
Влияние физических нагрузок на организм спортсмена. Гормоны и физическая нагрузка.
Гормоны играют крайне важную роль в работе человеческого организма. Эти вещества стимулируют работу определенных клеток и систем организма. Гормоны производятся эндокринными железами и определенными тканями.
Из широкого спектра гормонов особую важность имеют анаболические и катаболические гормоны. Катаболизм – это процесс метаболического распада клеток и тканей, а также разложения сложных структур с выделением энергии в виде тепла или в виде аденозинтрифосфата. Катаболические процессы обеспечивают высвобождение большого количества энергии.
Анаболические процессы противоположны катаболическим. Под анаболическими процессами подразумевают процессы создания клеток и тканей, а также веществ, необходимых для работы организма. Течение регенеративных процессов и анаболизм мышечной ткани во многом зависят от уровня гормона роста, инсулина и тестостерона в плазме крови.
Физическая активность существенно повышает концентрацию множества гормонов в плазме крови и не только непосредственно в момент нагрузки. С начала выполнения упражнения (напр. около максимальной мощности), за первые 4-10 минут концентрация различных гормонов и продуктов метаболизма меняется самопроизвольно. Так с началом упражнения растет концентрация молочной кислоты в крови. А концентрация глюкозы начинает меняться обратно пропорционально концентрации молочной кислоты. При увеличении времени нагрузки в крови растет уровень соматотропина.
Другие исследования продемонстрировали, что у людей преклонного возраста (65-75 лет) после занятий на велотренажере уровень тестостерона увеличивался на 40%. Специалисты геронтологии полагают, что именно сохранение нормальной концентрации тестостерона обеспечивает бодрое, энергичное состояние в преклонные годы и, вероятно, увеличивает продолжительность жизни.
Секрецию гормонов и их попадание в кровь при физических упражнениях можно представить в виде каскада реакций. Физическое напряжение как стресс провоцирует выделение в структурах мозга либеринов, которые, в свою очередь, запускают производство тропинов гипофизом. Через кровь тропины проникают в эндокринные железы, где и осуществляется секреция гормонов.
Физическое напряжение как стресс провоцирует выделение в структурах мозга либеринов, которые, в свою очередь, запускают производство тропинов гипофизом. Через кровь тропины проникают в эндокринные железы, где и осуществляется секреция гормонов.
Катаболизм обусловлен наличием в крови множества факторов, участвующих в высвобождении энергии. Один из этих факторов – 
Устранить катаболический эффект кортизола можно с применением анаболических стероидов. Но этот метод – крайне вреден для здоровья. Побочные явления столь опасны, что спортсмену следует найти другие эффективные анаболики, легальные и не вызывающие побочных эффектов. Получение организмом большого количества сахаридов в результате анаболической активности инсулина также благоприятствует быстрому восстановлению. Выяснилось, что и в данном случае эффект достигается ингибированием активности кортизола. Концентрация инсулина обратно пропорциональна концентрации кортизола в крови. Инсулин является полипептидным гормоном и необходим в объединении путей энергоснабжения. Анаболизм инсулина затрагивает мышечную, жировую ткань и печень. Инсулин стимулирует образование гликогена, алифатических кислот и протеинов.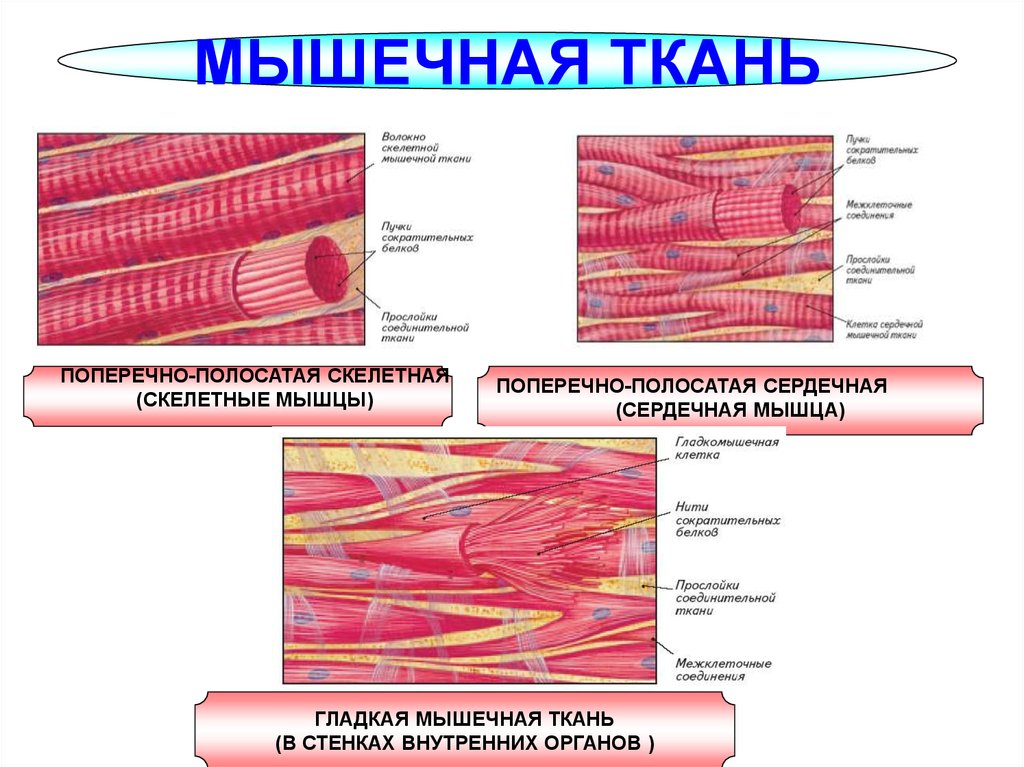
Сам механизм анаболизма инсулина состоит в ускорении попадания глюкозы и свободных аминокислот в клетки. Однако процессы образования гликогена, активируемые инсулином, провоцируют уменьшение концентрации глюкозы в крови (основной симптом гипогликемии). Инсулин замедляет катаболизм в организме, в т.ч. разложение гликогена и нейтрального жира. Ускорение анаболизма в организме, то, чего хотят большинство культуристов, возможно и без применения допинг-средств типа анаболических стероидов.
Одним из важнейших агентов, активирующих производство протеина, является прогормон – соматомедин С. Специалисты утверждают, что образование этого вещества стимулируется соматотропином и осуществляется в печени и мышечной ткани. Производство соматомедина С в определенной степени зависит от объёма аминокислот, получаемых организмом. Гормоны с анаболическим эффектом после физических упражнений выполняют еще одну задачу. В результате исследований было выяснено, что при физических нагрузках волокна мышц повреждаются.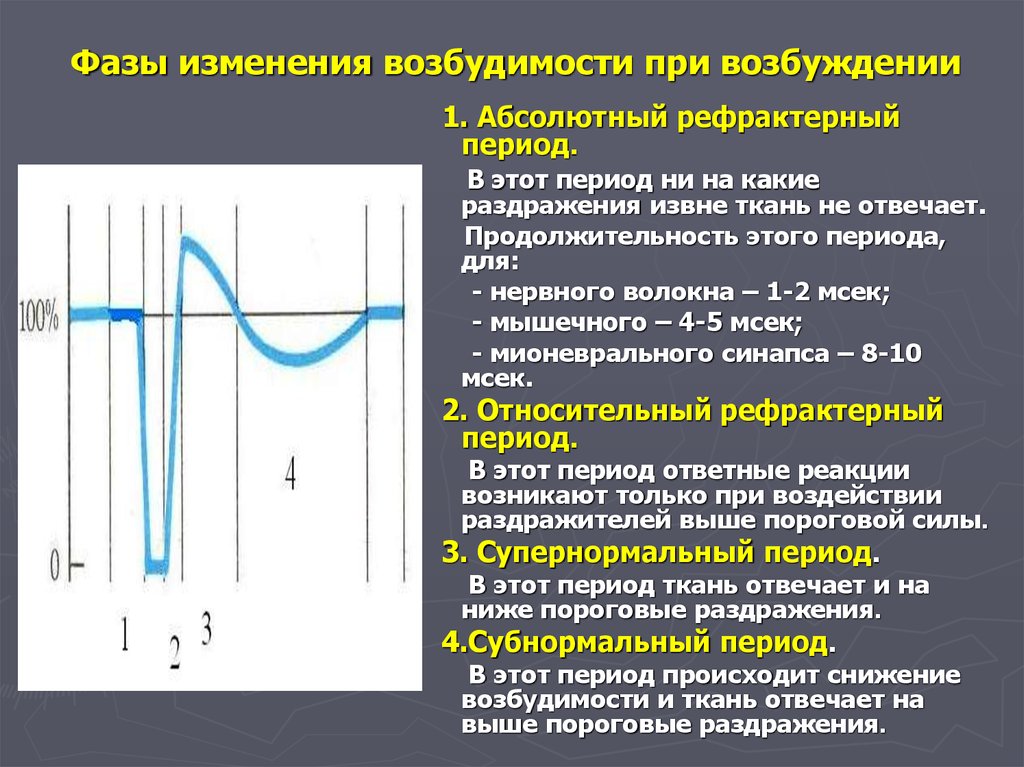
Эндокринная система управляет всеми видами метаболизма и, в зависимости от ситуации, может активировать резервные силы организма. Она же контролирует восстановление после тяжелых физических упражнений. Причем реакции гормональных систем сильно отличаются в соответствии со степенью нагрузки (большой или умеренной мощности).
При нагрузке умеренной мощности и долгой тренировке увеличивается уровень гормона роста и кортизола, падает уровень инсулина и увеличивается уровень трииодтиронина.
Нагрузке большой мощности сопутствует увеличение концентрации гормона роста, кортизола, инсулина и Т3. Гормон роста и кортизол обуславливают развитие специальной работоспособности, и поэтому увеличение их концентрации во время разных тренировочных циклов сопровождается улучшением спортивных показателей спортсмена.
В результате многих исследований специалистов было выяснено, что у профессиональных бегунов на сверхдальние дистанции в спокойном состоянии обнаруживается низкая или нормальная концентрация гормона роста. Однако при марафоновском забеге уровень гормона роста в крови сильно увеличивается, что обеспечивает высокую работоспособность на продолжительный срок. Гормон роста (соматотропин) – гормон, отвечающий за анаболизм в организме (рост, развитие, увеличение веса тела и различных органов). В организме взрослого человека воздействие гормона роста на функции роста в большей степени теряется, а на анаболические функции (образование протеина, сахаридный и жировой обмены) остается. Это и является причиной запрета соматотропного гормона как допинга.
Другим немаловажным гормоном адаптации служит кортизол, который отвечает за сахаридный и протеиновый метаболизм. Кортизол контролирует работоспособность путем катаболического процесса, при котором печень снабжается гликогеном и кетогенными аминокислотами. Вместе с катаболическим процессом (остановка производства протеина в лимфоидной и соединительной тканях) осуществляется сохранение концентрации глюкозы в плазме крови спортсмена на достаточном уровне. Данный гормон также запрещен в качестве допинга. Инсулин управляет концентрацией глюкозы и ее перемещением через мембраны мышечных и других клеток. Уровень инсулина в норме – 5-20 мкед/мл. Нехватка инсулина снижает работоспособность вследствие уменьшения количества глюкозы, доставляемой в клетки.
Вместе с катаболическим процессом (остановка производства протеина в лимфоидной и соединительной тканях) осуществляется сохранение концентрации глюкозы в плазме крови спортсмена на достаточном уровне. Данный гормон также запрещен в качестве допинга. Инсулин управляет концентрацией глюкозы и ее перемещением через мембраны мышечных и других клеток. Уровень инсулина в норме – 5-20 мкед/мл. Нехватка инсулина снижает работоспособность вследствие уменьшения количества глюкозы, доставляемой в клетки.
Выделение инсулина стимулируется при упражнениях большой мощности, что обеспечивает высокую проницаемость клеточных мембран для глюкозы (стимулируется гликолиз). Работоспособность достигается благодаря сахаридному обмену. При умеренной мощности упражнений уровень инсулина падает, что приводит к переходу с сахаридного метаболизма на липидный, что столь востребовано при продолжительной физической активности, когда резервы гликогена частично израсходованы.
Тиреоидные гормоны тироксин и трииодтиронин управляют основным метаболизмом, расходом кислорода и окислительным фосфорилированием. Изменение уровня тиреоидных гормонов определяет предел работоспособности и выносливости человека (возникает дисбаланс между получением кислорода и фосфорилированием, замедляется окислительное фосфорилирование в митохондриях мышечных клеток, замедляется ресинтез аденозинтрифосфата). Обследования бегунов на сверхдальние дистанции продемонстрировали связь между работоспособностью и соотношением гормона роста и кортизола.
Изменение уровня тиреоидных гормонов определяет предел работоспособности и выносливости человека (возникает дисбаланс между получением кислорода и фосфорилированием, замедляется окислительное фосфорилирование в митохондриях мышечных клеток, замедляется ресинтез аденозинтрифосфата). Обследования бегунов на сверхдальние дистанции продемонстрировали связь между работоспособностью и соотношением гормона роста и кортизола.
Обследование эндокринной системы определенного спортсмена позволяет определить его возможности и готовность выдержать физическую нагрузку с лучшими показателями. Другим существенным аспектом предсказания специальной работоспособности служит выявление способностей коры надпочечников производить кортизол в ответ на раздражение адренокортикотропным гормоном. Повышенное производство кортизола говорит о способности спортсмена работать в оптимальном режиме.
Спортивная работоспособность разных полов существенно зависит от тестостерона. Этот гормон обуславливает агрессию, темперамент и целеустремленность при исполнении задания. Гормональные средства (тестостерон и его вариации, анаболические стероиды, гормон роста, кортикотропин, гонадотропный гормон, эритропоэтин) искусственно увеличивают работоспособность человека, и поэтому считаются допингом и запрещены к употреблению в соревнованиях и на тренировках.
Гормональные средства (тестостерон и его вариации, анаболические стероиды, гормон роста, кортикотропин, гонадотропный гормон, эритропоэтин) искусственно увеличивают работоспособность человека, и поэтому считаются допингом и запрещены к употреблению в соревнованиях и на тренировках.
Зачастую употребление препаратов гормонов идет вразрез со здоровым образом жизни и в конечном счете может привести к тяжелым патологиям.
Статья подготовлена главным врачом ГУЗ «ОВФД»
Николаевой И.В.
404 Cтраница не найдена
Мы используем файлы cookies для улучшения работы сайта МГТУ и большего удобства его использования. Более подробную информацию об использовании файлов cookies можно найти здесь.
Продолжая пользоваться сайтом, вы подтверждаете, что были проинформированы об использовании файлов cookies сайтом ФГБОУ ВО «МГТУ» и согласны с нашими правилами обработки персональных данных.
Размер:
AAA
Изображения Вкл. Выкл.
Обычная версия сайта
К сожалению запрашиваемая страница не найдена.
Но вы можете воспользоваться поиском или картой сайта ниже
|
|
 Педагогический (научно-педагогический) состав
Педагогический (научно-педагогический) составРеакция мышц на одновременные стимулированные и физиологические серии потенциалов действия – имитационное исследование
- Список журналов
- Рукописи авторов HHS
- PMC4229042
В качестве библиотеки NLM предоставляет доступ к научной литературе.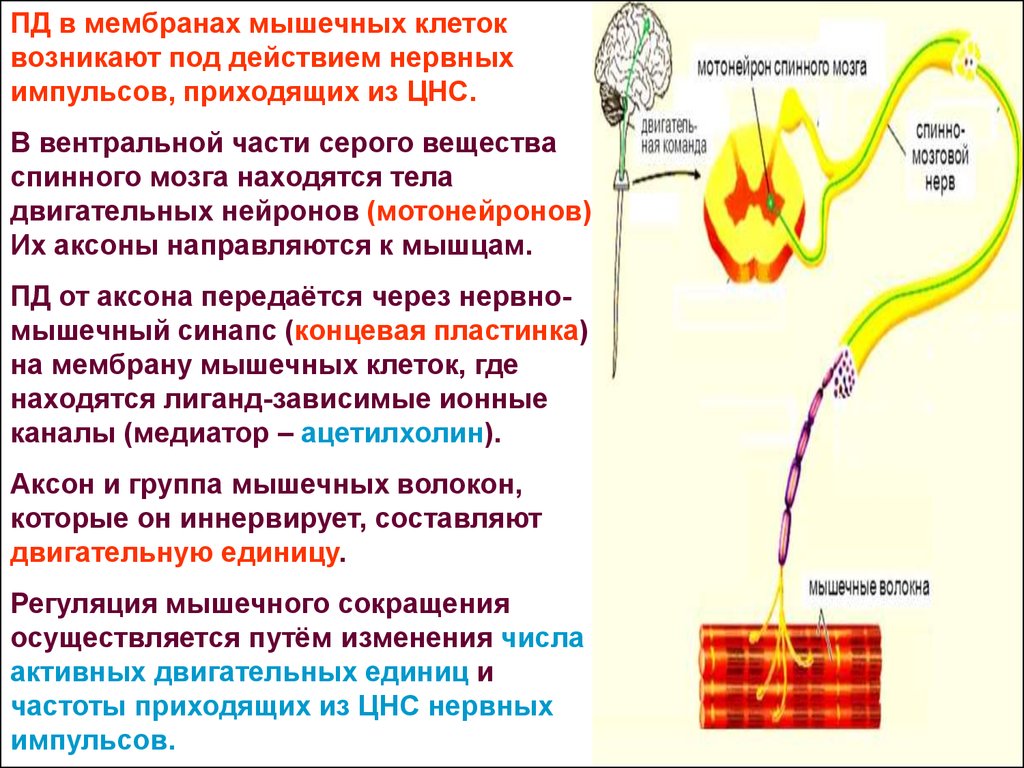 Включение в базу данных NLM не означает одобрения или согласия с
содержание NLM или Национальных институтов здравоохранения.
Узнайте больше о нашем отказе от ответственности.
Включение в базу данных NLM не означает одобрения или согласия с
содержание NLM или Национальных институтов здравоохранения.
Узнайте больше о нашем отказе от ответственности.
Conf Proc IEEE Eng Med Biol Soc. Авторская рукопись; доступно в PMC 2014 12 ноября.
Опубликовано в окончательной редакции как:
Conf Proc IEEE Eng Med Biol Soc. 2012 г.; 2012: 1839–1842 гг.
doi: 10.1109/EMBC.2012.6346309
PMCID: PMC4229042
NIHMSID: NIHMS620557
PMID: 23366270
900 Цель этого исследования состояла в том, чтобы оценить механизмы, ответственные за экспериментально наблюдаемое нелинейное сложение сил, вызванных произвольными сокращениями при наложенной электрической стимуляции той же мышцы. Модель взаимодействия потенциала действия предсказывает повышенную скорость возбуждения двигательных единиц во время наложенной стимуляции.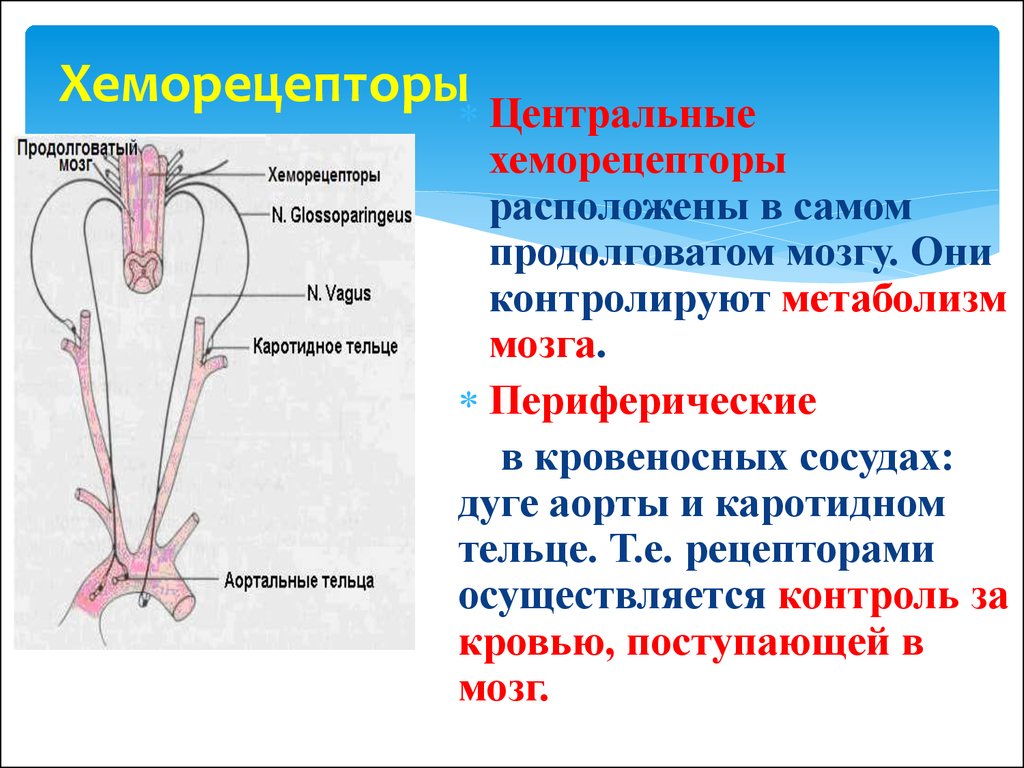 Результирующие эффекты на производство силы воспроизводят экспериментальные результаты, подтверждая, что насыщение двигательных единиц силой способствует нелинейному увеличению силы. Модель также предсказывает, что произвольная ЭМГ будет снижена при стимуляции из-за блокады столкновения и сброса фазы потенциалов действия двигательных единиц. Оба эффекта имеют значение для конструкции систем нейропротезов FES.
Результирующие эффекты на производство силы воспроизводят экспериментальные результаты, подтверждая, что насыщение двигательных единиц силой способствует нелинейному увеличению силы. Модель также предсказывает, что произвольная ЭМГ будет снижена при стимуляции из-за блокады столкновения и сброса фазы потенциалов действия двигательных единиц. Оба эффекта имеют значение для конструкции систем нейропротезов FES.
Электрическая стимуляция нервов широко используется в качестве терапевтического или восстановительного вмешательства, например, кожная стимуляция для облегчения боли или функциональная электрическая стимуляция (ФЭС) для восстановления функции руки и кисти в случаях тетраплегии после травмы спинного мозга. Однако систематического анализа ожидаемых моделей одновременных последовательностей потенциалов действия, создаваемых электрической стимуляцией нервов во время текущей физиологической активности, не проводилось. В зависимости от взаимодействия между внутренними и внешними источниками активации стимуляция на выходе будет вызывать разные физиологические эффекты.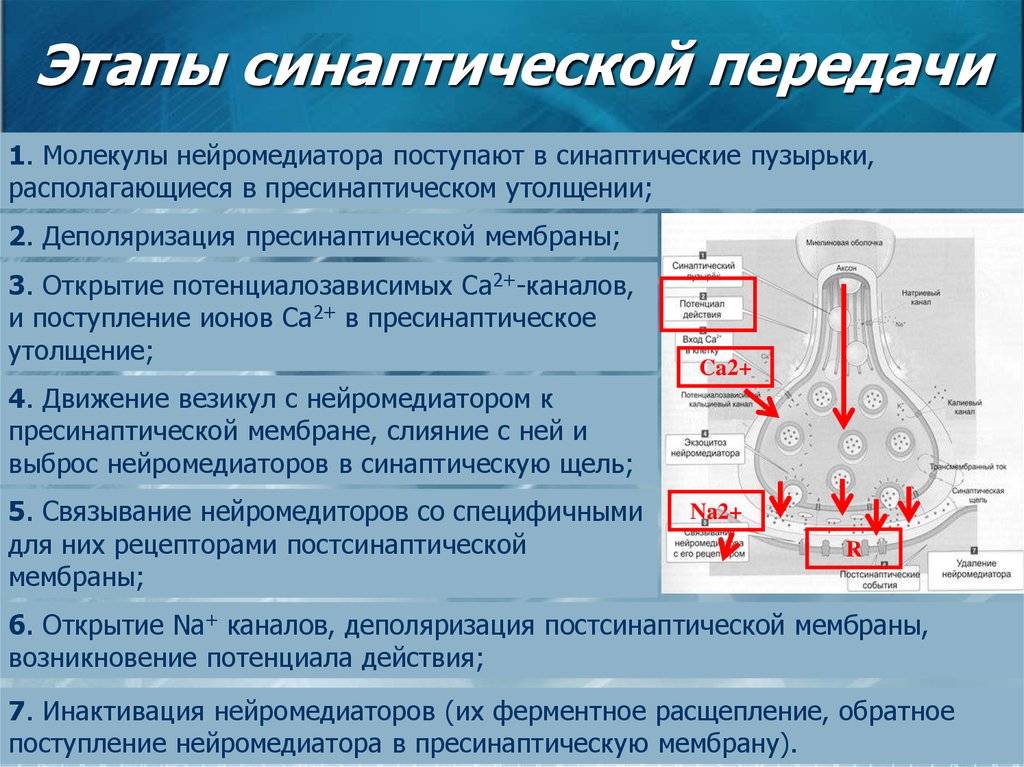 Мы исследовали взаимодействие между произвольной двигательной активностью и стимуляцией периферических двигательных аксонов на мышечную силу и генерацию ЭМГ. Это особенно важно при разработке нейропротезов для выживших после инсульта, поскольку они часто имеют слабость, которая может быть уменьшена с помощью электрической стимуляции мышц, но они также имеют постоянные физиологически генерируемые паттерны мышечного возбуждения, которые активируются остаточными произвольными усилиями и повышенными уровнями активности. тонус и рефлексы. Таким образом, способность наложенной стимуляции генерировать дополнительную силу может быть ограничена, как это наблюдалось экспериментально [1], [2]. Суммарная сила меньше суммы сил, измеренных по отдельности.
Мы исследовали взаимодействие между произвольной двигательной активностью и стимуляцией периферических двигательных аксонов на мышечную силу и генерацию ЭМГ. Это особенно важно при разработке нейропротезов для выживших после инсульта, поскольку они часто имеют слабость, которая может быть уменьшена с помощью электрической стимуляции мышц, но они также имеют постоянные физиологически генерируемые паттерны мышечного возбуждения, которые активируются остаточными произвольными усилиями и повышенными уровнями активности. тонус и рефлексы. Таким образом, способность наложенной стимуляции генерировать дополнительную силу может быть ограничена, как это наблюдалось экспериментально [1], [2]. Суммарная сила меньше суммы сил, измеренных по отдельности.
A. Моделирование взаимодействия между произвольными и стимулированными последовательностями потенциалов действия
Сила мышечного сокращения, вызванного источником произвольной или электрической стимуляции, модулируется процессами рекрутирования и модуляции частоты возбуждения, а в случае одновременного возбуждения, т.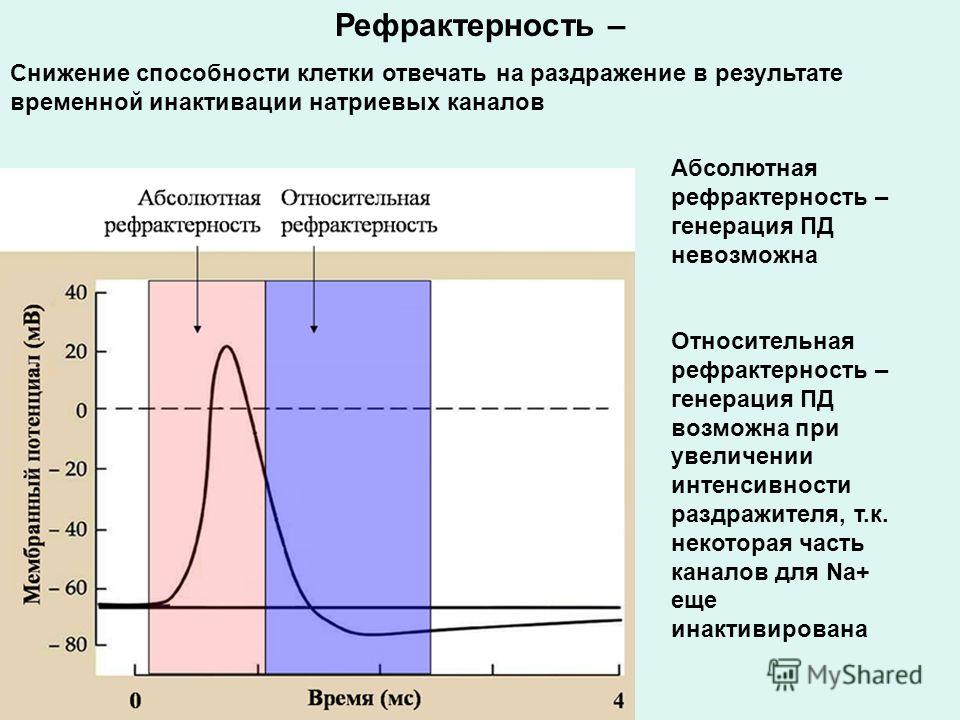 е. один и тот же аксон возбуждается обоими источниками в разных местах, результирующие потенциалы действия будут взаимодействовать, как показано на рис. Потенциалы действия от мотонейрона проводятся ортодромно к мышечным волокнам. Электрический стимул возбуждает аксон в какой-то точке между двигательным нейроном и мышцей и создает пары потенциалов действия, которые распространяются как в ортодромном, так и в антидромном направлениях. Взаимодействия между произвольно и электрически генерируемыми потенциалами действия в одиночных аксонах зависят от их относительного времени и времени проведения между двумя источниками.
е. один и тот же аксон возбуждается обоими источниками в разных местах, результирующие потенциалы действия будут взаимодействовать, как показано на рис. Потенциалы действия от мотонейрона проводятся ортодромно к мышечным волокнам. Электрический стимул возбуждает аксон в какой-то точке между двигательным нейроном и мышцей и создает пары потенциалов действия, которые распространяются как в ортодромном, так и в антидромном направлениях. Взаимодействия между произвольно и электрически генерируемыми потенциалами действия в одиночных аксонах зависят от их относительного времени и времени проведения между двумя источниками.
Открыть в отдельном окне
Взаимодействие между произвольными и электрическими потенциалами действия на одном моторном аксоне.
Предположим, что произвольный потенциал действия возникает в момент времени t v , а электрический стимул вызывает пару потенциалов действия через t с . с момента последнего стимулирующего импульса. Время проведения между двумя источниками составляет t c (расстояние, деленное на скорость), а время проведения от стимулирующего электрода до мышцы обозначается как т р .
Время проведения между двумя источниками составляет t c (расстояние, деленное на скорость), а время проведения от стимулирующего электрода до мышцы обозначается как т р .
Произвольно вызванный ортодромный потенциал действия будет сталкиваться с электрически вызванным антидромным потенциалом действия, если время между генерацией двух потенциалов действия меньше, чем время проведения между нейроном и источником электрической стимуляции, т. е. если либо t с < t v + t c или t v < t s + т с . Взаимная аннигиляция в точке столкновения двух потенциалов действия будет препятствовать тому, чтобы потенциал произвольного действия достиг мышцы, а электрически генерируемый потенциал антидромного действия достиг мотонейрона. Тем не менее, парный электрически генерируемый ортодромный потенциал действия достигнет мышцы, фактически заменив произвольный потенциал действия, но достигнув мышцы со смещением во времени (раньше или позже) на величину, равную разнице во времени двух возбуждений, t s – t v .
Если потенциал произвольного действия проходит мимо места стимуляции до момента стимуляции ( t v < t s – t c ), он продолжится в мышце. Напротив, если антидромный стимулированный потенциал действия достигает нейрона до его возбуждения ( t s < t v – t c ), мы предполагаем, что это сбрасывает время возбуждения двигательного нейрона, поскольку сообщалось, что антидромные потенциалы действия устраняют ранее существовавшие постсинаптические потенциалы [3]. Сброс по существу задерживает следующий потенциал произвольного действия на величину, равную периоду произвольного возбуждения.
Чтобы проиллюстрировать влияние этих различных условий на общую скорость поступления потенциалов действия на двигательную концевую пластинку, мы рассчитали скорость, наблюдаемую в мышце, как функцию периода стимуляции (1/частота). Эта взаимосвязь между периодом возбуждения мышц и периодами возбуждения двух источников потенциала действия показана на рис. В этом примере произвольный период был зафиксирован на уровне 0,05 с (20 Гц), а период стимула варьировался в диапазоне периодов, как длиннее, так и короче, чем период произвольного возбуждения. Каждая точка представляет собой средний период возбуждения мышцы, рассчитанный за 3-секундный период постоянной активности.
В этом примере произвольный период был зафиксирован на уровне 0,05 с (20 Гц), а период стимула варьировался в диапазоне периодов, как длиннее, так и короче, чем период произвольного возбуждения. Каждая точка представляет собой средний период возбуждения мышцы, рассчитанный за 3-секундный период постоянной активности.
Открыть в отдельном окне
Среднее время возбуждения мышц как функция периода стимула. Скорость произвольной стрельбы имела средний период 0,05 с с коэффициентом вариации 0,1. Красный символ отмечает точку, в которой два периода включения будут равными.
Одновременное возбуждение уменьшает средний период возбуждения, т. е. увеличивает среднюю частоту, независимо от того, является ли период стимула выше или ниже произвольного периода. Либо уменьшение, либо увеличение периода стимула приводит к среднему уменьшению периода возбуждения мышц. Дисперсия скорости мышечных импульсов существенно уменьшается, когда период стимула меньше произвольного периода (0,05 с), из-за синхронной перезагрузки двигательных нейронов.
B. Влияние взаимодействия на мышечную силу и ЭМГ
Мы решили исследовать произвольную и стимулированную силу и производство ЭМГ в первой дорсальной межкостной мышце человека из-за наличия ранее разработанных данных и моделей произвольного рекрутирования и частотной модуляции, а также ЭМГ -силовые отношения. Мы выбрали «луковичную» модель управления произвольными двигательными единицами, опубликованную De Luca и Contessa [4], и модели мышечной силы и генерации ЭМГ, разработанные Fuglevand [5] и впоследствии адаптированные Zhou и Rymer [6].
Детали модели хорошо описаны в вышеупомянутых документах и не будут повторяться в этом реферате. Общая структура представляет собой набор из 120 двигательных единиц с константами времени сокращения от 0,09 до 0,03 с и пиковой силой сокращения от 1 до 100 (условные единицы силы) соответственно. Один скалярный ввод в пул модулирует скорость как рекрутирования, так и произвольных импульсов с полным рекрутированием всех двигательных единиц при 67% максимального возбуждения [4].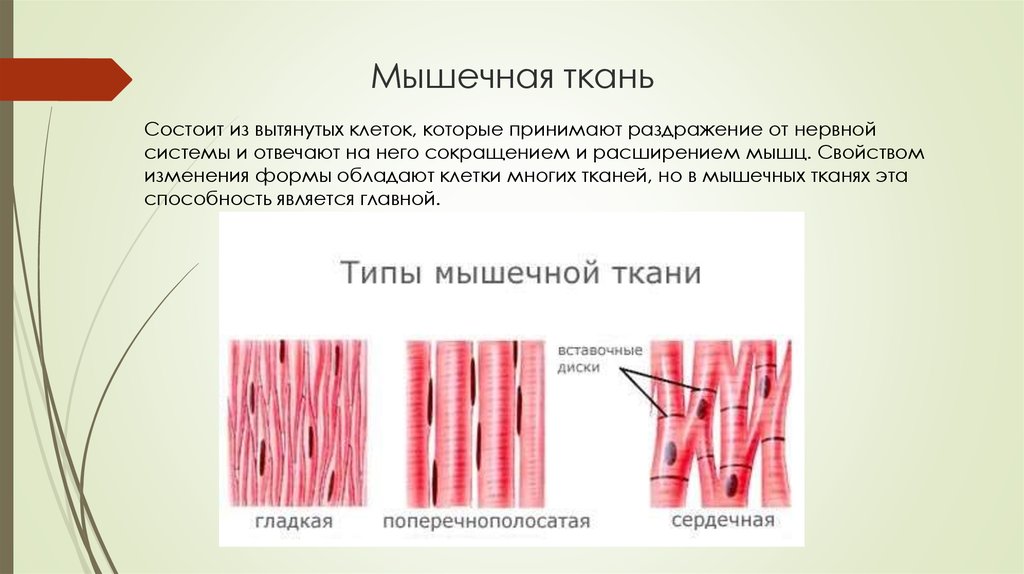 Коэффициент вариации периода произвольного срабатывания (1/частота) был зафиксирован на уровне 0,05 для описанных ниже силовых и ЭМГ-взаимодействий. Мы приняли время проведения 0,005 с и 0,001 с для t c и t p соответственно, поскольку они сравнимы с потенциалами действия, распространяющимися на 50 см и 10 см соответственно. Уровни произвольного и стимулированного сокращения были выбраны в процентах от максимальной произвольной силы, vFmax с шагом 10% в диапазоне от 0% до 50%. Мы модулировали силу стимуляции, варьируя количество задействованных двигательных единиц при фиксированной частоте стимула (20 или 35 Гц). Стимулируемые двигательные единицы были выбраны случайным образом с использованием равномерного распределения вероятностей в результатах, представленных здесь, отражающих экспериментальные исследования, которые подтверждают случайную схему рекрутирования с помощью электродов для поверхностной стимуляции [7] [1].
Коэффициент вариации периода произвольного срабатывания (1/частота) был зафиксирован на уровне 0,05 для описанных ниже силовых и ЭМГ-взаимодействий. Мы приняли время проведения 0,005 с и 0,001 с для t c и t p соответственно, поскольку они сравнимы с потенциалами действия, распространяющимися на 50 см и 10 см соответственно. Уровни произвольного и стимулированного сокращения были выбраны в процентах от максимальной произвольной силы, vFmax с шагом 10% в диапазоне от 0% до 50%. Мы модулировали силу стимуляции, варьируя количество задействованных двигательных единиц при фиксированной частоте стимула (20 или 35 Гц). Стимулируемые двигательные единицы были выбраны случайным образом с использованием равномерного распределения вероятностей в результатах, представленных здесь, отражающих экспериментальные исследования, которые подтверждают случайную схему рекрутирования с помощью электродов для поверхностной стимуляции [7] [1].
Мы смоделировали комбинированные произвольные и стимулированные сокращения, начиная с изолированного произвольного сокращения в течение первых 1,5 секунд, за которыми следовали произвольные сокращения с наложенной стимуляцией в течение последних 1,5 секунд каждой 3-секундной попытки, как показано на рис.
Открыть в отдельном окне
Симулированная сила (вверху), общая ЭМГ, включая М-зубцы (в центре), и произвольный компонент ЭМГ (внизу) с удаленными М-зубцами. Шаг произвольной активации в t = 0 с и шаг стимулированной активации в t = 1,5 с.
Силы отдельных двигательных единиц были рассчитаны путем суммирования импульсов силы, вызванных каждым потенциалом действия, нелинейно масштабированных по предшествующему интервалу потенциала действия. Общая сила рассчитывалась как линейная сумма всех активных сил двигательных единиц. ЭМГ отдельных двигательных единиц описывались импульсами второго порядка длительностью 0,012 с [6] и масштабировались (условные единицы) пропорционально силе сокращения двигательной единицы. Общая составная ЭМГ рассчитывалась как линейная сумма потенциалов действия отдельных двигательных единиц, независимо от того, были ли они генерированы произвольно или в результате стимуляции (М-волны очевидны после начала стимуляции в ). Поскольку источник каждого потенциала действия был известен, мы также рассчитывали М-волны отдельно и вычитали их из общей суммы, чтобы получить чистый произвольный компонент ЭМГ.
Поскольку источник каждого потенциала действия был известен, мы также рассчитывали М-волны отдельно и вычитали их из общей суммы, чтобы получить чистый произвольный компонент ЭМГ.
показывает частоту возбуждения и добавленную силу двигательных единиц во время произвольного сокращения 40% vFmax и наложенного 40% vFmax стимулированного (20 Гц) сокращения. В то время как средние частоты потенциала действия в мышце увеличиваются при стимуляции, приращение силы вследствие стимуляции ограничивается относительными значениями двух частот возбуждения и рабочей точки двигательной единицы на соотношении сила-частота. Двигательные единицы с низким порогом добровольного рекрутирования (низкое количество двигательных единиц) имеют низкую частоту слияния, и хотя стимуляция увеличивает скорость возбуждения почти на 50%, по существу нет дополнительной силы. Повышенная частота мышечных импульсов уменьшается с увеличением количества двигательных единиц до тех пор, пока произвольная частота не сравняется с частотой стимуляции, а затем остается неизменной на уровне частоты стимуляции. Для конкретного примера, показанного здесь, симуляция не производит заметной дополнительной силы до середины диапазона двигательных единиц, т. Е. Около половины стимулируемых двигательных единиц не создают никакой дополнительной силы из-за стимуляции. Это связано с двумя составляющими факторами: двигательные единицы с низким порогом уже активируются на частоте слияния или около нее, и эти двигательные единицы меньше, чем единицы с высоким порогом.
Для конкретного примера, показанного здесь, симуляция не производит заметной дополнительной силы до середины диапазона двигательных единиц, т. Е. Около половины стимулируемых двигательных единиц не создают никакой дополнительной силы из-за стимуляции. Это связано с двумя составляющими факторами: двигательные единицы с низким порогом уже активируются на частоте слияния или около нее, и эти двигательные единицы меньше, чем единицы с высоким порогом.
Открыть в отдельном окне
Слева — Частота возбуждения произвольно активированных и стимулированных двигательных единиц в зависимости от количества двигательных единиц, т. е. ранг произвольного включения. Это для той же симуляции, что и . Синие точки показывают среднюю частоту возбуждения двигательных нейронов, красные точки показывают частоту стимуляции, а черные символы + показывают среднюю частоту возбуждения мышечной единицы. Справа – увеличение силы двигательной единицы из-за стимуляции для каждой стимулируемой двигательной единицы. Символы «плюс» обозначают двигательные единицы, которые активируются как произвольно, так и при стимуляции.
Символы «плюс» обозначают двигательные единицы, которые активируются как произвольно, так и при стимуляции.
Были смоделированы все комбинации произвольных и стимулированных сокращений в диапазоне от 0% до 50% максимальной произвольной силы с шагом 10%. Произвольную силу рассчитывали как среднюю силу за одну секунду до начала стимуляции, а общую силу рассчитывали как среднюю за последнюю секунду наложенной стимуляции. Разница между этими двумя средними значениями была приращением за счет стимуляции. Моделирование повторялось шесть раз с другим набором случайно выбранных стимулируемых двигательных единиц, а результаты усреднялись.
Наложение стимуляции всегда увеличивало общую силу, но размер приращения уменьшался по мере увеличения произвольной силы (вверху). При фиксированном уровне стимуляции прирост силы, вызванный наложенной стимуляцией, уменьшался как прямолинейная функция произвольной силы с наклоном, пропорциональным произвольной силе. Таким образом, когда прирост стимулированной силы нормируется на стимулированную силу в отсутствие любого произвольного сокращения, зависимости от произвольной силы накладываются друг на друга. Зависимость от добровольной силы существенна. При частоте стимуляции 20 Гц способность стимуляции увеличивать произвольную силу снижается примерно на 40% при половинной максимальной произвольной силе.
Зависимость от добровольной силы существенна. При частоте стимуляции 20 Гц способность стимуляции увеличивать произвольную силу снижается примерно на 40% при половинной максимальной произвольной силе.
Открыть в отдельном окне.
Открыть в отдельном окне. Внизу — нормализованный произвольный компонент ЭМГ во время стимуляции как функция нормализованного произвольного ЭМГ в отсутствие стимуляции. ЭМГ количественно определялась как среднее абсолютное значение за те же 1-секундные периоды времени, что и сила.
Одновременная стимуляция также снижала произвольный компонент ЭМГ, что видно после начала стимуляции. По мере увеличения уровня стимуляции с 20% до 50% vFmax наклон зависимости нормированной ЭМГ от уровня произвольных сокращений уменьшается, что видно в нижней части графика. При максимальных изученных уровнях (50% произвольной и 50% стимулированной силы) произвольная ЭМГ снижалась примерно на 1/3. Два механизма способствуют уменьшению произвольной ЭМГ: блок прямого столкновения некоторых потенциалов произвольного действия и фазовый сброс двигательных нейронов, который непосредственно снижает частоту потенциалов произвольного действия, поступающих в мышцу.
Произвольные и электрически стимулированные сокращения, как было показано, складываются нелинейно в экспериментах на здоровых людях, а также у лиц, перенесших инсульт [1, 2]. Это неудивительно, учитывая, что два источника возбуждения активируют некоторые из одних и тех же двигательных единиц, а силы отдельных двигательных единиц насыщаются при высоких скоростях стимула. Вклад текущего отчета включает модель взаимодействия двух последовательностей потенциалов действия, количественную демонстрацию одного потенциального механизма для наблюдаемого нелинейного сложения и предсказание уменьшения амплитуды произвольной ЭМГ, о которой не сообщалось. ранее.
Уменьшение прироста стимулированной силы по мере увеличения произвольной силы означает, что существует убывающая польза от добавления стимуляции для увеличения силы в инсультном нейропротезе. Поскольку увеличение произвольной силы обычно приводит к нежелательным коактивационным синергиям [8, 9], разумным подходом было бы ограничение произвольных сокращений и увеличение мышечной силы путем выборочной стимуляции целевых мышц [10]. Стимуляцию можно контролировать с помощью ЭМГ, регистрируемой при произвольных сокращениях низкого уровня после удаления М-зубцов [11]. Наш прогноз снижения произвольной ЭМГ при стимуляции скромен, особенно при низких уровнях произвольной силы, что подтверждает осуществимость этого подхода. Мы не нашли убедительных прямых доказательств сброса фаз двигательных нейронов антидромными потенциалами действия. Если сброса фазы не происходит, снижение произвольной ЭМГ не будет таким драматичным, как здесь.
Стимуляцию можно контролировать с помощью ЭМГ, регистрируемой при произвольных сокращениях низкого уровня после удаления М-зубцов [11]. Наш прогноз снижения произвольной ЭМГ при стимуляции скромен, особенно при низких уровнях произвольной силы, что подтверждает осуществимость этого подхода. Мы не нашли убедительных прямых доказательств сброса фаз двигательных нейронов антидромными потенциалами действия. Если сброса фазы не происходит, снижение произвольной ЭМГ не будет таким драматичным, как здесь.
Сила модели заключается в ее способности количественно оценивать чувствительность результатов к допущениям моделирования, таким как рекрутирование пула двигательных нейронов и модели модуляции частоты, а также в качестве инструмента при разработке нейрореабилитационных вмешательств. Например, в настоящее время мы изучаем выбор схем и параметров стимуляции, в частности порядка рекрутирования и частоты стимуляции, при увеличении силы у выживших после инсульта с умеренными и тяжелыми нарушениями.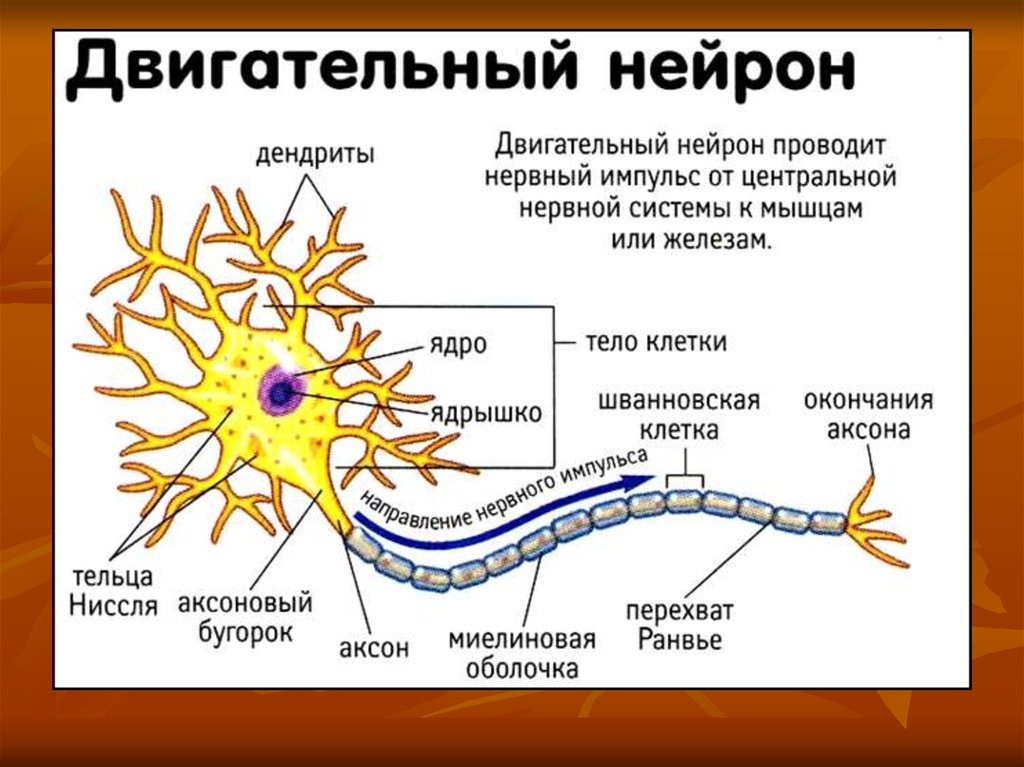
Мы с благодарностью признаем вклад Джули Мерфи и Дэнни Янга, которые осуществили динамическое моделирование произвольных и стимулированных потенциалов действия в одном аксоне, что помогло сформулировать наш подход. Мы также благодарим за полезные обсуждения с Эндрю Фуглевандом, Зев Раймер, Карло Де Лука и Паолой Контесса во время разработки и внедрения модели.
* Исследования частично поддерживаются Национальным институтом здравоохранения США, Национальным институтом детского здоровья и развития в рамках гранта R21HD05256 и гранта 11PRE6600000 Американской кардиологической ассоциации.
[1] Перумал Р., Векслер А.С., Кесар ТМ, Янкоско А., Лауфер Ю., Биндер-Маклауд С.А. Феноменологическая модель, предсказывающая силы, возникающие при наложении электрической стимуляции на субмаксимальные произвольные сокращения. Журнал прикладной физиологии. 2010 июнь; 108: 1595–604. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[2] Makowski NS, Knutson J, Chae J, Crago PE.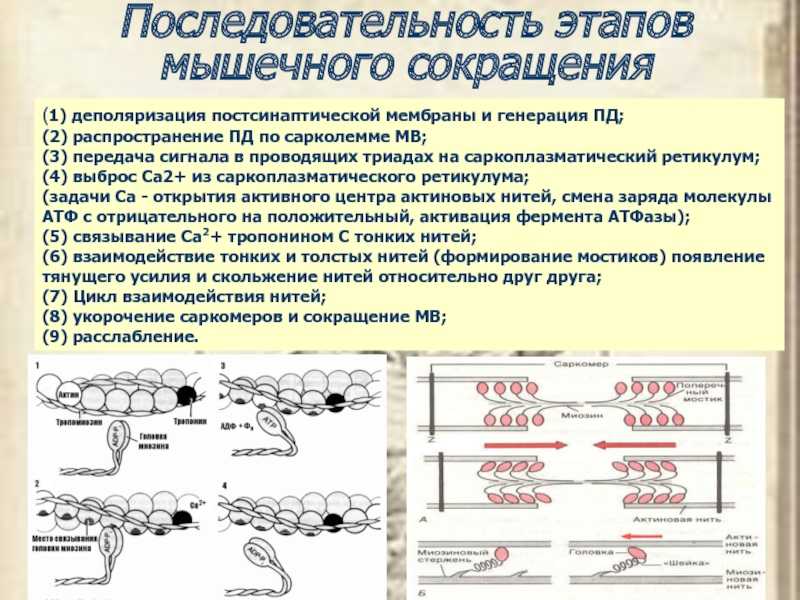 Взаимодействие ФЭС с произвольным усилием при хроническом инсульте; 39-я конференция по нейронным интерфейсам; Лонг-Бич, Калифорния. 2010.с. 62. [Google Академия]
Взаимодействие ФЭС с произвольным усилием при хроническом инсульте; 39-я конференция по нейронным интерфейсам; Лонг-Бич, Калифорния. 2010.с. 62. [Google Академия]
[3] Брок Л.Г., Кумбс Дж.С., Экклс Дж.К. Внутриклеточная запись с антидромно активированных мотонейронов. Журнал физиологии. 1953 г., 29 декабря; 122: 429–61. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[4] De Luca CJ, Contessa P. Иерархический контроль моторных единиц при произвольных сокращениях. J Нейрофизиол. 2012 1 января; 107: 178–195. 2012. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[5] Fuglevand AJ, Winter DA, Patla AE. Модели организации рекрутинга и кодирования скорости в моторно-единичных пулах. J Нейрофизиол. 1993;70:2470–2488. [PubMed] [Google Scholar]
[6] Zhou P, Suresh NL, Rymer WZ. Модельный анализ чувствительности отношения ЭМГ-силы к свойствам двигательных единиц: приложения к мышечному парезу при инсульте. Энн Биомед Инж. 2007 г., сен; 35: 1521–31. [PubMed] [Google Scholar]
[7] Кнафлиц М. , Мерлетти Р., Де Лука С.Дж. Вывод порядка рекрутирования двигательных единиц при произвольных и электрически вызванных сокращениях. Журнал прикладной физиологии. 1990 1 апреля; 68: 1657–1667. 1990. [PubMed] [Google Scholar]
, Мерлетти Р., Де Лука С.Дж. Вывод порядка рекрутирования двигательных единиц при произвольных и электрически вызванных сокращениях. Журнал прикладной физиологии. 1990 1 апреля; 68: 1657–1667. 1990. [PubMed] [Google Scholar]
[8] Dewald JP, Beer RF. Аномальный крутящий момент в паретичной верхней конечности у пациентов с гемипарезом. Мышечный нерв. 2001 февраль; 24: 273–83. [PubMed] [Google Scholar]
[9] Miller LC, Dewald JPA. Силы непроизвольного паретического сгибания запястья/пальцев и ЭМГ увеличиваются при нагрузке отведения плеча у лиц с хроническим инсультом. Клин Нейрофизиол. 2012 [Бесплатная статья PMC] [PubMed] [Google Scholar]
[10] Makowski NS, Knutson J, Chae J, Crago PE. Нервно-мышечная электрическая стимуляция для увеличения досягаемости и раскрытия рук после инсульта; 33-я ежегодная международная конференция IEEE EMBS; Бостон, Массачусетс. 2011. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[11] Yeom H, Chang y.-H. Аутогенная функциональная электрическая стимуляция под контролем ЭМГ для контроля тыльного сгибания голеностопного сустава. J Neurosci Методы. 2010; 193:118–125. [Бесплатная статья PMC] [PubMed] [Google Scholar]
J Neurosci Методы. 2010; 193:118–125. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Подергивание мышц и контроль | Биология для основных специальностей II
Результаты обучения
- Обсуждение мышечного напряжения и сокращения
Muscle Twitch
Мы можем улучшить наше понимание мышечного сокращения, исследуя сокращение одного мышечного волокна. Подергивание возникает, когда одно мышечное волокно сокращается в ответ на команду (стимул) нервной системы. Время между активацией мотонейрона и началом мышечного сокращения называется лаг-фазой (иногда называемой латентной фазой). Во время лаг-фазы сигнал, называемый потенциалом действия, перемещается к концу моторного нейрона (терминалу аксона). Это приводит к высвобождению ацетилхолина и деполяризации моторной концевой пластинки. Деполяризация приводит к высвобождению кальция саркоплазматическим ретикулумом и последующему связыванию кальция с тропонином, что приводит к обнажению сайта связывания миозина.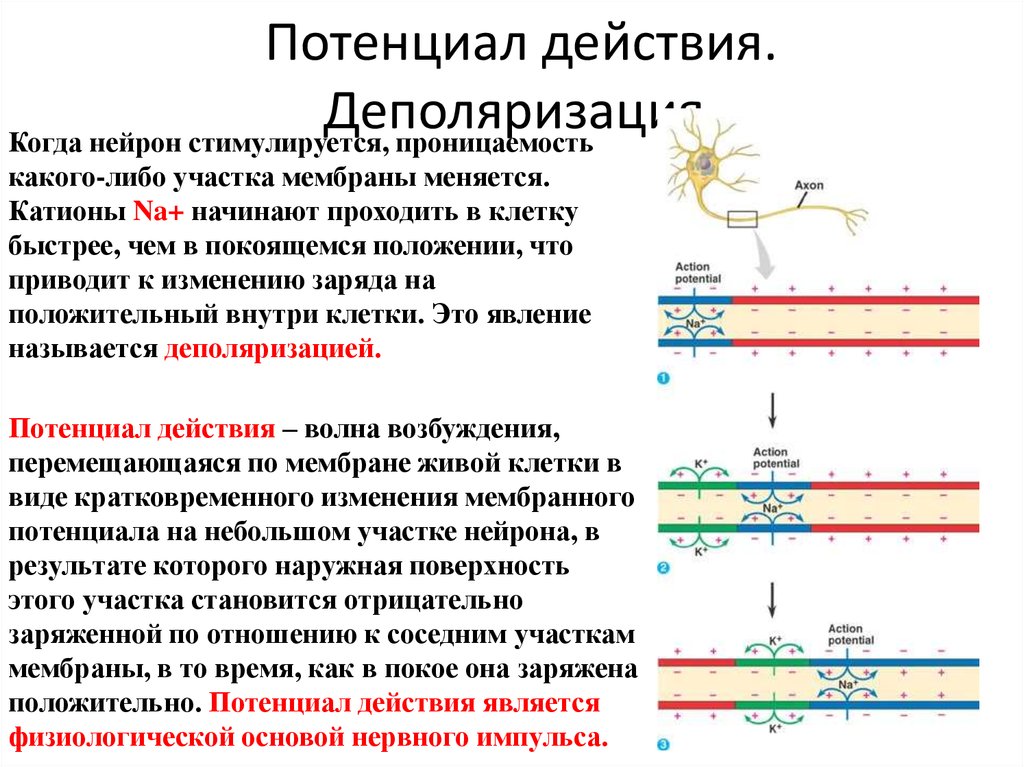 За этим следует фактическое мышечное сокращение, которое вызывает напряжение в мышцах. Эта следующая фаза называется фазой сокращения. Во время фазы сокращения образуются поперечные мостики между актином и миозином. Миозин перемещает актин, высвобождает и восстанавливает поперечные мостики много раз по мере того, как саркомер укорачивается и мышца сокращается. На этом этапе используется АТФ, и энергия выделяется в виде тепла. Миозин высвобождается из актина, когда к миозину присоединяется вторая АТФ. Миозин теперь доступен для формирования другого поперечного мостика. Когда мышца расслабляется, напряжение уменьшается. Эта фаза называется фазой релаксации. Во время этой фазы кальций активно транспортируется обратно в саркоплазматический ретикулум с помощью АТФ. Тропонин возвращается в положение, блокируя участок связывания миозина с актином, и мышца пассивно удлиняется.
За этим следует фактическое мышечное сокращение, которое вызывает напряжение в мышцах. Эта следующая фаза называется фазой сокращения. Во время фазы сокращения образуются поперечные мостики между актином и миозином. Миозин перемещает актин, высвобождает и восстанавливает поперечные мостики много раз по мере того, как саркомер укорачивается и мышца сокращается. На этом этапе используется АТФ, и энергия выделяется в виде тепла. Миозин высвобождается из актина, когда к миозину присоединяется вторая АТФ. Миозин теперь доступен для формирования другого поперечного мостика. Когда мышца расслабляется, напряжение уменьшается. Эта фаза называется фазой релаксации. Во время этой фазы кальций активно транспортируется обратно в саркоплазматический ретикулум с помощью АТФ. Тропонин возвращается в положение, блокируя участок связывания миозина с актином, и мышца пассивно удлиняется.
Мышечный стимул и сила сокращения
Скелетные мышечные волокна производят заданную силу, если стимул достаточно силен, чтобы достичь порога мышечного сокращения. Это называется законом «все или ничего». Допустим, мы электрически стимулируем мышечное волокно. Мы начинаем с небольшого количества стимуляции, которое не достигает порога, вызывающего сокращение. Мышечное волокно ответит тем, что останется расслабленным, оно не будет сокращаться. Теперь, если мы увеличим стимуляцию так, чтобы ее было достаточно для достижения порога, мышечное волокно ответит сокращением. Наконец, если мы продолжим усиливать раздражитель настолько, что он значительно превысит пороговое значение, волокно ответит сокращением с той же силой, что и в момент, когда мы только что достигли раздражителя. Мышца не будет сокращаться с большей силой, если стимул сильнее. Мышца реагирует на более сильные раздражители, производя ту же силу. В скелетных мышцах двигательный нейрон может иннервировать многие мышечные волокна. Это называется двигательной единицей. В скелетных мышцах имеется множество двигательных единиц. Двигательные единицы действуют согласованно. Один стимул воздействует на все мышечные волокна, иннервируемые данной двигательной единицей.
Это называется законом «все или ничего». Допустим, мы электрически стимулируем мышечное волокно. Мы начинаем с небольшого количества стимуляции, которое не достигает порога, вызывающего сокращение. Мышечное волокно ответит тем, что останется расслабленным, оно не будет сокращаться. Теперь, если мы увеличим стимуляцию так, чтобы ее было достаточно для достижения порога, мышечное волокно ответит сокращением. Наконец, если мы продолжим усиливать раздражитель настолько, что он значительно превысит пороговое значение, волокно ответит сокращением с той же силой, что и в момент, когда мы только что достигли раздражителя. Мышца не будет сокращаться с большей силой, если стимул сильнее. Мышца реагирует на более сильные раздражители, производя ту же силу. В скелетных мышцах двигательный нейрон может иннервировать многие мышечные волокна. Это называется двигательной единицей. В скелетных мышцах имеется множество двигательных единиц. Двигательные единицы действуют согласованно. Один стимул воздействует на все мышечные волокна, иннервируемые данной двигательной единицей.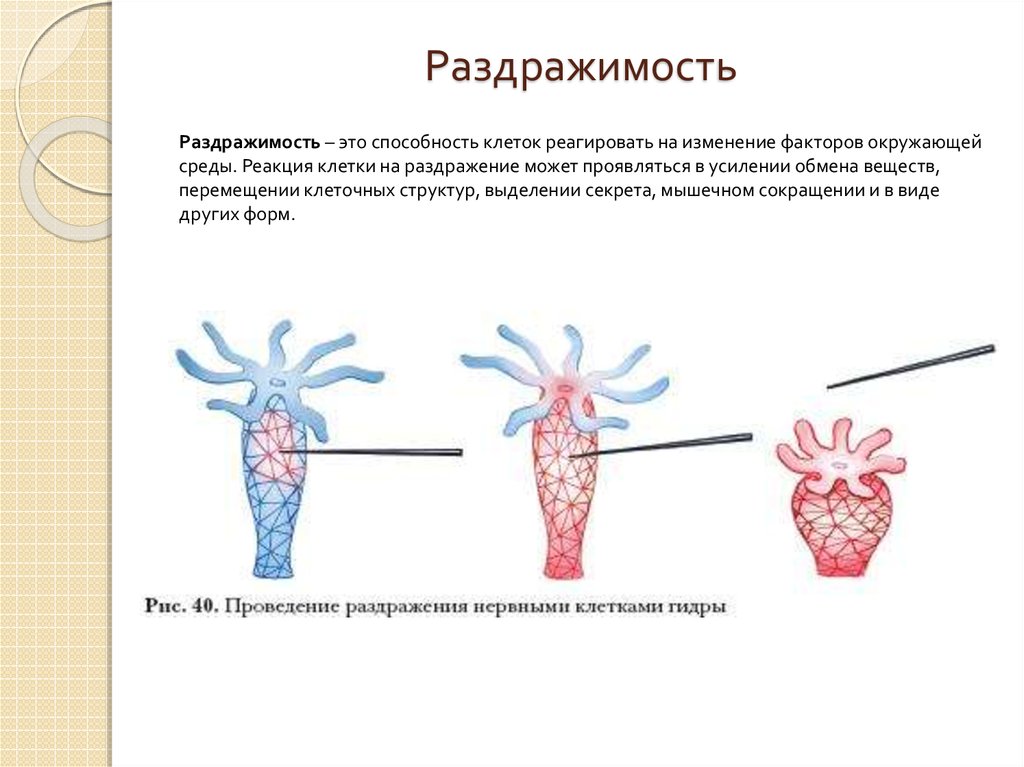
Зависимость длины мышцы от напряжения
Длина мышцы связана с напряжением, создаваемым мышцей. Мышцы будут генерировать больше силы, когда они растягиваются за пределы своей длины в состоянии покоя. Мышцы, растянутые за пределы этой точки, будут меньше напрягаться. Если мышца находится в состоянии покоя, она не будет производить максимального напряжения, потому что актиновые и миозиновые филаменты чрезмерно перекрываются. Нити миозина могут простираться в Z-диски, и обе нити мешают друг другу, ограничивая количество поперечных мостиков, которые могут образовываться. Если мышца растянута до точки, напряжение в мышце будет увеличиваться. Актиновые и миозиновые филаменты теперь могут оптимально перекрываться, так что может образоваться наибольшее количество поперечных мостиков. Если мышца перенапряжена, напряжение уменьшится. Нити актина и миозина не перекрываются, что приводит к уменьшению числа поперечных мостиков, которые могут образовываться. Идеальная длина саркомера при максимальном натяжении возникает, когда толстые и тонкие филаменты в наибольшей степени перекрываются.
Контроль мышечного напряжения
Нервный контроль инициирует образование актин-миозиновых поперечных мостиков, что приводит к укорочению саркомера, участвующему в мышечном сокращении. Эти сокращения распространяются от мышечного волокна через соединительную ткань, натягивая кости, вызывая движения скелета. Тяга, создаваемая мышцей, называется напряжением, и сила, создаваемая этим напряжением, может варьироваться. Это позволяет одним и тем же мышцам перемещать как очень легкие, так и очень тяжелые предметы. В отдельных мышечных волокнах величина производимого напряжения зависит от площади поперечного сечения мышечного волокна и частоты нервной стимуляции.
Количество поперечных мостиков, образованных между актином и миозином, определяет величину напряжения, которое может произвести мышечное волокно. Поперечные мостики могут образовываться только там, где толстые и тонкие филаменты перекрываются, позволяя миозину связываться с актином. Если образуется больше поперечных мостиков, больше миозина будет притягивать актин, и будет производиться большее напряжение.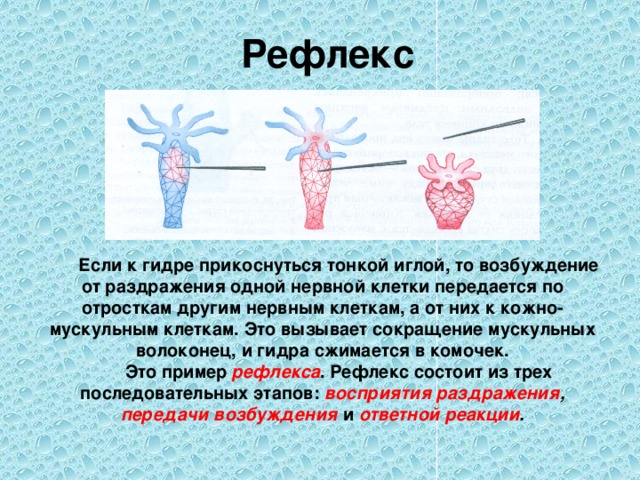
Идеальная длина саркомера при максимальном натяжении возникает, когда толстые и тонкие филаменты в наибольшей степени перекрываются. Если саркомер в состоянии покоя растянут за пределы идеальной длины покоя, толстые и тонкие филаменты не перекрываются в наибольшей степени, и может образовываться меньше поперечных мостиков. Это приводит к тому, что меньше миозиновых головок тянут актин, и возникает меньшее напряжение. Когда саркомер укорачивается, зона перекрытия уменьшается, поскольку тонкие филаменты достигают Н-зоны, состоящей из миозиновых хвостов. Поскольку именно головки миозина образуют поперечные мостики, актин не будет связываться с миозином в этой зоне, уменьшая натяжение, создаваемое этим миоволокном. Если саркомер укорачивается еще больше, тонкие филаменты начинают перекрываться друг с другом, еще больше уменьшая образование поперечных мостиков и вызывая еще меньшее натяжение. И наоборот, если саркомер растянут до такой степени, что толстые и тонкие нити совсем не перекрываются, поперечные мостики не образуются и натяжение не возникает. Такое растяжение обычно не происходит, потому что дополнительные белки, внутренние чувствительные нервы и соединительная ткань препятствуют чрезмерному растяжению.
Такое растяжение обычно не происходит, потому что дополнительные белки, внутренние чувствительные нервы и соединительная ткань препятствуют чрезмерному растяжению.
Первичной переменной, определяющей производство силы, является количество мышечных волокон в мышце, которые получают потенциал действия от нейрона, который контролирует это волокно. При использовании бицепса, чтобы взять карандаш, моторная кора головного мозга подает сигналы только нескольким нейронам бицепса, и только несколько миофибрилл реагируют. У позвоночных каждое мышечное волокно полностью отвечает на стимуляцию. Когда вы берете пианино, моторная кора подает сигнал всем нейронам бицепса, и в этом участвует каждое мышечное волокно. Это близко к максимальной силе, которую может произвести мышца. Как упоминалось выше, увеличение частоты потенциалов действия (количества сигналов в секунду) может еще немного увеличить силу, потому что тропомиозин заполнен кальцием.
Типы мышечных волокон
Существует три основных типа волокон скелетных мышц. Они называются быстрыми, медленными и промежуточными. Как правило, быстро сокращающиеся волокна генерируют большую силу в течение коротких промежутков времени. Медленно сокращающиеся волокна генерируют меньшую силу, но могут делать это в течение более длительных периодов времени. Промежуточные волокна обладают некоторыми характеристиками как быстрых, так и медленных волокон. Быстрые волокна также называют волокнами II типа. Быстрые волокна преобладают в организме. Они быстро реагируют на раздражители и могут генерировать большую силу. Они имеют большой диаметр за счет большого количества миофибрилл. Их активность подпитывается АТФ, образующимся в результате анаэробного метаболизма. Медленно сокращающиеся волокна реагируют на раздражители гораздо медленнее, чем быстрые волокна. Они меньше в диаметре и содержат большое количество митохондрий. Они способны поддерживать длительные сокращения и получать АТФ за счет аэробного метаболизма. Медленно сокращающиеся волокна окружены капиллярными сетями, которые снабжают кислородом кровь для использования в аэробных энергетических системах.
Они называются быстрыми, медленными и промежуточными. Как правило, быстро сокращающиеся волокна генерируют большую силу в течение коротких промежутков времени. Медленно сокращающиеся волокна генерируют меньшую силу, но могут делать это в течение более длительных периодов времени. Промежуточные волокна обладают некоторыми характеристиками как быстрых, так и медленных волокон. Быстрые волокна также называют волокнами II типа. Быстрые волокна преобладают в организме. Они быстро реагируют на раздражители и могут генерировать большую силу. Они имеют большой диаметр за счет большого количества миофибрилл. Их активность подпитывается АТФ, образующимся в результате анаэробного метаболизма. Медленно сокращающиеся волокна реагируют на раздражители гораздо медленнее, чем быстрые волокна. Они меньше в диаметре и содержат большое количество митохондрий. Они способны поддерживать длительные сокращения и получать АТФ за счет аэробного метаболизма. Медленно сокращающиеся волокна окружены капиллярными сетями, которые снабжают кислородом кровь для использования в аэробных энергетических системах. Они также содержат красный пигмент, называемый миоглобином. Миоглобин может связывать кислород (подобно гемоглобину) и обеспечивать значительный запас кислорода. Из-за красноватого цвета миоглобина эти волокна часто называют красными мышечными волокнами. Медленно сокращающиеся волокна также называют волокнами I типа. Промежуточные волокна напоминают быстро сокращающиеся волокна, потому что они содержат небольшое количество миоглобина. Они также имеют капиллярную сеть вокруг себя и не так быстро устают, как быстро сокращающиеся волокна. Они содержат больше митохондрий, чем быстросокращающиеся волокна, но не так много, как медленносокращающиеся волокна. Скорость сокращения и выносливость также находятся между быстрыми и медленными волокнами. Промежуточные волокна также называют волокнами типа IIa. Мышцы с преобладанием медленных волокон иногда называют красными мышцами, например, в области спины и ног. Точно так же мышцы, в которых преобладают быстрые волокна, называются белыми мышцами. Интересно отметить, что в глазных мышцах и мышцах рук нет медленно сокращающихся волокон.
Они также содержат красный пигмент, называемый миоглобином. Миоглобин может связывать кислород (подобно гемоглобину) и обеспечивать значительный запас кислорода. Из-за красноватого цвета миоглобина эти волокна часто называют красными мышечными волокнами. Медленно сокращающиеся волокна также называют волокнами I типа. Промежуточные волокна напоминают быстро сокращающиеся волокна, потому что они содержат небольшое количество миоглобина. Они также имеют капиллярную сеть вокруг себя и не так быстро устают, как быстро сокращающиеся волокна. Они содержат больше митохондрий, чем быстросокращающиеся волокна, но не так много, как медленносокращающиеся волокна. Скорость сокращения и выносливость также находятся между быстрыми и медленными волокнами. Промежуточные волокна также называют волокнами типа IIa. Мышцы с преобладанием медленных волокон иногда называют красными мышцами, например, в области спины и ног. Точно так же мышцы, в которых преобладают быстрые волокна, называются белыми мышцами. Интересно отметить, что в глазных мышцах и мышцах рук нет медленно сокращающихся волокон.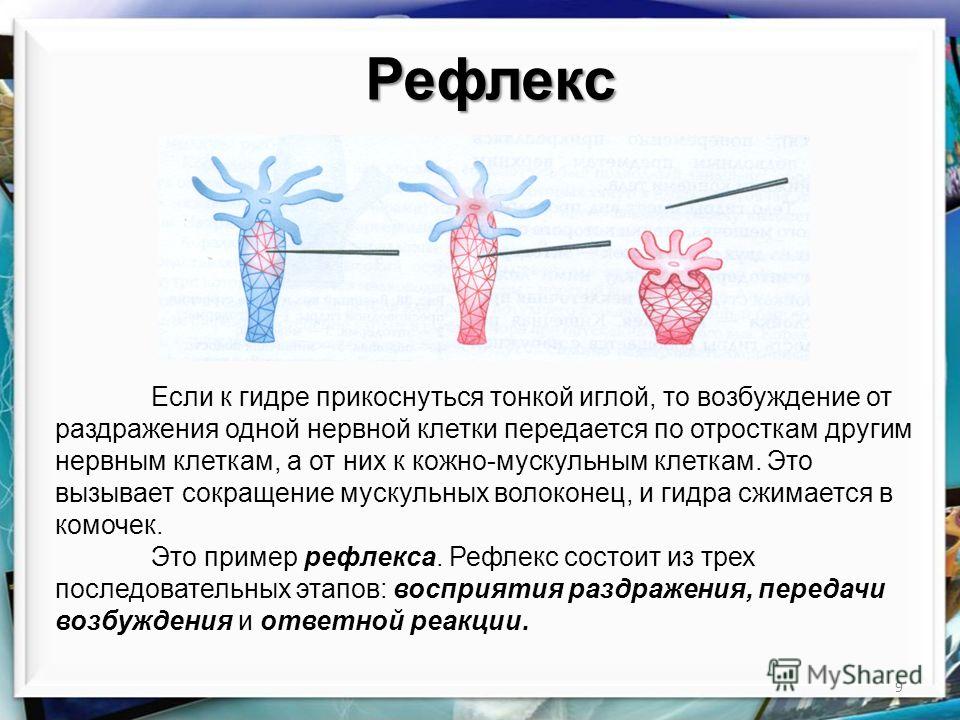
Leave A Comment