
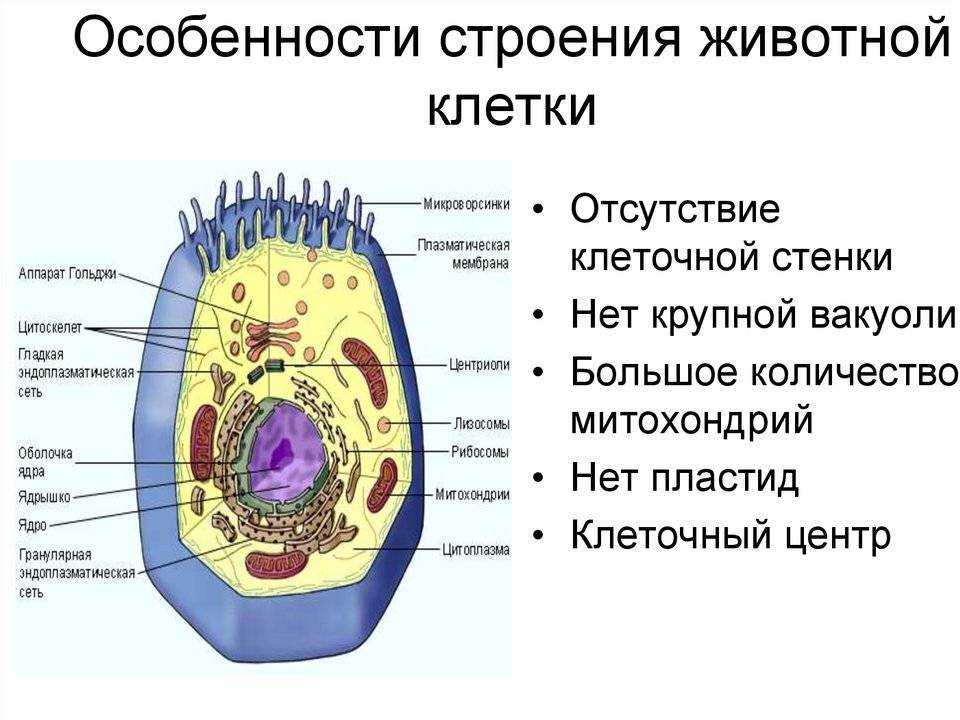
Связь между различными органоидами клетки осуществляется благодаря 1)митохондриям 2) аппарату Гольджи 3)лизосомам 4)эндоплазматической сети
Последние вопросы
Биология
2 минуты назад
Кістки передніх чи задніх кінцівок жаби більші за розмірами ? Як це пов’язано зі способом пересування? 7 клас помогитеБиология
17 минут назад
Завдання на фотоПредставники: тюльпан, суниця лісова, амеба звичайна, бджола, дріжджі, підберезник, конвалія, картопля, смородина, рак-самітник, інфузорія туфелька. Биология
1 час назад
1900 році три дослідники перевідкрили закономірності успадкування ознак у рослин, якi вiдкрив: а) г.
Биология
1 час назад
сколько раз в день нужно гладить котаБиология
1 час назад
Пожалуйста помогите!!! Даю 80 БАЛОВ.Биология
1 час назад
сколько нервных клеток выходит в состав коры головного мозга?Биология
1 час назад
Дайте відповідь на запитання: що для серця шкідливо, а що корисно? (Наведіть приклади)Биология
1 час назад
Что входит в выделительную систему плацентарных млекопитающих? 1. прямая кишка 2.клоака 3.почки 4. 5.мочевой пузырь 6.мочеточники 7.мочеиспускательный канал ответов несколькоБиология
2 часа назад
Наявність пігменту у волосеі людини домінус над альбінізмом (відсутністю пігменту). Чоловік і дружина гетерозиготні за піг метицісю волосся. Чи можливе народження у них дитини-альбіноса?Биология
2 часа назад
5. Однією з безпосередніх причин мутацій можуть бути: а) віруси; б) гриби; в) бактерії.Биология
2 часа назад
Якщо на початку року в популяції було 100 особин, за рік народилося 15, померло 20, вмігрувало 3, емігрувало 2, то природний приріст популяції становить:Биология
2 часа назад
наукове і практичне значення квітокОрхідеї-Бонсай-Антириум-Дроцена-будь ласка це срочно!2 часа назад
Особливим органом чуття у риб єБиология
2 часа назад
Охарактеризовать световые адаптации фотофилов.Биология
3 часа назад
СРОЧНО, ДАЮ 50 БАЛОВ Установи закономірність між розміщенням продихів та умовами росту рослин

 прямая кишка 2.клоака 3.почки 4. 5.мочевой пузырь 6.мочеточники 7.мочеиспускательный канал ответов несколько
прямая кишка 2.клоака 3.почки 4. 5.мочевой пузырь 6.мочеточники 7.мочеиспускательный канал ответов несколько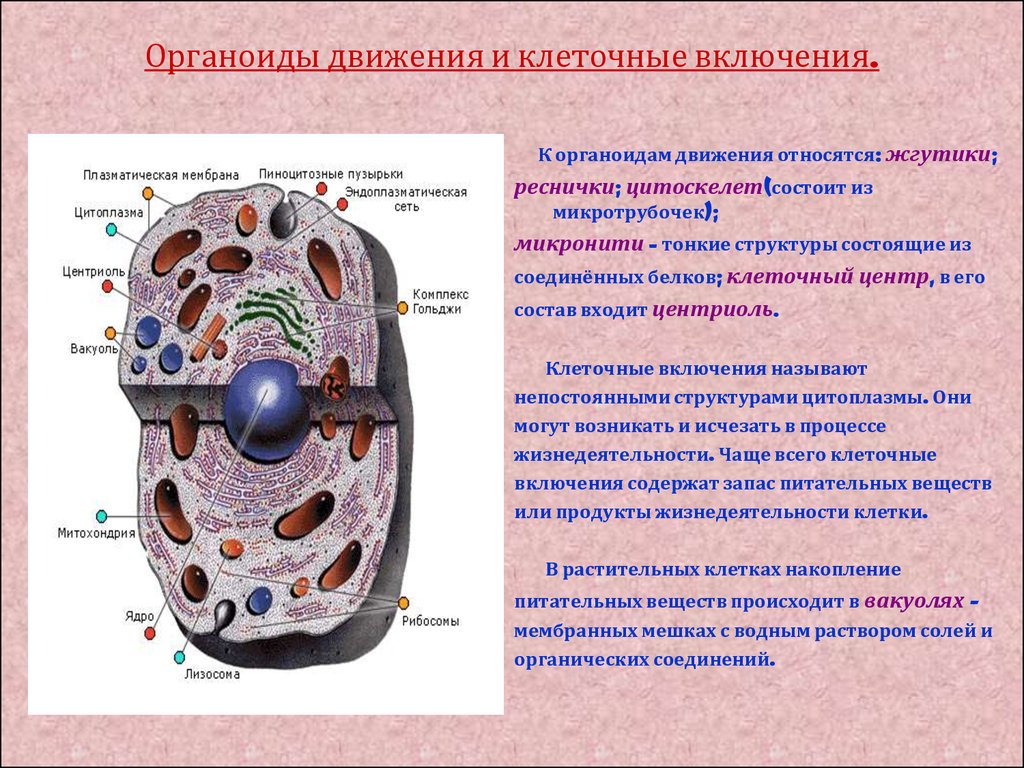
Все предметы
Выберите язык и регион
English
United States
Polski
Polska
Português
Brasil
English
India
Türkçe
Türkiye
English
Philippines
Español
España
Bahasa Indonesia
Indonesia
Русский
Россия
How much to ban the user?
1 hour 1 day 100 years
Цитология
Цитология- Рефераты на русском
- Медицина
- Цитология
Цитология
Цитология наука о клетке.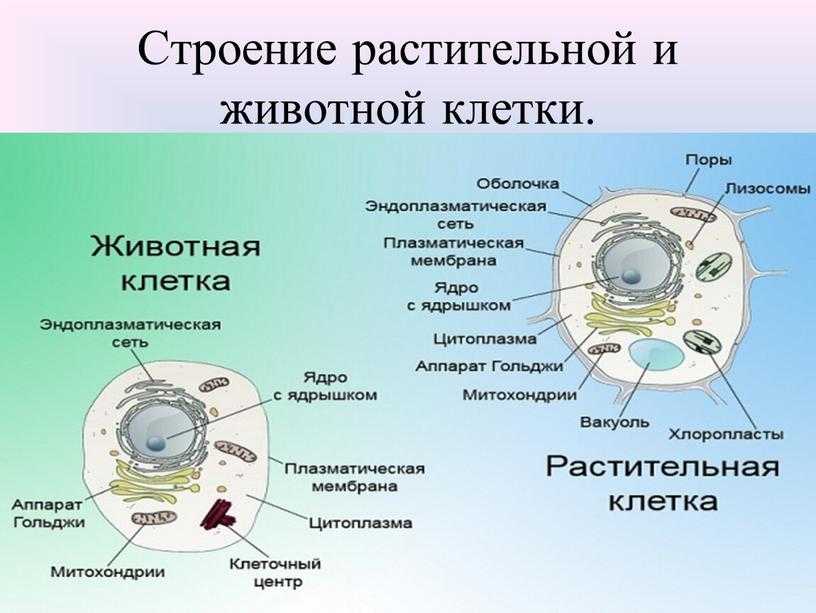 Наука о клетке называется цитологией (греч. -клетка, -наука) . Предмет цитологии — клетки многоклеточных животных и растений, а также одноклеточных организмов, к числу которых относятся бактерии, простейшие и одноклеточные водоросли. Цитология изучает строение и химический состав клеток, функции внутриклеточных структур, функции клеток в организме животных и растений, размножение и развитие клеток, приспособления клеток к условиям окружающей среды. Современная цитология — наука комплексная. Она имеет самые тесные связи с другими биологическими науками, например с ботаникой, зоологией, физиологией, учением об эволюции органического мира, а также с молекулярной биологией, химией, физикой, математикой. Цитология — одна из относительно молодых биологических наук, ее возраст около 100 лет. Возраст же термина «клетка» насчитывает свыше 300 лет. Впервые название в середине XVII в. применил Р. Гук. Рассматривая тонкий срез пробки с помощью микроскопа, Гук увидел, что пробка состоит из ячеек — клеток.
Наука о клетке называется цитологией (греч. -клетка, -наука) . Предмет цитологии — клетки многоклеточных животных и растений, а также одноклеточных организмов, к числу которых относятся бактерии, простейшие и одноклеточные водоросли. Цитология изучает строение и химический состав клеток, функции внутриклеточных структур, функции клеток в организме животных и растений, размножение и развитие клеток, приспособления клеток к условиям окружающей среды. Современная цитология — наука комплексная. Она имеет самые тесные связи с другими биологическими науками, например с ботаникой, зоологией, физиологией, учением об эволюции органического мира, а также с молекулярной биологией, химией, физикой, математикой. Цитология — одна из относительно молодых биологических наук, ее возраст около 100 лет. Возраст же термина «клетка» насчитывает свыше 300 лет. Впервые название в середине XVII в. применил Р. Гук. Рассматривая тонкий срез пробки с помощью микроскопа, Гук увидел, что пробка состоит из ячеек — клеток.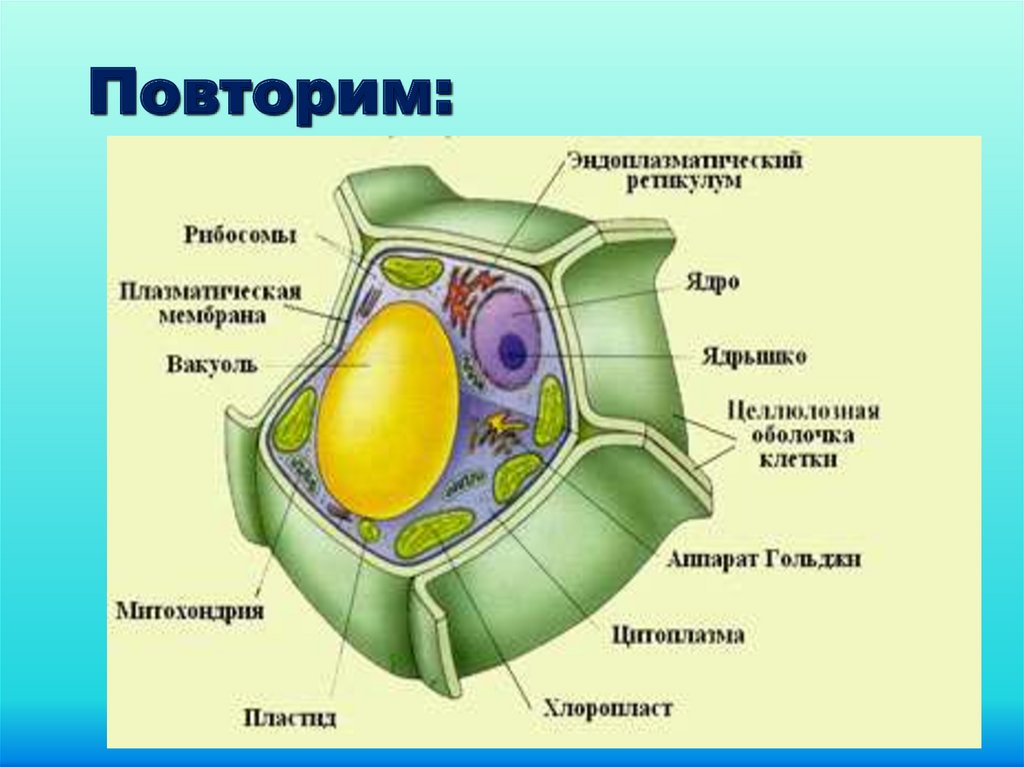
Клеточная теория. В середине XIX столетия на основе уже многочисленных знаний о клетке Т. Шванн сформулировал клеточную теорию (1838) . Он обобщил имевшиеся знания о клетке и показал, что клетка представляет основную единицу строения всех живых организмов, что клетки животных и растений сходны по своему строению. Эти положения явились важнейшими доказательствами единства происхождения всех живых организмов, единство всего органического мира. Т. Шван внес в науку правильное понимание клетки как самостоятельной единицы жизни, наименьшей единицы живого: вне клетки нет жизни.
Современная клеточная — теория включает следующие положения: клетка основная единица строения и развития всех живых организмов, наименьшая единица живого; клетки всех одноклеточных и многоклеточных организмов сходны (гомологичны) по своему строению, химическому составу, основным проявлениям жизнедеятельности и обмену веществ; размножение клеток происходит путем их деления, и каждая новая клетка образуется в результате деления исходной (материнской) клетки; в сложных многоклеточных организмах клетки специализированы по выполняемой ими функции и образуют ткани; из тканей состоят органы, которые тесно связаны между собой и подчинены нервным и гуморальным системам регуляции.
Исследования клетки имеют большое значение для разгадки заболеваний. Именно в клетках начинают развиваться патологические изменения, приводящие к возникновению заболеваний. Чтобы понять роль клеток в развитии заболеваний, приведем несколько примеров. Одно из серьезных заболеваний человека — сахарный диабет. Причина этого заболевания — недостаточная деятельность группы клеток поджелудочной железы, вырабатывающих гормон инсулин, который участвует в регуляции сахарного обмена организма. Злокачественные изменения, приводящие к развитию раковых опухолей, возникают также на уровне клеток. Возбудители кокцидиоза — опасного заболевания кроликов, кур, гусей и уток — паразитические простейшие — кокцидии проникают в клетки кишечного эпителия и печени, растут и размножаются в них, полностью нарушают обмен веществ, а затем разрушают эти клетки. У больных кокцидиозом животных сильно нарушается деятельность пищеварительной системы, и при отсутствии лечения животные погибают. Вот почему изучение строения, химического состава, обмена веществ и всех проявлений жизнедеятельности клеток необходимо не только в биологии, но также в медицине и ветеринарии.

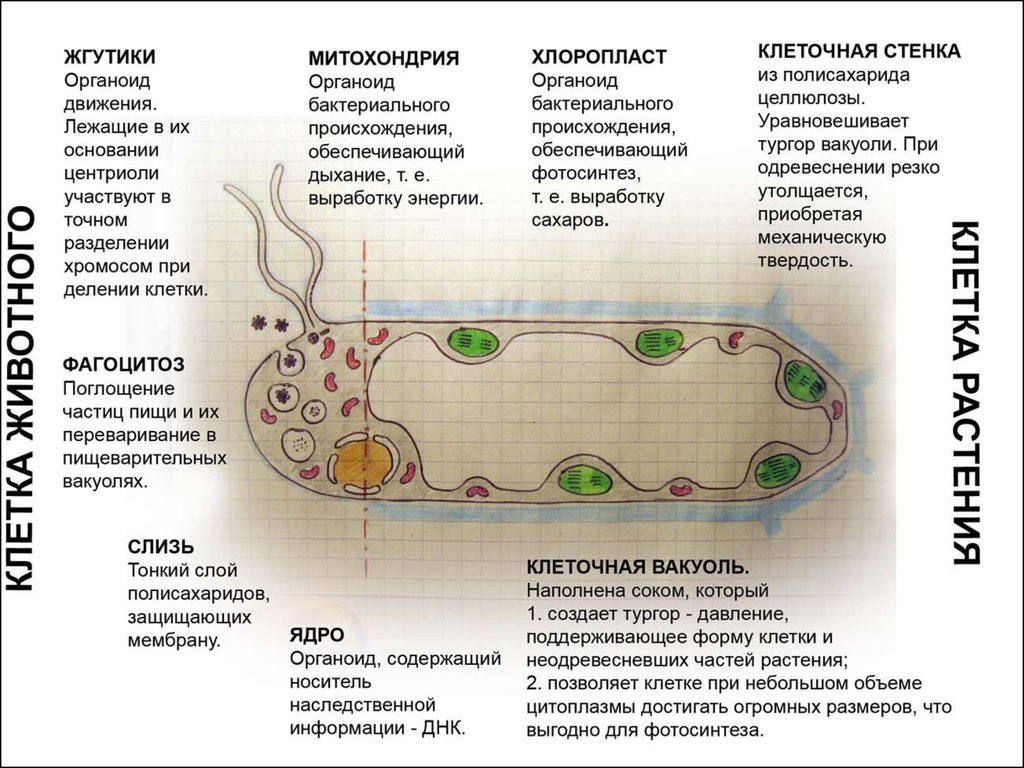
Изучение клеток разнообразных одноклеточных и многоклеточных организмов с помощью светооптического и электронного микроскопов показало, что по своему строению они разделяются на две группы. Одну группу составляют бактерии и сине-зеленые водоросли. Эти организмы имеют наиболее простое строение клеток. Их называют доеденными (прокариотами) , так как у них нет оформленного ядра (греч. -ядро) и нет многих структур, которые называют органоидами. Другую группу составляют все остальные организмы: от одноклеточных зеленых водорослей и простейших до высших цветковых растений, млекопитающих, в том числе и человека. Они имеют сложно устроенные клетки, которые называют ядерными (эукариотическими) . Эти клетки имеют ядро и органоиды, выполняющие специфические функции.
Особую, неклеточную форму жизни составляют вирусы, изучением которых занимается вирусология.
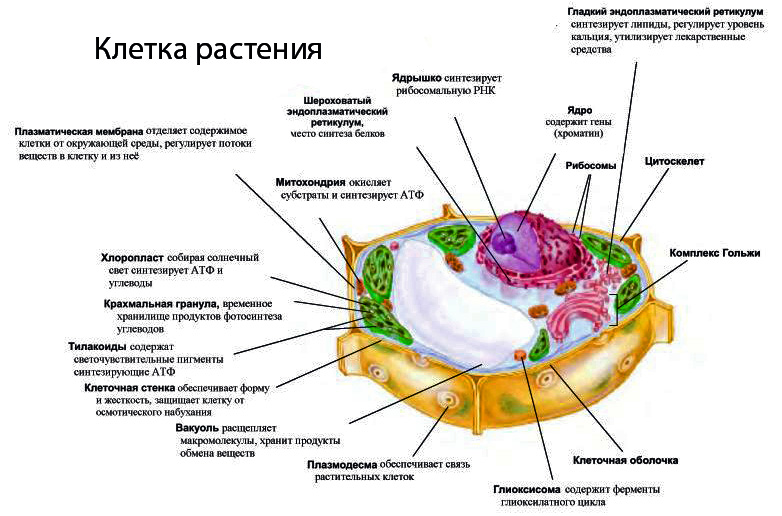
Строение и функции оболочки клетки Клетка любого организма, представляет собой целостную живую систему. Она состоит из трех неразрывно связанных между собой частей: оболочки, цитоплазмы и ядра.
 Оболочка клетка осуществляет непосредственное взаимодействие с внешней средой и взаимодействие с соседними клетками (в многоклеточных организмах) .
Оболочка клетка осуществляет непосредственное взаимодействие с внешней средой и взаимодействие с соседними клетками (в многоклеточных организмах) . Оболочка клеток. Оболочка клеток имеет сложное строение. Она состоит из наружного слоя и расположенной под ним плазматической мембраны. Клетки животных и растений различаются по строению их наружного слоя. У растений, а также у бактерий, сине-зеленых водорослей и грибов на поверхности клеток расположена плотная оболочка, или клеточная стенка. У большинства растений она состоит из клетчатки. Клеточная стенка играет исключительно важную роль: она представляет собой внешний каркас, защитную оболочку, обеспечивает тургор растительных клеток: через клеточную стенку проходит вода, соли, молекулы многих органических веществ.
Наружный слой поверхности клеток животных в отличие от клеточных стенок растений очень тонкий, эластичный. Он не виден в световой микроскоп и состоит из разнообразных полисахаридов и белков. Поверхностный слой животных клеток получил название гликокаликс.

Гликокаликс выполняет прежде всего функцию непосредственной связи клеток животных с внешней средой, со всеми окружающими ее веществами. Имея незначительную толщину (меньше 1 мкм) , наружный слой клетки животных не выполняет опорной роли, какая свойственна клеточным стенкам растений. Образование гликокаликса, так же как и клеточных стенок растений, происходит благодаря жизнедеятельности самих клеток.
Плазматическая мембрана. Под гликокаликсом и клеточной стенкой растений расположена плазматическая мембрана (лат. «мембрана>-кожица, пленка) , граничащая непосредственно с цитоплазмой. Толщина плазматической мембраны около 10 нм, изучение ее строения и функций возможно только с помощью электронного микроскопа.
В состав плазматической мембраны входят белки и липиды. Они упорядочено расположены и соединены друг с другом химическими взаимодействиями. По современным представлениям молекулы липидов в плазматической мембране расположены в два ряда и образуют сплошной слой. Молекулы белков не образуют сплошного слоя, они располагаются в слое липидов, погружаясь в него на разную глубину.
Молекулы белка и липидов подвижны, что обеспечивает динамичность плазматической мембраны.
Плазматическая мембрана выполняет много важных функций, от которых завидят жизнедеятельность клеток. Одна из таких функций заключается в том, что она образует барьер, отграничивающий внутреннее содержимое клетки от внешней среды. Но между клетками и внешней средой постоянно происходит обмен веществ. Из внешней среды в клетку поступает вода, разнообразные соли в форме отдельных ионов, неорганические и органические молекулы. Они проникают в клетку через очень тонкие каналы плазматической мембраны. Во внешнюю среду выводятся продукты, образованные в клетке. Транспорт веществ- одна из главных функций плазматической мембраны. Через плазматическую мембрану из клети выводятся продукты обмена, а также вещества, синтезированные в клетке. К числу их относятся разнообразные белки, углеводы, гормоны, которые вырабатываются в клетках различных желез и выводятся во внеклеточную среду в форме мелких капель.
Клетки, образующие у многоклеточных животных разнообразные ткани (эпителиальную, мышечную и др.) , соединяются друг с другом плазматической мембраной. В местах соединения двух клеток мембрана каждой из них может образовывать складки или выросты, которые придают соединениям особую прочность.
Соединение клеток растений обеспечивается путем образования тонких каналов, которые заполнены цитоплазмой и ограничены плазматической мембраной. По таким каналам, проходящим через клеточные оболочки, из одной клетки в другую поступают питательные вещества, ионы, углеводы и другие соединения.
На поверхности многих клеток животных, например, различных эпителиев, находятся очень мелкие тонкие выросты цитоплазмы, покрытые плазматической мембраной, микроворсинки. Наибольшее количество микроворсинок находится на поверхности клеток кишечника, где происходит интенсивное переваривание и всасывание переваренной пищи.
Фагоцитоз. Крупные молекулы органических веществ, например белков и полисахаридов, частицы пищи, бактерии поступают в клетку путем фагоцита (греч.
 «фагео» пожирать) . В фагоците непосредственное участие принимает плазматическая мембрана. В том месте, где поверхность клетки соприкасается с частицей какого-либо плотного вещества, мембрана прогибается, образует углубление и окружает частицу, которая в «мембранной упаковке» погружается внутрь клетки. Образуется пищеварительная вакуоль и в ней перевариваются поступившие в клетку органические вещества.
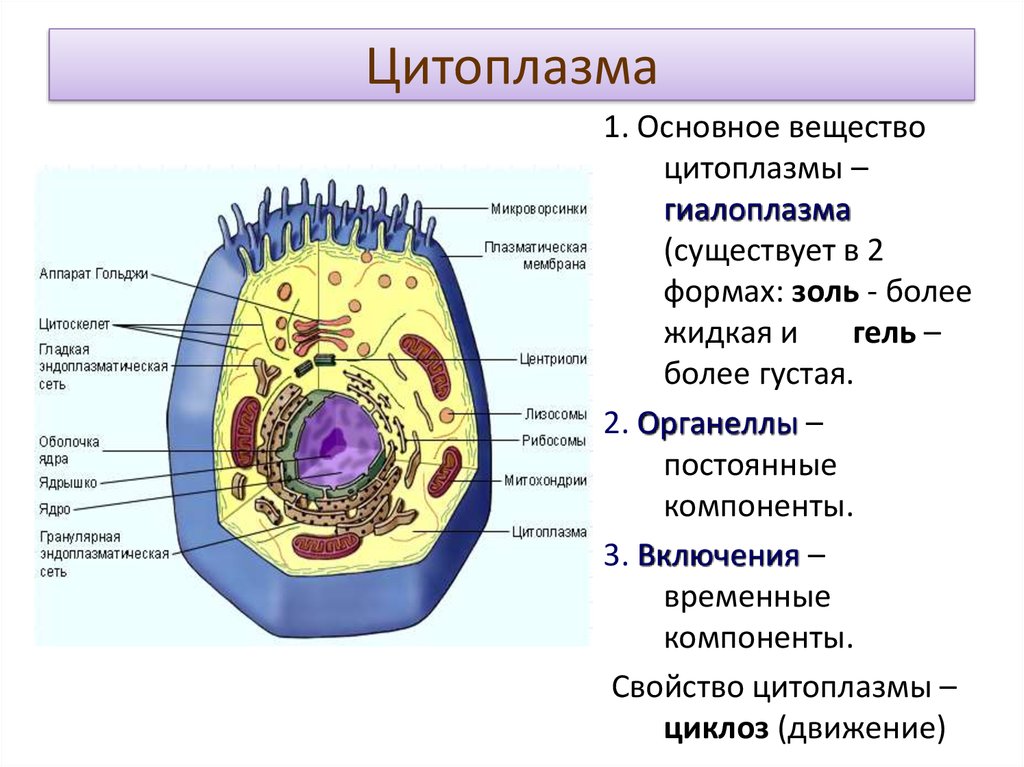
«фагео» пожирать) . В фагоците непосредственное участие принимает плазматическая мембрана. В том месте, где поверхность клетки соприкасается с частицей какого-либо плотного вещества, мембрана прогибается, образует углубление и окружает частицу, которая в «мембранной упаковке» погружается внутрь клетки. Образуется пищеварительная вакуоль и в ней перевариваются поступившие в клетку органические вещества. Цитоплазма. Отграниченная от внешней среды плазматической мембраной, цитоплазма представляет собой внутреннюю полужидкую среду клеток. В цитоплазму эукариотических клеток располагаются ядро и различные органоиды. Ядро располагается в центральной части цитоплазмы. В ней сосредоточены и разнообразные включения — продукты клеточной деятельности, вакуоли, а также мельчайшие трубочки и нити, образующие скелет клетки. В составе основного вещества цитоплазмы преобладают белки. В цитоплазме протекают основные процессы обмена веществ, она объединяет в одно целое ядро и все органоиды, обеспечивает их взаимодействие, деятельность клетки как единой целостной живой системы.
Эндоплазматическая сеть. Вся внутренняя зона цитоплазмы заполнена многочисленными мелкими каналами и полостями, стенки которых представляют собой мембраны, сходные по своей структуре с плазматической мембраной. Эти каналы ветвятся, соединяются друг с другом и образуют сеть, получившую название эндоплазматической сети.
Эндоплазматическая сеть неоднородна по своему строению. Известны два ее типа — гранулярная и гладкая. На мембранах каналов и полостей гранулярной сети располагается множество мелких округлых телец — рибосом, которые придают мембранам шероховатый вид. Мембраны гладкой эндоплазматической сети не несут рибосом на своей поверхности.
Эндоплазматическая сеть выполняет много разнообразных функций. Основная функция гранулярной эндоплазматической сети — участие в синтезе белка, который осуществляется в рибосомах.
На мембранах гладкой эндоплазматической сети происходит синтез липидов и углеводов. Все эти продукты синтеза накапливаются н каналах и полостях, а затем транспортируются к различным органоидам клетки, где потребляются или накапливаются в цитоплазме в качестве клеточных включений.
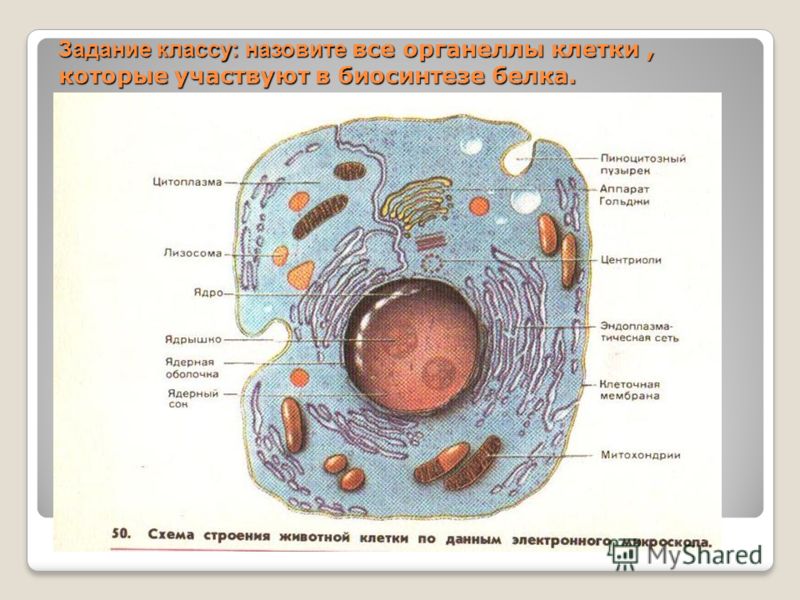
 Эндоплазматическая сеть связывает между собой основные органоиды клетки.
Эндоплазматическая сеть связывает между собой основные органоиды клетки. Рибосомы. Рибосомы обнаружены в клетках всех организмов. Это микроскопические тельца округлой формы диаметром 15-20 нм. Каждая рибосома состоит из двух неодинаковых по размерам частиц, малой и большой.
В одной клетке содержится много тысяч рибосом, они располагаются либо на мембранах гранулярной эндоплазматической сети, либо свободно лежат в цитоплазме. В состав рибосом входят белки и РНК. Функция рибосом — это синтез белка. Синтез белка сложный процесс, который осуществляется не одной рибосомой, а целой группой, включающей до нескольких десятков объединенных рибосом. Такую группу рибосом называют полисомой. Синтезированные белки сначала накапливаются в каналах и полостях эндоплазматической сети, а затем транспортируются к органоидам и участкам клетки, где они потребляютя. Эндоплазматическая сеть и рибосомы, расположенные на ее мембранах, представляют собой единый аппарат биосинтеза и транспортировки белков.

Митохондрии. В цитоплазме большинства клеток животных и растений содержатся мелкие тельца (0,2-7 мкм) — митохондрии (греч. — нить, — зерно, гранула) .
Митохондрии хорошо видны в световой микроскоп, с помощью которого можно рассмотреть их форму, расположение, сосчитать количество. Внутреннее строение митохондрий изучено с помощью электронного микроскопа. Оболочка митохондрии состоит из двух мембран — наружной и внутренней. Наружная мембрана гладкая, она не образует никаких складок и выростов. Внутренняя мембрана, напротив, образует многочисленные складки, которые направлены в полость митохондрии. Складки внутренней мембраны называют кристами (лат. — гребень, вырост) Число крист неодинаково в митохондриях разных клеток. Их может быть от нескольких десятков до нескольких сотен, причем особенно много крист в митохондриях активно функционирующих клеток, например мышечных.
Митохондрии называют клеток> так как их основная функция синтез аденозинтрифосфорной кислоты (АТФ) .
 Эта кислота синтезируется в митохондриях клеток всех организмов и представляет собой универсальный источник энергии, необходимый для осуществления процессов жизнедеятельности клетки и целого организма.
Эта кислота синтезируется в митохондриях клеток всех организмов и представляет собой универсальный источник энергии, необходимый для осуществления процессов жизнедеятельности клетки и целого организма. Новые митохондрии образуются делением уже существующих в клетке митохондрий.
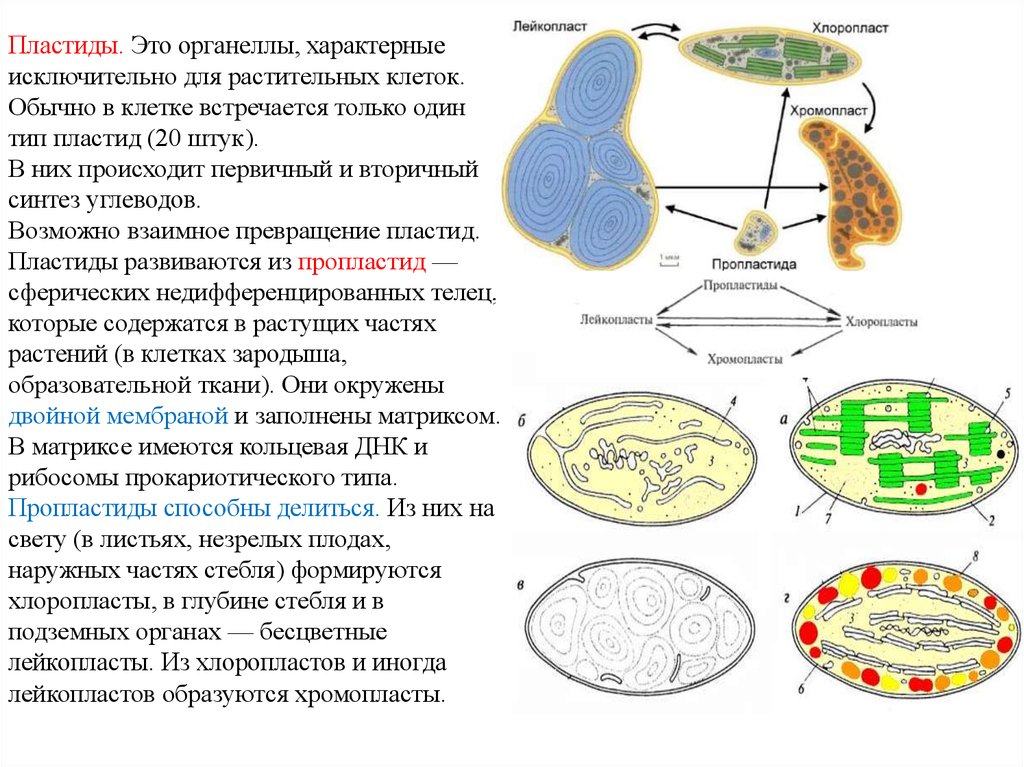
Пластиды. В цитоплазме клеток всех растений находятся пластиды. В клетках животных пластиды отсутствуют. Различают три основных типа пластид: зеленые — хлоропласты; красные, оранжевые и желтые — хромопласты; бесцветные — лейкопласты.
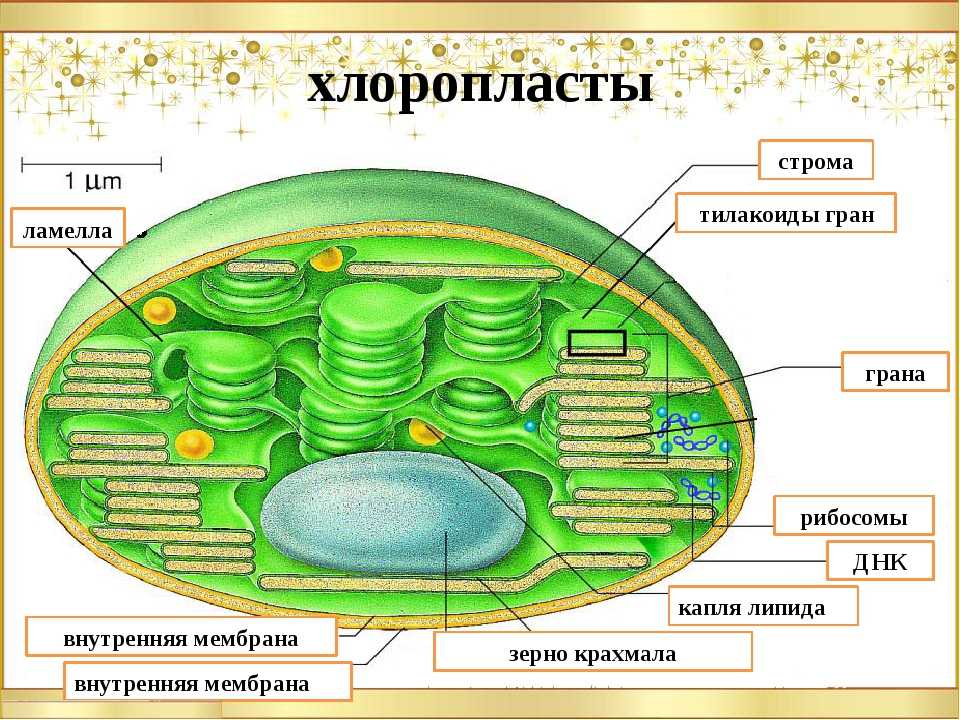
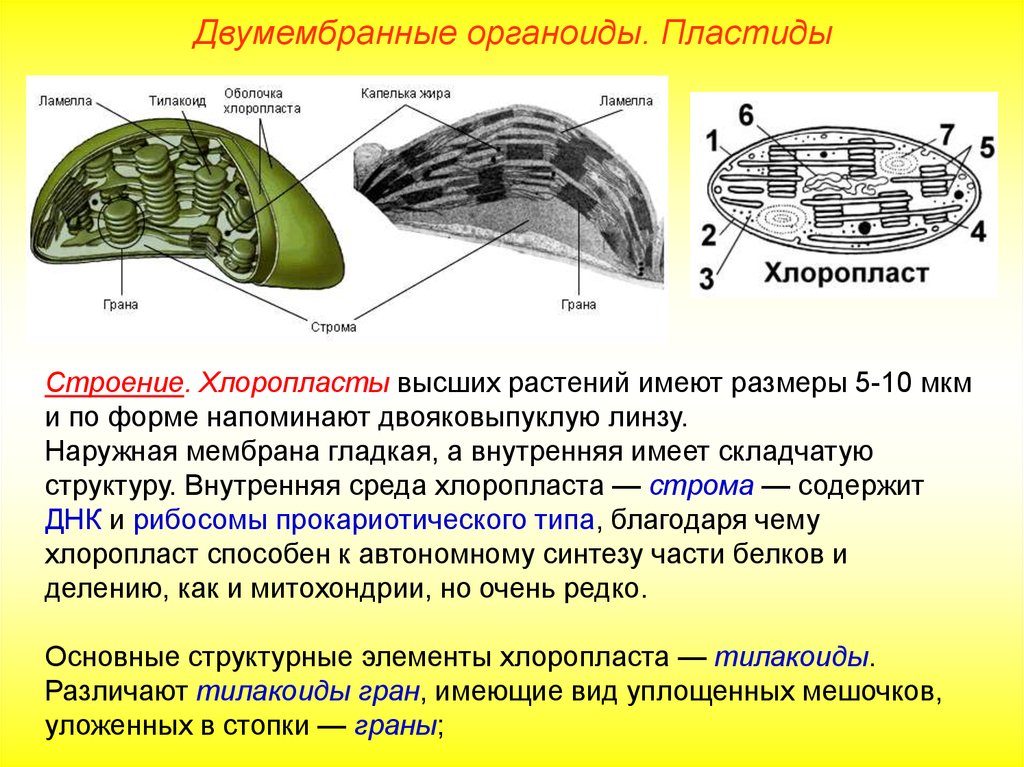
Хлоропласт. Эти органоиды содержатся в клетках листьев и других зеленых органов растений, а также у разнообразных водорослей. Размеры хлоропластов 4-6 мкм, наиболее часто они имеют овальную форму. У высших растений в одной клетке обычно бывает несколько десятков хлоропластов. Зеленый цвет хлоропластов зависит от содержания в них пигмента хлорофилла. Xлоропласт — основной органоид клеток растений, в котором происходит фотосинтез, т.е. образование органических веществ (углеводов) из неорганических (СО2 и Н2О) при использовании энергии солнечного света.

По строению хлоропласты сходны с митохондриями. От цитоплазмы хлоропласт отграничен двумя мембранами — наружной и внутренней. Наружная мембрана гладкая, без складок и выростов, а внутренняя образует много складчатых выростов, направленных внутрь хлоропласта. Поэтому внутри хлоропласта сосредоточено большое количество мембран, образующих особые структуры — граны. Они сложены наподобие стопки монет.
В мембранах гран располагаются молекулы хлорофилла, потому именно здесь происходит фотосинтез. В хлоропластах синтезируется и АТФ. Между внутренними мембранами хлоропласта содержатся ДНК, РНК и рибосомы. Следовательно, в хлоропластах, так же как и в митохондриях, происходит синтез белка, необходимого для деятельности этих органоидов. Хлоропласты размножаются делением.
Хромопласты находятся в цитоплазме клеток разных частей растений: в цветках, плодах, стеблях, листьях. Присутствием хромопластов объясняется желтая, оранжевая и красная окраска венчиков цветков, плодов, осенних листьев.
Лейкопласты. находятся в цитоплазме клеток неокрашенных частей растений, например в стеблях, корнях, клубнях. Форма лейкопластов разнообразна.
Хлоропласты, хромопласты и лейкопласты способны клетка взаимному переходу. Так при созревании плодов или изменении окраски листьев осенью хлоропласты превращаются в хромопласты, а лейкопласты могут превращаться в хлоропласты, например, при позеленении клубней картофеля.
Аппарат Гольджи. Во многих клетках животных, например, в нервных, он имеет форму сложной сети, расположенной вокруг ядра. В клетках растений и простейших аппарат Гольджи представлен отдельными тельцами серповидной или палочковидной формы. Строение этого органоида сходно в клетках растительных и животных организмов, несмотря на разнообразие его формы.
В состав аппарата Гольджи входят: полости, ограниченные мембранами и расположенные группами (по 5-10) ; крупные и мелкие пузырьки, расположенные на концах полостей. Все эти элементы составляют единый комплекс.

Аппарат Гольджи выполняет много важных функций. По каналам эндоплазматической сети к нему транспортируются продукты синтетической деятельности клетки — белки, углеводы и жиры. Все эти вещества сначала накапливаются, а затем в виде крупных и мелких пузырьков поступают в цитоплазму и либо используются в самой клетке в процессе ее жизнедеятельности, либо выводятся из нее и используются в организме. Например, в клетках поджелудочной железы млекопитающих синтезируются пищеварительные ферменты, которые накапливаются в полостях органоида. Затем образуются пузырьки, наполненные ферментами. Они выводятся из клеток в проток поджелудочной железы, откуда перетекают в полость кишечника. Еще одна важная функция этого органоида заключается в том, что на его мембранах происходит синтез жиров и углеводов (полисахаридов) , которые используются в клетке и которые входят в состав мембран. Благодаря деятельности аппарата Гольджи происходят обновление и рост плазматической мембраны.
Лизосомы. Представляют собой небольшие округлые тельца.
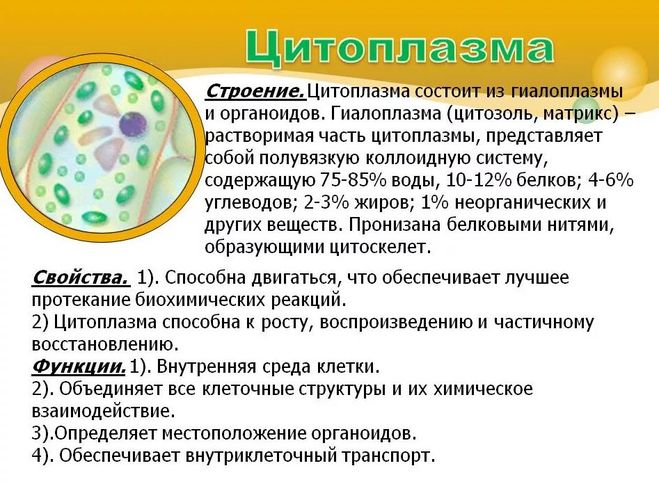 От Цитоплазмы каждая лизосома отграничена мембраной. Внутри лизосомы находятся ферменты, расщепляющие белки, жиры, углеводы, нуклеиновые кислоты.
От Цитоплазмы каждая лизосома отграничена мембраной. Внутри лизосомы находятся ферменты, расщепляющие белки, жиры, углеводы, нуклеиновые кислоты. К пищевой частице, поступившей в цитоплазму, подходят лизосомы, сливаются с ней, и образуется одна пищеварительная вакуоль, внутри которой находится пищевая частица, окруженная ферментами лизосом. Вещества, образовавшиеся в результате переваривания пищевой частицы, поступают в цитоплазму и используются клеткой.
Обладая способностью к активному перевариванию пищевых веществ, лизосомы участвуют в удалении отмирающих в процессе жизнедеятельности частей клеток, целых клеток и органов. Образование новых лизосом происходит в клетке постоянно. Ферменты, содержащиеся в лизосомах, как и всякие другие белки синтезируются на рибосомах цитоплазмы. Затем эти ферменты поступают по каналам эндоплазматической сети к аппарату Гольджи, в полостях которого формируются лизосомы. В таком виде лизосомы поступают в цитоплазму.
Клеточный центр. В клетках животных вблизи ядра находится органоид, который называют клеточным центром.
 Основную часть клеточного центра составляют два маленьких тельца — центриоли, расположенные в небольшом участке уплотненной цитоплазмы. Каждая центриоль имеет форму цилиндра длиной до 1 мкм. Центриоли играют важную роль при делении клетки; они участвуют в образовании веретена деления.
Основную часть клеточного центра составляют два маленьких тельца — центриоли, расположенные в небольшом участке уплотненной цитоплазмы. Каждая центриоль имеет форму цилиндра длиной до 1 мкм. Центриоли играют важную роль при делении клетки; они участвуют в образовании веретена деления. Клеточные включения. К клеточным включениям относятся углеводы, жиры и белки. Все эти вещества накапливаются в цитоплазме клетки в виде капель и зерен различной величины и формы. Они периодически синтезируются в клетке и используются в процессе обмена веществ.
Ядро. Каждая клетка одноклеточных и многоклеточных животных, а также растений содержит ядро. Форма и размеры ядра зависят от формы и размера клеток. В большинстве клеток имеется одно ядро, и такие клетки называют одноядерными. Существуют также клетки с двумя, тремя, с несколькими десятками и даже сотнями ядер. Это многоядерные клетки.
Ядерный сок — полужидкое вещество, которое находится под ядерной оболочкой и представляет внутреннюю среду ядра.
Химический состав клетки. Неорганические вещества Атомный и молекулярный состав клетки. В микроскопической клетке содержится несколько тысяч веществ, которые участвуют в разнообразных химических реакциях. Химические процессы, протекающие в клетке, — одно из основных условий ее жизни, развития и функционирования.
Все клетки животных и растительных организмов, а также микроорганизмов сходны по химическому составу, что свидетельствует о единстве органического мира.
Содержание химических элементов в клетке
Элементы Количество (в %)
Кислород 65-75
Кальций 0,04-2,00
Углерод 15-16
Магний 0,02-0,03
Водород 8-10
Натрий 0,02-0,03
Азот 1,5-3,0
Железо 0,01-0,015
Фосфор 0,2-1,0
Цинк 0,0003
Калий 0,15-0,4
Медь 0,0002
Сера 0,15-0,2
Йод 0,0001
Хлор 0,05-0,1
Фтор 0,0001
В таблице приведены данные об атомном составе клеток. Из 109 элементов периодической системы Менделеева в клетках обнаружено значительное их большинство.
 Особенно велико содержание в клетке четырех элементов — кислорода, углерода, азота и водорода. В сумме они составляют почти 98% всего содержимого клетки. Следующую группу составляют восемь элементов, содержание которых в клетке исчисляется десятыми и сотыми долями процента. Это сера, фосфор, хлор, калий, магний, натрий, кальций, железо. В сумме они составляют 1.9%. Все остальные элементы содержатся в клетке в исключительно малых количествах (меньше 0,01%) Таким образом, в клетке нет каких-нибудь особенных элементов, характерных только для живой природы. Это указывает на связь и единство живой и неживой природы. На атомном уровне различий между химическим составом органического и не органического мира нет. Различия обнаруживаются на более высоком уровне организации — молекулярном.
Особенно велико содержание в клетке четырех элементов — кислорода, углерода, азота и водорода. В сумме они составляют почти 98% всего содержимого клетки. Следующую группу составляют восемь элементов, содержание которых в клетке исчисляется десятыми и сотыми долями процента. Это сера, фосфор, хлор, калий, магний, натрий, кальций, железо. В сумме они составляют 1.9%. Все остальные элементы содержатся в клетке в исключительно малых количествах (меньше 0,01%) Таким образом, в клетке нет каких-нибудь особенных элементов, характерных только для живой природы. Это указывает на связь и единство живой и неживой природы. На атомном уровне различий между химическим составом органического и не органического мира нет. Различия обнаруживаются на более высоком уровне организации — молекулярном.Рефераты на русском языке — Медицина
Меню
| Главная |
| Новости |
| Краеведение История и современность Герои ВКО Почетные граждане Искусство Театр История театра Деятели театра Музыкальное искусство Народные композиторы Певцы и композиторы Изобразительное искусство Танцевальное искусство Киноискусство |
| Рефераты на русском |
| Рефераты на казахском |
| Литературная карта Усть-Каменогорск Катон-Карагай Курчум Тарбагатай Зайсан Улан Семей Глубокое Риддер Зыряновск Большенарым Аягуз Шемонаиха |
| Образцы документов |
Взаимодействующие органеллы — PMC
1. Хелле С.К., Канфер Г., Колар К., Ланг А., Мишель А.Х., Корнманн Б.: Организация и функция мест контакта с мембраной. Biochim Biophys Acta (2013) 1833 (11): 2526–2541. [PubMed] [Google Scholar]
Хелле С.К., Канфер Г., Колар К., Ланг А., Мишель А.Х., Корнманн Б.: Организация и функция мест контакта с мембраной. Biochim Biophys Acta (2013) 1833 (11): 2526–2541. [PubMed] [Google Scholar]
2. Айзенберг-Борд М., Шай Н., Шульдинер М., Бонерт М. Привязь есть привязь есть привязь: привязка в местах контакта с мембраной. Dev Cell (2016) 39 (4): 395–409. [PubMed] [Google Scholar]
3. Phillips MJ, Voeltz GK: Структура и функция участков контакта мембраны er с другими органеллами. Nat Rev Mol Cell Biol (2016) 17 (2): 69–82. [Бесплатная статья PMC] [PubMed] [Google Scholar]
4. Saheki Y, De Camilli P: Места контакта эндоплазматического ретикулума с плазматической мембраной. Annu Rev Biochem (2017) 86: 659–684. [PubMed] [Google Scholar]
5. Валм А.М., Коэн С., Легант В.Р., Мелунис Дж., Хершберг У., Уэйт Э., Коэн А.Р., Дэвидсон М.В., Бетциг Э., Липпинкотт-Шварц Дж.: Применение спектральных изображений и анализ для выявления интерактома органеллы. Природа (2017) 546 (7656): 162–167.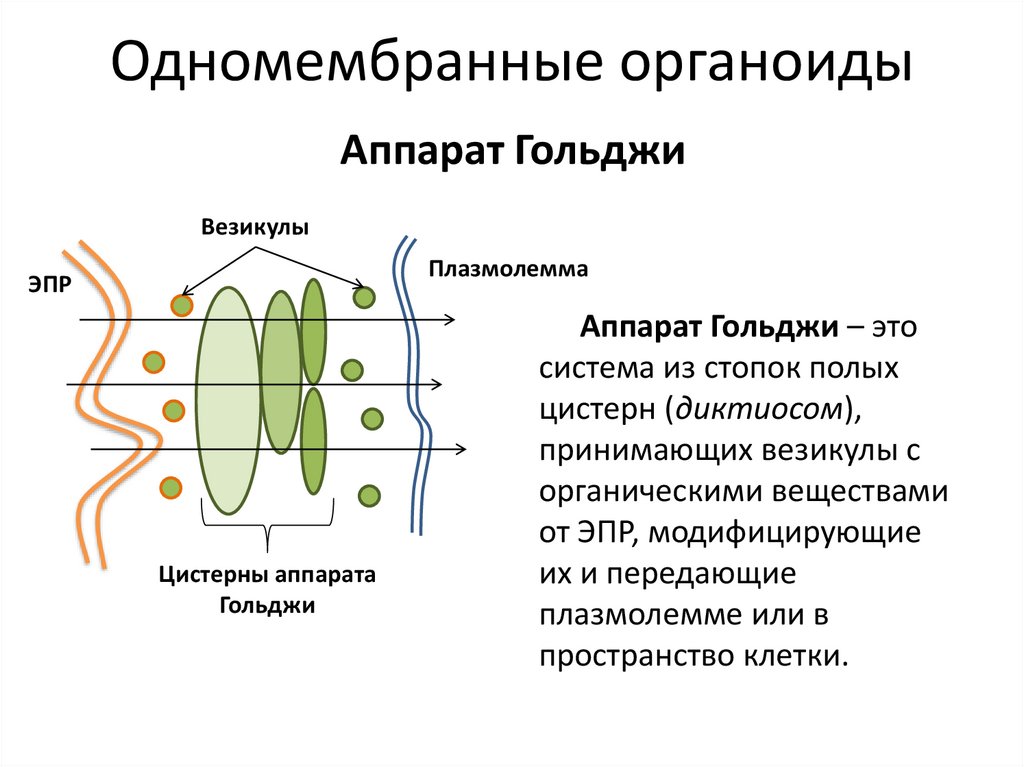 [Бесплатная статья PMC] [PubMed] [Google Scholar]
[Бесплатная статья PMC] [PubMed] [Google Scholar]
**Используя мультиспектральную визуализацию живых клеток, авторы характеризуют контакты между шестью органеллами и показывают, что картина контактов органелл стабильна.
6. Chang CL, Chen YJ, Liou J: соединения ER-плазматической мембраны: почему и как мы их изучаем? Biochim Biophys Acta (2017) 1864 (9): 1494–1506. [Бесплатная статья PMC] [PubMed] [Google Scholar]
7. Корнманн Б., Карри Э., Коллинз С.Р., Шульдинер М., Нуннари Дж., Вайсман Дж.С., Уолтер П.: Комплекс связывания ER-митохондрий, обнаруженный на экране синтетической биологии. Наука (2009 г.) 325(5939):477–481. [Бесплатная статья PMC] [PubMed] [Google Scholar]
8. Xue Y, Schmollinger S, Attar N, Campos OA, Vogelauer M, Carey MF, Merchant SS, Kurdistani SK: соединение эндоплазматического ретикулума и митохондрий необходимо для гомеостаза железа. . J Biol Chem (2017) 292 (32): 13197–13204. [Бесплатная статья PMC] [PubMed] [Google Scholar]
9. Szabadkai G, Bianchi K, Varnai P, De Stefani D, Wieckowski MR, Cavagna D, Nagy AI, Balla T, Rizzuto R: Опосредованное шапероном связывание эндоплазматического ретикулум и митохондриальные ca2+ каналы. J Cell Biol (2006) 175 (6): 901–911. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Szabadkai G, Bianchi K, Varnai P, De Stefani D, Wieckowski MR, Cavagna D, Nagy AI, Balla T, Rizzuto R: Опосредованное шапероном связывание эндоплазматического ретикулум и митохондриальные ca2+ каналы. J Cell Biol (2006) 175 (6): 901–911. [Бесплатная статья PMC] [PubMed] [Google Scholar]
10. De Vos KJ, Morotz GM, Stoica R, Tudor EL, Lau KF, Ackerley S, Warley A, Shaw CE, Miller CC: Vapb взаимодействует с митохондриальным белком ptpip51 для регулирования гомеостаза кальция. Hum Mol Genet (2012) 21 (6): 1299–1311. [Бесплатная статья PMC] [PubMed] [Google Scholar]
11. де Брито О.М., Скоррано Л.: Митофузин 2 привязывает эндоплазматический ретикулум к митохондриям. Природа (2008) 456 (7222): 605–610. [PubMed] [Google Scholar]
12. Наон Д., Занинелло М., Джакомелло М., Варанита Т., Греспи Ф., Лакшминаранаян С., Серафини А., Семензато М., Херкенне С., Эрнандес-Альварес М.И., Зорзано А.
и др.: Критическая переоценка подтверждает, что митофузин 2 является связующим звеном между эндоплазматическим ретикулумом и митохондриями. Proc Natl Acad Sci USA (2016) 113(40):11249–11254. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Proc Natl Acad Sci USA (2016) 113(40):11249–11254. [Бесплатная статья PMC] [PubMed] [Google Scholar]
13. Wang PT, Garcin PO, Fu M, Masoudi M, St-Pierre P, Pante N, Nabi IR: Различные механизмы, контролирующие шероховатые и гладкие контакты эндоплазматического ретикулума с митохондриями . J Cell Sci (2015) 128 (15): 2759–2765. [PubMed] [Google Scholar]
14. Филади Р., Греотти Э., Тураккио Г., Луини А., Поццан Т., Пиццо П. Абляция митофузина 2 увеличивает сцепление эндоплазматического ретикулума с митохондриями. Proc Natl Acad Sci USA (2015) 112(17):E2174–2181. [Бесплатная статья PMC] [PubMed] [Google Scholar]
15. Коссон П., Маркетти А., Раваццола М., Орчи Л.: Независимое сопоставление митофузина-2 эндоплазматического ретикулума и митохондрий: ультраструктурное исследование. PLoS One (2012) 7(9):e46293. [Бесплатная статья PMC] [PubMed] [Google Scholar]
16. Hirabayashi Y, Kwon SK, Paek H, Pernice WM, Paul MA, Lee J, Erfani P, Raczkowski A, Petrey DS, Pon LA, Polleux F: ER Связывание митохондрий с помощью pdzd8 регулирует динамику ca(2+) в нейронах млекопитающих.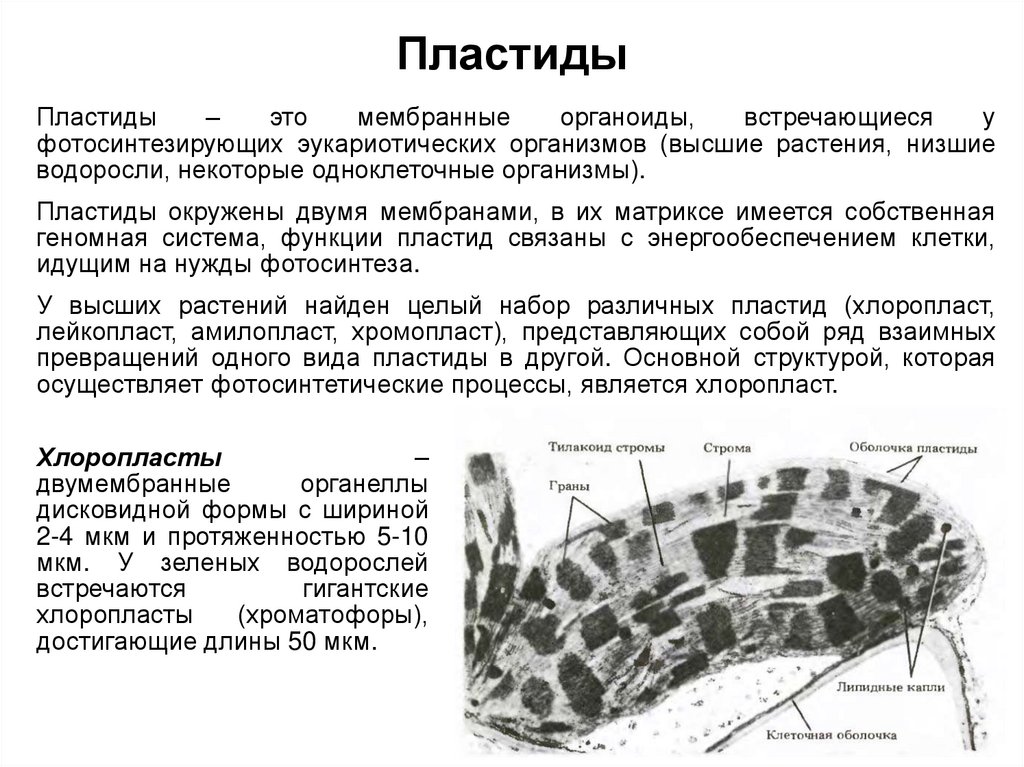 Наука (2017) 358 (6363): 623–630. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Наука (2017) 358 (6363): 623–630. [Бесплатная статья PMC] [PubMed] [Google Scholar]
*Авторы идентифицируют PDZD8 как белок, который регулирует связывание ER-митохондрий. Они показывают, что PDZD8 необходим для захвата митохондриями синаптически индуцированного Ca2+, высвобождаемого из ER.
17. Lahiri S, Chao JT, Tavassoli S, Wong AK, Choudhary V, Young BP, Loewen CJ, Prinz WA: Консервативный белковый комплекс мембраны эндоплазматического ретикулума (emc) облегчает перенос фосфолипидов из ER в митохондрии. PLoS Biol (2014) 12(10):e1001969. [Бесплатная статья PMC] [PubMed] [Google Scholar]
18. Markgraf DF, Klemm RW, Junker M, Hannibal-Bach HK, Ejsing CS, Rapoport TA: белок ER функционально связывает метаболизм нейтральных липидов в липидных каплях с мембранным синтезом липидов в er. Cell Rep (2014) 6 (1): 44–55. [Бесплатная статья PMC] [PubMed] [Google Scholar]
19. Gatta AT, Levine TP: Собираем воедино лоскутное одеяло из контактных сайтов. Тенденции Cell Biol (2017) 27(3):214–229.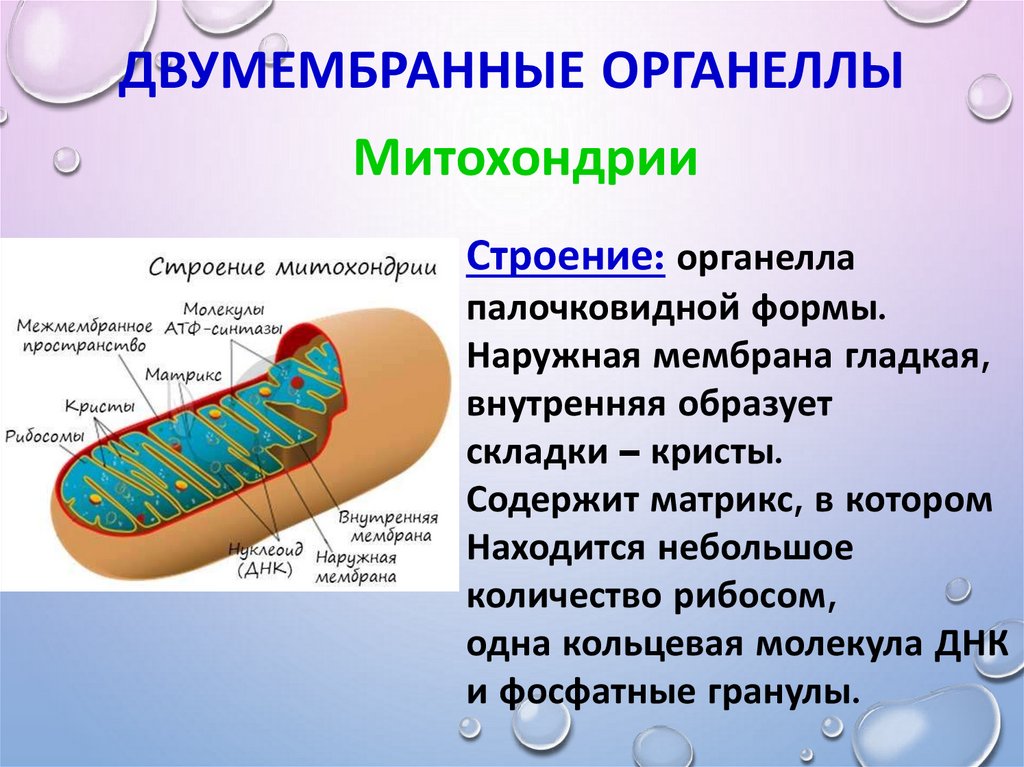 [PubMed] [Google Scholar]
[PubMed] [Google Scholar]
20. Xu N, Zhang SO, Cole RA, McKinney SA, Guo F, Haas JT, Bobba S, Farese RV Jr., Mak HY: Комплекс fatp1-dgat2 способствует расширению липидных капель при интерфейс ЭР-липидная капля. J Cell Biol (2012) 198(5):895–911. [Статья бесплатно PMC] [PubMed] [Google Scholar]
21. Рамбольд А.С., Коэн С., Липпинкотт-Шварц Дж.: Транспортировка жирных кислот в голодающих клетках: регуляция путем липолиза капель липидов, аутофагии и динамики слияния митохондрий. Dev Cell (2015) 32(6):678–692. [Бесплатная статья PMC] [PubMed] [Google Scholar]
22. Wang H, Sztalryd C: Окислительная ткань: Perilipin 5 связывает хранилище с печью. Trends Endocrinol Metab (2011) 22(6):197–203. [Бесплатная статья PMC] [PubMed] [Google Scholar]
23. Chu BB, Liao YC, Qi W, Xie C, Du X, Wang J, Yang H, Miao HH, Li BL, Song BL: транспорт холестерина через контакты лизосом-пероксисомной мембраны. Ячейка (2015) 161 (2): 291–306. [PubMed] [Google Scholar]
24. Пол А.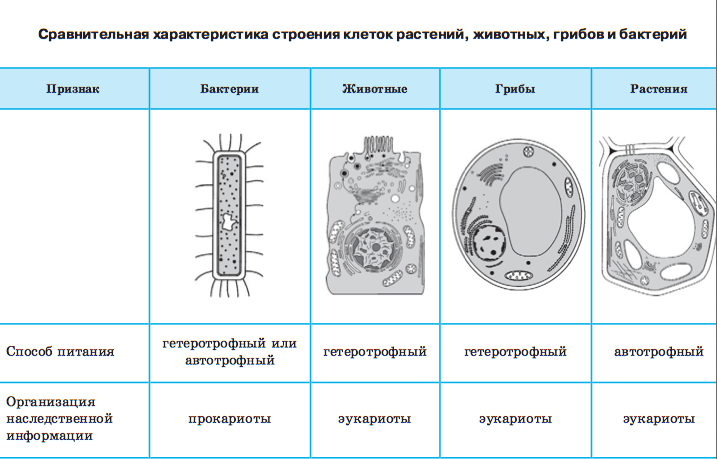 , Гросс С.П., Партон Р.Г.: Обзор: Биогенез многофункциональной липидной капли: липиды, белки и сайты. J Cell Biol (2014) 204(5):635–646. [Бесплатная статья PMC] [PubMed] [Google Scholar]
, Гросс С.П., Партон Р.Г.: Обзор: Биогенез многофункциональной липидной капли: липиды, белки и сайты. J Cell Biol (2014) 204(5):635–646. [Бесплатная статья PMC] [PubMed] [Google Scholar]
25. Walther TC, Chung J, Farese RV Jr.: Биогенез капель липидов. Annu Rev Cell Dev Biol (2017) 33 (49)1–510. [Бесплатная статья PMC] [PubMed] [Google Scholar]
26. Wilfling F, Wang H, Haas JT, Krahmer N, Gould TJ, Uchida A, Cheng JX, Graham M, Christiano R, Frohlich F, Liu X и др.: Ферменты синтеза триацилглицерина опосредуют рост липидных капель, перемещаясь из ER в липидные капли. Dev Cell (2013) 24 (4): 384–399. [Бесплатная статья PMC] [PubMed] [Google Scholar]
27. Wilfling F, Thiam AR, Olarte MJ, Wang J, Beck R, Gould TJ, Allgeyer ES, Pincet F, Bewersdorf J, Farese RV Jr., Walther TC : Механизм Arf1/copi действует непосредственно на липидные капли и делает возможным их соединение с er для нацеливания на белок. Elife (2014) 3(e01607. [Бесплатная статья PMC] [PubMed] [Google Scholar]
28. Salo VT, Belevich I, Li S, Karhinen L, Vihinen H, Vigouroux C, Magre J, Thiele C, Holtta-Vuori M, Jokitalo E, Ikonen E: Seipin регулирует контакты ER-липидных капель и доставку груза. EMBO J (2016) 35 (24): 2699–2716. [Статья бесплатно PMC] [PubMed] [Google Scholar]
Salo VT, Belevich I, Li S, Karhinen L, Vihinen H, Vigouroux C, Magre J, Thiele C, Holtta-Vuori M, Jokitalo E, Ikonen E: Seipin регулирует контакты ER-липидных капель и доставку груза. EMBO J (2016) 35 (24): 2699–2716. [Статья бесплатно PMC] [PubMed] [Google Scholar]
* Используя различные подходы к визуализации живых клеток, авторы показывают, что липиды и белки могут переноситься из ER в зрелые LD через сейпин-зависимые контакты.
29. Шульдинер М., Бонерт М.: Другой вид любви — места контакта липидных капель. Biochim Biophys Acta (2017) 1862 (10 Pt B): 1188–1196. [PubMed] [Google Scholar]
30. Rowland AA, Chitwood PJ, Phillips MJ, Voeltz GK: Контактные площадки ER определяют положение и время деления эндосомы. Ячейка (2014) 159 (5): 1027–1041. [Бесплатная статья PMC] [PubMed] [Google Scholar]
31. Lee JE, Westrate LM, Wu H, Page C, Voeltz GK: Несколько членов семейства динаминов сотрудничают, чтобы управлять делением митохондрий. Природа (2016) 540 (7631): 139–143. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[Бесплатная статья PMC] [PubMed] [Google Scholar]
32. Пернас Л., Скоррано Л.: Митоморфоз: митохондриальное слияние, деление и ремоделирование крист как ключевые медиаторы клеточной функции. Annu Rev Physiol (2016) 78: 505–531. [PubMed] [Академия Google]
33. Фридман Дж. Р., Лакнер Л. Л., Уэст М., ДиБенедетто Дж. Р., Нуннари Дж., Вольц Г. К.: канальцы ER отмечают участки митохондриального деления. Наука (2011) 334 (6054): 358–362. [Бесплатная статья PMC] [PubMed] [Google Scholar]
34. Коробова Ф., Рамабхадран В., Хиггс Х.Н.: Актин-зависимый этап деления митохондрий, опосредованный ER-ассоциированным формином inf2. Наука (2013) 339 (6118): 464–467. [Бесплатная статья PMC] [PubMed] [Google Scholar]
35. Manor U, Bartholomew S, Golani G, Christenson E, Kozlov M, Higgs H, Spudich J, Lippincott-Schwartz J: заякоренная в митохондриях изоформа актина Зародышеобразующий белок спирали регулирует деление митохондрий. Элиф (2015) 4:e08828. [Бесплатная статья PMC] [PubMed] [Google Scholar]
36. Chakrabarti R, Ji WK, Stan RV, de Juan Sanz J, Ryan TA, Higgs HN: Inf2-опосредованная полимеризация актина в er стимулирует поглощение митохондриальным кальцием, сужение внутренней мембраны и деление. J Cell Biol (2017). [Бесплатная статья PMC] [PubMed] [Google Scholar]
Chakrabarti R, Ji WK, Stan RV, de Juan Sanz J, Ryan TA, Higgs HN: Inf2-опосредованная полимеризация актина в er стимулирует поглощение митохондриальным кальцием, сужение внутренней мембраны и деление. J Cell Biol (2017). [Бесплатная статья PMC] [PubMed] [Google Scholar]
37. Wong YC, Ysselstein D, Krainc D: Контакты митохондрий и лизосом регулируют деление митохондрий посредством гидролиза rab7 gtp. Природа (2018) 554 (7692): 382–386. [Бесплатная статья PMC] [PubMed] [Google Scholar]
**Авторы показывают, что лизосомы играют роль в делении митохондрий. Это первый пример, показывающий, что две органеллы (лизосомы и ER) координируются, чтобы влиять на деление другой органеллы (митохондрии).
38. Helle SCJ, Feng Q, Aebersold MJ, Hirt L, Gruter RR, Vahid A, Sirianni A, Mostowy S, Snedeker JG, Saric A, Idema T и др.: Механическая сила вызывает деление митохондрий. Элиф (2017) 6:e30292. [Бесплатная статья PMC] [PubMed] [Google Scholar]
39. Galluzzi L, Baehrecke EH, Ballabio A, Boya P, Bravo-San Pedro JM, Cecconi F, Choi AM, Chu CT, Codogno P, Colombo MI, Cuervo ЯВЛЯЮСЬ
и др. : Молекулярные определения аутофагии и родственных процессов. EMBO J (2017) 36 (13): 1811–1836. [Бесплатная статья PMC] [PubMed] [Google Scholar]
: Молекулярные определения аутофагии и родственных процессов. EMBO J (2017) 36 (13): 1811–1836. [Бесплатная статья PMC] [PubMed] [Google Scholar]
40. Hailey DW, Rambold AS, Satpute-Krishnan P, Mitra K, Sougrat R, Kim PK, Lippincott-Schwartz J: Митохондрии поставляют мембраны для биогенеза аутофагосом во время голодания. Сотовый (2010) 141 (4): 656–667. [Бесплатная статья PMC] [PubMed] [Google Scholar]
41. Axe EL, Walker SA, Manifava M, Chandra P, Roderick HL, Habermann A, Griffiths G, Ktistakis NT: образование аутофагосом из мембранных компартментов, обогащенных фосфатидилинозитолом 3- фосфата и динамически связаны с эндоплазматическим ретикулумом. J Cell Biol (2008) 182(4):685–701. [Бесплатная статья PMC] [PubMed] [Google Scholar]
42. Хамасаки М., Фурута Н., Мацуда А., Незу А., Ямамото А., Фудзита Н., Оомори Х., Нода Т., Харагути Т., Хираока Ю., Амано А. и др.: Аутофагосомы образуются в местах контакта ER-митохондрий. Природа (2013) 495 (7441): 389–393. [PubMed] [Google Scholar]
43. Ravikumar B, Moreau K, Jahreiss L, Puri C, Rubinsztein DC: Плазматическая мембрана способствует образованию преаутофагосомных структур. Биология клеток природы (2010) 12 (8): 747–757. [Статья бесплатно PMC] [PubMed] [Google Scholar]
Ravikumar B, Moreau K, Jahreiss L, Puri C, Rubinsztein DC: Плазматическая мембрана способствует образованию преаутофагосомных структур. Биология клеток природы (2010) 12 (8): 747–757. [Статья бесплатно PMC] [PubMed] [Google Scholar]
44. Ge L, Melville D, Zhang M, Schekman R: Промежуточный компартмент ER-golgi является ключевым источником мембран для стадии липидизации lc3 биогенеза аутофагосом. Элиф (2013) 2:e00947. [Бесплатная статья PMC] [PubMed] [Google Scholar]
45. Guo Y, Chang C, Huang R, Liu B, Bao L, Liu W: Ap1 необходим для образования аутофагосом из сети транс-Гольджи. J Cell Sci (2012) 125 (Pt 7): 1706–1715. [PubMed] [Google Scholar]
46. Lamb CA, Yoshimori T, Tooze SA: Аутофагосома: происхождение неизвестно, комплекс биогенеза. Nat Rev Mol Cell Biol (2013) 14(12):759–774. [PubMed] [Google Scholar]
47. Gomez-Suaga P, Paillusson S, Stoica R, Noble W, Hanger DP, Miller CCJ: связывающий ER-митохондрии комплекс vapb-ptpip51 регулирует аутофагию. Curr Biol (2017) 27(3):371–385. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Curr Biol (2017) 27(3):371–385. [Бесплатная статья PMC] [PubMed] [Google Scholar]
48. Tripathi DN, Walker CL: Пероксисома как клеточная сигнальная органелла. Curr Opin Cell Biol (2016) 39: 109–112. [Бесплатная статья PMC] [PubMed] [Google Scholar]
49. Ким П. Биогенез пероксисом: союз двух органелл. Curr Biol (2017) 27(7):R271–R274. [PubMed] [Google Scholar]
50. Smith JJ, Aitchison JD: Пероксисомы обретают форму. Nat Rev Mol Cell Biol (2013) 14(12):803–817. [Бесплатная статья PMC] [PubMed] [Google Scholar]
51. Sugiura A, Mattie S, Prudent J, McBride HM: Новорожденные пероксисомы представляют собой гибрид митохондриальных и пре-пероксисом, полученных из эритроцитов. Природа (2017) 542 (7640): 251–254. [PubMed] [Академия Google]
**Используя элегантные анализы слияния клеток, авторы показывают, что везикулы, происходящие как из ER, так и из митохондрий, вносят вклад в биогенез пероксисом.
52. Soubannier V, McLelland GL, Zunino R, Braschi E, Rippstein P, Fon EA, McBride HM: везикулярный транспортный путь доставляет грузы из митохондрий в лизосомы.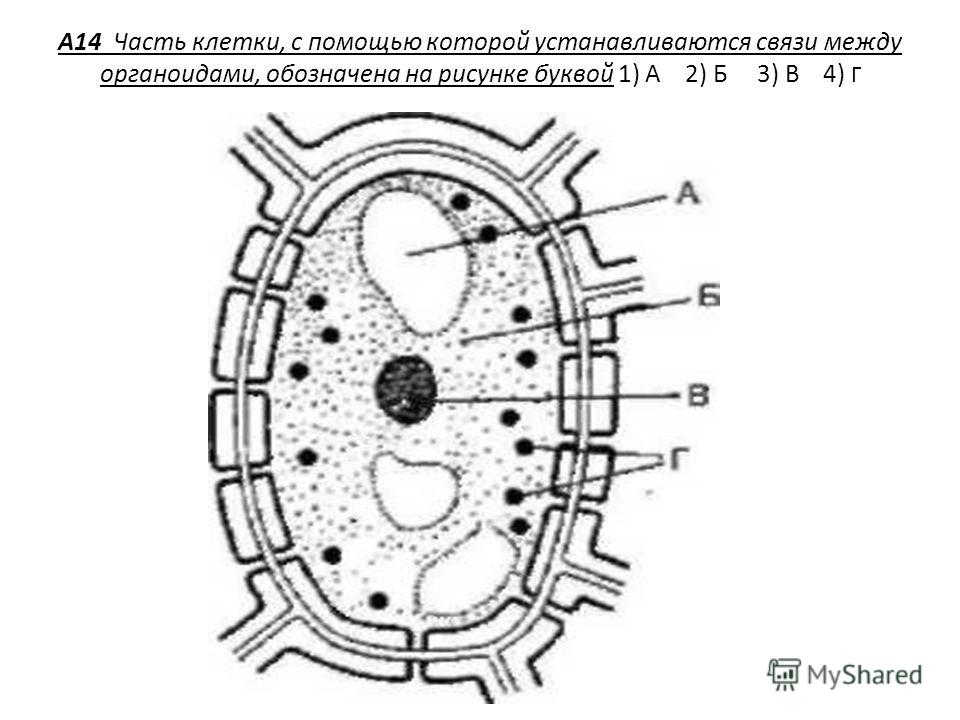 Curr Biol (2012) 22(2):135–141. [PubMed] [Google Scholar]
Curr Biol (2012) 22(2):135–141. [PubMed] [Google Scholar]
53. McFie PJ, Banman SL, Kary S, Stone SJ: Мышиная диацилглицерол-ацилтрансфераза-2 (dgat2) может катализировать синтез триацилглицерина и способствовать формированию липидных капель независимо от их локализации в эндоплазматическом ретикулуме. J Biol Chem (2011) 286(32):28235–28246. [Бесплатная статья PMC] [PubMed] [Google Scholar]
54. Rusinol AE, Cui Z, Chen MH, Vance JE: Уникальная митохондриально-ассоциированная мембранная фракция из печени крыс обладает высокой способностью к синтезу липидов и содержит секреторные белки пре-Гольджи, включая зарождающиеся липопротеины. J Biol Chem (1994) 269(44):27494–27502. [PubMed] [Google Scholar]
55. Wieckowski MR, Giorgi C, Lebedzinska M, Duszynski J, Pinton P: Изоляция митохондриально-ассоциированных мембран и митохондрий из тканей и клеток животных. Нат Проток (2009) 4 (11): 1582–1590. [PubMed] [Google Scholar]
56. Eden ER, White IJ, Tsapara A, Futter CE: Мембранные контакты между эндосомами и ER обеспечивают сайты для взаимодействия ptp1b-рецептора эпидермального фактора роста.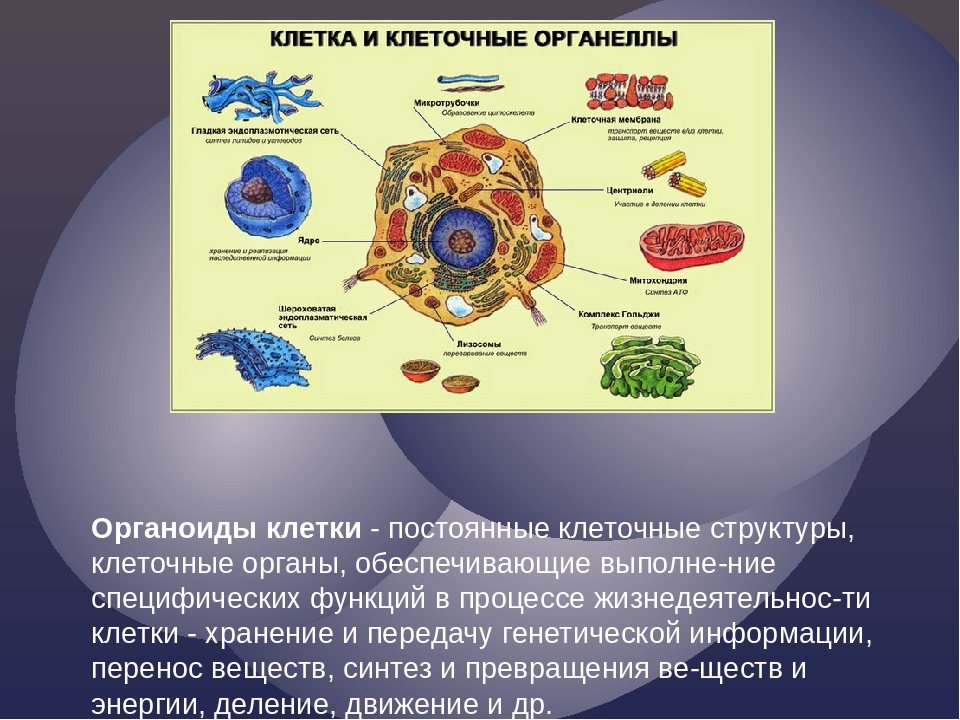 Nat Cell Biol (2010) 12(3):267–272. [PubMed] [Google Scholar]
Nat Cell Biol (2010) 12(3):267–272. [PubMed] [Google Scholar]
57. Costello JL, Castro IG, Hacker C, Schrader TA, Metz J, Zeuschner D, Azadi AS, Godinho LF, Costina V, Findeisen P, Manner A
и др.: Acbd5 и vapb опосредуют мембранные ассоциации между пероксисомами и ER. J Cell Biol (2017) 216 (2): 331–342. [Бесплатная статья PMC] [PubMed] [Google Scholar]
* Авторы используют пулл-даун и масс-спектрометрию для идентификации VAPA и VAPB на ER в качестве партнеров по связыванию пероксисомального белка ACBD5. Они показывают, что потеря ассоциации пероксисом-ER нарушает расширение и движение пероксисомной мембраны.
58. Hua R, Cheng D, Coyaud E, Freeman S, Di Pietro E, Wang Y, Vissa A, Yip CM, Fairn GD, Braverman N, Brumell JH
и др.: Vaps и acbd5 привязывают пероксисомы к ER для поддержания пероксисом и липидного гомеостаза. J Cell Biol (2017) 216 (2): 367–377. [Бесплатная статья PMC] [PubMed] [Google Scholar]
*Используя стратегию метки близости, авт. идентифицируют VAP на ER и ACBD5 на пероксисомах как часть связывающего комплекса ER-пероксисомы. Они показывают, что этот комплекс играет роль в подвижности пероксисом, росте и липидном гомеостазе.
Они показывают, что этот комплекс играет роль в подвижности пероксисом, росте и липидном гомеостазе.
59. Хунг В., Лам С.С., Удеши Н.Д., Свинкина Т., Гусман Г., Мутха В.К., Карр С.А., Тинг А.И.: Протеомное картирование обращенных к цитозолю наружных митохондриальных и ER мембран в живых клетках человека путем биотинилирования. Элиф (2017) 6: e24463. [Бесплатная статья PMC] [PubMed] [Google Scholar]
60. Портер К.Р., Палейд Г.Э.: Исследования эндоплазматического ретикулума. III. Его форма и распределение в поперечнополосатых мышечных клетках. J Biophys Biochem Cytol (1957) 3(2):269–300. [Бесплатная статья PMC] [PubMed] [Google Scholar]
61. Moore DH, Ruska H: Электронно-микроскопическое исследование клеток сердечной мышцы млекопитающих. J Biophys Biochem Cytol (1957) 3(2):261–268. [Бесплатная статья PMC] [PubMed] [Google Scholar]
62. Fernandez-Busnadiego R, Saheki Y, De Camilli P: Трехмерная архитектура расширенных синаптотагмин-опосредованных участков контакта эндоплазматического ретикулума с плазматической мембраной. Proc Natl Acad Sci USA (2015) 112(16):E2004–2013. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Proc Natl Acad Sci USA (2015) 112(16):E2004–2013. [Бесплатная статья PMC] [PubMed] [Google Scholar]
63. Охта К., Окаяма С., Того А., Накамура К.: Трехмерная организация мембраны эндоплазматического ретикулума вокруг места сужения митохондрий в клетках млекопитающих, выявленная с помощью томографии с фокусированным ионным пучком. Микроскопия (Oxf) (2014) 63 Suppl 1 (i34. [PubMed] [Google Scholar]
64. Wu Y, Whiteus C, Xu CS, Hayworth KJ, Weinberg RJ, Hess HF, De Camilli P: Контакты между эндоплазматическим ретикулумом и другими мембранами в нейронах. Proc. Национальная академия наук США (2017) 114 (24): E4859–E4867. [Бесплатная статья PMC] [PubMed] [Google Scholar]
** Используя FIB-SEM, авторы проводят подробный анализ мест контактов между ER и другими органеллами в нейронах.
65. Magliery TJ, Wilson CG, Pan W, Mishler D, Ghosh I, Hamilton AD, Regan L: Обнаружение белок-белковых взаимодействий с ловушкой повторной сборки зеленого флуоресцентного фрагмента белка: объем и механизм. J Am Chem Soc (2005) 127 (1): 146–157. [PubMed] [Google Scholar]
J Am Chem Soc (2005) 127 (1): 146–157. [PubMed] [Google Scholar]
66. Hu CD, Chinenov Y, Kerppola TK: Визуализация взаимодействий между белками семейства bzip и rel в живых клетках с использованием бимолекулярной флуоресцентной комплементации. Мол Селл (2002) 9(4): 789–798. [PubMed] [Google Scholar]
67. Bockler S, Westermann B: Митохондриальные контакты ER имеют решающее значение для митофагии у дрожжей. Dev Cell (2014) 28(4):450–458. [PubMed] [Google Scholar]
68. Хармон М., Ларкман П., Хардингем Г., Джексон М., Скехел П.: Бифлуоресцентная система комплементации для обнаружения связей между эндоплазматическим ретикулумом и митохондриями. Научный представитель (2017) 7 (1): 17467. [Бесплатная статья PMC] [PubMed] [Google Scholar]
69. Jing J, He L, Sun A, Quintana A, Ding Y, Ma G, Tan P, Liang X, Zheng X, Chen L, Shi X
и др.: Протеомное картирование соединений ER-pm идентифицирует стимат как регулятор притока ca(2)(+). Nat Cell Biol (2015) 17 (10): 1339–1347. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[Бесплатная статья PMC] [PubMed] [Google Scholar]
70. Benedetti L, Barentine AES, Messa M, Wheeler H, Bewersdorf J, De Camilli P: Светоактивируемое взаимодействие белков с высоким пространственным ограничением внутри клетки. Proc Natl Acad Sci USA (2018) 115(10):E2238–E2245. [Бесплатная статья PMC] [PubMed] [Google Scholar]
71. Shai N, Schuldiner M, Zalckvar E: Никакая пероксисома не является островом — места контакта пероксисом. Biochim Biophys Acta (2016) 1863 (5): 1061–1069. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Сотовая связь | Изучайте науку в Scitable
Изучение клеточной коммуникации сосредоточено на том, как клетка передает и получает сообщения с окружающей средой и самой собой. Действительно, клетки не живут изолированно. Их выживание зависит от получения и обработки информации из внешней среды, независимо от того, относится ли эта информация к наличию питательных веществ, изменениям температуры или уровням освещенности. Клетки также могут напрямую общаться друг с другом — и в ответ изменять свою внутреннюю работу — с помощью различных химических и механических сигналов. В многоклеточных организмах клеточная передача сигналов допускает специализацию групп клеток. Затем несколько типов клеток могут соединяться вместе, образуя ткани, такие как мышцы, кровь и ткань мозга. В одноклеточных организмах передача сигналов позволяет популяциям клеток координировать свои действия друг с другом и работать как команда для выполнения задач, которые ни одна клетка не может выполнить самостоятельно.
В многоклеточных организмах клеточная передача сигналов допускает специализацию групп клеток. Затем несколько типов клеток могут соединяться вместе, образуя ткани, такие как мышцы, кровь и ткань мозга. В одноклеточных организмах передача сигналов позволяет популяциям клеток координировать свои действия друг с другом и работать как команда для выполнения задач, которые ни одна клетка не может выполнить самостоятельно.
Изучение передачи сигналов клетками затрагивает несколько биологических дисциплин, таких как биология развития, нейробиология и эндокринология. Следовательно, актуальность клеточной коммуникации довольно велика, но основные области фундаментальных исследований часто делятся между изучением сигналов на клеточной мембране и изучением сигналов внутри и между внутриклеточными компартментами. Мембранная передача сигналов включает белки, сформированные в виде рецепторов, встроенных в клеточную мембрану, которые биофизически связывают триггеры во внешней среде с текущей динамической химией внутри клетки. Передача сигналов на мембране также включает ионные каналы, которые обеспечивают прямое прохождение молекул между внешними и внутренними компартментами клетки. Ученые спрашивают: какова структура рецептора, которая позволяет ему реагировать на внешний сигнал (например, на лиганд или даже на механическую силу)? Другие спрашивают: как после срабатывания сигнал обрабатывается внутри клетки?
Передача сигналов на мембране также включает ионные каналы, которые обеспечивают прямое прохождение молекул между внешними и внутренними компартментами клетки. Ученые спрашивают: какова структура рецептора, которая позволяет ему реагировать на внешний сигнал (например, на лиганд или даже на механическую силу)? Другие спрашивают: как после срабатывания сигнал обрабатывается внутри клетки?
Клетки развили множество сигнальных механизмов для передачи важной биологической информации. Некоторыми примерами этого разнообразия являются рецепторы, которые позволяют ионным потокам течь в ответ на фотоны, что эффективно переводит свет в химические мессенджеры внутри колбочек и палочек сетчатки; факторы роста, которые взаимодействуют с клеточной мембраной и могут запускать рецепторы, сильно влияющие на структуру хроматина и модуляцию экспрессии генов; метаболиты в крови, которые могут активировать клеточные рецепторы, вызывая высвобождение гормона, необходимого для регуляции уровня глюкозы; рецепторы адгезии, которые могут передавать силы, создаваемые напряжением, которые заставляют клетку оставаться на месте или изменять направление движения; и рецепторы, регулируемые развитием, которые могут строго направлять путь мигрирующей клетки, в конечном итоге контролируя, как весь организм связан вместе.
Как ученые изучают такую сложную сеть взаимодействий на стыке химии, физики и биологии? Один метод является редукционистским, при котором клетки выделяют и культивируют in vitro, чтобы можно было тщательно протестировать специфические сигналы с помощью химических веществ и измерить клеточные реакции. Другой более целостный метод включает измерение клеточной передачи сигналов в интактном организме ( in vivo ) путем применения специфических химических агентов, которые блокируют или активируют рецепторы в тщательно выбранной области ткани, а затем измеряют реакцию с помощью электрода, который передает активность ионных токов или путем забора жидкости из активированной области. Для обоих подходов жизненно важно измерение отклика, а измерение малых сотовых объектов действительно является сложной задачей. Ученые используют сложную покадровую микроскопию для отслеживания меченых молекул, которые перемещаются между субклеточными компартментами после сигнального события, или для отслеживания конформации рецептора, перешедшего из неактивного в активное состояние. Кроме того, методы масс-спектрометрии позволяют измерять пикомолярные количества, позволяя отслеживать внутриклеточные молекулы вторичных мессенджеров, которые имеют решающее значение для регуляции сигналов во внутриклеточной среде.
Кроме того, методы масс-спектрометрии позволяют измерять пикомолярные количества, позволяя отслеживать внутриклеточные молекулы вторичных мессенджеров, которые имеют решающее значение для регуляции сигналов во внутриклеточной среде.
Несмотря на технические достижения, глобальное понимание передачи сигнала, его внутренних иерархий, а также его высоко интегрированной и чрезвычайно динамичной природы остается в значительной степени загадочным. Потенциальный прорыв в этой области произошел недавно, когда ученые поняли, что существуют поразительные аналогии между сигнальными сетями в биологических системах и электронными схемами; оба они включают иерархии, переключатели, модульность, избыточность и наличие мощных механизмов обратной связи. Такая реализация дала толчок развитию области вычислительной биологии применительно к клеточной передаче сигналов. Сегодня изучение передачи клеточных сигналов не ограничивается биологами; благодаря вкладу инженеров и биофизиков ученые теперь могут создавать вычислительные алгоритмы, которые моделируют структуру сигнальной сети на основе биологических измерений, и эти модели можно использовать для прогнозирования результатов в физически невозможных экспериментальных условиях.
Leave A Comment