
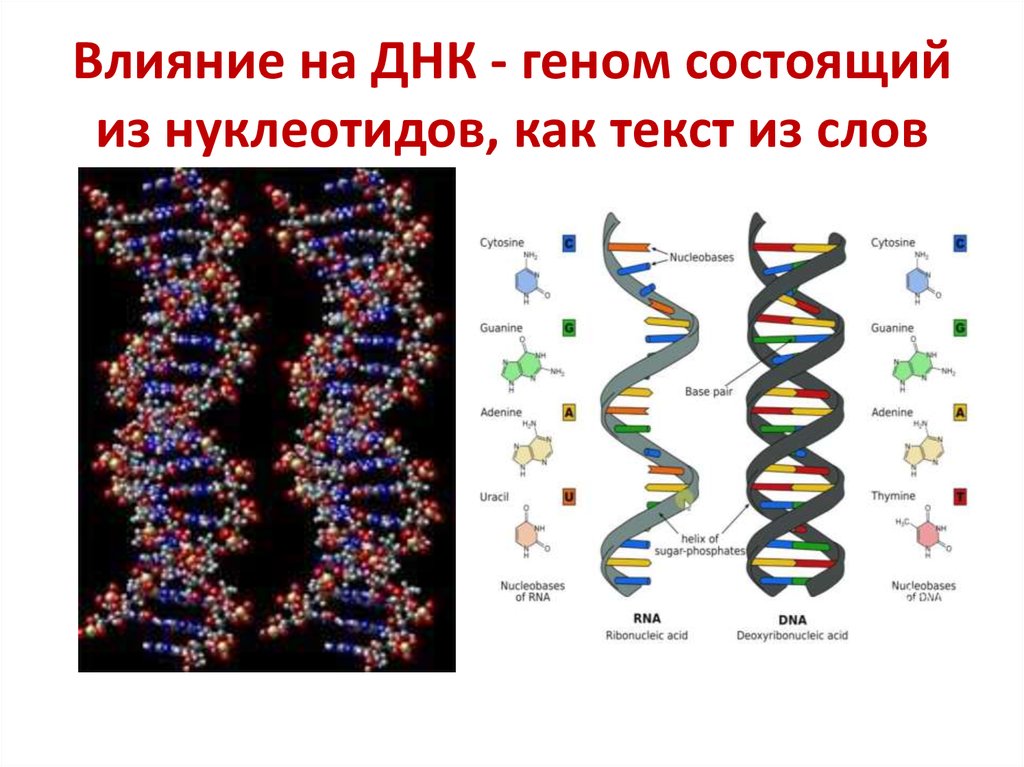
Мономер ДНК. Какие мономеры образуют молекулу ДНК?
Нуклеиновые кислоты, особенно ДНК, достаточно хорошо известны в науке. Объясняется это тем, что они являются веществами клетки, от которых зависит хранение и передача её наследственной информации. ДНК, открытое еще в 1868 году Ф. Мишером, представляет собой молекулы с ярко выраженными кислотными свойствами. Ученый выделил её из ядер лейкоцитов – клеток иммунной системы. В течение последующих 50 лет исследования нуклеиновых кислот проводились эпизодически, так как большинство ученых биохимиков считали главными органическими веществами, отвечающими в том числе и за наследственные признаки, белки.
С момента расшифровки строения ДНК, проведенной Уотсоном и Криком в 1953 году, начинаются серьёзные исследования, выяснившие что, дезоксирибонуклеиновая кислота – это полимер, а мономерами ДНК служат нуклеотиды. Их виды и строение будут изучены нами в данной работе.
Нуклеотиды как структурные единицы наследственной информации
Одно из фундаментальных свойств живой материи – это сохранение и передача информации о строении и функциях как клетки, так и всего организма в целом. Эту роль выполняет дезоксирибонуклеиновая кислота, а мономеры ДНК – нуклеотиды представляют собой своеобразные «кирпичики», из которых и построена уникальная конструкция вещества наследственности. Рассмотрим, какими же признаками руководствовалась живая природа, создавая суперспираль нуклеиновой кислоты.
Эту роль выполняет дезоксирибонуклеиновая кислота, а мономеры ДНК – нуклеотиды представляют собой своеобразные «кирпичики», из которых и построена уникальная конструкция вещества наследственности. Рассмотрим, какими же признаками руководствовалась живая природа, создавая суперспираль нуклеиновой кислоты.
Как образуются нуклеотиды
Чтобы ответить на этот вопрос, нам понадобятся некоторые знания из области химии органических соединений. В частности, мы напомним, что в природе существует группа азотсодержащих гетероциклических гликозидов, соединенных с моносахаридами – пентозами (дезоксирибозой или рибозой). Они называются нуклеозидами. Например, аденозин и другие виды нуклеозидов присутствуют в цитозоле клетки. Они вступают в реакцию этерификации с молекулами ортофосфорной кислоты. Продуктами этого процесса и будут нуклеотиды. Каждый мономер ДНК, а их четыре вида, имеет название, например, гуаниновый, тиминовый и цитозиновый нуклеотид.
Пуриновые мономеры ДНК
В биохимии принята классификация, разделяющая мономеры ДНК и их строение на две группы: так, пуриновыми являются адениновый и гуаниновый нуклеотиды.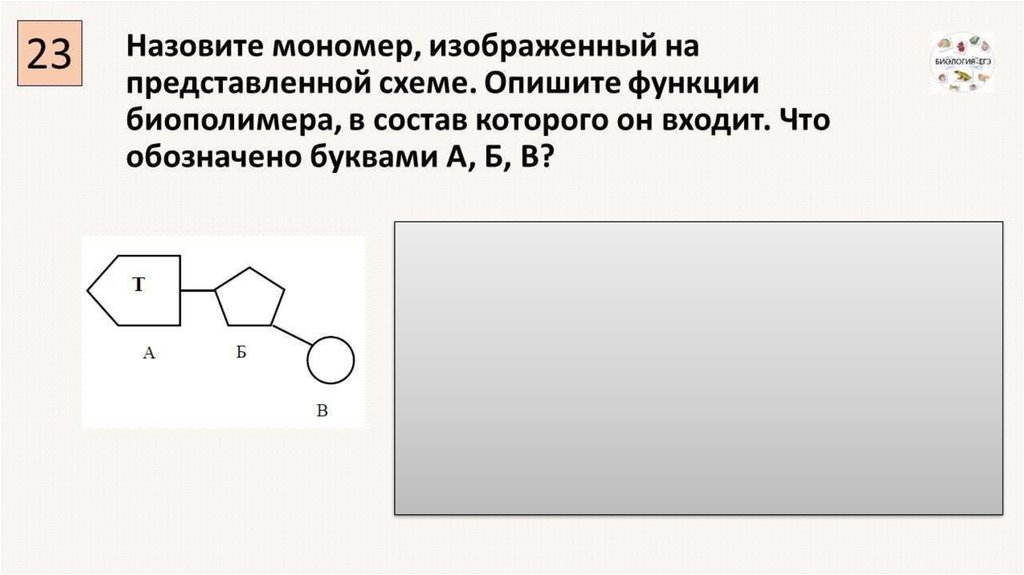 Они содержат в своем составе производные пурина – органического вещества, имеющего формулу C5H4N4. Мономер ДНК – гуаниновый нуклеотид, также содержит пуриновое азотистое основание, соединенное с дезоксирибозой N-гликозидной связью, находящейся в бетоконфигурации.
Они содержат в своем составе производные пурина – органического вещества, имеющего формулу C5H4N4. Мономер ДНК – гуаниновый нуклеотид, также содержит пуриновое азотистое основание, соединенное с дезоксирибозой N-гликозидной связью, находящейся в бетоконфигурации.
Пиримидиновые нуклеотиды
Азотистые основания, называемые цитидином и тимидином, являются производными органического вещества пиримидина. Его формула C4H4N2. Молекула представляет собой шестичленный плоский гетероцикл, содержащий два атома нитрогена. Известно, что вместо тиминового нуклеотида в молекулах рибонуклеиновой кислоты, таких как рРНК, тРНК, иРНК, содержится урациловый мономер. В процессе транскрипции, во время списывания информации с гена ДНК на молекулу иРНК, тиминовый нуклеотид замещается на адениновый, а адениновый нуклеотид – на урациловый в синтезируемой цепи иРНК. То есть справедливой будет следующая запись: А – У, Т – А.
Правило Чаргаффа
В предыдущем разделе мы уже частично коснулись принципов соответствия мономеров в цепях ДНК и в комплексе ген-иРНК. Известный биохимик Э. Чаргафф установил совершенно уникальное свойство молекул дезоксирибонуклеиновой кислоты, а именно, что количество адениновых нуклеотидов в ней всегда равно тиминовым, а гуаниновых – цитозиновым. Главной теоретической базой принципов Чаргаффа послужили исследования Уотсона и Крика, установившие, какие мономеры образуют молекулу ДНК и какую пространственную организацию они имеют. Еще одна закономерность, выведенная Чаргаффом и названная принципом комплементарности, указывает на химическое родство пуриновых и пиримидиновых оснований и их способность при взаимодействии между собой образовывать водородные связи. Это значит, что расположение мономеров в обеих цепях ДНК строго детерминировано: так, напротив А первой цепи ДНК может находиться только Т другой и между ними возникают две водородные связи. Напротив гуанинового нуклеотида может располагаться только цитозиновый. В этом случае между азотистыми основаниями образуются три водородные связи.
Известный биохимик Э. Чаргафф установил совершенно уникальное свойство молекул дезоксирибонуклеиновой кислоты, а именно, что количество адениновых нуклеотидов в ней всегда равно тиминовым, а гуаниновых – цитозиновым. Главной теоретической базой принципов Чаргаффа послужили исследования Уотсона и Крика, установившие, какие мономеры образуют молекулу ДНК и какую пространственную организацию они имеют. Еще одна закономерность, выведенная Чаргаффом и названная принципом комплементарности, указывает на химическое родство пуриновых и пиримидиновых оснований и их способность при взаимодействии между собой образовывать водородные связи. Это значит, что расположение мономеров в обеих цепях ДНК строго детерминировано: так, напротив А первой цепи ДНК может находиться только Т другой и между ними возникают две водородные связи. Напротив гуанинового нуклеотида может располагаться только цитозиновый. В этом случае между азотистыми основаниями образуются три водородные связи.
Роль нуклеотидов в генетическом коде
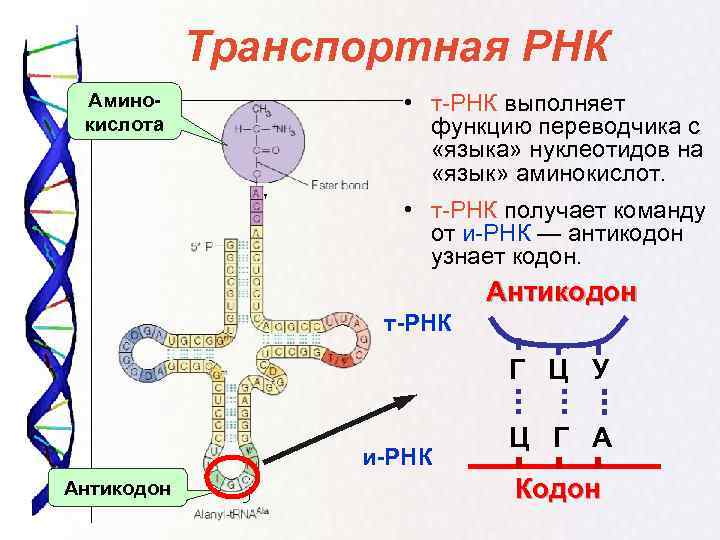
Для осуществления реакции биосинтеза белка, происходящей в рибосомах, существует механизм перевода информации об аминокислотном составе пептида из последовательности нуклеотидов иРНК в последовательность аминокислот.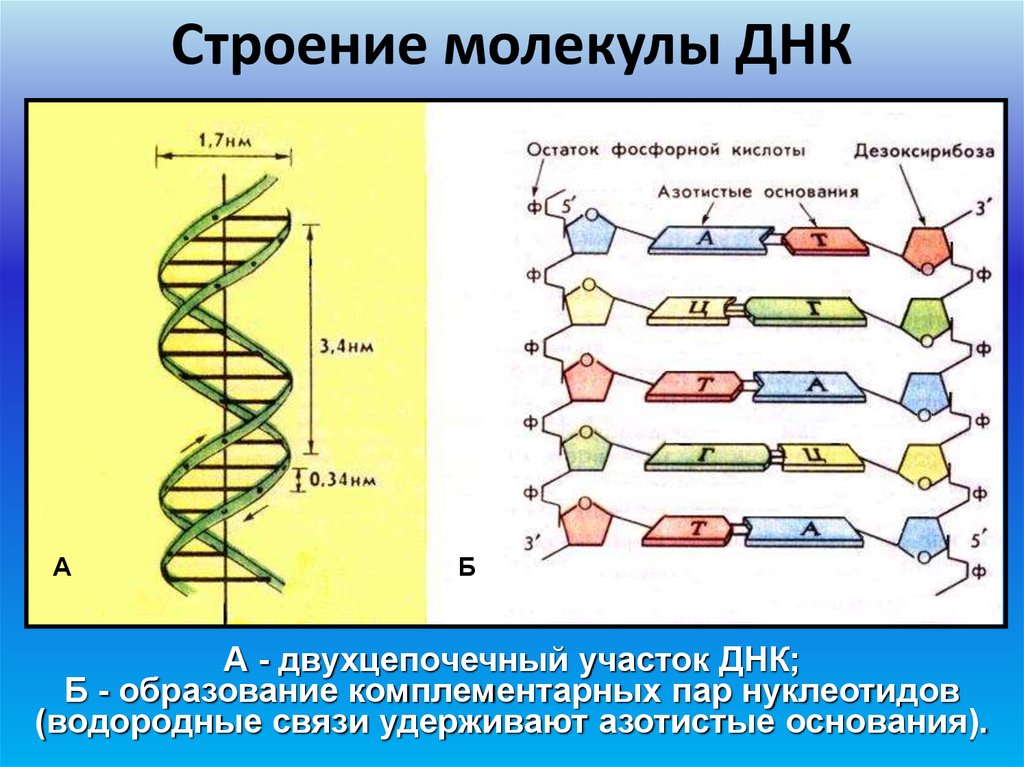 Оказалось, что три рядом расположенных мономера несут в себе информацию об одной из 20 возможных аминокислот. Это явление получило название генетический код. В решении задач по молекулярной биологии его применяют для определения как аминокислотного состава пептида, так и для выяснения вопроса: какие мономеры образуют молекулу ДНК, иными словами, каков состав соответствующего гена. Например, триплет (кодон) ААА в гене кодирует аминокислоту фенилаланин в молекуле белка, а в генетическом коде ей будет соответствовать триплет UUU в цепи иРНК.
Оказалось, что три рядом расположенных мономера несут в себе информацию об одной из 20 возможных аминокислот. Это явление получило название генетический код. В решении задач по молекулярной биологии его применяют для определения как аминокислотного состава пептида, так и для выяснения вопроса: какие мономеры образуют молекулу ДНК, иными словами, каков состав соответствующего гена. Например, триплет (кодон) ААА в гене кодирует аминокислоту фенилаланин в молекуле белка, а в генетическом коде ей будет соответствовать триплет UUU в цепи иРНК.
Взаимодействие нуклеотидов в процессе редупликации ДНК
Как было выяснено ранее, структурные единицы, мономеры ДНК – это нуклеотиды. Их определенная последовательность в цепях является матрицей для процесса синтеза дочерней молекулы дезоксирибонуклеиновой кислоты. Это явление происходит в S-стадии интерфазы клетки. Последовательность нуклеотидов новой молекулы ДНК собирается на материнских цепях под действием фермента ДНК-полимеразы с учетом принципа комплементарности (А – Т, Д – С). Репликация относится к реакциям матричного синтеза. Это значит, что мономеры ДНК и их строение в материнских цепях служат основой, то есть матрицей для её дочерней копии.
Репликация относится к реакциям матричного синтеза. Это значит, что мономеры ДНК и их строение в материнских цепях служат основой, то есть матрицей для её дочерней копии.
Может ли изменяться строение нуклеотида
К слову скажем, что дезоксирибонуклеиновая кислота – это очень консервативная структура клеточного ядра. Этому есть логическое объяснение: наследственная информация, хранящаяся в хроматине ядра, должна быть неизменной и копироваться без искажений. Ну а клеточный геном постоянно находится «под прицелом» факторов внешней среды. Например, таких агрессивных химических соединений, как алкоголь, лекарственное средство, радиоактивное излучение. Все они являются так называемыми мутагенами, под воздействием которых любой мономер ДНК может изменить свое химическое строение. Такое искажение в биохимии называют точковой мутацией. Частота возникновения их в геноме клетки достаточно высока. Мутации исправляются хорошо отлаженной работой клеточной репарационной системы, включающей в себя набор ферментов.
Одни из них, например рестриктазы, «вырезают» поврежденные нуклеотиды, полимеразы обеспечивают синтез нормальных мономеров, лигазы «сшивают» восстановленные участки гена. Если же вышеописанный механизм по какой-то причине в клетке не срабатывает и дефектный мономер ДНК остается в её молекуле, мутация подхватывается процессами матричного синтеза и фенотипически проявляется в виде белков с нарушенными свойствами, неспособных выполнять необходимые функции, присущие им в клеточном обмене веществ. Это является серьёзным негативным фактором, снижающим жизнеспособность клетки и сокращающим продолжительность её жизни.
ДНК (дезоксирибонуклеиновая кислота) | Молекулярная биология
ДНК (дезоксирибонуклеиновая кислота) относится к нуклеиновым кислотам (наряду с РНК), которые являются полимерами, а точнее — полинуклеотидами (мономер — нуклеотид).
ДНК отвечает за хранение и передачу при делении клеток генетического кода. Именно через молекулы ДНК реализуются наследственность и изменчивость.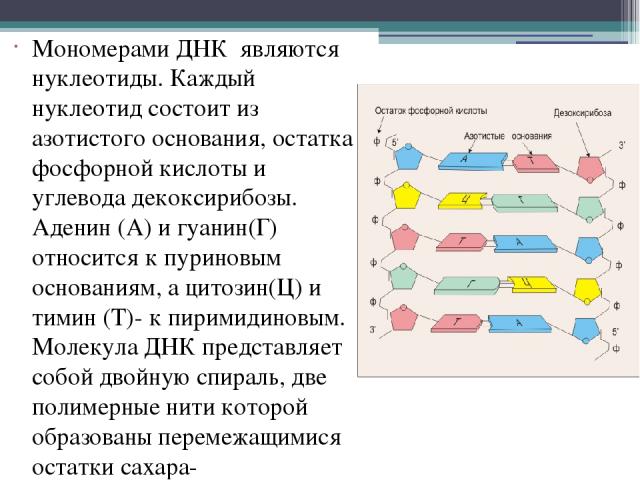 На ДНК синтезируются все виды РНК. Далее различные типы РНК совместно обеспечивают синтез белков клетки, т. е. реализуют генетическую информацию.
На ДНК синтезируются все виды РНК. Далее различные типы РНК совместно обеспечивают синтез белков клетки, т. е. реализуют генетическую информацию.
В клетках эукариот подавляющее количество ДНК находится в ядре, где они образуют комплексы с особыми белками, в результате чего образуются хромосомы. В клетках прокариот существует одна большая кольцевая (или линейная) молекула ДНК (также в комплексе с белками). Кроме того в клетках эукариот своя ДНК есть в митохондриях и хлоропластах.
В случае ДНК каждый нуклеотид состоит из 1) азотистого основания, которое может быть аденином, гуанином, цитозином или тимином, 2) дезоксирибозы, 3) фосфорной кислоты.
Последовательность нуклеотидов в цепочке ДНК определяет первичную структуру молекулы. Для ДНК характерна вторичная структура молекулы в форме двойной спирали (чаще всего правозакрученной). При этом две цепи ДНК соединяются между собой водородными связями, образуемыми между комплементарными азотистыми основаниями.
Аденин комплементарен тимину, а гуанин – цитозину.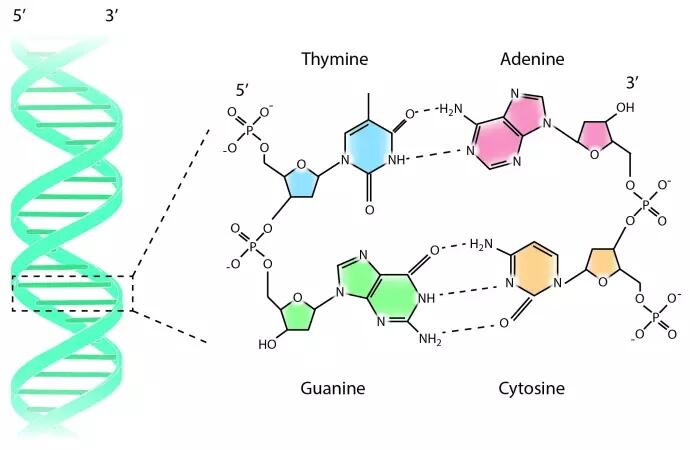 Между аденином и тимином образуются две водородные связи, а между гуанином и цитозином — три. Таким образом, гуанин и цитозин соединены между собой немного прочнее (хотя водородные связи в принципе слабые). Количество связей определяется особенностями строения молекул.
Между аденином и тимином образуются две водородные связи, а между гуанином и цитозином — три. Таким образом, гуанин и цитозин соединены между собой немного прочнее (хотя водородные связи в принципе слабые). Количество связей определяется особенностями строения молекул.
Аденин и гуанин относятся к пуринам и состоят из двух колец. Тимин и цитозин относятся к пиримидиновым основаниям, состоящим из одного кольца. Таким образом между остовами (состоящими из чередующихся дезоксирибозы и фосфорной кислоты) двух цепей ДНК при любой паре нуклеотидов разных цепей всегда существует три кольца (поскольку двухкольцовый пурин всегда комплементарен только определенному однокольцовому пиримидину). Это позволяет сохранять ширину между цепями молекулы ДНК одинаковой на всем протяжении (примерно 2,3 нм).
В одном витке спирали находится примерно 10 нуклеотидов. Длина одного нуклеотида примерно 0,34 нм. Длина же молекул ДНК обычно огромна, превышает миллионы нуклеотидов. Поэтому, чтобы более компактно разместиться в ядре клетки, ДНК подвергается различной степени «сверхспирализации».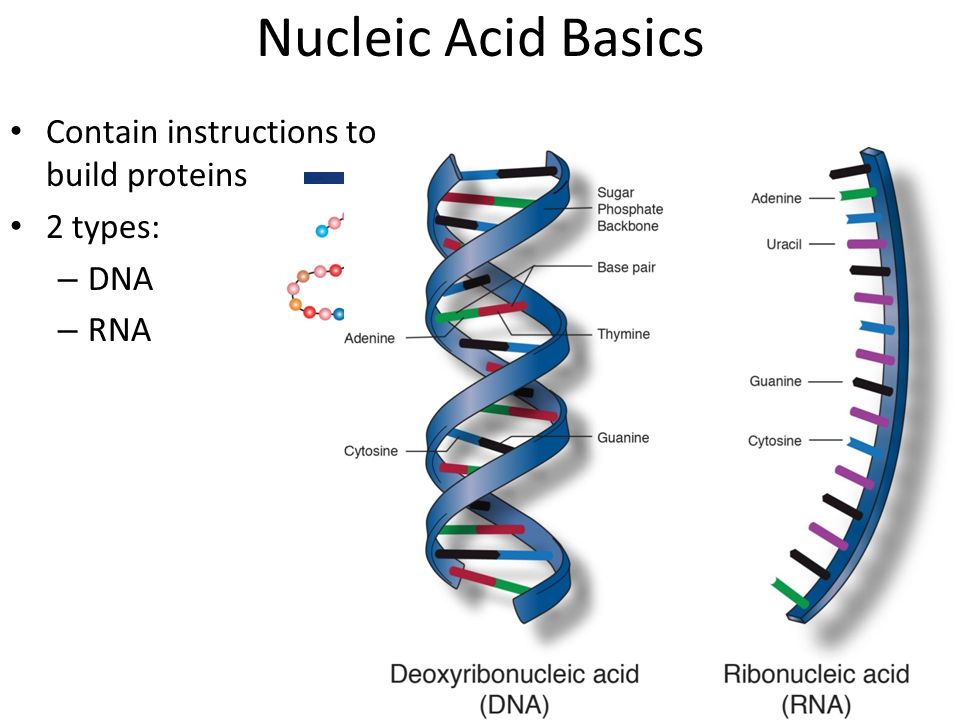
При считывании информации с ДНК (то есть синтезе на ней РНК, этот процесс называется транскрипцией) происходит деспирализация (процесс обратный спирализации), две цепочки расходятся под действием специального фермента. Водородные связи слабые, поэтому разделение и в последующем сшивание цепей происходит при малой затрате энергии. РНК синтезируется на ДНК согласно все тому же принципу комплементарности. Только вместо тимина в РНК аденину комплементарен урацил.
Генетический код, записанный на молекулах ДНК, состоит из триплетов (последовательностей трех нуклеотидов), которые обозначают одну аминокислоту (мономер белка). Однако большая часть ДНК не кодирует белок. Значение таких участков молекулы различно, во многом до конца не выяснено.
Перед делением клетки всегда происходит удвоение количества ДНК. Этот процесс называется репликацией. Она носит полуконсервативный характер: цепи одной молекулы ДНК расходятся, и на каждой достраивается своя новая комплементарная цепь.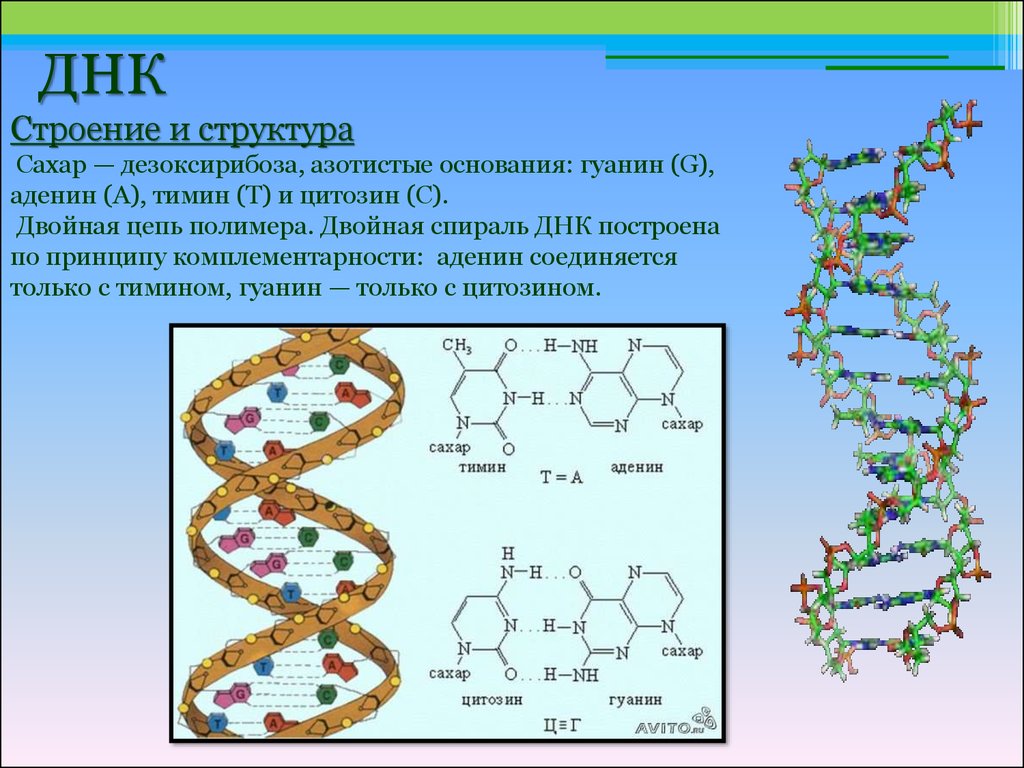
В ДНК полинуклеотидные цепи разнонаправлены, т. е. там где у одной цепи 5′-конец (остаток фосфорной кислоты присоединен к пятому атому углерода дезоксирибозы), у другой — 3′ (углерод, свободный от фосфорной кислоты).
При репликации и транскрипции синтез всегда идет в направлении от 5′-конца к 3′, так как новые нуклеотиды могут присоединяться только к свободному 3′ атому углерода.
Строение и роль ДНК как вещества, отвечающего за наследственную информацию, были выяснены в 40-50-х годах XX века. В 1953 году Д. Уотсон и Ф. Крик определили двухцепочечную структуру ДНК. Ранее Э. Чаргафф выяснил, что в ДНК количество тимина всегда соответствует аденину, а количество гуанина — цитозину.
9.1 Структура ДНК – Концепции биологии – 1-е канадское издание
Перейти к содержанию
Глава 9: Введение в молекулярную биологию
Цели обученияК концу этого раздела вы сможете:
- Описывать структуру ДНК
- Опишите, как устроена эукариотическая и прокариотическая ДНК в клетке
В 1950-х годах Фрэнсис Крик и Джеймс Уотсон вместе работали в Кембриджском университете в Англии над определением структуры ДНК.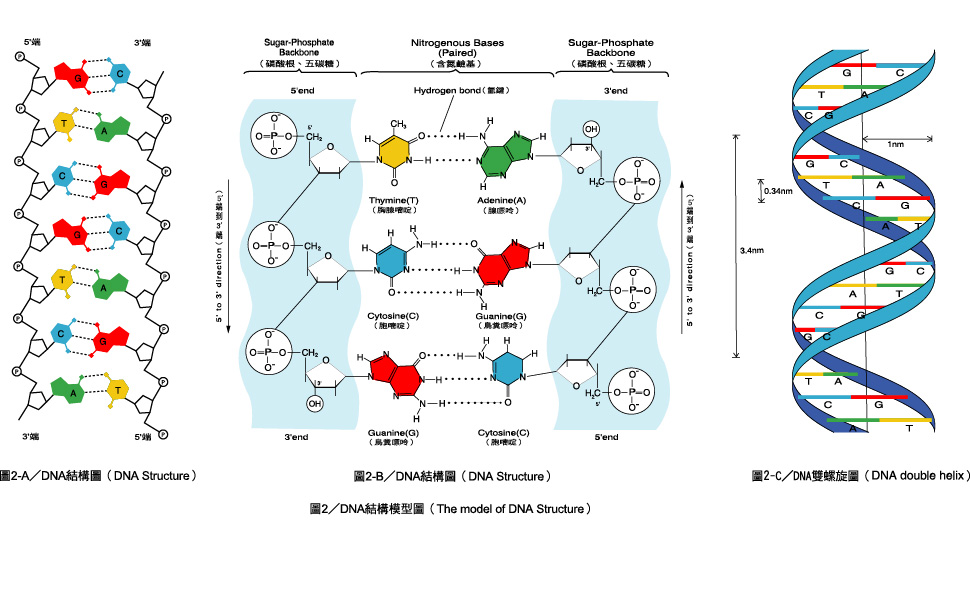 Другие ученые, такие как Лайнус Полинг и Морис Уилкинс, также активно исследовали эту область. Полинг открыл вторичную структуру белков с помощью рентгеновской кристаллографии. Рентгеновская кристаллография — это метод исследования молекулярной структуры путем наблюдения за узорами, образованными рентгеновскими лучами, пропущенными через кристалл вещества. Паттерны дают важную информацию о структуре интересующей молекулы. В лаборатории Уилкинса исследователь Розалинда Франклин использовала рентгеновскую кристаллографию, чтобы понять структуру ДНК. Уотсон и Крик смогли собрать воедино загадку молекулы ДНК, используя данные Франклина (рис. 9)..2). У Уотсона и Крика также была ключевая информация от других исследователей, например, правила Чаргаффа. Чаргафф показал, что из четырех типов мономеров (нуклеотидов), присутствующих в молекуле ДНК, два типа всегда присутствуют в равных количествах, а остальные два типа также всегда присутствуют в равных количествах. Это означало, что они всегда были в паре каким-то образом.
Другие ученые, такие как Лайнус Полинг и Морис Уилкинс, также активно исследовали эту область. Полинг открыл вторичную структуру белков с помощью рентгеновской кристаллографии. Рентгеновская кристаллография — это метод исследования молекулярной структуры путем наблюдения за узорами, образованными рентгеновскими лучами, пропущенными через кристалл вещества. Паттерны дают важную информацию о структуре интересующей молекулы. В лаборатории Уилкинса исследователь Розалинда Франклин использовала рентгеновскую кристаллографию, чтобы понять структуру ДНК. Уотсон и Крик смогли собрать воедино загадку молекулы ДНК, используя данные Франклина (рис. 9)..2). У Уотсона и Крика также была ключевая информация от других исследователей, например, правила Чаргаффа. Чаргафф показал, что из четырех типов мономеров (нуклеотидов), присутствующих в молекуле ДНК, два типа всегда присутствуют в равных количествах, а остальные два типа также всегда присутствуют в равных количествах. Это означало, что они всегда были в паре каким-то образом.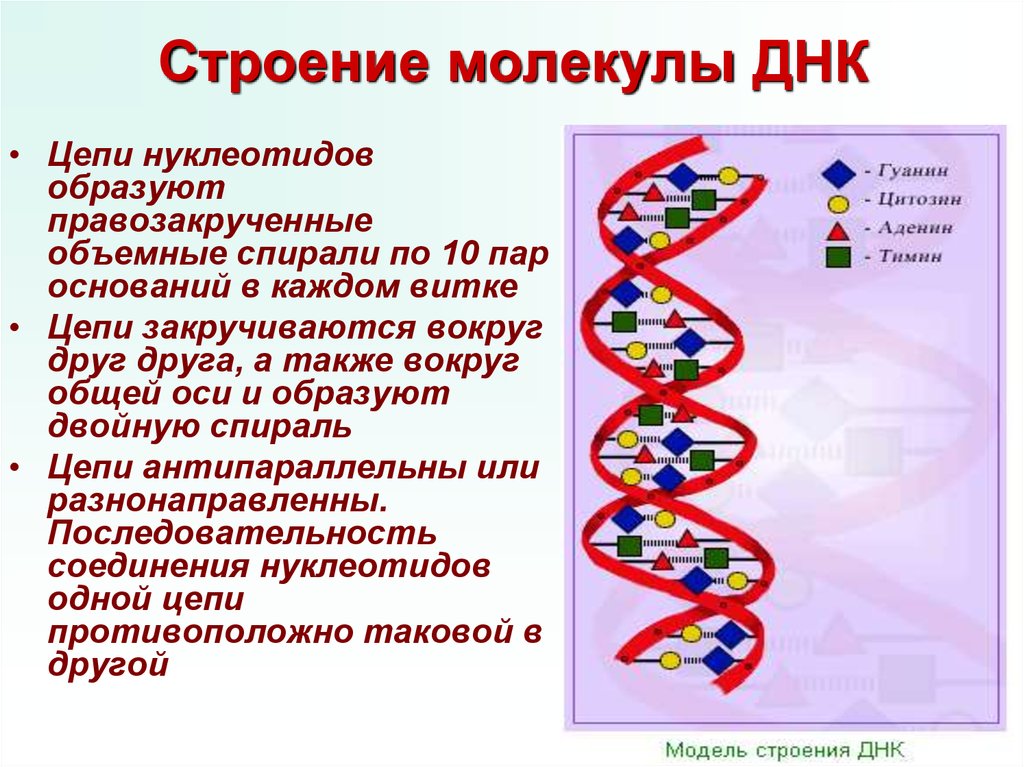
Фосфатная группа одного нуклеотида ковалентно связывается с молекулой сахара следующего нуклеотида и так далее, образуя длинный полимер нуклеотидных мономеров. Сахаро-фосфатные группы выстраиваются в «основу» каждой отдельной нити ДНК, а нуклеотидные основания выступают из этой основы. Атомы углерода пятиуглеродного сахара нумеруются по часовой стрелке от кислорода как 1′, 2′, 3′, 4′ и 5′ (1′ читается как «один штрих»). Фосфатная группа присоединена к 5′-углероду одного нуклеотида и 3′-углероду следующего нуклеотида. В своем естественном состоянии каждая молекула ДНК на самом деле состоит из двух одиночных нитей, соединенных по длине водородными связями между основаниями.
Уотсон и Крик предположили, что ДНК состоит из двух нитей, которые закручиваются друг вокруг друга, образуя правостороннюю спираль, называемую двойной спиралью.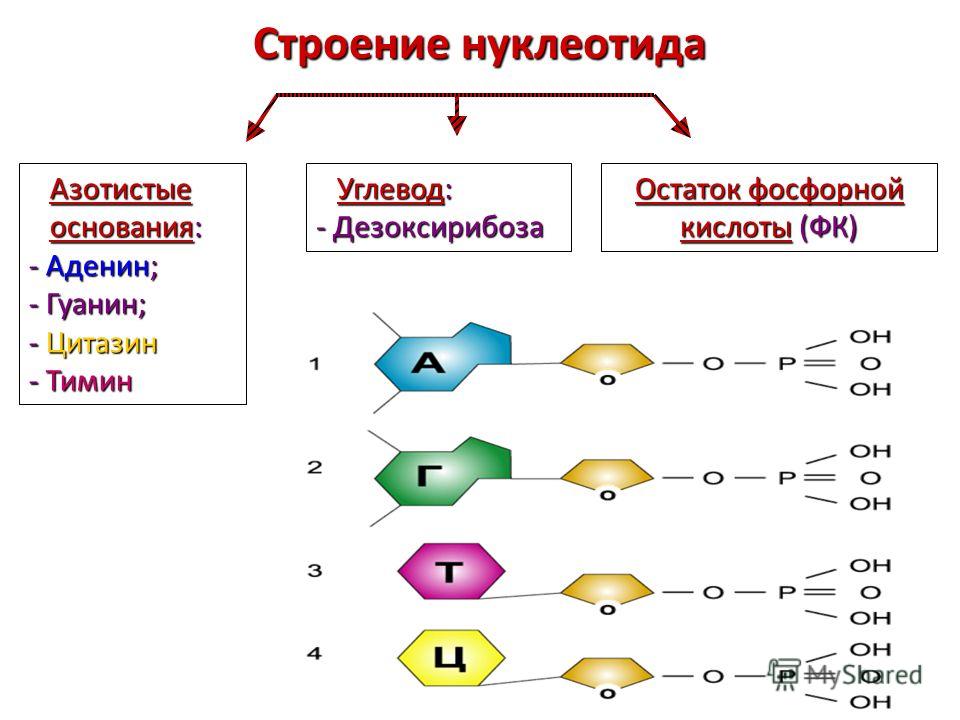 Спаривание оснований происходит между пурином и пиримидином: а именно, пар A с T и G пар с C . Другими словами, аденин и тимин являются комплементарными парами оснований, а цитозин и гуанин также являются комплементарными парами оснований. Это основа правила Чаргаффа; из-за их комплементарности в молекуле ДНК столько же аденина, сколько и тимина, и столько же гуанина, сколько цитозина. Аденин и тимин связаны двумя водородными связями, а цитозин и гуанин — тремя водородными связями. Две нити антипараллельны по своей природе; то есть одна цепь будет иметь 3′-углерод сахара в «верхнем» положении, тогда как другая цепь будет иметь 5′-углерод в верхнем положении. Диаметр двойной спирали ДНК везде одинаков, потому что пурин (два кольца) всегда сочетается с пиримидином (одно кольцо), а их общая длина всегда одинакова. (Рисунок 9.4).
Спаривание оснований происходит между пурином и пиримидином: а именно, пар A с T и G пар с C . Другими словами, аденин и тимин являются комплементарными парами оснований, а цитозин и гуанин также являются комплементарными парами оснований. Это основа правила Чаргаффа; из-за их комплементарности в молекуле ДНК столько же аденина, сколько и тимина, и столько же гуанина, сколько цитозина. Аденин и тимин связаны двумя водородными связями, а цитозин и гуанин — тремя водородными связями. Две нити антипараллельны по своей природе; то есть одна цепь будет иметь 3′-углерод сахара в «верхнем» положении, тогда как другая цепь будет иметь 5′-углерод в верхнем положении. Диаметр двойной спирали ДНК везде одинаков, потому что пурин (два кольца) всегда сочетается с пиримидином (одно кольцо), а их общая длина всегда одинакова. (Рисунок 9.4).
 Как и ДНК, РНК представляет собой полимер нуклеотидов. Каждый нуклеотид в РНК состоит из азотистого основания, пятиуглеродного сахара и фосфатной группы. В случае РНК пятиуглеродным сахаром является рибоза, а не дезоксирибоза. Рибоза имеет гидроксильную группу у 2′-углерода, в отличие от дезоксирибозы, которая имеет только атом водорода (рис. 9)..5). Рис. 9.5. Разница между рибозой, обнаруженной в РНК, и дезоксирибозой, обнаруженной в ДНК, заключается в том, что рибоза имеет гидроксильную группу у 2′-углерода.
Как и ДНК, РНК представляет собой полимер нуклеотидов. Каждый нуклеотид в РНК состоит из азотистого основания, пятиуглеродного сахара и фосфатной группы. В случае РНК пятиуглеродным сахаром является рибоза, а не дезоксирибоза. Рибоза имеет гидроксильную группу у 2′-углерода, в отличие от дезоксирибозы, которая имеет только атом водорода (рис. 9)..5). Рис. 9.5. Разница между рибозой, обнаруженной в РНК, и дезоксирибозой, обнаруженной в ДНК, заключается в том, что рибоза имеет гидроксильную группу у 2′-углерода. нуклеотида РНК содержат азотистые основания аденин, цитозин и гуанин. Однако они не содержат тимина , который вместо заменен урацилом , обозначенным буквой «U». РНК существует в виде одноцепочечной молекулы, а не двухцепочечной спирали. Молекулярные биологи назвали несколько видов РНК в зависимости от их функции. К ним относятся информационная РНК (мРНК), транспортная РНК (тРНК) и рибосомная РНК (рРНК) — молекулы, участвующие в производстве белков из кода ДНК.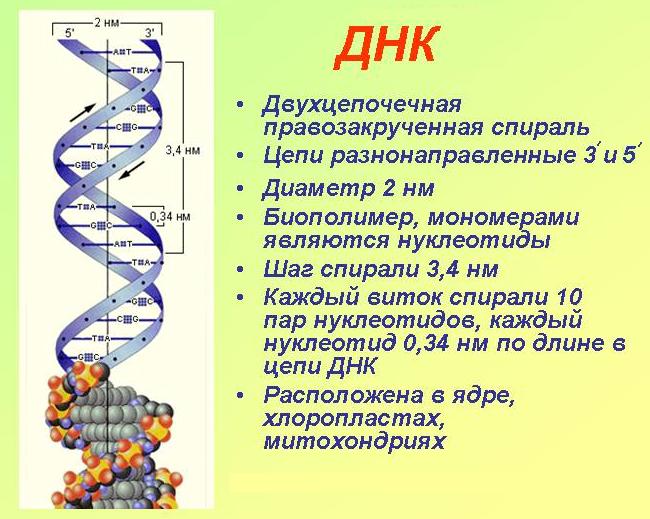
ДНК – рабочая молекула; он должен быть воспроизведен, когда клетка готова к делению, и его необходимо «прочитать», чтобы произвести молекулы, такие как белки, для выполнения функций клетки. По этой причине ДНК защищена и упакована очень специфическим образом. Кроме того, молекулы ДНК могут быть очень длинными. Растянутые из конца в конец, молекулы ДНК в одной человеческой клетке достигают длины около 2 метров . Таким образом, ДНК клетки должна быть упакована очень упорядоченным образом, чтобы соответствовать и функционировать внутри структуры (клетки), невидимой невооруженным глазом. Хромосомы прокариот намного проще, чем у эукариот, по многим своим признакам (рис. 9)..6). Большинство прокариот содержат одну кольцевую хромосому, которая находится в области цитоплазмы, называемой нуклеоидом.
Рис. 9.6. Эукариоты содержат четко очерченное ядро, тогда как у прокариот хромосома лежит в цитоплазме в области, называемой нуклеоидом. Размер генома одной из наиболее хорошо изученных прокариот, Escherichia coli, , составляет 4,6 миллиона пар оснований, которые при растяжении удлинились бы примерно на 1,6 мм.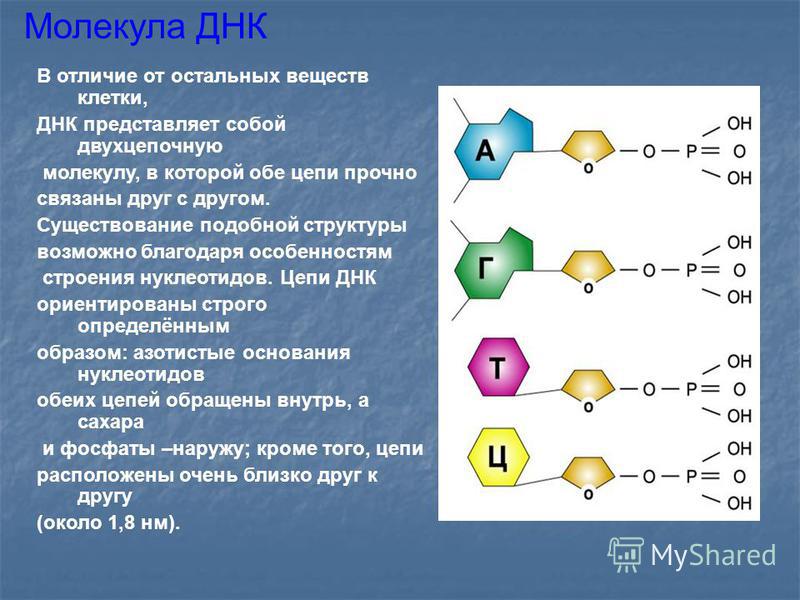 Так как же это помещается внутри маленькой бактериальной клетки? ДНК закручена за пределы двойной спирали в так называемой суперспирализации. Известно, что некоторые белки участвуют в суперспирализации; другие белки и ферменты помогают поддерживать сверхспиральную структуру.
Так как же это помещается внутри маленькой бактериальной клетки? ДНК закручена за пределы двойной спирали в так называемой суперспирализации. Известно, что некоторые белки участвуют в суперспирализации; другие белки и ферменты помогают поддерживать сверхспиральную структуру.
Эукариоты, каждая хромосома которых состоит из линейной молекулы ДНК, используют другой тип стратегии упаковки, чтобы поместить свою ДНК внутри ядра. На самом базовом уровне ДНК обернута вокруг белков, известных как гистоны, с образованием структур, называемых нуклеосомами. ДНК плотно обернута вокруг гистонового ядра. Эта нуклеосома связана со следующей короткой цепью ДНК, свободной от гистонов. Это также известно как структура «бусины на нитке»; нуклеосомы — это «бусинки», а короткие отрезки ДНК между ними — «нитка». Нуклеосомы со свернутой вокруг них ДНК компактно укладываются друг на друга, образуя волокно шириной 30 нм. Это волокно далее сворачивается в более толстую и компактную структуру. На стадии метафазы митоза, когда хромосомы выстраиваются в центр клетки, хромосомы наиболее компактны. Их ширина составляет примерно 700 нм, и они обнаруживаются в ассоциации с каркасными белками.
Их ширина составляет примерно 700 нм, и они обнаруживаются в ассоциации с каркасными белками.
В интерфазе, фазе клеточного цикла между митозами, при которой происходит деконденсация хромосом, хромосомы эукариот имеют две отдельные области, которые можно различить с помощью окрашивания. Есть плотно упакованная область, которая окрашивается в темный цвет, и менее плотная область. Темноокрашенные области обычно содержат неактивные гены и находятся в областях центромер и теломер. Слабоокрашенные области обычно содержат активные гены, при этом ДНК упаковывается вокруг нуклеосом, но не уплотняется дальше.
Рис. 9.7. Эти рисунки иллюстрируют уплотнение эукариотической хромосомы.
Концепция в действии
Посмотрите анимацию упаковки ДНК.
Модель двойной спирали ДНК была предложена Уотсоном и Криком. Молекула ДНК представляет собой полимер нуклеотидов. Каждый нуклеотид состоит из азотистого основания, пятиуглеродного сахара (дезоксирибозы) и фосфатной группы.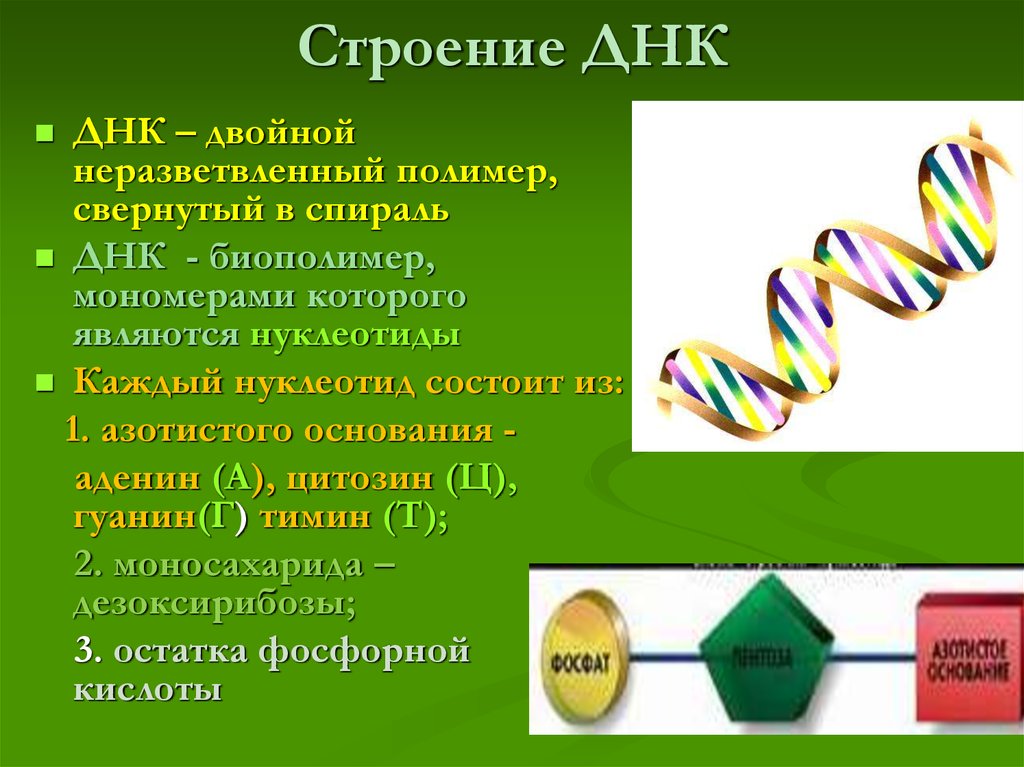 В ДНК четыре азотистых основания, два пуриновых (аденин и гуанин) и два пиримидиновых (цитозин и тимин). Молекула ДНК состоит из двух нитей. Каждая цепь состоит из нуклеотидов, ковалентно связанных друг с другом между фосфатной группой одного и дезоксирибозным сахаром следующего. От этого остова отходят основания. Основания одной нити связаны с основаниями второй нити водородными связями. Аденин всегда связывается с тимином, а цитозин всегда связывается с гуанином. Склеивание заставляет две нити закручиваться вокруг друг друга в форме, называемой двойной спиралью. Рибонуклеиновая кислота (РНК) является второй нуклеиновой кислотой, обнаруженной в клетках. РНК представляет собой одноцепочечный полимер нуклеотидов. Он также отличается от ДНК тем, что содержит сахар рибозу, а не дезоксирибозу, и нуклеотид урацил, а не тимин. Различные молекулы РНК функционируют в процессе формирования белков из генетического кода в ДНК.
В ДНК четыре азотистых основания, два пуриновых (аденин и гуанин) и два пиримидиновых (цитозин и тимин). Молекула ДНК состоит из двух нитей. Каждая цепь состоит из нуклеотидов, ковалентно связанных друг с другом между фосфатной группой одного и дезоксирибозным сахаром следующего. От этого остова отходят основания. Основания одной нити связаны с основаниями второй нити водородными связями. Аденин всегда связывается с тимином, а цитозин всегда связывается с гуанином. Склеивание заставляет две нити закручиваться вокруг друг друга в форме, называемой двойной спиралью. Рибонуклеиновая кислота (РНК) является второй нуклеиновой кислотой, обнаруженной в клетках. РНК представляет собой одноцепочечный полимер нуклеотидов. Он также отличается от ДНК тем, что содержит сахар рибозу, а не дезоксирибозу, и нуклеотид урацил, а не тимин. Различные молекулы РНК функционируют в процессе формирования белков из генетического кода в ДНК.
Прокариоты содержат одну двухцепочечную кольцевую хромосому. Эукариоты содержат двухцепочечные линейные молекулы ДНК, упакованные в хромосомы. Спираль ДНК оборачивается вокруг белков, образуя нуклеосомы. Белковые спирали еще больше скручены, и во время митоза и мейоза хромосомы еще больше скручены, чтобы облегчить их движение. Хромосомы имеют две отдельные области, которые можно различить по окрашиванию, отражающему различную степень упаковки и определяемому тем, экспрессируется ли ДНК в области (эухроматин) или нет (гетерохроматин).
Эукариоты содержат двухцепочечные линейные молекулы ДНК, упакованные в хромосомы. Спираль ДНК оборачивается вокруг белков, образуя нуклеосомы. Белковые спирали еще больше скручены, и во время митоза и мейоза хромосомы еще больше скручены, чтобы облегчить их движение. Хромосомы имеют две отдельные области, которые можно различить по окрашиванию, отражающему различную степень упаковки и определяемому тем, экспрессируется ли ДНК в области (эухроматин) или нет (гетерохроматин).
Глоссарий
дезоксирибоза: пятиуглеродная молекула сахара с атомом водорода, а не гидроксильной группой во 2′-положении; сахарный компонент ДНК нуклеотиды
двойная спираль: молекулярная форма ДНК, в которой две нити нуклеотидов закручиваются вокруг друг друга в форме спирали
азотистое основание: азотсодержащая молекула, действующая как основание; часто ссылаясь на один из пуриновых или пиримидиновых компонентов нуклеиновых кислот
фосфатная группа: молекулярная группа, состоящая из центрального атома фосфора, связанного с четырьмя атомами кислорода
Лицензия
Concepts of Biology — 1st Canadian Edition Чарльза Молнара и Джейн Гейр распространяется под лицензией Creative Commons Attribution 4.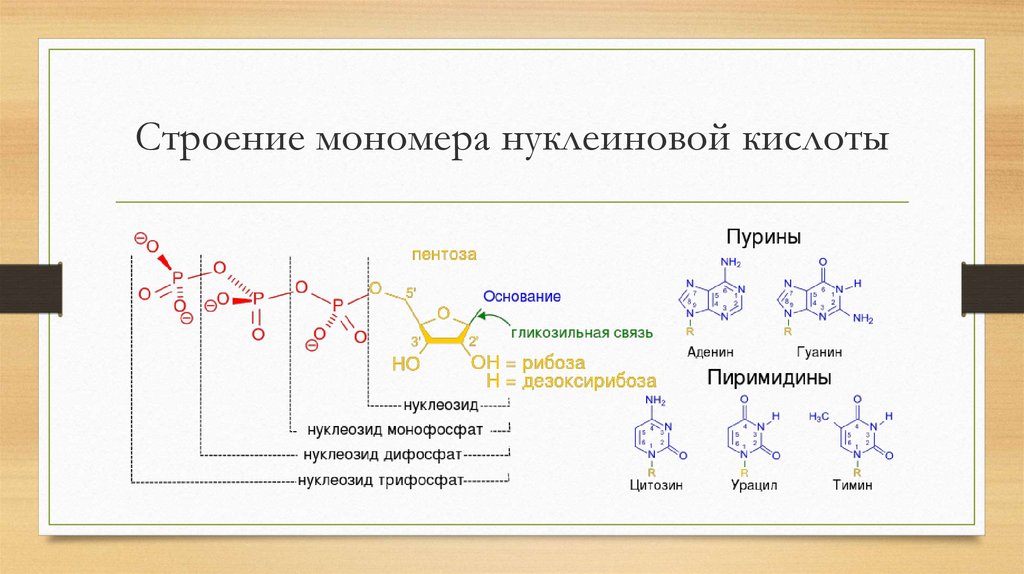 0 International License, если не указано иное.
0 International License, если не указано иное.
Поделиться этой книгой
Поделиться в Твиттере
Нуклеиновые кислоты Элементы и мономеры
Нуклеиновые кислоты представляют собой биосоединения, необходимые для живых организмов. Встречающиеся в двух формах — дезоксирибонуклеиновой кислоты (ДНК) и рибонуклеиновой кислоты (РНК) — эти полимерные цепи состоят из одних и тех же основных элементов и сходных мономерных нуклеотидов, но со специфическими различиями, касающимися формы и функции.
Элементы нуклеиновой кислоты
Каждый нуклеотидный мономер и, следовательно, каждый полимер нуклеиновой кислоты состоит из группы из пяти элементов. Эти элементы связываются с образованием моносахаридов, фосфатных групп и азотистых оснований, также известных как азотистые основания. И в РНК, и в ДНК фосфатная группа имеет одинаковую форму, но есть различия в азотистых основаниях и молекулах сахаров. Пять элементов, необходимых для построения цепи нуклеиновой кислоты, — это углерод, водород, кислород, азот и фосфор.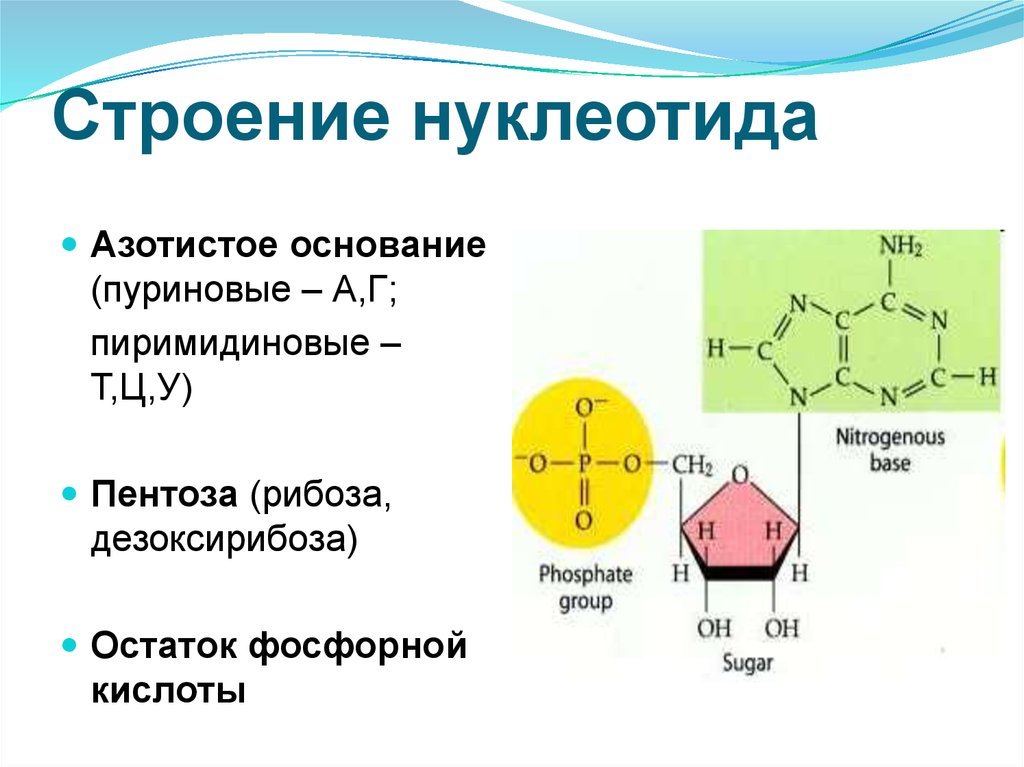 Добавление фосфора отличает нуклеиновую кислоту от других категорий биосоединений, а именно от углеводов, липидов и белков.
Добавление фосфора отличает нуклеиновую кислоту от других категорий биосоединений, а именно от углеводов, липидов и белков.
Мономеры нуклеиновой кислоты
Химические формулы мономера нуклеиновой кислоты показывают количество каждого элемента. Нуклеотидные мономеры называются в соответствии с типом азотистого основания, которое они содержат. В свободном состоянии эти мономеры могут иметь дополнительные фосфатные группы и находиться в форме дифосфатов, трифосфатов или полифосфатов. При образовании полимера РНК или ДНК высвобождаются дополнительные фосфатные группы, и к моносахариду остается только одна. Комбинация рибозы или дезоксирибозы и фосфатной группы образует сахарофосфатный остов. Азотистое основание присоединено к молекуле сахара. Добавление фосфатной группы к нуклеозиду, образованному сахаром и азотистым основанием, образует нуклеотид. Таким образом, нуклеотидный мономер имеет различные структуры с определенными названиями: сахаро-фосфатный остов, нуклеозид и отдельные молекулы азотистого основания, пентозного сахара и фосфатной группы.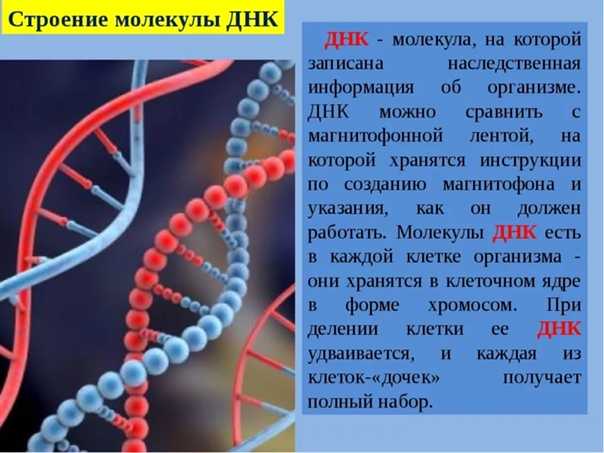
В нуклеиновых кислотах пентозные сахара представлены в двух различных формах: рибозе и дезоксирибозе. Первый обладает дополнительной молекулой кислорода, которая в соединении с водородом образует гидроксильную группу. Это свойство отсутствует у дезоксирибозы.
Азотистые основания классифицируются по размеру. Формы с двойным кольцом, называемые пуринами, крупнее и длиннее и содержат пять атомов азота. Однокольцевые формы, известные как пиримидины, содержат от двух до трех атомов азота, они меньше и короче. Это важно для двухцепочечной характеристики ДНК и процесса трансляции, поскольку возможны только определенные пары азотистых оснований (пары Уотсона-Крика). Они удерживают две нити на равном расстоянии друг от друга. Мнемоника, помогающая запомнить, какие нуклеотиды принадлежат к какой группе, представляет собой фразу «9».0027 Pur e A s G старый’; само собой разумеется, что остальные основания принадлежат пиримидиновой группе.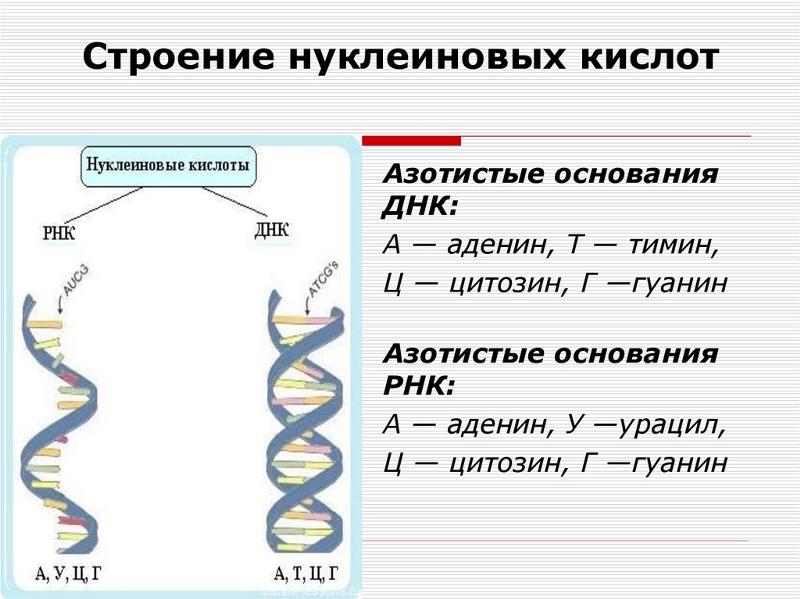 Это также говорит нам о том, что аденин и гуанин не могут вместе образовывать двухцепочечную связь. В РНК возможны и другие комбинации оснований, известные как пары, отличные от пар Уотсона-Крика.
Это также говорит нам о том, что аденин и гуанин не могут вместе образовывать двухцепочечную связь. В РНК возможны и другие комбинации оснований, известные как пары, отличные от пар Уотсона-Крика.
В парах Уотсона-Крика более крупные основания, аденин и гуанин никогда не образуют пары друг с другом. Точно так же пурины не связаны друг с другом (цитозин, тимин и урацил). В ДНК аденин образует пары только с тимином, а гуанин — с цитозином. В РНК аденин сочетается с урацилом, а гуанин с цитозином.
На следующих изображениях показана химическая структура каждого типа мономера, где четко определены пентагональная форма моносахарида и присоединенная к нему фосфатная группа и конкретное азотистое основание.
Adenosine Monophosphate (AMP): C
10 H 14 N 5 O 7 P This chemical formula represents the sum of the purine base adenine (C 5 H 5 N 5 ), рибоза (C 5 H 10 O 5 ) и фосфорной кислоты (H 3 PO 4 ), где в результате реакций конденсации в местах связывания молекул теряются две молекулы воды (2H 2 0).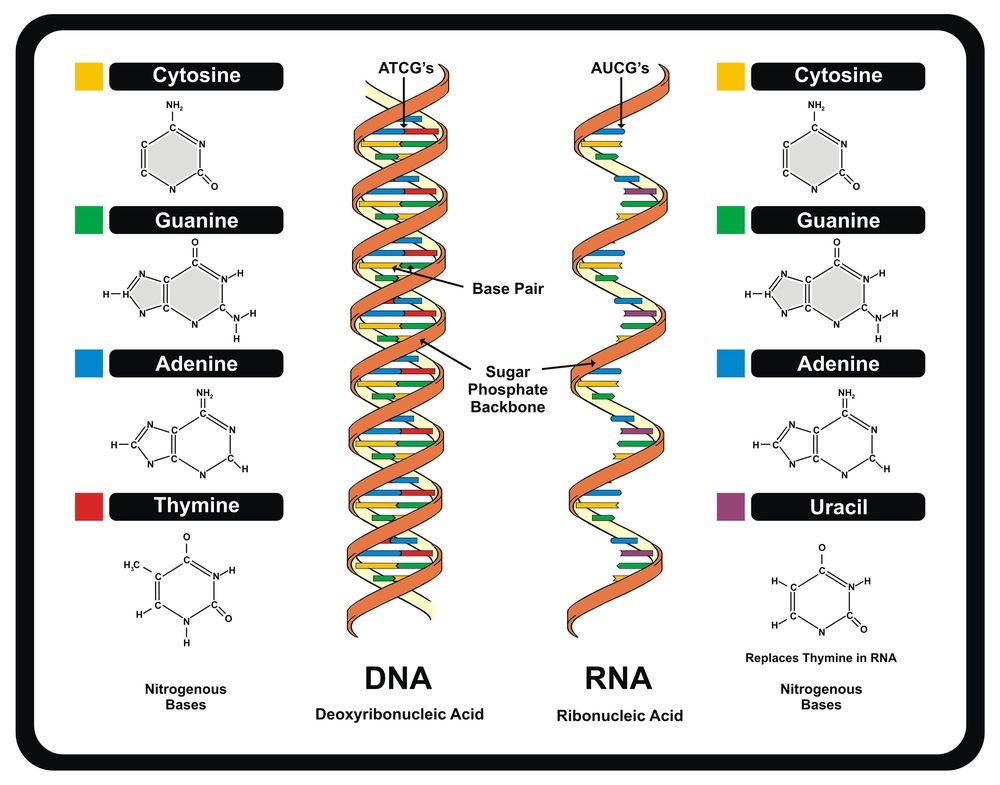 Это форма РНК.
Это форма РНК.
Дезоксиденозин -монофосфат (DEAM): C
10 H 14 N 5 O 6 P. Эта химическая формула представляет сумму базовой базы пурина (C 5 5 5 . 5 ), дезоксирибоза (C 5 H 10 O 4 ) и фосфорной кислоты (H 3 PO 4 ), где реакции конденсации в местах связывания молекул теряют две молекулы воды (2H 2 0). Это форма ДНК.
Guanosine Monophosphate (GMP): C
10 H 14 N 5 O 8 PThe sum of the purine base guanine (C 5 H 5 N 5 O ), рибозу (C 5 H 10 O 5 ) и фосфорную кислоту (H 3 PO 4 ), где реакции конденсации в местах связывания молекул теряют две молекулы воды (2H 2 0). Это форма РНК.
Дезоксигуанозин -монофосфат (DGMP): C
10 H 14 N 5 O 7 P Сумма пуриновой базы гуанина (C 5 5 5 5 5 5 5 5 5 5 5 5 5 5 5 5 5 5 5 5 5 5 5 5 5 5 . ), дезоксирибозу (C 5 H 10 O 4 ) и фосфорную кислоту (H 3 PO 4 ), где реакции конденсации в местах связи молекул теряют две молекулы воды (2H 2 0). Это форма ДНК.
), дезоксирибозу (C 5 H 10 O 4 ) и фосфорную кислоту (H 3 PO 4 ), где реакции конденсации в местах связи молекул теряют две молекулы воды (2H 2 0). Это форма ДНК.
Uridine Monophosphate (UMP): C
9 H 13 N 2 O 9 PThe sum of the pyrimidine base uracil (C 4 H 4 N 2 O 2 ), рибозу (C 5 H 10 O 5 ) и фосфорную кислоту (H 3 PO 4 ), где реакции конденсации в местах связывания молекул теряют две молекулы воды (2H 2 0). Встречается только в РНК.
Cytidine Monophosphate (CMP): C
9 H 14 N 3 O 8 P The sum of the pyrimidine base cytosine (C 4 H 5 N 3 O ), рибозу (C 5 H 10 O 5 ) и фосфорную кислоту (H 3 PO 4 ), где реакции конденсации в местах связывания молекул теряют две молекулы воды (2H 2 0 ).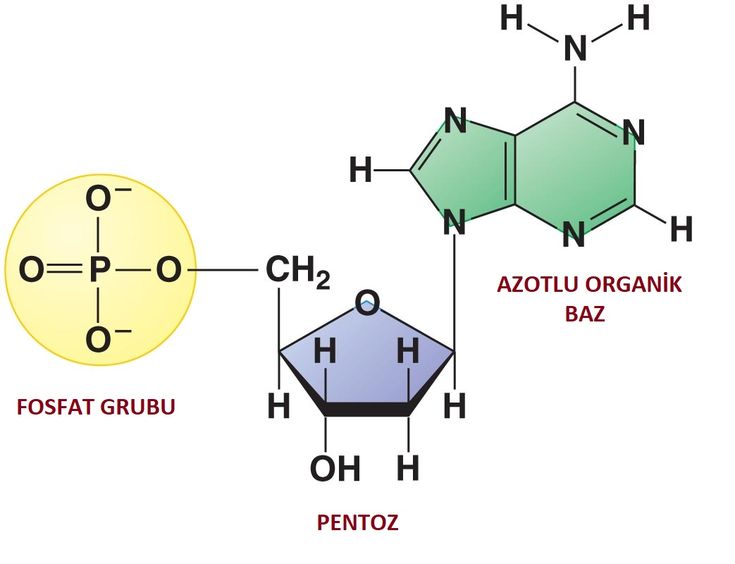
Leave A Comment