Переходные формы в биологии: Переходные формы в эволюции животных ( к теме «Докозательства эволюции)
Ископаемые переходные формы
Вступление
Что такое переходные формы? Это виды, в строении которых возникают новые элементы, характерные для их эволюционных потомков, но еще сохраняются старые элементы, доставшиеся в наследство от эволюционных предков. А еще это источник мифов о Теории эволюции, стабильно работающий уже полтора столетия.
Ученые из года в год ищут «переходное звено» от обезьяны к человеку и никак не могут найти. Противники эволюционизма утверждают, что переходных форм нет, а этот ваш археоптерикс – тупиковая ветвь. Журналисты пугают общественность тем, что у человека и банана 50% общей генетической информации. Если птицы и крокодилы родня, то где окаменелости мезозойской крокоутки? – вопрошают диванные скептики. Да и от самого термина несет какой-то недоделанностью. Что это за вид такой, который «ни то ни сё»?
Давайте разбираться.
Древо жизни
Для того чтобы получить представление о переходных формах, надо обратиться к биологической систематике. Идея «разложить по полочкам» все разнообразие живых организмов не нова. В XVIII веке очередной способ классификации был предложен шведским натуралистом Карлом Линнеем. Способ оказался настолько удачным, что после важного апгрейда в середине XIX века используется до сих пор.
В чем тут суть? Неожиданно все формы жизни устроены не абы как. Анатомические структуры – все эти органы и ткани – не разбросаны в хаотичном порядке, а подчиняются определенным закономерностям. Вот, к примеру, позвоночник. Он есть у рыб, птиц, рептилий, зверей и амфибий. Но ни одного жука с позвоночником вы не найдете. Как нет и птиц без позвоночника. А те, у кого есть позвоночник, делятся на рыб (с жабрами и без лап) и четвероногих (с легкими и лапами). А четвероногие, например, могут быть пернатыми, шерстистыми, чешуйчатыми или с голой слизистой кожей. Описание, конечно же, очень упрощенное, но принцип понятен.
Анатомические структуры выстраиваются в иерархическом порядке, от общего к частному, что позволяет разделить все организмы на соответствующие иерархические группы. Отсюда получаем биологическую систематику с известными всем типами, классами, отрядами, семействами, родами и видами.
Но почему живые организмы устроены именно так? Ключом к ответу на этот вопрос стала теория эволюции, предложенная Чарльзом Дарвином в 1859 году. Иерархия анатомических структур отражает ход эволюции, а их единство у разных организмов говорит об общем происхождении. К настоящему времени этот тезис многократно подтвержден фактами из сравнительной анатомии, эмбриологии, палеонтологии и генетики.
Систему биологической классификации можно представить в виде дерева (скорее куста), на котором каждый отдельный листик – это вид, а ветви и стволы – разные таксоны. Такое «дерево» называют филогенетическим, эволюционным, или же Древом жизни.
Из вышесказанного для нас важными будут шесть следствий.
Следствие первое: строение организмов можно предсказать
Могут ли у кошек быть копыта? Если кит плавает в воде, то почему у него нет жабр? Где искать цветок папоротника? Чтобы ответить на подобные вопросы, вовсе не нужно осматривать всех китов и кошек, и следить за всеми папоротниками в Купальскую ночь. Зная биологическую систематику можно заранее сказать, какие анатомические признаки должны присутствовать у той или иной группы организмов.
Такой инструмент очень удобен в палеонтологии. Имея всего один зуб можно классифицировать животное, иногда с точностью до рода. И на основании этого сделать вполне надежную реконструкцию. А уже потом, когда (и если) будет найден полный скелет, можно уточнять детали и делать нашу реконструкцию все более реалистичной.
Но можно обойтись и вовсе без останков. Например, если нас интересует общий предок всех кошачьих, мы можем составить абстрактный портрет, исходя из признаков, характерных для этого семейства. Получается, что окаменелостей нет, а мы уже знаем, кого искать.
Следствие второе: общих родственников можно найти
Древо жизни охватывает абсолютно все виды организмов. А значит мы все друг другу родственники. Можно взять два абсолютно любых вида, а затем спуститься вниз по веточкам нашего Древа, до точки пересечения. В этой точке будет находиться общий предок, а в его строении будет совмещены признаки, общие для обеих ветвей. Даже если идти придется очень далеко вниз.
Например, предок человека и банана по совместительству будет предком вообще всех растений и животных. Это одноклеточный организм, живший как минимум 3 млрд лет назад, то есть раньше самых древних следов растений в горных породах. И наш геном действительно хранит информацию, оставшуюся от этого далекого-далекого предка.
Следствие третье: переходные формы повсюду
От древних одноклеточных организмов до человека – очень долгий путь. Поколение за поколением, вид за видом наши предки менялись под действием эволюции. Древо жизни росло, давало множество новых побегов и в итоге получилось современное видовое разнообразие. Можно ли сказать, что на этом эволюция остановилась? Конечно, нет. Те современные виды, которые не вымрут, дадут свое эволюционное потомство. И человек тут не исключение.
А значит, каждый вид в любой момент времени является потенциально переходным между какими-то старыми и новыми видами. Мы все – переходное звено.
Но не стоит ждать, что наши потомки будут так уж сильно от нас отличаться. Эволюционные изменения накапливаются постепенно, дети все так же похожи на родителей, и заметить отличия можно изменив масштаб. На промежутке в 1 млн лет мы точно сможем различить два вида, но вот найти точку, в которой один вид стал другим – не получится.
Следствие четвертое: некоторые переходные формы интереснее других
Как и в истории, эволюционные события могут иметь разный масштаб. Например, процесс эволюции уток, поиск прото-утки и переходных форм между ними – это все очень интересно. Но на масштабные эволюционные события не тянет. Другое дело – происхождение птиц вообще.
Из современных животных ближайшая родня уток – это крокодилы. Спускаемся по веточкам нашего дерева, находим точку пересечения. Здесь, в этой точке, в триасовом периоде жил их общий предок. Внешне был похож на тощего голенастого крокодила. Имел четырехкамерное сердце, высокий метаболизм и откладывал яйца. Не то чтобы сильно похоже на птицу, но уже кое-что. Значит, в качестве переходной формы нам надо искать кого-то похожего, но с крыльями и перьями. В XIX веке про родство птиц с крокодилами не знали, искали просто пернатого птице-ящера. И нашли. Им оказался археоптерикс, вошедший во все учебники биологии как эталон переходной формы. Так вот он – родоначальник птиц? Вообще-то нет.
Следствие пятое: неуловимые предки
Вообще-то обнаружение переходной формы еще не обозначает обнаружение того самого вида-предка, который дал всю дальнейшую эволюционную ветвь. И чем шире область поиска, чем более древняя у нас переходная форма, тем меньше для этого шансов. Как так?
Переходная форма – это все равно, что фоторобот, набор анатомических структур, которые точно должны быть. Например: скелет рептилии (включая длинный хвост и зубы), легкие кости, перья, небольшой размер. Это сокращает поиск до одной из групп динозавров, живших в юрском периоде. А дальше начинаются проблемы.
В группе – сотни видов, не имеющих узкой специализации. Они жили на промежутке в несколько десятков млн лет, приходились друг другу какими-то родственниками и все обладают тем набором признаков, который нам нужен. То есть потенциально они все – переходные формы к птицам. Но какой шанс, что нам улыбнулась удача и из нескольких сотен видов мы нашли «того самого»?
Разумеется, изучая этих оперенных то-ли еще динозавров, то-ли уже птиц, мы будем накапливать информацию и отсеивать ненужные варианты. Археоптерикс окажется хоть и ранней птицей, но параллельной ветвью эволюции. Наш портрет птицы-первопредка будет становиться все точнее и точнее. Можно будет локализовать поиск до ранней юры Северного Китая, а таксономию до манирапторов. Но указать на конкретный вид-предок мы вряд ли сможем.
Следствие шестое: и все-таки их можно отыскать
Так ли уж важно знать, какой из десятка видов птицеподобных динозавров был точно первоптицей? Для специалистов важно, для широкого круга людей – пожалуй, не очень. Что уж говорить о предках жесткокрылых или голосеменных растений. А если задать вопрос о происхождении человека?
Про нас самих нам нужно знать как можно более точно. И в этом плане в сравнении с птицами нам крупно повезло. Все-таки мы изучаем молодые останки, большинству которых нет и 1 млн лет. Их достаточно много, сохранность хорошая, а если они моложе 100 тыс лет – есть возможность использовать палеогенетику. А значит, мы можем не просто указать на переходную форму от австралопитеков к нам, но и восстановить нашу эволюцию, шаг за шагом.
Сегодня вместо давно устаревшей линейной схемы, мы имеем разветвленное деревце, объединяющее наших родственников и предков. Это все равно что семейная фотография с несколькими поколениями. Мы с большой точностью можем назвать своих родителей и их родителей. А вот с остальными не всегда понятно, кто с кем в каком родстве состоит. Антропология развивается стремительно, создаются новые методы исследований и делаются замечательные открытия. Разумеется, восстановить всю цепочку поколений (как того иногда требуют креационисты) мы не сможем. Но серьезно увеличить детальность и сложность и без того непростой картины антропогенеза ученым вполне по силам.
Заключение
Подведем итог.
Переходные формы действительно существуют. И в большинстве своем они объединяют группу родственных видов, имеющих специфический набор анатомических особенностей. Часть этих особенностей новые и они получат развитие у потомков. Часть – старые, доставшиеся от предков и к потомкам они не перейдут. Особый интерес представляют переходные формы между крупными таксонами. Например, между рыбами и амфибиями. Или рептилиями и птицами, рептилиями и млекопитающими. На их примере можно видеть, как незначительные изменения приводят к серьезным результатам.
Благодаря развитию палеонтологии известно множество переходных форм. Эволюция позвоночных животных, например, изучена достаточно детально. И самая, пожалуй, изученная область – это эволюция человека. И все-таки, белых пятен хватит еще не на одно поколение палеонтологов. А учитывая неполноту геологической летописи, весь паззл мы не соберем никогда. Тем важнее вклад каждого, кто решил связать свою жизнь с палеонтологией. Возможно, и вы станете первооткрывателями недостающего звена.
Переходная форма | Info-Farm.RU
Переходные (промежуточные) формы — организмы, которые сочетают в своем строении признаки двух больших систематических групп.
Переходные формы характеризуются наличием более древних и примитивных (в смысле первичных) черт, чем более поздние формы, но, в то же время, наличием более прогрессивных (в смысле более поздних) черт, чем их предки. Как правило, термин «переходные формы» употребляют по отношению к ископаемых форм, хотя промежуточные виды совсем не обязательно должны умирать.
Переходные формы используют как одно из доказательств существования биологической эволюции.
История понятия
В 1859 г.., Когда была издана работа Ч. Дарвина «Происхождение видов», количество ископаемых остатков была крайне малой, науке не были известны переходные формы. Дарвин описал отсутствие промежуточных форм «как наиболее очевидное и тяжелое возражение, которое может быть против теории», но объяснил это крайней неполнотой геологической летописи. Он отмечал ограниченное количество доступных коллекций в то время, в то же время описал имеющуюся информацию об имеющихся ископаемые образцы с точки зрения эволюции и действия естественного отбора. Только два года спустя, в 1961 году. Был найден археоптерикс, который представлял классическую переходную форму между пресмыкающимися и птицами. Его находках, стала не только подтверждением теории Дарвина, а также знаковым фактом, подтверждающий реальность существования биологической эволюции. С тех пор было найдено большое количество ископаемых форм, которые показывают, что все классы позвоночных животных являются родственными между собой, причем большинство из них — через переходные формы.
С увеличением сведений о таксономическое разнообразие сосудистых растений в начале ХХ в., Начались исследования по поиску их возможного предка. В 1917 г.. Роберт Кидстон и Уильям Генри Ленд обнаружили остатки очень примитивной растения возле поселка
Rhynia в Шотландии. Это растение было названо Rhynia. Она сочетает в себе признаки зеленых водорослей и сосудистых растений.
Трактовка понятия
Переходные формы между двумя группами организмов не обязательно потомками одной группы и предком другой. По ископаемыми, как правило, невозможно точно установить является ли определенный организм предком другого. Кроме того, вероятность найти в палеонтологической летописи прямого предка определенной формы чрезвычайно мала. Гораздо больше вероятность обнаружить относительно близких родственников этого предка, которые сходны с ним по строению. Поэтому любая переходная форма автоматически интерпретируется как боковая ветвь эволюции, а не «участок филогенетического ствола».
Переходные формы и таксономия
Эволюционная таксономия оставалась доминирующей формой таксономии течение ХХ в. Выделение таксонов базируется на различных признаках, вследствие чего таксоны изображают в виде ветвей разветвленного эволюционного дерева. Переходные формы рассматриваются как «падающие» между различными группами в плане анатомии, они смесь характеристик от внутренней и внешней клади, что недавно разделилась.
С развитием кладистики в 1990-х гг. Взаимосвязи обычно изображают в виде кладограмы, иллюстрирующую дихотомическое ветвление эволюционных линий. Поэтому в кладистици переходные формы рассматриваются как более ранние ветви дерева, где еще не развились не все черты, характерные для ранее известных потомков на этой ветке. Такие ранние представители группы обычно называют основным таксоном (англ. Basal taxa) или сестринским таксоном (англ. Sister taxa), в зависимости от того, принадлежит ли ископаемый организм к данной клади или нет.
Проблемы выявления и интерпретации
Отсутствие переходных форм между многими группами организмов является предметом критики со стороны креационистов. Однако далеко не каждая переходная форма существует в виде окаменелостей из-за принципиальной неполноты палеонтологической летописи. Неполнота вызвана особенностями процесса фосилизации, то есть перехода в окаменевший состояние. Для образования окаменелости необходимо, чтобы организм, который погиб, был погребен под большим слоем осадочных пород. Из-за очень медленную скорость осадконакопления на суше, сухопутные виды редко переходят в окаменевший состояние и сохраняются. Кроме того, редко удается выявить виды, которые живут в глубинах океана через редкие случаи поднятия на поверхность больших массивов дна. Таким образом, большинство известных ископаемых (а, соответственно, и переходных форм) — это либо виды, обитающие на мелководье, в морях и реках, или наземные виды, которые ведут полуводный образ жизни, или живут у береговой линии. К упомянутым выше проблемам следует добавить чрезвычайно малую (в масштабах планеты) количество палеонтологов, которые осуществляют раскопки.
Переходные формы, как правило, не живут на больших территориях и не существуют в течение большого времени, иначе они были бы персистентный. Этот факт также снижает вероятность фосилизации и последующего обнаружения переходных форм.
Поэтому вероятность обнаружения промежуточных форм чрезвычайно мала.
Примеры среди животных
Древнейшими представителями земноводных считают ихтиостеги. Их считают переходным звеном между кистеперых рыбами и земноводными. Несмотря на то, что в ихтиостеги была пятипалая концовка, адаптированная к жизни на суше, значительную часть жизни они проводили как рыбы, имели хвостовой плавник, боковую линию и некоторые другие признаки рыб.
Батрахозавры, существовавшие в каменноугольный и пермский периоды, рассматривают как переходную форму между земноводными и пресмыкающимися. Батрахозавры, хоть и проводили жизнь во взрослой стадии на суше (подобно пресмыкающихся), были тесно связаны с водоемами и сохранили ряд признаков, присущих земноводным, в частности, откладывания икры и развитие личинок в воде, наличие жабр и тому подобное.
Обнаружено большое количество пресмыкающихся, которые выработали способность летать, часть из них имела перья, поэтому их рассматривают как переходные формы между пресмыкающимися и птицами. Наиболее известен археоптерикс. Он был размером примерно с современную ворону. Формой тела, строением конечностей и наличием оперения подобный современных птиц, возможно, летал. Общим с пресмыкающимися была особое строение таза и ребер, наличие клюва с коническими зубами, по три свободные пальцы на крыльях, подвийноувигнути позвонки, длинный хвост с 20-21 позвонка, кости могло не пневматизовани, грудная кость без киля. Другие известные переходные формы между пресмыкающимися и птицами — протоавис, конфуциусорниса.
Большое количество ископаемых форм звероподобных пресмыкающихся (синапсиды, терапсид, пеликозавров, различных динозавровых и др.), Найденных во многих районах земного шара, существовали в юрский и меловой периоды, сочетающие признаки пресмыкающихся и млекопитающих, раскрывают возможные направления и способы становления различных групп четвероногих , в частности млекопитающих. Например, звероподобный пресмыкающееся из группы терапсид — лиценопс
(Lycaenops) по развитию костей ротовой полости, дифференцированием зубов на клыки, резцы, по резцовые зубы и рядом других признаков строения тела напоминает хищных млекопитающих, хотя по другим признакам и образом жизни это были настоящие пресмыкающиеся.
Одной из форм, сохранилась в ископаемом состоянии является амбулоцетус
Ambulocetus natans («ходячий кит») — переходная форма между наземными млекопитающими и китообразными, которые являются вторинноводнимы формами. Внешне животное напоминало нечто среднее между крокодилом и дельфином. Кожа должна частично редуцированную шерсть. Животное имело лапы с перепонками; хвост и конечности приспособлены как вспомогательные органы передвижения в воде.
Примеры среди растений
Первые наземные растения из класса риниопсид, семей риниевих и псилофитовых, живших в силуре — девоне, сочетали признаки зеленых водорослей и примитивных форм высших растений. Их тело было безлистным, цилиндрический осевой орган — телом в верхней части дихотомически разветвленным на верхушках с спорангиями. Функцию минерального питания риниопсид выполняли ризоиды.
Ископаемые формы семенных папоротников, которые процветали в конце девона, сочетают в себе признаки папоротников и голосеменных. Они образовывали не только споры (как папоротники), но и семена (как семенах растения). Проводящая ткань их стеблей по строению напоминает древесину голосеменных (саговников).
Другой предшественник семенных растений был идентифицирован из отложений среднего девона. Рункария (Runcaria heinzelinii) существовала около 20 млн лет назад. Это была небольшая растение с радиальной симметрией; имела спорангий, окруженный интегументом и плюской. Рункария демонстрирует путь эволюции растений от споровых к семенных.
Переходные формы в эволюции человека
В наше время найдено большое количество ископаемых останков, которые раскрывают эволюционный путь человека разумного от ее человекообразных предков. К формам, которые в большей или меньшей степени можно отнести к переходным, относятся: сахелантропа, ардипитека, австралопитеки (африканский, афарский и другие), человек умелый, человек работающий, человек прямоходящий, человек-предшественник, гейдельбергский человек и кроманьонцы.
Среди упомянутых форм значительное внимание заслуживают австралопитеки. Австралопитек афарский с точки зрения эволюции находится между современными двуногими людьми и их четвероногими древними предками. Большое количество рис скелета этого австралопитека четко отражают двуногость, причем до такой степени, что некоторые исследователи считают, что это свойство возникла задолго до появления австралопитека афарского. Среди общих черт анатомии, его таз гораздо больше похож на этих костей у человека, чем у обезьян. Края подвздошных костей короче и шире, крестцовая кость широкая и расположена непосредственно позади тазобедренного сустава. Существует явное свидетельство о существовании мест крепления для мышц-разгибателей колена, предусматривает вертикальное положение этого организма. В то время, как таз австралопитека не совсем как у человека (заметно шире, с ориентацией края подвздошных костей наружу), эти особенности указывают на принципиальную перестройку, связанную с хождением на двух ногах. Бедренная кость образует угол в направлении колена. Эта черта позволяет ноге размещаться ближе к средней линии тела и является явным свидетельством привычный характер передвижения на двух ногах. В наше время человек разумный, орангутаны и коаты имеют такие же черты. Ноги австралопитека имели большие пальцы, что делает практически невозможным захват стопой ветвей деревьев. Кроме особенностей локомоции, в австралопитека был также значительно больше мозг, чем у современных шимпанзе и зубы были значительно больше подобными зубов современного человека, чем к обезьянам.
Филогенетические ряды
Филогенетические ряды — ряды ископаемых форм, связанные между собой в процессе эволюции и отражают постепенные изменения их исторического развития.
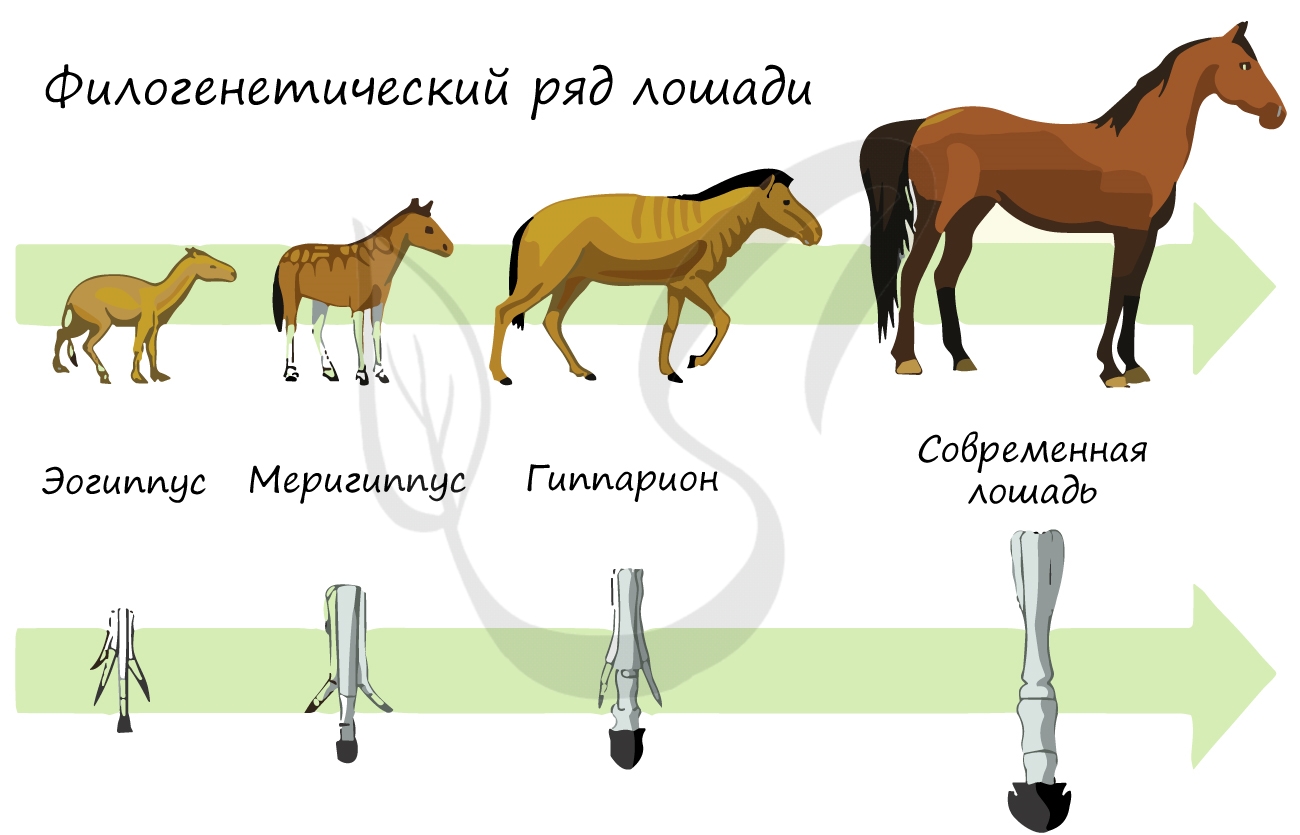
Были исследованы русским ученым А. Ковалевским и английским Дж. Симпсоном. Они показали, что современные однопалого копытные происходят от древних мелких всеядных животных. Анализ ископаемых лошадей помог установить постепенность процесса эволюции в пределах этой группы животных, в частности, как изменяясь во времени, ископаемые формы приобретали все большего сходства с современными лошадьми. Сравнивая эоценового еогипуса с современным конем, трудно доказать их филогенетическую родство. Однако наличие ряда переходных форм, которые последовательно сменяли друг друга на больших пространствах Евразии и Северной Америки, позволила восстановить филогенетический ряд лошадей и установить направление их эволюционных изменений. Он состоит из ряда следующих форм (в упрощенном виде): Phenacodus — Eohippus — Miohippus — Parahippus — Pliohippus — Equus.
Гильгендорф (1866) описал палеонтологический ряд брюхоногих моллюсков из миоценовых отложений, накопившихся в течение двух миллионов лет в озерных отложениях Штейнгеймського бассейна (Вюртемберг, Германия). Было обнаружено в последовательных слоях 29 различных форм, принадлежащих к ряду планорбис (Planorbis). Древние моллюски имели раковину в виде спираи, а более поздние — в виде турбоспирали. Ряд имел два ответвление. Предполагается, что изменение формы черепашки была вызвана повышением температуры и увеличением содержания карбоната кальция в результате горячих вулканических источников.
Таким образом, филогенетические ряды представляют собой историческую последовательность переходных форм.
В настоящее время известны филогенетические ряды для аммонитов (Вааген, 1869), брюхоногих моллюсков рода живородок (Viviparus) (Неймайром, 1875), носорогов, слонов, верблюдов, парнокопытных и других животных.
Изображения по теме
Доказательства эволюции, подготовка к ЕГЭ по биологии
Пути эволюции
В своих работах советский ученый Северцов А.Н. выделил понятия биологического прогресса и регресса.
Биологический прогресс подразумевает победу вида в борьбе за существование. Биологический прогресс характеризуется следующими признаками:
Численность вида увеличивается
Ареал расширяется
Смертность особей уменьшается
Рождаемость увеличивается
Происходит процветание вида
Основными направлениями биологического прогресса являются:
Ароморфоз (греч. airomorphosis — поднимаю форму)
Ароморфоз представляет собой прогрессивное эволюционное преобразование, повышающее уровень организации организмов.
В результате ароморфоза становится возможным освоение новых, ранее недоступных для жизни, территорий. К примеру,
теплокровность птиц позволила им заселить места с холодным климатом.
Идиоадаптация подразумевает незначительные, частные изменения в строении и функциях организма, которые помогают
приспособиться к условиям среды обитания. Идиоадаптации существенно не повышают уровень организации.
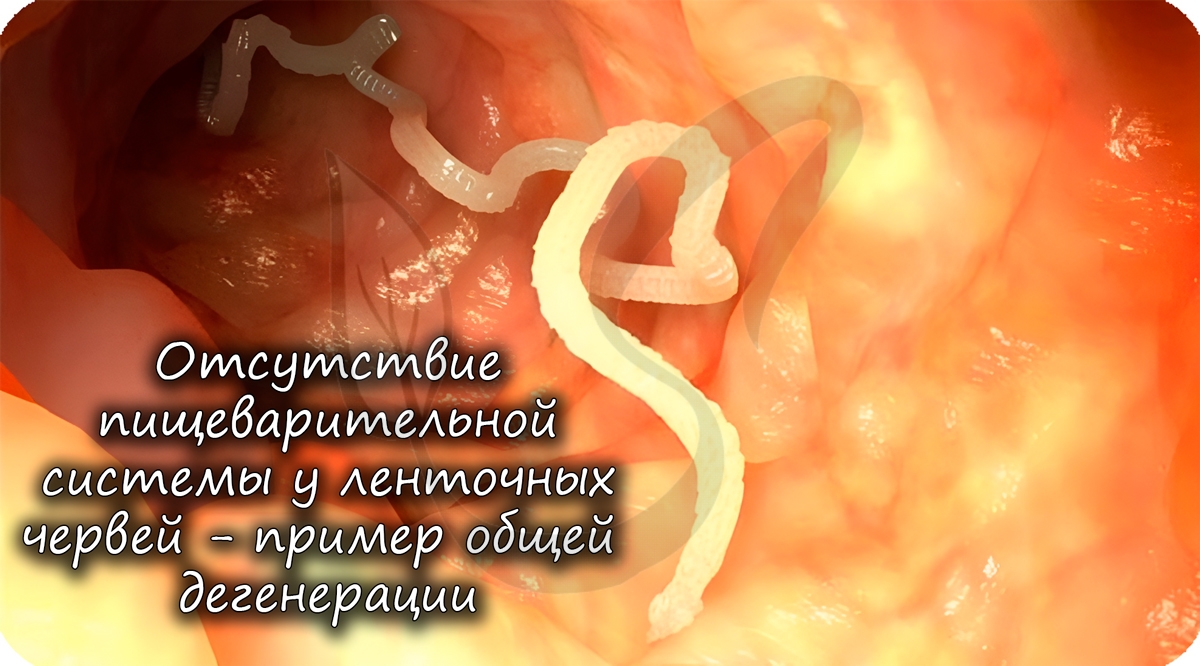
Общая дегенерация (лат. degenero — вырождаться, перерождаться)
Общей дегенерацией называют упрощение организации, которое заключается в утрате отдельных органов и систем органов.
У многих этот пункт вызывает внутреннее противоречие: как общая дегенерация может относиться к биологическому прогрессу?
На самом деле, если орган или система органов не нужна организму в его условиях обитания — то зачем она? Эта система
может исчезнуть и освободить место для других, более полезных в данных условиях, органов.
У многих паразитов отсутствуют различные органы, к примеру, у ленточных червей нет пищеварительной системы. А зачем она
им, когда пища в кишке, где они обитают, уже переварена и расщеплена организмом хозяина?
Биологический регресс характеризуется признаками, противоположными биологическому прогрессу:
Численность вида уменьшается
Ареал сужается
Смертность особей возрастает
Рождаемость уменьшается
Происходит вымирание вида
Главная причина биологического регресса в том, что скорость эволюции вида отстает от скорости изменения внешней среды, эволюции других видов: это несоответствие снижает приспособленность организмов. Часто деятельность человека молниеносно
меняет окружающую среду: далеко не все виды могут приспособиться к этому, происходит вымирание.
Изучение строения органов и их эволюционных изменений у различных групп организмов является основой выявления сравнительно-анатомических доказательств эволюции. Яркими примерами анатомических доказательств эволюции являются гомологичные
и аналогичные органы.
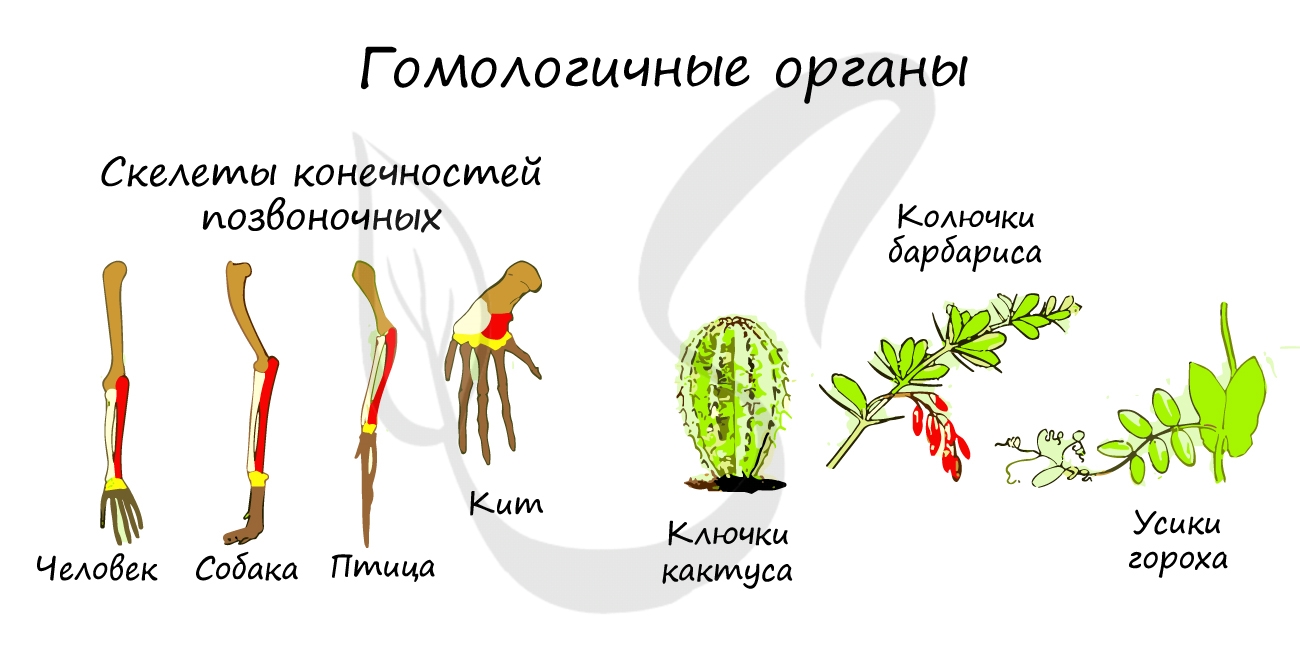
Гомологичные органы (гомология, от греч. homo(s) — равный, одинаковый)
Такие органы развиваются из одних и тех же зародышевых листков, имеют общий план строения, но выполняют разные функции.
Это связано с тем,
что животные освоили разные среды обитания, из-за чего происходит дивергенция (лат. divergo — отклоняюсь) —
расхождение признаков у первоначально близких животных в ходе эволюции.
Гомологичны между собой скелеты конечностей различных классов позвоночных: рука — ласт — крыло птицы, колючки кактуса
— усики гороха — листья растений.
Аналогичные органы (греч. análogos — соответственный)
Аналогичные органы развиваются из разных зародышевых листков, имеют различное строение, но выполняют схожие
функции. Такое сходство возникает в результате приспособления к одним и тем же условиям среды, из-за чего
происходит конвергенция (лат. convergo — сближаю) — схождение признаков у неблизкородственных видов в ходе эволюции.
Аналогичными органами являются крыло птицы — крыло бабочки, глаз человека — глаз кальмара, усики винограда — усики
гороха, жабры рака — жабры рыбы.
В строении нынешних животных можно найти признаки древних предковых форм, которые также свидетельствуют об эволюции. Сейчас
мы обсудим рудименты и атавизмы.
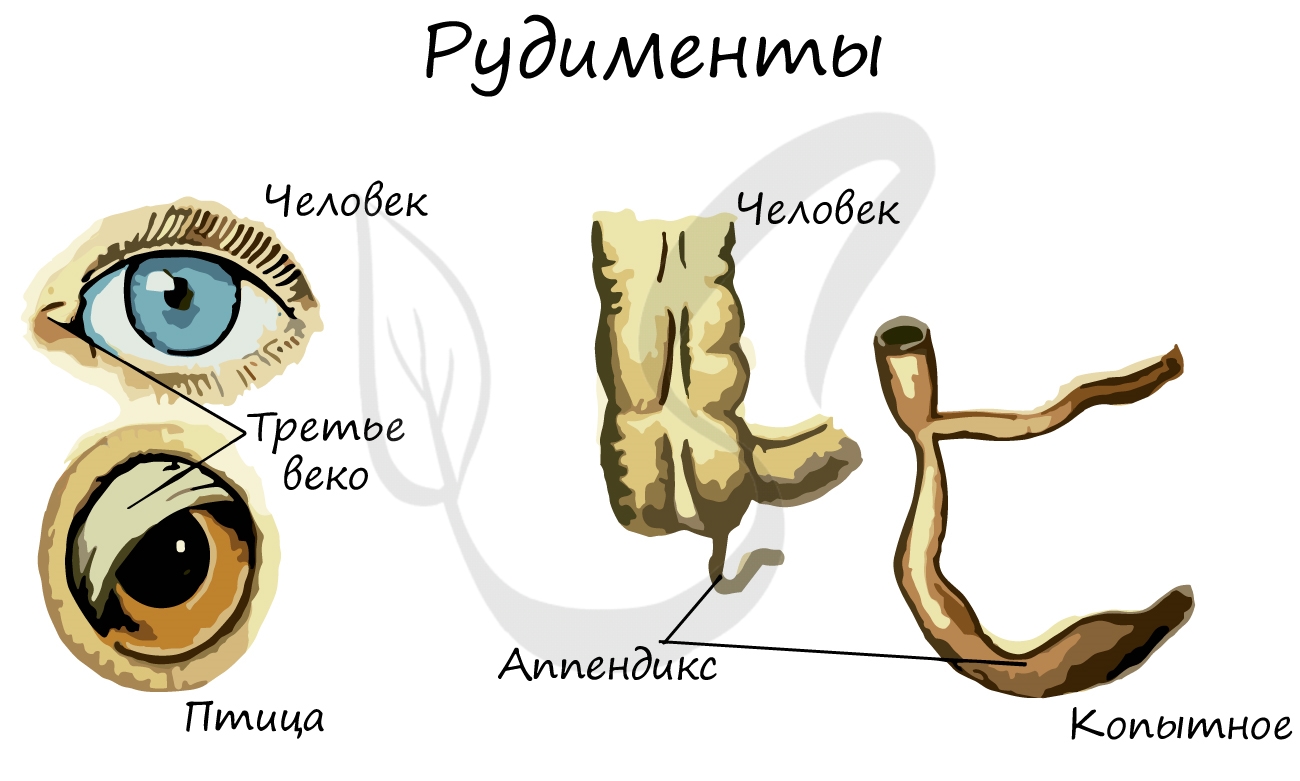
Рудименты (лат. rudimentum — зачаток) — органы, которые в ходе эволюции утратили свое функциональное значение. Они
сохраняются в течение всей жизни и в норме обнаруживаются у человека и животных.
У человека к рудиментарным органам относятся: зубы мудрости, копчик, ушные мышцы, аппендикс (червеобразный отросток),
третье веко (эпикантус).
Атавизмы (лат. atavus — отдалённый предок) — случаи проявления у отдельных особей признаков дальних предков. Атавизмы
сугубо индивидуальны и не являются нормой. Они также являются доказательством эволюции.
У человека атавизмами могут являться хвост, волосатое тело, добавочные молочные железы, незаращение межпредсердной перегородки.
Переходные формы
Переходные формы свидетельствуют о филогенетической преемственности, соединяя в своем строении черты высших и низших классов. Они —
наглядное, живое доказательство эволюции.
Такими формами являются, к примеру, утконос и ехидна из класса млекопитающих. При многих признаках млекопитающих, они откладывают яйца, тем самым подтверждают родство
млекопитающих с пресмыкающимися.
Эмбриологические доказательства
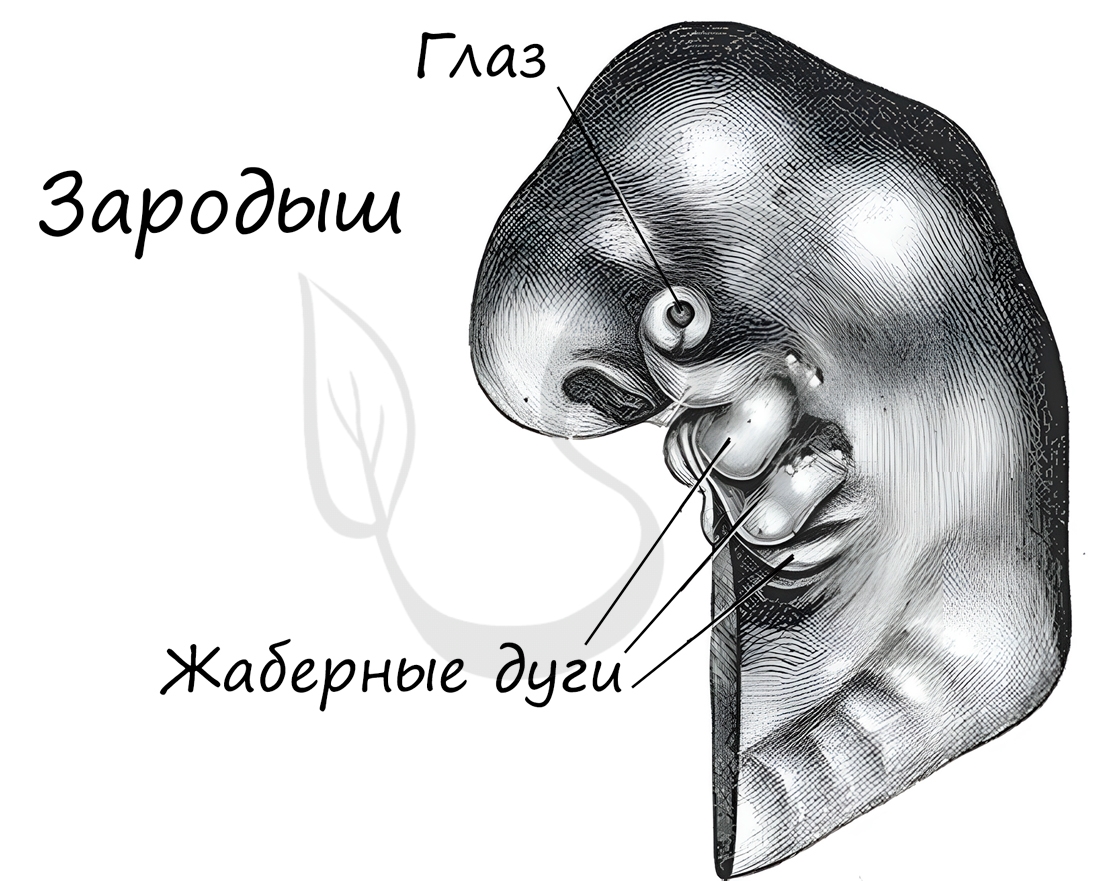
Эмбриология (греч. embryon — зародыш) — раздел биологии, изучающий строение эмбрионов. Только вдумайтесь: на этапе эмбриона,
через который мы с вами успешно прошли, у нас можно было найти закладку жаберных дуг, которые существуют непродолжительное время,
после чего исчезают.
А у рыб, например, жаберные дуги не исчезают — из них развиваются жабры.
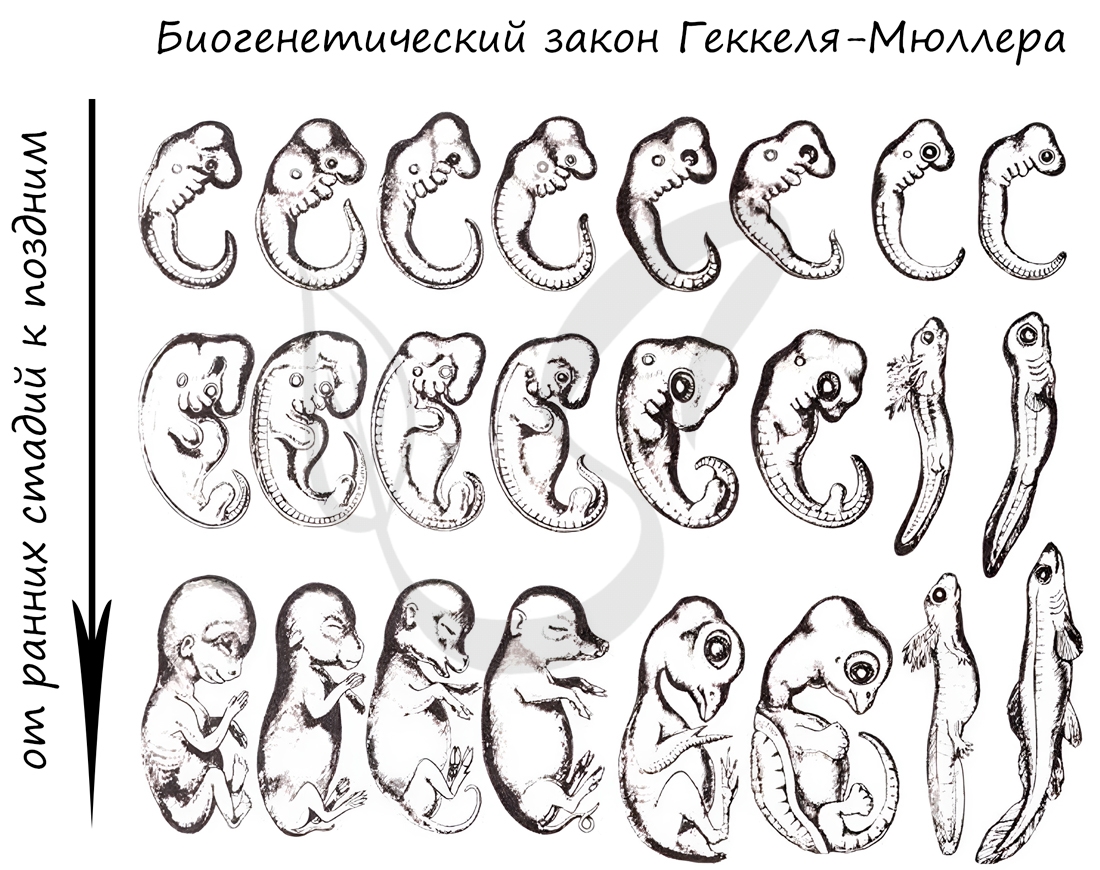
Немецкие ученые Ф. Мюллер и Э. Геккель во второй половине XIX века сформулировали биогенетический закон, гласящий, что
онтогенез (индивидуальное развитие) каждой особи есть краткое и быстрое повторение филогенеза (исторического развития вида).
Биогенетический закон Мюллера-Геккеля объясняет повторение этапов (на стадии зародыша), которые были свойственны нашим далеким
предкам. Таким образом, мы проходим их этапы, но, не останавливаясь на них, двигаемся дальше к более совершенным этапам.
У головастиков лягушек развивается плавник, есть жабры — это наглядное повторение признаков, которые характерны для их предков — рыб.
Карл Бэр сформулировал закон зародышевого сходства, который гласит, что на ранних стадиях развития зародыши позвоночных животных
настолько похожи друг на друга, что практически не различимы между собой. Это также указывает и подтверждает единство происхождения
животного мира.
Палеонтологические доказательства эволюции
Палеонтология (греч. palaios – древний) изучает ископаемые останки вымерших животных, их сходства и различия с ныне живущими
видами. Сопоставляя друг с другом ископаемые останки разных геологических эпох, можно увидеть как происходила эволюция различных
видов животных и растений.
В результате таких исследований иногда удается открыть переходные формы, а иногда — целые филогенетические ряды, то есть совокупность
последовательно сменяющих друг друга форм одного вида. Так, к примеру, был открыт филогенетический ряд лошади.
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение
(в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов
без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования,
обратитесь, пожалуйста, к Беллевичу Юрию.
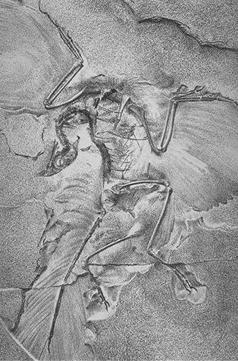
Переходная форма — это… Что такое Переходная форма? Лондонский экземпляр Археоптерикса, переходного звена между рептилиями и птицами.
Переходная форма — организм с промежуточным состоянием, обязательно существующим при состоявшемся постепенном переходе от одного биологического типа строения к другому. Переходные формы характеризуются наличием более древних и примитивных (в значении первичных) черт, чем их более поздние родственники, но, в то же время, наличием более прогрессивных (в значении более поздних) черт, чем их предки. Как правило, говоря о промежуточных формах, имеют в виду ископаемые виды, хотя промежуточные виды вовсе не должны непременно вымирать. Известно множество переходных форм, иллюстрирующих происхождение четвероногих от рыб, рептилий от земноводных, птиц от динозавров, млекопитающих от териодонтов, китообразных от наземных млекопитающих, лошадей от пятипалого предка и человека от древних гоминид.
Теория эволюции
Ко времени написания Чарльзом Дарвином «Происхождения видов» науке не были известны переходные формы жизни. Этот факт Дарвин считал одним из самых слабых мест своей теории. Однако уже через два года был обнаружен археоптерикс — переходное звено между рептилиями и птицами.
Переходные формы между двумя группами организмов не обязательно являются потомками одной группы и предками другой. По ископаемым останкам, как правило, невозможно достоверно установить, является ли один организм прямым предком другого. Кроме того, вероятность найти в палеонтологической летописи чьего-нибудь прямого предка очень мала, и по умолчанию среди биологов принято такую возможность вообще не рассматривать. Гораздо выше вероятность обнаружить относительно близких родственников этого предка, схожих с ним по строению. Поэтому любая переходная форма автоматически интерпретируется как боковая ветвь эволюции, а не «участок ствола».[1]
Примеры
Меритерий, ранний предок слонов, больше похож на свинью, хотя уже имеет маленький хобот и бивни, свойственные хоботным.
В качестве примеров переходных форм можно привести Амбулоцетуса — «ходячего кита» (переходная форма между китообразными и наземными млекопитающими), Тиктаалика и Ихтиостегу (переходная форма от рыб к земноводным), Мезогиппуса (между ранними лошадиными и современной лошадью).
Переходные формы в эволюции Человека Разумного
Черепа гоминид, от шимпанзе (A) до человека разумного (N). A — Шимпанзе, B,C — Австралопитек африканский, D,E — Человек умелый , F — Человек рудольфский, G — Человек прямоходящий, H — Человек работающий , I — Гейдельбергский человек, J,K,L — Неандерталец, M — Кроманьонец, N — Человек разумный
В настоящее время известно множество переходных форм между человеком разумным и его обезьяноподобными предками. Так, например, открыты останки Сахелантропа, Ардипитека, Австралопитеков (африканского, афарского и других), Человека умелого, Человека работающего, Человека прямоходящего, Человека-предшественника, Гейдельбергского человека, Неандертальца и раннего Человека разумного.
Спор о постепенности эволюционных преобразований
Переходные формы являются объектом исследований в споре между градуалистами и пунктуалистами. Градуалисты считают, что эволюция является постепенным, непрерывным процессом. Пунктуалисты, или сторонники теории прерывистого равновесия считают, что виды в течение долгого времени остаются неизменными, а изменения происходят в очень короткий промежуток времени. Переходные формы являются аргументом обеих сторон. Пунктуалисты утверждают, что большинство видов появляются внезапно и на протяжении всей своей истории меняются слабо. Градуалисты отмечают, что среди позвоночных нет ни одного бесспорного примера очень быстрого перехода от одного вида к другому, большинство переходов вообще плохо задокументированы ископаемыми. Также много хорошо описанных внутривидовых изменений. Например, описаны изменения среднекайнозойских кроликов, миоценовых грызунов, гиппарионов, эоценовых парнокопытных, переход между родами эоценовых приматов.[2]
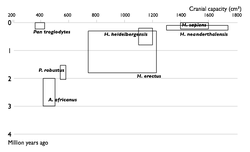
Недостающие звенья
1850
1900
1950
2002
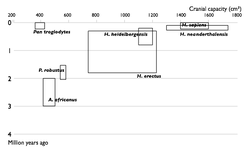
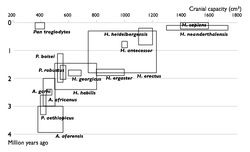
Прогресс в изучении эволюции человека. Увеличение числа известных науке видов гоминин по времени. Каждый вид изображен в виде прямоугольника, показывающего границы, в которых варьировал объём черепа, и место вида в палеонтологической летописи. Видно постепенное заполнение промежутков между видами.
Многие переходные формы между живыми организмами пока не найдены. Обычно их называют недостающими звеньями. Редкость переходных форм объясняется неполнотой палеонтологической летописи. Сторонники теории прерывистого равновесия также указывают на быстроту эволюционных преобразований. Однако палеонтологи постоянно находят новые ископаемые, заполняя пробелы (например, в 2004 году был найден Тиктаалик — ещё одно переходное звено между рыбами и земноводными)
Неполнота палеонтологической летописи
Далеко не каждая переходная форма существует в виде окаменелости из-за принципиальной неполноты палеонтологической летописи. Неполнота вызвана особенностями процесса фоссилизации, то есть перехода в окаменелое состояние. Для образования окаменелости необходимо, чтобы погибший организм был погребён под большим слоем осадочных пород. Из-за крайне медленной скорости осадконакопления на суше, чисто сухопутные виды редко переходят в окаменелое состояние и сохраняются. Кроме того, редко удаётся обнаружить виды, живущие в глубинах океана, из-за редкости поднятия на поверхность больших массивов дна. Таким образом, большинство известных ископаемых (а, соответственно, и переходных форм) — это либо виды, обитающие на мелководье, в морях и реках, либо наземные виды, ведущие полуводный образ жизни, либо обитающие недалеко от береговой линии. Закономерности процессов захоронения останков живых существ изучает особая отрасль палеонтологии — тафономия.
Филогенетические ряды
Филогенетический ряд лошади. Показана реконструкция ископаемых видов, полученных из последовательных слоёв породы. Справа — вид спереди передней левой ноги с выделенным третьим сегментом, а также зуб в продольном сечении[3].
Филогенетические ряды − ряды видов, последовательно сменявших друг друга в процессе эволюции различных групп животных и растений.
Впервые были открыты В. О. Ковалевским, который показал, что современные однопалые копытные происходят от древних пятипалых мелких всеядных животных.
Креационизм
Сторонники креационизма заявляют, что переходных форм не обнаружено. Научное сообщество считает подобные заявления ложью и намеренным введением в заблуждение.
См. также
Примечания
Литература
Дарвин, Чарльз. Происхождение видов путём естественного отбора: В 2 кн. — М.: ТЕРРА — Книжный клуб, 2009. — ISBN 978-5-275-02114-1
Кэрролл Р. Палеонтология и эволюция позвоночных: В 3-х т. — Мир, 1992. — 280 с. — ISBN 5-03-001819-0
Ссылки
Переходные формы — ТворенияВики, энциклопедия науки о сотворении
Переходные формы — формы, соединяющие в себе признаки двух соседних таксономических групп или единиц. Название является эволюционным термином, подразумевающим эволюцию как данность. Также используется близкий термин — промежуточные формы.
Гипотеза о существовании бесчисленного множества переходных форм была высказана ещё Чарльзом Дарвином в книге «Происхождение видов», однако до сих пор их наличие не бесспорно. Отсутствие переходных форм объясняется приверженцами теории эволюции тем, что громадное большинство форм, уже изолировалось вследствие вымирания соседних. Младоземельные креационисты считают, что значительная разница между живыми организмами и отсутствие явных переходных форм в летописи окаменелостей подтверждает теорию Сотворения.
В креационной биологии рассматриваются сотворённые роды живых организмов, которые имеют чёткие границы видовой изменчивости, заложенные в геноме. Появление принципиально новых видов животных, способных к выживанию и т. н. «победе в эволюционной борьбе» невозможно, так как единственный возможный вариант — мутации — всегда дегенеративны.
От одноклеточных к многоклеточным
Переходные формы не найдены. Многоклеточные в летописи окаменелостей появляются одномоментно, в большом разнообразии семейств и отрядов. Этот феномен, противоречащий всей теории эволюции, получил в среде эволюционистов наименование «Кембрийский взрыв».
От беспозвоночных к позвоночным
Ланцетник (фото с сайта comenius.susqu.edu))
Очередная огромная загадка эволюционной теории — это переход от экзоскелета (оболочка ракушек или хитиновый панцирь насекомых) к эндоскелету (внутреннему скелету). Нет ни найденных переходных форм, ни внятных и адекватных гипотез на этот счёт.
Следует четко представлять себе, какие колоссальные преобразования должны были бы произойти в строении животного, чтобы полностью преобразовались все системы организма.
В учебниках в качестве предка позвоночных животных до сих пор предлагаются ланцетники Branchiostoma, однако расшифровка их генома, законченная в 2008 году, показала, что ланцетники являются более дальними «родственниками» позвоночных, чем оболочники (прикреплённые ко дну организмы). В настоящее время учёные-эволюционисты рассматривают эту форму как побочную ветвь в эволюции позвоночных животных, происхождение которых, в свою очередь, остается загадочным[1]
От рыбы к амфибии
От амфибии к рептилии
От рептилии к птице
Эволюционное развитие рептилий (например динозавров) до птиц невозможно представить по следующим причинам:
Птицы в отличии от рептилий, являются теплокровными существами, что подразумевает, как минимум, иное строение кровеносной системы.
Живая птица обладает достаточным уровнем метаболизма, чтобы поддерживать температуру тела на постоянном уровне за счёт самостоятельного производства энергии из потребляемой пищи. Современные птицы относятся к истинно-гомойотермным существам. Помимо достаточных энергетических возможностей они имеют также различные механизмы, предназначенные для удержания тепла (перья, подкожный слой жировой ткани) и для защиты от перегрева при высокой температуре окружающей среды (потоотделение). Организм рептилий не обладает возможностью сохранять тепло. Инерциально-гомойотермное существо в периоды повышения температуры медленно нагревается, а в периоды похолодания — медленно остывает, то есть за счёт большой теплоёмкости колебания температуры организма сглаживаются. Так называемая инерциальная гомойотермия была характерна, по современным представлениям, для некоторых видов динозавров. Недостатком инерциальной гомойотермии является то, что она возможна только при определённом типе климата — когда средняя температура окружающей среды соответствует желаемой температуре тела и нет длительных периодов сильных похолоданий или потеплений.
Дыхательная система рептилий и птиц весьма различна. Птичье легкое является очень сложным и структурным в отличие от любого другого организма. Органы дыхания пернатых считается неприводимо сложной системой, в которой каждая деталь должна функционировать должным образом для того, чтобы работать.
Перья птиц и чешуя рептилий совершенно различны в устройстве, методах крепления на теле живого организма и внешнем виде
Различное число пальцев в конечностях динозавров и птиц. Превращаясь в птицу, динозавры потеряли бы один вид пальца, а затем должны были восстановить другой
Никаких остатков переходных форм между рептилиями, земноводными и птицами найдено не было.
От рептилии к млекоптающим
От сумчатых к плацентарным
От обезьяны к человеку
Первобытные люди — по теории эволюции, предки современных людей, отличающиеся по строению тела (ближе к обезъянам), с низким или отсутствующим уровнем культуры и языка и т. п.
Существование первобытных людей как переходной формы от «обезьяноподобного предка» до современного вида людей является неподтверждённой гипотезой, так как до сих пор не найдены соответствующие останки.
Креационисткий взгляд на происхождение людей, базируется на следующем положении Библии: «И сотворил Бог человека по образу Своему, по образу Божию сотворил его; мужчину и женщину сотворил их» (Быт.1:26-28 )
В Библии можно прочитать, что человек изначально был создан разумным. Первое поколение людей было способным к производству орудий труда для ремесла и сельского хозяйства, строительства, могло добывать огонь. Человек интересовался музыкой, а и как следствие обладал способностью создания музыкальных инструментов (см. 4 главу книги Бытие).
Эволюция лошади
Эволюция китообразных
Мозаичные формы
Живые ископаемые
Под живыми ископаемыми понимаются живые организмы, первоначально найденные только в виде окаменевших останков, а затем обнаруженные живыми и не изменившимися в течении предполагаемых эпох эволюции.
Среди сторонников эволюционной теории существует предубеждение, что живые ископаемые встречаются редко, так как в течении «миллионов лет», эти организмы эволюционировали в другие «более совершенные» формы жизни.
Однако современные фактические данные свидетельствуют о большом разнообразии живых организмов, которые могут быть отнесены к категории живых ископаемых. Одним из ярчайший примеров является несколько нынеживущих видов кистепёрой рыбы.
Совершенство ископаемых форм
Примечания
См. также
Палитра знаний: Ископаемые переходные формы
Ископаемые переходные формы — это формы организмов, сочетающие признаки более древних и молодых групп организмов.
Семенные папоротники сочетали признаки папоротников и семенных растений (семена, сидящие на безлистых осях или листоподобных органах, имели мегаспорангий — семязачаток; прототип стробилы; эустелу; окаймленные поры на трахеидах).
Ихтиостеги — рыболягушки, подотряд вымерших земноводных (лабиринтодонтов), представляющие собой переходную форму между кистеперыми рыбами и амфибиями. Известны из верхнедевонских отложений Гренландии и Австралии. Ихтиостеги имели такие признаки рыб:
Тело обтекаемой формы.
Имелся рудиментарный хвостовой плавник.
Органы боковой линии на черепе, как у рыб, заключены в костные каналы.
Сходное строение позвонков.
В черепе сохранились рудиментарные кости жаберных крышек.
Ноздри, как у двоякодышаших рыб, смещены к краю рта.
Признаки амфибий: имеются плечевой, тазовый пояса и грудная клетка, в конечностях имеются сходные кости.
Зверообразные рептилии — звероящеры, вымершие рептилии (синапсиды, тероморфы), сочетали признаки рептилий и зверей. Они были широко распространены в перми и триасе. Обособились от котилозавров в карбоне. В верхнем карбоне и перми преобладали примитивные звероящеры (пеликозавры). В верхней перми — триасе — преобладали терапсиды.
Звероящеры имели такие признаки млекопитающих:
одна височная ямка, ограниченная снизу скуловой дугой
нёбо вторичное костное
клыки хорошо развиты
нёбные зубы
развита большая зубная кость
нижняя челюсть подвешена непосредственно к черепу, а не при помощи квадратной кости, как у рептилий.
Это были хищники. Их потомки — териодонты. Обнаружены в Европе, Северной Америке, Африке.
Археоптерикс — промежуточная форма между птицами и рептилиями. Имела такие признаки птиц: тело покрыто перьями; маховые крылья крупные, асимметричные; кости конечностей частично полые; передние конечности — крылья, но с тремя пальцами.
Переходная форма — Википедия. Что такое Переходная форма Лондонский экземпляр археоптерикса, переходного звена между рептилиями и птицами.
Переходная форма — организм с промежуточным состоянием, обязательно существующим при состоявшемся постепенном переходе от одного биологического типа строения к другому. Переходные формы характеризуются наличием более древних и примитивных (в значении первичных) черт, чем их более поздние родственники, но, в то же время, наличием более прогрессивных (в значении более поздних) черт, чем их предки. Как правило, говоря о промежуточных формах, имеют в виду ископаемые виды, хотя промежуточные виды вовсе не должны непременно вымирать. Известно множество переходных форм, иллюстрирующих происхождение четвероногих от рыб, рептилий от земноводных, птиц от динозавров, млекопитающих от териодонтов, китообразных от наземных млекопитающих, лошадей от пятипалого предка и человека от древних гоминид[1].
Теория эволюции
Ко времени написания Чарльзом Дарвином «Происхождения видов» науке не были известны переходные формы жизни. Этот факт Дарвин считал одним из самых слабых мест своей теории. Однако уже через два года был обнаружен археоптерикс — переходное звено между рептилиями и птицами.
Переходные формы между двумя группами организмов не обязательно являются потомками одной группы и предками другой. По ископаемым останкам, как правило, невозможно достоверно установить, является ли один организм прямым предком другого. Кроме того, вероятность найти в палеонтологической летописи чьего-нибудь прямого предка очень мала, и по умолчанию среди биологов принято такую возможность вообще не рассматривать. Гораздо выше вероятность обнаружить относительно близких родственников этого предка, схожих с ним по строению. Поэтому любая переходная форма автоматически интерпретируется как боковая ветвь эволюции, а не «участок ствола».[2]
Примеры
Меритерий, ранний предок слонов, больше похож на свинью, хотя уже имеет маленький хобот и бивни, свойственные хоботным.
В качестве примеров переходных форм можно привести амбулоцетуса — «ходячего кита» (переходная форма между наземными млекопитающими и китообразными), тиктаалика и ихтиостегу (переходные формы от рыб к земноводным), мезогиппуса (между ранними лошадиными и современной лошадью).
Переходные формы в эволюции человека
В настоящее время известно множество переходных форм между человеком разумным и его обезьяноподобными предками. Так, например, открыты останки сахелантропа, ардипитека, австралопитеков (африканского, афарского и других), человека умелого, человека работающего, человека прямоходящего, человека-предшественника, гейдельбергского человека, неандертальца и раннего человека разумного.
Спор о постепенности эволюционных преобразований
Переходные формы являются объектом исследований в споре между градуалистами и пунктуалистами. Градуалисты считают, что эволюция является постепенным, непрерывным процессом. Пунктуалисты, или сторонники теории прерывистого равновесия считают, что виды в течение долгого времени остаются неизменными, а изменения происходят в очень короткий промежуток времени. Переходные формы являются аргументом обеих сторон. Пунктуалисты утверждают, что большинство видов появляются внезапно и на протяжении всей своей истории меняются слабо. Градуалисты отмечают, что среди позвоночных нет ни одного бесспорного примера очень быстрого перехода от одного вида к другому, большинство переходов вообще плохо задокументированы ископаемыми. Также много хорошо описанных внутривидовых изменений. Например, описаны изменения среднекайнозойских кроликов, миоценовых грызунов, гиппарионов, эоценовых парнокопытных, переход между родами эоценовых приматов.[3]
Недостающие звенья
1850
1900
1950
2002
Прогресс в изучении эволюции человека. Увеличение числа известных науке видов гоминин по времени. Каждый вид изображен в виде прямоугольника, показывающего границы, в которых варьировал объём черепа, и место вида в палеонтологической летописи. Видно постепенное заполнение промежутков между видами.
Многие переходные формы между живыми организмами пока не найдены. Обычно их называют недостающими звеньями. Редкость переходных форм объясняется неполнотой палеонтологической летописи. Сторонники теории прерывистого равновесия также указывают на быстроту эволюционных преобразований. Однако палеонтологи постоянно находят новые ископаемые, заполняя пробелы (например, в 2004 году был найден тиктаалик — ещё одно переходное звено между рыбами и земноводными).
Неполнота палеонтологической летописи
Далеко не каждая переходная форма существует в виде окаменелости из-за принципиальной неполноты палеонтологической летописи. Неполнота вызвана особенностями процесса фоссилизации, то есть перехода в окаменелое состояние. Для образования окаменелости необходимо, чтобы погибший организм был погребён под большим слоем осадочных пород. Из-за крайне медленной скорости осадконакопления на суше, чисто сухопутные виды редко переходят в окаменелое состояние и сохраняются. Кроме того, редко удаётся обнаружить виды, живущие в глубинах океана, из-за редкости поднятия на поверхность больших массивов дна. Таким образом, большинство известных ископаемых (а, соответственно, и переходных форм) — это либо виды, обитающие на мелководье, в морях и реках, либо наземные виды, ведущие полуводный образ жизни, либо обитающие недалеко от береговой линии. Закономерности процессов захоронения останков живых существ изучает особая отрасль палеонтологии — тафономия.
Филогенетические ряды
Филогенетический ряд лошади. Показана реконструкция ископаемых видов, полученных из последовательных слоёв породы. Справа — вид спереди передней левой ноги с выделенным третьим сегментом, а также зуб в продольном сечении[4].
Филогенетические ряды − ряды видов, последовательно сменявших друг друга в процессе эволюции различных групп животных и растений.
Впервые были открыты В. О. Ковалевским, который показал, что современные однопалые копытные происходят от древних пятипалых мелких всеядных животных.
Креационизм
Сторонники креационизма заявляют, что переходных форм не обнаружено. Научное сообщество считает подобные заявления ложью и намеренным введением в заблуждение.
См. также
Примечания
Литература
Дарвин, Чарльз. Происхождение видов путём естественного отбора: В 2 кн. — М.: ТЕРРА — Книжный клуб, 2009. — ISBN 978-5-275-02114-1.
Кэрролл Р. Палеонтология и эволюция позвоночных: В 3-х т. — Мир, 1992. — 280 с. — ISBN 5-03-001819-0.



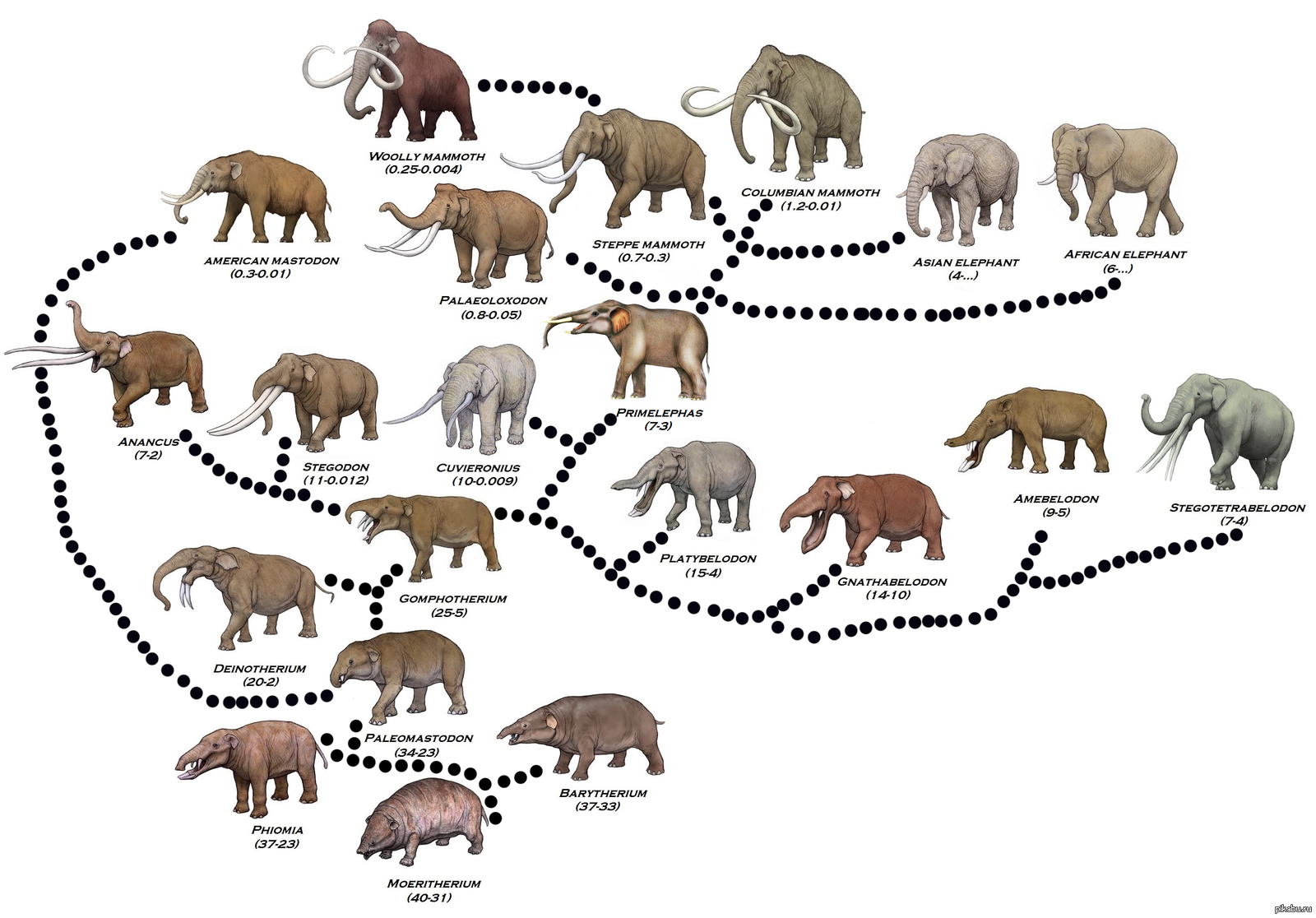











 Меритерий, ранний предок слонов, больше похож на свинью, хотя уже имеет маленький хобот и бивни, свойственные хоботным.
Меритерий, ранний предок слонов, больше похож на свинью, хотя уже имеет маленький хобот и бивни, свойственные хоботным.


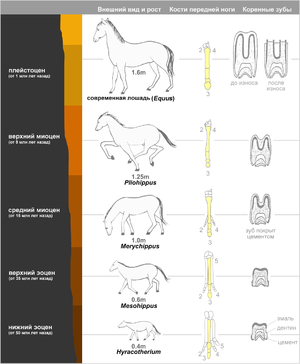 Филогенетический ряд лошади. Показана реконструкция ископаемых видов, полученных из последовательных слоёв породы. Справа — вид спереди передней левой ноги с выделенным третьим сегментом, а также зуб в продольном сечении[3].
Филогенетический ряд лошади. Показана реконструкция ископаемых видов, полученных из последовательных слоёв породы. Справа — вид спереди передней левой ноги с выделенным третьим сегментом, а также зуб в продольном сечении[3].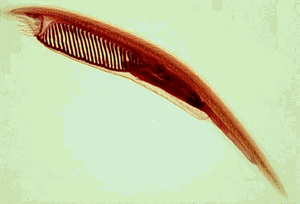 Ланцетник (фото с сайта comenius.susqu.edu))
Ланцетник (фото с сайта comenius.susqu.edu)) Меритерий, ранний предок слонов, больше похож на свинью, хотя уже имеет маленький хобот и бивни, свойственные хоботным.
Меритерий, ранний предок слонов, больше похож на свинью, хотя уже имеет маленький хобот и бивни, свойственные хоботным.

 Филогенетический ряд лошади. Показана реконструкция ископаемых видов, полученных из последовательных слоёв породы. Справа — вид спереди передней левой ноги с выделенным третьим сегментом, а также зуб в продольном сечении[4].
Филогенетический ряд лошади. Показана реконструкция ископаемых видов, полученных из последовательных слоёв породы. Справа — вид спереди передней левой ноги с выделенным третьим сегментом, а также зуб в продольном сечении[4].
Leave A Comment