
⇐ Предыдущая12345678910Следующая ⇒ 1. биосферный 2. биогеоценотический 3. популяционно-видовой 4. организменный А3. К микроэлементам относятся 1. O, N, H, C 2. Au, C, H, Ra 3. Zn, Mn, Cu, Co 4. Au, U, Ra, Hg А4. Анионы в клетке обеспечивают 1. тургор клеточных стенок 2. поддержание концентрации ионов водорода 3. работу клеточных мембран 4. равномерное распределение тепла в клетке А5. Какие связи удерживают молекулы воды друг возле друга 1. водородные 2. пептидные 3. ковалентные 4. дисульфидные А6. Значительную часть клетки составляет вода, которая 1. образует веретено деления 2. образует глобулы белка 3. растворяет жиры 4. придает клетке упругость А7. Карбоксильная группа в аминокислоте определяет 1. 2. кислотные свойства 3. вид аминокислоты 4. служит радикалом А8. Вторичная структура белка – это 1.спираль 2. цепочка из аминокислот 3. глобула 4. соединение нескольких глобул
А9. К белкам, выполняющим каталитическую функцию, относится 1. гемоглобин 2. альбумин 3. фибрин 4. пепсин А10. Недостаток каких органических соединений в пище является наиболее опасным для детей 1. белков 2. углеводов 3. животных жиров 4. растительных жиров А11. Что из перечисленного относится к полисахаридам 1. глюкоза 3. рибоза 4. гликоген А12. Какое из перечисленных ниже соединений не построено из моносахаридов 1. крахмал 2. гемоглобин 3. гликоген 4. целлюлоза ⇐ Предыдущая12345678910Следующая ⇒
|

 Нарушение авторских прав; Мы поможем в написании вашей работы!
Нарушение авторских прав; Мы поможем в написании вашей работы! ..
..

Тест: Наука Биология — Биология 10 класс
Тест: Наука Биология — Биология 10 классАнглийский язык
Астрономия
Белорусский язык
Биология
География
ИЗО
Информатика
История
Итальянский язык
Краеведение
Литература
Математика
Музыка
Немецкий язык
ОБЖ
Обществознание
Окружающий мир
ОРКСЭ
Русский язык
Технология
Физика
Физкультура
Химия
Черчение
Для учителей
Дошкольникам
VIP — доступ
- Предметы »
- Биология »
- 10 класс »
- Наука Биология
Наука Биология
Тест составлен по пособию 2012г. Лернер. С одним или несколькими ответами.
Лернер. С одним или несколькими ответами.
Биология 10 класс | Автор: Супрун З.М. | ID: 5841 | Дата: 25.10.2015
Помещать страницу в закладки могут только зарегистрированные пользователи
Зарегистрироваться
Вопрос №
1
Предметом изучения общей биологии является:
1) строение и функции организма
2) природные явления
3) закономерности развития и функционирования живых систем
Вопрос №
2
Наиболее правильно следующее из утверждений:
1) только живые системы построены из сложных молекул
2) все живые системы обладают высокой степенью организации
3) живые системы отличаются от неживых составом химических элементов
4) в неживой природе не встречается высокая сложность организации системы
Вопрос №
3
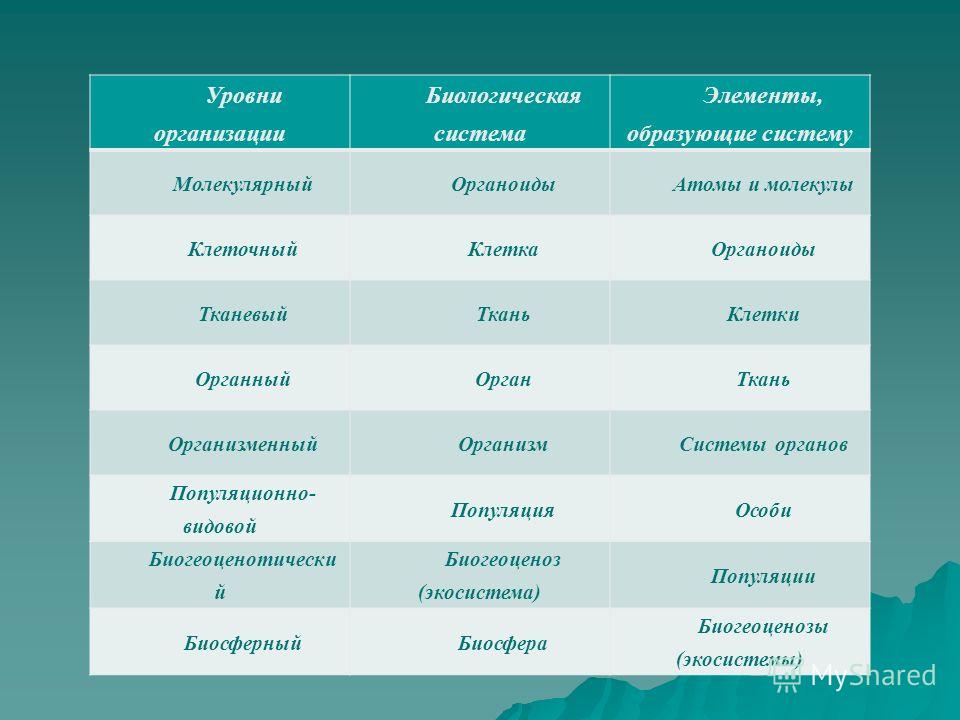
Минимальным уровнем организации жизни, на котором проявляется такое свойство живых систем, как способность к обмену веществ, энергии, информации, является:
1) биосферный
2) молекулярный
3) организменный
4) клеточный
Вопрос №
4
Высшим уровнем организации жизни является:
1) биосферный
2) биогеоценотический
3) популяционно-видовой
4) организменный
Вопрос №
5
Основным научным методом исследования в самый ранний период развития биологии был:
2) микроскопия
3) сравнительно — исторический
4) метод наблюдения и описания объектов
Вопрос №
6
Какое из приведенных утверждений наиболее правильно:
1) все организмы обладают одинаково сложным уровнем организации
2) все организмы обладают высоким уровнем обмена веществ
3) все организмы одинаково реагируют на окружающую среду
4) все организмы обладают одинаковым механизмом передачи наследственной информации
Вопрос №
7
Живые системы считаются открытыми потому, что они:
1) построены из тех же химических элементов, что и неживые системы
2) обмениваются веществом, энергией и информацией с внешней средой
3) обладают способностью к адаптациям
4) способны размножаться
Вопрос №
8
Уровень жизни, на котором начинают проявляться межвидовые отношения, называется:
1) биогеоценотическим
2) популяционно — видовым
3) организменным
4) биосферным
Вопрос №
9
Общим для всех уровней организации жизни свойством является:
1) сложность строения биологический системы
2) проявление закономерностей, действующих на каждом уровне
3) однородность элементов, составляющих систему
4) сходство качеств, которыми обладают разные системы
Вопрос №
10
Первым надорганизменным уровнем жизни считается:
1) биосферный
2) биогеоценотический
3) популяционно — видовой
4) организменный
Вопрос №
11
Факт сезонной линьки у животных установлен:
1) экспериментально
2) методом наблюдения
3) на основе литературных источников
4) на основе устных рассказов
Вопрос №
12
Изучение закономерностей появления и развития видов выясняется с помощью метода
1) наблюдения
2) экспериментального
3) моделирования
4) исторического
Вопрос №
13
Выберите три научные области, в которых основным методом является микроскопический.
А) Цитогенетика
Б) Гистология
В) Изучение родословных
Д) Микробиология
Е) Бионика
Ж) Биохимия
Вопрос №
14
Выберите только всеобщие свойства живых систем.
А) Способность к фотосинтезу
Б) Теплокровность
В) Обмен веществ
Г) Эукариотический тип строения клетки
Д) Наследственность
Е) Раздражимость
Показать ответы
Получение сертификата
о прохождении теста
Доступно только зарегистрированным пользователям
© TestEdu.ru 2013-2022
E-mail администратора: [email protected]
Суперорганизм — wikidoc
Суперорганизм — это организм, состоящий из множества организмов. Обычно это социальная единица эусоциальных животных, где разделение труда является узкоспециализированным и где особи не могут выживать самостоятельно в течение длительных периодов времени. Муравьи — самый известный пример такого сверхорганизма. Техническое определение суперорганизма — это «совокупность агентов, которые могут действовать согласованно, чтобы производить феномены, управляемые коллективом» 9.0005 [1] явления, представляющие собой любую деятельность, «нужную улью», например сбор муравьями пищи или выбор пчелами нового места для гнезда.
Муравьи — самый известный пример такого сверхорганизма. Техническое определение суперорганизма — это «совокупность агентов, которые могут действовать согласованно, чтобы производить феномены, управляемые коллективом» 9.0005 [1] явления, представляющие собой любую деятельность, «нужную улью», например сбор муравьями пищи или выбор пчелами нового места для гнезда.
Гипотеза Геи Джеймса Лавлока [2] и работы Джеймса Хаттона, Владимира Вернадского и Гая Мерчи предполагают, что биосферу можно рассматривать как сверхорганизм. Однако строгие экологические исследования обнаруживают, что внутри сообществ организмов мало или совсем нет самоконтроля, и такие сообщества обычно легко выходят из равновесия или превращаются в совершенно другие. Этому взгляду противостоит и уравновешивается Теория систем и динамика сложной системы.
В кибернетике, особенно в биокибернетике, суперорганизмы являются важной темой, поскольку они представляют собой форму «распределенного интеллекта», в которой множество отдельных агентов с ограниченным интеллектом и информацией могут объединять ресурсы для достижения цели, выходящей за пределы возможностей лица. Эта концепция имеет множество применений для военных и управленческих приложений и активно исследуется. [3]
Эта концепция имеет множество применений для военных и управленческих приложений и активно исследуется. [3]
Содержимое
- 1 Проблемы и замечания
- 2 Сверхорганическое в социальной теории
- 3 См. также
- 4 Каталожные номера
- 5 Внешние ссылки
Проблемы и критика
Концепция суперорганизма является спорной, поскольку многие биологи утверждают, что для того, чтобы социальная единица могла рассматриваться как организм сама по себе, индивидуумы должны находиться в постоянной физической связи друг с другом, и его эволюция должен регулироваться отбором ко всему обществу, а не к отдельным лицам. Хотя общепризнано, что общество эусоциальных животных является единицей естественного отбора, по крайней мере, до некоторой степени, большинство ученых-эволюционистов считают, что первичными единицами отбора по-прежнему остаются особи.
Остается вопрос «Что считать индивидуумом ?». Некоторые дарвинисты, такие как Ричард Докинз, предполагают, что выбранный индивидуум является эгоистичным геном. [4] Другие считают, что это весь геном организма. Э.О. Уилсон показал, что в случае с колониями муравьев и другими общественными насекомыми отбираются размножающиеся особи колонии, а не ее отдельные члены. Это может относиться к бактериальным членам строматолита, которые из-за общего генетического обмена каким-то образом составляют единый генофонд. Теоретики Геи, такие как Линн Маргулис, утверждают, что это в равной степени применимо к симбиогенезу бактериальных основ всей Земли.
Некоторые дарвинисты, такие как Ричард Докинз, предполагают, что выбранный индивидуум является эгоистичным геном. [4] Другие считают, что это весь геном организма. Э.О. Уилсон показал, что в случае с колониями муравьев и другими общественными насекомыми отбираются размножающиеся особи колонии, а не ее отдельные члены. Это может относиться к бактериальным членам строматолита, которые из-за общего генетического обмена каким-то образом составляют единый генофонд. Теоретики Геи, такие как Линн Маргулис, утверждают, что это в равной степени применимо к симбиогенезу бактериальных основ всей Земли.
Судя по компьютерным симуляциям, таким как Мир ромашек, биологический отбор происходит одновременно на нескольких уровнях.
Также считается, что люди (как зеркала Универсального Разума) являются сверхорганизмом, включающим микроорганизмы, такие как бактерии, и чье сознание состоит из совокупности процессов. Подсчитано, что «микробиота кишечника человека состоит из от 10 13 до 10 14 микроорганизмов, чей коллективный геном («микробиом») содержит как минимум в 100 раз больше генов, чем наш собственный[. ..] Наш микробиом значительно обогащенный метаболизм гликанов, аминокислот и ксенобиотиков, метаногенез и биосинтез витаминов и изопреноидов, опосредованный 2-метил-D-эритритол-4-фосфатным путем. [5] .
..] Наш микробиом значительно обогащенный метаболизм гликанов, аминокислот и ксенобиотиков, метаногенез и биосинтез витаминов и изопреноидов, опосредованный 2-метил-D-эритритол-4-фосфатным путем. [5] .
Тимоти Лири предположил, что на Земле существует только один организм: ДНК. Он описал все виды и физически независимые формы жизни как части этого организма, а их конечную цель — как рост за пределы планеты. Он также утверждал, что ДНК ранее росла и осела на Земле, а не возникла здесь (см. панспермию).
Сверхорганический в социальной теории
Эволюционист девятнадцатого века Герберт Спенсер ввел термин сверхорганический , чтобы сосредоточиться на социальной организации (первая глава его Принципов социологии озаглавлена «Сверхорганическая эволюция» [6] ), хотя это, по-видимому, было различием между органическим и социальным, а не идентичностью : Спенсер исследовал целостную природу общества как социального организма, выделяя при этом способы, которыми общество не ведет себя как организм. [7] Для Спенсера сверхорганическое было эмерджентным свойством взаимодействующих организмов, то есть людей. И, как утверждал Д. К. Филлипс, существует «разница между эмерджентностью и редукционизмом». [8]
[7] Для Спенсера сверхорганическое было эмерджентным свойством взаимодействующих организмов, то есть людей. И, как утверждал Д. К. Филлипс, существует «разница между эмерджентностью и редукционизмом». [8]
Точно так же экономист Карл Менгер расширил эволюционный характер социального роста, но никогда не отказывался от методологического индивидуализма. Менгер утверждал, что многие социальные институты возникли не как «результат социально-телеологических причин, а как непреднамеренный результат бесчисленных усилий экономических субъектов, преследующих «индивидуальные» интересы». [9]
Спенсер и Менгер оба утверждали, что, поскольку именно индивидуумы выбирают и действуют, любое социальное целое следует рассматривать меньше, чем организм, хотя Менгер подчеркивал это более решительно. Спенсер использовал органистическую идею для расширенного анализа социальной структуры, признавая, что это прежде всего аналогия. Итак, для Спенсера идея сверхорганического лучше всего обозначала определенный уровень социальной реальности выше уровня биологии и психологии, а не тождество один к одному с организмом.
Тем не менее, Спенсер также утверждал, что «каждый организм заметного размера является обществом», что кое-кто предположил, что проблема может быть терминологической. [10]
Термин сверхорганический был принят антропологом Альфредом Л. Кребером в 1917 году. [11]
Ссылки
- ↑ Келли, Кевин (1994). Из-под контроля: новая биология машин, социальных систем и экономического мира . Бостон: Аддисон-Уэсли. п. 98. ISBN 0-201-48340-8.
- ↑ Гайя: новый взгляд на жизнь на Земле , Джеймс Лавлок, Oxford University Press, 1979
- ↑ Келли, Кевин (1994). Из-под контроля: новая биология машин, социальных систем и экономического мира . Бостон: Аддисон-Уэсли. п. 251. ISBN 0-201-48340-8.
«Если полковник Торп [из DARPA США] добьется своего, четыре подразделения вооруженных сил США и сотни промышленных субподрядчиков станут единым взаимосвязанным суперорганизмом.
 Непосредственным шагом в этот мир распределенного интеллекта является разработанный инженерный протокол. консорциумом оборонных симуляционных центров в Орландо, Флорида…»
Непосредственным шагом в этот мир распределенного интеллекта является разработанный инженерный протокол. консорциумом оборонных симуляционных центров в Орландо, Флорида…» - ↑ Эгоистичный ген , Oxford University Press, 1976.
- ↑ Gill S.R., et al. Наука , 312 , 1355-1359 ( 2006 ). http://dx.doi.org/10.1126/science.1124234
- ↑ Принципы социологии , Vol. 1, часть 1. «Данные социологии», Герберт Спенсер, 1876 г.
- ↑ Принципы социологии , Vol. 1, часть 2, глава II, «Общество — это организм» (разделы 222 и 223), Герберт Спенсер, 1876 г.
- ↑ Холистическая мысль в социальных науках , DC Phillips, Stanford University Press, 1976, с. 123
- ↑ Исследования метода социальных наук с особым упором на экономику , Карл Менгер, Луи Шнайдер (переводчик), New York University Press, 1985
- ↑ Политическая философия Герберта Спенсера , Тим С. Грей, 1996, с. 211
- ↑ Образцы культуры , Рут Бенедикт, Хоутон Миффлин, 1934, с. 231
 Непосредственным шагом в этот мир распределенного интеллекта является разработанный инженерный протокол. консорциумом оборонных симуляционных центров в Орландо, Флорида…»
Непосредственным шагом в этот мир распределенного интеллекта является разработанный инженерный протокол. консорциумом оборонных симуляционных центров в Орландо, Флорида…» Грей, 1996, с. 211
Грей, 1996, с. 211Внешние ссылки
- Люди — гибрид человека и бактерии, журнал Wired, 11 октября 2004 г.
de:Сверхорганизм hu: Szuperorganizmus sv: Суперорганизм
Надорганизменные взаимодействия в кишечнике человека
- Список журналов
- Front Cell Infect Microbiol
- PMC4005949
Front Cell Infect Microbiol. 2014; 4: 47,
Опубликовано онлайн 2014 апреля 23. надорганизм» (который мы предпочитаем более распространенному, но несколько менее информативному «суперорганизму») относится к совокупности индивидуумов, которые ведут себя как единое целое с расширенными функциями. Первоначально он применялся к группам генетически идентичных особей, таких как колонии общественных насекомых (Мориц и Фукс, 19).98), но с тех пор он был расширен за счет включения систем, состоящих из таксономически разнообразных видов из всех областей жизни, а также вирусов (Salvucci, 2012). В кишечнике человека обитает до 10 14 бактерий, что примерно на порядок превышает количество собственных клеток хозяина: микробная концентрация в толстой кишке может достигать 10 12 клеток на грамм. Существует также большое количество вирусов, преимущественно бактериофагов, по крайней мере, 10 9 вирусных частиц на грамм человеческих фекалий. Текущие данные представляют картину преобладающего гомеостаза между хозяином, микробиомом и виромом, согласующуюся с описанием надорганизма, который, тем не менее, может входить в нарушенное альтернативное состояние, называемое «дисбиозом». Здесь мы рассматриваем эти доказательства и потенциал для принятия надорганизменных подходов к лечению и профилактике дисбактериоза в будущем.
Первоначально он применялся к группам генетически идентичных особей, таких как колонии общественных насекомых (Мориц и Фукс, 19).98), но с тех пор он был расширен за счет включения систем, состоящих из таксономически разнообразных видов из всех областей жизни, а также вирусов (Salvucci, 2012). В кишечнике человека обитает до 10 14 бактерий, что примерно на порядок превышает количество собственных клеток хозяина: микробная концентрация в толстой кишке может достигать 10 12 клеток на грамм. Существует также большое количество вирусов, преимущественно бактериофагов, по крайней мере, 10 9 вирусных частиц на грамм человеческих фекалий. Текущие данные представляют картину преобладающего гомеостаза между хозяином, микробиомом и виромом, согласующуюся с описанием надорганизма, который, тем не менее, может входить в нарушенное альтернативное состояние, называемое «дисбиозом». Здесь мы рассматриваем эти доказательства и потенциал для принятия надорганизменных подходов к лечению и профилактике дисбактериоза в будущем.
В желудочно-кишечном тракте (ЖКТ) взрослого человека обитает обширная и разнообразная популяция микроорганизмов, состоящая примерно из 10 14 особей с общим генетическим потенциалом примерно на два порядка выше, чем у хозяина (Sommer and Bäckhed, 2013). Анаэробные бактерии доминируют в этом микробном сообществе, которое также включает аэробные или факультативно анаэробные бактерии, археи и эукариоты, наряду с большим, но гораздо более плохо определенным вирусным сообществом (см. ниже). Несмотря на свое разнообразие, бактериальное сообщество включает лишь небольшое подмножество известных типов, что предполагает сильный отбор видов, адаптированных к среде хозяина и ориентированных на нейтральные или взаимовыгодные отношения с хозяином (Turnbaugh et al., 2009).; Консорциум проектов микробиома человека, 2012 г.). Типы Firmicutes и Bacteroidetes доминируют в микробиоме кишечника, также в изобилии присутствуют представители Proteobacteria, Actinobacteria и Fusobacteria. Колонизация желудочно-кишечного тракта младенцев начинается с рождения и инициируется факультативно анаэробными протеобактериями из-за изначально окислительной среды; анаэробные Bacteroidetes, Actinobacteria и Firmicutes колонизируются позже, а состав микробиома стабилизируется и начинает напоминать взрослый в возрасте от 1 до 2 лет (Секиров и др. , 2010).
, 2010).
О том, что кишечный микробиом приносит огромную пользу человеку-хозяину, свидетельствует его множество взаимодействий на протяжении всего развития (рис. ), а также негативные последствия его нарушения при дисбактериозе (Walker and Lawley, 2013). Ледерберг (2000) был первым, кто понял, что микробиом составляет часть человеческого «я» в теории надорганизма. Лучше всего определен вклад микробиоты желудочно-кишечного тракта в метаболический: эти микроорганизмы обладают совокупной метаболической способностью, эквивалентной таковой в печени, что оправдывает их описание в качестве дополнительного органа человека (Gill et al., 2006; Sommer and Bäckhed). , 2013). Их ключевые метаболические роли включают расщепление неперевариваемых полисахаридов на легкоусвояемые короткоцепочечные жирные кислоты (КЦЖК), производство необходимых витаминов и других метаболитов, а также детоксикацию вредных веществ (Walker and Lawley, 2013). Все эти действия дополняют метаболизм хозяина, расширяя его функциональные возможности без необходимости в дополнительном генетическом потенциале, кодируемом хозяином, в то время как другой стороной мутуалистических отношений является постоянная температура и богатая питательными веществами среда, предоставляемая микробиоте хозяином. Микробиота также играет роль в метаболических фенотипах, связанных с такими заболеваниями, как ожирение: наследственное ожирение ( ob / ob ) мышей имеют повышенный уровень SCFAs в слепой кишке, а их микробиота имеет повышенное содержание генов, участвующих в деградации полисахаридов, по сравнению с таковым у худых мышей (Tremaroli and Bäckhed, 2012), наблюдение повторено на людях. (Тернбо и др., 2009). Еще более поразительно, что трансплантация микробиоты от мыши-донора с ожирением стерильному реципиенту приводит к тому, что реципиент набирает вдвое больше жира, чем мышь, пересаженная микробиотой от худого донора (Turnbaugh et al., 2006), что подтверждает что микробиом может активно определять фенотипический ответ хозяина, а не просто пассивно реагировать на прием пищи. Кроме того, у реципиентов, пересаженных с «ожиревшей» микробиотой, не развивается фенотип ожирения, если они содержатся вместе с мышами, имеющими «худую» микробиоту. Это явление связано с переносом определенных видов Bacteroidetes из худой микробиоты в тучную, но зависит от диеты (Ridaura et al.
Микробиота также играет роль в метаболических фенотипах, связанных с такими заболеваниями, как ожирение: наследственное ожирение ( ob / ob ) мышей имеют повышенный уровень SCFAs в слепой кишке, а их микробиота имеет повышенное содержание генов, участвующих в деградации полисахаридов, по сравнению с таковым у худых мышей (Tremaroli and Bäckhed, 2012), наблюдение повторено на людях. (Тернбо и др., 2009). Еще более поразительно, что трансплантация микробиоты от мыши-донора с ожирением стерильному реципиенту приводит к тому, что реципиент набирает вдвое больше жира, чем мышь, пересаженная микробиотой от худого донора (Turnbaugh et al., 2006), что подтверждает что микробиом может активно определять фенотипический ответ хозяина, а не просто пассивно реагировать на прием пищи. Кроме того, у реципиентов, пересаженных с «ожиревшей» микробиотой, не развивается фенотип ожирения, если они содержатся вместе с мышами, имеющими «худую» микробиоту. Это явление связано с переносом определенных видов Bacteroidetes из худой микробиоты в тучную, но зависит от диеты (Ridaura et al. , 2013). Таким образом, существует сложное взаимодействие между геномом хозяина, микробиотой и окружающей средой, в котором все три компонента могут играть контролирующую роль (рис. ).
, 2013). Таким образом, существует сложное взаимодействие между геномом хозяина, микробиотой и окружающей средой, в котором все три компонента могут играть контролирующую роль (рис. ).
Открыть в отдельном окне
Взаимодействие между человеком-хозяином, кишечным микробиомом и виромом . Проиллюстрированы функциональные эффекты нормального кишечного микробиома, а также двунаправленные взаимодействия между микробиомом и виромом, а также между желудочно-кишечным трактом и мозгом через микробиом. На панелях справа показаны возмущения, связанные с различными измененными состояниями микробиома: безмикробным состоянием у лабораторных мышей, ожирением у мышей и людей и дисбиозом человека. Увеличенные параметры показаны желтым цветом и стрелками, указывающими вверх; уменьшенные параметры показаны красной штриховкой и стрелками, указывающими вниз.
Микробиом кишечника начинает формироваться с рождения и созревает вместе с иммунной системой хозяина; поэтому можно ожидать, что он внесет значительный вклад в развитие этой системы. Кишечник представляет собой уникальный биореактор с большой площадью поверхности, сверхплотным населенным биореактором с проницаемыми стенками, который должен сохранять биологическую целостность. Приобретение иммунной системы, способной переносить полезную микробиоту и эффективно реагировать на патогены, должно было стать решающим шагом в эволюция получившегося надорганизма. Это привело к предположению, что адаптивная иммунная система позвоночных могла эволюционировать, чтобы управлять сложными сообществами полезных микроорганизмов, обнаруживаемых в их желудочно-кишечном тракте, в отличие от врожденного иммунитета и либо очень простой, либо богатой микрофлорой беспозвоночных (McFall- Нгаи, 2007). Предположительно, это дало позвоночным преимущество в большей гибкости пищеварения, возможно, за счет восприимчивости к аутоиммунитету. Безусловно верно, что большинство взаимодействий иммунной системы связано с толерантностью к полезной микробиоте, а не с антагонизмом по отношению к патогенам (Arrietta and Finlay, 2012).
Кишечник представляет собой уникальный биореактор с большой площадью поверхности, сверхплотным населенным биореактором с проницаемыми стенками, который должен сохранять биологическую целостность. Приобретение иммунной системы, способной переносить полезную микробиоту и эффективно реагировать на патогены, должно было стать решающим шагом в эволюция получившегося надорганизма. Это привело к предположению, что адаптивная иммунная система позвоночных могла эволюционировать, чтобы управлять сложными сообществами полезных микроорганизмов, обнаруживаемых в их желудочно-кишечном тракте, в отличие от врожденного иммунитета и либо очень простой, либо богатой микрофлорой беспозвоночных (McFall- Нгаи, 2007). Предположительно, это дало позвоночным преимущество в большей гибкости пищеварения, возможно, за счет восприимчивости к аутоиммунитету. Безусловно верно, что большинство взаимодействий иммунной системы связано с толерантностью к полезной микробиоте, а не с антагонизмом по отношению к патогенам (Arrietta and Finlay, 2012). У стерильных мышей наблюдается недоразвитие связанной с кишечником лимфоидной ткани (GALT), измененная дифференцировка подтипов иммунных клеток и аномальный слой кишечной слизи, который можно восстановить путем лечения бактериальным липополисахаридом или пептидогликаном (Sommer and Bäckhed, 2013). Таким образом, взаимодействие между иммунной системой и микробиотой является взаимодействием взаимного усиления, а не контроля сверху вниз (рис. 1).
У стерильных мышей наблюдается недоразвитие связанной с кишечником лимфоидной ткани (GALT), измененная дифференцировка подтипов иммунных клеток и аномальный слой кишечной слизи, который можно восстановить путем лечения бактериальным липополисахаридом или пептидогликаном (Sommer and Bäckhed, 2013). Таким образом, взаимодействие между иммунной системой и микробиотой является взаимодействием взаимного усиления, а не контроля сверху вниз (рис. 1).
Наконец, влияние микробиома кишечника распространяется далеко за пределы самого желудочно-кишечного тракта. Одной из новых областей является взаимодействие между кишечной микробиотой и мозгом через так называемую «кишечно-мозговую ось» (Collins et al., 2012). Такие взаимодействия включают влияние кишечной микробиоты на уровни сигнальных молекул мозга, вызванное дисбактериозом усиление тревожного поведения у мышей, соответственно снижение тревожности у стерильных мышей (Sommer and Bäckhed, 2013) и одновременное возникновение психических расстройств.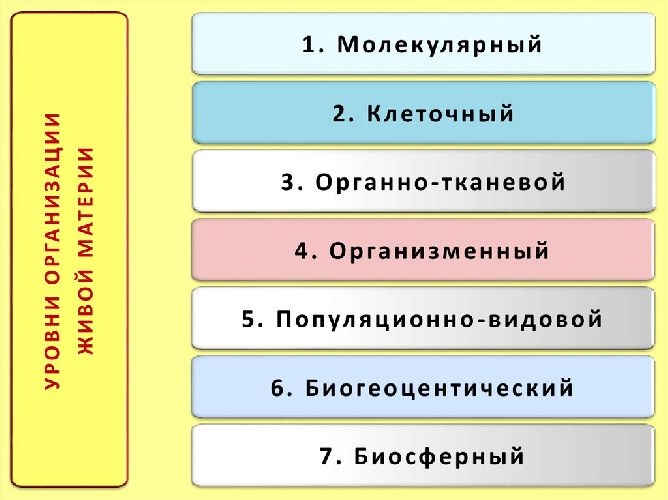 с дисбактериозами, такими как синдром раздраженного кишечника (Hsiao et al., 2013). Следовательно, бактериальные побочные продукты влияют на мозг напрямую или через эндокринные клетки или цитокины, в то время как гормоны или нейротрансмиттеры головного мозга модулируют микробиоту через физиологию кишечника или влияют на бактериальную передачу сигналов и экспрессию генов (Collins et al., 2012). Эти наблюдения подтверждают более широкую модель, в которой микробиота кишечника является интегрированным компонентом человеческого организма с множественными, широкомасштабными и двунаправленными взаимодействиями со многими другими его компонентами (рис. ).
с дисбактериозами, такими как синдром раздраженного кишечника (Hsiao et al., 2013). Следовательно, бактериальные побочные продукты влияют на мозг напрямую или через эндокринные клетки или цитокины, в то время как гормоны или нейротрансмиттеры головного мозга модулируют микробиоту через физиологию кишечника или влияют на бактериальную передачу сигналов и экспрессию генов (Collins et al., 2012). Эти наблюдения подтверждают более широкую модель, в которой микробиота кишечника является интегрированным компонентом человеческого организма с множественными, широкомасштабными и двунаправленными взаимодействиями со многими другими его компонентами (рис. ).
Численность и разнообразие вирусов превышает численность и разнообразие бактерий в большинстве экосистем, однако, за несколькими заметными исключениями (Dinsdale et al., 2008), их характеристики отстают от характеристик клеточных микроорганизмов. Подавляющее большинство вирусов представляют собой бактериофаги, нацеленные на прокариотические клетки, и в морских системах была проведена большая работа по изучению их влияния на структурирование микробиоты. Эти исследования показывают, что бактериофаги оказывают основное воздействие, воздействуя на самые быстрорастущие виды бактерий (гипотеза «Убить победителя»; Thingstad, 2000), вызывая резкую реструктуризацию микробного сообщества (Bouvier and del Giorgio, 2007) и увеличивая его численность. разнообразия (Hewson et al., 2003) с помощью процесса, сходного с отрицательным частотно-зависимым отбором (Ayala and Campbell, 19).74). Однако исследования вирома кишечника человека предполагают, что в нем может существовать довольно различная динамика бактериофагов. Хотя виром демонстрирует большую межличностную изменчивость, чем соответствующий микробиом, внутриличностное разнообразие невелико [отношение виротипов к бактериальным филотипам ~1:1 по сравнению с 10:1 в морских системах (Reyes et al., 2010)], вирусные частицы и бактериальные клетки присутствуют в сходных количествах, и большинство доминирующих виротипов стабильны во времени и демонстрируют умеренный (нелитический) образ жизни (Рисунок; Reyes et al.
Эти исследования показывают, что бактериофаги оказывают основное воздействие, воздействуя на самые быстрорастущие виды бактерий (гипотеза «Убить победителя»; Thingstad, 2000), вызывая резкую реструктуризацию микробного сообщества (Bouvier and del Giorgio, 2007) и увеличивая его численность. разнообразия (Hewson et al., 2003) с помощью процесса, сходного с отрицательным частотно-зависимым отбором (Ayala and Campbell, 19).74). Однако исследования вирома кишечника человека предполагают, что в нем может существовать довольно различная динамика бактериофагов. Хотя виром демонстрирует большую межличностную изменчивость, чем соответствующий микробиом, внутриличностное разнообразие невелико [отношение виротипов к бактериальным филотипам ~1:1 по сравнению с 10:1 в морских системах (Reyes et al., 2010)], вирусные частицы и бактериальные клетки присутствуют в сходных количествах, и большинство доминирующих виротипов стабильны во времени и демонстрируют умеренный (нелитический) образ жизни (Рисунок; Reyes et al. , 2012). Такая существующая временная вариация в значительной степени ограничивается литическими бактериофагами, такими как Microviridae, которые быстро эволюционируют (>10 -5 замен нуклеотидов -1 дней -1 ), что дает расхождение уровня, эквивалентного определению новых вирусных видов за 2,5-летний период (Minot et al., 2013). Вполне вероятно, что сочетание бактериального штамм-специфического отбора, долговременной стабильности большей части отобранного вирома и быстрой эволюции остальных приводит к крайней межличностной изменчивости кишечного вирома. Что еще более важно, мы предполагаем, что контраст между динамикой Лотка-Вольтерра взаимодействия вирус-бактерия в морских системах и очевидной совместной адаптацией вирома в кишечнике указывает на более сильную надорганизмальную организацию в последнем.
, 2012). Такая существующая временная вариация в значительной степени ограничивается литическими бактериофагами, такими как Microviridae, которые быстро эволюционируют (>10 -5 замен нуклеотидов -1 дней -1 ), что дает расхождение уровня, эквивалентного определению новых вирусных видов за 2,5-летний период (Minot et al., 2013). Вполне вероятно, что сочетание бактериального штамм-специфического отбора, долговременной стабильности большей части отобранного вирома и быстрой эволюции остальных приводит к крайней межличностной изменчивости кишечного вирома. Что еще более важно, мы предполагаем, что контраст между динамикой Лотка-Вольтерра взаимодействия вирус-бактерия в морских системах и очевидной совместной адаптацией вирома в кишечнике указывает на более сильную надорганизмальную организацию в последнем.
Вклад кодируемых фагами генов в бактериальные экосистемные функции хорошо задокументирован в морских системах. Цианофаги, инфицирующие виды Prochlorococcus и Synechococcus , несут ключевые функциональные гены, такие как гены, кодирующие компоненты фотосистемы, которые могут увеличивать первичную продукцию бактерий (Rohwer and Thurber, 2009). Сопоставимые эффекты вирома кишечника на функцию хозяина через кодируемый вирусом генетический или метаболический потенциал менее ясны. Многие из наиболее точно определенных примеров представляют собой факторы вирулентности, такие как фаг 9.33W Шига-подобный токсин (Plunkett et al., 1999), который способствует проникновению бактериальных патогенов в нормальную микробиоту; вирусы также участвуют в глобальном распространении генов устойчивости к антибиотикам посредством горизонтального переноса генов (Smillie et al., 2011). Однако есть признаки того, что виром может выполнять полезные кишечные функции для бактерий-хозяев, например, обнаружение многих генов анаэробного метаболизма в вирусных контигах фекального происхождения (Reyes et al., 2010) и возможность модификации утилизации углеводов бактериями. через фаговые рецепторы (Reyes et al., 2012).
Сопоставимые эффекты вирома кишечника на функцию хозяина через кодируемый вирусом генетический или метаболический потенциал менее ясны. Многие из наиболее точно определенных примеров представляют собой факторы вирулентности, такие как фаг 9.33W Шига-подобный токсин (Plunkett et al., 1999), который способствует проникновению бактериальных патогенов в нормальную микробиоту; вирусы также участвуют в глобальном распространении генов устойчивости к антибиотикам посредством горизонтального переноса генов (Smillie et al., 2011). Однако есть признаки того, что виром может выполнять полезные кишечные функции для бактерий-хозяев, например, обнаружение многих генов анаэробного метаболизма в вирусных контигах фекального происхождения (Reyes et al., 2010) и возможность модификации утилизации углеводов бактериями. через фаговые рецепторы (Reyes et al., 2012).
В вироме кишечника здоровых людей в значительной степени преобладают бактериофаги, при этом последовательности эукариотических вирусов либо не обнаруживаются, либо находятся на очень низком уровне (Minot et al.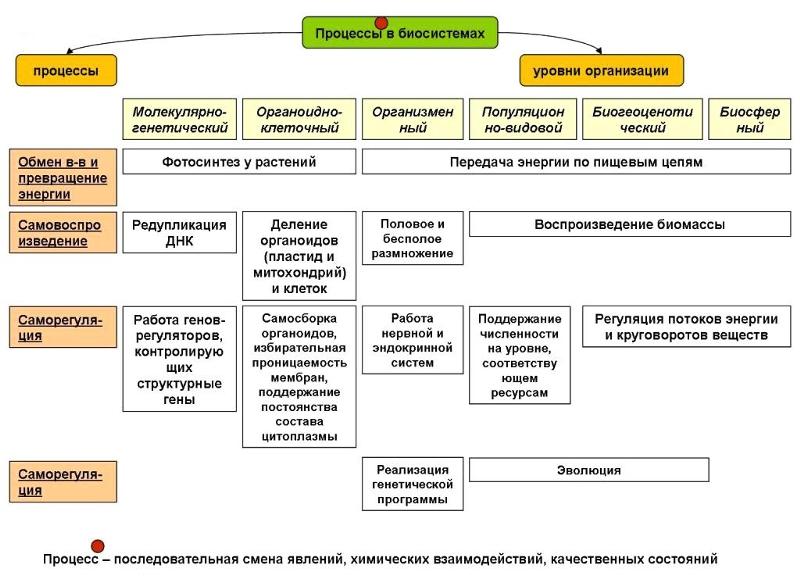 , 2013). Однако значительное появление эукариотических энтеровирусов особенно связано с заболеванием хозяина и нарушением микробиома дисбиоза, как обсуждается ниже.
, 2013). Однако значительное появление эукариотических энтеровирусов особенно связано с заболеванием хозяина и нарушением микробиома дисбиоза, как обсуждается ниже.
«Дисбиоз» — это довольно плохо определенный термин, который обычно используется для обозначения серьезного нарушения состава (кишечной) микробиоты, обычно связанного с влиянием на здоровье хозяина (Walker and Lawley, 2013). Он характеризуется снижением микробного разнообразия и преобладанием облигатных анаэробов, принадлежащих к типу Firmicutes, что часто связано со снижением количества Bacteroidetes и увеличением количества факультативно-анаэробных протеобактерий. Известные причины включают инфицирование патогенами желудочно-кишечного тракта, лечение антибиотиками широкого спектра действия и инициированные хозяином нарушения, такие как колоректальный рак (Walker and Lawley, 2013), но различают причинную и ответную роль микробиома при таких состояниях, как болезнь Крона. язвенный колит (ЯК) часто протекает тяжело.
Как анализ, так и, возможно, лечение дисбиоза могут выиграть от надорганизменного подхода. Помимо очевидного нарушения хозяина, вызванного диареей, при дисбиозе неблагоприятно воздействуют на зависящие от микробиома метаболические функции, такие как выработка SCFAs, что, в свою очередь, может усугубить диарею и способствовать хроническому воспалению (Ramakrishna and Mathan, 1993). Воспаление и нарушение эпителиального слизистого барьера, вероятно, сами по себе продлевают дисбактериоз, способствуя прикреплению и выживанию 9 клеток.0205 Enterobacteriaceae по сравнению с противовоспалительными видами Firmicutes (Willing et al., 2011). Эукариотический виром также может быть вовлечен в возникновение и/или поддержание дисбактериоза: диарея, вызванная такими агентами, как аденовирус, норовирус и ротавирус, связана с уменьшением разнообразия микробиомов, снижением уровня Bacteroidetes и увеличением количества Proteobacteria (Ma et al., 2011). ; Нельсон и др., 2012). Таким образом, существует сложный набор причинно-следственных взаимодействий, ведущих к сохранению дисбиотического состояния в надорганизме кишечника (рис.
Помимо очевидного нарушения хозяина, вызванного диареей, при дисбиозе неблагоприятно воздействуют на зависящие от микробиома метаболические функции, такие как выработка SCFAs, что, в свою очередь, может усугубить диарею и способствовать хроническому воспалению (Ramakrishna and Mathan, 1993). Воспаление и нарушение эпителиального слизистого барьера, вероятно, сами по себе продлевают дисбактериоз, способствуя прикреплению и выживанию 9 клеток.0205 Enterobacteriaceae по сравнению с противовоспалительными видами Firmicutes (Willing et al., 2011). Эукариотический виром также может быть вовлечен в возникновение и/или поддержание дисбактериоза: диарея, вызванная такими агентами, как аденовирус, норовирус и ротавирус, связана с уменьшением разнообразия микробиомов, снижением уровня Bacteroidetes и увеличением количества Proteobacteria (Ma et al., 2011). ; Нельсон и др., 2012). Таким образом, существует сложный набор причинно-следственных взаимодействий, ведущих к сохранению дисбиотического состояния в надорганизме кишечника (рис. 1).
1).
В последнее время наблюдается большой интерес к использованию трансплантации микробиоты, обычно с использованием фекалий здоровых доноров, для лечения дисбиоза кишечника (Palmer, 2011). Помимо клинических демонстраций этого подхода у пациентов с рецидивирующей инфекцией Clostridium difficile и язвенным колитом (Walker and Lawley, 2013), эксперименты на мышах демонстрируют, что это эффективный способ восстановления здорового микробиома у дисбиотических животных. устойчивы к последующему заражению патогенами (Lawley et al., 2012). Захватывающая перспектива использования бактериофаговых компонентов вирома для структурирования здорового микробиома и стимулирования полезных функций, таких как биосинтез питательных веществ (Reyes et al., 2012), была бы логическим продолжением такого подхода к лечению дисбактериоза. Наши быстро расширяющиеся знания о надорганизме, определяемом кишечником хозяина, его микробиоме и вироме, должны позволить нам воплотить в жизнь возможность такой терапии в ближайшие годы. Более того, общий подход связывания наших знаний об этом надорганизме с экологическими концепциями, касающимися гомеостаза и модуляции среды обитания ее резидентными организмами (Jones et al., 1994; Одлинг-Сми и др., 2003 г.; Dyke and Weaver, 2013) будет полезен для нашего понимания изменений состояния экосистемы в желудочно-кишечном тракте, которые мы теперь можем так легко отслеживать.
Более того, общий подход связывания наших знаний об этом надорганизме с экологическими концепциями, касающимися гомеостаза и модуляции среды обитания ее резидентными организмами (Jones et al., 1994; Одлинг-Сми и др., 2003 г.; Dyke and Weaver, 2013) будет полезен для нашего понимания изменений состояния экосистемы в желудочно-кишечном тракте, которые мы теперь можем так легко отслеживать.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Лаура Глендиннинг получает стипендию BBSRC EASTBIO. Мы благодарим Юлиана Петржика за комментарии к рукописи.
- Арриетта М.-К., Финли Б.Б. (2012). Комменсальная микробиота управляет иммунным гомеостазом. Передний. Иммунол. 3:33 10.3389/fimmu.2012.00033 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Ayala FJ, Campbell CA (1974). Частотно-зависимый отбор. Анну. Преподобный Экол. Сист. 5, 115–138
10.1146/annurev.es.05.110174.000555 [CrossRef] [Google Scholar]
- Bouvier T., del Giorgio P.A. (2007). Ключевая роль селективной вирусной смертности в определении состава морского бактериального сообщества. Окружающая среда. микробиол. 9, 287–297 10.1111/j.1462-2920.2006.01137.x [PubMed] [CrossRef] [Google Scholar]
- Collins S.M., Surette M., Bercik P. (2012). Взаимодействие микробиоты кишечника и головного мозга. Нац. Преподобный Микробиолог. 10, 735–742 10.1038/nrmicro2876 [PubMed] [CrossRef] [Google Scholar]
- Dinsdale E.A., Edwards R.A., Hall D., Angly F., Breitbart M., Brulc J.M., et al. (2008). Функциональное метагеномное профилирование девяти биомов. Природа 452, 629–632 10.1038/nature06810 [PubMed] [CrossRef] [Google Scholar]
- Дайк Дж. Г., Уивер И. С. (2013). Возникновение экологического гомеостаза в сложных экосистемах. PLoS-компьютер. биол. 9:e1003050
10.1371/journal.pcbi. 1003050 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Gill S. R., Pop M., Deboy R. T., Eckburg P. B., Turnbaugh P. J., Samuel B. S., et al. (2006). Метагеномный анализ микробиома дистального отдела кишечника человека. Наука 312, 1355–1359 гг. 10.1126/science.1124234 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Хьюсон И., Варго Г. А., Фурман Дж. А. (2003). Бактериальное разнообразие мелководного олиготрофного морского бентоса и вышележащих вод: последствия вирусной инфекции, сдерживание и обогащение питательными веществами. микроб. Экол. 46, 322–336 10.1007/s00248-002-1067-3 [PubMed] [CrossRef] [Google Scholar]
- Hsiao E.Y., McBride S.W., Hsien S., Sharon G., Hyde E.R., McCue T., et al. (2013). Микробиота модулирует поведенческие и физиологические отклонения, связанные с нарушениями развития нервной системы. Клетка 155, 1451–1463 гг. 10.1016/j.cell.2013.11.024 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Джонс К. Г., Лоутон Дж.Х., Шачак М. (1994). Организмы как инженеры экосистем. Ойкос
69, 373–386
10.2307/3545850 [CrossRef] [Google Scholar]
- Lawley T.D., Clare S., Walker A.W., Stares M.D., Connor T.R., Raisen C., et al. (2012). Целенаправленное восстановление кишечной микробиоты с помощью простой, определенной бактериотерапии устраняет рецидив болезни Clostridium difficile у мышей. PLoS Патог. 8:e1002995 10.1371/journal.ppat.1002995 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Ледерберг Дж. (2000). Инфекционная история. Наука 288, 287–293 10.1126/science.288.5464.287 [PubMed] [CrossRef] [Google Scholar]
- Ma C., Wu X., Nawaz M., Li J., Yu P., Moore J.E., et al. (2011). Молекулярная характеристика фекальной микробиоты у больных вирусной диареей. Курс. микробиол. 63, 259–266 10.1007/s00284-011-9972-7 [PubMed] [CrossRef] [Google Scholar]
- McFall-Ngai M. (2007). Адаптивный иммунитет: забота о сообществе. Природа 445, 153 10.1038/445153a [PubMed] [CrossRef] [Google Scholar]
- Майнот С. , Брайсон А., Чехуд С., Ву Г.Д., Льюис Дж.Д., Бушман Ф.Д. (2013). Быстрая эволюция вирома кишечника человека. проц. Натл. акад. науч. США 110, 12450–12455
10.1073/pnas.1300833110 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Moritz R. F. A., Fuchs S. (1998). Организация пчелиных семей: характеристики и последствия концепции надорганизма. Апидология 29, 7–21 10.1051/apido:19980101 [CrossRef] [Google Scholar]
- Нельсон А. М., Уок С. Т., Таубе С., Танючи М., Хоупт Э. Р., Вобус С. Э. и др. (2012). Нарушение микробиоты кишечника человека после норовирусной инфекции. ПЛОС ОДИН 7:e48224 10.1371/journal.pone.0048224 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Одлинг-Сми Ф.Дж., Лаланд К.Н., Фельдман М.В. (2003). Строительство ниши: забытый процесс эволюции, Vol. 37, Принстон, Нью-Джерси: Издательство Принстонского университета [Google Scholar]
- Палмер Р. (2011). Фекальные вещества. Нац. Мед. 17, 150–152 10.1038/nm0211-150 [PubMed] [CrossRef] [Google Scholar]
- Планкетт Г. , третий, Роуз Д. Дж., Дерфи Т. Дж., Блаттнер Ф. Р. (1999). Последовательность фага 933W шига-токсина 2 из Escherichia coli O157:H7: шига-токсин как продукт позднего гена фага. Дж. Бактериол. 181, 1767–1778 гг.
[Бесплатная статья PMC] [PubMed] [Google Scholar]
- Рамакришна Б.С., Матан В.И. (1993). Дисфункция толстой кишки при острой диарее: роль люминальных короткоцепочечных жирных кислот. кишки 34, 1215–1218 гг. [Бесплатная статья PMC] [PubMed] [Google Scholar]
- Reyes A., Haynes M., Hanson N., Angly F.E., Heath A.C., Rohwer F., et al. (2010). Вирусы в фекальной микробиоте монозиготных близнецов и их матерей. Природа 466, 334–338 10.1038/nature09199 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Рейес А., Семенкович Н. П., Уайтсон К., Ровер Ф., Гордон Дж. И. (2012). Становление вирусным: секвенирование нового поколения применяется к популяциям фагов в кишечнике человека. Нац. Преподобный Микробиолог. 10, 607–617 10.1038/nrmicro2853 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Ридаура В. К., Фейт Дж. Дж., Рей Ф. Э., Ченг Дж., Дункан А. Э., Кау А. Л. и др. (2013). Микробиота кишечника от близнецов, диссонирующих по ожирению, модулирует метаболизм у мышей. Наука
341:1241214
10.1126/science.1241214 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Rohwer F., Thurber RV (2009). Вирусы манипулируют морской средой. Природа 459, 207–212 10.1038/nature08060 [PubMed] [CrossRef] [Google Scholar]
- Salvucci E. (2012). Эгоизм, война и экономика; или интеграция, сотрудничество и биология. Передний. Клетка. Заразить. микробиол. 2:54 10.3389/fcimb.2012.00054 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Секиров И., Рассел С. Л., Антунес Л. К., Финлей Б. Б. (2010). Микробиота кишечника в норме и при патологии. Физиол. Откр. 90, 859–904. 10.1152/physrev.00045.2009 [PubMed] [CrossRef] [Google Scholar]
- Смилли К. С., Смит М. Б., Фридман Дж., Кордеро О. К., Дэвид Л. А., Алм Э. Дж. (2011). Экология управляет глобальной сетью обмена генами, соединяющей человеческий микробиом. Природа
480, 241–244
10.1038/nature10571 [PubMed] [CrossRef] [Google Scholar]
- Соммер Ф., Бекхед Ф. (2013). Кишечная микробиота — хозяева развития хозяина и физиологии. Нац. Преподобный Микробиолог. 11, 227–238 10.1038/nrmicro2974 [PubMed] [CrossRef] [Google Scholar]
- The Human Microbiome Project Consortium (2012). Структура, функции и разнообразие микробиома здорового человека. Природа 486, 207–214 10.1038/nature11234 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
- Thingstad TF (2000). Элементы теории механизмов контроля численности, разнообразия и биогеохимической роли литических бактериальных вирусов в водных системах. Лимнол. океаногр. 45, 1320–1328 гг. 10.4319/lo.2000.45.6.1320 [CrossRef] [Google Scholar]
- Tremaroli V., Bäckhed F. (2012). Функциональные взаимодействия между микробиотой кишечника и метаболизмом хозяина. Природа 489, 242–249 10.1038/nature11552 [PubMed] [CrossRef] [Google Scholar]
- Turnbaugh P.J., Hamady M.
 Частотно-зависимый отбор. Анну. Преподобный Экол. Сист. 5, 115–138
10.1146/annurev.es.05.110174.000555 [CrossRef] [Google Scholar]
Частотно-зависимый отбор. Анну. Преподобный Экол. Сист. 5, 115–138
10.1146/annurev.es.05.110174.000555 [CrossRef] [Google Scholar] 1003050 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
1003050 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] Г., Лоутон Дж.Х., Шачак М. (1994). Организмы как инженеры экосистем. Ойкос
69, 373–386
10.2307/3545850 [CrossRef] [Google Scholar]
Г., Лоутон Дж.Х., Шачак М. (1994). Организмы как инженеры экосистем. Ойкос
69, 373–386
10.2307/3545850 [CrossRef] [Google Scholar] , Брайсон А., Чехуд С., Ву Г.Д., Льюис Дж.Д., Бушман Ф.Д. (2013). Быстрая эволюция вирома кишечника человека. проц. Натл. акад. науч. США 110, 12450–12455
10.1073/pnas.1300833110 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
, Брайсон А., Чехуд С., Ву Г.Д., Льюис Дж.Д., Бушман Ф.Д. (2013). Быстрая эволюция вирома кишечника человека. проц. Натл. акад. науч. США 110, 12450–12455
10.1073/pnas.1300833110 [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] , третий, Роуз Д. Дж., Дерфи Т. Дж., Блаттнер Ф. Р. (1999). Последовательность фага 933W шига-токсина 2 из Escherichia coli O157:H7: шига-токсин как продукт позднего гена фага. Дж. Бактериол. 181, 1767–1778 гг.
[Бесплатная статья PMC] [PubMed] [Google Scholar]
, третий, Роуз Д. Дж., Дерфи Т. Дж., Блаттнер Ф. Р. (1999). Последовательность фага 933W шига-токсина 2 из Escherichia coli O157:H7: шига-токсин как продукт позднего гена фага. Дж. Бактериол. 181, 1767–1778 гг.
[Бесплатная статья PMC] [PubMed] [Google Scholar] К., Фейт Дж. Дж., Рей Ф. Э., Ченг Дж., Дункан А. Э., Кау А. Л. и др. (2013). Микробиота кишечника от близнецов, диссонирующих по ожирению, модулирует метаболизм у мышей. Наука
341:1241214
10.1126/science.1241214 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
К., Фейт Дж. Дж., Рей Ф. Э., Ченг Дж., Дункан А. Э., Кау А. Л. и др. (2013). Микробиота кишечника от близнецов, диссонирующих по ожирению, модулирует метаболизм у мышей. Наука
341:1241214
10.1126/science.1241214 [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar] Природа
480, 241–244
10.1038/nature10571 [PubMed] [CrossRef] [Google Scholar]
Природа
480, 241–244
10.1038/nature10571 [PubMed] [CrossRef] [Google Scholar]
Leave A Comment