
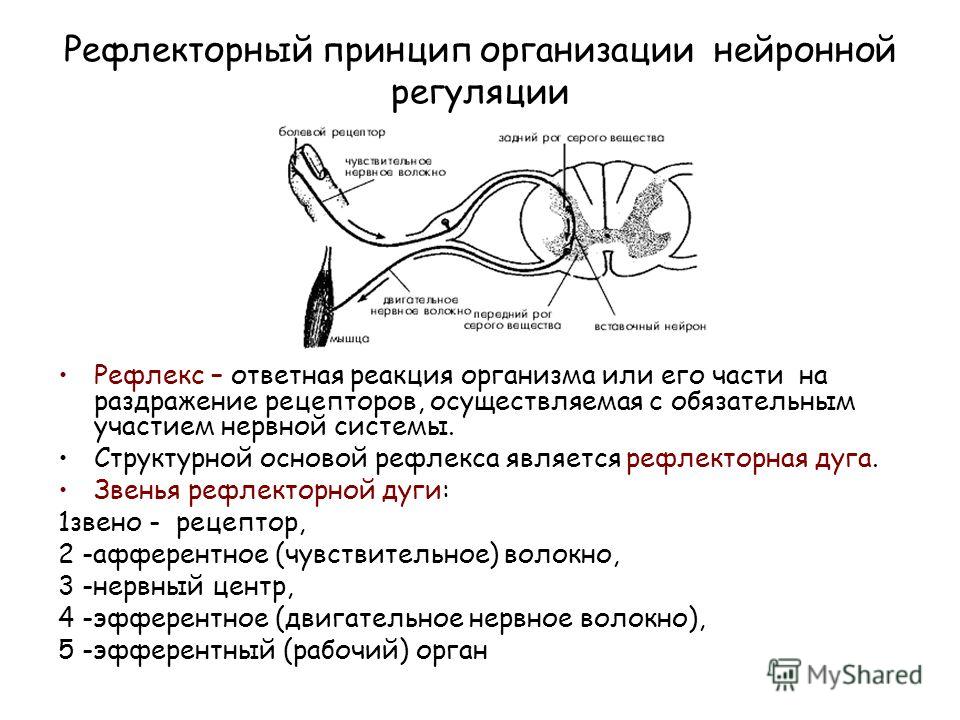
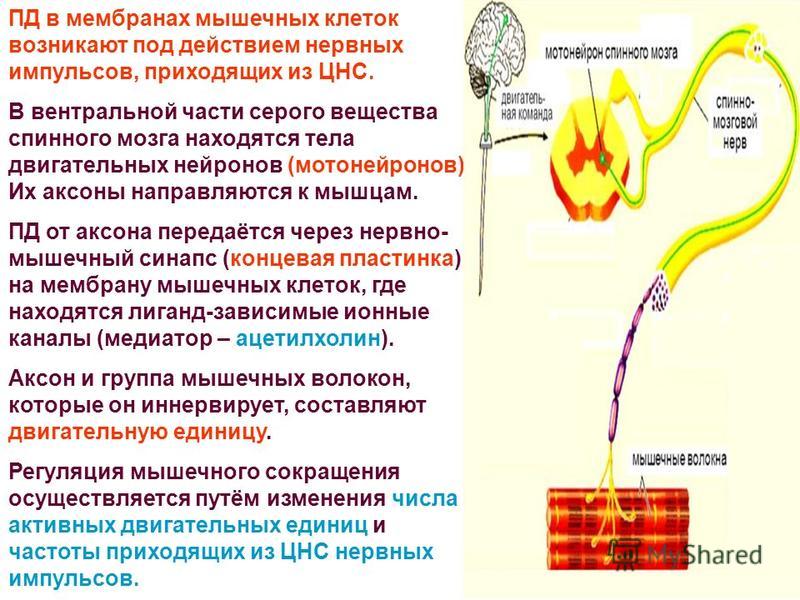
Нейрогуморальная регуляция
Импульсы от органа в ЦНС проводят:
чувствительные нейроны
вставочные нейроны
двигательные нейроны
Синапсом называется:
нервный узел
контакт между нейронами
нервное сплетеие
Нервные узлы образованы:
аксонами
нервами
телами нейронов
Нервный импульс — это результат:
электрохимических процессов
тепловых процессов, происходящих в клетках
механических процессов
Нервный импульс вызывает в соседнем нейроне:
возбуждение и торможение одновременно
возбуждение
возбуждение или торможение
В промежуточном мозге расположен центр регуляции:
пищеварения
терморегуляции
сердцебиения
В каком отделе головного мозга находятся центры регуляции дыхания, пищеварения:
мозжечок
кора головного мозга
продолговатый мозг
Возбуждение от ЦНС к органу или железам передается по:
исполнительным нейронам
чувствительным нейронам
вставочным нейронам
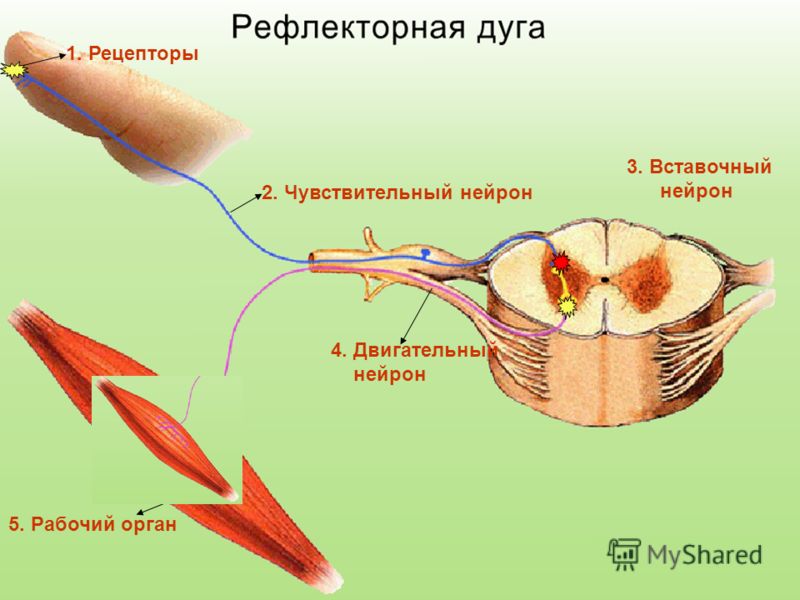
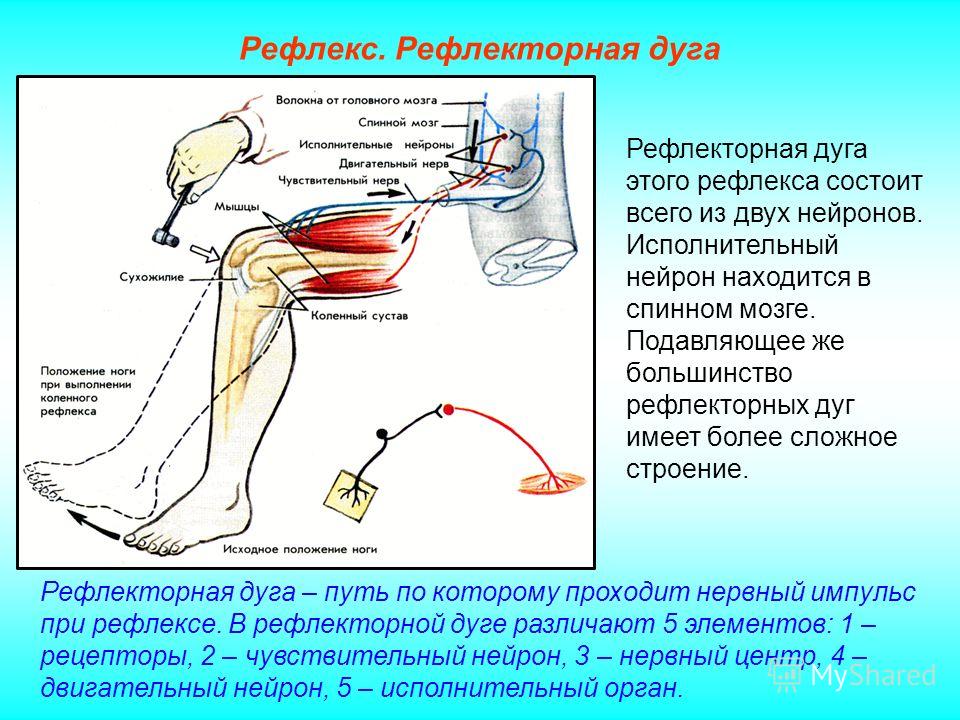
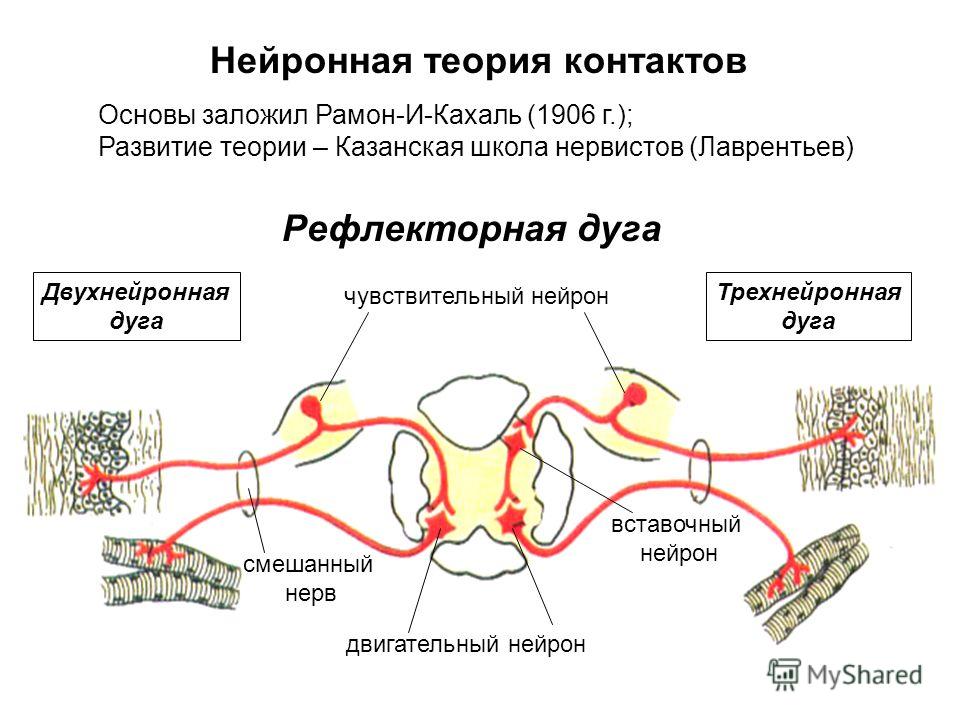
Дугу спинномозгового рефлекса составляют:
мышца — чувствительный нейрон — рецептор — вставочный нейрон — исполнительный нейрон
рецептор — исполнительный нейрон — вставочный нейрон — чувствительный нейрон — мышца
рецептор — чувствительный нейрон — вставочный нейрон — исполнительный нейрон — мышца
При ожоге возбуждение возникает в(во):
теле чувствительного нейрона
рецепторе чувствительного нейрона
теле исполнительного нейрона
Слюноотделение у человека будет безусловно-рефлекторной реакцией:
во время еды
при разговоре о еде
при виде любимого блюда
Учение об условных рефлексах создал:
Сеченов
Павлов
Мечников
Сколько из названных желез имеет специальные протоки: слюнная, щитовидная, гипофиз, потовая, надпочечник:
две
пять
три
К железам смешанной секреции относится:
гипофиз
надпочечники
поджелудочная железа
Гормоны — это:
биологически активные вещества, вырабатываемые организмом
белки, катализирующие химические реакции
соединения белков и витаминов
Тироксин — это гормон:
надпочечников
щитовидной железы
поджелудочной железы
Избыток или недостаток гормонов в крови воспринимается:
мозжечком
центрами спинного мозга
гипоталамусом
В клетках щитовидной железы больше, чем в остальных тканях, содержится:
глюкозы
йода
кальция
Гигантизм связан с нарушением функций:
щитовидной железы
вилочковой железы
гипофиза
Центральная нервная система образована:
спинным мозгом и спинномозговыми нервами
головным и спинным мозгом
нервами, нервными сплетениями и узлами
Основными свойствами нервной клетки являются:
возбудимость и проводимость
сократимость и проводимость
возбудимость и сократимость
Начальный этап рефлекторной дуги в слюноотделительном рефлексе:
исполнительный нейрон
рецептор
вставочный нейрон
Высшим центром контроля нейрогуморальной регуляции организма человека является:
спинной мозг
продолговатый мозг
гипоталамус
Выделение желудочного сока в ответ на раздражение пищей рецепторов ротовой полости:
саморегуляция
безусловный рефлекс
торможение
Условный рефлекс будет прочным, если условный раздражитель:
не подкрепляется безусловным раздражителем
нерегулярно подкрепляется безусловным раздражителем
постоянно подкрепляется безусловным раздражителем
По чувствительному нейрону возбуждение направляется:
к рецепторам
в центральную нервную систему
к исполнительному органу
Печень относят к железам внешней секреции, так как:
желчь выделяется из неё в полость кишечника
в ней глюкоза превращается в гликоген
в ней обезвреживаются ядовитые вещества
Какую функцию выполняет вилочковая железа:
вырабатывает адреналин
регулирует иммунитет
секретирует инсулин
В ЦНС импульс с чувствительного нейрона на двигательный передаётся:
через рецепторы
по нейроглии
по ставочному нейрону
Рефлекторная дуга заканчивается:
чувствительным нейроном
исполнительным органом
вставочным нейроном
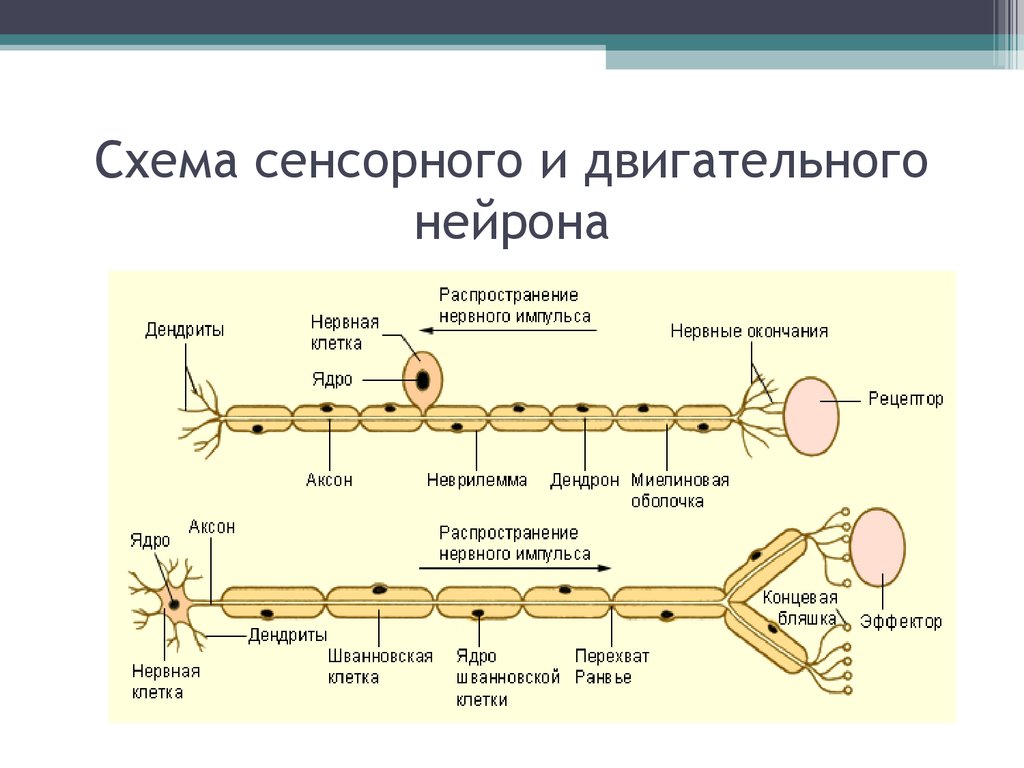
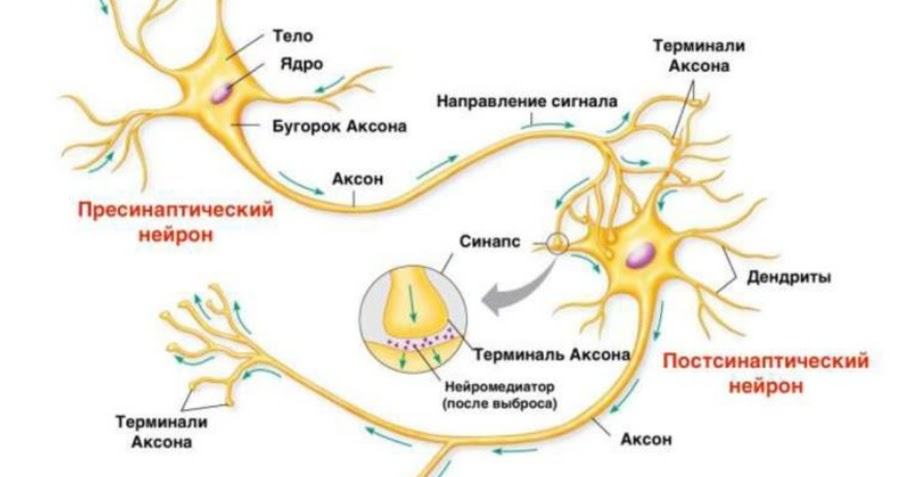
Распространение нервных импульсов • Джеймс Трефил, энциклопедия «Двести законов мироздания»
Нервные импульсы распространяются при перемещении ионов через мембрану нервной клетки и передаются из одной нервной клетки в другую с помощью нейромедиаторов.
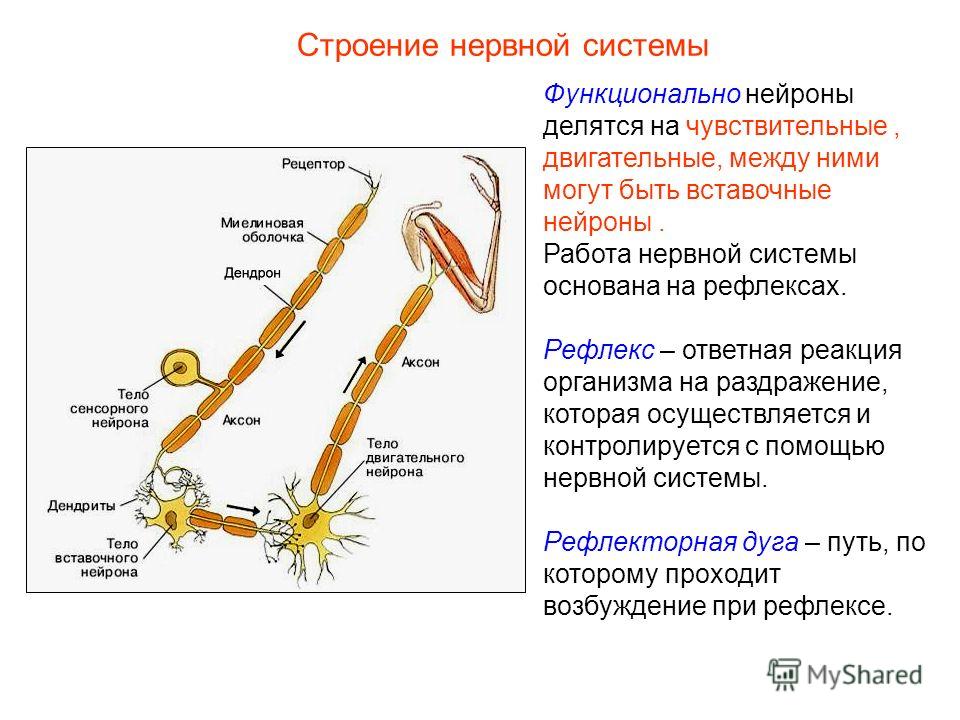
В результате эволюции нервной системы человека и других животных возникли сложные информационные сети, процессы в которых основаны на химических реакциях. Важнейшим элементом нервной системы являются специализированные клетки нейроны. Нейроны состоят из компактного тела клетки, содержащего ядро и другие органеллы. От этого тела отходит несколько разветвленных отростков. Большинство таких отростков, называемых дендритами, служат точками контакта для приема сигналов от других нейронов. Один отросток, как правило самый длинный, называется аксоном и передает сигналы на другие нейроны. Конец аксона может многократно ветвиться, и каждая из этих более мелких ветвей способна соединиться со следующим нейроном.
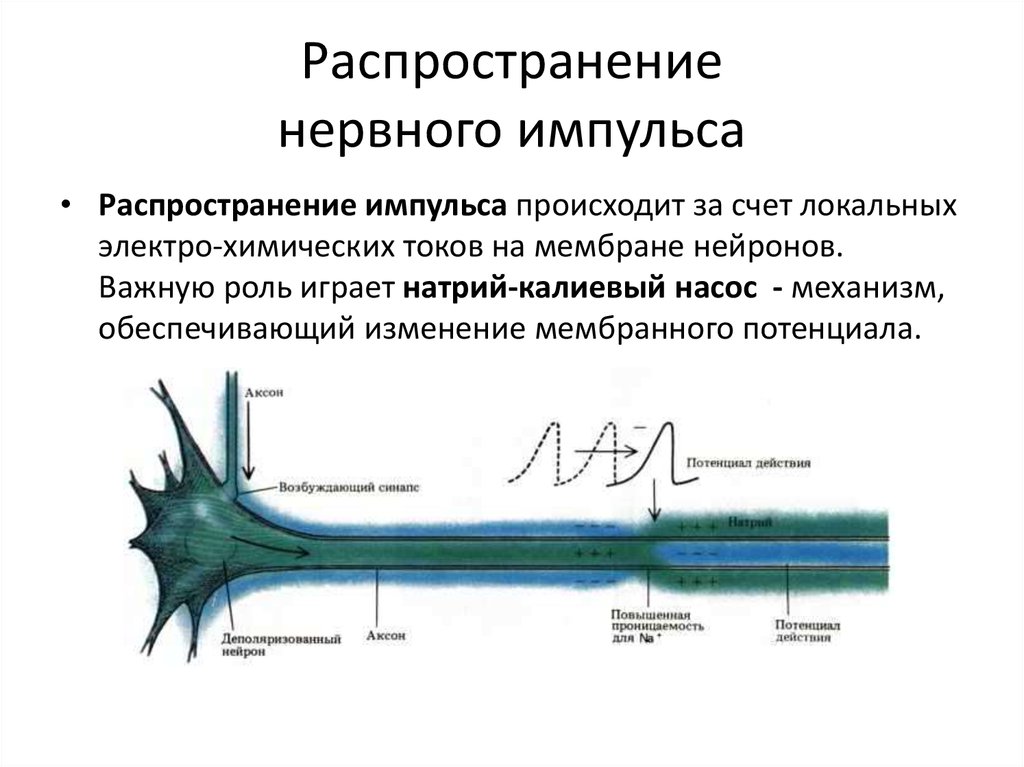
Во внешнем слое аксона находится сложная структура, образованная множеством молекул, выступающих в роли каналов, по которым могут поступать ионы — как внутрь, так и наружу клетки. Один конец этих молекул, отклоняясь, присоединяется к атому-мишени. После этого энергия других частей клетки используется на то, чтобы вытолкнуть этот атом за пределы клетки, тогда как процесс, действующий в обратном направлении, вводит внутрь клетки другую молекулу.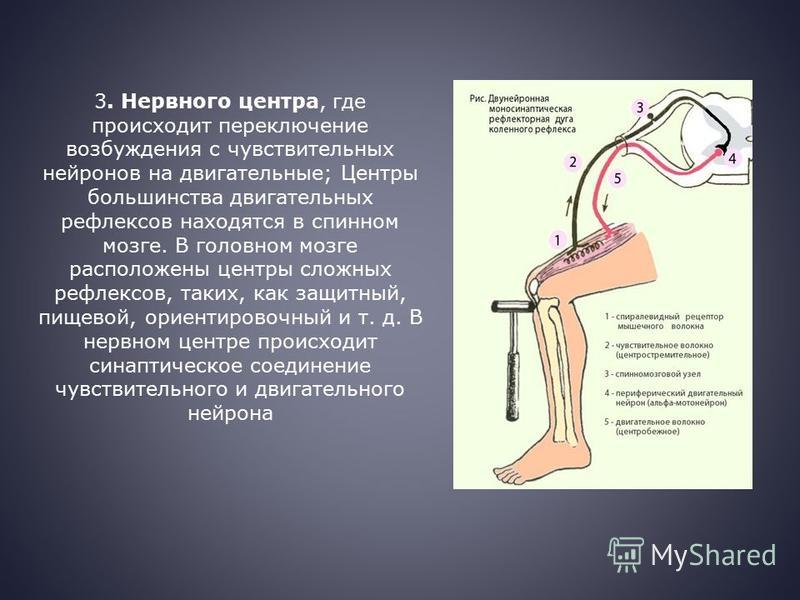 Наибольшее значение имеет молекулярный насос, который выводит из клетки ионы натрия и вводит в нее ионы калия (натрий-калиевый насос).
Наибольшее значение имеет молекулярный насос, который выводит из клетки ионы натрия и вводит в нее ионы калия (натрий-калиевый насос).
Когда клетка находится в покое и не проводит нервных импульсов, натрий-калиевый насос перемещает ионы калия внутрь клетки и выводит ионы натрия наружу (представьте себе клетку, содержащую пресную воду и окруженную соленой водой). Из-за такого дисбаланса разность потенциалов на мембране аксона достигает 70 милливольт (приблизительно 5% от напряжения обычной батарейки АА).
Однако при изменении состояния клетки и стимуляции аксона электрическим импульсом равновесие на мембране нарушается, и натрий-калиевый насос на короткое время начинает работать в обратном направлении. Положительно заряженные ионы натрия проникают внутрь аксона, а ионы калия откачиваются наружу. На мгновение внутренняя среда аксона приобретает положительный заряд. При этом каналы натрий-калиевого насоса деформируются, блокируя дальнейший приток натрия, а ионы калия продолжают выходить наружу, и исходная разность потенциалов восстанавливается.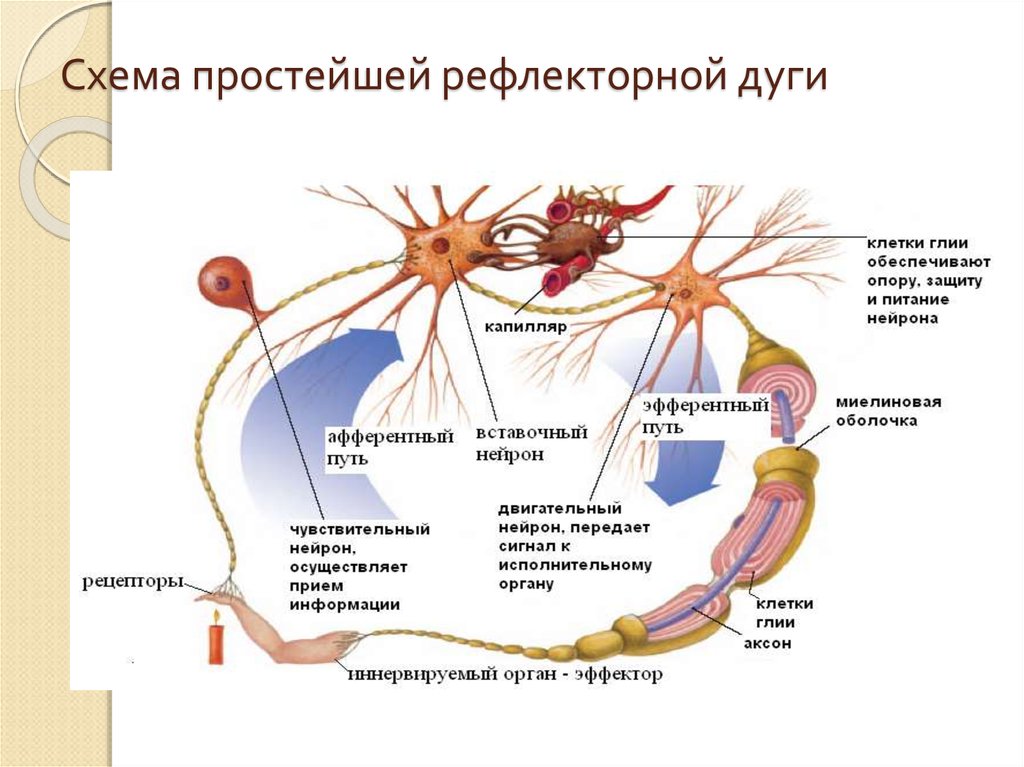 Тем временем ионы натрия распространяются внутри аксона, изменяя мембрану в нижней части аксона. При этом состояние расположенных ниже насосов меняется, способствуя дальнейшему распространению импульса. Резкое изменение напряжения, вызванное стремительными перемещения ионов натрия и калия, называют потенциалом действия. При прохождении потенциала действия через определенную точку аксона, насосы включаются и восстанавливают состояние покоя.
Тем временем ионы натрия распространяются внутри аксона, изменяя мембрану в нижней части аксона. При этом состояние расположенных ниже насосов меняется, способствуя дальнейшему распространению импульса. Резкое изменение напряжения, вызванное стремительными перемещения ионов натрия и калия, называют потенциалом действия. При прохождении потенциала действия через определенную точку аксона, насосы включаются и восстанавливают состояние покоя.
Потенциал действия распространяется довольно медленно — не более доли дюйма за секунду. Для того чтобы увеличить скорость передачи импульса (поскольку, в конце концов, не годится, чтобы сигнал, посланный мозгом, достигал руки лишь через минуту), аксоны окружены оболочкой из миелина, препятствующей притоку и оттоку калия и натрия. Миелиновая оболочка не непрерывна — через определенные интервалы в ней есть разрывы, и нервный импульс перескакивает из одного «окна» в другое, за счет этого скорость передачи импульса возрастает.
Когда импульс достигает конца основной части тела аксона, его необходимо передать либо следующему нижележащему нейрону, либо, если речь идет о нейронах головного мозга, по многочисленным ответвлениям многим другим нейронам. Для такой передачи используется абсолютно иной процесс, нежели для передачи импульса вдоль аксона. Каждый нейрон отделен от своего соседа небольшой щелью, называемой синапсом. Потенциал действия не может перескочить через эту щель, поэтому нужно найти какой-то другой способ для передачи импульса следующему нейрону. В конце каждого отростка имеются крошечные мешочки, называющие (пресинаптическими) пузырьками, в каждом из которых находятся особые соединения — нейромедиаторы. При поступлении потенциала действия из этих пузырьков высвобождаются молекулы нейромедиаторов, пересекающие синапс и присоединяющиеся к специфичным молекулярным рецепторам на мембране нижележащих нейронов. При присоединении нейромедиатора равновесие на мембране нейрона нарушается. Сейчас мы рассмотрим, возникает ли при таком нарушении равновесия новый потенциал действия (нейрофизиологи продолжают искать ответ на этот важный вопрос до сих пор).
Для такой передачи используется абсолютно иной процесс, нежели для передачи импульса вдоль аксона. Каждый нейрон отделен от своего соседа небольшой щелью, называемой синапсом. Потенциал действия не может перескочить через эту щель, поэтому нужно найти какой-то другой способ для передачи импульса следующему нейрону. В конце каждого отростка имеются крошечные мешочки, называющие (пресинаптическими) пузырьками, в каждом из которых находятся особые соединения — нейромедиаторы. При поступлении потенциала действия из этих пузырьков высвобождаются молекулы нейромедиаторов, пересекающие синапс и присоединяющиеся к специфичным молекулярным рецепторам на мембране нижележащих нейронов. При присоединении нейромедиатора равновесие на мембране нейрона нарушается. Сейчас мы рассмотрим, возникает ли при таком нарушении равновесия новый потенциал действия (нейрофизиологи продолжают искать ответ на этот важный вопрос до сих пор).
После того как нейромедиаторы передадут нервный импульс от одного нейрона на следующий, они могут просто диффундировать, или подвергнуться химическому расщеплению, или вернуться обратно в свои пузырьки (этот процесс нескладно называется обратным захватом). В конце XX века было сделано поразительное научное открытие — оказывается, лекарства, влияющие на выброс и обратный захват нейромедиаторов, могут коренным образом изменять психическое состояние человека. Прозак (Prozac*) и сходные с ним антидепрессанты блокируют обратный захват нейромедиатора серотонина. Складывается впечатление, что болезнь Паркинсона взаимосвязана с дефицитом нейромедиатора допамина в головном мозге. Исследователи, изучающие пограничные состояния в психиатрии, пытаются понять, как эти соединения влияют на человеческий рассудок.
В конце XX века было сделано поразительное научное открытие — оказывается, лекарства, влияющие на выброс и обратный захват нейромедиаторов, могут коренным образом изменять психическое состояние человека. Прозак (Prozac*) и сходные с ним антидепрессанты блокируют обратный захват нейромедиатора серотонина. Складывается впечатление, что болезнь Паркинсона взаимосвязана с дефицитом нейромедиатора допамина в головном мозге. Исследователи, изучающие пограничные состояния в психиатрии, пытаются понять, как эти соединения влияют на человеческий рассудок.
По-прежнему нет ответа на фундаментальный вопрос о том, что же заставляет нейрон инициировать потенциал действия — выражаясь профессиональным языком нейрофизиологов, неясен механизм «запуска» нейрона. В этом отношении особенно интересны нейроны головного мозга, которые могут принимать нейромедиаторы, посланные тысячей соседей. Об обработке и интеграции этих импульсов почти ничего не известно, хотя над этой проблемой работают многие исследовательские группы. Нам известно лишь, что в нейроне осуществляется процесс интеграции поступающих импульсов и выносится решение, следует или нет инициировать потенциал действия и передавать импульс дальше. Этот фундаментальный процесс управляет функционированием всего головного мозга. Неудивительно, что эта величайшая загадка природы остается, по крайней мере сегодня, загадкой и для науки!
Нам известно лишь, что в нейроне осуществляется процесс интеграции поступающих импульсов и выносится решение, следует или нет инициировать потенциал действия и передавать импульс дальше. Этот фундаментальный процесс управляет функционированием всего головного мозга. Неудивительно, что эта величайшая загадка природы остается, по крайней мере сегодня, загадкой и для науки!
Нервная система
Нервная система Нервная система — самая сложная система организма, но она
очень консервативен в плане изменений
Нервная система определяет реакции организма на изменения внутренних
и внешние среды
Он также действует как мессенджер и система координации для тела.
Основными компонентами нервной системы являются:
периферическая нервная система — черепные и спинномозговые нервы Все отделы нервной системы состоят из общей клеточной субъединицы — нейрон
Нейроны
Нейроны происходят из эктодермы нервной трубки, нервного гребня
клетки или эктодермальные плакоды.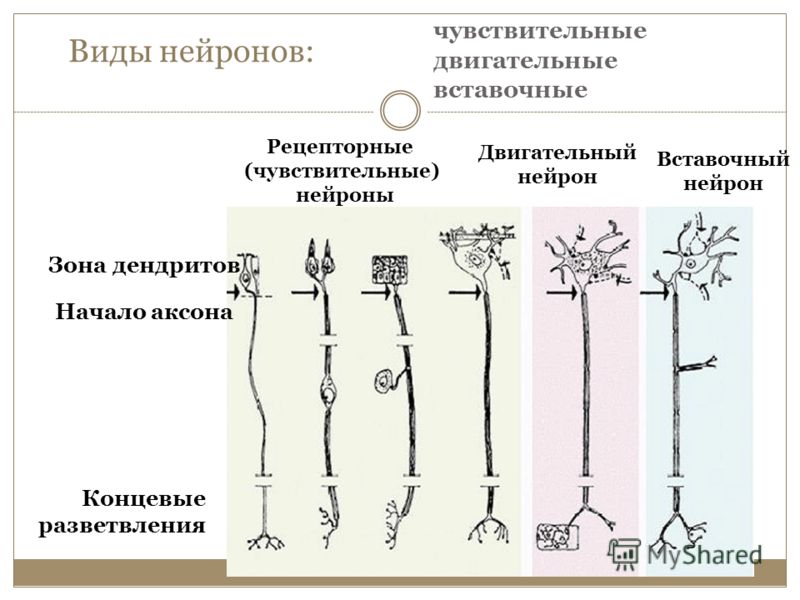
Частями нейрона являются (рис. 16.2, стр. 590):
Нервный импульс представляет собой электрическое явление, которое проходит как волна вдоль поверхностной оболочки нервного волокна
В норме нервная клетка находится в состоянии покоя, исходя из концентрации ионов натрия и калия внутри и вне клетки
Во время потенциала действия нейрон проходит несколько этапов: 1.
 Высокая концентрация ионов натрия снаружи; отрицательный заряд внутри
клетка по сравнению с внешней
Высокая концентрация ионов натрия снаружи; отрицательный заряд внутри
клетка по сравнению с внешней 3. Нервный импульс проходит по аксону в виде волны деполяризации.
4. Натрий выкачивается из клетки и восстанавливается потенциал покоя Между нейронами нервный импульс должен проходить через пространство или синапс. телодендрии содержат синаптические везикулы, содержащие нейротрансмиттера. (например, ацетилхолин, норадреналин, серотонин, дофамин)
Нейротрансмиттеры высвобождаются, когда нервный импульс достигает телодендриев, затем пересекают синаптическую щель и достигают дендритов следующего нейрона. в линии, вызывающей посылку импульса постсинаптической нервной клеткой
- Шванновские клетки (рис.
 16.4, стр. 591) — клетки происхождения нервного гребня
которые образуют тонкую оболочку, окружающую немиелинизированный аксон, или, после
миелинизировав аксон, лежит на поверхности миелиновой оболочки —
миелиновая оболочка действует как изолятор, увеличивая скорость передачи
проведения нервного импульса по аксону
16.4, стр. 591) — клетки происхождения нервного гребня
которые образуют тонкую оболочку, окружающую немиелинизированный аксон, или, после
миелинизировав аксон, лежит на поверхности миелиновой оболочки —
миелиновая оболочка действует как изолятор, увеличивая скорость передачи
проведения нервного импульса по аксону - Нейроглия — клетки центральной нервной системы, помогающие поддерживать, защищать, питать и поддерживать нейроны (рис. 16.1, стр. 590)
- Олигодендроциты — нейроглиальные клетки эктодермального происхождения, миелинизирующие аксонов в центральной нервной системе и образует белое вещество центральной нервная система (немиелиновые аксоны серого вещества
- Астроциты — звездчатые питательные и поддерживающие клетки глии центральная нервная система
- Микроглия — мелкие нейроглиальные клетки мезодермального происхождения, часть которые являются фагоцитами
 16.4, стр. 591) — клетки происхождения нервного гребня
которые образуют тонкую оболочку, окружающую немиелинизированный аксон, или, после
миелинизировав аксон, лежит на поверхности миелиновой оболочки —
миелиновая оболочка действует как изолятор, увеличивая скорость передачи
проведения нервного импульса по аксону
16.4, стр. 591) — клетки происхождения нервного гребня
которые образуют тонкую оболочку, окружающую немиелинизированный аксон, или, после
миелинизировав аксон, лежит на поверхности миелиновой оболочки —
миелиновая оболочка действует как изолятор, увеличивая скорость передачи
проведения нервного импульса по аксонуОбщие части нейронной схемы включают три основных типа нейронов: Первичные сенсорные нейроны или афферентные нейроны: несут импульсы из свободных нервных окончаний или рецепторных клеток в центральную нервную систему
Моторные или эфферентные нейроны: передают импульсы от центральной нервной системы к эффекторам, таким как мышцы или железы
Интернейроны: получают сигналы от сенсорных нейронов, интегрируются информацию и посылают сигналы двигательным нейронам
Соматические волокна относятся к коже и ее производным, а также к произвольные мышцы
Висцеральные волокна
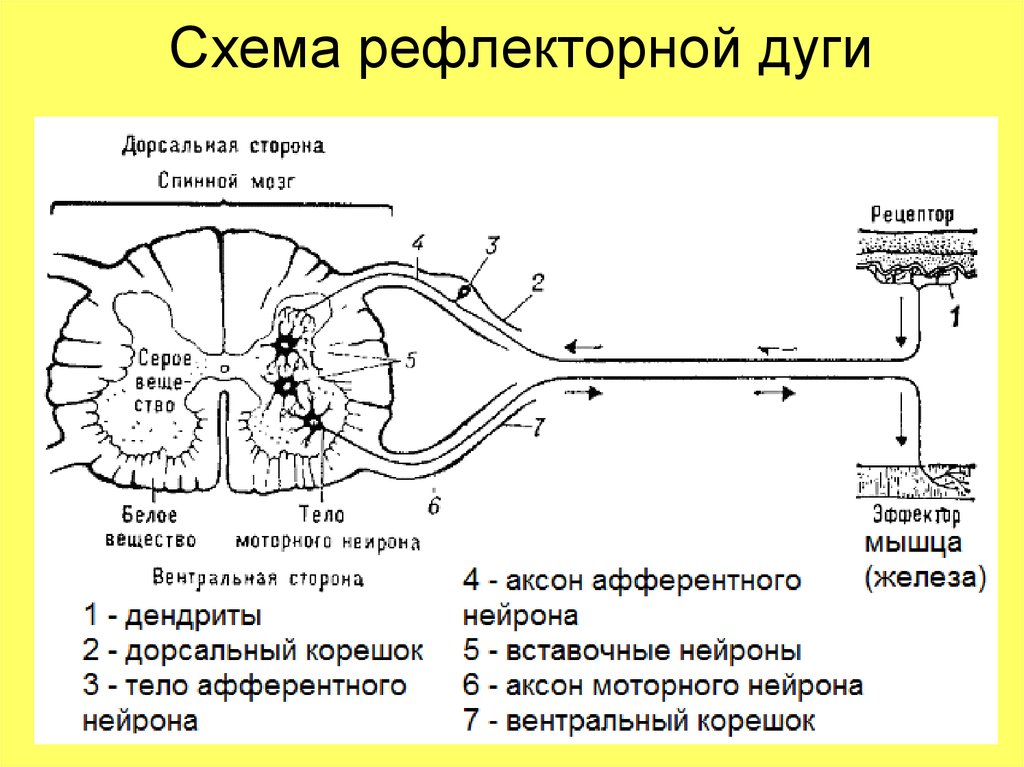
В простейшей рефлекторной дуге передаются сообщения от рецепторных органов внутри спинного мозга непосредственно от афферентных волокон к эфферентным волокнам, которые затем посылают соответствующие сообщения эффекторным органам
Функция спинного мозга – принимать входящие импульсы, интегрировать и координировать их, передавать их туда, куда они должны идти в пределах центральную нервную систему и посылают ответы в периферическую нервную систему по мере необходимости
Общая структура спинного мозга лучше всего видна на поперечном сечении спинного мозга амниоты (рис.
серое вещество напоминает букву Н, а плечи называются спинные столбы или рога, а нижние конечности называются брюшными столбами или рога
серая спайка образует поперечину буквы H и передает волокна с одной стороны спинного мозга на другую
внешнее белое вещество разделено на правую и левую части дорсомедиальная борозда и вентральная борозда
задний рог спинного мозга получает окончания первичных сенсорных нейроны
брюшной рог содержит дендриты и тела двигательных нейроны
Другие рефлексы включают межсегментарных рефлекса , которые включают нейроны, перекрещивающиеся на другой стороне тела — перекрест это пересечение нервных путей по средней линии центральной нервной система – межсегментарные рефлексы отвечают за поддержание координированных движений, таких как плавание или ходьба.
Условные рефлексы являются врожденными рефлексами, которые вырабатываются в результате повторяющихся переживаний животных Пути от низших к высшим центрам мозга Использование восходящих путей в спинном мозге — большинство восходящих импульсов перекрестить по пути Пути от мозга к нижним центрам Импульсы используют нисходящие пути в спинном мозге, которые могут перекрестить в головном мозге перед путешествием к соответствующей мышце
Генераторы центральных паттернов , группы нейронов в спинном мозге шнуре и в головном мозге, активность которого отвечает за врожденную цикличность движения частей тела — центральные генераторы паттернов не требуют продолжения сенсорный вход, чтобы вызвать реакцию
У более примитивных видов спинной и брюшной корешки образуют отдельные спинной и вентральный спинномозговые нервы
У всех других позвоночных спинной и брюшной нервы соединяются в один спинномозговой нерв с чувствительными волокнами, входящими через задний корешок (ветвь) и двигательные волокна, отходящие через передний корешок (ветвь)
Спинномозговые нервы определяются по их расположению и включают шейные, грудные, поясничный, крестцовый и хвостовой нервы — более хвостовые спинномозговые нервы образуют пучок, известный как
Сплетения представляют собой сети нервов или кровеносных сосудов, образованные до появления нервов распределяются по мышцам
шейное сплетение иннервирует брюшные мышцы шеиплечевое сплетение кровоснабжает грудной придаток
пояснично-крестцовое сплетение кровоснабжает тазовый придаток
копчиковое сплетение иннервирует некоторые другие мышцы таза Черепные нервы
В то время как спинномозговые нервы однородны по происхождению, конфигурация корешков и ветвей, компоненты нервных волокон и связь с центральной нервной система, черепные нервы не
Черепные нервы могут присутствовать у одних позвоночных и отсутствовать у других, могут разделиться в ходе эволюции, чтобы стать двумя, или слиться, чтобы стать один
Черепные нервы относятся к одной из трех основных категорий: 1) Последовательно с дорсальными корешками спинномозговых нервов присоединяются к стволу мозга на латеральном (не вентральном) уровне
включают смешанные нервы, содержащие комбинацию чувствительных и двигательных нервов.
 нейроны
нейроны включает нервы V, VII, IX, X, XI 2) Последовательно с вентральными спинномозговыми нервами присоединиться к стволу мозга на вентральном уровне
содержат соматические двигательные волокна и иннервируют бранхиометрические мышцы, так иногда называют бранхиометрические нервы
включает нервы III, IV, VI, XII 3) Нет аналога в ряду позвоночника, потому что его нервы обслуживают структуры присущие голове (нос, глаз, ухо, система боковой линии) сенсорные волокна
включает нервы 0, I, II, VIII Черепные нервы следующие:

Вегетативная нервная система
Вегетативная система структурно или функционально не изолирована от
центральной или периферической нервной системы.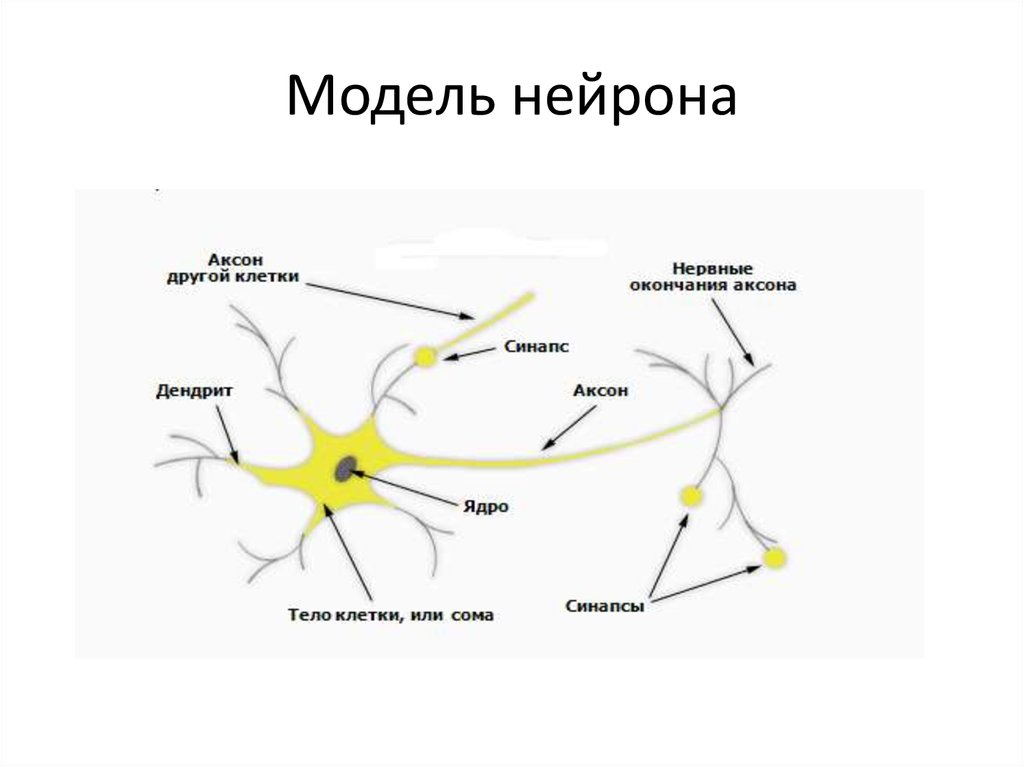 Однако это касается исключительно
к непроизвольным функциям организма. Включает только висцеральные волокна.
Однако это касается исключительно
к непроизвольным функциям организма. Включает только висцеральные волокна.
Отличительными признаками вегетативной нервной системы являются:
1. Каждый путь включает нейрон, имеющий тело клетки внутри ЦНС и тела нейрона вне ЦНС.У млекопитающих постганглионарные парасимпатические волокна секретируют ацетилхолин. и называются холинергическими волокнами. Постганглионарные симпатические волокна (которые вызывают реакцию «бей или беги») секретируют норадреналин (норэпинефрин) и называются адренергическими волокнами. Последствия стимуляции двух системы показаны в таблице 12-3, с. 481 в тексте и на рисунке вегетативной нервной системы млекопитающих в раздаточном материале.Волокна между ганглиями и ЦНС преганглионарные и миелинизированные. Волокна между ганглиями и конечными органами постганглионарные и немиелинизированные2. Делящиеся на несколько наборов волокон Симпатическая — часть вегетативной нервной системы, выходящая ЦНС из отделов спинного мозга. Активность симпатической нервной системы помогает животному приспособиться к стрессу, стимулируя физиологические процессы, которые увеличение энергии, доступной тканям организма. Также называется грудопоясничным оттоком.
Парасимпатическая — часть вегетативной нервной системы, покидает ЦНС от черепных и крестцовых нервов. Способствует метаболическим процессам которые производят и хранят энергию.
Enteric — Сложная сеть, образованная нейронами, расположенными внутри стенки кишки. Активируется непосредственно местными физическими и химическими раздражителями и промежуточные местные рефлексы.
 Также называется краниосакральным оттоком.
Также называется краниосакральным оттоком. Мозг
Мозг является самым сложным органом в организме и сложным,
полностью сформированный мозг является одним из производных признаков позвоночных.
Развитие
К тому времени, когда нервные складки смыкаются над нейроцелем, передняя
часть нервной трубки начала увеличиваться в диаметре и становиться
отличается от остальной части нервной трубки
Развивающийся мозг расширяется на трех уровнях по мере разделения пузырьков
друг от друга перетяжками:
передний мозг — передний мозгЭти три региона затем разделяются на несколько дополнительных регионов
средний мозг — средний мозг
ромбовидный мозг — задний мозг
Передняя часть прозэнцефалона развивается в
- конечный мозг за счет расширения боковых стенок будет расти для формирования полушарий головного мозга
- задняя часть прозэнцефалона, переходящая в промежуточный мозг
 и воронка, которая станет частью гипофиза или нейрогипофиз
и воронка, которая станет частью гипофиза или нейрогипофиз Конечный мозг также будет дифференцироваться спереди, чтобы сформировать обонятельный луковицы
Ромбовидный мозг образует передний средний мозг (который формирует взрослый мозжечок) и задний продолговатый мозг
Внутри отдела головного мозга находятся полости, называемые везикулы , которые впоследствии разрастаются, образуя расширения или желудочков .
- Боковые желудочки занимают полушария большого мозга.
- Третий желудочек находится в промежуточном мозге и соединяется с латеральным желудочки через отверстие Мунро.
- Четвертый желудочек располагается в пределах промежуточного и продолговатого мозга и соединяется с третьим желудочком сильвиевым водопроводом.
- Цереброспинальная жидкость содержится в желудочках и действует как дренажная система, а также источник питания для мозга.
- Головной изгиб находится в среднем мозге и вогнут вентрально
- Изгиб моста находится в среднем мозге и вогнут дорсально
- Шейный изгиб находится в задней части продолговатого мозга. и вогнут вентрально.
 и вогнут вентрально.
и вогнут вентрально.Ствол головного мозга – это первая область, формирующаяся в процессе развития, наименее переменная, и получает все черепные нервы (кроме конечного и обонятельные нервы). Часть взрослого среднего мозга и весь промежуточный мозг , mesencephalon и myelencephalon входят в состав ствола головного мозга. Ствол мозга контролирует большинство вегетативных функций организма и, таким образом, является жизненно важным для жизни.
мозжечок и мост (вентральная часть среднего мозга птиц и млекопитающих, имеющих полосу поперечных волокон) являются основными взрослые производные метэнцефалона. Мозжечок и мост участвуют координации двигательной функции.
Головной мозг является взрослой производной конечного мозга, и доминирует над мозгом как по размеру, так и по контролю.
Мозг взрослого человека окружают слои соединительной ткани мезодермального происхождения.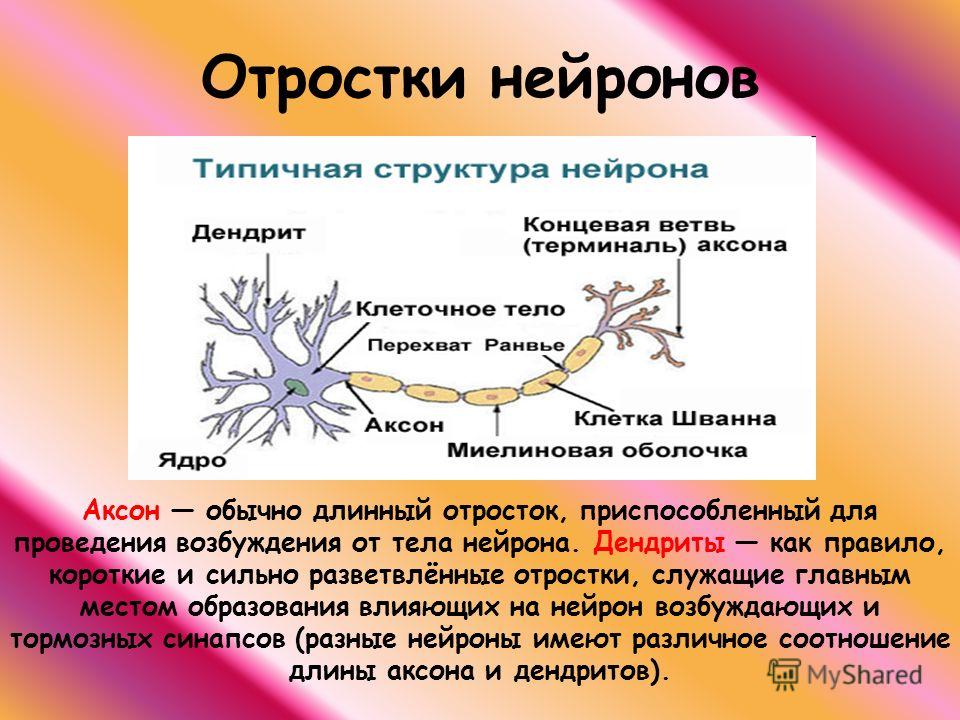 ткань под названием мозговые оболочки (единственное число: мозговые оболочки). В то время как круглоротые и
рыбы имеют только одну оболочку, называемую примитивной мозговой оболочкой ,
амфибии имеют два слоя, состоящих из наружной твердой мозговой оболочки , которая
чрезвычайно плотный и защитный, а мягкая паутинная оболочка или вторичная
мозговая оболочка более нежная и сосудистая. Млекопитающие имеют три мозговые оболочки: годовых
материя (которая следует за всеми извилинами мозга и является самой
интерьер), арахноидальный слой (нежный и направляет нити
к мягкой мозговой оболочке) и твердой мозговой оболочке (внешняя, более защитная
мозговые оболочки). Область между твердой мозговой оболочкой и паутинной оболочкой называется
субдуральное пространство; область между паутинной оболочкой называется
субарахноидальное пространство. Дополнительный слой ткани лежит между двумя
полушарий большого мозга и называется серпом большого мозга.
ткань под названием мозговые оболочки (единственное число: мозговые оболочки). В то время как круглоротые и
рыбы имеют только одну оболочку, называемую примитивной мозговой оболочкой ,
амфибии имеют два слоя, состоящих из наружной твердой мозговой оболочки , которая
чрезвычайно плотный и защитный, а мягкая паутинная оболочка или вторичная
мозговая оболочка более нежная и сосудистая. Млекопитающие имеют три мозговые оболочки: годовых
материя (которая следует за всеми извилинами мозга и является самой
интерьер), арахноидальный слой (нежный и направляет нити
к мягкой мозговой оболочке) и твердой мозговой оболочке (внешняя, более защитная
мозговые оболочки). Область между твердой мозговой оболочкой и паутинной оболочкой называется
субдуральное пространство; область между паутинной оболочкой называется
субарахноидальное пространство. Дополнительный слой ткани лежит между двумя
полушарий большого мозга и называется серпом большого мозга.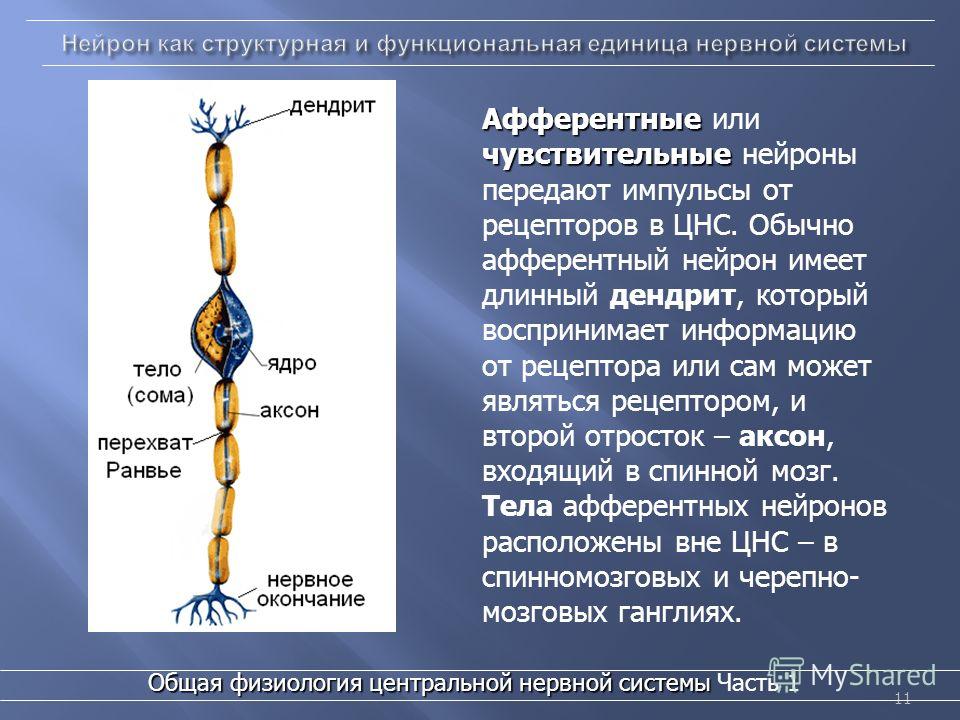
Задний ствол мозга: от продолговатого до среднего мозга
Задний ствол мозга является местом соединения черепных
нервов в центральную нервную систему. Каждый черепной нерв имеет ядро
в заднем стволе мозга для каждого типа волокон, которые он несет (например, соматических
сенсорная, висцеральная сенсорная, соматодвигательная, висцеромоторная).
Ретикулярная формация встречается у всех позвоночных и является сеть коротких интернейронов в стволе мозга, образующая примитивную интегрирующую система. Он проецируется в головной мозг, мозжечок, черепные ядра и спинной мозг и необходим для сознания, а также для контроля сердечно-сосудистой и дыхательной систем.
Ядро ruber и черная субстанция являются двумя другими важными
отделы головного мозга, расположенные в задней части ствола. Рубер
ядро играет роль в координации двигательных функций. Субстанция
nigra участвует в памяти о выученных задачах и гибели ее клеток
ассоциируется с болезнью Паркинсона.
Крыша среднего мозга называется тектумом . Тектум немлекопитающие позвоночные является местом для оптических долей, которые являются первичный центр восприятия зрения. У млекопитающих зрение воспринимается в головном мозге. Однако, в то время как головной мозг млекопитающих сообщает животному что такое объект, тектум сообщает млекопитающему, где в пространстве зрительный объект есть. В тектуме зрительные доли называются передними холмиками. Позади них находятся задние холмики, которые могут играть важную роль в координации. слуховых рефлексов. Вместе бугорки образуют четыре бугорка, называемые тела квадригемины.
Другие особенности заднего ствола мозга включают пирамидальные тракты , представляющие собой тракты двигательных волокон, идущие от мозгового коры в спинной мозг без перерыва. ножки головного мозга также важны, так как это места, где мозжечок соединяется с мозговой ствол.
Передний отдел ствола мозга: промежуточный мозг
Передний отдел ствола отличается от заднего тем, что не имеет
ядра для черепно-мозговых нервов и без ретикулярной формации, а также для связи
к более развитым функциям.
Дорсальной частью промежуточного мозга является эпиталамус, большая часть которого
по функциям ненервный. Включает в себя два выпячивания: теменной
орган и шишковидное тело , которые функционируют как эндокринная железа
и орган чувств.
Боковые отделы промежуточного мозга называются таламусом. таламус является ретрансляционным центром головного мозга и также функционирует в области осознания.
как в восприятии боли и удовольствия.
Вентральная часть промежуточного мозга — это гипоталамус . Это
контролирует большинство вегетативных функций организма, в том числе водный баланс,
регуляция температуры, аппетит и пищеварение, кровяное давление, сон и
бодрствование, сексуальное поведение и эмоции. На вентральной поверхности гипоталамуса
это перекрест зрительных нервов , где зрительные нервы сходятся и пересекаются.
Гипофиз (гипофиз) также лежит на вентральной стороне,
и функционирует как эндокринная железа
Мозжечок и мост
Мозжечок развивается из дорсальной части среднего мозга
в поперечном сечении, мозжечок состоит в основном из белого вещества.
 на коре и ветвящееся белое вещество в интерьере, придающее древовидное
внешний вид называется arbor vitae
на коре и ветвящееся белое вещество в интерьере, придающее древовидное
внешний вид называется arbor vitae функции для управления координацией движений и поддержания равновесия Головной мозг
Головной мозг является самой крупной частью головного мозга и развивается из телэнцефалон.
Каждая половина головного мозга называется полушарием головного мозга обонятельная луковица находится на переднем конце каждого полушария
полосатое тело — группа базальных ядер в основании головного мозга, через который проходят белые волокна и которые участвуют в движении мышечной массы и некоторое визуальное восприятие
кора — образует крышу и боковые стенки большого мозга; гиппокамп, играющий важную роль в пространственной памяти, может быть поврежден при лица с болезнью Альцгеймера. Определения: Астроциты – звездчатые питательные и поддерживающие глиальные клетки центральная нервная система Генераторы центральных паттернов – группы нейронов в спинной и головной мозг, деятельность которых отвечает за врожденные циклические движения частей тела
Ганглий — группа тел нейронов, расположенных периферически по отношению к центральному нервная система у позвоночных
Нейроглия – клетки центральной нервной системы, помогающие поддерживать, защищать и поддерживать нейроны
Микроглия — мелкие нейроглиальные клетки мезодермального происхождения, некоторые из которых фагоцитируют
Узел Ранвье — участки аксона, лежащие между шванновскими клетками, где плазматическая мембрана миелинизированного аксона близка к внеклеточной жидкость
Олигодендроциты — нейроглиальные клетки эктодермального происхождения, миелинизирующиеся аксонов в центральной нервной системе и образует белое вещество центральная нервная система (немиелиновые аксоны представляют собой серое вещество)
Сплетения — сети нервов или кровеносных сосудов, образованные до образования нервов распределяется по мышцам
Шванновские клетки, также называемые нейрилеммой. Клетки происхождения нервного гребня
которые образуют тонкую оболочку, окружающую немиелинизированный аксон, или, после
миелинизировав аксон, лежит на поверхности миелиновой оболочки
Клетки происхождения нервного гребня
которые образуют тонкую оболочку, окружающую немиелинизированный аксон, или, после
миелинизировав аксон, лежит на поверхности миелиновой оболочки
Телодендрии — конечные ветви аксона
Синаптическая передача в центральной нервной системе (раздел 1, глава 6) Неврология онлайн: электронный учебник по неврологии | Кафедра нейробиологии и анатомии
6.1 Синаптическая передача в простой рефлекторной цепи
Одним из простейших видов поведения, опосредуемых центральной нервной системой, является коленный рефлекс или рефлекс растяжения. В ответ на удар невролога молоточком по сухожилию надколенника возникает рефлекторное разгибание ноги. На рис. 6.1 показана нейросхема, контролирующая этот рефлекторный ответ. Растяжка сухожилия надколенника растягивает мышцу-разгибатель. В частности, он растягивает группу специфических рецепторов, известных как рецепторы мышечного веретена или просто рецепторы растяжения.
Рисунок 6. |
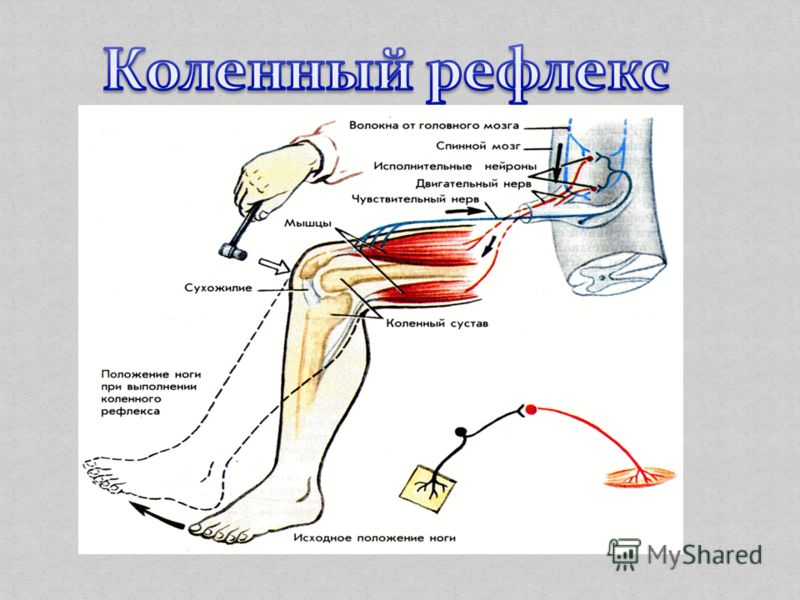 1
1Растяжение вызывает потенциалы действия в рецепторах растяжения, которые затем распространяются по афферентным волокнам типа 1А, соматы которых расположены в ганглии задних корешков. Затем отростки этих сенсорных нейронов входят в спинной мозг и образуют синаптические связи с двумя типами клеток. Сначала образуется синаптическая связь с разгибательным мотонейроном, расположенным в передних рогах спинного мозга. В результате синаптической активации этого мотонейрона потенциалы действия возникают в мотонейроне и распространяются от вентральных корешков, в конечном счете вторгаясь в конечные области моторного аксона (т.0468 нервно-мышечное соединение ), вызывая высвобождение ацетилхолина, деполяризацию мышечной клетки, формирование потенциала действия в мышечной клетке и последующее сокращение мышцы.
Сенсорные нейроны также образуют синаптические связи с другим типом нейронов в спинном мозге, называемым интернейроном.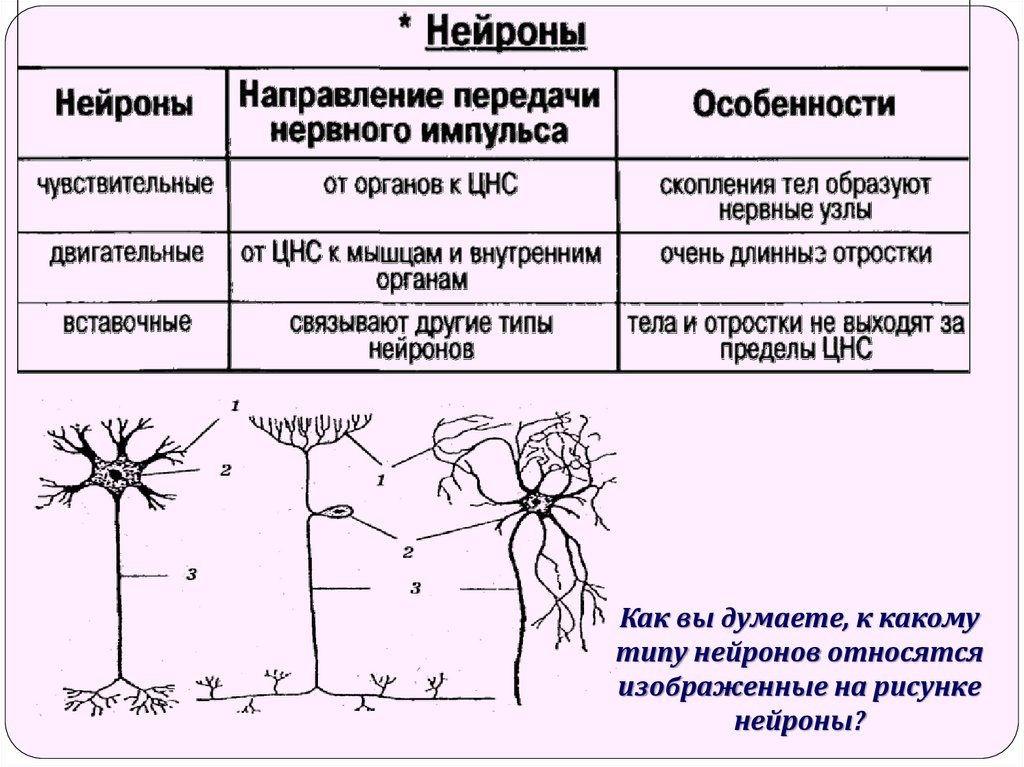 Интернейроны названы так потому, что они располагаются между одним типом нейрона и другим. Конкретный показанный интернейрон является тормозным интернейроном. В результате его активации в процессе синаптической передачи в интернейроне возникают потенциалы действия. Потенциал действия в тормозном нейроне приводит к высвобождению химического вещества-трансмиттера, которое ингибирует двигательный нейрон-сгибатель, тем самым предотвращая неправильное движение. Этот конкретный рефлекс известен как моносинаптический рефлекс растяжения, потому что этот рефлекс опосредуется одним возбуждающим синаптическим реле в центральной нервной системе.
Интернейроны названы так потому, что они располагаются между одним типом нейрона и другим. Конкретный показанный интернейрон является тормозным интернейроном. В результате его активации в процессе синаптической передачи в интернейроне возникают потенциалы действия. Потенциал действия в тормозном нейроне приводит к высвобождению химического вещества-трансмиттера, которое ингибирует двигательный нейрон-сгибатель, тем самым предотвращая неправильное движение. Этот конкретный рефлекс известен как моносинаптический рефлекс растяжения, потому что этот рефлекс опосредуется одним возбуждающим синаптическим реле в центральной нервной системе.
6.2. Ионные механизмы ВПСП
. Синаптические потенциалы
Обычно сенсорный нейрон активируется растяжением рецептора растяжения, но этот процесс можно обойти, вводя деполяризующий ток в сенсорный нейрон. Этот стимул инициирует потенциал действия в сенсорном нейроне, что приводит к изменению потенциала двигательного нейрона. Этот потенциал известен как возбуждающий постсинаптический потенциал (ВПСП) ; возбуждающим, потому что он имеет тенденцию деполяризовать клетку, тем самым увеличивая вероятность возбуждения потенциала действия в двигательном нейроне, и постсинаптическим, поскольку этот потенциал регистрируется на постсинаптической стороне синапса.
Ионные механизмы ВПСП в спинальном мотонейроне практически идентичны ионным механизмам ВПСП в нервно-мышечном синапсе. В частности, медиаторное вещество диффундирует через синаптическую щель и связывается со специфическими ионотропных рецепторов на постсинаптической мембране, что приводит к одновременному увеличению проницаемости для натрия и калия (см. рис. 4.10). Механизмы высвобождения также идентичны механизмам нервно-мышечного соединения. Потенциал действия в пресинаптических окончаниях приводит к открытию потенциалзависимых каналов Ca 2+ , а приток Ca 2+ вызывает высвобождение медиатора.
6.3 Различия между ВПСП в скелетном нервно-мышечном соединении и ВПСП в ЦНС
Существуют два принципиальных различия между процессом синаптической передачи в сенсомоторном синапсе в спинном мозге и процессом синаптической передачи в нервно-мышечном соединении. Первое , медиаторное вещество, высвобождаемое сенсорным нейроном, представляет собой не АХ, а глутамат аминокислоты. Действительно, в центральной нервной системе много разных передатчиков — до 50 и более, и с каждым годом этот список растет. К счастью, эти 50 или более веществ-трансмиттеров можно удобно сгруппировать в четыре основные категории: ацетилхолин , моноамины , пептиды и аминокислоты . Секунда , в отличие от амплитуды синаптического потенциала 50 мВ в нервно-мышечном соединении, амплитуда синаптического потенциала в спинальном мотонейроне в результате потенциала действия в афферентном волокне 1А составляет всего около 1 мВ.
Действительно, в центральной нервной системе много разных передатчиков — до 50 и более, и с каждым годом этот список растет. К счастью, эти 50 или более веществ-трансмиттеров можно удобно сгруппировать в четыре основные категории: ацетилхолин , моноамины , пептиды и аминокислоты . Секунда , в отличие от амплитуды синаптического потенциала 50 мВ в нервно-мышечном соединении, амплитуда синаптического потенциала в спинальном мотонейроне в результате потенциала действия в афферентном волокне 1А составляет всего около 1 мВ.
6.4 Временное и пространственное суммирование
Если амплитуда постсинаптического потенциала составляет всего 1 мВ, как может быть запущен потенциал действия в двигательном нейроне и рефлекторная функция? Обратите внимание, что ВПСП в 1 мВ вряд ли будет достаточно, чтобы довести мембранный потенциал двигательного нейрона до порогового значения, необходимого для срабатывания спайка.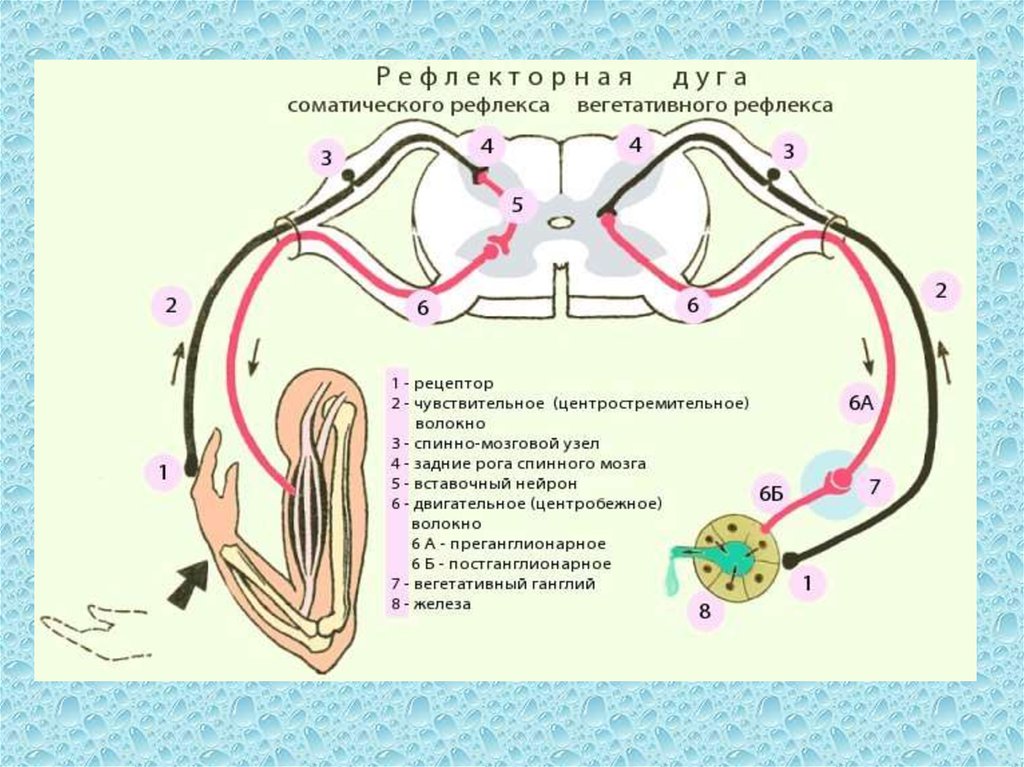 Если нет спайка, не будет и сокращения мышцы. Ответ заключается в том, что растяжение мышц запускает множественные потенциалы действия во многих различных рецепторах растяжения. На самом деле, чем больше растяжение, тем выше вероятность активации большего количества рецепторов растяжения. Этот процесс называется набор . Следовательно, множественные афференты 1А сойдутся на спинномозговом мотонейроне и будут участвовать в его активации. Однако это не весь ответ. Напомним, что чем больше интенсивность раздражителя, тем больше число потенциалов действия, вызываемых сенсорным рецептором. Чем больше растяжение, тем большее количество потенциалов действия вызывается в одном сенсорном нейроне и тем большее количество ВПСП продуцируется в двигательном нейроне из этой последовательности потенциалов действия в сенсорной клетке. Процессы, посредством которых множественные ВПСП от пресинаптических нейронов суммируются в пространстве и времени, называются временной и пространственной суммацией.
Если нет спайка, не будет и сокращения мышцы. Ответ заключается в том, что растяжение мышц запускает множественные потенциалы действия во многих различных рецепторах растяжения. На самом деле, чем больше растяжение, тем выше вероятность активации большего количества рецепторов растяжения. Этот процесс называется набор . Следовательно, множественные афференты 1А сойдутся на спинномозговом мотонейроне и будут участвовать в его активации. Однако это не весь ответ. Напомним, что чем больше интенсивность раздражителя, тем больше число потенциалов действия, вызываемых сенсорным рецептором. Чем больше растяжение, тем большее количество потенциалов действия вызывается в одном сенсорном нейроне и тем большее количество ВПСП продуцируется в двигательном нейроне из этой последовательности потенциалов действия в сенсорной клетке. Процессы, посредством которых множественные ВПСП от пресинаптических нейронов суммируются в пространстве и времени, называются временной и пространственной суммацией.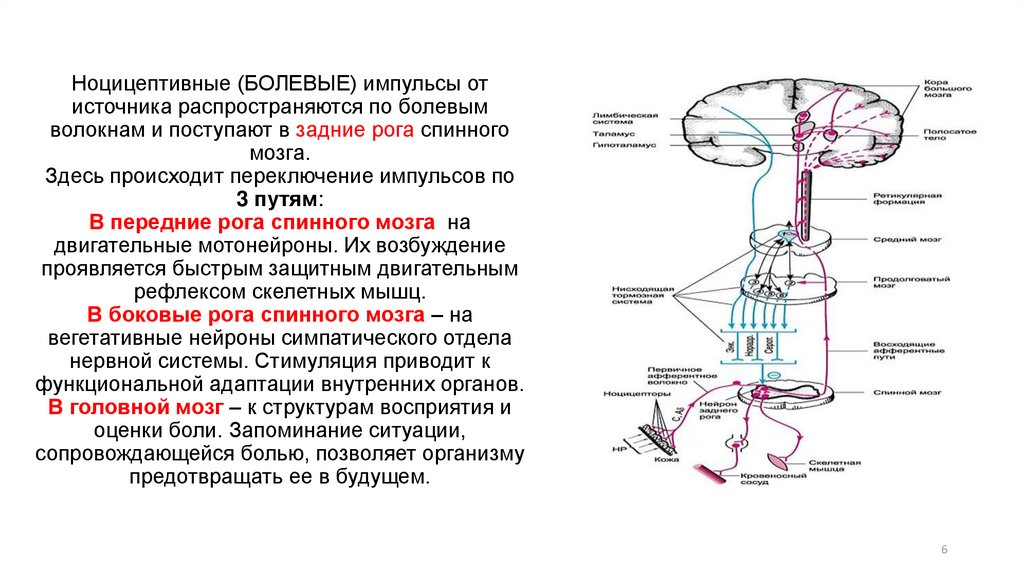
Рисунок 6.3 | |
Временное суммирование. Одиночный потенциал действия в сенсорном нейроне 1 вызывает ВПСП 1 мВ в моторном нейроне. Теперь рассмотрим последствия запуска двух потенциалов действия в быстрой последовательности (см. рисунок выше). Вызываются два ПКП, второе из которых суммируется по заднему фронту первого. В результате двух потенциалов действия возникает суммарный потенциал амплитудой около 2 мВ. Если бы пресинаптических потенциалов действия было три и они возникали достаточно быстро, то общий потенциал был бы около 3 мВ и т. д. Временная суммация является строго пассивным свойством нервных клеток. Для его объяснения не нужны специальные механизмы ионной проводимости. Потенциалы суммируются из-за пассивных свойств мембран нервных клеток, в частности способности мембран накапливать заряд. Мембрана временно хранит заряд первой ПСП, а затем к ней добавляется заряд второй ПСП, чтобы получить потенциал в два раза больше, чем сначала.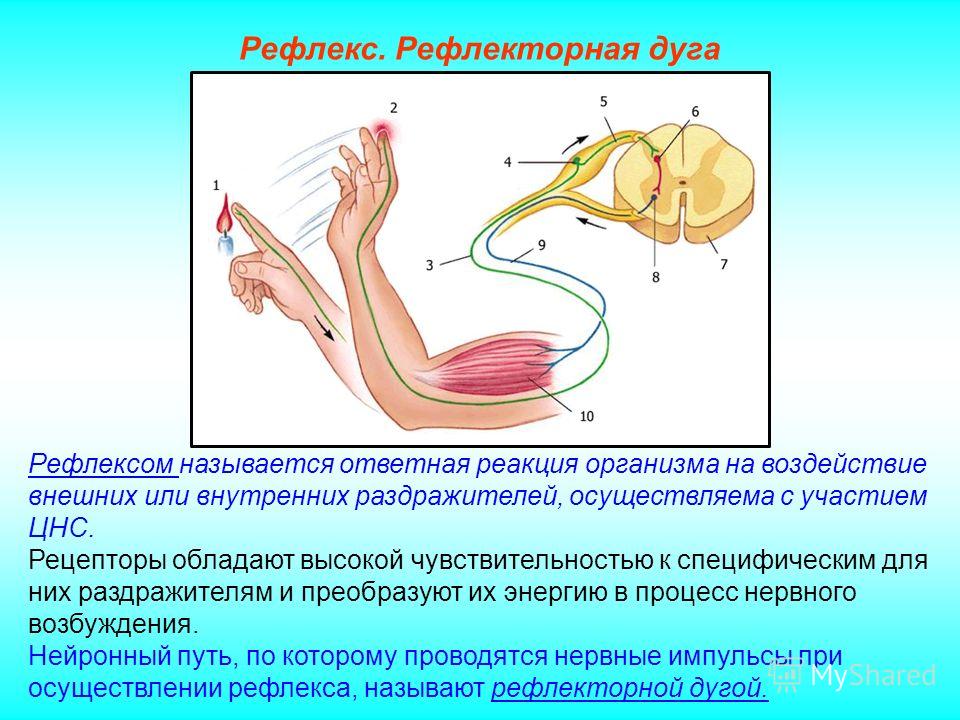 Этот процесс временной суммации очень сильно зависит от продолжительности синаптического потенциала. Временное суммирование происходит, когда пресинаптические потенциалы действия возникают в быстрой последовательности. Временные рамки зависят от пассивных свойств мембраны, особенно от постоянной времени.
Этот процесс временной суммации очень сильно зависит от продолжительности синаптического потенциала. Временное суммирование происходит, когда пресинаптические потенциалы действия возникают в быстрой последовательности. Временные рамки зависят от пассивных свойств мембраны, особенно от постоянной времени.
Пространственное суммирование. Теперь рассмотрим двигательный нейрон, который получает два входа. Потенциал действия, генерируемый в сенсорном нейроне 1, вызывает ВПСП с напряжением 1 мВ, а единичный потенциал действия в сенсорном нейроне 2 также вызывает ВПСП с напряжением 1 мВ. Если потенциалы действия генерируются одновременно в сенсорном нейроне 1 и в сенсорном нейроне 2, ВПСП суммируются, образуя суммарный ВПСП, который в два раза больше, чем отдельные ВПСП. Пространственная суммация в нервных клетках происходит из-за пространственной константы, способности заряда, произведенного в одной области клетки, распространяться на другие области клетки.
6.5 ТПСП
Активация нейрона в ответ на синаптический вход зависит от того, сколько потенциалов действия активируется на любом одном афферентном входе, а также от того, сколько отдельных афферентных путей активировано.
Решение о срабатывании также зависит от наличия тормозных синаптических входов. Искусственная деполяризация интернейрона для инициации потенциала действия вызывает временную гиперполяризацию мембранного потенциала двигательного нейрона (см. рис. 6.2). Временной ход этой гиперполяризации очень похож на ВПСП, но имеет обратный знак. Синаптический потенциал мотонейрона называется 9.0468 тормозной постсинаптический потенциал (ТПСП) , потому что он имеет тенденцию отодвигать мембранный потенциал от порога, тем самым уменьшая вероятность того, что этот нейрон инициирует потенциал действия.
6.6 Ионные механизмы для ТПСП
Мембранный потенциал сгибательного мотонейрона составляет около -65 мВ, поэтому можно предположить, что ТПСП будет вызван увеличением проницаемости или проводимости иона, равновесный потенциал которого больше отрицательный, чем -65 мВ. Одним из возможных вариантов является калий. Калий опосредует некоторые тормозные синаптические потенциалы в центральной нервной системе, но не в конкретном синапсе между спинальным интернейроном и спинальным мотонейроном. В этом конкретном синапсе IPSP происходит из-за избирательного увеличения проницаемости для хлоридов. Обратите внимание, что равновесный потенциал для хлорида составляет около -70 мВ. Медиатор, высвобождаемый спинальным интернейроном, связывается с особым классом ионотропных рецепторов, которые в норме закрыты, но открыты и становятся избирательно проницаемыми для ионов хлора в результате связывания медиатора. В результате увеличения Cl — проницаемости, мембранный потенциал перемещается от значения покоя -65 мВ к равновесному потенциалу Cl — . (Обратите внимание, что, в принципе, снижение проводимости Na + в состоянии покоя может также вызывать ТПСП.)
В этом конкретном синапсе IPSP происходит из-за избирательного увеличения проницаемости для хлоридов. Обратите внимание, что равновесный потенциал для хлорида составляет около -70 мВ. Медиатор, высвобождаемый спинальным интернейроном, связывается с особым классом ионотропных рецепторов, которые в норме закрыты, но открыты и становятся избирательно проницаемыми для ионов хлора в результате связывания медиатора. В результате увеличения Cl — проницаемости, мембранный потенциал перемещается от значения покоя -65 мВ к равновесному потенциалу Cl — . (Обратите внимание, что, в принципе, снижение проводимости Na + в состоянии покоя может также вызывать ТПСП.)
6.7 Передатчик вещества спинного тормозного нейрона
Что насчет вещества-передатчика, которое высвобождается тормозным интернейроном в спинном мозге? ? Передатчиком является глицин , аминокислота, которая часто используется в центральной нервной системе в качестве медиатора, оказывающего тормозящее действие. Однако он не самый распространенный. Наиболее распространенным передатчиком с тормозным действием является гамма-аминомасляная кислота (ГАМК) .
Однако он не самый распространенный. Наиболее распространенным передатчиком с тормозным действием является гамма-аминомасляная кислота (ГАМК) .
6.8 Метаботропные синаптические ответы
Помимо ответов, опосредованных ионотропными рецепторами, существует совершенно отдельный класс синаптических потенциалов, длительность которых на порядки превышает длительность классических ВПСП. Это так называемые медленные синаптические потенциалы, и они опосредованы метаботропными рецепторами. Медленные синаптические потенциалы наблюдаются не у каждого постсинаптического нейрона, но, безусловно, у многих. На рисунке ниже показан постсинаптический нейрон, который получает два входа. Потенциал действия в нейроне 1 вызывает возбуждающий постсинаптический потенциал или ВПСП в постсинаптической клетке, продолжительность которого составляет около 20 мсек. Нейрон 2 также может производить постсинаптический потенциал, но его продолжительность более чем на три порядка больше, чем у обычного типа синаптического потенциала.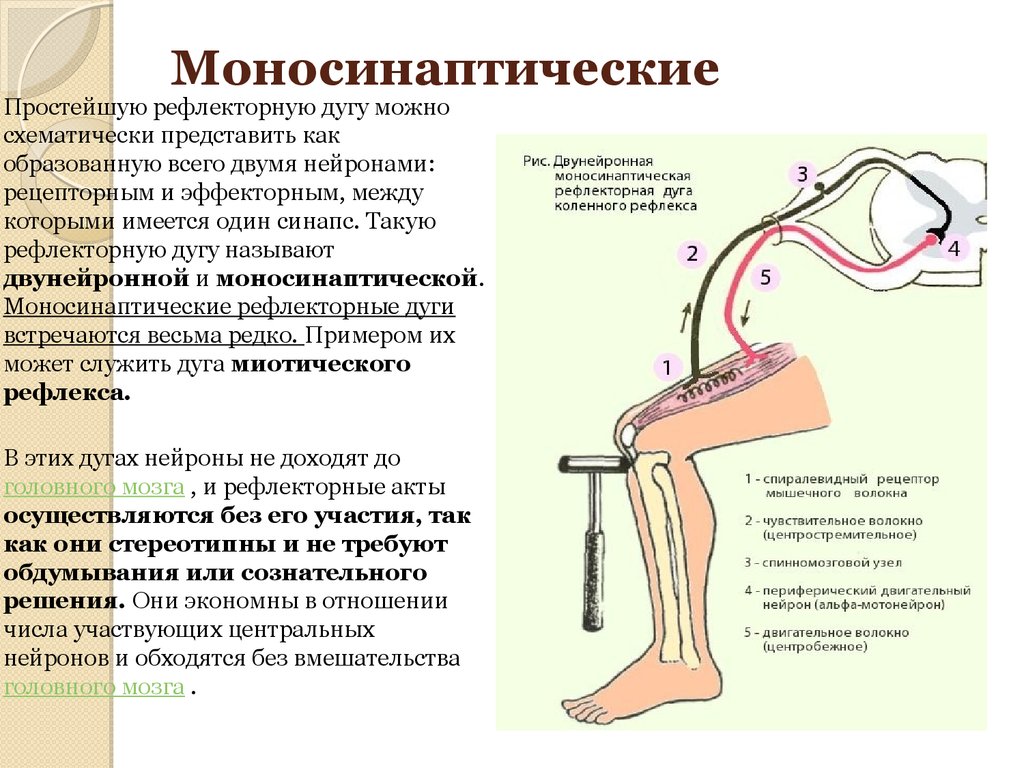 Механизм этих медленных синаптических ответов включает изменения в метаболизме клетки.
Механизм этих медленных синаптических ответов включает изменения в метаболизме клетки.
Рисунок 6.4 |
Рисунок 6.5 |
Один из механизмов медленного синаптического потенциала показан на иллюстрации слева (рис. 6.5) и на рис. 11.11. В отличие от ионотропных рецепторов, для которых рецепторы фактически являются частью комплекса каналов, каналы, продуцирующие медленные синаптические потенциалы, не связаны напрямую с рецепторами-медиаторами. Скорее, рецепторы отделены от канала. Эти рецепторы известны как метаботропные, потому что они связаны с изменениями метаболизма клетки и вообще с изменениями в активации специфических систем вторичных мессенджеров. Рисунок слева иллюстрирует пример одного типа ответа, который включает циклический каскад AMP. Медленные PSP в некоторых случаях опосредуются циклическим АМФ, но они также опосредуются другими протеинкиназами.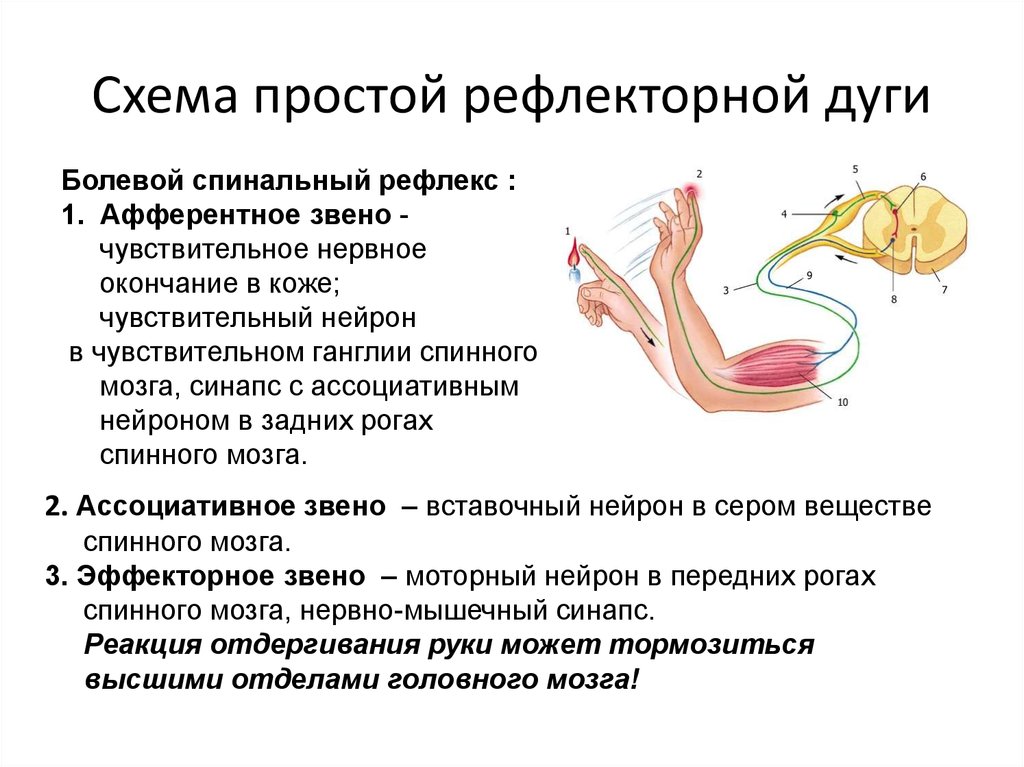 Для реакции, показанной на рис. 6.5, передатчик активирует G-белки, что приводит к усилению синтеза циклического АМФ. Затем циклический АМФ приводит к активации циклической АМФ-зависимой киназы (PKA), которая фосфорилирует белок канала или компонент канала, а затем вызывает конформационные изменения в канале и изменение его ионной проницаемости. В отличие от прямого конформационного изменения, вызываемого связыванием передатчика с комплексом рецепторных каналов (наблюдаемого в реакциях, опосредованных ионотропными рецепторами), конформационное изменение вызывается фосфорилированием. Конкретный канал избирательно проницаем для K + и нормально разомкнут. В результате фосфорилирования канала PKA канал закрывается и становится менее проницаемым для K + . Поскольку нормальный потенциал покоя обусловлен балансом Na + и K + , снижение проводимости K + способствует влиянию проводимости Na + и вызывает деполяризацию.
Для реакции, показанной на рис. 6.5, передатчик активирует G-белки, что приводит к усилению синтеза циклического АМФ. Затем циклический АМФ приводит к активации циклической АМФ-зависимой киназы (PKA), которая фосфорилирует белок канала или компонент канала, а затем вызывает конформационные изменения в канале и изменение его ионной проницаемости. В отличие от прямого конформационного изменения, вызываемого связыванием передатчика с комплексом рецепторных каналов (наблюдаемого в реакциях, опосредованных ионотропными рецепторами), конформационное изменение вызывается фосфорилированием. Конкретный канал избирательно проницаем для K + и нормально разомкнут. В результате фосфорилирования канала PKA канал закрывается и становится менее проницаемым для K + . Поскольку нормальный потенциал покоя обусловлен балансом Na + и K + , снижение проводимости K + способствует влиянию проводимости Na + и вызывает деполяризацию.
Интересно отметить, что активация метаботропных рецепторов может вызывать эффекты, длящиеся гораздо дольше нескольких сотен секунд.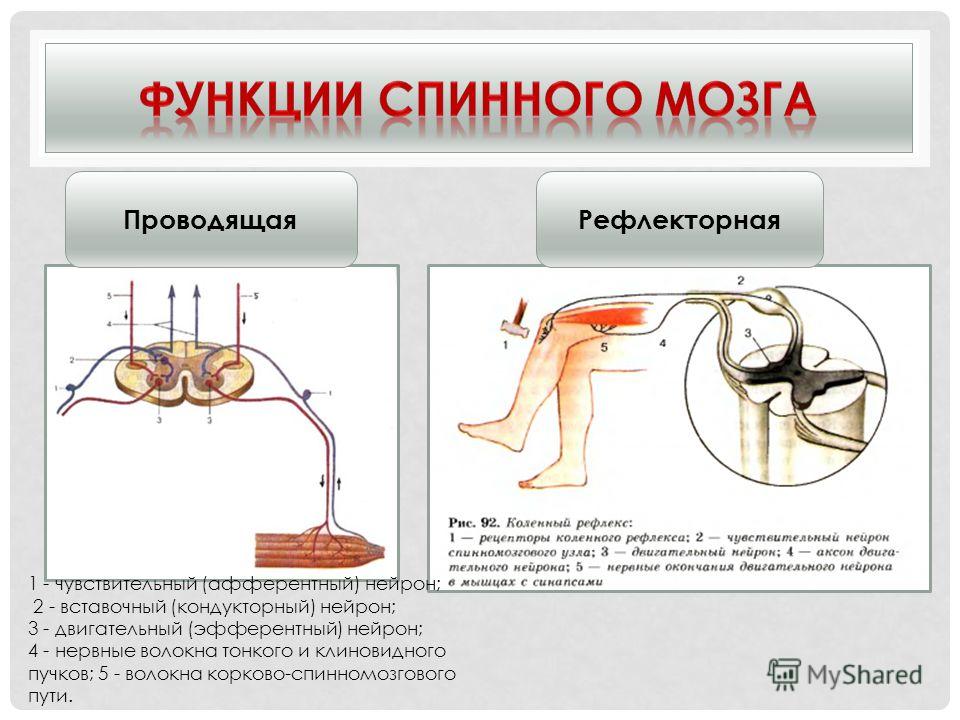 Например, протеинкиназа А может диффундировать в ядро, где она может фосфорилировать белки (то есть факторы транскрипции), которые регулируют экспрессию генов.
Например, протеинкиназа А может диффундировать в ядро, где она может фосфорилировать белки (то есть факторы транскрипции), которые регулируют экспрессию генов.
6.9 Типы синаптической передачи
Эта и две предыдущие главы посвящены химической синаптической передаче. Как вы видели в химических синапсах, существует четко выраженный цитоплазматический разрыв, разделяющий пресинаптические и постсинаптические мембраны (рис. 6.6А).
Рисунок 6.6A |
Рисунок 6.6B |
Этот разрыв известен как синаптическая щель. Пресинаптическое окончание химических синапсов содержит высокую концентрацию митохондрий и синаптических пузырьков, имеется характерное утолщение постсинаптической мембраны. В результате деполяризации или потенциала действия в пресинаптических окончаниях из пресинаптических окончаний высвобождаются химические медиаторы, которые диффундируют через синаптическую щель и связываются с рецепторами на постсинаптической мембране. Это приводит к изменению проницаемости, которое создает постсинаптический потенциал. Для химических синапсов существует задержка (обычно длительностью примерно 0,5—1 мс) между инициацией потенциала действия в пресинаптической терминали и изменением потенциала в постсинаптической клетке. Синаптическая задержка связана со временем, необходимым для высвобождения медиатора, диффузии через щель и связывания с рецепторами на постсинаптической мембране. Химическая синаптическая передача, как правило, однонаправленная. Изменение потенциала в пресинаптической клетке высвобождает медиатор, который создает постсинаптический потенциал, но деполяризация в постсинаптической клетке не вызывает никаких эффектов в пресинаптической клетке, потому что из постсинаптической клетки в синаптической области медиатор не высвобождается.
В результате деполяризации или потенциала действия в пресинаптических окончаниях из пресинаптических окончаний высвобождаются химические медиаторы, которые диффундируют через синаптическую щель и связываются с рецепторами на постсинаптической мембране. Это приводит к изменению проницаемости, которое создает постсинаптический потенциал. Для химических синапсов существует задержка (обычно длительностью примерно 0,5—1 мс) между инициацией потенциала действия в пресинаптической терминали и изменением потенциала в постсинаптической клетке. Синаптическая задержка связана со временем, необходимым для высвобождения медиатора, диффузии через щель и связывания с рецепторами на постсинаптической мембране. Химическая синаптическая передача, как правило, однонаправленная. Изменение потенциала в пресинаптической клетке высвобождает медиатор, который создает постсинаптический потенциал, но деполяризация в постсинаптической клетке не вызывает никаких эффектов в пресинаптической клетке, потому что из постсинаптической клетки в синаптической области медиатор не высвобождается. Наиболее преобладающим типом синапсов являются химические синапсы, и по этой причине они были в центре внимания этой и предыдущих глав.
Наиболее преобладающим типом синапсов являются химические синапсы, и по этой причине они были в центре внимания этой и предыдущих глав.
Однако другая категория синапсов связана с электрической синаптической передачей. Электрическая синаптическая передача обеспечивается специализированными структурами, известными как щелевые соединения (рис. 6.6В), которые обеспечивают путь цитоплазматической непрерывности между пресинаптическими и постсинаптическими клетками. Следовательно, деполяризация (или гиперполяризация), вызываемая в пресинаптическом терминале, вызывает изменение потенциала постсинаптического терминала, что обусловлено прямым ионным путем между клетками. Для электрических синапсов присутствует минимальная синаптическая задержка; как только в пресинаптических окончаниях возникает потенциальное изменение, в постсинаптической клетке возникает отражение этого потенциального изменения. Электрические контакты находятся как в нервной системе, так и между другими возбудимыми мембранами, такими как гладкомышечные клетки и клетки сердечной мышцы.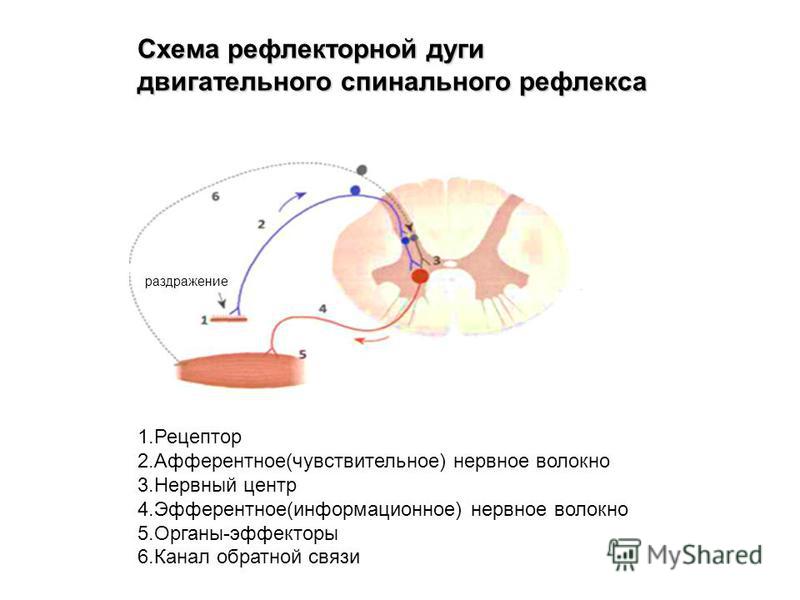 В этих мышечных клетках они обеспечивают важный путь для распространения потенциалов действия от одной мышечной клетки к другой.
В этих мышечных клетках они обеспечивают важный путь для распространения потенциалов действия от одной мышечной клетки к другой.
6.10 Нейротоксины
Открытие некоторых токсинов значительно облегчило анализ потенциалов и химически управляемых каналов, а также процесс синаптической передачи. В следующей таблице показаны некоторые из них, которые оказались особенно полезными.

Leave A Comment