
Поступление веществ в клетки
Благодаря содержанию растворов солей, сахаров и других осмотически активных веществ, клетки характеризуются наличием в них определенного осмотического давления.
Например:
давление в клетках животных (морских и океанических форм) достигает 30 атм и более.
В оптимальных условиях осмотическое давление клеточного сока наземных органов болотных растений колеблется от 2 до 16 ат, у степных — от 8 до 40 ат. В разных клетках растения О. д. может резко различаться (так, у мангровых О. д. клеточного сока около 60 ат, а О. д. в сосудах ксилемы не превышает 1—2 ат). У гомойосмотических организмов, т. е. способных поддерживать относительное постоянство О. д., средней величины и диапазон колебаний О. д. различны (дождевой червь — 3,6—4,8 ат, пресноводные рыбы — 6,0—6,6, океанические костистые рыбы — 7,8—8,5, акуловые — 22,3—23,2, млекопитающие — 6,6—8,0 ат). У млекопитающих О. д. большинства биологических жидкостей равно О. д. крови (исключение составляют жидкости, выделяемые некоторыми железами, — слюна, пот, моча и др.). О. д., создаваемое в клетках животных высокомолекулярными соединениями (белки, полисахариды и др.), незначительно, но играет важную роль в обмене веществ
Градиент концентрации — разность концентрации веществ внутри и снаружи клетки
Поступление веществ в клетки животных, равно как и удаление их из клеток, связано с проницаемостью клеточной мембраны для молекул или ионов, а также со свойствами веществ. Клеточная мембрана регулирует обмен различными веществами между клеткой и средой. Поддержание мембраны и ее проницаемость обеспечиваются клеточной энергией.
Основные пути поступления веществ в клетки:
| пассивный транспорт | обеспечивается диффузией через мембрану по градиенту концентрации. Молекулы обычно переходят из области высокой концентрации в область более низкой концентрации. Вещества в клетку проникают через поры, имеющиеся в клеточной мембране. Не зависит от энергии, обеспечиваемой АТФ. | Этим путем в клетку проникают вода, двуокись углерода и молекулы органических веществ, способные хорошо растворяться в жирах | обеспечивают проникновение в клетки лишь малых молекул |
| катализируемый транспорт «облегченная диффузия» | Повышение скорости диффузии разных веществ, например, сахаров, аминокислот и нуклеозидов через мембрану под воздействием ферментов. Зависит от градиента концентрации. Является переносом по градиенту концентрации, она тоже непосредственно не зависит от энергии, обеспечиваемой АТФ. | Некоторые сахара, в частности, глюкоза | |
| активный транспорт | Перенос вещества против градиента концентрации, т. е. из области низкой концентрации в область более высокой концентрации. Связан со способностью мембраны поддерживать разность электрических потенциалов (помимо поддержания разности в концентрациях веществ внутри и снаружи клетки), под которыми понимают различия между электрическими потенциалами внутри и вне клетки, а также с затратами энергии на работу в виде перемещения веществ против электрохимического градиента, т. е. «вверх» Ферменты, катализирующие эти реакции | эффективен в случае переноса ионов | |
| эндоцитоз: | ответственен за поступление в клетки макромолекул (белков, по-линуклеотидов, полисахаридов) и разных твердых частиц, включая бактерии | ||
| фагоцитоз | процесс активного захватывания и поглощения живых и неживых частиц одноклеточными организмами или особыми клетками (фагоцитами) многоклеточных животных организмов. Обеспечивается путем выпячиваний своей клеточной мембраны и образования пузырьков, сливающихся затем с плазматической мембраной и открывающихся внутрь клетки. Вошедшие внутрь клеток частицы поступают в лизосомы, где с помощью клеточных (лизосомных) ферментов разрушаются и усваиваются затем клетками. | У многоклеточных (млекопитающих) выполняется специализированными клетками (лейкоцитами). У простейших является формой питания. Биологическое значение фагоцитоза у млекопитающих заключается в том, что он обеспечивает иммунную (фагоцитарную) защиту организма | |
| пиноцитоз | захват клеточной поверхностью жидкости с содержащимися в ней веществами путем впячиваний плазматической мембраны и образования пузырьков (канальцев), куда поступает жидкость. Канальцы после заполнения жидкостью отшнуровываются, поступают в цитоплазму и доходят до лизосом, где их стенки перевариваются, в результате чего содержимое (жидкость) канальцев освобождается и подвергается дальнейшей обработке лизосомными ферментами. | Один из основных механизмов проникновения в клетку высокомолекулярных соединений, в частности белков и углеводно-белковых комплексов. Наиболее активный П. наблюдается у амёб, в эпителиальных клетках кишечника и почечных канальцев, в эндотелии сосудов и растущих ооцитах, в клетках кровеносной и лимфатической систем, в клетках злокачественных опухолей, а также в клетках тканей, для которых характерен повышенный уровень обмена веществ. |
Экзоцитоз —процесс секретирования клетками различных веществ
Различают:
· регулируемый экзоцитоз
· конститутивный экзоцитоз
Клетки способны секретировать различные вещества в окружающую их среду. Этот процесс называют экзоцитозом.
Можно сказать, что пассивный транспорт веществ в клетки осуществляется обычной диффузией через клеточную мембрану, причем скорость диффузии вещества зависит от его растворимости в мембране, коэффициента диффузии в мембране и от разности концентрации веществ в клетке и за ее пределами (в среде).
Катализируемая, или так называемая «облегченная» диффузия, при которой скорость диффузии разных веществ, например, сахаров, аминокислот и нуклеозидов через мембрану повышается с помощью ферментов. Как и обычная, «облегченная» диффузия тоже зависит от градиента концентрации, однако здесь имеются подвижные «переносчики», роль которых выполняют ферменты. Находясь в составе мембраны, ферменты действуют в качестве «переносчиков» молекул веществ, проникая (диффундируя) на противоположную сторону мембраны, где они освобождаются от переносимых веществ. Поскольку «облегченная» диффузия веществ является переносом по градиенту концентрации, она тоже непосредственно не зависит от энергии, обеспечиваемой АТФ.
Примеры
Такие вещества, как глюкоза, почти нерастворимы в липидах, и размеры их частиц больше 0,8 нм; тем не менее они довольно быстро проходят через плазматическую мембрану, например через оболочку эритроцита. По-видимому, это обусловлено «облегченной диффузией» — присоединением к специфической молекуле-переносчику, представляющей собой пептид или белок. Молекула глюкозы G соединяется с молекулой-переносчиком X у наружной поверхности мембраны, и образовавшийся комплекс GX, растворимый в липидах, может диффундировать через мембрану к ее внутренней стороне, где он диссоциирует, и освобожденная глюкоза оказывается внутри клетки. Затем переносчик диффундирует обратно к наружной поверхности и сразу же может присоединить к себе другую молекулу глюкозы. При такой системе максимальная скорость переноса глюкозы определяется общим числом молекул-переносчиков, имеющихся в мембране, и возможными скоростями образования и расщепления комплекса GX. Для подобного процесса характерна «кинетика насыщения»: при малой концентрации глюкозы в наружном растворе скорость ее проникновения в клетку пропорциональна этой концентрации; однако при более высоких концентрациях пропорциональность исчезает, так как все молекулы переносчика уже «насыщены» глюкозой. Переносчики специфичны — они могут присоединять только глюкозу и некоторые очень близкие к ней по структуре сахара. Молекулы сахаров, сходные по своему химическому строению, будут конкурировать между собой за связывающие участки в молекулах-переносчиках.
Из клеточных мембран кишечной палочки Escherichia coli был выделен и частично очищен липопротеид, который, по-видимому, служит переносчиком, или пермеазой, для лактозы. На одну клетку приходится около 9000 молекул пермеазы. Как полагают, этот белок присоединяет лактозу у наружной поверхности мембраны, а затем образовавшийся комплекс диффундирует к ее внутренней поверхности, где лактоза отделяется от переносчика. Этот гипотетический механизм в принципе сходен с предполагаемым механизмом натриевого насоса.
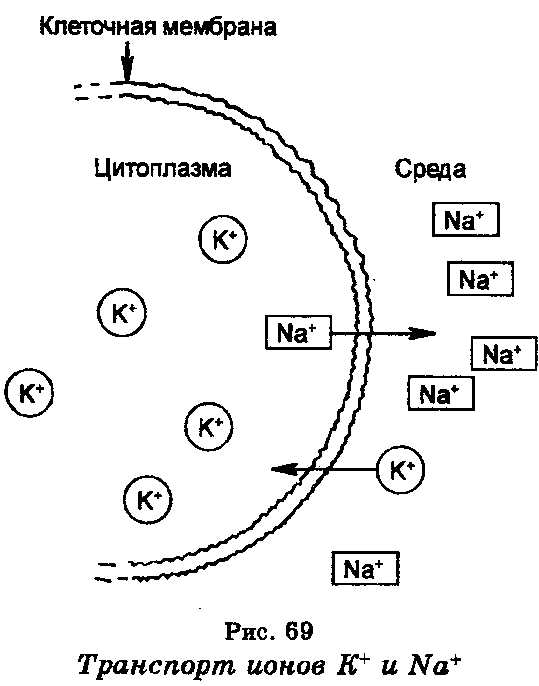
Активный транспорт.
Пример:
транспорт ионов натрия и калия, который определяет клеточный мембранный потенциал. Концентрация ионов натрия (Na+) внутри большинства клеток является меньшей, чем в среде, тогда как концентрация ионов калия (К+) внутри клеток является в 10—20 раз большей, чем в среде. В результате этого ионы Nа+ стремятся проникнуть из среды в клетку, а ионы К+, наоборот, выйти из клетки в среду. Поддержание концентрации этих ионов в клетке и в окружающей среде обеспечивается благодаря наличию в клеточной мембране системы, которая является ионным «насосом» и которая откачивает ионы Na+ из клетки в среду и накачивает ионы К+ в клетку из среды. Работа этой системы, т.е. движение ионов против электрохимического градиента, обеспечивается энергией, которая генерируется гидролизом АТФ, причем фермент АТФ-аза, катализирующий эту реакцию, содержится в самой мембране и, как считают, выполняет роль натриево-калиевого «насоса», генерирующего мембранный потенциал. Энергия, освобождаемая при гидролизе одной молекулы АТФ, обеспечивает транспорт за пределы клетки трех ионов Na+ и внутрь клетки двух ионов К+.
Система Na+ + К+ —АТФ-аза помогает поддерживать ассиметрическое распределение ионов калия при высокой концентрации последнего в клетках. Ионы калия участвуют в регуляции многих клеточных функций, включая поток солей и воды из почечных клеток, освобождение инсулина из панкреатических клеток, частоту сердцебиений.
Установлено, что энергетически выгодный транспорт ионов Na+ внутрь клеток оказывает также влияние на транспорт сахаров и аминокислот в клетки. В частности, с транспортом ионов Na+ сопряжен транспорт глюкозы. Чтобы создать градиент концентрации ионов Na+, благоприятный для транспорта ионов К+ и глюкозы внутрь клеток, ионная «насосная» система благодаря энергии активно откачивает ионы Na+ из клетки за ее пределы.
Определенная роль в транспорте веществ принадлежит белоксвязывающим системам, представляющим четвертый способ транспорта. Речь идет о белках, локализованных в периплазматическом пространстве. Эти белки специфически связывают сахара, аминокислоты и ионы, перенося их затем к специфическим молекулам-носителям, локализованным в клеточной мембране. Источником энергии для этих систем является АТФ.
Экзоцитоз
Конститутивный экзоцитоз осуществляется периодически, по мере накоплениях секрета, без видимого воздействия внеклеточных факторов, при колебаниях гидратированности клетки, без участия аппарата Гольджи и Са2+—независимо (Инсулин и ряд других гормонов, нейротрансмиттеров и многих ферментов.)
Нейромедиатор (Neurotransmitter) — химический посредник, освобождающийся из пресинаптического нервного окончания и передающий нервный импульс в синапсе постсинаптичсскому окончанию, мышечному волокну или железе, которые эти нервы иннервируют. Основными нейромедиаторами в периферической нервной системе являются ацетилхолин и норадреналин (сскретируются нервными окончаниями симпатической нервной системы). В центральной нервной системе наряду с ацетилхолином и норадреналином, нейромедиаторами являются дофамин, серотонин, гаммааминобутировая кислота и некоторые другие вещества.
Регулируемый экзоцитоз отличается от предыдущего необходимостью для запуска внеклеточного воздействия и четкой стадийностью процесса. Каждая из стадий регулируется дифференцированно. Специфика экзоцитоза определяется химической природой выделяеммых веществ, невезикулярной или везикулярной формами его выведения.
С помощью экзоцитоза из клетки удаляются также частицы, оказавшиеся непереваренными путем фагоцитоза. У большинства клеток циклы эндоцитоз-экзоцитоз непрерывны.
Фотосинтез
Фотосинтез — это синтез органических соединений в листьях зеленых растений из воды и углекислого газа атмосферы с использованием солнечной (световой) энергии, адсорбируемой хлорофиллом в хлоропластах.
Благодаря фотосинтезу происходит улавливание энергии видимого света и превращение ее в химическую энергию, сохраняемую (запасаемую) в органических веществах, образуемых при фотосинтезе.
Датой открытия процесса фотосинтеза можно считать 1771 г. Английский ученый Дж. Пристли обратил внимание на изменение состава воздуха вследствие жизнедеятельности животных. В присутствии зеленых растений воздух вновь становился пригодным как для дыхания, так и для горения. В дальнейшем работами ряда ученых (Я. Ингенгауз, Ж. Сенебье, Т. Соссюр, Ж.Б. Буссенго) было установлено, что зеленые растения из воздуха поглощают С02, из которого при участии воды на свету образуется органическое вещество. Именно этот процесс в 1877 г. немецкий ученый В. Пфеффер назвал фотосинтезом. Большое значение для раскрытия сущности фотосинтеза имел закон сохранения энергии, сформулированный Р. Майером. В 1845 г. Р. Майер выдвинул предположение, что энергия, используемая растениями, — это энергия Солнца, которую растения в процессе фотосинтеза превращают в химическую энергию. Это положение было развито и экспериментально подтверждено в исследованиях замечательного русского ученого К.А. Тимирязева.
Основная роль фотосинтезирующих организмов:
1) трансформация энергии солнечного света в энергию химических связей органических соединений;
2) насыщение атмосферы кислородом;
В результате фотосинтеза на Земле образуется 150 млрд. т. органического вещества и выделяется около 200 млрд. т свободного кислорода в год. Он препятствует увеличению концентрации CO2 в атмосфере, предотвращая перегрев Земли (парниковый эффект).
Созданная фотосинтезом атмосфера защищает живое от губительного коротковолнового УФ-излучения (кислородно-озоновый экран атмосферы).
В урожай сельскохозяйственных растений переходит лишь 1-2% солнечной энергии, потери обусловлены неполным поглощением света. Поэтому имеется огромная перспектива повышения урожайности благодаря селекции сортов с высокой эффективностью фотосинтеза, созданию благоприятной для светопоглощения структуры посевов. В связи с этим особенно актуальными становятся разработка теоретических основ управления фотосинтезом
Значение фотосинтеза гигантское. Отметим лишь, что он поставляет топливо (энергию) и атмосферный кислород, необходимые для существования всего живого. Следовательно, роль фотосинтеза является планетарной.
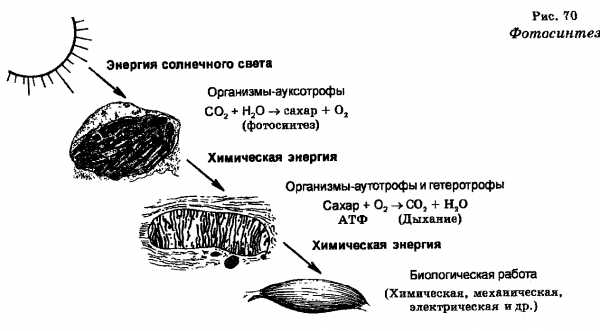
Планетарность фотосинтеза определяется также тем, что благодаря круговороту кислорода и углерода (в основном) поддерживается современный состав атмосферы, что в свою очередь определяет дальнейшее поддержание жизни на Земле. Можно сказать далее, что энергия, которая запасается в продуктах фотосинтеза, есть по существу основной источник энергии, которым сейчас располагает человечество.
Похожие статьи:
poznayka.org
Способы поступления питательных веществ в клетку
Питание бактерий
Бактерии представляют обширную группу мельчайших и в большинстве одноклеточных организмов. Основными элементами, влияющими на рост и развитие бактерий, являются: азот, углерод, кислород, водород, фосфор, сера, которые входят в состав белков, углеводов, жиров, нуклеиновых кислот. Эти элементы требуются в значительных количествах – макроэлементы. Микроэлементы, необходимые в микромолярных количествах, — это ионы таких металлов, как хром, кобальт, медь, цинк, молибден, марганец, никель, селен, вольфрам, ванадий, обычно входящие в состав ферментов и кофакторов.
Способы поступления питательных веществ в клетку
Поступление веществ в клетку и выделение продуктов обмена в окружающую среду происходит у микроорганизмов через всю поверхность тела. У микроорганизмов очень большая по сравнению с объемом всасывающая пищу поверхность клетки, что обусловливает весьма активный обмен веществ. Поступление питательных веществ в клетку сложный процесс.
Возможность проникновения веществ извне в клетку обусловлена многими факторами: величиной и структурой их молекул; способностью растворяться в компонентах цитоплазматической мембраны или вступать с ними в химические соединения; концентрацией веществ в клетке и в среде; электрическим зарядом клетки и другое.
Вещества питательной среды могут поступать в клетку только в растворенном состоянии. Нерастворимые сложные органические соединения должны подвергнуться расщеплению на более простые вне клетки, что происходит с помощью экзоферментов микроорганизмов.
Клеточная стенка проницаема и задерживает лишь макромолекулы. Цитоплазматическая мембрана обладает полупроницаемостью. Она служит осмотическим барьером, проницаемость её для различных веществ неодинакова.
Наиболее известны два пути проникновения веществ в клетку: осмос и адсорбция (специфический перенос). Активная роль в этих процессах принадлежит цитоплазматической мембране.
О с м о с представляет собой диффузию веществ в растворах через полупроницаемую перепонку (мембрану). Как известно, через такие мембраны могут диффундировать вещества, находящиеся в состоянии истинных растворов. Возникает осмос под действием разности осмотических давлений в растворах по обе стороны полупроницаемой мембраны.
Величина осмотического давления раствора зависит от молярной концентрации растворённых в нём веществ. В относительно слабых растворах осмотическое давление изменяется пропорционально концентрации растворённых веществ. Чем больше разность осмотических давлений (концентраций растворов) по обе стороны полупроницаемой мембраны, тем с большей интенсивностью осмотирует растворитель (вода) в раствор с бо¢льшим осмотическим давлением. Осмотируют растворённые в воде вещества; при этом каждое диффундирует в соответствии с его собственным (парциальным) осмотическим давлением, то есть в раствор с его меньшей концентрацией.
Оболочка клетки проницаема и задерживает лишь макромолекулы. Цитоплазматическая мембрана клетки обладает полупроницаемостью; она является осмотическим барьером, регулируя поступление в клетку и выход из неё растворённых веществ. Вещества, не растворимые в воде или образующие коллоидные растворы (например, белки, крахмал), непосредственно не могут быть использованы клеткой. Они могут проникнуть в неё лишь после расщепления на более простые вне клетки, что и происходит в среде с помощью экзоферментов микробов.
Таким образом, при осмотическом проникновении питательных веществ в клетку движущей силой служит разность осмотических давлений между средой и клеткой. Такой пассивный перенос веществ не требует затраты энергии и протекает до выравнивания концентрации с наружным раствором. Поскольку поступившие в клетку вещества включаются в реакции конструктивного и энергетического обмена, концентрация некоторых из них будет ниже, чем в среде, и поступление данных веществ возможно до полного исчерпания их из субстрата.
Если осмотическое давление микробных клеток, обусловленное растворёнными в клеточном соке веществами, несколько выше, чем в среде, то за счёт притока из неё воды в клетке создаётся определённое упругое напряжение, называемое тургором. Протопласт клетки при этом прижимается к клеточной оболочке, слегка растягивая её.
Если микроорганизм попадает в субстрат, осмотическое давление которого выше, чем в клетке, то цитоплазма отдает воду во внешнюю среду. Питательные вещества в клетку не поступают, содержимое клетки уменьшается в объёме, и протопласт отстаёт от клеточной оболочки. Это явление называется плазмолизом клетки.
При чрезмерно низком осмотическом давлении внешней среды может наступить плазмоптис клетки – явление, обратное плазмолису, когда вследствие высокой разности осмотических давлений цитоплазма быстро переполняется водой. Это может привести к разрыву клеточной оболочки, что наблюдается, например, при помещении бактерий в дистиллированную воду.
Второй путь поступления веществ в клетку – активный – путём переноса их особыми, локализованными в цитоплазматической мембране веществами ферментной природы. Эти переносчики, называемые пермеазами, обладают субстратной специфичностью. Каждый транспортирует только определённое вещество, имеющее сходную с белком-переносчиком стереохимическую структуру молекулы. На внешней стороне цитоплазматической мембраны переносчик адсорбирует вещество – вступает с ним во временную связь и диффундирует комплексно через мембрану, отдавая на внутренней стороне её транспортируемое вещество в цитоплазму. Вещество может поступать и тогда, когда концентрация его в клетке больше, чем в среде. При таком переносе веществ затрачивается энергия. При этом транспортируемое вещество может подвергнуться изменению, например из не растворимого в мембране переходит в растворимое состояние.
Цитоплазматическая мембрана, таким образом, является не только осмотическим барьером, но и обладает избирательной проницаемостью.
Структурные компоненты клетки, энергетический заряд
Ядерная оболочка клетки
Ядрышко, хроматин, элементы, цитозоль, цитоскелет, центриоли, митохондрии, пероксисомы, микротрубочки, органеллы клетки
Взаимодействие различных органоидов клетки
Функции клеточной стенки, лизосом, рибосом, комплекса гольджи, вакуоли
biofile.ru
Поступление веществ в клетки
Благодаря содержанию растворов солей, сахаров и других осмотически активных веществ, клетки характеризуются наличием в них определенного осмотического давления.
Например:
давление в клетках животных (морских и океанических форм) достигает 30 атм и более.
В оптимальных условиях осмотическое давление клеточного сока наземных органов болотных растений колеблется от 2 до 16 ат, у степных — от 8 до 40 ат. В разных клетках растения О. д. может резко различаться (так, у мангровых О. д. клеточного сока около 60 ат, а О. д. в сосудах ксилемы не превышает 1—2 ат). У гомойосмотических организмов, т. е. способных поддерживать относительное постоянство О. д., средней величины и диапазон колебаний О. д. различны (дождевой червь — 3,6—4,8 ат, пресноводные рыбы — 6,0—6,6, океанические костистые рыбы — 7,8—8,5, акуловые — 22,3—23,2, млекопитающие — 6,6—8,0 ат). У млекопитающих О. д. большинства биологических жидкостей равно О. д. крови (исключение составляют жидкости, выделяемые некоторыми железами, — слюна, пот, моча и др.). О. д., создаваемое в клетках животных высокомолекулярными соединениями (белки, полисахариды и др.), незначительно, но играет важную роль в обмене веществ
Градиент концентрации — разность концентрации веществ внутри и снаружи клетки
Поступление веществ в клетки животных, равно как и удаление их из клеток, связано с проницаемостью клеточной мембраны для молекул или ионов, а также со свойствами веществ. Клеточная мембрана регулирует обмен различными веществами между клеткой и средой. Поддержание мембраны и ее проницаемость обеспечиваются клеточной энергией.
Основные пути поступления веществ в клетки:
пассивный транспорт | обеспечивается диффузией через мембрану по градиенту концентрации. Молекулы обычно переходят из области высокой концентрации в область более низкой концентрации. Вещества в клетку проникают через поры, имеющиеся в клеточной мембране. Не зависит от энергии, обеспечиваемой АТФ. | Этим путем в клетку проникают вода, двуокись углерода и молекулы органических веществ, способные хорошо растворяться в жирах | обеспечивают проникновение в клетки лишь малых молекул |
катализируемый транспорт «облегченная диффузия» | Повышение скорости диффузии разных веществ, например, сахаров, аминокислот и нуклеозидов через мембрану под воздействием ферментов. Зависит от градиента концентрации. Является переносом по градиенту концентрации, она тоже непосредственно не зависит от энергии, обеспечиваемой АТФ. | Некоторые сахара, в частности, глюкоза | |
активный транспорт | Перенос вещества против градиента концентрации, т. е. из области низкой концентрации в область более высокой концентрации. Связан со способностью мембраны поддерживать разность электрических потенциалов (помимо поддержания разности в концентрациях веществ внутри и снаружи клетки), под которыми понимают различия между электрическими потенциалами внутри и вне клетки, а также с затратами энергии на работу в виде перемещения веществ против электрохимического градиента, т. е. «вверх» Ферменты, катализирующие эти реакции | эффективен в случае переноса ионов | |
эндоцитоз: | ответственен за поступление в клетки макромолекул (белков, по-линуклеотидов, полисахаридов) и разных твердых частиц, включая бактерии | ||
фагоцитоз | процесс активного захватывания и поглощения живых и неживых частиц одноклеточными организмами или особыми клетками (фагоцитами) многоклеточных животных организмов. Обеспечивается путем выпячиваний своей клеточной мембраны и образования пузырьков, сливающихся затем с плазматической мембраной и открывающихся внутрь клетки. Вошедшие внутрь клеток частицы поступают в лизосомы, где с помощью клеточных (лизосомных) ферментов разрушаются и усваиваются затем клетками. | У многоклеточных (млекопитающих) выполняется специализированными клетками (лейкоцитами). У простейших является формой питания. Биологическое значение фагоцитоза у млекопитающих заключается в том, что он обеспечивает иммунную (фагоцитарную) защиту организма | |
пиноцитоз | захват клеточной поверхностью жидкости с содержащимися в ней веществами путем впячиваний плазматической мембраны и образования пузырьков (канальцев), куда поступает жидкость. Канальцы после заполнения жидкостью отшнуровываются, поступают в цитоплазму и доходят до лизосом, где их стенки перевариваются, в результате чего содержимое (жидкость) канальцев освобождается и подвергается дальнейшей обработке лизосомными ферментами. | Один из основных механизмов проникновения в клетку высокомолекулярных соединений, в частности белков и углеводно-белковых комплексов. Наиболее активный П. наблюдается у амёб, в эпителиальных клетках кишечника и почечных канальцев, в эндотелии сосудов и растущих ооцитах, в клетках кровеносной и лимфатической систем, в клетках злокачественных опухолей, а также в клетках тканей, для которых характерен повышенный уровень обмена веществ. |
Экзоцитоз —процесс секретирования клетками различных веществ
Различают:
Клетки способны секретировать различные вещества в окружающую их среду. Этот процесс называют экзоцитозом.
Пассивный транспорт. Когда транспортируются незаряженные молекулы, то пассивный транспорт определяется только градиентом концентрации, т. е. разностью концентрации вещества на разных сторонах мембраны. Если же молекулы транспортируемого вещества заряжены, то к влиянию градиента концентрации добавляется влияние электических потенциалов по обе стороны мембраны. Градиент концентрации и электрический градиент в совокупности составляют электрохимический потенциал, который позволяет транспорт в клетку только положительно заряженных ионов.
Можно сказать, что пассивный транспорт веществ в клетки осуществляется обычной диффузией через клеточную мембрану, причем скорость диффузии вещества зависит от его растворимости в мембране, коэффициента диффузии в мембране и от разности концентрации веществ в клетке и за ее пределами (в среде).
Катализируемая, или так называемая «облегченная» диффузия, при которой скорость диффузии разных веществ, например, сахаров, аминокислот и нуклеозидов через мембрану повышается с помощью ферментов. Как и обычная, «облегченная» диффузия тоже зависит от градиента концентрации, однако здесь имеются подвижные «переносчики», роль которых выполняют ферменты. Находясь в составе мембраны, ферменты действуют в качестве «переносчиков» молекул веществ, проникая (диффундируя) на противоположную сторону мембраны, где они освобождаются от переносимых веществ. Поскольку «облегченная» диффузия веществ является переносом по градиенту концентрации, она тоже непосредственно не зависит от энергии, обеспечиваемой АТФ.
Примеры
Такие вещества, как глюкоза, почти нерастворимы в липидах, и размеры их частиц больше 0,8 нм; тем не менее они довольно быстро проходят через плазматическую мембрану, например через оболочку эритроцита. По-видимому, это обусловлено «облегченной диффузией» — присоединением к специфической молекуле-переносчику, представляющей собой пептид или белок. Молекула глюкозы G соединяется с молекулой-переносчиком X у наружной поверхности мембраны, и образовавшийся комплекс GX, растворимый в липидах, может диффундировать через мембрану к ее внутренней стороне, где он диссоциирует, и освобожденная глюкоза оказывается внутри клетки. Затем переносчик диффундирует обратно к наружной поверхности и сразу же может присоединить к себе другую молекулу глюкозы. При такой системе максимальная скорость переноса глюкозы определяется общим числом молекул-переносчиков, имеющихся в мембране, и возможными скоростями образования и расщепления комплекса GX. Для подобного процесса характерна «кинетика насыщения»: при малой концентрации глюкозы в наружном растворе скорость ее проникновения в клетку пропорциональна этой концентрации; однако при более высоких концентрациях пропорциональность исчезает, так как все молекулы переносчика уже «насыщены» глюкозой. Переносчики специфичны — они могут присоединять только глюкозу и некоторые очень близкие к ней по структуре сахара. Молекулы сахаров, сходные по своему химическому строению, будут конкурировать между собой за связывающие участки в молекулах-переносчиках. Облегченная диффузия не требует расхода энергии, если наружная концентрация глюкозы выше внутренней и глюкоза, таким образом, перемещается «вниз» по химическому градиенту. Однако некоторые клетки, например клетки кишечного эпителия и внутренней выстилки почечных канальцев, способны концентрировать глюкозу, заставляя ее перемещаться «вверх» по химическому градиенту (или, как говорят, против градиента концентрации), и для этого уже требуется затрата энергии. Гормон инсулин резко усиливает поглощение глюкозы скелетными мышцами и некоторыми клетками нашего организма. Пока еще не ясно, вызывает ли он увеличение числа эффективных молекул-переносчиков или же просто ускоряет реакции, с которыми связано образование и расщепление комплекса переносчика с глюкозой. Полагают, что не только облегченная диффузия, но и активный перенос глюкозы происходят с участием специфического переносчика, образующего с глюкозой комплекс, синтез или расщепление которого требует затраты энергии.
Из клеточных мембран кишечной палочки Escherichia coli был выделен и частично очищен липопротеид, который, по-видимому, служит переносчиком, или пермеазой, для лактозы. На одну клетку приходится около 9000 молекул пермеазы. Как полагают, этот белок присоединяет лактозу у наружной поверхности мембраны, а затем образовавшийся комплекс диффундирует к ее внутренней поверхности, где лактоза отделяется от переносчика. Этот гипотетический механизм в принципе сходен с предполагаемым механизмом натриевого насоса.
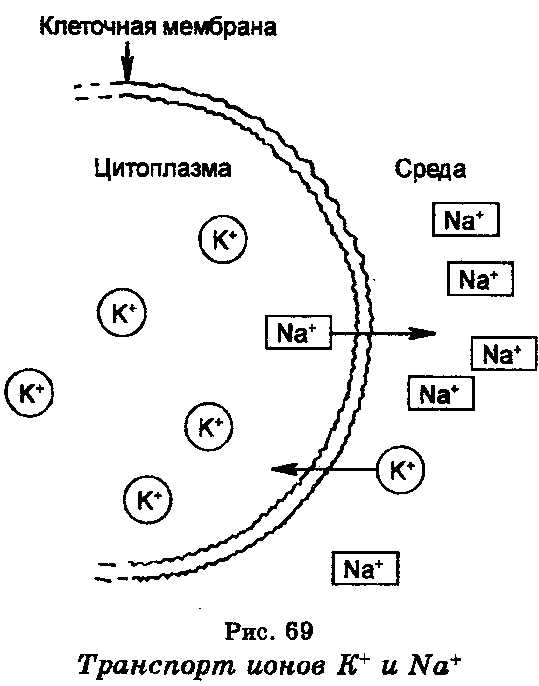
Активный транспорт.
Пример:
транспорт ионов натрия и калия, который определяет клеточный мембранный потенциал. Концентрация ионов натрия (Na+) внутри большинства клеток является меньшей, чем в среде, тогда как концентрация ионов калия (К+) внутри клеток является в 10—20 раз большей, чем в среде. В результате этого ионы Nа+ стремятся проникнуть из среды в клетку, а ионы К+, наоборот, выйти из клетки в среду. Поддержание концентрации этих ионов в клетке и в окружающей среде обеспечивается благодаря наличию в клеточной мембране системы, которая является ионным «насосом» и которая откачивает ионы Na+ из клетки в среду и накачивает ионы К+ в клетку из среды. Работа этой системы, т.е. движение ионов против электрохимического градиента, обеспечивается энергией, которая генерируется гидролизом АТФ, причем фермент АТФ-аза, катализирующий эту реакцию, содержится в самой мембране и, как считают, выполняет роль натриево-калиевого «насоса», генерирующего мембранный потенциал. Энергия, освобождаемая при гидролизе одной молекулы АТФ, обеспечивает транспорт за пределы клетки трех ионов Na+ и внутрь клетки двух ионов К+.
Система Na+ + К+ —АТФ-аза помогает поддерживать ассиметрическое распределение ионов калия при высокой концентрации последнего в клетках. Ионы калия участвуют в регуляции многих клеточных функций, включая поток солей и воды из почечных клеток, освобождение инсулина из панкреатических клеток, частоту сердцебиений.
Установлено, что энергетически выгодный транспорт ионов Na+ внутрь клеток оказывает также влияние на транспорт сахаров и аминокислот в клетки. В частности, с транспортом ионов Na+ сопряжен транспорт глюкозы. Чтобы создать градиент концентрации ионов Na+, благоприятный для транспорта ионов К+ и глюкозы внутрь клеток, ионная «насосная» система благодаря энергии активно откачивает ионы Na+ из клетки за ее пределы.
Определенная роль в транспорте веществ принадлежит белоксвязывающим системам, представляющим четвертый способ транспорта. Речь идет о белках, локализованных в периплазматическом пространстве. Эти белки специфически связывают сахара, аминокислоты и ионы, перенося их затем к специфическим молекулам-носителям, локализованным в клеточной мембране. Источником энергии для этих систем является АТФ.
Экзоцитоз
Конститутивный экзоцитоз осуществляется периодически, по мере накоплениях секрета, без видимого воздействия внеклеточных факторов, при колебаниях гидратированности клетки, без участия аппарата Гольджи и Са2+—независимо (Инсулин и ряд других гормонов, нейротрансмиттеров и многих ферментов.)
Нейромедиатор (Neurotransmitter) — химический посредник, освобождающийся из пресинаптического нервного окончания и передающий нервный импульс в синапсе постсинаптичсскому окончанию, мышечному волокну или железе, которые эти нервы иннервируют. Основными нейромедиаторами в периферической нервной системе являются ацетилхолин и норадреналин (сскретируются нервными окончаниями симпатической нервной системы). В центральной нервной системе наряду с ацетилхолином и норадреналином, нейромедиаторами являются дофамин, серотонин, гаммааминобутировая кислота и некоторые другие вещества.
Регулируемый экзоцитоз отличается от предыдущего необходимостью для запуска внеклеточного воздействия и четкой стадийностью процесса. Каждая из стадий регулируется дифференцированно. Специфика экзоцитоза определяется химической природой выделяеммых веществ, невезикулярной или везикулярной формами его выведения.
С помощью экзоцитоза из клетки удаляются также частицы, оказавшиеся непереваренными путем фагоцитоза. У большинства клеток циклы эндоцитоз-экзоцитоз непрерывны.
studfiles.net
1) Питание микроорганизмов. Способы поступления питательных веществ в бактериальную клетку. Классификация по типам питания. Зависимость от источника углерода и энергии. Факторы роста.
По типу питания микроорганизмы делятся на:
Различают голозойный (поглощение питательных веществ в виде твердых частиц, переваривание в специальных пищеварительных органах, у животных) и голофитный (всасывание через поверхность в виде растворов, у растений, грибов и бактерий) способы питания. Затем питательные вещества должны поступить в клетки.
Механизмы поступления питательных веществ в клетку могут быть следующими.
1. Пассивная (простая) диффузия – за счет разности концентраций веществ в клетке и в среде (очень редко, только вода, кислород и некоторые ионы, чаще другой механизм – облегченная диффузия).
2.Облегченная диффузия – тоже за счет разности концентраций, но с использованием специальных ферментов-переносчиков в мембране (транслоказ), выход продуктов метаболизма осуществляется так же.
3. Активный транспорт (АТ) — против градиента концентраций, с использованием ферментов-переносчиков и затратой энергии,. Различают первичный АТ с использованием химической энергии и вторичный АТ с использованием энергии протонного потенциала (симпорт, антипорт)
В зависимости от источников энергии природы доноров электронов:
фототрофы (фотосинтезирующие), способные использовать солнечную энергию – сапрофиты; хемотрофы (хемосинтезирующие), способные использовать энергию за счет окислительно-восстановительных реакций – патогенные микробы.
В зависимости от доноров электронов хемотрофы подразделяют: хемолитотрофы (хемоавтотрофы) хемоорганотрофы (хемогетеротрофы)
Факторы роста:
Аминокислоты. Многие микроорганизмы нуждаются в тех или иных аминокислотах (одной или нескольких), поскольку они не могут их самостоятельно синтезировать, например, клостридии – в лейцине, тирозине; стрептококки – в лейцине, аргинине и др.
Пуриновые и пиримидиновые основания и их производные. (аденин, гуанин, цитозин, урацил, тимин и др.) являются факторами роста для разных видов стрептококков; некоторые азотистые основания нужны для роста стафилококков и других бактерий.
Липиды (жирные кислоты – компоненты фосфолипидов) нужны для роста некоторых стрептококков и микоплазм.
Витамины, главным образом группы В, входят в состав коферментов. Например: Коринебактерии дифтерии, шигеллы нуждаются в никотиновой кислоте или ее амиде, который входит в состав НАД и НАДФ;
Золотистый стафилококк, пневмококк, бруцеллы –в тиамине (витамин В1), входящего в состав пирофосфата;
Стрептококки некоторые виды, бациллы столбняка –пантотеновой кислоте, являющейся составной частью кофермента КоА и т.д.;
Для многих бактерий факторами роста является фолиевая кислота, биотин, гемы –компоненты цитохромов (гемы необходимы гемофильным бактериям, МКБtbc
2) Особенности метаболизма бактерий: интенсивность обмена веществ, разнообразие типов метаболизма, метаболическая пластичность. Питание бактерий. Классификация бактерий по типам питания. Понятие об аутотрофах, гетеротрофах, сапрофитах, абсолютных и факультативных паразитах, прототрофах, ауксотрофах.
Метаболизм (обмен веществ) — это совокупность биохимических
реакций, протекающих в микробной клетке и направленных на построение ее компонентов и обеспечение ее энергией. Клеточный метаболизм состоит их двух потоков реакций, тесно связанных между собой и лротекающих одновременно: энергетического и конструктивного.
Конструктивный метаболизмом (анаболизмом) — называют поток реакций, направленный на построение вещества клетки (биосинтез) из мелких и простых молекул, в том числе и из поступивших извне с использованием запасенной энергии.
Энергетический метаболизм (катаболизм) — это поток реакций, сопровождаюициися распадом крупных молекул на более мелкие с выделением энергии и запасанием получаемой энергии в форме макроэргических молекул — АТФ, АДФ. ЦТФ (цитидинтрифосфат). УТФ
(уридинтрифосфат) и других, главными из них являются АТФ, которые в дальнейшем расходуются для биосинтеза, для активного движения. осморегуляции и других энергозатратных процессов.
Основными способами получения энергии для большинства микробных клеток являются дыхание (окислительное фосфорилирование) и брожение (субстратное фосфорилирование). При этом в обоих случаях происходит перенос фосфорильной группы от фосфорной кислоты на АДФ с образованием АТФ.
Дыхание по энергетической эффективности во много раз превосходит брожение. В процессе дыхания при полном окислении одной молекулы глюкозы образуется 38 молекул АТФ, А, например, при молочнокислом брожении одна молекула глюкозы даст только 2 молекулы АТФ.
Типы питания. Микроорганизмы нуждаются в углеводе, азоте, сере, фосфоре, калии и других элементах. В зависимости от источников углерода для питания бактерии делятся на аутотрофы, использующие для построения своих клеток диоксид углерода С02 и другие неорганические соединения, и гетеротрофы, питающиеся за счет готовых органических соединений. Аутотрофными бактериями являются нитрифицирующие бактерии, находящиеся в почве; серобактерии, обитающие в воде с сероводородом; железобактерии, живущие в воде с закисным железом, и др.
Гетеротрофы, утилизирующие органические остатки отмерших организмов в окружающей среде, называются сапрофитами. Гетеротрофы, вызывающие заболевания у человека или животных, относят к патогенным и условно-патогенным. Среди патогенных микроорганизмов встречаются облигатные и факультативные паразиты (от греч. parasitos — нахлебник). Облигатные паразиты способны существовать только внутри клетки, например риккетсии, вирусы и некоторые простейшие.
В зависимости от окисляемого субстрата, называемого донором электронов или водорода, микроорганизмы делят на две группы. Микроорганизмы, использующие в качестве доноров водорода неорганические соединения, называют литотрофны-ми (от греч. lithos — камень), а микроорганизмы, использующие в качестве доноров водорода органические соединения, — органотрофами.
Учитывая источник энергии, среди бактерий различают фототрофы, т.е. фотосинтезирующие (например, сине-зеленые водоросли, использующие энергию света), и хемотрофы, нуждающиеся в химических источниках энергии.
3) Физиология бактерий: рост и размножение. Пути реализации генетической информации у бактерий (особенности процессов репликации; биосинтез белка, как реализация первичной генетической информации).
Жизнедеятельность бактерий характеризуется ростом — формированием структурно-функциональных компонентов клетки и увеличением самой бактериальной клетки, а также размножением — самовоспроизведением, приводящим к увеличению количества бактериальных клеток в популяции.
Бактерии размножаются путем бинарного деления пополам, реже путем почкования. Актиномицеты, как и грибы, могут размножаться спорами. Актиномицеты, являясь ветвящимися бактериями, размножаются путем фрагментации нитевидных клеток.
Рост – это согласованное увеличение всех компонентов клетки. Результатом роста является размножение бактерий.
Размножение бактерий – увеличение числа бактерий, популяции. В процессе роста бактериальная клетка увеличивается в 2-3 раза, она интенсивно окрашивается и накапливается РНК. В благоприятных условиях рост заканчивается размножением. У бактерий размножение происходит делением пополам – бинарное деление. Это основной способ размножения у бактерий.
Размножение бактерий. Начинается с репликации — удвоения генома, а затем происходит деление. У бактерий вегетативная репликация т.е. информация передается от родительской клетки к дочерней.
У бактерий репликация саморегулируемая, так в геноме имеются гены, ответственные за репликацию, т.е. саморегулиремый процесс.
Репликация носит полуконсервативный характер. То есть дочерние клетки получают равномерно распределяющийся генетический материал. Репликация начинается с определенной точки – локус ДНК. От этой точки происходит раскручивание нитей ДНК, образуется репликационная вилка, синтезируется SSB белок, который препятствует повторному скручиванию нитей. Процесс осуществляется ДНК полимеразой, которая способна присоединять комплиментарные нуклеотиды к свободному 3”концу. Синтез комплиментарных участков запускается заправкой праймером. Это участок РНК, комплиментарный матричной ДНК и у праймера имеется свободный 3’ конец. Заправка праймером запускает синтез ДНК, на матрице строятся фрагменты ДНК(оказаки), которые сшиваются в единую нить ДНКлигазами. В бактериальной клетке образуются 2 идентичные нити ДНК, которые растаскиваются по полюсам клетки и после репликации запускается деление бактерий. Оно начинается с удлинения цитоплазматической мембраны, она удлиняется, формируется межклеточная перегородка по экватору, по которой бактерия бинарно делится и образуются 2 идентичные дочерние клетки
studfiles.net
Способы питания и поступления в клетку различных веществ — МегаЛекции
Питание микроорганизмов
Микроорганизмы, как и все другие живые существа, нуждаются в пище, которая поступает в их клетки из окружающей среды. Пищей обычно называют вещества, которые, попав в живой организм, служат либо источником энергии для процессов жизнедеятельности, либо материалом для построения составных частей клетки.
Способы питания и поступления в клетку различных веществ
Потребность в питательных веществах микроорганизмы могут удовлетворять непосредственно усваивая их или предварительно преобразуя в доступную форму. Известны два способа питания живых существ — голозойный и голофитный.
При голозойном способе питания живой организм захватывает или заглатывает плотные частицы пищи, которая затем переваривается в пищеварительном тракте. Указанный способ питания характерен для животных (от простейших до высших). При голофитном способе питания живые существа, не имеющие специальных органов для заглатывания и пищеварения, используют питательные вещества, всасывая их в виде относительно небольших молекул из водного раствора. Данный способ питания свойствен растениям и микроорганизмам.
Полимерные органические соединения (полисахариды, белки и др.) микроорганизмы не могут поглощать и использовать непосредственно в обмене веществ клетки. Такие вещества должны быть вначале расщеплены на простые соединения, для которых клеточная мембрана проницаема. Крупные молекулы расщепляются экзо-ферментами, экскретируемыми клетками микроорганизмов в среду. Такое внешнее, или внеклеточное, переваривание свойственно только микроорганизмам.
Поступление воды и растворенных в ней питательных веществ из окружающей среды внутрь микробной клетки, а также выход продуктов обмена происходят через клеточную стенку, капсулу и слизистые слои. Капсула и слизистые слои представляют собой достаточно рыхлые образования и, возможно, не оказывают значительного влияния на транспорт веществ, тогда как клеточная стенка служит существенным барьером для поступления питательных соединений в клетку.
Активную роль в поступлении в клетку питательных веществ играет цитоплазматическая мембрана. Чтобы обеспечить нормальную жизнедеятельность микроорганизма, последняя должна быть проницаемой для питательных веществ и кислорода, поступающих в клетку, а также для продуктов обмена, выходящих наружу. Поступление воды и растворенных в ней веществ через цитоплазматическую мембрану — активный процесс: живая микробная клетка никогда не находится в равновесии с веществами окружающей среды, проходящими через ее мембрану.
Транспорт питательных веществ. Выделяют несколько типов транспортных систем, при помощи которых вещества из окружающей среды проходят через цитоплазматическую мембрану: пассивную диффузию, облегченную диффузию, активный транспорт и перенос групп (радикалов). Причем два из них (пассивная и облегченная диффузия) обеспечивают только транспорт, но не накопление веществ в микробной клетке, в то время как активный транспорт способствует аккумуляции веществ внутри клетки.
При пассивной диффузии транспорт вещества происходит через цитоплазматическую мембрану под действием разности концентраций (для неэлектролитов) или разности электрических потенциалов (для ионов) по обе стороны мембраны. Экспериментами показано, что, за исключением воды, только кислород и некоторые ионы проходят через цитоплазматическую мембрану в результате пассивной диффузии. Скорость такого переноса веществ весьма незначительна. Определенное значение пассивная диффузия приобретает при нарушениях жизнедеятельности бактериальной клетки.
Транспорт большинства растворенных веществ осуществляется через мембрану при действии специальных механизмов переноса, того служат молекулы-переносчики, циркулирующие между внешним и внутренним пограничными слоями цитоплазматической мембраны. Считают, что данные переносчики связывают молекулы растворенных веществ на ее внешней стороне, транспортируют их к внутренней, где освобождают, и молекулы питательного вещества поступают в цитоплазму без изменения. Такие связанные с цитоплазматической мембраной переносчики, представляющие собой субстратспецифичные связывающие белки, называют пермеазами, или транслоказами.
Транспорт растворенных веществ, осуществляемый переносчиками, может быть в виде облегченной диффузии и активного транспорта. Движущей силой облегченной диффузии служит разница в концентрации какого-либо вещества по обе стороны мембраны. Молекула вещества соединяется с молекулой-переносчиком у наружной поверхности мембраны, и образовавшийся комплекс диффундирует через мембрану к ее внутренней стороне. Там он диссоциирует, и освобожденное вещество оказывается внутри клетки. Затем переносчик диффундирует к наружной поверхности и может присоединить новую молекулу вещества.
Облегченная диффузия не требует расхода энергии, если наружная концентрация вещества выше внутренней, так как оно перемещается «вниз» по химическому градиенту. Скорость процесса зависит от концентрации вещества в наружном растворе. Предполагают, что выход продуктов обмена веществ из микробной клетки также происходит по типу облегченной диффузии при участии переносчиков.
Активный транспорт связан с работой специфических транспортных белков (пермеаз, транслоказ и др.), которые также находятся в цитоплазматической мембране. В этом случае растворенные вещества переносятся в клетки микроорганизмов «вверх» по химическому градиенту (или против градиента концентрации). Считают, что большинство веществ проникает в клетку микроорганизма в результате активного транспорта. Источниками энергии для транспортных процессов служат АТФ, протонный потенциал и фосфоенолпируват. Существуют системы «первичного» и «вторичного» активного транспорта.
В системах «первичного» активного транспорта используется химическая энергия. Необходимость использования энергии для поддержания активного транспорта объясняется теми изменениями, которые претерпевает переносчик: когда он обращен к внешней поверхности мембраны, то обладает высоким сродством к субстрату, а когда к внутренней — низким сродством к субстрату. Возможность транспортировать вещества против градиентов концентраций часто используется клетками бактерий для получения этих веществ из окружающей среды, где их концентрация мала, что обычно для природных условий. При отсутствии источников энергии накопления веществ внутри не происходит.
Подсчитано, что перенос молекулы тиогалактозида через цитоплазматическую мембрану кишечной палочки требует затраты одной молекулы АТФ. Предполагая, что активный перенос других соединений связан с подобным же расходом АТФ, можно понять, какое значительное количество энергии на транспорт веществ в клетку потребляет растущий и размножающийся микроорганизм. В отдельных случаях на активный транспорт может затрачиваться почти вся энергия, вырабатываемая в микробной клетке.
В системах «вторичного» активного транспорта для переноса против градиента концентрации через мембрану многих веществ, в том числе неорганических и органических ионов, Сахаров, используется энергия протонного потенциала. В процессе дыхания в локализованной в мембране дыхательной цепи осуществляется вывод протонов. В результате перемещения протонов через мембрану за счет энергии дыхания создается градиент электрохимического потенциала (называемый также протонным потенциалом, или протондвижущей силой) между наружной и внутренней сторонами мембраны. Протонный потенциал обусловливает фосфорилирование, т. е. синтез АТФ, или используется непосредственно транспортными системами. Протонный потенциал (Δр) определяется мембранным потенциалом (Δψ), имеющим отрицательное значение внутри клетки, и градиентом протонов (ΔрН), имеющим внутри клетки щелочной показатель в соответствии с уравнением
Δр = Δψ — z • ΔрН (мВ),
где z = 59 мВ при 25 °С.
Для поддержания протонного потенциала микроорганизм непрерывно выкачивает за пределы своей клетки протоны и другие ионы (Na+). В этих целях используются специфичные транспортные белки, имеющиеся в мембране. Каждому транспортному белку присуща определенная функция. Способность белка катализировать одновременный и однонаправленный транспорт одного протона и одной молекулы субстрата, например сахара (лактозы, глюкозы и др.), называют симпортом. Унипорт наблюдается, когда белок осуществляет перенос только одного субстрата (без протона), антипорт — когда он осуществляет перенос двух разных субстратов, обычно ионов, в противоположных направлениях.
У многих микроорганизмов сахара (фруктоза, глюкоза и другие родственные вещества) транспортируются в клетку при помощи фосфотрансферазной системы переноса групп (радикалов). Данный процесс отличается от активного транспорта тем, что субстрат попадает внутрь бактериальной клетки в химически модифицированной форме — чаще всего в виде фосфатного эфира. Движущая сила рассматриваемого процесса состоит в том, что внутри цитоплазматической мембраны сахар связывается в результате реакции с фосфорилированным ферментом (фосфотрансферазная система, источником энергии в этой реакции служит фосфоенолпируват), образующийся в итоге фосфорный эфир освобождается и поступает в цитоплазму. Химическая природа транспортируемого вещества при переносе претерпевает определенные изменения.
Таким образом, удовлетворение пищевых потребностей микроорганизмов зависит не только от внутреннего комплекса ферментов, необходимого для утилизации определенных соединений, но и от действия специфических транспортных механизмов.
Рекомендуемые страницы:
Воспользуйтесь поиском по сайту:
megalektsii.ru
Поступление вещества в клетки
Благодаря содержанию растворов солей, сахаров и других осмотически активных веществ, клетки характеризуются наличием в них определенного осмотического давления. Например, давление в клетках животных (морских и океанических форм) достигает 30 атм и более. В клетках растений осмотическое давление является еще большим. Разность концентрации веществ внутри и снаружи клетки называют градиентом концентрации.
Поступление веществ в клетки животных, равно как и удаление их из клеток, связано с проницаемостью клеточной мембраны для молекул или ионов, а также со свойствами веществ. Клеточная мембрана регулирует обмен различными веществами между клеткой и средой. Поддержание мембраны и ее проницаемость обеспечиваются клеточной энергией.
Известно несколько путей поступления веществ в клетки. В частности, различают пассивный, катализируемый и активный транспорт веществ в клетки, а также проникновение веществ в клетки путем эндоцитоза в виде фагоцитоза и пиноцитоза. Пассивный, катализируемый и активный транспорт обеспечивают проникновение в клетки лишь малых молекул, тогда как эндоцитоз ответственен за поступление в клетки макромолекул (белков, полинуклеотидов, полисахаридов) и разных твердых частиц, включая бактерии. В то же время клетки способны секретировать различные вещества в окружающую их среду. Этот процесс называют экзоцитозом.
Пассивный транспорт веществ в клетки обеспечивается диффузией через мембрану по градиенту концентрации. Молекулы обычно переходят из области высокой концентрации в область более низкой концентрации.
Когда транспортируются незаряженные молекулы, то пассивный транспорт определяется только градиентом концентрации, т. е. разностью концентрации вещества на разных сторонах мембраны. Если же молекулы транспортируемого вещества заряжены, то к влиянию градиента концентрации добавляется влияние электрических потенциалов по обе стороны мембраны. Градиент концентрации и электрический градиент в совокупности составляют электрохимический потенциал, который позволяет транспорт в клетку только положительно заряженных ионов.
Можно сказать, что пассивный транспорт веществ в клетки осуществляется обычной диффузией через клеточную мембрану, причем скорость диффузии вещества зависит от его растворимости в мембране, коэффициента диффузии в мембране и от разности концентрации веществ в клетке и за ее пределами (в среде). Этим путем в клетку проникают вода, двуокись углерода и молекулы органических веществ, способные хорошо растворяться в жирах. Вещества в клетку проникают через поры, имеющиеся в клеточной мембране. Пассивный транспорт не зависит от энергии, обеспечиваемой АТФ.
Известна катализируемая, или так называемая «облегченная» диффузия, при которой скорость диффузии разных веществ, например, Сахаров, аминокислот и нуклеозидов через мембрану повышается с помощью белков (ферментов). Как и обычная диффузия, «облегченная» диффузия тоже зависит от градиента концентрации, однако здесь имеются подвижные «переносчики», роль которых выполняют ферменты. Находясь в составе мембраны, ферменты действуют в качестве «переносчиков» молекул веществ, проникая (диффундируя) на противоположную сторону мембраны, где они освобождаются от переносимых веществ. Поскольку «облегченная» диффузия веществ является переносом по градиенту концентрации, она тоже непосредственно не зависит от энергии, обеспечиваемой АТФ.
Активный транспорт веществ в клетку отличается от пассивного (диффузии) тем, что вещество переносится против градиента концентрации, т. е. из области низкой концентрации в область более высокой концентрации. Активный транспорт связан со способностью мембраны поддерживать разность электрических потенциалов (помимо поддержания разности в концентрациях веществ внутри и снаружи клетки), под которыми понимают различия между электрическими потенциалами внутри и вне клетки, а также с затратами энергии на работу в виде перемещения веществ против электрохимического градиента, т. е. «вверх».
Энергия для транспорта обеспечивается фосфоэнолпируватом, фосфатная группа которого и часть химической энергии которого передаются белкам, часть которых используется всеми сахарами, транспортируемыми фосфотрансферазной системой, а часть специфична для отдельных Сахаров. Конечный белок содержится в мембране и ответственен за транспорт и фосфорилирование Сахаров.
Активный транспорт особенно эффективен в случае переноса ионов. Реакции, обеспечивающие активный транспорт, происходят в мембране и сопряжены с реакциями, дающими свободную энергию. Ферменты, катализирующие эти реакции, также локализованы в мембране. Примером активного транспорта веществ является транспорт ионов натрия и калия, который определяет клеточный мембранный потенциал. Концентрация ионов натрия (Na+) внутри большинства клеток является меньшей, чем в среде, тогда как концентрация ионов калия (К+) внутри клеток является в 10—20 раз большей, чем в среде. В результате этого ионы Nа+ стремятся проникнуть из среды в клетку, а ионы К+, наоборот, выйти из клетки в среду.
Поддержание концентрации этих ионов в клетке и в окружающей среде обеспечивается благодаря наличию в клеточной мембране системы, которая является ионным «насосом» и которая откачивает ионы Na+ из клетки в среду и накачивает ионы К+ в клетку из среды. Работа этой системы, т.е. движение ионов против электрохимического градиента, обеспечивается энергией, которая генерируется гидролизом АТФ, причем фермент АТФ-аза, катализирующий эту реакцию, содержится в самой мембране и, как считают, выполняет роль натриево-калиевого «насоса», генерирующего мембранный потенциал. Энергия, освобождаемая при гидролизе одной молекулы АТФ, обеспечивает транспорт за пределы клетки трех ионов Na+ и внутрь клетки двух ионов К+.
Система Na+ + К+ —АТФ-аза помогает поддерживать ассиметрическое распределение ионов калия при высокой концентрации последнего в клетках. Ионы калия участвуют в регуляции многих клеточных функций, включая поток солей и воды из почечных клеток, освобождение инсулина из панкреатических клеток, частоту сердцебиений.
Установлено, что энергетически выгодный транспорт ионов Na+ внутрь клеток оказывает также влияние на транспорт сахаров и аминокислот в клетки. В частности, с транспортом ионов Na+ сопряжен транспорт глюкозы. Чтобы создать градиент концентрации ионов Na+, благоприятный для транспорта ионов К+ и глюкозы внутрь клеток, ионная «насосная» система благодаря энергии активно откачивает ионы Na+ из клетки за ее пределы.
Определенная роль в транспорте веществ принадлежит белоксвязывающим системам, представляющим четвертый способ транспорта. Речь идет о белках, локализованных в периплазматическом пространстве. Эти белки специфически связывают сахара, аминокислоты и ионы, перенося их затем к специфическим молекулам-носителям, локализованным в клеточной мембране. Источником энергии для этих систем является АТФ.
Эндоцитоз, как отмечено выше, обеспечивает перенос в клетки крупных частиц и молекул. В рамках эндоцитоза различают фагоцитоз и пиноцитоз.
Фагоцитоз (от греч. phagos — пожирающий и cytos — клетка) представляет собой процесс, заключающийся в том, что клетки-лейкоциты (макрофаги и нейтрофилы) захватывают (обволакивают) твердые частицы (фрагменты клеток, бактерии) путем выпячиваний своей клеточной мембраны и образования пузырьков, сливающихся затем с плазматической мембраной и открывающихся внутрь клетки. Вошедшие внутрь клеток частицы поступают в лизосомы, где с помощью клеточных (лизосомных) ферментов разрушаются и усваиваются затем клетками. Фагоцитоз широко распространен среди одноклеточных организмов. У многоклеточных (млекопитающих) он выполняется специализированными клетками (лейкоцитами).
У простейших фагоцитоз является формой питания, в результате которого твердые частицы проникают в лизосомы, где и перевариваются, образуя продукты, служащие пищей. Биологическое значение фагоцитоза у млекопитающих заключается в том, что он обеспечивает иммунную (фагоцитарную) защиту организма
Пиноцитоз (от греч. pino — пить и cytos — клетка) представляет собой процесс, при котором клетки поглощают жидкости и находящиеся в них высокомолекулярные вещества путем впячиваний плазматической мембраны и образования пузырьков (канальцев), куда поступает жидкость. Канальцы после заполнения жидкостью отшнуровываются, поступают в цитоплазму и доходят до лизосом, где их стенки перевариваются, в результате чего содержимое (жидкость) канальцев освобождается и подвергается дальнейшей обработке лизосомными ферментами.
Пиноцитоз часто встречается у одноклеточных животных, у многоклеточных он наблюдается в клетках кровеносной и лимфатической систем, в клетках злокачественных опухолей, а также в клетках тканей, для которых характерен повышенный уровень обмена веществ.
Экзоцитоз — это процесс секретирования клетками различных веществ, причем известны регулируемый и конститутивный пути экзоцитоза. Примером регулируемого экзоцитоза является экзоцитоз инсулина. Клетки поджелудочной железы, продуцирующие инсулин, упаковывают его вначале в так называемые секреторные пузырьки, которые после внеклеточного сигнала сливаются с плазматической мембраной, а затем открываются в межклеточное пространство, освобождая гормон. Подобным образом происходит экзоцитоз других гормонов, нейротрансмиттеров и многих ферментов. Напротив, конститутивный путь экзоцитоза присущ многим белкам, непрерывно синтезируемым клетками и упаковываемым в эк-зоцитозные пузырьки в комплексе Гольджи, после чего эти пузырьки перемещаются к плазматической мембране, где и открываются в межклеточное пространство, освобождаясь от белкового содержимого. С помощью экзоцитоза из клетки удаляются также частицы, оказавшиеся непереваренными путем фагоцитоза. У большинства клеток циклы эндоцитоз-экзоцитоз непрерывны.
biofile.ru
1. Особенности анаболизма у разных групп микробов. Способы поступления питательных веществ в клетку, классификация микробов по этому принципу.
Поступление питательных веществ в микробную клетку может происходить за счёт:
облегченная диффузия – не требует метаболической энергии. Так поступает вода, электролиты (все классы микроорганизмов)
активный транспорт питательных веществ – явл. энергозависимым процессом. В этом процессе должны учавствовать экзоферменты бактерий. Это голофитный тип питания (грибы, бактерии).
Эндоцитоз состоит из фагоцитоза и пиноцитоза. Этот тип характерен только для эукариот. Они имеют мягкую оболочку и процессы пищеварения протекают с участием лизосом внутри клетки. Это голозойный тип питания (простейшие).
Анаболизм – конструктивный метаболизм, ассимиляция, способ питания. В ферментативных реакциях происходит синтез молекулярных компонентов, из которых состоит тело клетки. эти процессы связаны с потреблением энергии. В зависимости от использования углерода и азота для митания микробы делят на АУТОТРОФЫ( сами синтезируют компоненты клетки из простых соединений и со2 как источника углерода), ГЕТЕРОТРОФЫ(получают углерод из различных хим-х соед), ГИПОТРОФЫ(за счет клетки-хозяина).
2. Внутрикожные аллергические пробы в диагностике инфекционных болезней. Проба Манту.
Туберкулиновая проба (реакция Манту, проба Пирке) представляет собой кожную пробу, направленную на выявление наличия специфического иммунного ответана введение туберкулина. Наличие выраженной кожной реакции свидетельствует о наличии напряжённогоиммунитета, то есть, что организм активно взаимодействует с возбудителем. Реакция Манту — это своего рода иммунологический тест, который показывает, есть ли в организме туберкулёзная инфекция.
Туберкулин — общее название экстрактов микобактерийM. tuberculosis, M. bovis или M. avium, используемых для проведения внутрикожных диагностических проб на туберкулёз у человека и животных.
Внутрикожная туберкулиновая проба применяется для диагностики туберкулёзной инфекциина основании появляющейся местной индурации (уплотнения кожи), а также для определения местоположения аллергии (перед прививкойБЦЖ) и для контроля конверсии (Инверсии) после прививки.
Проверка результата пробы производится не ранее чем через 48 часов, лучше всего на третий день, самое позднее — через одну неделю после аппликации. Индурация отмечается, измеряется, документируется и оценивается.
Индурация< 5 мм в основном не имеет значения;
10 мм указывает на возможное заражение туберкулезом в группах риска и при контакте с пациентами с открытыми формами туберкулеза
при индурации15 мм или язвенной реакции кожи (образование гнойников) очень вероятно заражение туберкулезом.
Туберкулиновая проба Манту показывает реакцию организма — антиген-антителона возбудителя туберкулёза. Позитивная реакция кожи показывает, что исследуемый пациент имел контакт с возбудителями туберкулёза. Это, однако, не означает, что данный пациент болен туберкулезом.
2, 4, 6, 24,26, 28, 33,34, 45,47, 48, 49, 51, 52, 54, 61, 62, 66
studfiles.net
Leave A Comment