
Пластиды
Пластиды — органоиды, присущие только растительным клеткам. Обычно это крупные тельца, хорошо видимые под световым микроскопом.[ …]
Пластиды, характеризующиеся сложной мембранной системой, содержат крахмал. Оболочка яйцеклетки пронизана плазмодес-мами; в нижней части яйцеклетки она представлена плазмалем-мой, которая непосредственно контактирует с плазмалеммой синергид и центральной клетки. Ядро яйцеклетки также покрыто оболочкой, имеющей поры.[ …]
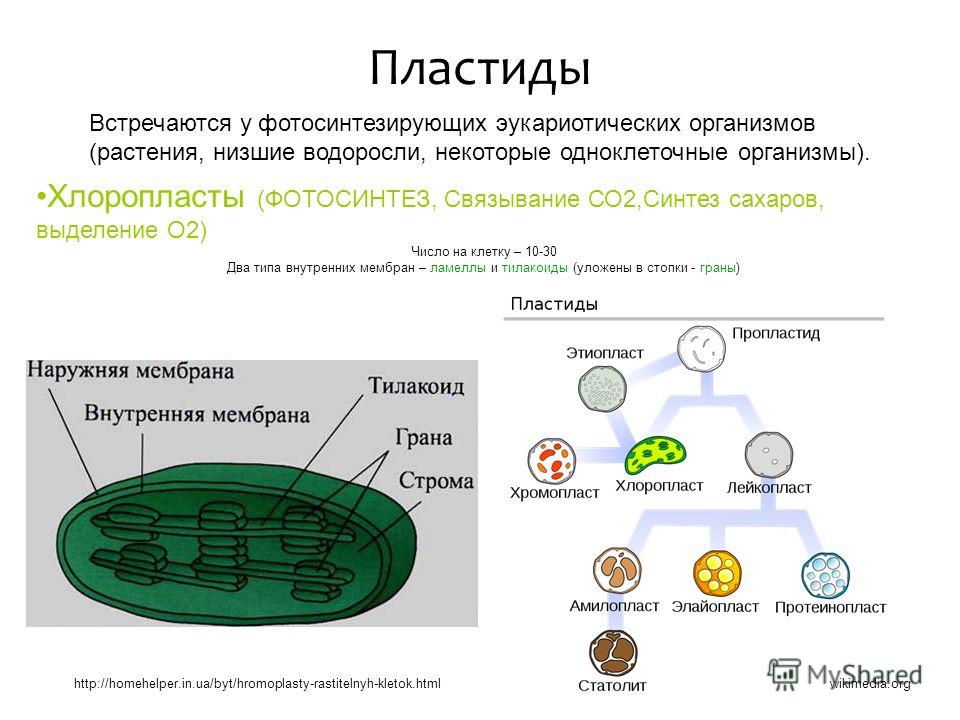
Пластиды — специализированные дискообразные тела, содержащиеся в цитоплазме и свойственные только растительным клеткам. По наличию или отсутствию пигмента пластиды делятся на лейкопласты (бесцветные) и хромопласты (окрашенные). Лейкопласты присутствуют во взрослых клетках, не подвергающихся воздействию света, а некоторые типы включены в запасной крахмал. Из окрашенных наибольшее значение имеют те пластиды, которые содержат хлорофилл (хлоропласты), так как они являются структурно и функционально полными элементами фотосинтеза, процесса, в котором углекислый газ и вода превращаются в углеродсодержащие соединения. Эти реакции приводят к образованию крахмальных зерен в хлоропла-стах.[ …]
Пластиды (от греч. р1а 1оБ — вылепленный и е1с1о5— подобный). Так называют постоянные мембранные органеллы растительных клеток, в которых осуществляется первичный синтез органических веществ. Грибы, бактерии, слизевики, а также сине-зеленые водоросли пластид не имеют.[ …]
Для пластид характерны переходы из одного типа в другой. Например, зеленые хлоропласты листьев переходят в хромопласты (осенью, когда изменяется окраска листьев), а лейкопласты зеленеющих клубней картофеля переходят в хлоропласты.[ …]
Тело пластид состоит из стромы, содержащей в основном протеины и липиды, а также пигменты и минеральные элементы, и ограничено двумя липопротеидиыми мембранами.[ …]
Форма пластид высших растений довольно однообразна: дискообразная (округло-эллиптическая). У водорослей хроматофо-ры бывают палочкообразными, лентовидными, спиралевидными, чашеобразными и т.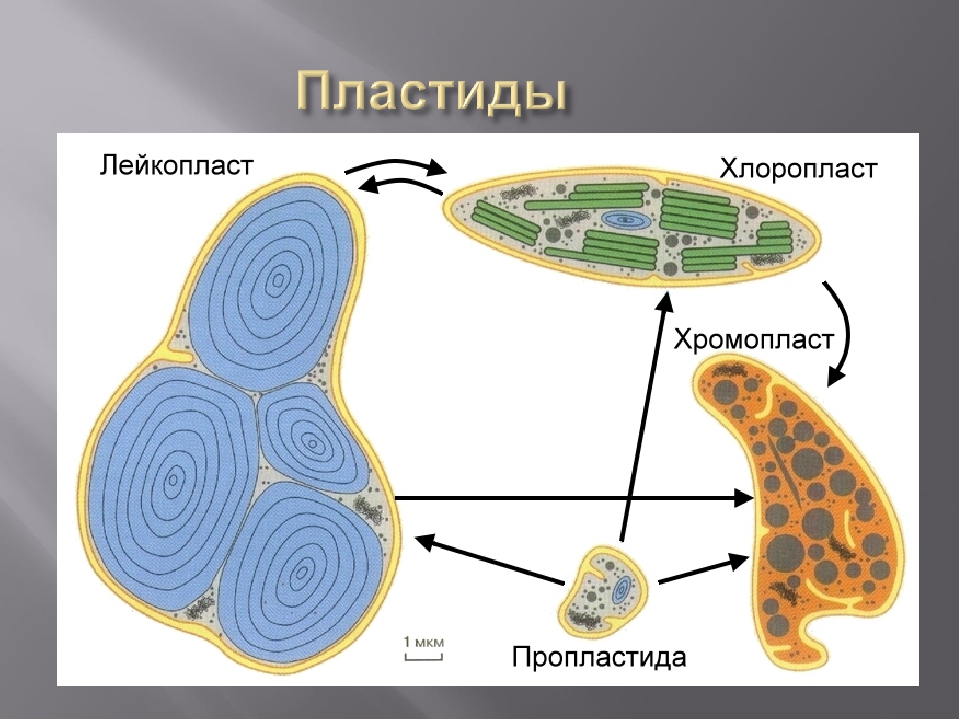 д. Величина пластид у покрытосеменных растений колеблется от 3 до 10 мкм, среди них самые мелкие — лейкопласты.[ …]
д. Величина пластид у покрытосеменных растений колеблется от 3 до 10 мкм, среди них самые мелкие — лейкопласты.[ …]
Зеленые пластиды чутко реагируют на освещение, и хлорофилл образуется в них только на свету. Молекулы хлорофилла поглощают свет. Энергия его преобразуется в процессе синтеза органического вещества в энергию химической связи органических (углеродистых) соединений.[ …]
Все типы пластид генетически связаны между собой, хотя функции их строго специфичны. Исходной формой пластид являются пропластиды, по внешнему виду сходные с митохондриями, но более крупных размеров, удлиненной формы, с неупорядоченным расположением крист. Митохондрии от пластид можно отличить по способности их прижизненно окрашиваться янусом зеленым Б, в то время как пропластиды этим веществом не окрашиваются.[ …]
Общее число пластид в клетках высших растений варьирует в довольно значительных пределах. Так, в клетках листьев высших растений их от 20 до 100, а у низших одноклеточных организмов — всего одна.[ …]
У разных типов пластид наблюдаются значительные колебания в составе химических компонентов. Содержание воды в них зависит от водного режима растений и колеблется от 60 до 75%.[ …]
Различают 3 типа пластид: бесцветные — лейкопласты, зеленые — х л о р о-пласты, окрашенные в другие цвета — хромопласты. Пластиды каждого типа имеют свое строение и несут свои, им присущие функции. Однако возможны переходы пластид из одного типа в другой. Так, позеленение клубней картофеля вызывается перестройкой их лейкопластов в хлоропласты. В корнеплоде моркови лейкопласты переходят в хромопласты. Пластиды всех трех типов образуются из пропластид.[ …]
Помимо хлорофилла, пластидам свойственны и другие пигменты, обладающие способностью поглощать лучистую энергию. Вообще все растения, способные к фотосинтезу, содержат, кроме хлорофилла, группу желтых пигментов — каротиноидов (это каротины, ксантофиллы том числе фуко ксантин бурых и некоторых других водорослей).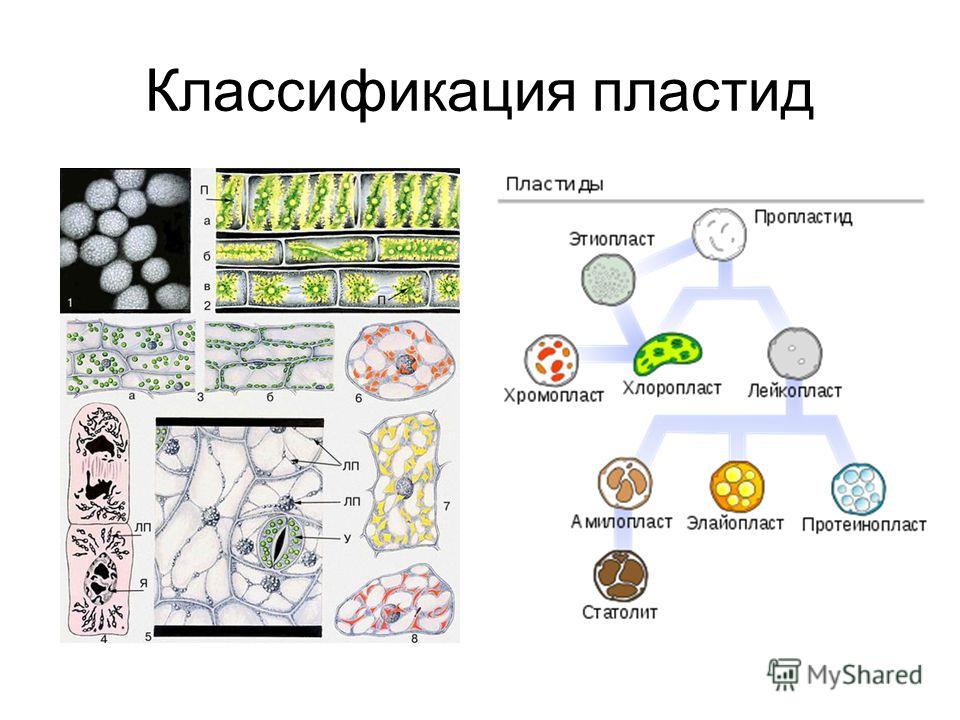
Хлоропласты (зеленые пластиды)—органеллы клетки, обусловливающие накопление углеводов в процессе фотосинтеза. В них содержится большое количество ферментов, контролирующих фотосинтез, а также белков, жирных кислот и фосфолипидов. Эти важнейшие мембранные органеллы встречаются в клетках эукариотических организмов: высших растений и некоторых одноклеточных.[ …]
Клетки имеют одну или две пластиды золотистого или желто-ко-р ичневого цвета, один или два жгутика и иногда одну стигму. Способ защиты от неблагоприятных условий — образование окременелых цист.[ …]
В живой растительной клетке в пластидах хлорофилл находится в виде комплекса его с белком и липоидами, и одновременно с обновлением белка происходит и обновление хлорофилла. Таким образом, молекулы хлорофилла, так же как и белковые молекулы, постоянно разрушаются и вновь синтезируются. Скорость обновления молекул белка и хлорофилла выражается величинами одного порядка.[ …]
Часть нуклеиновых кислот, связанная с более крупными фрагментами клетки, оставалась в осадке. Центрифугат доводился до определенного объема и разливался при постоянном взбалтывании равными порциями по колбочкам. Содержимое каждой колбочки соответствовало 1 г сырых листьев исходной пробы. В качестве антисептика в колбочки добавлялся 1 мл толуола.. В каждой серии в 4 колбочках ферментативные процессы в центрифугате останавливались нагреванием до 90—100°; эти пробы служили контролем. Автолиз в опытных колбах проводился в термостате при 25°; через определенные интервалы (60 и 190 часов) автолиз останавливался в оставшейся части проб также путем нагревания.[ …]
Для выяснения локализации фермента в пластидах было проведено разделение хлоропластов на фракции в градиенте плотности сахарозы. Холинэстеразная активность была обнаружена во фракциях наружных мембран и тилакоидов. При этом активность холинэстеразы в тилакоидах была примерно в 7 раз выше, чем в оболочке хлоропластов.[ …]
У цветных жгутиконосцев обнаруживаются пластиды, цветные вещества, митохондрии (Pringsheim, 1963, рис. 1), светочувствительный органоид — стигма.[ …]
1), светочувствительный органоид — стигма.[ …]
В сформировавшейся клетке растущих листьев пластиды размножаются путем деления. Деление может происходить или на стадии пропластид (у высших растепий). или на стадии уже сформировавшихся хлоропластов (у водорослей и папоротников). Пластиды делятся или путем возникновения поперечных перегородок, или, реже, путем почкования. Поперечные перегородки образуются за счет образования складок на внутренней мембране. До тех пор, пока складка внутренней мембраны же поделила тело пластид, наружная мембрана сохраняется и объединяет обе дочерние пластиды. Она либо разрывается, либо образует складки.[ …]
Различают три вида пластид: лейкопласты — бесцветные, хромопласты — оранжевые, хлоропласты — зеленые. Именно в хлоро-пластах сосредоточен зеленый пигмент хлорофилл.[ …]
Для растительной клетки характерно присутствие пластид. Важнейшие пластиды — это хлоропласты. Они осуществляют трансформацию световой энергии в химическую. Другой важнейший энергетический процесс (синтез АТФ за счет энергии окисления) происходит в митохондриях. Они представляют собой овальные или палочковидные структуры длиной 1…2 мкм. Система канальцев и цистерн (диктиосом), ограниченных однослойной мембраной, составляет аппарат Гольджи, основная функция которого — внутриклеточная секреция веществ, необходимых для построения клеточной оболочки и др. В округлых тельцах — лизосомах сконцентрированы гидролитические ферменты. С помощью сферосом идет синтез липидов.[ …]
Для количественной характеристики состояния ультраструктуры пластиды промеряли на фотографиях планиметром и полученные величины относили к 100 мкм2 зоны пластид, т. е. к тому участку протоплазмы клетки, который окружен двойной мембраной. Серийные срезы, гарантирующие типичность результатов, не делали, и в связи с этим, как это также подчеркивает Шнепф (Schnepf, 1963), представленные данные являются относительными. Тем не менее при сравнении с неподвергав-шимися газации объектами удалось выявить структурные изменения, связанные с воздействием загрязнителей воздуха.
Во время деления клетки происходит репликация различных органелл, в том числе пластид и митохондрий. Простейшим типом пластиды является пропластида, из которой развиваются все типы пластид, включая хлоропласта. Пластиды представляют собой полуавтономные оргаиеллы, способные к удвоению путем деления или почкования. В клетках высших растений может содержаться от нескольких до большого числа пластид, и клетки различных типов значительно отличаются друг от друга по содержанию в них пластид. Число пластид в клетках какого-либо одного типа обычно остается приблизительно постоянным, и это наводит на мысль, что репликация пластид происходит одновременно с делением клетки. Однако распределение пластид материнской клетки между дочерними происходит, по-видимому, случайным образом.[ …]
В. Chrysophyta. Этот тип включает три больших группы водорослей, никогда не образующих крахмал. Пластиды желто-зелено-го, желтого или коричневого цвета; запасным продуктом являются лейкозин или жир и масла. Эти водоросли образуют либо кремнистый скелет (диатомеи), либо оболочку, иногда покрытую кремнистыми чешуйками (Chrysophyceae), или образуют кремнистые цисты (форма защиты) (Chrysophyceae и Xanthophyceae).[ …]
Фотосинтетические реакции в клетках эукариот происходят в высокоспециализированных органеллах — пластидах или хлоро-пластах, которые состоят из мелких мембранных образований (гран), имеющих слоистую (ламеллярную) структуру. В ламеллах и вокруг них локализованы фотосинтетические пигменты. Мембраны состоят из субединиц или физиологических единиц фотосинтеза, включающих определенное количество молекул хлорофиллов, каротиноидов, а также белки, липиды, цитохромы и некоторые металлы. У синезеленых водорослей, которые относятся к прокариотам, фотосинтезирующие структуры организованы проще: они представляют собой сферические частицы хромато-форы, содержащие пигменты, фосфолипиды и весь набор ферментов, необходимых для фотосинтеза.[ …]
Одноклеточные планктические водоросли с многочисленными коричневыми пластидами; тело их часто покрыто оболочкой, состоящей из целлюлозных пластинок. [ …]
[ …]
У водорослей хлоропласты могут иметь самую разнообразную форму и размеры, а у высших растений они очень однородны по своей форме. Эти пластиды окружены двойной мембраной и состоят из бесцветного жидкого матрикса (стромы) с находящейся в нем сложной системой уплощенных мембранных пузырьков, известных как тилакоиды, которые объединяются в стопки (граны). Тилакоиды имеют округлые очертания и соединяются с тилакоидами в другой гране посредством связующих мембран (рис. 1.8). Мембраны тилакоидов .состоят из-липидов и белков, к которым присоединяется хлорофилл.[ …]
Не1егоз1рЬопа1е5 включает нитчатые, ветвящиеся и несептиро-ванные формы. Наиболее распространенной является Vau.ch.er ¡а, обладающая множеством мелких пластид (рис. 28.37).[ …]
Наряду с минеральным питанием основой жизнедеятельности растений является усвоение углекислоты, происходящее в определенных частях клетки — пластидах (лейкопласты, хлоропласта и хромопласты).[ …]
В других случаях окраска растений (со сходными оттенками) обусловлена другой большой группой пигментов. Это — каротинои-ды, содержащиеся преимущественно в пластидах и обуславливающие гамму расцветок желтого, оранжевого и красного цвета (вспомните цвет календулы — ноготков, а также расцветку осенних листьев). В сравнительно больших количествах каротиноиды содержатся в тканях и структурах, связанных с размножением. Это спорофиллы и спорангии плаунов, хвощей, папоротников, хвойных, цветковых растений (тычинки и пестики), у которых преобладает желтая, бурая и красная окраски. Эта группа пигментов распространена у представителей семейств лютиковых, лилейных, кувшинковых, крестоцветных, пасленовых, тыквенных, норичниковых, сложноцветных; они содержатся в листьях крапивы, люцерны, клевера, шпината, в кожице красного перца, в плодах абрикоса, рябины, облепихи, апельсинов, мандаринов, хурмы, банана, дыни, томатов, в цветках календулы, нарцисса, одуванчиков. В зеленых растениях каротиноиды часто замаскированы хлорофиллом и появляются только при его разрушении в период созревания плодов, осеннего расцвечивания листьев, стрессах.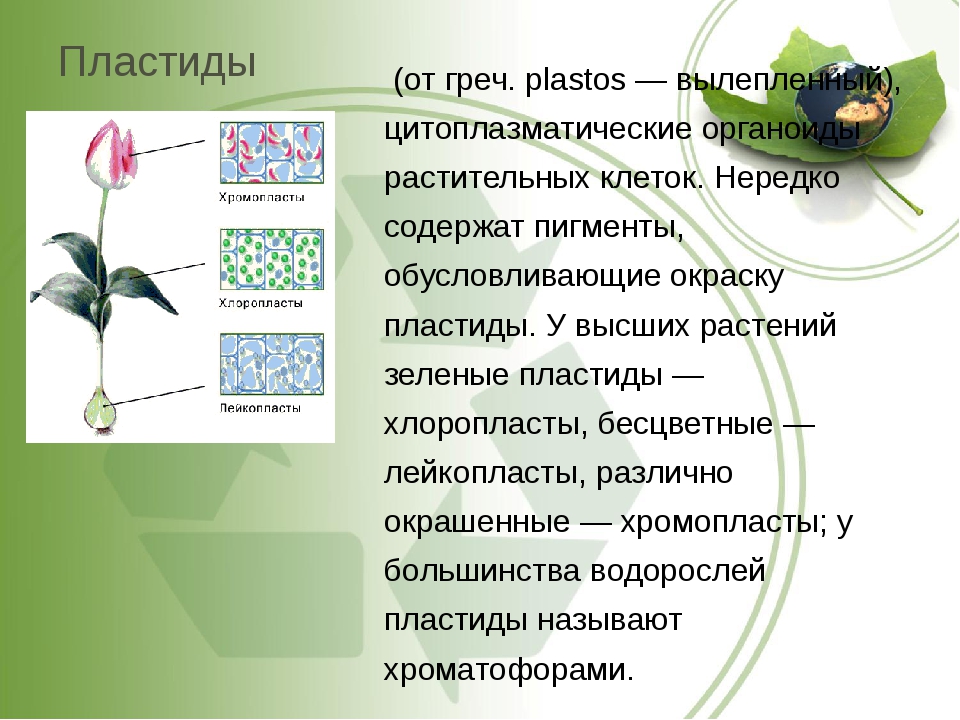 [ …]
[ …]
Все содержимое клетки, за исключением ядра и клеточной стенки, называется цитоплазмой. В жидкой, бесструктурной фазе цитоплазмы (матриксе) находятся рибосомы, мембранные системы, митохондрии, пластиды и другие структуры, а также запасные питательные вещества. Цитоплазма обладает чрезвычайно сложной, тонкой структурой (слоистая, гранулярная). С помощью электронного микроскопа раскрыты многие интересные детали строения клетки.[ …]
Как уже отмечалось, под влиянием экзогенного ацетилхолина наблюдается выход в среду ионов №+ и К+ из интактных хлоропластов. Внутри хлоропластов гороха концентрация этих ионов выше, чем в цитоплазме, но оболочка пластиды непроницаема для них. Открывание ионных каналов под действием ацетилхолина, по-видимому, способствует пассивному транспорту ряда ионов по градиенту электрохимического потенциала. Однако протон в этом случае по-разному поступает в отдельные компартменты органелл.[ …]
Делению плетки предшествует деление ядра. Каждое ядро дочерней клетки получает ровно такое же число хромосом и такое жеяо-личество ДНК, как и материнская. Во время митоза движение цитоплазмы прекращается, митохондрии в пластиды распределяются примерно поровну между дочершгмп клетками.[ …]
Для дальнейшего развития структуры пропластид необходим свет. Молекулы хлорофилла локализуются в мембранах. Короткие тилакоиды располагаются стопкой друг над другом и образуют тилакоиды гран. Пластиды достигают окончательного размера (рис. 30). Непосредственно из пропластид могут образовываться и бесцветные пластиды (лейкопласты—амилопласты) (рис. 31), Лейкопласты чаще всего локализованы в клетках запасающих ткапей. Подобно пропластидам они характеризуются слабо развитой ламеллярной структурой. Во многих случаях в лейкопластах ламеллы сохраняют связь с внутренней оболочкой. Хромопласты — это, по-ви-димому, результат деградации хлоропластов, образовавшиеся за счет частичного разрушения ламеллярной структуры. Эти глобулы располагаются сплошным слоем под оболочкой пластид.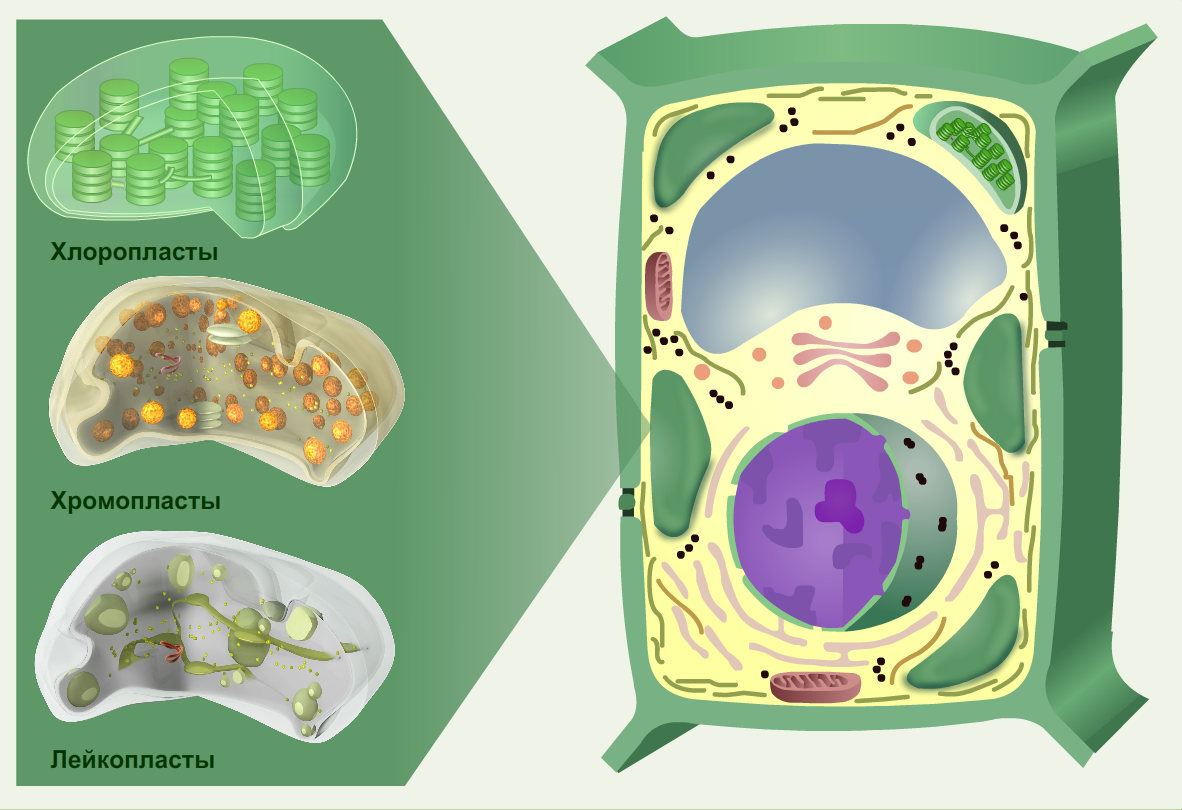 [ …]
[ …]
Вблизи яйцевого аппарата видна центральная клетка зародышевого мешка, включающая два полярных ядра. Электронномикроскопические исследования позволили выявить интересные особенности ее структуры. Цитоплазма центральной клетки ограничена плазмалеммой и содержит много митохондрий, пластид, развитый аппарат Гольджи и многочисленные каналы эндоплазматической сети в виде длинных тяжей, окутывающих полярные ядра. Рибосомы в изобилии располагаются свободно или прикреплены к мембранам эндоплазматической сети. Пластиды имеют хорошо выраженную ламеллярную структуру и содержат крупные крахмальные зерна. Митохондрии физиологически высокоактивны, что соответствует существующим представлениям о трофической функции центральной клетки.[ …]
Внутренняя организация ситовидных элементов в той мере, как она вскрывается с помощью электронного микроскопа, также не лишена своеобразия. Как правило, в зрелых ситовидных элементах рибосомы полностью отсутствуют, аппарат Гольджи отмечается только на первых стадиях их формирования. Напротив, хорошо развиты митохондрии, пластиды и эндоплазматическая сеть. Вакуоль в этих клетках также отсутствует, что приводит к значительному разжижению цитоплазмы. Сильно развитая эндоплазматическая сеть пронизывает не только цитоплазму ситовидных элементов, но и цитоплазматические тяжи в каналах ситовидных полей. Пластиды также обильны, причем у цветковых растений и некоторых голосеменных можно различать два типа пластид — содержащие белок (Б-тип) и содержащие крахмал (К-тип). Это различие имеет известное таксономическое значение, поскольку большинство двудольных имеет пластиды К-типа, а однодольные — Б-типа. Впрочем, различие это не очень четкое, и нет уверенности, что оно не имеет исключений. Для большинства голосеменных характерно наличие пластид двух типов, тогда как пластиды исследованных в этом отношении папоротников и плаунов не имеют ни белка, ни крахмала.[ …]
Основными элементами клетки являются цитоплазма и ядро.. Цитоплазма представляет собой густую полужидкую массу. Ядро имеет более плотную консистенцию. Растительные клетки заключены в прочную клеточную оболочку. Все содержимое клетки, лишенное клеточной оболочки, называется протопластом. Помимо ядра, в цитоплазме клетки обнаруживаются и другие крупные органеллы, видимые под световым микроскопом — пластиды и митохондрии1 (рис. 6). Кроме того, в ней находятся также многочисленные субмикроскопические структуры, такие, как аппарат Гольджи, эндоплазматическая сеть, рибосомы, микротрубочки и др. Все они погружены в гиалоплазму, служащую матриксом цитоплазмы.[ …]
Ядро имеет более плотную консистенцию. Растительные клетки заключены в прочную клеточную оболочку. Все содержимое клетки, лишенное клеточной оболочки, называется протопластом. Помимо ядра, в цитоплазме клетки обнаруживаются и другие крупные органеллы, видимые под световым микроскопом — пластиды и митохондрии1 (рис. 6). Кроме того, в ней находятся также многочисленные субмикроскопические структуры, такие, как аппарат Гольджи, эндоплазматическая сеть, рибосомы, микротрубочки и др. Все они погружены в гиалоплазму, служащую матриксом цитоплазмы.[ …]
Однако результаты многих наших наблюдений не могут быть объяснены с точки ирония классической схемы распределения слоев ГЛ, 1Л1 и ЫН. Результаты недавней работы Барка и Стюарта (личное сообщение), проведенной с пластидиыми мутантами табака, позволяют предполагать, что нормальный процесс развития листа, вероятно, гораздо болео сложен и изменчив, нежели это считали ранее. Для того чтобы объяснить присутствие небольших островков нормальных клеток в палисадной ткани, мутантной по слоям Ы1 и ЫН, ученые предположили, что нормальные эпидермальные клетки (Ы) могут мигрировать в палисадный слой (Ы1) па некоторой стадии развития листа. Эти клетки (Ы) приобретают свойстиа палисадных, и таким образом возникают занявшие чужое владение островки клоток с нормальными хлоропластами. Мы предположили, что островки: темно-золепых клеток в ткани пунктирного тина образуются именно в результате такого внедрения. Ткань приобретает характерный «пунктирный» вид после того, как слой Ы1, содержащий вирус в высокой концентрации, покрывается эпидермисом Ы темно-зеленого типа. Таким образом, островки темно-зеленых клеток во взрослом листе представляют собой потомство клеток слоя Ы, мигрировавших в слой 1Л1. Микроскопические и небольшие макроскопические островки темно-зеленой ткани, появляющиеся в других участках мозаичного листа, почти всегда связаны с верхним эпидермисом. Причина этого неясна, однако Барк и Стюарт (личное сообщение) обнаружили аналогачиое распределение островков клеток мезофилла с нормальными хлоропластами в пластидных мутантах табака. [ …]
[ …]
Кроме биохимических изменений на молекулярном уровне и структурных изменений, видимых в обычный световой микроскоп, с помощью электронного микроскопа можно обнаружить изменения, происходящие на ультраструктурном уровне. Однако есть и исключения, например в клетках ситовидных трубок во время дифференцировки большинство органелл подвергается дезинтеграции. Наибольшая вариабельность характерна для пластид. Их структура чрезвычайно разнообразна в зависимости от того, находятся ли они в тканях листа, запасающих тканях, плодах (например, томата) или, частях цветка, таких, как лепестки.[ …]
В соответствии с другой гипотезой считают, что эукариотическая клетка произошла от прокариотической клетки, содержавшей несколько геномов, прикрепленных к клеточной мембране. В результате инвагинаций клеточной мембраны образовывались мезо-сомы, способные первоночально к фотосинтезу. Однако в дальнейшем произошла специализация этих органелл, в результате чего одна из них, утратив дыхательную и фотосинтетическую функцию, развилась в ядро, другие, наоборот, развив эти функции, стали митохондриями у животных и пластидами у растений. В пользу этой гипотезы свидетельствуют данные о двойном строении мембран ядра, митохондрий и пластид.[ …]
Пожалуй, самым удивительным является содержимое мюллеровских телец: оно состоит главным образом из гликогена (животного крахмала) — основного запасного углевода животных п грибов. У цекропии (как и у других высших растений) основные запасные углеводы представлены в форме крахмала, гликоген же синтезируется только и мюллеровских тельцах, причем па ранних стадиях их развития, как показали недавние исследования с помощью электронной микроскопии (Ф. Риксон, 1971, 1974), в этих образованиях гликогена нет. Небольшое число гли-когеповых пластид образуется также к жемчужных железках — крохотных беловатых выростах, изредка появляющиеся на черешках и нижней поверхности листьев цекропии м также поедаемых муравьями.[ …]
Все организмы, способные к фотосинтезу, содержат хлорофилл (немногие бактерии, способные к фотосинтезу, имеют пигменты, отличающиеся от хлорофилла).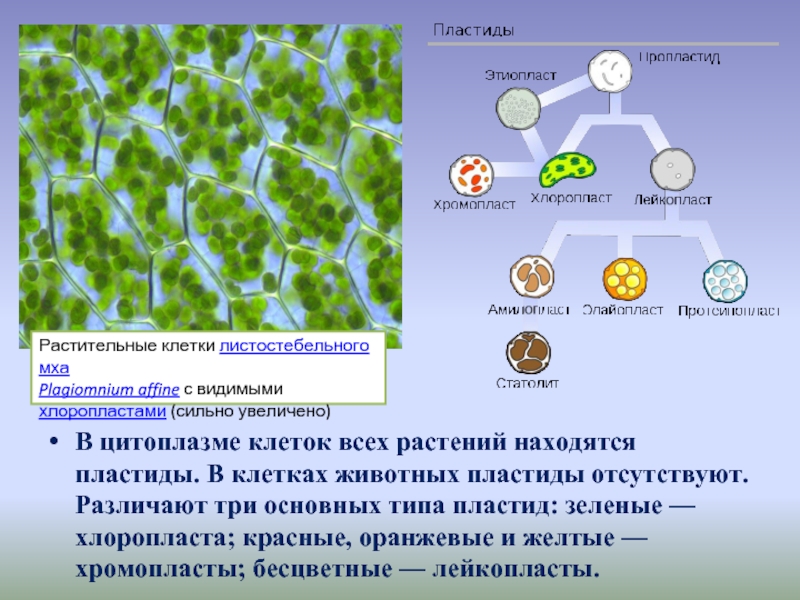 Это зеленый пигмент, содержащийся в особых органоидах растительной клетки — хлоропластах. Они взвешены в цитоплазме клетки в виде мельчайших зеленых гранул и обладают способностью движения в зависимости от освещения. В зеленых листьях высших растений хлоропласты представляют собой мелкие белково-липоидные тельца. В клетке их содержится от 20 до 100. Общее количество хлоропластов во взрослом дереве достигает десятков и сотен миллиардов, но у низших одноклеточных водорослей бывает несколько или даже одна зеленая пластида. Величина зеленой пластиды у высших растений обычно 3—5 мк.[ …]
Это зеленый пигмент, содержащийся в особых органоидах растительной клетки — хлоропластах. Они взвешены в цитоплазме клетки в виде мельчайших зеленых гранул и обладают способностью движения в зависимости от освещения. В зеленых листьях высших растений хлоропласты представляют собой мелкие белково-липоидные тельца. В клетке их содержится от 20 до 100. Общее количество хлоропластов во взрослом дереве достигает десятков и сотен миллиардов, но у низших одноклеточных водорослей бывает несколько или даже одна зеленая пластида. Величина зеленой пластиды у высших растений обычно 3—5 мк.[ …]
Цитоплазмой (протоплазмой) называется все содержимое клетки, за исключением ядра и оболочки. Термин «цитоплазма» был предложен в 1882 г. Э. Страсбургером. По своему значению он более точно указывает на то, что речь идет именно о плазме клетки, а не обо всем содержимом клетки — протопласте, или протоплазме в ее широком понимании. В молодой растительной клетке цитоплазма занимает большую часть ее объема. В эмбриональных клетках растений и животных цитоплазма отличается слабо развитой системой внутриклеточных мембран, почти полностью состоит из гиалоплазмы (основного матрикса) и рибосом. В процессе эволюции клетки возникли внутриклеточные мембраны, а также некоторые клеточные органеллы, например митохондрии, пластиды и центри-оли, составляющие большую часть цитоплазмы (рис. 9).[ …]
Бактерии и сине-зеленые водоросли (циа-неи) — две филогенетически родственные группы — резко отличаются от всех остальных живых существ (в том числе от грибов) отсутствием истинного ядра и тем, что ДНК лежит в их клетке свободно, погруженная в так называемую нуклеоплазму, которая не отделена от цитоплазмы ядерной мембраной. У них нет также митохондрий и сложных жгутиков. Жгутики у них (когда они имеются) устроены проще и имеют принципиально иное строение, чем у остальных организмов; их клеточная стенка состоит из гетерополимерного вещества му-реина, которое не было обнаружено ни у одной другой группы организмов. Эти организмы называют прокариотами (Ргосагуо-1а — доядерные). У всех остальных организмов, как одноклеточных, так и многоклеточных, имеется настоящее ядро, окруженное ядерной мембраной и тем самым резко отграниченное от цитоплазмы. Такие организмы называют эукариотами (Еисагуо1а — ядерные). Кроме ясно дифференцированного ядра и цитоплазмы, у них имеются также митохондрии, а у многих также пластиды и сложные жгутики. Постепенно стало выясняться, что различия между прокариотами и эукариотами гораздо более глубокие и фундаментальные, чем, например, различия между высшими животными и высшими растениями (те и другие—эукариоты).[ …]
Эти организмы называют прокариотами (Ргосагуо-1а — доядерные). У всех остальных организмов, как одноклеточных, так и многоклеточных, имеется настоящее ядро, окруженное ядерной мембраной и тем самым резко отграниченное от цитоплазмы. Такие организмы называют эукариотами (Еисагуо1а — ядерные). Кроме ясно дифференцированного ядра и цитоплазмы, у них имеются также митохондрии, а у многих также пластиды и сложные жгутики. Постепенно стало выясняться, что различия между прокариотами и эукариотами гораздо более глубокие и фундаментальные, чем, например, различия между высшими животными и высшими растениями (те и другие—эукариоты).[ …]
Поскольку в этом случае первый продукт карбоксилирования — ЩУК содержит четыре атома углерода, его называют «С-4» путь, в отличие от цикла Кальвина, где образуется ФГК, содержащая три атома углерода («С-3 путь). Щавелевоуксусная кислота преобразуется в яблочную кислоту. В последующем происходит реакция транскарбоксилирования, при которой СО2 снова отщепляется от органических кислот и вступает в цикл Кальвина — присоединяется к рибулезодифосфату (рис. 44). Таким образом, сущность «С-4» пути заключается в том, что реакция карбоксилирования происходит дважды. Это позволяет растению создавать запасы углерода в клетках. Как и во всяком биохимическом цикле, акцепторы (ФЕП о РДФ) регенерируют, что и создает возможность его непрерывного функционирования. Исследования показали, что в растениях, в которых процесс фотосинтеза протекает по «С-4» пути, имеются два типа хлоропластов; 1) крупные пластиды, часто лишенные гран, в клетках обкладки, окружающих сосудистые пучки; 2) мелкие гра-нальжые пластиды в клетках мезофилла листа.[ …]
какие виды пластид вам известны ? могут ли они перходить друг в друга ?
Пластиды — типы, происхождение, строение, функции
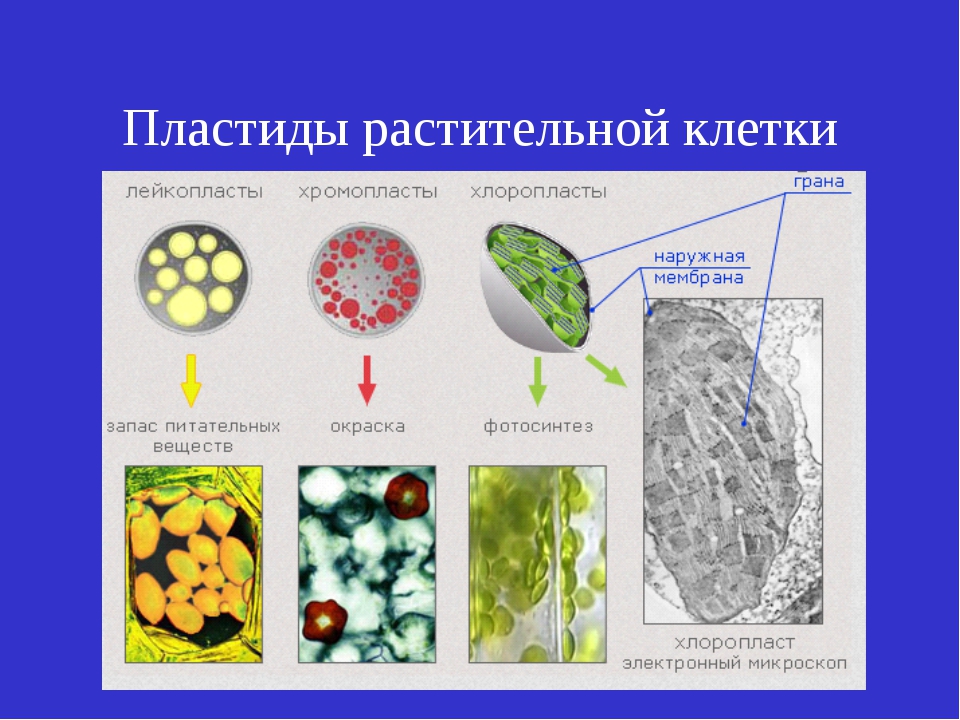
Пластиды – органоиды, специфичные для клеток растений (они имеются в клетках всех растений, за исключением большинства бактерий, грибов и некоторых водорослей). В клетках высших растений находится обычно от 10 до 200 пластид размером 3–10 мкм, чаще всего имеющих форму двояковыпуклой линзы. У водорослей зеленые пластиды, называемые хроматофорами, очень разнообразны по форме и величине. Они могут иметь звездчатую, лентовидную, сетчатую и другие формы.Различают бесцветные пластиды – лейкопласты, и окрашенные – хлоропласты (зеленого цвета), хромопласты (желтого, красного и других цветов). Эти виды пластид до известной степени способны превращаться друг в друга – лейкопласты при накоплении хлорофилла переходят в хлоропласты, а последние при появлении красных, бурых и других пигментов – в хромопласты.Внутреннее строение пластид очень сложно. В хлоропластах есть свои рибосомы, ДНК, РНК, включения жира, зерна крахмала. Снаружи хлоропласты покрыты двумя белково-липидными мембранами, а в их полужидкую строму (основное вещество) погружены мелкие тельца – граны и мембранные каналы. Граны (размером около 1 мкм) – пакеты круглых плоских мешочков (тилакоидов), сложенных подобно столбику монет. Располагаются они перпендикулярно поверхности хлоропласта. Тилакоиды соседних гран соединены между собой мембранными каналами, образуя единую систему. Число гран в хлоропластах различно. Например, в клетках шпината каждый хлоропласт содержит 40–60 гран. Хлоропласты внутри клетки могут двигаться пассивно, увлекаемые током цитоплазмы, либо активно перемещаться с места на место. Если свет очень интенсивен, они поворачиваются ребром к ярким лучам солнца и выстраиваются вдоль стенок, параллельных свету. При слабом освещении хлоропласты перемещаются на стенки клетки, обращенные к свету, и поворачиваются к нему своей большой поверхностью. При средней освещенности они занимают среднее положение. Этим достигаются наиболее благоприятные для процесса фотосинтеза условия освещения.В гранах содержится хлорофилл, упакованный с белковыми и фосфолипидными молекулами так, чтобы обеспечить способность улавливать световую энергию,Молекула хлорофилла очень сходна с молекулой гемоглобина и отличается главным образом тем, что расположенный в центре молекулы гемоглобина атом железа заменен в хлорофилле на атом магния.
У водорослей зеленые пластиды, называемые хроматофорами, очень разнообразны по форме и величине. Они могут иметь звездчатую, лентовидную, сетчатую и другие формы.Различают бесцветные пластиды – лейкопласты, и окрашенные – хлоропласты (зеленого цвета), хромопласты (желтого, красного и других цветов). Эти виды пластид до известной степени способны превращаться друг в друга – лейкопласты при накоплении хлорофилла переходят в хлоропласты, а последние при появлении красных, бурых и других пигментов – в хромопласты.Внутреннее строение пластид очень сложно. В хлоропластах есть свои рибосомы, ДНК, РНК, включения жира, зерна крахмала. Снаружи хлоропласты покрыты двумя белково-липидными мембранами, а в их полужидкую строму (основное вещество) погружены мелкие тельца – граны и мембранные каналы. Граны (размером около 1 мкм) – пакеты круглых плоских мешочков (тилакоидов), сложенных подобно столбику монет. Располагаются они перпендикулярно поверхности хлоропласта. Тилакоиды соседних гран соединены между собой мембранными каналами, образуя единую систему. Число гран в хлоропластах различно. Например, в клетках шпината каждый хлоропласт содержит 40–60 гран. Хлоропласты внутри клетки могут двигаться пассивно, увлекаемые током цитоплазмы, либо активно перемещаться с места на место. Если свет очень интенсивен, они поворачиваются ребром к ярким лучам солнца и выстраиваются вдоль стенок, параллельных свету. При слабом освещении хлоропласты перемещаются на стенки клетки, обращенные к свету, и поворачиваются к нему своей большой поверхностью. При средней освещенности они занимают среднее положение. Этим достигаются наиболее благоприятные для процесса фотосинтеза условия освещения.В гранах содержится хлорофилл, упакованный с белковыми и фосфолипидными молекулами так, чтобы обеспечить способность улавливать световую энергию,Молекула хлорофилла очень сходна с молекулой гемоглобина и отличается главным образом тем, что расположенный в центре молекулы гемоглобина атом железа заменен в хлорофилле на атом магния. В природе встречается четыре типа хлорофилла: а, b, с, е. Хлорофиллы а и b содержат высшие растения и зеленые водоросли, диатомовые водоросли содержат а и с, красные – а и <1. Лучше других изучены хлорофиллы а и b (их впервые разделил русский ученый М. С. Цвет в начале XX в.).Кроме них существуют четыре вида бактериохлорофиллов – зеленых пигментов пурпурных и зеленых бактерий: а, b, с, с1. Большинство фотосинтезирующих бактерий содержат бактериохлорофилл а, некоторые – бактериохлорофилл b, зеленые бактерии – с и с1. Хлорофилл обладает способностью очень эффективно поглощать солнечную энергию и передавать ее другим молекулам. Благодаря этой способности хлорофилл – единственная структура на Земле, которая обеспечивает процесс фотосинтеза. Пластидам, так же, как и митохондриям, свойственна до некоторой степени автономность внутри клетки. Они размножаются путем деления.Наряду с фотосинтезом, в пластидах происходит процесс биосинтеза белка.Благодаря содержанию ДНК пластиды играют определенную роль в передаче признаков по наследству (цитоплазматическая наследственность).
В природе встречается четыре типа хлорофилла: а, b, с, е. Хлорофиллы а и b содержат высшие растения и зеленые водоросли, диатомовые водоросли содержат а и с, красные – а и <1. Лучше других изучены хлорофиллы а и b (их впервые разделил русский ученый М. С. Цвет в начале XX в.).Кроме них существуют четыре вида бактериохлорофиллов – зеленых пигментов пурпурных и зеленых бактерий: а, b, с, с1. Большинство фотосинтезирующих бактерий содержат бактериохлорофилл а, некоторые – бактериохлорофилл b, зеленые бактерии – с и с1. Хлорофилл обладает способностью очень эффективно поглощать солнечную энергию и передавать ее другим молекулам. Благодаря этой способности хлорофилл – единственная структура на Земле, которая обеспечивает процесс фотосинтеза. Пластидам, так же, как и митохондриям, свойственна до некоторой степени автономность внутри клетки. Они размножаются путем деления.Наряду с фотосинтезом, в пластидах происходит процесс биосинтеза белка.Благодаря содержанию ДНК пластиды играют определенную роль в передаче признаков по наследству (цитоплазматическая наследственность).
Строение пластидов
Одной из важных живых частей (органелл) клетки являются пластиды. Различают три вида пластид: бесцветные лейкопласты, оранжевые или красно-оранжевые хромопласты и зеленые хлоропласты.
Лейкопласты могут быть родоначальниками хлоропластов. Известно, что выращенные в темноте растения бывают бледно-желтого цвета. Они содержат лейкопласты, которые на свету зеленеют, т. е. переходят в хлоропласты. Хлоропласта в созревающих плодах переходят в хромопласты.
Окраска пластид обусловлена наличием особых красящих веществ — пигментов. У зеленых пластид — хлоропластов окраска обусловлена наличием хлорофилла, а у хромопластов — наличием оранжевого пигмента каротина.
Существуют, однако, лейкопласты, не зеленеющие на свету. Такие лейкопласты можно найти в
У высших растений пластиды имеют большей частью округлую форму. Хромопласты нередко бывают палочковидными, например в клетках корня моркови.
Хлоропласты играют основную роль в питании зеленого растения. На солнечном свету зеленое растение в своих хлоропластах строит из воды и углекислого газа углеводы, выделяя в окружающую атмосферу кислород.
Хлоропласты у высших растений представлены в виде хлорофилловых зерен. У водорослей пластиды обычно цельные, имеют вид пластинок, спиральных лент, чашевидных телец и называются хроматофорами (носителями окраски).
Тело пластиды имеет плазменную белковую основу (строму), на поверхности которой адсорбирован пигмент.
По данным Н. М. Сисакяна (1956), все виды пластид играют еще одну весьма важную роль в клетке, так как содержат целый ряд ферментов. Сисакян считает, что пластиды являются своеобразными депо ферментов в растении.
У водорослей, мхов и папоротников хлоропласты возникают заново путем их деления. В яйцеклетке многих мхов содержатся хлоропласты. Однако у высших растений они возникают из митохондрий. Крупный русский ученый С. Г. Навашин считал, что митохондрии возникают из микроскопических гранул, находящихся в цитоплазме клеток. В последнее время немецкий ученый З. Штруггер (1956) высказывает мнение, что хлоропласты возникают не из митохондрий, а из особых образований — пропластид.
5.Пластиды, строение и их биологическое значение. Типы пластид.
От греческого слова — ищменчивый, пластичный.
Пластиды – органоиды, содержащиеся только в растительной клетке. Каждая пластида ограничена двумя элементарными мембранами. Пластиды разнообразны по форме, строению и функциям. В зависимости от окраски, различают три основных типа пластид: хлоропласты – зеленые, хромопласты – желто-оранжевые или красные, лейкопласты – бесцветные.
Пластиды
имеют единое происхождение от пропластид
меристематических клеток. Возможны
взаимные превращения пластид в онтогенезе
вида. Обычно в клетке содержится только
один из типов пластид.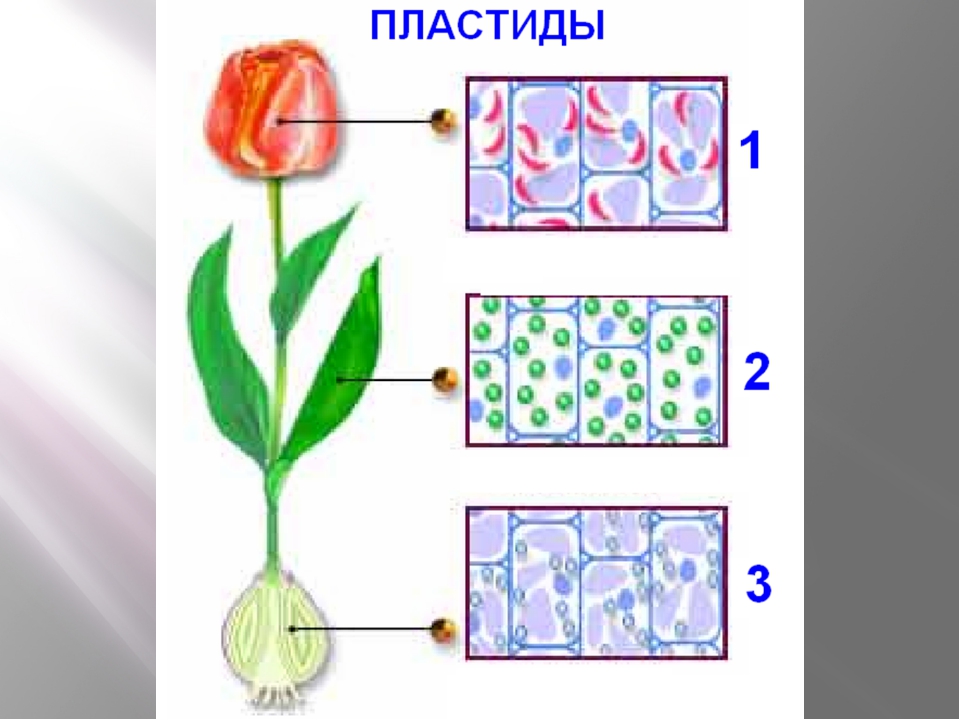
Хлоропласты широко распространены у низших и высших растений. Зеленая окраска хлоропластов обусловлена присутствием зеленого пигмента – хлорофилла. В хлоропластах при участии солнечной энергии идет фотосинтез – процесс образования органических веществ из воды и углекислого газа.
Кроме хлорофилла в состав хлоропластов входят каротиноиды: каротин – пигмент оранжево-красный; ксантофилл – желтый. В зеленых листьях они маскируются хлорофиллом и становятся заметными при его разрушении (например, осенью или при заболеваниях). Форма относительно постоянна – линзообразная. Чаще всего хлоропласты равномерно располагаются по цитоплазме, но способны к движению. Они меняют своё положение в зависимости от условий освещения так, чтобы наилучшим образом улавливать свет.
Хромопласты – пластиды желтого или красно-оранжевого цвета. Хромопласты встречаются в созревающих плодах (томаты, шиповник, рябина, арбуз), в клетках лепестков (роза, лютик, одуванчик), в корнеплодах (морковь), в осенних листьях.
В отличие от хлоропластов форма хромопластов очень изменчива: глобулярная, фибриллярная, кристаллическая. Например, в плодах рябины хромопласты имеют вытянутую, заостренную, слегка изогнутую форму, в клетках плодов шиповника и перца красного – овальную.
Лейкопласты пигментов не содержат. Это обычно довольно мелкие пластиды. Встречаются в клетках корней, корневищ и клубней, а также в семенах, других органах, скрытых от солнечного света. Лейкопласты не имеют строго определенной формы: они бывают округлые, яйцевидные, веретенообразные, палочковидные, амебовидные, чашевидные и т.д.; причем форма их даже в одной клетке может меняться несколько раз. В клетке они скапливаются вокруг ядра.
Главная
функция пластид – синтез органических
веществ, благодаря наличию собственных
ДНК и РНК и структур белкового синтеза.
В пластидах также содержатся пигменты,
обусловливающие их цвет. Все виды данных
органелл имеют сложное внутреннее
строение. Снаружи пластиду покрывают
две элементарные мембраны, имеется
система внутренних мембран, погруженных
в строму или матрикс.
Главная функция пластид – синтез органических веществ, благодаря наличию собственных ДНК и РНК и структур белкового синтеза. В пластидах также содержатся пигменты, обусловливающие их цвет. Все виды данных органелл имеют сложное внутреннее строение. Снаружи пластиду покрывают две элементарные мембраны, имеется система внутренних мембран, погруженных в строму или матрикс.
6.Клеточная
стенка, ее биологическое значение. Клеточная
стенка (нередко в качестве синонима
термина «клеточная стенка» в учебной
и научной литературе используется
термин «клеточная оболочка».) у
растений — это структурное образование,
располагающееся по периферии клетки,
за пределами плазмалеммы ,
придающее клетке прочность, сохраняющее
ее форму и защищающее протопласт .
Клеточная
стенка растений противостоит высокому
осмотическому давлению большой
центральной вакуоли и
препятствует разрыву клетки. Кроме
того, совокупность прочных клеточных
стенок выполняет роль своеобразного
внешнего скелета, поддерживающего форму
растения и придающего ему механическую
прочность. Клеточная стенка, обладая
большой прочностью, в то же время способна
к росту, и прежде всего к росту растяжением. Клеточная
стенка, как правило, прозрачна и хорошо
пропускает солнечный свет. Через нее
легко проникают вода и низкомолекулярные
вещества, но для высокомолекулярных
веществ она полностью или частично
непроницаема. У многоклеточных организмов
стенки соседних клеток скреплены между
собой пектиновыми веществами, образующими
срединную пластинку.
В
результате тургорного давления стенки
соседних клеток в углах могут округляться
и между ними образуются межклетники.
Стенка
клетки представляет собой продукт
жизнедеятельности ее протопласта .
Поэтому стенка может расти, только
находясь в контакте с протопластом.
Однако при отмирании протопласта стенка
сохраняется и мертвая клетка может
продолжать выполнять функции проведения
воды или играть роль механической
опоры.
Основу
клеточной стенки составляют высокополимерные
углеводы: молекулы целлюлозы
(клетчатки) ,
собранные в сложные пучки — фибриллы,
образующие каркас, погруженный в основу
(матрикс), состоящий
из гемицеллюлоз , пектинов и гликопротеидов ( рис. 21 ).
Молекулы целлюлозы состоят
из большого числа линейно расположенных
мономеров — остатков глюкозы .
Целлюлоза очень стойка, не растворяется
в разбавленных кислотах и даже в
концентрированных щелочах. Эластичный
целлюлозный скелет придает клеточной
оболочке механическую прочность.
Первоначально число микрофибрилл,
образованных молекулами целлюлозы, в
клеточной стенке относительно невелико,
но с возрастом оно увеличивается и
клетка теряет способность к
растяжению.
Гемицеллюлозы отличаются
от целлюлозы составом
мономеров и разветвленным их расположением
в молекулах. Являясь одним из компонентов
пластичного матрикса, гемицеллюлозы придают
клеточной стенке дополнительную
прочность, но почти не препятствуют ее
росту. Гемицеллюлозы могут
быть и запасными веществами, так как
легко гидролизуются. Кроме гемицеллюлоз
в матрикс, а также в срединную пластинку
входят пектиновые вещества, или пектины ,
и полисахариды ,
образованные мономерами — уроновыми
кислотами .
Эти вещества скрепляют, склеивают
оболочки соседних клеток. Молекулы
гемицеллюлоз, пектина и гликопротеидов
соединяют целлюлозные
микрофибриллы.
Помимо полисахаридов ,
в матриксе стенок многих клеток часто
обнаруживаются неуглеводные компоненты.
Наиболее обычен из них лигнин —
полимерное вещество полифенольной
природы. Содержание его в стенках
некоторых видов клеток может достигать
30%. Лигнин откладывается
при завершении роста стенки. Процесс
отложения лигнина получил
название одревеснения,
или лигнификации .
Стенка, пропитанная лигнином, очень
прочна и тверда. Лигнифицируются чаще
всего оболочки клеток, подвергающихся
механическим нагрузкам.
Стенки
некоторых типов клеток могут включать
слои липидов : воска , кутина и суберина . Кутин и воскобычно
покрывают наружные стенки клеток
эпидермы. Слой кутина создает на
поверхности растения водо- и
воздухонепроницаемый слой кутикулы .
Суберин пропитывает стенки. Он непроницаем
для воды и газов, поэтому такая
суберинизированная, или опробковевшая,
клетка быстро отмирает.
21 ).
Молекулы целлюлозы состоят
из большого числа линейно расположенных
мономеров — остатков глюкозы .
Целлюлоза очень стойка, не растворяется
в разбавленных кислотах и даже в
концентрированных щелочах. Эластичный
целлюлозный скелет придает клеточной
оболочке механическую прочность.
Первоначально число микрофибрилл,
образованных молекулами целлюлозы, в
клеточной стенке относительно невелико,
но с возрастом оно увеличивается и
клетка теряет способность к
растяжению.
Гемицеллюлозы отличаются
от целлюлозы составом
мономеров и разветвленным их расположением
в молекулах. Являясь одним из компонентов
пластичного матрикса, гемицеллюлозы придают
клеточной стенке дополнительную
прочность, но почти не препятствуют ее
росту. Гемицеллюлозы могут
быть и запасными веществами, так как
легко гидролизуются. Кроме гемицеллюлоз
в матрикс, а также в срединную пластинку
входят пектиновые вещества, или пектины ,
и полисахариды ,
образованные мономерами — уроновыми
кислотами .
Эти вещества скрепляют, склеивают
оболочки соседних клеток. Молекулы
гемицеллюлоз, пектина и гликопротеидов
соединяют целлюлозные
микрофибриллы.
Помимо полисахаридов ,
в матриксе стенок многих клеток часто
обнаруживаются неуглеводные компоненты.
Наиболее обычен из них лигнин —
полимерное вещество полифенольной
природы. Содержание его в стенках
некоторых видов клеток может достигать
30%. Лигнин откладывается
при завершении роста стенки. Процесс
отложения лигнина получил
название одревеснения,
или лигнификации .
Стенка, пропитанная лигнином, очень
прочна и тверда. Лигнифицируются чаще
всего оболочки клеток, подвергающихся
механическим нагрузкам.
Стенки
некоторых типов клеток могут включать
слои липидов : воска , кутина и суберина . Кутин и воскобычно
покрывают наружные стенки клеток
эпидермы. Слой кутина создает на
поверхности растения водо- и
воздухонепроницаемый слой кутикулы .
Суберин пропитывает стенки. Он непроницаем
для воды и газов, поэтому такая
суберинизированная, или опробковевшая,
клетка быстро отмирает.
Биология для студентов — 21.
 Пластиды высших и низших растений
Пластиды высших и низших растенийПластиды – это мембранные органоиды, встречающиеся у фотосинтезирующих эукариотических организмов (высшие растения, низшие водоросли, некоторые одноклеточные организмы).
Они выполняют различные функции, связанные, главным образом, с синтезом органических веществ. В зависимости от окраски, обусловленной наличием пигментов, различают три основных типа пластид:
- хлоропласты,
- хромопласты,
- лейкопласты.
Хлоропласты — зеленые пластиды, содержащие зеленый пигмент хлорофилл и небольшое количество каротина и ксантофилла. Главная функция хлоропластов — фотосинтез, в результате которого происходит образование богатых энергией органических веществ. Синтез хлорофилла обычно происходит только на свету, поэтому растения, выращенные в темноте или при недостатке света, становятся бледно-желтыми и называются этиолированными. Вместо типичных хлоропластов в них образуются этиопласты. В клетках низших растений (водорослей) хлоропласты крупные и немногочисленные (один или несколько). Они имеют разнообразную форму (пластинчатую, звездчатую, ленточную и др.). Такие хлоропласты называются хроматофорами.
Хромопласты представляют собой пластиды, содержащие пигменты из группы каротиноидов, имеют желтую, оранжевую или красную окраску. К каротиноидам относят широко распространенные каротины (оранжевые) и ксантофиллы (желтые). Хромопласты имеют разнообразную форму. Они образуются в осенних листьях, корнеплодах (морковь), зрелых плодах и т.д. В отличие от хлоропластов, форма хромопластов очень изменчива, но видоспецифична, что объясняется их происхождением и состоянием в них пигментов.
Лейкопласты — это мелкие бесцветные пластиды шаровидной, яйцевидной или веретеновидной формы. Они обычно встречаются в клетках органов, скрытых от солнечного света: в корневищах, клубнях, корнях, семенах, сердцевине стеблей и очень редко — в клетках освещенных частей растения (в клетках эпидермы). Часто лейкопласты собираются вокруг ядра, окружая его со всех сторон. Деятельность лейкопластов специализирована и связана с образованием запасных веществ. Одни из них накапливают преимущественно крахмал (амилопласты), другие — белки (протеопласты или алейронопласты), а третьи — масла (олеопласты).
Часто лейкопласты собираются вокруг ядра, окружая его со всех сторон. Деятельность лейкопластов специализирована и связана с образованием запасных веществ. Одни из них накапливают преимущественно крахмал (амилопласты), другие — белки (протеопласты или алейронопласты), а третьи — масла (олеопласты).
Пластиды окружены двумя мембранами, в их матриксе имеется собственная геномная система, функции пластид связаны с энергообеспечением клетки, идущим на нужды фотосинтеза. У высших растений найден целый набор различных пластид (хлоропласт, лейкопласт, амилопласт, хромопласт), представляющих собой ряд взаимных превращений одного вида пластиды в другой. Основной структурой, которая осуществляет фотосинтетические процессы, является хлоропласт. У высших растений также встречается деление зрелых хлоропластов, но очень редко.
Увеличение числа хлоропластов и образование других форм пластид (лейкопластов и хромопластов) следует рассматривать как путь превращения структур-предшественников, пропластид. Весь же процесс развития различных пластид можно представить в виде монотропного (идущего в одном направлении) ряда смены форм:
Многими исследованиями был установлен необратимый характер онтогенетических переходов пластид. У высших растений возникновение и развитие хлоропластов происходят через изменения пропластид. Пропластиды представляют собой мелкие (0,4-1 мкм) двумембранные пузырьки, не имеющие отличительных черт их внутреннего строения. Они отличаются от вакуолей цитоплазмы более плотным содержимым и наличием двух отграничивающих мембран, внешней и внутренней. Внутренняя мембрана может давать небольшие складки или образовывать мелкие вакуоли. Пропластиды чаще всего встречаются в делящихся тканях растений (клетки меристемы корня, листьев, в точки роста стеблей и др.). По всей вероятности, увеличение их числа происходит путем деления или почкования, отделения от тела пропластиды мелких двумембранных пузырьков.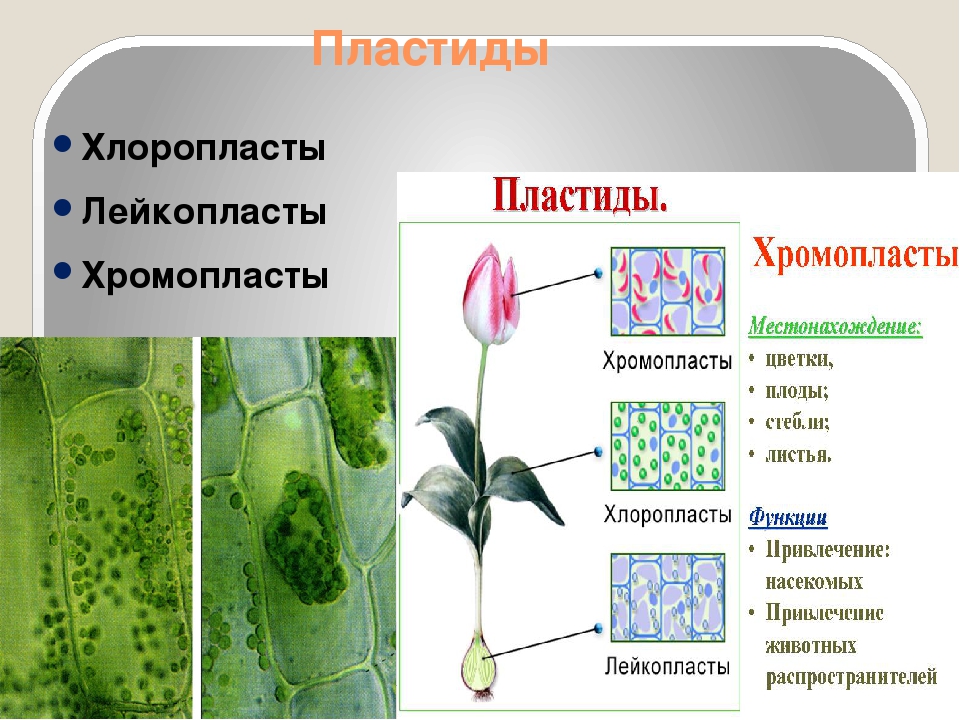
Урок биологии в 5 «А» классе ФГОС «Пластиды в клетках растений»
Урок биологии в 5 «А» классе ФГОС «Пластиды в клетках растений»
Автор: О.В.Высоцкая
Биология
Задачи урока:
1. Образовательная: создание условий для самостоятельной деятельности, направленной на изучение пластид в клетках растений.
2. Развивающая: продолжить формирование умений работать с микроскопом; научиться готовить микропрепараты клеток листа элодеи, клеток томата; распознавать основные части клетки на микропрепарате и таблице.
3. Воспитательная: воспитание положительной мотивации учения, умение работать индивидуально и в группе .
Методы и методические приѐмы
Методы, способствующие первичному усвоению учебного материала: информационно-развивающие, поисковые. Методы, способствующие закреплению и совершенствованию приобретѐнных знаний (упражнения, практическая работа, решение проблемных задач).
Тип урока: изучение нового материала, лабораторная работа.
Оборудование: ПК, медиапроектор, микроскопы , оборудование для лабораторной работы, прозеленённые клубни картофеля, растение колеус ,презентация по теме «Пластиды».
Содержательная часть урока:
|
Этапы урока |
Деятельность учителя |
Деятельность ученика |
Развиваемые УУД |
|
1.Организационный момент |
Взаимные приветствия учителя и учащихся; организация внимания и внутренней готовности. |
Учащиеся настраиваются на работу на уроке ,проверяют наличие учебных принадлежностей. |
Сформировать умение слушать и понимать речь других людей. |
|
2. Актуализация |
Заглянём ка на часок В нашу клетку –теремок Там такое происходит Цитоплазма кругом ходит, Помогает то движенье В клетке чудным превращеньям. Аещё в ней там и тут органоиды живут Формулировка темы и цели урока: Выяснить степень усвоения заданного на дом материала; определить типичные недостатки в знаниях и их причины; ликвидировать обнаруженные пробелы. 1) устный ответ с места : — назовите известные вам увеличительные приборы -как приготовить микропрепарат клеток кожицы лука? 2) ответ у доски: назовите и покажите основные органоиды клетки ? 1 ученик оценивает ответы товарищей (показывая смалики). 1 и 2 парты каждого ряда выполняют письменную работу на карточках(заранее раздать по этим вопросам): 1. назовите основные органоиды растительной клетки. 2.Соотнесите название органоида клетки и выполняемую функцию (Слайд). |
Предполагают тему урока и формулируют цели урока. Отвечают на вопросы Работа в карточках. Работают у доски. Взаимопроверка, оценка и обсуждение результата ( что необходимо сделать, чтобы лучше усвоить учебный материал?). |
Целеполагание построение логической цепи рассуждений Логические:подведение под понятие, выведение следствий. |
|
3. Мотивация |
Слайды на доске « Органоиды растительной и животной клеток». Сравните схемы строения растительной и животной клетки. Какие вы увидели отличия? Как называются эти мелкие тельца? Ответ найдите в тексте учебника на стр.37. Мелкие тельца –пластиды,органоиды придающие окраску листьям, цветкам, плодам и они могут быть не только зелёного цвета. |
Предполагаемый ответ: в растительных клетках находятся мелкие тельца зелёного цвета. |
Общеучебные: знаково-символические (работа с рисунками ). прогнозирование (предвосхищение результата и уровня усвоения). Формирование умения ориентироваться в учебнике, находить и использовать нужную информацию |
|
4. Изучение нового материала. |
Добиться от учащихся восприятия, осознания, первичного обобщения и систематизации новых знаний, усвоения учащимися способов, путей, средств, которые привели к данному обобщению; на основе приобретаемых знаний вырабатывать соответствующие УУД. Формулируем тему и цель урока . Объяснение учителя об особенностях строения пластид с привлечением дополнительного материала и демонстрацией слайдов. Цвет пластидам обеспечивают пигменты. Хлоропласты содержат зелёный пигмент хлорофилл.(обеспечивает зелёную окраску) Слайд Хромопласты содержат оранжевый пигмент обеспечивающий окраску плодов, лепестков, листьев осенью. Лейкопласты-бесцветные пластиды находятся в клетках чешуй луковиц, в клубнях картофеля. Демонстрация: пластиды могут взаимопереходить друг в друга ( демонстрация позеленённого клубня картофеля и осенних листьев. Физкультминутка. Лабораторная работа «Пластиды в клетках листа элодеи в плодах томата и яблока». Рассматривание микропрепарата пластид в клетках томата. |
Записывают тему в тетрадь. Формулируют цель урока, знакомятся с планом достижения цели. Слушают объяснение учителя. Анализ текста в учебнике стр.37 Самостоятельно составляют схему в тетрадях. Слушают обьяснение учителя Выполняют работу по инструкции в учебнике стр. 37, 39 и инструктивной карточке. (приложение № 1). Самостоятельно рассматривают микропрепарат ,продолжают заполнение таблицы по слайду. |
Целеполагание :выражение своих мыслей Логические:подведение под понятие, выведение следствий; работа в группе) планирование :(определение цели, функций участников, способов взаимодействия) постановка вопросов (инициативное сотрудничество в поиске и сборе информации) Формирование умения строить логическое рассуждение, включающее установление причинно-следственных связей. |
|
5. Первичное закрепление |
Установить, усвоили или нет учащиеся связь между фактами, содержание новых понятий, закономерностей, устранить обнаруженные пробелы 1. 2. Бесцветные пластиды…. 3. Окраску плодам, лепесткам придают… 4. Лейкопласты на свету превращаются в…. 5. Хлоропласты на свету могут превращаться в…. 6.Осенью когда дни становятся короче хлоропласты листьев превращаются в…… 7. Красные или жёлтые пластиды…. 8 В каких пластидах содержится хлорофилл….. 9. Какие пластиды встречаются в клубнях картофеля… |
Поднимают заготовки (квадратики, вырезанные из цветной бумаги: хлоропласты-зелёный цвет,хромопласты-оранжевый, жёлтый цвет, лейкопласты-белый цвет). |
оценка выделение и осознание учащимися того, что уже усвоено и что ещѐ подлежит усвоению, осознание качества и уровня усвоения |
|
6. Рефлексия учебной деятель ности. |
Слайд: Сегодня я узнал…. Было интересно… Я научился… Я понял,что… Меня удивило… |
Выбирают начало фразы и формулируют ответ |
Логические: построение цепи рассуждений, обобщение, анализ, синтез. Формирование умения в диалоге с учителем совершенствовать самостоятельно выработанные критерии оценки. |
|
7. Домашнее задание |
1. В учебнике стр.37-40., вопросы №1-4. 2. |

 В клетках листа элодеи вы увидели пластиды…
В клетках листа элодеи вы увидели пластиды… Творческое:сделать модель-аппликацию растительной клетки.
Творческое:сделать модель-аппликацию растительной клетки.
1 настроим микроскоп
2 направим луч света в отверстие предметного столика
3 на предметное стекло нанесём капельку воды.
4 положим немного препарата и разровняем препаравальной иглой до тонкого слоя
5 накроем микрпрепарат покровным стеклом, и положим на предметный столик микроскопа
6 вращая винты найдём изображение препарата.
Строение растительной клетки — Kid-mama
Клетка-это элементарная живая система, тот кирпичик, из которого состоят все живые организмы, только у многоклеточных клетки различаются по строению и функциям, а у одноклеточных клетка сама-целый организм.
Клеткам присущи все свойства живого организма — обмен веществ, а значит и дыхание, питание и выделение; выработка необходимой для жизни энергии , размножение с передачей наследственной информации и др.Вся живая материя на Земле делится на два надцарства — прокариоты (доядерные) и эукариоты (ядерные). К прокариотам относятся бактерии и цианобактерии. Все остальные организмы являются эукариотами, т.е. имеют ядро.
Рассмотрим подробно строение растительной клетки.
Клетки растений, так же как и клетки животных, состоят из наружной мембраны, цитоплазмы (протоплазмы) и ядра. Цитоплазма в свою очередь состоит из гиалоплазмы — желеобразного белкового вещества, и различных органелл, расположенных в ней.
У растений, грибов и бактерий, в отличие от животных и простейших, снаружи от цитоплазматической мембраны имеется еще одна оболочка — так называемая клеточная стенка. Она жесткая, прочная, и выполняет защитную, механическую и транспортную функции. У растений она состоит из целлюлозы (клетчатки) и пектинов. (у грибов -из хитина, а у бактерий из полисахаридов). Со временем происходит одревеснение клеточной стенки и отмирание клетки.
Она жесткая, прочная, и выполняет защитную, механическую и транспортную функции. У растений она состоит из целлюлозы (клетчатки) и пектинов. (у грибов -из хитина, а у бактерий из полисахаридов). Со временем происходит одревеснение клеточной стенки и отмирание клетки.
У злаков и некоторых других растений в клеточной стенке откладываются минеральные вещества, благодаря чему растение становится жестче — это защита от поедания животными.
В клеточной стенке имеются отверстия, через которые соседние клетки сообщаются друг с другом с помощью выростов — плазмодесм.
Гиалоплазма представляет собой густой раствор различных неорганических и органических молекул в воде. Она движется внутри клетки, увлекая за собой органеллы. Также она может пропускать через себя одни вещества и не пропускать другие, участвуя в обмене веществ как внутри клетки, так и между клетками.
Кроме того в растительной клетке находятся полости, ограниченные мембраной, и заполненные вязким клеточным соком — вакуоли. Клеточный сок — это вода с растворенными в ней минеральными солями и органическими веществами -глюкозой, фруктозой, пектинами и др., а также продуктами обмена. Иногда клеточный сок окрашен, и придает окраску растению, Например, у краснокочанной капусты. В старых клетках вакуоль может занимать почти весь объем клетки. Вакуоли служат хранилищем запасов воды в клетке, и придают тургор (плотность) тканям растения.
Цитоскелет. Цитоплазма клеток эукариотов пронизана трехмерной сетью белковых нитей, образующих так называемый цитоскелет. Цитоскелет выполняет три основные функции:
- Служит механическим каркасом, придающим клетке форму, а также связывает мембрану с органеллами
- Работает как мотор при движении или работе клетки (у животных — работа мышц, у растений — движение листьев, раскрывание лепестков и т.д)
- Служит «рельсами» для передвижения органелл, например, митохондрий или пластидов, внутри клетки
Митохондрии
Митохондрии — энергетические станции клетки. В них происходит образование энергии, необходимой для всех жизненных процессов в клетке. Поглощая из цитоплазмы органические вещества, митохондрии расщепляют их при участии кислорода (иначе говоря, «сжигают» ), и синтезируют из них молекулы АТФ — основной источник энергии в клетке. При этом также образуется вода и углекислый газ. Таким образом митохондрии-основные потребители кислорода в клетке, и первыми страдают при его недостатке. Митохондрии состоят из двух мембран, гладкой наружной и складчатой внутренней. Складки внутренней мембраны образуют перегородки — кристы.
Пластиды
Пластиды — органеллы, состоящие из мембран, и содержащие особые пигменты. Пластиды имеются только у растений.
Выделяют три вида пластид : хлоропласты, хромопласты и лейкопласты.
Хлоропласты — содержат зеленый пигмент хлорофилл, они присутствуют в листьях и других зеленых частях растения. Хлоропласты участвуют в фотосинтезе — образовании органических веществ из воды и углекислого газа , используя энергию солнечного света.
Хромопласты — пластиды, содержащие другой пигмент — красного или желтого цвета. Хромопласты придают окраску цветам, осенним листьям. Они создают все разнообразие и красоту растительного мира.
И, наконец, лейкопласты — бесцветные пластиды, они содержатся в клетках луковиц, корней, стеблей и других неокрашенных частей растений. Зато в них накапливаются некоторые органические вещества.
При определенных условиях различные виды пластид могут превращаться друг в друга.
Ядро
Ядро — это хранилище наследственной информации — ДНК, а также главный регулятор синтеза белка. В ядре можно увидеть ядрышки, они исчезают, когда клетка начинает делиться. В ядрышках образуются рибосомы, органеллы, ответственные за синтез белка. ДНК в ядре находится в виде хромосом.
Эндоплазматическая сеть
Эндоплазматическая сеть — это система мембран и цистерн, которые сообщаются с ядром, переходя в его мембрану. Это вызвано общими функциями этих органелл — синтезом белка и других веществ. Выделяют гладкую и шероховатую эндоплазматическую сеть. На мембранах шероховатой эндоплазматической сети находятся рибосомы. Ее функция — синтез белка. Гладкая эндоплазматическая сеть занимается синтезом липидов, участвует в углеводном обмене и др.
Аппарат Гольджи — Это еще одна мембранная органелла клетки. Выглядит Аппарат Гольджи как стопка цистерн, от которых отрываются пузырьки — секреторные гранулы. Аппарат Гольджи связан с эндоплазматической сетью, и занимается секрецией и транспортом белков, ферментов и всего того, что синтезировалось в ней. Образующиеся пузырьки с секретом — лизосомы — могут как перемещаться по клетке, так и выводить секрет наружу, в межклеточное пространство.
Далее осталось еще два вида немембранных органелл — рибосомы и клеточный центр.
Рибосомы — маленькие, но очень важные органеллы, отвечающие за синтез белка . Они находятся в клетке как в свободном состоянии, так и прикрепленными к мембранам шероховатой эндоплазматической сети.
Клеточный центр (или центросома) — имеется не у всех растений. Это немембранная органелла, которая участвует в клеточном цикле — делении клетки, а также в формировании жгутиков и ресничек у некоторых растений.
Итак, мы разобрали основные органеллы растительной клетки, и теперь подведем итоги, за что же они отвечают:
Органелла | Функция в клетке |
| Ядро | Хранение и передача наследственной информации, синтез белка |
| Митохондрия | Выработка энергии для клетки и запасание ее в виде АТФ |
| Пластиды | Хлоропласты — Фотосинтез — образование органических веществ из углекислого газа и воды на светуХромопласты — Придают окраску, привлекательный для насекомых и животных видЛейкопласты — запасают питательные вещества |
| Эндоплазматическая сеть | Синтез белка, жиров и др. веществ |
| Аппарат Гольджи | Транспорт и секреция веществ |
| Рибосомы | Синтез белка |
| Клеточный центр | Участвует в делении клетки, формировании жгутиков и ресничек |
| Вакуоль | Запасает воду, минеральные и органические вещества |
Plastids: определение, структура, типы и функции — видео и стенограмма урока
Типы и функции пластидов
Хлоропласты , вероятно, самые известные из пластид. Они отвечают за фотосинтез . Хлоропласт заполнен тилакоидами, в которых происходит фотосинтез, и хлорофиллом.
Хромопласты — это то, что описывает название, место для хранения и синтеза пигментов в растении. Они содержатся в цветущих растениях, фруктах и стареющих листьях. Хлоропласты фактически превращаются в хромопласты. Здесь есть каротиноидные пигменты, которые позволяют придать разные цвета фруктам и осенним листьям. Одна из основных причин появления этих структур и цветов — привлечение опылителей.
Геронтопласты — это в основном хлоропласты, которые проходят процесс старения. Это хлоропласты листьев, которые начинают превращаться в различные органеллы или перестраиваются, поскольку лист больше не использует фотосинтез (например, в осенние месяцы).
Лейкопласты — это непигментированные органеллы. В отличие от других, о которых мы говорили, лейкопласты вообще не имеют цвета. Они находятся в нефотосинтезирующих частях растения, таких как корни.В зависимости от того, что нужно растению, они могут стать, по сути, просто навесами для хранения крахмала, липидов и белков. Их чаще используют для синтеза аминокислот и жирных кислот.
Лейкопласты подразделяются на три различных пластида:
- Амилопласты
- Протеинопласты
- и Элайопласты
Амилопласты являются самыми большими из трех и отвечают за хранение крахмала.Затем идут протеинопласты , которые помогают накапливать белки, которые необходимы растению и обычно содержатся в семенах. Наконец, элайопласты используются для хранения жиров и масел, которые необходимы растению, особенно в семенах.
Эволюция пластидов
Пластиды возникли из того, что, по мнению ученых, было небольшими прокариотическими организмами, живущими внутри других прокариотических организмов. Они считают, что эти органеллы вместе с митохондриями начали симбиотический или взаимовыгодные отношения с более крупной прокариотической клеткой, прежде чем в конечном итоге стать частью клетки.
Ученые даже смогли определить, что митохондрии и хлоропласты были приблизительными размерами некоторых древних бактерий и даже имели мембраны, похожие на эти бактерии. Эти органеллы размножаются аналогично древним бактериям и несут в своих мембранах некоторые из тех же ферментов.
Короче говоря, ученые признали, что митохондрии и пластиды произошли от древних бактерий, которые находились в симбиотических отношениях с более крупной прокариотической клеткой.В конце концов, эти бактерии превратились в эукариотические клетки с органеллами, которые мы видим и изучаем сегодня. Пластиды эволюционировали вместе с ними.
Резюме урока
Пластиды — это органеллы с двойной мембраной, обнаруженные в растениях. Они произошли от крошечных древних бактерий, которые потреблялись другими крупными прокариотическими клетками. У них развились симбиотические отношения, и по мере продолжения эволюции они превратились в пластиды, существующие в клетках наших эукариотических растений.
Существует четыре основных типа пластид:
- Хлоропласты
- Хромопласты
- Геронтопласты
- и Лейкопласты
Хлоропласты находятся в зеленых частях растения и являются эпицентрами фотосинтеза. Хромопласты встречаются в ярких цветках и плодах растения. Они содержат другие цветные пигменты, предназначенные для привлечения опылителей. Геронтопласты — это стареющие хлоропласты, когда лист отмирает, готовясь к зиме.
Лейкопласты разбиты на три подгруппы:
- Амилопласты
- Протеинопласты
- и Элайопласты
Все находятся в корнях растения. Амилопласты и накапливают крахмал, а протеинопласты и накапливают белок.Элайопласты хранят другие жиры и масла. Пластиды необходимы для функционирования растительных клеток.
Пластиды — Основные термины
- Пластиды : двойные мембраносвязанные органеллы, обнаруженные в растениях
- Хлоропласты : ответственные за фотосинтез
- фотосинтез : процесс, с помощью которого растения дышат, едят и пьют
- Хромопласты : обнаружены в цветковых растениях, плодах и стареющих листьях
- Геронтопласты : стареющие хлоропласты
- Лейкопласты : нефотосинтезирующие части растений, такие как корни, амилопласты, протеинопласты и элайопласты
- Симбиотический : взаимно зависимые отношения
Результаты обучения
После того, как вы запомните приведенные выше концепции и факты, вы сможете:
- Определить пластиды и распознать наиболее распространенные
- Подчеркнуть типы и обязанности пластидов
- Обсудить эволюцию пластид
Пластиды — определение, структура, типы, функции и схема
Главная »Клеточная биология» Пластиды — определение, структура, типы, функции и схемы
Обучающие видео по биологииПоследнее обновление 15 февраля 2020 г. Автор: Sagar Aryal
Пластиды Определение
- Пластид — это органелла с двойной мембраной, участвующая в синтезе и хранении пищи, обычно обнаруживаемая в клетках фотосинтезирующих растений.
- Пластиды были открыты и названы Эрнстом Геккелем, но А. Ф. В. Шимпер был первым, кто дал четкое определение.
- Они необходимы для жизненно важных процессов, таких как фотосинтез и хранение пищи.
- Пластида, содержащая зеленый пигмент (хлорофилл), называется хлоропластом, тогда как пластида, содержащая пигменты, кроме зеленого, называется хромопластом. Пластида, в которой отсутствуют пигменты, называется лейкопластом и участвует в основном в хранении пищи.
Недифференцированный пластид называется пропластидом .Позже он может развиться в любую из других пластид.
A. Хлоропласты
- Хлоропласты, вероятно, являются наиболее известными пластидами.
- Они отвечают за фотосинтез.
- Хлоропласт заполнен тилакоидами, в которых происходит фотосинтез, а хлорофилл остается.
B. Хромопласты
- Хромопласты — это единицы, в которых пигменты хранятся и синтезируются в растении.
- Они содержатся в цветущих растениях, фруктах и стареющих листьях.
- Хлоропласты фактически превращаются в хромопласты.
- Каротиноидные пигменты позволяют добиться различного цвета фруктов и осенних листьев. Одна из основных причин появления этих структур и цветов — привлечение опылителей.
C. Лейкопласты
- Лейкопласты представляют собой непигментированные органеллы.
- Они находятся в нефотосинтезирующих частях растения, таких как корни.
- В зависимости от того, что нужно растению, они могут стать, по сути, просто навесами для хранения крахмала, липидов и белков.
- Их чаще используют для синтеза аминокислот и жирных кислот.
- Лейкопласт может быть амилопластом, накапливающим крахмал, элайопластом, накапливающим жир, или протеинопластом, накапливающим белки.
D. Геронтопласты
- Геронтопласты — это в основном хлоропласты, которые проходят процесс старения.
- Это хлоропласты листьев, которые начинают превращаться в различные органеллы или перестраиваются, поскольку лист больше не использует фотосинтез (например, в осенние месяцы).
В зависимости от своей морфологии и функции пластиды обладают способностью дифференцировать или повторно дифференцироваться между этими и другими формами.
Структура пластидов- Хлоропласты могут быть сферическими, яйцевидными или дискообразными у высших растений и звездчатыми, чашевидными или спиралевидными, как у некоторых водорослей.
- Обычно они имеют диаметр 4-6 мкм и количество в каждой клетке высших растений от 20 до 40, равномерно распределенных по цитоплазме.
- Хлоропласт ограничен двумя липопротеидными мембранами, внешней и внутренней мембранами, с межмембранным пространством между ними.
- Внутренняя мембрана окружает матрикс, строму которого содержат небольшие цилиндрические структуры, называемые грана. Большинство хлоропластов содержат 10-100 гран.
Грана и тилакоиды
- Каждая гранум имеет множество дискообразных перепончатых мешочков, называемых гранными пластинками или тилакоидами (80–120 Å в поперечнике), наложенных друг на друга.
- Граны соединены сетью анастомозирующих канальцев, называемых межгранами или ламеллами стромы.
- Отдельные тилакоиды, называемые тилакоидами стромы, также обнаруживаются в хлоропластах.
- Электронные плотные тела, осмофильные гранулы наряду с рибосомами (70S), кольцевая ДНК, РНК и растворимые ферменты циклов Кальвина также присутствуют в матриксе стромы.
- Таким образом, хлоропласты имеют три разные мембраны: внешнюю, внутреннюю и тилакоидную.
- Тилакоидная мембрана состоит из липопротеина с большим количеством липидов: галактолипидов, сульфолипидов, фосфолипидов.
- Внутренняя поверхность тилакоидной мембраны гранулярна в организации из-за небольших сфероидальных квантосом.
- Квантосомы являются фотосинтетическими единицами и состоят из двух структурно различных фотосистем, ФС I и ФС II, содержащих около 250 молекул хлорофилла. Каждая фотосистема имеет антенные комплексы хлорофилла и один реакционный центр, в котором происходит преобразование энергии.У высших растений присутствуют пигменты хлорофилла-а, хлорофилла-b, каротина и ксантофилла.
- Две фотосистемы и компоненты цепи переноса электронов асимметрично распределены по тилакоидной мембране. Акцепторы электронов как ФС I, так и ФС II находятся на внешней (стромальной) поверхности тилакоидной мембраны. Доноры электронов ФС I находятся на внутренней (тилакоидное пространство) поверхности.
Все клетки растений содержат пластиды в той или иной форме.Эта перекличка указывает на их функциональное разнообразие и демонстрирует, что пластиды лежат в самой основе клеточной функции растений.
- Пластиды — это место производства и хранения важных химических соединений, используемых клетками автотрофных эукариот.
- Тилакоидная мембрана содержит все ферментные компоненты, необходимые для фотосинтеза. Взаимодействие между хлорофиллом, переносчиками электронов, факторами связи и другими компонентами происходит внутри тилакоидной мембраны.Таким образом, тилакоидная мембрана представляет собой специализированную структуру, которая играет ключевую роль в захвате света и транспорте электронов.
- Таким образом, хлоропласты являются центрами синтеза и метаболизма углеводов.
- Они имеют решающее значение не только в фотосинтезе, но и в хранении основных пищевых продуктов, особенно крахмала.
- Его функция во многом зависит от наличия пигментов. Пластида, участвующая в синтезе пищи, обычно содержит пигменты, которые также отвечают за цвет структуры растения (например,грамм. зеленый лист, красный цветок, желтые плоды и т. д.).
- Подобно митохондриям, пластиды имеют собственную ДНК и рибосомы. Следовательно, они могут быть использованы в филогенетических исследованиях.
- Смит, К. М., Маркс, А. Д., Либерман, М. А., Маркс, Д. Б., и Маркс, Д. Б. (2005). Базовая медицинская биохимия Марка: клинический подход. Филадельфия: Липпинкотт Уильямс и Уилкинс.
- http://www.biologydiscussion.com/plastids/plastids-types-structure-and-function-with-diagram/38591
- Koolman, J., & Röhm, K.-H. (2005). Цветной атлас биохимии. Штутгарт: Тиме.
- Альбертс Б. (2004). Существенная клеточная биология. Нью-Йорк, штат Нью-Йорк: научный паб Garland.
- Verma, P. S., & Agrawal, V. K. (2006). Клеточная биология, генетика, молекулярная биология, эволюция и экология (1-е изд.). S.Chand and Company Ltd.
- https://www.biology-online.org/dictionary/Plastid
- https://study.com/academy/lesson/plastids-definition-structure-types-functions.html
- Лигуд Р.С. (2010). Пластидная биология. Анналы ботаники, 106 (1), ix.
- https://www.britannica.com/science/plastid
Пластиды — определение, структура, типы, функции и диаграмма
Категории Клеточная биология Теги Хлоропласты, Хромопласты, Геронтопласты, Лейкопласты, Пластиды, Функции пластидов, Пластиды , Типы пластидов, proplastidPost navigationТипы пластидов: хромопласты и лукопласты
Двумя важными типами пластидов являются следующие:
Пластиды содержат пигменты, которые могут быть разных типов.
Иногда пигменты отсутствуют. По наличию пигментов пластиды бывают двух типов: (i) хромопласты (хроматофоры), содержащие пигмент, и (ii) лейкопласты, которые представляют собой бесцветные пластиды.
Изображение предоставлено: www.ck12.org/ck12/images?id=346038
Хромопласты:
Хромопласты могут быть далее разделены в зависимости от цвета пигмента, и они бывают следующих типов —
1. Хлоропласты:
Наиболее распространенная пластида (гр.Chlor, green, plast и living), который содержит пигменты хлорофилла a и b, а также ДНК и РНК. Хлоропласты находятся в основном в клетках листьев высших растений и водорослей. Это наиболее биологически важная пластида. В процессе фотосинтеза они производят кислород и большую часть химической энергии, используемой живыми организмами.
2. Phaeoplast:
Это желтые или коричневые пластиды, обнаруженные у бурых водорослей, диатомовых водорослей и динофлагеллят. Фукоксантин — это каротиноидный пигмент, который маскирует цвет хлорофилла а, который также присутствует.Он также поглощает свет и передает энергию хлорофиллу а.
3. Родопласты:
Пластиды красного цвета. Он содержится в красных водорослях, а его красный цвет обусловлен фикоэритрином. Он также поглощает свет.
4. Хроматофоры:
Они присутствуют в сине-зеленых водорослях. Термин хроматофор используется вместо пластида, поскольку пигменты не организованы в отдельном пластидном теле, а часто расположены на ламеллярных структурах концентрическими кольцами или пластинами внутри водорослевой клетки.Сине-зеленый цвет этих водорослей обусловлен фикоцианином и фикобилинами. Эти вспомогательные пигменты не участвуют в фотосинтезе.
5. Пурпурные фотосинтезирующие бактерии имеют хлорофилл, бактериохлорофилл. Их фиолетовый цвет обусловлен особыми каротиноидами.
Не фотосинтетические хромопласты:
Также обнаружено множество дополнительных пигментов, которые, по-видимому, не принимают непосредственного участия в фотосинтезе или передаче энергии.
Хромопласты могут развиваться из хлоропластов путем накопления нефотосинтетических пигментов, например.г., красный каротиноид, ликопин в помидорах. Гены синтеза пигментов лежат в ядре.
1. Каротиноиды и ксантофиллы:
Каротин защищает от фотоокисления. Они красно-желтого или коричневого цвета. Они придают окраску цветам и фруктам.
2. Лейкопласты:
Эти пластиды лишены пигмента и представляют собой мембранные структуры. Они служат для хранения крахмала, масел и белков. Это следующие типы —
(а) Амилопласты:
Это ячейки для хранения пищевых продуктов и крахмала.Обычно они содержатся в клубнях, семядолях и эндосперме. Они находятся в областях с низким или нулевым освещением. Амилопласты содержат нуклеоиды и рибосомы.
(б) Элайопласты:
Они содержатся в некоторых однодольных растениях и предназначены для хранения масел.
(c) Протеинопласты (алейронопласты):
Они содержатся в семенах рицинуса и бразильского ореха и хранят белки. Эпидермальные клетки Helleborus также обладают протеинопластами.
Дифференцировка пластид зависит от метаболических потребностей клетки. Хлоропласты могут развиваться из лейкопластов, а хромопласты, которые считаются конечными формами дифференцировки пластид, могут развиваться как из лейкопластов, так и из хлоропластов.
Пропластиды могут дифференцироваться на один из трех типов пластид, и поскольку в некоторых случаях один тип пластид может дифференцироваться в другой, обычно предполагалось, что все пластиды по существу одинаковы по структуре, имея способность дифференцироваться по-разному. , в зависимости от требований ячеек.
Растительная жизнь: хлоропласты и другие пластиды
Пластиды — это узкоспециализированные органеллы с двойной мембраной, обнаруженные в клетках всех растений и водорослей. Тип пластиды, называемый хлоропластом, является клеточным участком процесса фотосинтеза.
Пластиды демонстрируют удивительное разнообразие в отношении своего развития, морфологии, функции, а также физиологической и генетической регуляции. Хлоропласты, тип пластид, вероятно, в значительной степени ответственны за поддержание и сохранение большинства основных форм жизни на Земле посредством фотосинтеза.
Процесс фотосинтеза использует видимый свет в качестве источника энергии для преобразования атмосферного углекислого газа в органические молекулы, которые могут использоваться живыми организмами.
В качестве побочного продукта фотосинтеза кислород выделяется в атмосферу и используется живыми организмами в процессе получения энергии клеточного дыхания. Другие типы пластид специализируются на синтезе и хранении пигментов, крахмала и других вторичных метаболитов.
Пластидная структура
Типичная пластида из клетки цветущего растения окружена двойной мембранной системой, состоящей из внутренней и внешней мембран, с межмембранным пространством между ними.
В хлоропластах фотосинтетические пигменты, отвечающие за поглощение солнечного света, расположены в системе мембран тилакоидов. Эта непрерывная внутренняя мембранная система находится по всей строме хлоропласта, внутренней жидкой матрице, аналогичной клеточному цитозолю.
Гранальные тилакоиды организованы в стопки, а стромальные тилакоиды разложены и выставлены на стромальный матрикс. Внутреннее пространство внутри тилакоидной мембранной системы называется просветом.
Пигменты и белки, участвующие в световых реакциях фотосинтеза, процессах, посредством которых световая энергия преобразуется в химическую энергию, встроены в мембранную систему тилакоидов. В строме происходят темные реакции, или цикл Кальвина, который представляет собой путь фиксации углерода, ведущий к образованию простых углеводов.
Маленькие гранулы крахмала и масляные тельца, называемые пластоглобулами, часто встречаются в хлоропластах. Они служат запасами энергии для растительной клетки.В других пластидах, кроме хлоропластов, тикалоиды обычно отсутствуют.
Пропластиды
Предшественником развития всех типов пластид является пропластид. Пропластиды — это относительно недифференцированные пластиды, обычно обнаруживаемые в молодых недифференцированных меристематических клетках и тканях.
В соответствующих клеточных условиях и условиях окружающей среды пропластиды могут развиваться и дифференцироваться до любого из трех основных типов пластид: хлоропластов, хромопластов или лейкопластов.
Хлоропласты
Хлоропласты обычно содержат один или несколько из трех типов пластидных хлорофиллов (хлорофилл a, b или c) и, часто, члены двух классов фотосинтетических дополнительных пигментов: каротиноиды и фикобилины.
Самым очевидным и важным физиологическим процессом, уникальным для хлоропластов, является фотосинтез. В реакциях преобразования энергии (световых реакциях) лучистая энергия в виде видимого света (в основном с длинами волн фиолетового, синего и красного цветов) используется главным образом зеленым пигментом хлорофиллом.
Собранная энергия затем используется для фосфорилирования аденозиндифосфата (АДФ) с образованием аденозинтрифосфата (АТФ) в процессе, называемом нециклическим фотофосфорилированием, и восстановления электронного носителя никотинамидадениндинуклеотидфосфата (НАДФ) до НАДФН. Кислород высвобождается в результате окислительного расщепления воды в зависимости от света.
В реакциях связывания углерода (часто называемых реакциями темноты, хотя они могут происходить в присутствии света) АТФ используется в качестве источника энергии для присоединения атмосферного углекислого газа к простому сахарному рибулозо-1,5-бисфосфату. (RuBP).НАДФН используется для облегчения восстановления RuBP с помощью ряда простых сахаров в биохимическом наборе реакций, известном как цикл Кальвина.
Один из продуктов этого цикла, глицеральдегид-3-фосфат (G3P), используется хлоропластом для производства глюкозы и других углеводов. G3P также необходим для продолжения цикла Кальвина, поэтому только один из каждых трех произведенных организмов используется для синтеза углеводов.
| Хлоропласты в растении |
Хлоропласты также являются местом синтеза трех ароматических аминокислот: фенилаланина, тирозина и триптофана.
Соединение-предшественник аспартата импортируется в хлоропласты из цитозоля клетки и используется для синтеза аминокислот лизина, треонина и изолейцина. Промежуточный продукт синтеза треонина, называемый гомосерин-4-фосфатом, экспортируется из хлоропласта в цитозоль в качестве предшественника метионина.
Таким образом, существует сильная интеграция функций между хлоропластом, цитозолем и ядром, поскольку ферменты, участвующие в этих путях биосинтеза аминокислот, кодируются ядром, их мРНК транслируются с использованием цитозольных рибосом, а большинство ферментов биосинтеза импортируются в хлоропласт.
Биосинтез жирных кислот — еще одна биохимическая функция, которая происходит в хлоропластах. Жирные кислоты, как предшественники липидов, могут либо непосредственно включаться в липиды хлоропластов посредством биохимических путей, локализованных в пластидах, либо экспортироваться в цитоплазму для преобразования в липиды эндоплазматического ретикулума.
Липиды, обнаруженные во внутренней пластидной мембране, синтезируются пластидами, тогда как липиды внешней пластидной мембраны синтезируются в эндоплазматическом ретикулуме.
Другие пластиды
Другие типы пластид включают хромопласты, которые обычно содержат пигменты каротин или ксантофилл и отвечают за окраску многих фруктов, цветов и корней.При некоторых условиях хромопласты могут дифференцироваться в хлоропласты. Лейкопласты бесцветны и лишены сложных внутренних мембран.
Один из типов лейкопластов, амилопласт, синтезирует и хранит крахмал. Другие лейкопласты синтезируют широкий спектр продуктов, включая масла и белки. Пропластиды, которые задерживаются во время своего нормального развития в хлоропласты, называются этиопластами. Обычно они образуются, когда развивающиеся ткани растений лишены света.
История эволюции
Пластиды обладают рядом особенностей, которые позволяют понять их замечательную историю эволюции.Хлоропласты эукариотических клеток фотосинтезируют аналогично более древним прокариотическим цианобактериям, используя связанный с мембраной хлорофилл для захвата лучистой энергии.
Некоторые пластиды даже имеют сильное морфологическое сходство с цианобактериями, будучи схожими по размеру и имеющими схожую внутреннюю структуру. Пластиды делятся на две части аналогично размножению бактерий.
Пластиды также обладают определенной степенью автономии с точки зрения их генетической системы.Как правило, большинство пластид цветковых растений содержат множественные копии (50-100) кольцевой хромосомы, размер которых варьируется от 130 до 180 пар оснований (kb) у высших растений. Размер хромосом у водорослей гораздо более вариабельный и колеблется от 57 кб до 1500 кб.
Пластидная хромосома содержит гены РНК, такие как рРНК (рибосомная РНК) и тРНК (транспортная РНК), а также структурные гены, которые кодируют полипептиды, участвующие в фотосинтезе, транскрипции, синтезе белка, трансдукции энергии и некоторых других функциях.Многие гены на хромосоме хлоропласта организованы в кластеры, называемые оперонами, подобно тому, как это происходит у эубактерий.
Нуклеотидные последовательности многих пластидных генов, особенно генов рибосомной РНК, очень похожи на таковые у эубактерий, а рибосомы, обнаруженные в пластидах, имеют такой же состав и размер, что и эубактериальные рибосомы.
Кодируемые пластидом гены транскрибируются либо кодируемой ядром, либо кодируемой пластидом РНК-полимеразой, и полученные мРНК транслируются пластидными рибосомами, обнаруженными в строме.Большинство биохимических процессов в пластиде зависит как от ядер, так и от кодируемых пластидом генов.
Некоторые белки, такие как карбоксилаза / оксигеназа RuBP (Rubisco), состоят из белковых субъединиц, кодируемых как ядром, так и пластидом, что снова демонстрирует замечательную координацию биогенеза и развития между органеллой и цитозолем.
Это свидетельство убедительно подтверждает эндосимбиотическую теорию происхождения пластид. Эта теория, по сути, утверждает, что пластиды когда-то были свободноживущими, автотрофными (имеющими способность получать углерод из углекислого газа) прокариотическими клетками, которые были поглощены фагоцитозом наследственной гетеротрофной ядросодержащей клеткой (клетка, метаболизм которой требует углерода быть полученным из органических молекул), называемый протоэукариотом.
Обычно это поглощение приводит к проглатыванию и последующему разрушению поглощенной прокариотической клетки. Однако в одном — или, возможно, нескольких — независимых инцидентах постепенно установились симбиотические отношения между поглощенной фотосинтезирующей бактерией и протоэукариотом.
Захваченная бактерия обеспечивала внутренний источник производства пищи для гетеротрофной эукариотической клетки посредством фотосинтеза. Бактерии, в свою очередь, были обеспечены защита и стабильная внешняя среда в цитозоле эукариотической клетки.
Для дальнейшей координации физиологических и генетических взаимодействий между ними со временем произошел массовый перенос генов из генома фотосинтетической бактерии в ядерный геном протоэукариота, что привело к генетическому захвату и контролю фотосинтетического эндосимбионта.
Недавние исследования показали, что это событие переноса гена — непрерывный процесс, с примерами переноса, задокументированными в недавнее эволюционное время как для пластид, так и для митохондрий в нескольких различных эволюционных линиях цветковых растений.
ПЛАСТИДЫ
ПЛАСТИНЫПЛАСТИДЫ
Пластиды крупные цитоплазматические органеллы. Пластиды — это основные органеллы, обнаруженные в клетках растения и водоросли. Пластиды — это место производства и хранения важных химические соединения, используемые клеткой. Пластиды часто содержат пигменты, используемые в фотосинтез, и типы присутствующих пигментов могут изменить или определить цвет ячейки. Термин пластида произошел от греческого слова пластикас. значение сформировано или сформировано.Этот термин был введен Шимпером в 1885 году.
У растений пластиды могут дифференцируются на несколько форм, в зависимости от того, какую функцию они должны выполнять. играть в клетке. Пластиды можно разделить на два основных типа. а именно хромопласты и лейкопласты.
Хромопласт:
Это цветные пластиды. (хромо = цвет; пластик = живая).Они содержат различные пигменты. Они синтезируют пищевые материалы путем фотосинтеза. Они содержат желтый, оранжевый или красный пигменты. Хромопласты обычно встречаются в цветах и фруктах. Хромопласты также делятся на три типа в зависимости от цвета, а именно хлоропласты, феопласт и родопласт.
Хлоропласт: зеленый цвет. Он содержит пигменты хлорофилла. Он содержится в высших растениях и зеленые водоросли.
Phaeoplast: Темно-коричневый в цвет.Он содержит пигменты фукоксантина. Встречается в бурых водорослях, диатомовых водорослях. и динофлагелаты.
Родопласт: Красный в цвет. Он содержит фикоэритрин. Он содержится в красных водорослях.
Лейкопластов:
Они — это непигментированные пластиды (Leuco = белый; plast = живой). Их основная функция — хранить продукты материалы. Они не занимаются синтетической деятельностью.Лейкопласты подразделяется на три типа, а именно амлиопласт, элайопласт и протеинопласт.
Амлилопласт: накапливает крахмал и содержится в клубнях, семядолях и эндосперме
Элайопласт: хранит масло и обнаруживается в клетках эпидермиса.
Протеинопласт: хранит белок и содержится в семенах и орехах.
ХЛОРОПЛАСТЫ
Хлоропласты — органеллы содержится в клетках растений и других эукариотических организмах, которые проводят фотосинтез.Слово хлоропласт происходит от греческого слова chloros , что означает зеленый, и пластик, , что означает форма или сущность. Хлоропласты являются членами класса органелл, известных как пластиды.
Форма:
Хлоропласт различается по форме. У высших растений они имеют сфероидальную, яйцевидную или дискообразную форму. Они имеют чашевидную форму в хламидомонады и спиралевидно свернутой в спирогиру.
Размер:
Размер пластид варьируется. от вида к виду. Но размер остается постоянным для данного типа ячеек. У высших растений он составляет 4-5 мкм в длину и 1-3 мкм в толщину. Обычно хролопласты растений, произрастающих в тени, больше по размеру.
Номер:
Количество хлоропластов варьируется от растения к растению, но остается неизменным для данного растения.В у высших растений имеется от 20 до 40 хлоропластов на клетку или до 1000 хлоропласты.
Состав:
Хлоропласты растений крупные органеллы (от 5 до 10 мкм длиной м), которые, как и митохондрии, ограничены двойная мембрана, называемая оболочкой хлоропласта. Помимо внутреннего и внешние мембраны оболочки, хлоропласты имеют третью внутреннюю мембрану система, называемая тилакоидной мембраной.Тилакоидная мембрана образует сеть из уплощенные диски, называемые тилакоидами, которые часто собираются в стопки называется грана. Грана соединены между собой разветвляющимися мембранными канальцами, которые называются лады (стромальные ламели). Благодаря этой трехмембранной структуре внутренняя организация хлоропластов сложнее, чем митохондрий. В частности, их три мембраны делят хлоропласты на три отдельных внутренние отсеки: (1) межмембранное пространство между двумя мембранами оболочка хлоропласта; (2) строма, которая находится внутри оболочки, но вне тилакоидной мембраны; и (3) просвет тилакоида.
Тилакоид имеет уплощенный форма диска. Внутри него находится пустое пространство, называемое тилакоидным пространством или просветом. Фотосинтез происходит на тилакоидной мембране; как в митохондриях окислительное фосфорилирование, оно включает взаимодействие межмембранных потоков с биосинтезом через диссипацию протонного электрохимического градиента. В тилакоидную мембрану встроены антенные комплексы, каждый из которых состоит светопоглощающих пигментов, включая хлорофилл и каротиноиды, а также как белки, связывающие пигменты.Эти комплексы называются квантосомами. Этот комплекс как увеличивает площадь поверхности для захвата света, так и позволяет захват фотонов с более широким диапазоном длин волн. Энергия инцидента фотоны поглощаются пигментами и направляются в реакционный центр этого сложные за счет резонансной передачи энергии. Тогда две молекулы хлорофилла ионизируется, образуя возбужденный электрон, который затем переходит на фотохимический центр реакции.
Хлоропласт содержит белки, липиды, углеводы, ДНК, РНК, каротиноиды, хлорофилл и минералы. Состав этих химикатов указан в следующей таблице:
S. № | Химическая промышленность | В процентах |
1. | Белки | 35 55% |
2. | Липиды | 20 30% |
3. | Углеводы | Переменная |
4. | Хлорофилл | 9% |
5. | Каротиноиды | 4,5% |
6. | РНК | 3 4% |
7. | ДНК | 0,5% |
8. | Минералы | 0,2% |
Фотосинтетические пигменты:
Производство пигментов фотосинтез называют фотосинтетическими пигментами. Хлоропласт также содержит фотосинтетические пигменты. Это хлорофиллы, каротиноиды и фикобилины. Хлорофилл а хорошо поглощает на длине волны около 400-450 нм и на длине волны 650-700 нм. нм; хлорофилл b при 450-500 нм и 600-650 нм.Ксантофилл хорошо всасывается при 400-530 нм. Однако ни один из пигментов хорошо не впитывается в желто-зеленый цвет. регион, который отвечает за обильную зелень, которую мы видим в природе.
Хлорофиллы ар зеленоватые пигменты, содержащие порфириновое кольцо. Это стабильный кольцеобразный молекула, вокруг которой могут свободно перемещаться электроны. Потому что электроны движутся свободно, кольцо может легко получать или терять электроны, и, таким образом, потенциал для передачи заряженных электронов другим молекулам.Это фундаментальный процесс, с помощью которого хлорофилл «улавливает» энергию солнечного света.
Существует несколько видов хлорофилла, самые важен хлорофилл «а». Это молекула, которая производит фотосинтез. возможно, путем передачи заряженных электронов молекулам, которые будут производить сахар. Все растения, водоросли и цианобактерии, которые фотосинтезируют содержат хлорофилл «а». Второй вид хлорофилла — это хлорофилл «b», который встречается только в «зеленых водорослях» и в растениях.Третья форма хлорофилла который является обычным (что неудивительно) называется хлорофиллом «с» и встречается только у фотосинтезирующих представителей Chromista, а также у динофлагеллят. Различия между хлорофиллами этих основных групп были одним из первые признаки того, что они не были так тесно связаны, как считалось ранее.
Каротиноиды обычно красные, оранжевые или желтые пигменты и включают знакомый составной каротин, который придает цвет моркови.Эти соединения состоят из двух небольших шестиуглеродные кольца, соединенные «цепочкой» атомов углерода. В результате они делают не растворяются в воде и должны прикрепляться к мембранам внутри клетки. Каротиноиды не могут передавать энергию солнечного света непосредственно фотосинтетикам. пути, но должны передавать свою поглощенную энергию хлорофиллу. По этой причине, их называют вспомогательными пигментами. Один очень заметный дополнительный пигмент — это фукоксантин — коричневый пигмент, который окрашивает ламинарии и другие коричневые водоросли как диатомовые водоросли.
Phycobilins являются водорастворимые пигменты, поэтому они обнаруживаются в цитоплазме или в строма хлоропласта. Встречаются только у Cyanobacteria и Rhodophyta. В на рисунке справа показаны два класса фикобилинов, которые могут быть извлечены из этих «водорослей». Флакон слева содержит голубоватый пигмент фикоцианин, что и дало название цианобактериям. Флакон справа содержит красноватый пигмент фикоэритрин, который дал красным водорослям их общее название.
Фикобилины полезны не только для организмов. которые используют их для впитывания световой энергии; они также нашли применение в качестве исследования инструменты. И пикоцианин, и фикоэритрин флуоресцируют на определенной длине волны. То есть, когда они подвергаются воздействию яркого света, они поглощают световую энергию, и высвободить его, испуская свет очень узкого диапазона длин волн. В свет, производимый этой флуоресценцией, настолько отличительный и надежный, что фикобилины могут использоваться как химические «метки».Пигменты химически связаны к антителам, которые затем помещаются в раствор клеток. Когда решение распыляется потоком мелких капель мимо лазерного и компьютерного датчика, машины может определить, были ли клетки в каплях «помечены» антитела. Это нашло широкое применение в исследованиях рака, для «маркировки» опухоли. клетки.
Транспортировка хлоропластов:
Большинство белков хлоропластов синтезируются на свободных рибосомах в цитозоле и предназначены для импорта в хлоропласты амино-концевыми транзитными пептидами.Белки, входящие в состав просвет тилакоидов сначала импортируется в строму хлоропласта, а затем нацелен для транспорта через тилакоидную мембрану вторым гидрофобным сигналом последовательность.
ФУНКЦИИ ХЛОРОПЛАСТА
Хлоропласт выполняет следующие функции:
1. Синтез белка:
Поскольку хлоропласт содержит собственный ДНК и рибосмес, он синтезирует определенное количество белков для своего метаболизм.
2. Startch Storage:
Хлоропласты временно хранятся крахмал в дневное время в пиреноиде, который представляет собой органеллы, образующие крахмал. Ночью крахмал переносится в районы роста и хранения.
4. Подача кислорода:
Хлоропласты используют углекислый газ для фотосинтеза и выделения кислорода. Этот кислород используют все животные и растения для дыхания.Выделение кислорода происходит во время фотофосфорилирование световых реакций. При легкой реакции вода литический центр или центр выделения кислорода, высвобождает кислород, разрушая воду молекулы в присутствии света фотосистемой-II.
5. Фотосинтез:
Фотосинтез (от греч. фото , «свет» и синтез , «соединение») — это процесс. который преобразует углекислый газ в органические соединения, особенно сахара, используя энергия солнечного света.Фотосинтез происходит у растений, водорослей и многих других виды бактерий, но не у архей. Фотосинтезирующие организмы называются . photoautotrophs , поскольку он позволяет им создавать свою собственную пищу. У растений, фотосинтез водорослей и цианобактерий использует углекислый газ и воду, высвобождая кислород как отходы. Фотосинтез жизненно необходим для жизни на Земле.
2n CO 2 + 2n H 2 O + фотоны → 2 (CH 2 O) n + 2н О 2
Диоксид углерода + вода + световая энергия → углевод + кислород
6.Фотодыхание:
Фотодыхание — это процесс когда рибулозобифосфаткарбоксилаза / оксигеназа (RUBisco) вместо этого фиксируют кислород диоксида углерода. Фотодыхание может возникнуть, когда уровень углекислого газа низкий. Фотодыхание не производит АТФ (энергию для клеток) и приводит к чистому потеря углерода и азота (в виде аммиака), что замедляет рост растений. Фотодыхание считается эволюционным реликтом. Снижается фотодыхание эффективность фотосинтеза за счет удаления молекул углерода из кальвина Цикл.
При фотодыхании фермент вовлеченных (рибулозобисфосфаткарбоксилаза или рубиско) иногда катализирует добавление O 2 вместо CO 2 с образованием одной молекулы 3-фосфоглицерат и одна молекула фосфогликолата (два атома углерода). Это побочная реакция, и фосфогликолят не является полезным метаболитом. Это первый превращается в гликолят, а затем переносится в пероксисомы, где окисляется и превращается в глицин.Затем глицин переносится в митохондрии, где два молекулы глицина превращаются в одну молекулу серина с потерей CO 2 и NH 3 . Затем серин возвращается в пероксисомы, где он превращается в глицерат. Наконец, глицерат переносится обратно. в хлоропласты, где он снова входит в цикл Кальвина. Фотодыхание не делает оказываются полезными для растения, так как это, по сути, противоположность фотосинтез O 2 потребляется, а CO 2 выделяется без любой прирост АТФ.Однако периодическое использование O 2 на месте CO 2 , по-видимому, является неотъемлемым свойством rubisco, поэтому фотодыхание — общее сопровождение фотосинтеза.
ФОТОСИНТЕЗ
Фотосинтез проходит в хлоропласты зеленых растений и эукариотических водорослей, а также фотосинтезирующие мембраны у прокариот.Процесс фотосинтеза происходит в двух шаги. Это Светлая реакция Темная реакция. В легких реакциях, энергия солнечного света стимулирует синтез АТФ и НАДФН, связанных с образование O 2 из H 2 O. В темноте реакции, названные так поскольку им не нужен солнечный свет, АТФ и НАДФН, производимые светом реакции приводят к синтезу глюкозы. В эукариотических клетках как светлые, так и темные реакции фотосинтеза происходят внутри хлоропластов; световые реакции в тилакоидная мембрана и темные реакции в строме.
СВЕТОВАЯ РЕАКЦИЯ:
В световых реакциях одна Молекула пигмента хлорофилла поглощает один фотон и теряет один электрон. Это происходит только при наличии света, отсюда и название световой реакции. Во время световой реакции световая энергия (фотон) преобразуется в химическую энергию. (АТФ). В световых реакциях есть две стадии: фотолиз и фотофосфорилирование. При фотолизе вода расщепляется на электроны, водород и кислород.При фотофосфорилировании синтезируется АТФ. Фотофосфорилирование происходит двумя путями, а именно нециклическим фотофосфорилированием и циклическим фотофосфорилирование.
Нециклическое фотофосфорилирование:
Путь электронного потока начинается в фотосистема II, которая гомологична фотосинтетическому реакционному центру R . viridis уже описано. Однако в фотосистеме II энергия полученный в результате поглощения фотонов, используется для разделения молекул воды на молекулярные кислород и протоны.Реакция протекает в просвете тилакоида, поэтому высвобождение протонов из H 2 O устанавливает протонный градиент поперек тилакоидная мембрана. Электроны высоких энергий, полученные в результате этого процесса: переносится через ряд носителей в пластохинон, жирорастворимый переносчик, аналогичный коферменту Q (убихинону) митохондрий. Пластохинон переносит электроны из фотосистемы II в комплекс цитохрома bf , в котором электроны переходят на пластоцианин и дополнительные протоны закачиваются в просвет тилакоида.Электронный транспорт через фотосистему II таким образом, связано с установлением протонного градиента, который приводит в движение хемиосмотический синтез АТФ.
Из фотосистемы II электроны уносятся пластоцианин (белок периферической мембраны) в фотосистему I, где поглощение дополнительных фотонов снова генерирует электроны высокой энергии. Фотосистема I, однако, не действует как протонный насос; вместо этого он использует эти электроны высокой энергии для восстановления NADP + до NADPH.Центр реакции хлорофилл фотосистемы I переносит возбужденные электроны через серию переносчики ферродоксина, небольшого белка на стромальной стороне тилакоида мембрана. Затем фермент НАДФ-редуктаза переносит электроны от ферродоксина к НАДФ + , образуя НАДФН. Прохождение электронов через фотосистемы I и II, таким образом, генерируют как АТФ, так и НАДФН, которые используются Ферменты цикла Кальвина в строме хлоропластов превращают CO 2 в углеводы.Электроны перемещаются по цепи переноса электронов, как Буква Z, так что процесс, называемый Z-схемой, изначально функционирует для генерируют хемиосмотический потенциал через мембрану.
Циклическое фотофосфорилирование:
Второй путь переноса электронов, называемый циклический поток электронов, производит АТФ без синтеза НАДФН, тем самым снабжение дополнительным АТФ для других метаболических процессов. В циклическом электронном потоке световая энергия, собранная в фотосистеме I, используется для синтеза АТФ, а не Синтез НАДФН.Вместо перевода на NADP + , высокоэнергетический электроны из фотосистемы I переносятся на цитохром bf сложный. Затем осуществляется перенос электрона через комплекс цитохрома bf . связаны, как в фотосистеме II, с установлением протонного градиента поперек тилакоидная мембрана. Затем пластоцианин возвращает эти электроны в фотосистему. I в состоянии с более низкой энергией, завершая цикл переноса электронов, в котором световая энергия, собранная в фотосистеме I, используется для накачки протонов в цитохром bf комплекс.Перенос электронов из фотосистемы я могу таким образом генерируют либо АТФ, либо НАДФН, в зависимости от метаболических потребностей клетки.
Синтез АТФ:
АТФ-синтаза тилакоидной мембраны похож на митохондриальный фермент. Однако энергия, запасенная в протоне градиент через тилакоидную мембрану, в отличие от внутренней митохондриальной мембрана, почти полностью химическая по своей природе. Это потому, что тилакоид мембрана хоть и непроницаема для протонов, но отличается от внутренней митохондриальной мембрана проницаема для других ионов, особенно Mg 2+ и Cl —.Свободный проход этих ионов нейтрализует составляющую напряжения протона. градиент, поэтому энергия, полученная в результате фотосинтеза, сохраняется в основном как разница в концентрации протонов (pH) на тилакоидной мембране. Тем не мение, поскольку просвет тилакоида представляет собой закрытый отсек, эта разница в протонном концентрация может быть довольно большой, что соответствует разнице более три единицы pH между стромой и просветом тилакоида. Из-за величина этой разницы pH, полная свободная энергия, запасенная через тилакоидная мембрана похожа на мембрану внутренней митохондриальной мембрана.
На каждую переносимую пару электронов по два протоны переносятся через тилакоидную мембрану в фотосистеме II и двух до четырех протонов в комплексе цитохрома bf . Поскольку четыре протона необходим для управления синтезом одной молекулы АТФ, прохождение каждой пары электронов через фотосистемы I и II нециклическим потоком электронов дает от 1 до 1,5 молекул АТФ. Циклический поток электронов имеет меньший выход, соответствует между 0.5 и 1 молекула АТФ на пару электронов.
ТЕМНЫЕ РЕАКЦИИ:
Темные реакции или Световые независимые реакции, превращает корбондиоксид в углевод с помощью АТФ и НАДФН. Этот процесс также называется фиксацией углерода или углеродной реакцией, поскольку углерод фиксируется во время эта реакция. Встречается в строме хлоропласта. Есть три пути происходит при темновой реакции, а именно циле Кальвина, пути C 4 и пути CAM.
Цикл Кальвина или путь C-3:
Путь углерода в темновой реакции отработан Мелвин Кэлвин.Отсюда его название — цикл Кальвина. Как три углерода соединение 3-фосфоглицерат является первым стабильным продуктом в этом цикле, это обычно называемый C-3 путь. В этом цикле каждая молекула CO 2 является добавлен к рибулозо-1,5-бисфосфату, чтобы получить две молекулы 3-фосфоглицерата. Таким образом, шесть молекул CO 2 приводят к образованию 12 молекул 3-фосфоглицерат, которые превращаются в 12 молекул глицеральдегид-3-фосфат ценой по 12 молекул АТФ и НАДФН.Затем две молекулы глицеральдегид-3-фосфата используются для синтеза глюкоза и десять молекул продолжают цикл Кальвина, чтобы сформировать шесть молекул рибулозо-5-фосфат. Затем цикл завершается использованием шести дополнительных Молекулы АТФ для синтеза рибулозо-1,5-бисфосфата.
C-4 Pathway или Hatch-Slack pathway:
В жарких и засушливых условиях растения закроются. их устьица, чтобы предотвратить потерю воды.В этих условиях CO 2 уменьшится, и газообразный кислород, образующийся в результате световых реакций фотосинтез, уменьшится в стебле, а не в листьях, что приведет к увеличению фотодыхание за счет оксигеназной активности рибулозо-1,5-бисфосфата карбоксилаза / оксигеназа и снижение фиксации углерода. Некоторые растения эволюционировали механизмы повышения концентрации CO 2 в листьях под эти условия.
Название «C 4 » происходит от того факта, что первый продукт фиксации CO 2 в этих растениях имеет четыре углерода атомы, а именно оксалоацетат.Путь C 4 был открыт M.D. Хэтча и К. Р. Слэка в Австралии в 1966 году, поэтому его иногда называют Путь Хетча-Слэка. C 4 растения химически фиксируют углекислый газ в клетки мезофилла, добавив его к трехуглеродной молекуле фосфоенолпируват (PEP), реакция, катализируемая ферментом PEP карбоксилазы и которая создает четырехуглеродную органическую кислоту, щавелевоуксусную кислоту. Щавелевоуксусная кислота или малат, синтезированные этим способом, затем перемещаются в специализированные клетки оболочки пучка, где фермент, рубиско и другие клетки Кальвина ферменты цикла расположены, и где CO 2 высвобождается декарбоксилированием четырехуглеродных кислот затем связывается рубисковой активностью с трехуглеродными сахар 3-фосфоглицериновой кислоты.Физическое отделение рубиско от световые реакции, генерирующие кислород, уменьшают фотодыхание и увеличивают CO 2 фиксация и, следовательно, фотосинтетическая способность листа. C 4 растения могут производят больше сахара, чем растения C 3 в условиях высокой освещенности и температура. Многие важные культурные растения — это растения C 4 , включая кукуруза, сорго, сахарный тростник и просо.
Есть несколько вариантов этого пути:
- 4-углеродная кислота, переносимая из клеток мезофилла, может быть малатом, как указано выше, или может быть аспартатом.
- 3-углеродная кислота, перенесенная обратно из пучка-оболочки. клетки могут быть пируватом, как указано выше, или аланином.
- Фермент, катализирующий декарбоксилирование в пучково-оболочковые клетки различаются. В кукурузе и сахарном тростнике фермент НАДФ-яблочный. фермент, у проса это НАД-яблочный фермент, а у паникума максимум — PEP карбоксикиназа.
Метаболизм CAM или крассуловой кислоты:
Ксерофиты, такие как кактусы и большинство суккулентов, также использовать карбоксилазу PEP для захвата диоксида углерода в процессе, называемом Crassulacean кислотный метаболизм (CAM).В отличие от метаболизма C4, который физически отделяет фиксацию CO 2 на PEP от цикла Кальвина, только CAM временно разделяет эти два процесса. У растений САМ другой лист анатомии, чем растения C 4 , и зафиксируйте CO 2 ночью, когда их устьица открыты. Заводы CAM хранят CO 2 в основном в форме яблочной кислоты путем карбоксилирования фосфоенолпирувата до оксалоацетата, который является затем уменьшили до малата.Декарбоксилирование малата в течение дня высвобождает CO 2 внутри листьев, что позволяет углероду связывать 3-фосфоглицерат рубиско. Путь CAM позволяет устьицам оставаться закрытыми в течение дня; поэтому это особенно часто встречается у растений, адаптированных к засушливым условиям.
Ночью
CAM растения открывают устьица во время охлаждения и более влажные ночные часы, позволяющие поглощать углекислый газ с минимальная потеря воды.
Углекислый газ превращается в растворимый молекулы, которые могут быть легко сохранены растением в разумной концентрации.
Химический путь включает трехуглеродный соединение фосфоенолпируват (PEP), к которому добавлена молекула CO 2 . При этом образуется новая молекула оксалоацетат, которая, в свою очередь, образует малат. Оксалоацетат и малат построены вокруг скелета из четырех атомов углерода, поэтому термин C 4 . Малат может легко храниться растением в вакуолях внутри отдельные клетки.
Днем
Малат может быть разбит по требованию, высвободив молекула CO 2 , поскольку она превращается в пируват. Пируват может быть фосфорилированы (т.е. имеют фосфатную группу, добавленную «энергоносителем» АТФ) для регенерирования PEP, с которого растение было запущено, готового к введению в действие на следующую ночь. Но именно выпуск CO 2 делает цикл стоит растения. Направлен на строму хлоропластов: сайты, на которых фотосинтез наиболее активен.Там он предоставляется RuBisCO в большей концентрации, увеличивая эффективность молекулы, и следовательно, производство большего количества сахаров на единицу фотосинтеза.
МИТОХОНДРИЯ
Слово митохондрия из греческого mitos , нить + chondrion , гранула. В камере биологии, митохондрия (множественное число митохондрий) представляет собой органеллу, заключенную в мембрану обнаружен в большинстве эукариотических клеток.Митохондрии иногда описывают как «сотовые электростанции», потому что они производят большую часть запаса клеточной энергии аденозинтрифосфат (АТФ), используемый в качестве источника химической энергии. В митохондии были впервые открыты Флеммингом и Колликером. Период, термин Митохондрии были представлены Бендой.
Номер:
Количество митохондрий в клетка широко варьируется в зависимости от организма и типа ткани.Многие ячейки имеют только одну митохондрии, тогда как другие могут содержать несколько тысяч митохондрий.
Размер:
Размер митохондрий составляет сильно изменчивый. Эти органеллы варьируются от 0,510 микрометров (мкм) в диаметр. Наименьшие митохондрии наблюдаются у дрожжей (1 мкм). Самый большой митохондрии, обнаруженные в ооцитах земноводных (20–40 мкм).
Распределение:
В большинстве клеток митохондрии равномерно распределяются по цитоплазме.Но в некоторых случаях они собраны вокруг ядра. В Парамециуме они расположены прямо под поверхность клеток. В канальцах почек они располагаются в складках базальных областей. возле плазматической мембраны.
Форма:
Митохондрии могут быть по форме нитевидные или зернистые. Форма митохондрий может меняться от одной к другому в зависимости от физиологических условий клетки.Они могут быть палочковидная, булавовидная, кольцеобразная, округлая или везикулярная.
Состав:
Митохонстин содержит внешнюю и внутренние мембраны, состоящие из фосфолипидных бислоев и белков. Два мембраны, однако, обладают другими свойствами. Из-за этого двухмембранного В митохондрии есть пять отдельных отделов. Есть внешняя митохондриальная мембрана, межмембранное пространство (пространство между внешней и внутренней мембранами), внутренняя митохондриальная мембрана, пространство крист (образовано складками внутренней мембраны) и матрикс (пространство внутри внутренней мембраны).
Внешний митохондриальный мембрана, которая охватывает всю органеллу, имеет белок-фосфолипид соотношение аналогично плазматической мембране эукариот (около 1: 1 по весу). Он содержит большое количество интегральных белков, называемых поринами. Эти порины образуют каналы, которые позволяют молекулам с молекулярной массой 5000 Дальтон или меньше свободно перемещаться распространяются с одной стороны мембраны на другую.
Межмембранное пространство пространство между внешней мембраной и внутренней мембраной.Потому что внешний мембрана свободно проницаема для малых молекул, концентрации малых молекулы, такие как ионы и сахара в межмембранном пространстве, такие же, как цитозоль. Однако, поскольку большие белки должны иметь специфическую сигнальную последовательность для транспортироваться через внешнюю мембрану, белковый состав этого пространства отличается от белкового состава цитозоля. Один белок, который Таким образом, в межмембранном пространстве локализован цитохром с.
Содержит более 100 различных полипептидов и имеет очень высокое соотношение белка к фосфолипидам (более чем 3: 1 по массе, что составляет примерно 1 белок на 15 фосфолипидов). Внутренний В мембране содержится около 1/5 всего белка митохондрии. В Кроме того, внутренняя мембрана богата необычным фосфолипидом — кардиолипином. Кардиолипин содержит четыре жирных кислоты, а не две, и может помочь внутренняя мембрана непроницаема.В отличие от внешней мембраны, внутренняя мембрана делает не содержит поринов и очень непроницаем для всех молекул. Почти все ионы а молекулы требуют специальных мембранных транспортеров для входа в матрицу или выхода из нее. Белки переносятся в матрикс через транслоказу внутренней мембраны. (TIM) или через Oxa1. Кроме того, существует мембранный потенциал через внутренняя мембрана, образованная действием ферментов переноса электронов цепь.
Внутренняя митохондрия мембрана разделена на многочисленные кристы, которые расширяют поверхность область внутренней митохондриальной мембраны, повышая ее способность производить АТФ.Это не просто случайные складки, а инвагинации внутренних мембрана, которая может влиять на общую хемиосмотическую функцию. В типичной печени митохондрии, например, площадь поверхности, включая кристы, составляет около пяти раз больше внешней мембраны. Митохондрии клеток, пользующихся повышенным спросом для АТФ, таких как мышечные клетки, содержат больше крист, чем типичная печень Эти складки усеяны небольшими круглыми тельцами, известными как F 1 . частицы или оксисомы.
Матрица пробел заключен внутренней мембраной. Он содержит около 2/3 общего белка в митохондрия. Матрица важна для производства АТФ с помощью АТФ-синтаза, содержащаяся во внутренней мембране. Матрица содержит высококонцентрированная смесь сотен ферментов, особая митохондриальная рибосомы, тРНК и несколько копий генома митохондриальной ДНК. Из ферменты, основные функции которых включают окисление пирувата и жирных кислот, а также цикл лимонной кислоты.Митохондрии имеют собственный генетический материал, а оборудование для производства собственных РНК и белков. Опубликованный человек Последовательность митохондриальной ДНК выявила 16 569 пар оснований, кодирующих всего 37 генов: 22 тРНК, 2 рРНК и 13 пептидных генов. 13 митохондриальных пептидов у человека интегрированы во внутреннюю митохондриальную мембрану вместе с белками кодируется генами, находящимися в ядре клетки-хозяина.
Митохондрии обнаруживают содержат от 65 до 75% белка, от 25 до 30% липидов, 0.5% РНК и небольшое количество ДНК. Липидная часть митохондрий состоит из 90% фосфолипидов, 5% холестерина. и 5% свободных жирных кислот. Небольшое количество серы, железа, меди и некоторых витамины присутствуют. В составе более 70 ферментов и коферментов. митохондрии.
ФУНКЦИИ МИТОХОНДРИИ:
Энергоснабжение:
Доминирующая роль митохондрии — это производство АТФ, о чем свидетельствует большое количество белки внутренней мембраны для этой задачи.Это делается путем окисления основные продукты глюкозы, пирувата и НАДН, которые производятся в цитозоле.
Клеточное дыхание:
Этот процесс сотовой дыхание, также известное как аэробное дыхание, зависит от наличия кислород. Когда кислород ограничен, гликолитические продукты будут метаболизироваться анаэробное дыхание — процесс, не зависящий от митохондрий.В производство АТФ из глюкозы дает примерно в 13 раз больший выход во время аэробное дыхание по сравнению с анаэробным дыханием.
Производство тепла:
При определенных условиях, протоны могут повторно входить в митохондриальный матрикс, не внося вклад в АТФ синтез. Этот процесс известен как протонная утечка или митохондриальная разобщение и связано с облегченной диффузией протонов в матрица.Результатом этого процесса является высвобождение потенциальной энергии протона. электрохимический градиент выделяется в виде тепла. Процесс опосредуется протонный канал называется термогенином или UCP1. Термогенин — это первый белок с молекулярной массой 33 кДа. открыт в 1973 году. Термогенин в основном содержится в коричневой жировой ткани или бурый жир и отвечает за термогенез без дрожи. Коричневый жир ткань встречается у млекопитающих и находится на самом высоком уровне в раннем периоде жизни и в зимующие животные.У людей коричневая жировая ткань присутствует при рождении и уменьшается с возрастом.
Хранение кальция:
Концентрации свободных кальций в клетке может регулировать множество реакций и важен для передача сигнала в клетке. Митохондрии могут временно накапливать кальций, способствует процессу гомеостаза кальция в клетке. Фактически, их способность быстро усваивать кальций для последующего высвобождения делает их очень хорошими «цитозольными» буферы »для кальция.Эндоплазматический ретикулум (ЭР) является наиболее значимым место хранения кальция, и существует значительная взаимосвязь между митохондрии и ER относительно кальция. Кальций попадает в матрикс унипортером кальция на внутренней митохондриальной мембране. это в первую очередь за счет потенциала митохондриальной мембраны. Выпуск этого кальций обратно внутрь клетки может происходить через обмен натрия и кальция. белка или через пути «кальций-индуцированного высвобождения кальция».Это может инициировать кальциевые шипы или кальциевые волны с большими изменениями мембранного потенциала. Они могут активировать ряд белков системы вторичных мессенджеров, которые могут координировать процессы, такие как высвобождение нейротрансмиттера в нервных клетках и высвобождение гормонов в эндокринных клетках.
Синтез белка:
От 5 до 10% белков митохондрии синтезируются митохондриальной ДНК.Митохондрии синтезируют субъединицы АТФазы, часть редуктазы и три субъединицы цитохрома оксидаза.
Синтез стероидов гормоны:
Первый шаг в превращение холестерина в стероидные гормоны в коре надпочечников, катализируется митохондриальными ферментами.
Цикл мочевины:
Цикл мочевины играет решающую роль в детоксикации аммиака.В этом цикле синтезируется мочевина. Первый шаг этого цикла, то есть превращения орнитина в цитруллин, происходит в митохондрии.
Биосинтез гема:
Heme, который нужен как простетическая группа в нескольких важных белках, таких как гемоглобин, миоглобин и цитохром С частично синтезируется в митохондриях.Конденсация сукцинил-КоА и глицин в δ-аминолевулиновую кислоту является ключевым этапом синтеза гема. и происходит в митохондрии. δ-Аминолевулиновая кислота затем доставляется в цитозоль, в котором копропорфириноген III образуется после ряда реакций. Эта молекула позже возвращается в митохондрию и превращается в гем.
Окисление жирных кислот:
Карнитин-зависимый транспорт жирных кислот и их окисление β является еще одним важным метаболический путь расположен в митохондрии.Большинство жирных кислот должны быть окисленные для производства энергии путем внутримитохондриального β-окисления, должны быть транспортируется из цитозоля в митохондрию. Для транспорта жирный кислоты сначала этерифицируются коферментом A (CoA) для активации, а затем соединен с карнитином, чтобы пересечь двойную мембрану митохондрий. Все Ферменты β-окисления — это митохондриальные ферменты. Ацетил-КоА, НАДН и ФАДН 2 , которые образуются в каждом раунде окисления жирных кислот, позже будут направляется либо в цикл лимонной кислоты, либо непосредственно в дыхательные пути цепь для производства АТФ.
КЛЕТОЧНОЕ ДЫХАНИЕ:
Клеточное дыхание, также известный как «окислительный метаболизм», является одним из ключевых способов получения клетками полезных энергия. Это совокупность происходящих метаболических реакций и процессов. в клетках организмов для преобразования биохимической энергии из питательных веществ в аденозин трифосфат (АТФ), а затем выделяют продукты жизнедеятельности. Реакции, связанные с дыхание — это катаболические реакции, которые включают окисление одной молекулы и сокращение другого.
Питательные вещества, обычно используемые дыхательные клетки животных и растений включают глюкозу, аминокислоты и жирные кислоты. кислоты, а общий окислитель (акцептор электронов) — молекулярный кислород (O2). Бактерии и археи также могут быть литотрофами, и эти организмы могут дышать, используя широкий спектр неорганических молекул в качестве доноров электронов и акцепторы, такие как сера, ионы металлов, метан или водород. Организмы, использующие кислород как конечный акцептор электронов при дыхании описывается как аэробный, в то время как те, которые этого не делают, называются анаэробными.
Энергия, выделяемая в дыхание используется для синтеза АТФ для хранения этой энергии. Энергия, хранящаяся в Затем АТФ можно использовать для управления процессами, требующими энергии, в том числе биосинтез, перемещение или транспортировка молекул через клеточные мембраны.
Аэробное дыхание:
Для аэробного дыхания требуется кислород для выработки энергии (АТФ).Это предпочтительный метод пирувата. распад в результате гликолиза и требует, чтобы пируват проник в митохондрии в чтобы полностью окислиться по циклу Кребса.
Гликолиз:
Гликолиз — это метаболический пути, который находится в цитоплазме клеток всех живых организмов и является анаэробный (то есть кислород не требуется). Процесс превращает одну молекулу глюкозы в две молекулы пирувата, и производит энергию в виде двух чистые молекулы АТФ.Фактически производится четыре молекулы АТФ на глюкозу; однако два потребляются на подготовительном этапе. Начальное фосфорилирование глюкозы требуется для дестабилизации молекулы для расщепления на две триозы сахара. Во время фазы окупаемости гликолиза четыре фосфатные группы переносится на АДФ путем фосфорилирования на уровне субстрата с образованием четырех АТФ и двух НАДН образуется при окислении триозных сахаров.
Окислительное декарбоксилирование Пируват:
Пируват окисляется до ацетил-КоА и CO 2 комплексом пируватдегидрогеназы, кластер ферментов: несколько копий каждого из трех ферментов, расположенных в митохондриях. эукариотических клеток и в цитозоле прокариот.В процессе один молекула НАДН образуется на окисленный пируват, и образуются 3 моля АТФ на каждую моль пирувата. Этот шаг также известен как реакция связи , поскольку он связывает гликолиз и цикл Кребса.
Цикл лимонной кислоты:
Это также называется циклом Кребса или цикл трикарбоновых кислот . Когда присутствует кислород, образуется ацетил-КоА. из молекул пирувата, созданных в результате гликолиза.Как только ацетил-КоА образуется, могут происходить два процесса: аэробное или анаэробное дыхание. Когда кислород в настоящее время митохондрии будут подвергаться аэробному дыханию, что приводит к Цикл Кребса. Однако, если кислорода нет, ферментация пирувата молекула произойдет. В присутствии кислорода, когда образуется ацетил-КоА, молекула затем входит в цикл лимонной кислоты (цикл Кребса) внутри митохондриальный матрикс и окисляется до CO 2 , в то же время восстановление НАД до НАДН.НАДН может быть использован цепочкой переноса электронов для создания далее АТФ как часть окислительного фосфорилирования. Чтобы полностью окислить эквивалент одной молекулы глюкозы, два ацетил-КоА должны метаболизироваться Цикл Кребса. Образуются два отхода: H 2 O и CO 2 . во время этого цикла.
Цикл лимонной кислоты — это 8-этапный процесс. с участием 8 различных ферментов. На протяжении всего цикла ацетил-КоА меняет в цитрат, изоцитрат, α-кетоглутарат, сукцинил-КоА, сукцинат, фумарат, малат и, наконец, оксалоацетат.Чистая выгода от одного цикла составляет 3 НАДН, 1 ФАДГ и 1 АТФ. Таким образом, общий выход энергии из одной цельной глюкозы молекула (2 молекулы пирувата) — это 6 НАДН, 2 ФАДН и 2 АТФ.
Окислительное фосфорилирование:
У эукариот происходит окислительное фосфорилирование. в митохондриальных кристах. Он включает цепь переноса электронов, которая устанавливает протонный градиент (хемиосмотический потенциал) через внутреннюю мембрану путем окисления НАДН, образующегося в цикле Кребса.АТФ синтезируется Фермент АТФ-синтаза, когда хемиосмотический градиент используется для управления фосфорилирование АДФ. В конечном итоге электроны переходят в экзогенный кислород и, при добавлении двух протонов, образуется вода.
Электронная транспортная цепь (ETC):
При окислительном фосфорилировании электроны полученный из NADH и FADH 2 в сочетании с O 2 , и энергия, высвобождаемая в результате этих реакций окисления / восстановления, используется для приведения в действие синтез АТФ из АДФ.
Электроны из НАДН попадают в электронный транспорт. цепь в комплексе I, который состоит из почти 40 полипептидных цепей. Эти электроны сначала передаются от НАДН к флавинмононуклеотиду, а затем, через носитель железо-сера к коферменту Ql. Коэнзим Q (также называемый убихинон) представляет собой небольшую липидорастворимую молекулу, которая переносит электроны от комплекс I через мембрану до комплекса III, который состоит из примерно десяти полипептиды. В комплексе III электроны переносятся с цитохрома b к цитохрому c an энергоемкая реакция.Цитохром c , белок периферической мембраны связаны с внешней стороной внутренней мембраны, а затем переносят электроны в сложные IV (цитохромоксидаза), где они окончательно переносятся на O 2 .
Отдельный белковый комплекс (II), состоящий из из четырех полипептидов, получает электроны из цикла лимонной кислоты промежуточный, сукцинатный. Эти электроны переносятся на FADH 2 , а не к НАДН, а затем к коферменту Q.Из кофермента Q электроны переходят в комплекс III, а затем в комплекс IV, как уже описано. В в отличие от переноса электронов от НАДН к коферменту Q в комплексе I, перенос электронов от FADH 2 к коферменту Q не связан с значительное уменьшение свободной энергии и, следовательно, не связывается с АТФ синтез. Следовательно, прохождение электронов происходит от FADH 2 через цепь переноса электронов дает свободную энергию только у комплексов III и IV.
Синтез АТФ:
Механизм сопряжения транспорта электронов с Генерация АТФ, хемиосмотическое соединение, является ярким примером взаимосвязи между структурой и функцией в клеточной биологии. Гипотеза хемиосмотического соединение было впервые предложено в 1961 году Питером Митчеллом, который предположил, что АТФ является генерируется за счет использования энергии, накопленной в виде протонных градиентов через биологические мембраны, а не путем прямого химического переноса высокоэнергетических группы.
Электронный транспорт через комплексы I, III и IV связан с переносом протонов изнутри митохондрия. Таким образом, энергоемкие реакции переноса электронов имеют вид связанный с переносом протонов из матрицы в межмембранное пространство, который устанавливает градиент протонов через внутреннюю мембрану. Комплексы I и III каждый переносит четыре протона через мембрану на пару электронов. п комплекс IV, два протона на пару электронов перекачиваются через мембрану и еще два протона на пару электронов объединяются с O 2 , чтобы образуют H 2 O внутри матрицы.Таким образом, эквивалент четырех протонов на пара электронов переносится из митохондриального матрикса в каждом из эти три комплекса.
Поскольку бислой фосфолипидов непроницаем к ионам протоны могут пересекать мембрану только через белковый канал. Это ограничение позволяет энергии в электрохимическом градиенте быть собираются и превращаются в АТФ в результате действия пятого комплекса участвует в окислительном фосфорилировании, комплексе V или АТФ-синтазе.АТФ-синтаза состоит из двух структурно различных компонентов: F 0 и F 1 , которые связаны тонким стеблем. Часть F 0 охватывает внутреннюю мембрана и обеспечивает канал, по которому протоны могут течь обратно из межмембранное пространство к матрице. Энергетически выгодное возвращение протоны к матрице связаны с синтезом АТФ субъединицей F 1 , который катализирует синтез АТФ из АДФ и ионов фосфата (P i ).
Транспорт через митохондрии:
Помимо стимулирования синтеза АТФ, потенциальная энергия, накопленная в электрохимическом градиенте, управляет переносом небольшие молекулы в митохондрии и из них. Например, синтезированный АТФ внутри митохондрий должен экспортироваться в цитозоль, в то время как АДФ и P i необходимо импортировать из цитозоля для продолжения синтеза АТФ.В электрохимический градиент, создаваемый протонной накачкой, обеспечивает необходимую энергию для транспорта этих молекул и других метаболитов, которые необходимо сосредоточены в митохондриях.
Большинство митохондриальных белков кодируется ядерным геномом. Эти белки транслируются на свободные рибосомы. и импортируется в митохондрии в виде законченных полипептидных цепей. Положительно заряженные предследовательности целевых белков для импорта в митохондриальный матрикс.Фосфолипиды переносятся в митохондрии из эндоплазматического ретикулума посредством белки-переносчики фосфолипидов.
ТЕОРЕТИЧЕСКИЙ ВЫХОД ЭНЕРГИИ:
Теперь можно рассчитать общий выход АТФ от окисления глюкозы. Чистая прибыль от гликолиз — это две молекулы АТФ и две молекулы НАДН. Преобразование пирувата в ацетил-КоА и его метаболизм через цикл лимонной кислоты дает два дополнительные молекулы АТФ, восемь NADH и две FADH 2 .Предполагая, что три молекулы АТФ образуются в результате окисления каждого НАДН и по два от каждого FADH 2 , общий выход составляет 38 молекул АТФ на молекула глюкозы. Однако в некоторых ячейках этот выход ниже, потому что два молекулы НАДН, образующиеся при гликолизе в цитозоле, не могут проникать митохондрии напрямую. Вместо этого их электроны должны быть переведены в митохондрии через челночную систему. В зависимости от используемой системы этот перевод может привести к тому, что эти электроны попадут в цепь переноса электронов на уровне из FADH 2 .В таких случаях две молекулы НАДН, полученные из гликолиз дают две, а не три молекулы АТФ, уменьшая общий выход до 36, а не 38 АТФ на молекулу глюкозы.
Ферментация:
Ферментация — это процесс получение энергии от окисления органических соединений, таких как углеводы, с использованием эндогенного акцептора электронов, которым обычно является органическое соединение.Без кислорода пируват не метаболизируется посредством клеточного дыхания, а не метаболизируется. подвергается процессу брожения. Пируват не попадает в митохондрии, но остается в цитоплазме, где превращается в отходы продукты, которые могут быть удалены из ячейки. Это служит для окисления носители водорода, чтобы они могли снова выполнить гликолиз и удалить избыток пирувата. Этот продукт жизнедеятельности варьируется в зависимости от организма. В скелетные мышцы, продукт жизнедеятельности — молочная кислота.Этот вид брожения называется молочнокислым брожением. В дрожжах отходами являются этанол и углекислый газ. Этот тип ферментации известен как алкогольный или этанольный. ферментация. АТФ, образующийся в этом процессе, производится на уровне субстрата. фосфорилирование, не требующее кислорода
Анаэробное дыхание:
Используется анаэробное дыхание некоторыми микроорганизмами, у которых нет ни кислорода (аэробное дыхание), ни пируват или производное пирувата (ферментация) является конечным акцептором электронов.Вместо этого используется неорганический акцептор (например, сера).
23.1E: Эволюция пластид
Пластиды могут происходить от цианобактерий, поглощенных посредством эндосимбиоза ранними эукариотами, что дает клеткам способность проводить фотосинтез.
Задачи обучения
- Объяснить связь между эндосимбиозом и пластидами и эволюцией эукариот
Ключевые моменты
- Хлоропласты, хромопласты и лейкопласты — каждый тип пластиды.
- Пластиды у эукариот происходят от первичного эндосимбиоза с участием древних цианобактерий.
- Chlorarachniophytes — это тип водорослей, которые возникли в результате вторичного эндосимбиоза, когда эукариот поглотил зеленую водоросль (которая сама была продуктом первичного эндосимбиоза с цианобактериями).
- Пластиды имеют несколько общих черт с митохондриями, в том числе наличие собственной ДНК и способность к репликации путем бинарного деления.
Ключевые термины
- хлоропласт : органелла, обнаруженная в клетках зеленых растений и фотосинтезирующих водорослей, где происходит фотосинтез
- тилакоид : складчатая мембрана в хлоропластах растений, из которой образуются граны, используемая в фотосинтезе
- пластида : любая из различных органелл, обнаруженных в клетках растений и водорослей, часто связанных с фотосинтезом
Пластиды
Некоторые группы эукариот фотосинтезируют: их клетки содержат, помимо стандартных эукариотических органелл, органеллы другого типа, называемые пластидами.Есть три типа пластид: хлоропласты, хромопласты и лейкопласты. Хлоропласты — это пластиды, которые проводят фотосинтез. Хромопласты — это пластиды, которые синтезируют и хранят пигменты. Лейкопласты — это пластиды, расположенные в несинтетических тканях растения (, например, , корни) и обычно хранящие непигментные молекулы.
Подобно митохондриям, пластиды, по-видимому, имеют первичное эндосимбиотическое происхождение, но отличаются тем, что происходят от цианобактерий, а не от альфа-протеобактерий.Цианобактерии — это группа фотосинтезирующих бактерий со всеми обычными структурами прокариот. Однако, в отличие от большинства прокариот, у них есть обширные внутренние мембранные компартменты, называемые тилакоидами, которые содержат хлорофилл и являются местом светозависимых реакций фотосинтеза. Помимо тилакоидов, хлоропласты, обнаруженные у эукариот, имеют кольцевую хромосому ДНК и рибосомы, аналогичные таковым у цианобактерий. Каждый хлоропласт окружен двумя мембранами, что указывает на первичный эндосимбиоз.Считается, что внешняя мембрана, окружающая пластиду, произошла от вакуоли хозяина, а внутренняя мембрана, как полагают, произошла от плазматической мембраны эндосимбионта.
Рисунок \ (\ PageIndex {1} \): Хлоропласт : (a) Это поперечное сечение хлоропласта иллюстрирует его сложную внутреннюю мембранную организацию. Стеки тилакоидных мембран разделяют фотосинтетические ферменты и обеспечивают основу для ДНК хлоропластов. (б) Хлоропласты можно увидеть как маленькие зеленые сферы.Также, как и в случае с митохондриями, есть убедительные доказательства того, что многие из генов эндосимбионта передаются в ядро. Пластиды, как и митохондрии, не могут жить независимо вне хозяина. Кроме того, как и митохондрии, пластиды образуются в результате бинарного деления других пластид. Исследователи предположили, что эндосимбиотическое событие, которое привело к появлению Archaeplastida (наземные растения, красные и зеленые водоросли), произошло от 1 до 1,5 миллиарда лет назад, по крайней мере, через 500 миллионов лет после летописи окаменелостей, что предполагает присутствие эукариот.
Вторичный эндосимбиоз Chlorarachniophytes
Эндосимбиоз вовлекает одну клетку, поглощающую другую, чтобы со временем произвести коэволюционирующие отношения, в которых ни одна клетка не могла бы выжить в одиночку. Например, хлоропласты красных и зеленых водорослей образуются в результате поглощения фотосинтетической цианобактерии ранним прокариотом. Это приводит к вопросу о возможности клетки, содержащей эндосимбионт, поглотить себя, что приведет к вторичному эндосимбиозу.Не все пластиды эукариот происходят непосредственно от первичного эндосимбиоза. Некоторые из основных групп водорослей стали фотосинтезировать в результате вторичного эндосимбиоза; то есть, поглощая либо зеленые водоросли, либо красные водоросли в качестве эндосимбионтов. Этот вывод подтверждают многочисленные микроскопические и генетические исследования; вторичные пластиды окружены тремя и более мембранами; некоторые вторичные пластиды даже имеют явные остатки ядра эндосимбиотических водорослей.
Рисунок \ (\ PageIndex {1} \): Первичный и вторичный эндосимбиоз : показан предполагаемый процесс эндосимбиотических событий, ведущих к эволюции хлорарахниофитов.При первичном эндосимбиотическом событии гетеротрофный эукариот потреблял цианобактерию. При вторичном эндосимбиозе клетка, возникшая в результате первичного эндосимбиоза, была поглощена второй клеткой. Образовавшаяся органелла стала пластидой у современных хлорарахниофитов. Рисунок \ (\ PageIndex {1} \): Красные и зеленые водоросли : (а) красные водоросли и (б) зеленые водоросли (визуализированные с помощью световой микроскопии) имеют сходные последовательности ДНК с фотосинтетические цианобактерии. Ученые предполагают, что в процессе, называемом эндосимбиозом, предковые прокариоты поглотили фотосинтетические цианобактерии, которые превратились в современные хлоропласты.Молекулярные и морфологические данные свидетельствуют о том, что простейшие хлорахниофиты являются производными вторичного эндосимбиотического события. Хлорарахниофиты — это редкие водоросли, обитающие в тропических морях и песках. Считается, что эти протисты возникли, когда эукариот поглотил зеленую водоросль, последняя из которых уже установила эндосимбиотические отношения с фотосинтетической цианобактерией. Несколько линий доказательств подтверждают, что хлорарахниофиты произошли от вторичного эндосимбиоза.Хлоропласты, содержащиеся в эндосимбионтах зеленых водорослей, способны к фотосинтезу, заставляя хлорарахниофиты фотосинтезировать. Эндосимбионт зеленых водорослей также имеет низкорослое рудиментарное ядро. Фактически, кажется, что хлорахниофиты являются продуктами недавнего (в масштабе эволюции) вторичного эндосимбиотического события. Пластиды хлорахниофитов окружены четырьмя мембранами: первые две соответствуют внутренней и внешней мембранам фотосинтетической цианобактерии, третья соответствует зеленой водоросли, а четвертая соответствует вакуоли, окружавшей зеленую водоросль, когда она была поглощена. предок хлорарахниофитов.
Процесс вторичного эндосимбиоза не уникален для хлорарахниофитов. Фактически, вторичный эндосимбиоз зеленых водорослей также привел к появлению простейших эвгленид, тогда как вторичный эндосимбиоз красных водорослей привел к эволюции динофлагеллят, апикомплексанов и страменопилов.
ЛИЦЕНЗИИ И АТРИБУЦИИ
CC ЛИЦЕНЗИОННОЕ СОДЕРЖАНИЕ, ПРЕДЫДУЩЕЕ РАЗМЕЩЕНИЕ
- Кураторство и пересмотр. Источник : Boundless.com. Лицензия : CC BY-SA: Attribution-ShareAlike
CC ЛИЦЕНЗИОННОЕ СОДЕРЖАНИЕ, СПЕЦИАЛЬНАЯ АТРИБУЦИЯ
- Колледж OpenStax, Биология.16 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44612/latest…ol11448/latest . Лицензия : CC BY: Атрибуция
- Колледж OpenStax, Биология. 12 ноября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44614/latest…ol11448/latest . Лицензия : CC BY: Атрибуция
- Общая биология / Классификация живых существ / Эукариоты / Протисты. Источник : Викиучебники. Расположен по адресу : en.wikibooks.org/wiki/General…yotes/Protists . Лицензия : CC BY-SA: Attribution-ShareAlike
- эндомембрана. Источник : Викисловарь. Расположен по адресу : en.wiktionary.org/wiki/endomembrane . Лицензия : CC BY-SA: Attribution-ShareAlike
- аэробный. Источник : Викисловарь. Расположен по адресу : en.wiktionary.org/wiki/aerobic . Лицензия : CC BY-SA: Attribution-ShareAlike
- цианобактерий. Источник : Викисловарь. Адрес: : en.wiktionary.org/wiki/cyanobacteria . Лицензия : CC BY-SA: Attribution-ShareAlike
- Колледж OpenStax, Введение.16 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44612/latest…3_00_01abc.jpg . Лицензия : CC BY: Атрибуция
- Колледж OpenStax, Биология. 16 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44614/latest…ol11448/latest . Лицензия : CC BY: Атрибуция
- Структурная биохимия / Прокариоты и эукариоты. Источник : Викиучебники. Расположен по адресу : en.wikibooks.org/wiki/Structu…and_Eukaryotes . Лицензия : CC BY-SA: Attribution-ShareAlike
- Структурная биохимия / Клеточная основа / Эукариотическая клетка. Источник : Викиучебники. Расположен по адресу : en.wikibooks.org/wiki/Structu…ukaryotic_Cell . Лицензия : CC BY-SA: Attribution-ShareAlike
- Структурная биохимия / Митохондриальная ДНК. Источник : Викиучебники. Расположен по адресу : en.wikibooks.org/wiki/Structural_Biochemistry/Mitochondrial_DNA . Лицензия : CC BY-SA: Attribution-ShareAlike Плазмида
- . Источник : Викисловарь. Расположен по адресу : en.wiktionary.org/wiki/plasmid . Лицензия : CC BY-SA: Attribution-ShareAlike
- теломер. Источник : Викисловарь. Адрес: : en.wiktionary.org/wiki/telomere . Лицензия : CC BY-SA: Attribution-ShareAlike
- Колледж OpenStax, Введение. 16 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44612/latest…3_00_01abc.jpg . Лицензия : CC BY: Атрибуция
- Структурная биохимия / Прокариоты и эукариоты. Источник : Викиучебники. Расположен по адресу : en.wikibooks.org/wiki/Structu…and_Eukaryotes . Лицензия : CC BY-SA: Attribution-ShareAlike
- цианобактерий. Источник : Викисловарь. Адрес: : en.wiktionary.org/wiki/cyanobacteria . Лицензия : CC BY-SA: Attribution-ShareAlike
- Колледж OpenStax, Биология.16 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44614/latest…ol11448/latest . Лицензия : CC BY: Атрибуция
- Структурная биохимия / Теория эндосимбиотиков. Источник : Викиучебники. Расположен по адресу : en.wikibooks.org/wiki/Structu…mbiotic_Theory . Лицензия : CC BY-SA: Attribution-ShareAlike
- Введение в молекулярную биологию / макромолекулы и клетки. Источник : Викиучебники. Расположен по адресу : en.wikibooks.org/wiki/An_Intr…mbiotic_theory . Лицензия : CC BY-SA: Attribution-ShareAlike
- пероксисома. Источник : Викисловарь. Адрес: : en.wiktionary.org/wiki/peroxisome . Лицензия : CC BY-SA: Attribution-ShareAlike
- эндосимбионт. Источник : Викисловарь. Адрес: : en.wiktionary.org/wiki/endosymbiont . Лицензия : CC BY-SA: Attribution-ShareAlike
- Колледж OpenStax, Введение. 16 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44612/latest…3_00_01abc.jpg . Лицензия : CC BY: Атрибуция
- Структурная биохимия / Прокариоты и эукариоты. Источник : Викиучебники. Расположен по адресу : en.wikibooks.org/wiki/Structural_Biochemistry/Prokaryotes_and_Eukaryotes . Лицензия : CC BY-SA: Attribution-ShareAlike
- Plagiomnium affine ламиназеллен. Источник : Википедия. Расположен по адресу : en.Wikipedia.org/wiki/File:Pl…inazellen.jpeg . Лицензия : CC BY: Атрибуция
- Эндосимбиоз. Источник : Викимедиа. Адрес: : commons.wikimedia.org/wiki/Fi…osymbiosis.svg . Лицензия : CC BY-SA: Attribution-ShareAlike
- Колледж OpenStax, Биология. 16 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44614/latest…ol11448/latest . Лицензия : CC BY: Атрибуция
- Структурная биохимия / Теория эндосимбиотиков. Источник : Викиучебники. Расположен по адресу : en.wikibooks.org/wiki/Structu…mbiotic_Theory . Лицензия : CC BY-SA: Attribution-ShareAlike
- вакуоль. Источник : Викисловарь. Адрес: : en.wiktionary.org/wiki/vacuole . Лицензия : CC BY-SA: Attribution-ShareAlike
- crista. Источник : Википедия. Расположен по адресу : en.Wikipedia.org/wiki/crista . Лицензия : CC BY-SA: Attribution-ShareAlike
- эндосимбиоз. Источник : Викисловарь. Расположен по адресу : en.wiktionary.org/wiki/endosymbiosis . Лицензия : CC BY-SA: Attribution-ShareAlike
- Колледж OpenStax, Введение.16 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44612/latest…3_00_01abc.jpg . Лицензия : CC BY: Атрибуция
- Структурная биохимия / Прокариоты и эукариоты. Источник : Викиучебники. Расположен по адресу : en.wikibooks.org/wiki/Structural_Biochemistry/Prokaryotes_and_Eukaryotes . Лицензия : CC BY-SA: Attribution-ShareAlike
- Plagiomnium affine ламиназеллен. Источник : Википедия. Расположен по адресу : en.Wikipedia.org/wiki/File:Plagiomnium_affine_laminazellen.jpeg . Лицензия : CC BY: Атрибуция
- Эндосимбиоз. Источник : Викимедиа. Расположен по адресу : commons.wikimedia.org/wiki/File:Endosymbiosis.svg . Лицензия : CC BY-SA: Attribution-ShareAlike
- Колледж OpenStax, Эукариотическое происхождение.16 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44614/latest/Figure_23_01_01.jpg . Лицензия : CC BY: Атрибуция
- Колледж OpenStax, Биология. 16 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44614/latest/?collection=col11448/latest . Лицензия : CC BY: Атрибуция
- Структурная биохимия / Клеточные органеллы / Растительная клетка. Источник : Викиучебники. Расположен по адресу : en.wikibooks.org/wiki/Structural_Biochemistry/Cell_Organelles/Plant_Cell%23Plastid . Лицензия : CC BY-SA: Attribution-ShareAlike
- пластид. Источник : Викисловарь. Адрес: : en.wiktionary.org/wiki/plastid . Лицензия : CC BY-SA: Attribution-ShareAlike
- тилакоид. Источник : Викисловарь. Адрес: : en.wiktionary.org/wiki/thylakoid . Лицензия : CC BY-SA: Attribution-ShareAlike
- хлоропласт. Источник : Викисловарь. Адрес: : en.wiktionary.org/wiki/chloroplast . Лицензия : CC BY-SA: Attribution-ShareAlike
- Колледж OpenStax, Введение.16 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44612/latest/Figure_B23_00_01abc.jpg . Лицензия : CC BY: Атрибуция
- Структурная биохимия / Прокариоты и эукариоты. Источник : Викиучебники. Расположен по адресу : en.wikibooks.org/wiki/Structural_Biochemistry/Prokaryotes_and_Eukaryotes . Лицензия : CC BY-SA: Attribution-ShareAlike
- Plagiomnium affine ламиназеллен. Источник : Википедия. Расположен по адресу : en.Wikipedia.org/wiki/File:Plagiomnium_affine_laminazellen.jpeg . Лицензия : CC BY: Атрибуция
- Эндосимбиоз. Источник : Викимедиа. Расположен по адресу : commons.wikimedia.org/wiki/File:Endosymbiosis.svg . Лицензия : CC BY-SA: Attribution-ShareAlike
- Колледж OpenStax, Эукариотическое происхождение.16 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44614/latest/Figure_23_01_01.jpg . Лицензия : CC BY: Атрибуция
- Колледж OpenStax, Эукариотическое происхождение. 16 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44614/latest/Figure_23_01_03ab.jpg . Лицензия : CC BY: Атрибуция
- Колледж OpenStax, Эукариотическое происхождение.16 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44614/latest/Figure_23_01_02.jpg . Лицензия : CC BY: Атрибуция
- Колледж OpenStax, Эукариотическое происхождение. 16 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44614/latest/Figure_23_01_05.jpg . Лицензия : CC BY: Атрибуция
Ячейка.6. Не везикулярный. Пластиды. Атлас гистологии растений и животных.
Пластиды — это органеллы, обнаруженные в клетках растений и водорослей, хотя они также встречаются у некоторых морских животных. Эти органеллы возникли в процессе эволюции после процесса эндосимбиоза, когда бактерия с фотосинтетическими способностями, подобная нынешним цианобактериям, была поглощена клеткой эукариот и стала эндосимбионтом вместо того, чтобы перевариваться. В ходе эволюции большинство генов захваченных цианобактерий были перенесены в ядро хозяина, а эндосимбионт превратился в пластиды, которые развили разнообразный набор функций: фотосинтез, синтез аминокислот и липидов, хранение липидов, углеводов и белков, обеспечение окраски различных частей. тела растения, ощущение силы тяжести, регулирование поведения устьиц и многие другие.
Пластиды ограничены двойной мембраной и межмембранным пространством (рис. 1). Внутри они содержат другие ограниченные мембраной компартменты, такие как тилакоиды хлоропластов и канальцы хромопластов. Подобно митохондриям, пластиды также содержат ДНК и молекулярные механизмы для роста и деления, причем оба процесса находятся под контролем ядерных генов клетки. Каждая пластида в каждой клетке происходит из другой существующей пластиды, и они передаются следующему поколению растений, включенных в гаметы во время оплодотворения.Таким образом, все пластиды растения являются потомками пластид эмбриона, известных как пропластиды. Пропластиды также обнаруживаются в меристематических клетках взрослых растений, где они делятся перед делением клеток, чтобы обеспечить надлежащее количество пропластидов в двух новых клетках. Когда начинается дифференцировка клеток, пропластиды также запускают свою собственную программу дифференцировки, так что разные типы зрелых пластид генерируются в соответствии с типом клеток. Это могут быть лейкопластиды (элайопласты, амилопластиды и протеопластиды), хлоропласты и хромопласты.Хлоропласты могут также дифференцироваться в другие типы пластид, а другие пластиды могут дифференцироваться в хлоропласты. Это двунаправленный путь дифференциации (рис. 1).
Рисунок 1. Типы пластид и пути дифференцировки (адаптировано из Jarvis and López-Juez, 2013)1. Пропластиды
Пропластиды — это небольшие пластиды, около 1 мкм в диаметре, менее сложные на структурном уровне, чем другие пластиды растения. Они бесцветны, могут изменять свою морфологию и могут содержать различное количество трубчатых внутренних мембранных компартментов, а также депо крахмала.Эти особенности присущи двум типам пропластидов: зародышевым и клубеньковым пропластидам. Зародышевые пропластиды обнаружены в зародышах растений (в семенах) и меристематических клетках. Делением и дифференциацией они дают начало остальным пластидам растения. Они также могут выполнять некоторые метаболические функции, такие как синтез гибберелловой кислоты, что очень важно для меристематического метаболизма. Пропластиды клубеньков, как следует из названия, находятся в клубеньках корней и участвуют в фиксации азота.
Этиопласты, другой тип пластид, находятся в стеблях, но не в корнях. Они представляют собой длительную промежуточную стадию на пути дифференциации от пропластидов до хлоропластов при очень низкой интенсивности света или темноты. Этиопласты возобновляют дифференцировку в сторону хлоропластов, как только свет становится достаточно интенсивным.
2. Лейкопласты
Лейкопласты — это бесцветные пластиды (без пигментов), которые функционируют как хранящие органеллы.Лейкопласты включают амилопласты, элайопласты (или олеопласты) и протеинопласты. Они хранят крахмал, липиды и белки соответственно.
В клетках растений амилопласты синтезируют крахмал (рис. 2). Весь крахмал, хранящийся в клетке, можно найти в пластидах в виде гранул крахмала. Амилопласты специализируются на этой роли и содержат большие депо крахмала. Майелопласты не только хранят крахмал, но и являются датчиками силы тяжести в клетках корня. Гранулы крахмала плотнее воды, поэтому амилопласты падают на дно клетки, и это то, что нужно клеткам для обнаружения вектора гравитации.Амилопласты также участвуют в метаболизме азота.
Элайопласты представляют собой пластиды небольшого размера, содержащие масло и липиды, образующие жировые капли. В клетках растений существует два пути синтеза липидов. Эукариотический путь зависит от гладкой эндоплазматической сети, тогда как так называемый прокариотический путь включает элайопластиды. Эти два пути производят разные типы липидов. Элайопласты также участвуют в созревании пыльцы. Некоторые растения способны накапливать липиды в других органеллах, известных как элайосомы, которые происходят из эндоплазматического ретикулума.
Протеинопласты содержат высокую концентрацию белков, настолько высокую, что белки образуют кристаллы или очень плотный аморфный материал. Однако пока не ясно, существует ли тип пластид, предназначенный исключительно для хранения белка в растительных клетках.
3. Хромопласты
Хромопласты содержат каротиноидные пигменты, которые придают красный, оранжевый и желтый цвета структуре растения, где они присутствуют (рис. 2. Эти пластиды изобилуют цветами, фруктами, старыми листьями и некоторыми корнями.Считается, что одна из их основных функций — привлечение животных для опыления или распространения семян. Хромопласты метаболически активны, хотя они содержат меньше копий ДНК, чем хлоропласты.
Рисунок 2. Пластиды. Хлоропласты (А и Б). А) Фотосинтетическая паренхима. Стома показана на B). Хромопласты (C и D) томата. Амилопласты (E и F) из картофеля.Хромопласты содержат липидные капли с каротиноидами и сложные молекулы, известные как фибриллы, которые также содержат ядро каротиноидов.Хромопласты отличаются от хлоропластов, а также от пропластидов. Во время дифференциации от хлоропластов механизмы фотосинтеза и тилакоиды разрушаются, и каротиноиды синтезируются вместе с компартментами, в которые они будут включены. Эти компартменты, известные как пластоглобулы, представляют собой липидные капли с большим количеством триацилглицеридов, расположенных в строме пластиды. Пластоглобулы также можно найти в других пластидах. Каротиноиды, в основном ксантофиллы, также хранятся в других местах стромы пластид, настолько концентрированные, что могут образовывать нити или кристаллы.Хромопласты образуют внутреннюю слоистую мембранную систему во внешней части стромы. Эти новые мембраны возникают из-за инвагинации внутренней мембраны, а не из старых деградированных тилакоидов. Каротиноиды, такие как лютеин, бета-каротиноид и другие, также могут быть связаны с этими новыми мембранами. Иногда внутренние оболочки имеют сетчатое расположение.
Во время созревания хромопластов концентрация пигментов может быть настолько высокой, что они могут образовывать кристаллы, такие как бета-каротиноиды в корне моркови или ликопин в томате.Каротиноиды также могут концентрироваться в трубчатых структурах. Хромопласты могут содержать другие компоненты, такие как гранулы крахмала и агрегаты белка.
Хотя хромопласты считаются продвинутой стадией развития хлоропластов, было замечено, что они могут снова стать хлоропластами. Некоторые ткани корней и плодов могут снова стать зелеными. Например, плоды лимона, оставленные на дереве, меняют цвет с желтого на зеленый, а корни моркови становятся зелеными под воздействием света.
4. Хлоропласты
Хлоропласты — самые многочисленные и изученные пластиды. Они будут изучены на следующей странице.
5. Пластиды другие виды
В периоды старения и смерти клетки содержат геронтопласты, которые дифференцируются от хлоропастов. Муропласты обнаружены в водорослях глаукоцистофитных и содержат рудиментарную стенку пептидогликана, расположенную между внутренней и внешней мембранами органеллы. Пластиды типов S и T были обнаружены в ситовидных клетках флоэмы и могут реагировать на раны.Родопласты представляют собой фотосинтетические пластиды, обнаруженные в красных водорослях, и содержат хлорофилл типа А, но не хлорофиллы b или c. Родопласты содержат тилакоиды, не расположенные в стопки, а также молекулярные агрегаты, содержащие красные пигменты, известные как фикобилисомы, которые способны улавливать световые волны, которые достигают больших расстояний под морской водой. Наконец, некоторые животные едят водоросли, а хлоропласты не перевариваются, а включаются в клетки животных. Эти пластиды способны фотосинтезировать в течение месяцев, питая таким образом животное (например, Elysia chlorotica ).Эти пластиды известны как клептопласты. Кроме того, некоторые паразиты, такие как Plasmodium , могут содержать пластиды, известные как апикопласты.
Библиография
Jarvis P, López-Juez E . 2013. Биогенез и гомеостаз хлоропластов и других пластид. Обзоры природы в молекулярной и клеточной биологии. 14: 787-802.
Ljubesic N, Wrischer M, Devidé Z . 1991. Хромопласты — последние стадии развития пластид. Международный журнал биологии развития.35: 251-258.
Мудрый RR . 2006. Разнообразие форм и функций пластид. В Строении и функциях пластид. Springer Нидерланды.
Leave A Comment