
Эндоплазматическая сеть: основные понятия
Что такое эндоплазматическая сеть
Определение
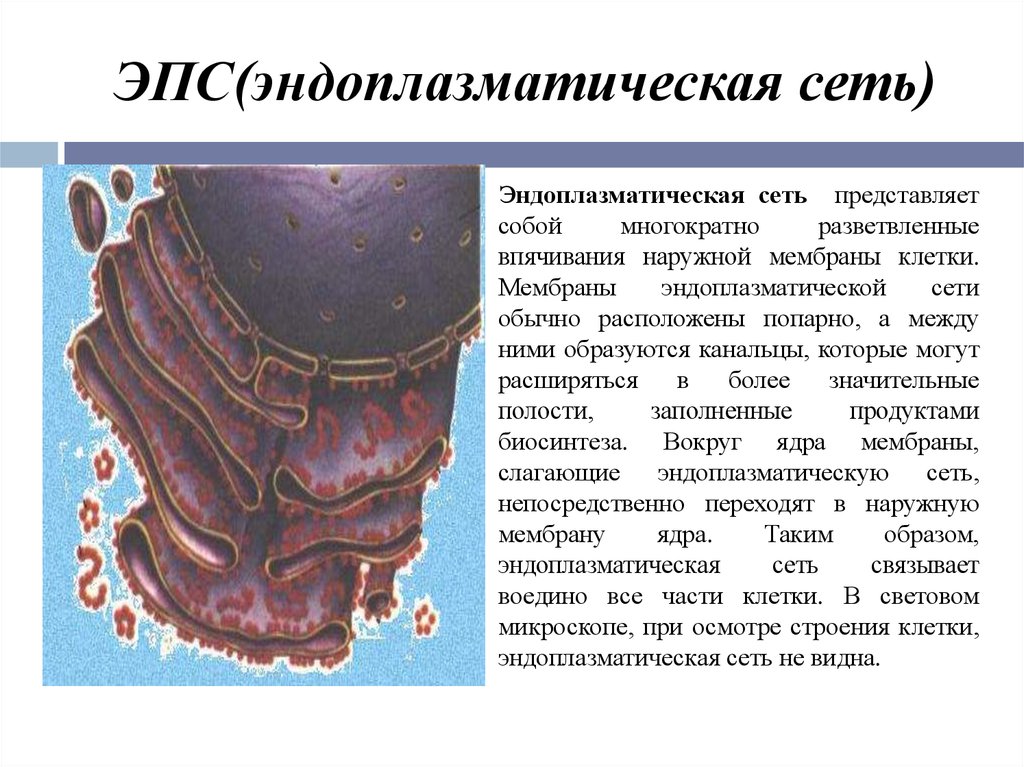
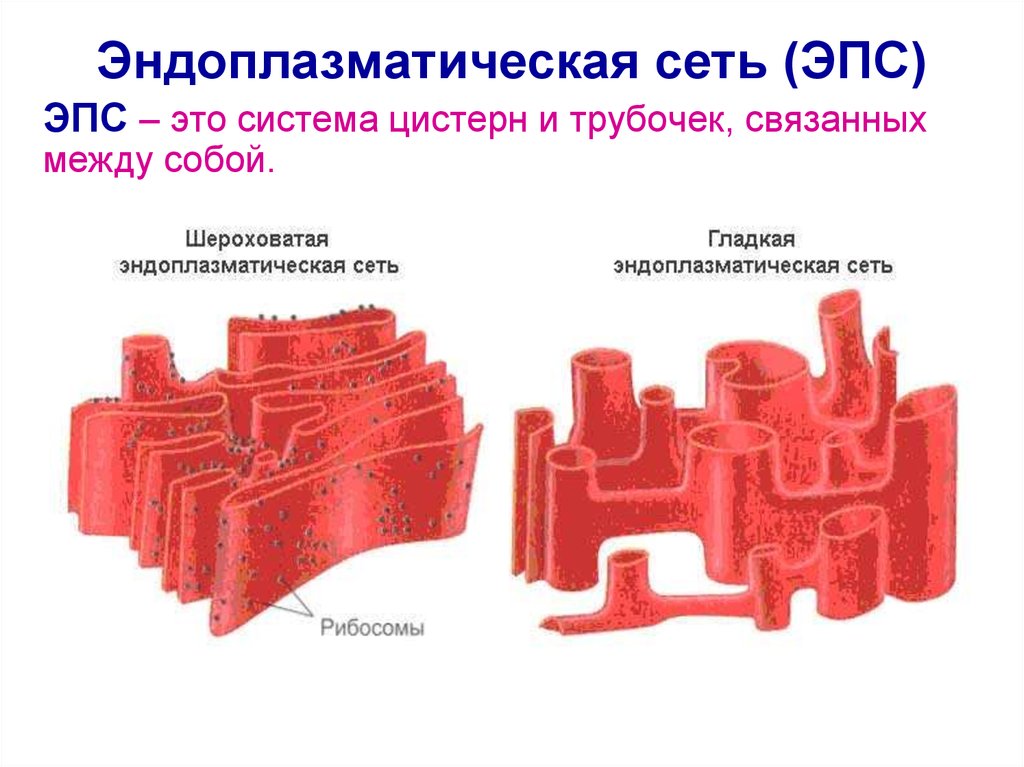
Эндоплазматическая сеть — внутриклеточный органоид, именуемый иначе эндоплазмати́ческим рети́кулумом. В сокращении обозначают ЭПС либо ЭПР. Представлен в виде разветвлённой ультрамикроскопической величины сетки. Компонентами которой служит система: уплощённых полостей, особого рода пузырьков, канальцев, имеющих мембранное окружение.
Сетка имеет непростую структуру и достаточно сложные взаимосвязи. Кроме того, ретикулярные составные в значительной мере не стабильны, они подвержены частым переменам. ЭПР находится в цитоплазме, разделяет её практически равномерно. Делит на секции, структурируя содержимое. Присутствует исключительно в клетках эукариотического типа.
Основные понятия
Определения
Эукариот — это клеточное образование, в котором имеются такие структуры как: оформленное ядро и мембранного типа органоиды.
Клетка — элементарная структурная единица в системе организма, наделённая способностью к самовозобновлению, возможностью к саморегуляции и самовоспроизведению.
Цитолемма (плазмолемма) — окружающая клетку биологическая мембрана.
Ретикулум даёт возможность значимо увеличить суммарные значения площади внутриклеточных поверхностей. Внутренне содержимое органоида — матрикс — материал, являющийся продуктом местного синтеза. Имеет умеренную плотность. На мембранах ЭПР ведётся большое число первичных синтезов. Производятся востребованные для жизнедеятельности клетки продукты.
Первичными их именуют условно, поскольку молекулы синтезированных компонентов станут подвергаться в последующем химическим преобразованиям. Эти синтетические превращения веществ идут как одновременно, так и опосредованно. Поэтому объём и состав химических включений внутри секций неодинаков. Открываются полости ретикулума в перинуклеарное пространство. Расположена последняя между соприкасающимися участками кариолеммы — ядерной оболочки.
Пример:
Площадь сетки эндоплазматического ретикулума занимает более ½ суммарной площади всех клеточных мембранных конструкций.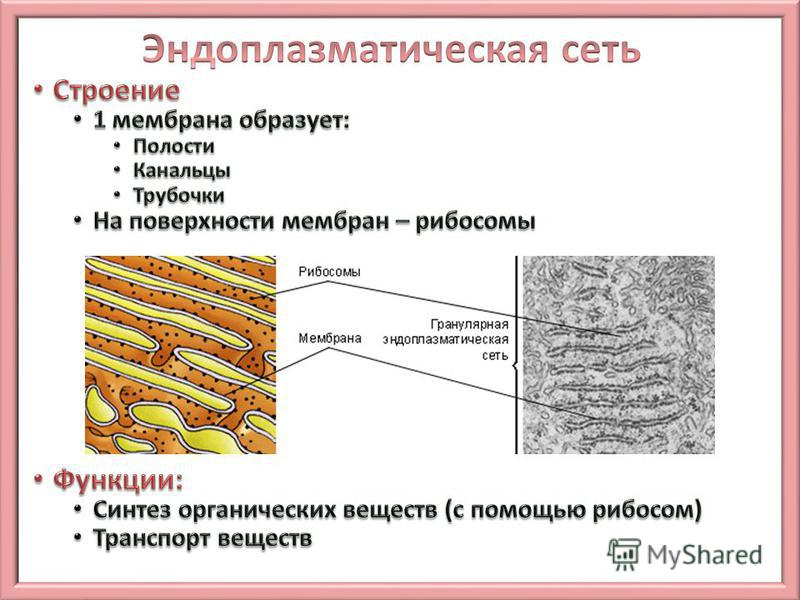 По морфологии она равнозначна покрову клеточного ядра, с которым составляет единое целое. Мембранный аппарат ЭПС позволяют вести активный транспорт ряда компонентов по градиенту концентрации, то есть от меньшей концентрации к большей.
По морфологии она равнозначна покрову клеточного ядра, с которым составляет единое целое. Мембранный аппарат ЭПС позволяют вести активный транспорт ряда компонентов по градиенту концентрации, то есть от меньшей концентрации к большей.
Продукцией ЭПР служат белки, липиды и ферменты: аденозинфосфатазы и синтезированные мембранные липиды.
Строение ретикулума
Органелла имеет следующие варианты строения:
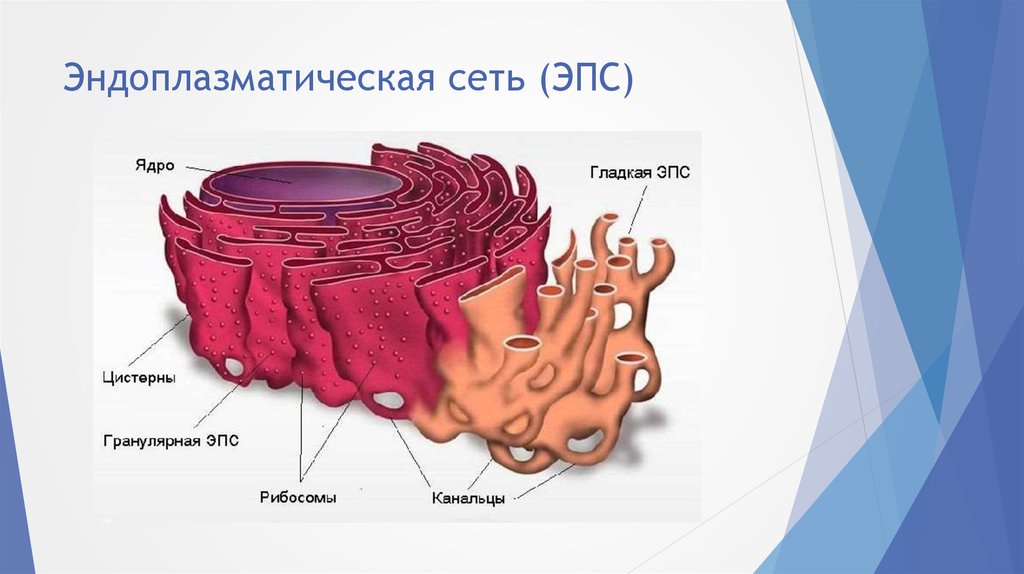
- Шероховатый, иначе гранулярный — грЭС. Имеющий в составе соединённые цистерны, на поверхности несущие рибосомы. Поверхностное их расположение положено в основу наименования грЭС,
- Гладкий, иначе агранулярный — аЭС. Органоид представлен анастомозирующими между собой трубочками, не содержащими на поверхности рибосом.
Выделяют также переходящий или транзиторный вариант ЭПС располагающийся в зоне перехода между вышеуказанными видами клеточного ретикулума.
Агранулярная эндоплазматическая сеть
Расположена в клеточных образованиях: надпочечников — производящих стероидные гормоны, мышечных структур — участвующих в обмене кальция, железистых конструкций желудка — вырабатывающих ионы хлора. Среди прочих видов мембран ЭПР включает: разветвлённые мембранные трубочки, а также везикулы, цистерны, обеспечивающие перемещение синтезированных веществ.
Среди прочих видов мембран ЭПР включает: разветвлённые мембранные трубочки, а также везикулы, цистерны, обеспечивающие перемещение синтезированных веществ.
Гладкий ретикулум весьма чувствителен к факторам внешней среды. По этой причине легко подвергается повреждению. Последнее ведёт к ослаблению клетки, а впоследствии и всего организма. Это может повлечь развитие различных болезненных процессов.
Гранулярная эндоплазматическая сеть
Величина её развитости, зависит напрямую от специализации клетки. Не содержится ЭПР исключительно в сперматозоидах. Особенно развита грЭС в клеточных образованиях следующих органов: поджелудочной железе — производящей ферменты для пищеварения, печени — восполняющей сывороточные белки-альбумины, фибробластах — продуцирующих клеточный коллаген, плазматических клетках — неустанно синтезирующих иммуноглобулины, пр. На поверхности грЭС (гранулярной ЭС) прикрепляются рибосомы — это молекулы шарообразной формы, которые образованы особыми рибонуклеиновыми кислотами.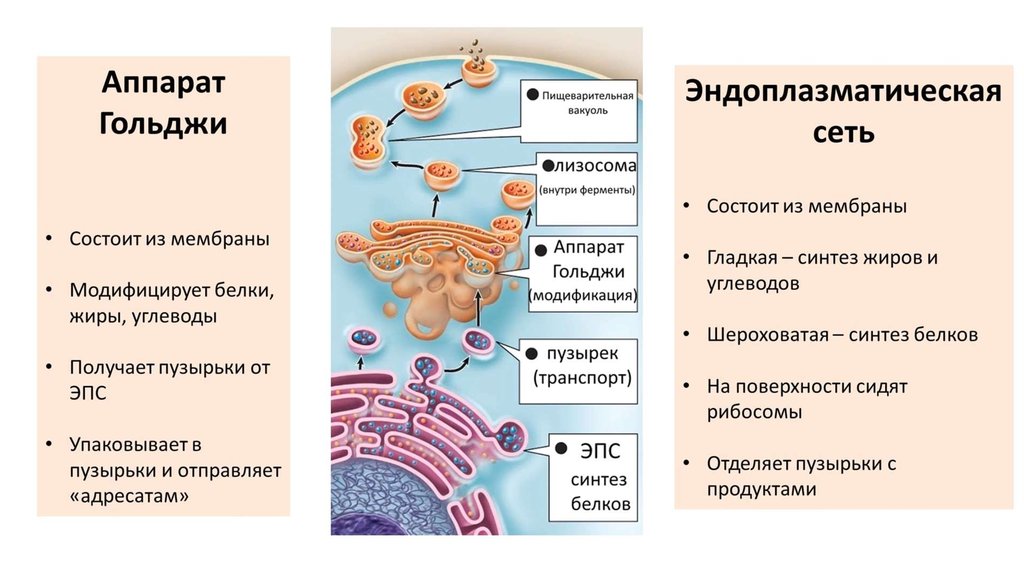
Функции органоида — эндоплазматическая сеть
Ретикулум выполняет достаточно сложные задачи по внутриклеточному синтезу и перемещению полезных компонентов цитоплазмы. Он также участвует в накоплении созданных продуктов. Активно участвует и в построении ядерной оболочки, к примеру, после митоза.
Оба варианта эндоплазматической сети выполняют равные задачи. Синтезируют и перемещают вещества, а, следовательно, ЭС является на клеточном уровне транспортной системой, которая в значительной степени универсальна.
Гладкая и шероховатая ЭС, имеющимся мембранным аппаратом, а также матриксом выполняют равноценные функциональные обязанности. Задачи ЭПР следующие:
- Синтез мембранных липидов. В производстве принимают участие ферменты мембран, именно они содействуют, среди прочего, репродукции ретикулума.
- Разделительная, иначе структурирующая. Позволяет упорядочить цитоплазму, предотвратить её смешивание. Делает невозможным проникновение в органеллы случайных компонентов.

- Проводящая. Разность потенциалов возникающий между двумя мембранами при жизнедеятельности клетки представляет возможность для проведения импульсов.
- Трансмембранный транспорт. Перемещение через мембранный аппарат стенки клеточного образования определённых продуктов.
Помимо перечисленных каждый вариант ЭС выполняет специфические задачи.
Функции шероховатой эндоплазматической сети
Специфические функции характерны также и для шероховатой (гранулярной) ЭПС. В их числе следующие:
- Синтез белков. Отличается процесс синтеза в грЭС тем, что начинается он исключительно на свободных полисомах, которые далее связываются с мембранной системой. Здесь синтезируются все белковые конструкции клеточного мембранного аппарата. Имеются исключения: часть гидрофобных белков, часть протеинов внутренних митохондриальных мембранных конструкций. Также не синтезируются в гранулярной сетке: внутренние мембранные образования хлоропластов, продукт внутренней фазы органелл конструкции мембраны, а также секреторные протеины, переносимые внутриклеточно и за её пределы во внеклеточный промежуток.
- Транспорт веществ. Ведётся по внутренней зоне сетки. По промежуткам ЭС вновь синтезированные белки проводятся к комплексу Гольджи, который, в свою очередь, выводит вещества из клеточных образований.
- Модификация белков пострансляционная. В их числе: гидроксилирование, сульфатирование, фосфорилирование. Происходит гликозилирование во взаимодействии с фермента гликозилтрансферазы. Указанные процессы идут перед секрецией, проводятся перед перемещением компонентов к некоторым органоидам. Таким, как лизосомы, комплекс Гольджи и, конечно, плазмолемма.
- Создание органоидов. Благодаря участию грЭС в сетке образуется комплекс Гольджи.
Базовой задачей сетки зернистого типа становится перемещение протеинов, синтезированных в рибосомах. Последние расположены на поверхности ретикулярной системы. При этом синтезированный протеин поступает внутрь конструкции ЭПС, где скручиваясь приобретает третичную структуру, пр.
Протеины, транспортируемые к цистернам значительно трансформируются по ходу перемещения. Проходят фосфорилирование, гликозилирование, пр. Обычный вариант перемещения для протеинов — это движение через шероховатый тип и эндоплазматическую сеть в комплекс Гольджи, через который они выводятся из клетки либо в ней же поступают к прочим органеллам, к примеру, лизосомам. Могут также откладываться внутри клеточной структуры в виде запасных гранул.
Проходят фосфорилирование, гликозилирование, пр. Обычный вариант перемещения для протеинов — это движение через шероховатый тип и эндоплазматическую сеть в комплекс Гольджи, через который они выводятся из клетки либо в ней же поступают к прочим органеллам, к примеру, лизосомам. Могут также откладываться внутри клеточной структуры в виде запасных гранул.
В гепатоцитах обе ЭПС: зернистого и гладкого типов принимают непосредственное участие в ходе детоксикации многочисленных ядовитых веществ, которые впоследствии подлежат выведению из клетки.
Внешняя мембрана и ретикулярная имеют избирательную проницаемость. По этой причине концентрация компонентов внутри и снаружи конструкций каналов сетки не равны, что непосредственно оказывает влияние на функционирование клетки.
Пример:
Сетка содержащаяся в мышечных волоконных структурах содержит значительно большее количество ионов кальция, нежели цитоплазма. Между внутренней и наружной поверхностями сетки создаётся разница потенциалов.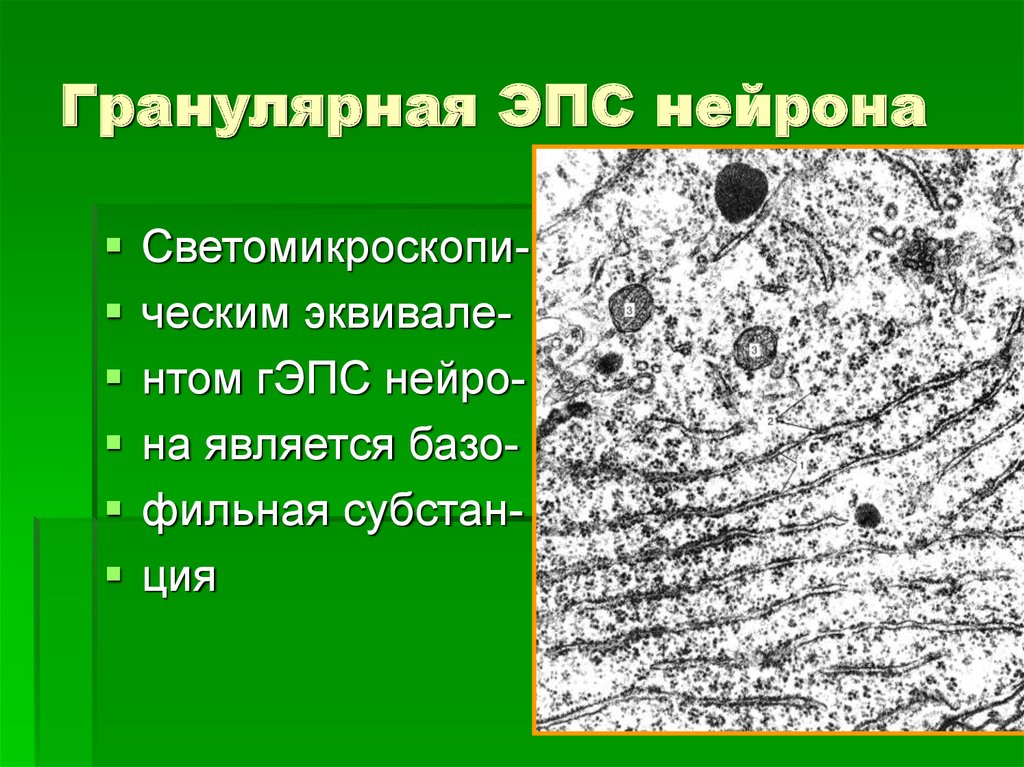 Такая система позволяет проводить импульсы. Так происходит процесс сокращения волоконных структур мышц.
Такая система позволяет проводить импульсы. Так происходит процесс сокращения волоконных структур мышц.
Нет времени решать самому?
Наши эксперты помогут!
Контрольная
| от 300 ₽ |
Реферат
| от 500 ₽ |
Курсовая
| от 1 000 ₽ |
Функции гладкой эндоплазматической сети
Индивидуальными задачами сетки гладкого типа являются следующие:
- Синтез компонентов. Создание в клетке эндоплазматической сетью образований липидного спектра: холестерина и стероидов. Стероидные гормоны производятся большей частью в надпочечниках, половых железах, продуцируются в печени, почках. В структурах кишечника производятся липиды, которые впоследствии выводятся в лимфу и кровеносное русло. Органоиды клеток печени — гепатоциты активно продуцируют гликоген.
- Депо кальция. Обеспечивает накопление и изменение концентрации кальция. Размещается в клетках мышц скелета, сердца, в яйцеклетках, а также в нейронах. Кроме этого, поперечно-полосатые волоконца мышц содержат особую модификацию ЭПС, именуемую саркоплазматическим ретикулумом. Он становится резервуаром кальций-ионов. В конструкции сети имеются мощные кальциевые насосы, которые способны выбрасывать внутриклеточно в цитоплазму значительное число кальция либо перемещать его за сотые доли секунды в систему полостей и канальцев сетки.
- Детоксикация. Производится обезвреживание токсинов всех типов: экзогенного и эндогенного.
 Он становится резервуаром кальций-ионов. В конструкции сети имеются мощные кальциевые насосы, которые способны выбрасывать внутриклеточно в цитоплазму значительное число кальция либо перемещать его за сотые доли секунды в систему полостей и канальцев сетки.
Он становится резервуаром кальций-ионов. В конструкции сети имеются мощные кальциевые насосы, которые способны выбрасывать внутриклеточно в цитоплазму значительное число кальция либо перемещать его за сотые доли секунды в систему полостей и канальцев сетки. Пример:
Гладкого типа ЭПР внутри гепатоцитов активно обезвреживает всевозможные яды. При этом ферменты органоида присоединяют к молекулам токсичных компонентов специфические гидрофильные радикалы. Вредоносные для организма токсичные вещества становятся растворимыми в крови и моче. Поэтому довольно скоро выводятся из тела.
Эндоплазматическая сеть в клетке
Рассматриваемая нами органелла учёными выявлена в цитоплазме, она же окружает содержимое ядра. Покровы органоида-ЭПР включают рибосомы. Из мембранных конструкций структурируются митохондрии клеточных субстанций.
Синтез эндоплазматической сети
Внутренними ферментами самой же сетки производятся липидные её составляющие.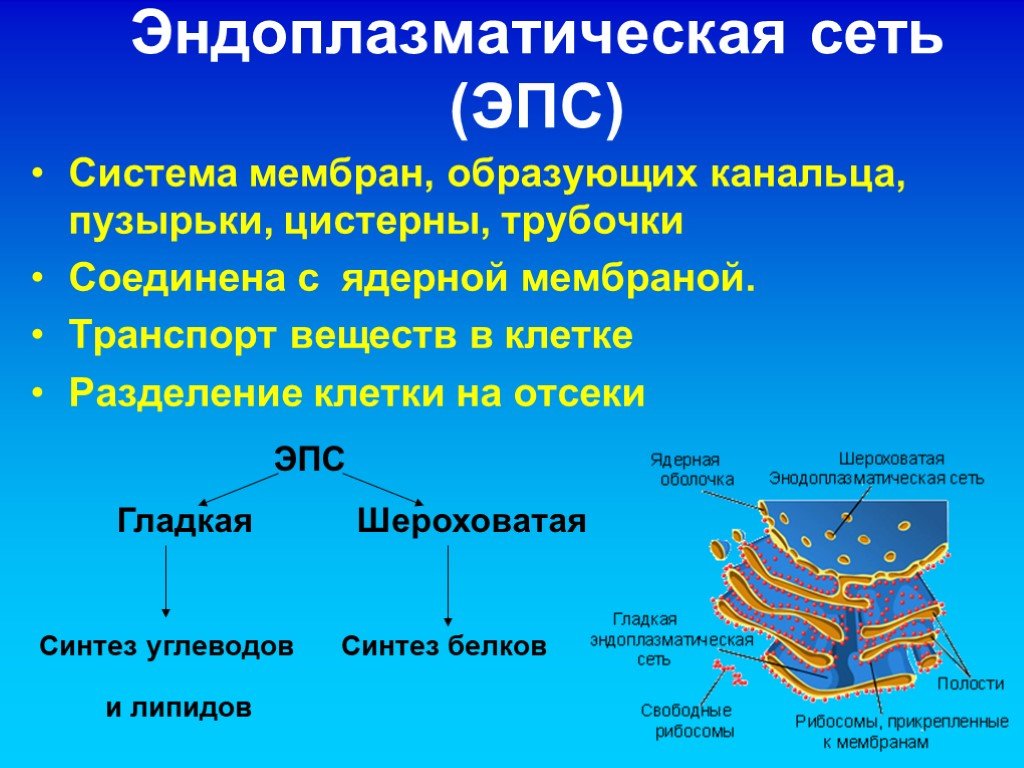 Белковые составляющие поступают из рибосом, размещённых на её мембранах. При этом аЭС собственные протеины производить не имеет возможности, поскольку в своём составе не содержит предназначенные для этих целей структуры. Синтез рассматриваемой органеллы производится за счёт использования резервов рибосомального аппарата грЭС — так характеризуют этот процесс исследователи.
Белковые составляющие поступают из рибосом, размещённых на её мембранах. При этом аЭС собственные протеины производить не имеет возможности, поскольку в своём составе не содержит предназначенные для этих целей структуры. Синтез рассматриваемой органеллы производится за счёт использования резервов рибосомального аппарата грЭС — так характеризуют этот процесс исследователи.
В таблице представляем читателю особенности строения и функций эндоплазматической сети:
Эндоплазматическая сеть | Цитология | Биология
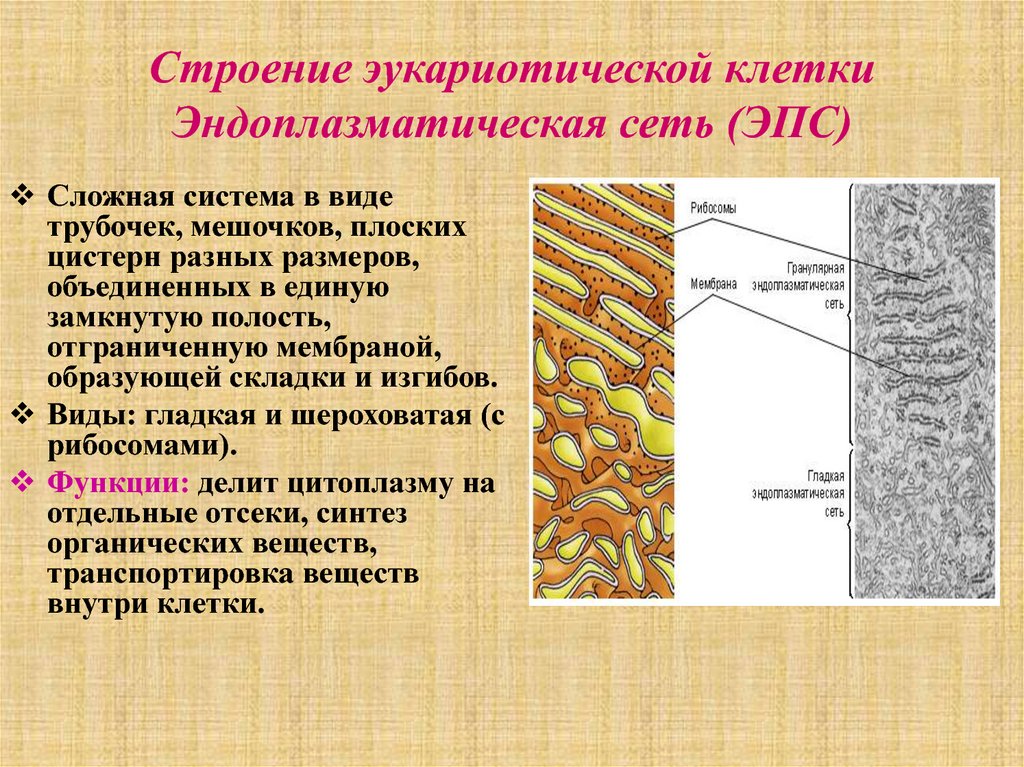
Эндоплазматическую сеть (ЭПС), или эндоплазматический ретикулум (ЭПР), удалось обнаружить только с появлением электронного микроскопа. ЭПС есть только в эукариотических клетках и представляет собой сложную систему мембран, образующих уплощенные полости и трубочки. Все вместе это выглядит как сеть. ЭПС относится к одномембранным органоидам клетки.
Микрофотография эндоплазматической сетиМембраны ЭПС отходят от внешней мембраны ядра и по строению сходны с ней.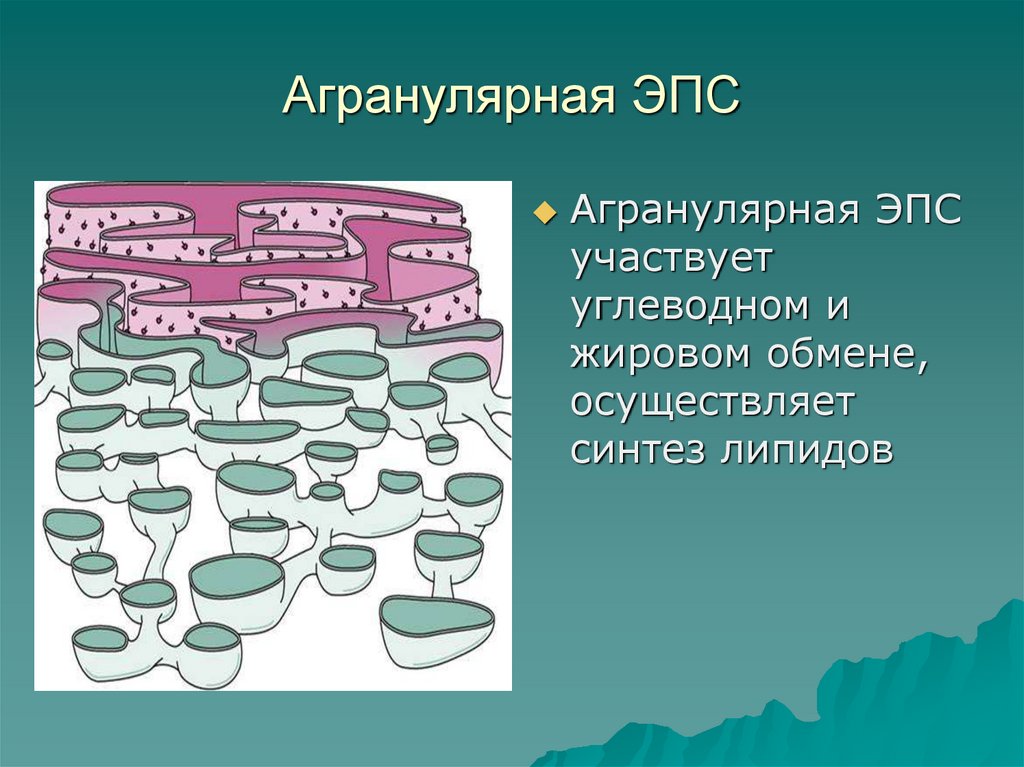
Эндоплазматическая сеть делится на гладкую (агранулярную) и шероховатую (гранулярную). Последняя усеяна прикрепленными к ней рибосомами (из-за этого и возникает «шероховатость»). Основная функция обоих типов связана с синтезом и транспортом веществ. Только шероховатая отвечает за белок, а гладкая — за углеводы и жиры.
По своему строению ЭПС представляет собой множество парных параллельных мембран, пронизывающих почти всю цитоплазму. Пара мембран образует пластинку (полость внутри имеет разную ширину и высоту), однако гладкая эндоплазматическая сеть в большей степени имеет трубчатое строение. Такие уплощенные мембранные мешочки называют
Рибосомы, расположенные на шероховатой ЭПС, синтезируют белки, которые поступают в каналы ЭПС, созревают (приобретают третичную структуру) там и транспортируются. У таких белков сначала синтезируется сигнальная последовательность (состоящая преимущественно из неполярных аминокислот), конфигурация которой соответствует специфическому рецептору ЭПС.
После того, как белок оказывается в канале эндоплазматического ретикулума сигнальная последовательность от него отделяется. После этого он свертывается в свою третичную структуру. При транспортировке по ЭПС белок приобретает ряд других изменений (фосфорилирование, образование связи с углеводом, т. е. превращение в гликопротеин).
Большинство белков, оказавшихся в шероховатой ЭПС, далее попадают в аппарат (комплекс) Гольджи. Оттуда белки либо секретируются из клетки, либо поступают в другие органоиды (обычно лизосомы), либо откладываются как гранулы запасных веществ.
Следует иметь в виду, что не все белки клетки синтезируются на шероховатой ЭПС. Часть (обычно меньшая) синтезируется свободными рибосомами в гиалоплазме, такие белки используются самой клеткой. У них сигнальная последовательность не синтезируется за ненужностью.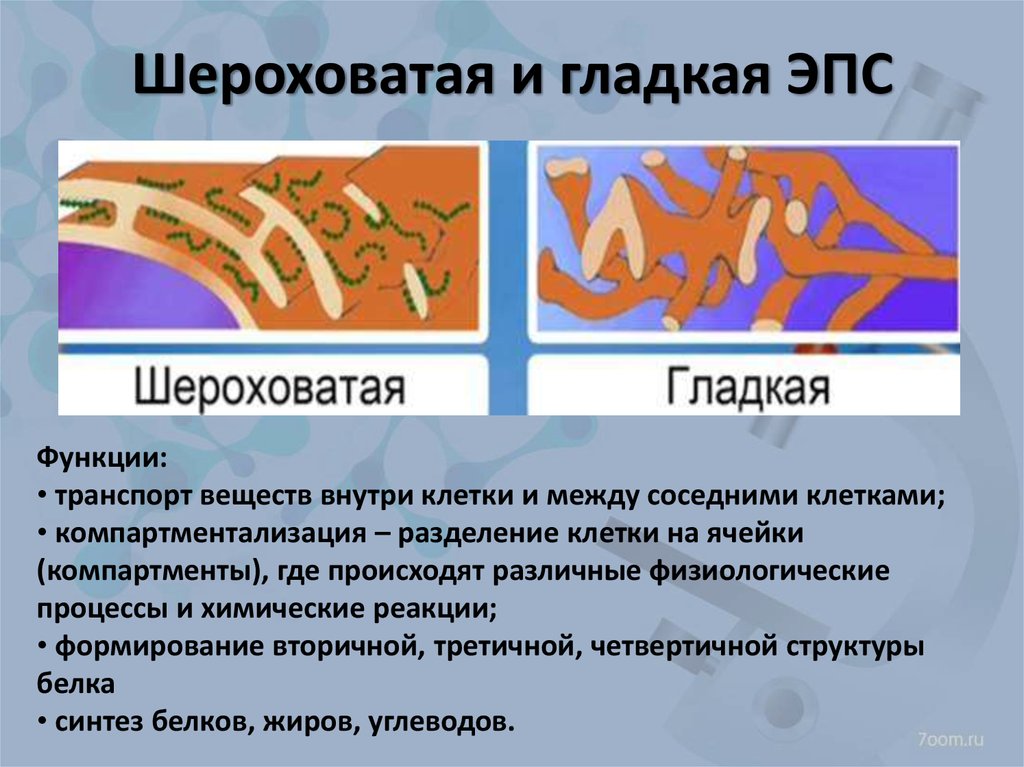
Основной функцией гладкой эндоплазматической сети является синтез липидов (жиров). Например, ЭПС эпителия кишечника синтезирует их из жирных кислот и глицерола, всасывающихся из кишечника. Затем липиды попадают в комплекс Гольджи. Кроме клеток кишечника, гладкая ЭПС хорошо развита в клетках, секретирующих стероидные гормоны (стероиды относятся к липидам). Например, в клетках надпочечников, интерстициальных клетках семенников.
Синтез и транспорт белков, жиров и углеводов не единственные функции ЭПС. В печение эндоплазматический ретикулум участвует в процессах детоксикации. Особая форма гладкой ЭПС — саркоплазматический ретикулум – присутствует в мышечных клетках и обеспечивает сокращение за счет перекачки ионов кальция.
Структура, объем и функциональность эндоплазматической сети клетки не является постоянной на протяжении клеточного цикла, а подвержены тем или иным изменениям.
Роль внеклеточных полимерных веществ ЭПС в аэробной грануляции
Последнее обновление пятница, 07 января 2022 г.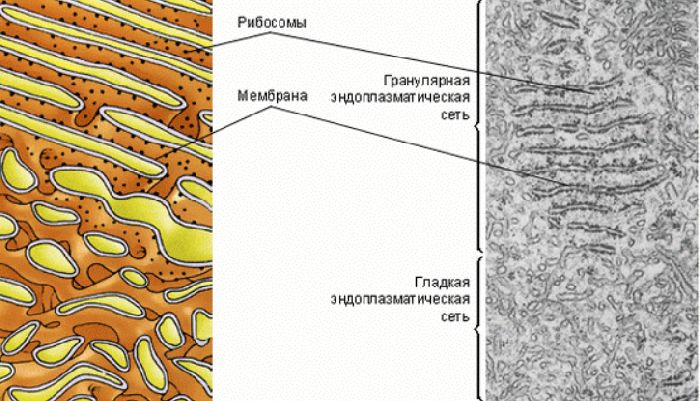 |
Аэробные гранулы
|
Аэробные гранулы
Внеклеточные полимерные вещества (ВПС) представляют собой липкие материалы, секретируемые клетками, и могут играть важную роль в явлениях клеточной адгезии, формировании структуры матрикса, микробной физиологии и улучшении долговременной стабильности гранул (Schmidt and Ahring, 1994; Tay et al., 2001c; Liu et al., 2004b; McSwain et al., 2005). Высокое содержание полисахаридов может способствовать межклеточному взаимодействию и дополнительно укреплять микробную структуру за счет образования полимерной матрицы. Накопление ЭПС в виде капсульного материала и периферической слизи коррелирует с процессами биологической адгезии и агрегации (Costerton et al., 19).81; Тай и др., 2001c; Лю и др., 2002). Было обнаружено, что метаболическая блокировка синтеза экзополисахарида предотвращает агрегацию микробов (Cammarota and Sant’Anna, 1998; Yang et al., 2004). Было высказано предположение, что ЭПС в гранулах соединяют две соседние бактериальные клетки физически друг с другом, а также с другими инертными твердыми частицами и оседают в виде агрегатов (Liu et al.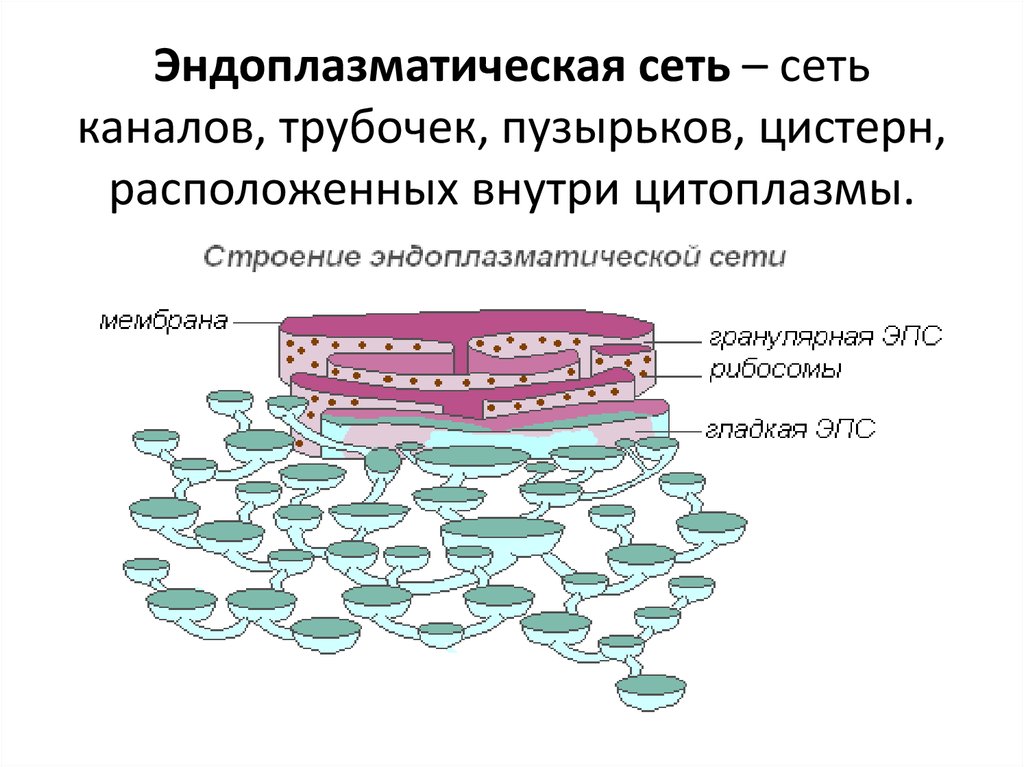
Индивидуальная бактерия
Индивидуальная бактерия
Полимерная цепь EPS
Рис. 4.3. Схематическое изображение биогрануляции, усиленной внеклеточным полимерным веществом (Liu et al., 2004b).
Полимерная цепь EPS
Рис. 4.3. Схематическое изображение биогрануляции, усиленной внеклеточным полимерным веществом (Liu et al., 2004b).
ЭПС наблюдали в различных типах биогранул с помощью сканирующей электронной микроскопии и просвечивающей электронной микроскопии. В процессе биогрануляции пенополистирол может обеспечить обширную поверхность для связывания бактерий (рис. 4.3). Кроме того, внеклеточный полисахаридный матрикс, окружающий агрегированные бактерии, может предоставлять сайты, доступные для привлечения органических и неорганических материалов (Yu et al., 2001; Sponza, 2002; Liu et al., 2004b). Имеющиеся данные показывают, что образование биогранул является микробной эволюцией, а не случайным скоплением взвешенных микробов (El-Mamouni et al.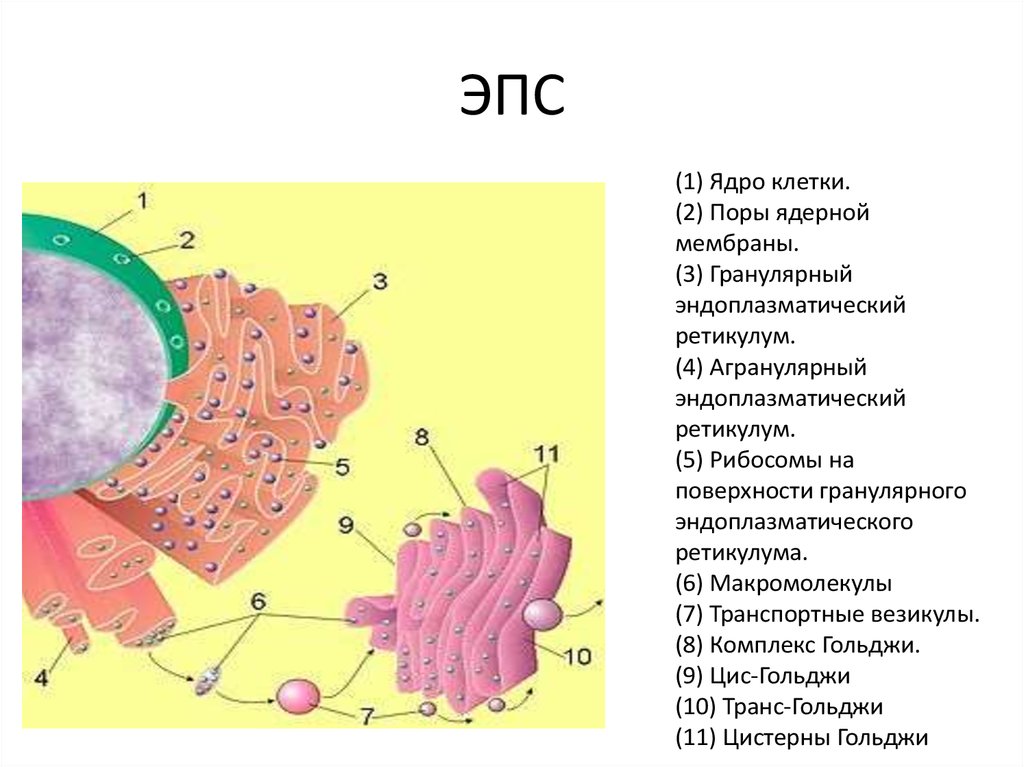
Пространственное распределение ЭПС в биогранулах должно коррелировать с микробной эволюцией и распределением во время образования биогранул. Исследование пространственного распределения ЭПС по глубине в гетеротрофных биопленках показало, что выход ЭПС имеет тенденцию к снижению с глубиной биопленки (Zhang and Bishop, 2001). Вероятно, это связано с тем, что жизнеспособная биомасса теряет способность продуцировать ЭПС в более глубоких отделах биопленок из-за более низкой микробной активности в результате меньшей доступности питательных веществ. Совсем недавно Wang et al. (2005b) обнаружили, что внешняя оболочка аэробной гранулы состоит из плохо растворимого и трудно биоразлагаемого ЭПС, тогда как ее сердцевинная часть заполнена легко растворимым и биоразлагаемым ЭПС. На рис. 4.4 показано, что флуоресцентный 9м; (в) профиль распределения интенсивности флуоресценции красителя по радиусу гранулы от поверхности к центру (стрелка) (Wang et al., 2005b).
(калькофлуор белый) в основном был прикреплен к внешней оболочке гранулы, в то время как в центре гранулы флуоресценция была очень слабой. Кроме того, ЭПС, продуцируемый бактериями, может быть использован в качестве вторичного субстрата в более глубоких слоях или зонах аэробных гранул, где легкоразлагаемые субстраты либо недоступны, либо ограничены (Chi, 2005; Wang et al., 2005b). Представляется, что пространственное распределение и свойства ЭПС, а не его абсолютное количество в аэробных гранулах, играют существенную роль в стабилизации структуры и поддержании прочности микробных агрегатов (Wang et al., 2005b). 9-связанный ЭПС будет в основном располагаться во внешней оболочке гранулы. Считается, что нерастворимый ЭПС, присутствующий в оболочке гранулы, будет играть защитную роль в отношении структурной стабильности и целостности аэробных гранул, т.е. нерастворимый ЭПС может служить основой агрегированной структуры, а легко биоразлагаемый ЭПС, расположенный в ядре гранулы будут играть менее важную роль.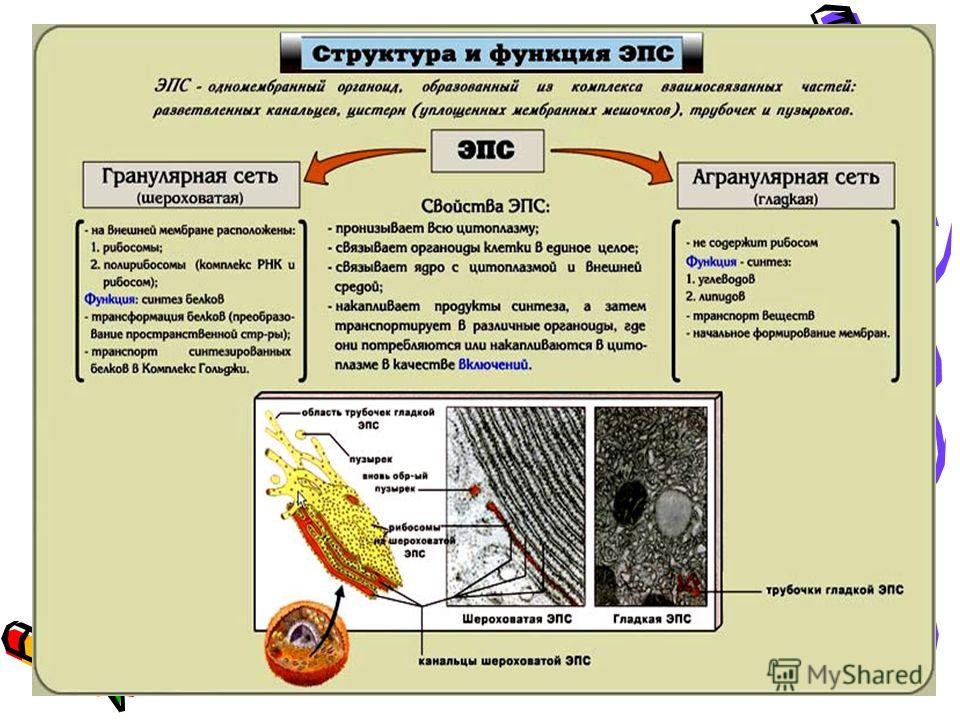
Продолжить чтение здесь: Состав субстрата
Была ли эта статья полезной?
Эффекты субстратного шока на экскрецию внеклеточного полимерного вещества (EPS) и характеристики прикрепленных биопленочных гранул анаммокса
Влияние шока субстрата на экскрецию внеклеточного полимерного вещества (EPS) и характеристики прикрепленных к биопленке гранул анаммокса†
Яньлун Чжан, и Хайюань Ма, и Цигуи Ниу, и Ронг Чен, аб Тосимаса Ходжо и и Ю-Ю Ли* ак
Принадлежности автора
* Соответствующие авторы
и Кафедра гражданской и экологической инженерии, Высшая инженерная школа, Университет Тохоку, 6-6-06 Аоба, Арамаки, Аоба-ку, Сендай, Япония
Электронная почта: Gyokuyu. [email protected]
[email protected]
б Key Lab of Northwest Water Resource, Environment and Ecology, MOE, Xi’an University of Architecture and Technology, No.13 Yanta Road, Сиань 710055, Китай
с Высшая школа экологических исследований, Университет Тохоку, 6-6-06 Аоба, Арамаки, Аоба-ку, Сендай, Япония
Аннотация
 org/schema/rscart38″> Предполагается, что стрессы окружающей среды оказывают значительное влияние на содержание и концентрацию образующихся внеклеточных полимерных веществ (ВПС) и, следовательно, влияют на характеристики реактора с вспененным слоем с прикрепленной пленкой ананммокс (AAFEB). В этом исследовании преходящая высокая концентрация субстрата 2500 мг N л −1 (рассчитана как сумма NN 4 + –N и NO 2 2 7 − –N) в течение 24 часов стимулировал обильное выделение ЭПС, а также порчу гранул анаммокса. Результаты показали, что высокая концентрация ЭПС, составляющая 89,6 ± 48,3 мг/г -1 ВСС, приводила к снижению осаждаемости гранул на 35,0 ± 0,8% и уменьшению общего количества ВСС на 30,5 ± 0,9%. Производство EPS было обоснованно связано с воздействием субстрата, связанным с использованием и стрессом. Результаты серии периодических экспериментов показали, что быстрое увеличение содержания слабосвязанного ЭПС (LB-EPS) с 41,2 до 114,6 мг/г -1 VSS наблюдался, когда концентрация субстрата неуклонно увеличивалась с 400 до 1000 мг N л -1 , напротив, прочно связанный ЭПС (ТБ-ЭПС) оставался стабильным на уровне 32,5 ± 2,8 мг г −1 ВСС.
org/schema/rscart38″> Предполагается, что стрессы окружающей среды оказывают значительное влияние на содержание и концентрацию образующихся внеклеточных полимерных веществ (ВПС) и, следовательно, влияют на характеристики реактора с вспененным слоем с прикрепленной пленкой ананммокс (AAFEB). В этом исследовании преходящая высокая концентрация субстрата 2500 мг N л −1 (рассчитана как сумма NN 4 + –N и NO 2 2 7 − –N) в течение 24 часов стимулировал обильное выделение ЭПС, а также порчу гранул анаммокса. Результаты показали, что высокая концентрация ЭПС, составляющая 89,6 ± 48,3 мг/г -1 ВСС, приводила к снижению осаждаемости гранул на 35,0 ± 0,8% и уменьшению общего количества ВСС на 30,5 ± 0,9%. Производство EPS было обоснованно связано с воздействием субстрата, связанным с использованием и стрессом. Результаты серии периодических экспериментов показали, что быстрое увеличение содержания слабосвязанного ЭПС (LB-EPS) с 41,2 до 114,6 мг/г -1 VSS наблюдался, когда концентрация субстрата неуклонно увеличивалась с 400 до 1000 мг N л -1 , напротив, прочно связанный ЭПС (ТБ-ЭПС) оставался стабильным на уровне 32,5 ± 2,8 мг г −1 ВСС.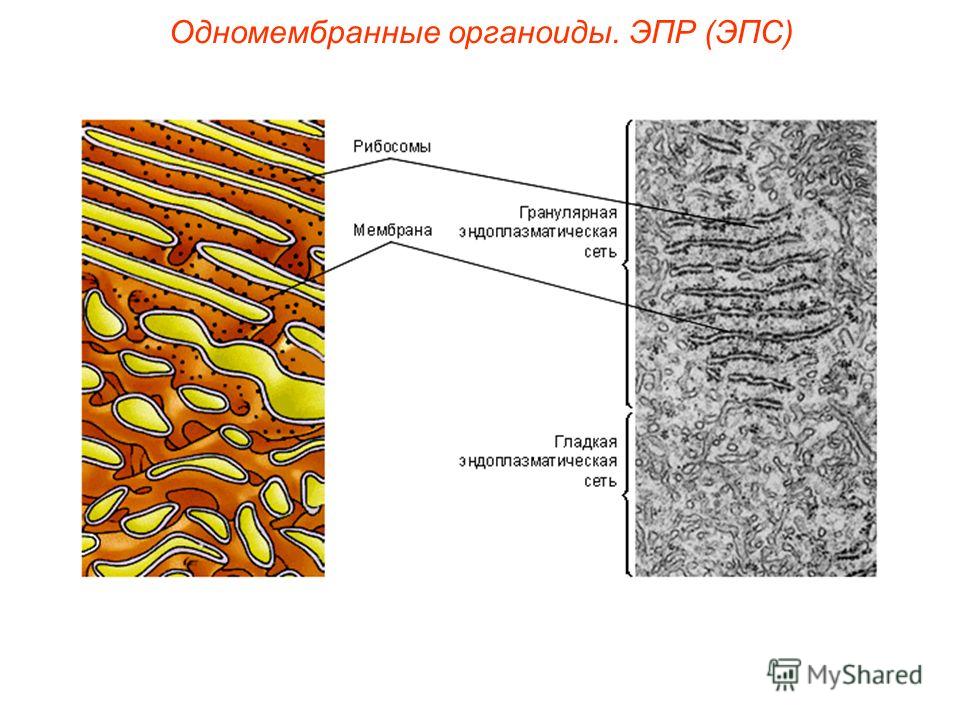
Leave A Comment