
Бактерии автотрофы или гетеротрофы? 11 фактов, которые вы должны знать
В этой статье мы подробно обсудим «бактерии автотрофы или гетеротрофы» с 11 фактами.
Бактерии делятся на две категории: автотрофы означают «самопитающихся», которые готовят себе пищу самостоятельно, или гетеротрофы означают «других питающихся», все они полагаются на других, не способных самостоятельно готовить пищу..
Теперь взгляните на 11 фактов о том, являются ли бактерии автотрофами или гетеротрофами в деталях.
Каким образом бактерии являются автотрофами?Слово «автотроф» означает один из видов первичных продуцентов в основе пищевой цепи, другими словами, это организмы, которые могут производить себе пищу, принимая в свое тело какое-то неорганическое вещество и используя некоторую химическую энергию или собирая некоторую энергию, которая накапливается в организме. природа превращает его в органическое вещество.
Проще говоря, большинство автотрофных бактерий используют процесс фотосинтеза для производства пищи. Автотрофы вообще очень важны, потому что они являются основными первичными производителями свободноживущих и встречаются в океанах, водоемах или почве, они могут выполнять несколько реакций и могут поддерживать свою жизнь в суровых или экстремальных условиях или окружающей среде.
Автотрофы вообще очень важны, потому что они являются основными первичными производителями свободноживущих и встречаются в океанах, водоемах или почве, они могут выполнять несколько реакций и могут поддерживать свою жизнь в суровых или экстремальных условиях или окружающей среде.
Все бактерии не автотрофы, некоторые из них гетеротрофы.Некоторые виды бактерий используют энергию в виде света для приготовления пищи так же, как и в процессе фотосинтеза. Эти бактерии не относятся к категории автотрофов, потому что все они полагаются на других, не способных производить пищу самостоятельно..
Бактерии могут быть паразитами или сапрофитами. Также это могут быть хемосинтетические автотрофы или фотосинтетики. К большинству живых организмов относятся грибы, протисты или бактерии, простейшие гетеротрофны, все они зависят от автотрофов, так как у них нет хлорофилла, поэтому они также известны как консументы.
Бактерии могут быть и тем и другим, но большинство распространенных бактерий являются гетеротрофами. Гетеротрофные бактерии повсеместно присутствуют во всех типах растительности, пищевых продуктах, почве, воде и воздухе. Такие бактерии получают органический материал из окружающей среды для роста.
Гетеротроф означает «другой фидер, либопотребителейЭто означает, что он не может производить свою пищу, как автотроф, он может получать пищу из другого источника или из окружающей среды. в то время как автотроф означает «автокормушка” и выступает в качестве основного производитель. Автотрофы находятся на начальном уровне пищевой цепи. Он является членом некоторых одноклеточных организмов, таких как цианобактерии а хлорофилл или хлоропласт в основном отвечает за приготовление пищи
Какой тип бактерий является автотрофным?Два разных типа бактерий являются автотрофными.
Уникальной характеристикой этих бактерий является то, что они процветают при достаточно высоких температурах, чтобы убить другие организмы. Они живут в глубоководных жерлах океана в очень экстремальных условиях, также называемых «экстремофилами». Этот тип автотрофных бактерий получает энергию от окисления неорганических соединений, таких как железо и аммиак, сероводород.
Он также может поглощать воду и углекислый газ из окружающей среды и превращать их в соединение серы для создания сахаров и углеводов посредством метаболического процесса.
Примеры включают, Erwinia, Agrobacterium, Pseudomonas syringae, Stigmatella, Streptomyces, азотфиксирующие бактерии, такие как Nitrosomonas, Nitrobacter, Rhizobium, Frankiaсероокисляющие хемоавтотрофы, такие как Thermothrix, Thiobacillus,Цианобактерии как носток, ананбаэна, осциллятория, спирулина,Археи: Метанококки, метаноспириллы, пиролобусы, термоплазмы,Водородные бактерии как Helicobacter pylori, термофилы Hydrogenobacter
Фотоавтотроф:Этот тип автотрофных бактерий получает энергию непосредственно от солнца и создает себе пищу, используя неорганические соединения серы для осуществления различных клеточных метаболических процессов, таких как дыхание и биосинтез.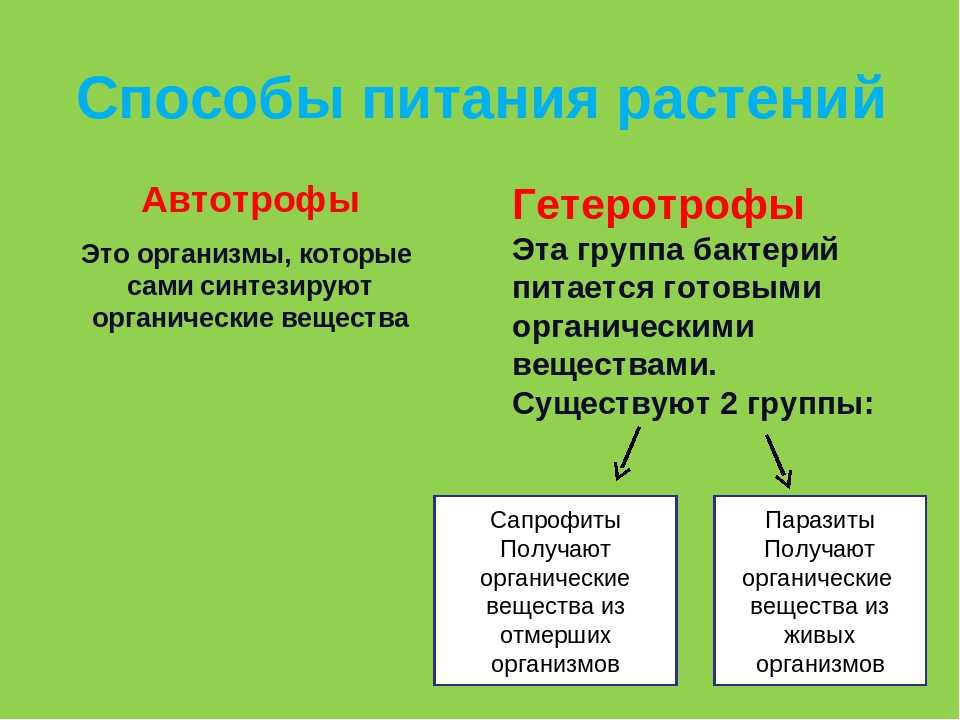 такой тип бактерий также называют голофитными.
такой тип бактерий также называют голофитными.
Они являются наиболее распространенными первичными производителями в большинстве морских и наземных экосистем. Зеленые растения или некоторые цианобактерии в присутствии солнечного света превращают углекислый газ в углеводы. фотоавтотроф, включают пурпурные серные бактерии, пурпурные несерные бактерии, зеленые серные бактерии, оксигенные и аноксигенные фотосинтетические бактерии, некоторые высшие растения, такие как травы или растения кукурузы, некоторые цианобактерии, эвглену и зеленые водоросли.
Все ли бактерии гетеротрофны?Не все бактерии являются гетеротрофами. Если сравнивать их с автотрофами, то большинство бактерий являются гетеротрофами в окружающей среде. В окружающей среде большое количество гетеротрофных бактерий..
Гетеротрофы не могут производить пищу, как автотрофы, они могут получать пищу из другого источника, потому что все они действуют как потребители.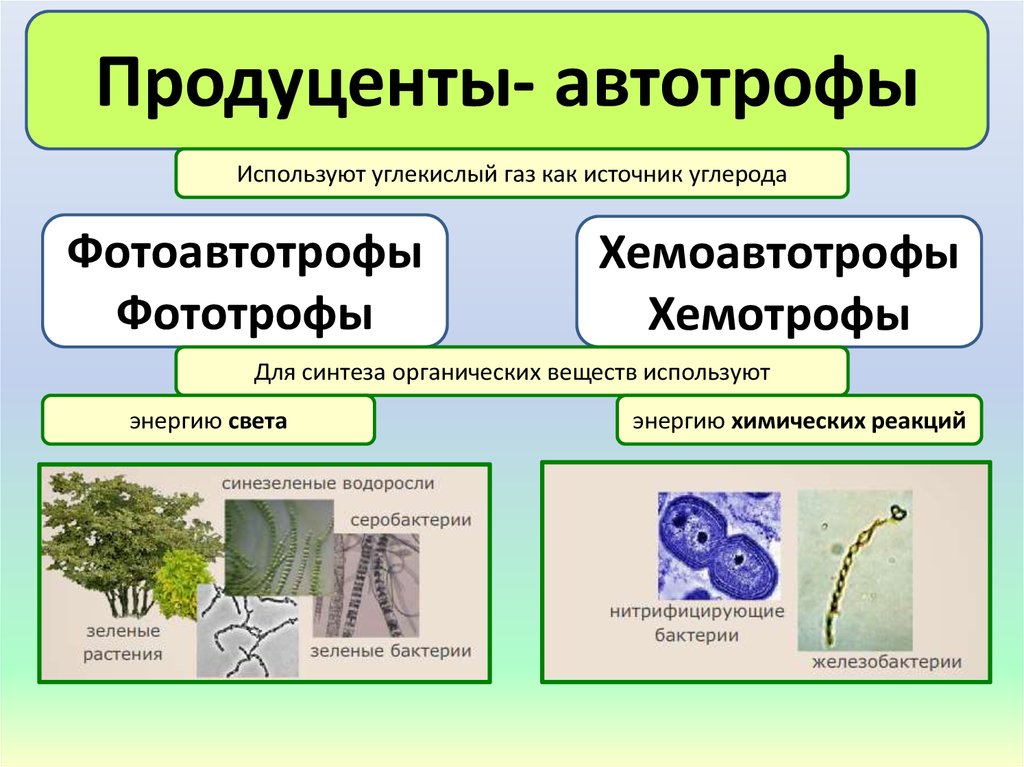 Эти бактерии в основном ответственны за процесс разложения органического вещества. Бактерии также помогают в фиксации азота и переработке питательных веществ. Некоторые наиболее распространенные гетеротрофные бактерии включают: Lactobacillus acidophilus, Desulfovibrio desulfuricans, Escherichia coli, Agarobacterium, Xanthomonas, Rhizobum, Salmonella и pseudomonas
Эти бактерии в основном ответственны за процесс разложения органического вещества. Бактерии также помогают в фиксации азота и переработке питательных веществ. Некоторые наиболее распространенные гетеротрофные бактерии включают: Lactobacillus acidophilus, Desulfovibrio desulfuricans, Escherichia coli, Agarobacterium, Xanthomonas, Rhizobum, Salmonella и pseudomonas
Большинство гетеротрофных бактерий существуют как вездесущие симбионты или паразиты. В зависимости от среды обитания, источника пищи и ассоциации с другими организмами эти бактерии классифицируются по различным типам категорий, включая симбиотические, паразитарные и сапрофитные..
Есть два типа гетеротрофов, Хемогетеротрофы-При использовании углекислого газа и окислении органических соединений они получают энергию.Фотогетеротрофы. Эти гетеротрофы не могут использовать углекислый газ, но могут получать углерод из различных типов соединений, включая жирные кислоты, и углеводы. В зависимости от источника энергии гетеротрофы можно разделить на литотропы или органотрофы, а другой способ их классификации — хемолитогетеротрофы и фотоорганогетеротрофы. Вот некоторые примеры гетеротрофных бактерий, включая пурпурные несерные бактерии, Родоспирилловые, Oceanithermus profundus Pseudomonas syringae, Neisseria gonorrhoeae, Clostridium tetani, Bordetella pertussis, Corynebacterium diphtheria, Xanthomonas campestris, Xanthomonas axonopodis, Pseudomonas solanacearum.
В зависимости от источника энергии гетеротрофы можно разделить на литотропы или органотрофы, а другой способ их классификации — хемолитогетеротрофы и фотоорганогетеротрофы. Вот некоторые примеры гетеротрофных бактерий, включая пурпурные несерные бактерии, Родоспирилловые, Oceanithermus profundus Pseudomonas syringae, Neisseria gonorrhoeae, Clostridium tetani, Bordetella pertussis, Corynebacterium diphtheria, Xanthomonas campestris, Xanthomonas axonopodis, Pseudomonas solanacearum.
Как бактерии становятся гетеротрофами? Основные характеристики этого бактерии имеют клеточное и простое строение с клеточными стенками, Цитоплазма, Пилли, Жгутик, Капсула, рибосомы и хромосомы, в которых отсутствует ядро, связанное с мембраной. Это тип бактерий, которым требуется сахар для роста, выживания и размножения.. Эти бактерии наиболее многочисленны и широко распространены и противостоят автотрофам. Большинство зеленых или зеленых серобактерий шаровидные или палочковидные, мезофильные, неподвижные и преимущественно фотолитоавтотрофные. Все зеленые бактерии в основном относятся к категории автотрофных бактерий, потому что они используют некоторое количество восстановленного соединения серы и световую энергию в качестве источника электронов. Зеленые автотрофные бактерии также содержат некоторые Бактериохлорофиллы и в основном требуют анаэробных условий, предпочтительна умеренная температура, а в условиях низкой освещенности они в основном способны к фотосинтезу. Все патогенных бактерий являются гетеротрофными, потому что они получают энергию и питание за счет поглощения биомассы и окисления органических молекул, таких как липиды, белки и углеводы, из окружающей среды. Они могут напрямую атаковать, чтобы получить питательные вещества для своего роста, что наносит ущерб хозяину. а разложение органического вещества — основная функция гетеротрофных бактерий. Таким образом, патогенные гетеротрофные бактерии также известны как редуценты. Они могут быть либо анаэробный или аэробный. Архебактерии могут быть как автотрофами, так и гетеротрофами, но в большинстве своем археи классифицируются как автотрофы, облигатные анаэробы, и они получают пищу из простых органических молекул, таких как сахар. Они также используют некоторые различные виды основных веществ, таких как металлы, углерод, водород и кислород, или они могут использовать процессы хемосинтеза и фотосинтеза для производства пищи, питания и роста. Они используют свет посредством фотосинтеза для производства пищи. Они очень метаболически разнообразны. Автотрофный архебактерии были также известны как экстремофилы поскольку они живут в очень суровых условиях, они представляют собой несколько термофильных бактерий-архей, которые могут жить на очень горячем морском дне и в горячих источниках. эти бактерии также делятся на две группы, включающие органотроф который использует органические соединения, в то время как другие литотрофы которые используют неорганическое соединение для получения энергии. Некоторые примеры включают Archaebacteria, Halobacterium, Pyrolobus, Acidilobus saccharovorans, Staphylothermus Hellenic, Sulfolobus solfataricus, Thermoproteus tenax. Хемосинтетические бактерии представляют собой группу автотрофных бактерий, потому что они производят свою пищу, используя химическую энергию для производства. Хемосинтезирующие бактерии также известны как группа фотосинтезирующих бактерий, потому что все они могут окислять различные неорганические вещества. Эти бактерии могут выживать в суровых или экстремальных условиях окружающей среды или различных средах обитания. Большинство этих альтерий зависят от кислорода, что означает, что они аэробы. но некоторые виды также связаны с анаэробным хемосинтезом. Например, Sulfuricurvum kujienese, Некоторые другие примеры автотрофных хемосинтетических бактерий включают Ferrooxidans, Venenivibrio stagnispumantis, серобактерии, включая паракокки, азотфиксирующие бактерии, водородные бактерии.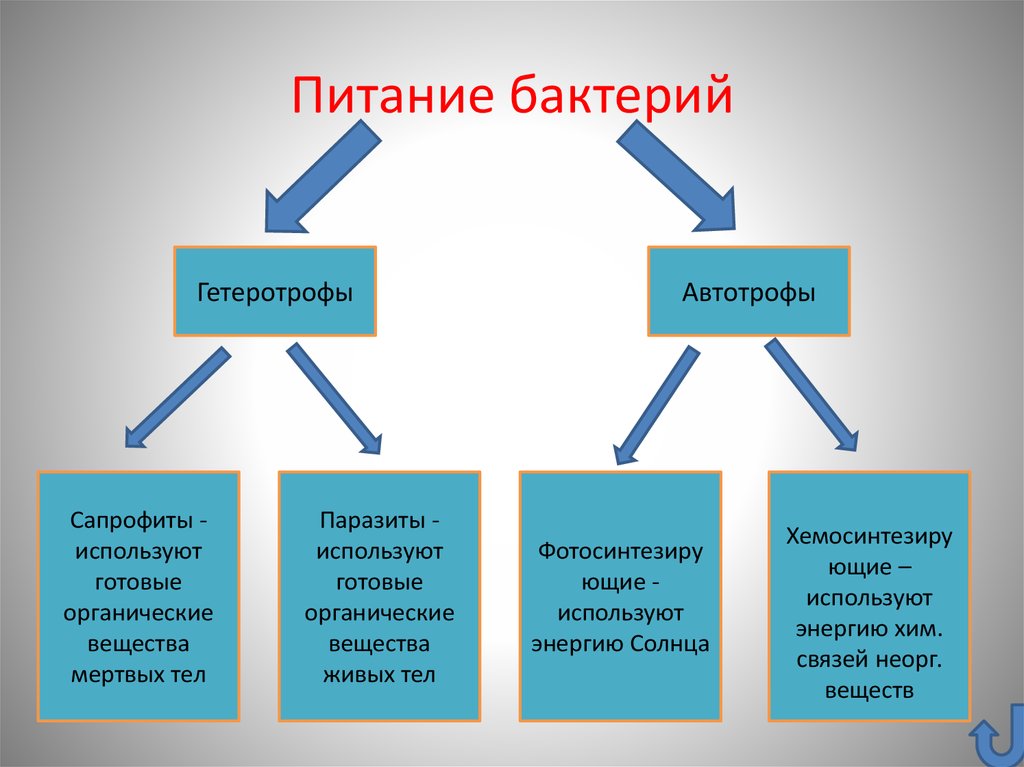 Форма бактерий зависит от размера и вида бактерий. Гетеротрофные бактерии, наиболее часто встречающиеся в каждой области жизни, включают Бактерии, археи и эукарии. Примеры гетеротрофов включают стрептококки, стрептомицеты, лактобациллы,Альканиворакс, Метилоцелла сильвестрис и многие другие имеют различные экономические применения
Форма бактерий зависит от размера и вида бактерий. Гетеротрофные бактерии, наиболее часто встречающиеся в каждой области жизни, включают Бактерии, археи и эукарии. Примеры гетеротрофов включают стрептококки, стрептомицеты, лактобациллы,Альканиворакс, Метилоцелла сильвестрис и многие другие имеют различные экономические применения эти бактерии могут метаболизировать серу. Они могут жить в анаэробных условиях или в водной среде. Некоторые другие пример автотрофных зеленых бактерий is Хлоробий тепидум некоторые другие включают хеликобактерии, фототрофные кислые бактерии, пурпурные серные бактерии, пурпурные несерные бактерии и нитевидные аноксигенные фототрофы.
эти бактерии могут метаболизировать серу. Они могут жить в анаэробных условиях или в водной среде. Некоторые другие пример автотрофных зеленых бактерий is Хлоробий тепидум некоторые другие включают хеликобактерии, фототрофные кислые бактерии, пурпурные серные бактерии, пурпурные несерные бактерии и нитевидные аноксигенные фототрофы. Две распространенные гетеротрофные бактерии, а именно Аэромонас и Pseudomonas действуют как патогенные. Эти бактерии действуют как потребители, чтобы зависеть от других микроорганизмов для своего питания, роста и выживания они также способны выделять или выделять некоторые токсичные вещества, которые могут влиять на других людей в природе. Некоторые примеры: Escherichia coli, Pseudomonas, Agrobacterium, Salmonella и Xanthomonas, Acinetobacter, Corynebacteria, Klebsillea, Listeria, Micrococcus, Staphylococcus
Две распространенные гетеротрофные бактерии, а именно Аэромонас и Pseudomonas действуют как патогенные. Эти бактерии действуют как потребители, чтобы зависеть от других микроорганизмов для своего питания, роста и выживания они также способны выделять или выделять некоторые токсичные вещества, которые могут влиять на других людей в природе. Некоторые примеры: Escherichia coli, Pseudomonas, Agrobacterium, Salmonella и Xanthomonas, Acinetobacter, Corynebacteria, Klebsillea, Listeria, Micrococcus, Staphylococcus
 . Им нужен некоторый источник энергии, а также источник углерода для производства продуктов питания. Они играют очень важную экономическую роль в переработке различных видов питательных веществ, включая железо, фосфор и азот..
. Им нужен некоторый источник энергии, а также источник углерода для производства продуктов питания. Они играют очень важную экономическую роль в переработке различных видов питательных веществ, включая железо, фосфор и азот..
Что значит гетеротрофы — Значения слов
 org/BreadcrumbList»>
org/BreadcrumbList»>Раздел очень прост в использовании. В предложенное поле достаточно ввести нужное слово, и мы вам выдадим список его значений. Хочется отметить, что наш сайт предоставляет данные из разных источников – энциклопедического, толкового, словообразовательного словарей. Также здесь можно познакомиться с примерами употребления введенного вами слова.
Энциклопедический словарь, 1998 г.

гетеротрофы
ГЕТЕРОТРОФЫ (от гетеро… и греч. trophe — пища) (гетеротрофные организмы) используют для своего питания готовые органические вещества. К гетеротрофам относятся человек, все животные, некоторые растения, большинство бактерий, грибы. Ср. Автотрофы.
Википедия
Гетеротрофы
Гетеротро́фы — организмы, которые не способны синтезировать органические вещества из неорганических путём фотосинтеза или хемосинтеза . Для синтеза необходимых для своей жизнедеятельности органических веществ им требуются экзогенные органические вещества, то есть произведённые другими организмами. В процессе пищеварения пищеварительные ферменты расщепляют полимеры органических веществ на мономеры .
Растения-гетеротрофы полностью ( заразиха , раффлезия ) или почти полностью ( повилика ) лишены хлорофилла и питаются, прорастая в тело растения-хозяина.
Граница между автотрофами и гетеротрофами достаточно условна, так как существует множество видов, обладающих переходной формой питания — миксотрофией , либо использующие наиболее удобный в данных условиях тип питания.
Гетеротрофы подразделяются по способу поглощения пищи на фаготрофов и осмотрофов. По виду получаемой пищи подразделяются на биотрофов и сапротрофов.
Гетеротрофы
Гетеротрофы – это организмы, которые поедают другие растения или животных для получения энергии и питательных веществ.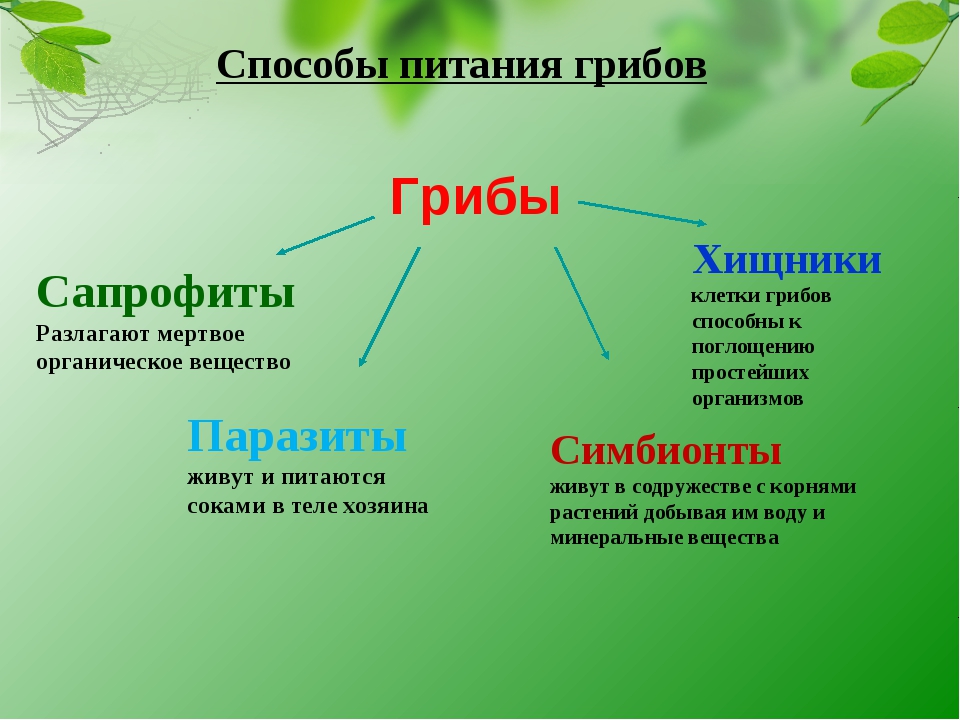 Термин происходит от греческих слов гетеро для «другого» и трофе для «питание».
Термин происходит от греческих слов гетеро для «другого» и трофе для «питание».
Организмы можно разделить на две большие категории в зависимости от того, как они получают энергию и питательные вещества: автотрофы и гетеротрофы. Автотрофы известны как производители, потому что они способны производить себе пищу из сырья и энергии. Примеры включают растения, водоросли и некоторые виды бактерий. Гетеротрофы известны как консументы, потому что они потребляют продуцентов или других консументов. Собаки, птицы, рыбы и люди — все это примеры гетеротрофов.
Гетеротрофы занимают второй и третий уровни в пищевой цепи, последовательности организмов, которые обеспечивают энергию и питательные вещества для других организмов. Каждая пищевая цепь состоит из трех трофических уровней, описывающих роль организма в экосистеме. Первый трофический уровень занимают автотрофы, такие как растения и водоросли. Травоядные – организмы, питающиеся растениями, – занимают второй уровень. Плотоядные (организмы, питающиеся мясом) и всеядные (организмы, питающиеся растениями и мясом) занимают третий уровень. И первичные (травоядные), и вторичные (хищники и всеядные) консументы являются гетеротрофами, а первичные продуценты — автотрофами.
Плотоядные (организмы, питающиеся мясом) и всеядные (организмы, питающиеся растениями и мясом) занимают третий уровень. И первичные (травоядные), и вторичные (хищники и всеядные) консументы являются гетеротрофами, а первичные продуценты — автотрофами.
Третий тип гетеротрофных потребителей — детритофаги. Эти организмы получают пищу, питаясь остатками растений и животных, а также фекалиями. Детритофаги играют важную роль в поддержании здоровой экосистемы путем переработки отходов. Примеры детритофагов включают грибы, черви и насекомые.
Существует две подкатегории гетеротрофов: фотогетеротрофы и хемогетеротрофы. Фотогетеротрофы — это организмы, которые получают энергию от света, но все же должны потреблять углерод от других организмов, поскольку они не могут утилизировать углекислый газ из воздуха. Хемогетеротрофы, напротив, получают как энергию, так и углерод от других организмов.
Основное различие между автотрофами и гетеротрофами заключается в том, что первые способны производить себе пищу путем фотосинтеза, тогда как вторые не могут. Фотосинтез — это процесс, который включает в себя образование глюкозы (сахара) и кислорода из воды и углекислого газа с использованием энергии солнечного света. Автотрофы могут производить энергию от солнца, но гетеротрофы должны полагаться на другие организмы для получения энергии.
Фотосинтез — это процесс, который включает в себя образование глюкозы (сахара) и кислорода из воды и углекислого газа с использованием энергии солнечного света. Автотрофы могут производить энергию от солнца, но гетеротрофы должны полагаться на другие организмы для получения энергии.
Еще одно важное различие между автотрофами и гетеротрофами заключается в том, что у автотрофов есть важный пигмент, называемый хлорофиллом, который позволяет им улавливать энергию солнечного света во время фотосинтеза, тогда как у гетеротрофов этого нет. Без этого пигмента фотосинтез невозможен.
Гетеротрофы получают пользу от фотосинтеза различными способами. Они зависят от процесса получения кислорода, который образуется как побочный продукт фотосинтеза. Более того, фотосинтез поддерживает автотрофов, от которых зависит выживание гетеротрофов. Хотя выживание плотоядных плотоядных может не зависеть напрямую от фотосинтезирующих растений, они зависят от других животных, которые потребляют фотосинтезирующие растения в качестве источника пищи.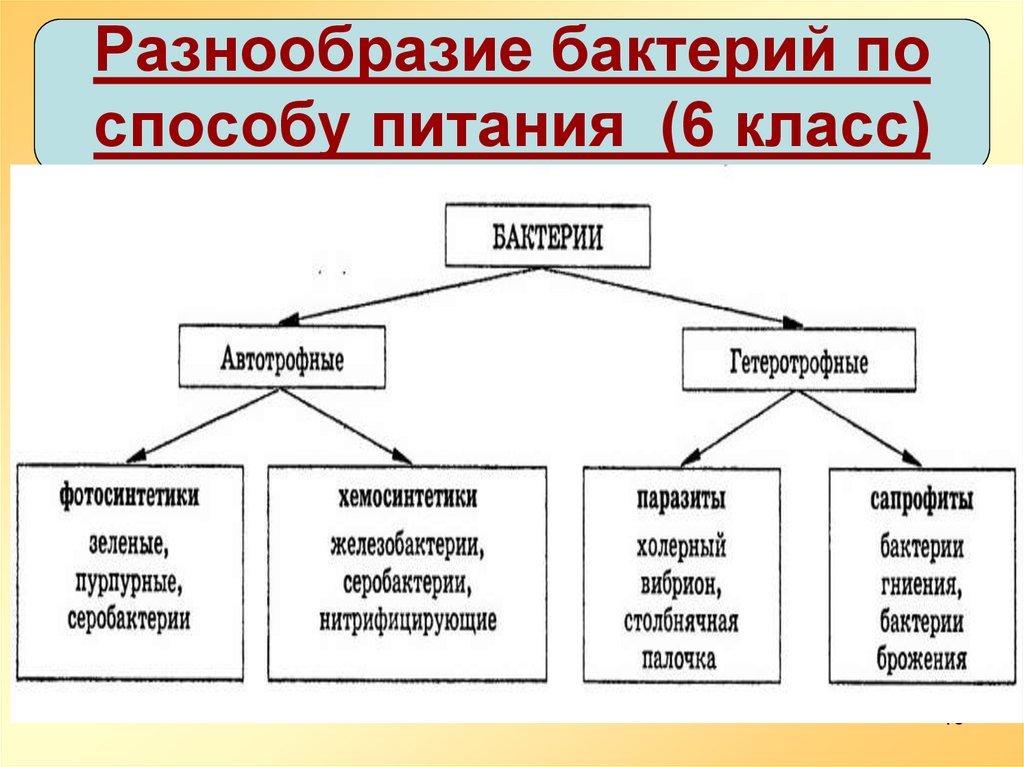
Эффективность роста гетеротрофных бактерий и структура сообщества при различных концентрациях природного органического углерода
1. Альтшул, С. Ф., В. Гиш, В. Миллер, Э. В. Майерс и Д. Дж. Липман. 1990. Базовый инструмент локального поиска выравнивания. Дж. Мол. биол. 215 : 403-410. [PubMed] [Google Scholar]
2. Амон, Р. М. У. и Р. Беннер. 1996. Бактериальная утилизация растворенных органических веществ различных классов размера. Лимнол. океаногр. 41 : 41-51. [Google Scholar].
4. Bano, N., M.A. Moran, and R.E. Hodson. 1997. Бактериальная утилизация растворенных гуминовых веществ из пресноводного болота. Аква. микроб. Экол. 12 : 233-238. [Google Scholar]
5. Бертилссон С. и Л. Дж. Транвик. 2000. Фотохимическая трансформация растворенного органического вещества в озерах. Лимнол. океаногр. 45 : 753-762. [Академия Google]
Лимнол. океаногр. 45 : 753-762. [Академия Google]
6. Bjørnsen, P. K. 1986. Рост бактериопланктона в непрерывных культурах морской воды. Мар. Экол. прог. сер. 30
7. Бьёрнсен П. К. и Купаринен Дж. 1991. Определение биомассы бактериопланктона, чистой продукции и эффективности роста в Южном океане. Мар. Экол. прог. сер. 71 : 185-194. [Google Scholar]
8. Carlson, C.A., and H.W. Ducklow. 1996. Рост бактериопланктона и потребление растворенного органического углерода в Саргосском море. Аква. микроб. Экол. 10 : 69-85. [Google Scholar]
9. Casamayor, E. O., H. Schäfer, L. Bañeras, C. Pedrós-Alio, and G. Muyzer. 2000. Идентификация и пространственно-временные различия между микробными сообществами из двух соседних сернистых озер: сравнение с помощью микроскопии и денатурирующего градиентного гель-электрофореза.
10. Casamayor, EO, R. Massana, S. Benlloch, L. Øvreås, B. Diez, VJ Goddard, JM Gasol, I. Joint, F. Rodríguez -Валера и К. Педрос-Алио. 2002. Изменения в архейных бактериальных и эукариальных сообществах вдоль градиента солености путем сравнения методов генетического фингерпринтинга в многопрудовой солнечной солончаке. Окружающая среда. микробиол. 4
11. Church, MJ, D.A. Hutchins, and H.W. Ducklow. 2000. Ограничение роста бактерий растворенными органическими веществами и железом в Южном океане. заявл. Окружающая среда. микробиол. 66 : 455-466. [Бесплатная статья PMC] [PubMed] [Google Scholar]
12. Коффин, Р. Б., Дж. П. Коннели и П. С. Харрис. 1993. Доступность растворенного органического углерода бактериопланктону исследовали по использованию кислорода.
13. Дель Джорджио, П. А., А. Ф. Берд, Ю. Т. Прейри и Д. Планас. 1996. Проточное цитометрическое определение численности бактерий в озерном планктоне с использованием красителя зеленой кислоты SYTO 13. Limnol. океаногр. 41 : 783-789. [Академия Google]
14. Дель Джорджио, П. А., и Дж. Дж. Коул. 1998. Эффективность роста бактерий в природных системах. Анну. Преподобный Экол. Сист. 29 : 503-541. [Google Scholar]
15. Fogel, G. B., C. R. Collins, J. Li, and C. F. Brunk. 1999. Размер прокариотического генома и количество копий рДНК SSU: оценка относительной численности микроорганизмов в смешанной популяции. микроб. Экол. 38
16. Джованни С. и М. Раппе. 2000. Эволюция, разнообразие и молекулярная экология морских прокариот, с. 47-84. В Д. Л. Кирхман (ред.), Микробная экология океанов. Wiley-Liss, New York, NY
47-84. В Д. Л. Кирхман (ред.), Микробная экология океанов. Wiley-Liss, New York, NY
17. Glöckner, F.O., B.M. Fuchs, and R. Amann. 1999. Состав бактериопланктона озер и океанов: первое сравнение, основанное на флуоресцентной гибридизации in situ. заявл. Окружающая среда. микробиол. 65
18. Глекнер Ф. О., Зайчиков Е., Белькова Н., Денисова Л., Пернталер Дж., Пернталер А., Аманн Р. 2000. Сравнительный анализ 16S рРНК озерного бактериопланктона выявил глобально распределенные филогенетические кластеры, в том числе многочисленную группу актинобактерий. заявл. Окружающая среда. микробиол. 66 : 5053-5065. [Бесплатная статья PMC] [PubMed] [Google Scholar]
19. Goldman, JC, and M.R. Dennett. 2000. Рост морских бактерий в периодических и непрерывных культурах при ограничении содержания углерода и азота. Лимнол.
20. Harder, J. 1997. Независимая от вида энергия поддержания и естественная численность населения. ФЭМС микробиол. Экол. 23 : 39-44. [Google Scholar]
21. Хобби, Дж. Э., Р. Дж. Дейли и С. Джаспер. 1977. Использование фильтров Nuclepore для подсчета бактерий с помощью флуоресцентной микроскопии. заявл. Окружающая среда. микробиол. 33 : 1225-1228. [Бесплатная статья PMC] [PubMed] [Google Scholar]
22. Kirchman, D.L., Y. Suzuki, C. Garside, and H.W. Ducklow.
23. Kirchman, D. L. 2002. Экология Cytophaga-Flavobacteria в водной среде. ФЭМС микроб. Экол. 39 : 91-100. [PubMed] [Google Scholar]
24. Кристиансен К., Х. Нильсен, Б. Риманн и Дж. А. Фурман. 1992. Эффективность роста пресноводного бактериопланктона. микроб. Экол. 24 : 145-160. [PubMed] [Google Scholar]
Кристиансен К., Х. Нильсен, Б. Риманн и Дж. А. Фурман. 1992. Эффективность роста пресноводного бактериопланктона. микроб. Экол. 24 : 145-160. [PubMed] [Google Scholar]
25. Kroer, N. 1993. Эффективность роста бактерий на естественном растворенном органическом веществе. Лимнол. океаногр. 38 : 1282-1290. [Google Scholar]
26. Lehman, JT 1980. Высвобождение и круговорот питательных веществ между планктонными водорослями и травоядными. Лимнол. океаногр. 25 : 620-632. [Google Scholar]
27. Лайонел Р., П. Франк и Н. Сильви. 2000. Мониторинг сложных бактериальных сообществ с использованием культурально-независимых молекулярных методов: применение в почвенной среде. Рез. микробиол. 1551 : 167-177. [PubMed] [Google Scholar]
28. Лоферер-Кроссбахер, М., Дж. Клима и Р. Псеннер. 1998. Определение сухой массы бактериальных клеток с помощью просвечивающей электронной микроскопии и денситометрического анализа изображений. заявл. Окружающая среда. микробиол. 64 : 668-694. [Бесплатная статья PMC] [PubMed] [Google Scholar]
заявл. Окружающая среда. микробиол. 64 : 668-694. [Бесплатная статья PMC] [PubMed] [Google Scholar]
29. Marden, P., T. Nystrom, and S. Kjelleberg. 1987. Поглощение лейцина морской грамотрицательной гетеротрофной бактерией при голодании. ФЭМС микробиол. Экол. 45 : 233-241. [Google Scholar]
30. McMannus, G.B. 1993. Темпы роста естественных популяций гетеротрофного нанопланктона, с. 557-562. В П. Ф. Кемп, Э. Б. Шерр и Дж. Дж. Коул (ред.), Современные методы водной микробной экологии. Lewis Publishers, Boca Raton, Fla.
31. Méthe, B.A., W.D. Hiorns, and J.P. Zehr. 1998. Различия между составом морских и пресноводных бактериальных сообществ: анализ сообществ в озере Джордж и шести других озерах Адирондак. Лимнол. океаногр. 43 : 368-374. [Google Scholar]
32. Миддельбо М., Б. Нильсен и М. Сондергаард. 1992. Использование бактериями растворенного органического углерода (РОУ) в прибрежных водах – определение урожайности. Арка гидробиол. Эрг. Лимнол. 37 : 51-61. [Google Scholar]
Арка гидробиол. Эрг. Лимнол. 37 : 51-61. [Google Scholar]
33. Миддельбо М. и М. Сондергаард. 1993. Урожайность бактериопланктона: сезонные колебания и связь с лабильностью субстрата и активностью β-глюкозидазы. заявл. Окружающая среда. микробиол. 59 : 3916-3921. [Бесплатная статья PMC] [PubMed] [Google Scholar]
34. Mille-Lindblom, C., and L. J. Tranvik. 2003. Антагонизм между бактериями и грибами при разложении опада водных растений. микроб. Экол. 45 : 173-182. [PubMed] [Google Scholar]
35. Morita, RY 1997. Бактерии в олиготрофных средах. Chapman & Hall, New York, NY
36. Muyzer, G., E.C. de Waal, and A.G. Uitterlinden. 1993. Профилирование сложных микробных популяций с помощью денатурирующего градиентного гель-электрофореза анализа амплифицированных полимеразной цепной реакцией генов, кодирующих 16S рРНК. заявл. Окружающая среда. микробиол. 59 : 695-700. [Бесплатная статья PMC] [PubMed] [Google Scholar]
микробиол. 59 : 695-700. [Бесплатная статья PMC] [PubMed] [Google Scholar]
37. Muyzer, G. 1998. Структура, функции и динамика микробных сообществ: молекулярно-биологический подход, с. 157-187. В Г. Р. Карвальо (ред.), Достижения в области молекулярной экологии. IOS Press, Амстердам, Нидерланды.
38. Pinhassi, J., F. Azam, J. Hemphälä, R.A. Long, J. Martinez, U.L. Zweifel, and Å. Хагстрем. 1999. Связь между видовым составом бактериопланктона, динамикой популяций и деградацией органического вещества. Аква. микроб. Экол. 17 : 13-26. [Google Scholar]
39. Рассел, Дж. Б. и Г. М. Кук. 1995. Энергетика бактериального роста: баланс анаболических и катаболических реакций. микробиол. Ред. 59 : 48-62. [Бесплатная статья PMC] [PubMed] [Google Scholar]
40. Schweitzer, B., and M. Simon. 1995. Ограничение роста планктонных бактерий в крупном мезотрофном озере. микроб. Экол. 30 : 89-104. [PubMed] [Google Scholar]
микроб. Экол. 30 : 89-104. [PubMed] [Google Scholar]
41. Секигучи Х., Н. Томиока, Т. Накахара и Х. Утияма. 2001. Одна полоса не всегда представляет отдельные бактериальные штаммы в анализе денатурирующего градиентного гель-электрофореза. Биотехнолог. лат. 23 : 1205-1208. [Google Scholar]
42. Серкиз С. М. и Э. М. Пердью. 1990. Выделение растворенного органического вещества из реки Суванни с помощью обратного осмоса. Вода Res. 24 : 911-916. [Google Scholar]
43. Søndergaard, M., and M. Middelboe. 1995. Межсистемный анализ лабильного растворенного органического углерода. Мар. Экол. прог. сер. 118 : 283-294. [Google Scholar]
44. Søndergaard, M., and J. Theil-Nielsen. 1997. Эффективность роста бактерий в культурах озерной воды. Аква. микроб. Экол. 12 : 115-122. [Google Scholar]
45. Степанаускас Р., Л. Леонардсон и Л. Дж. Транвик. 1999. Биодоступность ДОН, полученного из водно-болотных угодий, для пресноводного и морского бактериопланктона. Лимнол. океаногр. 44 : 1477-1485. [Google Scholar]
Степанаускас Р., Л. Леонардсон и Л. Дж. Транвик. 1999. Биодоступность ДОН, полученного из водно-болотных угодий, для пресноводного и морского бактериопланктона. Лимнол. океаногр. 44 : 1477-1485. [Google Scholar]
46. Stouthamer, A.H., and C. Bettenhaussen. 1973. Использование энергии для роста и поддержания в непрерывных и периодических культурах микроорганизмов. Биохим. Биоф. Acta 301 : 53-70. [PubMed] [Google Scholar]
47. Судзуки М. Т. и С. Дж. Джованнони. 1996. Смещение, вызванное отжигом матрицы при амплификации смесей генов 16S рРНК методом ПЦР. заявл. Окружающая среда. микробиол. 62 : 625-630. [Бесплатная статья PMC] [PubMed] [Google Scholar]
48. Teske, A., C. Wawer, G. Muyzer, and NB Ramsing. 1996. Распределение сульфатредуцирующих бактерий в стратифицированном фьорде (Мариагер-фьорд, Дания) по оценке наиболее вероятного количества и денатурирующего градиентного гель-электрофореза ПЦР-амплифицированных фрагментов рибосомной ДНК. заявл. Окружающая среда. микробиол. 62 : 1405-1415. [Бесплатная статья PMC] [PubMed] [Google Scholar]
заявл. Окружающая среда. микробиол. 62 : 1405-1415. [Бесплатная статья PMC] [PubMed] [Google Scholar]
49. Торсвик В., Даае Ф., Р.-А. Сандаа и Л. Овреас. 1998. Новые методы анализа микробного разнообразия в естественной и нарушенной среде. Дж. Биотехнология. 64 : 53-62. [PubMed] [Google Scholar]
50. Tranvik, LJ, and MG Höfle. 1987. Бактериальный рост в смешанных культурах на растворенном органическом углероде из гуминовых и чистых вод. заявл. Окружающая среда. микробиол. 53 : 482-488. [Бесплатная статья PMC] [PubMed] [Google Scholar]
51. Tranvik, LJ 1988. Доступность растворенного органического углерода для планктонных бактерий в олиготрофных озерах с различным содержанием гумуса. микроб. Экол. 16 : 311-322. [PubMed] [Академия Google]
52. Валлино, Дж. Дж., К. С. Хоппкинсон и Дж. Э. Хобби. 1996. Моделирование использования бактериями растворенного органического вещества: оптимизация заменяет кинетику роста Моно. Лимнол. океаногр. 41 : 1591-1609. [Google Scholar]
Лимнол. океаногр. 41 : 1591-1609. [Google Scholar]
53. Ван Ханнен, Э. Дж., М. П. Ван Агтервельд, Х. Дж. Гонс и Х. Дж. Лаанбрук. 1998. Выявление генетического разнообразия эукариотических микроорганизмов в водной среде с помощью денатурирующего градиентного гель-электрофореза. Дж. Фикол. 34 : 206-213. [Google Scholar]
54. Ван Ханнен, Э. Дж., Г. Зварт, М. П. Ван Агтервельд, Х. Дж. Гонс, Дж. Эберт и Х. Дж. Лаанбрук. 1999. Изменения в структуре бактериального и эукариотического сообщества после массового лизиса нитчатых цианобактерий, ассоциированных с вирусами. заявл. Окружающая среда. микробиол. 65 : 795-801. [Бесплатная статья PMC] [PubMed] [Google Scholar]
55. Van Hannen, EJ, W. Mooij, MP Van Agterveld, HJ Gons и HJ Laanbroek. 1999. Детритозависимое развитие микробного сообщества в экспериментальной системе: качественный анализ методом денатурирующего градиентного гель-электрофореза.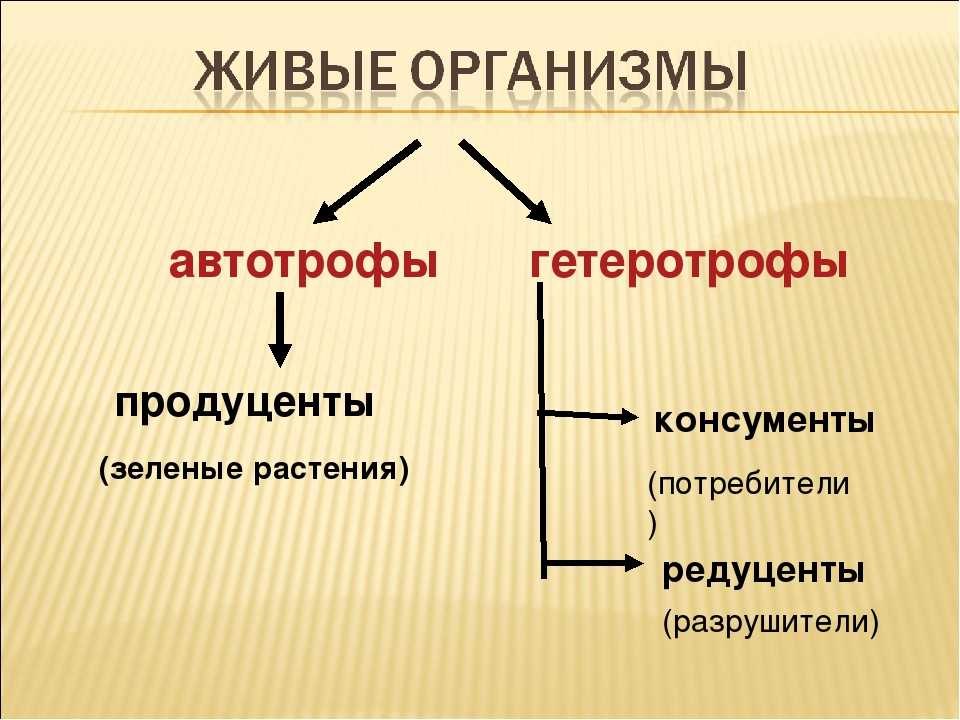
Leave A Comment