
Экзаменационные вопросы по курсу «Ботаника. Высшие растения»
Высшие растения (ФББ)
Экзаменационные вопросы по курсу «Ботаника. Высшие растения»
для студентов 1 курса факультета биоинформатики и биоинженерии
(2022/2023 уч. год)
скачать файл.doc
- Уровни морфологической организации растений. Общая характеристика высших растений. Особенности жизни растений в наземных условиях. Теломные представления происхождения побеговой организации тела растения.
- Слоевищный и побеговый типы организации тела растений. Представленность у современных таксонов высших растений. Возможное возникновение корня в филогенезе.
- Особенности строения растительной клетки. Пластиды. Фотосинтез, его значение в биосфере и эволюции жизни. Эргастические вещества. Их место в растительной клетке, формы отложения, биологическое и практическое значение.
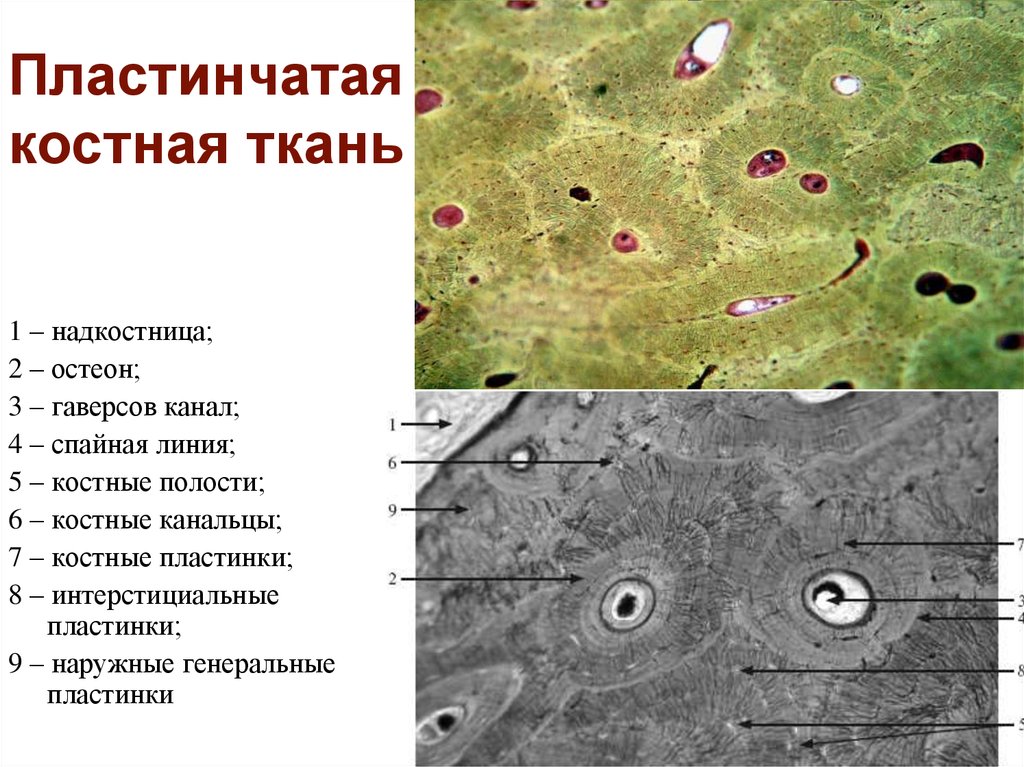
- Клеточная оболочка. Химический состав и молекулярная организация. Первичная и вторичная оболочка.
 Физические и химические свойства. Формирование клеточной стенки при цитокинезе. Плазмодесмы, первичные поровые поля, поры. Химические изменения оболочки: лигнификация, суберинизация, кутинизация, кутикуляризация, минерализация.
Физические и химические свойства. Формирование клеточной стенки при цитокинезе. Плазмодесмы, первичные поровые поля, поры. Химические изменения оболочки: лигнификация, суберинизация, кутинизация, кутикуляризация, минерализация. - Ткани: различные подходы к классификации тканей. Основные типы растительных тканей. Первичные и вторичные постоянные ткани. Понятие об идиобластах.
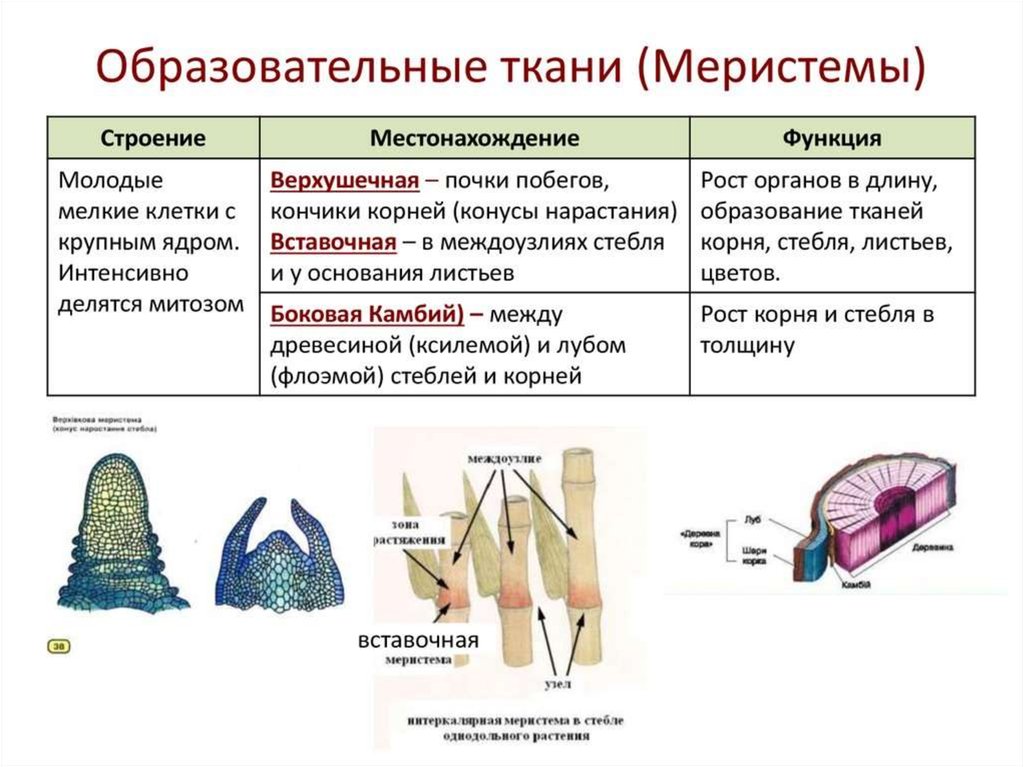
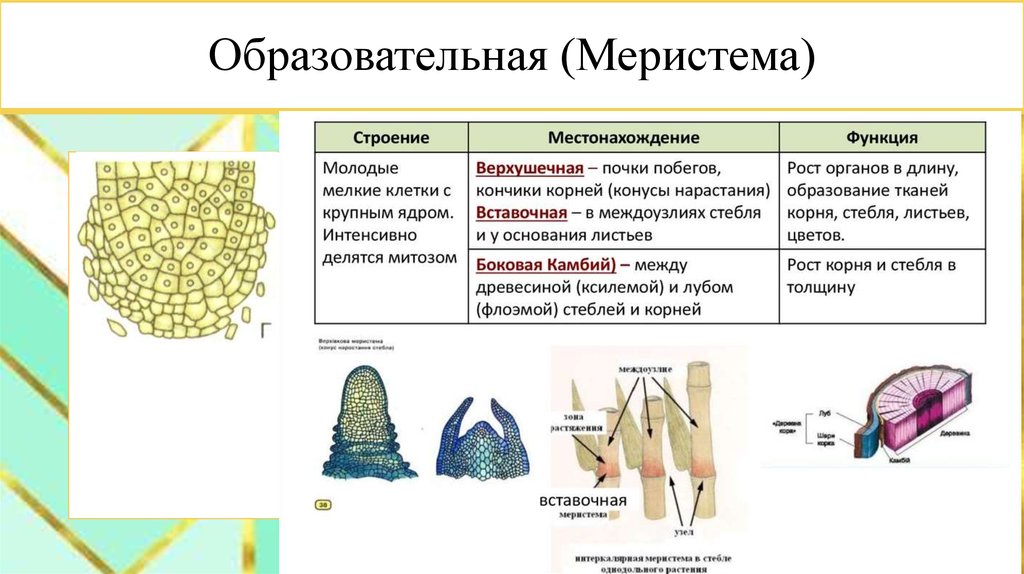
- Меристемы, подходы к классификации. Общие принципы функционирования. Первичные меристемы, их организация. Понятие о покоящемся центре. Интеркалярный рост. Прокамбий.
- Боковые меристемы, устройство и функционирование. Меристема первичного утолщения. Вторичный рост растений.
- Механические ткани: колленхима и склеренхима. Опорная система древесины. Строение стебля как механической опорной конструкции, противодействие деформации.
- Проводящие ткани высших растений. Общие принципы организации. Возможная эволюция водопроводящих элементов ксилемы. Проводящие ткани мохобразных.
- Ксилема, первичная и вторичная, особенности строения и функционирования.
- Флоэма, первичная и вторичная, особенности строения и функционирования.
- Покровные ткани (эпидерма, экзодерма, феллема, полидерма), их образование, строение. Перидерма. Эндодерма, ее строение и функции.
- Организация газообмена с внешней средой у высших растений. Ближний и дальний транспорт газов в растении. Хлоренхима, ее строение и локализация в теле спорофитов и гаметофитов высших растений.
- Корень. Основные функции корня. Топографические зоны молодого корня: деления, растяжения, всасывания, проведения. Корневой чехлик. Корневые волоски. Морфологические типы корней: главный, боковые и придаточные. Гомо- и аллоризия. Строение корневой системы в зависимости от условий среды и осо¬бенностей растения.
- Корень. Основные и дополнительные функции корня. Метаморфозы корня: корнеплоды, корневые клубни, втягивающие, столбовидные, досковидные, ходульные, дыхатель¬ные корни, гаустории, микориза, ризобиальные и актиноризные клубеньки.
- Внутреннее строение корня: первичная кора, центральный цилиндр.
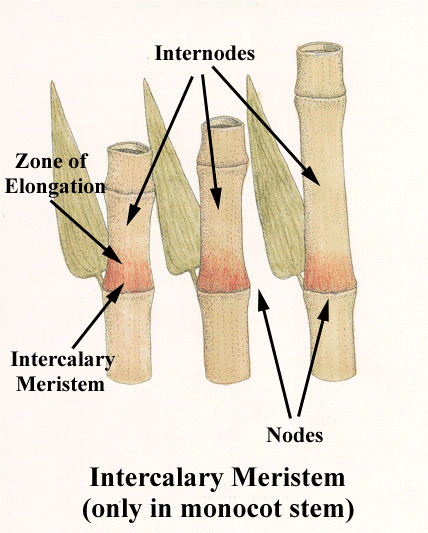
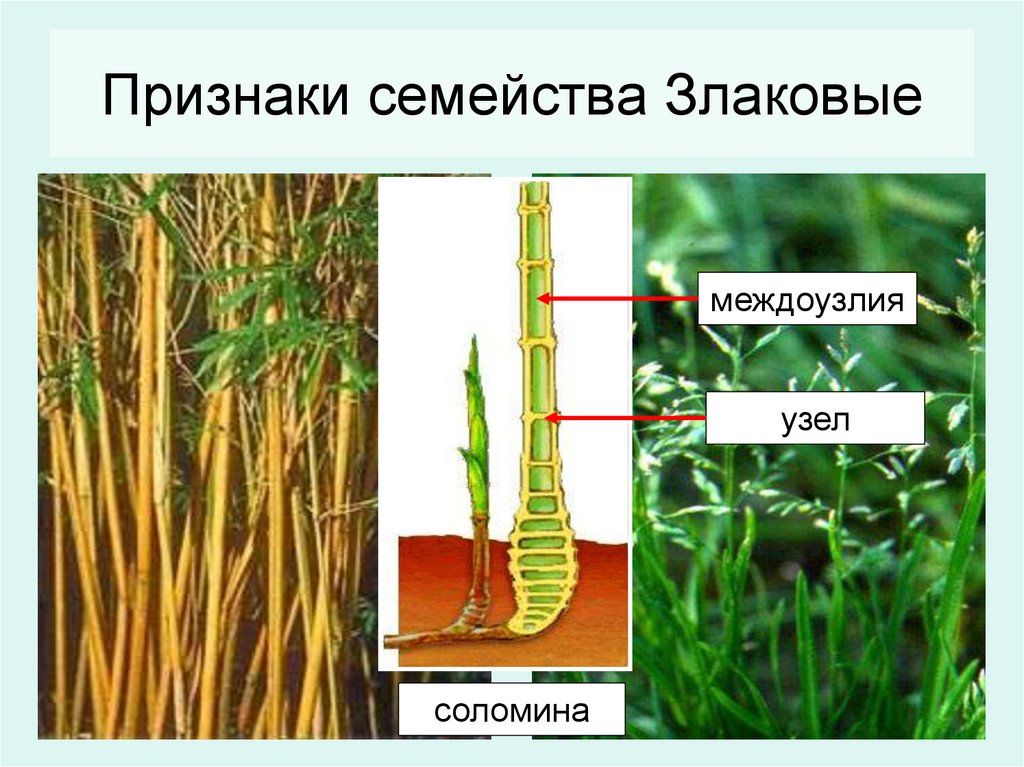
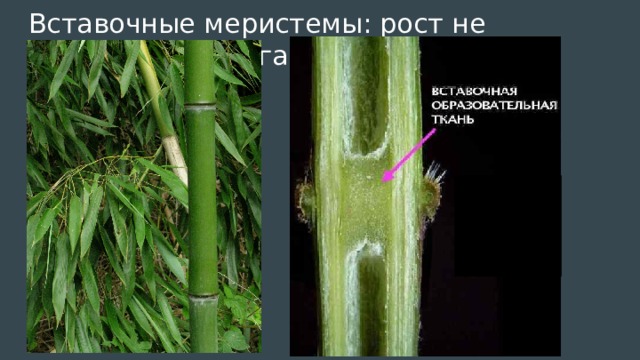
- Понятие о побеге. Узел, междоузлие. Понятие о метамерности побега. Морфологические типы побегов. Строение почки как зачатка побега. Верхушеч¬ный и вставочный рост побега.
- Ветвление побега: верхушечное и боковое. Нарастание побеговых осей: дихоподиальное, моноподиальное и симподиальное. Рост и формирование побеговых систем (однотактный, годичный, силлептический, пролептический, ортотропный, плагиотропный, анизотропный побеги, ауксибласты и брахибласты).
- Соцветие как особый тип побеговой системы цветковых растений. Принципы классификации соцветий. Цимозные и рацемозные соцветия. Тирсы. Понятие о синфлорисценции.
- Метаморфозы побега. Особенности строения, функциональное значение различных метаморфозов побега. Критерии гомологии.
- Стела. Типы стел и их возможная эволюция. Разнообразие устройства стелы у высших растений.
- Стебель и его основные функции. Анатомическое строение стебля травянистых растений. Черты сходства и различия у однодольных и двудольных цветковых растений. Особенности заложе¬ния и дифференциации ксилемы в стебле. Типы строе¬ния пучков. Камбий и вторичное утолщение травянистых стеблей.
- Стебель древесных растений. Нарастание древесины и луба. Структурные элементы древесины и луба и их функции. Годичные кольца древесины. Возрастные изменения древесины и луба. Исполь¬зование древесины и луба в хозяйстве. Особенности вторичного утолщения у древесных цветковых однодольных растений.
- Особенности вторичного утолщения стебля и корня. Отличия анатомического строения корня от стебля. Переход от анатомического строения корня к анатомическому строению стебля.
- Морфологическое и гистологическое строение апексов побега и корня: разнообразие, сходство и различия.
- Лист и его основные функции и развитие. Типы листорасположения. Основные части листа. Разнообразие листьев. Простые и сложные листья. Формации (ярусные категории) листьев. Гетерофиллия и анизофиллия. Жилкование. Влияние усло¬вий на строение листьев. Метаморфозы листа. Листопад, его значение в жизни растений, механизм листопада.
- Анатомическое строение листа на уровне листовой пластинки. Особенности функционирования хлоренхимы. Столбчатый и губчатый мезофилл. Разнообразие анатомического строения листа у растений разных экологических групп.
- Воспроизведение высших растений: бесполое и по¬ловое. Вегетативное размножение растений. Значение вегетативного размножения растений в природе и сельскохозяйственной практике.
- Бесполое воспроизведение. Спорообразование у высших растений. Строение спор. Равно- и разноспоровость. Половое воспроизведение. Особенности полового процесса у высших растений. Зоидо- и сифоногамия. Строение половых органов (антеридиев и архегониев) и гамет.
- Понятие о спорофите и гаметофите. Чередование поколений и ядерных фаз у высших растений.
- Разнообразие строения гаметофитов у современных высших растений. Эндо- и экзоспорическое развитие гаметофитов. Редукция гаметофитов у семенных растений.
- Споры и пыльца. Приспособление к биотическим и абиотическим агентам переноса.
- Надотдел Моховидные (Bryomorphae). Общая характеристика. Особенности жизненного цикла. Деление на отделы: антоцеротовые (Anthocerotophyta), печеночники (Marchantiophyta), листостебельные мхи (Bryophyta).
- Отдел антоцеротовые (Anthocerotophyta). Особенности их строения, размноже¬ния и образа жизни.
- Отдел печеночники (Marchantiophyta). Общая характеристика. Талломная и листостебельная организация тела печеночника. Гаметофит, спорофит, их строение. Маршанциевые и юнгерманниевые печеночники. Экология, географическое распространение, значение в природе.
- Отдел листостебельные мхи (Bryophyta). Общая характеристика. Строение гаметофита и спорофита. Устройство перистома. Особен¬ности строения зеленых и сфагновых мхов. Проблема происхождения мохообразных. Экология, географическое распространение, значение в природе и хозяйстве.
- «Отдел» Риниофиты (Rhiniophyta). Общая характеристи¬ка и анатомическая структура основных представителей: куксония, аглаофитон, риния и др. Возможный жизненный цикл. Разные точки зрения на положение отдельных представителей в системе высших растений.
- Отдел Плауновидные (Lycopodiophyta). Общая характеристика. Микрофилльная линия эволюции. Происхождение листа. Особенности строения и жизненный цикл представителей класса Плауновые (Lycopodiopsida). Стелярная организация.
- Разноспоровые плауновидные. Особенности строе¬ния плаунка (Selaginella) и полушника (Isoёtes): стебель, проводящая система, листья. Спороношение. Половое поколение, редукция гаметофита у разноспоровых представителей. Ископаемые лигульные плауновидные, их роль в растительном покрове палеозойской эры и в образовании каменного угля.
- Отдел Папоротниковидные (Pteridophyta). Общая характеристика. Жизнен¬ный цикл. Макрофилльная линия эволюции. Происхождение листа. Разнообразие строения спорофита и гаметофита. Типы стелы. Особенности формирования спорангиев (эвспорангиатные и лептоспорангиатные формы).
- Отдел Папоротниковидные (Pteridophyta). Класс хвощевые (Equisetopsida). Общая характеристика. Распространение и экологические особенности. Жизненный цикл. Особенности строения спорофита: листья, ветвление, проводящая система. Гаметофит. Ископаемые представители хвощевых.
- Отдел Папоротниковидные (Pteridophyta). Классы псилотовые (Psilotopsida), ужовниковые (Ophioglossopsida) и мараттиевые (Marattiopsida). Строение их спорофитов и гаметофитов.
- Отдел Папоротниковидные (Pteridophyta). Красс Лептоспорангиатные папоротники (Polypodiopsida). Строение их спорофитов и гаметофитов, цикл развития. Экологическое разнообразие папоротников, значение современных папоротниковидных в природе.
- Отдел Голосеменные (Gymnospermae). Общая характеристика. Жизненный цикл (на примере сосны). Возникновение семязачатка в филогенезе. Значение семени для эволюции наземных растений. Разделение на классы: саговниковые (Cycadopsida), гинкговые (Ginkgoopsida), хвойные (Pinopsida), оболочкосеменные (Gnetopsida).
- Класс саговниковые (Cycadopsida). Общая характеристика. Строение листьев, стебля, органов спороношения. Особенности строения гаметофитов. Половой процесс. Семя.
- Класс гинкговые (Ginkgoopsida). Общая характеристика. Современный пред¬ставитель — гинкго. Строение его вегетативных органов. Органы размножения, половой процесс, формирование семени. Класс оболочкосеменные (Gnetopsida). Общая характеристика. Строение вегетативных органов. Органы размножения, гаметофиты, семена.
- Класс шишконосные (Pinopsida). Общая характеристика. Строение листьев, стебля. Органы размножения. Развитие гаметофитов. Половой процесс и образова¬ние семян.
- Отдел Покрытосеменные (Angiopsermae). Общая характеристика. Разнообразие строения. Цветок. Двойное оплодотворение. Семя и плод. Жизненный цикл покрытосеменных растений. Значение покрытосеменных для человечества.
- Строение цветка покрытосеменных: цветоложе, околоцветник и многообразие его строения (симметрия, срастание, редукция). Гинецей. Пестик. Апокарпия, ценокарпия. Положение завязи в цветке.
- Андроцей; строение тычинки, пыльник, образование микроспор и пыльцы. Разнообразие строения андроцея. Абиотическое и биотическое опыление.
- Двойное оплодотворение. Развитие семени. Морфологические типы семян. Прорастание семян. Типы прорастания семян. Морфологическое разнообразие проростков.
- Развитие плода. Основные типы плодов: апокарпии, синкарпии, паракарпии, лизикарпии. Мно-гообразие плодов.
- Плоды и семена – элементы расселения растений. Способы распространения диаспор у семенных растений.
- Взгляды на морфологическую природу и происхождение цветка покрыто¬семенных растений. Эвантовая, псевдантовая и теломная теории происхождения цветка.
- Разделение покрытосеменных на классы. Классы двудольные (Dicotyledones, Magnoliopsida) и однодольные (Monocotyledones, Liliopsida). Общая характеристика. Морфологические особенности вегетативных органов, цветка, семян. Признаки классов в строении вегетативных органов и цветка.
- Понятие о жизненной форме растений. Основные классификации жизненных форм.
- Понятие о факторах внешней среды. Климатические и эдафические факторы. Экологические группы растений по отношению к воде. Анатомо-морфологические особенности растений разных экологических групп по отношению к воде.
- Понятие о факторах внешней среды. Климатические и эдафические факторы. Экологические группы растений по отношению к свету. Анатомо-морфологические особенности растений разных экологических групп по отношению к свету.
- Понятие о факторах внешней среды. Климатические и эдафические факторы. Экологические группы растений по отношению к субстрату. Анатомо-морфологические особенности растений разных экологических групп по отношению к субстрату.
- Растительный покров. Подходы к его изучению: флора и растительность. Структура флоры и принципы классификации растительности.
 Физические и химические свойства. Формирование клеточной стенки при цитокинезе. Плазмодесмы, первичные поровые поля, поры. Химические изменения оболочки: лигнификация, суберинизация, кутинизация, кутикуляризация, минерализация.
Физические и химические свойства. Формирование клеточной стенки при цитокинезе. Плазмодесмы, первичные поровые поля, поры. Химические изменения оболочки: лигнификация, суберинизация, кутинизация, кутикуляризация, минерализация.
 Разнообразие устройства стелы у высших растений.
Разнообразие устройства стелы у высших растений. Основные части листа. Разнообразие листьев. Простые и сложные листья. Формации (ярусные категории) листьев. Гетерофиллия и анизофиллия. Жилкование. Влияние усло¬вий на строение листьев. Метаморфозы листа. Листопад, его значение в жизни растений, механизм листопада.
Основные части листа. Разнообразие листьев. Простые и сложные листья. Формации (ярусные категории) листьев. Гетерофиллия и анизофиллия. Жилкование. Влияние усло¬вий на строение листьев. Метаморфозы листа. Листопад, его значение в жизни растений, механизм листопада.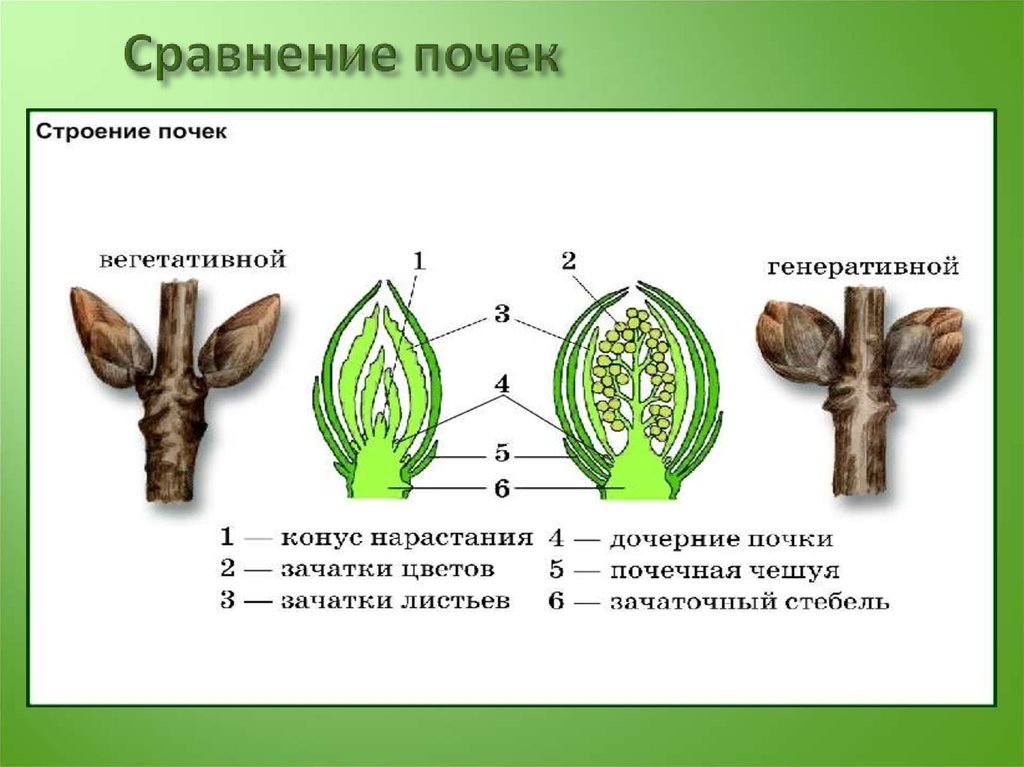
 Общая характеристика. Строение гаметофита и спорофита. Устройство перистома. Особен¬ности строения зеленых и сфагновых мхов. Проблема происхождения мохообразных. Экология, географическое распространение, значение в природе и хозяйстве.
Общая характеристика. Строение гаметофита и спорофита. Устройство перистома. Особен¬ности строения зеленых и сфагновых мхов. Проблема происхождения мохообразных. Экология, географическое распространение, значение в природе и хозяйстве.
 Общая характеристика. Жизненный цикл (на примере сосны). Возникновение семязачатка в филогенезе. Значение семени для эволюции наземных растений. Разделение на классы: саговниковые (Cycadopsida), гинкговые (Ginkgoopsida), хвойные (Pinopsida), оболочкосеменные (Gnetopsida).
Общая характеристика. Жизненный цикл (на примере сосны). Возникновение семязачатка в филогенезе. Значение семени для эволюции наземных растений. Разделение на классы: саговниковые (Cycadopsida), гинкговые (Ginkgoopsida), хвойные (Pinopsida), оболочкосеменные (Gnetopsida). Разнообразие строения. Цветок. Двойное оплодотворение. Семя и плод. Жизненный цикл покрытосеменных растений. Значение покрытосеменных для человечества.
Разнообразие строения. Цветок. Двойное оплодотворение. Семя и плод. Жизненный цикл покрытосеменных растений. Значение покрытосеменных для человечества.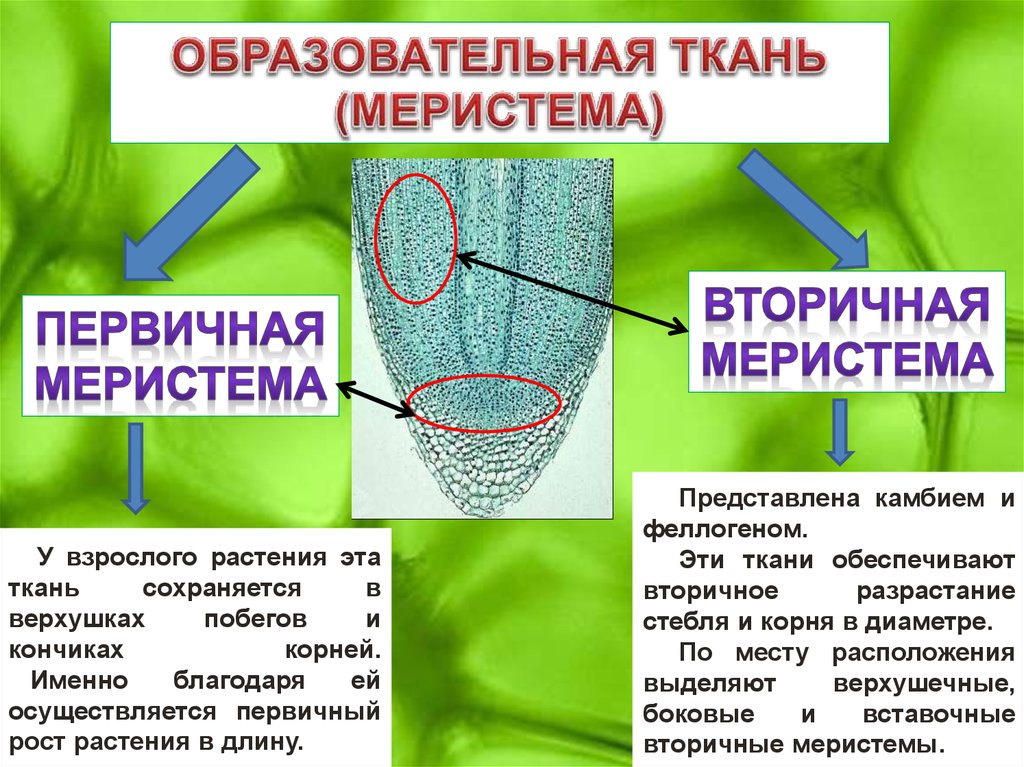
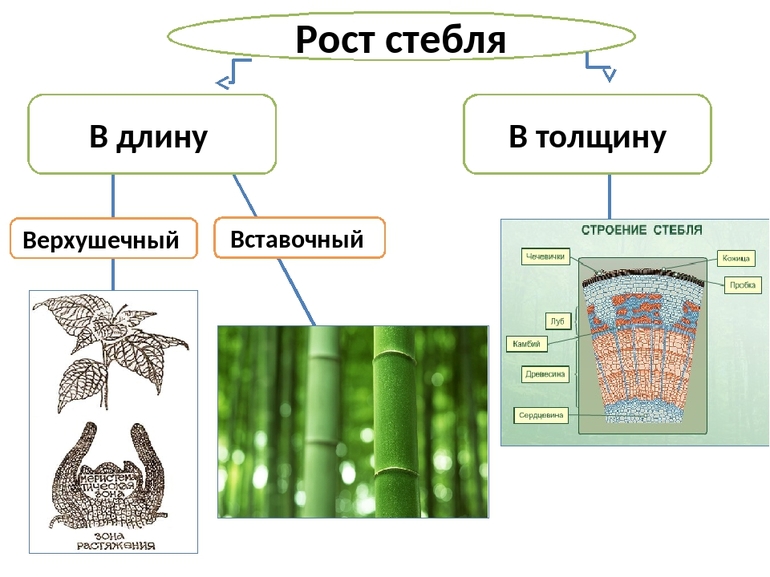
Рост стебля | Растения | Биология
Рост стебля в длину
Стебель вырастает из зачаточного стебля почки. Если это главный стебель растения, то он развивается из зародышевой почечки семени.
После того, как почки набухают и у них раздвигаются защитные чешуйки, начинает расти стебель и разворачиваться листочки. У растущего из почки стебля постепенно увеличивается длина междоузлий.
На самом верху побега находится так называемая верхушечная почка. В ней есть конус нарастания. Деление клеток конуса нарастания приводит к росту стебля в длину.
Конус нарастания состоит из образовательной ткани. Ее клетки способны к постоянному делению.
На более нижних клетках конуса нарастания появляются зачаточные листья, клетки стебля перестают делится и начинают расти. В результате растет сам стебель, и получается что он растет своей верхней частью. Так если нанести на всю длину стебля метки, то через некоторое время расстояние между метками на его верхушке увеличится, так как здесь клетки продолжают расти в длину. В то время как ниже по стеблю расстояние между метками может не измениться.
В результате растет сам стебель, и получается что он растет своей верхней частью. Так если нанести на всю длину стебля метки, то через некоторое время расстояние между метками на его верхушке увеличится, так как здесь клетки продолжают расти в длину. В то время как ниже по стеблю расстояние между метками может не измениться.
Однако стебли растут в длину не всегда только за счет конуса нарастания. У многих растений наблюдается вставочный рост, при котором удлиняются междоузлия побега. Обычно при этом делятся и растут клетки оснований междоузлий.
Если у стебля удалить его верхушку вместе с конусом нарастания, то прекратится его рост в длину. Но при этом стебель начнет ветвится, т. е. начнут вырастать боковые побеги.
Рост стебля в толщину
Рост стебля в толщину обеспечивается за счет деления клеток камбия. Рост в толщину наблюдается у деревьев и кустарников, а также у многолетних трав. У деревьев камбий находится под корой. Камбий состоит из образовательной ткани.
Камбий состоит из образовательной ткани.
Рост стебля в толщину происходит в благоприятный период года. В умеренных широтах это происходит в теплый период. В это время клетки камбия активно делятся.
У деревьев те клетки камбия, которые находятся ближе к коре, становятся клетками луба. Те, что ближе к древесине, становятся древесиной. При этом за вегетационный сезон у дерева клеток древесины образуется больше, чем клеток луба.
В древесине, которая нарастает весной, развиваются достаточно толстые сосуды с тонкими стенками. Сосуды же осенней древесины, наоборот, тонкие с более толстыми оболочками.
Поскольку зимой стебель в толщину не растет, а весной начинают образовываться снова крупные клетки, то на спиле ствола видны отчетливые переходы от мелких клеток к крупным. Клетки древесины одного года называются годичным кольцом. По количеству годовых колец можно определить возраст дерева.
Годичные кольца разных лет могут отличаться между собой.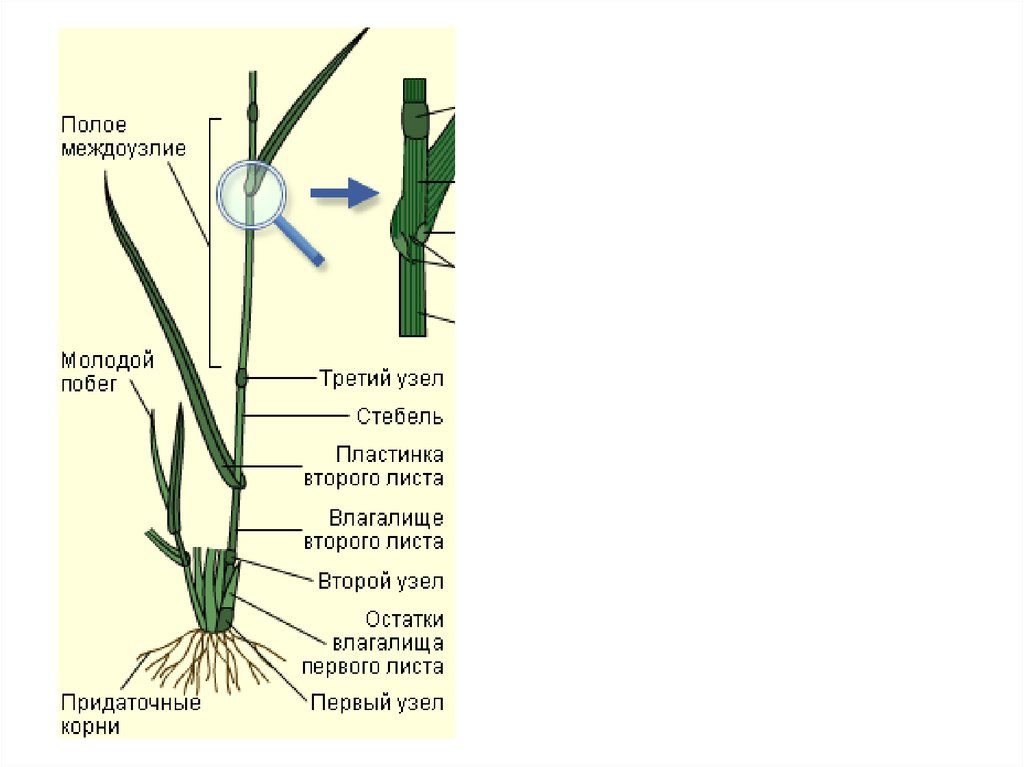 Одни могут быть более узкие, другие — более широкие. Такое различие обусловлено разными погодными условиями. Если год был хороший, дерево получало достаточно влаги и света, то годовое кольцо будет широким. Также ширина каждого отдельно взятого годового кольца не одинакова. С южной стороны кольца обычно шире, чем с северной. Это связано с тем, что с северной стороны камбий обычно меньше прогревается, и поэтому хуже делятся его клетки.
Одни могут быть более узкие, другие — более широкие. Такое различие обусловлено разными погодными условиями. Если год был хороший, дерево получало достаточно влаги и света, то годовое кольцо будет широким. Также ширина каждого отдельно взятого годового кольца не одинакова. С южной стороны кольца обычно шире, чем с северной. Это связано с тем, что с северной стороны камбий обычно меньше прогревается, и поэтому хуже делятся его клетки.
Интеркалярная меристема – определение и примеры
Интеркалярная меристема
сущ., множественное число: вставочные меристемы
[in′tər·kə‚ler·ē ′mer·ə‚ствол]
Определение: ( ботаника ) Тип меристематической ткани в основании узлов и листовых пластинок однодольных
Содержание
Основной структурный каркас растений состоит из различных типов тканей. Основываясь на способности делиться, ткани растений можно разделить на два основных типа, а именно постоянные ткани и меристематические ткани . В отличие от постоянных тканей клетки меристематических тканей не теряют способности к делению. Они не дифференцируются, чтобы приобрести постоянную форму, размер или функцию; скорее, они остаются относительно маленькими, незрелыми и цитоплазматически плотными с высокой скоростью дыхания.
В отличие от постоянных тканей клетки меристематических тканей не теряют способности к делению. Они не дифференцируются, чтобы приобрести постоянную форму, размер или функцию; скорее, они остаются относительно маленькими, незрелыми и цитоплазматически плотными с высокой скоростью дыхания.
По происхождению меристематические ткани бывают двух типов, а именно первичные и вторичные меристемы . первичные меристематические ткани происходят непосредственно из меристем эмбрионов, тогда как вторичные меристематические ткани происходят из постоянных тканей в процессе дедифференцировки . Среди первичных меристематических тканей различают три типа в зависимости от положения ткани. Это:
1) Верхушечный
2) Вставочный
3) Боковой
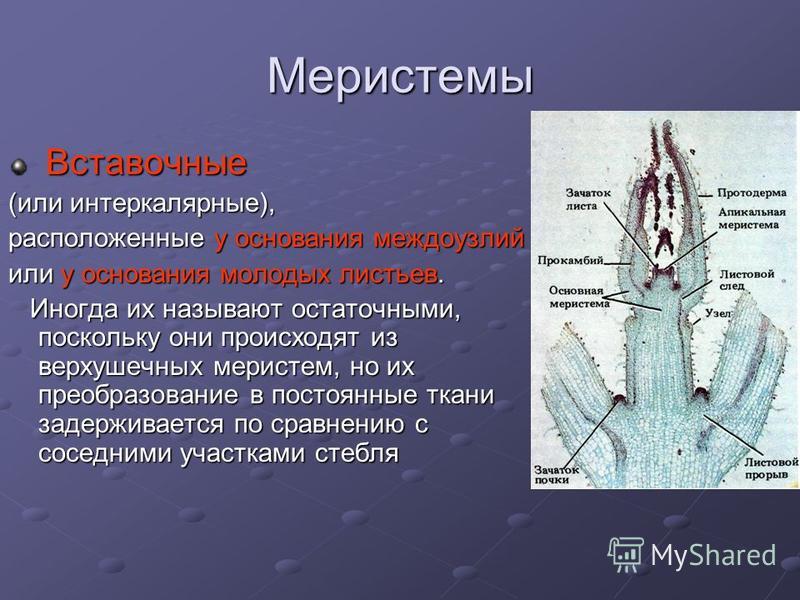
Если спросить у растений, где происходит рост? Мы можем объяснить, что это происходит на верхушках (как корней, так и верхушек побегов) с помощью «Апикальной меристемы» . Это происходит в междоузлиях тела растения с помощью «Интеркалярной меристемы» . И это происходит вместе с обхватом тела растения с помощью « Боковая меристема» . Давайте посмотрим на диаграмму ниже, чтобы помочь определить и отличить вставочную меристему.
Это происходит в междоузлиях тела растения с помощью «Интеркалярной меристемы» . И это происходит вместе с обхватом тела растения с помощью « Боковая меристема» . Давайте посмотрим на диаграмму ниже, чтобы помочь определить и отличить вставочную меристему.
Меристематические ткани состоят из активно делящихся клеток. Они ответственны за неопределенный рост растений. Они дают начало постоянным тканям растений, таким как сосудистые ткани, эпидермис, феллема, наземные ткани и т. Д. В зависимости от расположения меристематической ткани выделяют три различных типа: (1) апикальная меристема (терминальные части), (2) интеркалярная меристема. (в узлах некоторых однодольных) и (3) латеральная меристема (в сторону или в стороны).
(в узлах некоторых однодольных) и (3) латеральная меристема (в сторону или в стороны).
Интеркалярная меристема Определение
Интеркалярная меристема — это первичная меристематическая ткань, которая помогает растению или дереву расти вертикально за счет увеличения его длины. Клетки в этой меристематической области маленькие, тотипотентные, тонкостенные и полные протоплазмы. Отличительными признаками интеркалярной меристемы являются:
- делящаяся природа
- первичные (возникающие непосредственно из эмбриона) по происхождению
- вставочный (между постоянными тканями) по положению
Поскольку интеркалярная меристема является митотически (делительной) активной, она участвует в вертикальном или интеркалярном росте растения, тем самым давая свое название.
В отличие от других первичных меристематических тканей, интеркалярная меристема в конечном итоге имеет пределы; в конечном итоге он достигнет предела и созреет в постоянные ткани.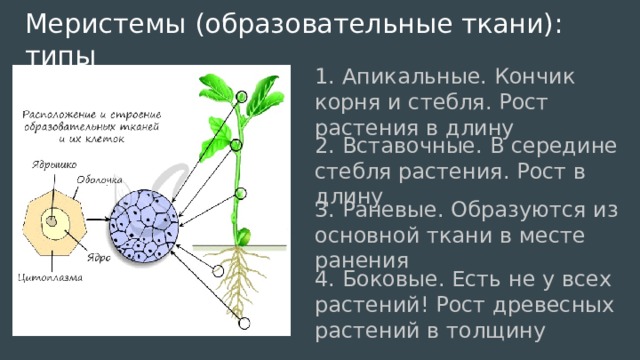
В некоторых научных источниках также указывается, что интеркалярная меристема на самом деле является меристемной тканью, встроенной в зрелую ткань. В такой теории апикальная меристема была указана как источник интеркалярной меристемы. Говорят, что после прикрепления апикальная ткань разделяется последовательно образующимися узлами на ряд вставочных отделов, которые в конечном итоге образуют ряд вставочных меристем. (Кришнамурти К.В. и др., 2015, Сюн В.Ю., 1980 and Schweingruber F.H., 2018)
Рисунок 2: Схема типичного газона. Предоставлено: Kelvinsong, CC BY-SA 3.0 Биологическое определение ( ботаника ):
Интеркалярная меристема — это тип меристематической ткани, которая находится в основании узлов и листовых пластинок однодольных растений. Интеркалярная меристема — это тип меристематической ткани, связанный с ростом в длину в среднем положении, отсюда и название. В частности, он состоит из меристематических клеток, которые митотически делятся в стебле у основания узлов и листовых пластинок. Рост в этой точке обозначается как вставочный рост . Это важно для этих групп растений, поскольку служит адаптивным механизмом против травоядных. Благодаря вставочному росту эти растения способны быстро отращивать листья и удлинять стебли, чтобы компенсировать части растений, поедаемые травоядными животными.
Рост в этой точке обозначается как вставочный рост . Это важно для этих групп растений, поскольку служит адаптивным механизмом против травоядных. Благодаря вставочному росту эти растения способны быстро отращивать листья и удлинять стебли, чтобы компенсировать части растений, поедаемые травоядными животными.
Синоним: интеркалярная меристематическая ткань; интеркалярная ростовая ткань.
Два возможных источника интеркалярных меристем
Интеркалярная меристема может возникать из двух источников, которые находятся далеко позади кончиков побегов и корней. Один — субапикальная меристема , которая лежит в основании апикальной меристемы и способствует « непосредственно » образованию интеркалярной меристемы. Другой – процессом дедифференцировки зрелых тканей, который «косвенно» приводит к образованию интеркалярной меристемы в позднем периоде жизни растения.
- Субапикальная меристема
Это происходит путем постепенного ограничения активности деления любой областью междоузлия/оси цветков/оси колоса, где она очень сильно продлевается, внося значительный вклад в междоузлия. - Дедифференцировка зрелых тканей
Интеркалярная меристема часто созревает в развитые ткани, но некоторая ее часть остается «молча консервированной» даже после созревания. Когда какая-то среда, условия роста или развития вызывают эту покоящуюся ткань, она переходит в состояние дедифференцировки. Дедифференцировка зрелых тканей представляет собой трансформацию клеток из заданного высокодифференцированного состояния в менее дифференцированное или стволово-подобное состояние). По этому пути вносит вклад в активность деления в течение очень долгого времени и, как правило, вносит основной вклад в продольный рост в последней части жизни растений.
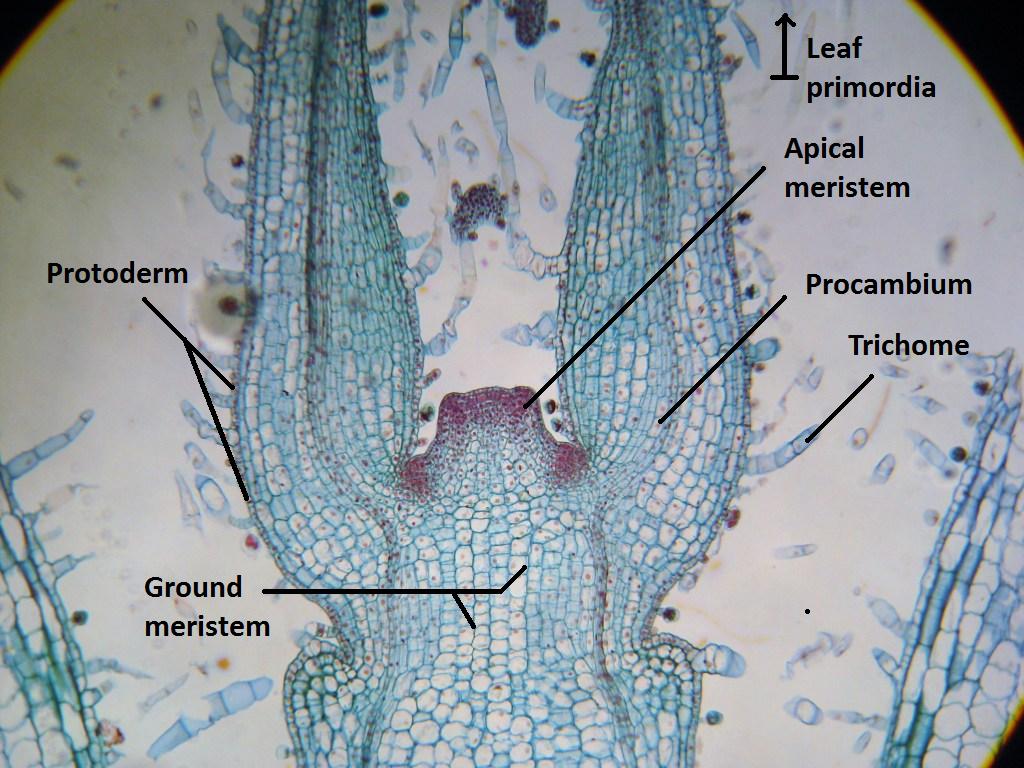
Рисунок 4: Интеркалярная меристема сахарного тростника расположена у основания междоузлия и над узлами.
 Множественные вставочные меристемы в каждой вставочной секции могут синхронизироваться или действовать независимо, удлиняя стебель сахарного тростника. Источник: Биззо, 2014 г.
Множественные вставочные меристемы в каждой вставочной секции могут синхронизироваться или действовать независимо, удлиняя стебель сахарного тростника. Источник: Биззо, 2014 г.Присутствие в различных растительных линиях
Различные линии растений, начиная с Thallophytes (водоросли), Bryophytes (Mosses), Pteridophytes (папоротники) до Gymnosperms (Conifers, Conifers, Conifers, Conifers, Confers, и Angios. ) показали бесчисленное количество вставочных меристем.
Хотя положение вставочных меристем и их функции сильно различаются по всему спектру, значение в общем росте и развитии растений остается прежним.
Покрытосеменные
Среди покрытосеменных интеркалярная меристема обычно встречается у однодольных. Хотя и однодольные, и двудольные имеют апикальную меристему побега, она делится во время развития с образованием дополнительной меристемы ниже верхушки, т. Е. Вставочной меристемы, только у однодольных, а не у двудольных.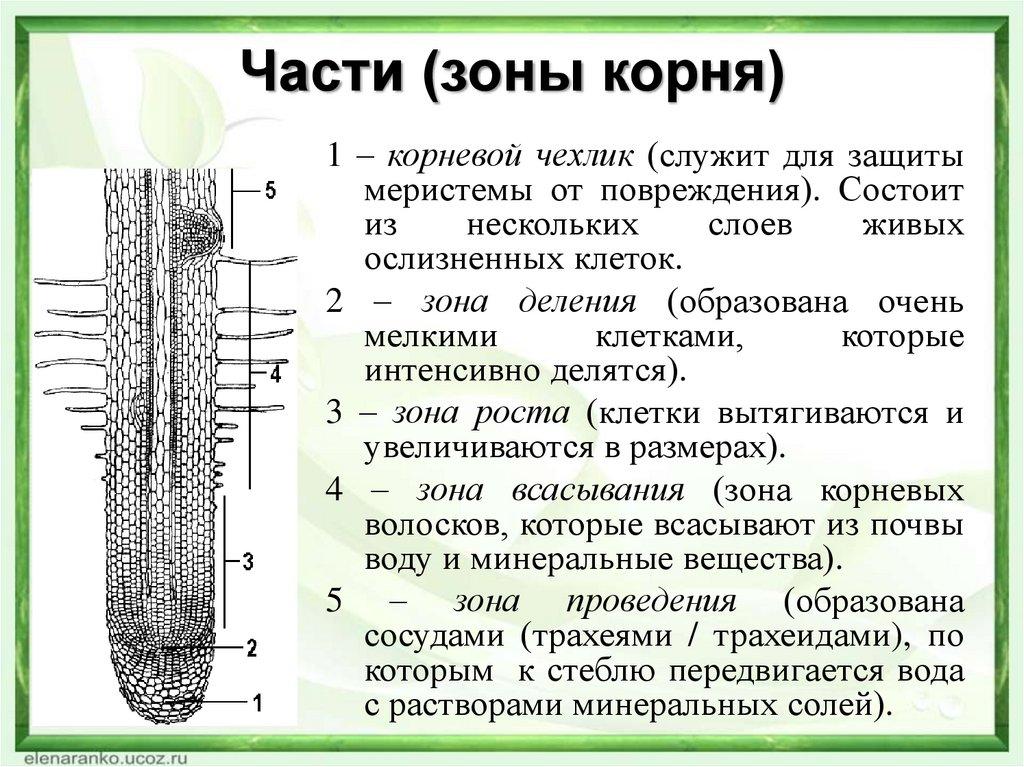 Этот процесс продолжается до начала цветения и создает интеркалярные меристемы, расположенные в узлах злаков.
Этот процесс продолжается до начала цветения и создает интеркалярные меристемы, расположенные в узлах злаков.
Исключения:
- В мяте обнаружена интеркалярная меристема ( Mentha arvensis ), который относится к Lamiaceae, и растениям арахиса ( Arachis hypogea ), который относится к Fabaceae. (И Lamiaceae, и Fabaceae относятся к семействам двудольных покрытосеменных.)
- Интеркалярная меристема обнаружена также у некоторых двудольных растений. Пример: Ipomoea spp., Thunbergia grandiflora .
Рисунок 5: Макроскопический аспект интеркалярных меристем покрытосеменных растений. Стрелка указывает на узел стебля Poaceae. Интеркалярная меристема располагается над узлом. Источник: Schweingruber F.H., 2018.
Рисунок 6: Микроскопический вид интеркалярных меристем покрытосеменных
Зона удлинения с неодревесневшими меристемными клетками во влагалище листьев Poaceae.
Источник: Schweingruber F.H., 2018
Голосеменные
Среди голосеменных интеркалярная меристема обнаружена у Пинус . В развитии Pinus cataphyl (редуцированных листьев голосеменных растений) решающую роль играет интеркалярная меристема.
Таллофиты
Среди таллофитов/водорослей сообщалось о интеркалярных меристемах в пластинке ламинарии (Phaeophyta/бурые водоросли). Интеркалярная меристема у ламинарии присутствует между ножкой и пластинкой, увеличивая длину слоевища.
Птеридофиты
Среди папоротников/папоротников интеркалярная меристема обнаружена у хвощей (виды хвоща). Воздушные побеги Equisetum spp. имеют интеркалярные меристемы, но нет данных об их наличии в корневищах.
Как интеркалярная меристема способствует вегетативному росту Equisetum spp. стрелять?
Последовательность событий, которые приводят к образованию интеркалярной меристемы у Equisetum spp.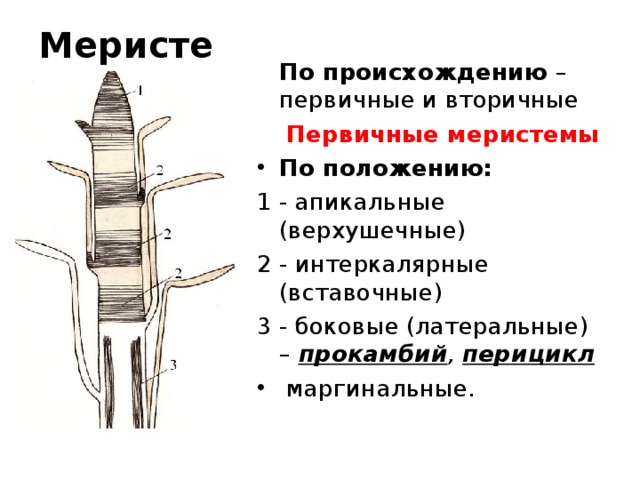 . объясняется ниже. После образования интеркалярной меристемы у Equisetum spp. , способствует вегетативному росту вместе с апикальной меристемой.
. объясняется ниже. После образования интеркалярной меристемы у Equisetum spp. , способствует вегетативному росту вместе с апикальной меристемой.
- Верхушечные клетки вегетативного побега делятся и приводят к образованию «мерофитов».
- Эти мерофиты работают синхронно и приводят к развитию единиц, называемых «фитомерами».
- Фитомер можно разделить на два отдельных уровня.
- верхний узловой ярус
- базальный междоузлий ярус
- Именно этот базальный междоузлий сохраняет меристематическую активность и приводит к образованию интеркалярной меристемы Виды хвоща.
Функции интеркалярных меристем
Как и у животных, орган или ткань могут выполнять различные функции в различных линиях, родах и видах; точно так же у растений интеркалярная меристематическая ткань действует по разным направлениям.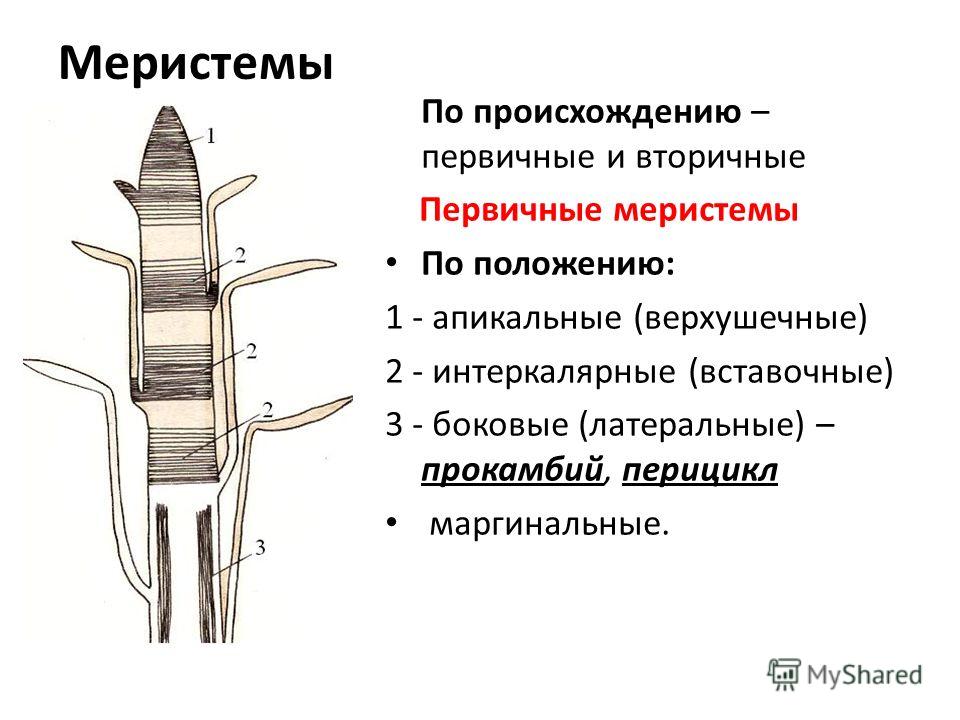 Начиная от вертикального (продольного) роста и заканчивая пастбищной устойчивостью и образованием гинофоров, он отвечает некоторым безупречным требованиям растения для выживания и размножения. Некоторые из наиболее важных ролей интеркалярной меристемы перечислены ниже:
Начиная от вертикального (продольного) роста и заканчивая пастбищной устойчивостью и образованием гинофоров, он отвечает некоторым безупречным требованиям растения для выживания и размножения. Некоторые из наиболее важных ролей интеркалярной меристемы перечислены ниже:
1) Помогает упавшим стеблям злаков стать прямостоячими.
2) Помогает в регенерации газонной травы, удаленной в результате выпаса травоядных животных или повторного кошения.
3) Помогает росту бамбука в высоту, удлинению междоузлий.
4) Он играет огромную роль в морфологии развития и физиологии трав. При росте злаков сначала весь зачаток листа обладает меристематической активностью, но со временем клеточное деление ограничивается интеркалярной меристемой. Когда происходит формирование язычка (рис. 8), интеркалярная меристематическая деятельность внутри пластинки прекращается. Когда язычок обнажается, интеркалярная меристематическая активность внутри оболочки прекращается. Таким образом, базальное расположение интеркалярной меристематической ткани в двух основных частях травы — лезвии и оболочке объясняет, почему удлинение листа может происходить после дефолиации/поедания/повторного скашивания без замены кончика листа.
8), интеркалярная меристематическая деятельность внутри пластинки прекращается. Когда язычок обнажается, интеркалярная меристематическая активность внутри оболочки прекращается. Таким образом, базальное расположение интеркалярной меристематической ткани в двух основных частях травы — лезвии и оболочке объясняет, почему удлинение листа может происходить после дефолиации/поедания/повторного скашивания без замены кончика листа.
Если мы сравним меристематическую активность разных меристем у злаков, мы, по существу, выделим два основных момента:
- Скорость роста листьев ремонтных трав самая высокая за счет интеркалярной меристемы и самая медленная за счет пазушной меристемы. Поэтому интеркалярную меристему злаков часто называют меристемой злаков.
Интеркалярная меристема > Апикальная меристема > Подмышечная меристемаРисунок 9: Сравнение меристематической активности различных меристем растений: интеркалярной, верхушечной и пазушной меристемы почек.
Источник Бриске Д.Д., 19 лет86 - Пазухные почки придают растению многолетник, в то время как интеркалярные меристемы недолговечны.
 Источник Бриске Д.Д., 19 лет86
Источник Бриске Д.Д., 19 лет86 5) В стеблевидных травах, таких как сорго, просо и сахарный тростник, интеркалярная меристема играет ключевую роль в удлинении стебля. При приготовлении солода из этих трав солома составляет важную часть солодового продукта. Чем более удлинена солома, тем больше количество извлекаемого солода. Кроме того, из-за их высокой пищевой ценности они используются в качестве полезных кормов для животных.
6) У арахиса ( Arachis hypogea ) образование гинофора происходит вблизи завязи из-за наличия интеркалярной меристемы в стебле завязи. Эта вставочная меристема активируется только после опыления и успешного оплодотворения. Роль вставочной меристемы здесь заключается в том, чтобы закапывать оплодотворенную завязь глубоко в почву.
7) У ламинарии (Phaeophyta/бурые водоросли) интеркалярная меристема способствует увеличению длины слоевища.
Примеры интеркалярной меристемы
Таким образом, примерами интеркалярных меристем являются следующие автотрофы и их расположение:
- Присутствуют у представителей Poaceae, т.е. трав и злаков (у основания междоузлий и над узлами)
- В наличии (ниже узлов)
- Присутствует в Pinus (у основания листьев)
- Присутствует в бамбуке (в междоузлиях)
- Присутствует в сахарном тростнике (у основания нескольких самых верхних междоузлий)
- Присутствует в Equisetum (расположен над каждым узлом)
- Присутствует в Arachis hypogea (в стебле яичника)
- Присутствует в ламинарии, Хордовая нить (как в вегетативной, так и в спорофитной фазах)
 Источник: Когаме, 19 лет.96.
Источник: Когаме, 19 лет.96.Рисунок 11: Интеркалярная меристема присутствует в стебле завязи растения арахиса (Arachis hypogea). Эта вставочная меристема активируется только после опыления и успешного оплодотворения. Его деятельность производит «гвоздь», ось, которая уносит оплодотворенную завязь, будущий плод, вниз, зарывая его в почву, где он развивается. Источник: Morfología de Plantas Vasculares
Интересный факт о вставочной меристеме
Найдите время для отдыха и прогуляйтесь ночью по кукурузному полю. Вы услышите несколько хлопающих звуков; громко и четко!! Угадайте, что это может быть?? Какое-то насекомое или вредитель???
Нет, это звук внезапного роста стеблей кукурузы из-за быстрого удлинения интеркалярной меристемы. Поскольку удлинение кукурузы происходит очень спонтанно, вы можете услышать, как оно растет… (Источник: Losos J.B., 2010).
Попробуйте ответить на приведенный ниже тест, чтобы проверить, что вы уже узнали об интеркалярной меристеме.
Викторина
Выберите лучший ответ.
1. Что такое интеркалярная меристема?
Ткань растения, состоящая из дифференцированных клеток
Ткань растения, состоящая из клеток, способных к делению клеток
Постоянная ткань растения
2. Где находится интеркалярная меристема?
Верхушка побега растения
Кончик корня растения
Стебель у основания узла
3. Интеркалярная меристема – _________ тип меристематической ткани
Первичная
Вторичная
Третичная
4. Все нижеперечисленные содержат интеркалярную меристему, кроме _____________.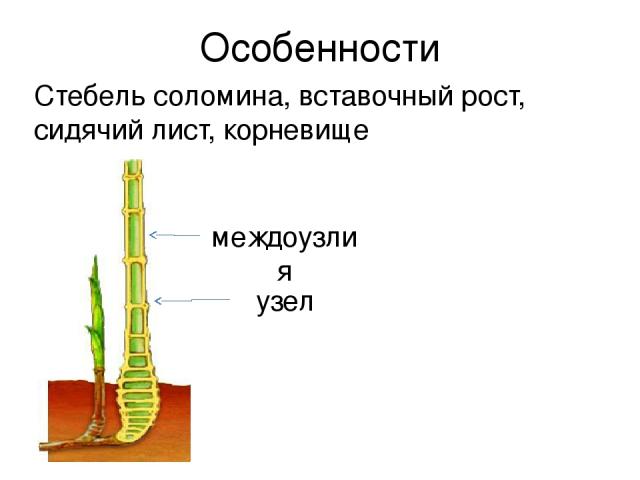
Основание узлов
Основание листовых пластин
Кончики корней
5. Какова общая функция интеркалярной меристемы?
Рост
Размножение
Хранение
Отправьте результаты (необязательно)
Ваше имя
На электронную почту
Next
Расширение интеркалярных гиф: новый механизм роста эндофитов у растений | Информационная система кормов
Поле поиска
You are here
Home » Овсяница тростниковая Онлайн Монография » Глава 14: Овсяница тростниковая — симбиоз эндофитов
Расширение интеркалярных гиф: новый механизм роста эндофитов у растений
Рост листьев травы в значительной степени зависит от деления клеток в зачатке листа и клеток непосредственно над ним, которые затем перестают делиться и проходят фазу быстрого расширения. Клетки травы увеличиваются в зоне расширения листа, области, которая охватывает несколько сантиметров от основания формирующегося листа (Snyder et al., 19).90). Растительные клетки в этой зоне расширяются в большей или меньшей степени, одновременно смещаясь от основания листа за счет продолжающегося производства клеток под ним. Лист эффективно растет у основания за счет вставочного расширения. Гифы эндофитов, присутствующие в зоне экспансии листа, по-видимому, прикреплены к клеточным стенкам хозяина (Christensen et al., 2008). Кроме того, гифы могут иметь боковые ответвления (рис. 14-25), некоторые из которых соединяют продольные гифы по обе стороны от рядов расширяющихся растительных клеток. Учитывая, что считается, что грибы растут исключительно за счет удлинения апикального кончика, до недавнего времени было неясно, как нити гиф и боковые ветви остаются интактными при прикреплении к увеличивающимся клеткам листа (Christensen et al., 2008; Christensen and Voisey, 2007). Гифы в зоне расширения листа распространяются не за счет апикального роста, а за счет интеркалярного роста по длине нити (Christensen et al.
Клетки травы увеличиваются в зоне расширения листа, области, которая охватывает несколько сантиметров от основания формирующегося листа (Snyder et al., 19).90). Растительные клетки в этой зоне расширяются в большей или меньшей степени, одновременно смещаясь от основания листа за счет продолжающегося производства клеток под ним. Лист эффективно растет у основания за счет вставочного расширения. Гифы эндофитов, присутствующие в зоне экспансии листа, по-видимому, прикреплены к клеточным стенкам хозяина (Christensen et al., 2008). Кроме того, гифы могут иметь боковые ответвления (рис. 14-25), некоторые из которых соединяют продольные гифы по обе стороны от рядов расширяющихся растительных клеток. Учитывая, что считается, что грибы растут исключительно за счет удлинения апикального кончика, до недавнего времени было неясно, как нити гиф и боковые ветви остаются интактными при прикреплении к увеличивающимся клеткам листа (Christensen et al., 2008; Christensen and Voisey, 2007). Гифы в зоне расширения листа распространяются не за счет апикального роста, а за счет интеркалярного роста по длине нити (Christensen et al.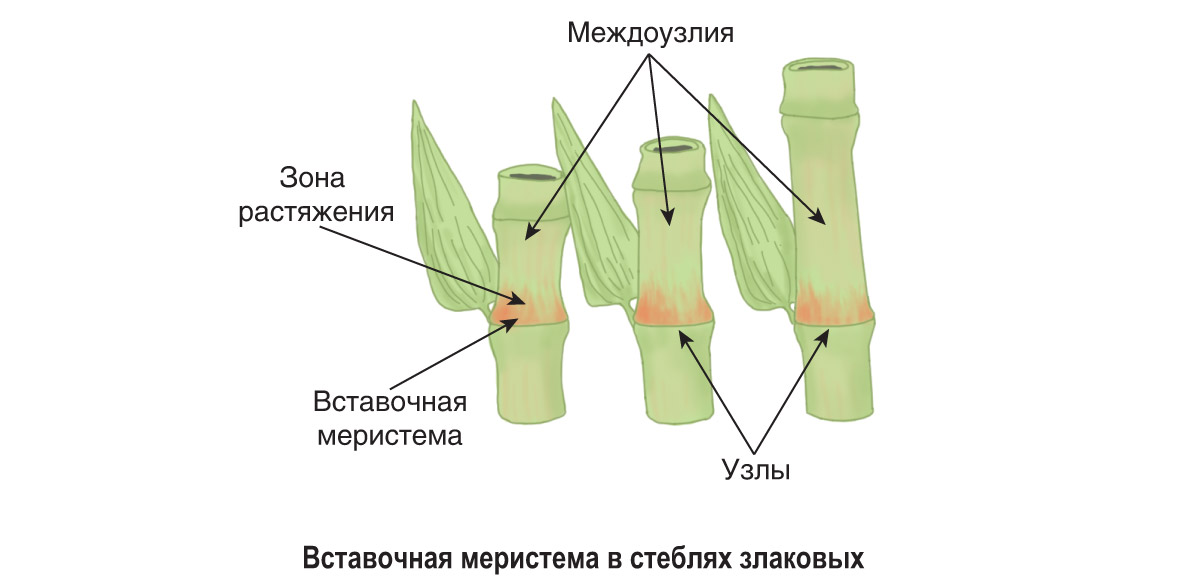 , 2008). Кумулятивное увеличение интеркалярной длины гиф позволяет эндофиту расти в зоне экспансии аналогично хозяину. Эта адаптация позволяет эндофитам, передаваемым семенами, преодолевать силы физического растяжения во время расширения листьев, которые в противном случае вызвали бы фрагментацию гиф.
, 2008). Кумулятивное увеличение интеркалярной длины гиф позволяет эндофиту расти в зоне экспансии аналогично хозяину. Эта адаптация позволяет эндофитам, передаваемым семенами, преодолевать силы физического растяжения во время расширения листьев, которые в противном случае вызвали бы фрагментацию гиф.
Построение стенки вдали от кончика гифы, существенное требование расширения интеркалярной гифы, является непрерывным процессом, о чем свидетельствует увеличение толщины стенки по мере старения гифы (Christensen et al., 2002).
Следующие данные подтверждают гипотезу о том, что гифы Epichloë и Neotyphodium растут интеркалярным расширением как в растущих листьях, так и в междоузлиях развивающихся репродуктивных побегов и, возможно, в других частях растений, где растительные клетки увеличиваются в размерах.
1. Боковые ветви и гифы, на которых они образовались, остаются нетронутыми внутри удлиняющихся листьев (Christensen et al., 2008).
2. Расстояние между ветвями на GFP-экспрессирующих гифах в зоне расширения листа удлиняющихся листьев увеличивается со временем, что измеряется с помощью покадровой фотографии (Christensen et al., 2008).
Расстояние между ветвями на GFP-экспрессирующих гифах в зоне расширения листа удлиняющихся листьев увеличивается со временем, что измеряется с помощью покадровой фотографии (Christensen et al., 2008).
3. Ультраструктура гиф в удлиняющихся листьях связана с расстоянием отделов от основания листа (возраст ткани листа), а не с расстоянием от кончика гифы, что позволяет предположить, что все гифы в одной и той же области листа находятся на одном уровне. аналогичная стадия развития (Christensen et al., 2008). Доказательства того, что гифы колонизируют каждую секцию листа в одно и то же время, согласуются с интеркалярным ростом гиф, обусловленным их прикреплением к расширяющимся вверх тканям листа.
Рис. 14-25. Разветвленные гифы штамма Epichloë festucae , экспрессирующего GFP, в зоне разрастания листа райграса пастбищного. Две темные продольные полосы указывают на положение сосудистых пучков. |
Leave A Comment