
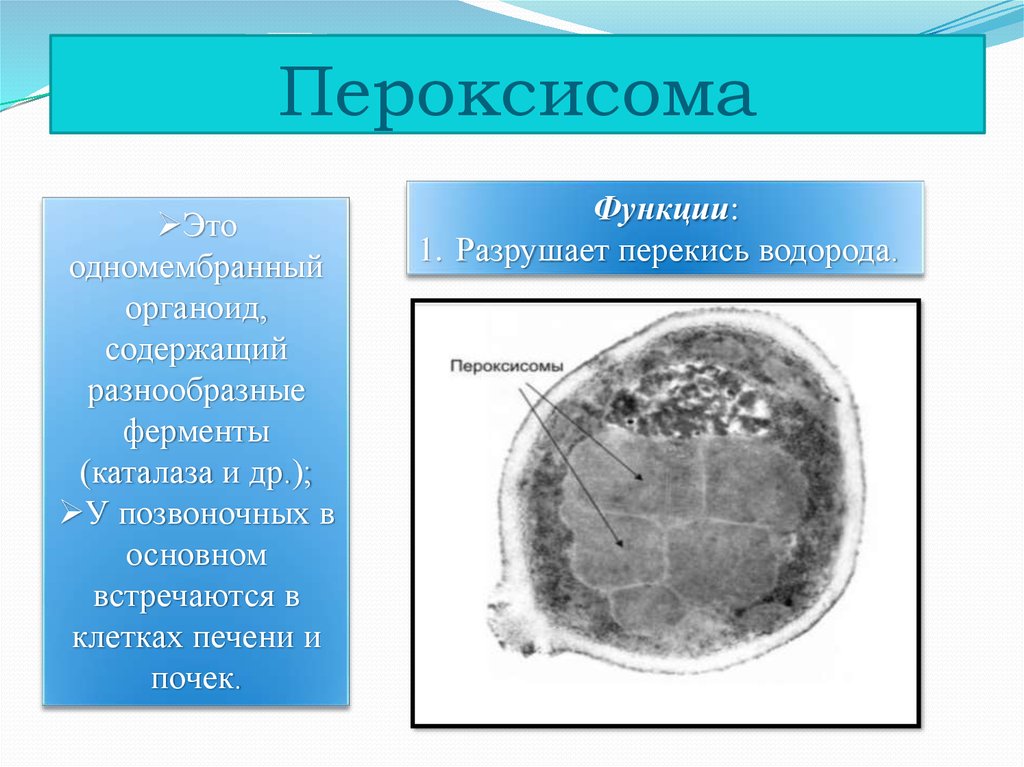
8. Пероксисомы. Строение и функции.
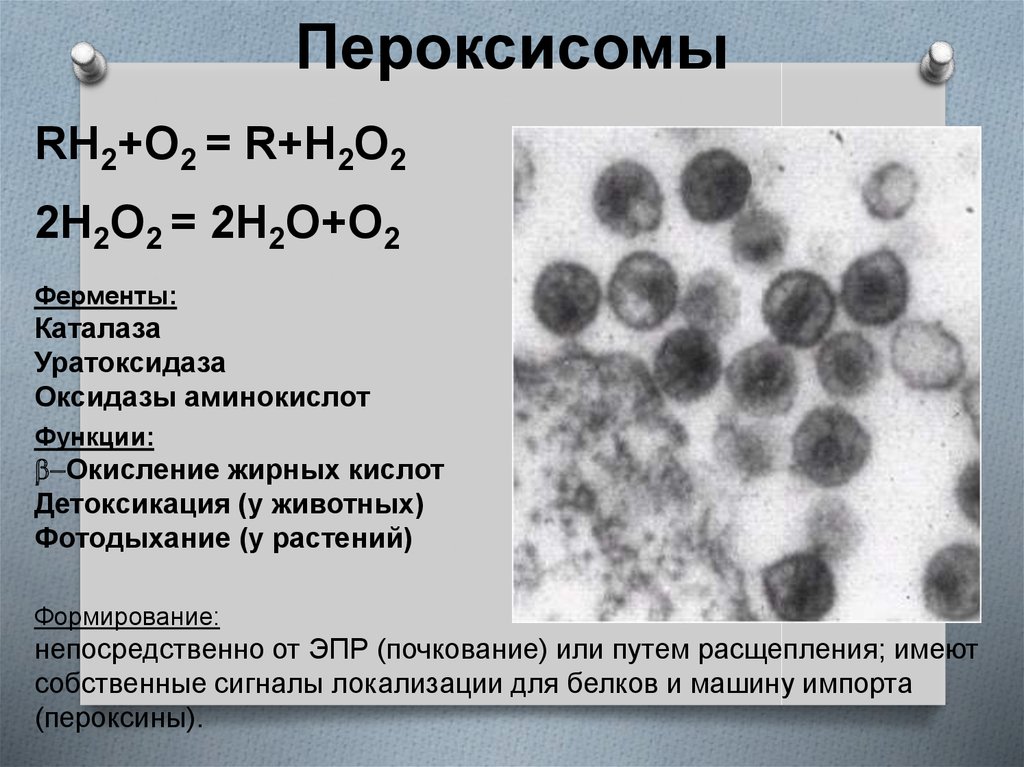
Пероксисомы (микротельца) по строению сходны с лизосомами. Они состоят из матирикса и нуклеотида. Матрикс пероксисом содержит до 15 ферментов. Наиболее важные из них пероксидаза и каталаза, оксидаза D-аминокислот и уратроксидаза. Нуклеотид пероксисомы соответствует области конденсации ферментов. Пероксисомы образуются в ЭПС, отпочковываясь от агранулярной ЭПС, их ферменты частично синтезируются в гранулярной ЭПС, частично в гиалоплазме. Мембрана пероксисом непроницаема для ионов и низкомолекулярных субстратов.
Пероксисомы – главный центр образования кислорода клетки. В результате окисления аминокислот, углеводов образуется Н2О2, которая благодаря каталазе распадается на воду и О2. Крупные пероксисомы печени и почек играют важную роль в обезвреживании ряда веществ. Помимо этого они участвуют в катаболизме (в обмене аминокислот, оксалата и полиаминов).
В
настоящее время открыт класс наследственных
болезней – пероксисомные болезни,
развитие которых обусловлекнно дефектом
пероксисом.
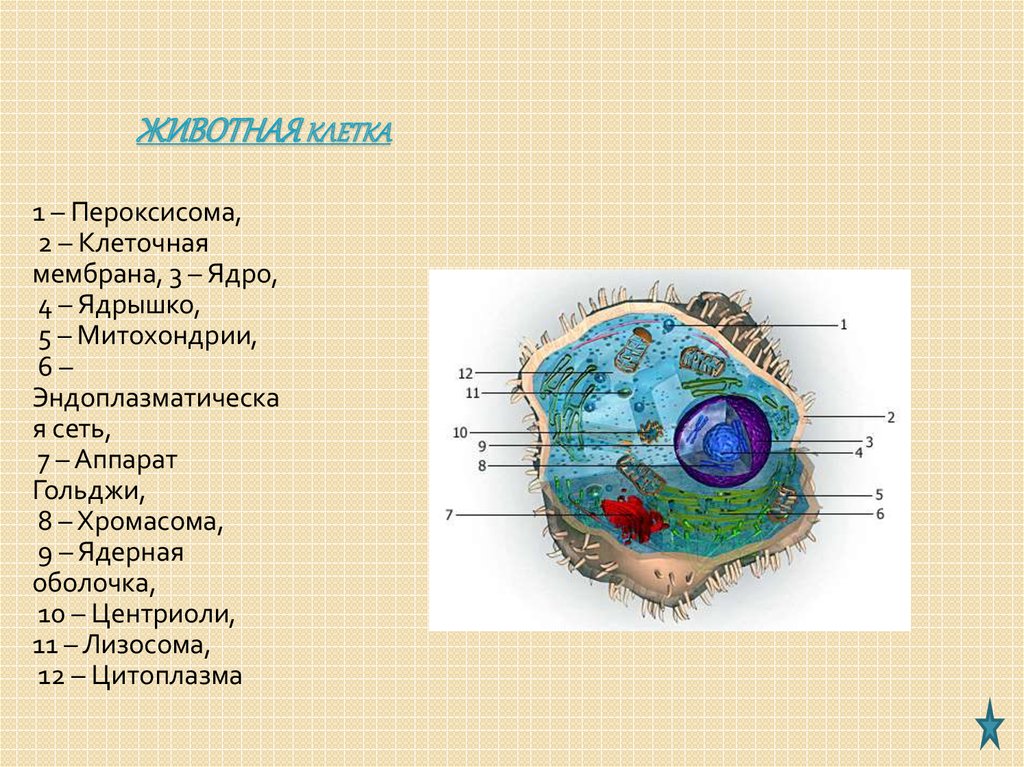
Митохондрии являются универсальным мембранными органоидами клеток. Митохондрии имеют 2 мембраны – наружную и внутреннюю. Между этими мембранами нахлдится межмембранное пространство. В некоторых участках мембраны образуют контактные сайты. В митохондрии находится митохондриальный матрикс. В нем локализуются молекулы митохондриальной ДНК, собственные рибосомы, РНК, белки, низкомолекулярные метаболиты.
В
наружной мембране содержится более 80%
липидов и менее 20% белков, а во внутренней
– наоборот. Среди белков наружной
мембраны имеются порины, формирующие
поры. Через них из гиалоплазмы поступают
молекулы определенного размера. В
результате этого наружная мембрана
имеет неспецифическую проницаемость.
В зоне контактных сайтов локализуются
специальные рецепторы и канальные
белки. Внутренняя мембрана образует
кристы. На них со стороны митохондриального
матрикса локализуются грибовидные
тельца – белковые компоненты, которые
осуществляют синтез АТФ.
Симптомы
большинства митохондриальных болезней
проявляются с возрастом, что вероятно,
обусловлено накоплением мутаций,
осуществляемыми Н
Жизненный цикл митохондрий около 10 суток, их разрушение происходит путем аутофагии, а гибнущие органеллы замещаются новыми, которые формируются путем пеершнуровки предшествующих. Репликация митохондриальной ДНК происходит в любые фазы клеточного цикла независимо от ядерной ДНК.
Функции митохондрий:
Дыхательный и энергетический центр клетки – в них усваивается кислород необходимый для третьего (аэробного) этапа диссимиляции.
Синтез своих ДНК, РНК, части белков.
Строение и образование пероксисом — презентация онлайн
Похожие презентации:
Эндокринная система
Анатомо — физиологические особенности сердечно — сосудистой системы детей
Хронический панкреатит
Топографическая анатомия верхних конечностей
Анатомия и физиология сердца
Мышцы головы и шеи
Эхинококкоз человека
Черепно-мозговые нервы
Анатомия и физиология печени
Топографическая анатомия и оперативная хирургия таза и промежности
1.
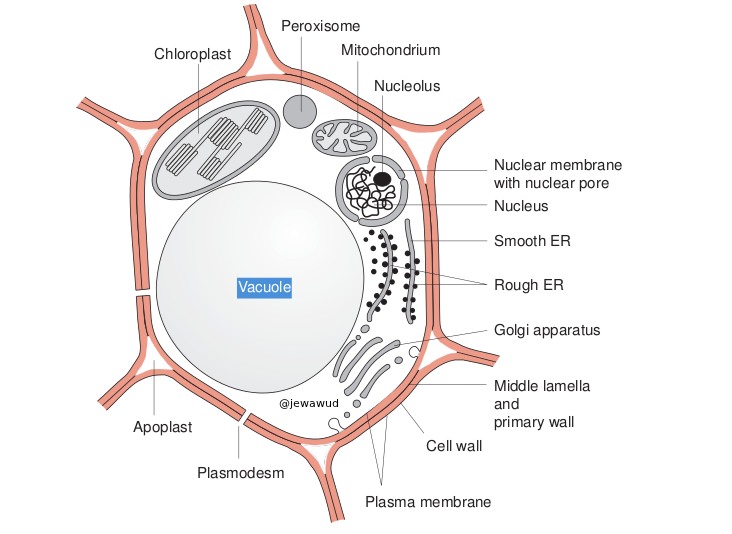 Строение и функции пероксисом
Строение и функции пероксисом2. Открытие пероксисом
Открытыв 1965 году
Кристианом де Дювом бельгийским цитологом,
биохимиком, лауреатом
Нобелевской премии по
физиологии и медицине
в 1974 году «за
открытия, касающиеся
структурной и
функциональной
организации клетки».
3. Строение пероксисом
Пероксисома — обязательная органеллаэукариотической клетки, ограниченная мембраной,
содержащая по разным источникам от 15 до 50
различных ферментов, катализирующих
окислительно-восстановительные реакции. Имеет
размер от 0,2 до 1,5 мкм.
Набор функций пероксисом различается в клетках
разных типов. Среди них: окисление жирных кислот,
фотодыхание, разрушение токсичных соединений,
синтез желчных кислот, холестерина, а также
эфиросодержащих липидов, построение миелиновой
оболочки нервных волокон, метаболизм фитановой
кислоты и т. д. Наряду с митохондриями являются
главными потребителями O2 в клетке.

5. Образование пероксисом
Длительность жизниорганоида — 5-6 суток.
Новые пероксисомы
образуются чаще всего в
результате деления
предшествующих, как
митохондрии и хлоропласты,
но не содержат ДНК и рибосом,
поэтому предположение об их
эндосимбиотическом
происхождении не может быть
верно. Также могут
формироваться из ЭПР.
6. Ферментный состав и функции
1. Пероксисомы получили такое названиеблагодаря тому, что обычно в их состав входит
один или несколько ферментов,
использующих молекулярный кислород для
отщепления атомов водорода от некоторых
органических субстратов в окислительной
реакции с образованием пероксида водорода:
7. Ферментный состав и функции
2. Использует образующийся пероксиддля окисления множества субстратов (фенолов,
муравьиной кислоты, формальдегида,этанола)
фермент каталаза:
Этот тип окислительных реакций особенно важен
в клетках печени и почек , пероксисомы
которых обезвреживают множество ядовитых
веществ , попадающих в кровоток.
 Почти
Почтиполовина этанола , попадающего в организм,
окисляется этим способом.
8. Ферментный состав и функции
В пероксисомах происходит окислениедлинноцепочечных жирных кислот с образованием
повторного использования в метаболических реакциях
(является важным звеном в цикле трикарбоновых
кислот). Известно, что жирные кислоты длиной более чем
22 углеродных атома окисляются исключительно в этом
органоиде.
В результате дефекта функционирования пероксисом
происходит накопление этих кислот в клетках организма.
У пациентов с пероксисомными болезнями биохимические
нарушения главным образом заключаются в дефектах α- и
β-окисления жирных кислот и синтеза
плазмалогенов (структурных фосфолипидов миелиновых
волокон)
English Русский Правила
Образование и поддержание пероксисом зависят от эндоплазматического ретикулума
Обзор
.
doi: 10.1146/annurev-biochem-081111-125123. Epub 2013, 14 февраля.
Хенк Ф Табак 1 , Инеке Браакман, Адабелла ван дер Занд
принадлежность
- 1 Секция химии клеточных белков факультета естественных наук Утрехтского университета, NL-3584 CH Утрехт, Нидерланды.
- PMID: 23414306
- DOI: 10.1146/annurev-biochem-081111-125123
Обзор
Henk F Tabak et al.
Анну Рев Биохим.
2013.
. 2013;82:723-44.
doi: 10.1146/annurev-biochem-081111-125123. Epub 2013, 14 февраля.
Авторы
Хенк Ф Табак 1 , Инеке Браакман, Адабелла ван дер Занд
принадлежность
- 1 Секция химии клеточных белков факультета естественных наук Утрехтского университета, NL-3584 CH Утрехт, Нидерланды.
- PMID: 23414306
- DOI: 10.1146/annurev-biochem-081111-125123
Абстрактный
Внешность может быть обманчива.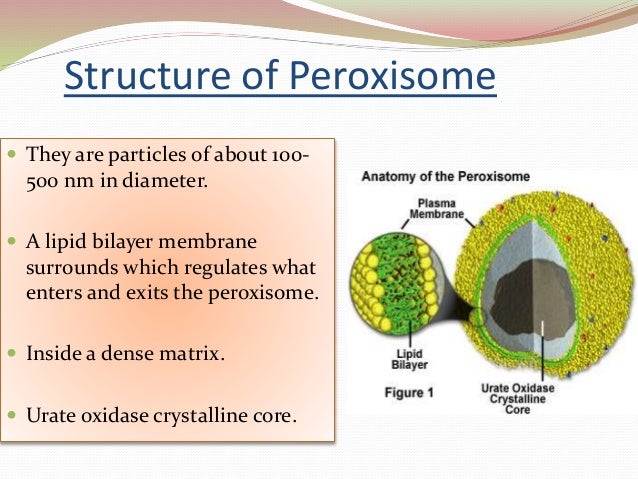 Хотя пероксисомы кажутся простыми органеллами, их формирование и поддержание представляют собой уникальные проблемы для клетки. Рождение новых пероксисом начинается в эндоплазматическом ретикулуме (ЭР), который доставляет липиды и мембранные белки. Чтобы сформировать новый пероксисомальный компартмент, препероксисомальные везикулы, происходящие из ER, несущие разные мембранные белки, сливаются, что позволяет собрать пероксисомальный транслокон. Для завершения формирования пероксисомы импортируют свои растворимые белки непосредственно из цитозоля, используя только что собранный транслокон. Вместе с биогенным путем, происходящим из ER, деление пероксисом и сегрегация впоследствии поддерживают клеточную популяцию пероксисом. В этом обзоре мы освещаем последние данные о жизненном цикле пероксисом и показываем, как новая концепция клеточной биологии образования пероксисом влияет на наши представления о заболеваниях, связанных с пероксисомами, и их эволюционном прошлом. Будущая задача заключается в идентификации всех белков, участвующих в этом сложном биогенном процессе, и в анализе механизма их действия.
Хотя пероксисомы кажутся простыми органеллами, их формирование и поддержание представляют собой уникальные проблемы для клетки. Рождение новых пероксисом начинается в эндоплазматическом ретикулуме (ЭР), который доставляет липиды и мембранные белки. Чтобы сформировать новый пероксисомальный компартмент, препероксисомальные везикулы, происходящие из ER, несущие разные мембранные белки, сливаются, что позволяет собрать пероксисомальный транслокон. Для завершения формирования пероксисомы импортируют свои растворимые белки непосредственно из цитозоля, используя только что собранный транслокон. Вместе с биогенным путем, происходящим из ER, деление пероксисом и сегрегация впоследствии поддерживают клеточную популяцию пероксисом. В этом обзоре мы освещаем последние данные о жизненном цикле пероксисом и показываем, как новая концепция клеточной биологии образования пероксисом влияет на наши представления о заболеваниях, связанных с пероксисомами, и их эволюционном прошлом. Будущая задача заключается в идентификации всех белков, участвующих в этом сложном биогенном процессе, и в анализе механизма их действия.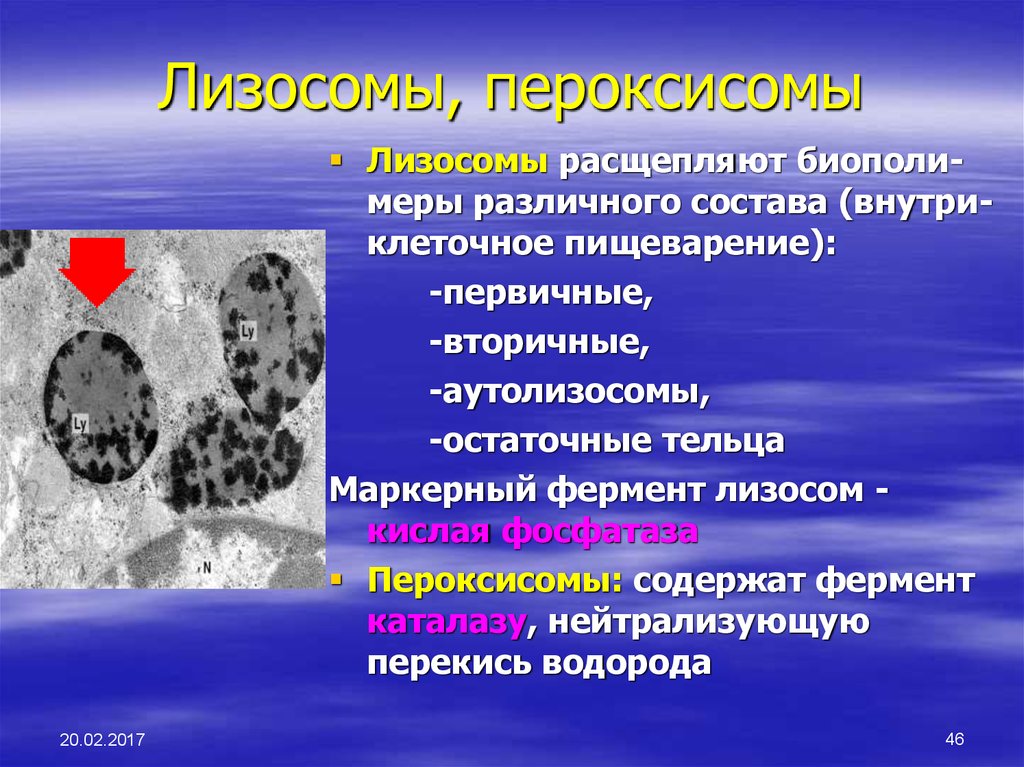
Похожие статьи
Биохимически отличные везикулы из эндоплазматического ретикулума сливаются с образованием пероксисом.
ван дер Занд А., Гент Дж., Браакман И., Табак Х.Ф. ван дер Занд А. и др. Клетка. 13 апреля 2012 г.; 149(2):397-409. doi: 10.1016/j.cell.2012.01.054. Клетка. 2012. PMID: 22500805
Пероксисомы: ответвления ER.
ван дер Занд А., Табак Х.Ф. ван дер Занд А. и др. Curr Opin Cell Biol. 2013 авг; 25 (4): 449-54. doi: 10.1016/j.ceb.2013.05.004. Epub 2013 14 июня. Curr Opin Cell Biol. 2013. PMID: 23773570 Обзор.
Роль эндоплазматического ретикулума в биогенезе пероксисом.

Димитров Л., Лам С.К., Шекман Р. Димитров Л. и соавт. Колд Спринг Харб Перспект Биол. 2013 1 мая; 5(5):a013243. doi: 10.1101/cshperspect.a013243. Колд Спринг Харб Перспект Биол. 2013. PMID: 23637287 Бесплатная статья ЧВК. Обзор.
Биогенез пероксисом: пероксисомальная эндомембранная система и роль ER.
Титоренко В.И., Маллен Р.Т. Титоренко В.И. и соавт. Джей Селл Биол. 2006 3 июля; 174 (1): 11-7. doi: 10.1083/jcb.200604036. Epub 2006 26 июня. Джей Селл Биол. 2006. PMID: 16801391 Бесплатная статья ЧВК. Обзор.
Биогенез пероксисом de novo: развитие концепций и загадок.
Агравал Г., Субрамани С. Агравал Г.
и соавт.
Биохим Биофиз Акта. 2016 май; 1863 (5): 892-901. doi: 10.1016/j.bbamcr.2015.09.014. Epub 2015 14 сентября.
Биохим Биофиз Акта. 2016.
PMID: 26381541
Бесплатная статья ЧВК.
Обзор.
 и соавт.
Биохим Биофиз Акта. 2016 май; 1863 (5): 892-901. doi: 10.1016/j.bbamcr.2015.09.014. Epub 2015 14 сентября.
Биохим Биофиз Акта. 2016.
PMID: 26381541
Бесплатная статья ЧВК.
Обзор.
и соавт.
Биохим Биофиз Акта. 2016 май; 1863 (5): 892-901. doi: 10.1016/j.bbamcr.2015.09.014. Epub 2015 14 сентября.
Биохим Биофиз Акта. 2016.
PMID: 26381541
Бесплатная статья ЧВК.
Обзор.Посмотреть все похожие статьи
Цитируется
Биогенез пероксисом и коммуникация между органеллами: незаменимая роль белков семейства Pex11 и Pex30 в дрожжах.
Деори Н.М., Наготу С. Деори Н.М. и соавт. Карр Жене. 2022 Декабрь; 68 (5-6): 537-550. doi: 10.1007/s00294-022-01254-y. Epub 2022 15 октября. Карр Жене. 2022. PMID: 36242632 Обзор.
Места контакта пероксисом с мембраной у дрожжей.
Джоши А.С. Джоши АС.
Front Cell Dev Biol. 2021 19 ноя;9:735031. doi: 10.3389/fcell.2021.735031. Электронная коллекция 2021.
Front Cell Dev Biol. 2021.
PMID: 34869317
Бесплатная статья ЧВК.
Обзор.Биохимическая основа митохондриальной дисфункции при расстройствах спектра Зеллвегера.
Нубель Э., Морган Дж. Т., Фогарти С., Винтер Дж. М., Леттлова С., Берг Дж. А., Чен Ю. К., Кидвелл К. Ю., Машек Дж. А., Клоуэрс К. Дж., Аргириу С., Чен Л., Виттиг И., Кокс Дж. Э., Рох-Джонсон М., Браверман Н., Бонковски Дж., Гиги С.П., Раттер Дж. Нубель Э. и др. Представитель EMBO 5 октября 2021 г .; 22 (10): e51991. doi: 10.15252/embr.202051991. Epub 2021 5 августа. Представитель EMBO 2021. PMID: 34351705 Бесплатная статья ЧВК.
Гликосомная гетерогенность кинетопластид.
Кроу Л.П., Моррис МТ. Кроу Л.П. и др. Биохим Сок Транс. 2021 февраль 26;49(1):29-39. дои: 10.1042/BST20190517. Биохим Сок Транс. 2021. PMID: 33439256 Бесплатная статья ЧВК. Обзор.
Колебание липидов в пероксисомах и эндолизосомах при активации Т-клеток.
Досиль С.Г., Рохас-Гомес А., Санчес-Мадрид Ф., Мартин-Кофресес Н.Б. Досил С.Г. и соавт. Int J Mol Sci. 2020 19 апреля; 21 (8): 2859. дои: 10.3390/ijms21082859. Int J Mol Sci. 2020. PMID: 32325900 Бесплатная статья ЧВК. Обзор.
 Front Cell Dev Biol. 2021 19 ноя;9:735031. doi: 10.3389/fcell.2021.735031. Электронная коллекция 2021.
Front Cell Dev Biol. 2021.
PMID: 34869317
Бесплатная статья ЧВК.
Обзор.
Front Cell Dev Biol. 2021 19 ноя;9:735031. doi: 10.3389/fcell.2021.735031. Электронная коллекция 2021.
Front Cell Dev Biol. 2021.
PMID: 34869317
Бесплатная статья ЧВК.
Обзор.
Просмотреть все статьи «Цитируется по»
Типы публикаций
термины MeSH
вещества
Пероксисомы — Клетка — Книжная полка NCBI
Пероксисомы представляют собой небольшие, окруженные мембраной органеллы (), которые содержат ферменты, участвующие в различных метаболических реакциях, включая несколько аспектов энергетического метаболизма.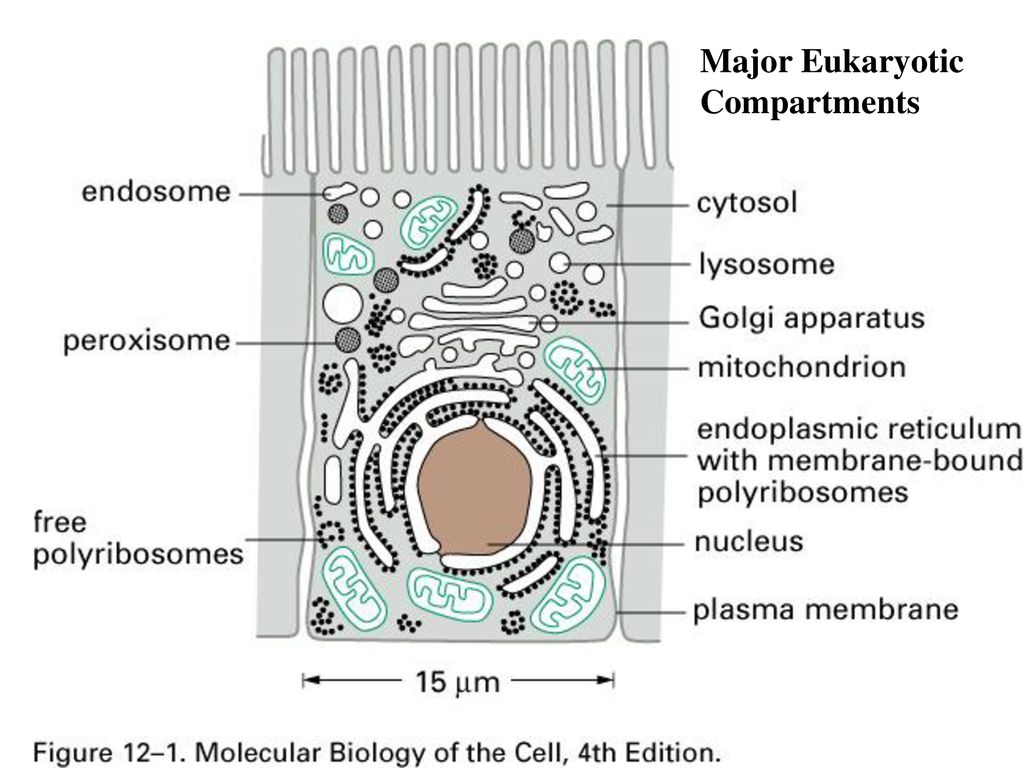 Хотя пероксисомы морфологически сходны с лизосомами, они собираются, подобно митохондриям и хлоропластам, из белков, которые синтезируются на свободных рибосомах и затем импортируются в пероксисомы в виде завершенных полипептидных цепей. Хотя пероксисомы не содержат собственных геномов, они сходны с митохондриями и хлоропластами в том, что реплицируются путем деления.
Хотя пероксисомы морфологически сходны с лизосомами, они собираются, подобно митохондриям и хлоропластам, из белков, которые синтезируются на свободных рибосомах и затем импортируются в пероксисомы в виде завершенных полипептидных цепей. Хотя пероксисомы не содержат собственных геномов, они сходны с митохондриями и хлоропластами в том, что реплицируются путем деления.
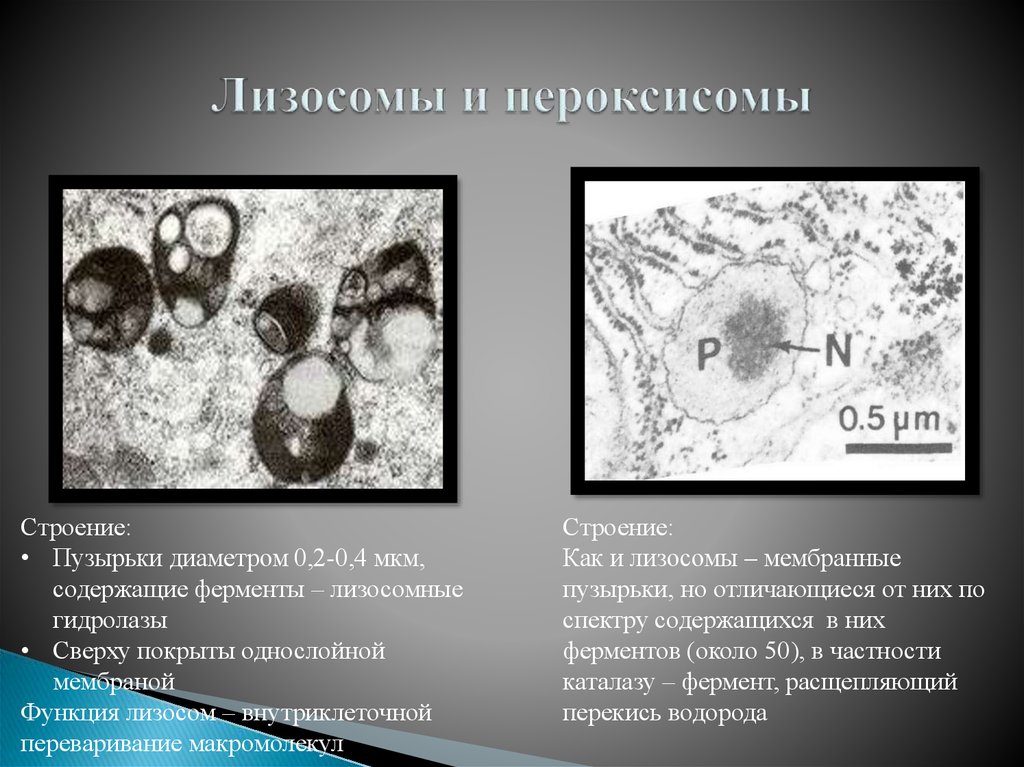
Рисунок 10.24
Электронная микрофотография пероксисом. Показаны три пероксисомы из печени крысы. Два содержат плотные области, которые представляют собой паракристаллические массивы фермента уратоксидазы. (Don Fawcett/Photo Researchers, Inc.)
Функции пероксисом
Пероксисомы содержат не менее 50 различных ферментов, которые участвуют в различных биохимических процессах в различных типах клеток. Первоначально пероксисомы определялись как органеллы, которые осуществляют реакции окисления, приводящие к образованию перекиси водорода. Поскольку перекись водорода вредна для клетки, пероксисомы также содержат фермент каталазу, которая разлагает перекись водорода, либо превращая ее в воду, либо используя ее для окисления другого органического соединения. В ходе таких окислительных реакций в пероксисомах расщепляются различные субстраты, включая мочевую кислоту, аминокислоты и жирные кислоты. Окисление жирных кислот () является особенно важным примером, поскольку оно обеспечивает основной источник метаболической энергии. В клетках животных жирные кислоты окисляются как в пероксисомах, так и в митохондриях, но в дрожжах и растениях окисление жирных кислот ограничивается пероксисомами.
В ходе таких окислительных реакций в пероксисомах расщепляются различные субстраты, включая мочевую кислоту, аминокислоты и жирные кислоты. Окисление жирных кислот () является особенно важным примером, поскольку оно обеспечивает основной источник метаболической энергии. В клетках животных жирные кислоты окисляются как в пероксисомах, так и в митохондриях, но в дрожжах и растениях окисление жирных кислот ограничивается пероксисомами.
Рисунок 10.25
Окисление жирных кислот в пероксисомах. Окисление жирной кислоты сопровождается образованием пероксида водорода (H 2 O 2 ) из кислорода. Перекись водорода разлагается каталазой либо путем превращения в воду, либо путем окисления другого органического вещества (подробнее…)
Помимо обеспечения компартмента для реакций окисления, пероксисомы участвуют в биосинтезе липидов. В клетках животных холестерин и долихол синтезируются как в пероксисомах, так и в ЭР. В печени пероксисомы также участвуют в синтезе желчных кислот, которые образуются из холестерина. Кроме того, пероксисомы содержат ферменты, необходимые для синтеза плазмалогенов — семейства фосфолипидов, в которых одна из углеводородных цепей соединена с глицерином простой эфирной, а не сложноэфирной связью. Плазмалогены являются важными мембранными компонентами в некоторых тканях, особенно в сердце и головном мозге, хотя в других они отсутствуют.
Кроме того, пероксисомы содержат ферменты, необходимые для синтеза плазмалогенов — семейства фосфолипидов, в которых одна из углеводородных цепей соединена с глицерином простой эфирной, а не сложноэфирной связью. Плазмалогены являются важными мембранными компонентами в некоторых тканях, особенно в сердце и головном мозге, хотя в других они отсутствуют.
Рисунок 10.26
Структура плазмалогена. Показанный плазмалоген аналогичен фосфатидилхолину. Однако одна из цепей жирных кислот соединена с глицерином эфирной, а не сложноэфирной связью.
Пероксисомы играют две особенно важные роли в растениях. Во-первых, пероксисомы в семенах ответственны за преобразование накопленных жирных кислот в углеводы, что имеет решающее значение для обеспечения энергией и сырьем для роста прорастающего растения. Это происходит посредством ряда реакций, называемых циклом глиоксилата, который является вариантом цикла лимонной кислоты (16). Пероксисомы, в которых это происходит, иногда называют глиоксисомы .
Рисунок 10.27
Глиоксилатный цикл. Растения способны синтезировать углеводы из жирных кислот посредством глиоксилатного цикла, который является вариантом цикла лимонной кислоты (см. рис. 2.34). Как и в цикле лимонной кислоты, ацетил-КоА соединяется с оксалоацетатом с образованием (подробнее…)
Во-вторых, пероксисомы в листьях участвуют в фотодыхании , которое служит для метаболизма побочного продукта, образующегося в процессе фотосинтеза (). СО 2 превращается в углеводы во время фотосинтеза посредством ряда реакций, называемых циклом Кальвина (см. Ресурсы). Первый шаг — добавление CO 2 к пятиуглеродному сахару рибулозо-1,5-бисфосфату с получением двух молекул 3-фосфоглицерата (по три атома углерода каждая). Однако задействованный фермент (рибулозобисфосфаткарбоксилаза или рубиско) иногда катализирует присоединение O 2 вместо CO 2 с образованием одной молекулы 3-фосфоглицерата и одной молекулы фосфогликолята (два атома углерода). Это побочная реакция, а фосфогликолят не является полезным метаболитом. Сначала он превращается в гликолят, а затем переносится на пероксисомы, где окисляется и превращается в глицин. Затем глицин переносится в митохондрии, где две молекулы глицина превращаются в одну молекулу серина с потерей СО 2 и NH 3 . Затем серин возвращается к пероксисомам, где он превращается в глицерат. Наконец, глицерат переносится обратно в хлоропласты, где снова входит в цикл Кальвина. Фотодыхание, по-видимому, не приносит пользы растению, поскольку оно, по сути, является обратным фотосинтезу: O 2 потребляется, а CO 2 высвобождается без какого-либо увеличения АТФ. Однако случайное использование O 2 вместо CO 2 , по-видимому, является неотъемлемым свойством рубиско, поэтому фотодыхание является общим сопровождением фотосинтеза. Таким образом, пероксисомы играют важную роль, позволяя восстанавливать и использовать большую часть углерода в гликоляте.
Это побочная реакция, а фосфогликолят не является полезным метаболитом. Сначала он превращается в гликолят, а затем переносится на пероксисомы, где окисляется и превращается в глицин. Затем глицин переносится в митохондрии, где две молекулы глицина превращаются в одну молекулу серина с потерей СО 2 и NH 3 . Затем серин возвращается к пероксисомам, где он превращается в глицерат. Наконец, глицерат переносится обратно в хлоропласты, где снова входит в цикл Кальвина. Фотодыхание, по-видимому, не приносит пользы растению, поскольку оно, по сути, является обратным фотосинтезу: O 2 потребляется, а CO 2 высвобождается без какого-либо увеличения АТФ. Однако случайное использование O 2 вместо CO 2 , по-видимому, является неотъемлемым свойством рубиско, поэтому фотодыхание является общим сопровождением фотосинтеза. Таким образом, пероксисомы играют важную роль, позволяя восстанавливать и использовать большую часть углерода в гликоляте.
Рисунок 10.28
Роль пероксисом в фотодыхании. Во время фотосинтеза CO 2 превращается в углеводы по циклу Кальвина, который начинается с присоединения CO 2 к пятиуглеродному сахару рибулозо-1,5-бисфосфату. Однако фермент иногда участвует (подробнее…)
Сборка пероксисом
Как уже отмечалось, сборка пероксисом принципиально аналогична сборке митохондрий и хлоропластов, а не эндоплазматического ретикулума, аппарата Гольджи и лизосом . Белки, предназначенные для пероксисом, транслируются на свободных цитозольных рибосомах, а затем транспортируются в пероксисомы в виде законченных полипептидных цепей. Фосфолипиды также импортируются в пероксисомы через белки-переносчики фосфолипидов из их основного места синтеза в ER. Импорт белков и фосфолипидов приводит к росту пероксисом, а затем путем деления старых образуются новые пероксисомы.
Рисунок 10.29
Сборка пероксисом. Белки, предназначенные для пероксисом, синтезируются на свободных рибосомах и импортируются в уже существующие пероксисомы в виде завершенных полипептидных цепей. Импорт белка приводит к росту пероксисом и образованию новых пероксисом с помощью (далее…)
Импорт белка приводит к росту пероксисом и образованию новых пероксисом с помощью (далее…)
Белки направляются внутрь пероксисом по крайней мере двумя путями, которые сохраняются от дрожжей до человека. Большинство белков нацелены на пероксисомы с помощью простой аминокислотной последовательности Ser-Lys-Leu на их карбокси-конце (сигнал 1 нацеливания на пероксисомы, или PTS1). Другие белки нацелены на последовательность из девяти аминокислот (PTS2) на их амино-конце, а некоторые белки могут быть нацелены на альтернативные сигналы, которые еще точно не определены.
PTS1 и PTS2 распознаются разными рецепторами и затем переносятся в транслокационный комплекс, который обеспечивает их транспорт через мембрану пероксисом. Однако механизм импорта белков в пероксисомы изучен хуже, чем механизмы транслокации белков через мембраны других субклеточных органелл. В отличие от транслокации полипептидных цепей через мембраны эндоплазматического ретикулума, митохондрий и хлоропластов, таргетинговые сигналы обычно не расщепляются при импорте белков в пероксисомы. Цитозольный Hsp70 вовлечен в импорт белка в пероксисомы, но возможная роль молекулярных шаперонов внутри пероксисом неясна. Более того, оказалось, что белки могут транспортироваться в пероксисомы, по крайней мере, в частично свернутых конформациях, а не в виде вытянутых полипептидных цепей.
Цитозольный Hsp70 вовлечен в импорт белка в пероксисомы, но возможная роль молекулярных шаперонов внутри пероксисом неясна. Более того, оказалось, что белки могут транспортироваться в пероксисомы, по крайней мере, в частично свернутых конформациях, а не в виде вытянутых полипептидных цепей.
Некоторые белки мембран пероксисом сходным образом синтезируются на цитозольных рибосомах и направляются к мембране пероксисом с помощью различных внутренних сигналов. Однако другие эксперименты предполагают, что некоторые пероксисомальные мембранные белки могут быть синтезированы на мембраносвязанных полисомах эндоплазматического ретикулума и затем транспортироваться к пероксисомам, что указывает на роль эндоплазматического ретикулума в поддержании пероксисом. Таким образом, импорт белков в пероксисомы, по-видимому, имеет несколько новых особенностей, что делает его активной областью исследований.
Интересно, что некоторые компоненты путей импорта пероксисом были идентифицированы не только как мутанты дрожжей, но и как мутации, связанные с серьезными заболеваниями человека, включающими нарушения пероксисом.
Leave A Comment