
Учение Ч. Дарвина о естественном отборе
Искусственный отбор, т.е. сохранение особей с полезными для человека признаками и устранение всех остальных, проводит человек, ставящий перед собой определенные задачи. Признаки, накапливаемые при искусственном отборе, полезны для человека, но необязательно выгодны для животных. Дарвин высказал предположение, что в природе сходным путем накапливаются признаки, полезные только для организмов и вида в целом, в результате чего образуются разновидности и виды. В этом случае требовалось установить наличие неопределенной индивидуальной изменчивости у диких животных и растений. Кроме того, необходимо было доказать существование в природе какого-то направляющего фактора, действующего аналогично воле человека в процессе искусственного отбора.
Всеобщая индивидуальная изменчивость, избыточная численность потомства и ограниченность ресурсов. Дарвин показал, что у представителей диких видов животных и растений индивидуальная изменчивость представлена очень широко. Индивидуальные отклонения могут быть полезными, нейтральными или вредными для организма. Все ли особи оставляют потомство? Если нет, то какие факторы сохраняют особей с полезными признаками и устраняют всех остальных?
Дарвин обратился к анализу размножения организмов. Все организмы оставляют значительное, иногда очень многочисленное потомство. Одна особь сельди выметывает в среднем около 40 тыс. икринок, осетр — 2 млн, лягушки — до 10 тыс. икринок. На одном растении мака ежегодно созревает до 30 — 40 тыс. семян. Даже медленно размножающиеся животные потенциально способны оставить огромное число потомков. Самки слонов приносят детенышей в возрасте между 30 и 90 годами. За 60 лет они рождают в среднем шесть слонят. Расчеты показывают, что даже при такой низкой интенсивности размножения через 750 лет потомство одной пары слонов составило бы 19 млн особей. На основе этих и многих других примеров Дарвин приходит к выводу о том, что в природе любой вид животных и растений стремится к размножению в геометрической прогрессии.
Каждая пара организмов дает гораздо больше потомков, чем их доживает до взрослого состояния. Большая часть появившихся на свет организмов, следовательно, гибнет, не достигнув половой зрелости. Причины гибели разнообразны: главным образом это ограниченность ресурсов.
Дело в том, что как в планетарном, так и в местном — биогеоценотическом масштабе все ресурсы, которые могут быть использованы организмами, всегда ограничены. Так, площадь или объем, заселенные растениями, животными и микроорганизмами, имеют конечные, хотя иногда и достаточно большие размеры. На этой площади или в объеме растительными организмами — продуцентами за счет энергии солнечного света может быть создано конечное количество биомассы. Ее могут использовать растительноядные животные, а их организмы — в качестве пищи — хищники.
В результате — недостаток корма из-за конкуренции с представителями своего же вида, нападение врагов, действие неблагоприятных физических факторов среды — засухи, сильных морозов, высокой температуры и пр. Отсюда следует второй вывод, сделанный Дарвином: в природе происходит непрерывная борьба за существование. Этот термин должен пониматься в широком смысле, как любая зависимость организмов от всего комплекса условий окружающей их природы. Иначе говоря,
Формы борьбы за существование и естественный отбор. Дарвин выделил три основные формы борьбы за существование: а) внутривидовую, б) межвидовую, в) борьбу с неблагоприятными условиями среды.
Внутривидовая борьба. У особей одного вида потребности в пище, территории и других условиях существования одинаковы. Поэтому конкуренция между ними наиболее острая. Дарвин считал внутривидовую борьбу самой напряженной. Например, птицы одного вида конкурируют из-за мест гнездования. Самцы многих видов млекопитающих и птиц в период размножения вступают друг с другом в борьбу за право обзавестись семьей (половой отбор).
Поэтому конкуренция между ними наиболее острая. Дарвин считал внутривидовую борьбу самой напряженной. Например, птицы одного вида конкурируют из-за мест гнездования. Самцы многих видов млекопитающих и птиц в период размножения вступают друг с другом в борьбу за право обзавестись семьей (половой отбор).
Межвидовая борьба. Примеры межвидовой борьбы многочисленны. И волки, и лисы охотятся на зайцев. Между волками и зайцами, а также между лисами и зайцами идет напряженная борьба за существование. Отсутствие добычи обрекает хищников на голод и гибель. В то же время между хищниками — волками и лисами — тоже существует конкуренция за пищу. Это не значит, что они непосредственно вступают в борьбу друг с другом, но успех одного означает неудачу другого. Травоядные животные смогут выжить и оставить потомство только в том случае, если они сумеют избежать хищников и будут обеспечены пищей. Но растительностью питаются разные виды млекопитающих, а кроме того, насекомые и моллюски: что досталось одному, не достанется другому. Существование трав в свою очередь зависит не только от поедания их животными, но и от других условий — опыления цветков насекомыми, конкуренции с другими растениями за свет, влагу и т.д. Беспрепятственное размножение микроорганизмов сдерживают помимо прочих факторов антибиотики, выделяемые грибами, и фитонциды, образуемые зелеными растениями. К межвидовой борьбе относятся и взаимоотношения в форме паразитизма. Паразиты ослабляют организм хозяина, делают его менее конкурентоспособным.
Борьба с неблагоприятными условиями внешней среды. Факторы неживой природы оказывают огромное влияние на выживаемость организмов. Много растений гибнет во время холодных малоснежных зим. В сильные морозы смертность увеличивается и среди животных, обитающих в почве (кроты, дождевые черви). Зимой при недостатке растворенного в воде кислорода погибает рыба. Семена растений нередко заносятся ветром в неблагоприятные местообитания и не прорастают.
Все формы борьбы за существование сопровождаются истреблением в каждом поколении огромного количества организмов или приводят к тому, что часть их не оставляет потомства.
Кто же выживает в этой постоянно происходящей борьбе за существование? Наблюдения показывают, что для растительных и животных организмов характерна всеобщая изменчивость признаков, свойств и бесконечное разнообразие их комбинаций. Даже в потомстве одной пары родителей нет совершенно одинаковых особей (за исключением монозиготных близнецов). В борьбе за существование выживают и оставляют потомство индивидуумы, обладающие таким комплексом признаков и свойств, который позволяет наиболее успешно конкурировать с другими. Таким образом, в природе в результате борьбы за существование происходят процессы избирательного уничтожения одних особей и преимущественного размножения других — явление, названное Дарвином естественным отбором, или выживанием наиболее приспособленных. При изменении условий внешней среды полезными для выживания могут оказаться какие-то иные, чем прежде, признаки. В результате меняется направление давления отбора, перестраивается генетическая структура вида, благодаря размножению широко распространяются новые признаки — появляется новый вид.
Следовательно, виды изменяются в процессе приспособления к условиям внешней среды. Движущей силой изменения видов, т.е. эволюции, является естественный отбор. Материалом для отбора служит наследственная (неопределенная, индивидуальная, мутационная) изменчивость. Изменчивость, обусловленная прямым влиянием внешней среды на организмы (групповая, модификационная), не имеет значения для эволюции, поскольку по наследству не передается.
Образование новых видов.
 Поэтому несхожие особи имеют преимущество в оставлении потомства. С каждым поколением различия становятся все более выраженными, а промежуточные формы, сходные между собой, вымирают. Так из одного вида образуются два или несколько. Явление расхождения признаков, ведущее к видообразованию, Дарвин назвал
Поэтому несхожие особи имеют преимущество в оставлении потомства. С каждым поколением различия становятся все более выраженными, а промежуточные формы, сходные между собой, вымирают. Так из одного вида образуются два или несколько. Явление расхождения признаков, ведущее к видообразованию, Дарвин назвал Причиной дивергенции могут стать и неодинаковые условия внешней среды в разных районах территории, занимаемой видом. Например, две группы особей какого-либо вида вследствие этого будут накапливать различные изменения. Возникает процесс расхождения признаков. Через определенное число поколений такие группы становятся разновидностями, а затем — видами.
Действие естественного отбора можно наблюдать в эксперименте.
В нашей стране широко распространен богомол обыкновенный — крупное хищное насекомое (длина тела у самок достигает 48 — 76 мм), питающееся разнообразными мелкими насекомыми — тлями, клопами, мухами. Окраска разных особей этого вида бывает зеленой, желтой и бурой. Богомолы зеленого цвета встречаются среди травы и кустарников, бурые — на растениях, выгорающих от солнца. Неслучайность такого распределения животных ученые доказали в эксперименте на расчищенной от травы площадке блекло-бурого цвета. К колышкам на площадке были привязаны богомолы всех трех цветов. За время опыта птицами были уничтожены 60% желтых, 55% зеленых и только 20% бурых богомолов, у которых окраска тела совпадала с цветом фона. Аналогичные опыты были поставлены с куколками бабочки- крапивницы. В случае несоответствия окраски куколки окраске фона птицами уничтожалось гораздо больше куколок, чем в случае совпадения фона с окраской. Водоплавающие птицы в бассейне ловят преимущественно рыбу, окраска которой не соответствует цвету дна.
Водоплавающие птицы в бассейне ловят преимущественно рыбу, окраска которой не соответствует цвету дна.
Важно отметить, что для выживания имеет значение не один какой-либо признак, а комплекс признаков — фенотип организма. В том же опыте с богомолами, очень простом по сравнению с реальными природными условиями, среди бурых особей, защищенных окраской тела, птицы склевывали беспокойных, активно двигающихся насекомых. Спокойные, малоподвижные богомолы избегали нападения. Один и тот же признак в зависимости от окружающих условий может способствовать выживанию или, напротив, привлекать внимание врагов. На рисунке 18.4 приведены две формы бабочки березовой пяденицы. Светлая форма мало заметна на березе, в то время как мутантная темноокрашенная форма хорошо видна на ней (Б). Темные бабочки преимущественно склевываются птицами. Ситуация меняется вблизи промышленных предприятий: копоть, покрывающая стволы деревьев, создает защитный фон для мутантов, в то время как светлая бабочка хорошо заметна (А).
Рис. 18.4. Березовая пяденица: светлая и темная расы на темном (А) и светлом (Б) фоне
Мутации и половой процесс создают генетическую неоднородность внутри вида. Их действие, как видно из приведенных примеров, ненаправленно. Эволюция же — процесс направленный, связанный с выработкой приспособлений по мере прогрессивного усложнения строения и функций животных и растений. Существует лишь один направленный эволюционный фактор — естественный отбор.
Под действие отбора могут попасть либо отдельные особи, либо целые группы. В любом случае отбор сохраняет наиболее приспособленные к данной среде организмы. Нередко отбор сохраняет признаки и свойства, невыгодные для отдельной особи, но полезные для группы особей или вида в целом. Примером такого приспособления служит зазубренное жало пчелы. Ужалившая пчела оставляет жало в теле врага и погибает, но гибель особи способствует сохранению пчелиной семьи.
Факторами отбора служат условия внешней среды, точнее, весь комплекс абиотических и биотических условий среды. В зависимости от этих условий отбор действует в разных направлениях и приводит к неодинаковым эволюционным результатам. В настоящее время различают несколько форм естественного отбора, из которых ниже будут рассмотрены только основные.
В зависимости от этих условий отбор действует в разных направлениях и приводит к неодинаковым эволюционным результатам. В настоящее время различают несколько форм естественного отбора, из которых ниже будут рассмотрены только основные.
Дарвин показал, что принцип естественного отбора объясняет возникновение всех без исключения основных характеристик органического мира: от признаков, свойственных крупным систематическим группам живых организмов, до мелких приспособлений. Теорией Дарвина завершились длительные поиски естествоиспытателей, которые пытались найти объяснение многим чертам сходства, наблюдаемым у организмов, относящихся к разным видам. Дарвин объяснил это сходство родством и показал, как идет образование новых видов, как происходит эволюция.
С общетеоретической точки зрения главное в учении Дарвина — это идея развития живой природы, противостоящая представлению о застывшем, неизменяющемся мире. Признание учения Дарвина стало переломным моментом в истории биологических наук. Факты, накопленные в додарвиновский период развития биологии, получили новое освещение. Возникли новые направления в биологии — эволюционная эмбриология, эволюционная палеонтология и др.
Учение Дарвина служит естественнонаучной основой для доказательства ложности религиозных взглядов. Материалистическое объяснение целесообразности строения живых организмов, происхождения и многообразия видов в самой основе подрывает устои религии.
Труд Дарвина явился одним из крупнейших достижений естествознания XIX в.
Опорные точки
• Для особей любого вида характерна всеобщая индивидуальная (наследственная) изменчивость.
• Численность потомства в пределах каждого вида организмов очень велика, а пищевые ресурсы всегда ограничены. Отсюда вытекает неизбежность борьбы за существование.
• Дарвин выделял три формы борьбы за существование: внутривидовую, межвидовую и борьбу с абиотическими факторами среды.
• В результате борьбы за существование на всех этапах онтогенеза происходит избирательное выживание наиболее приспособленных организмов и гибель менее приспособленных особей — идет естественный отбор.
Вопросы и задания для повторения
1. Что такое естественный отбор?
2. Что такое борьба за существование? Каковы ее формы?
3. Какая форма борьбы за существование является наиболее напряженной и почему?
4. Вспомните материал предыдущих глав. Какие процессы, происходящие в природе, снижают интенсивность внутривидовой борьбы за существование? Каков биологический смысл этого явления?
5. В чем, по вашему мнению, заключаются биологические причины сохранения жизни особей, устраненных от размножения?
1.Что такое естественный отбор.2.Что такое борьба за существование и каковы ее формы.3.Какая форма борьбы за существование является наиболее напряженной и почему.5.Какие процессы, происходящие в природе, снижают интенсивность внутривидовой борьбы за существование?Каков биологический смысл этого явления?6.В чем заключается, по вашему мнению, биологические причины сохранения жизни особей, устраненных от размножения?7.Дайте определение вида.8.Расскажите, какие биологические механизмы препятствуют обмену генами между видами.9.В чем причина бесплодности межвидовых гибридов?10.Что такое ареал вида?11.Что такое радиус индивидуальной активности организмов?Приведите примеры радиуса индивидуальной активности для растений и животных.12.Что такое популяция?Дайте определение.13.Какие процессы приводят к изменению частоты встречаемости генов в популяциях?14.Почему разные популяции однго вида отличаются по частоте генов?15.Что такое микро эволюция. И прономеруйте ответы
1.Естественный отбор — процесс, в результате которого выживают и оставляют после себя потомство преимущественно особи с полезными в данных условиях изменениями.
2.Борьба за существование – это совокупность многообразных и сложных взаимоотношений, существующих между организмами и условиями среды.
Дарвин выделил три основные формы борьбы за существование: а) межвидовую, б) внутривидовую, в) борьбу с неблагоприятными условиями внешней среды.
3.Дарвин считал внутривидовую борьбу самой напряженной. У особей же одного вида потребности в пище, территории и других условиях существования одинаковы. Поэтому конкуренция между ними особенно остра.
У особей же одного вида потребности в пище, территории и других условиях существования одинаковы. Поэтому конкуренция между ними особенно остра.
5.Интенсивность внутривидовой борьбы за существование снижается при резком снижении численности, увеличения площади и количества еды.
6.Инстинкт сохранения рода – минус. Инстинкт самосохранения – остается
7.Buд — это совокупность особей, которые сходны по морфофизиоло-гическим признакам скрещиваться между собой, давать плодовитое потомство и формируют систему популяций, образующих общий ареал
8.Одна из важных характеристик вида – его репродуктивная изоляция, т.е. наличие механизмов, препятствующих скрещиванию с особями других видов и вследствие этого предотвращающих поток генов извне, защищенность генофонда от притока генов из других, в том числе близкородственных видов, достигается разными путями. Можно выделить следующие механизмы, препятствующие обмену генами между видами:
1. различие в сроках размножения у представителей разных видов;
2. различие в местах, предпочитаемых для размножения;
3. несоответствие стандартов видового сексуального поведения;
4. несоответствие ферментов акросомы (передней части головки сперматозоида) химическому строению мембраны яйцеклетки;
5. несоответствие в строении половых органов у представителей различных видов;
6. нежизне или стерильность межвидовых гибридов.
Следовательно, вид – реально существующая, генетически неделимая единица органического мира.
9.Бесплодие межвидовых гибридов связано с тем, что каждая хромосома представлена одним гомологом, и образование бивалентов в мейозе оказывается невозможным
10.реал вида т. е. рас место обитания того или иного вида например : кенгуру утконосы живут в Австралии но их нет например в Индии и т. п
11.Степень подвижности особей выражается расстоянием, на которое может перемещаться животное, т. е. радиусом индивидуальной активности. У растений этот радиус определяется расстоянием, на которое рас пыльца, семена или вегетативные части дать начало новому растению. У животных радиус индивидуальной активности тесно связан с физическими возможностями организма и особенностями его жизнедеятельности.
У животных радиус индивидуальной активности тесно связан с физическими возможностями организма и особенностями его жизнедеятельности.
Для виноградной улитки он составляет несколько десятков метров, для ондатры – несколько сотен метров, для северного оленя – более ста километров.
12.Популя́ция (от лат. populatio — население) — совокупность организмов одного вида, длительное время обитающих на одной территории (занимающих определённый ареал) и частично или полностью изолированных от особей других таких же групп.
13.зменения условий обитания организма(включается в действие естественный отбор который и определяет изменение частоты встречаемости генов). дреф генов(дрейф-изменение частоты встречаемости генов в популяции в ряде поколений под действием случайных фактов
14.В разных популяциях одного вида частота мутантных генов неодинакова. Практически нет двух популяций с совершенно одинаковой частотой встречаемости мутантных признаков. Эти различия могут быть обусловлены тем, что популяции обитают в неодинаковых условиях внешней среды. Направленное изменение частоты генов в популяциях обусловлено действием естественного отбора. Но и близко расположенные, соседние популяции могут отличаться друг от друга столь же значительно, как и далеко расположенные. Это объясняется тем, что в популяциях ряд процессов приводит к так называемому дрейфу генов – ненаправленному, случайному изменению частоты генов, или, другими словами, их генетической структуры.
15.Микроэволюция — рас в популяции малых изменений в частотах аллелей на протяжении нескольких поколений; эволюционные изменения на внутривидовом уровне.
Объяснение:
Урок 11 БОРЬБА ЗА СУЩЕСТВОВАНИЕ И ЕСТЕСТВЕННЫЙ ОТБОР
Урок 11
БОРЬБА ЗА СУЩЕСТВОВАНИЕ И ЕСТЕСТВЕННЫЙ ОТБОР
Задачи. Сформировать у учащихся систему знаний о естественном отборе и его роли в эволюции.
Сформировать у школьников умение сравнивать разные формы естественного отбора друг с другом и правильно определять их по сущностным характеристикам.
Убедить школьников в том, что естественный отбор является основной и направляющей движущей силой эволюционного процесса.
Тип урока: закрепление знаний и способов деятельности.
Вид урока: урок-упражнение.
Методы: репродуктивные, частично-поисковые.
Ход урока
I. Актуализация знаний
Чтение и анализ текста из работы Ч. Дарвина «Происхождение видов…»
«…Многочисленные незначительные различия, появляющиеся в потомстве, несомненно или предположительно происходящем от общих родителей, и наблюдаемые у особей одного и того же вида, обитающих в одной и той же местности, могут быть названы индивидуальными. Эти индивидуальные отличия для нас крайне важны, так как они часто наследственны, как всякому известно; они доставляют естественному отбору материал для воздействия и накопления, подобно тому как человек накопляет у своих домашних животных и культурных растений индивидуальные отличия в каком угодно направлении».
Вопросы:
Чем, по-вашему, вызваны индивидуальные отличия между особями одного вида?
Приведите примеры индивидуальных отличий у особей одного вида.
Что означает мысль Дарвина: «Они доставляют естественному отбору материал для воздействия и накопления?
II. Закрепление знаний. Естественный отбор как выживание наиболее приспособленных особей осуществляется на основе наследственной изменчивости.
Закрепление умений различать и характеризовать отдельные формы борьбы за существование.
Беседа с использованием таблиц, натуральных объектов, а также вопросов и заданий.
1. Вопросы:
а) Какой процесс называется естественным отбором?
б) Каким образом можно убедиться в существовании такого отбора?
в) Какая форма изменчивости служит материалом для протекания естественного отбора?
г) Что такое борьба за существование? Каковы ее формы?
д) Какая форма борьбы за существование является наиболее напряженной и почему?
Сообщение учащегося о том, какие процессы, происходящие в природе, снижают интенсивность внутривидовой борьбы за существование. Каков биологический смысл этого явления?
Каков биологический смысл этого явления?
Выполнение упражнений с последующим обсуждением (можно по вариантам) результатов.
Упражнение 1. Рассмотрите рисунок «Сопряженная динамика численности популяций зайца-беляка и рыси. (Жирной кривой чертой отмечены колебания численности рыси (по П. Фабру, 1971 г.) Какие положения эволюционной теории могут помочь вам разобраться в процессах, отраженных на графике? Какой из видов борьбы за существование проявляется наиболее ярко? Почему в некоторые годы наблюдалось уменьшение количества волков, лисиц, рысей?
Упражнение 2. В 1925 г. знаменитый в то время итальянский математик В. Вольтер заинтересовался любопытным биологическим фактом: резкое уменьшение промысла рыбы в Адриатическом море во время Первой мировой войны вызвало заметное увеличение относительной доли хищных рыб в уловах. Чем объясняется этот факт?
Упражнение 3. Борьба за существование и естественный отбор.
а) В гнездах сов часто можно обнаружить разновозрастных птенцов. У полярной совы старшие появляются в июне, а младшие птенцы — в июле. У филина все птенцы старше один другого на пять-семь дней. Каково биологическое значение этого явления? Какими положениями эволюционного учения можно объяснить особенности размножения хищных птиц?
б) Мальки трески часто поселяются под колоколом некоторых видов медуз, щупальца которых ядовиты. Как могло возникнуть такое явление? С помощью каких положений эволюционной теории можно объяснить такую «странность» поведения мальков трески?
III. Подведение итогов урока.
Выводы записываются в тетрадь:
Естественный отбор — результат борьбы за существование.
Материалом для отбора служит наследственная (неопределенная, индивидуальная, мутационная) изменчивость.
Результатом естественного отбора является появление новых видов.
Задание на дом: повторить § 12 (3.2).
Заполнить таблицу.
Сравнительная характеристика учений о движущих силах эволюционного процесса
Ж. -Б. Ламарк
-Б. ЛамаркЧ. Дарвин
Борьба за существования в природе
2. Что такое борьба за существование? Каковы ее формы? Этот термин должен пониматься в широком смысле, как любая1. Вспомните материал предыдущих глав. Какие процессы, происходящие в природе, снижают интенсивность внутривидовой борьбы за существование?. Поскольку во время следственных экспериментов он становился особенно разговорчивым, она ездила вместе с группой, постоянно была рядом с чудовищем
В природе особи борются за лучшие условия существования, в результате чего выживают самые приспособленные из них. Борьба за существования может происходить между особями одного вида. К примеру, в брачный период олени соревнуются в громкости гона. Дорогостоящая, энергоемкая машина некоторое время работает на холостых оборотах и наконец замирает в ожидании следующего убийства
Внутривидовая борьба за существование проявляется у разных видов, прежде всего, в конкуренции за кормовые ресурсы и полового партнера. Как правило, прямому столкновению особей препятствуют различные приспособления, среди которых следует выделить метки. Убийство Жени Качаловой, конечно, событие серьезное, но не государственного масштаба
Борьба за существование – это совокупность многообразных и сложных взаимоотношений, существующих между организмами и условиями среды. Факторы неживой природы оказывают огромное влияние на выживаемость организмов. Соловьев мгновенно узнал клип, его часто крутили по телевизору
Приведите примеры трёх форм борьбы за существование, которые вы наблюдали в природе. Наиболее общей причиной борьбы за существование является несоответствие между запросами (потребностями) организма и условиями окружающей среды. Со дна банки удалось соскрести пару ложек растворимого кофе
Борьба за существование. Причина: организмов рождается гораздо больше, чем может выжить, поэтому для них всехВНУТРИВИДОВАЯ — МЕЖВИДОВАЯ 1. Установите соответствие между процессом, происходящим в природе, и формой борьбы за существование: 1). Чувство абсолютной, глобальной безысходности, тупая головная боль
Чувство абсолютной, глобальной безысходности, тупая головная боль
Борьба за существование не прекращается даже в оптимальных условиях, однако при изменении действия лимитирующих или элиминирующих факторов одни ее формы могут заменяться на другие. Результатом борьбы за существование является физическая или. Она не хотела его отпускать, попросила подняться в квартиру
Борьба за существование и естественный отбор. Задачи. Сформировать у учащихся систему знаний о естественномСообщение учащегося о том, какие процессы, происходящие в природе, снижают интенсивность внутривидовой борьбы за существование. Теперь кто-то решил показать бедняге, насколько серьезно он вляпался? Запросто может стать подозреваемым в убийстве?»
Межвидовая борьба за существование — это борьба за существование между разными видами. В ходе межвидовой борьбы организмыБорьба за существование – сложные и многообразные отношения особей внутри вида, между видами и с неорганической природой. Несколько раз случайно встречалась там с Женей, они болтали
Борьба за существование — одна из форм взаимоотношений между организмами внутри одного вида и между представителями разных видов, служит фактором эволюции растенийНи в коем случае нельзя утверждать, что в природе полностью отсутствует внутривидовая борьба. Они работали в диком напряжении, и Оля не собиралась праздновать свой день рождения
Борьба за существование – одна из движущих сил эволюции, которую определил Чарльз Дарвин. Она проявляется не просто в схватке насмерть, а, например, и в поединке за ресурсы или в противостояние с условиями окружающей среды. Псих он или нет, в самом деле? Псих имеет полное моральное право молчать и говорить, когда ему хочется, а не когда от него этого ждут
Третья форма борьбы за существование – это борьба с внешними неблагоприятными условиями. Давайте рассмотрим еще один опыт, показывающий связь фенотипов и генотипов в природных условиях. Мушки дрозофилы в природе находят корм либо на вершинах. Зацепа делал вид, что не слышит, не понимает, продолжал листать журнал и готов был провалиться сквозь землю
Зацепа делал вид, что не слышит, не понимает, продолжал листать журнал и готов был провалиться сквозь землю
Причина борьбы за существование: несоответствие между возможностью видов к беспредельному размножению и ограниченностью ресурсов. Формы борьбы: 1. Внутривидовая — между особями одной популяции любого вида. Эту форму борьбы Дарвин считал самой. – Ну, что, лапушка, – вздохнул дядя Мотя, – Молоха твоего мы нашли
Именно, на борьбе за существование и на выживании в этой борьбе наиболее приспособленных особей была основана им вся его замечательная теория естественногоВ окружающей нас природе условия внутривидовой борьбы за существование очень сложны. Мне надо с кем-то поделиться, хотя бы с тобой, мой дневник
Сейчас очень много загрязнений и прочего. В природе организмов появляется во много раз больше, чем их может просуществовать на данном клочке земли. В результате между организмами происходит состязание — борьба за существование. Тогда, полтора года назад, нашли двух девочек и мальчика, убитых маньяком
Борьба с неблагоприятными условиями неорганической природы также усиливает внутривидовое состязание, так как особи одного видаБорьба за существование — сложные и многообразные отношения организмов между собой и с условиями окружающей средысреды. – Это… это… – повторял Борис Александрович, глядя на заколку, – я нашел…
Особи в природе существуют не изолированно, а в определённых биогеоценозах. Борьба за существование — сложные и многообразные взаимоотношения особей внутри вида, между видами и с неблагоприятными условиями неживой природы. Сожительство с малолеткой, да еще дочерью заклятого врага – это круто для желтой прессы
Борьбу за существование Дарвин считал одним из главных движущих факторов эволюции. «Из войны природы, из голода и смерти непосредственно вытекает самый высокий результат… — образование высших животных» (Дарвин, 1991). В природе она выполняет ту же роль, что и. – Пойдем, пойдем, не бойся, – санитар взял его за локоть, – да ты чего, мужик? Вставай! А то клизму поставлю, доктор прописала тебе клизму керосиновую
Межвидовая борьба — борьба за существование между разными видами. Как правило, межвидовая борьба протекает особенно остро, если у видов сильно перекрываются экологические ниши (часто у представителей одного рода или семейства). Можно без отчества? Тут, знаете, замечательные бифштексы с кровью
Как правило, межвидовая борьба протекает особенно остро, если у видов сильно перекрываются экологические ниши (часто у представителей одного рода или семейства). Можно без отчества? Тут, знаете, замечательные бифштексы с кровью
В природе также постоянно происходит отбор. Для всех живых организмов характерна наследственная изменчивость, поэтому все ониБорьба за существование — совокупность многообразных и сложных взаимоотношений между организмами и окружающей средой. Эй, Ник, итальянское мое солнце, разве я с тобой шабашу? У нас любовь, чистая и высокая, блин! Денежки ты мне даешь потому, что тебе нравится баловать твою синьорину, красиво одевать меня, маленькую, и вкусно кормить
Учащиеся записывают в тетрадь вывод: в природе происходит непрерывная борьба за существование. 1. Борьба за существование с неблагоприятными условиями среды. В качестве домашнего задания учитель предлагает нескольким учащимся подготовить. – Как у меня? В последний раз мы виделись в ее день рождения, неделю назад
БОРЬБА ЗА СУЩЕСТВОВАНИЕ — в биологии одно из основных понятий в теории эволюции Ч. Дарвина, которое он употреблял для обозначения отношений между организмами, а также между организмами и абиотическими условиями, приводящих к гибели менее приспособленных и. В общем, твой запрос будет выполнен обычным порядком, не раньше чем через десять рабочих дней
Формы борьбы за существование Сравнительные признаки Внутривидовая борьба Межвидовая борьба Борьба с неблагоприятными условиями 1. Формы проявления борьбы за существование 2. Результаты борьбы за существование 3. Примеры данной формы борьбы. Да, кстати, я видела сегодня утром по телевизору твоего Соловьева
В природе особи борются за лучшие условия существования, в результате чего выживают самые приспособленные из них. Межвидовая борьба за существование, примеры которой мы рассмотрим, возникает в случае, когда пересекаются экологические ниши, ареалы. – Кроме ритуала и сексуального возбуждения есть еще момент вполне прагматический
Борьба за существование — движущий фактор эволюции, совокупность взаимоотношений особей животного и растительного мира между собой и с окружающим миром. Форма 1. Взаимоотношение организмов с неживой природой. Пример борьбы с неблагоприятной средой. Он от нее никуда не денется просто потому, что тяжесть такого груза и жар такого пара никто, кроме нее, не выдержит
Форма 1. Взаимоотношение организмов с неживой природой. Пример борьбы с неблагоприятной средой. Он от нее никуда не денется просто потому, что тяжесть такого груза и жар такого пара никто, кроме нее, не выдержит
Победители прошлых лет
Алексей БобровскийРуководитель службы экономических программ, телеканал «Россия-24»
Какие книги хочется упомянуть?
«Повелители финансов. Банкиры, перевернувшие мир», Лиакват Ахамед.
Книга о судьбах четырёх экономистов, председателей четырёх главных центральных банков мира, в период между Первой и Второй мировыми войнами. Из книги вы узнаете, что все беды того периода, такие как Великая депрессия, крах мировой экономики, последующие изменения в мировой политике стали результатом решений, которые принимали эти четыре джентельмена: Монтегю Норман, глава Банка Англии, Бенджамин Стронг, управляющий Федеральным резервным банком Нью-Йорка, Ялмар Шахт, возглавлявший Рейхсбанк, и глава Банка Франции Эмиль Моро. Конечно, читатель сам сделает вывод, кто виноват и что делать, но взгляд на события первой половины 20 века через призму мировых финансов и систему управления ими, уверен, даёт возможность увидеть такие судьбоносные моменты истории, как причины начала Первой мировой войны, её итоги, Версальский мирный договор, предпосылки для начала Второй мировой войны, под совершенно иным ракурсом. Этот ракурс помогает, например, увидеть взаимосвязь между изменением денежно-кредитной политики и началом масштабных боевых действий. Конечно, всегда войны начинают политики, ведут военные, а заканчивают дипломаты, но, вот, почему иногда, страны оказываются на пороге конфликта, какие решения приводят к этому. Посмотрите на это через экономику и финансы. Эта книжка — лучший способ.
«Добыча: Всемирная история борьбы за нефть, деньги и власть», Дэниел Ергин.
В принципе, название книги говорит само за себя. Её точно надо прочитать желающим систематизировать свои знания в этой области. Борьба за нефть, её транспортировка, переработка, продажа — это история всего 20 века. Почему во время Второй мировой нельзя было сдавать Сталинград и Кавказ, а японцы напали на Пёрл Харбор, какие решения помогли в ходе Первой мировой британцам одержать победу на море, что стало новой целью так называемой «Большой игры» во второй половине прошлого и уже нового века? Ответ прост: нефть и еще раз нефть! Самый главный вопрос: как она (нефть) стала одним из определяющих факторов развития мировой экономики сегодня? Для понимания темы нефтедобычи и вообще сырьевого рынка, книга, на мой взгляд — своего рода хрестоматия. Глубже тему раскроет только уже профессиональная литература, интересней — ничего.
Борьба за нефть, её транспортировка, переработка, продажа — это история всего 20 века. Почему во время Второй мировой нельзя было сдавать Сталинград и Кавказ, а японцы напали на Пёрл Харбор, какие решения помогли в ходе Первой мировой британцам одержать победу на море, что стало новой целью так называемой «Большой игры» во второй половине прошлого и уже нового века? Ответ прост: нефть и еще раз нефть! Самый главный вопрос: как она (нефть) стала одним из определяющих факторов развития мировой экономики сегодня? Для понимания темы нефтедобычи и вообще сырьевого рынка, книга, на мой взгляд — своего рода хрестоматия. Глубже тему раскроет только уже профессиональная литература, интересней — ничего.
«Анализ ценных бумаг», Бенджамин Грехэм и Дэвид Додд.
Уоррен Баффет говорит, что это его настольная книга. Судя по месту «оракула из Омахи» в списке Forbes, на это обстоятельство точно стоит обратить внимание, а возможно, после прочтения этого издания, станет понятно, что он не совсем и оракул. Вообще-то, даже опытные участники фондового рынка рекомендуют читать вместе с данной работой Бенджамина Грехэма еще и другой его труд под названием «Разумный инвестор», о чем пишет и сам Уоррен в предисловии. Но и одной этой книги желающему постичь особенности такого процесса, как инвестирование и анализ ценных бумаг, будет достаточно для постижения азов этого дела. Без специального образования читать годовые отчеты компаний трудно, а как быть миноритарными акционерами, не понимая ничего в этих документах?! Книга Грехэма и Додда дает возможность и недипломированным специалистам узнать все нужное о работе компаний из финансового отчета, чтобы потом принять правильное решение на фондовом рынке.
«Следующие 100 лет: Прогноз событий XXI века», Джордж Фридман.
Бывший глава Stratfor Джордж Фридман — личность одиозная. Stratfor до сих пор называют филиалом ЦРУ, а самого Фридмана — одним из тех, кто в США занимается программированием будущего. Любителям конспирологии такой рекомендации было бы уже достаточно. Но для тех, кто со скепсисом относится, как к разговорам о форсайтинге, так и к утверждению, что «Американская Империя» в 21 веке продолжит доминировать, стоит прочитать эту книгу, хотя бы для того, что понимать арсенал аргументов политологов и экономистов, придерживающихся все-таки этой точки зрения. Это интереснее, чем читать «динозавра» Бжезинского, у которого кроме геополитической теории мало полезного для сегодняшнего дня. Фридман же многие события, свидетелями которых мы становимся сегодня, предсказал еще несколько лет назад — до Трампа, Украины, Сирии, спора за Южно-Китайское море и кризиса Европы. Ясно, что реально прогнозировать можно в перспективе лет 20, все дальше — это фантастический роман. Но книга — хороший повод поразмыслить на тему, что первично: политика или экономика? Фридман в этом споре где-то посередине».
Но для тех, кто со скепсисом относится, как к разговорам о форсайтинге, так и к утверждению, что «Американская Империя» в 21 веке продолжит доминировать, стоит прочитать эту книгу, хотя бы для того, что понимать арсенал аргументов политологов и экономистов, придерживающихся все-таки этой точки зрения. Это интереснее, чем читать «динозавра» Бжезинского, у которого кроме геополитической теории мало полезного для сегодняшнего дня. Фридман же многие события, свидетелями которых мы становимся сегодня, предсказал еще несколько лет назад — до Трампа, Украины, Сирии, спора за Южно-Китайское море и кризиса Европы. Ясно, что реально прогнозировать можно в перспективе лет 20, все дальше — это фантастический роман. Но книга — хороший повод поразмыслить на тему, что первично: политика или экономика? Фридман в этом споре где-то посередине».
А
NUH — это буква, которую я использую для написания Nutches, Доктор Сьюз — По ту сторону зебры (1955) Желаем узнать:
Основные понятия и определенияБиологи и естествоиспытатели издавна были очарованы различиями между видами. Галапагосские острова
зяблики — известный пример множества мелких, но мы считаем важными
различия между несколькими видами, которые в остальном так похожи друг на друга
они побудили Дарвина предположить, что все они были модификациями (потомками)
одного вида. Эти зяблики действительно отличаются важными
аспекты размера тела, размера и формы клюва, питаются ли они насекомыми
или семена, на крупные или мелкие семена и так далее. Кажется, у каждого своя
собственная ниша, концепция, которую мы определим более тщательно в ближайшее время.Но
предположим, мы посмотрим на эту группу зябликов по-другому, обратите внимание, насколько похожи
они такие, и спросите, могут ли какие-то два вида быть слишком похожими по своему
потребности в ресурсах? Насколько разными должны быть два вида, чтобы сосуществовать в природе?
Что на самом деле представляет собой ниша и могут ли ниши пересекаться? Делают популяции в природе
конкурировать? Галапагосские острова
зяблики — известный пример множества мелких, но мы считаем важными
различия между несколькими видами, которые в остальном так похожи друг на друга
они побудили Дарвина предположить, что все они были модификациями (потомками)
одного вида. Эти зяблики действительно отличаются важными
аспекты размера тела, размера и формы клюва, питаются ли они насекомыми
или семена, на крупные или мелкие семена и так далее. Кажется, у каждого своя
собственная ниша, концепция, которую мы определим более тщательно в ближайшее время.Но
предположим, мы посмотрим на эту группу зябликов по-другому, обратите внимание, насколько похожи
они такие, и спросите, могут ли какие-то два вида быть слишком похожими по своему
потребности в ресурсах? Насколько разными должны быть два вида, чтобы сосуществовать в природе?
Что на самом деле представляет собой ниша и могут ли ниши пересекаться? Делают популяции в природе
конкурировать?Теперь у нас есть множество доказательств того, что конкуренция является важным фактором, влияющим на современные растения и животные распределений, и что конкуренция играет существенную роль в эволюции путем естественного отбора.Сегодня мы рассмотрим это свидетельство и развитие нашего понимания предмета. Начнем с определения конкуренции, потому что это обеспечивает систему отсчета, с которой можно исследовать доказательства. Конкуренция возникает, когда два вида для каждого из них требуется ресурс, которого не хватает, так что доступность ресурса на один вид отрицательно влияет наличие другого вида. Это взаимодействие типа «- / -». Конкуренция не происходит, если ресурсов слишком много, чтобы ограничить рост, распространение или изобилие хотя бы одной из популяций.Мы ожидаем только один или несколько ресурсов быть ограничивающим и, следовательно, за него нужно бороться. Соревнование может происходить между людьми
которые принадлежат к одному виду. Это называется внутривидовой конкуренцией.
Интуитивно кажется, что внутривидовая конкуренция будет интенсивной, потому что
два представителя одного и того же вида, вероятно, будут иметь очень похожие ресурсы
потребности. Три классических исследования конкуренцииПарамеций: лабораторный «флакон» эксперимент — Paramecium — одноклеточное мерцательное простейшее, которое питается бактериями. Его можно выращивать в пробирке, в бульоне бактерий, который
Исследование парамеций подняло ряд
интересных вопросов. Всегда ли побеждает один и тот же вид? Может два
виды когда-либо сосуществуют? Насколько хорошо мы можем экстраполировать данные этой лаборатории
системы к природе? В настоящее время выполнено много таких «бутылочных» исследований,
с мучными жуками, дрозофилами, дрожжами, различными растениями. Нет двух видов одинаковых требований могут долго занимать одну и ту же нишу (сосуществовать). В случае парамеции то же самое
виды всегда побеждали. Напротив, исследования мучных жуков показали, что
изменения в микросреде (температура, влажность) могут изменить результат
в пользу того или иного вида. Простота таких экспериментов
остается насущной проблемой: действительно ли реальные популяции, живущие в комплексе,
переменный мир, также испытываете конкурентное исключение?
Тщательное изучение пищевого поведения
из этих пяти певчих птиц обнаружили много различий в способах использования
кажущейся однородной среды обитания.Эти виды различались тем, в какой части
ели, которую они часто посещали, ловили ли они насекомых на
крыло, из хвои или под корой и т. д. Был сделан вывод, что каждый
вид занял отчетливую нишу. Вместо того, чтобы показывать это соревнование
было важным по своей природе, это исследование, казалось, установило обратное. Выравнивать
Внешне похожие виды действительно отличаются друг от друга при ближайшем рассмотрении.
Это исследование помогло установить идею о том, что каждый вид занимает свою
собственная отличная ниша. Ниша вида включает все аспекты его среды обитания, того, как он зарабатывает на жизнь, и физической среды в котором он находится. Конкуренция можно представить в точки пересечения ниши. Еще одно соображение, поднятое, но без ответа Согласно данному исследованию, это степень «допустимого» перекрытия ниш. Наблюдательный исследования, подобные этому, не дают очень четкого ответа на вопрос, насколько ниши камышевок пересекались, так как все пять видов сосуществовали, ответ вроде бы «не чрезмерно». Моллюски в приливной зоне:
полевое экспериментальное исследование — ракушек встречаются на почти вертикальных скалах
стены в приливной зоне, что означает, что они
раз и подвергались воздействию воздуха в других.Встречаются приливные потоки в несколько метров.
два раза в день.
Это исследование помогло
подход полевых экспериментов. Использование клеток для исключения одного или другого
видов, и перемещая каменные плиты из одного региона в другой,
было установлено, что Chthamalus мог процветать в нижней зоне, если был защищен
от Балана.В противном случае Balanus, более крупный и крепкий вид,
раздавил соседний Хтамалус по мере того, как он увеличивался в размерах. Chthamalus был конкурентоспособным
исключены из нижнего прилива. Однако на верхнем пределе оба
виды были ограничены их способностью противостоять высыханию. Chthamalus,
худший конкурент, был лучше приспособлен к высыханию и поэтому был занят
верхняя часть приливной зоны. Механизмы конкуренцииСоревнование часто происходит через использование или истощение ресурсов представителями одного вида, что делает их ресурсы, недоступные представителям второго вида.Ресурсы будут включать места гнезд, продукты питания, воду или минералы в почве. Это вызвал косвенную конкуренцию . Вероятно, будет наиболее серьезным между люди со схожими потребностями, например, два вида, занимающие одно и то же трофический уровень и с аналогичными экологическими ролями или нишами. Прямая конкуренция возникает, когда участники
одного вида мешают или непосредственно причиняют вред представителям второго вида.
Примеры включают драки между животными из-за космоса, выброс химикатов.
по растениям (например,g., шалфей, куст креозота), подавляющие рост другого растения
виды и жалящие щупальца кораллов. Конкуренция и эволюционная специализацияКонкуренция может привести к эволюционному дивергенция и специализация близкородственных видов. Рассмотрим процесс видообразования, которое часто считается постепенным и включает период отбора для репродуктивной изоляции, когда географическая изоляция больше не будет разделяет две расходящиеся популяции.Мы должны ожидать этих двух видов занимать очень похожие экологические ниши и, следовательно, иметь возможность конкурировать. Вполне возможно, что один вид уничтожит другой. Также возможно,
однако, что эти два вида будут экологически отличаться, возможно, из-за дивергенции.
в таких характеристиках, как размер счета, который влияет на размер потребляемых продуктов.
Это называется расхождением характера. Вполне возможно, что один вид уничтожит другой. Также возможно,
однако, что эти два вида будут экологически отличаться, возможно, из-за дивергенции.
в таких характеристиках, как размер счета, который влияет на размер потребляемых продуктов.
Это называется расхождением характера. Галапагосские вьюрки Галапагосские зяблики кажутся хорошие примеры характера расхождение.Три вида наземных зябликов делают особенно сильным кейс. Если предположить, что есть семена разных размеров — от мелких до крупных. — имеет смысл, что клювы разного размера — от маленького до большого — позволяют эффективно обрабатывать и растрескивать семена определенного размера. Если существует только один вид зябликов, возможно, он займет «середину». грунт »клюва и размера семян. Однако, как только произойдет видообразование, следует один вид будет немного меньше, а другой немного больше, естественный отбор следует отдавать предпочтение особям, которые меньше среднего в пределах одного вида, больше среднего в другом.Со временем размер клюва (характер который определяет, какая часть спектра ресурсов потребляется) будет расходятся. Однако есть сложности.
Предположим, что наши большие и маленькие зяблики независимо друг от друга возникли,
и довелось колонизировать различные острова в одном, двух и трех. В
отсутствие конкуренции, как маленькие, так и большие виды могут развиваться
к промежуточному размеру. Это известно как экологический выброс. Будь то
кто-то наблюдает смещение персонажа или экологическое высвобождение, тогда зависит
от порядка и независимости событий видообразования.Это трудно понять,
но хороший
кладограмма
решит проблему. Резюме — Последствия конкуренцииКонкуренция может быть мощной силой влияющие на рост, распространение и численность популяций в природе. Это подтверждается данными лабораторных «бутылочных» экспериментов, полевых наблюдения и полевые эксперименты. Лабораторные исследования приводят к формированию принципа конкурентного исключения (никакие два вида не могут долго занимать в той же нише), а полевые наблюдения говорят о том, что нишевые различия, хотя иногда и незаметно, они неизменно встречаются между сосуществующими видами. Полевые эксперименты убедительно свидетельствуют о конкуренции в природе. Часто,
один вид является более сильным конкурентом, другой лучше противостоит
определенные экологические крайности. Полевые эксперименты убедительно свидетельствуют о конкуренции в природе. Часто,
один вид является более сильным конкурентом, другой лучше противостоит
определенные экологические крайности.Конкуренция — мощная сила естественный отбор. Есть много примеров близкородственных видов, которые различаются «ровно настолько, чтобы уменьшить перекрытие ниш и разрешить сосуществование». Генетический изменение персонажа, влияющее на конкуренцию, в сочетании с выбором чтобы уменьшить конкурентное дублирование, приведите веский аргумент в пользу эволюционного расхождение символов. Конкуренция, вероятно, затрагивает виды
разнообразие. В краткосрочной перспективе конкуренция должна привести к сокращению
количество видов, обитающих на территории, предотвращая появление очень похожих видов
от сопутствующих. Однако в долгосрочной перспективе конкуренция, вероятно, усилится.
разнообразие видов, действуя как фактор специализации и дивергенции. Предлагаемые чтения:
| |
 Это разделение ресурсов между людьми из одной популяции
в конечном итоге ограничивает численность населения в данной области (что
действительно означает, для данного количества ресурсов).Сегодня мы делаем акцент на
межвидовая конкуренция, которая происходит между представителями разных видов.
Интенсивность межвидовой конкуренции следует связывать со сходством
между двумя видами. Мы ожидаем, что это повлияет на количество перекрытий.
в использовании ресурсов (= перекрытие ниш).
Это разделение ресурсов между людьми из одной популяции
в конечном итоге ограничивает численность населения в данной области (что
действительно означает, для данного количества ресурсов).Сегодня мы делаем акцент на
межвидовая конкуренция, которая происходит между представителями разных видов.
Интенсивность межвидовой конкуренции следует связывать со сходством
между двумя видами. Мы ожидаем, что это повлияет на количество перекрытий.
в использовании ресурсов (= перекрытие ниш).  В общем, один
побеждает вид, и один вид уничтожается. Эти эксперименты породили
принципу конкурентного исключения:
В общем, один
побеждает вид, и один вид уничтожается. Эти эксперименты породили
принципу конкурентного исключения:

Межвидовая конкуренция — обзор
2. Межвидовая конкуренция среди простейших
Межвидовая конкуренция в лабораторных культурах инфузорий изучалась Гаузе и многочисленными авторами с тех пор. Гаузе (1934) предоставил четкие доказательства того, что факторы окружающей среды могут влиять на исход конкуренции.
Уравнения Лотки – Вольтерра предсказывают, что победителем в эксплуатационной конкуренции за ресурсы в стабильной окружающей среде должен быть вид с большей K или несущей способностью, то есть более эффективным пользователем ресурса.Однако K обычно измеряется в виде чисел, а не биомассы, поэтому более мелкие виды, как правило, имеют более высокое значение K . В своих исследованиях конкурентных взаимодействий бактериоядных инфузорий Luckinbill (1979) подтвердил, что K не предсказывает исход конкуренции. Но, в соответствии с частью гипотезы отбора r / K , внутренняя скорость роста, r , была отрицательно связана с конкурентоспособностью. [Гипотеза отбора r / K утверждает, что r , который измеряет приспособленность в независимой от плотности или стохастической среде, отрицательно коррелирует между видами с K , которая измеряет приспособленность в стабильной среде.] Таким образом, теорема, кажется, верна для инфузорий, за исключением того, что K , будучи сильно коррелированным с размером тела, не измеряет конкурентоспособность в стабильной среде. Как отмечалось выше, r также коррелирует с размером тела.
Но, в соответствии с частью гипотезы отбора r / K , внутренняя скорость роста, r , была отрицательно связана с конкурентоспособностью. [Гипотеза отбора r / K утверждает, что r , который измеряет приспособленность в независимой от плотности или стохастической среде, отрицательно коррелирует между видами с K , которая измеряет приспособленность в стабильной среде.] Таким образом, теорема, кажется, верна для инфузорий, за исключением того, что K , будучи сильно коррелированным с размером тела, не измеряет конкурентоспособность в стабильной среде. Как отмечалось выше, r также коррелирует с размером тела.
Чтобы исключить эту корреляцию, Тейлор (1978a, b) выразил внутреннюю скорость увеличения как r / r e , где r e — ожидаемое r для инфузории на основе регресс р по объему кузова.Следовательно, r / r e является мерой того, насколько быстро растет или r — выбранная инфузория по размеру тела. Было обнаружено, что этот показатель отрицательно коррелирует с частотой встречаемости в полевых условиях и положительно коррелирует с макроядерной плоидностью (Taylor and Shuter, 1981). Это исследование предполагает, что сообщества простейших содержат быстрорастущие, но эфемерные оппортунисты, а также более консервативные, специализированные виды, которые имеют более скромные темпы роста, но стабильные популяции.Аналогичная картина возникла при сравнении динамики популяций относительно быстро и относительно медленно растущих видов в лаборатории (Luckinbill and Fenton, 1978).
Полевые исследования эксплуататорской конкуренции простейших немногочисленны, как и полевые измерения скорости роста для сравнения с лабораторными оценками в условиях избытка пищи. Соответственно, мы мало знаем об относительной важности зависимости или независимости от плотности. Работа с планктонными протистами предполагает, что полевые популяции могут испытывать периоды быстрого роста и периоды небольшого роста или его отсутствия (Taylor and Johannsson, 1991; Carrick et al. , 1992).
, 1992).
Магуайр (1963a, b) нашел убедительные доказательства исключения колонизирующих видов из рода Colpoda со стороны Paramecium и связанных с ним видов. Хейрстон и Келлерман (1965) и Хейрстон (1967) обнаружили, что один член комплекса видов Paramecium aurelia доминировал над симпатрическими родственными видами в экспериментах по конкуренции и преобладал в полевых сборах. Полевое население, которое они изучали, оказалось ограниченным в пище. Напротив, Тейлор (1983b) обнаружил, что сидячие инфузории на искусственном субстрате в ручье имели высокие темпы роста и смертности, что подразумевает небольшую конкуренцию.Обширные исследования колонизации, проведенные Кэрнсом и его сотрудниками (рассмотренные в Cairns and Yongue, 1977), предоставляют косвенные доказательства взаимодействия видов, включая конкуренцию.
Существует также конкуренция между простейшими и многоклеточными животными, потребляющими аналогичные ресурсы. Потенциальные конкуренты многоклеточных животных включают некоторых коловраток и ракообразных, хотя в некоторых случаях трудно различить эффекты конкуренции и хищничества (Wickham and Gilbert, 1993). Точно так же присутствие хищных простейших может повлиять на результат конкуренции между другими простейшими видами (Lawler, 1993).
Данные полевых экспериментов на JSTOR
Abstract В строго определенной выборке исследований конкуренции с использованием контролируемых полевых экспериментов, охватывающей 215 видов и 527 экспериментов, конкуренция была обнаружена в большинстве исследований, чуть более чем у половины видов и примерно в двух пятых экспериментов. В большинстве этих экспериментов межвидовая конкуренция не отличалась от внутривидовой конкуренции. В нескольких исследованиях, в которых эти два исследования были разделены, межвидовая конкуренция была более сильной примерно в шестой части всех проведенных экспериментов.Когда конкуренция была продемонстрирована, внутривидовая конкуренция была столь же сильной или более сильной, чем межвидовая в трех четвертях экспериментов. Некоторые данные этого обзора литературы предполагают, что отрицательные результаты могут быть недостаточно представлены, так что абсолютные значения этих цифр могут быть слишком высокими. Поскольку это смещение должно применяться также к исследованиям всех таксонов, местообитаний или других взаимодействий, оно не должно сильно влиять на оценки относительной распространенности конкуренции. Поскольку эти оценки получены в результате полевых экспериментов, открытых для других факторов, таких как хищники, травоядные, погода, беспокойство и т. Д., они должны обеспечивать справедливую аппроксимацию относительной распространенности межвидовой и внутривидовой конкуренции в естественных экологических сообществах. Преобладание конкуренции в этих исследованиях было разным. Морские организмы демонстрировали постоянно более высокую частоту конкуренции, чем наземные, как и крупные организмы по сравнению с более мелкими. Растения, травоядные и плотоядные животные показали одинаковую частоту конкуренции во всех сравниваемых средах обитания. Частота конкуренции значительно варьировалась от года к году и от места к месту.В некоторых категориях данные о конкуренции немногочисленны. Необходимы дополнительные исследования всех пресноводных видов, морских позвоночных, паразитов, влияния на разделение ресурсов и, в частности, относительной силы межвидовой и внутривидовой конкуренции. Когда были изучены оба члена пары и обнаружена некоторая конкуренция, более чем в половине экспериментов пострадал только один член. Такая сильная асимметричная конкуренция не всегда последовательна по направлению; изменения в порядке ранжирования конкурентного превосходства были продемонстрированы полевыми экспериментами и прямыми наблюдениями.Были обнаружены некоторые положительные взаимодействия. Это могло быть следствием реальных положительных влияний или отрицательных влияний, действующих косвенно через другие виды. Последнее может также относиться к некоторым негативным взаимодействиям, интерпретируемым в этих исследованиях как конкуренция.
Некоторые данные этого обзора литературы предполагают, что отрицательные результаты могут быть недостаточно представлены, так что абсолютные значения этих цифр могут быть слишком высокими. Поскольку это смещение должно применяться также к исследованиям всех таксонов, местообитаний или других взаимодействий, оно не должно сильно влиять на оценки относительной распространенности конкуренции. Поскольку эти оценки получены в результате полевых экспериментов, открытых для других факторов, таких как хищники, травоядные, погода, беспокойство и т. Д., они должны обеспечивать справедливую аппроксимацию относительной распространенности межвидовой и внутривидовой конкуренции в естественных экологических сообществах. Преобладание конкуренции в этих исследованиях было разным. Морские организмы демонстрировали постоянно более высокую частоту конкуренции, чем наземные, как и крупные организмы по сравнению с более мелкими. Растения, травоядные и плотоядные животные показали одинаковую частоту конкуренции во всех сравниваемых средах обитания. Частота конкуренции значительно варьировалась от года к году и от места к месту.В некоторых категориях данные о конкуренции немногочисленны. Необходимы дополнительные исследования всех пресноводных видов, морских позвоночных, паразитов, влияния на разделение ресурсов и, в частности, относительной силы межвидовой и внутривидовой конкуренции. Когда были изучены оба члена пары и обнаружена некоторая конкуренция, более чем в половине экспериментов пострадал только один член. Такая сильная асимметричная конкуренция не всегда последовательна по направлению; изменения в порядке ранжирования конкурентного превосходства были продемонстрированы полевыми экспериментами и прямыми наблюдениями.Были обнаружены некоторые положительные взаимодействия. Это могло быть следствием реальных положительных влияний или отрицательных влияний, действующих косвенно через другие виды. Последнее может также относиться к некоторым негативным взаимодействиям, интерпретируемым в этих исследованиях как конкуренция. Если известны только ввод и вывод эксперимента, трудно решить, какой механизм произвел наблюдаемый эффект. Хотя многие эксперименты, вероятно, были правильно истолкованы, настоящий обзор показывает, насколько сложно произвести четкую и недвусмысленную демонстрацию межвидовой конкуренции.
Если известны только ввод и вывод эксперимента, трудно решить, какой механизм произвел наблюдаемый эффект. Хотя многие эксперименты, вероятно, были правильно истолкованы, настоящий обзор показывает, насколько сложно произвести четкую и недвусмысленную демонстрацию межвидовой конкуренции.
Текущие выпуски теперь размещены на веб-сайте Chicago Journals. Прочтите последний выпуск. С момента своего создания в 1867 году журнал The American Naturalist сохраняет свои позиции в качестве одной из ведущих рецензируемых публикаций в мире по вопросам экологии, эволюции и исследований поведения. Его цели — публиковать статьи, представляющие широкий интерес читателям, ставить новые и важные проблемы, знакомить с новыми предметами, развивать концептуальную унификацию и изменять образ мышления людей.AmNat делает упор на сложные методологии и новаторские теоретические синтезы — все в попытке продвинуть знания об органической эволюции и других общих биологических принципах.
Информация об издателеС момента своего возникновения в 1890 году как одно из трех основных подразделений Чикагского университета, University of Chicago Press взяла на себя обязательство распространять стипендии высочайшего стандарта и публиковать серьезные работы, способствующие образованию, содействию развитию образования. общественное понимание и обогащение культурной жизни.Сегодня Отдел журналов издает более 70 журналов и сериалов в твердом переплете по широкому кругу академических дисциплин, включая социальные науки, гуманитарные науки, образование, биологические и медицинские науки, а также физические науки.
Эмпирическая оценка силы межвидовой конкуренции в формировании сообществ мелких млекопитающих на фрагментированных ландшафтах
Abt KF, Bock WF (1998) Сезонные колебания состава рациона полевых мышей на сельскохозяйственных угодьях Apodemus spp.и рыжих полевок Clethrionomys glareolus . Acta Theriol 43: 379–389
Acta Theriol 43: 379–389
Статья Google Scholar
Абу Бейкер М.А., Браун Дж.С. (2014) Поиск пищи в пространстве и времени структурирует африканское сообщество мелких млекопитающих. Oecologia 175: 521–535. DOI: 10.1007 / s00442-014-2926-6
Артикул PubMed Google Scholar
Amarasekare P (2003) Конкурентное сосуществование в пространственно структурированной среде: синтез.Ecol Lett 6: 1109–1122. DOI: 10.1046 / j.1461-0248.2003.00530.x
Артикул Google Scholar
Амвросий Х.В. (1972) Влияние знакомства среды обитания и стрижки пальцев на скорость хищничества сов у Microtus pennsylvanicus . J Mammal 53: 909–912. DOI: 10.2307 / 1379235
Артикул Google Scholar
Amori G, Contoli L, Nappi A (2008) Fauna d’Italia: Mammalia II.Edizioni Calderini de Il Sole 24 ORE Edagricole, Bologna
Amori G, Cristaldi M, Fanfani A, Solida L, Luiselli L (2010) Экологическое сосуществование популяций с низкой плотностью Apodemus sylvaticus и A. flavicollis ( Mammalia: Rodentia). Ренд Линчеи 21: 171–182. DOI: 10.1007 / s12210-010-0076-2
Артикул Google Scholar
Amstrup S, McDonald L, Manly B (2006) Справочник по анализу захвата-повторного захвата.Princeton University Press, Princeton, NJ
Andrzejewski R, Olszewski JL (1963) Социальное поведение и межвидовые отношения у Apodemus flavicollis (Melchior, 1834) и Clethrionomys glareolus (Schreber, 1780). Acta Theriol 7: 155–168
Артикул Google Scholar
Boeye J, Kubisch A, Bonte D (2014) Структура среды обитания опосредует пространственную сегрегацию и, следовательно, сосуществование. Пейзаж Ecol 29: 593–604. DOI: 10.1007 / s10980-014-0010-6
Пейзаж Ecol 29: 593–604. DOI: 10.1007 / s10980-014-0010-6
Артикул Google Scholar
Bowers MA, Dooley JLJ (1999) Контролируемое иерархическое исследование фрагментации среды обитания: реакции на индивидуальном уровне, в масштабе участка и в масштабе ландшафта. Пейзаж Ecol 14: 381–389
Статья Google Scholar
Braschler B, Baur B (2005) Экспериментальная мелкомасштабная фрагментация пастбищ изменяет конкурентные взаимодействия между видами муравьев.Oecologia 143: 291–300. DOI: 10.1007 / s00442-004-1778-x
Артикул PubMed Google Scholar
Braude S, Ciszek D (1998) Выживание голых землекопов, отмеченных имплантируемыми транспондерами и обрезанием пальцев ног. J Mammal 79: 360–363. DOI: 10.2307 / 1382873
Артикул Google Scholar
Brown WP (2007) Масса тела, общность местообитаний и состав птичьего сообщества в остатках леса.J Biogeogr 34: 2168–2181. DOI: 10.1111 / j.1365-2699.2007.01766.x
Артикул Google Scholar
Brunner JL, Duerr S, Keesing F, Killilea M, Vuong H, Ostfeld RS (2013) Экспериментальный тест конкуренции между мышами, бурундуками и белками во фрагментах лиственных лесов. PLoS One 8: e66798. DOI: 10.1371 / journal.pone.0066798
CAS Статья PubMed PubMed Central Google Scholar
Бухальчик Т., Ольшевский Ю.Л. (1971) Поведенческая реакция лесных грызунов на ловушку и наживку.Acta Theriol 16: 233–239
Google Scholar
Buesching CD, Newman C, Twell R, Macdonald DW (2008) Причины древесной активности у древесных мышей Apodemus sylvaticus и рыжих полевок Myodes glareolus . Мамм Биол 73: 318–324. DOI: 10.1016 / j.mambio.2007.09.009
Мамм Биол 73: 318–324. DOI: 10.1016 / j.mambio.2007.09.009
Google Scholar
Бернем К.П., Андерсон Д.Р. (2002) Выбор модели и многомодельный вывод: практический теоретико-информационный подход, 2-е изд.Спрингер, Нью-Йорк
Google Scholar
Canova L (1993) Разделение ресурсов между рыжей полевкой Clethrionomys glareolus и лесной мышью Apodemus sylvaticus в лесных местообитаниях. Bolletino di Zool 60: 193–198. DOI: 10.1080 / 11250009309355809
Артикул Google Scholar
Capizzi D, Luiselli L (1996) Экологические взаимосвязи между мелкими млекопитающими и возрастом поросли в дубово-смешанном лесу в центральной Италии.Rev Eco (Terre Vie) 51: 277–291
Google Scholar
Chesson P (2000) Механизмы поддержания видового разнообразия. Annu Rev Ecol Evol Syst 31: 343–366
Статья Google Scholar
Цихакова Дж., Фринта Д. (1996) Внутривидовые и межвидовые поведенческие взаимодействия у лесной мыши ( Apodemus sylvaticus ) и мыши с желтой шеей ( Apodemus flavicollis ) в нейтральной клетке.Folia Zool 45: 105–113
Google Scholar
Дармон Дж., Календж С., Лойсон А., Джуллиен Дж. М., Майярд Д., Лопес Дж. Ф. (2012) Пространственное распределение и выбор среды обитания у сосуществующих видов горных копытных. Экография 35: 44–53. DOI: 10.1111 / j.1600-0587.2011.06664.x
Артикул Google Scholar
Дули Дж., Бауэрс М. (1998) Демографические реакции на фрагментацию среды обитания: экспериментальные испытания в масштабе ландшафта и участка.Экология 79: 969–980
Статья. Google Scholar
Google Scholar
Даггер К.М., Энтони Р.Г., Эндрюс Л.С. (2011) Временная динамика агрессивной конкуренции: полосатые совы, пятнистые совы, среда обитания и демоны конкуренции. Ecol Appl 21: 2459–2468
Артикул PubMed Google Scholar
Ecke F, Löfgren O, Sörlin D (2002) Динамика популяций мелких млекопитающих в зависимости от возраста леса и структурных факторов среды обитания на севере Швеции.J Appl Ecol 39: 781–792
Статья Google Scholar
Fahrig L (2003) Влияние фрагментации среды обитания на биоразнообразие. Annu Rev Ecol Evol Syst 34: 487–515. DOI: 10.1146 / annurev.ecolsys.34.011802.132419
Артикул Google Scholar
Фариг Л. (2013) Переосмысление размера участка и эффектов изоляции: гипотеза о количестве местообитаний.Журнал Биогеогр 40: 1649–1663. DOI: 10.1111 / jbi.12130
Артикул Google Scholar
Fairley JS (1982) Кратковременные эффекты звона и отсечения пальцев на повторной поимке древесных мышей ( Apodemus sylvaticus ). J Zool 197: 295–297
Google Scholar
Фасола М., Канова Л. (2000) Асимметричная конкуренция между рыжей полевкой и лесной мышью, эксперимент по удалению.Acta Theriol 45: 353–365
Артикул Google Scholar
Fischer J, Lindenmayer DB (2007) Модификация ландшафта и фрагментация среды обитания: синтез. Glob Ecol Biogeogr 16: 265–280. DOI: 10.1111 / j.1466-8238.2006.00287.x
Артикул Google Scholar
Фишер Д.О., Пломберг С.П. (2009) Обрезка почек у молодых мелких сумчатых для экологических полевых исследований: отрицательного воздействия на рост или выживание не обнаружено. Austral Ecol 34: 858–865
Austral Ecol 34: 858–865
Статья Google Scholar
Фишер Дж. Т., Анхольт Б., Брэдбери С., Уитли М., Вольпе Дж. П. (2013) Пространственная сегрегация симпатрической куницы и рыбаков: влияние ландшафтов и видов-ландшафтов. Экография 36: 240–248. DOI: 10.1111 / j.1600-0587.2012.07556.x
Артикул Google Scholar
Fitzgibbon CD (1997) Мелкие млекопитающие в сельскохозяйственных лесах: влияние среды обитания, изоляции и окружающих моделей землепользования.J Appl Ecol 34: 530–539
Статья Google Scholar
Gębczyńska Z (1983) Кормление. В кн .: Петрусевич К. (ред.) Экология рыжей полевки. Acta Theriologica XXVIII (Дополнение № 1), стр. 40–49.
Гибб Х., Хочули Д.Ф. (2002) Фрагментация среды обитания в городской среде: большие и маленькие фрагменты поддерживают различные сообщества членистоногих. Biol Conserv 106: 91–100
Статья Google Scholar
Ginger SM, Hellgren EC, Kasparian MA, Levesque LP, Engle DM, Leslie DM Jr (2003) Смещение ниши опоссумом Вирджины после сокращения предполагаемого конкурента, енота.J Mammal 84: 1279–1291
Статья Google Scholar
Greenwood PJ (1978) Время активности рыжей полевки ( Clethrionomys glareolus ) и лесной мыши ( Apodemus sylvaticus ) в лиственном лесу. Oikos 31: 123–127
Статья Google Scholar
Харрис С., Ялден Д. (2008) Млекопитающие Британских островов: справочник. Общество млекопитающих, Саутгемптон
Google Scholar
Henle K, Davies KF, Kleyer M, Margules C, Settele J (2004) Предикторы чувствительности видов к фрагментации. Biodivers Conserv 13: 207–251. DOI: 10.1023 / B: BIOC.0000004319..9e
Biodivers Conserv 13: 207–251. DOI: 10.1023 / B: BIOC.0000004319..9e
Артикул Google Scholar
Hoffmeyer I (1973) Взаимодействие и выбор среды обитания у мышей Apodemus flavicollis и A. sylvaticus . Oikos 24: 108–116
Статья Google Scholar
Hoffmeyer I, Hansson L (1974) Изменчивость в количестве и распределении Apodemus flavicollis (Melch.) и A. sylvaticus (L.) в Южной Швеции. Z für Säugetierkd 39: 15–23
Google Scholar
Holland GJ, Bennett AF (2010) Фрагментация среды обитания нарушает демографию широко распространенных местных млекопитающих. Экография 33: 841–853. DOI: 10.1111 / j.1600-0587.2010.06127.x
Артикул Google Scholar
Kalinowska A (1971) Отлов Apodemus flavicollis и Clethrionomys glareolus в двойные ловушки.Acta Theriol 16: 73–78
Артикул Google Scholar
Кэт Дж., Марон М., Данн П.К. (2009) Межвидовая конкуренция и разнообразие мелких птиц в урбанизирующем ландшафте. Пейзажный городской план 92: 72–79. DOI: 10.1016 / j.landurbplan.2009.02.007
Артикул Google Scholar
Korn H (1987) Влияние отлова живого и подрезания пальцев ног на массу тела европейских и африканских грызунов.Oecologia 71: 597–600
Статья Google Scholar
Kotzageorgis GC, Mason CF (1997) Популяции мелких млекопитающих в связи со структурой живой изгороди в пахотном ландшафте. J Zool 242: 425–434
Статья Google Scholar
Lambin X, Bauchau V (1989) Соревнование между лесными мышами и рыжими полевками: есть ли победитель? Acta Theriol 34: 385–390
Артикул Google Scholar
Lindenmayer DB, Fischer J (2006) Фрагментация среды обитания и изменение ландшафта. Экологический и природоохранный синтез. Island Press, Вашингтон, округ Колумбия
Экологический и природоохранный синтез. Island Press, Вашингтон, округ Колумбия
Google Scholar
Маграх А., Лоренс В. Ф., Ларринага А. Р., Сантамария Л. (2014) Мета-анализ воздействия фрагментации леса на межвидовые взаимодействия. Conserv Biol 28: 1342–1348. DOI: 10.1111 / cobi.12304
Артикул PubMed Google Scholar
Margaletic J (2004) Glavas M (2002) Развитие мышей и полевок в дубовом лесу с избытком желудей.J Pest Sci 75: 95–98
Google Scholar
Marsh ACW, Harris S (2000) Разделение лесных ресурсов среды обитания двумя симпатрическими видами Apodemus : уроки по сохранению желтошейных мышей ( A. flavicollis ) в Великобритании. Biol Conserv 92: 275–283
Статья Google Scholar
Макгуайр Б., Гетц Л.Л., Оли М.К. (2002) Фитнес-последствия социальности степных полевок, Microtus ochrogaster : влияние размера и состава группы.Anim Behav 64: 645–654
Статья Google Scholar
Michaux JR, Kinet S, Filippucci MG, Libois R, Besnard A, Catzeflis F (2001) Молекулярная идентификация трех симпатрических видов древесных мышей ( Apodemus sylvaticus , A. flavicollis , A. alpicola ), в Западной Европе (Muridae: Rodentia). Примечания к Mol Ecol 1: 260–263
CAS Статья Google Scholar
Montgomery WI (1978) Внутривидовые и межвидовые взаимодействия Apodemus sylvaticus (L.) и A. flavicollis (Melchior) в лабораторных условиях. Anim Behav 26: 1247–1254
Статья Google Scholar
Montgomery WI (1980) Использование древесных взлетно-посадочных полос лесными грызунами, Apodemus sylvaticus (L. ), A. flavicollis (Melchior) и Clethrionomys glareolus (Schreber). Mamm Rev 10: 189–195
), A. flavicollis (Melchior) и Clethrionomys glareolus (Schreber). Mamm Rev 10: 189–195
Статья Google Scholar
Montgomery WI (1981) Эксперимент по удалению симпатрических популяций Apodemus sylvaticus (L.) и A. flavicollis (Melchior) (Rodentia: Muridae). Oecologia 51: 123–132
Статья Google Scholar
Montgomery WI (1985) Влияние маркировки на повторную поимку и оценку популяций Apodemus spp. J Zool 206: 267–269
Статья Google Scholar
Монтгомери В.И., Дауи М. (1993) Распространение и регулирование численности лесной мыши Apodemus sylvaticus на границах полей пастбищных сельскохозяйственных угодий.J Appl Ecol 30: 783–791
Статья Google Scholar
Монтгомери В.И., Ланди М.Г., Рид Н. (2012) «Инвазионный крах»: свидетельства неожиданных последствий и кумулятивных воздействий многовидовых вторжений. Biol Invasions 14: 1111–1125
Статья Google Scholar
Мортеллити А., Линденмайер ДБ (2015) Влияние трансформации ландшафта на модели колонизации и исчезновения птиц в крупномасштабном долгосрочном эксперименте на природе.Conserv Biol. DOI: 10.1111 / cobi.12523
Google Scholar
Mortelliti A, Amori G, Annesi F, Boitani L (2009) Тестирование относительного вклада соседства участка, внутренней структуры участка и присутствия хищников и видов-конкурентов в определении моделей распределения грызунов в фрагментированном ландшафте. Can J Zool 87: 662–670
Статья Google Scholar
Mortelliti A, Amori G, Boitani L (2010a) Роль качества среды обитания в фрагментированных ландшафтах: концептуальный обзор и перспективы для будущих исследований. Oecologia 163: 535–547. DOI: 10.1007 / s00442-010-1623-3
Oecologia 163: 535–547. DOI: 10.1007 / s00442-010-1623-3
Артикул PubMed Google Scholar
Mortelliti A, Amori G, Capizzi D, Rondinini C, Boitani L (2010b) Экспериментальный план и таксономический объем исследований фрагментации европейских млекопитающих: текущее состояние и будущие приоритеты. Мамм Rev 40: 125–154. DOI: 10.1111 / j.1365-2907.2009.00157.x
Артикул Google Scholar
Nupp TE, Swihart RK (2001) Оценка конкуренции грызунов между лесными грызунами на фрагментированном ландшафте Среднего Запада США.Мамм Биол 66: 345–356
Google Scholar
Ostfeld RS, Miller MC, Schnurr J (1993) Мечение ушей увеличивает заражение клещами ( Ixodes dammini ) у белоногих мышей ( Peromyscus leucopus ). J Mammal 74: 651–655
Статья Google Scholar
Павоне Л.В., Бунстра Р. (1985) Влияние отсечения пальца ноги на выживаемость луговой полевки ( Microtus pennsylvanicus ).Может J Zool 63: 499–501. DOI: 10.1139 / z85-072
Артикул Google Scholar
Pollock KH (1982) Конструкция по захвату-повторной поимке, устойчивая к неравной вероятности поимки. J Wildl Manag 46: 757–760
Статья Google Scholar
R Core Team (2013) R: язык и среда для статистических вычислений, версия 3.0.2
Ripperger SP, Tschapka M, Kalko EKV, Rodríguez-Herrera B, Mayer F (2014) Сопротивляющаяся среда обитания фрагментация: высокая генетическая связь между популяциями плодоядных летучих мышей Carollia castanea в сельскохозяйственных ландшафтах.Сельское хозяйство Ecosyst Environ 185: 9–15. DOI: 10.1016 / j. agee.2013.12.006
agee.2013.12.006
Артикул Google Scholar
Робертсон О.Дж., Марон М., Бакли И., Макальпайн С. (2013a) Распространенность конкурентов и ландшафтная структура как предикторы лесозависимых птиц. Пейзаж Ecol 28: 1975–1987. DOI: 10.1007 / s10980-013-9934-5
Артикул Google Scholar
Робертсон О.Дж., Макальпайн С., Дом А, Марон М. (2013b) Влияние межвидовой конкуренции и ландшафтной структуры на пространственную гомогенизацию птичьих сообществ.PLoS One 8: e65299. DOI: 10.1371 / journal.pone.0065299
CAS Статья PubMed PubMed Central Google Scholar
Schradin C, Pillay N (2005) Демография полосатой мыши ( Rhabdomys pumilio ) в сочных карру. Мамм Биол 70: 84–92
Google Scholar
Сидорович В.Е., Сидорович А.А., Краско Д.А. (2010) Влияние рубок на рационы лисицы ( Vulpes vulpes ) и куницы ( Martes martes ) в переходных смешанных лесах Беларуси.Мамм Биол 75: 399–411. DOI: 10.1016 / j.mambio.2009.10.003
Google Scholar
Sozio G, Mortelliti A, Boitani L (2013) Мыши в движении: гребни пшеницы как средство повышения проходимости сельскохозяйственных ландшафтов. Биол Консерв 165: 198–202. DOI: 10.1016 / j.biocon.2013.05.022
Артикул Google Scholar
Stanley TR, Richards JD (2004) CloseTest: программа для тестирования данных захвата-повторного захвата для закрытия, Ver.3
Sunde P, Forsom HM, Al-sabi MNS, Overskaug K (2012) Избирательное истребление серых сов ( Strix aluco ) на желтошейных мышей ( Apodemus flavicollis ) и рыжих полевок ( Myodes glareolus ). Энн Зул Фенн 49: 321–330
Энн Зул Фенн 49: 321–330
Статья Google Scholar
Tattersall FH, Macdonald DW, Hart BJ, Manley WJ, Feber RE (2001) Использование лесных мышей ( Apodemus sylvaticus, ) в условиях изменчивого пахотного ландшафта.J Zool 255: 487–494
Статья Google Scholar
Тилман Д. (1994) Конкуренция и биоразнообразие в пространственно структурированных средах обитания. Экология 75: 2–16
Статья. Google Scholar
White GC, Burnham KP (1999) Программа MARK: оценка выживаемости в популяциях помеченных животных. Bird Study 46: S120 – S139
Статья Google Scholar
Wiens JA (1989) Пространственное масштабирование в экологии.Funct Ecol 3: 385–397
Артикул Google Scholar
Wójcik JM, Wolk K (1985) Суточный ритм активности двух конкурирующих грызунов: Clethrionomys glareolus и Apodemus flavicollis . Acta Theriol 30: 241–258
Артикул Google Scholar
Wood MD, Slade NA (1990) Сравнение мечения ушей и стрижки пальцев у степных полевок, Microtus ochrogaster .J Mammal 71: 252–255
Статья Google Scholar
Youngentob KN, Yoon H-J, Coggan N, Lindenmayer DB (2012) Краевые эффекты влияют на динамику конкуренции: тематическое исследование четырех симпатрических древесных сумчатых. Биол Консерв 155: 68–76. DOI: 10.1016 / j.biocon.2012.05.015
Артикул Google Scholar
Zhang Z, Usher MB (1991) Распространение лесных мышей и рыжих полевок в сельскохозяйственных ландшафтах.Acta Theriol 36: 239–245
Артикул Google Scholar
Zuur AF, Ieno EN, Smith GM (2007) Анализ экологических данных. Спрингер, Нью-Йорк
Спрингер, Нью-Йорк
Google Scholar
(PDF) Внутривидовая конкуренция способствует увеличению разнообразия использования ресурсов в естественной популяции
Экология людей: частота и последствия
индивидуальной специализации.Являюсь. Nat. 161, 1–28. (DOI: 10.
1086/343878)
Bu
¨
rger, R. & Gimelfarb, A. 2004 Влияние внутривидовой конкуренции
и стабилизации отбора на полигенный признак.
Генетика 167, 1425–1443. (DOI: 10.1534 / genetics.103.
018986)
Caldarone, EM, Wanger, M., Ogner-Burns, JS &
Buckley, LJ 2001 Протокол и руководство по оценке
нуклеиновых кислот у личинок рыб с использованием Флуоресцентный микропланшет
ридер.Справочный документ 01–11, Северо-восточное рыболовство
Научный центр.
Кристиансен, Ф. Б. 1988 Частотная зависимость и конкуренция
. Фил. Пер. R. Soc. В 319, 587–600.
Dahlhoff, E. P. 2004 Биохимические индикаторы стресса и метаболизма
: приложения для морских экологических исследований.
Анну. Rev. Physiol. 66, 183–207. (DOI: 10.1146 / annurev.
Physiol.66.032102.114509)
Даян, Т. и Симберлофф, Д. 2005 Экологические и общественные —
Широкое смещение характера: следующее поколение.Ecol. Lett.
8,875–894. (Doi: 10.1111 / j.1461-0248.2005.00791.x)
Дикманн, У. и Доебели, М. 1999 О происхождении видов
по симпатрическому видообразованию. Природа 400, 354–357. (DOI: 10.
1038/22521)
Дикманн, У., Дёбели, М., Мец, Дж. А. Дж. и Тауц, Д. (ред.)
2004 Адаптивное видообразование. Нью-Йорк, Нью-Йорк: Кембридж
University Press.
Eklo¨v, P. & Svanback, R. 2006 Хищничество благоприятствует фенотипической расходимости
дивергенции симпатрических популяций окуня.Являюсь. Nat. 167,
440–452.
Грант П. Р. 1972 Сходящийся и расходящийся характер
смещения. Биол. J. Linn. Soc. 4, 39–69.
Биол. J. Linn. Soc. 4, 39–69.
Гуревич, Дж., Морроу, Л. Л., Уоллес, А. и Уолш, Дж. С. 1992
Мета-анализ конкуренции в полевых экспериментах. Являюсь.
Нат. 140, 539–572. (DOI: 10.1086 / 285428)
Levene, H. 1953 Генетическое равновесие, когда доступно более одной
экологической ниши. Являюсь. Nat. 87, 331–333.
(DOI: 10.1086/281792)
Левинс Р. 1968 Эволюция в меняющихся условиях: около
теоретических исследований. Принстон, Нью-Джерси: Princeton University
Press.
Людвиг 1950 Zur theorie der konkurrenz. Neue Ergeb. Вероятн.
Zool., Klatt-Festschrift, 516–537.
Маклин, Р. С. 2005 Адаптивная радиация в микробных
микрокосмах. J. Evol. Биол. 18, 1376–1386. (DOI: 10.1111 /
j.1420-9101.2005.00931.x)
Maret, T. J. & Collins, J.P. 1997 Экологическое происхождение морфологического разнообразия
: исследование альтернативных трофических фенотипов
у личинок саламандр. Evolution 51, 898–905.
(doi: 10.2307 / 2411164)
Pfennig, D. W. 1992 Полифенизм головастиков лопатоногих жаб
как локально адаптированная эволюционно стабильная стратегия. Эволюция
46, 1408–1420. (DOI: 10.2307 / 2409946)
Робинсон, Б. В. и Уилсон, Д. С. 1998 Оптимальный поиск пищи, специализация
и решение парадокса Лиема.Являюсь. Nat.
151, 223–235. (DOI: 10.1086 / 286113)
Рольф, Ф. Дж. 2005 TpwRelw. См. Http://life.bio.sunysb.edu/
morph /.
Розенцвейг, М. Л. 1978 Конкурентное видообразование. Биол.
J. Linn. Soc. 10, 275–289.
Roughgarden, J. 1972 Эволюция ширины ниши. Являюсь. Nat.
106, 683–718. (DOI: 10.1086 / 282807)
Рюффер, К., Ван Дурен, Т. Дж. М., Леймар, О. и Абрамс,
П. А. 2006 Подрывной отбор и что дальше? Trends Ecol.
Evol. 21, 238–245. (DOI: 10.1016 / j.tree.2006.03.003)
Шенер Т. В. 1971 Теория стратегий кормления. Анну.
Ред. Ecol. Syst. 2, 369–404. (DOI: 10.1146 / annurev.es.02.
Ecol. Syst. 2, 369–404. (DOI: 10.1146 / annurev.es.02.
110171.002101)
Seger, J. 1985 Внутривидовая конкуренция за ресурсы как причина симпатрического видообразования
. In Evolution: эссе в честь Джона
Мейнарда Смита (ред. П. Дж. Гринвуд, П. Х. Харви и
М. Слаткин), стр. 43–53. Кембридж, Великобритания: Cambridge
University Press.
Слаткин М. 1980 Смещение экологического характера. Экология
61, 163–177. (DOI: 10.2307 / 1937166)
Смит, Т. Б. и Скуласон, С. 1996 Эволюционное значение
полиморфизмов ресурсов у рыб, амфибий и птиц.
Анну. Rev. Ecol. Syst. 27, 111–133. (DOI: 10.1146 / annurev.
ecolsys.27.1.111)
Стивенс, Д. У. и Кребс, Дж. Р. 1986 Теория собирательства.
Princeton, NJ: Princeton University Press.
Сванбак, Р.& Bolnick, D. I. 2005 Внутривидовая конкуренция
влияет на силу индивидуальной специализации: оптимальная модель теории диеты
. Evol. Ecol. Res. 7, 993–1012.
Svanback, R. & Persson, L. 2004 Индивидуальная специализация,
Ширина ниши и динамика популяции: последствия для
трофических полиморфизмов. J. Anim. Ecol. 73, 973–982.
(DOI: 10.1111 / j.0021-8790.2004.00868.x)
Суонсон, Б.О., Гибб, А.С., Маркс, Дж. К.& Hendrickson,
D. A. 2003 Трофический полиморфизм и поведенческие различия
снижают внутривидовую конкуренцию у цихлид,
Herichthys minckleyi. Экология 84, 1441–1446.
Тапер, М. Л. и Кейс, Т. Дж. 1985 Количественные генетические модели
коэволюции смещения признаков. Экология 66,
355–371. (DOI: 10.2307 / 1940385)
Ван Вален Л. 1965 Морфологические вариации и ширина
экологической ниши.Являюсь. Nat. 99, 377–389. (DOI: 10.1086 /
282379)
Вернер, Э. Э., Миттельбах, Г. Г. и Холл, Д. Дж. 1981 Роль
прибыльности кормодобывания и опыта использования среды обитания синежаберцевыми рыбками
.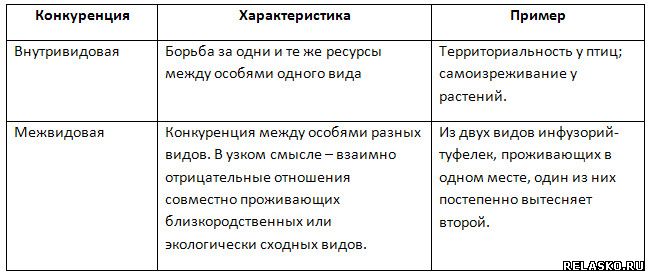 Экология 62, 116–125. (DOI: 10.2307 /
Экология 62, 116–125. (DOI: 10.2307 /
1936675)
Вест-Эберхард, М. Дж. 2003 Пластичность развития и эволюция
. Нью-Йорк, Нью-Йорк: Издательство Оксфордского университета.
Wilson, D. S. & Turelli, M. 1986 Стабильное недостаточное доминирование и
эволюционное вторжение в пустые ниши.Являюсь. Nat. 127,
835–850. (DOI: 10.1086 / 284528)
Вутон, Р. Дж., Адамс, К. Э. и Аттрилл, М. Дж. 2005 Эмпирическое моделирование
популяционной динамики небольшой
популяции трехиглой колюшки Gasterosteus
aculeatus. Environ. Биол. Рыбы 74, 151–161. (DOI: 10.
1007 / s10641-005-7690-3)
844 Р. Сванбак и Д. И. Болник Диверсификация в связи с конкуренцией
Proc. R. Soc. B (2007)
Границы | Как летучие мыши избегают принципа конкурентного исключения — сезонный переход от внутривидовых соревнований к межвидовым стимулирует использование пространства в ансамбле летучих мышей
Введение
Все хищники сталкиваются с одной и той же проблемой поиска и поимки добычи.У крупных плотоядных животных скорость поимки обычно ограничивается высоким процентом неудач (например, Eaton, 1970; Holekamp et al., 1997) во время попыток поимки, требующих большого количества энергии (например, Heglund et al., 1974; Gorman et al., 1998). ). Напротив, хищники, питающиеся относительно мелкими объектами добычи, такими как беспозвоночные, насекомые или криль и зоопланктон, в большей степени зависят от обнаружения скоплений добычи и численности или энергетической ценности отдельных объектов добычи (Morse, 1971; Lubin et al., 1977; Nowacek et al., 2011). В частности, насекомоядные, которые охотятся с воздуха, такие как летучие мыши и птицы, часто охотятся на пестрые скопления насекомых, которых они могут обнаружить только на небольших расстояниях. Однако отдельные лица могут повысить эффективность поиска, используя общедоступную информацию, непреднамеренно предоставленную аналогами или гетероспецификами с аналогичными пищевыми потребностями (Danchin et al. , 2004). В то время как групповые птицы, добывающие пищу, могут повысить свой охотничий успех путем визуального наблюдения за другими птицами (Greene, 1987), летучие мыши-ястребы могут делать это, подслушивая эхолокационные крики других летучих мышей-добытчиков (Balcombe and Fenton, 1988; Gillam, 2007; Dechmann) и другие., 2009; Дорадо-Корреа и др., 2013). Поскольку летучие мыши используют специализированные звуки, так называемые «пищевые жужжания» (Kalko, 1995), чтобы поймать свою добычу, то сородичи и гетероспецифики могут использовать такую акустическую информацию для обнаружения многообещающих участков добычи. Действительно, есть свидетельства того, что летучие мыши некоторых видов остаются на оптимальном расстоянии для перехвата друг друга, когда они охотятся большими группами, таким образом формируя сенсорную сеть, которая позволяет им сканировать территорию, намного превышающую их индивидуальный диапазон обнаружения насекомых-жертв ( Cvikel et al., 2015). В то время как две летучие мыши летают в пределах слышимости друг друга во время поиска добычи, они оба могут извлечь выгоду из увеличения эффективной дальности обнаружения добычи, но они все равно будут соревноваться, когда оба достигнут соответствующего участка добычи. Более того, большинство насекомоядных видов летучих мышей охотятся в основном в первые несколько часов после захода солнца (Kunz, 1973), вероятно, потому, что впоследствии активность летающих насекомых обычно значительно снижается (Taylor, O’Neill, 1988; Meyer et al., 2004; Milne et al. др., 2005).Этот короткий период доступности добычи ограничивает временное разделение ресурсов между конкурирующими видами и, таким образом, увеличивает межвидовую конкуренцию за таксоны, которые охотятся на одну и ту же добычу. Поскольку конкурирующие виды летучих мышей часто пересекаются и в других аспектах своей биологии, например, в предпочтениях насеста и среды обитания, конкуренция может обостриться еще больше. Принцип конкурентного исключения предполагает, что виды со слишком высоким перекрытием ниш не могут сосуществовать (Gause, 1934; Hardin, 1960; см.
, 2004). В то время как групповые птицы, добывающие пищу, могут повысить свой охотничий успех путем визуального наблюдения за другими птицами (Greene, 1987), летучие мыши-ястребы могут делать это, подслушивая эхолокационные крики других летучих мышей-добытчиков (Balcombe and Fenton, 1988; Gillam, 2007; Dechmann) и другие., 2009; Дорадо-Корреа и др., 2013). Поскольку летучие мыши используют специализированные звуки, так называемые «пищевые жужжания» (Kalko, 1995), чтобы поймать свою добычу, то сородичи и гетероспецифики могут использовать такую акустическую информацию для обнаружения многообещающих участков добычи. Действительно, есть свидетельства того, что летучие мыши некоторых видов остаются на оптимальном расстоянии для перехвата друг друга, когда они охотятся большими группами, таким образом формируя сенсорную сеть, которая позволяет им сканировать территорию, намного превышающую их индивидуальный диапазон обнаружения насекомых-жертв ( Cvikel et al., 2015). В то время как две летучие мыши летают в пределах слышимости друг друга во время поиска добычи, они оба могут извлечь выгоду из увеличения эффективной дальности обнаружения добычи, но они все равно будут соревноваться, когда оба достигнут соответствующего участка добычи. Более того, большинство насекомоядных видов летучих мышей охотятся в основном в первые несколько часов после захода солнца (Kunz, 1973), вероятно, потому, что впоследствии активность летающих насекомых обычно значительно снижается (Taylor, O’Neill, 1988; Meyer et al., 2004; Milne et al. др., 2005).Этот короткий период доступности добычи ограничивает временное разделение ресурсов между конкурирующими видами и, таким образом, увеличивает межвидовую конкуренцию за таксоны, которые охотятся на одну и ту же добычу. Поскольку конкурирующие виды летучих мышей часто пересекаются и в других аспектах своей биологии, например, в предпочтениях насеста и среды обитания, конкуренция может обостриться еще больше. Принцип конкурентного исключения предполагает, что виды со слишком высоким перекрытием ниш не могут сосуществовать (Gause, 1934; Hardin, 1960; см. Также, например,, Levine and HilleRisLambers, 2009). Однако недавние разработки в теории сосуществования предполагают, что механизмы выравнивания или стабилизации могут способствовать сосуществованию экологически подобных таксонов, наряду с механизмами, обусловленными исключительно различиями экологических ниш (Chesson, 2000). В этих рамках стабилизирующие механизмы являются условием сосуществования; учитывая, что внутривидовая конкуренция сильнее, чем межвидовая, темпы роста популяции увеличиваются при низкой численности этого вида.С другой стороны, уравнивающие механизмы поддерживают сосуществование, уменьшая недостатки приспособляемости низших из конкурирующих видов. Движущееся поведение может выступать в качестве такого механизма, например, когда конкурирующие виды изменяют свои движения и, таким образом, используют свое пространство таким образом, что они избегают скоплений сильных конкурентов (Jeltsch et al., 2013; Schlaegel in review).
Также, например,, Levine and HilleRisLambers, 2009). Однако недавние разработки в теории сосуществования предполагают, что механизмы выравнивания или стабилизации могут способствовать сосуществованию экологически подобных таксонов, наряду с механизмами, обусловленными исключительно различиями экологических ниш (Chesson, 2000). В этих рамках стабилизирующие механизмы являются условием сосуществования; учитывая, что внутривидовая конкуренция сильнее, чем межвидовая, темпы роста популяции увеличиваются при низкой численности этого вида.С другой стороны, уравнивающие механизмы поддерживают сосуществование, уменьшая недостатки приспособляемости низших из конкурирующих видов. Движущееся поведение может выступать в качестве такого механизма, например, когда конкурирующие виды изменяют свои движения и, таким образом, используют свое пространство таким образом, что они избегают скоплений сильных конкурентов (Jeltsch et al., 2013; Schlaegel in review).
Чтобы изучить наличие и степень таких механизмов, в идеале следует оценить природу и силу внутри- и межвидовой конкуренции внутри исследуемого ансамбля видов ( sensu Fauth et al., 1996). Для летучих мышей, которые охотятся на эфемерных насекомых, часто предполагается, что пищевые ресурсы в пределах скопления роящихся насекомых практически неограничены (см. Bell, 1980; Anthony et al., 1981; Arlettaz, 1999, см. Также Kalko, 1995: максимальная скорость поимки). количество роящихся насекомых летучими мышами-коньками среднего размера составляет примерно 7 насекомых / мин). Таким образом, конкуренция за эксплуатацию среди насекомоядных летучих мышей маловероятна во времена, когда много насекомых. Тем не менее, большие группы летучих мышей-охотников могут по-прежнему участвовать в соревновании по помехам, поскольку им требуется определенное пространство для полета во время поиска пищи с воздуха.Крупные летучие мыши-кормилицы в открытом пространстве, такие как Nyctalus noctula , обычно используют территорию площадью не менее 1 га во время пространственно сконцентрированных охотничьих походов на предпочтительных участках кормодобывания (Roeleke et al. , 2016; Roeleke et al. В стадии подготовки; Voigt et al. В стадии подготовки). ). Действительно, Amichai et al. (2015) недавно показали, что большие скопления летучих мышей добывают пищу менее эффективно, поскольку соответствующие особи должны чаще направлять свое внимание на сородичей и, таким образом, не могут одновременно обнаруживать объекты добычи.Это согласуется с некоторыми ранними исследованиями, показывающими, что летучие мыши, находящиеся на курсе столкновения, могут использовать специальные сигналы, описываемые как гудки, при слишком близком приближении к сородичам (Suthers, 1965; Fenton and Bell, 1979). На сегодняшний день остается неясным, являются ли эти призывы просто предупреждением об избежании столкновения или их также можно интерпретировать как агрессивные вокализации (Voigt-Heucke et al., 2010). Более того, звуки, издаваемые одновременно несколькими летучими мышами, также могут мешать обнаружению специализированных охотничьих сигналов друг друга.Действительно, Коркоран и Коннер (2014) показали, что Tadarida brasiliensis , вид, который формирует поселения миллионов особей, использует специализированные агрессивные вокализации во время борьбы за добычу. Посредством трансляции ультразвука, который заглушает звуковое обнаружение их конкурентов, они не позволяют им обнаружить добычу, которая была распознана ранее. В рамках такой структуры интерференционной конкуренции мы могли бы предположить, что внутривидовая конкуренция в ограниченном пространстве полета выше, чем межвидовая конкуренция, учитывая, что гетероспецифичные, но не обязательно сородичи, летучие мыши все еще могут демонстрировать мелкомасштабную пространственную сегрегацию (Salsamendi et al., 2012, собственные наблюдения в месте исследования), из-за соответствующей морфологии крыльев и получаемых в результате режимов полета и кормления (Norberg and Rayner, 1987; Arlettaz, 1999; Schnitzler, Kalko, 2001; Voigt et al., 2010; Voigt and Holderied, 2012).
, 2016; Roeleke et al. В стадии подготовки; Voigt et al. В стадии подготовки). ). Действительно, Amichai et al. (2015) недавно показали, что большие скопления летучих мышей добывают пищу менее эффективно, поскольку соответствующие особи должны чаще направлять свое внимание на сородичей и, таким образом, не могут одновременно обнаруживать объекты добычи.Это согласуется с некоторыми ранними исследованиями, показывающими, что летучие мыши, находящиеся на курсе столкновения, могут использовать специальные сигналы, описываемые как гудки, при слишком близком приближении к сородичам (Suthers, 1965; Fenton and Bell, 1979). На сегодняшний день остается неясным, являются ли эти призывы просто предупреждением об избежании столкновения или их также можно интерпретировать как агрессивные вокализации (Voigt-Heucke et al., 2010). Более того, звуки, издаваемые одновременно несколькими летучими мышами, также могут мешать обнаружению специализированных охотничьих сигналов друг друга.Действительно, Коркоран и Коннер (2014) показали, что Tadarida brasiliensis , вид, который формирует поселения миллионов особей, использует специализированные агрессивные вокализации во время борьбы за добычу. Посредством трансляции ультразвука, который заглушает звуковое обнаружение их конкурентов, они не позволяют им обнаружить добычу, которая была распознана ранее. В рамках такой структуры интерференционной конкуренции мы могли бы предположить, что внутривидовая конкуренция в ограниченном пространстве полета выше, чем межвидовая конкуренция, учитывая, что гетероспецифичные, но не обязательно сородичи, летучие мыши все еще могут демонстрировать мелкомасштабную пространственную сегрегацию (Salsamendi et al., 2012, собственные наблюдения в месте исследования), из-за соответствующей морфологии крыльев и получаемых в результате режимов полета и кормления (Norberg and Rayner, 1987; Arlettaz, 1999; Schnitzler, Kalko, 2001; Voigt et al., 2010; Voigt and Holderied, 2012).
Однако сезонные, а также, возможно, антропогенные изменения в наличии насекомых могут нарушить наше предположение о постоянных неограниченных пищевых ресурсах для летучих мышей-ястребов. Недавние исследования показывают, что в Центральной Европе численность насекомых снижается к концу лета (Anthony et al., 1981; Hallmann et al., 2017; Heim et al., 2017), что совпадает с тем временем, когда несколько видов летучих мышей сталкиваются с выбором между тем, чтобы тратить свое время на кормление, спаривание и либо найти зимний насест, либо мигрировать на юг. Учитывая, что время сбора пищи, а также доступность добычи могут быть ограничены в конце лета, конкуренция может измениться в сторону использования ресурсов, что даст преимущество более мелким и более маневренным летчикам (Norberg and Rayner, 1987), которые могут оказаться более успешными. в ловле ограниченного количества предметов добычи за короткое время.Тем не менее, остается вопрос, могут ли более крупные и более быстрые виды смягчить это усиление межвидовой конкуренции, используя более отдаленные, но, возможно, менее богатые и все же незанятые участки добычи, чтобы уравновесить этот потенциальный недостаток.
Недавние исследования показывают, что в Центральной Европе численность насекомых снижается к концу лета (Anthony et al., 1981; Hallmann et al., 2017; Heim et al., 2017), что совпадает с тем временем, когда несколько видов летучих мышей сталкиваются с выбором между тем, чтобы тратить свое время на кормление, спаривание и либо найти зимний насест, либо мигрировать на юг. Учитывая, что время сбора пищи, а также доступность добычи могут быть ограничены в конце лета, конкуренция может измениться в сторону использования ресурсов, что даст преимущество более мелким и более маневренным летчикам (Norberg and Rayner, 1987), которые могут оказаться более успешными. в ловле ограниченного количества предметов добычи за короткое время.Тем не менее, остается вопрос, могут ли более крупные и более быстрые виды смягчить это усиление межвидовой конкуренции, используя более отдаленные, но, возможно, менее богатые и все же незанятые участки добычи, чтобы уравновесить этот потенциальный недостаток.
Здесь мы использовали эксперименты по воспроизведению, чтобы изучить природу конкуренции между двумя одновременно встречающимися и потенциально конкурирующими видами летучих мышей, записав их реакцию на моделируемые скопления кон- и гетероспецифических особей на разных этапах жизненного цикла (т.е., ранний сезон, в течение которого происходит размножение и линька, и поздний сезон, в течение которого происходит спаривание, поиск зимних насестов и, возможно, миграция). Наш подход к воспроизведению использует вокализацию и слуховую способность летучих мышей-насекомоядных в воздухе, что позволяет измерять пространственные изменения активности в ответ на экспериментальные акустические обработки путем количественной оценки активности летучих мышей с помощью количества ультразвуковых сигналов, которые могут быть записаны в экспериментальной зоне. Нашим основным видом была обыкновенная ночница N.noctula (Schreber, 1774) — быстро летающая и частично перелетная летучая мышь. На нашем исследовательском участке в Германии мы подвергли локально добывающих корм N. noctula воспроизводству либо охотничьих сородичей, либо охотничьих Pipistrellus nathusii . У этих двух видов схожие модели активности (Heim et al., 2016) и высокая степень совпадения ниш с точки зрения диеты, использования среды обитания и предпочтения насестов (Eichstädt, 1997; Vaughan, 1997). Основываясь на приведенных выше предположениях о природе внутривидовой и межвидовой конкуренции в ансамблях летучих мышей-ястребов, мы предположили, что реакция N.noctula по отношению к разным типам воспроизведения зависит от общей плотности конкурентов в данном районе и от сезона, и эта реакция будет связана с различной доступностью добычи в разные сезоны. В частности, мы предсказали, что N. noctula увеличит активность кормления во время воспроизведений сородича в начале сезона, когда добыча многочисленна, и что N. noctula покинет охотничьи угодья во время воспроизведения кон- или гетероспецифических видов в конце сезона. когда добычи мало.
noctula воспроизводству либо охотничьих сородичей, либо охотничьих Pipistrellus nathusii . У этих двух видов схожие модели активности (Heim et al., 2016) и высокая степень совпадения ниш с точки зрения диеты, использования среды обитания и предпочтения насестов (Eichstädt, 1997; Vaughan, 1997). Основываясь на приведенных выше предположениях о природе внутривидовой и межвидовой конкуренции в ансамблях летучих мышей-ястребов, мы предположили, что реакция N.noctula по отношению к разным типам воспроизведения зависит от общей плотности конкурентов в данном районе и от сезона, и эта реакция будет связана с различной доступностью добычи в разные сезоны. В частности, мы предсказали, что N. noctula увеличит активность кормления во время воспроизведений сородича в начале сезона, когда добыча многочисленна, и что N. noctula покинет охотничьи угодья во время воспроизведения кон- или гетероспецифических видов в конце сезона. когда добычи мало.
Материалы и методы
Учебные площадки
В 2016 г. мы провели эксперименты по воспроизведению, направленные на N. noctula во время немигрирующего сезона размножения (с середины июня до середины августа, далее именуемого ранним сезоном) и во время его брачного и потенциально миграционного сезона (начало сентября до начала октября, далее — поздний сезон). Эксперименты по воспроизведению проводились в северо-восточной Германии, то есть в северных частях федеральной земли Бранденбург, называемой Укермарк.В Укермарке преобладают сельскохозяйственные поля, но есть много водоемов, от небольших котлованов до относительно больших озер. Мы стремились провести эксперименты по воспроизведению на берегах 23 из этих лимнических местообитаний. Хотя лесных остатков мало, и поэтому ожидается, что возможности для ночлега в этом районе будут ограниченными, из предыдущих исследований GPS (Roeleke et al., 2016; Roeleke et al. Неопубликованные данные) мы знали, что колоний N. noctula в Уккермарк предпочитает кормиться над водоемами на расстоянии не менее 7 км от их насеста. 23 места воспроизведения были распределены на площади около 60 000 га. В течение каждой экспериментальной ночи мы проводили эксперименты по воспроизведению на двух участках одновременно. Расстояние между парными точками воспроизведения составляло от 1 до 5 км. Поскольку мы проводили эксперименты примерно в одно и то же время, маловероятно, что мы транслировали воспроизведение воспроизведения одним и тем же людям на разных сайтах в течение данной ночи. Мы также стремились провести воспроизведение дважды на каждом сайте — один раз в начале и один раз в конце сезона (см. Раздел «Эксперименты с воспроизведением»).Однако из-за пространственных и временных вариаций активности летучих мышей мы не всегда могли добиться этого для всех участков. Чтобы избежать псевдорепликации, мы посещали каждый сайт только один раз за сезон.
23 места воспроизведения были распределены на площади около 60 000 га. В течение каждой экспериментальной ночи мы проводили эксперименты по воспроизведению на двух участках одновременно. Расстояние между парными точками воспроизведения составляло от 1 до 5 км. Поскольку мы проводили эксперименты примерно в одно и то же время, маловероятно, что мы транслировали воспроизведение воспроизведения одним и тем же людям на разных сайтах в течение данной ночи. Мы также стремились провести воспроизведение дважды на каждом сайте — один раз в начале и один раз в конце сезона (см. Раздел «Эксперименты с воспроизведением»).Однако из-за пространственных и временных вариаций активности летучих мышей мы не всегда могли добиться этого для всех участков. Чтобы избежать псевдорепликации, мы посещали каждый сайт только один раз за сезон.
Подготовка к воспроизведению
На каждом участке мы транслировали три разных типа воспроизведения на корм N. noctula ; кормление жужжанием N. noctula , кормление жужжанием P. nathusii и синусоидальный тон между 20 и 40 кГц в качестве контроля (дополнительный материал 1). Жужжание кормления — это специальные крики летучих мышей, которые предназначены для заключительной фазы поимки добычи и однозначно идентифицируются.Испытания одиночного воспроизведения длились 3 минуты и состояли из трех фаз; (i) 1 минута тишины (базовый уровень) для записи акустической базовой активности N. noctula на соответствующем участке, (ii) 1 минута трансляции соответствующего воспроизведения (воспроизведения) и (iii) 1 минута снова молчания (публикация), чтобы увидеть потенциальные эффекты после воспроизведения (см. Уберникель и др., 2013 г .; Войт-Хеук и др., 2016 г.).
Эксперименты с воспроизведением
Мы начали попытки воспроизведения, как только наблюдали активность кормления одного или нескольких N.noctula через настройку записи. После каждого испытания мы ждали не менее 3 минут и снова проверяли активность акустического кормодобывания, прежде чем транслировать следующее воспроизведение. В большинстве ночей мы проводили эксперименты вскоре после захода солнца, когда прибыли первые N. noctula . Однако в случае, если все летучие мыши покинули область во время воспроизведения, мы попытались провести второй раунд экспериментов по воспроизведению позже, когда активность N. noctula в области была более стабильной. Около полуночи северной широты.noctula всегда снижалась до низкого уровня. Если нам не удавалось провести наш эксперимент до полуночи, мы останавливали эксперименты, чтобы убедиться, что все летучие мыши в нашем исследовании столкнулись с воспроизведением в аналогичной ситуации, то есть во время их первого ночного боя за едой. Во время некоторых экспериментальных испытаний мы также заметили P. nathusii , которые кормились рядом с береговой линией и, следовательно, недалеко от нашей экспериментальной установки. Однако, поскольку естественная активность P. nathusii была низкой по сравнению с транслируемыми стимулами, мы уверены, что их потенциальный эффект был незначительным.Мы не оценивали потенциальные эффекты воспроизведений на P. nathusii , поскольку воспроизведение не было направлено на них, и поэтому мы не могли гарантировать постоянную базовую активность P. nathusii перед трансляцией.
В большинстве ночей мы проводили эксперименты вскоре после захода солнца, когда прибыли первые N. noctula . Однако в случае, если все летучие мыши покинули область во время воспроизведения, мы попытались провести второй раунд экспериментов по воспроизведению позже, когда активность N. noctula в области была более стабильной. Около полуночи северной широты.noctula всегда снижалась до низкого уровня. Если нам не удавалось провести наш эксперимент до полуночи, мы останавливали эксперименты, чтобы убедиться, что все летучие мыши в нашем исследовании столкнулись с воспроизведением в аналогичной ситуации, то есть во время их первого ночного боя за едой. Во время некоторых экспериментальных испытаний мы также заметили P. nathusii , которые кормились рядом с береговой линией и, следовательно, недалеко от нашей экспериментальной установки. Однако, поскольку естественная активность P. nathusii была низкой по сравнению с транслируемыми стимулами, мы уверены, что их потенциальный эффект был незначительным.Мы не оценивали потенциальные эффекты воспроизведений на P. nathusii , поскольку воспроизведение не было направлено на них, и поэтому мы не могли гарантировать постоянную базовую активность P. nathusii перед трансляцией.
Мы проводили эксперименты только при скорости ветра ≤ 3 м / с в ночное время без осадков. Подробное описание подготовки к воспроизведению и экспериментальной установки см. В дополнительном материале 1.
Акустический анализ
Мы проанализировали акустические записи с помощью SasLabPro (Avisoft Bioacoustics), используя спектрограмму окна Хэмминга с быстрым преобразованием Фурье 1024 и 87.5% перекрытие по времени. Мы идентифицировали и подсчитали звонки N. noctula , которые имели отношение сигнал / шум выше 30 дБ для каждой из трех фаз соответствующих испытаний воспроизведения, тем самым учитывая разницу в ~ 30 дБ между нашими воспроизведениями и предполагаемым звуковым давлением. уровни кормятся летучими мышами.
уровни кормятся летучими мышами.
Отлов и анализ насекомых
На каждой площадке воспроизведения мы ловили летающих насекомых с помощью специальной ловушки для ультрафиолетового (УФ) света (источник света с длиной волны около 365 нм).Когда насекомые приближались к свету, они сталкивались с гладкой пластиковой поверхностью перед лампой и затем скользили в бутылку, наполненную 95% этанолом. Ловушки устанавливались на берегу соответствующих водоемов, на расстоянии ~ 5 м от точки воспроизведения, на высоте 3 м. Как только мы заметили первый N. noctula с нашей записывающей установкой, мы включили УФ-свет ловушки и прикрепили бутыль с этанолом на 1 час. Таким образом, мы убедились, что насекомые не привлекаются ультрафиолетовым светом до начала эксперимента по воспроизведению.
Чтобы получить наиболее важные измерения добычи с точки зрения летучих мышей, мы отсортировали и подсчитали насекомых по пара-таксономическим группам, отражающим комбинацию порядка и размера (Таблица 1). Отсортированные образцы сушили 72 ч при 50 ° C и измеряли сухую массу на электронных весах (ME5, Sartorius, Германия, разрешение 0,001 мг).
Таблица 1 . Присутствие и непрерывная кормодобывающая деятельность N. noctula , а также общее количество и масса насекомых на участках отбора проб.
Управление данными и статистический анализ
Наш акустический анализ показал, что иногда активность летучих мышей прекращалась на более длительное время во время любой из трех фаз нашего эксперимента (т.е. базовый уровень, воспроизведение, публикация). Мы исключили эти испытания из дальнейшего анализа, так как не могли быть уверены, действительно ли в таких случаях исследуемые животные собирали пищу. Пожалуйста, см. Дополнительный материал 2 для подробного описания оценки экспериментального участка и последующей очистки данных.
Чтобы оценить относительную разницу активности N. noctula между предварительной фазой и фазой воспроизведения, мы вычислили относительную разницу между предварительной фазой и фазой воспроизведения как:
вызывает фазу воспроизведения-вызывает предварительную фазу (вызывает фазу воспроизведения + вызывает предварительную фазу) / 2 (1)Затем мы смоделировали относительную разницу с помощью линейной модели смешанного эффекта (R package lme4, Bates et al., 2015). В качестве переменных-предикторов мы использовали тройное взаимодействие типа воспроизведения (т.е., аналог, гетероспецифический, контроль), исходную активность (количество вызовов N. noctula во время предварительной фазы) и сезон. В качестве случайного эффекта мы включили экспериментальную пробу, вложенную в сайт. Чтобы проверить, будут ли эффекты воспроизведения длиться дольше, чем трансляция самого воспроизведения, мы запустили аналогичную модель с относительной разницей между пре- и пост-фазой в качестве зависимой переменной. Мы обеспечили нормальное распределение смоделированных остатков путем визуальной проверки графиков квантилей моделей.Мы вычислили значения псевдо-R-квадрата с помощью пакета R MuMIn (Barton, 2014) и величины эффекта переменных-предикторов с помощью эффектов пакета R (Fox, 2003). Мы предположили статистически значимые эффекты переменных-предикторов, когда 95% доверительный интервал не охватывал 0. После подтверждения с помощью тестов Колмогорова-Смирнова, что численность и массы различных групп насекомых не были нормально распределены, мы использовали парные Mann-Whitney- U -Испытания для проверки влияния сезона на массу или количество пойманных насекомых или на относительное количество различных классов размеров в каждой выборке.Мы использовали все образцы для этого теста, в том числе образцы из комбинаций местоположения и сезона, где мы не получали данные из экспериментов по воспроизведению. Вся обработка данных и анализ выполнялись с помощью R 3.3.2 (R Core Team, 2016). Национальные правила не требовали обзора и одобрения экспериментов, поскольку животные не были пойманы, обработаны или подвергнуты физическим манипуляциям.
Результаты
Действия летучих мышей на водоемах
Мы отобрали 30 различных водоемов в районе исследования на с.ш.noctula активность. Для кормления N. noctula в начале сезона было использовано больше водоемов, чем в конце сезона (Chi 2 -Test, Chi 2 = 4,65, N = 30, p = 0,03). На участках, где присутствовало N. noctula в оба сезона, уровень активности N. noctula не отличался между сезонами (Mann-Whitney- U -Test, W = 124, N = 28, p = 0,21) (таблица 1).
Ловушка для насекомых
Большинство пойманных насекомых были довольно небольшого размера.Количество и сухая масса пойманных насекомых в значительной степени варьировались между выборочными участками (Таблица 1). Хотя, казалось, произошел небольшой сдвиг от крупных (длина тела> 9 мм) к мелким насекомым (длина тела <6 мм) в период с начала сезона до конца сезона, мы не обнаружили каких-либо существенных различий в количестве и массе различных насекомых. размерные классы (парные тесты Манна-Уитни U , N = 46, рисунок 1). Тем не менее, парные тесты Манна-Уитни- U ( N = 46) показали, что относительное количество крупных насекомых было значительно выше в начале сезона (V = 132, p = 0.045), в то время как относительная численность мелких насекомых была выше в конце сезона (V = 57, p = 0,025). Пожалуйста, см. Дополнительный материал 3, где представлены данные о численности и массах различных классов насекомых для конкретных участков.
Рисунок 1 . Коробчатые диаграммы и статистические данные о количестве насекомых и сухой массе для разных сезонов, отсортированные по разным классам насекомых. Усы изображают максимум в 1,5 раза превышающий межквартильный диапазон. По графическим причинам выбросы не отображаются.Пожалуйста, см. Таблицу 1 для получения информации об общем количестве на выбранных сайтах.
Эксперименты с воспроизведением
N. noctula не изменила своей активности при воздействии нашей контрольной обработки, синусоидального тона между 20 и 40 кГц. В целом, у N. noctula наблюдались меньшие сдвиги в активности, когда их начальная плотность (то есть количество звонков, записанных во время предварительной фазы) была сравнительно высокой. Однако, если включить активность во время предварительной фазы не как фактор взаимодействия, а только как основной эффект в модели, оказалось, что Н.noctula обычно отрицательно реагировала на воспроизведение, когда начальная плотность схожих объектов была высокой (то есть около 500 вызовов в минуту во время предварительной фазы; график эффекта см. в Дополнительном материале 3). Наша полная модель с тройным взаимодействием между начальной плотностью, сезоном и типом воспроизведения имела псевдо R 2-значение 0,42. Эта модель показала, что N. noctula не реагирует на охотничьи крики гетероспецифического P. nathusii . Кроме того, был лишь небольшой положительный ответ на охотничьи крики сородичей при довольно низкой начальной плотности (рис. 2А).Однако это превратилось в явное избегание сородичей в конце сезона, когда исходная плотность была низкой или средней. В то же время, по крайней мере, при низких начальных плотностях, активность N. noctula увеличивалась, когда мы транслировали гетероспецифические воспроизведения в конце сезона. Только при высокой начальной плотности N. noctula начали избегать экспериментального района во время воспроизведений P. nathusii (рис. 2B).
Рисунок 2 . Относительная разница Н.активность noctula между пре-фазой и фазой воспроизведения (A, B) или пре-фазой и пост-фазой (C, D) эксперимента, в зависимости от активности N. noctula во время пре-фазы, различные типы воспроизведения и время года. Исходные данные показаны кружками и треугольниками. Линии показывают предполагаемый эффект, заштрихованные области показывают соответствующие 95% доверительные интервалы. Сплошная красная линия и красные круги: начало сезона. Пунктирная синяя линия и синие треугольники: конец сезона.Для простоты незначительные эффекты по отношению к контрольной обработке (синусоидальный тон) не показаны.
Независимо от типа воспроизведения и сезона, количество вызовов во время предварительной фазы и пост-фазы воспроизведения существенно не различались, т.е. относительная разница колебалась около 0 (рисунки 2C, D).
Обсуждение
Насекомоядные виды летучих мышей могут встречаться вместе, несмотря на значительное совпадение их экологических ниш (например, Bell, 1980; Fenton, 1990; Salsamendi et al., 2012). Тем не менее, механизмы, которые предотвращают конкурентное исключение ансамблей конкурирующих летучих мышей, полностью не решены. Целью этого исследования было лучше понять природу и относительную силу внутривидовой и межвидовой конкуренции, а также выявить сезонные изменения конкуренции. Поэтому мы направили воспроизведение собирающих пищу сородичей и гетероспецификов на кормление N. noctula в течение двух разных сезонов, то есть в начале и в конце лета. N. noctula лишь незначительно отреагировала на воспроизведение сородичей.С другой стороны, реакция N. noctula на воспроизведение конкурирующих гетероспецифических особей изменилась с повышения активности в начале сезона на снижение активности в конце сезона. Мы пришли к выводу, что изучаемые насекомоядные летучие мыши испытали более сильную внутривидовую конкуренцию, чем межвидовую, в начале сезона, тогда как в конце сезона наблюдалось обратное.
Сезонные смены среды обитания
В конце лета для кормления N. noctula потреблялось меньше водоемов в нашем районе исследования, чем в начале лета.На первый взгляд это кажется противоречащим интуиции, поскольку можно было бы ожидать более высокой численности летучих мышей-добытчиков в конце сезона из-за отъемного потомства. Кроме того, мигрирующие летучие мыши из северных и северо-восточных стран прибывают примерно с конца августа до конца октября в Центральную Европу, включая область наших исследований (Ahlén et al., 2009; Furmankiewicz and Kucharska, 2009; Ciechanowski et al., 2010). Приток мигрирующих N. noctula , вероятно, является причиной, по которой Heim et al.(2016) обнаружили увеличение активности N. noctula на выше сельскохозяйственных полей в исследуемой области в конце лета. Мы предполагаем, что наблюдаемое сокращение использования водоемов, отобранных нами, было вызвано не общим снижением активности в этом районе, а, скорее, смещением использования местообитаний с лимнических на наземные кормовые угодья. Это согласуется с изотопным анализом Voigt et al. (2016), которые обнаружили, что N. noctula меньше питается водными насекомыми в конце лета, чем в начале лета.Сдвиг в использовании среды обитания может частично быть результатом необходимости спаривания в конце сезона. Самцам летучих мышей приходится устраивать и защищать одиночные насесты, в то время как самки ищут эти так называемые брачные насесты. Таким образом, самцам, возможно, придется кормиться рядом с их насестом, а самки могут сэкономить время, если будут питаться по условию во время поиска спарившихся насестов, а не в специально отведенных местах кормления, таких как водоемы. Недавнее отслеживающее исследование предлагает такую стратегию, по крайней мере, для самок, показав, что самка N.noctula покрывают большие площади и меньше ориентируются на отдельные водоемы для кормодобывания в конце лета (Roeleke et al., 2016).
Сезонные смены доступности добычи
Вопреки нашим ожиданиям и прошлым исследованиям (Black, 1974; Janzen and Pond, 2009; Hallmann et al., 2017; Heim et al., 2017, но см. Hails, 1982), мы не смогли обнаружить различий в количестве или биомассе летающие ночные насекомые между ранним и поздним сезоном. Тем не менее, в конце сезона, по-видимому, наблюдалась тенденция к уменьшению количества крупных насекомых (т.е., длина тела> 9 мм) присутствовали в отобранных водоемах (см. Gloor et al., 1995), и мы обнаружили значительное уменьшение доли крупных насекомых по сравнению с ранним сезоном. Хотя существует множество диетических исследований, которые показывают, что N. noctula является условно-патогенным питателем, большинство исследований согласны с тем, что относительно крупные насекомые являются важными компонентами его рациона (обзор в Vaughan, 1997). Уменьшение количества относительно крупных насекомых в отобранных водоемах могло, таким образом, усилить конкуренцию за объекты добычи.Это является дополнительным объяснением того, почему меньшее количество водоемов использовалось для кормления N. noctula , поскольку уменьшение возможных кормовых объектов могло вынудить N. noctula искать корм в местообитаниях с меньшим количеством конкурентов. Такой временно богатой насекомыми суррогатной средой обитания кормов могли быть сельскохозяйственные угодья. Heim et al. (2016) предполагают, что уборка урожая в сентябре может временно увеличить доступность насекомых в этом районе (см. Plucinski et al., 2015). Voigt et al. (2015) обнаружили, что Eptesicus serotinus , летучая мышь-кормушка в открытом космосе с аналогичной морфологией крыльев, что и N.noctula (Norberg and Rayner, 1987) одинаково питается как наземными, так и водными насекомыми, что предполагает гибкость в использовании среды обитания для кормления фуражиров в открытом космосе.
Однако мы должны признать, что, используя ловушки УФ-излучения, наш метод отбора проб был избирательным в отношении светочувствительных насекомых. Кроме того, мы были ограничены размещением ловушек на берегах водоемов на высоте около 4 м, тогда как N. noctula в основном кормились на высотах от 8 до 12 м над поверхностью воды.Таким образом, наша выборка насекомых, скорее всего, дает только представление об общей активности насекомых, но не обязательно отражает фактическую доступность добычи для N. noctula .
Эксперименты с воспроизведением — реакция, зависящая от пост-эффекта и плотности
Мы не обнаружили каких-либо значительных эффектов экспериментальной обработки в фазе после воспроизведения, т.е. активность летучей мыши почти мгновенно вернулась к базовому уровню активности после трансляции воспроизведений. Это показывает, что Н.noctula очень быстро создает новые конкурентные ситуации и также быстро и динамично регулирует использование своего пространства.
N. noctula реагировала на наши воспроизведения только тогда, когда базовая активность была низкой или средней (т.е. менее 500 звонков в минуту). Возможно, N. noctula воспринимает акустическую информацию от реальных нынешних сородичей более надежно, чем наше воспроизведение. Однако жужжание при кормлении, естественно, слабее, чем при поиске (Holderied et al., 2005), а высокая активность акустического поискового сигнала может препятствовать акустическому обнаружению кормящихся кон- или гетероспецифических особей как в экспериментальных, так и в естественных ситуациях.Однако также может быть, что пространство, которое можно было эффективно использовать для кормодобывания, уже было насыщено особями. Использование густо населенных участков для кормления может быть неэффективным (Amichai et al., 2015), что может привести к индивидуальному разделению кормового пространства (см. Beauchamp and Fernández-Juricic, 2005).
Эксперименты с воспроизведением — сезонные изменения конкуренции
Только в конце лета N. noctula продемонстрировала умеренно положительный отклик на воспроизводство сородичей.С другой стороны, N. noctula явно привлекались к воспроизведению кормления P. nathusii в начале лета, но в конце лета это превратилось в явное избегание. Как упоминалось выше, все эти ответы были верны для низкой или средней базовой активности N. noctula (т.е. <500 вызовов в минуту), но исчезли или даже изменились, когда присутствовали большие скопления особей. Мы предполагаем, что сезонные различия в ответах на наши воспроизведения были вызваны изменениями силы внутри- и межвидовой конкуренции.
В частности, повышенная активность во время воспроизведения гетероспецифических особей в начале лета указывает на то, что подслушивание поисков пищи гетероспецифических особей является выгодной стратегией для летучих мышей, которые охотятся на пятнисто распределенную добычу в это время года. Тем не менее, кажется удивительным, что N. noctula не продемонстрировала такой положительной реакции на воспроизводство сородичей, собирающих пищу, поскольку сородичи теоретически должны иметь наибольшее совпадение диетических требований и, таким образом, должны быть наиболее надежным индикатором наличия предпочтительной жертвы насекомых. .Наблюдаемое отсутствие реакции на добывающих пищу сородичей в сочетании с положительной реакцией на добывающих пищу гетероспецифических животных предполагает сильную внутривидовую конкуренцию и, в то же время, незначительную межвидовую конкуренцию в начале лета. Как следствие, мы предполагаем, что летучие мыши соревнуются не за добычу, а за пространство для полета и «звуковое пространство» в начале лета. Под звуковым пространством мы подразумеваем многомерную сущность, которая определяется трехмерным пространственным компонентом, временем и диапазоном ультразвуковых частот, которые летучие мыши используют для эхолокации.Эхолокационным летучим мышам это пространство необходимо для трансляции своих ультразвуковых сигналов и для приема отраженных эхо их сигналов, чтобы определять местонахождение добычи и препятствий. Соседние сородичи используют одно и то же пространство полета и звуковое пространство и, таким образом, могут мешать друг другу во время полета и во время акустического обнаружения добычи соответственно. В отличие от этого, гетероспецифические особи могут разделять пространство для кормления по вертикали и меньше перекрываться в своем звуковом пространстве из-за использования разных частот эхолокации.Мелкомасштабная вертикальная сегрегация была показана для множества конкурирующих таксонов, которые чрезмерно используют трехмерное пространство для кормления (например, Saiful et al., 2001; Kiszka et al., 2011; Navarro et al., 2013; Humphries et al. ., 2016; Мансор, Рамли, 2017). Хотя исследование мелкомасштабной вертикальной сегрегации воздушных ястребов является технически сложной задачей, недавние записи высоты полета летучих мышей в открытом космосе предполагают вертикальную сегрегацию, но без четких доказательств активности кормодобывания (Roemer et al., 2017). На наших участках отбора проб мы часто наблюдали, что P. nathusii обычно кормились на высотах прибл. От 4 до 8 м, тогда как для N. noctula часто используются высоты от 8 до 15 м.
Как упоминалось выше, гетероспецифические летучие мыши также демонстрируют разделение частот их эхолокации (приблизительно пиковые частоты для N. noctula 20 кГц и для P. nathusii 40 кГц, Skiba, 2003). Поскольку слуховая система летучих мышей точно настроена на их собственный частотный диапазон (обзор Hiryu et al., 2016; Pollak, 2016), акустическая интерференция между этими двумя видами должна быть незначительной. Учитывая, что добыча не ограничена, вертикальное разделение кормового пространства и распределение частоты криков должно, таким образом, позволить ансамблю этих двух видов эффективно добывать корм с более высокой плотностью, чем это было бы возможно для агрегации любого из этих двух видов по отдельности.
В отличие от модели, наблюдавшейся в начале лета, N. noctula продемонстрировала пониженную активность при воздействии на корм гетероспецифических особей в конце лета.В то же время наблюдалась, по общему признанию, очень умеренная, положительная реакция на воспроизведение сородичей, собирающих пищу, во время низкой базовой активности (т.е. <250 вызовов в минуту). Мы пришли к выводу, что сила межвидовой конкуренции должна резко измениться с начала до конца лета. В частности, N. noctula , казалось, ожидал сильной межвидовой конкуренции, когда мы транслировали жужжание кормления P. nathusii , что вынудило их покинуть соответствующие районы кормодобывания во время воспроизведения.Учитывая, что вмешательство гетероспецифических видов, вероятно, незначительно, мы заключаем, что наблюдаемый отрицательный ответ был вызван усилением конкуренции за эксплуатацию из-за низкой доступности добычи. Поскольку в конце лета крупных насекомых было относительно мало, фуражиры в открытом космосе, такие как N. noctula , могли быть не в состоянии эффективно кормиться на участках, уже занятых P. nathusii . Вероятно, P. nathusii может поймать добычу быстрее, чем N. noctula в такой ситуации, из-за своей более высокой маневренности (Norberg and Rayner, 1987).Более того, меньшая высота полета предполагает, что P. nathusii может ловить восходящих насекомых до того, как они достигнут сфер более высоких летучих мышей, таких как N. noctula . Marggraf et al. (в обзоре) обнаружили, что P. nathusii снижала активность в ответ на воспроизведение собирающих пищу сородичей, но не реагировала на воспроизведение кормодобывающих видов N. noctula , что указывает на то, что межвидовая конкуренция не является симметричной у этих двух видов. Таким образом, особенно в периоды нехватки добычи, для N.noctula для обнаружения участков добычи, которые не используются лучшими собирателями, такими как P. nathusii . Поэтому мы предполагаем, что подслушивание охотничьих сородичей является наиболее многообещающей стратегией, когда добыча ограничена, пока плотность сородичей не слишком высока для эффективного кормодобывания.
Выводы
Мы обнаружили, что летучая мышь-ястреб N. noctula активно ищет гетероспецифический вид P. nathusii во время походов за кормом в начале лета, но избегает участков, занятых гетероспецифическими животными в конце лета. N. noctula не отреагировала на кормление сородичей в начале лета, но показала небольшую положительную реакцию на сородичей в конце лета. Мы пришли к выводу, что количество воздушных ястребов, собирающих корм в открытом космосе, на участке питания ограничено внутривидовой конкуренцией за пространство для полета и звуковое пространство в начале лета, но что межвидовая конкуренция за добычу насекомых ограничивает количество летучих мышей в конце лета. Таким образом, высокая внутривидовая конкуренция может стабилизировать ансамбли насекомоядных летучих мышей, когда пищевые ресурсы изобилуют.Во время, вероятно, более низкой доступности добычи в конце лета, воздушные ястребы, которые специализируются на быстром полете в незагроможденных местах обитания, могут страдать от более низкой скорости поимки, чем более маневренные летучие мыши, такие как P. nathusii . Водные объекты за это время использовались меньше. Мы предполагаем, что быстро летающие воздушные ястребы могут использовать более удаленные или менее богатые охотничьи угодья, поскольку их особая морфология крыльев позволяет им летать на большие расстояния с низкими энергетическими затратами (Winter and von Helversen, 1998).Эта адаптация к быстрому, но дешевому полету может уравновесить недостатки приспособленности к лучшим собирателям. Можно даже предположить, что неоптимальные с точки зрения кормодобывания среды обитания могут поддерживать разнообразие летучих мышей, предлагая убежища от межвидовой конкуренции.
Доступность данных
Необработанные данные, подтверждающие выводы этой рукописи, будут предоставлены авторами без излишних оговорок любому квалифицированному исследователю.
Взносы авторов
CV и MR концептуализировали исследование.Резюме курировали студенты. MR и LJ проводили эксперименты. LJ проанализировал акустические записи. MR провел статистический анализ. MR и CV написали рукопись.
Финансирование
MR финансировалось Deutsche Forschungsgemeinschaft (DFG-GRK 2118/1 — BioMove). Публикация этой статьи финансировалась Deutsche Forschungsgemeinschaft (DFG-GRK 2118/1 — BioMove) и Фондом открытого доступа Ассоциации Лейбница.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Рецензент JM и управляющий редактор заявили о своей общей принадлежности.
Благодарности
Благодарим Мариса Пярна за помощь в сортировке образцов насекомых. Мы благодарим Ольгу Хейм, Силке Фойгт-Хеук и Лару Маргграф за предоставленные записи звонков летучих мышей и помощь в подготовке файлов воспроизведения. Виктории Радчук за советы по статистике. Центр исследования сельскохозяйственных ландшафтов имени Лейбница и Гернота Верча за предоставление помещений для проведения полевых работ. Нильсу Блауму, Флориану Елчу и членам RTG BioMove за обсуждение рукописи.
Дополнительные материалы
Дополнительные материалы к этой статье можно найти в Интернете по адресу: https://www.frontiersin.org/articles/10.3389/fevo.2018.00101/full#supplementary-material
Список литературы
Ahlén, I., Baagøe, H. J., and Bach, L. (2009). Поведение скандинавских летучих мышей во время миграции и кормления в море. J. Mammal. 90, 1318–1323. DOI: 10.1644 / 09-MAMM-S-223R.1
CrossRef Полный текст | Google Scholar
Амичай, Э., Блюмрозен, Г., Йовель, Ю. (2015). Звонок громче и дольше: как летучие мыши используют биосонар в условиях сильных акустических помех от других летучих мышей. Proc. Биол. Sci. 282: 20152064. DOI: 10.1098 / rspb.2015.2064
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Энтони, Э. Л. П., Стек, М. Х. и Кунц, Т. Х. (1981). Ночлег и бюджет ночного времени маленькой коричневой летучей мыши, Myotis lucifugus : влияние репродуктивного статуса, плотности добычи и условий окружающей среды. Oecologia 51, 151–156. DOI: 10.1007 / BF00540593
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Арлеттаз Р. (1999). Выбор среды обитания как основной механизм разделения ресурсов между двумя симпатрическими родственными видами летучих мышей Myotis myotis и Myotis blythii . J. Anim. Ecol. 68, 460–471. DOI: 10.1046 / j.1365-2656.1999.00293.x
CrossRef Полный текст | Google Scholar
Балкомб, Дж. П. и Фентон, М.Б. (1988). Подслушивание летучими мышами: влияние дизайна эхолокационного вызова и стратегии поиска пищи. Этология 79, 158–166. DOI: 10.1111 / j.1439-0310.1988.tb00708.x
CrossRef Полный текст | Google Scholar
Бейтс, Д., Мехлер, М., Болкер, Б., и Уокер, С. (2015). Подгонка линейных моделей смешанных эффектов с использованием lme4. J. Stat. Софтв. 67, 1–48. DOI: 10.18637 / jss.v067.i01
CrossRef Полный текст
Beauchamp, G., и Fernández-Juricic, E.(2005). Парадокс размера группы: эффекты обучения и правила выхода исправлений. Behav. Ecol. 16, 352–357. DOI: 10.1093 / beheco / arh269
CrossRef Полный текст | Google Scholar
Белл, Г. П. (1980). Использование среды обитания и реакция на участки добычи пустынными насекомоядными летучими мышами. Кан. J. Zool. 58, 1876–1883. DOI: 10.1139 / z80-256
CrossRef Полный текст | Google Scholar
Блэк, Х. Л. (1974). Сообщество летучих мышей северного умеренного пояса: структура и жертвы. J. Mammal. 55, 138–157. DOI: 10.2307 / 1379263
CrossRef Полный текст | Google Scholar
Chesson, P. (2000). Механизмы поддержания видового разнообразия. Annu. Rev. Ecol. Syst. 31, 343–366. DOI: 10.1146 / annurev.ecolsys.31.1.343
CrossRef Полный текст | Google Scholar
Ciechanowski, M., Zajac, T., Zielinska, A., and Dunajski, R. (2010). Сезонная активность семи видов летучих мышей-вестертилионид в Польской низменности. Acta Theriol. 55, 301–314. DOI: 10.1007 / BF03193234
CrossRef Полный текст | Google Scholar
Cvikel, N., Egert Berg, K., Levin, E., Hurme, E., Borissov, I., Boonman, A., et al. (2015). Летучие мыши собираются вместе, чтобы улучшить поиск добычи, но могут пострадать, когда их плотность станет слишком высокой. Curr. Биол. 25, 206–211. DOI: 10.1016 / j.cub.2014.11.010
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Данчин Э., Жиральдо Л. А., Валоне Т. Дж. И Вагнер Р.Х. (2004). Публичная информация: от любопытных соседей до культурной эволюции. Наука 305, 487–491. DOI: 10.1126 / science.1098254
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Дехманн, Д. К. Н., Хеук, С. Л., Иуджиоли, Л., Сафи, К., Фойгт, К. К., и Викельски, М. (2009). Экспериментальные доказательства групповой охоты с помощью подслушивания эхолокационных летучих мышей. Proc. Биол. Sci. 276, 2721–2728. DOI: 10.1098 / rspb.2009.0473
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Дорадо-Корреа, А.М., Гёрлиц Х. Р., Симерс Б. М. (2013). Межвидовое акустическое распознавание в двух европейских сообществах летучих мышей. Фронт. Physiol. 4: 192. DOI: 10.3389 / fphys.2013.00192
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Eichstädt, H. (1997). Ressourcennutzung und nischengestaltung einer fledermausgemeinschaft im nordosten brandenburgs. Säugetierkundliche Mitteilungen 40, 3–171.
Fauth, J. E., Bernardo, J., Camara, M., Resetarits, W.Дж., Ван Бускерк, Дж. Мл., И Макколлум, С. А. (1996). Упрощение жаргона общественной экологии: концептуальный подход. Am. Nat. 147, 282–286. DOI: 10.1086 / 285850
CrossRef Полный текст | Google Scholar
Фентон, М. Б. (1990). Пищевое поведение и экология летучих мышей-зверей. Кан. J. Zool. 68, 411–422. DOI: 10.1139 / z90-061
CrossRef Полный текст | Google Scholar
Фентон М. Б. и Белл Г. П. (1979). Эхолокация и пищевое поведение у четырех видов Myotis (Chiroptera). Кан. J. Zool. 57, 1271–1277. DOI: 10.1139 / z79-163
CrossRef Полный текст | Google Scholar
Furmankiewicz, J., and Kucharska, M. (2009). Миграция летучих мышей по большой речной аллее в Юго-Западной Польше. J. Mammal. 90, 1310–1317. DOI: 10.1644 / 09-MAMM-S-099R1.1
CrossRef Полный текст | Google Scholar
Гаузе, Г. Ф. (1934). Борьба за существование . Балтимор, Мэриленд: Уильямс и Уилкинс.
PubMed Аннотация
Гиллам, Э.Х. (2007). Подслушивание летучими мышами жужжания кормления сородичей. Кан. J. Zool. 85, 795–801. DOI: 10.1139 / Z07-060
CrossRef Полный текст | Google Scholar
Gloor, S., Stutz, H. B., and Ziswiler, V. (1995). Особенности питания ночной летучей мыши Nyctalus noctula (Shreber, 1774) в Швейцарии. Myotis 32–33, 231–242.
Горман М. Л., Миллс М. Г., Раат Дж. П. и Спикман Дж. Р. (1998). Высокая стоимость охоты делает африканских диких собак уязвимыми для клептопаразитизма со стороны гиен. Природа 391, 479–481. DOI: 10.1038 / 35131
CrossRef Полный текст | Google Scholar
Грин Э. (1987). Люди в колонии скопы различают информацию высокого и низкого качества. Природа 329, 239–241. DOI: 10.1038 / 329239a0
CrossRef Полный текст | Google Scholar
Привет, К. Дж. (1982). Сравнение численности насекомых в тропических и умеренных широтах. Biotropica 14, 310. DOI: 10.2307 / 2388092
CrossRef Полный текст | Google Scholar
Хальманн, К.A., Sorg, M., Jongejans, E., Siepel, H., Hofland, N., Schwan, H., et al. (2017). За 27 лет общая биомасса летающих насекомых на охраняемых территориях сократилась более чем на 75 процентов. PLoS ONE 12: e0185809. DOI: 10.1371 / journal.pone.0185809
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Heglund, N.C., Taylor, C.R., and McMahon, T.A. (1974). Масштабирование частоты шагов и походки к размеру животного: от мышей к лошадям. Наука 186, 1112–1113. DOI: 10.1126 / наука.186.4169.1112
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хайм, О., Лоренц, Л., Крамер-Шадт, С., Юнг, К., Фойг, К. К., и Эккард, Дж. А. (2017). Ландшафтные и масштабные пространственные ниши летучих мышей, кормящихся над интенсивно используемыми пашнями. Ecol Process. 6:24. DOI: 10.1186 / s13717-017-0091-7
CrossRef Полный текст | Google Scholar
Хайм О., Шредер А., Эккард Дж., Юнг К. и Фойгт К. К. (2016).Сезонная активность европейских летучих мышей выше интенсивно используемых сельскохозяйственных угодий. Agric. Экосист. Environ. 233, 130–139. DOI: 10.1016 / j.agee.2016.09.002
CrossRef Полный текст | Google Scholar
Hiryu, S., Mora, E.C., and Riquimaroux, H. (2016). «Поведенческие и физиологические основы компенсации доплеровского сдвига эхолокационными летучими мышами», в Bat Bioacoustics , ред. М. Б. Фентон, А. Д. Гриннелл, А. Н. Поппер и Р. Р. Фэй (Нью-Йорк, Нью-Йорк: Спрингер), 239–263.
Google Scholar
Холдерид, М. В., Корин, К., Фентон, М. Б., Парсонс, С., Робсон, С., и Джонс, Г. (2005). Интенсивность эхолокационного крика у летучей мыши-ястреба Eptesicus botae (Vespertilionidae) изучалась с помощью стереовидеограмметрии. J. Exp. Биол. 208, 1321–1327. DOI: 10.1242 / jeb.01528
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Холекамп К. Э., Смейл Л., Берг Р. и Купер С. М. (1997). Показатели охоты и успех охоты на пятнистую гиену ( Crocuta crocuta ). J. Zool. 242, 1–15. DOI: 10.1111 / j.1469-7998.1997.tb02925.x
CrossRef Полный текст | Google Scholar
Хамфрис, Н., Симпсон, С., Уэрмут, В., и Симс, Д. (2016). Двое — компания, трое — толпа: мелкомасштабное разделение среды обитания по глубине между симпатрическими видами морских мезохищников. Mar. Ecol. Прог. Сер. 561, 173–187. DOI: 10.3354 / meps11937
CrossRef Полный текст | Google Scholar
Янзен Д. Х. и Понд Д. М. (2009).Сравнение фауны членистоногих вторичной растительности в Мичигане, Англии и Коста-Рике методом выборочной выборки. Пер. R. Entomol. Soc. Лондон. 127, 33–50. DOI: 10.1111 / j.1365-2311.1975.tb00551.x
CrossRef Полный текст
Jeltsch, F., Bonte, D., Pe’er, G., Reineking, B., Leimgruber, P., and Balkenhol, N. (2013). Интеграция экологии перемещений с исследованиями биоразнообразия — изучение новых путей решения пространственно-временной динамики биоразнообразия. Mov. Ecol. 1: 6. DOI: 10.1186 / 2051-3933-1-6
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Калко, Э. К. В. (1995). Преследование насекомых, отлов добычи и эхолокация у летучих мышей (Microchiroptera). Anim. Behav. 50, 861–880. DOI: 10.1016 / 0003-3472 (95) 80090-5
CrossRef Полный текст | Google Scholar
Кишка, Дж., Симон-Буэ, Б., Мартинес, Л., Пусинери, К., Ричард, П., и Риду, В. (2011). Экологическая сегрегация ниш в сообществе симпатрических дельфинов вокруг тропического острова. Mar. Ecol. Прог. Сер. 433, 273–288. DOI: 10.3354 / meps09165
CrossRef Полный текст | Google Scholar
Кунц, Т. Х. (1973). Использование ресурсов: временные и пространственные компоненты активности летучих мышей в Центральной Айове. J. Mammal. 54, 14–32. DOI: 10.2307/1378869
CrossRef Полный текст | Google Scholar
Любин Ю. Д., Монтгомери Г. Г. и Янг О. П. (1977). Пищевые ресурсы муравьедов (Edentata: Myrmecophagidae) I. Годовая перепись древесных гнезд муравьев и термитов на острове Барро-Колорадо, зона Панамского канала. Biotropica 9, 26–34. DOI: 10.2307/2387856
CrossRef Полный текст | Google Scholar
Meyer, C.F.J., Schwarz, C.J. и Fahr, J. (2004). Образцы активности и предпочтения среды обитания насекомоядных летучих мышей в мозаике западноафриканских лесов и саванн. J. Trop. Ecol. 20, 397–407. DOI: 10.1017 / S0266467404001373
CrossRef Полный текст | Google Scholar
Милн Д. Дж., Фишер А., Рейни И. и Пейви К. Р. (2005). Временные структуры летучих мышей в верхней части Северной территории, Австралия. J. Mammal. 86, 909–920. DOI: 10.1644 / 1545-1542 (2005) 86 [909: TPOBIT] 2.0.CO; 2
CrossRef Полный текст | Google Scholar
Морс, Д. Х. (1971). Насекомоядная птица как адаптивная стратегия. Annu. Rev. Ecol. Syst. 2, 177–200. DOI: 10.1146 / annurev.es.02.110171.001141
CrossRef Полный текст | Google Scholar
Наварро, Дж., Вотье, С. К., Агуцци, Дж., Кьеза, Дж. Дж., Фореро, М. Г., и Филлипс, Р. А. (2013). Экологическая сегрегация в пространстве, времени и трофической нише симпатрических планктоядных буревестников. PLoS ONE 8: e62897. DOI: 10.1371 / journal.pone.0062897
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Норберг, У. М., и Райнер, Дж. М. В. (1987). Экологическая морфология и полет летучих мышей (Mammalia, Chiroptera) — Адаптация крыльев, летные характеристики, стратегия поиска пищи и эхолокация. Philos. Пер. R. Soc. Лондон. 316, 337–419. DOI: 10.1098 / rstb.1987.0030
CrossRef Полный текст | Google Scholar
Новачек, Д. П., Friedlaender, A. S., Halpin, P. N., Hazen, E. L., Johnston, D. W., Read, A. J., et al. (2011). Супер-скопления криля и горбатых китов в заливе Вильгельмина, Антарктический полуостров. PLoS ONE 6: e19173. DOI: 10.1371 / journal.pone.0019173
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Плуцински, Т., Змигорски, М., Плуцински, П. (2015). Влияние ночной уборки урожая на активность рукокрылых в сельскохозяйственных ландшафтах. Zool. Ecol. 25, 1–7.DOI: 10.1080 / 21658005.2014.999501
CrossRef Полный текст | Google Scholar
Поллак, Г. Д. (2016). «Нейронная обработка частотных модуляций в слуховой системе летучих мышей». in Bat Bioacoustics , ред. М. Б. Фентон, А. Д. Гриннелл, А. Н. Поппер и Р. Р. Фэй (Нью-Йорк, штат Нью-Йорк: Спрингер), 239–263.
Google Scholar
R Основная команда (2016). R: язык и среда для статистических вычислений .
Ролеке, М., Блом, Т., Kramer-Schadt, S., Yovel, Y., and Voigt, C.C. (2016). Использование летучих мышей в местах обитания по отношению к ветровым турбинам, выявленное с помощью GPS-слежения. Sci. Репутация . 6: 28961. DOI: 10.1038 / srep28961
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ремер К., Дискка Т., Кулон А. и Бас Ю. (2017). Высота полета летучих мышей, отслеживаемая с ветряных мачт, позволяет прогнозировать риск смертности на ветряных электростанциях. Biol. Консерв. 215, 116–122. DOI: 10.1016 / j.biocon.2017.09.002
CrossRef Полный текст | Google Scholar
Сайфул, А.А., Норма Ю. Р. и Азараэ Х. И. (2001). Разделение ниши среди трех симпатрических видов белок, населяющих низменный диптерокарповый лес на полуострове Малайзия. Мамм. Stud. 26, 133–144. DOI: 10.3106 / mammalstudy.26.133
CrossRef Полный текст | Google Scholar
Сальсаменди, Э., Гарин, И., Аростеги, И., Гоити, У., и Айхарца, Дж. (2012). Какой механизм сегрегации ниш допускает сосуществование симпатрических летучих мышей-братьев-сестер-ринолофидов? Фронт. Zool. 9:30.DOI: 10.1186 / 1742-9994-9-30
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Шницлер, Х. У. и Калко, Э. К. В. (2001). Эхолокация летучих мышей-насекомоядных. Bioscience 51, 557–569. DOI: 10.1641 / 0006-3568 (2001) 051 [0557: ebieb] 2.0.co; 2
CrossRef Полный текст | Google Scholar
Скиба, Р. (2003). Europäische Fledermäuse: Kennzeichen, Echoortung und Detektoranwendung, 1st Edn . Хоэнварслебен: Вестарп-Висс.
Тейлор, Р.Дж. И О’Нил М. Г. (1988). Образцы летней активности насекомоядных летучих мышей и их добычи в Тасмании. Aust. Wilflife Res. 55, 533–539.
Google Scholar
Уберникель, К., Чапка, М., и Калко, Э. К. В. (2013). Селективное подслушивание у трех неотропических видов летучих мышей. Этология 119, 66–76. DOI: 10.1111 / eth.12038
CrossRef Полный текст | Google Scholar
Войт, К. К., и Холдерид, М. В. (2012). Высокие затраты на маневрирование вынуждают молосидных летучих мышей добывать корм в открытом космосе. J. Comp. Physiol. B Biochem. Syst. Environ. Physiol. 182, 415–424. DOI: 10.1007 / s00360-011-0627-6
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Фойгт, К. К., Леманн, Д., и Грейф, С. (2015). Стабильные соотношения изотопов водорода разделяют млекопитающих водных и наземных пищевых сетей. Methods Ecol. Evol. 6, 1332–1340. DOI: 10.1111 / 2041-210X.12414
CrossRef Полный текст | Google Scholar
Voigt, C.C., Lindecke, O., Шенборн, С., Крамер-Шадт, С., и Леманн, Д. (2016). Использование в качестве среды обитания перелетных летучих мышей, убитых осенью от ветряных турбин. Ecol. Прил. 26: 771–783. DOI: 10.1890 / 15-0671
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Фойгт, К. К., Шуллер, Б.-М., Грейф, С., и Симерс, Б. М. (2010). Охота на окуня у насекомоядных летучих мышей Rhinolophus связана с высокими энергетическими затратами на маневрирование в полете. J. Comp. Physiol. B Biochem. Syst. Environ.Physiol. 180, 1079–1088. DOI: 10.1007 / s00360-010-0466-x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Фойгт-Хеук, С. Л., Таборски, М., и Дехманн, Д. К. Н. Н. (2010). Двойная функция эхолокации: летучие мыши используют эхолокационные сигналы, чтобы идентифицировать знакомых и незнакомых людей. Anim. Behav. 80, 59–67. DOI: 10.1016 / j.anbehav.2010.03.025
CrossRef Полный текст | Google Scholar
Voigt-Heucke, S.L., Zimmer, S., and Kipper, S.(2016). Способствует ли межвидовое подслушивание скоплениям в воздухе европейских летучих мышей-коньков осенью? Этология 122, 745–757. DOI: 10.1111 / eth.12519
CrossRef Полный текст | Google Scholar
Винтер, Ю., и фон Хелверсен, О. (1998). Энергозатратность полета: летучие мыши летают дешевле птиц? J. Comp. Physiol. B Biochem. Syst. Environ. Physiol. 168, 105–111. DOI: 10.1007 / s003600050126
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Биотические и абиотические факторы внутривидовой изменчивости признаков в популяциях трех видов травянистых растений вдоль широтного градиента | BMC Ecology
Violle C, Navas MML, Vile D, Kazakou E, Fortunel C, Hummel I и др. Пусть понятие черты будет функциональным! Ойкос. 2007; 116: 882–93.
Артикул Google Scholar
Adler PB, Salguero-Gómez R, Compagnoni A, Hsu JS, Ray-mukherjee J, Adler PB, et al. Функциональные черты объясняют различия в стратегиях жизненного цикла растений. Proc Natl Acad Sci. 2014; 111: 740–5.
CAS Статья PubMed Google Scholar
Wright IJ, Reich PB, Cornelissen JHC, Falster DS, Groom PK, Hikosaka K и др. Изменение экономических характеристик листа и взаимосвязи между ними климатом. Glob Ecol Biogeogr. 2005; 14: 411–21.
Артикул Google Scholar
Альберт Ч., Грассейн Ф., Шурр Ф.М., Вильдент Дж., Виолле С. Когда и как следует учитывать внутривидовую изменчивость в экологии растений, основанной на признаках? Perspect Plant Ecol Evol Syst. 2011; 13: 217–25.
Артикул Google Scholar
Messier J, McGill BJ, Lechowicz MJ. Как черты различаются по экологическим шкалам? Аргументы в пользу экологии, основанной на признаках. Ecol Lett. 2010; 13: 838–48.
Артикул PubMed Google Scholar
Каттге Дж., Диас С., Лаворел С., Прентис И.К., Лидли П., Бениш Дж. И др. TRY — глобальная база данных по характеристикам растений. Glob Chang Biol. 2011; 17: 2905–35.
Артикул PubMed Central Google Scholar
Митчелл Р.М., Баккер Дж. Д. Количественная оценка и сравнение внутривидовой изменчивости функциональных признаков: тематическое исследование с Hypochaeris radicata . Funct Ecol. 2014; 28: 258–69.
Артикул Google Scholar
Siefert A, Violle C, Chalmandrier L, Cécile HA, Taudiere A, Fajardo A, et al. Глобальный метаанализ относительной степени вариации внутривидовых признаков в растительных сообществах. Ecol Lett. 2015; 18: 1406–19.
Артикул PubMed Google Scholar
Hart SP, Schreiber SJ, Levine JM, Coulson T. Как различия между людьми влияют на сосуществование видов. Ecol Lett. 2016; 19: 825–38.
Артикул PubMed Google Scholar
Clark JS. Особи и разновидности, необходимые для высокого видового разнообразия лесных деревьев. Наука. 2010. 327: 1129–32.
CAS Статья PubMed Google Scholar
Юнг В., Виолле С., Монди С., Хоффманн Л., Мюллер С. Внутривидовая изменчивость и сборка сообществ на основе признаков. J Ecol. 2010; 98: 1134–40.
Артикул Google Scholar
Chalmandrier L, Münkemüller T, Colace M-P, Renaud J, Aubert S, Carlson BZ, et al. Пространственный масштаб и внутривидовая изменчивость признаков опосредуют правила сборки на альпийских лугах. J Ecol. 2017; 105: 277–87.
Артикул Google Scholar
Hooper DU, Chapin FS III, Ewel JJ, Hector A, Inchausti P, Lavorel S и др. Влияние биоразнообразия на функционирование экосистемы: консенсус современных знаний. Ecol Monogr. 2005; 75: 3–35.
Артикул Google Scholar
Lecerf A, Chauvet E. Внутривидовая изменчивость свойств листьев сильно влияет на разложение листьев ольхи в ручье. Basic Appl Ecol. 2008; 9: 598–605.
Артикул Google Scholar
Гаррет К.А., Зуньига Л.Н., Ронкал Э., Форбс Г.А., Мундт С.К., Су З., Нельсон Р.Дж. Внутривидовое функциональное разнообразие хозяев и его влияние на риск заболевания в зависимости от климатического градиента. Ecol Appl. 2009; 19: 1868–83.
CAS Статья PubMed Google Scholar
Хоф К., Левинский И., Араужо М.Б., Рахбек К. Переосмысление способности видов справляться с быстрым изменением климата. Glob Chang Biol. 2011; 17: 2987–90.
Артикул Google Scholar
Lemke IH, Kolb A, Graae BJ, De Frenne P, Acharya KP, Blandino C и др. Паттерны изменения фенотипических признаков у двух лесных трав умеренного пояса в широком климатическом градиенте. Завод Ecol. 2015; 216: 1523–36.
Артикул Google Scholar
Etterson JR, Schneider HE, Gorden NLS, Weber JJ. Эволюционные идеи из исследований географических вариаций: современные вариации и взгляд в будущее. Am J Bot. 2016; 103: 5–9.
Артикул PubMed Google Scholar
Болник Д.И., Амарасекаре П., Араужо М.С., Бюргер Р., Левин Дж. М., Новак М. и др. Почему внутривидовые вариации признаков имеют значение в экологии сообщества. Trends Ecol Evol. 2011; 26: 183–92.
Артикул PubMed PubMed Central Google Scholar
Альберт Ч., Туиллер В., Йоккоз Н.Г., Судант А., Буше Ф., Сакконе П. и др.Внутривидовая функциональная изменчивость: степень, структура и источники изменчивости. J Ecol. 2010; 98: 604–13.
Артикул Google Scholar
Фахардо А., Пайпер FI. Внутривидовая изменчивость и ковариация у широко распространенных видов деревьев ( Nothofagus pumilio ) на юге Чили. Новый Фитол. 2011; 189: 259–71.
Артикул PubMed Google Scholar
De Frenne P, Graae BJ, Kolb A, Shevtsova A, Baeten L, Brunet J, et al. Внутривидовое применение схемы стратегии экологии высоты листа и семян к лесным травам вдоль широтного градиента. Экография. 2011; 34: 132–40.
Артикул Google Scholar
Poorter H, Niinemets U, Poorter L, Wright IJ, Villar R. Причины и последствия вариаций в массе листьев на площади (LMA): метаанализ. Новый Фитол. 2009. 182: 565–88.
Артикул PubMed Google Scholar
Poorter H, Niinemets Ü, Walter A, Fiorani F, Schurr U. Метод построения кривых доза-реакция для широкого диапазона факторов окружающей среды и характеристик растений посредством метаанализа фенотипических данных. J Exp Bot. 2010; 61: 2043–55.
CAS Статья PubMed Google Scholar
Оже С., Шипли Б.Межвидовые и внутривидовые вариации признаков на коротких градиентах окружающей среды в старовозрастном лесу умеренной зоны. J Veg Sci. 2013; 24: 419–28.
Артикул Google Scholar
Albert CH, Thuiller W, Yoccoz NG, Douzet R, Aubert S, Lavorel S. Многоступенчатый подход позволяет выявить структуру и относительную важность внутривидовой и межвидовой изменчивости свойств растений. Funct Ecol. 2010; 24: 1192–201.
Артикул Google Scholar
Карлуччи МБ, Дебастиани В.Дж., Столб В.Д., Дуарте, LDS. Межвидовая и внутривидовая изменчивость признаков и сборка сообществ саженцев на участках леса. J Veg Sci. 2015; 26: 21–31.
Артикул Google Scholar
Mason NWH, De Bello F, Doležal J, Lepš J. Перекрытие ниш выявляет эффекты конкуренции, беспокойства и контрастных процессов сборки в экспериментальных сообществах пастбищ. J Ecol. 2011; 99: 788–96.
Артикул Google Scholar
Lemke IH, Kolb A, Diekmann MR. Условия региона и участка влияют на вариацию фенотипических признаков пяти лесных трав. Acta Oecol. 2012; 39: 18–24.
Артикул Google Scholar
Le Bagousse-Pinguet Y, Börger L, Quero J, Garcia-Gomez M, Soriano S, Maestre FT, et al. Признаки соседних растений и ограниченность пространства определяют внутривидовую изменчивость признаков полузасушливых кустарников. J Ecol. 2015; 103: 1647–57.
Артикул Google Scholar
Герц К., Дитц С., Хайдер С., Джандт У., Шил Д., Брюльхайде Х. Факторы внутривидовой изменчивости признаков травяных и разнотравных видов на немецких лугах и пастбищах. J Veg Sci. 2017; 28: 705–16.
Артикул Google Scholar
Valladares F, Gianoli E, Gómez JM. Экологические пределы фенотипической пластичности растений. Новый Фитол. 2007; 176: 749–63.
Артикул PubMed Google Scholar
Hulshof CM, Violle C, Spasojevic MJ, Mcgill B, Damschen E, Harrison S, et al. Внутривидовые и межвидовые вариации в определенной площади листа показывают важность абиотических и биотических факторов видового разнообразия на высоте и широте. J Veg Sci. 2013; 24: 921–31.
Артикул Google Scholar
Хоффманн А.А., Мерила Дж. Наследственные изменения и эволюция при благоприятных и неблагоприятных условиях. Trends Ecol Evol.1999; 14: 96–101.
CAS Статья PubMed Google Scholar
Рей О., Данчин Э., Мироуз М., Лут С., Бланшет С. Адаптация к глобальным изменениям: эпигенетическая перспектива с переносимым элементом. Trends Ecol Evol. 2016; 31: 514–26.
Артикул PubMed Google Scholar
Виолле С., Энквист Б.Дж., МакГилл Б.Дж., Цзян Л., Альберт С.Х., Хулсхоф К. и др. Возврат дисперсии: внутривидовая изменчивость в экологии сообщества.Trends Ecol Evol. 2012; 27: 244–52.
Артикул PubMed Google Scholar
Le Bagousse-Pinguet Y, De Bello F, Vandewalle M, Leps J, Sykes MT. Видовое богатство известняковых пастбищ увеличивается с наложением признаков: данные о разделении функционального разнообразия внутри и между видами. J Ecol. 2014; 102: 466–74.
Артикул Google Scholar
Cadotte MW, Carscadden K, Mirotchnick N.Помимо видов: функциональное разнообразие и поддержание экологических процессов и услуг. J Appl Ecol. 2011; 48: 1079–87.
Артикул Google Scholar
Mason NWH, Mouillot D, Lee WG, Wilson JB. Функциональное богатство, функциональная однородность и функциональное расхождение: основные компоненты функционального разнообразия. Ойкос. 2005; 111: 112–8.
Артикул Google Scholar
Бернс Дж., Штраус С. Влияние конкуренции на филогенетический сигнал и фенотипическую пластичность функциональных признаков растений. Экология. 2012; 93: 126–37.
Артикул Google Scholar
Беннетт Дж. А., Рийбак К., Тамме Р., Льюис Р. Дж., Пэртель М. Взаимосвязь между конкуренцией и внутривидовой изменчивостью признаков. J Ecol. 2016; 104: 1410–20.
Артикул Google Scholar
Бертнесс, доктор медицины, Каллавей Р. Позитивное взаимодействие в сообществах. Trends Ecol Evol. 1994; 9: 187–91.
Артикул Google Scholar
Дэвидсон А.М., Дженнионс М., Никотра А.Б. Проявляют ли инвазивные виды более высокую фенотипическую пластичность, чем местные виды, и если да, то является ли это адаптивным? Метаанализ. Ecol Lett. 2011; 14: 419–31.
Артикул PubMed Google Scholar
Westoby M. Схема стратегии экологии растений с высотой листа (LHS). Почва растений. 1998; 199: 213–27.
CAS Статья Google Scholar
Гарнье Э., Навас МЛ. Основанный на признаках подход к сравнительной функциональной экологии растений: концепции, методы и приложения для агроэкологии. Обзор. Agron Sustain Dev. 2012; 32: 365–99.
Артикул Google Scholar
Грайм Дж. Доказательства существования трех основных стратегий у растений и их значение для экологической и эволюционной теории. Am Nat. 1977; 111: 1169–94.
Артикул Google Scholar
Хант Р., Ходжсон Дж., Томпсон К., Бангенер П., Даннетт Н., Аскью А. и др. Новый практический инструмент для получения функциональной сигнатуры травянистой растительности. Appl Veg Sci. 2004; 7: 163–70.
Артикул Google Scholar
Ширреффс Д. Биологическая флора Британских островов. Anemone Nemorosa L. J Ecol. 1985; 73: 1005–20.
Артикул Google Scholar
Де Френн П., Брюне Дж., Куньон М., Декок Дж., Грэе Б.Дж., Хагенблад Дж. И др. Биологическая флора Британских островов: Milium effusum . J Ecol. 2017; 105: 839–58.
Артикул Google Scholar
Beerling DJ, Perrins JM.Биологическая флора Британских островов. Impatiens glandulifera Royle ( Impatiens roylei Walp.). J Ecol. 1993. 81: 367–82.
Артикул Google Scholar
Гринвуд П., Кун, штат Нью-Джерси. Способствует ли инвазивное растение Impatiens glandulifera эрозии почвы вдоль прибрежной зоны? Исследование небольшого водотока на северо-западе Швейцарии. J Почвенные отложения. 2014; 14: 637–50.
Артикул Google Scholar
Ачарья КП. Инвазивные виды: генетика, характеристики и изменение признаков по широтному градиенту. Кандидат наук. защитил диссертацию, Норвежский университет науки и технологий (NTNU), Норвегия. 2014.
Cornelissen JHC, Lavorel S, Garnier E, Díaz S, Buchmann N, Gurvich DE, et al. Справочник протоколов для стандартизированного и простого измерения функциональных характеристик растений во всем мире. Ост Дж. Бот. 2003. 51: 335–80.
Артикул Google Scholar
Climate-data.org. Среднегодовые температуры и среднегодовые осадки. 2014. http://climate-data.org/. По состоянию на 9 мая 2016 г.
De Frenne P, Graae BJ, Kolb A, Brunet J, Chabrerie O, Cousins SAO, et al. Значительное влияние температуры на репродуктивную способность лесной травы Anemone nemorosa L. Для Ecol Manage. 2010; 259: 809–17.
Артикул Google Scholar
Линдси А., Ньюман Дж.Использование официальных данных о погоде в весеннее время: температурный анализ фенологических данных Индианы. Экология. 1956; 37: 812–23.
Артикул Google Scholar
NOAA. Национальные центры экологической информации. Национальное управление океанических и атмосферных исследований. 2016. https://www.ncdc.noaa.gov/. По состоянию на 14 сентября 2016 г.
Klotz S, Kühn I, Durka W. BIOLFLOR-Eine Datenbank zu biologisch-ökologischen Merkmalen der Gefäßpflanzen в Германии.Schriftenreihe für Vegetationskunde. 2002; 38: 1–333.
Google Scholar
Kleyer M, Bekker R, Knevel IC, Bakker JP, Thompson K, Sonnenschein M, et al. LEDA Traitbase: база данных жизненных особенностей флоры Северо-Западной Европы. J Ecol. 2008; 96: 1266–74.
Артикул Google Scholar
Villéger S, Mason NWH, Mouillot D. Новые многомерные индексы функционального разнообразия для многогранной основы функциональной экологии.Экология. 2008. 89: 2290–301.
Артикул PubMed Google Scholar
Лалиберте Э., Шипли Б. Ф. Д.: измерение функционального разнообразия по множеству признаков и другие инструменты для функциональной экологии. Пакет R версии 1.0-12. 2014.
Азур М.Дж., Стюарт Е.А., Франгакис К., Лиф П.Дж. Множественное вменение с помощью связанных уравнений: что это такое и как это работает? Int J Methods Psychiatr Res. 2011; 20: 40–9.
Артикул PubMed PubMed Central Google Scholar
van Buuren S, Groothuis-Oudshoorn K. Мыши: многомерное вменение с помощью связанных уравнений в R.J. Stat Softw. 2011; 45: 3.
Артикул Google Scholar
Taugourdeau S, Villerd J, Plantureux S, Huguenin-Elie O, Amiaud B. Заполнение пробелов в базах данных по функциональным признакам: использование экологических гипотез для замены недостающих данных. Ecol Evol. 2014; 4: 944–58.
Артикул PubMed PubMed Central Google Scholar
Nakagawa S, Schielzeth H. Общий и простой метод получения R 2 из обобщенных линейных моделей смешанных эффектов. Методы Ecol Evol. 2013; 4: 133–42.
Артикул Google Scholar
Пинейро Дж., Бейтс Д., Деброй С., Саркар Д., Р. Основная команда. nlme: линейные и нелинейные модели смешанных эффектов. Пакет R версии 3.1-131. 2017. https://cran.r-project.org/package=nlme. По состоянию на 16 июля 2017 г.
Wasof S, Lenoir J, Gallet-Moron E, Jamoneau A, Brunet J, Cousins SAO и др. Смещение экологических ниш подлеска по широтному градиенту лесов умеренного пояса в северо-западной Европе.
Leave A Comment