
Синтез липидов из углеводов
Если количество углеводов, поглощенных с пищей за один прием, больше того, чем может быть запасено в виде гликогена, то избыток углеводов превращается в жиры. Начальная последовательность реакций совпадает при этом с обычным окислительным путем, т.е. сначала из глюкозы образуется ацетил-КоА, который используется в цитоплазме клетки для синтеза длинноцепочечных жирных кислот. Затем жирные кислоты запасаются в виде нейтральных жиров (триглицеридов), которые откладываются в разных частях тела. Когда требуется энергия, нейтральные жиры подвергаются гидролизу и жирные кислоты поступают в кровь, адсорбируются молекулами плазменных белков (альбуминов и глобулинов) и затем поглощаются различными клетками.
Циклы Кори и аланина
В клетках, не содержащих митохондрий (например, в эритроцитах), или в тканях при недостаточном снабжении кислородом (например, в активно работающих мышцах) АТФ (АТР) синтезируется за счет процесса превращений глюкозы в лактат (

Лактат переносится кровью в печень, где в процессе глюконеогенеза с затратой АТФ вновь конвертируется в глюкозу (цикл Кори).
При
интенсивной работе мышц максимально
активируется гликолиз.
Продукт гликолиза,
пировиноградная
кислота
(пируват) накапливается в цитоплазме
и недостаточно быстро поступает в
митохондрии,
если они из-за недостатка кислорода
не готовы к окислению
пирувата. В анаэробных условиях пируват
в реакции,
катализируемой лактатдегидрогеназой
(заключительный этап гликолиза)
восстанавливается до лактата. Одновременно
НАДН (NADH), кофермент
лактатдегидрогеназы,
окисляется до НАД+ (NAD+),
который вновь используется на окислительном
этапе гликолитического пути. Этой
реакции
способствует относительно высокое
отношение НАДН/НАД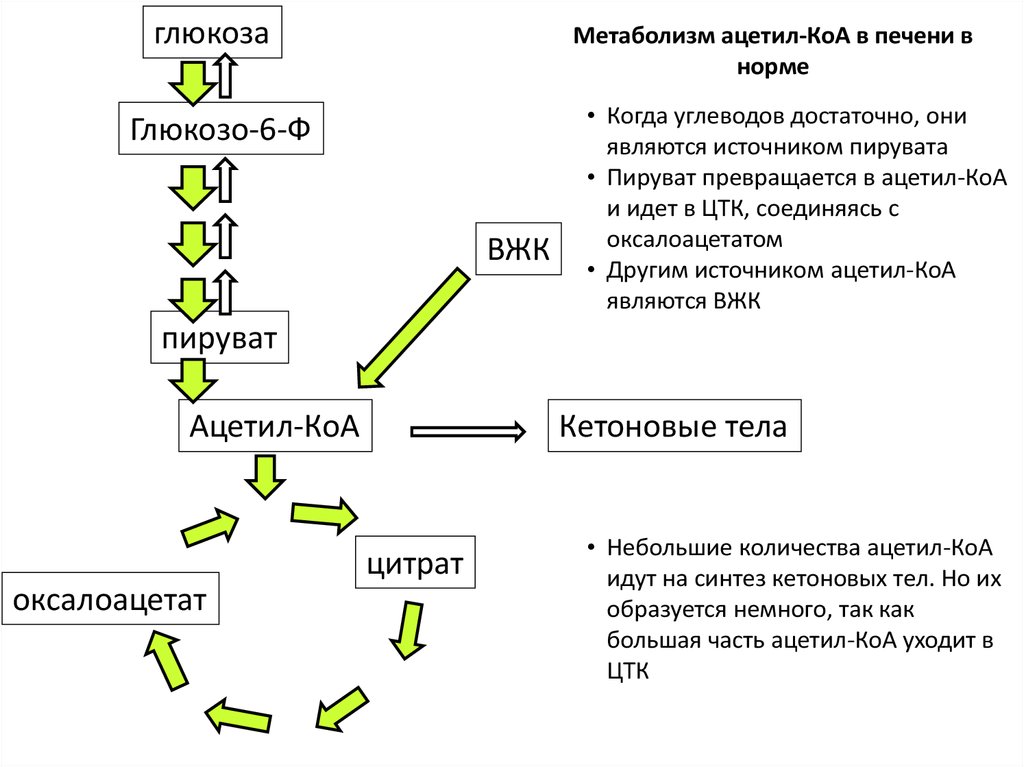
Обратная связь, подобная циклу Кори, существует в глюкозо-аланиновом цикле, в котором также участвует пируват. Глюкозо-аланиновый цикл берет начало с протеолиза белков. Образующиеся аминокислоты в результате трансаминирования в присутствии ферментов превращаются в α-кетокислоты, которые в основном включаются в цикл трикарбоновых кислот (цитратный цикл). Одновременно в реакции, катализируемой аланинтрансаминазой, аминогруппы из разных аминокислот переносятся на имеющийся субстрат, пируват. Образующийся аланин поступает в кровь и переносится в печень. Таким образом, цикл аланина служит каналом передачи азота и предшественников глюкозы в печень, которая является местом синтеза конечных продуктов азотистого обмена, например мочевины.
Роль инсулина в регуляции обмена веществ
Инсулин
участвует в регуляции всех видов обмена
веществ. Это основной гормон, обладающий
гипогликемическим действием (снижению
сахара крови также способствует
соматостатин).
Он является полипептидным гормоном, образующимся в β-клетках поджелудочной железы. Главным стимулятором синтеза и секреции инсулина является глюкоза. Образующийся инсулин поступает в сосудистое русло, где он частично остается в свободном виде, а частично образует комплексы с белками крови.
Инсулин опосредует поступление и метаболизм глюкозы в ткани, имеющие рецепторы к инсулину (инсулинзависимые ткани), рис. 7.
К инсулинзависимым тканям относятся мышечная, жировая ткань, печень и островковый аппарат поджелудочной железы. В эти ткани глюкоза поступает путем пассивного переноса или облегченной диффузии. Остальные ткани являются инсулиннезависимыми и поступление глюкозы в них происходит без участия инсулина по градиенту концентрации.
Активируя
гексокиназу, в клетках инсулинзависимых
тканей инсулин обеспечивает фосфорилирование
глюкозы, превращение ее в Гл-6-фосфат и
дальнейшее превращение. Инсулин
стимулирует гликолиз, цикл Кребса,
пентозофосфатный путь обмена глюкозы
и угнетает глюконеогенез. Инсулин
усиливает гликогенез в результате
активации гликоген-синтетазы и тормозит
гликогенолиз.
Инсулин
стимулирует гликолиз, цикл Кребса,
пентозофосфатный путь обмена глюкозы
и угнетает глюконеогенез. Инсулин
усиливает гликогенез в результате
активации гликоген-синтетазы и тормозит
гликогенолиз.
Кроме глюкозы инсулин также опосредует поступление в клетки аминокислот и электролитов. Он активирует синтез белка, жира и тормозит их распад, препятствует избыточному холестерол- и кетообразованию. Инсулин также участвует в регуляции водно-электролитного обмена (обладает водо- и солесберегающим действием), участвует в регуляции кислотно-основного состояния (препятствует развитию ацидоза).
Суточная потребность в инсулине – 40 единиц, а его содержание в поджелудочной железе здорового человека составляет 150-250 Ед. Инактивация инсулина происходит преимущественно в печени и почках под влиянием инсулиназы.
По
влиянию на уровень глюкозы другие
гормоны (глюкагон, соматотропный гормон
(СТГ), пролактин, адреналин и норадреналин,
глюкокортикоиды) относятся к группе
контринсулярных гормонов. Глюкагон
синтезируется в α-клетках поджелудочной
железы. Механизм гипергиликемического
действия глюкагона связан с усилением
гликогенолиза в печени. Аналогичным
действием обладают СТГ, пролактин,
адреналин и норадреналин. Гипергликемическое
действие глюкокортикоидов преимущественно
связано с их стимулирующим влиянием на
глюконеогенез.
Глюкагон
синтезируется в α-клетках поджелудочной
железы. Механизм гипергиликемического
действия глюкагона связан с усилением
гликогенолиза в печени. Аналогичным
действием обладают СТГ, пролактин,
адреналин и норадреналин. Гипергликемическое
действие глюкокортикоидов преимущественно
связано с их стимулирующим влиянием на
глюконеогенез.
10 класс. Биология. Строение клетки. Комплекс Гольджи. Эндоплазматическая сеть. Лизосомы. Клеточные включения — Строение клетки. Комплекс Гольджи. Эндоплазматическая сеть. Лизосомы. Клеточные включения
Комментарии преподавателяМы продолжаем изучать органоиды клетки.
Все органоиды делятся на мембранные и немембранные.
Немембранные органоиды мы рассмотрели на предыдущем занятии, напомним, что к ним относятся рибосомы, клеточный центр и органоиды движения.
Среди мембранных органоидов различают одномембранные и двумембранные.
В этой части курса мы рассмотрим одномембранные органоиды: эндоплазматическую сеть, аппарат Гольджи и лизосомы.
Кроме этого, мы рассмотрим включения – непостоянные образования клетки, которые возникают и исчезают в процессе жизнедеятельности клетки.
Эндоплазматическая сеть
Одним из самых важных открытий, сделанных с помощью электронного микроскопа, было обнаружение сложной системы мембран, пронизывающей цитоплазму всех эукариотических клеток. Эта сеть мембран в дальнейшем получила название ЭПС (эндоплазматической сети) (рис. 1) или ЭПР (эндоплазматического ретикулума). ЭПС представляет систему трубочек и полостей, пронизывающей цитоплазму клетки.
Рис. 1. Эндоплазматическая сеть
Слева – среди других органоидов клетки. Справа – отдельно выделенная
Справа – отдельно выделенная
Мембраны ЭПС (рис. 2) имеют такое же строение, как и клеточная или плазматическая мембрана (плазмалемма). ЭПС занимает до 50% объема клетки. Она нигде не обрывается и не открывается в цитоплазму.
Различают гладкую ЭПС и шероховатую, или гранулярную ЭПС (рис. 2). На внутренних мембранах шероховатой ЭПС располагаются рибосомы – здесь идет синтез белков.
Рис. 2. Виды ЭПС
Шероховатая ЭПС (слева) несет на мембранах рибосомы и отвечает за синтез белка в клетке. Гладкая ЭПС (справа) не содержит рибосом и отвечает за синтез углеводов и липидов.
На поверхности гладкой ЭПС (рис. 2) идет синтез углеводов и липидов. Вещества, синтезированные на мембранах ЭПС, переносятся в трубочки и затем транспортируются к местам назначения, где депонируются или используются в биохимических процессах.
Шероховатая ЭПС лучше развита в клетках, которые синтезируют белки для нужд организма, например, белковые гормоны эндокринной системы человека. А гладкая ЭПС – в тех клетках, которые синтезируют сахара и липиды.
В гладкой ЭПС накапливаются ионы кальция (важные для регуляции всей функций клеток и целого организма).
Структуру, известную сегодня как комплекс или аппарат Гольджи (АГ) (рис. 3), впервые обнаружил в 1898 году итальянский ученый Камилло Гольджи (Источник).
Подробно изучить строение комплекса Гольджи удалось значительно позже с помощью электронного микроскопа. Эта структура содержится практически во всех эукариотических клетках, и представляет собой стопку уплощенных мембранных мешочков, т. н. цистерн, и связанную с ними систему пузырьков, называемых пузырьками Гольджи.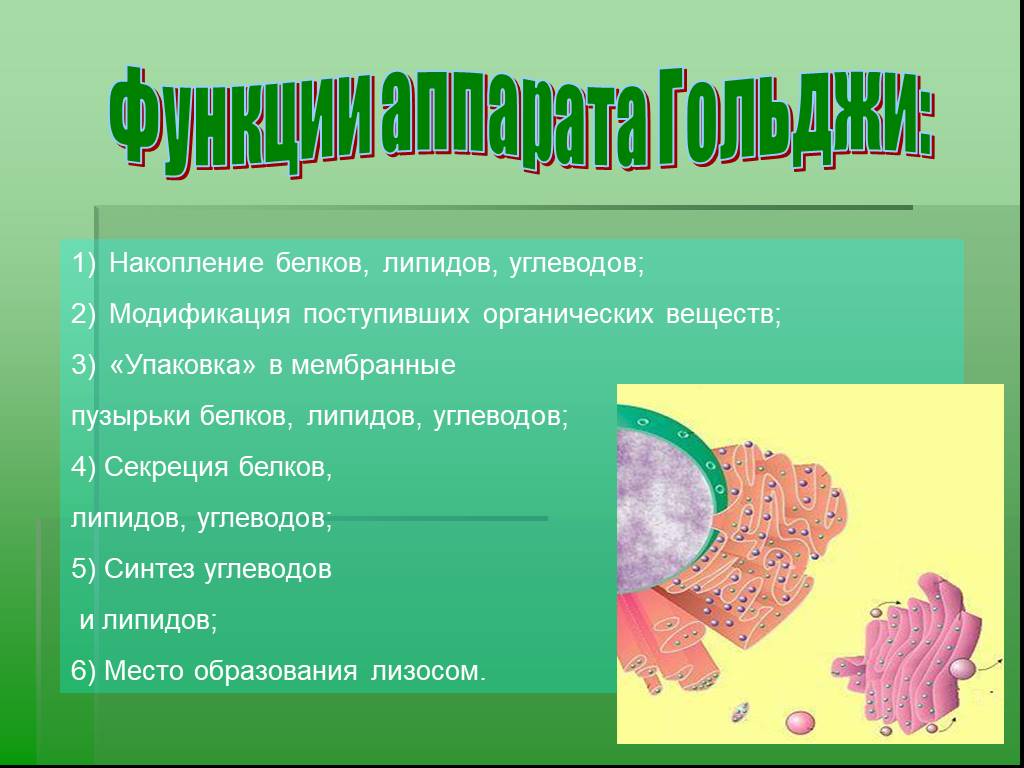
Рис. 3. Комплекс Гольджи
Слева – в клетке, среди других органоидов.
Справа – комплекс Гольджи с отделяющимися от него мембранными пузырьками
Во внутриклеточных цистернах накапливаются вещества, синтезированные клеткой, т. е. белки, углеводы, липиды.
В этих же цистернах вещества, поступившие из ЭПС, претерпевают дальнейшие биохимические превращения, упаковываются в мембранные пузырьки и доставляются к тем местам клетки, где они необходимы. Они участвуют в достройке клеточной мембраны или выделяются наружу (секретируются) из клетки.
Комплекс Гольджи построен из мембран и расположен рядом с ЭПС, но не сообщается с её каналами.
Все вещества, синтезированные на мембранах ЭПС (рис. 2), переносятся в комплекс Гольджи в мембранных пузырьках, которые отпочковываются от ЭПС и сливаются затем с комплексом Гольджи, где они претерпевают дальнейшие изменения.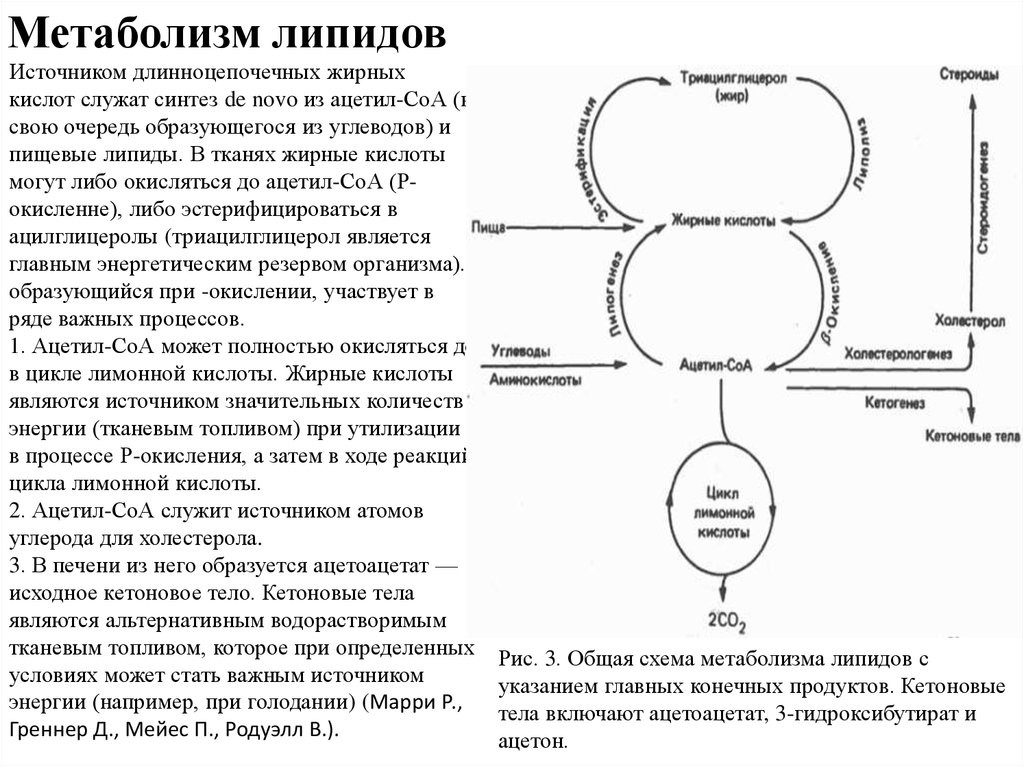
Одна из функций комплекса Гольджи – сборка мембран. Вещества, из которых состоят мембраны – белки и липиды, как вы уже знаете, – поступают в комплекс Гольджи из ЭПС.
В полостях комплекса собираются участки мембран, из которых образуются особые мембранные пузырьки (рис. 4), они передвигаются по цитоплазме в те места, где необходима достройка мембраны.
Рис. 4. Синтез мембран в клетке комплексом Гольджи (см. видео)
В комплексе Гольджи синтезируются практически все полисахариды, необходимые для построения клеточной стенки клеток растений и грибов. Здесь они упаковываются в мембранные пузырьки, доставляются к клеточной стенке и сливаются с ней.
Таким образом, основные функция комплекса (аппарата) Гольджи – химическое превращение синтезированных в ЭПС веществ, синтез полисахаридов, упаковка и транспорт органических веществ в клетке, формирование лизосомы.
Лизосомы (рис. 5) обнаружены у большинства эукариотических организмов, но особенно много их в клетках, которые способны к фагоцитозу. Они представляют собой одномембранные мешочки, наполненные гидролитическими или пищеварительными ферментами, такими как липазы, протеазы и нуклеазы, т. е. ферменты, которые расщепляют жиры, белки и нуклеиновые кислоты.
Рис. 5. Лизосома – мембранный пузырек, содержащий гидролитические ферменты
Содержимое лизосом имеет кислую реакцию – для их ферментов характерен низкий оптимум pH. Мембраны лизосомы изолируют гидролитические ферменты, не давая им разрушать другие компоненты клетки. В клетках животных лизосомы имеют округлую форму, их диаметр – от 0,2 до 0,4 микрон.
В растительных клетках функцию лизосом выполняют крупные вакуоли. В некоторых растительных клетках, особенно погибающих, можно заметить небольшие тельца, напоминающие лизосомы.
В некоторых растительных клетках, особенно погибающих, можно заметить небольшие тельца, напоминающие лизосомы.
Скопление веществ, которые клетка депонирует, использует для своих нужд, или хранит для выделения вовне, называютклеточными включениями.
Среди них зерна крахмала (запасной углевод растительного происхождения) или гликогена (запасной углевод животного происхождения), капли жира, а также гранулы белков.
Эти запасные питательные вещества располагаются в цитоплазме свободно и не отделены от неё мембраной.
Функции ЭПС
Одна из самых важных функций ЭПС – синтез липидов. Поэтому ЭПС обычно представлена в тех клетках, где интенсивно происходит этот процесс.
Как происходит синтез липидов? В клетках животных липиды синтезируются из жирных кислот и глицерина, которые поступают с пищей (в клетках растений они синтезируются из глюкозы).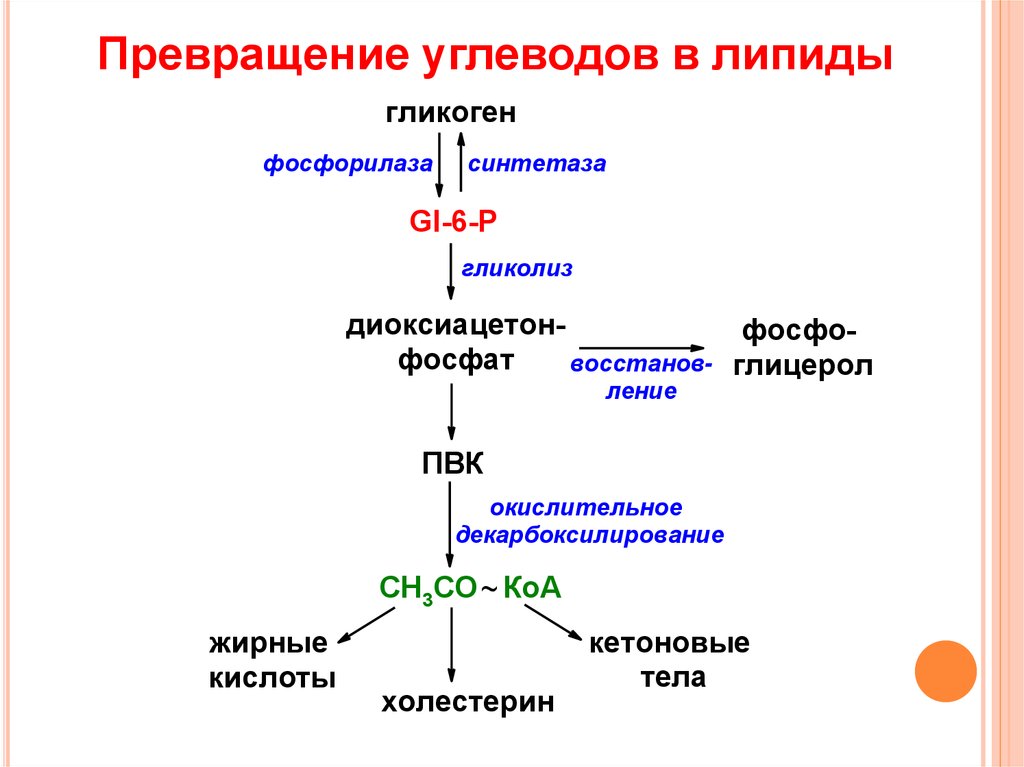 Синтезированные в ЭПС липиды передаются в комплекс Гольджи, где «дозревают».
Синтезированные в ЭПС липиды передаются в комплекс Гольджи, где «дозревают».
ЭПС представлена в клетках коры надпочечников и в половых железах, поскольку здесь синтезируются стероиды, а стероиды – гормоны липидной природы. К стероидам относится мужской гормон тестостерон, и женский гормон эстрадиол.
Ещё одна функция ЭПС – участие в процессах детоксикации. В клетках печени шероховатая и гладкая ЭПС участвуют в процессах обезвреживания вредных веществ, поступающих в организм. ЭПС удаляет яды из нашего организма.
В мышечных клетках присутствуют особые формы ЭПС – саркоплазматический ретикулум. Саркоплазматический ретикулум – один из видов эндоплазматической сети, который присутствует в поперечнополосатой мышечной ткани. Его основной функцией является хранение ионов кальция, и введение их в саркоплазму – среду миофибрилл.
Секреторная функция комплекса Гольджи
Функцией комплекса Гольджи является транспорт и химическая модификация веществ. Особенно хорошо это видно в секреторных клетках.
В качестве примера можно привести клетки поджелудочной железы, синтезирующие ферменты панкреатического сока, который затем выходит в проток железы, открывающийся в двенадцатиперстную железу.
Исходным субстратом для ферментов служат белки, поступающие в комплекс Гольджи из ЭПС. Здесь с ними происходят биохимические превращения, они концентрируются, упаковываются в мембранные пузырьки и перемещаются к плазматической мембране секреторной клетки. Затем они выделяются наружу посредством экзоцитоза.
Ферменты поджелудочной железы секретируются в неактивной форме, чтобы они не разрушали клетку, в которой образуются. Неактивная форма фермента называется проферментом или энзимогеном. Например, фермент трипсин, образуется в неактивной форме в виде трипсиногена в поджелудочной железе и переходит в свою активную форму – трипсин в кишечнике.
Например, фермент трипсин, образуется в неактивной форме в виде трипсиногена в поджелудочной железе и переходит в свою активную форму – трипсин в кишечнике.
Комплексом Гольджи синтезируется также важный гликопротеин – муцин. Муцин синтезируется бокаловидными клетками эпителия, слизистой оболочки желудочно-кишечного тракта и дыхательных путей. Муцин служит барьером, защищающим расположенные под ним эпителиальные клетки от разных повреждений, в первую очередь, механических.
В желудочно-кишечном тракте эта слизь защищает нежную поверхность эпителиальных клеток от действия грубого комка пищи. В дыхательных путях и желудочно-кишечном тракте муцин защищает наш организм от проникновения патогенов – бактерий и вирусов.
В клетках кончика корня растений комплекс Гольджи секретирует мукополисахаридную слизь, которая облегчает продвижение корня в почве.
В железах на листьях насекомоядных растений, росянки и жирянки (рис. 6), аппарат Гольджи производит клейкую слизь и ферменты, с помощью которых эти растения ловят и переваривают добычу.
Рис. 6. Клейкие листья насекомоядных растений
В клетках растений комплекс Гольджи также участвует в образовании смол, камедей и восков.
Автолиз
Автолиз – это саморазрушение клеток, возникающее вследствие высвобождения содержимого лизосом внутри клетки.
Благодаря этому лизосомы в шутку называют «орудиями самоубийства». Автолиз представляет собой нормальное явление онтогенеза, он может распространяться как на отдельные клетки, так и на всю ткань или орган, как это происходит при резорбции хвоста головастика во время метаморфоза, т. е. при превращении головастика в лягушку (рис. 7).
7).
Рис. 7. Резорбция хвоста лягушки благодаря автолизу в ходе онтогенеза
Автолиз происходит в мышечной ткани, остающейся долго без работы.
Кроме этого, автолиз наблюдается у клеток после гибели, поэтому вы могли наблюдать, как продукты питания сами портятся, если они не были заморожены.
Таким образом, мы рассмотрели основные одномембранные органоиды клетки: ЭПС, комплекс Гольджи и лизосомы, выяснили их функции в процессах жизнедеятельности отдельной клетки и организма в целом. Установили связь между синтезом веществ в ЭПС, транспортом их в мембранных пузырьках в комплекс Гольджи, «дозреванием» веществ в комплексе Гольджи и выделением их из клетки при помощи мембранных пузырьков, в том числе лизосом. Также мы говорили о включениях – непостоянных структурах клетки, которые представляют собой скопления органических веществ (крахмала, гликогена, капель масла или гранул белка).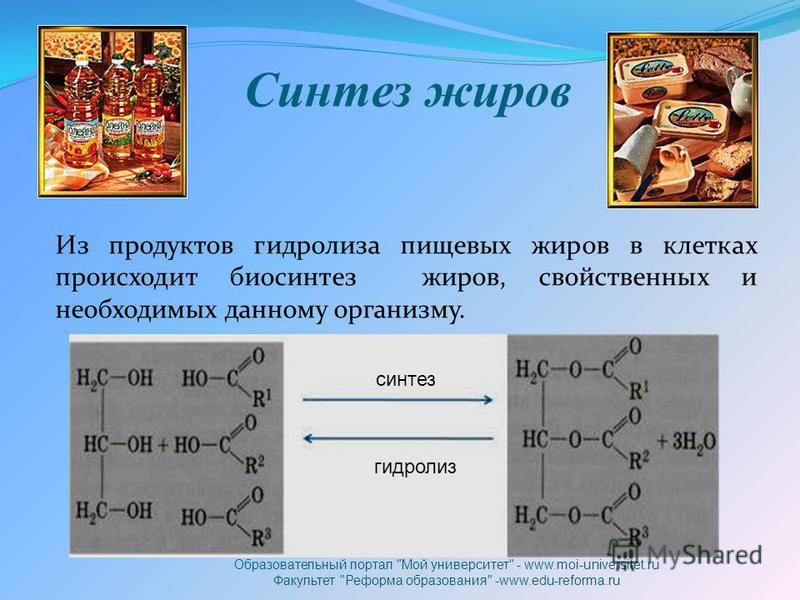 Из приведенных в тексте примеров мы можем сделать вывод о том, что процессы жизнедеятельности, которые происходят на клеточном уровне, отражаются на функционировании целого организма (синтез гормонов, автолиз, накопление питательных веществ).
Из приведенных в тексте примеров мы можем сделать вывод о том, что процессы жизнедеятельности, которые происходят на клеточном уровне, отражаются на функционировании целого организма (синтез гормонов, автолиз, накопление питательных веществ).
источник конспекта — http://interneturok.ru/ru/school/biology/10-klass/bosnovy-citologii-b/stroenie-kletki-kompleks-goldzhi-endoplazmaticheskaya-set-lizosomy-kletochnye-vklyucheniya
источник видео — http://www.youtube.com/watch?v=ROvPqrK8LCw
источник видео — http://www.youtube.com/watch?v=paGe-9Dpee8
источник видео — http://www.youtube.com/watch?v=n-RlcLRuJfM
источник видео — http://www.youtube.com/watch?v=WSdDlT9CrGc
источник видео — http://www.youtube.com/watch?v=k1xbMzVe7Zo
источник видео — http://www.youtube.com/watch?v=v8Fkks2A7js
источник видео — http://www.youtube.com/watch?v=9VeNL-wVTZU
источник видео — http://www.youtube.com/watch?v=8Ku4D_4B9hk
источник презентации — http://www. myshared.ru/slide/151741
myshared.ru/slide/151741
4.12: Эндомембранная система и белки — эндоплазматический ретикулум
- Последнее обновление
- Сохранить как PDF
- Идентификатор страницы
- 12722
- Boundless (теперь LumenLearning)
- Безбрежный
Цели обучения
- Описать структуру эндоплазматического ретикулума и его роль в синтезе и метаболизме
Эндоплазматический ретикулум
Эндоплазматический ретикулум (ЭР) представляет собой серию взаимосвязанных мембранных мешочков и канальцев, которые совместно модифицируют белки и синтезируют липиды. Однако эти две функции выполняются в отдельных областях ЭР: шероховатой ЭР и гладкой ЭР. Полая часть канальцев ЭПР называется просветом или цистернальным пространством. Мембрана ER, представляющая собой двойной слой фосфолипидов, окруженный белками, непрерывна с ядерной оболочкой.
Полая часть канальцев ЭПР называется просветом или цистернальным пространством. Мембрана ER, представляющая собой двойной слой фосфолипидов, окруженный белками, непрерывна с ядерной оболочкой.
Шероховатый ER
Шероховатый эндоплазматический ретикулум (RER) назван так потому, что рибосомы, прикрепленные к его цитоплазматической поверхности, придают ему шиповатый вид при рассмотрении в электронном микроскопе. Рибосомы переносят свои вновь синтезированные белки в просвет RER, где они претерпевают структурные модификации, такие как сворачивание или приобретение боковых цепей. Эти модифицированные белки будут включены в клеточные мембраны — мембраны ER или других органелл — или секретироваться из клетки (например, белковые гормоны, ферменты и т. д.). RER также производит фосфолипиды для клеточных мембран. Если фосфолипидам или модифицированным белкам не суждено остаться в RER, они достигнут места назначения через транспортные везикулы, которые отпочковываются от мембраны RER.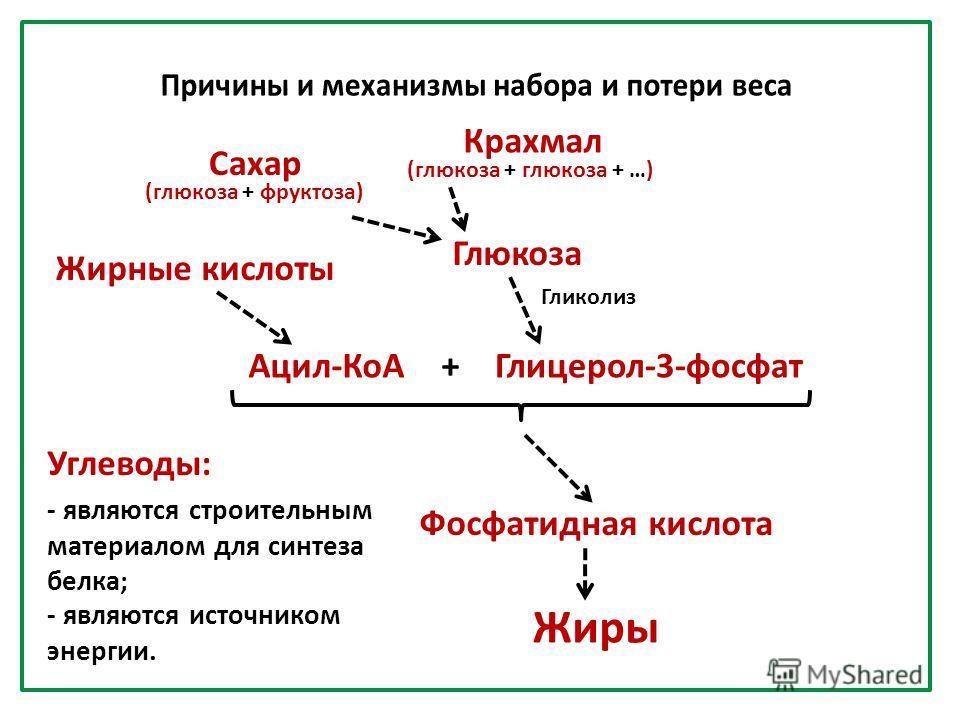 Поскольку RER участвует в модификации белков (таких как, например, ферменты), которые будут секретироваться из клетки, RER в избытке присутствует в клетках, секретирующих белки. Так обстоит дело, например, с клетками печени.
Поскольку RER участвует в модификации белков (таких как, например, ферменты), которые будут секретироваться из клетки, RER в избытке присутствует в клетках, секретирующих белки. Так обстоит дело, например, с клетками печени.
Smooth ER
Гладкий эндоплазматический ретикулум (SER) является продолжением RER, но на его цитоплазматической поверхности имеется мало или совсем нет рибосом. Функции SER включают синтез углеводов, липидов и стероидных гормонов; дезинтоксикация от лекарств и ядов; и хранения ионов кальция. В мышечных клетках специализированный SER, называемый саркоплазматическим ретикулумом, отвечает за хранение ионов кальция, которые необходимы для запуска скоординированных сокращений мышечных клеток.
Ключевые моменты
- Если к эндоплазматическому ретикулуму (ЭР) прикреплены рибосомы, он называется шероховатым ЭР; если это не так, то это называется гладким ER.

- Белки, вырабатываемые шероховатой эндоплазматической сетью, предназначены для использования вне клетки.
- Функции гладкой эндоплазматической сети включают синтез углеводов, липидов и стероидных гормонов; дезинтоксикация от лекарств и ядов; и хранения ионов кальция.

Основные термины
- просвет : полость или канал внутри трубки или трубчатого органа.
- ретикулум : Сеть
- Наверх
- Была ли эта статья полезной?
- Тип изделия
- Раздел или страница
- Автор
- Безграничный
- Количество столбцов печати
- Два
- Печать CSS
- Плотный
- Лицензия
- CC BY-SA
- Версия лицензии
- 4,0
- Показать оглавление
- нет
- Теги
- эндоплазматический ретикулум
Клетка.
 3. Клеточная мембрана. Углеводы. Атлас гистологии растений и животных.
3. Клеточная мембрана. Углеводы. Атлас гистологии растений и животных.Темный
Домашний / Клетка / Клеточная мембрана / Углеводы
- Содержание сайта
- Клетка
- Типы ячеек
- Ткани животных
- Ткани растений
- Органы животных
- Органы растений
- Гистологические методы
- Виртуальная микроскопия
- СОДЕРЖАНИЕ. КЛЕТКА.
- 1. Введение
- Разнообразие клеток
- Открытие клеток
- Клеточная теория
- Происхождение клетки
- Происхождение эукариот
- Эндосимбиоз
- 2. Внеклеточный матрикс
- Структурные белки
- гликопротеины
- Типы
- 3. Клеточная мембрана
- Липиды
- Белки
- Углеводы
- Проницаемость, текучесть
- Асимметрия, ремонт
- Синтез
- Транспорт
- Адгезия
- Клеточные соединения
- 4. Ядро
- Ядерная оболочка
- Ядерная пора
- хроматин
- ядрышко
- 5. Везикулярный трафик
- Эндоплазматический ретикулум
- От ретикулума к Гольджи
- аппарат Гольджи
- экзоцитоз
- эндоцитоз
- эндосомы
- лизосомы
- В растительных клетках
- Вакуоли
- 6. Невезикулярный
- пероксисомы
- Митохондрии
- пластиды
- Хлоропласты
- Липидные капли
- 7. Цитозоль
- Цитоскелет
- Актиновые филаменты
- Микротрубочки
- Промежуточные филаменты
- 8. Клеточный цикл
- Фаза G1
- S-фаза
- Фаза G2
- М фаза. Митоз.
- 9. Мейоз
- Больше информации
- Викторины
- Глоссарий
- Библиография
 Клеточная мембрана
Клеточная мембрана Митоз.
Митоз.УГЛЕВОДЫ
Мембранные углеводы химически связаны с липидами, образуя гликолипиды, и с белками, образуя гликопротеины. Другими мембранными углеводами являются гликозаминогликаны в составе протеогликанов, которые встраивают свою аминокислотную цепь в липидные цепи жирных кислот (рис. 1). Хотя некоторые углеводы можно обнаружить во внутриклеточных мембранах, большинство из них находится во внешнем полуслое плазматической мембраны, обращенном во внеклеточное пространство. Синтез мембранных углеводов начинается в эндоплазматическом ретикулуме, но именно в аппарате Гольджи, где они модифицируются и выращиваются путем добавления множества новых мономеров с образованием сложных углеводных полимеров.
Рисунок 1. Углеводы в плазматической мембране. Гликолипиды в основном представляют собой сфинголипиды с различным углеводным составом. Некоторые протеогликаны имеют часть своей аминокислотной последовательности, вставленную среди цепей липидов жирных кислот. Большинство углеводов химически связаны с белками, известными как гликопротеины, либо посредством О-гликозилирования (через сериновую аминокислоту), либо посредством N-гликозилирования (через аминокислоту аспарагина) (с изменениями из Fuster and Esko, 2005).
Углеводы в плазматической мембране. Гликолипиды в основном представляют собой сфинголипиды с различным углеводным составом. Некоторые протеогликаны имеют часть своей аминокислотной последовательности, вставленную среди цепей липидов жирных кислот. Большинство углеводов химически связаны с белками, известными как гликопротеины, либо посредством О-гликозилирования (через сериновую аминокислоту), либо посредством N-гликозилирования (через аминокислоту аспарагина) (с изменениями из Fuster and Esko, 2005). В мембранах обнаружены три типа гликолипидов: гликосфинголипиды, наиболее распространенные в клетках животных, гликоглицеролипиды и гликофосфатидилинозитол. Гликоглицеролипиды чаще встречаются в плазматической мембране растительных клеток. Однако большинство мембранных углеводов связаны с белками, известными как гликопротеины. Почти все мембранные белки содержат углеводы, но только 5 % липидов составляют гликолипиды. Углеводы плазматической мембраны в целом называются гликокаликсом. В некоторых типах клеток гликокаликс развит настолько, что его можно наблюдать в электронный микроскоп. Например, в энтероцитах гликокаликс может простираться более чем на 1 мкм в длину от поверхности плазмы. Таким образом, клетка покрывается оболочкой из углеводов, которая может составлять от 2 до 10 % массы мембраны. Развитие гликокаликса зависит от типа клеток.
В некоторых типах клеток гликокаликс развит настолько, что его можно наблюдать в электронный микроскоп. Например, в энтероцитах гликокаликс может простираться более чем на 1 мкм в длину от поверхности плазмы. Таким образом, клетка покрывается оболочкой из углеводов, которая может составлять от 2 до 10 % массы мембраны. Развитие гликокаликса зависит от типа клеток.
Мембранные углеводы являются местами клеточного узнавания и адгезии, либо межклеточного узнавания, либо межклеточного взаимодействия с патогеном. Например, группы крови определяются углеводами клеточной поверхности эритроцитов, и они также обладают способностью запускать иммунологические реакции. После инфекции эндотелиальные клетки вблизи поврежденной ткани высвобождают в своих плазматических мембранах тип белков, известных как селектины. Селектины распознают и связывают углеводы плазматической мембраны лимфоцитов, циркулирующих в кровотоке. Таким образом, лимфоциты прикрепляются к стенкам кровеносных сосудов, могут преодолевать эндотелий и перемещаться к очагу инфекции. Углеводы как распознающие молекулы важны во время развития эмбриона. Они также играют структурную роль в качестве физического барьера.
Углеводы как распознающие молекулы важны во время развития эмбриона. Они также играют структурную роль в качестве физического барьера.
Иногда мембранные углеводы модифицируются. Например, раковые клетки демонстрируют различный углеводный набор, и тип раковой клетки можно идентифицировать по различному поверхностному набору углеводов. Эти углеводы могут модулировать способность раковой клетки получать и понимать молекулярную информацию, поступающую от других клеток. Кроме того, тандемное повторение сиаловой кислоты усиливает злокачественные признаки раковых клеток, такие как пролиферация, инвазия, миграция, адгезия и метастазирование. Ганглиозид GD3 обнаружен в меланомах и является мишенью для лечения.
Углеводы плазматической мембраны являются основными местами распознавания и прикрепления патогенов во время инфекции.
Leave A Comment