
Виды и примеры бесполого размножения организмов
Все живые существа должны размножаться, чтобы передавать гены потомству и обеспечивать выживание вида. Естественный отбор, процесс эволюции, который выбирает, какие черты организмов являются благоприятными для данной среды, и какие из них неблагоприятные. Те особи, которые имеют нежелательные черты, в конечном итоге исчезнут, а организмы с «хорошими» чертами будут жить достаточно долго, чтобы размножаться и передавать эти гены следующему поколению.
Существует два типа размножения: половое и бесполое. Половое размножение предусматривает слияние мужских и женских половых клеток во время оплодотворения, которые в итоге создадут потомство частично похожее на родителей. Бесполое размножение требует только одного родителя, который передаст все свои гены потомству. Это означает, что нет смешивания генов, и потомство на самом деле является клоном родителя (запрещающим какие-либо мутации).
Бесполое размножение обычно распространено в менее сложных видах и является довольно эффективным.
Существует несколько различных видов бесполого размножения. Давайте рассмотрим некоторые наиболее распространенные.
Бинарное деление
Почти все прокариоты размножаются бинарным делением. Этот вид размножения очень похож на процесс митоза эукариот. Однако, поскольку ядра нет, а ДНК прокариот обычно находится только в одном кольце, этот процесс не такой сложный, как митоз. Бинарное деление начинается с одной клетки, которая копирует ее ДНК и затем делится на две идентичные клетки.
Это очень быстрый и эффективный способ создания потомства для бактерий, и подобных типов клеток. Однако, если бы в процессе размножения происходила мутация ДНК, это могло бы изменить генетику потомства, и они больше не были бы идентичными клонами.
Почкование
Другой вид бесполого размножения называется почкованием. Почкование происходит когда новый организм или потомство вырастает со стороны родителя через часть, называемую почкой. Потомок остается привязан к предку, пока не достигнет зрелости и не станет независимым организмом. У одного родителя может быть много почек и много потомков одновременно.
С помощью почкования могут размножаться как одноклеточные организмы, такие как дрожжи, и многоклеточные, такие как гидры. Опять же, потомство является клоном родителя, если не происходит какая-либо мутация во время копирования ДНК или размножения клеток.
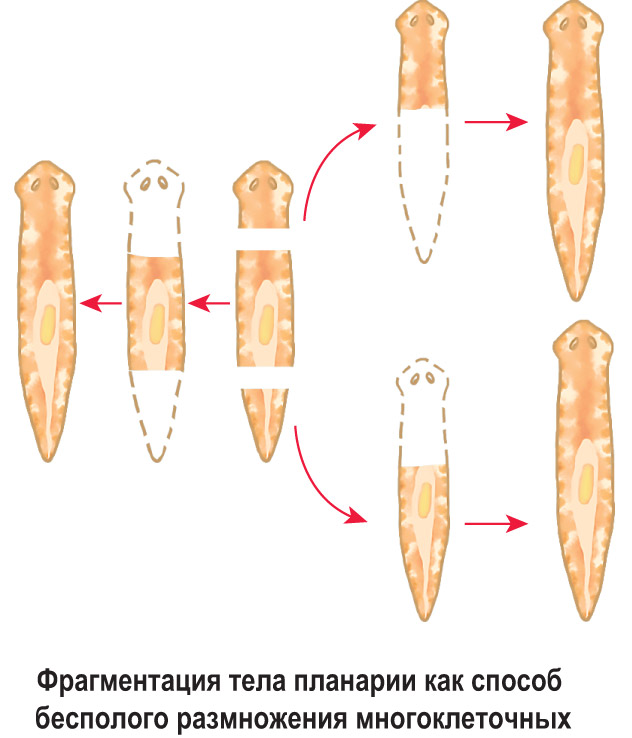
Фрагментация
Некоторые виды организмов имеют множество жизнеспособных частей, которые могут жить независимо от одной особи. Эти виды способны размножаться бесполым способом размножения, известном как фрагментация. Она происходит, когда часть индивидуума отделяется, и из нее образуется совершенно новый организм. Исходный организм также восстанавливает часть тела, которая отделилась.
Наиболее известным организмом, который подвергается фрагментации, является морская звезда. Морские звезды могут отделить от тела любую из своих пяти рук, которые затем станут их потомством. Это в основном связано с их радиальной симметрией. У них центральное нервное кольцо посередине, которое разветвляется на пять лучей или рук. Каждая рука имеет все элементы, необходимые для создания совершенно новой особи путем фрагментации. Губки, некоторые плоские черви и грибы также могут размножаться с помощью фрагментации.
Партеногенез
Чем сложнее организмы, тем более вероятно, что они будут подвергаться половому, а не бесполому размножению. Однако есть некоторые сложные животные и растения, которые способны размножаться через партеногенез, когда это необходимо. Это не является предпочтительным методом размножения для большинства этих видов, но он может стать единственным способом оставить после себя потомство по разным причинам.
Партеногенез — это вид размножения, когда потомство появляется из неоплодотворенного яйца. Отсутствие доступных партнеров, непосредственная угроза жизни самки или другие подобные ситуации могут привести к тому, что партеногенез будет необходим для сохранения вида. Разумеется, это не будет идеальным вариантом, потому что потомок станет клоном матери.
Некоторые животные, которые могут размножатся с помощью партеногенеза, включают насекомых (пчелы и кузнечики), ящериц (комодский варан), и очень редко встречаются у птиц.
Размножение спорами
Многие растения и грибы используют споры как вид бесполого размножения. Эти типы организмов подвергаются жизненному циклу, называемому чередованием поколений, при котором они проходят разные фазы своей жизни, характеризующиеся наличием диплоидных или гаплоидных клеток. Во время диплоидной фазы они называются спорофитами и производят диплоидные споры, которые используются для бесполого размножения. Виды, которые образуют споры, не нуждаются в партнере или оплодотворении, чтобы произвести потомство.
Не нашли, то что искали? Используйте форму поиска по сайту
Понравилась статья? Оставь комментарий и поделись с друзьями
🤰 Фрагментация ДНК сперматозоидов — цена, сдать тест на фрагментацию ДНК сперматозоидов
Первые полгода без предохранения — полет нормальный. Потом начали волноваться. Потом перепробовали все — от диет и стояния вверх ногами после секса, до обследования в лучших клиниках. Врачи разводят руками. Дальше 4 протокола ЭКО за полтора года в 2 клиниках. На выходе — одна замершая беременность во втором протоколе.
И тут у меня опустились руки. Начались проблемы с мужем — он гораздо раньше, чем я, предлагал родить детей, и вот теперь стал пилить меня, мол, из-за тебя упустили время… Короче, неизвестно чем бы все это кончилось, если бы не Его Величество Случай.
Начались проблемы с мужем — он гораздо раньше, чем я, предлагал родить детей, и вот теперь стал пилить меня, мол, из-за тебя упустили время… Короче, неизвестно чем бы все это кончилось, если бы не Его Величество Случай.
Как-то летом приехала к маме на дачу, а там старая знакомая с мужем и двумя карапузами ковыряется. Я ее знала если не с пеленок, то с песочницы точно. У нас дачи рядом, поэтому нас на лето родители все время к бабушкам туда отвозили. А потом уже стали сами туда к родителям ездить или с компаниями. В общем, не близкие подруги, конечно, но и не чужие. Позвала она меня вечерком в гости. Посидели мы с ней, выпили вина, смотрю я на Ирку, на ее малышей, и тут меня «развезло». Расплакалась, рассказала ей все. Оказалось, что у Ирки тоже были проблемы по женской части, и дети, оказывается, «Экошки», двойняшки. Она посоветовала мне своего врача.
Первый прием. Репродуктолог Анна Владиславовна Морозова. Мне как-то сразу с ней было психологически комфортно и спокойно. Как говорится, на одной волне, нет этого обычного врачебного проглядывающего «если ничего не понимаешь, молчи и делай как я тебе говорю». Все обсудили. Она сказала правильную вещь — «Бесплодие неясного генеза — это не значит, что причины нет. Это значит что ее пока не нашли. Будем искать». Протокол, 11 клеточек, 8 эмбрионов, 5 с хорошей морфологией. А дальше понятно — отбор эмбрионов по полу, и с первой попытки успешная беременность. И вот я, 39-летняя «молодая мамаша», сейчас сижу на кухне, варю обед и приглядываю за своей Анькой…
Все обсудили. Она сказала правильную вещь — «Бесплодие неясного генеза — это не значит, что причины нет. Это значит что ее пока не нашли. Будем искать». Протокол, 11 клеточек, 8 эмбрионов, 5 с хорошей морфологией. А дальше понятно — отбор эмбрионов по полу, и с первой попытки успешная беременность. И вот я, 39-летняя «молодая мамаша», сейчас сижу на кухне, варю обед и приглядываю за своей Анькой…
Анна Владиславовна, слов нет, чтобы выразить, как я Вам благодарна. А всем девчонкам совет — не затягивайте. А то времени может и не хватить.
Фрагментация
Для таких слож-ных случаев при работе с библиотекой NBS (Национальное бюро стандартов США) можно применять разработанный Мак-Лафферти метод вероятностного поиска и сравнения (Probability Based Matching, РВМ) (рис. 14).[ …]
Фрагментация (ландшафта) (fragmoitation (of landscape). Степень мозаичности территории, число различимых классов, расчленение единой территории линейными структурами, например дорогами.
Фрагментация алкиланилинов аналогична распаду алкил-бензолов под действием электронного удара и заключается в обычном р-разрыве боковой алкильной цепочки.[ …]
ФРАГМЕНТАЦИЯ (ф.) — процесс расчленения единой экосистемы на изолированные участки, как правило, в результате хозяйственной деятельности человека. Примеры Ф. — небольшие пятна естественных экосистем, степных или лесных, в массиве пашни, полянки луговой растительности в лесу. Ф. превращает единые экосистемы в «острова» со своим биологическим разнообразием (см. Островная биогеография). На сохранение биоразнообразия «островов» влияет и краевой эффект, так как внешняя часть «острова» по своим условиям отличается от внутренней и на ней формируются особые типы сообществ экотонов, например, лесных опушек, в которых представлены виды лесов, лугов и опушечные виды растений и их спутников — насекомых.
Фрагментация мест обитания угрожает сущестюванию видов и более сложным образом. Прежде всего, фрагментация ограничивает возможности видов к расселению. Многие виды птиц, млекопитающих и насекомых, обитающие в глубине леса, не могут пересечь даже узкие полосы открытого пространства из-за опасности попасться хищнику. В результате некоторые виды после исчезновения популяции во фрагменте не имеют возможности заселить его вновь. Более того, если из-за фрагментации исчезают животные, ответственные за распространение мясистых и липких плодов, то страдают и соответствующие виды растений. В конечном итоге изолированные фрагменты местообитаний не заселяются многими исходно характерными для них видами. А поскольку внутри отдельных фрагментов происходит естественное исчезновение видов из-за закономерных сукцессионных и популяционных процессов, а новые виды из-за барьеров не могут пополнить их убыль, следовательно, во фрагменте происходит постепенное видовое обеднение.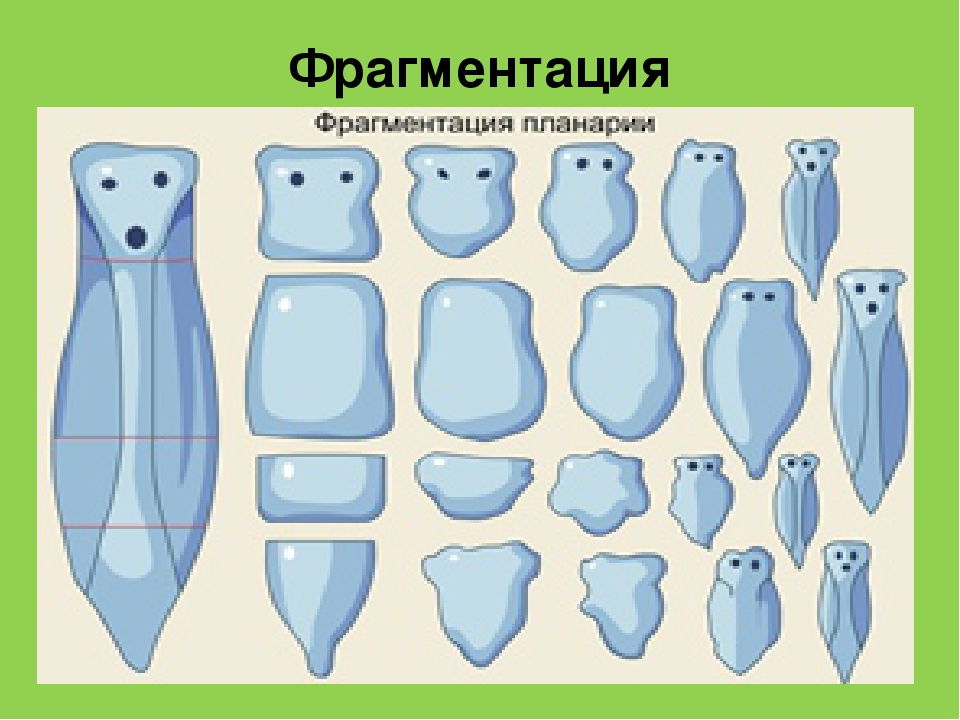 [ …]
[ …]
Фрагментация п -оксибензальдегида начинается с отрыва атома водорода (путь Б) при карбонильной группе и продолжается отщеплением СО(т = 71,5; 45,5) и С2Н2 (т =23,4).[ …]
Фрагментация (ландшафта) (fragmentation (of landscape). Степень мозаичности территории, число различимых классов, расчленение единой территории линейными структурами, например дорогами.[ …]
При фрагментации ванилина наблюдались процессы расщепления, протекающие и путем А, и путем Б. В отличие от п -оксибензаль— дегида в данном случае в схему расщепления путем Б входят и такие реакции, при которых отщепляется СН20.[ …]
Считается, что фрагментация муравьиной кислоты с образованием устойчивых продуктов (Н2 и С02) не играет значительной роли. В условиях нижней атмосферы сильную конкуренцию прямому фотоокислению создают процессы с участием реакционноспособных кислородсодержащих частиц.[ …]
Как показано выпе, фрагментация мест обитания сильно увеличивает долю пограничных местообитаний по отношению к внутренним (рис.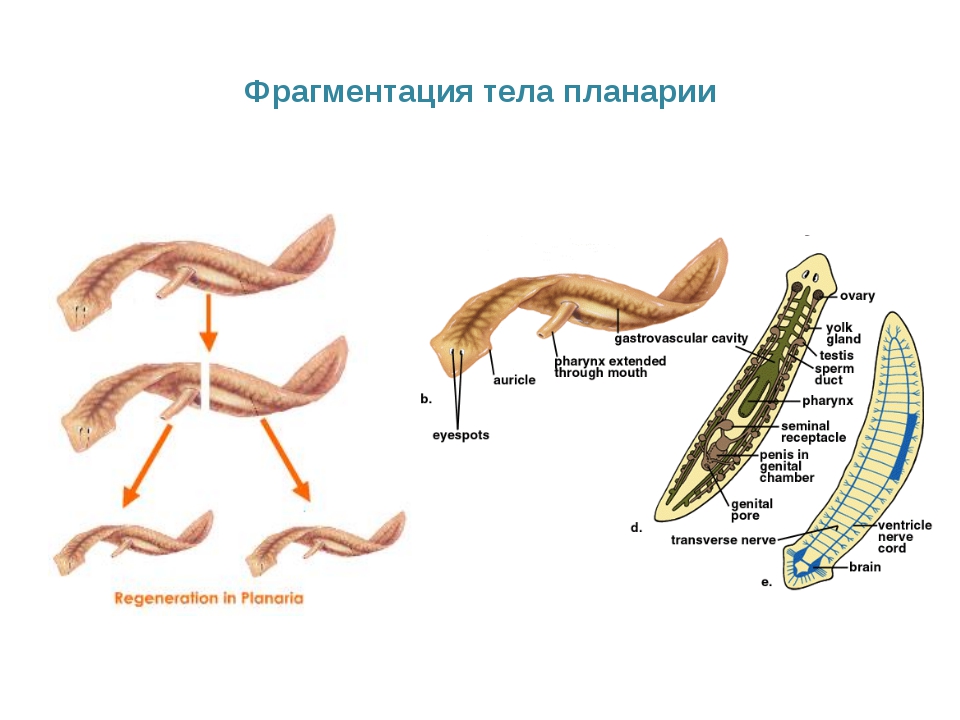 2.8).[ …]
2.8).[ …]
Аналогичные схемы фрагментации Ковачик и сотр. /112,113/ предложили для всех вышеперечисленных исследованных веществ.[ …]
Чтобы предотвратить фрагментацию или агрегацию вируса в ходе реакции, необходимо тщательно контролировать такие факторы, как тепемпера-тура, pH и величина ионной силы.[ …]
Углубление процесса фрагментации ареала — дальнейшее сокращение площади ареала; — снижение успешности размножения; — нарушение половой, возрастной и социальной структуры популяций; — разрушение межпопуляционных связей; — возрастание возможностей межподвидовой гибридизации; — общее ослабление жизненности популций.[ …]
Структура и площадь ареала фрагментация ареала под влиянием антропогенных факторов; стабильное сокращение общей площади ареала; реликтовый характер ареала; — ареал эндемичного вида (узкоареальный вид).[ …]
Рост размеров коацерватов и их фрагментация (деление), возможно, вели к образованию одинаковых коацерватов, и таким образом процесс мог продолжаться. Описанная последовательность событий должна была привести к возникновению примитивного гетеротрофного организма, питающегося органическими веществами первичного бульона.[ …]
Описанная последовательность событий должна была привести к возникновению примитивного гетеротрофного организма, питающегося органическими веществами первичного бульона.[ …]
Несколько иной подход к анализу фрагментации обычно известен как спектрометрия кинетических энергий ионов. Для проведения такого вида анализа применяют масс-спектрометр с двойной фокусировкой. При этом используют ионы Ам+, образующиеся в бесполевой области между электрическим анализатором и секторным магнитом [475]. В приборах этого типа возможно использование одного электрического сектора для анализа по энергиям. Легкость калибровки энергетической шкалы и отсутствие эффекта гистерезиса при легко контролируемом изменении поля позволяют избежать наложений сигналов нормальных ионов. Использование прибора с двойной фокусировкой делает возможным при анализе выделить какую-то определенную реакцию, индуцированную столкновениями. Располагая камеру столкновений между двумя анализаторами, получают возможность выделить какой-либо определенный ион для реакции и разделить образовавшиеся в результате столкновений ионы во втором анализаторе. Разные методы согласованного сканирования электрического и магнитного полей позволяют наблюдать продукты метастабильных или индуцированных столкновениями процессов без наложения сигналов «нормальных» ионов.[ …]
Разные методы согласованного сканирования электрического и магнитного полей позволяют наблюдать продукты метастабильных или индуцированных столкновениями процессов без наложения сигналов «нормальных» ионов.[ …]
Несколько иной подход к анализу фрагментации обычно известен как спектрометрия кинетических энергий ионов. Для проведения такого вида анализа применяют масс-спектрометр с двойной фокусировкой. При этом используют ионы Ам+, образующиеся в бесполевой области между электрическим анализатором и секторным магнитом [475]. В приборах этого типа возможно использование одного электрического сектора для анализа по энергиям. Легкость калибровки энергетической шкалы и отсутствие эффекта гистерезиса при легко контролируемом изменении поля позволяют избежать наложений сигналов нормальных ионов. Использование прибора с двойной фокусировкой делает возможным при анализе выделить какую-то определенную реакцию, индуцированную столкновениями. Располагая камеру столкновений между двумя анализаторами, получают возможность выделить какой-либо определенный ион для реакции и разделить образовавшиеся в результате столкновений ионы во втором анализаторе.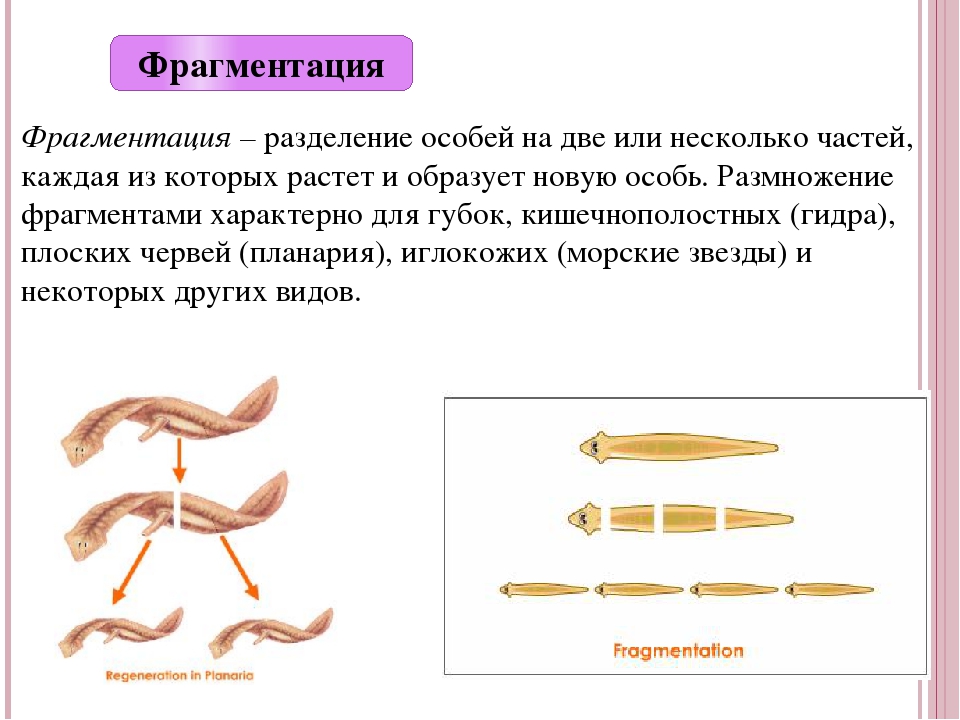 Разные методы согласованного сканирования электрического и магнитного полей позволяют наблюдать продукты метастабильных или индуцированных столкновениями процессов без наложения сигналов «нормальных» ионов.[ …]
Разные методы согласованного сканирования электрического и магнитного полей позволяют наблюдать продукты метастабильных или индуцированных столкновениями процессов без наложения сигналов «нормальных» ионов.[ …]
У многоклеточных животных вегетативное размножение происходит путем фрагментации их тела на части, после чего каждая часть развивается в новое животное. Такое размножение характерно для губок, кишечнополостных (гидр), немертин, плоских червей, иглокожих (морских звезд) и некоторых других организмов. Близкой формой к вегетативному размножений животных фрагментацией является полиэмбриония животных, заключающаяся в том, что на определенной стадии развития эмбрион разделяется на несколько частей, каждая из которых развивается в самостоятельный организм. Полиэмбриония встречается у броненосцев. Однако последние размножаются половым путем. Поэтому полиэмбриония является скорее своеобразной стадией в половом размножении, а потомство, возникающее в результате полиэмбрионии, представлено монозиготными близнецами. [ …]
[ …]
Новые пиреноиды могут возникать тремя различными способами: делением надвое, фрагментацией крупного родительского пиреноида на большое число мелких дочерних и в результате новообразования. Причем в первом и во втором случаях пиреноид не всегда распадается на равновеликие части, что является еще одной причиной наличия в клетке пиреноидов разного размера.[ …]
Трансформация и разрушение необходимых местообитаний — сокращение площади и дальнейшая фрагментация ареала; — сокращение репродуктивного потенциала; — сокращение или утрата кормовой базы; — увеличение пресса хищников; — общее сокращение численности вида; — нарушение структуры популяций — прямое негативное воздействие интродуцентов.[ …]
В последние годы широко изучали масс-спектрометрию убихинонов [43, 44]. Главной особенностью фрагментации молекулы убихинонов явилось наличие интенсивного пика с т/е 235, отвечающего пириллиевому иону (XXXVI), свойственному всем убихинонам. Между этим пиком и молекулярным ионом проявляются только пики низкой интенсивности, ответственные за ритмичный распад изо-преноидной цепи. [ …]
[ …]
Вследствие кислотно-основного катализа растворителя на первой стадии процесса происходят фрагментация и функциолизация структурных звеньев лигнина, что приводит к увеличению количества структур со свободным фенольным гидроксилом. Молекулы растворителя атакуют органический субстрат, облегчая его диссоциацию, и гидратируют богатые энергией и легко подвергающиеся электростатическому взаимодействию ионы. Кроме того, в результате кислотно-основного катализа растворителя при варьировании условий (концентрация оксида серы (IV), активность протонов, температура) изменяется ион-молекулярный состав сульфитных варочных растворов, который характеризуется концентрацией равновесных форм окси-соединений серы, различающихся по редокс-свойствам и также способных образовывать ОВС.[ …]
Вторая область является пограничной между зоной развития ПТС и отступающей первичной биосферой. Это область ее фрагментации и антропогенного экоцида, в которой при расширенном антропогенном воздействии уничтожаются первичные (гомеостатические) экосистемы, главным образом в процессе расширенных изъятий исчерпаемых биосферных ресурсов: лесных ресурсов, освоений земельных ресурсов и бесконтрольной экспансии рекреационного воздействия. Здесь происходит фронтальная деградация первичной биосферы с замещением первичных экосистем вторичными и последующими экосистемами в разных стадиях незавершенных циклов сукцессии на опустыненных пространствах после вырубок, неоднократных антропогенных выжиганий, на остатках военных баз и лагерных пунктов.[ …]
Здесь происходит фронтальная деградация первичной биосферы с замещением первичных экосистем вторичными и последующими экосистемами в разных стадиях незавершенных циклов сукцессии на опустыненных пространствах после вырубок, неоднократных антропогенных выжиганий, на остатках военных баз и лагерных пунктов.[ …]
Таким образом, передача зарядов осуществляется ион-кристалличес-кими ассоциатами воды как в виде пакетов зарядов при их фрагментации на электродах системы, так и при замыкании электродов стабилизированными ассоциатами.[ …]
В южной части Восточно-Сибирского региона техносферный метаболизм в биосфере имеет очагово-линейное распространение и создает область фрагментации в полосе от Присаянья с выходом на правобережье р. Ангары шириной от 30 до 100 км, вытянутую на северо-запад вдоль Транссибирской магистрали. В ней деструктивное техносферное воздействие вызывает обратные связи в виде материальных следствий биотического характера — истощения биосферных ресурсов и воздействий абиотических погодно-климатического и физико-географического факторов, постепенно усиливающих отрицательное влияние на физическую устойчивость техносферы и вызывающих неблагоприятные изменения в искусственной СО населяющего социума. [ …]
[ …]
Программное обеспечение состоит из двух приложений: SX Receiver и ScanViewer. Оно обеспечивает полностью автоматический прием, просмотр полученных изображений и их предварительную обработку — как общую (привязку, фрагментацию и т.д.), так и специфическую для AVHRR.[ …]
Еще один класс инвазивных видов — это те виды, которые расширили свой ареал в континентальных областях потому, что им оказались выгодны вызванные человеком нарушения окружающей среды. В Северной Америке из-за фрагментации лесов, развития системы пригородов и появления легко доступных пищевых отходов увеличились численность и область распространения койотов, рыжих лисиц и серебристых чаек. Эти агрессивные виды теснят местные, менее конкурентоспособные, отступающие перед хищниками виды. Поэтому очередной задачей в деле сохранения редких видов и охраняемых территорий становится управление местными видами, которые прекрасно адаптировались к деятельности человека и достигли необычно высокой численности. Другой особый класс инвазивных видов — это те, которые имеют близких родственников среди местной биоты.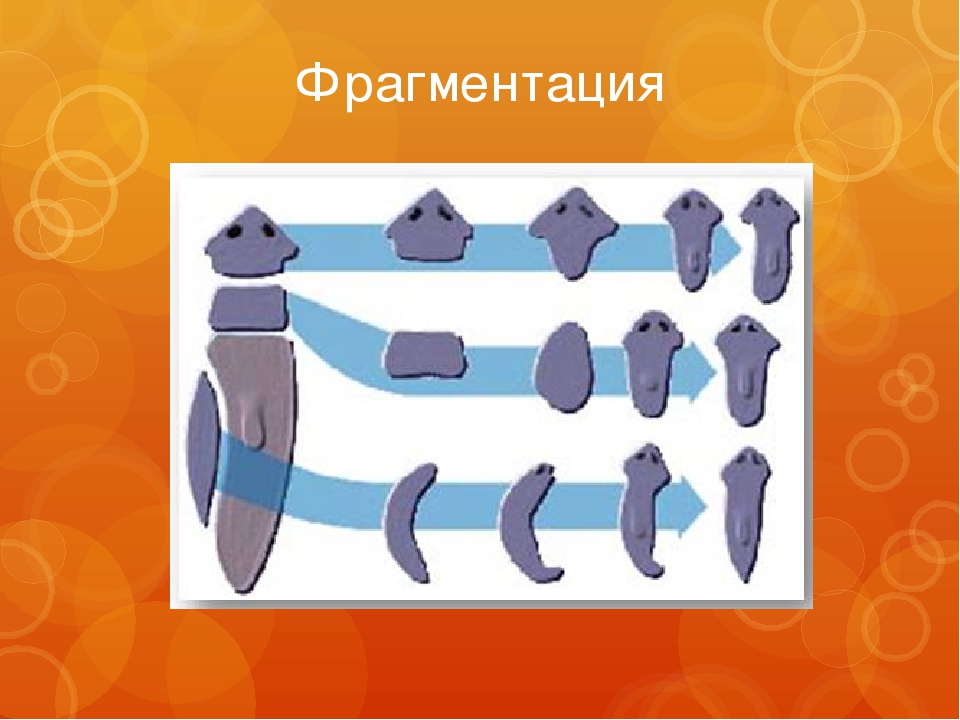 Когда такие экзотические виды скрещиваются с местными видами и подвидами, из местных популяций могут исчезнуть уникальные генотипы, а таксономические границы окажутся размытыми. Похоже такая судьба постигнет некоторые локальные виды лососей, столкнувшихся с промышленными видами. На юго-западе Америки из-за разрушения местообитаний и конкуренции с интродуцированными видами рыб сократился ареал лосося Апачей (Oncorhynchus apaché). Помимо этого, вид еще и широко скрещивался с радужной форелью (О. mykiss), интродуцированной для спортивной рыбалки [Dowling, Childs, 1992].[ …]
Когда такие экзотические виды скрещиваются с местными видами и подвидами, из местных популяций могут исчезнуть уникальные генотипы, а таксономические границы окажутся размытыми. Похоже такая судьба постигнет некоторые локальные виды лососей, столкнувшихся с промышленными видами. На юго-западе Америки из-за разрушения местообитаний и конкуренции с интродуцированными видами рыб сократился ареал лосося Апачей (Oncorhynchus apaché). Помимо этого, вид еще и широко скрещивался с радужной форелью (О. mykiss), интродуцированной для спортивной рыбалки [Dowling, Childs, 1992].[ …]
Ароматические эфиры. Ароматические эфиры дают в масс-спектрах интенсивные пики молекулярных ионов, которые нередко являются максимальными. Наиболее характерным путем распада незамещенных эфиров является разрыв С—О-связи. В случае замещенных ароматических эфиров альтернативные пути фрагментации аналогичны диссоциации алкилбензолов. Характерными опорными пиками в масс-спектрах ароматических эфиров может являться пик молекулярного иона, а также пики фрагментов с т/е=77 и 91. [ …]
[ …]
Практически неограниченная возможность любых контактов и обмена информацией имеет огромное социально-экономическое значение. Питер Хьюбер, автор статьи под названием «Чем больше у людей компьютеров, тем свободнее общество» по этому поводу пишет: «Политические и культурные последствия этого поистине огромны. Фрагментация и децентрализация возвестили конец монополии — будь то монополия западного капитализма или восточного коммунизма. Децентрализация означает наступление конкуренции и свободы» [82 .[ …]
Приборы с ионизацией полем имеют чувствительность на порядок ниже чем, например, электроноударные источники, однако с их помощью можно регистрировать пики молекулярных ионов лабильных соединений. В этом смысле можно сравнить спектры полевой ионизации с низкоэнергетическими электроноударными спектрами. Отсутствие части информации о фрагментации целиком восполняется преимуществом получения пика молекулярного иона почти для всех соединений. К недостаткам метода следует отнести его несколько пониженную чувствительность по сравнению с таковой при химической ионизации и электронном ударе.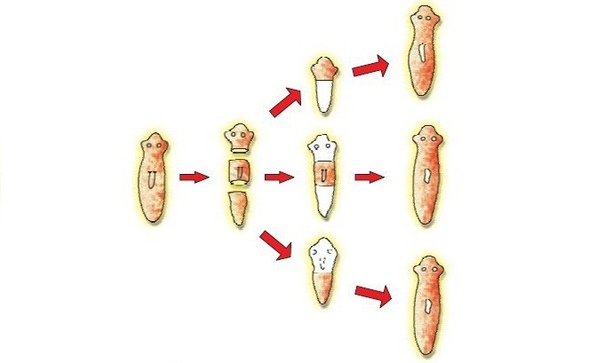 Ионизация полем в комбинации ГХ — МС [438] позволяет получать рабочий спектр при анализе 100 нг чистого компонента в смеси.[ …]
Ионизация полем в комбинации ГХ — МС [438] позволяет получать рабочий спектр при анализе 100 нг чистого компонента в смеси.[ …]
Начальная стадия формирования локальной авифауны совпадает с проведением мероприятий по землеустройству и началу строительных работ — разметкой участков, возведением заборов, складированием стройматериалов и началом мозаичной застройки. В этих условиях видовое разнообразие птиц, населяющих локальный участок ландшафта, заметно возрастает как за счет фрагментации ранее существовавших биотопов, так и за счет появления новых видов птиц, «втягивающихся» из соседних биотопов. На территории вновь создаваемых поселков, расположенных вблизи леса, увеличивается представительство птиц древесного комплекса-большой синицы, мухоловки-пеструшки, зарянки, славки-завирушки, чечевицы, овсянки обыкновенной. Для дачных участков, окруженных полями, увеличивается численность и разнообразие птиц открытого пространства: лугового чекана, серой славки, реже — перепела, полевого жаворонка, коростеля.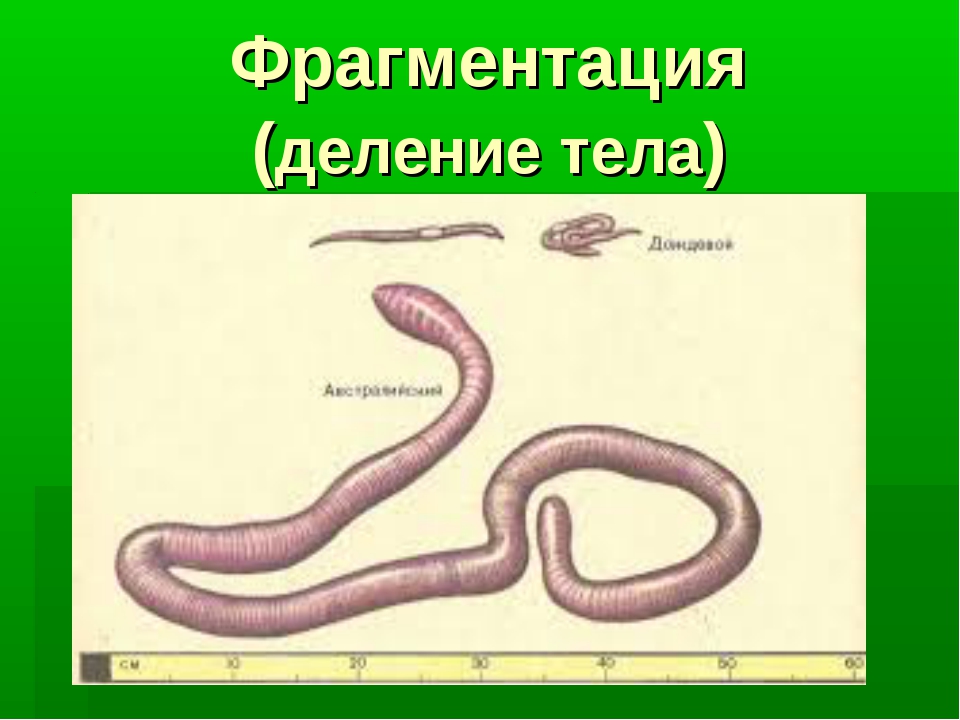 Застраиваемые территории привлекают на гнездование и постоянных спутников человека — птиц-синантропов (скворца, галку, сизого голубя и др.). Появление новых поселений человека в целом способствует возрастанию численности и некоторых видов, размножающихся на сопредельных территориях: так, в древостоях, подступающих к трансформированным территориям, заметно увеличивается плотность гнездования врановых, ушастой совы и неясыти, более частыми визитерами становятся ворон, большой пестрый дятел, кукушка и др. Таким образом, по разнообразию птиц вновь создаваемые садово-дачные поселки заметно отличаются от традиционных поселений человека — сел и деревень.[ …]
Застраиваемые территории привлекают на гнездование и постоянных спутников человека — птиц-синантропов (скворца, галку, сизого голубя и др.). Появление новых поселений человека в целом способствует возрастанию численности и некоторых видов, размножающихся на сопредельных территориях: так, в древостоях, подступающих к трансформированным территориям, заметно увеличивается плотность гнездования врановых, ушастой совы и неясыти, более частыми визитерами становятся ворон, большой пестрый дятел, кукушка и др. Таким образом, по разнообразию птиц вновь создаваемые садово-дачные поселки заметно отличаются от традиционных поселений человека — сел и деревень.[ …]
Анализ популяционной жизнеспособности (АПЖ) — раздел демографического анализа, направленный на понимание того, насколько данный вид способен сохраниться в окружающей среде [Ruggiero et al., 1994; Akcakaya et al, 1999]. АПЖ определяет потребности вида и ресурсы, присутствующие в окружающей его среде, для выявления уязвимых моментов в его естественной истории. АПЖ полезен для понимания последствий фрагментации или деградации местообитания редкого вида. Хотя АПЖ до сих пор развивается как подход для прогноза устойчивости видов, он не является стандартным методом в рамках статистических исследований [Lacy, Lindemayer, 1995], применяемые для него методы систематического и последовательного сбора данных о видах являются логическим продолжением изучения естественной истории и демографических исследований. Попытки использования статистики для прогноза будущих тенденций в размерах популяции нужно делать с осторожностью, вместе с большой долей здравого смысла, поскольку условия могут измениться и меры по сохранению могут оказаться как успешными, таки неэффективными.[ …]
АПЖ полезен для понимания последствий фрагментации или деградации местообитания редкого вида. Хотя АПЖ до сих пор развивается как подход для прогноза устойчивости видов, он не является стандартным методом в рамках статистических исследований [Lacy, Lindemayer, 1995], применяемые для него методы систематического и последовательного сбора данных о видах являются логическим продолжением изучения естественной истории и демографических исследований. Попытки использования статистики для прогноза будущих тенденций в размерах популяции нужно делать с осторожностью, вместе с большой долей здравого смысла, поскольку условия могут измениться и меры по сохранению могут оказаться как успешными, таки неэффективными.[ …]
Амитозом называется деление клетки, находящейся в состоянии интерфазы. К амитозу иногда относят все случаи немитотического деления клетки (рис. 66). При этом не происходит конденсации хромосом, распада ядерной оболочки и образования веретена деления; амитоз осуществляется при вытягивании ядра и его последующем делении на две части. Еще более неупорядоченное дробление ядра на два или более неидентичных комка получило название фрагментации; оно, безусловно, носит патологический характер. Однако между амитозом и фрагментацией резкой и принципиальной границы провести нельзя.[ …]
Еще более неупорядоченное дробление ядра на два или более неидентичных комка получило название фрагментации; оно, безусловно, носит патологический характер. Однако между амитозом и фрагментацией резкой и принципиальной границы провести нельзя.[ …]
Подводя итоги вышеизложенному, следует отметить, что объем информации, которую можно получить, меняя условия работы различных узлов масс-спектрометра, невозможно описать в ограниченном объеме настоящей работы. Система ввода образца может быть использована для селективного выделения компонентов смесей в газовой фазе или для пиролиза больших молекул при программировании температуры. Соответствующие способы ионизации обеспечивают получение интенсивных пиков молекулярных ионов, интенсивной фрагментации исходных ионов, многозарядных ионов. Масс-анализатор» позволяет получать масс-спектры по отношению массы к заряду, энергии к заряду, анализировать положительные, отрицательные и метастабильные ионы. Масс-спектрометр может содержать камеру столкновений, в которой выбранные ионы подвергаются фрагментации с целью изучения их структуры, к т. д. Все виды полученной информации дополняют друг друга при определении структуры органических соединений и идентификации компонентов сложных смесей.[ …]
д. Все виды полученной информации дополняют друг друга при определении структуры органических соединений и идентификации компонентов сложных смесей.[ …]
Нафтеновые углеводороды. Повышенная интенсивность пиков молекулярных ионов является характерным отличием масс-спектров нафтеновых углеводородов от спектров парафиновых углеводородов, содержащих такое же число углеродных атомов. Фрагментация осуществляется путем разрыва С—С-связей. Интенсивность пиков молекулярных ионов нафтеновых углеводородов зависит от строения молекулы. Для циклогексанов она несколько выше, чем для циклопентанов. Кроме того, наличие боковых цепей приводит к снижению интенсивности пиков молекулярных ионов в масс-спектрах соединений с насыщенной циклической структурой. Наличие разветвления в боковых цепях еще более понижает интенсивность пика молекулярного иона и приводит к появлению в спектрах пиков ионов, соответствующих разрыву в месте присоединения заместителя. К этому можно добавить, что пики ионов т/е=41; 55; 69; 83, имея значительную интенсивность, присутствуют во всех масс-спектрах нафтеновых углеводородов, и сумма их интенсивностей служит характеристикой данного класса соединений при групповом анализе. [ …]
[ …]
Многие национальные парки и другие охраняемые территории были созданы для охраны “харизматической мегафауны”, в которой такие животные, как львы, тигры, медведи, являются национальными символами и привлекают туристов. Однако если территории, на которых обитают эти виды в составе определенного сообщества, выделить в качестве охраняемых чисто формально, это может не остановить их исчезновение. Часто охраняемые территории создаются лишь тогда, когда большинство популяций, находящихся под угрозой видов, уже сильно сократилось из-за деградации и уничтожения местообитаний, их фрагментации и чрезмерной эксплуатации. В таких обстоятельствах численность видов продолжает сокращаться, вплоть до их исчезновения. Кроме того, за границами охраняемой тфритории животные остаются незащищенными и подвергаются риску. Для того, чтобы создать полноценную программу сохранения, способную защитить и даже восстановить находящиеся в опасности виды, жизненно необходимо понять состояние их популяций в дикой природе, выявить, как реагируют эти популяции на различные условия, знать естественную историю и экологию этих видов. В этой главе показано, как для защиты и восстановления видов биологи используют подход на популяционном уровне.[ …]
В этой главе показано, как для защиты и восстановления видов биологи используют подход на популяционном уровне.[ …]
Характер связи с местообитаниями. Реакция вида на изменение эволюционно обусловленных местообитаний, как и многие другие биологические параметры, неоднозначна и варьирует в сравнительно широких пределах. Для одних видов изменение или полное разрушение типичных естественных (исходных) местообитаний под воздействием антропогенных или иных факторов влечет за собой потерю кормовой базы, мест и условий размножения, обострение межвидовых отношений, изменение других необходимых для существования экологических факторов. Следствием этого является в первую очередь сокращение численности и фрагментация и/или сокращение площади ареала, означающие, как правило, начало процесса деградации вида. Для других видов утрата типичных местообитаний не влечет за собой таких катастрофических последствий, а в отдельных случаях создает дополнительные возможности для экспансии и расширения ареала.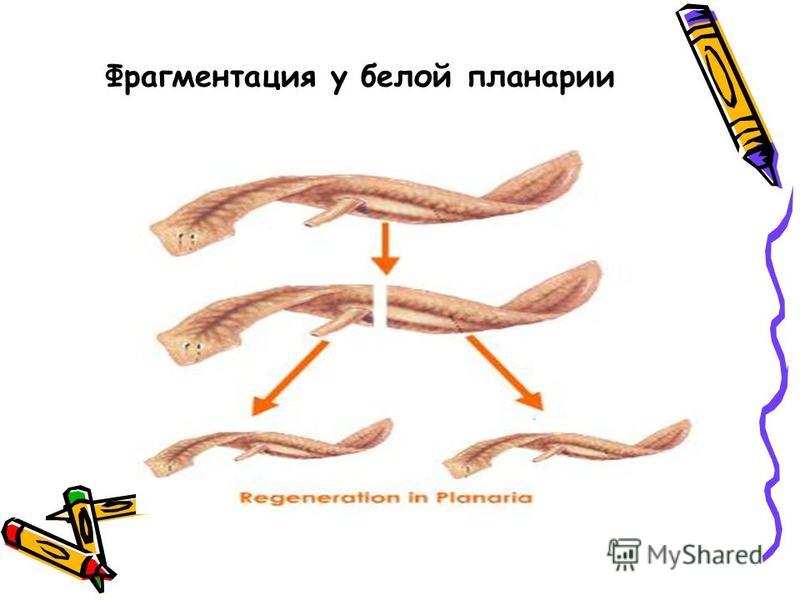 Принципиальные основы этого феномена рассмотрены в разделе, посвященном экологической специализации и экологической валентности видов. Здесь же следует лишь подчеркнуть, что утрата (разрушение) типичных местообитаний, вопреки широко бытующему представлению, далеко не всегда выступает как главный лимитирующий фактор. Это положение особенно важно учитывать при разработке проектов по восстановлению угасших или созданию новых популяций видов, находящихся на грани исчезновения. Будущее вида в этом случае зависит от степени его экологической специализации, а не от наличия местообитаний, абсолютно идентичных исчезнувшим.[ …]
Принципиальные основы этого феномена рассмотрены в разделе, посвященном экологической специализации и экологической валентности видов. Здесь же следует лишь подчеркнуть, что утрата (разрушение) типичных местообитаний, вопреки широко бытующему представлению, далеко не всегда выступает как главный лимитирующий фактор. Это положение особенно важно учитывать при разработке проектов по восстановлению угасших или созданию новых популяций видов, находящихся на грани исчезновения. Будущее вида в этом случае зависит от степени его экологической специализации, а не от наличия местообитаний, абсолютно идентичных исчезнувшим.[ …]
В свете рассмотренных выше данных, характеризующих кинетику сульфитной варки при температуре 90 С и pH [ …]
Территория любого города с ландшафтной точки зрения представляет собой сочетание большего или меньшего набора фаций, отличных своими условиями для городских птиц. Большая часть городских территорий застроена, в том числе селитебная часть, представленная одноэтажной и многоэтажной застройкой, обычно с высокой степенью асфальтового покрытия и однорядным озеленением улиц, а также территории промышленных объектов. Долю своеобразия вносят в городскую черту участки естественных местообитаний (река и пойменные участки, озера, болота, островки леса и пр.) или их аналоги (парки, полосы отчуждения вдоль железных дорог, лесополосы, пустыри). Количество и качество таких включений, а также степень фрагментации (или, наоборот, компактности) ландшафтных разностей в основном и определяют своеобразие «орнитофаунистичес-кого лица» города.[ …]
Долю своеобразия вносят в городскую черту участки естественных местообитаний (река и пойменные участки, озера, болота, островки леса и пр.) или их аналоги (парки, полосы отчуждения вдоль железных дорог, лесополосы, пустыри). Количество и качество таких включений, а также степень фрагментации (или, наоборот, компактности) ландшафтных разностей в основном и определяют своеобразие «орнитофаунистичес-кого лица» города.[ …]
Прежде всего, среди редких видов животных немало таких, которые уже не сохранились в природе или численность которых перешла низший допустимый предел, обеспечивающий все параметры стабильного существования. Среди таких видов нужно в первую очередь назвать лошадь Пржевальского, зубра, амурского леопарда, стерха, дрофу и ряд других видов, отнесенных к категории I Красной книги России. В мировом масштабе таких видов десятки и сотни. Второе обстоятельство: каждая охраняемая территория, будь то заповедник, заказник, резерват или национальный парк, это прежде всего «остров» среди плотно обступивших его антропогенных, экологически нетождественных и даже чуждых ландшафтов.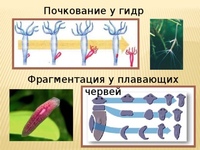 А остров — это как раз и есть результат фрагментации ареала, т.е. фрагмент ареала, где обязательно разрушаются нормальные экологические и генетические процессы. И наконец, третье обстоятельство, сущность которого заключается в том, что большинство редких видов животных относится к категории мигрирующих и часть жизненного цикла проводят не только вне охраняемых территорий, но даже за рубежами нашей страны. Длительные миграции и лишенные охраны зимовки — это один из самых опасных периодов в жизни таких мигрирующих видов, именно на него падает основная смертность в популяциях. Все это свидетельствует в пользу уже высказанной ранее мысли: стратегия сохранения редких видов должна быть поливекторной, комплексной, гармонично включать все необходимые стратегические элементы. Основная задача заповедников, заказников различного ранга и других ООПТ- сохранение экосистем, где редкие виды выступают в роли компонентов. Поэтому для этих видов заповедание территории -только более или менее дисперсная охрана.
А остров — это как раз и есть результат фрагментации ареала, т.е. фрагмент ареала, где обязательно разрушаются нормальные экологические и генетические процессы. И наконец, третье обстоятельство, сущность которого заключается в том, что большинство редких видов животных относится к категории мигрирующих и часть жизненного цикла проводят не только вне охраняемых территорий, но даже за рубежами нашей страны. Длительные миграции и лишенные охраны зимовки — это один из самых опасных периодов в жизни таких мигрирующих видов, именно на него падает основная смертность в популяциях. Все это свидетельствует в пользу уже высказанной ранее мысли: стратегия сохранения редких видов должна быть поливекторной, комплексной, гармонично включать все необходимые стратегические элементы. Основная задача заповедников, заказников различного ранга и других ООПТ- сохранение экосистем, где редкие виды выступают в роли компонентов. Поэтому для этих видов заповедание территории -только более или менее дисперсная охрана. Важная, но не всеохватывающая функция.[ …]
Важная, но не всеохватывающая функция.[ …]
Несмотря на то, что в органической масс-спектрометрии до сих пор не существует единого мнения по вопросу образования фрагментов при действии электронного удара, в ряде случаев при интерпретации масс-спектров удобно считать, что распад молекулярного иона с большей вероятностью может протекать с разрывом связей, находящихся вблизи от вероятного места локализации положительного заряда, например, на каком-либо атоме, имеющем я- или р-электроны, способные покидать молекулу при возбуждении. Однако следует иметь в виду, что при распаде молекулярного иона совершенно не исключается возможность разрыва и других связей. Кроме того, не всегда можно установить даже предполагаемое место локализации заряда. Следует иметь в виду, что схематическое изображение путей фрагментации с помощью фрагментов, представленных в виде структурных формул, является весьма условным, однако эти схемы наглядны и облегчают установление строения соединений по его масс-спектру. В схемах распада обычная стрелка обозначает перенос электронной пары, а «крючок» означает перенос одного электрона или атома с одним неспаренным электроном.[ …]
В схемах распада обычная стрелка обозначает перенос электронной пары, а «крючок» означает перенос одного электрона или атома с одним неспаренным электроном.[ …]
Полевая ионизация. Ионизация полем осуществляется путем удаления электрона из молекулы под действием электрического поля с высоким потенциалом порядка 8-103 В, которое создается у поверхностей с максимальной кривизной (острие, лезвие, тонкая проволока диаметром 10 мкм). В этом случае под действием значительного градиента потенциала (107— 108 В/см) происходит деформация атомного потенциального ■барьера, что приводит к квантовому туннельному переходу валентного электрона молекулы к положительно заряженному электроду. В анализатор поступают положительные ионы, выталкиваемые и ускоряемые анодом. Молекулярный ион, образовавшийся под действием поля, обладает некоторой избыточной энергией, но поскольку она невелика, например, по сравнению с электронным ударом, то происходят в основном первичные процессы фрагментации [437].[ .. .]
.]
Второй подход является реальной теоретической основой создания эко-нетов, которые призваны обепсечивать сохранение биоразнообразия при достаточно интенсивном хозяйственном использовании территории. Еще одна функция эконета — поддержание экологического баланса крупных регионов за счет создания сети природных и природно-антро-погенных территорий различного функционального назначения (и с различными режимами использования). Таким образом, эконет — это оптимальный инструмент территориальной охраны природы. Исходя из указанных особенностей эконетов, они наиболее эффективны в условиях регионов интенсивного старого освоение, где естественные природные комплексы составляют незначительный процент площади и сильно изменены. В данном случае эконет позволяет до определенной степени снизить эффект фрагментации местообитаний и природных “островов”, что часто не требуется в малоосвоенных регионах, где сохраняются естетственные пространственные связи между ключевыми местообитаниями. В таких регионах создание экосетей может иметь превентивный характер для опережающего создания условий пространственного сохранения биоразнообразия до начала массового освоения и трансформации региона. [ …]
[ …]
Полученные результаты говорят о том, что между бурозубками из разных биотопов, удаленных друг от друга незначительно, существуют некоторые различия в морфологии черепа. Они едва улавливаются довольно чувствительным методом дискриминантного анализа, но вряд ли являются случайными, так как использовано большое число признаков. Возможно, что эти различия вызваны не разными условиями обитания, а колебаниями численности обыкновенной бурозубки. В весенний период происходит массовая гибель особей. Резкое снижение численности может дать эффект «бутылочного горлышка», когда соотношения долей разных фенотипов в удаленных друг от друга биотопах будет неодинаковым. Данное утверждение требует дальнейшей проработки на более массовом материале, особенно из предгорного района Печоро-Илычского заповедника, где фрагментация рельефа выше и подобная картина может проявиться более четко.[ …]
Изолированные местообитания более подвержены инвазии как экзотических видов, так и паразитов исходных видов. Край леса -это нарушенная среда, в ней могут легко без конкуренции обосноваться паразитические виды, увеличиться в численности и затем заселить внутреннюю часть фрагмента Т акие всеядные животные, как еноты, скунсы, голубые сойки могут расплодиться вдоль опушки леса, одинаково успешно эксплуатируя в поисках корма как первичные, так и нарушенные местообитания. Эти агрессивные виды поедают яйца и разоряют гнезда лесных птиц, часто препятствуя успешному воспроизводству многих видов на сотни метров от ближайшей опушки леса. Гнездовой паразит воловья птица Molothrus ater), обитающий в полях и в пограничных местообитаниях, использует последние как стартовую позицию для инвазии внутрь леса, где их птенцы уничтожают яйца и птенцов лесных певчих птиц. Сочетание фрагментации мест обитания, усиление пресса хищников, уничтожающих гнезда и разрушения тропических мест зимовки, возможно, стало причиной катастрофического спада численности некоторых видов мигрирующих певчих птиц Северной Америки, таких как голубой певун (Dendroica cerúlea) на востоке США.
Край леса -это нарушенная среда, в ней могут легко без конкуренции обосноваться паразитические виды, увеличиться в численности и затем заселить внутреннюю часть фрагмента Т акие всеядные животные, как еноты, скунсы, голубые сойки могут расплодиться вдоль опушки леса, одинаково успешно эксплуатируя в поисках корма как первичные, так и нарушенные местообитания. Эти агрессивные виды поедают яйца и разоряют гнезда лесных птиц, часто препятствуя успешному воспроизводству многих видов на сотни метров от ближайшей опушки леса. Гнездовой паразит воловья птица Molothrus ater), обитающий в полях и в пограничных местообитаниях, использует последние как стартовую позицию для инвазии внутрь леса, где их птенцы уничтожают яйца и птенцов лесных певчих птиц. Сочетание фрагментации мест обитания, усиление пресса хищников, уничтожающих гнезда и разрушения тропических мест зимовки, возможно, стало причиной катастрофического спада численности некоторых видов мигрирующих певчих птиц Северной Америки, таких как голубой певун (Dendroica cerúlea) на востоке США. Некоторые виды птиц в Северной Америке и в Европе в ответ на изменяющиеся схемы землепользования (сельскохозяйственная или лесохозяйственная деятельность) на конкретном региональном уровне могут либо увеличивать, либо уменьшать свою численность (James, et al., 1996).[ …]
Некоторые виды птиц в Северной Америке и в Европе в ответ на изменяющиеся схемы землепользования (сельскохозяйственная или лесохозяйственная деятельность) на конкретном региональном уровне могут либо увеличивать, либо уменьшать свою численность (James, et al., 1996).[ …]
Также представлены гипоаркто-монтанный (9) и не моральный (Anaptichia ciliaris, Lobaria pulmonaria, Physcia adscendens, Physconia perisidiosa) элементы. Оченырд, а участие бореальных видов уменьшается с 41 до 28%. Среди субстратных групп преобладают эпигейные лишайники (101 вид). Далее стоят эпилиты (40) и эпифиты (27). Отмечено четыре вида эпиксильных лишайников (Cladonia botrytes, С. carneóla, С. cenotea, С. digitata). При продвижении вдоль широтного градиента на север увеличивается доля видов размножающихся фрагментацией таллома, с 36 до 44-45%, и уменьшается процент лишайников, репродуцирующихся посредством соредий или изидий, с 28 до 18%.[ …]
Идентификация отдельных компонентов сложной матрицы включает предварительное отнесение неизвестных веществ к тому или иному классу соединений. Для этой цели используют пики молекулярных и основных осколочных ионов. В ряде случаев в дополнение к характеристическим пикам масс-спектров целесообразно рассматривать массы нейтральных фрагментов, образующихся из молекулярных ионов. Наиболее информативным для установления класса соединения и его идентификации является пик молекулярного иона, характерным признаком которого является наибольшее значение массового числа в «чистом» масс-спектре. Помимо этого, разность между значениями массового числа пика молекулярного иона и первичных осколочных ионов не может быть равной числам от 4 до 14. Если в масс-спектре отсутствует пик молекулярного иона, то при отнесении неизвестного соединения к тому или иному классу следует рассматривать наиболее интенсивные характеристические пики осколочных ионов, соответствующих наиболее характерным путям фрагментации молекулы.[ …]
Для этой цели используют пики молекулярных и основных осколочных ионов. В ряде случаев в дополнение к характеристическим пикам масс-спектров целесообразно рассматривать массы нейтральных фрагментов, образующихся из молекулярных ионов. Наиболее информативным для установления класса соединения и его идентификации является пик молекулярного иона, характерным признаком которого является наибольшее значение массового числа в «чистом» масс-спектре. Помимо этого, разность между значениями массового числа пика молекулярного иона и первичных осколочных ионов не может быть равной числам от 4 до 14. Если в масс-спектре отсутствует пик молекулярного иона, то при отнесении неизвестного соединения к тому или иному классу следует рассматривать наиболее интенсивные характеристические пики осколочных ионов, соответствующих наиболее характерным путям фрагментации молекулы.[ …]
Программа ООН по окружающей среде (ЮНЕП) выделила четыре важные причины потери биоразнообразия, связанные с хозяйственной деятельностью человека. Это, во-первых, нынешняя неэффективная структура землепользования: примерно 1-2% поверхности суши подвержено урбанизации и изъято из природной активности, около 25% — занято сельскохозяйственной деятельностью. В результате примерно 40% первичной продукции биоты Земли изъято деятельностью человека. Эти экономические процессы связаны как с ростом народонаселения (и нагрузки на природу), так и с формами собственности на землю и другие природные ресурсы. Вырубка тропических лесов в 80-е годы XX веке в Южной Америке, Африке и Юго-Восточной Азии способствовала снижению биоразнообразия. Это приводит к ускоренному исчезновению биологических видов темпами, которые уже сейчас в 50-100 раз превышают природные фоновые процессы и могут в ближайшее время достигнуть 10 000 раз. Вызывает озабоченность наблюдаемая в США тенденция к малым земельным участкам, что расчленяет и фрагментирует ландшафт, вызывает проблемы с местообитанием видов флоры и фауны. Такая же фрагментация происходит в Южной Америке.[ .
Это, во-первых, нынешняя неэффективная структура землепользования: примерно 1-2% поверхности суши подвержено урбанизации и изъято из природной активности, около 25% — занято сельскохозяйственной деятельностью. В результате примерно 40% первичной продукции биоты Земли изъято деятельностью человека. Эти экономические процессы связаны как с ростом народонаселения (и нагрузки на природу), так и с формами собственности на землю и другие природные ресурсы. Вырубка тропических лесов в 80-е годы XX веке в Южной Америке, Африке и Юго-Восточной Азии способствовала снижению биоразнообразия. Это приводит к ускоренному исчезновению биологических видов темпами, которые уже сейчас в 50-100 раз превышают природные фоновые процессы и могут в ближайшее время достигнуть 10 000 раз. Вызывает озабоченность наблюдаемая в США тенденция к малым земельным участкам, что расчленяет и фрагментирует ландшафт, вызывает проблемы с местообитанием видов флоры и фауны. Такая же фрагментация происходит в Южной Америке.[ . ..]
..]
Ученые запечатлели процесс рождения тройной звезды
B. Saxton / NRAO/AUI/NSF; ALMA (ESO/NAOJ/NRAO)
Астрономам удалось запечатлеть процесс рождения тройной звездной системы. Это наблюдение подтверждает гипотезу о том, что молодые кратные звезды могут формироваться в результате «разрыва» крупного протозвездного диска из-за гравитационной нестабильности. Об этом сообщает статья, опубликованная в журнале Nature.
Двойные и тройные (кратные) звездные системы довольно часто встречаются в нашей Вселенной — почти половина всех солнцеподобных звезд предположительно имеют компаньона. Ученые предполагают, что существует несколько механизмов, которые могут приводить к формированию многокомпонентных систем. Одна из теорий заключается в том, что двойные и тройные звезды появляются в результате гравитационной нестабильности в газопылевом диске вокруг протозвезды. Из-за этой неустойчивости происходит
фрагментация диска и образуется сгусток
материи, масса которой выше критической,
что впоследствии позволит ему стать
еще одной протозвездой. Однако это предположение до сих пор было основано на исследованиях тесно расположенных протозвезд, которые уже почти вышли на главную последовательность, то есть скоро станут полноценными звездами.
Ученые предполагают, что существует несколько механизмов, которые могут приводить к формированию многокомпонентных систем. Одна из теорий заключается в том, что двойные и тройные звезды появляются в результате гравитационной нестабильности в газопылевом диске вокруг протозвезды. Из-за этой неустойчивости происходит
фрагментация диска и образуется сгусток
материи, масса которой выше критической,
что впоследствии позволит ему стать
еще одной протозвездой. Однако это предположение до сих пор было основано на исследованиях тесно расположенных протозвезд, которые уже почти вышли на главную последовательность, то есть скоро станут полноценными звездами.
Авторам новой работы удалось заглянуть в саму звездную колыбель, где протозвезды находятся на более раннем этапе эволюции. С помощью телескопа ALMA астрономы изучили процессы в системе L1448 IRS3B, которая находится на расстоянии 750 световых лет от Земли в созвездии Персея. Ее возраст предположительно не превышает 150 тысяч лет, что по космическим меркам очень мало — для сравнения, Солнцу сейчас около 4,5 миллиардов лет.
Система L1448 IRS3B состоит из трех протозвезд. В ее центре находится протозвезда IRS3B-a, у которой есть компаньон, расположенный на расстоянии 61 астрономической единицы от нее (одна астрономическая единица равна среднему расстоянию между Землей и Солнцем). Еще одна протозвезда находится в отдалении от двух других в одном из спиральных рукавов газопылевого диска. Расстояние между ней и IRS3B-a достигает уже 183 астрономических единиц. Протозвезды центральной пары предположительно имеют такую же массу, как и наше Солнце, в то время как масса третьего, более далекого небесного тела оценивается в 0,085 солнечной массы.
Наблюдение ученых стало прямым доказательством того, что в результате фрагментации протозвездного диска из-за гравитационной нестабильности могут рождаться молодые кратные звездные системы. «Разрыв» в диске произошел совсем недавно: возраст самой далекой от IRS3B-a протозвезды, которая находится в одном из газопылевых рукавов, оценивается всего в 10-20 тысяч лет.
Недавно исследователям удалось впервые увидеть рукава в протопланетном диске вокруг очень молодой звезды. Их удалось обнаружить благодаря тепловому излучению частиц пыли, которые составляют от 1 до 10 процентов всего вещества в диске. По данным исследователей, длина газопылевых рукавов, отходящих от материнской звезды, достигает более десяти миллиардов километров, что почти в два раза больше среднего расстояния от Плутона до Солнца.
Кристина Уласович
Тестовая работа по теме «Размножение и развитие живых организмов», 10 класс
Тестовая работа по теме: «Размножение и индивидуальное развитие организмов».
( работа рассчитана на профильный уровень преподавания биологии в 10 классе при
3 часах в неделю)
1 вариант
Часть А: (выберите один правильный ответ из четырех возможных)
1. Способ бесполого размножения при котором дочерняя особь появляется из группы клеток, отделившихся от материнской особи.
Способ бесполого размножения при котором дочерняя особь появляется из группы клеток, отделившихся от материнской особи.
а) споруляция б) почкование
в) фрагментация г) клонирование
2. Какой из организмов размножается путем бинарного деления клетки.
а) земляника б) еж
в) инфузория г) гидра
3. Как называется способ полового размножения из неоплодотворенного яйца?
а) коньюгация б) почкование
в) фрагментация г) партеногенез
4. Как называется стадия интерфазы, где происходит синтез АТФ для мейоза?
а) синтетическая б) телофаза
в) постсинтетическая г) пресинтетическая
5. В какой фазе мейоза происходит расположение хромосом по экватору клетки в один ряд?
а) метафаза 2 б) телофаза 2
в) анафаза 2 г) метафаза 1
6. В какую фазу мейоза происходит кроссинговер?
а) профаза 2 б) профаза 1
в) анафаза 2 г) метафаза 1
7. Какой фазе мейоза соответствует процесс 2n4c → 1n2c ?
а) анафаза 1 б) телофаза 2
в) анафаза 2 г) метафаза 1
8. В телофазе 2 набор хромосом в дочерних клетках характеризуется формулой:
В телофазе 2 набор хромосом в дочерних клетках характеризуется формулой:
а) 2n2c б) 1n2c
в) 2n4c г) 1n1c
9. В результате мейоза у водорослей формируются:
а) гаметы б) споры
в) зигота г) гаметофит
10. В зоне созревания при развитии половых клеток происходит:
а) митоз б) рост
в) мейоз г) ничего не происходит
11. Двуслойный зародыш образуется на стадии:
а) нейруляции б) гаструляции
в) бластуляции г) органогенеза
12. Дыхательная система организма формируется из:
а) эктодермы б) энтодермы
в) зиготы г) мезодермы
13. Ограниченный рост характерен для:
а) берёзы б) щуки
в) улитки г) собаки
14. Спорофитом в жизненном цикле мхов является:
а) протонема б) взрослое растение
в) коробочка г) заросток
Часть В: Найдите неверные утверждения и опровергните их.
1. У семенных растений преобладающим поколением является спорофит.
2. Мужской гаметофит у цветковых растений представлен зародышевым мешком.
3. После трех митотических делений мегаспоры образуется двуядерный зародышевый мешок.
4. Первый спермий оплодотворяет яйцеклетку, а второй центральную клетку.
5. Зародыш и эндосперм цветковых растений имеют диплоидные наборы хромосом.
6. Рост растения в толщину происходит за счет клеток камбия.
Часть С:
1. Назовите фазу мейоза и опишите процессы.
2. Запишите схему непрямого развития с неполным метаморфозом и приведите примеры.
Тестовая работа по теме: «Размножение и индивидуальное развитие организмов».
2 вариант
Часть А: (выберите один правильный ответ из четырех возможных)
1.Как называется способ бесполого размножения отдельными органами.
а) споруляция б) почкование
в) фрагментация г) вегетативное
2. Какой из организмов размножается с помощью спор.
а) земляника б) папоротник
в) инфузория в) гидра
3. Как называется способ полового размножения, при котором происходит слияние ядерного
содержимого?
а) коньюгация б) почкование
в) фрагментация г) партеногенез
4. Как называется стадия интерфазы, где набор хромосом 2n2c?
Как называется стадия интерфазы, где набор хромосом 2n2c?
а) синтетическая б) телофаза
в) постсинтетическая г) пресинтетическая
5. В какой фазе мейоза происходит расхождение парных хромосом к полюсам клетки?
а) анафаза 1 б) телофаза 2
в) анафаза 2 г) метафаза 1
6. В результате какого процесса происходит образование бивалента?
а) кроссинговер б) спирализация
в) деспирализация г) коньюгация
7. Какой фазе мейоза соответствует процесс 1n2c → 1n1c ?
а) анафаза 1 б) телофаза 2
в) анафаза 2 г) метафаза 1
8. В метафазе 2 набор хромосом характеризуется формулой:
а) 2n2c б) 1n2c
в) 2n4c г) 1n1c
9. В результате мейоза у животных формируются:
а) гаметы б) споры
в) зигота г) гаметофит
10. В зоне размножения при развитии половых клеток происходит:
а) митоз б) рост
в) мейоз г) ничего не происходит
11. Осевые структуры зародыша образуются на стадии:
а) нейруляции б) гаструляции
в) бластуляции г) органогенеза
12. Покровная система организма формируется из:
Покровная система организма формируется из:
а) эктодермы б) энтодермы
в) зиготы г) мезодермы
13. Неограниченный рост характерен для:
а) вороны б) березы
в) человека г) собаки
14. Взрослое растение папоротника – это…
а) спорофит б) зигота
в) гаметофит г) заросток
Часть В: Соотнесите названия половых клеток и их характеристики.
А – яйцеклетка 1. Имеет головку и хвостик
Б – сперматозоид 2. Имеет крупные размеры
3. Имеет запас питательных веществ
4. Есть акросома
5. Имеет гаплоидный набор хромосом
6. В шейке находятся митохондрии
7. Открыта А.Левенгуком
8. Открыта К.Бэром
Открыта К.Бэром
9. Подвижна
10. Неподвижна
Часть С:
1. Назовите фазу мейоза и опишите процессы.
2. Запишите схему непрямого развития с полным метаморфозом и приведите примеры.
Ответы на задания 1 варианта:
Часть А
1 — б
2 – г
3 – г
4 – в
5 – а
6 – б
7 – а
8 – г
9 – б
10 – в
11 – б
12 – б
13 – г
14 – в
Часть В
Неверные утверждения:
2 – Мужской гаметофит у цветковых растений представлен не зародышевым мешком, а пыльцевым зерном.
3 — После трех митотических делений мегаспоры образуется не двуядерный зародышевый мешок, а восьмиядерный.
5 — Зародыш цветковых растений имеет диплоидный набор хромосом, а эндосперм – триплоидный.
Часть С
1 – это анафаза второго деления мейоза. Процессы: нити ахроматинового веретена сокращаются и к полюсам клетки расходятся хроматиды, которые впоследствии станут самостоятельными хромосомами.
Процессы: нити ахроматинового веретена сокращаются и к полюсам клетки расходятся хроматиды, которые впоследствии станут самостоятельными хромосомами.
2 – яйцо, личинка, взрослая особь
Примеры: земноводные, прямокрылые и т.д.
Ответы на задания 1 варианта:
Часть А
1 – г
2 – б
3 – а
4 – г
5 – а
6 – г
7 – в
8 – б
9 – а
10 – а
11 – а
12 – а
13 – б
14 – а
Часть В
А – 2, 3, 5, 8, 10
В – 1, 4, 5, 6, 7, 9
Часть С
1 – это метафаза первого деления мейоза. Процессы: пары гомологичных хромосом (биваленты) располагаются по экватору клетки, к каждой хромосоме присоединяется нить веретена деления только от одного полюса, материнские и отцовские по происхождению хромосомы ориентированы к полюсам произвольно.
2 – яйцо, личинка, куколка, взрослая особь
Примеры: чешуекрылые, жуки и т.д.
%d1%84%d1%80%d0%b0%d0%b3%d0%bc%d0%b5%d0%bd%d1%82%d0%b0%d1%86%d0%b8%d1%8f 🎓 ⚗ перевод со всех языков на все языки
Все языкиАбхазскийАдыгейскийАзербайджанскийАймараАйнский языкАканАлбанскийАлтайскийАнглийскийАрабскийАрагонскийАрмянскийАрумынскийАстурийскийАфрикаансБагобоБаскскийБашкирскийБелорусскийБолгарскийБретонскийБурятскийВаллийскийВарайскийВенгерскийВепсскийВерхнелужицкийВьетнамскийГаитянскийГреческийГрузинскийГуараниГэльскийДатскийДолганскийДревнерусский языкИвритИдишИнгушскийИндонезийскийИнупиакИрландскийИсландскийИспанскийИтальянскийЙорубаКазахскийКарачаевскийКаталанскийКвеньяКечуаКиргизскийКитайскийКлингонскийКомиКорейскийКриКрымскотатарскийКумыкскийКурдскийКхмерскийЛатинскийЛатышскийЛингалаЛитовскийЛожбанЛюксембургскийМайяМакедонскийМалайскийМаньчжурскийМаориМарийскийМикенскийМокшанскийМонгольскийНауатльНемецкийНидерландскийНогайскийНорвежскийОрокскийОсетинскийОсманскийПалиПапьяментоПенджабскийПерсидскийПольскийПортугальскийРумынский, МолдавскийРусскийСанскритСеверносаамскийСербскийСефардскийСилезскийСловацкийСловенскийСуахилиТагальскийТаджикскийТайскийТатарскийТвиТибетскийТофаларскийТувинскийТурецкийТуркменскийУдмурдскийУзбекскийУйгурскийУкраинскийУрдуУрумскийФарерскийФинскийФранцузскийХиндиХорватскийЦерковнославянский (Старославянский)ЧеркесскийЧерокиЧеченскийЧешскийЧувашскийШайенскогоШведскийШорскийШумерскийЭвенкийскийЭльзасскийЭрзянскийЭсперантоЭстонскийЮпийскийЯкутскийЯпонский
Все языкиАбхазскийАварскийАдыгейскийАзербайджанскийАймараАйнский языкАлбанскийАлтайскийАнглийскийАрабскийАрмянскийАфрикаансБаскскийБашкирскийБелорусскийБолгарскийВенгерскийВепсскийВодскийВьетнамскийГаитянскийГалисийскийГреческийГрузинскийДатскийДревнерусский языкИвритИдишИжорскийИнгушскийИндонезийскийИрландскийИсландскийИспанскийИтальянскийЙорубаКазахскийКарачаевскийКаталанскийКвеньяКечуаКитайскийКлингонскийКорейскийКрымскотатарскийКумыкскийКурдскийКхмерскийЛатинскийЛатышскийЛингалаЛитовскийЛожбанМайяМакедонскийМалайскийМальтийскийМаориМарийскийМокшанскийМонгольскийНемецкийНидерландскийНорвежскийОсетинскийПалиПапьяментоПенджабскийПерсидскийПольскийПортугальскийПуштуРумынский, МолдавскийРусскийСербскийСловацкийСловенскийСуахилиТагальскийТаджикскийТайскийТамильскийТатарскийТурецкийТуркменскийУдмурдскийУзбекскийУйгурскийУкраинскийУрдуУрумскийФарерскийФинскийФранцузскийХиндиХорватскийЦерковнославянский (Старославянский)ЧаморроЧерокиЧеченскийЧешскийЧувашскийШведскийШорскийЭвенкийскийЭльзасскийЭрзянскийЭсперантоЭстонскийЯкутскийЯпонский
Почкование, Фрагментация — Справочник химика 21
У архебактерий описаны разные способы размножения равновеликое бинарное деление, почкование, фрагментация. Все они имеются и у эубактерий. [c.409]
Все они имеются и у эубактерий. [c.409]У многоклеточных животных (почкование, фрагментация) [c.87]
Бесполое размножение осуществляется обычно при помощи спор, путем почкования или фрагментации. Наиболее широко распространено [c.155]
Способ деления данной бактерии обычно не выявляется при первом поверхностном осмотре препарата для этого требуются продолжительные наблюдения живых клеток в подходящей среде [5, 10]. Методики простого окрашивания не позволяют с достаточной точностью определить, к какому типу относится деление — делению на две или на три клетки, делению почкованием или фрагментацией. Для этой цели может оказаться необходимым окрашивание клеточной стенки или даже электронная микроскопия. [c.65]
Для микоплазм описаны. различные способы размножения бинарное деление, фрагментация крупных тел и нитей, сопровождающаяся освобождением большого числа кокковидных форм процесс, сходный с почкованием.
 [c.157]
[c.157]Относятся прокариоты, у которых отсутствует клеточная стенка и не синтезируются предшественники пептидогликана. Клетки окружены ЦПМ, чрезвычайно плеоморфны. Размножение бинарным делением, почкованием, фрагментацией. Окрашивание по Граму отрицательное. Характерно образование мелких, врастающих в агар колоний. Могут быть сапрофитами, паразитами или патогенами. Представлены одним классом Мо111си1е5 [c.159]
Бесполое размножение осуществляется обычно при помощи спор, путем почкования или фрагментации. Наиболее широко распространено и наиболее дифференцировано спорообразование. Если споры отшну- [c.55]
Грибы размножаются вегетативным, т. е. бесполым путем, но у них есть и половое размножение. Бесполое раампожекке возможно- спорами, почкованием клеток, фрагментацией гиф с образованием оидий или артроспор, а также в результате механического разрыва мицелия. Грибам свойственна высокая регенеративная способность любой обрывок мицелия в благоприятных условиях дает рост, превращаясь в организм следующего поколения. Половое размножение грибов происходит в результате слияния двух клеток — гамет (плазмогамия), сопро- [c.72]
Половое размножение грибов происходит в результате слияния двух клеток — гамет (плазмогамия), сопро- [c.72]
Грибы размножаются спорами половым и бесполым способами, а также вегетативным путем (почкование или фрагментация гиф). Грибы, размножающиеся половым и бесполым путем, относятся к совершенным. Несовершенными называют фибы, у которых отсутствует или еще не описан половой путь размножения. Бесполое размножение осуществляется у фибов с помощью эндогенных спор, созревающих внутри круглой структуры — спорангия (рис. 2.6,а), и экзогенных спор — конидий, формирующихся на кончиках плодоносящих гиф (рис. 2.6,б,в). [c.33]
Фрагментация — Определение и примеры
Фрагментация
сущ., Множественное число: фрагментации
[ˌfræɡmɛnˈteɪʃən]
Определение: разбиение на более мелкие части, как при бесполом размножении, когда родительский организм разбивается на фрагменты
Определение фрагментации
? В общем, фрагментация относится к состоянию или процессу разделения на более мелкие части, называемые фрагментами . В биологии это может относиться к процессу репродуктивной фрагментации как к форме бесполого размножения или к этапу определенных клеточных активностей, таких как апоптоз и клонирование ДНК.
В биологии это может относиться к процессу репродуктивной фрагментации как к форме бесполого размножения или к этапу определенных клеточных активностей, таких как апоптоз и клонирование ДНК.
Фрагментация (определение биологии): (1) форма бесполого размножения, при которой родительский организм распадается на фрагменты, каждый из которых способен независимо расти в новый организм, (2) распадаться на более мелкие части или фрагменты. Синонимы: скалывание.
Фрагментация в репродуктивной биологии
Процесс фрагментации является одним из наиболее важных механизмов, которые чаще всего происходят в многоклеточном организме (т. Е. В организме, который состоит из более чем одной клетки).В репродуктивной биологии фрагментация относится к процессу, при котором живой организм разделяется на части, которые впоследствии становятся идентичными исходному организму. Различные тела или части, которые образуются в результате такого процесса, называются фрагментами , и иллюстрируют бесполое размножение.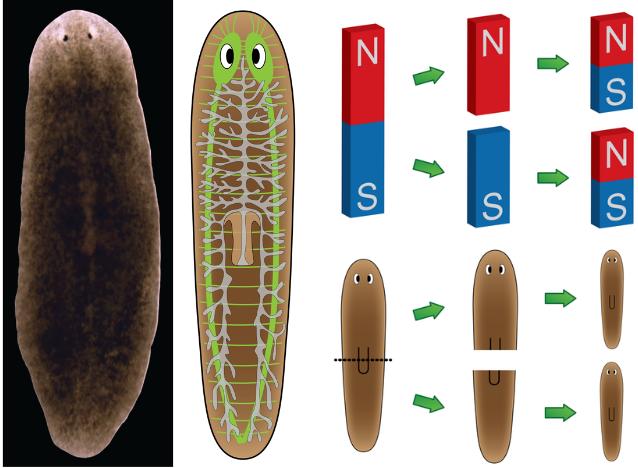
При фрагментации фрагменты образуются в результате расщепления различных живых организмов. Каждый из сформированных фрагментов после прохождения цикла роста превращается в зрелую, полностью выросшую особь, идентичную своему родителю по форме, размеру и другим параметрам.(Ссылка 1)
Основной характеристикой процесса фрагментации является то, что этот процесс может происходить как намеренно, так и непреднамеренно. Например, искусственные причины или изменения окружающей среды могут привести к фрагментации организма и, в конечном итоге, к его развитию в полные и зрелые организмы, похожие на родителя.
Фрагментация встречается как у животных, так и у растений. Грибы, лишайники, плесень, черви, морские звезды, плоские черви acoel и губки — вот некоторые из распространенных примеров, когда способ размножения происходит путем фрагментации.На изображении ниже показана фрагментация морской звезды. Можно увидеть, что фрагментирующее тело отделяется от руки морской звезды, и через некоторое время оно начинает развиваться в новую морскую звезду, как показано на рисунке 1.
. Фрагментация и расщепление.
. Процесс фрагментации — это действительно процесс разрушения организма на различные фрагменты, которые со временем развиваются в целостный организм.Таким образом, этот процесс также называют процессом расщепления.
Процесс воспроизводства
Биологический процесс, в результате которого от родителей образуются новые особи / организмы, называется воспроизводством. Половое и бесполое размножение — это две формы размножения, которые широко изучаются многими исследователями. Процесс, посредством которого потомство рождается в результате спаривания двух родителей, называется половым воспроизводством . Каждый из родителей вносит яйцеклетку от самки и сперму от самца, и после объединения этих двух гамет в процессе оплодотворения образуется зигота, которая превращается в эмбрион и, наконец, в полностью выросший организм.Аллогамия, внутреннее оплодотворение, внешнее оплодотворение и автогамия — вот некоторые из типов полового размножения. Процесс полового размножения подробно описан на Рисунке 2.
Процесс полового размножения подробно описан на Рисунке 2.
Процесс, посредством которого цикл воспроизводства достигается только одним родителем, называется бесполым воспроизводством .Примеры включают бинарное деление, почкование, фрагментацию и партеногенез. Процесс бинарного деления наблюдается у бактерий, в которых одна бактерия имеет тенденцию делиться на две клетки. Во время бутонизации в телах родителей образуются очень маленькие органы, и поэтому через некоторое время они имеют тенденцию ломаться и, таким образом, в конечном итоге превращаться в двух особей. Этот тип бесполого размножения встречается у дрожжей и некоторых других гидр. (Ссылка 2) При фрагментации размножение происходит через разрушение родительского организма на различные фрагменты, которые могут расти так же, как их родители. Обычно он встречается в кораллах, губках и морских звездах. В партеногенезе этот тип бесполого размножения встречается у многих рыб, рептилий и амфибий.
Обычно он встречается в кораллах, губках и морских звездах. В партеногенезе этот тип бесполого размножения встречается у многих рыб, рептилий и амфибий.
Для сравнения полового и бесполого размножения см. Таблицу ниже:
Таблица 1: Сравнение полового и бесполого размножения
| Половое размножение | Бесполое размножение |
|---|---|
| Участвуют два родителя | Один родитель участвует |
| Яйцеклетка оплодотворяется спермой | Новый организм образуется из родительского организма, и оба они идентичны друг другу |
| Встречается у людей, многих растений и многих животных | Встречается в бактериях , рептилии, некоторые животные и многие растения |
| Потомки отличаются от своих родителей во многих основных аспектах | Потомки идентичны своим родителям и предкам |
Процесс занимает много времени, но приводит к большим генетическим изменениям. вариации в следующем поколении вариации в следующем поколении | Процесс относительно быстрый г, но в следующем поколении нет / ограничены генетические вариации. |
Фрагментация — общие шаги
Процесс фрагментации очень важен в биологии для бесполого размножения.Как описано ранее, это форма воспроизводства, при которой небольшие органы или части тела родительской особи отделяются и, наконец, превращаются в полностью зрелый организм. Давайте возьмем для примера Planaria , одного из многих плоских червей, принадлежащих к классу Rhabditophora и филюму Platyhelminthes. См. Рис. 3 для изображения фрагментации и этапов фрагментации в Planaria.
Рис. 3. Тело организма делится на более мелкие органы или часть тела отделяется от родительского тела.Затем фрагменты начинают врастать в зрелый организм. После завершения цикла роста новые организмы будут идентичны родительским. Предоставлено: Шилпа Нагпал, Classnotes.org.inФрагментация и регенерация
Слова «фрагментация» и «регенерация» часто называют словами, используемыми при воспроизведении. Тем не менее, в них есть некоторые общие различия, которые вполне очевидны. Процесс фрагментации часто происходит, когда часть организма разбивается на более мелкие фрагменты, которые могут вырасти в отдельные идентичные организмы, тогда как регенерация — это процесс, при котором организм способен восстанавливать определенные части тела, которые были потеряны.По сути, новые виды образуются в результате фрагментации, тогда как новые органы образуются во время регенерации. Считается, что все организмы способны восстанавливать утраченные части тела, в то время как лишь некоторые из них обладают способностью воспроизводиться посредством фрагментации. Процесс регенерации наблюдается у ящериц и осьминогов. Подробности фрагментации и регенерации можно найти на Рисунке 4. (Ссылка 3)
Тем не менее, в них есть некоторые общие различия, которые вполне очевидны. Процесс фрагментации часто происходит, когда часть организма разбивается на более мелкие фрагменты, которые могут вырасти в отдельные идентичные организмы, тогда как регенерация — это процесс, при котором организм способен восстанавливать определенные части тела, которые были потеряны.По сути, новые виды образуются в результате фрагментации, тогда как новые органы образуются во время регенерации. Считается, что все организмы способны восстанавливать утраченные части тела, в то время как лишь некоторые из них обладают способностью воспроизводиться посредством фрагментации. Процесс регенерации наблюдается у ящериц и осьминогов. Подробности фрагментации и регенерации можно найти на Рисунке 4. (Ссылка 3)
Фрагментация — преимущества
Основным преимуществом фрагментации является то, что она может быть достигнута только с одним родителем, поскольку спаривание между самцом и самкой не является обязательным. Полный цикл воспроизводства может быть легко достигнут путем образования фрагментов из тел родительского организма, а затем их роста и превращения в новые организмы, идентичные их династиям. Второе важное преимущество процесса фрагментации заключается в том, что его цикл воспроизведения может быть достигнут за относительно меньшее количество времени.Все время, которое тратится впустую во время брачного сезона и условий полового размножения, избегается при размножении посредством бесполых систем. Таким образом, фрагментация как разновидность бесполого размножения — очень быстрый процесс. Более того, поскольку фрагментация — это очень быстрый процесс получения потомства, считается, что относительно большее количество организмов может быть получено за более короткое время, поскольку цепочка воспроизводства идет от одной особи к двум и от двух к четырем. Следовательно, за меньшее время целое новое поколение идентичных организмов может быть получено от одного родителя посредством фрагментации.
Полный цикл воспроизводства может быть легко достигнут путем образования фрагментов из тел родительского организма, а затем их роста и превращения в новые организмы, идентичные их династиям. Второе важное преимущество процесса фрагментации заключается в том, что его цикл воспроизведения может быть достигнут за относительно меньшее количество времени.Все время, которое тратится впустую во время брачного сезона и условий полового размножения, избегается при размножении посредством бесполых систем. Таким образом, фрагментация как разновидность бесполого размножения — очень быстрый процесс. Более того, поскольку фрагментация — это очень быстрый процесс получения потомства, считается, что относительно большее количество организмов может быть получено за более короткое время, поскольку цепочка воспроизводства идет от одной особи к двум и от двух к четырем. Следовательно, за меньшее время целое новое поколение идентичных организмов может быть получено от одного родителя посредством фрагментации. Наконец, еще одно преимущество, которое делает процесс фрагментации очень важным в биологии, — это его способность происходить и развиваться в любой среде и любых климатических условиях. (Ссылка 4)
Наконец, еще одно преимущество, которое делает процесс фрагментации очень важным в биологии, — это его способность происходить и развиваться в любой среде и любых климатических условиях. (Ссылка 4)
Фрагментация — недостатки
Хотя из вышеприведенного обсуждения можно понять, что бесполое размножение, особенно фрагментация, играет очень важную роль в поддержании равновесия в экосистеме, производя более крупные организмы за относительно меньшее время, но все же есть много недостатков, связанных с этим процессом.Главный недостаток фрагментации связан с биоразнообразием.
Бесполое размножение осуществляется одним родителем, и в связи с этим новому поколению передаются только его черты, гены и характеристики. Без особого генетического разнообразия следующее поколение, как правило, очень идентично друг другу. И наоборот, при половом размножении смешиваются два гена от обоих родителей. Это дает начало генетически разнообразному потомству.
Второй недостаток фрагментации заключается в том, что те же проблемы наследия, которые однажды были обнаружены у родителей, скорее всего, проявятся в следующем поколении. Этот процесс возникает постоянно при бесполом размножении, одни и те же черты и хромосомы копируются и передаются всему потомству. В результате потомство может быть подвержено раннему исчезновению из-за некоторых болезней, которые они унаследовали.
Этот процесс возникает постоянно при бесполом размножении, одни и те же черты и хромосомы копируются и передаются всему потомству. В результате потомство может быть подвержено раннему исчезновению из-за некоторых болезней, которые они унаследовали.
Виды, полученные путем фрагментации, как правило, идентичны. Таким образом, слабость их тела может сделать их уязвимыми для атак паразитов и хищников. Это может заставить их бороться за выживание. Если возникнет дополнительное давление, например, внезапный сдвиг в окружающей среде или среде обитания, виды могут бороться еще больше и в конечном итоге могут быть уничтожены и вымерли.Напротив, такие проблемы с меньшей вероятностью возникнут при половом размножении, потому что это способствует большему биоразнообразию.
По сути, фрагментация в биологии связана с проблемами контроля в популяции, поскольку это цепной процесс, в котором организм делится на два, а затем из этих фрагментов образуются еще два. Это приводит к воспроизводству массовых популяций от одного родителя.
Примеры фрагментации
Процесс фрагментации встречается у растений, животных и различных микроскопических организмов как форма воспроизводства.
Фрагментация грибов и водорослей
Фрагментация очень часто наблюдается как у грибов, так и у лишайников. Плесень, дрожжи и грибы — это некоторые из видов грибов, размножение которых достигается за счет бесполой фрагментации. Особый тип структуры, которая достигается в процессе расщепления, называется гифами. Они представляют собой нитчатые ветви, из которых состоит мицелий грибов. Это ветви материнских грибов, откуда они легко отделяются и трансформируются.Перед процессом расщепления гифы растут на грибах, откуда они получают пищу и воду, необходимые для роста и питания. Когда гифы становятся достаточно зрелыми, чтобы питаться самостоятельно и размножаться, они отделяются от родительского тела и живут независимо. Некоторые водоросли также размножаются путем фрагментации. Например, нитчатые водоросли, такие как Spirogyra , размножаются, разрезая себя на фрагменты.
Фрагментация у растений
Фрагментация у растений обычно описывается как вегетативный тип фрагментации. У растений в основном диаметр их колоний увеличивается за счет образования новых систем корней и побегов за счет распространения корневищ или столонов старых деревьев, кустарников, папоротников и многолетних растений. Таким образом, как только корень отделен от колонии, он начинает расти сам по себе, давая начало совершенно новой корневой системе. Другими распространенными примерами естественной фрагментации являются следующие:
У растений в основном диаметр их колоний увеличивается за счет образования новых систем корней и побегов за счет распространения корневищ или столонов старых деревьев, кустарников, папоротников и многолетних растений. Таким образом, как только корень отделен от колонии, он начинает расти сам по себе, давая начало совершенно новой корневой системе. Другими распространенными примерами естественной фрагментации являются следующие:
- Древесные растения, например ивы, ветки опадают довольно легко и при соответствующих условиях окружающей среды могут вырасти в новое растение.Это естественное опадение веточек известно как кладоптоз .
- Отделение частей растения с образованием нового организма также наблюдается у недревесных растений, таких как Kalanchoe daigremontiana.
 На фото ниже обратите внимание на придаточные ростки на листе K. daigremontiana. Растения могут упасть и впоследствии пустить корни в землю, чтобы стать новым и независимым растением.
На фото ниже обратите внимание на придаточные ростки на листе K. daigremontiana. Растения могут упасть и впоследствии пустить корни в землю, чтобы стать новым и независимым растением.
 На фото ниже обратите внимание на придаточные ростки на листе K. daigremontiana. Растения могут упасть и впоследствии пустить корни в землю, чтобы стать новым и независимым растением.
На фото ниже обратите внимание на придаточные ростки на листе K. daigremontiana. Растения могут упасть и впоследствии пустить корни в землю, чтобы стать новым и независимым растением.- Несосудистые растения, такие как мхи и печеночники, размножение путем фрагментации также является обычным явлением. Merchantia polymorpha (печеночник обыкновенный) является примером несосудистого растения, которое фрагментирует, производя геммы, которые отламываются от исходного растения и превращаются в новое и независимое растение.
Ниже приведено видео, на котором показано возникновение фрагментации растений и ее значение для них.
Фрагментация у животных
Фрагментация наблюдается во многих колониях кораллов. И твердые кораллы, и мягкие кораллы легко разрушаются. Точно так же такие породы, как Montipora, Acropora, Pocillopora, Euphyllia, и Caulastrea , способны к фрагментации.Многие морские анемоны воспроизводят себя в процессе расщепления или фрагментации. Другими примерами фрагментации животных являются морские звезды и Planaria , которые были подробно описаны в предыдущих разделах. (Ссылка 5)
И твердые кораллы, и мягкие кораллы легко разрушаются. Точно так же такие породы, как Montipora, Acropora, Pocillopora, Euphyllia, и Caulastrea , способны к фрагментации.Многие морские анемоны воспроизводят себя в процессе расщепления или фрагментации. Другими примерами фрагментации животных являются морские звезды и Planaria , которые были подробно описаны в предыдущих разделах. (Ссылка 5)
Фрагментация в клеточной и молекулярной биологии: определение и биологическое значение
Давайте посмотрим на клеточный уровень процесса репродуктивной фрагментации. Клонирование ДНК относится к дублированию молекул ДНК — созданию точной копии или «клона» ДНК, особенно для бесполого размножения.Естественным путем организм создает идентичные молекулы ДНК для своего «клона» (потомства). Слово клонирование здесь относится к тому факту, что репродуцированные особи демонстрируют ДНК, идентичную ДНК их предков. Это происходит при фрагментации. Однако это не исключительный процесс, поскольку клонирование ДНК происходит и при других формах бесполого размножения, таких как бинарное деление. Таким образом, клонирование ДНК — важный клеточный процесс бесполого размножения.
Клонирование ДНК относится к дублированию молекул ДНК — созданию точной копии или «клона» ДНК, особенно для бесполого размножения.Естественным путем организм создает идентичные молекулы ДНК для своего «клона» (потомства). Слово клонирование здесь относится к тому факту, что репродуцированные особи демонстрируют ДНК, идентичную ДНК их предков. Это происходит при фрагментации. Однако это не исключительный процесс, поскольку клонирование ДНК происходит и при других формах бесполого размножения, таких как бинарное деление. Таким образом, клонирование ДНК — важный клеточный процесс бесполого размножения.
Клонирование ДНК может быть достигнуто как естественными, так и искусственными методами.Исследователи использовали этот процесс в лабораториях для исследовательских и экспериментальных исследований. Они придумали так называемую процедуру «фрагментации ДНК», которая представляет собой молекулярно-генетический метод, который позволяет им расщеплять и клонировать определенные фрагменты ДНК из генома организма с помощью технологии рекомбинантной ДНК.
Помимо этого молекулярного аспекта использования фрагментации, этот термин также используется для описания другого клеточного процесса, при котором части клетки подвергаются фрагментации. Это особенно заметно при апоптозе.Апоптоз — это процесс, в котором происходит запрограммированное разрушение клеток, и достигается он за счет очень организованной последовательности морфологических изменений. Более того, морфологические изменения характеризуются сжатием клеток и ядер, фрагментацией и конденсацией хроматина, образованием апоптотических телец и процессом фагоцитоза всех доступных соседних клеток. Кроме того, апоптоз включает однородную активацию эндонуклеаз с расщеплением ДНК во фрагментах либо ста восьмидесяти пар оснований, либо кратных ста восьмидесяти пар оснований.Таким образом, связь между апоптозом и фрагментацией ДНК играет очень важную роль в поддержании клеточной организации. Развитие апоптоза клеток играет ключевую роль в спасении здоровых клеток и тканей, работающих в организме человека, от опасного воздействия клеток, которые не работают стабильно и находятся в процессе гибели.
Резюме
Биологический процесс, при котором организмы разделяются на два или более фрагмента, чтобы стать новыми особями (потомками), называется фрагментацией.Эти фрагменты вырастают и превращаются в целое новое поколение с характеристиками, идентичными своим родителям. Фрагментация также называется расщеплением в различной литературе и является синонимом друг друга. Процесс фрагментации является бесполым, что означает, что участвует только один родитель. Следовательно, потомство, скорее всего, будет иметь одинаковые генетические характеристики. С другой стороны, размножение, в котором участвуют родители как мужского, так и женского пола, известно как половое размножение.Оба типа воспроизводства имеют свои плюсы и минусы.
Фрагментация происходит в три этапа. Процесс начинается с образования осколков. Далее следует развитие фрагментов, а затем заканчивается превращением фрагментов в развитые виды. Весь цикл бесполого размножения, особенно фрагментация, имеет большое биологическое значение. Формирование идентичных видов, меньшее время размножения и передача положительных генов от родителей к потомству являются одними из ключевых преимуществ фрагментации, в то время как отсутствие или сокращение генетического разнообразия, те же проблемы наследия и неспособность справиться с окружающей средой. изменения — это некоторые из его недостатков.
Весь цикл бесполого размножения, особенно фрагментация, имеет большое биологическое значение. Формирование идентичных видов, меньшее время размножения и передача положительных генов от родителей к потомству являются одними из ключевых преимуществ фрагментации, в то время как отсутствие или сокращение генетического разнообразия, те же проблемы наследия и неспособность справиться с окружающей средой. изменения — это некоторые из его недостатков.
Фрагментация также используется для обозначения определенных клеточных процессов. Поскольку фрагментация обычно относится к разрыву или разрезанию большого объекта на более мелкие части, она используется для определения клеточных процессов, таких как фрагментация ДНК (происходящая во время клонирования ДНК) и фрагментация клеток, которая, в частности, является последней стадией апоптоза.
Двумя основными областями, в которых фрагментация играет очень важную роль в биологических клетках, являются апоптоз и клонирование ДНК. При клонировании ДНК часть генов переносится из основного тела во фрагменты, тогда как при апоптозе происходит распад мертвых клеток.
Ссылки
1. Миллер-Рашинг, А., Примак, Р., Девиктор, В., и Либа Пейчар. (2019, апрель). Как фрагментация среды обитания влияет на биоразнообразие? Спорный вопрос, лежащий в основе сохранения… ResearchGate; Эльзевир. https://www.researchgate.net/publication/330445263_How_does_habitat_fragmentation_affect_biodiversity_A_controversial_question_at_the_core_of_conservation_biology
2. Веданту. (2019, 24 апреля). Фрагментация. Vedantu.Com; Веданту.https://www.vedantu.com/biology/fragmentation#:~:text=Fragmentation%20in%20Plants%20and%20Animals,a%20form%20of%20asexual%20reproduction.
3. Админ. (2018, август). Разница между фрагментацией и регенерацией. БЮС; BYJU’S. https://byjus.com/biology/difference-between-fragmentation-and-regeneration/#:~:text=Fragmentation%20occurs%20when%20an%20organism,body%20when%20it%20loses%20them.
4. Джессен, Т., Ван, Ю., и Уилмерс, К. С. (2017). Фрагментация среды обитания дает конкурентное преимущество инвазивной древесной белке Sciurus carolinensis. Биологические вторжения, 20 (3), 607–618. https://doi.org/10.1007/s10530-017-1560-8
Биологические вторжения, 20 (3), 607–618. https://doi.org/10.1007/s10530-017-1560-8
5. Хехт, Л., и Олкок, М. (18 июня 2020 г.). Возможные последствия фрагментации среды обитания для благополучия диких животных. ResearchGate; неизвестный. https://www.researchgate.net/publication/342292473_Potential_effects_of_habitat_fragmentation_on_wild_animal_welfare
© BiologyOnline.com Материалы предоставлены и модерированы редакторами Biology Online.
СледующийФрагментация (репродукция) Факты для детей
Кораллы можно размножить в аквариумах, прикрепив «фрагменты» материнской колонии к подходящему субстрату, например, керамической пробке или куску живого камня.Этот аквариум создан специально для выращивания колоний кораллов из фрагов. В биологии фрагментация — это когда тело распадается на две или более частей, и эти части превращаются в новых особей. Фрагментация наблюдается у некоторых водорослей, таких как спирогира.
У многоклеточных или колониальных организмов — это форма бесполого размножения или клонирования, при которой организм разделяется на фрагменты. Каждый из этих фрагментов превращается в зрелых, полностью выросших особей, которые являются клонами исходного организма.
Расщепление может быть или не быть преднамеренным — оно может произойти или не произойти из-за антропогенного или естественного ущерба, нанесенного окружающей средой или хищниками. У этого типа организмов могут развиваться определенные органы или зоны, которые могут быть отслоены или легко отломаны. Если расщепление происходит без предварительной подготовки организма, оба фрагмента должны иметь возможность регенерировать весь организм, чтобы он мог функционировать как воспроизводство.
Фрагментация, также известная как расщепление, как метод размножения наблюдается у многих организмов, таких как нитчатые цианобактерии, плесени, лишайники, многих растений и животных, таких как губки, плоские черви acoel, некоторые кольчатые черви и морские звезды.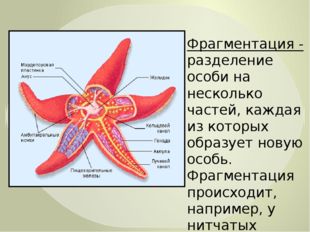
Фрагментация в различных организмах
Плесени, дрожжи и грибы, которые являются частью царства грибов, производят крошечные волокна, называемые гифами. Эти гифы получают пищу и питательные вещества из организма других организмов для роста и оплодотворения. Затем кусок гифы отламывается и превращается в новую особь, и цикл продолжается.
Многие лишайники образуют специализированные структуры, которые могут легко отколоться и разойтись. Эти структуры содержат гифы микобионта и водоросли (фикобионт) (см. Соредии и изидии.Более крупные фрагменты слоевища могут отколоться при высыхании лишайника или из-за механических нарушений (см. Раздел о размножении лишайников).
Растения
Фрагментация — очень распространенный тип вегетативного размножения растений. Многие деревья, кустарники, недревесные многолетние растения и папоротники образуют клональные колонии, создавая новые укорененные побеги корневищами или столонами, что увеличивает диаметр колонии. Если укоренившийся побег отделяется от колонии, значит, произошла фрагментация.Есть несколько других механизмов естественной фрагментации растений.
Если укоренившийся побег отделяется от колонии, значит, произошла фрагментация.Есть несколько других механизмов естественной фрагментации растений.
Люди используют фрагментацию для искусственного размножения многих растений путем деления, отводки, черенкования, прививки, микроразмножения и органов хранения, таких как луковицы, клубнелуковицы, клубни и корневища.
Животные
Животные, такие как губки и колонии кораллов, естественным образом фрагментируются и воспроизводятся. Этим методом размножаются многие виды кольчатых червей и плоских червей.
Фрагментация — Определение | Фрагментация в растениях, грибах и животных
Этот механизм может быть преднамеренным, а может и не быть.Это одна из главных его характеристик. Фрагментация может произойти из-за естественного ущерба, например ущерба окружающей среде от хищников. Это может быть рукотворное. Из-за этого у этих организмов есть тенденция к развитию определенных органов, которые могут легко отколоться или сломаться. Даже если фрагментация может произойти без подготовки организма, все фрагменты должны быть способны регенерировать весь организм, чтобы они могли воспроизводиться и нормально функционировать.
Даже если фрагментация может произойти без подготовки организма, все фрагменты должны быть способны регенерировать весь организм, чтобы они могли воспроизводиться и нормально функционировать.
Как обсуждалось выше, фрагментация — это процесс разделения.Следовательно, это также известно как расщепление. Этот метод размножения наблюдается у многих организмов, таких как нитчатые цианобактерии, лишайники, многие растения и плесень. Он также встречается у таких животных, как губки, плоские черви acoel, морские звезды и некоторые виды кольчатых червей.
Фрагментация в различных организмах
1. Фрагментация в грибах
Фрагментация наблюдается в различных типах грибов, таких как плесень, дрожжи, грибы. Они размножаются путем фрагментации, используя особый тип структуры, известный как гифы.Гифы можно определить как каждую из ветвящихся нитей, которые составляют мицелий гриба. Это разветвленная часть тела родительского грибка, и от него легко избавиться.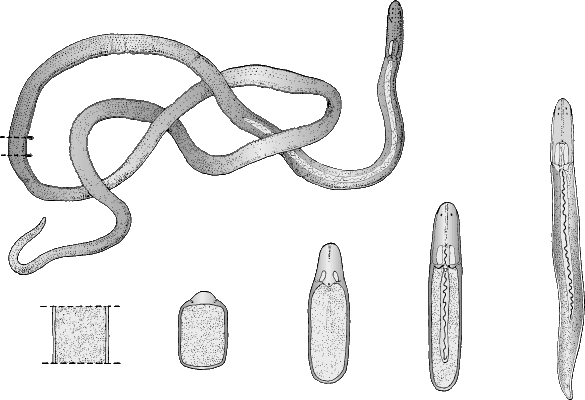 В течение жизненного цикла гиф они получают пищу и другие питательные вещества из тела родительских грибов. Поступая таким образом, гифы в конечном итоге растут и становятся зрелыми, и в конечном итоге они становятся готовыми к оплодотворению. Теперь кусок гиф отрывается от родительского тела и вступает в фазу роста как индивидуальное тело. В конце концов, они также созревают и вырастают гифы, и таким образом цикл продолжается.
В течение жизненного цикла гиф они получают пищу и другие питательные вещества из тела родительских грибов. Поступая таким образом, гифы в конечном итоге растут и становятся зрелыми, и в конечном итоге они становятся готовыми к оплодотворению. Теперь кусок гиф отрывается от родительского тела и вступает в фазу роста как индивидуальное тело. В конце концов, они также созревают и вырастают гифы, и таким образом цикл продолжается.
2. Фрагментация лишайников
Лишайник — это сложный организм, образованный водорослями или цианобактериями, живущими во взаимных отношениях между нитями нескольких видов грибов. Комбинированный лишай имеет свойства, отличные от свойств организмов, из которых он состоит. Лишайники бывают самых разных цветов, размеров и форм. Многие лишайники создают специализированные структуры, которые могут легко разрушаться и рассеиваться. Эти структуры включают гифы микобионтов и водоросли (фикобионты) (см. Соредии и изидии). Более крупные фрагменты слоевища могут разрушаться при высыхании лишайника или из-за механических нарушений (см. Раздел «Размножение лишайников»).
Более крупные фрагменты слоевища могут разрушаться при высыхании лишайника или из-за механических нарушений (см. Раздел «Размножение лишайников»).
Фрагментация у растений
Фрагментация у растений — очень распространенный тип вегетативного размножения. Многие деревья, кустарники, недревесные многолетние растения и папоротники образуют клональные колонии, производя корневища или столоны, чтобы произвести новые укорененные побеги, увеличивающие диаметр колонии. Если укоренившийся побег отделяется от колонии, значит, произошла фрагментация.У растений существует несколько других естественных механизмов фрагментации.
Производство узкоспециализированных репродуктивных структур: например, на листьях некоторых растений появляются придаточные ростки, которые опадают и образуют независимые растения. Tolmieamenziesii и daigremontianakalanchoe. Другие производят такие органы, как луковицы и турионы.
Части, у которых есть высокий потенциал для развития целого растения, легко теряются: некоторые древесные растения, такие как ива, естественным образом опадают.Это известно как кладоптоз. Потерянные ветки могут стать корнями для создания нового растения в подходящей среде. Речные течения часто разрушают обломки ветвей некоторых видов тополя, растущие на берегах реки. Фрагменты могут укореняться и образовывать новые растения, достигающие подходящей среды. У некоторых кактусов и других растений стебли соединены вместе. Он может укорениться и сформировать новое растение, когда отваливается сегмент стебля, называемый подушечкой. Некоторые растения легко оставляют корни при опадании, например.грамм. Эхеверия и Очиток.
У несосудистых растений, например, фрагментация наблюдается также у печеночников и мхов. Небольшие кусочки «штаммов» или «листьев» мха часто разлетаются ветром, водой или животными.
Если фрагмент лося попадает в подходящую среду, он может создать новое растение. Они также производят геммы, например в брызгах Marchantiapolymorpha — чашки, которые легко разбиваются и разбиваются.

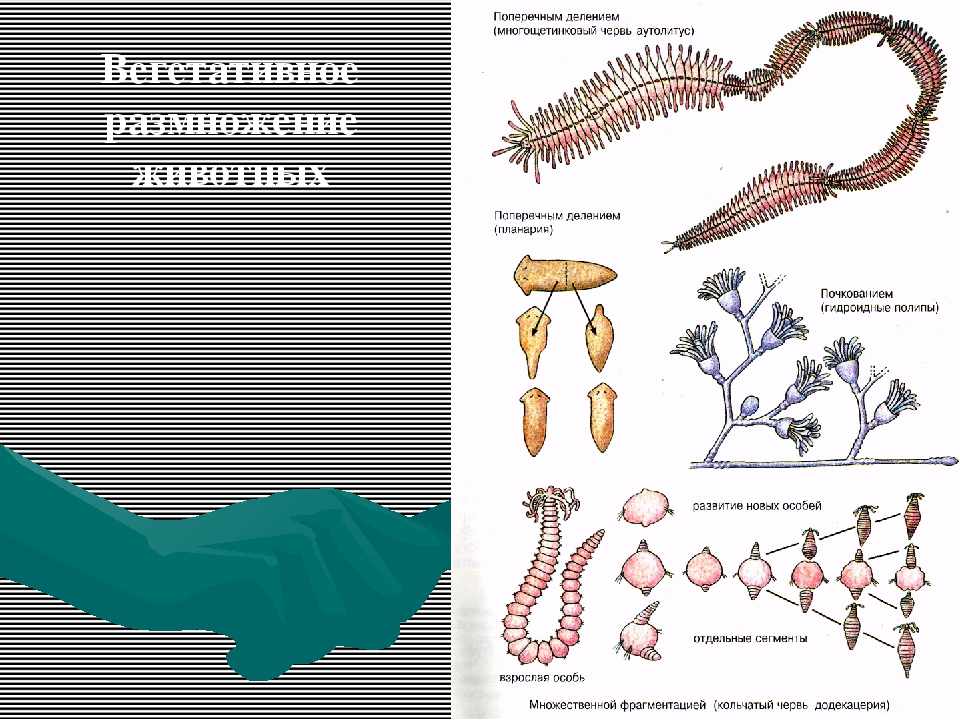 Если фрагмент лося попадает в подходящую среду, он может создать новое растение. Они также производят геммы, например в брызгах Marchantiapolymorpha — чашки, которые легко разбиваются и разбиваются.
Если фрагмент лося попадает в подходящую среду, он может создать новое растение. Они также производят геммы, например в брызгах Marchantiapolymorpha — чашки, которые легко разбиваются и разбиваются.Фрагментация используется людьми для искусственного распространения различных растений.Это делается путем отводов, деления, прививки, срезания и микроразмножения.
Фрагментация у животных
Животные, такие как губки и колонии кораллов, фрагментируются и воспроизводятся естественным образом. Этот метод воспроизводит многие виды кольчатых червей и плоских червей.
Термины «архитомия», «паратомия» и «почкование» используются, когда расщепление происходит из-за определенных изменений в развитии. При архитомии животное раскалывается в определенной точке, и два фрагмента регенерируют отсутствующие органы и ткани.Развитие теряемых тканей не предшествует расщеплению. Перед расколом у животного могут образоваться борозды в зоне расщепления. Полная голова должна быть восстановлена безголовым фрагментом.
Полная голова должна быть восстановлена безголовым фрагментом.
При паратомии расщепление происходит перпендикулярно переднезадней оси, а в задней части расщеплению предшествует «прегенерация» передних структур. Два организма выстраивают оси своего тела, то есть они развиваются от головы до хвоста. Почкование можно рассматривать как паратомию, за исключением того, что оси тела не нужно выравнивать: новая голова может расти вбок или даже указывать назад (например,грамм. Конволутрилобаретрогемма плоского червя acoel).
Коралл
Многие типы колоний кораллов могут увеличиваться в количестве за счет естественной или искусственной фрагментации. Энтузиасты регулярно фрагментируют кораллы внутри аквариума для различных целей, включая контроль формы; продажа, обмен или обмен с другими; регенерационные эксперименты; и сведение к минимуму ущерба естественным коралловым рифам. Раздробить можно как твердые, так и мягкие кораллы. Среди многих других Acropora, Montipora, Pocillopora, Euphyllia и Caulastraea показали высокую толерантность к фрагментации.Большинство морских анемонов размножаются путем фрагментации. Продольное деление, при котором исходная ветреница расщепляется по центру, образуя две анемоны равного размера и разрывая основание, где небольшие части животного отделяются от основания и образуют новую ветреницу.
Среди многих других Acropora, Montipora, Pocillopora, Euphyllia и Caulastraea показали высокую толерантность к фрагментации.Большинство морских анемонов размножаются путем фрагментации. Продольное деление, при котором исходная ветреница расщепляется по центру, образуя две анемоны равного размера и разрывая основание, где небольшие части животного отделяются от основания и образуют новую ветреницу.
Иглокожие
Этот процесс обычно называют делением у иглокожих (термин, который также редко используется для деления в целом). Некоторые виды могут намеренно воспроизводиться таким способом посредством аутотомии.
Теперь бесполое размножение дает несколько преимуществ и этих преимуществ. Прежде чем использовать такой процесс, очень важно знать обе стороны медали, чтобы получить максимальную отдачу от его результатов. Если вам это понадобится больше всего в будущем, вы сможете использовать эти знания.
Список недостатков этого метода воспроизводства
1. Сдерживает разнообразие
Сдерживает разнообразие
Поскольку характеристики и черты только одного из родителей передаются их потомству, бесполое размножение будет препятствовать их поколению генетического разнообразия.Это заставляет популяцию организма быть точно такой же. Это было огромным преимуществом при половом размножении, поскольку мы можем смешивать генофонды, чтобы обеспечить разнообразную экосистему.
2. Это создает некоторые проблемы наследия
Практически всегда требуется только один бесполый родитель, у которого мы можем копировать хромосомы и гены, а это означает, что генетические дефекты или мутации, возникающие в результате бесполого размножения, все еще будут существовать в потомство без исключения. Этот недостаток может даже привести к гораздо более неблагоприятным мутациям, которые делают организмы, полученные бесполым путем, восприимчивыми к болезням, что также может привести к гибели большого количества потомства.
3. На самом деле это может привести к вымиранию — предрасположенный организм
На самом деле это может привести к вымиранию — предрасположенный организм
Все те же характеристики и черты также включают в себя все те же слабости, поэтому мы можем предположить, что определенный паразит или хищник, который эволюционировал, чтобы убить конкретный бесполый организм сможет удалить все его население. Проще говоря, бесполое размножение может привести к борьбе за существование.
4. Он вызывает проблемы с контролем популяции
По сути, эта форма воспроизводства не контролирует быстрое увеличение популяции рассматриваемых организмов.Поскольку в процессе эволюции нет конкуренции, каждый организм обладает высокой способностью к воспроизводству, а это означает, что его популяция даже удваивается в каждом репродуктивном цикле. Однако ученые обнаружили, что этот процесс остановится в тот момент, когда его количество станет чрезмерным.
5. Это приводит к тому, что организмы не могут адаптироваться
Организмы могут реагировать на определенные вещи бесполым размножением, которое затем передается их потомству. А поскольку потомство производит меньше изменений, они станут менее адаптированы к определенным изменениям окружающей среды, что приведет к тому, что они не выживут.На стадии редактирования личинок этот метод более распространен.
А поскольку потомство производит меньше изменений, они станут менее адаптированы к определенным изменениям окружающей среды, что приведет к тому, что они не выживут.На стадии редактирования личинок этот метод более распространен.
24.1. Методы воспроизведения — Концепции биологии — 1-е канадское издание
Раздел 4: Строение и функции животных
Цели обучения
К концу этого раздела вы сможете:
- Опишите преимущества и недостатки бесполого и полового размножения
- Обсудить методы бесполого размножения
- Обсудить методы полового размножения
Животные производят потомство путем бесполого и / или полового размножения.У обоих методов есть достоинства и недостатки. Бесполое размножение дает потомство, которое генетически идентично родителю, поскольку все потомство является клонами исходного родителя. Одна особь может произвести потомство бесполым путем, и большое количество потомства может быть произведено быстро.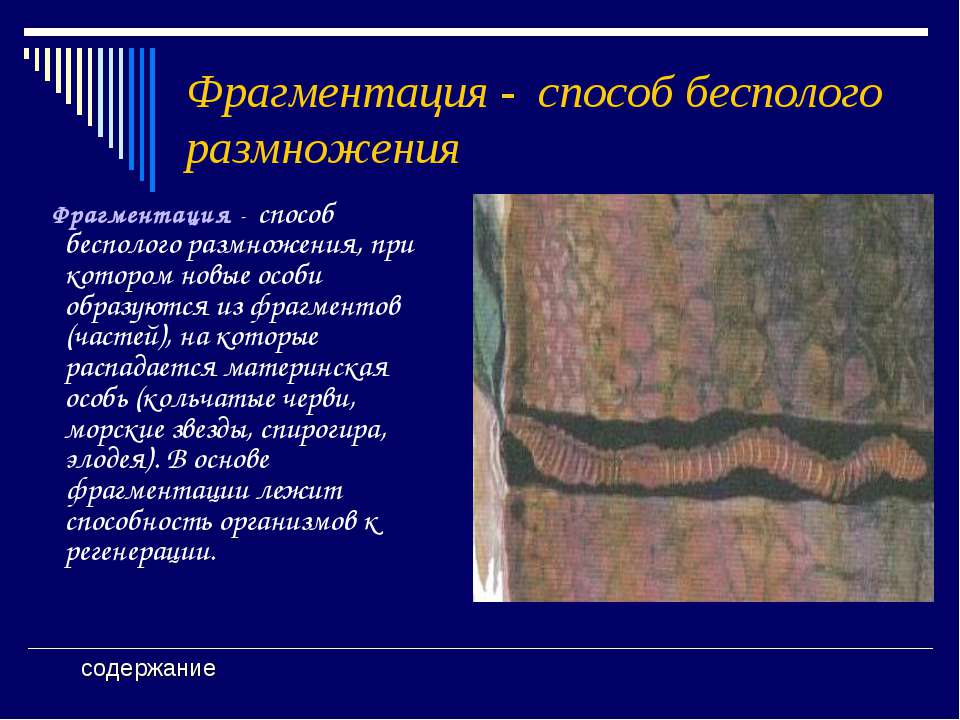 В стабильной или предсказуемой среде бесполое размножение является эффективным средством воспроизводства, потому что все потомство будет адаптировано к этой среде. В нестабильной или непредсказуемой среде виды, размножающиеся бесполым путем, могут оказаться в невыгодном положении, потому что все потомки генетически идентичны и могут не иметь генетической изменчивости для выживания в новых или иных условиях.С другой стороны, высокие темпы бесполого размножения могут позволить быстро отреагировать на изменения окружающей среды, если у людей есть мутации. Дополнительным преимуществом бесполого размножения является то, что заселение новых мест обитания может быть проще, когда человеку не нужно искать себе пару для размножения.
В стабильной или предсказуемой среде бесполое размножение является эффективным средством воспроизводства, потому что все потомство будет адаптировано к этой среде. В нестабильной или непредсказуемой среде виды, размножающиеся бесполым путем, могут оказаться в невыгодном положении, потому что все потомки генетически идентичны и могут не иметь генетической изменчивости для выживания в новых или иных условиях.С другой стороны, высокие темпы бесполого размножения могут позволить быстро отреагировать на изменения окружающей среды, если у людей есть мутации. Дополнительным преимуществом бесполого размножения является то, что заселение новых мест обитания может быть проще, когда человеку не нужно искать себе пару для размножения.
Во время полового размножения генетический материал двух особей объединяется, чтобы произвести генетически различное потомство, которое отличается от своих родителей. Считается, что генетическое разнообразие потомства, произведенного половым путем, дает видам больше шансов на выживание в непредсказуемой или меняющейся среде. Виды, которые размножаются половым путем, должны поддерживать два разных типа особей, самцов и самок, что может ограничивать возможность колонизации новых местообитаний, поскольку должны присутствовать оба пола.
Виды, которые размножаются половым путем, должны поддерживать два разных типа особей, самцов и самок, что может ограничивать возможность колонизации новых местообитаний, поскольку должны присутствовать оба пола.
Бесполое размножение происходит у прокариотических микроорганизмов (бактерий) и некоторых эукариотических одноклеточных и многоклеточных организмов. Есть несколько способов бесполого размножения животных.
Деление , также называемое бинарным делением, происходит у прокариотических микроорганизмов и у некоторых беспозвоночных, многоклеточных организмов.После периода роста организм разделяется на два отдельных организма. Некоторые одноклеточные эукариотические организмы подвергаются бинарному делению путем митоза. У других организмов часть особи отделяется и образует вторую особь. Этот процесс происходит, например, у многих астероидных иглокожих через расщепление центрального диска. Некоторые морские анемоны и некоторые коралловые полипы (рис. 24.2) также размножаются путем деления.
24.2) также размножаются путем деления.
Почкование — это форма бесполого размножения, которая возникает в результате разрастания части клетки или области тела, приводящей к отделению от исходного организма на двух особей.Почкование обычно происходит у некоторых беспозвоночных животных, таких как кораллы и гидры. У гидр образуется почка, которая развивается во взрослую особь и отделяется от основной части тела, как показано на рис. 24.3, тогда как у кораллов бутон не отделяется и размножается в составе новой колонии.
Рисунок 24.3. Размножаются гидры бесполым путём бутонизации.Посмотрите видео о распускании бутонов гидры.
Фрагментация — разделение тела на две части с последующей регенерацией.Если животное способно к фрагментации и часть достаточно большая, отдельная особь вырастет заново.
Например, у многих морских звезд бесполое размножение осуществляется путем фрагментации. На рис. 24.4 изображена морская звезда, у которой рука человека оторвана и возрождает новую морскую звезду. Работники рыболовства, как известно, пытались убить морских звезд, поедающих своих моллюсков или устриц, разрезая их пополам и бросая обратно в океан. К несчастью для рабочих, каждая из двух частей может регенерировать новую половину, в результате чего вдвое больше морских звезд охотятся на устриц и моллюсков.Фрагментация также наблюдается у кольчатых червей, турбеллярий и порифер.
На рис. 24.4 изображена морская звезда, у которой рука человека оторвана и возрождает новую морскую звезду. Работники рыболовства, как известно, пытались убить морских звезд, поедающих своих моллюсков или устриц, разрезая их пополам и бросая обратно в океан. К несчастью для рабочих, каждая из двух частей может регенерировать новую половину, в результате чего вдвое больше морских звезд охотятся на устриц и моллюсков.Фрагментация также наблюдается у кольчатых червей, турбеллярий и порифер.
Обратите внимание, что при фрагментации обычно наблюдается заметная разница в размерах особей, тогда как при делении образуются две особи приблизительного размера.
Партеногенез — это форма бесполого размножения, при которой яйцеклетка развивается в полноценную особь без оплодотворения.Полученное потомство может быть гаплоидным или диплоидным, в зависимости от процесса и вида. Партеногенез происходит у беспозвоночных, таких как водяные бегуны, коловратки, тли, палочники, некоторые муравьи, осы и пчелы. Пчелы используют партеногенез для производства гаплоидных самцов (трутней) и диплоидных самок (рабочих). Если яйцо оплодотворяется, получается матка. Пчелиная матка контролирует воспроизводство пчел в улье, чтобы регулировать тип производимой пчелы.
Партеногенез происходит у беспозвоночных, таких как водяные бегуны, коловратки, тли, палочники, некоторые муравьи, осы и пчелы. Пчелы используют партеногенез для производства гаплоидных самцов (трутней) и диплоидных самок (рабочих). Если яйцо оплодотворяется, получается матка. Пчелиная матка контролирует воспроизводство пчел в улье, чтобы регулировать тип производимой пчелы.
Некоторые позвоночные животные, такие как некоторые рептилии, земноводные и рыбы, также размножаются посредством партеногенеза.Хотя партеногенез чаще встречается у растений, он наблюдался у видов животных, которые были разделены по полу в наземных или морских зоопарках. Две самки драконов Комодо, акула-молот и черная акула дали партеногенное потомство, когда самки были изолированы от самцов.
Половое размножение — это сочетание (обычно гаплоидных) репродуктивных клеток двух особей с образованием третьего (обычно диплоидного) уникального потомства. Половое размножение дает потомство с новыми комбинациями генов. Это может быть адаптивным преимуществом в нестабильных или непредсказуемых средах. Как люди, мы привыкли думать о животных как о двух разных полах — мужском и женском, — определенных при зачатии. Однако в животном мире существует множество вариаций на эту тему.
Это может быть адаптивным преимуществом в нестабильных или непредсказуемых средах. Как люди, мы привыкли думать о животных как о двух разных полах — мужском и женском, — определенных при зачатии. Однако в животном мире существует множество вариаций на эту тему.
Гермафродитизм встречается у животных, у которых одна особь имеет как мужские, так и женские репродуктивные части. Беспозвоночные, такие как дождевые черви, слизни, ленточные черви и улитки, показанные на рис. 24.5, часто являются гермафродитами.Гермафродиты могут самооплодотворяться или спариваться с другими представителями своего вида, оплодотворяя друг друга и производя потомство. Самооплодотворение часто встречается у животных с ограниченной подвижностью или неподвижных, таких как ракушки и моллюски.
Рисунок 24.5. Многие улитки — гермафродиты. Когда две особи спариваются, они могут произвести до ста яиц каждая. (Источник: Ассаф Штильман) Определение пола у млекопитающих определяется генетически по наличию X- и Y-хромосом. Гомозиготные по X (XX) особи — женские, а гетерозиготные (XY) — мужские.Наличие Y-хромосомы вызывает развитие мужских качеств, а ее отсутствие приводит к женским характеристикам. Система XY также встречается у некоторых насекомых и растений.
Гомозиготные по X (XX) особи — женские, а гетерозиготные (XY) — мужские.Наличие Y-хромосомы вызывает развитие мужских качеств, а ее отсутствие приводит к женским характеристикам. Система XY также встречается у некоторых насекомых и растений.
Определение пола птиц зависит от наличия Z- и W-хромосом. Гомозиготный по Z (ZZ) приводит к мужскому полу, а гетерозиготный (ZW) результат к женскому. W, по-видимому, важен для определения пола человека, как и Y-хромосома у млекопитающих. Некоторые рыбы, ракообразные, насекомые (например, бабочки и моль) и рептилии используют эту систему.
Пол некоторых видов определяется не генетикой, а некоторыми аспектами окружающей среды. Например, определение пола у некоторых крокодилов и черепах часто зависит от температуры в критические периоды развития яиц. Это называется определением пола по окружающей среде или, более конкретно, определением пола в зависимости от температуры. У многих черепах более низкие температуры во время инкубации яиц приводят к появлению самцов, а более высокие температуры — к самкам. У некоторых крокодилов при умеренных температурах рождаются самцы, а при высоких и низких температурах — самки.У некоторых видов пол зависит как от генетики, так и от температуры.
У некоторых крокодилов при умеренных температурах рождаются самцы, а при высоких и низких температурах — самки.У некоторых видов пол зависит как от генетики, так и от температуры.
Особи некоторых видов меняют пол в течение жизни, попеременно то самец, то самец. Если индивид в первую очередь женщина, это называется протогиния или «первая женщина», если в первую очередь самец, это называется протандрией или «первым самцом». Например, устрицы рождаются самцами, растут, становятся самками и откладывают яйца; некоторые виды устриц меняют пол несколько раз.
Сводка
Размножение может быть бесполым, когда одна особь производит генетически идентичное потомство, или половым, когда генетический материал от двух особей объединяется для получения генетически разнообразного потомства.Бесполое размножение происходит путем деления, бутонизации и фрагментации. Половое размножение может означать соединение сперматозоидов и яйцеклеток внутри тела животных или может означать выброс спермы и яйцеклеток в окружающую среду.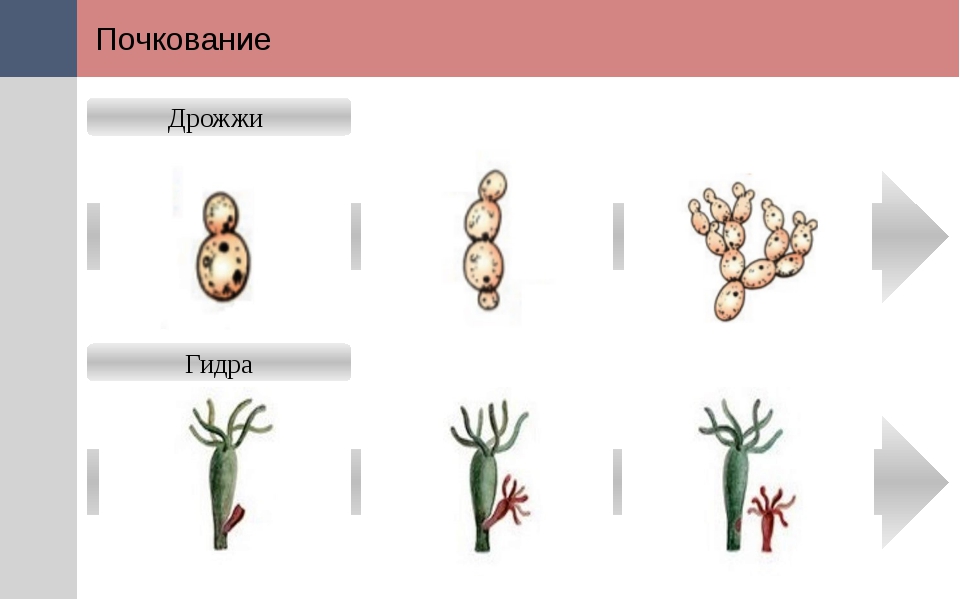 Человек может быть одного или обоих полов; он может начаться как один пол и переключаться в течение жизни, или он может оставаться мужчиной или женщиной.
Человек может быть одного или обоих полов; он может начаться как один пол и переключаться в течение жизни, или он может оставаться мужчиной или женщиной.
Упражнения
- Какая форма воспроизводства считается лучшей в стабильной среде?
- бесполые
- половой
- бутонизация
- партеногенез
- Какая форма воспроизводства может возникнуть в результате повреждения исходного животного?
- бесполые
- фрагментация
- бутонизация
- партеногенез
- Какая форма размножения полезна для малоподвижного животного, которое размножается половым путем?
- деление
- бутонизация
- партеногенез
- гермафродитизм
- Генетически уникальные особи получены через ________.
- половое размножение
- партеногенез
- бутонизация
- фрагментация
- Почему половое размножение полезно, если только половина животных может производить потомство, а две отдельные клетки должны быть объединены, чтобы образовать третью?
- От чего зависит, какой пол даст потомство птиц и млекопитающих?
Ответы
- А
- В
- D
- А
- Половое размножение производит новую комбинацию генов в потомстве, которая может лучше помочь им пережить изменения в окружающей среде и помочь в выживании вида.
- Наличие W-хромосомы у птиц определяет женское начало, а наличие Y-хромосомы у млекопитающих определяет мужской пол. Отсутствие этих хромосом и однородность потомства (ZZ или XX) приводит к развитию другого пола.

Глоссарий
- бесполое размножение
- Форма воспроизводства, дающая потомство, генетически идентичное родителю
- бутонизация
- форма бесполого размножения, которая возникает в результате роста части клетки, приводящей к разделению от исходного животного на двух особей
- деление
- (также бинарное деление) метод увеличения размеров многоклеточных организмов или бесполое размножение, при котором одноклеточный организм разделяется на два отдельных организма путем митоза
- фрагментация
- разрезание или фрагментация исходного животного на части и выращивание отдельного животного из каждой части
- гермафродитизм
- Состояние наличия мужских и женских репродуктивных органов у одного и того же человека
- партеногенез
- форма бесполого размножения, при которой яйцеклетка развивается в полноценную особь без оплодотворения
- половое размножение
- смешение генетического материала от двух особей для получения генетически уникального потомства
методов воспроизведения | Безграничная биология
Методы воспроизведения
Воспроизводство животных необходимо для выживания вида; это может происходить либо бесполым, либо половым путем.
Цели обучения
Опишите воспроизводство животных
Основные выводы
Ключевые моменты
- Репродукция (или продолжение рода) — это биологический процесс, посредством которого новое «потомство» производится от их «родителей».
- Бесполое размножение дает генетически идентичные организмы, потому что один человек размножается без другого.
- При половом размножении генетический материал двух особей одного и того же вида объединяется, чтобы произвести генетически различное потомство; это обеспечивает смешение генофонда видов.
- Организмы, которые размножаются путем бесполого размножения, имеют тенденцию к экспоненциальному росту и зависят от мутаций для изменения ДНК, в то время как те, которые размножаются половым путем, дают меньшее количество потомков, но имеют большую генетическую изменчивость.
Ключевые термины
- воспроизводство : биологическое производство новых особей
- клон : живой организм, полученный бесполым путем от одного предка, которому он генетически идентичен
Репродукция животных
Репродукция (или размножение) — это биологический процесс, посредством которого новое «потомство» (отдельные организмы) производится от их «родителей». «Фундаментальной чертой всей известной жизни является то, что каждый отдельный организм существует в результате воспроизводства. Самое главное, воспроизводство необходимо для выживания вида. Известные методы размножения в целом можно разделить на два основных типа: половое и бесполое.
«Фундаментальной чертой всей известной жизни является то, что каждый отдельный организм существует в результате воспроизводства. Самое главное, воспроизводство необходимо для выживания вида. Известные методы размножения в целом можно разделить на два основных типа: половое и бесполое.
При бесполом размножении особь может воспроизводить потомство без участия другой особи этого вида. Деление бактериальной клетки на две дочерние клетки является примером бесполого размножения.Этот тип воспроизводства производит генетически идентичные организмы (клоны), тогда как при половом размножении генетический материал двух особей объединяется, чтобы произвести потомство, которое генетически отличается от своих родителей.
Во время полового размножения мужская гамета (сперматозоид) может быть помещена внутрь тела самки для внутреннего оплодотворения, или сперма и яйца могут быть выпущены в окружающую среду для внешнего оплодотворения. Люди являются примером первого, а морские коньки — вторым. После брачного танца самка морского конька откладывает яйца в брюшной мешок самца морского конька, где они оплодотворяются. Яйца вылупляются, и потомство в течение нескольких недель развивается в сумке.
После брачного танца самка морского конька откладывает яйца в брюшной мешок самца морского конька, где они оплодотворяются. Яйца вылупляются, и потомство в течение нескольких недель развивается в сумке.
Половое размножение морских коньков : Самки морских коньков откладывают яйца для размножения, которые затем оплодотворяются самцами. В отличие от почти всех других животных, самец морского конька вынашивает детенышей до рождения.
Бесполое и половое размножение
Количество организмов, размножающихся бесполым путем, увеличивается в геометрической прогрессии.Однако, поскольку они полагаются на мутации для изменения своей ДНК, все представители этого вида имеют схожие уязвимости. Организмы, которые размножаются половым путем, дают меньшее количество потомков, но большое количество вариаций в их генах делает их менее восприимчивыми к болезням.
Многие организмы могут размножаться как половым, так и бесполым путем. Тля, слизистая плесень, актинии и некоторые виды морских звезд являются примерами видов животных, обладающих этой способностью. Когда факторы окружающей среды благоприятны, используется бесполое размножение, чтобы использовать подходящие условия для выживания, такие как изобилие пищи, адекватное жилище, благоприятный климат, болезни, оптимальный pH или правильное сочетание других требований образа жизни.Популяции этих организмов экспоненциально увеличиваются за счет бесполых репродуктивных стратегий, позволяющих в полной мере использовать богатые ресурсы снабжения. Когда источники пищи истощаются, климат становится враждебным или индивидуальное выживание подвергается опасности из-за некоторых других неблагоприятных изменений условий жизни, эти организмы переключаются на половые формы воспроизводства.
Когда факторы окружающей среды благоприятны, используется бесполое размножение, чтобы использовать подходящие условия для выживания, такие как изобилие пищи, адекватное жилище, благоприятный климат, болезни, оптимальный pH или правильное сочетание других требований образа жизни.Популяции этих организмов экспоненциально увеличиваются за счет бесполых репродуктивных стратегий, позволяющих в полной мере использовать богатые ресурсы снабжения. Когда источники пищи истощаются, климат становится враждебным или индивидуальное выживание подвергается опасности из-за некоторых других неблагоприятных изменений условий жизни, эти организмы переключаются на половые формы воспроизводства.
Половое размножение обеспечивает смешение генофонда видов. Изменения, обнаруженные у потомства при половом размножении, позволяют некоторым особям лучше приспособиться к выживанию и обеспечивают механизм избирательной адаптации.Кроме того, половое размножение обычно приводит к формированию стадии жизни, способной выдержать условия, угрожающие потомству бесполого родителя. Таким образом, семена, споры, яйца, куколки, цисты или другие «зимующие» стадии полового размножения обеспечивают выживание в неблагоприятные времена, поскольку организм может «переждать» неблагоприятные ситуации, пока не произойдет возврат к пригодности.
Таким образом, семена, споры, яйца, куколки, цисты или другие «зимующие» стадии полового размножения обеспечивают выживание в неблагоприятные времена, поскольку организм может «переждать» неблагоприятные ситуации, пока не произойдет возврат к пригодности.
Типы полового и бесполого размножения
Бесполое и половое размножение, два метода воспроизводства среди животных, дают потомство, которое является клонированным или генетически уникальным.
Цели обучения
Обсудить методы полового и бесполого размножения
Основные выводы
Ключевые моменты
- Бесполое размножение включает деление, почкование, фрагментацию и партеногенез, в то время как половое размножение достигается за счет комбинации репродуктивных клеток двух особей.
- Способность вида к размножению посредством фрагментации зависит от размера части, которая отламывается, в то время как при бинарном делении особь отделяется и образует двух особей одинакового размера.
- Почкование может привести к появлению полностью новой взрослой особи, которая формируется отдельно от исходного тела или может оставаться прикрепленной к исходному телу.
- Наблюдаемый у беспозвоночных и некоторых позвоночных, партеногенез дает потомство, которое может быть гаплоидным или диплоидным.
- Половое размножение, производство потомства с новой комбинацией генов, может также включать гермафродитизм, при котором организм может самооплодотворяться или спариваться с другим особью того же вида.

Ключевые термины
- бинарное деление : процесс, при котором клетка делится бесполым путем с образованием двух дочерних клеток
- гермафродитизм : наличие половых органов у обоих полов
- партеногенез : форма бесполого размножения, при которой рост и развитие эмбрионов происходят без оплодотворения
Способы размножения: бесполое и половое
Бесполое размножение
Бесполое размножение дает потомство, которое генетически идентично родителю, поскольку все потомство является клонами первоначального родителя. Этот тип воспроизводства встречается у прокариотических микроорганизмов (бактерий) и у некоторых эукариотических одноклеточных и многоклеточных организмов. Животные могут размножаться бесполым путем путем деления, почкования, фрагментации или партеногенеза.
Этот тип воспроизводства встречается у прокариотических микроорганизмов (бактерий) и у некоторых эукариотических одноклеточных и многоклеточных организмов. Животные могут размножаться бесполым путем путем деления, почкования, фрагментации или партеногенеза.
Деление
Деление, также называемое бинарным делением, происходит у прокариотических микроорганизмов и у некоторых беспозвоночных, многоклеточных организмов. После периода роста организм разделяется на два отдельных организма. Некоторые одноклеточные эукариотические организмы подвергаются бинарному делению путем митоза.У других организмов часть особи отделяется, образуя вторую особь. Этот процесс происходит, например, у многих астероидных иглокожих через расщепление центрального диска. Некоторые морские анемоны и некоторые коралловые полипы также размножаются путем деления.
Деление : Коралловые полипы размножаются бесполым путем деления, когда организм разделяется на два отдельных организма.
Бутонирование
Почкование — это форма бесполого размножения, которая возникает в результате разрастания части клетки или области тела, приводящей к разделению от исходного организма на двух особей.Почкование обычно происходит у некоторых беспозвоночных животных, таких как кораллы и гидры. У гидр образуется почка, которая развивается во взрослую особь, которая отрывается от основного тела; тогда как у кораллов бутон не отделяется, а размножается как часть новой колонии.
Бутонирование : Гидра размножается бесполым путем за счет бутонизации, когда образуется почка, которая развивается во взрослую особь и отделяется от основного тела.
Фрагментация
Фрагментация — это разделение тела на две части с последующей регенерацией.Если животное способно к фрагментации и часть достаточно большая, отдельная особь вырастет заново.
Многие морские звезды размножаются бесполым путем путем фрагментации. Например, если рука отдельной морской звезды сломана, она возродит новую морскую звезду. Известно, что работники рыболовства пытались убить морских звезд, поедающих их моллюсков или устриц, разрезая их пополам и бросая обратно в океан. К несчастью для рабочих, каждая из двух частей может регенерировать новую половину, в результате чего вдвое больше морских звезд охотятся на устриц и моллюсков.Фрагментация также наблюдается у кольчатых червей, турбеллярий и порифер.
Известно, что работники рыболовства пытались убить морских звезд, поедающих их моллюсков или устриц, разрезая их пополам и бросая обратно в океан. К несчастью для рабочих, каждая из двух частей может регенерировать новую половину, в результате чего вдвое больше морских звезд охотятся на устриц и моллюсков.Фрагментация также наблюдается у кольчатых червей, турбеллярий и порифер.
Фрагментация : Морские звезды могут воспроизводиться путем фрагментации. Большая рука, фрагмент другой морской звезды, превращается в новую личность.
Обратите внимание, что при фрагментации обычно наблюдается заметная разница в размере особей, тогда как при делении образуются две особи примерно одинакового размера.
Партеногенез
Партеногенез — это форма бесполого размножения, при которой яйцеклетка превращается в полноценную особь без оплодотворения.Полученное потомство может быть гаплоидным или диплоидным, в зависимости от процесса и вида. Партеногенез происходит у беспозвоночных, таких как водяные блохи, коловратки, тли, палочники, некоторые муравьи, осы и пчелы. Пчелы используют партеногенез для производства гаплоидных самцов (трутней) и диплоидных самок (рабочих). Если яйцо оплодотворяется, получается матка. Пчелиная матка контролирует воспроизводство пчел в улье, чтобы регулировать тип производимой пчелы.
Пчелы используют партеногенез для производства гаплоидных самцов (трутней) и диплоидных самок (рабочих). Если яйцо оплодотворяется, получается матка. Пчелиная матка контролирует воспроизводство пчел в улье, чтобы регулировать тип производимой пчелы.
Некоторые позвоночные животные, такие как рептилии, земноводные и рыбы, также размножаются посредством партеногенеза.Хотя партеногенез чаще встречается у растений, он наблюдался у видов животных, которые были разделены по полу в наземных или морских зоопарках. Два дракона Комодо, капотоголовая акула и черноперая акула дали партеногенное потомство, когда самки были изолированы от самцов.
Половое размножение
Половое размножение — это сочетание (обычно гаплоидных или имеющих один набор непарных хромосом) репродуктивных клеток от двух особей с образованием третьего (обычно диплоидного или имеющего пару хромосом каждого типа) уникального потомства.Половое размножение дает потомство с новыми комбинациями генов. Это может быть адаптивным преимуществом в нестабильных или непредсказуемых средах.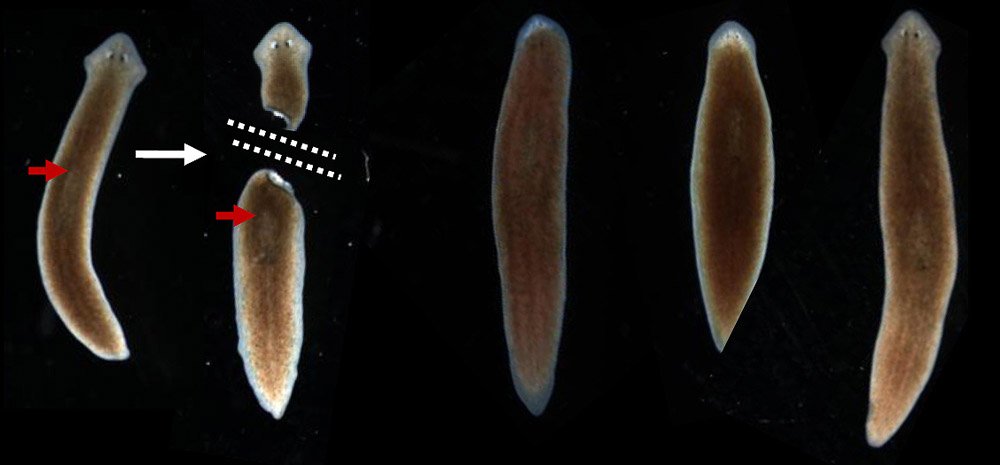 Как люди, мы привыкли думать о животных как о двух разных полах, мужском и женском, определенных при зачатии. Однако в животном мире существует множество вариаций на эту тему.
Как люди, мы привыкли думать о животных как о двух разных полах, мужском и женском, определенных при зачатии. Однако в животном мире существует множество вариаций на эту тему.
Гермафродитизм
Гермафродитизм встречается у животных, у которых одна особь имеет как мужские, так и женские репродуктивные части. Беспозвоночные, такие как дождевые черви, слизни, ленточные черви и улитки, часто являются гермафродитами.Гермафродиты могут самооплодотворяться или спариваться с другими представителями своего вида, оплодотворяя друг друга и производя потомство. Самооплодотворение часто встречается у животных с ограниченной подвижностью или неподвижных, таких как ракушки и моллюски.
Определение пола
Определение пола у животных может регулироваться наличием хромосом или влиянием факторов окружающей среды.
Цели обучения
Различать различные способы, которыми животные определяют пол потомства
Основные выводы
Ключевые моменты
- Млекопитающие, птицы и некоторые другие виды животных зависят от гетерозиготных или гомозиготных комбинаций хромосом для определения пола.
- Прохладные или теплые температуры влияют на определение пола у таких видов, как крокодилы и черепахи.
- Некоторые виды, например, устрицы, способны менять пол несколько раз в течение своей жизни.

Ключевые термины
- protandry : состояние, при котором организм начинает жизнь как мужчина, а затем превращается в женщину
- протогиния : состояние, при котором организм начинает жизнь как женщина, а затем превращается в мужчину
- гомозиготный : организма, в котором обе копии данного гена имеют один и тот же аллель
- гетерозиготный : организм, имеющий два разных аллеля данного гена
Определение пола
Пол млекопитающих определяется генетически по наличию X- и Y-хромосом.Гомозиготные по X (XX) особи — женские, а гетерозиготные (XY) — мужские. Наличие Y-хромосомы вызывает развитие мужских качеств, а ее отсутствие приводит к женским характеристикам.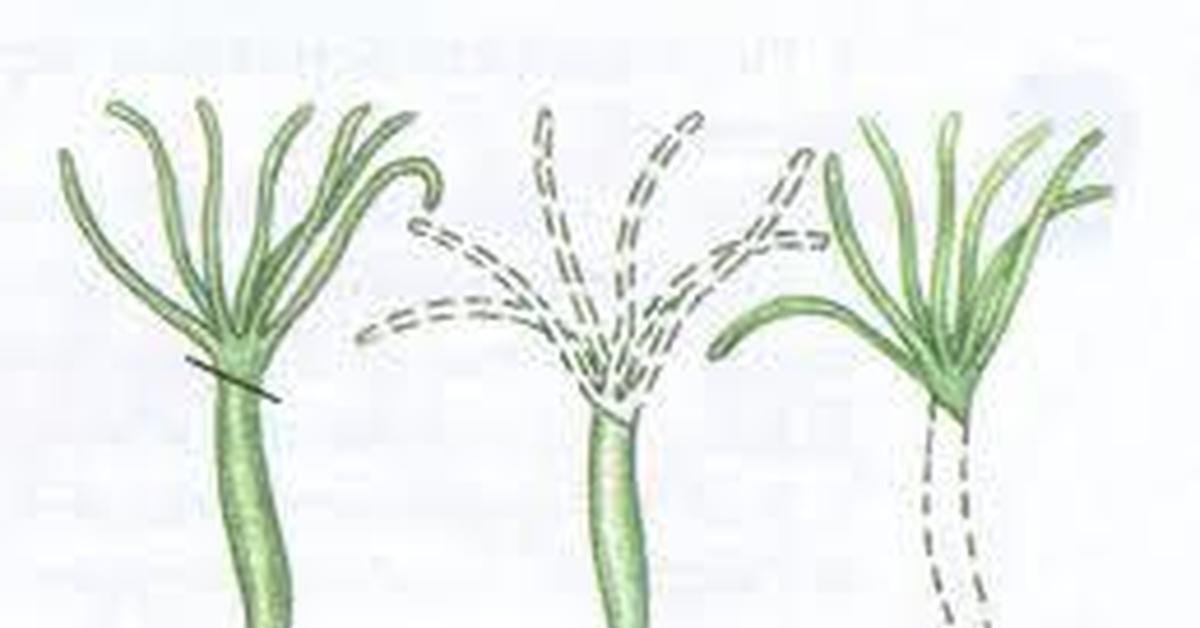 Система XY также встречается у некоторых насекомых и растений.
Система XY также встречается у некоторых насекомых и растений.
Определение пола : Наличие X- и Y-хромосом является одним из факторов, ответственных за определение пола у млекопитающих, причем самцы являются гетерозиготным полом. У птиц Z и W хромосомы определяют пол, а самки являются гетерозиготным полом.
Определение пола птиц зависит от наличия Z- и W-хромосом. Гомозиготный по Z (ZZ) приводит к самцу, а гетерозиготный (ZW) — к самке. W, по-видимому, важен для определения пола человека, как и Y-хромосома у млекопитающих. Некоторые рыбы, ракообразные, насекомые (например, бабочки и моль) и рептилии используют эту систему.
Пол некоторых видов определяется не генетикой, а некоторыми аспектами окружающей среды. Например, определение пола у некоторых крокодилов и черепах часто зависит от температуры в критические периоды развития яиц.Это называется определением пола по окружающей среде или, более конкретно, определением пола в зависимости от температуры. У многих черепах при более низких температурах во время инкубации яиц появляются самцы, а при более высоких температурах — самки. У некоторых крокодилов при умеренных температурах рождаются самцы, а в теплых и холодных — самки. У некоторых видов пол зависит как от генетики, так и от температуры.
У многих черепах при более низких температурах во время инкубации яиц появляются самцы, а при более высоких температурах — самки. У некоторых крокодилов при умеренных температурах рождаются самцы, а в теплых и холодных — самки. У некоторых видов пол зависит как от генетики, так и от температуры.
Особи некоторых видов меняют пол в течение жизни, попеременно то самец, то самец.Если особь — это прежде всего женщина, ее называют протогиния или «первая женщина»; если это прежде всего самец, его называют протандрием или «первым самцом». Например, устрицы рождаются самцами, растут, становятся самками и откладывают яйца; некоторые виды устриц меняют пол несколько раз.
43.1B: Типы полового и бесполого размножения
Бесполое и половое размножение, два метода воспроизводства среди животных, дают потомство, которое является клонами или генетически уникальным.
Задачи обучения
- Обсудить методы полового и бесполого размножения
Ключевые моменты
- Бесполое размножение включает деление, почкование, фрагментацию и партеногенез, в то время как половое размножение достигается за счет комбинации репродуктивных клеток двух особей.
- Способность вида к размножению посредством фрагментации зависит от размера части, которая отламывается, в то время как при бинарном делении особь отделяется и образует двух особей одинакового размера.
- Почкование может привести к появлению полностью новой взрослой особи, которая формируется отдельно от исходного тела или может оставаться прикрепленной к исходному телу.
- Наблюдаемый у беспозвоночных и некоторых позвоночных, партеногенез дает потомство, которое может быть гаплоидным или диплоидным.
- Половое размножение, производство потомства с новой комбинацией генов, может также включать гермафродитизм, при котором организм может самооплодотворяться или спариваться с другим особью того же вида.

Ключевые термины
- бинарное деление : процесс, при котором клетка делится бесполым путем с образованием двух дочерних клеток
- гермафродитизм : наличие половых органов у обоих полов
- партеногенез : форма бесполого размножения, при которой рост и развитие эмбрионов происходят без оплодотворения
Способы размножения: бесполое и половое
Бесполое воспроизведение
Бесполое размножение дает потомство, которое генетически идентично родителю, поскольку все потомство является клонами первоначального родителя. Этот тип воспроизводства встречается у прокариотических микроорганизмов (бактерий) и у некоторых эукариотических одноклеточных и многоклеточных организмов. Животные могут размножаться бесполым путем путем деления, почкования, фрагментации или партеногенеза.
Этот тип воспроизводства встречается у прокариотических микроорганизмов (бактерий) и у некоторых эукариотических одноклеточных и многоклеточных организмов. Животные могут размножаться бесполым путем путем деления, почкования, фрагментации или партеногенеза.
Деление
Деление, также называемое бинарным делением, происходит у прокариотических микроорганизмов и у некоторых беспозвоночных, многоклеточных организмов. После периода роста организм разделяется на два отдельных организма. Некоторые одноклеточные эукариотические организмы подвергаются бинарному делению путем митоза.У других организмов часть особи отделяется, образуя вторую особь. Этот процесс происходит, например, у многих астероидных иглокожих через расщепление центрального диска. Некоторые морские анемоны и некоторые коралловые полипы также размножаются путем деления.
Рисунок \ (\ PageIndex {1} \): Деление : Коралловые полипы размножаются бесполым путем деления, когда организм разделяется на два отдельных организма.
Почкование
Почкование — это форма бесполого размножения, которая возникает в результате разрастания части клетки или области тела, приводящей к разделению от исходного организма на двух особей.Почкование обычно происходит у некоторых беспозвоночных животных, таких как кораллы и гидры. У гидр образуется почка, которая развивается во взрослую особь, которая отрывается от основного тела; тогда как у кораллов бутон не отделяется, а размножается как часть новой колонии.
Рисунок \ (\ PageIndex {1} \): Бутонирование : Гидра размножается бесполым путем за счет бутонизации, при этом образуется почка, которая развивается во взрослую особь и отделяется от основного тела.Фрагментация
Фрагментация — это разделение тела на две части с последующей регенерацией.Если животное способно к фрагментации и часть достаточно большая, отдельная особь вырастет заново.
Многие морские звезды размножаются бесполым путем путем фрагментации. Например, если рука отдельной морской звезды сломана, она возродит новую морскую звезду. Известно, что работники рыболовства пытались убить морских звезд, поедающих их моллюсков или устриц, разрезая их пополам и бросая обратно в океан. К несчастью для рабочих, каждая из двух частей может регенерировать новую половину, в результате чего вдвое больше морских звезд охотятся на устриц и моллюсков.Фрагментация также наблюдается у кольчатых червей, турбеллярий и порифер.
Известно, что работники рыболовства пытались убить морских звезд, поедающих их моллюсков или устриц, разрезая их пополам и бросая обратно в океан. К несчастью для рабочих, каждая из двух частей может регенерировать новую половину, в результате чего вдвое больше морских звезд охотятся на устриц и моллюсков.Фрагментация также наблюдается у кольчатых червей, турбеллярий и порифер.
Обратите внимание, что при фрагментации обычно наблюдается заметная разница в размере особей, тогда как при делении образуются две особи примерно одинакового размера.
Партеногенез
Партеногенез — это форма бесполого размножения, при которой яйцеклетка превращается в полноценную особь без оплодотворения.Полученное потомство может быть гаплоидным или диплоидным, в зависимости от процесса и вида. Партеногенез происходит у беспозвоночных, таких как водяные блохи, коловратки, тли, палочники, некоторые муравьи, осы и пчелы. Пчелы используют партеногенез для производства гаплоидных самцов (трутней) и диплоидных самок (рабочих). Если яйцо оплодотворяется, получается матка. Пчелиная матка контролирует воспроизводство пчел в улье, чтобы регулировать тип производимой пчелы.
Партеногенез происходит у беспозвоночных, таких как водяные блохи, коловратки, тли, палочники, некоторые муравьи, осы и пчелы. Пчелы используют партеногенез для производства гаплоидных самцов (трутней) и диплоидных самок (рабочих). Если яйцо оплодотворяется, получается матка. Пчелиная матка контролирует воспроизводство пчел в улье, чтобы регулировать тип производимой пчелы.
Некоторые позвоночные животные, такие как рептилии, земноводные и рыбы, также размножаются посредством партеногенеза.Хотя партеногенез чаще встречается у растений, он наблюдался у видов животных, которые были разделены по полу в наземных или морских зоопарках. Два дракона Комодо, капотоголовая акула и черноперая акула дали партеногенное потомство, когда самки были изолированы от самцов.
Половое размножение
Половое размножение — это сочетание (обычно гаплоидных или имеющих один набор непарных хромосом) репродуктивных клеток от двух особей с образованием третьего (обычно диплоидного или имеющего пару хромосом каждого типа) уникального потомства. Половое размножение дает потомство с новыми комбинациями генов. Это может быть адаптивным преимуществом в нестабильных или непредсказуемых средах. Как люди, мы привыкли думать о животных как о двух разных полах, мужском и женском, определенных при зачатии. Однако в животном мире существует множество вариаций на эту тему.
Половое размножение дает потомство с новыми комбинациями генов. Это может быть адаптивным преимуществом в нестабильных или непредсказуемых средах. Как люди, мы привыкли думать о животных как о двух разных полах, мужском и женском, определенных при зачатии. Однако в животном мире существует множество вариаций на эту тему.
Гермафродитизм
Гермафродитизм встречается у животных, у которых одна особь имеет как мужские, так и женские репродуктивные части. Беспозвоночные, такие как дождевые черви, слизни, ленточные черви и улитки, часто являются гермафродитами.Гермафродиты могут самооплодотворяться или спариваться с другими представителями своего вида, оплодотворяя друг друга и производя потомство. Самооплодотворение часто встречается у животных с ограниченной подвижностью или неподвижных, таких как ракушки и моллюски.
Фрагментация рибосомной РНК на короткие РНК (rRF) модулируется в зависимости от пола и популяции в зависимости от происхождения | BMC Biology
Обзор конвейера анализа rRF RNA-seq
После удаления всех образцов, поступивших из учреждения «шесть» (см.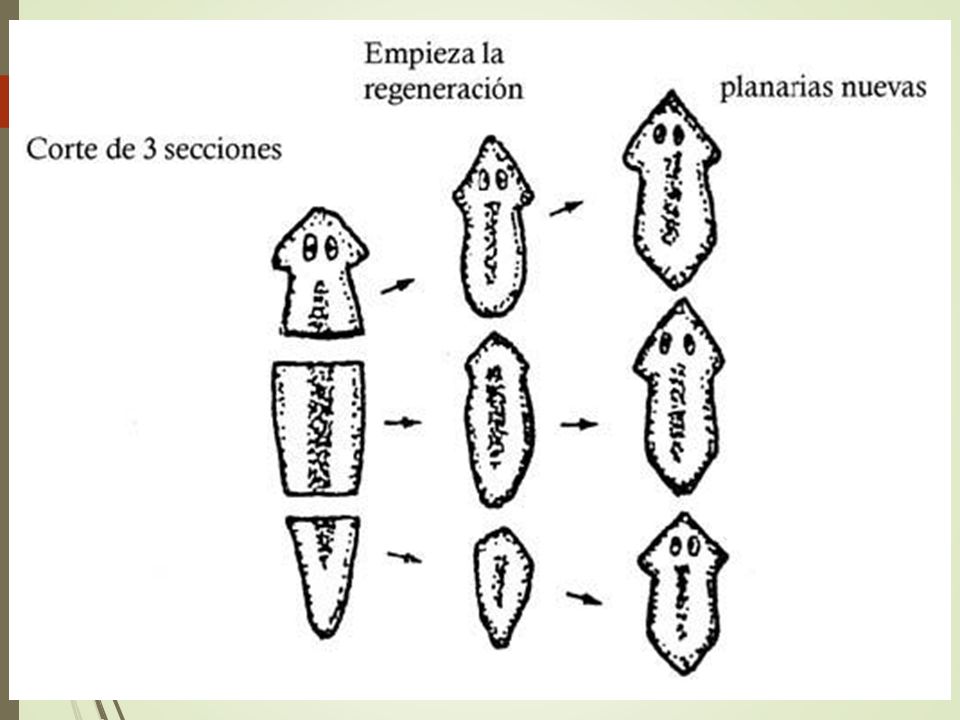 Раздел «Методы»), осталось 434 набора данных RNA-seq из проекта «1000 геномов» (1 кг) [32] для нашего последующего анализа.Наборы данных представляют людей, принадлежащих к пяти группам населения: жители штата Юта с северным и западноевропейским происхождением (CEU), финны в Финляндии (FIN), британцы в Англии и Шотландии (GBR), Toscani в Италии (TSI) и Yoruba в Ибадане, Нигерия (YRI). На рис. 1a мы представляем наглядное изображение конвейера, который мы использовали для анализа 434 наборов данных LCL. Конвейер начинается с грубого, детерминированного и исчерпывающего картирования всех секвенированных чтений на 6 рРНК. Сохраняются только те чтения, которые точно соответствуют рРНК.Конвейер также отслеживает rRF, которые охватывают 5′- или 3′-конец шести эталонных рРНК (см. Раздел «Методы»). Любые rRF, численность которых не удовлетворяет порогу для конкретного образца, определенному Threshold-seq [35] (см. Раздел «Методы»), отбрасываются. На рисунке 1b в столбце 3 показано количество уникальных rRF, продуцируемых каждой рРНК, прошедшей пороговую секцию.
Раздел «Методы»), осталось 434 набора данных RNA-seq из проекта «1000 геномов» (1 кг) [32] для нашего последующего анализа.Наборы данных представляют людей, принадлежащих к пяти группам населения: жители штата Юта с северным и западноевропейским происхождением (CEU), финны в Финляндии (FIN), британцы в Англии и Шотландии (GBR), Toscani в Италии (TSI) и Yoruba в Ибадане, Нигерия (YRI). На рис. 1a мы представляем наглядное изображение конвейера, который мы использовали для анализа 434 наборов данных LCL. Конвейер начинается с грубого, детерминированного и исчерпывающего картирования всех секвенированных чтений на 6 рРНК. Сохраняются только те чтения, которые точно соответствуют рРНК.Конвейер также отслеживает rRF, которые охватывают 5′- или 3′-конец шести эталонных рРНК (см. Раздел «Методы»). Любые rRF, численность которых не удовлетворяет порогу для конкретного образца, определенному Threshold-seq [35] (см. Раздел «Методы»), отбрасываются. На рисунке 1b в столбце 3 показано количество уникальных rRF, продуцируемых каждой рРНК, прошедшей пороговую секцию.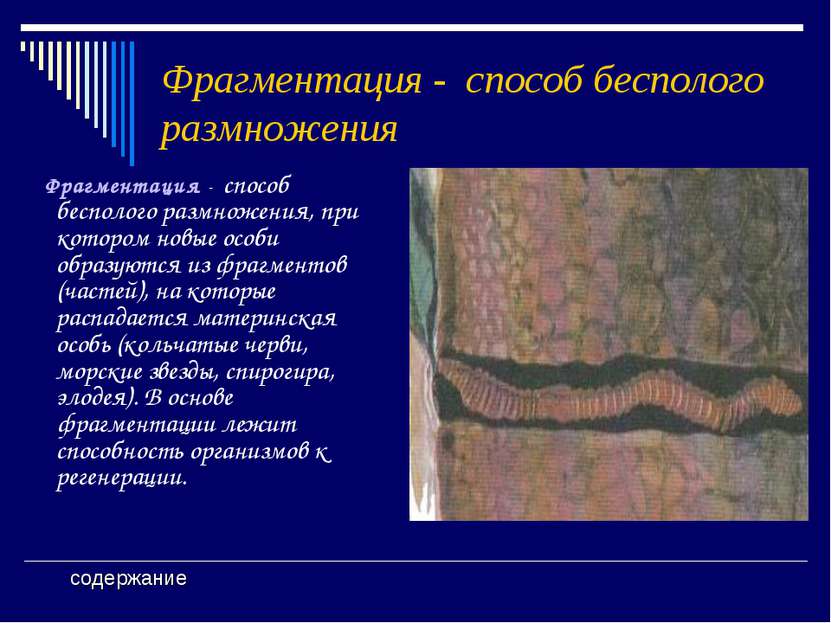 В дополнение к Threshold-seq, мы нормализуем количество каждого rRF до числа операций чтения на миллион (RPM), а для дополнительной строгости мы устанавливаем более строгий минимальный порог ≥ 10 RPM.На рисунке 1b столбец 5 показывает количество уникальных rRFs, продуцируемых каждой рРНК, которые проходят порог отсечки 10 об / мин. На заключительном этапе фильтрации мы отбрасывали те rRFs, экземпляры которых за пределами пространства рРНК превышали 2% от всех их геномных экземпляров, и отношение сигнал / шум (S / N) ≥ 50 (см. Раздел «Методы»).
В дополнение к Threshold-seq, мы нормализуем количество каждого rRF до числа операций чтения на миллион (RPM), а для дополнительной строгости мы устанавливаем более строгий минимальный порог ≥ 10 RPM.На рисунке 1b столбец 5 показывает количество уникальных rRFs, продуцируемых каждой рРНК, которые проходят порог отсечки 10 об / мин. На заключительном этапе фильтрации мы отбрасывали те rRFs, экземпляры которых за пределами пространства рРНК превышали 2% от всех их геномных экземпляров, и отношение сигнал / шум (S / N) ≥ 50 (см. Раздел «Методы»).
Конвейер анализа rRF выявляет множество уникальных фрагментов. a Рабочий процесс конвейера. b Таблица, показывающая количество уникальных rRF, которые картируются с каждой рРНК, и количество rRF на единицу длины для 434 образцов LCL.РРНК были дополнены по 50 нт с каждой стороны. Обратите внимание, что многие rRF имеют частоту ≥ 10 об / мин. c Распределение количества образцов LCL (из 434), в которых данный isomiR, tRF или rRF может быть обнаружен при численности ≥ 10 об / мин. Коробчатые диаграммы сгруппированы по геному происхождения, ядерному или митохондриальному. d Коробчатые диаграммы показывают распределение среднего содержания (в об / мин) для молекул, принадлежащих к каждой категории. В нашем последующем анализе рассматривались только молекулы с содержанием ≥ 10 об / мин.Коробчатые диаграммы сгруппированы по геному происхождения. c , d Ширина прямоугольников пропорциональна количеству уникальных молекул в каждой категории, а горизонтальные полосы в каждом прямоугольнике представляют собой медианное значение числа оборотов в минуту
Коробчатые диаграммы сгруппированы по геному происхождения, ядерному или митохондриальному. d Коробчатые диаграммы показывают распределение среднего содержания (в об / мин) для молекул, принадлежащих к каждой категории. В нашем последующем анализе рассматривались только молекулы с содержанием ≥ 10 об / мин.Коробчатые диаграммы сгруппированы по геному происхождения. c , d Ширина прямоугольников пропорциональна количеству уникальных молекул в каждой категории, а горизонтальные полосы в каждом прямоугольнике представляют собой медианное значение числа оборотов в минуту
Все шесть рРНК производят множество обильных коротких фрагментов
Мы находим, что все шесть рРНК продуцируют множество обильных rRF. В столбце 3 на рис. 1b показано количество уникальных rRFs от каждой рРНК, которые превышают адаптивный порог для конкретного образца, определенный алгоритмом Threshold-seq [35] (см. Раздел «Методы») в 434 проанализированных наборах данных.В столбце 4 показано то же число, нормированное на длину родительской рРНК. Также на той же панели указано количество уникальных rRF (столбец 5), которые сопоставляются с каждой рРНК и превышают пороговое значение 10 об / мин. В столбце 6 показаны те же числа после того, как они были нормализованы по разной длине рРНК. Только 18 981 rRF выдерживают этот строгий порог для всех рРНК. Как также можно видеть на рис. 1b, для всех шести рРНК количество rRFs на единицу длины находится в диапазоне от 6,26 до 9,87, т.е. попадает в узкое окно значений, что предполагает, что rRFs обрабатываются с постоянной скоростью.В целях сравнения мы также исследовали isomiR (используя то же отображение грубой силы, что и с rRF) и tRF (используя алгоритм MINTmap [36]) и вычислили их нормализованное количество RPM. Для isomiR, tRF и rRF мы использовали в качестве знаменателя общее количество секвенированных считываний каждого образца. Было 1522 tRF и 1212 изомиР, численность которых составляла не менее 10 об / мин.
Также на той же панели указано количество уникальных rRF (столбец 5), которые сопоставляются с каждой рРНК и превышают пороговое значение 10 об / мин. В столбце 6 показаны те же числа после того, как они были нормализованы по разной длине рРНК. Только 18 981 rRF выдерживают этот строгий порог для всех рРНК. Как также можно видеть на рис. 1b, для всех шести рРНК количество rRFs на единицу длины находится в диапазоне от 6,26 до 9,87, т.е. попадает в узкое окно значений, что предполагает, что rRFs обрабатываются с постоянной скоростью.В целях сравнения мы также исследовали isomiR (используя то же отображение грубой силы, что и с rRF) и tRF (используя алгоритм MINTmap [36]) и вычислили их нормализованное количество RPM. Для isomiR, tRF и rRF мы использовали в качестве знаменателя общее количество секвенированных считываний каждого образца. Было 1522 tRF и 1212 изомиР, численность которых составляла не менее 10 об / мин.
Специфические rRF присутствуют во всех образцах, в то время как другие являются специфичными для группы
Учитывая, что все 434 образца принадлежат к одному и тому же типу клеток (иммортализованные B-клетки), мы хотели знать, повторяются ли rRF, которые отображаются на каждую из шести рРНК. по этим образцам.С этой целью и отдельно для каждого rRF мы подсчитали количество выборок (среди 434), в которых каждый rRF превысил строгий порог в 10 об / мин. Мы объединили результаты для rRF из той же рРНК в единую коробчатую диаграмму (рис. 1c). В целях сравнения мы также создали соответствующие коробчатые диаграммы для 1522 tRF и 1212 isomiR, которые также удовлетворяют порогу 10 об / мин. Полученные распределения показаны рядом на рис. 1c. Ширина каждой прямоугольной диаграммы с надрезом пропорциональна количеству уникальных фрагментов в пределах соответствующего типа короткой нкРНК; это число также указано на каждой диаграмме.
по этим образцам.С этой целью и отдельно для каждого rRF мы подсчитали количество выборок (среди 434), в которых каждый rRF превысил строгий порог в 10 об / мин. Мы объединили результаты для rRF из той же рРНК в единую коробчатую диаграмму (рис. 1c). В целях сравнения мы также создали соответствующие коробчатые диаграммы для 1522 tRF и 1212 isomiR, которые также удовлетворяют порогу 10 об / мин. Полученные распределения показаны рядом на рис. 1c. Ширина каждой прямоугольной диаграммы с надрезом пропорциональна количеству уникальных фрагментов в пределах соответствующего типа короткой нкРНК; это число также указано на каждой диаграмме.
Несмотря на то, что образцы представляют один тип клеток, прямоугольные диаграммы на рис. 1c показывают, что основная часть rRFs, связанных с каждой из шести рРНК, появляется только в части образцов. Распределение rRF для 18S и 28S особенно примечательно: хотя эти две рРНК продуцируют множество rRF (4545 и 9263 соответственно), большинство из них присутствует только в небольшой части из 434 образцов (рис.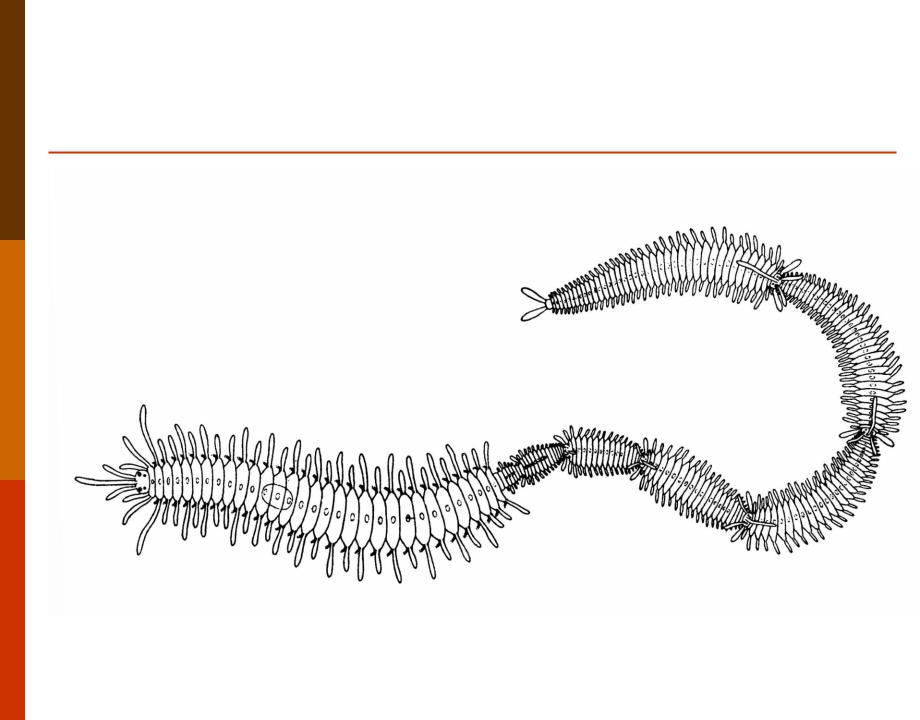 1c). Это предполагает зависимость от других переменных, о чем мы поговорим ниже.
1c). Это предполагает зависимость от других переменных, о чем мы поговорим ниже.
Важно подчеркнуть, что эти наблюдения относительно численности rRF аналогичны нашим предыдущим результатам для isomiRs и tRFs [23, 25, 26, 29], которые резюмируются соответствующими распределениями на рис. 1c. Например, обратите внимание, что типичная isomiR существует только в 21,9% из 434 образцов (медиана = 95 образцов). Из всех имеющихся изомий только небольшое количество присутствует в большинстве из 434 образцов. Аналогичное наблюдение можно сделать для tRF: здесь, опять же, только очень небольшое количество tRF появляется в большинстве из 434 выборок.
rRFs столь же многочисленны, как и ранее установленные типы ncRNA.
Затем мы исследовали численность rRFs во всех 434 наборах данных LCL, сопоставив их нормализованные количества с таковыми изомиРов и tRFs (рис. 1d). Хотя значения медианного содержания для всех трех типов РНК по существу одинаковы, стоит отметить, что содержания отдельных РНК охватывают очень широкий диапазон: от 10 до более 16000 об / мин с rRF от 10 до ~ 8000 об / мин. . Выше мы упоминали, что количество уникальных rRF зависит от родительской рРНК (рис.1б). Однако ясно, что количество уникальных rRF не коррелирует с численностью rRF. В самом деле, как видно из рис. 1d, rRFs и tRFs имеют сходные распределения численности, даже несмотря на то, что их соответствующие молекулярные категории содержат отчетливо разное количество фрагментов. Обратите также внимание на то, что все изображенные категории имеют выбросы с чрезвычайно высокими значениями частоты вращения.
. Выше мы упоминали, что количество уникальных rRF зависит от родительской рРНК (рис.1б). Однако ясно, что количество уникальных rRF не коррелирует с численностью rRF. В самом деле, как видно из рис. 1d, rRFs и tRFs имеют сходные распределения численности, даже несмотря на то, что их соответствующие молекулярные категории содержат отчетливо разное количество фрагментов. Обратите также внимание на то, что все изображенные категории имеют выбросы с чрезвычайно высокими значениями частоты вращения.
Важны как длина, так и состав последовательности rRF.
Во всех наших анализах мы исключили секвенированные считывания короче 16 нуклеотидов (нуклеотидов).Это связано с тем, что такие короткие последовательности, которые могут картироваться, например, с тРНК или мРНК, также с большей вероятностью будут картироваться в другом месте генома [37, 38]. Эта неоднозначность затрудняет точное определение истинного геномного происхождения соответствующих РНК.
Образцы 434 LCL содержат множество обильных фрагментов длиной от 16 до 33 нт, которые отображаются на шесть эталонных рРНК. Чтобы исследовать, какие из этих предполагаемых rRFs также можно найти за пределами «пространства рРНК» (см. Раздел «Методы»), мы провели поиск каждой такой последовательности по всему геному, используя грубый детерминированный подход.Это позволило нам рассчитать для каждого k -mer следующее отношение «сигнал-шум» (S / N) (см. Раздел «Методы»): количество экземпляров k-mer внутри пространства рРНК, больше количество экземпляров k-mer за пределами пространства рРНК . В дальнейшем были рассмотрены только k -меров с S / N ≥ 50.
Чтобы исследовать, какие из этих предполагаемых rRFs также можно найти за пределами «пространства рРНК» (см. Раздел «Методы»), мы провели поиск каждой такой последовательности по всему геному, используя грубый детерминированный подход.Это позволило нам рассчитать для каждого k -mer следующее отношение «сигнал-шум» (S / N) (см. Раздел «Методы»): количество экземпляров k-mer внутри пространства рРНК, больше количество экземпляров k-mer за пределами пространства рРНК . В дальнейшем были рассмотрены только k -меров с S / N ≥ 50.
Мы обнаружили, что состав последовательностей 5S rRFs довольно уникален среди шести рРНК. Даже 5S rRFs только с 16 нуклеотидами имеют S / N ≥ 63. S / N для всех комбинаций длины и источника родительской рРНК перечислены в Таблице 1.Как показано в таблице, минимальная длина rRF, удовлетворяющая этому отсечению, различается для каждой из шести рРНК. Минимальные длины следующие: 16 нт для 5S; 18 нт для 12S, 18S и 5,8S; и 19 нт для 16S и 28S. В последующих анализах использовались только те rRF, которые удовлетворяли этим ограничениям по минимальной длине и соответствующее отношение S / N ≥ 50.
В последующих анализах использовались только те rRF, которые удовлетворяли этим ограничениям по минимальной длине и соответствующее отношение S / N ≥ 50.
rRF возникают из «горячих точек» в пределах каждого участка рРНК.
Затем мы попытались определить, где различные rRFs отображаются вдоль каждой рРНК.Мы обнаружили, что rRFs могут возникать из любой части диапазона родительской рРНК. Чтобы соответствовать обозначениям, используемым для tRF [26,27,28,29,30,31, 36], мы называем те rRF, которые возникают из 5′-конца рРНК, как «5′-rRFs, »Те, которые возникают изнутри рРНК как« i-rRF », те, которые возникают из 3′-конца рРНК как« 3′-rRF », и те, которые охватывают 5′- или 3′-концы рРНК как «x-rRFs». Последовательности из 16 279 rRF, которые выдерживают строгие пороговые значения (Threshold-seq, ≥ 10 об / мин, ограничения длины), перечислены в дополнительном файле 4.Для каждого rRF мы указываем его тип (5′-rRF, i-rRF, 3′-rRF или конец, пересекающий x-rRF). Мы также перечисляем «номерные знаки» каждого RFR, распространяющиеся на RFR схему маркировки, которую мы ввели для tRF в 2016 году [27] и использовали для маркировки tRF, которые в настоящее время находятся в базе MINT [28]. Схема маркировки номерных знаков гарантирует уникальную метку для каждого rRF и , наоборот, , и особенно подходит для маркировки rRF, учитывая многочисленные копии, которые они имеют в геноме.
Тепловые карты на рис.2a – c показывают несколько примеров относительной численности rRFs, которые картируются в выделенные области 28S, 16S и 5S рРНК. В каждом случае мы сгруппировали образцы из одной и той же популяции в последовательные ряды, которые мы раскрасили по-разному для каждой популяции: CEU — фиолетовый; FIN — оранжевый; GBR — голубой; TSI — серый; и YRI — желтый. К каждой тепловой карте примыкают прямоугольные диаграммы, показывающие распределение начальных и конечных местоположений для показанных rRF.
Рис. 2Примеры горячих точек, генерирующих РФЧ. a — c Показаны области из 28S рРНК ( a ), 16S рРНК ( b ) и 5S рРНК ( c ). Каждая тепловая карта отображает относительное количество rRF во всех 434 образцах LCL, отображаемых в интересующей области. На каждой смежной прямоугольной диаграмме отображается распределение начальных и конечных положений rRF для области, показанной на тепловой карте. Тепловые карты масштабируются по строкам (выборкам). d Каждая эталонная рРНК (GenBank) показана в виде прямоугольника со сплошным синим контуром.Сплошная черная вертикальная линия обозначает положение 1 референсных рРНК. Наблюдаемые конечные точки x-rRF, которые отображаются непосредственно выше или ниже эталонных последовательностей рРНК, представлены желтыми прямоугольниками. Места расположения эталонных транскриптов рРНК, которые примыкают к номинальным конечным точкам и не имеют сопоставления rRFs с ними, показаны пустыми прямоугольниками с пунктирными линиями. Не в масштабе
Можно легко сделать несколько наблюдений. Например, на рис. 2а показано, что 28S продуцирует 5′-rRFs, которые начинаются в положении +1 во всех 434 образцах (все пять популяций и оба пола).Примечательно, что во многих из 434 образцов 28S рРНК также продуцирует x-rRF, которые начинаются в положении -1 (данные не показаны). Другое наблюдение состоит в том, что область 16S, показанная на рис. 2b, продуцирует i-rRFs преимущественно в четырех европейских популяциях (CEU, FIN, GBR и TSI), но не в африканской популяции (YRI).
3′-область 5S рРНК показана на рис. 2c. Многие из rRFs, которые отображаются на эту часть рРНК, имеют длину 18 или 19 нуклеотидов. Из всех rRFs, которые картируются с 5S рРНК, ~ 30% начинаются в позиции 103 и заканчиваются в основном в позиции 120 или позиции 121 (i-rRFs и 3′-rRFs соответственно).
Конечные точки rRF не всегда совпадают с конечными точками родительских рРНК.
Наш анализ конечных точек rRFs привел к любопытному наблюдению. Мы обнаружили, что ряд наблюдаемых rRF с численностью ≥ 10 об / мин могут либо выходить за границы нескольких эталонных рРНК, либо вообще избегать их (см. Раздел «Методы» для эталонных идентификаторов). Рисунок 2d суммирует это наблюдение. Прямоугольники со сплошной линией обозначают шесть транскриптов рРНК. Положения, которые присутствуют в анализируемых rRF и соответствуют положениям за контрольными границами рРНК (выше или ниже), обозначены желтым, где желтые прямоугольники представляют дополнительные нуклеотиды.Позиции, проксимальные к 5′- или 3′-концам рРНК и не присутствующие ни в одном из проанализированных rRF, обозначены пунктирными линиями, тогда как пустые прямоугольники обозначают пропуск нуклеотидов.
Здесь стоит сделать несколько замечаний. Во-первых, rRF, продуцируемые из 5.8S рРНК в LCL, начинаются на 6 нт выше номинального 5′-конца, указанного в GenBank. Интересно, что известно, что существует две изоформы 5.8S: более короткая из двух указана в записи GenBank NR_145819.1 (см. Раздел «Методы»), тогда как более длинный простирается на несколько нуклеотидов вверх (рис. 2d). Недавно сообщалось [19], что в предыдущих исследованиях более короткая из двух изоформ была наиболее распространенной. Хотя это вполне может иметь место и в LCL — наш анализ изучал короткие и не длинные наборы данных RNA-seq — наши данные показывают, что более длинная изоформа производит наибольшее количество 5′-rRFs. В дополнение к вариантам транскрипта 5.8S существуют rRF, продуцируемые из 28S и 5S рРНК, которые начинаются на один и два нуклеотида выше номинальных 5′-концов рРНК, соответственно.Похоже, что rRF от 12S до избегают первой позиции этой рРНК: вместо этого все они начинаются со второй позиции. Для 18S ни один из проанализированных rRFs не включает ни одно из последних 24 положений рРНК: действительно, крайний правый rRF заканчивается в положении 1845, тогда как длина этой рРНК составляет 1869 нт. Наконец, мы отмечаем, что и 12S, и 16S продуцируют x-rRFs, которые охватывают соответствующие аннотированные 3′-концы и заимствуют нуклеотиды от нижестоящих тРНК ValTAC и тРНК LeuTAA , соответственно.
Длина и численность преобладающих rRFs различаются для каждой рРНК.
Далее мы попытались определить, квантованы ли длины rRFs. Линейные графики на рис. 3 показывают, отдельно для каждой из шести рРНК, распределения длин rRF, которые выживают при использовании фильтров численности, S / N и длины. Как можно видеть, картированные rRFs имеют профили длины, специфичные для каждой из шести рРНК. Митохондриальные 12S и 16S рРНК генерируют в основном более длинные rRF (26-29 нт).В одной трети образцов 12S также дает более короткие (20–21 нт) rRF. 5S рРНК продуцирует короткие (18-19 нуклеотидов), а также промежуточные (24-25 нуклеотидов) и длинные rRF (32-33 нуклеотида). Однако обратите внимание, что в тех образцах, где преобладают короткие rRF, промежуточные и более длинные rRF обычно отсутствуют, и наоборот .
Рис. 3Профили длины и численности rRFs повторяются в глобальном масштабе и зависят от популяции. На линейных графиках показано отношение (число оборотов в минуту каждой длины / общее число оборотов в минуту) для каждой длины rRF и каждой из 6 рРНК, где черная кривая представляет среднее отношение длин по всем 434 образцам LCL, а серая область представляет собой стандартное отклонение отношения .Каждая тепловая карта показывает отношения длин rRF для каждой длины последовательности отдельно для мужских (207) и женских (227) образцов. Тепловые карты иерархически сгруппированы по строкам. Темно-зеленый соответствует наименьшей численности ( z- баллов — 3), а темно-пурпурный — наибольшему обилию ( z- баллов + 3)
rRF, которые продуцируются из трех производных 45S рРНК ( 18S, 5.8S и 28S) довольно интригуют. Что касается длины, они охватывают широкий диапазон. 5.8S рРНК продуцирует в основном rRF длиной 18 нт (35.7% всех rRFs картируются на эту рРНК). 18S рРНК продуцирует rRF любой длины. Однако наблюдается очевидное бимодальное поведение: в тех образцах, где преобладают 18S rRF с длиной от 18 до 23 нт включительно, более длинные rRF с длинами 29 и 30 нт отсутствуют, и наоборот . Наконец, очень немногие rRF с промежуточной длиной (21–26 нт) отображаются в 28S. Интересно, что примерно в одной трети образцов практически нет rRF короче 28 нт, которые соответствуют 28S. Ниже мы еще раз рассмотрим эти распределения, исследуя их отдельно для различных подмножеств из 434 выборок.
Профили длин rRF показывают сходство и различия между полами и популяциями.
По замыслу, 434 выборки были отобраны так, чтобы охватить два измерения: пол и происхождение населения. В частности, образцы представляют пять групп населения: из них четыре группы являются европейскими популяциями (CEU, FIN, GBR, TSI), а пятая группа — африканским населением (YRI). В популяции мужчины и женщины представлены равномерно. На рисунке 3 показаны тепловые карты распределения длины различных rRF, отдельно для каждого пола и помеченные популяциями.Схема цветовой кодировки для пяти популяций такая же, как на рис. 2. Образцы иерархически сгруппированы с использованием профилей длины каждого образца. В целом эти тепловые карты не выявляют какой-либо сильной зависимости от пола или происхождения населения. Тем не менее, тщательный осмотр показывает, что такие различия действительно существуют.
На самом деле, самки YRI постоянно производят больше 33-меров и меньше 26- / 27-меров из 16S рРНК по сравнению с другими четырьмя популяциями, что указывает на популяционно-специфический сигнал, который различает европейцев (CEU, FIN , GBR, TSI) и африканское (YRI) население.С другой стороны, европейские самцы стабильно производят больше 26/27 меров, чем 33-меров.
Многочисленные rRF по-разному распространены по полу и происхождению из популяции
Несмотря на то, что длины rRF, отображаемые на различные рРНК, в значительной степени согласованы, мы хотели знать, проявляют ли количества этих rRFs различия, которые связаны с полом. или для конкретной популяции. Для этого мы использовали два разных подхода: SAM и PLS-DA (см. Раздел «Методы»).При пороге FDR, равном 0,01, SAM идентифицировал 3103 уникальных rRF, которые дифференциально многочисленны, по крайней мере, при одном попарном сравнении популяций для выборок, принадлежащих к одному полу. SAM также идентифицировал 88 уникальных rRF, которые по-разному многочисленны по полу для выборок, принадлежащих к одной и той же популяции. Второй метод, PLS-DA, выявил 1697 уникальных rRF, специфичных для популяции (сравнение однополых выборок), и 2264 уникальных rRF, специфичных для пола (сравнение одних и тех же выборок населения). Рисунок 4c и дополнительный файл 1: Рисунок S1A показывают попарные rRF, которые обнаруживаются SAM и PLS-DA для каждой рРНК между всеми популяциями и двумя полами.
Рис. 4Многочисленные rRFs имеют разную численность в зависимости от пола и происхождения населения. a Количество случаев, когда SAM идентифицировал rRF как дифференциально распространенный, показано желтым (FDR ≤ 0,01). Число случаев, когда PLS-DA идентифицировал rRF как дифференциально распространенный, показано зеленым (VIP ≥ 1,5). Пересечение двух кружков показывает количество сравнений, в которых как SAM, так и PLS-DA обнаружили, что один и тот же rRF является дифференциально распространенным. Также показан индекс Жаккара для сравнений, которые, как было установлено, являются дифференциально многочисленными с помощью двух методов. b Индекс Жаккара для каждой рРНК наносят на график в зависимости от медианного числа оборотов в минуту iDAR. c Для iDAR было нанесено log 2 -кратное изменение как функция значимости ( q -значение) для каждой из 10 комбинаций популяций и отдельно для мужчин и женщин. Ключ внизу показывает распределение значений q в каждом из 10 интервалов, которые представляют собой сравнение популяции с популяцией
Для строгости мы пересекли две коллекции дифференциально распространенных rRF, в результате чего осталось 549 уникальных популяций. специфические rRF и 39 уникальных специфичных для пола rRF (данные не показаны).Мы называем rRF, которые SAM и PLS-DA определяют как дифференциально распространенные в данном сравнении, как «пересечение дифференциально распространенных rRF» или «iDAR». Путем вычисления индекса Жаккара для сравнений, полученных из SAM и PLS-DA отдельно для каждой рРНК, мы обнаружили, что rRF 5S и 5.8S имеют самые высокие индексы (0,32 и 0,34, соответственно) для попарно специфичных для популяции . сравнения (рис. 4а). 5.8S также имеет наивысший индекс Жаккара (0,17) для парных сравнений с учетом пола (дополнительный файл 1: рисунок S1A).На рисунке 4b показано, что 5.8S производит iDAR с высоким медианным числом оборотов в минуту (31,4), которые также участвуют в сравнении с наивысшим индексом Жаккара (0,34), показанным красным прямоугольником на рисунке 4a. Между тем, 5S производит iDAR с третьим по величине средним значением RPM (19,4), которые также участвуют в сравнении со вторым по величине индексом Жаккара (0,32), показанным красным прямоугольником на рисунке 4a. Кроме того, на рисунке S1B показано, что iDAR для конкретного пола из 5.8S имеют второй по величине средний RPM (48,4) и участвуют в сравнении с наивысшим индексом Жаккара (0.17). Вместе это указывает на то, что rRFs из 5.8S рРНК могут быть специфичными для популяции и пола.
Когда мы смотрим на дифференциальную численность iDAR, сразу становятся очевидными резкие различия по полу и происхождению популяций (рис. 4c, дополнительный файл 1: рис. S1C). Чтобы визуализировать кратные различия iDAR, мы построили логарифм 2 кратных изменений для всех парных сравнений и отдельно для каждого пола (рис. 4c). Беглый взгляд на эту панель выявляет поразительную общность: независимо от того, сколько rRF являются iDAR в каждом случае, все шесть рРНК продуцируют множество rRF с постоянно более высокой численностью в четырех европейских популяциях (CEU, FIN, GBR и TSI), чем в африканских популяциях. население (YRI).Это верно как для мужчин, так и для женщин.
Более того, внутри четырех европейских популяций существуют популяционные различия. Например, популяция CEU продуцирует rRFs из 5S и 5.8S рРНК в меньшем количестве, чем популяции FIN, GBR и TSI, как у мужчин, так и у женщин. Самцы CEU производят больше 18S rRF, чем их аналоги FIN, GBR и TSI, тогда как самки CEU производят различные rRF 18S по сравнению с другими европейскими самками (FIN, GBR и TSI). Интересно, что и самцы, и самки из популяции CEU продуцируют множественные «активированные» и «подавленные» 28S rRFs по сравнению с другими четырьмя популяциями (FIN, GBR, TSI и YRI).
Мы также более подробно изучили дифференциальную численность 88 уникальных iDAR, зависящих от пола (дополнительный файл 1: Рисунок S1). Примечательно, что почти все эти rRFs обнаруживают более высокую численность у самок, чем у самцов. Большинство этих rRFs отображается на 5.8S рРНК (16 уникальных iDAR). Удивительно, но 5.8S rRFs демонстрируют специфичные для пола различия в численности только в трех из четырех европейских популяций: CEU, GBR и TSI. Самки FIN и YRI производят больше iDAR из 16S, 18S и 5S, чем самцы.И один 28S iDAR, который зависит от пола, присутствует в большей степени у мужчин, чем у женщин в популяции GBR.
Половые и популяционные различия в численности rRF являются определяющими для группы.
rRF с самым высоким изменением численности — это 24-мерный GGGCUACGCCUGUCUGAGCGUCGC с 5.8S. Этот i-rRF отображается на 3′-первичный конец 5.8S рРНК (дополнительный файл 2: Рисунок S2) и оканчивается двумя нуклеотидами, отстающими от номинального транскрипта 5.8S. Этот i-rRF дифференциально представлен в следующих сравнениях: YRI / CEU (- 2.31 log 2 кратных изменений), YRI / FIN (- 2,37 log 2 кратных изменений), YRI / GBR (- 2,49 log 2 кратных изменений) и YRI / TSI (- 2,47 log 2 кратных изменений ) . На рис. 5a численность этого 24-мерного i-rRF стратифицирована коробчатыми диаграммами. Мы наблюдаем статистически значимую тенденцию, которая показывает, что все 4 европейские популяции (CEU, FIN, GBR и TSI), независимо от пола, производят больше этого фрагмента, чем мужчины и женщины из популяции YRI. Поскольку этот i-rRF по-разному распространен между YRI и всеми четырьмя европейскими популяциями, rRF может отражать географические различия на уровне континента (т.е.е., европейские и африканские РФС).
Рис. 5Независимые эксперименты подтверждают наличие rRF в LCL. a Коробчатые диаграммы показывают распространенность 5,8S 24-мерного i-rRF (GGGCUACGCCUGUCUGAGCGUCGC) в пяти популяциях LCL (всего 434 образца). Самцы представлены серыми прямоугольниками, а женщины — оранжевыми. Численность показана на прямоугольных диаграммах, где существует статистически значимая разница между каждой популяцией в соответствии с тестом Велча t .b Два нозерн-блоттинга, исследующих 5,8S 24-мерный i-rRF в РНК из девяти мужских и девяти женских LCL. Для каждого пола использовали по три клеточные линии от CEU, GBR и YRI. Каждая дорожка содержит 5 мкг РНК из каждого образца, а первая дорожка в каждом блоте содержит 5 пмоль 5,8S 24-мерной целевой кДНК (нижняя полоса). Полноразмерный 5S (ACGUCUGAUCUGAGGUCGCGU) является верхней полосой и служит для контроля загрузки. Полосы rRF были нормализованы к полосе контроля нагрузки 5S, состоящей из 121 н. Под каждым блотом приводится количественная оценка 5.Зонд 8S rRF, где CEU представлен фиолетовым цветом, GBR представлен голубым, а YRI представлен желтым
. Мы также наблюдаем, что 5.8S i-rRF 19-мерный UAAUGUGAAUUGCAGGACA, который продуцируется из центра 5.8S рРНК транскрипт, дифференциально распространен (значение p- = 0,03) в зависимости от пола (дополнительный файл 3: верхний рисунок S3A), что позволяет предположить, что это потенциально специфичный для пола rRF. Помимо значительных половых различий, мы распаковали полы по группам населения и обнаружили, что, хотя существует общая тенденция, специфичная для пола, существует также сильная популяционная зависимость (дополнительный файл 3: нижний рисунок S3A) с CEU. и GBR, имеющий статистически значимые различия между полами с p -значениями, равными 0.014 и 0,010 соответственно. Интересно, что в то время как самки CEU, GBR и TSI производят на больше этого 19-мерного i-rRF на , чем их соответствующие самцы, самки FIN фактически производят на меньше этого фрагмента, чем самцы FIN. Самцы и самки YRI производят фрагмент примерно в одинаковом количестве. Мы также исследовали численность 5,8S 21-мерного i-rRF UAAUGAGAAUUGCAGGACACA, который имеет такое же стартовое положение на 5′-конце, что и 19-мерный i-rRF, но содержит два дополнительных нуклеотида на 3′-конце UAAUGUGAAUUGCAGGACA (дополнительная файл 3: Рисунок S3).Интересно, что мы наблюдали, что так же, как и 19-мер, 21-мерный i-rRF является дифференциально многочисленным ( p -значение = 0,007) и сохраняет сходные популяционные и половые различия. Однако этот i-rRF присутствует в более низком количестве, чем 19-мерный i-rRF, что предполагает получение селективной последовательности.
Анализы независимо полученных образцов подтверждают присутствие rRF в LCL.
На рис. 1c мы видели, что только небольшое количество rRF, isomiR и tRF присутствует во всех образцах LCL, что указывает на специфичность фрагментов на основе подгрупп.Кроме того, рис. 5a показывает, что даже несмотря на то, что различная численность отдельных rRFs значительна между популяциями, они все же демонстрируют большие вариации численности «в группе». Имея это в виду, мы экспериментально исследовали результаты, специфичные для пола и популяции, с помощью LCL, полученных от 18 здоровых людей в рамках проекта 1KG, которые составили три популяции (CEU, GBR и YRI) (см. Раздел «Методы». для идентификаторов клеточных линий). Эти 18 образцов составляют , а не среди образцов, секвенированных в рамках проекта 1KG.
Мы провели нозерн-блоттинг с РНК из этих клеточных линий, используя в качестве зонда обратный комплемент 5.8S 24-мерного i-rRF GGGCUACGCCUGUCUGAGCGUCGC. Нозерн-блоты на фиг. 5b обеспечивают независимую экспериментальную проверку присутствия этого i-rRF. Имеется широкое распределение численности по 18 выборкам, включая выборки от самцов и самок (рис. 5b). Обратите внимание, как это распределение согласуется с результатами нашего компьютерного анализа (рис. 5a): действительно, хотя медианная численность этого i-rRF характерно и значительно выше для четырех европейских популяций по сравнению с африканской популяцией, фактические значения численности охватывают очень широкий диапазон в каждой популяции.Хотя мы не ожидаем, что разница в средних значениях будет зафиксирована в нескольких проанализированных нами выборках, мы можем увидеть тенденции, характерные для пола и населения. Количественная оценка северных блотов на фиг. 5b показывает, что в целом самки CEU и GBR продуцируют больше фрагмента, чем их коллеги-самцы. Самки YRI продуцируют такое же количество i-rRF при низкой численности. Кроме того, заметна популяционная тенденция различий между двумя европейскими женскими популяциями (CEU и GBR) и одной женской африканской популяцией (YRI) (рис.5b внизу). Эту тенденцию нельзя наблюдать среди нескольких проанализированных нами мужских выборок.
Идентичность и профили численности rRF могут также различаться в разных тканях.
Далее мы хотели узнать, существуют ли наиболее широко распространенные и очень распространенные rRF, которые мы наблюдаем в наборах данных 434 LCL, в других тканях человека. Для этой цели мы выбрали 80 наборов данных RNA-seq из образцов первичной увеальной меланомы (UVM) из Атласа генома рака (TCGA) [33], а также три набора данных RNA-seq клеток 293T и их три соответствующих 293T EV RNA-seq. наборы данных из общедоступных данных из Gen Expression Omnibus (GEO) [34] (см. раздел «Методы» и Дополнительный файл 4).На рисунке 6 отслеживаются наиболее распространенные rRF из наборов данных LCL в трех других коллекциях. Наиболее распространенные rRF, ACCGGUGCUGUAGGCUU an i-rRF и ACCGGGUGCUGUAGGCUU U a 3′-rRF, происходят от 3′-конца 5S рРНК и отличаются одним нуклеотидом. Эти i- и 3′-rRF имеют длину 18 нт и 19 нт, соответственно, и среднюю численность 1434 и 1334 об / мин соответственно. Как видно из рис. 6a, rRFs, которые соответствуют более коротким или более длинным экземплярам этих двух обильных rRFs, продуцируются либо в более низком количестве, либо не образуются вообще в коллекции LCL.Наборы данных UVM и 293T cell и 293T EV демонстрируют аналогичный шаблон распределения с наборами данных LCL. Однако количество этих двух rRF значительно ниже в наборах данных 293T EV и UVM. Более того, ни UVM, ни наборы данных ячеек 293T или 293T EV не содержат 17-мерных i-rRF ACCGGGUGCUGUAGGCU в сколько-нибудь заметном количестве.
Рис. 6Предварительные доказательства того, что профили rRF зависят от ткани. Коробчатые диаграммы показывают количество rRF на каждой длине для 434 образцов LCL, всех 80 образцов TCGA UVM, трех повторов клеток 293T и трех повторов 293T EV.Желтые прямоугольники указывают максимальные значения оси и , представленные на каждой прямоугольной диаграмме. Последовательность, общая для всех rRF, показана подчеркнутой. Каждый rRF отличается от соседнего rRF на один нуклеотид. — i-, 3′- и x-rRF, продуцируемые с 3′-конца транскрипта 5S рРНК. b 5′-rRF, продуцируемых из транскрипта 28S рРНК. Значения данных для 5S и 28S rRF для трех клеток 293T и трех 293T EV находятся в дополнительном файле 4
Рисунок 6b отслеживает CGCGACCUCAGAUCAGACG U и CGCGACCUCAGAUCAGACG UGGCGACC, два 5′-rSFNA — их общие 28RFACC. сегмент последовательности показан подчеркнутым.5′-rRFs имеют длину 20 и 27 нт, соответственно, и очень распространены в наборах данных LCL (медианная численность составляет 1136 и 707 об / мин, соответственно). В наборах данных UVM эти 5′-rRF не очень распространены. Однако распространенность 22-мерного варианта CGCGACCUCAGAUCAGACG UGG составляет 676 об / мин. Интересно, что не только клетки 293T и 293T EV продуцируют 20-мерный CGCGACCUCAGAUCAGACG U с чрезвычайно высокой численностью (медиана численности составляет 9030 и 27,593 об / мин соответственно), но и 19-мерный CGCGACCUCAGAUCAGACG также очень распространен (медиана численность составляет 3870 и 5807 об / мин, соответственно — обратите внимание на различный диапазон оси и на этих двух графиках).Кроме того, клетки 293T продуцируют 23-мерный CGCGACCUCAGAUCAGACG UGGC со средней численностью 1376 об / мин, тогда как в 293T EV тот же 23-мер присутствует с еще более высокой средней концентрацией (4192 об / мин), что позволяет предположить, что это предпочтительно секретный. Этот 23-мерный 5′-rRF менее распространен в коллекциях UVM и LCL.
Стабильность профилей rRF в лабораториях и различных методах подготовки библиотек
Вполне возможно, что различия в rRF, которые мы показали, существуют среди наборов данных LCL, являются результатом различий в протоколах сбора или выращивания или межлабораторных различий.Консорциум Geuvadis, который сгенерировал анализируемые нами наборы данных, отобрал пять образцов (по одному из каждой из пяти популяций), которые они секвенировали независимо в семи различных европейских центрах секвенирования [39], создав в общей сложности 35 наборов данных. Когда мы сравниваем 35 наборов данных с использованием 1000 лучших rRF из каждой рРНК, мы обнаруживаем высокие корреляции между центрами секвенирования (дополнительный файл 5: рисунок S4A).
Также возможно, что наблюдаемые нами различия rRF являются артефактами приготовления библиотеки кДНК.Чтобы обратиться к этим возможностям, мы глубоко секвенировали два сопоставимых по возрасту и полу коммерчески доступных LCL (которые не были частью образцов проекта 1 кг, чьи профили RNA-seq были описаны в литературе): ND02672, полученный из 63-летний мужчина афроамериканец, и ND07114, который происходит от 66-летнего американца европеоидной расы. Мы глубоко секвенировали каждую клеточную линию с использованием двух различных наборов для подготовки библиотек: TruSeq от Illumina и NEBNext от NEB (см. Раздел «Методы»).В каждом из четырех наборов данных мы идентифицировали РФС, используя методологию, описанную в нашей рукописи. Для каждой из шести рРНК мы определили 1000 самых распространенных rRF и использовали их для вычисления попарных корреляций Пирсона (дополнительный файл 5: рисунок S4B). Как можно увидеть в Дополнительном файле 5: Рисунок S4B, сравнения между наборами показывают высокую корреляцию (корреляция Пирсона ≥ 0,76), что указывает на то, что одинаковые rRF идентифицировались двумя наборами и с сопоставимой численностью.
Дальнейшие доказательства и присутствие rRF в контексте родительских структур рРНК
Чтобы дополнительно подтвердить наши данные о стойких, очень распространенных и дифференциально распространенных rRF, мы показываем нозерн-блоты для четырех rRF, которые многочисленны в наборах данных LCL ( Дополнительный файл 6: Рисунок S5): GGGCUACGCCUGUCUGAGCGUCGC (5.8S 24-мерный i-rRF, средний RPM = 489), UACGCCUGUCUGAGCGUCGCU (5,8S 21-мерный i-rRF, средний RPM = 642), ACCGGGUGCUGUAGGCUU (5S 18-мерный i-rRF, средний RPM = 1752) и CGUCAGCAGAC 28S 20-мер 5′-rRF, средняя частота вращения = 1473) (см. Раздел «Методы»). К ним относятся 24-мерный i-rRF, который был в большом количестве (рис. 5), а также три новых rRF. Опять же, rRF сравнивали с их соответствующими синтетическими ДНК-положительными контролями. Мы видим в северных пятнах в дополнительном файле 6: рис. S5A-D, что rRF, для которых мы зондировали, являются частью большей коллекции различных rRF.Интересно, что 5,8S 24-мер и 21-мер и 5S 18-мер i-rRF сильно детектируются даже в контексте более длинных фрагментов, тогда как 28S 20-мер гораздо менее распространен по сравнению с более длинными фрагментами 28S.
Затем мы хотели узнать, имеют ли эти короткие фрагменты рРНК свою собственную вторичную структуру, которая может указывать на потенциальные механизмы биогенеза. В Дополнительном файле 6: Рисунок S5 мы видим, что два из трех 3′-rRF имеют согласованные шпильковые структуры, в то время как оставшийся частично структурирован, а 5′-rRF 28S вообще не имеет структуры.Мы также исследовали расположение четырех rRF в предполагаемой вторичной структуре полноразмерных рРНК (дополнительный файл 7: рисунок S6). Мы наблюдаем, что хотя два 5.8S i-rRFs (дополнительный файл 7: Рисунок S6A) отображаются в одном и том же общем местоположении родительской 5.8S рРНК, они обрабатываются из разных структурных местоположений. 24-мерный 5.8S i-rRF расщепляется на своем 5′-конце в середине двухцепочечной шпилечной структуры 5.8S рРНК (между положениями 131-132), в то время как его 3′-конец находится на расстоянии двух нуклеотидов от 3′-конец полноразмерной рРНК (положение 155).5′-конец 21-мерного 5.8S i-rRF отщеплен прямо на пересечении двух шпилек РНК (положение 136) и простирается до предпоследнего положения 3′-конца полноразмерного 5.8S. рРНК (положение 156). В дополнительном файле 7: Рисунок S6B, мы можем наблюдать, что 5′-конец 18-мерного 5S i-rRF расщеплен в основании петли (между положениями 102-103) и простирается до предпоследнего нуклеотида в полноразмерная 5S рРНК (позиция 120). Наконец, в Дополнительном файле 7: Рисунок S6C, 20-мерный 28S 5′-rRF имеет 3′-конец в основании петли (позиция 20).
В совокупности выводы и результаты, показанные на рис.
Leave A Comment