
Особенности жизненного цикла клетки и три периода интерфазы
Жизненный цикл клетки
Определение жизненного цикла
Почти все клетки характеризуются наличием определенного жизненного цикла.
Что такое жизненный цикл клетки?
Определение 1Жизненный цикл — это промежуток жизни клетки, который начинается в момент ее возникновения и заканчивается, когда завершается деление или клетка гибнет.
На протяжении жизненного цикла в клетке происходит множество различных процессов:
- рост;
- развитие;
- дифференциация;
- функционирование и др.
Из каких этапов состоит клеточный цикл?
Клеточный цикл включает продолжительный период интерфазы, а также короткие периоды митоза и цитокинеза.
Пример 1К примеру, лейкоциты характеризуются 10-минутными митозом и цитокинезом, а также более чем 24-часовым промежутком интерфазы.
Определение 2Интерфаза — это период жизни клетки, на протяжении которого она не делится.
В этот промежуток времени клетки нацелена на поддержание своего гомеостаза и выполнение определенных функций.
В ходе исследований различных групп клеток отдельного организма ученые пришли к выводу, что почти все они находятся в интерфазе. Только 1% клеток задействуется в процессе митоза.
Цикл клетки с делением в конце — характерная особенность большинства разновидностей клеток многоклеточного организма и всех одноклеточных организмов.
Клетки различаются длительностью не только всего цикла, но и отдельных его периодов. Причем эти различия встречаются даже в различных тканях одного организма.
Пример 2Продолжительность клеточного цикла у людей для эпителиальных клеток — от 10 до 20 суток, для лейкоцитов — 4-5 суток, для клеток костного мозга — от 8 до 12 часов.
Сколько будет жить клетка закладывается генетически и передается по наследству.
Существует определенный этап жизнедеятельности клетки, когда происходит образование специальных белковых молекул.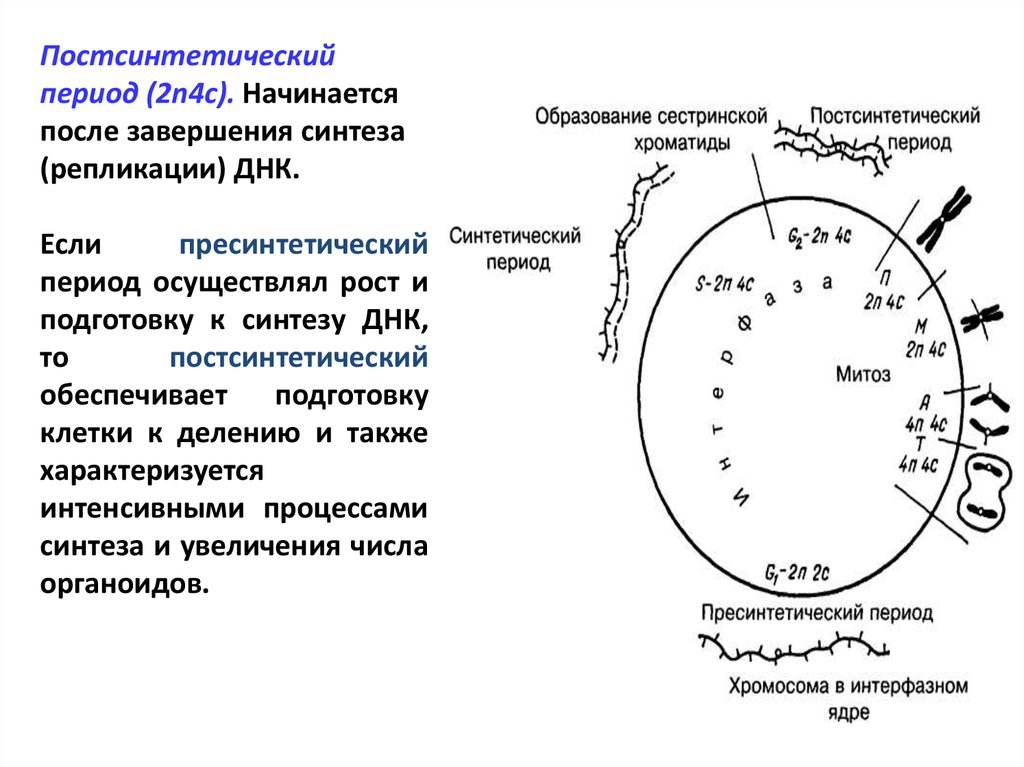 Определенная их концентрация указывает на необходимость деления или гибели.
Определенная их концентрация указывает на необходимость деления или гибели.
Периоды интерфазы
Интерфаза является периодом жизненного цикла клетки, который характеризуется ее жизнью, функционированием и подготовкой к делению.
Замечание 1Окончание предыдущего цитокенеза — это начало интерфазы и всего клеточного цикла.
Пресинтетический период
Первый период интерфазы называется пресинтетическим или G₁. Он характеризуется тем, что закодированная в ДНК генетическая информация находится в состоянии максимального функционирования. ДНК отвечает за синтез РНК и белков. Это наиболее длительный период, во время которого происходит рост клеток, дифференциация с выполнением соответствующих функций.
Ядра клеток содержат диплоидный набор хромосом: в каждой из них находится одна молекула ДНК. Генетическая формула клетки в этот период выглядит так: 2n2c (n обозначает гаплоидный набор хромосом, а с — количество копий ДНК).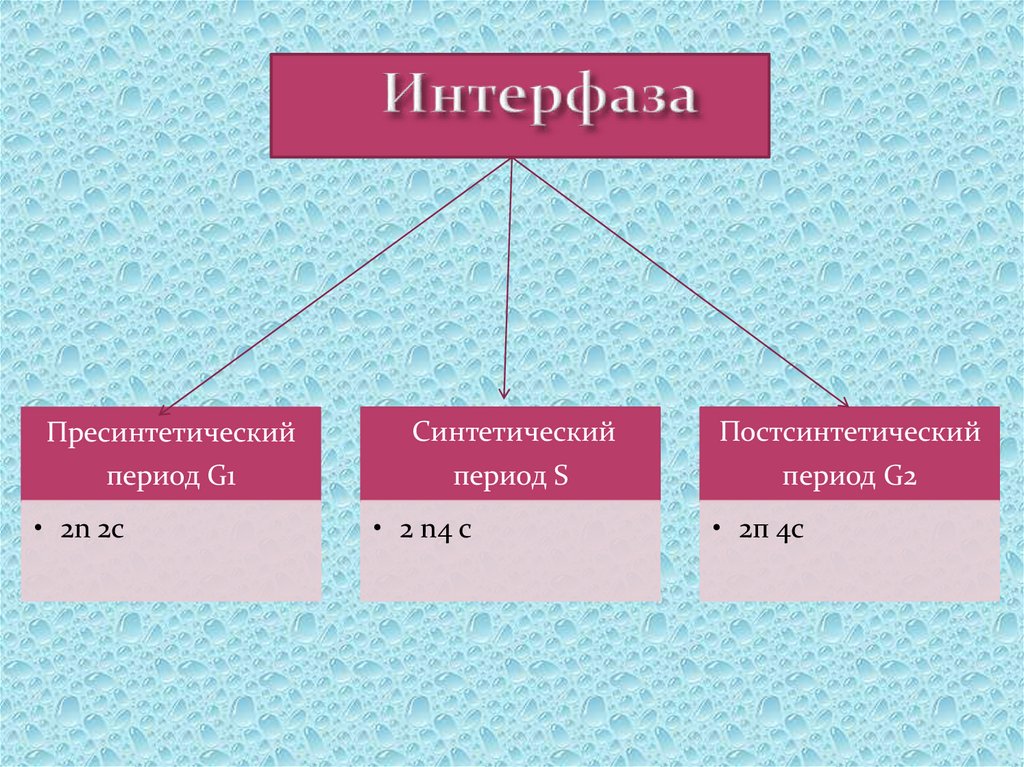
Синтетический период
Следующий период — синтетический или S
Важный момент во время подготовки клетки к делению — репликация ДНК. Именно репликация составляет основу и бесполого, и полового размножения. Поэтому только в этом случае можно говорить о непрерывности жизни.
Начало синтетического периода связано с началом синтеза ДНК. Как только начинается удвоение ДНК, клетка теряет возможность вернуться на предыдущий период и имеет только один вариант — деление.
Начало синтетического периода также называют точкой рестрикции. Запуск синтеза ДНК происходит после появления специальных сигнальных молекул белков-активаторов этой фазы. В конце периода, после полной репликации ДНК, происходит разрушение белка-активатора и переход клетки к следующему периоду.
В конце периода, после полной репликации ДНК, происходит разрушение белка-активатора и переход клетки к следующему периоду.
Клетки, у которых «разрешение» на деление отсутствует, не могут пройти точку рестрикции. Поэтому такие клетки на некоторое время переходят в состояние «покоя» или в G₀-фазу. В это время они поддерживают свой метаболизм и выполняют определенные функции.
Нейроны и мышечные клетки способны функционировать на протяжении всей жизни организма.
Постсинтетический период
Следующий этап — постсинтетический или G₂. На этом этапе клетки готовятся к процессу митоза. Цитоскелет постепенно разрушается, осуществляется конденсация и спирализация хроматина. Отмечается усиление синтеза АТФ, белков, РНК, липидов и углеводов. Также происходит формирование новых клеточных органелл.
Клетки существенно увеличиваются в размерах. Кроме того, осуществляется синтез специальных белков-регуляторов, которые способствуют тому, чтобы клетка из этого этапа перешла к делению.
Постсинтетический период плавно перетекает в профазу митоза. Это момент, когда с помощью светового микроскопа впервые можно заметить хромосомы, сформированные из хроматина.
Окружающие клетки и гуморальные факторы организма контролирует жизненный цикл клеток многоклеточного организма. Большая роль в процессе регуляции отводится также специальным белкам, которые образует сама клетка под влиянием собственной генетической программы.
Какие процессы происходят в клетке в период интерфазы?
Если перечислять, какие процессы происходят в клетке в период интерфазы и готовят клетку к делению, то обязательно стоит упомянуть спирализацию и сокращение половинок хромосом (хроматид), удвоение центриолей, синтез белков будущего ахроматинового веретена, синтез высокоэнергетических соединений (в частности, АТФ).
Рост клетки завершается. Клетка готова вступить в профазу следующего митоза.
Цитокенез
За митозом следует такой этап клеточного цикла как цитокенез или деление цитоплазмы.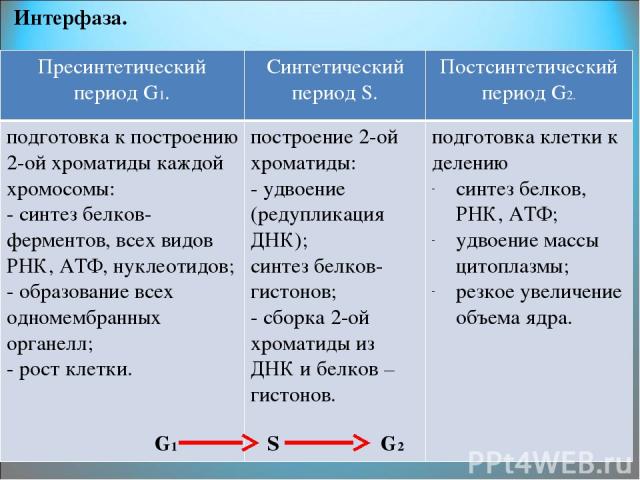
Начинается все с образования по экватору материнской клетки животных организмов перетяжки. Она, в частности, формируется еще в телофазе митоза. Микрофилламенты цитоскелета, образующие сократительное кольцо, формируют перетяжку деления. Кольцо постепенно становится меньше, а перетяжка все больше углубляется по всему периметру. Через определенный промежуток времени происходит деление материнской клетки на две дочерние.
Цитоскелет принимает активное участие в образовании перетяжки и ее углублении, в полном делении дочерних клеток. По окончании цитокенеза в дочерних клетках есть все компоненты материнской клетки.
В случае, если после митоза не происходит цитокенез, образуются многоядерные клетки.
1.1. Пресинтетический период (g1)
Пресинтетический
или постмитотический (G1) период (от англ.
gap — промежуток) наступает сразу же после
митотического деления клетки и
характеризуется активным ростом клетки
и синтезом белка и РНК, благодаря чему
клетка достигает нормальных размеров
и восстанавливает необходимый набор
органелл. G1 -период длится от нескольких
часов до нескольких дней. В течение
этого периода синтезируются особые
«запускающие» белки (trigger proteins), или
активаторы S-периода. Они обеспечивают
достижение клеткой определенного порога
(точки R — рестрикции или ограничения),
после которого она вступает в S-период.
G1 -период длится от нескольких
часов до нескольких дней. В течение
этого периода синтезируются особые
«запускающие» белки (trigger proteins), или
активаторы S-периода. Они обеспечивают
достижение клеткой определенного порога
(точки R — рестрикции или ограничения),
после которого она вступает в S-период.
Контроль, осуществляемый на уровне точки R (при переходе из G1 в S), ограничивает возможность нерегулируемого размножения клеток. Проходя эту точку, клетка переключается на последующую регуляцию внутренними факторами клеточного цикла, которая обеспечивает закономерное завершение ее деления.
Если клетка не
достигает точки R, она выходит из цикла
и вступает в период репродуктивного
покоя (G0) для того, чтобы (в зависимости
от причин остановки): дифференцироваться
и выполнять свои специфической функции,
выжить в условиях недостаточности
питательных веществ или факторов роста,
осуществить репарацию поврежденной
ДНК. Клетки одних тканей при соответствующей
стимуляции вновь способны возвращаться
из периода (G0) в клеточный цикл, других
— утрачивают эту способность по мере
дифференцировки.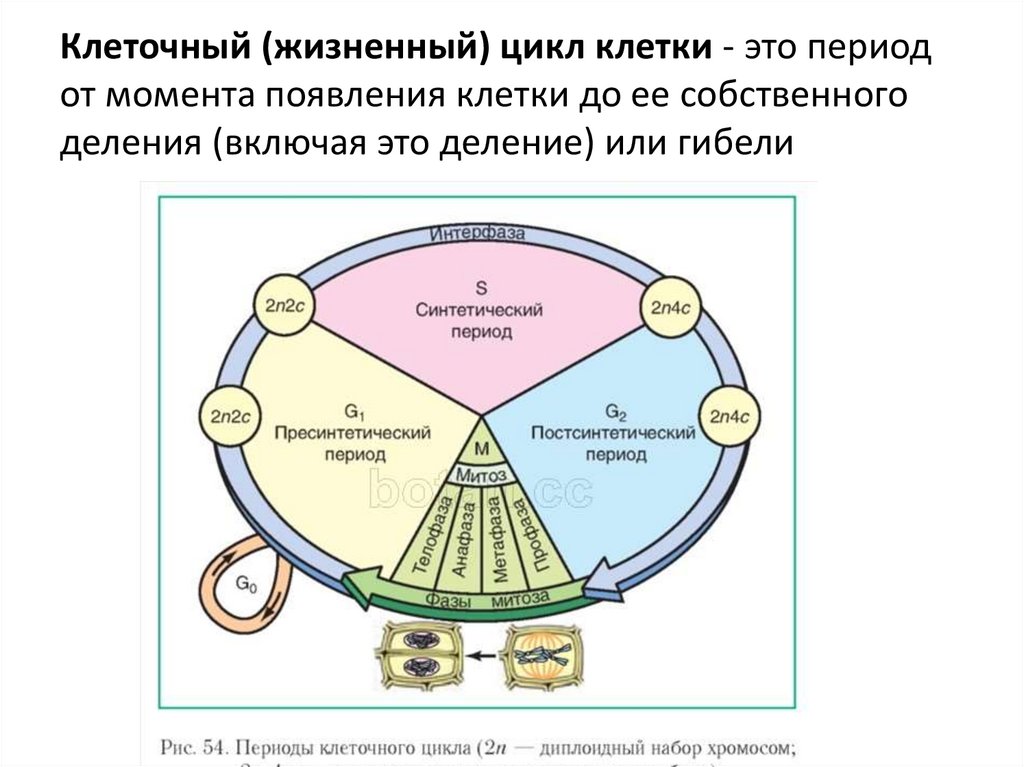
1.2. Синтетический период (s)
Синтетический (S-) период характеризуется удвоением содержания (репликацией) ДНК и синтезом белков, в частности, гистонов, которые поступают в ядро из цитоплазмы и обеспечивают нуклеосомную упаковку вновь синтезированной ДНК. В результате происходит удвоение числа хромосом. Одновременно удваивается число центриолей. S-период длится у большинства клеток 8-12 часов.
1.3. Постсинтетический период (g2)
Постсинтетический
или премитотический (G2-)период следует
за S-периодом и продолжается вплоть до
митоза (часто обозначаемого буквой М).
В течение этого периода клетка осуществляет
непосредственную подготовку к делению.
Происходит созревание центриолей,
запасается энергия, синтезируются РНК
и белки (в частности, тубу-лин), необходимые
для процесса деления. Длительность
G2-периода составляет 2-4 часа. Возможность
выхода клетки из G2-периода в G0-neриод с
последующим возвращением в G2-период в
настоящее время большинством авторов
отрицается.
1.4. Деление клетки
Итак, если всё в порядке, ДНК реплицировалась удачно, то наступает G2-период, финишная прямая. Во время него опять проверяется качество ДНК, клетка увеличивает свою массу. И при условии, что все внешние и внутренние факторы способствуют переходу G2/M, наступает митоз.
Для того, чтобы две дочерние клетки поровну получили генетический материал, организация процесса деления должна быть удобной и четкой. Хромосомы вне деления находятся в деконденсированном виде, т.е. представляют собой чрезвычайно длинные нити, рассредоточенные по всему ядру. Естественно, удобно разделить ДНК у каждой из них поровну просто не получится, поэтому первым делом хромосомы спирализуются и приобретают характерный вид, в это же время образуются абсолютно необходимые для деления органеллы — центросомы. Затем происходит разрушение ядерной оболочки, что знаменует начало следующей по списку стадии, прометафазы.
Микротрубочки,
растущие из центросом, соединяются с
хромосомами, образуется веретено деления
и какое-то время совершается серия
сложных движений, названных конгрессией —
прикрепленные к микротрубочкам хромосомы
движутся от и к полюсам, до тех пор, пока
не выстроятся ровно на экваторе клетки,
образуя метафазную пластинку.
Веретено деления — это уникальная структура, существующая только во время деления клетки, совершенно необходимая как для разделения хромосом, так и для разделения цитоплазмы дочерних клеток. Состоит она из центросом, состоящих из двух центриолей каждая, располагающихся в норме у человека на двух противоположных полюсах, микротрубочек, растущих от них в направлении к центру клетки — к хромосомам, астральных микротрубочек, идущих от центросом во всех направлениях, но не к хромосомам, белковых моторов, а также из самих хромосом.
В делении задействован
особый участок хромосомы — центромера,
представляющая из себя комплекс из ДНК
и хроматина, к которой прикрепляется
кинетохор, сложная структура, служащая
для связывания с микротрубочкам веретена.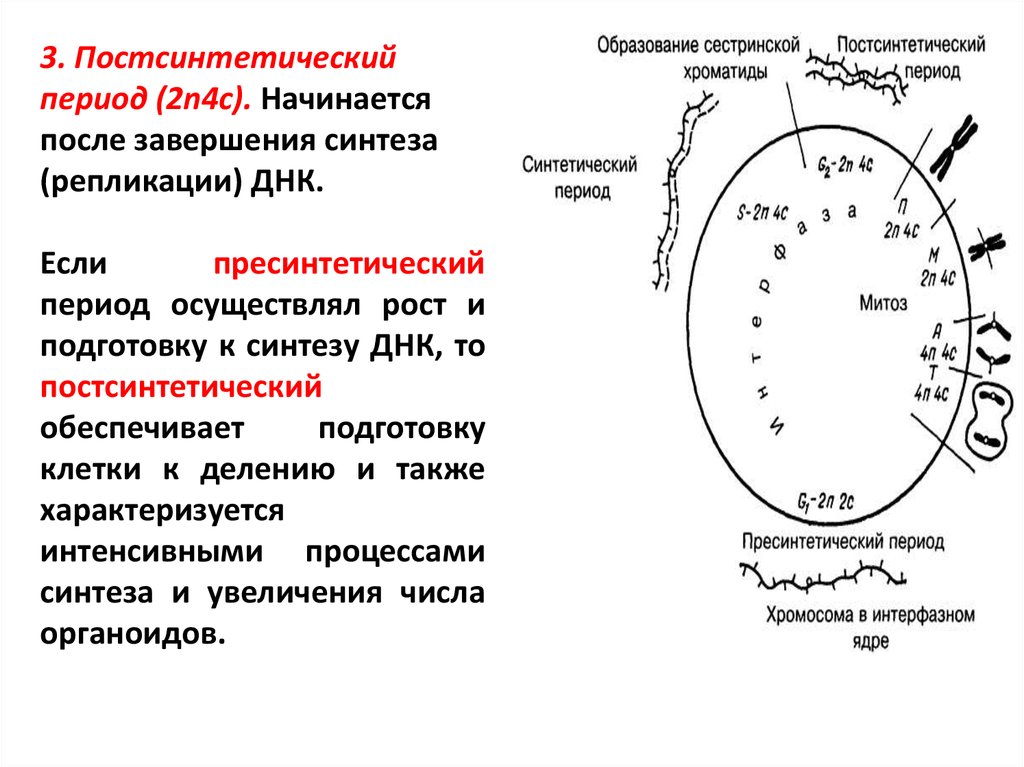 Центромера состоит из специфических
участков, названных CEN, и с ними связываются
соответствующие белки, CENP. Последние
остаются с хромосомами до самого конца,
но есть ещё и «хромосомные пассажиры»,
меняющие свою локализацию в ходе деления,
к ним относят в первую очередь некоторые
киназы. Кинетохоров у каждой хромосомы
два, и они находятся на противоположных
сторонах центромеры, повернуты друг от
друга, и основная их функция — захват
и стабилизация микротрубочек. Первое
осуществляет наружная часть кинетохора,
корона, имеющая вид щупалец, а второе —
наружная пластинка (См. Рисунок 3).
Центромера состоит из специфических
участков, названных CEN, и с ними связываются
соответствующие белки, CENP. Последние
остаются с хромосомами до самого конца,
но есть ещё и «хромосомные пассажиры»,
меняющие свою локализацию в ходе деления,
к ним относят в первую очередь некоторые
киназы. Кинетохоров у каждой хромосомы
два, и они находятся на противоположных
сторонах центромеры, повернуты друг от
друга, и основная их функция — захват
и стабилизация микротрубочек. Первое
осуществляет наружная часть кинетохора,
корона, имеющая вид щупалец, а второе —
наружная пластинка (См. Рисунок 3).
После того как
ядро разрушилось, хромосомы вырываются
на волю и норовят рассредоточиться по
всей цитоплазме. В это же время центросомы
начинают во всех направлениях сразу
активно синтезировать микротрубочки,
чтобы точно не промахнуться. Если
микротрубочка не нашла свой кинетохор,
она диссоциирует, если же нашла, то
последний захватывает ее, и начинает
перемещаться в направлении полюса,
попутно связывая и другие, в результате
чего образуется нить кинетохора.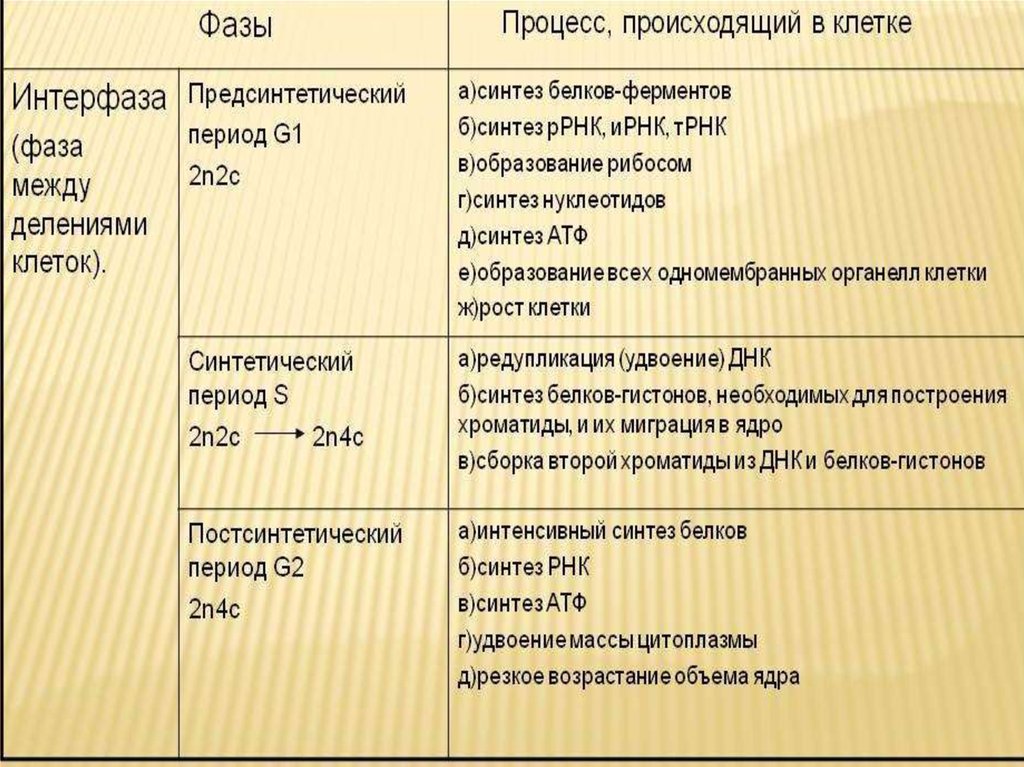 В
какой-то момент кинетохор противоположной
стороны также связывает какую-нибудь
микротрубочку.
В
какой-то момент кинетохор противоположной
стороны также связывает какую-нибудь
микротрубочку.
Движение хромосом,
вообще, очень сложный процесс, и даже
на данный момент неизвестно точно, как
это происходит, но есть несколько теорий,
а также свеженькая (2017 года) статья,
авторы которой предлагают объединить
оба главных предположения в одно:
гипотезу Остергрена, что кинетохор
находится на конце микротрубочки, а
моторы на полюсах подтягивают ее к себе,
причем усилие прямо пропорционально
длине микротрубочки — таким образом
хромосомы стремятся к экватору, где
будет наблюдаться равновесие сил; и
модель «пакман», в которой основное
усилие создают моторы на кинетохорах,
а концы микротрубочек, связанные с ними,
диссоциируют, т.е. кинетохор как бы
«поедает» кинетохорную нить. Также
в движении участвуют моторные белки,
хромокинезины, находящиеся на плечах
хромосом, а также отталкивающая сила
микротрубочек, неприкреплённых к
кинетохорам (таким образом, при движении
хромосомы к полюсу, её плечи будут
отталкиваться растущими навстречу
микротрубочками). В принципе. найдено
более ста белков, участвующих в конгрессии
хромосом, но точная функция установлена
лишь для немногих.
В принципе. найдено
более ста белков, участвующих в конгрессии
хромосом, но точная функция установлена
лишь для немногих.
Когда все хромосомы выстроились на экваторе клетки, белки-пассажиры, проверяют, достаточно ли микротрубочек прикрепилось к каждому кинетохору. И до тех пор, пока «достаточно» не станет, анафаза не не начнётся.
Анафазу условно
делят на анафазу А и анафазу В. Но это
не стадии этапа митоза, а скорее процессы,
происходящие в неё. В анафазу А расходятся
к полюсам хромосомы, в анафазу В — сами
полюса. Но процессы эти идут практически
одновременно. Первым делом хромосомы
разделяются на две хроматиды, как раз
в области центромеры, таким образом,
что каждая хроматида обладает одним
кинетохором. Те же силы, что управляют
движением хромосом во время прометафазы,
управляют движением и во время анафазы,
но теперь ничто не мешает хроматиде
спокойно двигаться к полюсу. В этом
также участвуют многочисленные моторы,
располагающиеся между микротрубочками,
связанными с кинетохорами, между ними
и микротрубочками, с ними не связанными,
моторы полюсов.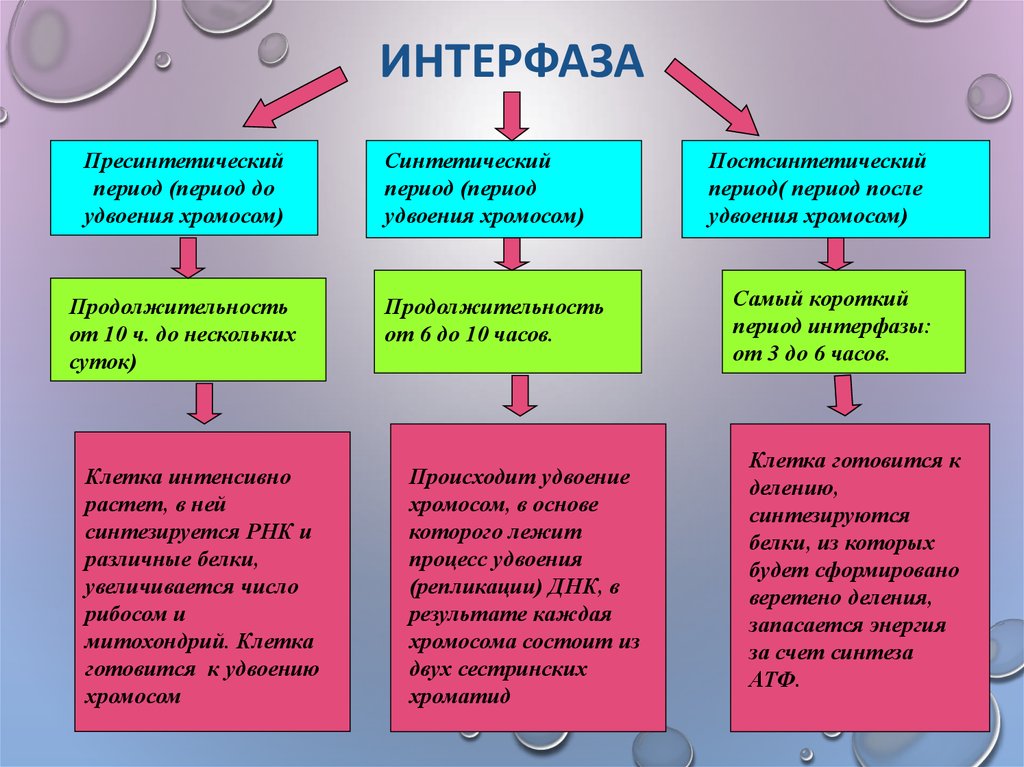 (См. Рисунок 4)
(См. Рисунок 4)
Во время анафазы В моторы, находящиеся на закрепленных у цитолеммы концах астральных микротрубочек, притягивают полюс к себе, а кинезиноподобные белки вроде Eg5, соединяясь с микротрубочками, идущими от противоположных полюсов, отталкивают микротрубочки от центра, и, таким образом, расходятся и сами полюса. В анафазу происходит и образование толстого пучка микротрубочек между расходящимися хромосомами, который после получит название остаточного тельца.
Далее наступает
телофаза, в которую образуются две
дочерние клетки. Процесс непосредственного
разделения цитоплазмы на две части
носит названия цитокинеза и является
завершающим этапом митоза. Для него
необходимо три элемента: веретено
деления (оно никуда не делось после
расхождения хромосом), остаточное тельце
и сократимое кольцо. Последнее представляет
собой пучок актиновых филаментов,
связанных между собой миозинами, и
находится оно прям под цитолеммой,
образуя кольцо в области экватора.
Веретено деления необходимо для
образования в нужном месте остаточного
тельца, а оно уже, в свою очередь, даёт
клетке понять, где именно синтезировать
сократимое кольцо.
Образовавшись, кольцо тут же начинает сокращение, благодаря взаимодействию актина и миозина II. Плазмалемма втягивается, образуется «бороздка деления». Интересен тот факт, что толщина сократимого кольца не увличивается при сокращениии, а остаётся неизменной до самого конца, что достигается за счёт постоянной диссоциации белков, его образующих. Итак, кольцо сжимается-сжимается, и в итоге дочерние клетки становятся, связаны друг с другом лишь перемычкой — остаточным тельцем, которое может существовать ещё долгое время. В случае, если какой-то генетический материал не до конца разошёлся, а остался посреди клетки, бороздка деления пропадает, и образуется двухядерная клетка, если же всё пошло по плану, то перемычка разрушается, так или иначе, и деление тем самым завершается.
определение слова синтез+период по Медицинскому словарю
Синтез+период | определение синтез+период по Медицинскому словарюСинтез+период | определение синтез+период по Медицинскому словарю
Слово, не найденное в Словаре и Энциклопедии.
Возможно, Вы имели в виду:
Пожалуйста, попробуйте слова по отдельности:
синтез период
Некоторые статьи, соответствующие вашему запросу:
Не можете найти то, что ищете? Попробуйте выполнить поиск по сайту Google или помогите нам улучшить его, отправив свое определение.Полный браузер ?
- ▲
- Рабочая группа по синтезу и моделированию
- Синтез и системная интеграция смешанной информации
- Отжиг стренги в зависимости от синтеза
- Synthesis Design Group, Inc.
- Коалиция инженерного образования Synthesis
- синтез-газ
- синтез-газ
- синтез-газ
- Синтез изображений
- Синтез изображений
- Синтез самолетов
- синтез непрерывности
- Синтез приложений баз данных и мультимедийных приложений
- Синтез алгоритмов обработки изображений и речи на кремнии
- синтез обучения
- синтез обучения
- синтез обучения
- синтез обучения
- Синтез механизмов
- Синтез благородных металлов
- Синтез драгоценных металлов
- Синтез технологий моделирования
- Синтез доктрины расы
- период синтеза
- Фаза синтеза
- Реакция синтеза
- Синтез Солюшнс, Инк.

- Синтез Телескоп
- Синтез Телескоп
- Набор инструментов для синтеза
- синтез+период
- Синтез, художественный
- Синтез, химический
- Синтез-Городское Святилище
- синтез
- синтез
- синтез
- синтез
- синтез
- синтез
- Синтезированный

- Синтезированный
- Синтезированный
- синтезатор
- синтезатор
- синтезатор
- Синтезаторы
- Синтезаторы
- синтезирует
- синтезирует
- синтезирует
- синтезирующий
- синтезирующий
- синтезирующий
- синтез
- синтез
- синтез
- синтез
- синтезаторы
- синтезаторы
- синтезаторы
- ▼
Сайт: Следовать:
Делиться:
Открыть / Закрыть
Эукариотический клеточный цикл — Клетка
Цикл деления большинства клеток состоит из четырех скоординированных процессов: клеточного роста, репликации ДНК, распространения дуплицированных хромосом в дочерние клетки и клеточного деления.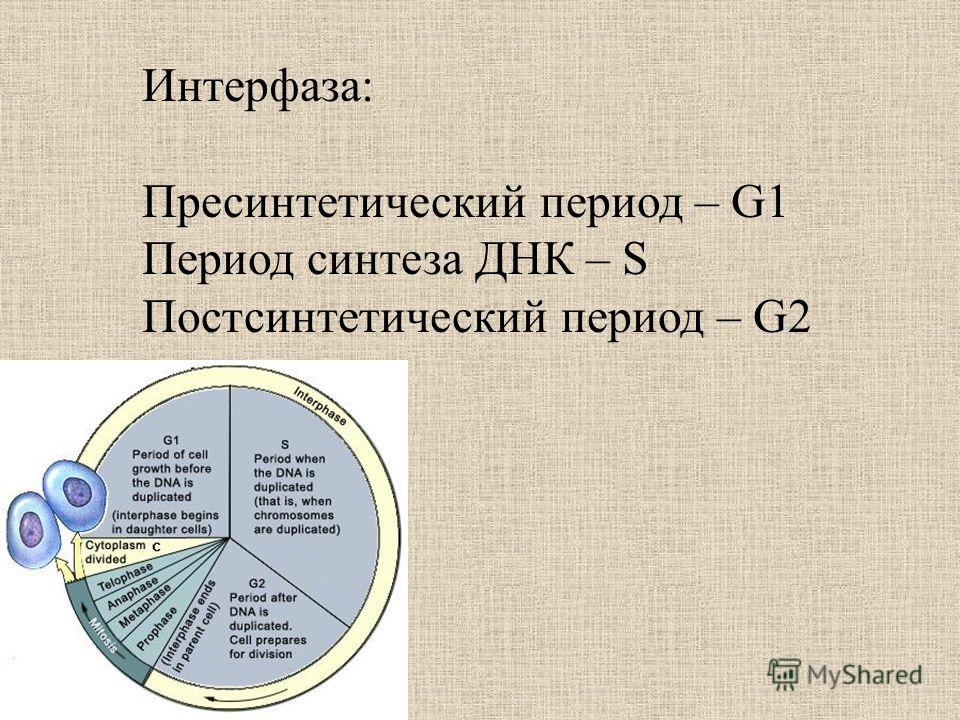 У бактерий клеточный рост и репликация ДНК происходят на протяжении большей части клеточного цикла, а дублированные хромосомы распределяются между дочерними клетками вместе с плазматической мембраной. Однако у эукариот клеточный цикл более сложен и состоит из четырех дискретных фаз. Хотя клеточный рост обычно представляет собой непрерывный процесс, ДНК синтезируется только во время одной фазы клеточного цикла, а затем реплицированные хромосомы распределяются по дочерним ядрам в результате сложной серии событий, предшествующих клеточному делению. Переход между этими стадиями клеточного цикла контролируется консервативным регуляторным аппаратом, который не только координирует различные события клеточного цикла, но также связывает клеточный цикл с внеклеточными сигналами, которые контролируют клеточную пролиферацию.
У бактерий клеточный рост и репликация ДНК происходят на протяжении большей части клеточного цикла, а дублированные хромосомы распределяются между дочерними клетками вместе с плазматической мембраной. Однако у эукариот клеточный цикл более сложен и состоит из четырех дискретных фаз. Хотя клеточный рост обычно представляет собой непрерывный процесс, ДНК синтезируется только во время одной фазы клеточного цикла, а затем реплицированные хромосомы распределяются по дочерним ядрам в результате сложной серии событий, предшествующих клеточному делению. Переход между этими стадиями клеточного цикла контролируется консервативным регуляторным аппаратом, который не только координирует различные события клеточного цикла, но также связывает клеточный цикл с внеклеточными сигналами, которые контролируют клеточную пролиферацию.
Фазы клеточного цикла
Типичный эукариотический клеточный цикл иллюстрируется клетками человека в культуре, которые делятся примерно каждые 24 часа. С точки зрения микроскопа клеточный цикл делится на две основные части: митоз и интерфазу. Митоз (деление ядра) — наиболее драматичная стадия клеточного цикла, соответствующая расхождению дочерних хромосом и обычно заканчивающаяся клеточным делением (цитокинез). Однако митоз и цитокинез длятся всего около часа, т. е. примерно 95% клеточного цикла проходит в интерфазе — периоде между митозами. Во время интерфазы хромосомы деконденсируются и распределяются по ядру, поэтому ядро выглядит морфологически однородным. Однако на молекулярном уровне интерфаза — это время, в течение которого и рост клеток, и репликация ДНК происходят упорядоченным образом при подготовке к клеточному делению.
Митоз (деление ядра) — наиболее драматичная стадия клеточного цикла, соответствующая расхождению дочерних хромосом и обычно заканчивающаяся клеточным делением (цитокинез). Однако митоз и цитокинез длятся всего около часа, т. е. примерно 95% клеточного цикла проходит в интерфазе — периоде между митозами. Во время интерфазы хромосомы деконденсируются и распределяются по ядру, поэтому ядро выглядит морфологически однородным. Однако на молекулярном уровне интерфаза — это время, в течение которого и рост клеток, и репликация ДНК происходят упорядоченным образом при подготовке к клеточному делению.
Клетка растет с постоянной скоростью на протяжении всей интерфазы, при этом большинство делящихся клеток удваиваются в размере между одним митозом и другим. Напротив, ДНК синтезируется только в течение части интерфазы. Таким образом, синхронизация синтеза ДНК делит цикл эукариотических клеток на четыре дискретные фазы (4). Фаза М цикла соответствует митозу, за которым обычно следует цитокинез. За этой фазой следует G 1 фаза (пробел 1), которая соответствует интервалу (пробелу) между митозом и инициацией репликации ДНК. Во время G 1 клетка метаболически активна и постоянно растет, но не реплицирует свою ДНК. За G 1 следует S-фаза (синтез), во время которой происходит репликация ДНК. За завершением синтеза ДНК следует фаза (пробел 2), во время которой продолжается рост клеток и происходит синтез белков для подготовки к митозу.
За этой фазой следует G 1 фаза (пробел 1), которая соответствует интервалу (пробелу) между митозом и инициацией репликации ДНК. Во время G 1 клетка метаболически активна и постоянно растет, но не реплицирует свою ДНК. За G 1 следует S-фаза (синтез), во время которой происходит репликация ДНК. За завершением синтеза ДНК следует фаза (пробел 2), во время которой продолжается рост клеток и происходит синтез белков для подготовки к митозу.
Рисунок 14.1
Фазы клеточного цикла. Цикл деления большинства эукариотических клеток делится на четыре отдельные фазы: M, G 1 , S и G 2 . Фаза М (митоз) обычно сопровождается цитокинезом. S-фаза – это период, в течение которого происходит репликация ДНК. Клетка растет (подробнее…)
Продолжительность этих фаз клеточного цикла значительно различается у разных типов клеток. Для типичной быстро пролиферирующей клетки человека с общим временем цикла 24 часа G 9Фаза 0174 1 может длиться около 11 часов, фаза S — около 8 часов, фаза G 2 — около 4 часов, а фаза M — около 1 часа. Однако другие типы клеток могут делиться намного быстрее. Почкующиеся дрожжи, например, могут пройти все четыре стадии клеточного цикла всего за 90 минут. Еще более короткие клеточные циклы (30 минут или меньше) встречаются в ранних эмбриональных клетках вскоре после оплодотворения яйцеклетки (14). Однако в этом случае роста клеток не происходит. Вместо этого эти ранние эмбриональные клеточные циклы быстро делят цитоплазму яйца на более мелкие клетки. Г 9 нет0174 1 или G 2 , и репликация ДНК происходит очень быстро в этих ранних эмбриональных клеточных циклах, которые поэтому состоят из очень коротких S фаз, чередующихся с M фазами.
Однако другие типы клеток могут делиться намного быстрее. Почкующиеся дрожжи, например, могут пройти все четыре стадии клеточного цикла всего за 90 минут. Еще более короткие клеточные циклы (30 минут или меньше) встречаются в ранних эмбриональных клетках вскоре после оплодотворения яйцеклетки (14). Однако в этом случае роста клеток не происходит. Вместо этого эти ранние эмбриональные клеточные циклы быстро делят цитоплазму яйца на более мелкие клетки. Г 9 нет0174 1 или G 2 , и репликация ДНК происходит очень быстро в этих ранних эмбриональных клеточных циклах, которые поэтому состоят из очень коротких S фаз, чередующихся с M фазами.
Рисунок 14.2
Эмбриональные клеточные циклы. Ранние эмбриональные клеточные циклы быстро делят цитоплазму яйца на более мелкие клетки. Клетки не растут во время этих циклов, в которых отсутствуют G 1 и G 2 и которые состоят просто из коротких S фаз, чередующихся с M фазами.
В отличие от быстрой пролиферации эмбриональных клеток, некоторые клетки у взрослых животных полностью прекращают деление (например, нервные клетки), а многие другие клетки делятся только изредка, по мере необходимости, чтобы заменить клетки, которые были потеряны из-за повреждения или гибели клеток. К клеткам последнего типа относятся кожные фибробласты, а также клетки многих внутренних органов, таких как печень, почки и легкие. Как обсуждается далее в следующем разделе, эти клетки выходят из G 1 , чтобы войти в стадию покоя цикла, называемую 9.0172 G 0 , где они остаются метаболически активными, но больше не размножаются, если к этому не призывают соответствующие внеклеточные сигналы.
К клеткам последнего типа относятся кожные фибробласты, а также клетки многих внутренних органов, таких как печень, почки и легкие. Как обсуждается далее в следующем разделе, эти клетки выходят из G 1 , чтобы войти в стадию покоя цикла, называемую 9.0172 G 0 , где они остаются метаболически активными, но больше не размножаются, если к этому не призывают соответствующие внеклеточные сигналы.
Анализ клеточного цикла требует идентификации клеток на различных стадиях, описанных выше. Хотя митотические клетки можно различить под микроскопом, клетки других фаз цикла (G 1 , S и G 2 ) необходимо идентифицировать по биохимическим критериям. Клетки в S-фазе можно легко идентифицировать, поскольку они содержат радиоактивный тимидин, который используется исключительно для синтеза ДНК (4). Например, если популяция быстро пролиферирующих клеток человека в культуре подвергается воздействию радиоактивного тимидина в течение короткого периода времени (например, 15 минут), а затем анализируется радиоавтографией, то обнаруживается, что примерно треть клеток имеет радиоактивную метку.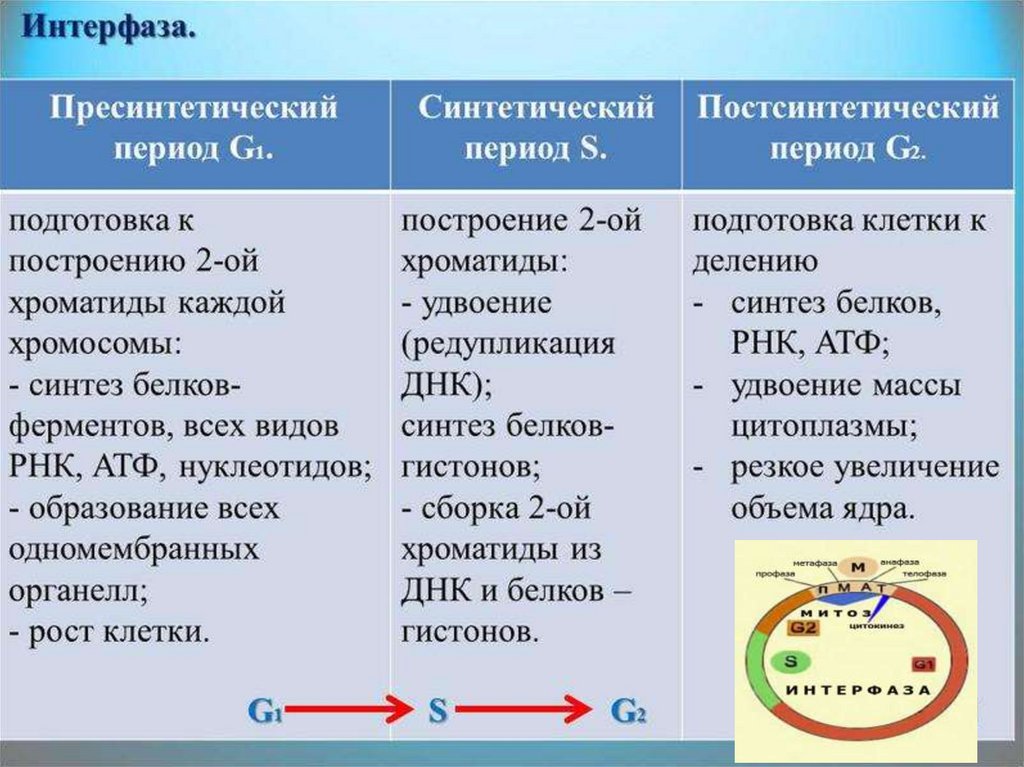 соответствует доле клеток в S-фазе.
соответствует доле клеток в S-фазе.
Рисунок 14.3
Идентификация клеток S фазы путем включения радиоактивного тимидина. Клетки подвергали воздействию радиоактивного тимидина и анализировали авторадиографией. Помеченные ячейки указаны стрелками. (Из D.W. Stacey et al., 1991. Mol. Cell Biol. 11: 4053.) (подробнее…)
Вариации таких экспериментов по маркировке клеток также можно использовать для определения длины различных стадий клетки. цикл. Например, рассмотрим эксперимент, в котором клетки подвергают воздействию радиоактивного тимидина в течение 15 минут, после чего радиоактивный тимидин удаляют и клетки культивируют в течение различных периодов времени перед авторадиографией. Радиоактивно меченые интерфазные клетки, находившиеся в S-фазе во время воздействия радиоактивного тимидина, будут наблюдаться в течение нескольких часов по мере их прохождения через остаток S и G 2 . Напротив, радиоактивно меченные митотические клетки не будут наблюдаться до 4 часов после мечения. Это 4-часовое запаздывание соответствует продолжительности G 2 — минимальному времени, необходимому для того, чтобы клетка, включившая радиоактивный тимидин в конце S-фазы, вступила в митоз.
Это 4-часовое запаздывание соответствует продолжительности G 2 — минимальному времени, необходимому для того, чтобы клетка, включившая радиоактивный тимидин в конце S-фазы, вступила в митоз.
Клетки на разных стадиях клеточного цикла также можно отличить по содержанию ДНК (). Например, клетки животных в G 1 являются диплоидными (содержат две копии каждой хромосомы), поэтому содержание их ДНК обозначается как 2 n ( n обозначает содержание гаплоидной ДНК в геноме). Во время фазы S репликация увеличивает содержание ДНК в клетке с 2 90 238 n 90 239 до 4 90 238 n 90 239 , поэтому клетки в S имеют содержание ДНК в диапазоне от 2 90 238 n 90 239 до 4 90 238 n 90 239 . Затем содержание ДНК остается на уровне 4 н для клеток G 2 и М, снижаясь до 2 н после цитокинеза. Экспериментально содержание клеточной ДНК можно определить путем инкубации клеток с флуоресцентным красителем, связывающимся с ДНК, с последующим анализом интенсивности флуоресценции отдельных клеток в проточном цитометре или активируемый флуоресценцией клеточный сортировщик , тем самым различая клетки в фазах G 1 , S и G 2 /M клеточного цикла.
Рисунок 14.4
Определение содержания клеточной ДНК. Популяцию клеток метят флуоресцентным красителем, который связывает ДНК. Затем клетки пропускают через проточный цитометр, который измеряет интенсивность флуоресценции отдельных клеток. Данные представлены в виде клетки (подробнее…)
Регуляция клеточного цикла с помощью роста клеток и внеклеточных сигналов
Прохождение клеток через цикл деления регулируется внеклеточными сигналами из окружающей среды, а также внутренними сигналами, которые отслеживают и координируют различные процессы, происходящие на разных фазах клеточного цикла. Примером регуляции клеточного цикла внеклеточными сигналами может служить влияние факторов роста на пролиферацию клеток животных. Кроме того, различные клеточные процессы, такие как клеточный рост, репликация ДНК и митоз, должны координироваться во время развития клеточного цикла. Это достигается серией контрольных точек, которые регулируют прохождение через различные фазы клеточного цикла.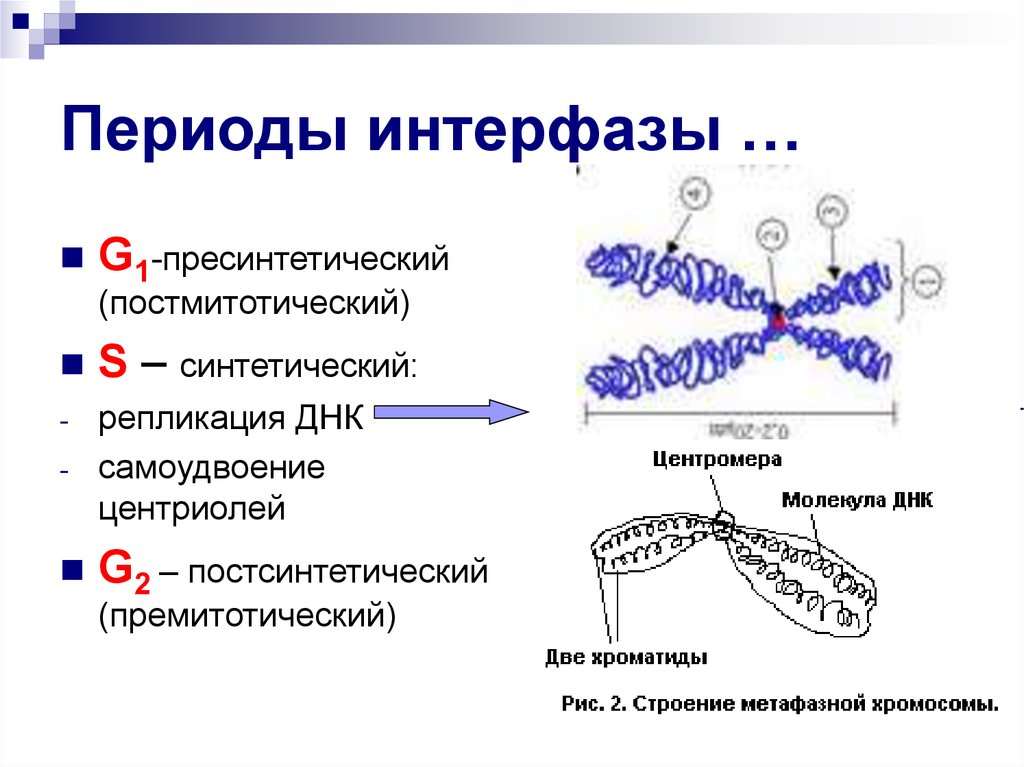
Основная точка регуляции клеточного цикла во многих типах клеток возникает поздно в G 1 и контролирует переход от G 1 к S. Эта регуляторная точка была впервые определена в исследованиях почкующихся дрожжей ( Saccharomyces cerevisiae ), где он известен как СТАРТ (). После того, как клетки прошли СТАРТ, они готовы вступить в S-фазу и пройти один цикл клеточного деления. Однако прохождение через START является строго регулируемым событием в клеточном цикле дрожжей, где оно контролируется внешними сигналами, такими как доступность питательных веществ, а также размером клетки. Например, если дрожжи сталкиваются с нехваткой питательных веществ, они останавливают свой клеточный цикл в момент НАЧАЛА и переходят в состояние покоя, а не в S-фазу. Таким образом, СТАРТ представляет собой точку принятия решения, в которой клетка определяет, доступно ли достаточное количество питательных веществ для поддержки прохождения оставшейся части цикла деления. Полипептидные факторы, которые сигнализируют о спаривании дрожжей, также останавливают клеточный цикл в START, позволяя гаплоидным дрожжевым клеткам сливаться друг с другом вместо перехода в S-фазу.
Рисунок 14.5
Регуляция клеточного цикла почкующихся дрожжей. (A) Клеточный цикл Saccharomyces cerevisiae регулируется главным образом в конце G 1 , называемом START. Прохождение через START контролируется наличием питательных веществ, факторами спаривания и размером клеток. (подробнее…)
Помимо использования в качестве точки принятия решения для мониторинга внеклеточных сигналов, START является точкой, в которой клеточный рост координируется с репликацией ДНК и клеточным делением. Важность этой регуляции особенно очевидна у почкующихся дрожжей, у которых при делении клеток образуются клетки-потомки самых разных размеров: большая материнская клетка и маленькая дочерняя клетка. Чтобы дрожжевые клетки сохраняли постоянный размер, маленькая дочерняя клетка должна расти больше, чем крупная материнская клетка, прежде чем снова делиться. Таким образом, необходимо контролировать размер клеток, чтобы координировать рост клеток с другими событиями клеточного цикла.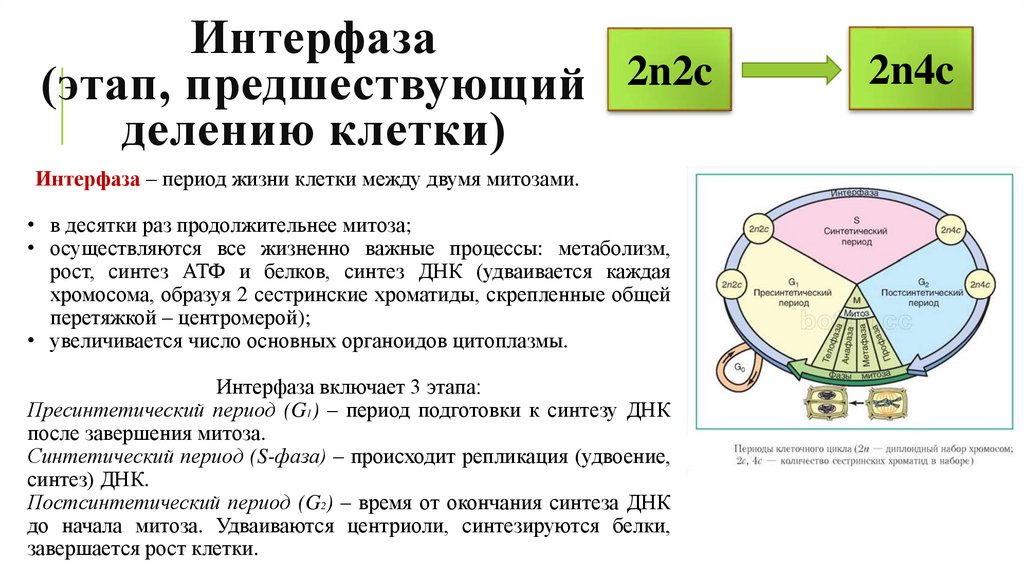 Это регулирование осуществляется с помощью механизма управления, который требует, чтобы каждая ячейка достигла минимального размера, прежде чем она сможет пройти START. Следовательно, маленькая дочерняя клетка дольше находится в G9.0174 1 и растет больше, чем материнская клетка.
Это регулирование осуществляется с помощью механизма управления, который требует, чтобы каждая ячейка достигла минимального размера, прежде чем она сможет пройти START. Следовательно, маленькая дочерняя клетка дольше находится в G9.0174 1 и растет больше, чем материнская клетка.
Пролиферация большинства клеток животных аналогичным образом регулируется в фазе G 1 клеточного цикла. В частности, точка принятия решения в конце G 1 , называемая точкой рестрикции в клетках животных, функционирует аналогично START у дрожжей (4). Однако, в отличие от дрожжей, прохождение животных клеток через клеточный цикл регулируется главным образом внеклеточными факторами роста, которые сигнализируют о пролиферации клеток, а не наличием питательных веществ. В присутствии соответствующих факторов роста клетки проходят точку рестрикции и вступают в S-фазу. После прохождения точки рестрикции клетка обязуется пройти через S-фазу и остальную часть клеточного цикла, даже в отсутствие дальнейшей стимуляции фактором роста. С другой стороны, если соответствующие факторы роста недоступны в G 1 , продвижение по клеточному циклу останавливается в точке рестрикции. Такие арестованные клетки затем вступают в стадию покоя клеточного цикла, называемую G 0 , в которой они могут оставаться в течение длительного периода времени без пролиферации. Клетки G 0 метаболически активны, хотя прекращают рост и снижают скорость синтеза белка. Как уже отмечалось, многие клетки у животных остаются в G 0 , если только соответствующие факторы роста или другие внеклеточные сигналы не призывают к пролиферации. Например, фибробласты кожи задерживаются в G9.0174 0 до тех пор, пока они не будут стимулированы к делению, необходимому для восстановления повреждений, полученных в результате ранения. Пролиферация этих клеток запускается тромбоцитарным фактором роста, который высвобождается из тромбоцитов во время свертывания крови и сигнализирует о пролиферации фибробластов вблизи поврежденной ткани.
С другой стороны, если соответствующие факторы роста недоступны в G 1 , продвижение по клеточному циклу останавливается в точке рестрикции. Такие арестованные клетки затем вступают в стадию покоя клеточного цикла, называемую G 0 , в которой они могут оставаться в течение длительного периода времени без пролиферации. Клетки G 0 метаболически активны, хотя прекращают рост и снижают скорость синтеза белка. Как уже отмечалось, многие клетки у животных остаются в G 0 , если только соответствующие факторы роста или другие внеклеточные сигналы не призывают к пролиферации. Например, фибробласты кожи задерживаются в G9.0174 0 до тех пор, пока они не будут стимулированы к делению, необходимому для восстановления повреждений, полученных в результате ранения. Пролиферация этих клеток запускается тромбоцитарным фактором роста, который высвобождается из тромбоцитов во время свертывания крови и сигнализирует о пролиферации фибробластов вблизи поврежденной ткани.
Рисунок 14.6
Регуляция клеточных циклов животных факторами роста. Доступность факторов роста контролирует клеточный цикл животных в конце G 1 позвонил в пункт ограничения. Если факторы роста недоступны во время G 1 , клетки переходят в стадию покоя (подробнее…)
Хотя пролиферация большинства клеток регулируется главным образом в G 1 , некоторые клеточные циклы вместо этого контролируются главным образом в G 2 . Одним из примеров является клеточный цикл делящихся дрожжей Schizosaccharomyces pombe (). В отличие от Saccharomyces cerevisiae , клеточный цикл S . pombe регулируется главным образом путем контроля перехода от G 2 к М, что является основной точкой, в которой отслеживается размер клеток и доступность питательных веществ. У животных основным примером контроля клеточного цикла в G 2 являются ооциты. Ооциты позвоночных могут оставаться задержанными в G 2 в течение длительных периодов времени (несколько десятилетий у человека), пока их переход в М-фазу не будет вызван гормональной стимуляцией.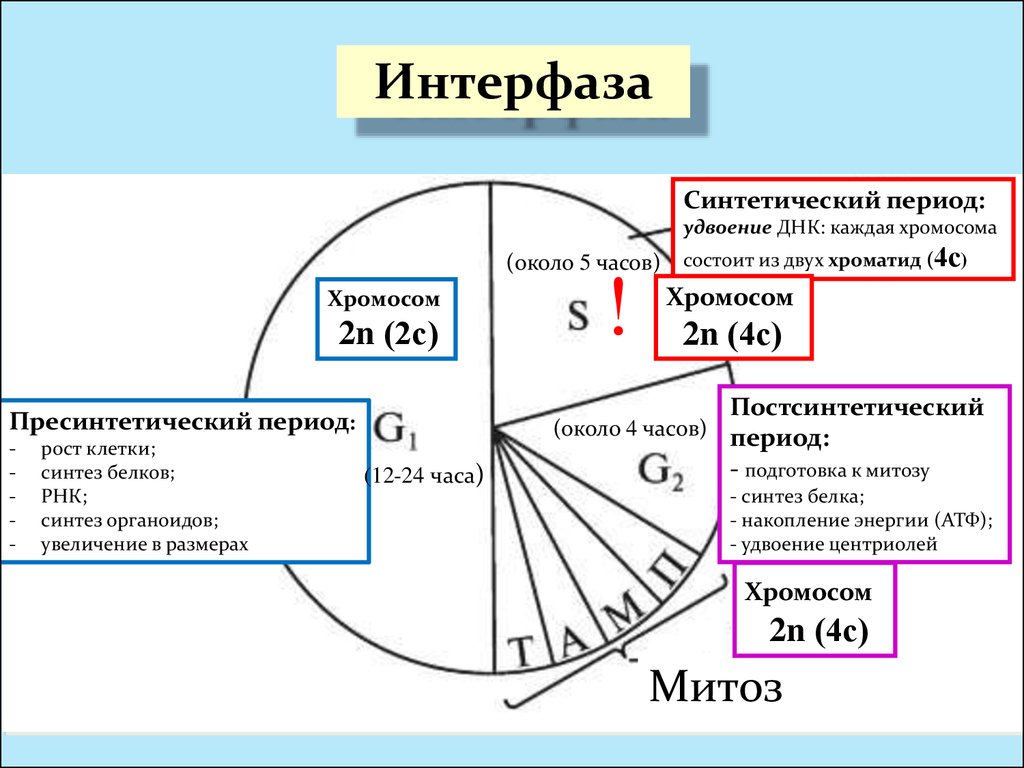 Таким образом, внеклеточные сигналы могут контролировать пролиферацию клеток, регулируя прогрессирование от G 2 до M, а также G 1 до S фазы клеточного цикла.
Таким образом, внеклеточные сигналы могут контролировать пролиферацию клеток, регулируя прогрессирование от G 2 до M, а также G 1 до S фазы клеточного цикла.
Рисунок 14.7
Клеточный цикл делящихся дрожжей. (A) Делящиеся дрожжи растут, удлиняясь с обоих концов, и делятся, образуя стенку через середину клетки. В отличие от цикла почкующихся дрожжей клеточный цикл делящихся дрожжей имеет нормальные G 1 , S, G 2 и М фазы. (подробнее…)
Контрольные точки клеточного цикла
Элементы управления, описанные в предыдущем разделе, регулируют ход клеточного цикла в зависимости от размера клеток и внеклеточных сигналов, таких как питательные вещества и факторы роста. Кроме того, события, происходящие на разных стадиях клеточного цикла, должны быть согласованы друг с другом, чтобы они происходили в соответствующем порядке. Например, критически важно, чтобы клетка не начинала митоз до завершения репликации генома. Альтернативой могло бы стать катастрофическое деление клетки, при котором дочерние клетки не смогли бы унаследовать полные копии генетического материала. В большинстве клеток эта координация между различными фазами клеточного цикла зависит от системы контрольных точек и контроля с обратной связью, которые предотвращают переход к следующей фазе клеточного цикла до тех пор, пока не будут завершены события предыдущей фазы.
Альтернативой могло бы стать катастрофическое деление клетки, при котором дочерние клетки не смогли бы унаследовать полные копии генетического материала. В большинстве клеток эта координация между различными фазами клеточного цикла зависит от системы контрольных точек и контроля с обратной связью, которые предотвращают переход к следующей фазе клеточного цикла до тех пор, пока не будут завершены события предыдущей фазы.
Несколько контрольных точек клеточного цикла функционируют, чтобы гарантировать, что неполные или поврежденные хромосомы не реплицируются и не передаются дочерним клеткам (). Одна из наиболее четко определенных контрольных точек встречается в G 2 и предотвращает инициацию митоза до завершения репликации ДНК. Эта контрольная точка G 2 обнаруживает нереплицированную ДНК, которая генерирует сигнал, приводящий к остановке клеточного цикла. Таким образом, работа контрольной точки G 2 предотвращает начало фазы M до завершения фазы S, поэтому ячейки остаются в G 9. 0174 2 до полной репликации генома. Только тогда ингибирование прогрессии G 2 снимается, позволяя клетке инициировать митоз и распределять полностью реплицированные хромосомы по дочерним клеткам.
0174 2 до полной репликации генома. Только тогда ингибирование прогрессии G 2 снимается, позволяя клетке инициировать митоз и распределять полностью реплицированные хромосомы по дочерним клеткам.
Рисунок 14.8
Контрольные точки клеточного цикла. Несколько контрольных точек обеспечивают передачу полных геномов дочерним клеткам. Одна из основных контрольных точек арестовывает клетки в G 2 в ответ на поврежденную или нереплицированную ДНК. Наличие поврежденной ДНК также приводит к клеточному (далее…)
Прохождение клеточного цикла также останавливается в контрольной точке G 2 в ответ на повреждение ДНК, например, в результате облучения. Эта остановка дает время для устранения повреждений, а не для передачи их дочерним клеткам. Исследования мутантов дрожжей показали, что одна и та же контрольная точка клеточного цикла отвечает за остановку G 2 , вызванную либо нереплицированной, либо поврежденной ДНК, обе из которых сигнализируют об остановке клеточного цикла через родственные пути.
 главу 15), иллюстрируя критическую важность регуляции клеточного цикла в жизни многоклеточных организмов.
главу 15), иллюстрируя критическую важность регуляции клеточного цикла в жизни многоклеточных организмов.Рисунок 14.9
Роль p53 в G 1 остановке, вызванной повреждением ДНК. Повреждение ДНК, например, в результате облучения, приводит к быстрому повышению уровня р53. Затем белок p53 сигнализирует об остановке клеточного цикла в контрольной точке G 1 .
Другая важная контрольная точка клеточного цикла, которая поддерживает целостность генома, происходит ближе к концу митоза (см. ). Эта контрольная точка отслеживает выравнивание хромосом в митотическом веретене, гарантируя тем самым точное распределение полного набора хромосом по дочерним клеткам. Например, неспособность одной или нескольких хромосом правильно выровняться на веретене приводит к остановке митоза в метафазе до сегрегации вновь реплицированных хромосом в дочерние ядра. В результате этой контрольной точки хромосомы не разделяются до тех пор, пока не будет организован полный набор хромосом для распределения по каждой дочерней клетке.
Связывание S-фазы с M-фазой
Контрольная точка G 2 предотвращает инициацию митоза до завершения S-фазы, тем самым гарантируя, что не полностью реплицированная ДНК не будет распределена по дочерним клеткам. Не менее важно обеспечить, чтобы геном реплицировался только один раз за клеточный цикл. Таким образом, после репликации ДНК должны существовать механизмы контроля, чтобы предотвратить инициацию новой S-фазы до митоза. Эти элементы управления предотвращают появление ячеек в G 2 от повторного входа в S-фазу и заблокировать инициацию другого раунда репликации ДНК до завершения митоза, после чего клетка войдет в фазу G 1 следующего клеточного цикла.
Первоначальное понимание этой зависимости S-фазы от M-фазы пришло из экспериментов по слиянию клеток Поту Рао и Роберта Джонсона в 1970 году (). Эти исследователи изолировали клетки в разных фазах цикла, а затем сливали эти клетки друг с другом, чтобы сформировать клеточные гибриды.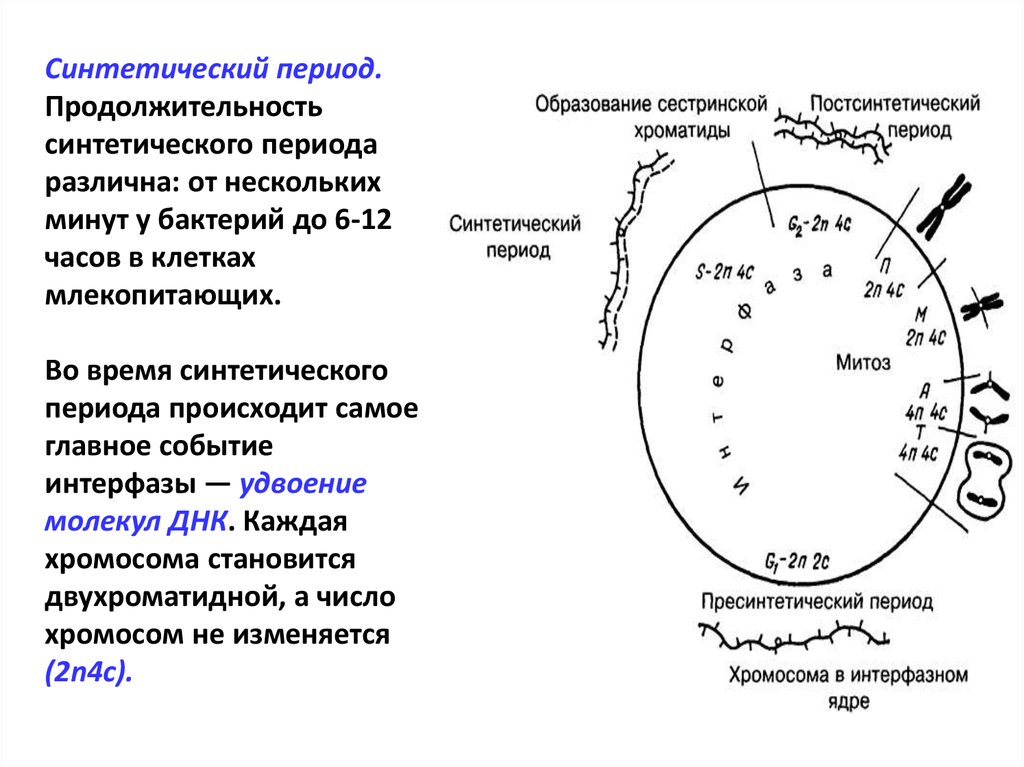 Когда клетки G 1 были слиты с клетками S фазы, G 1 ядро немедленно начало синтезировать ДНК. Таким образом, цитоплазма клеток S фазы содержала факторы, инициирующие синтез ДНК в ядре G 1 . Однако слияние клеток G 2 с клетками S фазы дало совершенно другой результат: ядро G 2 не могло инициировать синтез ДНК даже в присутствии цитоплазмы S фазы. Таким образом, оказалось, что синтез ДНК в ядре G 2 предотвращается механизмом, который блокирует повторную репликацию генома до тех пор, пока не произойдет митоз.
Когда клетки G 1 были слиты с клетками S фазы, G 1 ядро немедленно начало синтезировать ДНК. Таким образом, цитоплазма клеток S фазы содержала факторы, инициирующие синтез ДНК в ядре G 1 . Однако слияние клеток G 2 с клетками S фазы дало совершенно другой результат: ядро G 2 не могло инициировать синтез ДНК даже в присутствии цитоплазмы S фазы. Таким образом, оказалось, что синтез ДНК в ядре G 2 предотвращается механизмом, который блокирует повторную репликацию генома до тех пор, пока не произойдет митоз.
Рис. 14.10
Эксперименты по слиянию клеток, демонстрирующие зависимость S-фазы от M-фазы. Клетки в S-фазе сливали либо с клетками G 1 , либо с клетками G 2 . Когда клетки G 1 сливались с клетками S фазы, ядро G 1 немедленно начинало реплицировать ДНК. Напротив, (подробнее…)
Молекулярный механизм, который ограничивает репликацию ДНК до одного раза за клеточный цикл, включает действие семейства белков (называемых белками MCM), которые связываются с точками начала репликации вместе с комплексом репликации начала (ORC).
Leave A Comment