммер, Питер. 1963. «Чарльз Дарвин и смешанное наследование». Исида 54: 371–90.
—— 1970. Чарльз Дарвин: годы противоречий. Происхождение видов и его критики
1859–1882. Филадельфия: издательство Temple University Press.
Вейсманн, август. 1868. Über die Berechtigung der Darwin’schen Theorie. Лейпциг:
Вильгельм Энгельманн.
—— 1875.«Über den Saison-Dimorphismus der Schmetterlinge». In Studien zur Descendenz-
Theorie, 2 vols., Pp. 1–94. Лейпциг: Вильгельм Энгельманн.
—— 1882. «О сезонном диморфизме сливочных масел». В «Исследованиях теории происхождения»,
, перевод Рафаэля Мелдола, стр. 1–160. Лондон: Сэмпсон Лоу, Марстон, Сирл и
Ривингтон.
Винтер, Расмус. В стадии подготовки. «Вейсман о вариации зародышевой плазмы». Представлено в 1999 г. на встрече
Международного общества истории, философии и социальных исследований биологии
в Оахаке, Мексика, 7–11 июля 1999 г.
Циркле, Конвей. 1946. «Ранняя история идеи наследования приобретенных
персонажей и пангенезиса». Труды Американского философского общества 35:
91–151.
Генетическая вариация | Национальное географическое общество
Под генетическими вариациями понимаются различия между геномами представителей одного и того же вида. Геном — это вся наследственная информация — все гены — организма. Например, геном человека содержит от двадцати до двадцати пяти тысяч генов.
Гены представляют собой единицы наследственной информации, и они несут инструкции для построения белков. Гены, которые закодированы в этих белках, позволяют клеткам функционировать. Большинство организмов, которые размножаются половым путем, имеют по две копии каждого гена, потому что каждая родительская клетка или организм жертвует одну копию своих генов своему потомству.
Leave A Comment