
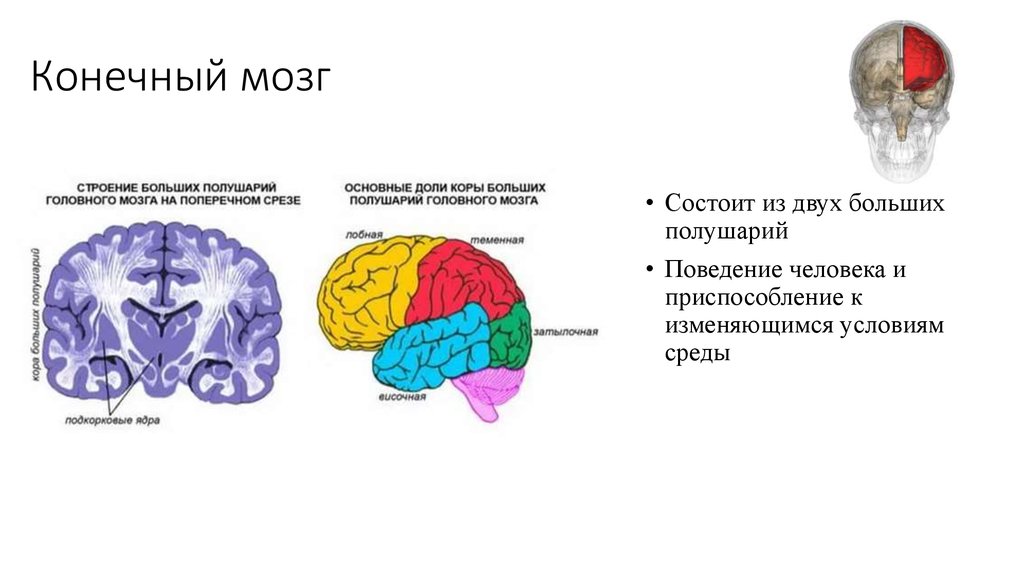
Кора больших полушарий головного мозга.
Кора мозга – пласт серого вещества на поверхности больших полушарий, толщиной 2-5 мм, образующий многочисленные борозды, извилины значительно увеличивающие ее площадь. Кора образована телами нейронов и глиальных клеток, расположенных послойно («экранный» тип организации). Под ней лежит белое вещество, представленное нервными волокнами.
Кора представляет собой наиболее молодой филогенетически и наиболее сложный по морфофункциональной организации отдел мозга. Это место высшего анализа и синтеза всей информации поступающей в мозг. Здесь происходит интеграция всех сложных форм поведения. Кора мозга отвечает за сознание, мышление, память, «эвристическую деятельность» (способность к обобщениям, открытиям). В коре содержится более 10 млрд. нейронов и 100 млрд. глиальных клеток.
Нейроны коры по количеству отростков только
мультиполярные, а по их месту в
рефлекторных дугах и выполняемым
функциям все они вставочные, ассоциативные.
Среди непирамидных нейронов
наиболее часто встречаются звёздчатые
и веретеновидные клетки.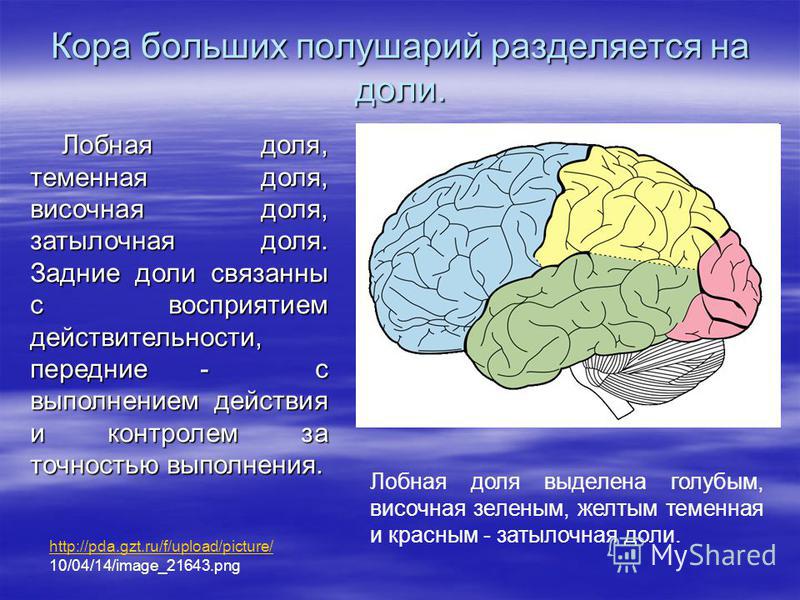 Звёздчатые нейроны — это мелкие клетки с короткими
сильно ветвящимися дендритами и аксонами,
образующими внутрикорковые связи. Одни
из них оказывают тормозное, а другие —
возбуждающее влияние на пирамидные
нейроны. Веретеновидные нейроны имеют длинный аксон, который
может идти в вертикальном, или
горизонтальном направлении.
Звёздчатые нейроны — это мелкие клетки с короткими
сильно ветвящимися дендритами и аксонами,
образующими внутрикорковые связи. Одни
из них оказывают тормозное, а другие —
возбуждающее влияние на пирамидные
нейроны. Веретеновидные нейроны имеют длинный аксон, который
может идти в вертикальном, или
горизонтальном направлении.
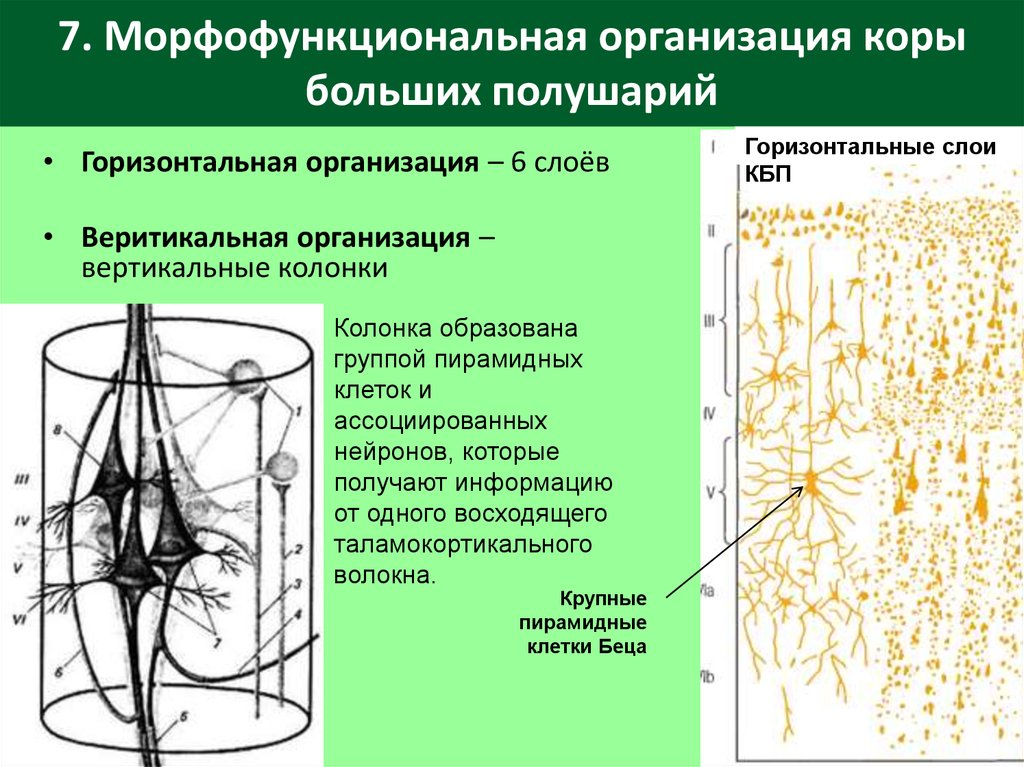
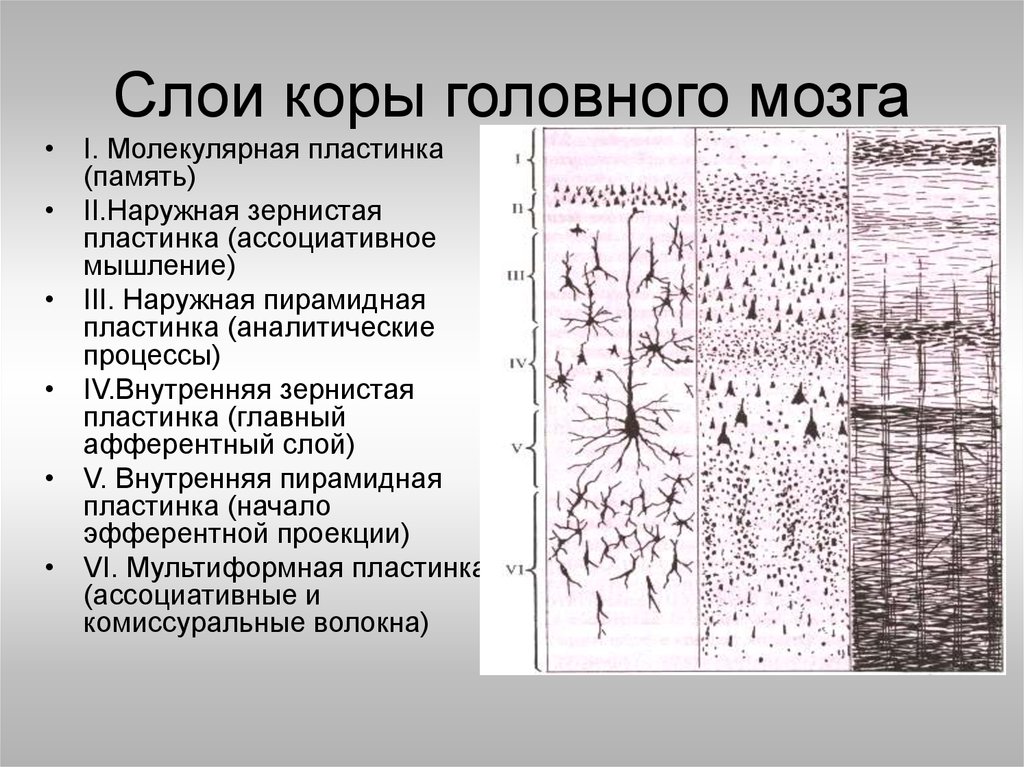
Кора построена по экранному типу, то есть нейроны, сходные по структуре и функции расположены слоями (рис. 9-7). Таких слоёв в коре шесть:
1. Молекулярный слой – самый наружный. В нём находится сплетение нервных волокон, расположенных параллельно поверхности коры. Основную массу этих волокон составляют ветвления апикальных дендритов пирамидных нейронов нижележащих слоёв коры. Сюда же приходят афферентные волокна от зрительных бугров, регулирующих возбудимость корковых нейронов. Нейроны в молекулярном слое в основном мелкие, веретеновидные.
2. Наружный
зернистый слой.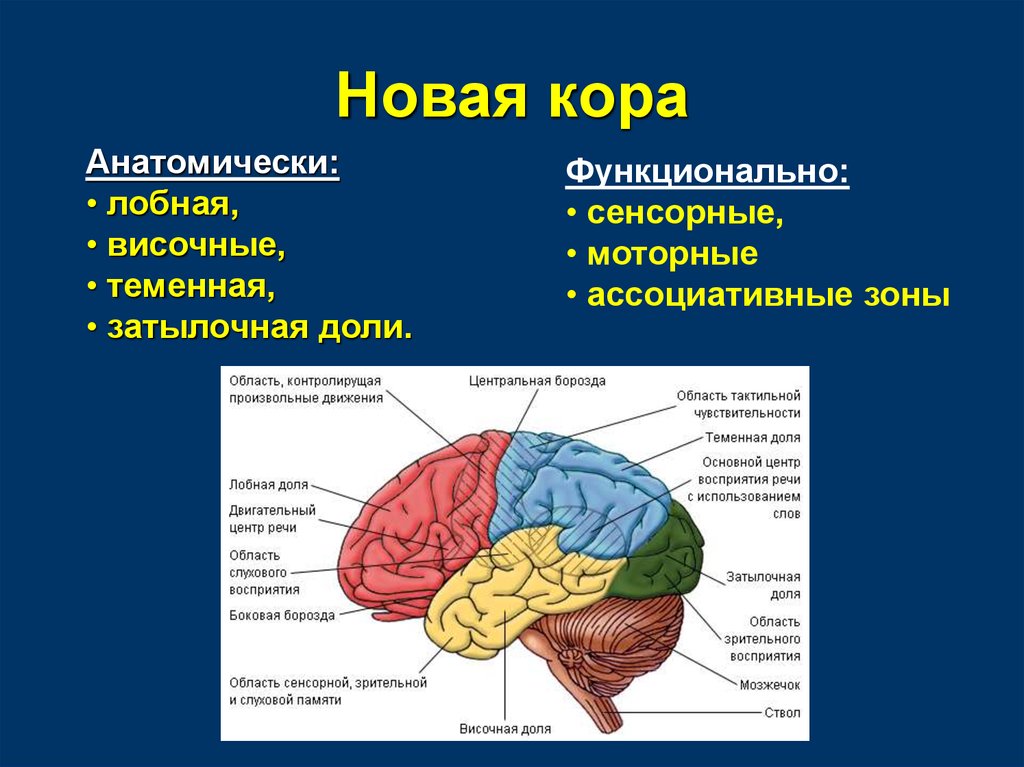 Состоит
из большого числа звёздчатых клеток.
Их дендриты идут в молекулярный слой и
образуют синапсы с таламо-кортикальными
афферентными нервными волокнами.
Боковые дендриты связываются с
соседними нейронами этого же слоя.
Аксоны образуют ассоциативные
волокна, которые идут через белое
вещество в соседние участки коры и
там образуют синапсы.
Состоит
из большого числа звёздчатых клеток.
Их дендриты идут в молекулярный слой и
образуют синапсы с таламо-кортикальными
афферентными нервными волокнами.
Боковые дендриты связываются с
соседними нейронами этого же слоя.
Аксоны образуют ассоциативные
волокна, которые идут через белое
вещество в соседние участки коры и
там образуют синапсы.
3. Наружный слой пирамидных нейронов (пирамидный слой). Он образован пирамидными нейронами средней величины. Так же, как у нейронов второго слоя, их дендриты идут в молекулярный слой, а аксоны – в белое вещество.
4. Внутренний зернистый слой. Он содержит много звёздчатых нейронов. Это ассоциативные, афферентные нейроны. Они образуют многочисленные связи с другими нейронами коры. Здесь расположен ещё один слой горизонтальных волокон.
5. Внутренний
слой пирамидных нейронов (ганглионарный слой). Он образован
крупными пирамидными нейронами.
Последние особенно велики в моторной
коре (прецентральной извилине), где
имеют размеры до 140 мкм и называются
клетками Беца. Их апикальные дендриты
поднимаются в молекулярный слой,
боковые дендриты образуют связи с
соседними клетками Беца, а аксоны –
проекционные эфферентные волокна,
идущие в продолговатый и спинной мозг.
Их апикальные дендриты
поднимаются в молекулярный слой,
боковые дендриты образуют связи с
соседними клетками Беца, а аксоны –
проекционные эфферентные волокна,
идущие в продолговатый и спинной мозг.
6. Слой веретеновидных нейронов (слой полиморфных клеток) состоит в основном из веретеновидных нейронов. Их дендриты идут в молекулярный слой, а аксоны – к зрительным буграм.
Шестислойный тип строения коры характерен для всей коры, однако в разных её участках выраженность слоёв, а также форма и расположение нейронов, нервных волокон значительно различаются. По этим признакам К. Бродман выделил в коре 50 цитоархитектонических полей. Эти поля также различаются по функции и обмену веществ.
Специфическую
организацию нейронов называют цитоархитектоникой. Так, в сенсорных
зонах коры пирамидный и ганглиозный
слои выражены слабо, а зернистые слои
— хорошо. Такой тип коры называется гранулярным. В моторных
зонах, напротив, зернистые слои развиты
плохо, а пирамидные хорошо.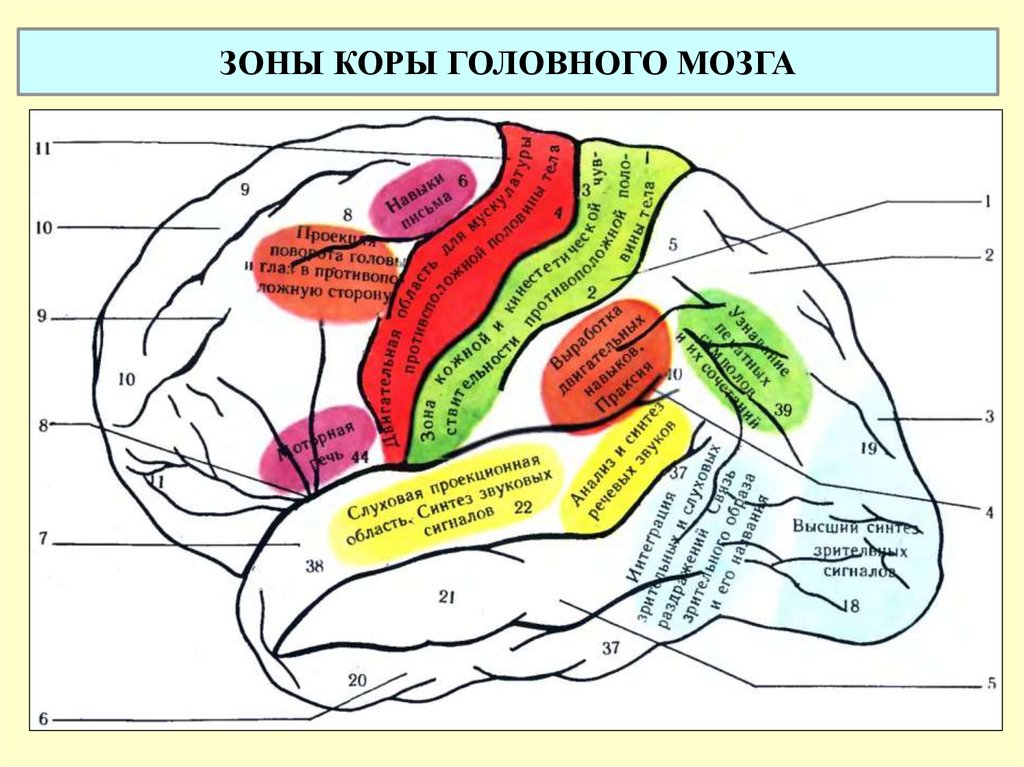 Это агранулярный
тип коры.
Это агранулярный
тип коры.
Кроме того, существует понятиемиелоархитектоника. Это определённая организация нервных волокон. Так, в коре мозга различают вертикальные и три горизонтальных пучка миелиновых нервных волокон. Среди нервных волокон коры мозга различают
Рис. 9-7. Кора больших полуша-рий головного моз-га человека.
А, Б. Расположение клеток (цитоархитектоника).
В. Расположение миелиновых волокон (миелоархитектоника).
1. Молекулярный слой.
2. Наружный зернистый слой.
3. Пирамидный слой.
5. Ганглионарный
слой.
Ганглионарный
слой.
6. Слой полиморф-ных клеток.
7. Наружная полоска Байарже.8 Внутренняя полоска Байарже. (По Ю. И. Афанасьеву).
Все нейроны коры по их месту в рефлекторных дугах являются ассоциативными — расположены между другими нейронами. Однако по направлению передачи нервного импульса нейроны 1, 2 и 4-го слоёв можно условно назвать афферентными, т. к. они воспринимают сигналы от нижележащих отделов мозга, а нейроны 3, 5 и 6-го слоёв – эфферентными, поскольку по их аксонам импульсы идут в другие участки коры и нижележащие отделы мозга.
Строение нервной системы
Единая нервная система для удобства изучения подразделяется на центральную (головной и спинной мозг) и периферическую (черепно- и спинномозговые нервы, их сплетения и узлы), а также соматическую и вегетативную (или автономную).
Соматическая нервная система осуществляет преимущественно связь организма с внешней средой: восприятие раздражений, регуляцию движений поперечнополосатой мускулатуры скелета и др.
Вегетативная — регулирует обмен веществ и работу внутренних органов: биение сердца, перистальтическое сокращение кишечника, секрецию различных желез и т. п. Обе они функционируют в тесном взаимодействии, однако вегетативная система обладает некоторой самостоятельностью (автономностью), управляя многими непроизвольными функциями.
Спинной мозг: слева — общий план строения;
справа — поперечные разрезы разных отделов
Спинной мозг находится в позвоночном канале и имеет вид белого тяжа, протянувшегося от затылочного отверстия до поясницы. На поперечном разрезе видно, что спинной мозг состоит из белого (снаружи) и серого (внутри) вещества. Серое вещество состоит из тел нервных клеток и имеет на поперечном слое форму бабочки, от расправленных «крыльев» которой отходят два передних и два задних рога. В передних рогах находятся центробежные нейроны, от которых отходят двигательные нервы. Задние рога включают нервные клетки (промежуточные нейроны), к которым подходят отростки чувствительных нейронов, лежащих в утолщениях задних корешков.
Каждая пара нервов иннервирует определенную группу мышц и соответствующий участок кожи.
Белое вещество образовано отростками нервных клеток (нервными волокнами), объединенными в проводящие пути, которые тянутся вдоль спинного мозга» соединяя как отдельные его сегменты друг с другом, так и спинной мозг с головным. Одни проводящие пути называются восходящими, или чувствительными, передающими возбуждение в головной мозг, другие — нисходящими, или двигательными, которые проводят импульсы от головного мозга к определенным сегментам спинного мозга.
Спинной мозг выполняет две функции: рефлекторную и проводниковую. Деятельность спинного мозга находится под контролем головного мозга.
Головной мозг расположен в мозговом отделе черепа. Средняя его масса 1300–1400 г. После рождения человека рост мозга продолжается до 20 лет. Состоит из пяти отделов; переднего (большие полушария), промежуточного, среднего, заднего и продолговатого мозга.
Состоит из пяти отделов; переднего (большие полушария), промежуточного, среднего, заднего и продолговатого мозга.
Полушария (наиболее новая в эволюционном отношении часть) достигают у человека высокого развития, составляя 80% массы мозга.
Филогенетически более древняя часть — ствол головного мозга. Ствол включает продолговатый мозг, мозговой (варолиев) мост, средний и промежуточный мозг. В белом веществе ствола залегают многочисленные ядра серого вещества. Ядра 12–ти пар черепно-мозговых нервов также лежат в стволе мозга. Стволовая часть мозга покрыта полушариями головного мозга.
Продолговатый мозг — продолжение спинного и повторяет его строение: на передней и задней поверхности здесь также залегают борозды. Он состоит из белого вещества (проводящих пучков), где рассеяны скопления серого вещества — ядра, от которых берут начало черепные нервы. Сверху и с боков почти весь продолговатый мозг покрыт большими полушариями и мозжечком. В сером веществе продолговатого мозга залегают жизненно важные центры, регулирующие сердечную деятельность, дыхание, глотание, осуществляющие защитные рефлексы (чихание, кашель, рвота, слезоотделение), секрецию слюны, желудочного и поджелудочного сока и др. Повреждение продолговатого мозга может быть причиной смерти вследствие прекращения сердечной деятельности и дыхания.
В сером веществе продолговатого мозга залегают жизненно важные центры, регулирующие сердечную деятельность, дыхание, глотание, осуществляющие защитные рефлексы (чихание, кашель, рвота, слезоотделение), секрецию слюны, желудочного и поджелудочного сока и др. Повреждение продолговатого мозга может быть причиной смерти вследствие прекращения сердечной деятельности и дыхания.
Задний мозг включает варолиев мост и мозжечок. В веществе варолиева моста находятся ядра тройничного, отводящего, лицевого и слухового нервов.
Мозжечок — его поверхность покрыта серым веществом, под ним находится белое вещество, в котором имеются ядра — скопления белого вещества. Основная функция мозжечка — координация движений, определяющая их четкость, плавность и сохранение равновесия тела, а также поддержание тонуса мышц. Контролирует деятельность мозжечка кора больших полушарий.
Средний мозг расположен впереди варолиева моста и представлен четверохолмием и ножками мозга. В ножках мозга продолжаются проводящие пути от продолговатого мозга и варолиева моста к большим полушариям.
В ножках мозга продолжаются проводящие пути от продолговатого мозга и варолиева моста к большим полушариям.
Средний мозг играет важную роль в регуляции тонуса и в осуществлении рефлексов, благодаря которым возможны стояние и ходьба.
Промежуточный мозг занимает в стволе самое высокое положение. Состоит из зрительных бугров (таламус) и подбугровой области (гипоталамус). Зрительные бугры регулируют ритм корковой активности и участвуют в образовании условных рефлексов, эмоций и т. Д.
Подбугровая область связана со всеми отделами центральной нервной системы и с железами внутренней секреции. Она является регулятором обмена веществ и температуры тела, постоянства внутренней среды организма и функций пищеварительной, сердечнососудистой, мочеполовой систем, а также желез внутренней секреции.
Передний мозг у человека состоит из сильно развитых полушарий и соединяющей их средней части. Правое и левое полушария отделены друг от друга глубокой щелью, на дне которой лежит мозолистое тело.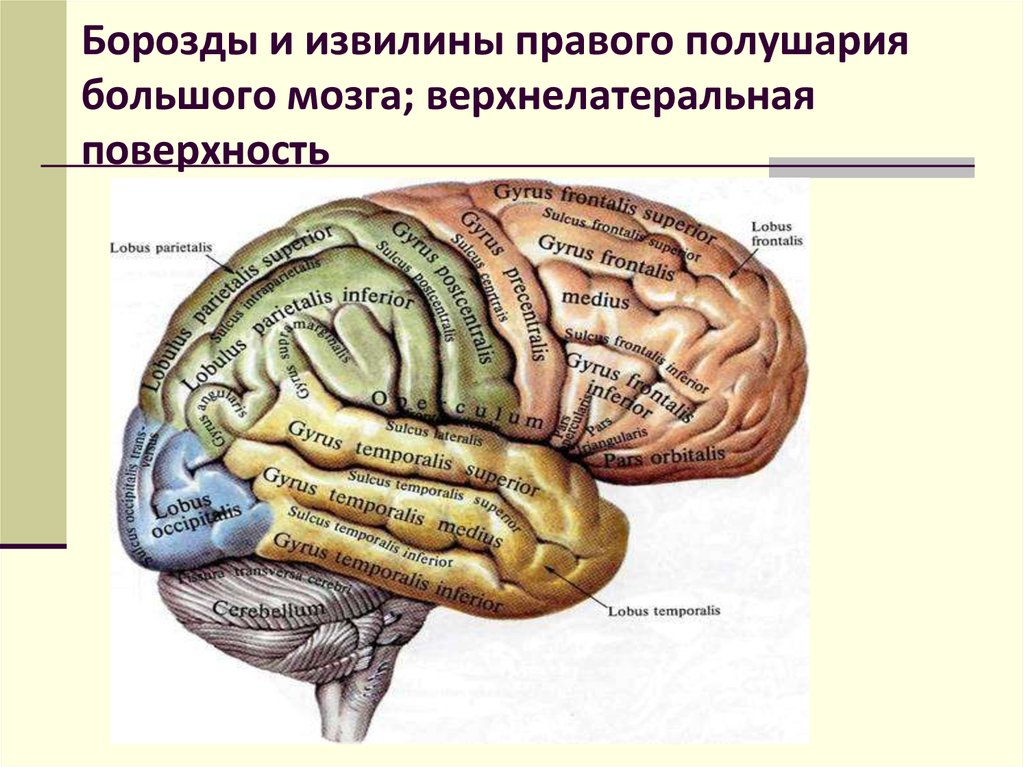 Поверхность больших полушарий образована серым веществом — корой, под которой находится белое вещество с подкорковыми ядрами. Общая поверхность коры больших полушарий составляет 2000–2500 см2, толщина ее 2,5–3 мм. В ней насчитывается от 12 до 18 млрд нейронов, расположенных шестью слоями. Больше 2/3 поверхности коры скрыто в глубоких бороздах между выпуклыми извилинами. Три главные борозды — центральная, боковая и теменно-затылочная — делят каждое полушарие на четыре доли: лобную, теменную, затылочную и височную .
Поверхность больших полушарий образована серым веществом — корой, под которой находится белое вещество с подкорковыми ядрами. Общая поверхность коры больших полушарий составляет 2000–2500 см2, толщина ее 2,5–3 мм. В ней насчитывается от 12 до 18 млрд нейронов, расположенных шестью слоями. Больше 2/3 поверхности коры скрыто в глубоких бороздах между выпуклыми извилинами. Три главные борозды — центральная, боковая и теменно-затылочная — делят каждое полушарие на четыре доли: лобную, теменную, затылочную и височную .
Большие полушария головного мозга
Нижняя поверхность полушарий и стволовая часть мозга называется основанием мозга.
Чтобы понять, как функционирует кора больших полушарий головного мозга, нужно вспомнить, что в организме человека имеется большое количество разнообразных рецепторов, способных улавливать самые незначительные изменения во внешней и внутренней среде.
Рецепторы, расположенные в коже, реагируют на изменения во внешней среде.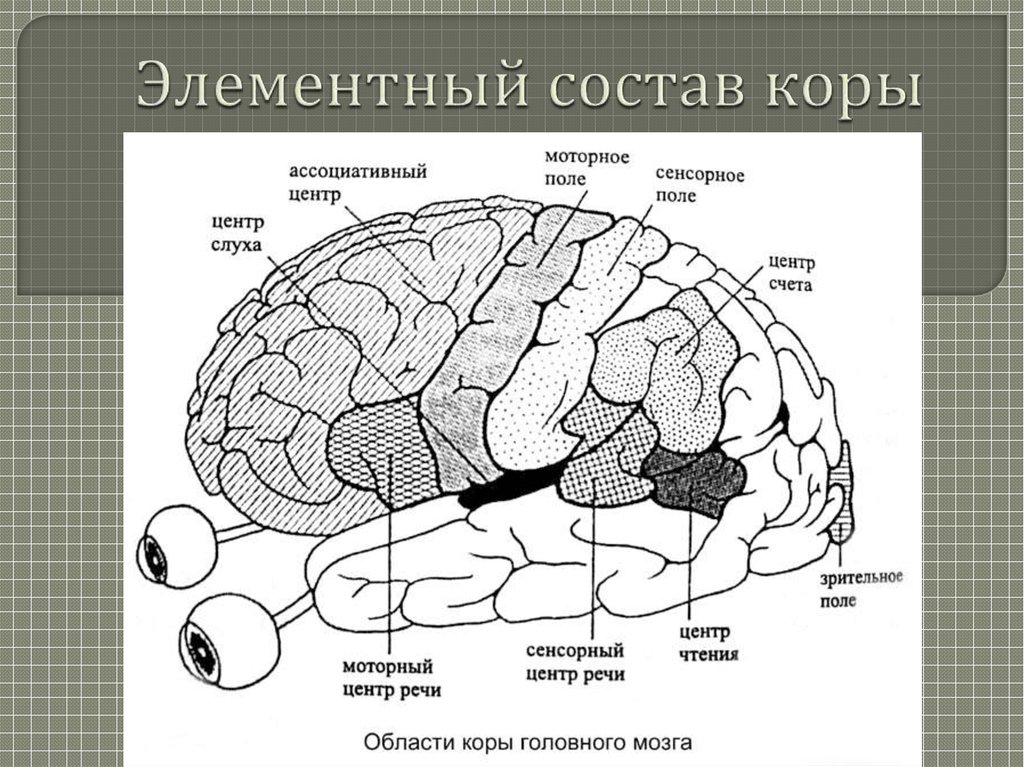 В мышцах и сухожилиях находятся рецепторы, сигнализирующие в мозг о степени натяжения мышц, движениях суставов. Имеются рецепторы, реагирующие на изменения химического и газового состава крови, осмотического давления, температуры и др. В рецепторе раздражение преобразуется в нервные импульсы. По чувствительным нервным путям импульсы проводятся к соответствующим чувствительным зонам коры головного мозга, где и формируется специфическое ощущение — зрительное, обонятельное и др.
В мышцах и сухожилиях находятся рецепторы, сигнализирующие в мозг о степени натяжения мышц, движениях суставов. Имеются рецепторы, реагирующие на изменения химического и газового состава крови, осмотического давления, температуры и др. В рецепторе раздражение преобразуется в нервные импульсы. По чувствительным нервным путям импульсы проводятся к соответствующим чувствительным зонам коры головного мозга, где и формируется специфическое ощущение — зрительное, обонятельное и др.
Функциональную систему, состоящую из рецептора, чувствительного проводящего пути и зоны коры, куда проецируется данный вид чувствительности, И. П. Павлов назвал анализатором.
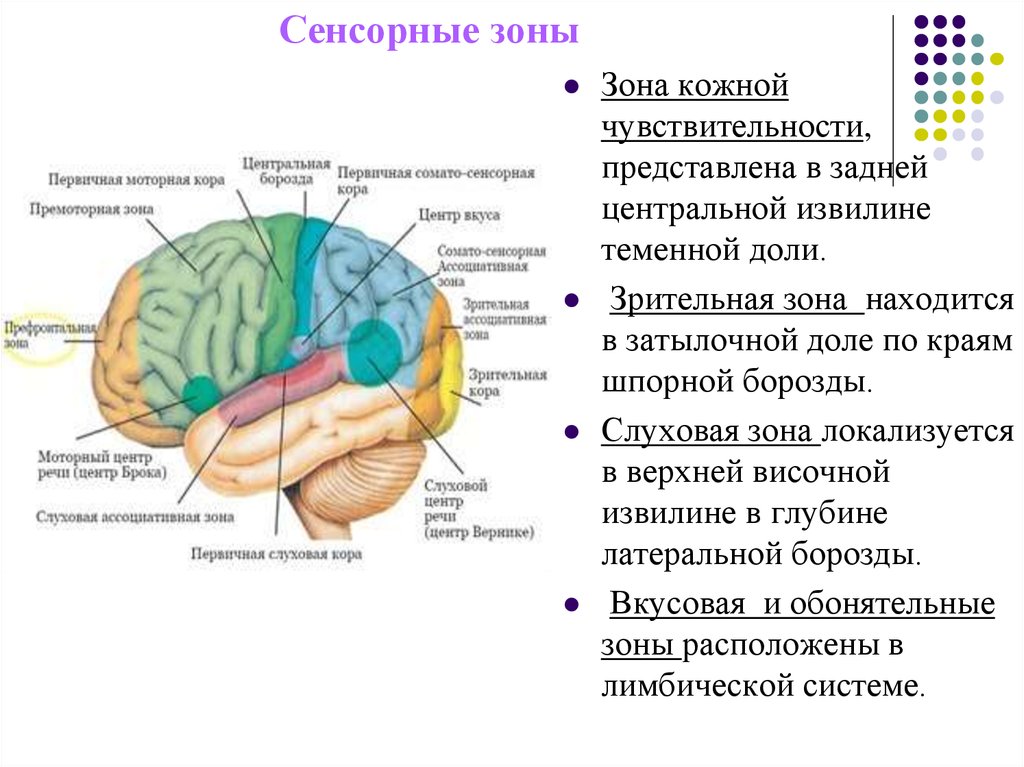
Анализ и синтез полученной информации осуществляется в строго определенном участке — зоне коры боль-
Важнейшие зоны коры — двигательная, чувствительная, зрительная, слуховая, обонятельная.
Двигательная зона расположена в передней центральной извилине впереди центральной борозды лобной доли, зона кожно-мышечной чувствительности — позади центральной борозды, в задней центральной извилине теменной доли. Зрительная зона сосредоточена в затылочной зоне, слуховая — в верхней височной извилине височной доли, обонятельная и вкусовая — в переднем отделе височной доли.
Зрительная зона сосредоточена в затылочной зоне, слуховая — в верхней височной извилине височной доли, обонятельная и вкусовая — в переднем отделе височной доли.
Деятельность анализаторов отражает в нашем сознании внешний материальный мир. Это дает возможность млекопитающим приспосабливаться к условиям путем изменения поведения. Человек, познавая природные явления, законы природы и создавая орудия труда, активно изменяет внешнюю среду, приспосабливая ее к своим потребностям.
Кора больших полушарий выполняет функцию высшего анализатора сигналов от всех рецепторов тела и синтеза ответных реакций в биологически целесообразный акт. Она является высшим органом координации рефлекторной деятельности и органом приобретения временных связей — условных рефлексов. Кора выполняет ассоциативную функцию и является материальной основой психологической деятельности человека — памяти, мышления, эмоций, речи и регуляции поведения.
Проводящие пути головного мозга связывают его части между собой, а также со спинным мозгом (восходящие и нисходящие нервные пути), так что вся центральная нервная система функционирует как единое целое.
Запись опубликована в рубрике Биология человека с метками нервная система, система органов, человек. Добавьте в закладки постоянную ссылку.
Нормальное и нарушенное формирование коры головного мозга: нормальная эмбриология, родственные молекулы, типы миграции, нарушения миграции
1. Андраде Д.М. Генетическая основа эпилепсии, вызванной пороками развития коры головного мозга и у лиц со структурно нормальным мозгом. Хам Жене. 2009; 126: 173–193. [PubMed] [Google Scholar]
2. Антон Э.С., Маркионни М.А., Ли К.Ф., Ракик П. Роль передачи сигналов GGF/нейрегулина во взаимодействиях между мигрирующими нейронами и радиальной глией в развивающейся коре головного мозга. Разработка. 1997;124:3501–3510. [PubMed] [Google Scholar]
3. Баркович А.Дж., Геррини Р., Кузнецкий Р.И., Джексон Г.Д., Добинс В.Б. Эволюционная и генетическая классификация пороков развития коры: обновление 2012 г. Мозг. 2012; 135:1348–1369. [бесплатная статья PMC] [PubMed] [Google Scholar]
4. Баркович А.Ю., Кузнецкий Р.И. Нейровизуализация очаговых аномалий развития коры. Дж. Клин Нейрофизиол. 1996; 13: 481–494. [PubMed] [Google Scholar]
Баркович А.Ю., Кузнецкий Р.И. Нейровизуализация очаговых аномалий развития коры. Дж. Клин Нейрофизиол. 1996; 13: 481–494. [PubMed] [Google Scholar]
5. Баркович А.Дж., Кузнецкий Р.И., Добинс В.Б., Джексон Г.Д., Беккер Л.Е., Эврард П. Схема классификации пороков развития коры головного мозга. нейропедиатрия. 1996;27:59–63. [PubMed] [Google Scholar]
6. Баркович А.Дж., Кузнецкий Р.И., Джексон Г.Д., Геррини Р., Добинс В.Б. Эволюционная и генетическая классификация пороков развития коры головного мозга. Неврология. 2005; 65: 1873–1887. [PubMed] [Google Scholar]
7. Баркович А.Дж., Рейбо К.А. Пороки развития коры. Нейровизуализация Clin N Am. 2004; 14:401–423. [PubMed] [Google Scholar]
8. Buysse K, Riemersma M, Powell G, van Reeuwijk J, Chitayat D, Roscioli T, et al. Миссенс-мутации в бета-1,3-N-ацетилглюкозаминилтрансферазе 1 (B3GNT1) вызывают синдром Уокера-Варбурга. Хум Мол Жене. 2013; 22:1746–1754. [Бесплатная статья PMC] [PubMed] [Google Scholar]
9. Bystron I, Blakemore C, Rakic P. Развитие коры головного мозга человека: пересмотр комитета Боулдера. Нат Рев Нейроски. 2008; 9: 110–122. [PubMed] [Google Scholar]
Bystron I, Blakemore C, Rakic P. Развитие коры головного мозга человека: пересмотр комитета Боулдера. Нат Рев Нейроски. 2008; 9: 110–122. [PubMed] [Google Scholar]
10. Crino PB, Nathanson KL, Henske EP. Комплекс туберозного склероза. N Engl J Med. 2006; 355:1345–1356. [PubMed] [Google Scholar]
11. D’Arcangelo G, Miao GG, Chen SC, Soares HD, Morgan JI, Curran T. Белок, связанный с белками внеклеточного матрикса, делетированный в мышином мутантном рилере. Природа. 1995;374:719–723. [PubMed] [Google Scholar]
12. Dehay C, Kennedy H. Контроль клеточного цикла и развитие коры головного мозга. Нат Рев Нейроски. 2007; 8: 438–450. [PubMed] [Google Scholar]
13. DiLiberti JH. Наследственные синдромы макроцефалии-гамартомы. Am J Med Genet. 1998; 79: 284–290. [PubMed] [Google Scholar]
14. Dulabon L, Olson EC, Taglienti MG, Eisenhuth S, McGrath B, Walsh CA, et al. Рилин связывает интегрин альфа3бета1 и ингибирует миграцию нейронов. Нейрон. 2000; 27:33–44. [PubMed] [Академия Google]
[PubMed] [Академия Google]
15. Элиас Л.А., Ван Д.Д., Кригштейн А.Р. Адгезия щелевого соединения необходима для радиальной миграции в неокортексе. Природа. 2007; 448: 901–907. [PubMed] [Google Scholar]
16. Ferland RJ, Batiz LF, Neal J, Lian G, Bundock E, Lu J, et al. Нарушение нейронных предшественников вдоль вентрикулярной и субвентрикулярной зон при перивентрикулярной гетеротопии. Хум Мол Жене. 2009; 18: 497–516. [Бесплатная статья PMC] [PubMed] [Google Scholar]
17. Fernandez V, Llinares-Benadero C, Borrell V. Расширение и свертывание коры головного мозга: чему мы научились? EMBO J. 2016; 35:1021–1044. [Бесплатная статья PMC] [PubMed] [Google Scholar]
18. Fox JW, Lamperti ED, Eksioglu YZ, Hong SE, Feng Y, Graham DA, et al. Мутации в филамине 1 предотвращают миграцию нейронов коры головного мозга при перивентрикулярной гетеротопии человека. Нейрон. 1998; 21:1315–1325. [PubMed] [Google Scholar]
19. Гросс Р.Е., Мелер М.Ф., Маби П.С., Занг З., Санчи Л. , Кесслер Дж.А. Костные морфогенетические белки способствуют фиксации астроглиального клона клетками-предшественниками субвентрикулярной зоны млекопитающих. Нейрон. 1996; 17: 595–606. [PubMed] [Академия Google]
, Кесслер Дж.А. Костные морфогенетические белки способствуют фиксации астроглиального клона клетками-предшественниками субвентрикулярной зоны млекопитающих. Нейрон. 1996; 17: 595–606. [PubMed] [Академия Google]
20. Грубер Р., Чжоу З., Сукчев М., Йоерс Т., Фраппарт П.О., Ван З.К. MCPh2 регулирует способ деления нейропредшественников путем связывания центросомного цикла с митотическим входом через путь Chk1-Cdc25. Nat Cell Biol. 2011;13:1325–1334. [PubMed] [Google Scholar]
21. Guerrini R, Dobyns WB, Barkovich AJ. Аномальное развитие коры головного мозга человека: генетика, функциональные последствия и варианты лечения. Тренды Нейроси. 2008; 31: 154–162. [PubMed] [Google Scholar]
22. Guerrini R, Marini C. Генетические пороки развития коры головного мозга. Опыт Мозг Res. 2006; 173:322–333. [PubMed] [Академия Google]
23. Гуль А., Хассан М.Дж., Махмуд С., Чен В., Рахмани С., Насир М.И. и др. Генетические исследования аутосомно-рецессивной первичной микроцефалии в 33 пакистанских семьях: новые варианты последовательности в гене ASPM.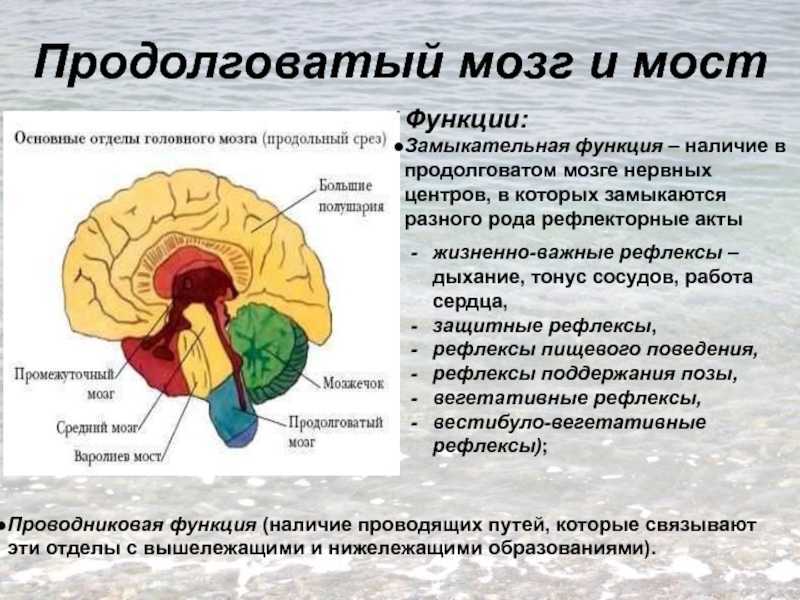 Нейрогенетика. 2006; 7: 105–110. [PubMed] [Google Scholar]
Нейрогенетика. 2006; 7: 105–110. [PubMed] [Google Scholar]
24. Hansen DV, Lui JH, Parker PR, Kriegstein AR. Нейрогенная радиальная глия в наружной субвентрикулярной зоне неокортекса человека. Природа. 2010; 464: 554–561. [PubMed] [Google Scholar]
25. Jackson AP, Eastwood H, Bell SM, Adu J, Toomes C, Carr IM, et al. Идентификация микроцефалина, белка, участвующего в определении размера человеческого мозга. Am J Hum Genet. 2002; 71: 136–142. [Бесплатная статья PMC] [PubMed] [Google Scholar]
26. Клячко В.А., Стивенс С.Ф. Оптимизация связности и позиционирование областей коры головного мозга. Proc Natl Acad Sci U S A. 2003;100:7937–7941. [Бесплатная статья PMC] [PubMed] [Google Scholar]
27. Kriegstein AR. Построение цепей: нейрогенез и миграция в развивающемся неокортексе. Эпилепсия 46 Приложение. 2005; 7:15–21. [PubMed] [Google Scholar]
28. Kriegstein AR, Noctor SC. Паттерны миграции нейронов в эмбриональной коре. Тренды Нейроси. 2004; 27: 392–399. [PubMed] [Академия Google]
[PubMed] [Академия Google]
29. Кумар А., Блэнтон С.Х., Бабу М., Маркандая М., Гиримаджи С.К. Генетический анализ первичной микроцефалии в индийских семьях: новые мутации ASPM. Клин Жене. 2004; 66: 341–348. [PubMed] [Google Scholar]
30. Lee JH, Huynh M, Silhavy JL, Kim S, Dixon-Salazar T, Heiberg A, et al. Соматические мутации de novo в компонентах пути PI3K-AKT3-mTOR вызывают гемимегалэнцефалию. Нат Жене. 2012;44:941–945. [Статья бесплатно PMC] [PubMed] [Google Scholar]
31. Luo R, Jeong SJ, Jin Z, Strokes N, Li S, Piao X. G-белковый рецептор 56 и коллаген III, пара рецептор-лиганд, регулирует корковое развитие и слоистость. Proc Natl Acad Sci U S A. 2011; 108:12925–12930. [Бесплатная статья PMC] [PubMed] [Google Scholar]
32. Махон О., ван ден Бут С.Дж., Бэкман М., Кемлер Р., Краусс С. Роль бета-катенина в развитии нейроэпителия коры и гиппокампа. Неврология. 2003; 122:129–143. [PubMed] [Google Scholar]
33. Morris-Rosendahl DJ, Najm J, Lachmeijer AM, Sztriha L, Martins M, Kuechler A, et al.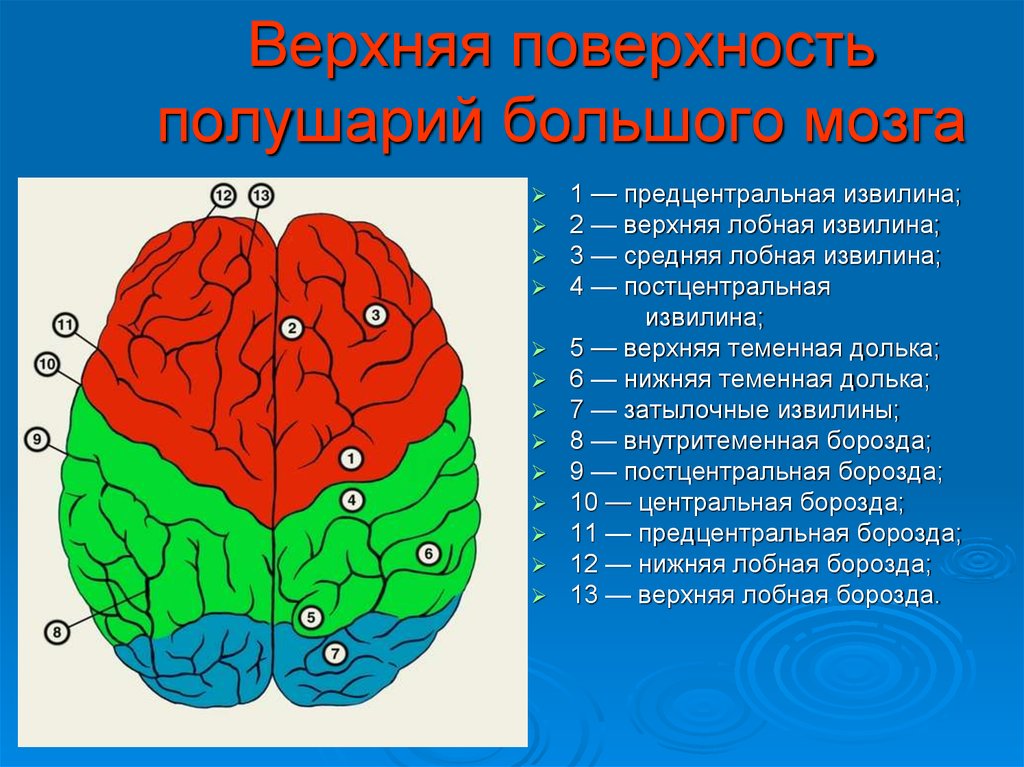 Уточнение фенотипа мутации тубулина альфа-1а (TUBA1A) у пациентов с классической лиссэнцефалией. Клин Жене. 2008; 74: 425–433. [PubMed] [Академия Google]
Уточнение фенотипа мутации тубулина альфа-1а (TUBA1A) у пациентов с классической лиссэнцефалией. Клин Жене. 2008; 74: 425–433. [PubMed] [Академия Google]
34. Надараджа Б., Парнавелас Дж.Г. Способы миграции нейронов в развивающейся коре головного мозга. Нат Рев Нейроски. 2002; 3: 423–432. [PubMed] [Google Scholar]
35. O’Leary DD, Borngasser D. Прогениторы кортикальной вентрикулярной зоны и их потомство сохраняют пространственные отношения и радиальное формирование паттерна во время развития препластинки, что указывает на раннюю протокарту. Приложение Cortex 16 Cereb. 2006; 1: i46–i56. [PubMed] [Google Scholar]
36. Olson EC, Walsh CA. Гладкое, грубое и перевернутое развитие новой коры. Curr Opin Genet Dev. 2002; 12: 320–327. [PubMed] [Академия Google]
37. Palmini A, Najm I, Avanzini G, Babb T, Guerrini R, Foldvary-Schaefer N, et al. Терминология и классификация кортикальных дисплазий. Неврология. 2004;62(6 Приложение 3):S2–S8. [PubMed] [Google Scholar]
38. Phoenix TN, Temple S. Spred1, отрицательный регулятор Ras-MAPK-ERK, накапливается в зародышевых зонах ЦНС, подавляет пролиферацию НСК и поддерживает структуру желудочковой зоны. Гены Дев. 2010; 24:45–56. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Phoenix TN, Temple S. Spred1, отрицательный регулятор Ras-MAPK-ERK, накапливается в зародышевых зонах ЦНС, подавляет пролиферацию НСК и поддерживает структуру желудочковой зоны. Гены Дев. 2010; 24:45–56. [Бесплатная статья PMC] [PubMed] [Google Scholar]
39. Pilz DT, Matsumoto N, Minnerath S, Mills P, Gleeson JG, Allen KM, et al. Мутации LIS1 и XLIS (DCX) вызывают большую часть классической лиссэнцефалии, но разные модели пороков развития. Хум Мол Жене. 1998;7:2029–2037. [PubMed] [Google Scholar]
40. Pontious A, Kowalczyk T, Englund C, Hevner RF. Роль промежуточных клеток-предшественников в развитии коры головного мозга. Дев Нейроски. 2008; 30:24–32. [PubMed] [Google Scholar]
41. Ракич П. Маленький шаг для клетки, гигантский скачок для человечества: гипотеза расширения неокортекса в процессе эволюции. Тренды Нейроси. 1995; 18: 383–388. [PubMed] [Google Scholar]
42. Rash BG, Lim HD, Breunig JJ, Vaccarino FM. Передача сигналов FGF расширяет площадь поверхности коры эмбриона, регулируя Notch-зависимый нейрогенез. Дж. Нейроски. 2011;31:15604–15617. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Дж. Нейроски. 2011;31:15604–15617. [Бесплатная статья PMC] [PubMed] [Google Scholar]
43. Раш Б.Г., Томази С., Лим Х.Д., Сух С.И., Ваккарино FM. Корковая гирификация, индуцированная фактором роста фибробластов 2 в мозге мыши. Дж. Нейроски. 2013;33:10802–10814. [Бесплатная статья PMC] [PubMed] [Google Scholar]
44. Raybaud C, Widjaja E. Развитие и дисгенезия коры головного мозга: пороки развития коры. Нейровизуализация Clin N Am. vii, 2011; 21:483–543. [PubMed] [Google Scholar]
45. Rivière JB, Mirzaa GM, O’Roak BJ, Beddaoui M, Alcantara D, Conway RL, et al. Зародышевые и постзиготические мутации de novo в AKT3, PIK3R2 и PIK3CA вызывают спектр родственных синдромов мегалэнцефалии. Нат Жене. 2012;44:934–940. [Бесплатная статья PMC] [PubMed] [Google Scholar]
46. Roll P, Rudolf G, Pereira S, Royer B, Scheffer IE, Massacrier A и др. Мутации SRPX2 при нарушениях языковой коры и когнитивных функций. Хум Мол Жене. 2006; 15:1195–1207. [PubMed] [Google Scholar]
47. Roscioli T, Kamsteeg EJ, Buysse K, Maystadt I, van Reeuwijk J, van den Elzen C, et al. Мутации в ISPD вызывают синдром Уокера-Варбурга и нарушение гликозилирования альфа-дистрогликана. Нат Жене. 2012; 44: 581–585. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Roscioli T, Kamsteeg EJ, Buysse K, Maystadt I, van Reeuwijk J, van den Elzen C, et al. Мутации в ISPD вызывают синдром Уокера-Варбурга и нарушение гликозилирования альфа-дистрогликана. Нат Жене. 2012; 44: 581–585. [Бесплатная статья PMC] [PubMed] [Google Scholar]
48. Сахара С., О’Лири Д.Д. Fgf10 регулирует переходный период дифференцировки стволовых клеток коры в радиальную глию, контролирующую генерацию нейронов и базальных предшественников. Нейрон. 2009; 63:48–62. [Бесплатная статья PMC] [PubMed] [Google Scholar]
49. Sarkisian MR, Bartley CM, Chi H, Nakamura F, Hashimoto-Torii K, Torii M, et al. Передача сигналов MEKK4 регулирует экспрессию филаминов и миграцию нейронов. Нейрон. 2006; 52: 789–801. [Бесплатная статья PMC] [PubMed] [Google Scholar]
50. Sheen VL, Dixon PH, Fox JW, Hong SE, Kinton L, Sisodiya SM, et al. Мутации в Х-сцепленном гене филамина 1 вызывают перивентрикулярную узловую гетеротопию как у мужчин, так и у женщин. Хум Мол Жене. 2001; 10: 1775–1783. [PubMed] [Академия Google]
[PubMed] [Академия Google]
51. Шин В.Л., Уолш, Калифорния. Перивентрикулярная гетеротопия: новый взгляд на синдром Элерса-Данлоса. Клин Мед Рез. 2005; 3: 229–233. [Бесплатная статья PMC] [PubMed] [Google Scholar]
52. Siegenthaler JA, Ashique AM, Zarbalis K, Patterson KP, Hecht JH, Kane MA, et al. Ретиноевая кислота из мозговых оболочек регулирует образование корковых нейронов. Клетка. 2009; 139: 597–609. [Бесплатная статья PMC] [PubMed] [Google Scholar]
53. Sun T, Hevner RF. Рост и сворачивание коры головного мозга млекопитающих: от молекул к порокам развития. Нат Рев Нейроски. 2014;15:217–232. [Бесплатная статья PMC] [PubMed] [Google Scholar]
54. Супер Х., Сориано Э., Уйлингс Х.Б. Функции предпластинки в развитии и эволюции неокортекса и гиппокампа. Res Brain Res Rev. 1998; 27:40–64. [PubMed] [Google Scholar]
55. van Reeuwijk J, Brunner HG, van Bokhoven H. Glyc-O-генетика синдрома Уокера-Варбурга. Клин Жене. 2005; 67: 281–289. [PubMed] [Google Scholar]
56. Уолш, Калифорния. Генетические аномалии коры головного мозга человека. Нейрон. 1999; 23:19–29. [PubMed] [Академия Google]
Уолш, Калифорния. Генетические аномалии коры головного мозга человека. Нейрон. 1999; 23:19–29. [PubMed] [Академия Google]
57. Уолш, Калифорния. Неврология в постгеномную эпоху: обзор. Тренды Нейроси. 2001; 24: 363–364. [PubMed] [Google Scholar]
58. Wang X, Tsai JW, LaMonica B, Kriegstein AR. Новый подтип клеток-предшественников в неокортексе эмбриона мыши. Нат Нейроски. 2011; 14: 555–561. [Бесплатная статья PMC] [PubMed] [Google Scholar]
59. Woods CG, Bond J, Enard W. Аутосомно-рецессивная первичная микроцефалия (MCPH): обзор клинических, молекулярных и эволюционных данных. Am J Hum Genet. 2005; 76: 717–728. [Бесплатная статья PMC] [PubMed] [Google Scholar]
60. Yamamoto T, Kato Y, Karita M, Kawaguchi M, Shibata N, Kobayashi M. Экспрессия генов, связанных с мышечной дистрофией с лиссэнцефалией. Педиатр Нейрол. 2004; 31: 183–190. [PubMed] [Google Scholar]
Что это такое, функции и местонахождение
Анатомия коры головного мозга.Что такое кора головного мозга?
Кора головного мозга — это самый внешний слой вашего мозга. Его поверхность имеет множество складок, что придает ему морщинистый вид. Складки состоят из множества глубоких борозд, называемых бороздами, и приподнятых областей, называемых извилинами. Эти складки увеличивают площадь поверхности коры головного мозга, позволяя обрабатывать большие объемы информации большим количеством нервных клеток. Кора головного мозга составляет около половины общей массы вашего мозга.
Кора головного мозга состоит из шести слоев нервных клеток, содержащих от 14 до 16 миллиардов нервных клеток. Его толщина составляет от двух миллиметров (мм) до четырех мм (от 0,08 до 0,16 дюйма).
Кора головного мозга разделена на четыре доли: лобную, теменную, височную и затылочную. Каждая из этих долей отвечает за обработку различных типов информации. В совокупности кора головного мозга отвечает за процессы более высокого уровня человеческого мозга, включая язык, память, рассуждение, мышление, обучение, принятие решений, эмоции, интеллект и личность.
Почему кору головного мозга называют серым веществом?
Серое вещество во внешнем слое головного мозга состоит из тел нервных клеток, включая конечные части нервов, называемые дендритами. Дендриты — это часть нервной клетки, которая получает химическое сообщение от другой клетки. Ваша кора головного мозга серая, потому что в этой части нерва отсутствует жировой покровный материал, называемый миелином.
Дендриты — это часть нервной клетки, которая получает химическое сообщение от другой клетки. Ваша кора головного мозга серая, потому что в этой части нерва отсутствует жировой покровный материал, называемый миелином.
Белое вещество головного мозга состоит из пучков аксонов, длинной центральной части нервной клетки, обернутой миелином. Беловатый цвет ткани придает миелин.
В чем разница между корой головного мозга и головным мозгом?
Кора головного мозга — это внешний слой, лежащий поверх головного мозга. Головной мозг — самая большая область вашего мозга. Ваш головной мозг делит ваш мозг на две половины, называемые полушариями. Полушария соединены пучком нервных волокон, называемым мозолистым телом. Мозолистое тело позволяет вашим полушариям общаться друг с другом.
Что такое неокортекс?
Большая часть коры головного мозга считается неокортексом. «Нео» означает новый. Ваш неокортекс назван так потому, что его появление считается относительно новым в эволюции позвоночных. У человека 90% коры головного мозга составляет неокортекс.
У человека 90% коры головного мозга составляет неокортекс.
Каковы функции коры головного мозга?
Кора головного мозга участвует во многих высокоуровневых функциях, таких как рассуждение, эмоции, мышление, память, язык и сознание. Каждая доля вашего мозга связана с различными функциями.
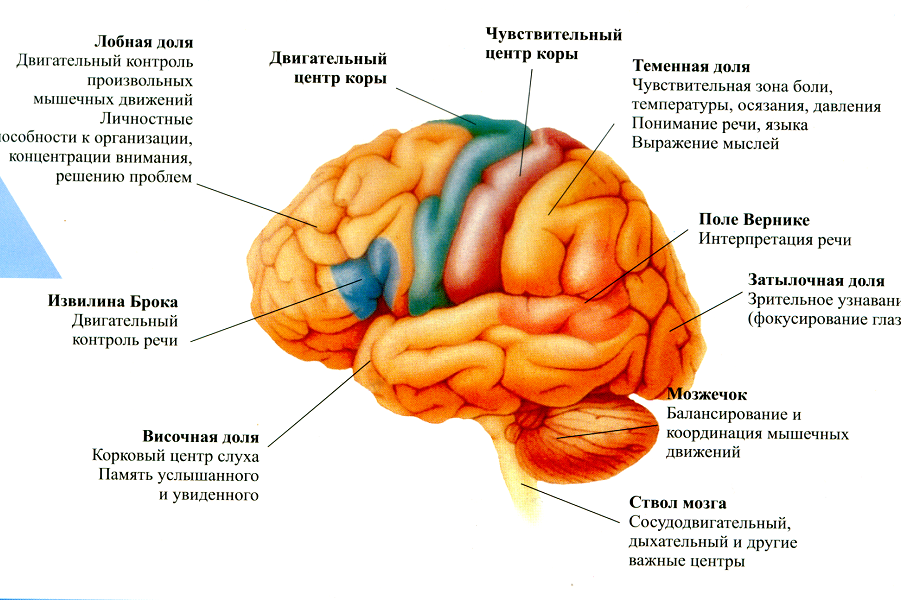
Функции лобной доли
Лобная доля находится в передней части мозга за лбом. Функции вашей лобной доли включают:
- Принятие решений, решение проблем.
- Сознательная мысль.
- Внимание.
- Эмоциональный и поведенческий контроль.
- Производство речи.
- Личность.
- Интеллект.
- Движение тела.
В этой доле следует особо отметить моторную кору, префронтальную кору и зону Брока. Ваша моторная кора отвечает за движение тела. Ваша префронтальная кора отвечает за «исполнительные функции», такие как мышление и решение проблем. Он также контролирует и направляет другие области вашего мозга. Зона Брока — это часть лобной доли, которая отвечает за производство речи.
Зона Брока — это часть лобной доли, которая отвечает за производство речи.
Функции затылочной доли
Затылочная доля находится в задней части мозга. Функции вашей затылочной доли включают:
- Визуальная обработка и интерпретация.
- Сбор визуальных данных о цвете, движении и ориентации.
- Распознавание объектов и лиц.
- Восприятие глубины и расстояния.
- Картографирование визуального мира.
Функции теменной доли
Темная доля расположена между лобной и затылочной долями и над височной долей. Функции вашей теменной доли включают в себя:
- Обработка сенсорной информации (прикосновение, давление, боль, положение, вибрация, температура).
- Пространственная обработка и пространственные манипуляции. Это способность понимать, где вы находитесь в трехмерном пространстве, например, как перемещаться по дому или городу.
В этой доле следует особо отметить соматосенсорную кору. Он получает сенсорную информацию («чувственную» информацию) со всего тела. Вот пример совместной работы долей мозга:
Он получает сенсорную информацию («чувственную» информацию) со всего тела. Вот пример совместной работы долей мозга:
Моторная кора в лобной доле вашего мозга посылает сообщение, которое предписывает мышцам руки и кисти потянуться к чашке супа на кухонном столе. Соматосенсорная кора теменной доли оценивает информацию, полученную при прикосновении к чашке, в том числе оценивает ее температуру. Пространственная обработка в теменной доле позволяет вам схватить чашку, безупречно ориентируясь на расстоянии от руки до чашки относительно стола и других окружающих предметов.
Функции височной доли
Височная доля расположена между лобной и затылочной долями и ниже теменной доли. Функции вашей височной доли включают:
- Понимание языка, формирование речи, обучение.
- Память.
- Слушание.
- Невербальная интерпретация.
- Преобразование звукового изображения в визуальное.
Особое место в этой доле занимает зона Вернике.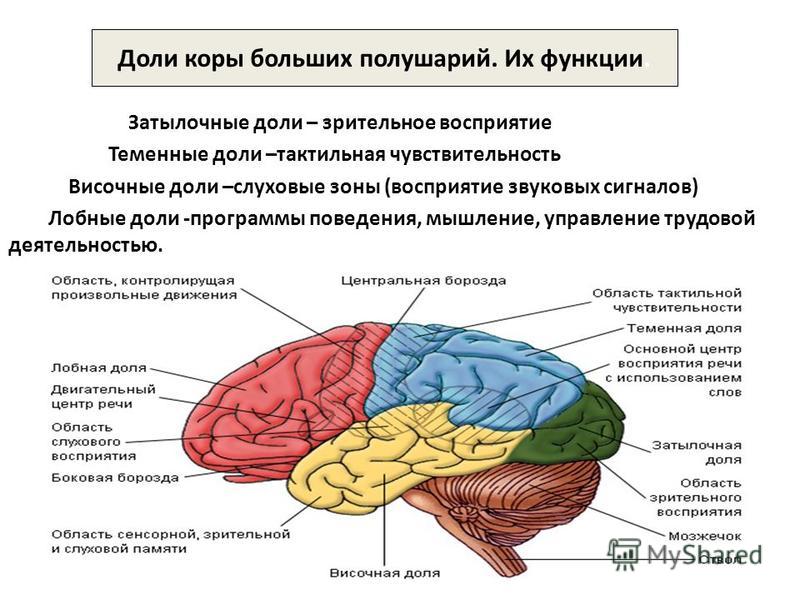 Совсем недавно было обнаружено, что эта область участвует в языковом комплексе, основанном на тонах и звуках речи, связывая их с ранее изученными звуками.
Совсем недавно было обнаружено, что эта область участвует в языковом комплексе, основанном на тонах и звуках речи, связывая их с ранее изученными звуками.
Каковы области коры головного мозга?
Некоторые исследователи смотрят на мозг по-другому и классифицируют области коры головного мозга по их трем основным типам функций: сенсорные, моторные и ассоциативные области.
Сенсорные области: Эти области коры головного мозга получают сенсорную информацию от ваших органов чувств и окружающей среды. Функции включают в себя:
- Осмысление визуальной информации и распознавание объектов. Эти функции обрабатываются областью затылочной доли, называемой зрительной корой.
- Оценка информации о прикосновении, температуре, положении, вибрации, давлении и боли вашего тела. Эти функции обрабатываются областью вашей теменной доли, называемой соматосенсорной корой.
- Обработка слуховой информации. Эта функция обрабатывается областью височной доли, называемой слуховой корой.

- Обработка вкуса и аромата. Эти функции обрабатываются областью лобной доли, называемой вкусовой корой.
Моторные зоны: Эти области коры головного мозга участвуют в произвольных движениях мышц. Эти функции обрабатываются в основном вашей лобной долей. Функции включают в себя:
- Координация движения мышц.
- Планирование сложных движений.
- Обучение через подражание и эмпатию.
Ассоциативные области: Эти области разбросаны по всем четырем долям и соединяются и усложняют функции. Функции включают в себя:
- Организация и придание смысла информации из сенсорных и моторных областей.
- Личность и контроль над эмоциональным поведением.
- Пространственное восприятие и мышление.
- Обработка памяти.
- Думайте образно и сохраняйте зрительную память.
- Создавайте визуальную информацию с помощью воспоминаний, звука и языка.
Как может быть повреждена кора головного мозга?
Повреждение любой области коры головного мозга обычно возникает в результате опухолей, травм, аутоиммунных заболеваний или нарушения мозгового кровообращения (кровоизлияние в мозг или инсульт).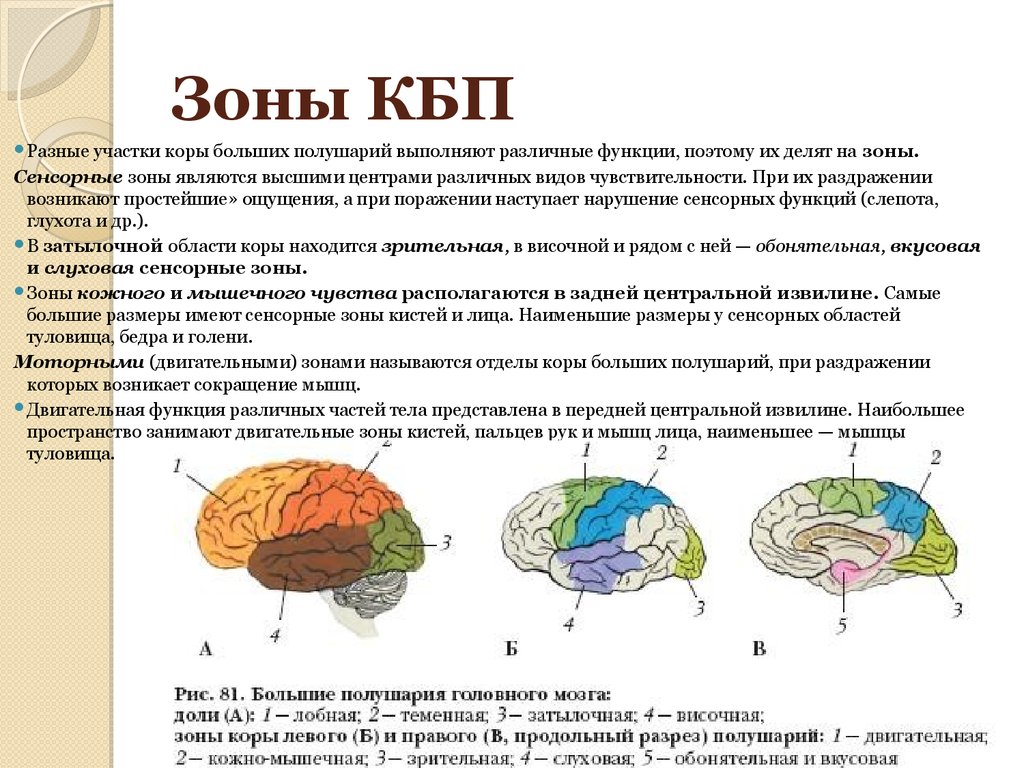
Каковы симптомы повреждения коры головного мозга?
Симптомы зависят от пораженной области коры головного мозга.
Травма лобной доли
Симптомы повреждения или травмы лобной доли включают:
- Проблемы с памятью.
- Изменения личности.
- Решение проблем, вопросы принятия решений.
- Проблемы с вниманием.
- Эмоциональный дефицит, социально неадекватное поведение, изменения поведения.
- Отсутствие способности понимать или выражать речь (афазия).
- Затрудненная речь (апраксия).
- Слабость, паралич, потеря мышечного контроля на одной стороне тела (вялая гемиплегия).
Дополнительной причиной поражения лобных долей является деменция.
Травма теменной доли
Симптомы повреждения теменной доли включают:
- Генерация памяти.
- Проблемы с письмом или невозможность писать (аграфия).
- Трудности с математикой.
- Онемение.
Leave A Comment