
комплементарність, епістаз, полімерія, плейотропія. Реакції між генами
Новоутворення є демонстрацією прояву взаємодії генів типу. Взаємодія неалельних генів: комплементарність, епістаз, полімерія, плейотропія. Реакції між генами
В окремих випадках дія різних генів відносно незалежна, але, як правило, прояв ознак є результатом взаємодії продуктів різних генів. Ці взаємодії можуть бути пов’язані як з алельними, так і з неалельнимигенами.
Взаємодія між алельнимигенами здійснюється у вигляді трьох форм: повне домінування, неповне домінування та незалежний прояв (кодомінування).
Раніше були розглянуті досліди Менделя, які виявили повне домінування одного алеля та рецесивність іншого. Неповне домінування спостерігається у тому випадку, коли один ген з пари алелей не забезпечує утворення в достатньому для нормального прояву ознаки його білкового продукту. При цій формі взаємодії генів усі гетерозиготи та гомозиготи значно відрізняються за фенотипом один від одного. При кодомініруванняу гетерозиготних організмів кожен з алельних генів викликає формування у фенотипі контрольованої їм ознаки. Прикладом цієї форми взаємодії алелів служить успадкування груп крові людини за системою АВО, детермінованих геном I. Існує три алелі цього гена Iо, Iа, IЬ, що визначають антигени груп крові. Спадкування груп крові ілюструє також явище
При кодомініруванняу гетерозиготних організмів кожен з алельних генів викликає формування у фенотипі контрольованої їм ознаки. Прикладом цієї форми взаємодії алелів служить успадкування груп крові людини за системою АВО, детермінованих геном I. Існує три алелі цього гена Iо, Iа, IЬ, що визначають антигени груп крові. Спадкування груп крові ілюструє також явище
Взаємодія неалельних генів. У ряді випадків на одну ознаку організму можуть впливати дві (або більше) пари неалельних генів. Це призводить до значних чисельних відхилень фенотипічних (але не генотипних) класів від встановлених Менделем при дигібридному схрещуванні. Взаємодія неалельних генів поділяють на основні форми: комплементарність, епістаз, полімерію.
При комплементарному
 Прикладом комплементарної взаємодії може бути схрещування двох різних сортів запашного горошку з білими пелюстками квіток.
Прикладом комплементарної взаємодії може бути схрещування двох різних сортів запашного горошку з білими пелюстками квіток.Наступним видом взаємодії неалельних генів є епістаз, при якому ген однієї алельної пари пригнічує дію гена іншої пари. Ген, який пригнічує дію іншого, називається епістатичним геном(або супресором).Пригнічений ген називається гіпостатичного.Епістаз може бути домінантним та рецесивним. Прикладом домінантного епістазу є спадкування забарвлення оперення курей. Ген С у домінантній формі визначає нормальну продукцію пігменту, але домінантний алель іншого гена I є його супресором. Внаслідок цього кури, що мають у генотипі домінантний алель гена забарвлення, у присутності супресора виявляються білими. Епістатична дія рецесивного гена ілюструє успадкування забарвлення шерсті у будинкових мишей. Забарвлення агуті (рудувато-сіре забарвлення шерсті) визначається домінантним геном А. Його рецесивний аллель а в гомозиготному стані зумовлює чорне забарвлення.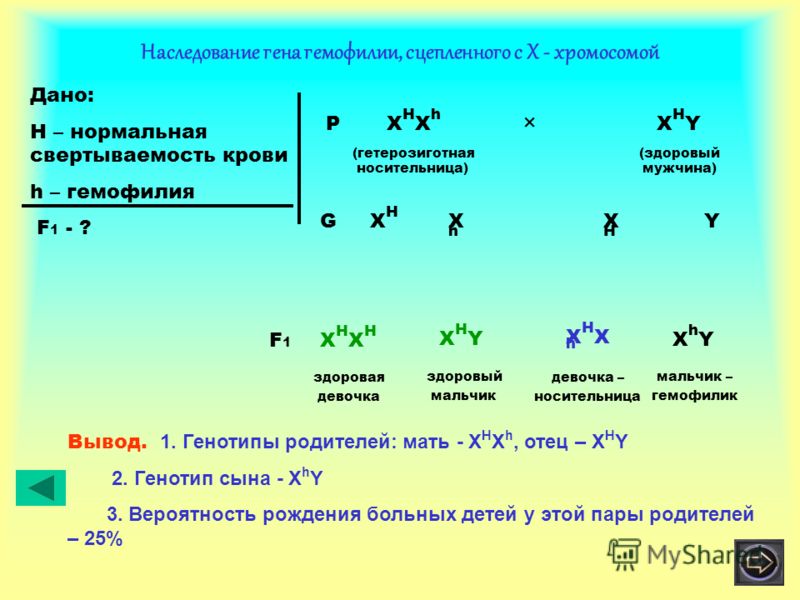
Спадкування ознаки, передача та розвиток якого, обумовлені, як правило, двома алелями одного гена, називають моногенним. Крім того відомі гени з різних алельних пар (їх називають полімерними або полігенами), приблизно однаково впливають ознака.
Явище одночасного на ознаку кількох неалельных однотипних генів одержало назву полімерії. Хоча полімерні гени є алельними, але оскільки визначають розвиток однієї ознаки, їх зазвичай позначають однією літерою А (а), цифрами вказуючи число алельних пар. Дія полігенів найчастіше буває підсумовуючим.
Зчеплене успадкування
Аналіз успадкування одночасно кількох ознак у дрозофіли, проведений Т. Морганом, показав, що результати аналізуючого схрещування гібридів F1 іноді відрізняються від очікуваних у разі їхнього незалежного успадкування. У нащадків такого схрещування замість вільного комбінування ознак різних пар спостерігали тенденцію до спадкування переважно батьківських поєднань ознак. Таке успадкування ознак було названо зчепленим.Зчеплене успадкування пояснюється розташуванням відповідних генів у тому ж хромосомі. У складі останньої вони передаються з покоління до покоління клітин та організмів, зберігаючи поєднання алелей батьків.
У нащадків такого схрещування замість вільного комбінування ознак різних пар спостерігали тенденцію до спадкування переважно батьківських поєднань ознак. Таке успадкування ознак було названо зчепленим.Зчеплене успадкування пояснюється розташуванням відповідних генів у тому ж хромосомі. У складі останньої вони передаються з покоління до покоління клітин та організмів, зберігаючи поєднання алелей батьків.
Залежність зчепленого успадкування ознак від локалізації генів в одній хромосомі дає підставу розглядати хромосоми як окремі групи зчеплення.Аналіз успадкування ознаки забарвлення очей у дрозофіли в лабораторії Т. Моргана виявив деякі особливості, що змусили виділити як окремий тип успадкування ознак зчеплене зі статтю успадкування.
Залежність результатів експерименту від того, хто з батьків був носієм домінантного варіанта ознаки, дозволила висловити припущення, що ген, що визначає забарвлення очей у дрозофіли, розташований у Х-хромосомі та не має гомолога в У-хромосомі. Всі особливості зчепленого зі статтю спадкування пояснюються неоднаковою дозою відповідних генів у представників різної — гомо-і гетерогаметної статі. Х-хромосома присутня в каріотипі кожної особини, тому ознаки, які визначають гени цієї хромосоми, формуються у представників як жіночої, так і чоловічої статі. Особини гомогаметної статі отримують ці гени від обох батьків і через свої гамети передають їх усім нащадкам. Представники гетерогаметної статі отримують єдину X-хромосому від гомогаметного батька та передають її своєму гомогаметному потомству. У ссавців (зокрема і людини) чоловіча стать отримує Х-зчеплені гени від матері і передає їх дочкам. При цьому чоловіча стать ніколи не успадковує батьківського Х-зчепленого ознаки і не передає його своїм синам
Всі особливості зчепленого зі статтю спадкування пояснюються неоднаковою дозою відповідних генів у представників різної — гомо-і гетерогаметної статі. Х-хромосома присутня в каріотипі кожної особини, тому ознаки, які визначають гени цієї хромосоми, формуються у представників як жіночої, так і чоловічої статі. Особини гомогаметної статі отримують ці гени від обох батьків і через свої гамети передають їх усім нащадкам. Представники гетерогаметної статі отримують єдину X-хромосому від гомогаметного батька та передають її своєму гомогаметному потомству. У ссавців (зокрема і людини) чоловіча стать отримує Х-зчеплені гени від матері і передає їх дочкам. При цьому чоловіча стать ніколи не успадковує батьківського Х-зчепленого ознаки і не передає його своїм синам
Активно функціонуючі гени У-хромосоми, що не мають алелів у Х-хромосомі, присутні в генотипі тільки гетерогаметної статі, причому в гемізиготному стані. Тому вони виявляються фенотипно і передаються з покоління до покоління лише у представників гетерогаметної статі. Так, у людини ознака гіпертрихозу вушної раковини (волосаті вуха) спостерігається виключно у чоловіків і успадковується від батька до сина.
Так, у людини ознака гіпертрихозу вушної раковини (волосаті вуха) спостерігається виключно у чоловіків і успадковується від батька до сина.
Цитоплазматичне успадкування
Наявність деякої кількості спадкового матеріалу в цитоплазмі у вигляді кільцевих молекул ДНК мітохондрій та пластид, а також інших позаядерних генетичних елементів дає підстави спеціально зупинитися на їхній участі у формуванні фенотипу в процесі індивідуального розвитку. Цитоплазматичні гени не підкоряються менделівським закономірностям спадкування, які визначаються поведінкою хромосом при мітозі, мейозі та заплідненні. У зв’язку з тим, що організм, що утворюється внаслідок запліднення, отримує цитоплазматичні структури головним чином з яйцеклітиною, цитоплазматичне успадкування ознак здійснюється по материнській лінії. Такий тип успадкування був вперше описаний у 1908 р. К. Корренсом щодо ознаки строкатого листя у деяких рослин.
Генетика людини з основами загальної генетики [Навчальний посібник] Курчанов Микола Анатолійович
4. 3. Взаємодія генів
3. Взаємодія генів
В організмі одночасно функціонує безліч генів. У процесах реалізації генетичної інформації на ознаку можливі численні «пункти» взаємодії різних генів лише на рівні біохімічних реакцій. Такі взаємодії неминуче відбиваються на формуванні фенотипу.
Алельні гени визначають альтернативні ознаки, тому що лежать у гомологічних локусах гомологічних хромосом. Між алелями гетерозиготи можливі певні взаємодії, що лежать в основі прояву ознаки фенотипу. Відомі три основні форми міжалельних взаємодій.
Повне домінування – у гетерозиготі один аллель (домінантний) пригнічує прояв другого алелю (рецесивного).
Неповне домінування – у гетерозиготі спостерігається проміжний прояв ознак алелей.
Кодомінування – незалежний прояв алелей у гетерозиготі.
У деяких випадках механізм взаємодії алелів розшифровано. Найкраще їх взаємини ілюструють білки-ферменти. Дані біохімічної генетики показують, що домінування часто пов’язані з активністю певного ферменту.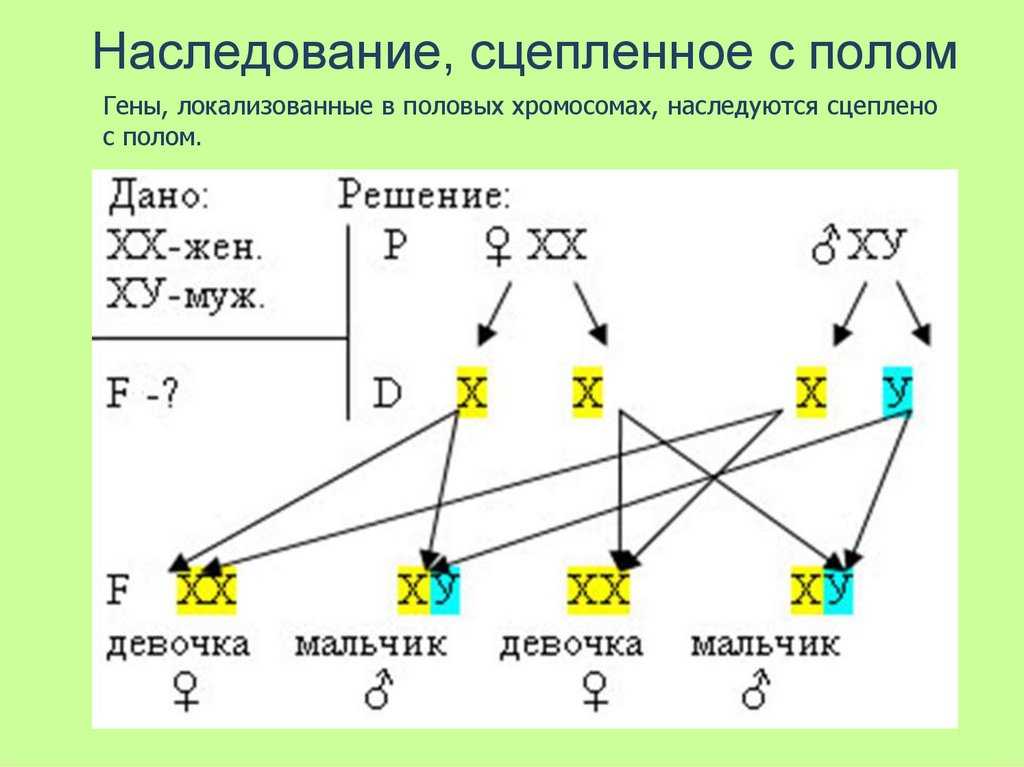
Менш зрозуміле явище моногенного гетерозису, як у гетерозиготі ознака проявляється сильніше, ніж у гомозиготі по домінантному алелю.
Феномен моногенного гетерозису, коли показано залежність ознаки від однієї алельної пари, є лише одним аспектом теорії гетерозису. За пропозицією Ф. Добжанського це явище отримало назву
 Воно має велике значення для еволюційної теорії, оскільки демонструє перевагу гетерозиготу у популяціях. Однак це поняття швидше застосовується до адаптивного рівня, тому до з’ясування генетичних механізмів наддомінування розглядати його як особливий вид міжалельних взаємодій передчасно.
Воно має велике значення для еволюційної теорії, оскільки демонструє перевагу гетерозиготу у популяціях. Однак це поняття швидше застосовується до адаптивного рівня, тому до з’ясування генетичних механізмів наддомінування розглядати його як особливий вид міжалельних взаємодій передчасно.У живих організмах часто взаємодіють не тільки алелі одного гена, але й алелі різних генів, даючи різні варіанти розщеплення. Розрізняють три основні типи взаємодії неалельних генів.
Комплементарність – взаємодія різних домінантних алелей зумовлює появу нової ознаки.
За типом комплементарності зазвичай взаємодіють гени, що контролюють різні етапи одного й того ж метаболічного шляху. Для деяких морфологічних ознак біохімічний механізм реалізації невідомий.
Епістаз – один ген пригнічує прояв іншого, неалельного йому гена.
Гени, що пригнічують дію інших генів, називаються епістатичними (або генами-супресорами). Можливі два варіанти епістазу: домінантний епістаз– епістатичний ген є домінантним у своїй алельній парі та рецесивний епістаз– епістатичний ген є рецесивним у своїй алельній парі.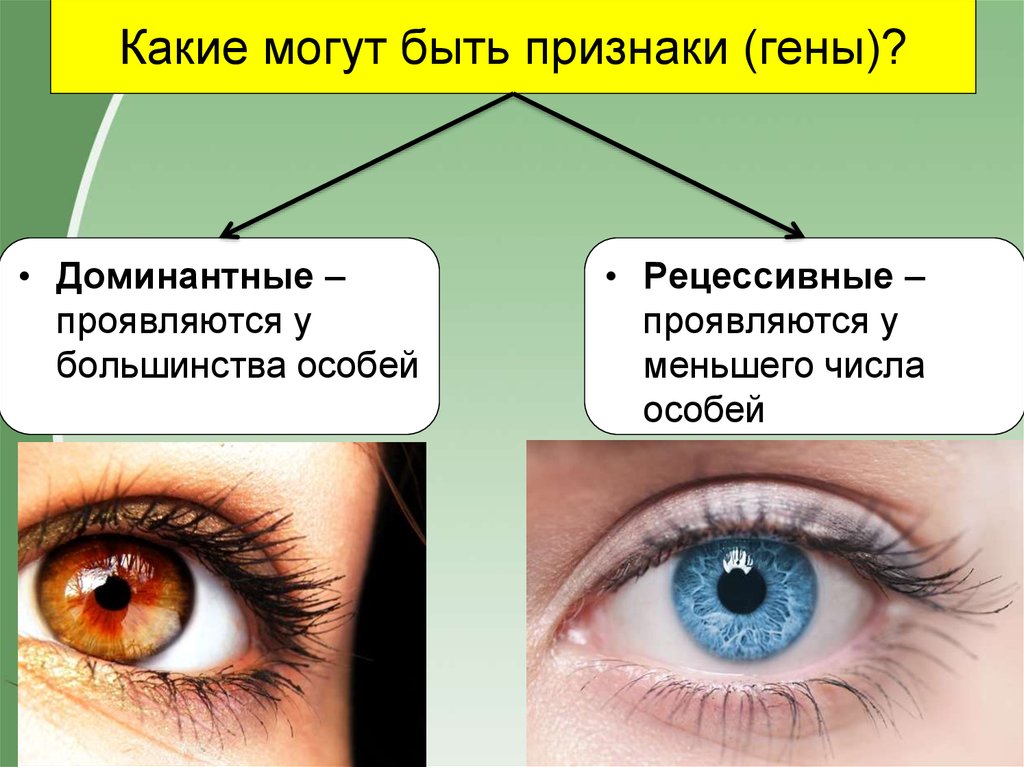
Полімерія – однозначна дія неалельних генів. Полімерія пов’язана з контролем ознаки декількома неалельних генів. Полігенний контроль дуже поширений у генетиці. Полімерні гени зазвичай позначаються однаковими літерами з нижнім індексом — А1, А2, А3 і т.д.
Полімерія також зустрічається у двох варіантах. При кумулятивної полімеріїінтенсивність ознаки пропорційна числу домінантних алелей серед полімерних генів, а при некумулятивної полімеріїрізні полімерні гени дублюють один одного і для прояву ознаки достатньо однієї з домінантних алелей.
Численні випадки взаємодії генів заповнюють основний обсяг всіх задачників з генетики. У типовому випадку при схрещуванні дигетерозигот при взаємодії генів утворюються різні відносини фенотипічних класів у поколіннях — 9: 3: 4; 9: 7; 13: 3; 12: 3: 1; 15: 1 та інші. Генетичний аналіз показує, що вони є видозміною класичної менделевской формули дигібридного розщеплення 9: 3: 3: 1. Вирішення великої кількості завдань з генетики є необхідним етапом у підготовці студентів, які вивчають генетику.
Словосполучення «взаємодія генів» дещо умовне, оскільки зазвичай взаємодіють не самі гени, які продукти. Проте не можна погодитись з терміном «взаємодія фенів», який неточно відображає сенс явища. На мій погляд, у навчальній літературі краще залишити традиційний термін «взаємодія генів» (алельних та неалельних).
Цей текст є ознайомлювальним фрагментом.З книги Генетика забарвлень собак автора Робінсон Рой
Взаємодія локусів А та Е Важливість цих двох серій алелей буде показана у цьому розділі. Обидві серії контролюють розподіл чорного та жовтого пігментів по шерсті у більшості порід собак. У взаємодії ці локуси утворюють безліч добре відомих
З книги Мікробіологія: конспект лекцій автора Ткаченко Ксенія Вікторівна
2. Взаємодія вірусів з клітиною господаря Взаємодія йде в єдиній біологічній системі на генетичному рівні. Існує чотири типи взаємодії: 1) продуктивна вірусна інфекція (взаємодія, в результаті якої відбувається репродукція вірусу, а
З книги Мураха, сім’я, колонія автора Захаров Анатолій Олександрович
Сім’я мурах — об’єднання сотень, тисяч, а іноді й мільйонів індивідів, зусиллями яких споруджується гніздо, вирощується численне потомство, охороняється мурашник та його кормова ділянка від посягань агресивних.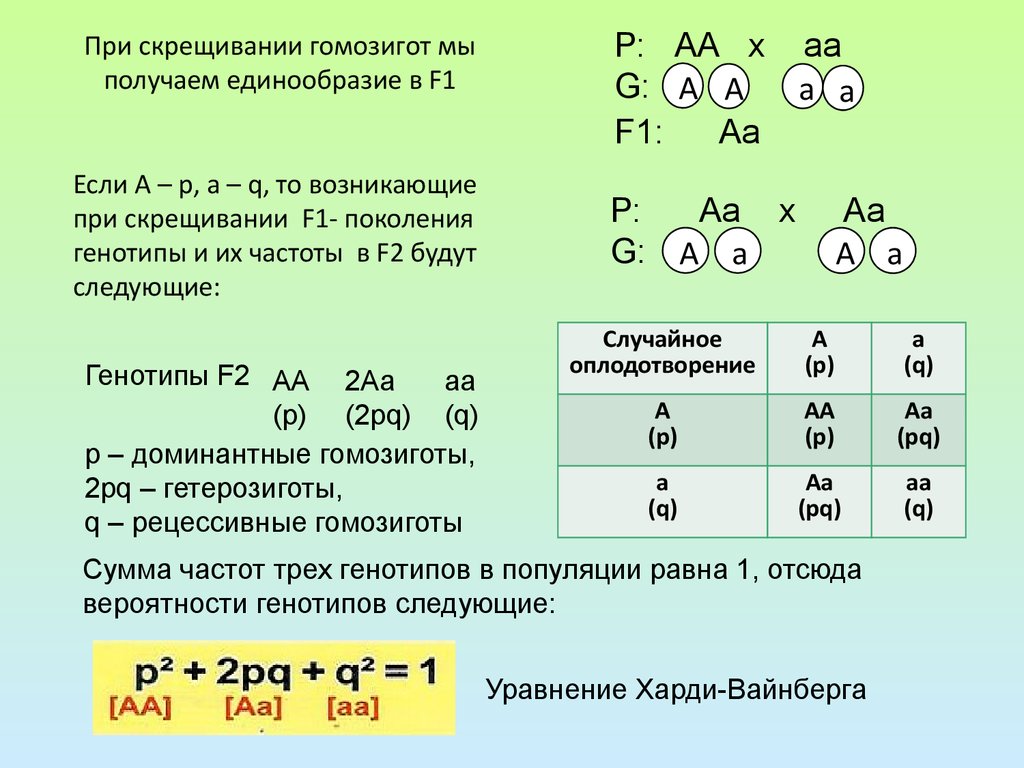
З книги Еволюція автора Дженкінс Мортон
Дрейф генів Іноді ця концепція називається «ефект Сьюелла — Райта», на честь двох популяційних генетиків, що її запропонували. Після того як Мендель довів, що гени є одиницями спадковості, а Харді та Вайнберг продемонстрували механізм їхньої поведінки,
З книги Рід людський автора Барнетт Ентоні
Взаємодія спадковості та середовища Іноді запитують: що важливіше – спадковість чи навколишнє середовище? На це питання не так просто відповісти. Якщо під цим мати на увазі, чт? має найбільшу силу впливу, і тоді слід обмежитися окремими випадками.
З книги Основи психофізіології автора Олександров Юрій
7. ВЗАЄМОДІЯ СЕНСОРНИХ СИСТЕМ Взаємодія сенсорних систем здійснюється на спинальному, ретикулярному, таламічному та кірковому рівні. Особливо широка інтеграція сигналів у ретикулярній формації. У корі мозку відбувається інтеграція сигналів найвищого порядку.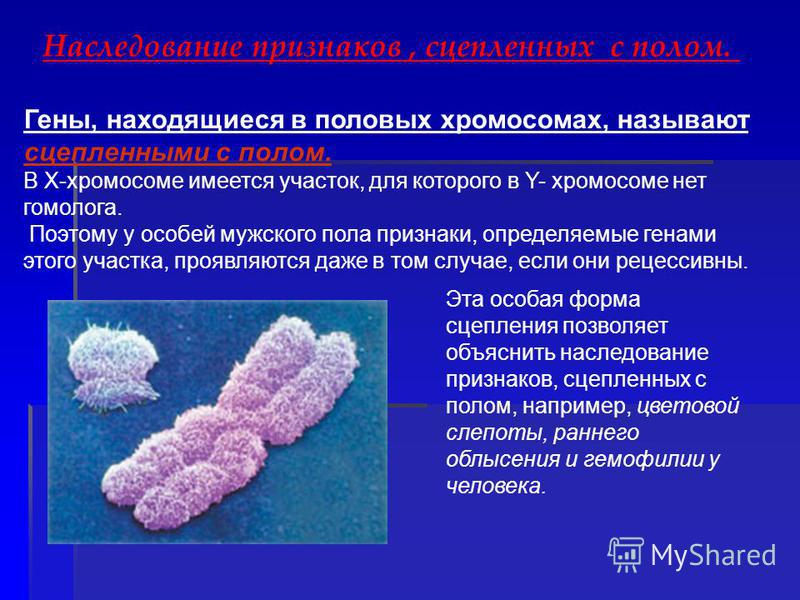 В
В
З книги Ембріони, гени та еволюція автора Рефф Рудольф А
В наш час відвідувачі музеїв сприймають виставлені в них змонтовані скелети величезних вимерлих тварин як щось само собою зрозуміле. Але так було не завжди. Наукові відомості про такі імпозантні форми, як мамонти та
З книги Теорія адекватного харчування та трофологія [таблиці текстом] автора
З книги Стій, хто веде? [Біологія поведінки людини та інших звірів] автора Жуків. Дмитро Анатолійович
Як вже було згадано, окремі ендокринні залози та їх гормони взаємодіють один з одним, забезпечуючи функціональну єдність організму. між
З книги Теорія адекватного харчування та трофологія [таблиці картинками] автора Уголєв Олександр Михайлович
9.7. Заключні зауваження (взаємодія у біоценозах) Розглянемо біохімічні взаємодії живих організмів у природних умовах, які поширені надзвичайно широко та стали об’єктом спеціальної науки – алелепатії (див. гл. 1). Приклади таких
гл. 1). Приклади таких
З книги Фенетика [Еволюція, популяція, ознака] автора Яблуків Олексій Володимирович
Взаємодія еволюційних сил — механізм еволюції Пусковий механізм еволюції функціонує внаслідок спільної дії еволюційних факторів у межах популяції як еволюційної одиниці. Будь-яка популяція у будь-якого виду схильна до того чи іншого тиску
З книги Еволюція [Класичні ідеї у світлі нових відкриттів] автора
Думка про те, що дуплікація генів служить найважливішим джерелом еволюційних нововведень, була висловлена ще в 1930-і роки видатним біологом Джоном Холдейном. Сьогодні в цьому немає жодних
З книги Гени та розвиток організму автора Нейфах Олександр Олександрович
1. Промотори генів У цьому розділі ми коротко розповімо у тому, які нуклеотидні послідовності, прилеглі до генів, котрий іноді всередині гена, відповідальні за процес транскрипції.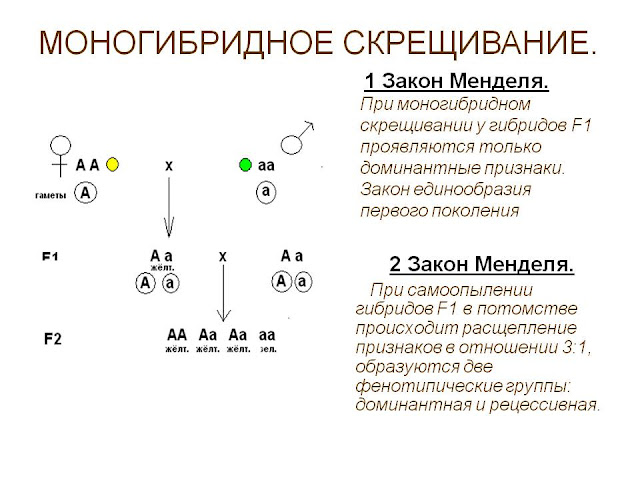 У прокаріотів ці ділянки, з якими зв’язується молекула РНК-полімерази та звідки
У прокаріотів ці ділянки, з якими зв’язується молекула РНК-полімерази та звідки
З книги Еволюція людини. Книга 1. Мавпи, кістки та гени автора Марков Олександр Володимирович
Зміни активності генів Еволюція тварин загалом і приматів зокрема протікає й не так з допомогою зміни структури білок-кодирующих генів, як з допомогою зміни їх активності. Невелика зміна у верхніх поверхах ієрархічно організованих
З книги Генетика людини з основами загальної генетики [Навчальний посібник] автора Курчанов Микола Анатолійович
4.4. Взаємодія генотипу та середовища Природа прояву дії генів набагато складніша, ніж у описаних вище варіантах. Розглядаючи дію генів та їх алелей, необхідно враховувати вплив довкілля на прояв ознак, і навіть модифицирующее дію інших
З книги автора
7.1. Виділення генів Можливе використання кількох шляхів виділення генів. Кожен з них має свої переваги та недоліки. Хімічний синтез генів, тобто синтез нуклеотидів із заданою послідовністю, що відповідає одному гену, вперше був здійснений у
Кожен з них має свої переваги та недоліки. Хімічний синтез генів, тобто синтез нуклеотидів із заданою послідовністю, що відповідає одному гену, вперше був здійснений у
Це гени, розташовані в однакових місцях (локусах) гомологічних хромосом, які відповідають за розвиток альтернативних ознак. Взаємодія алельних генів відбувається лише у гетерозиготному стані (Аа).
Варіанти взаємодії алельних генів:
а) повне домінування,
б) неповне домінування,
в) кодомінування,
г) наддомінування,
д) плейотропна дія гена.
1 . Повне домінування. Виявляється у тих випадках, коли один аллель
гена (домінантний) повністю приховує присутність іншого (рецесивного)
алеля. Наприклад:
А — карі очі
а – блакитні очі
Людина з генотипом Аа має карі очі.
2. Неповне домінування. При неповному домінуванні фенотип
гібридів першого покоління (Аа) зовні відрізняються від батьківських особин
(АА) та (аа).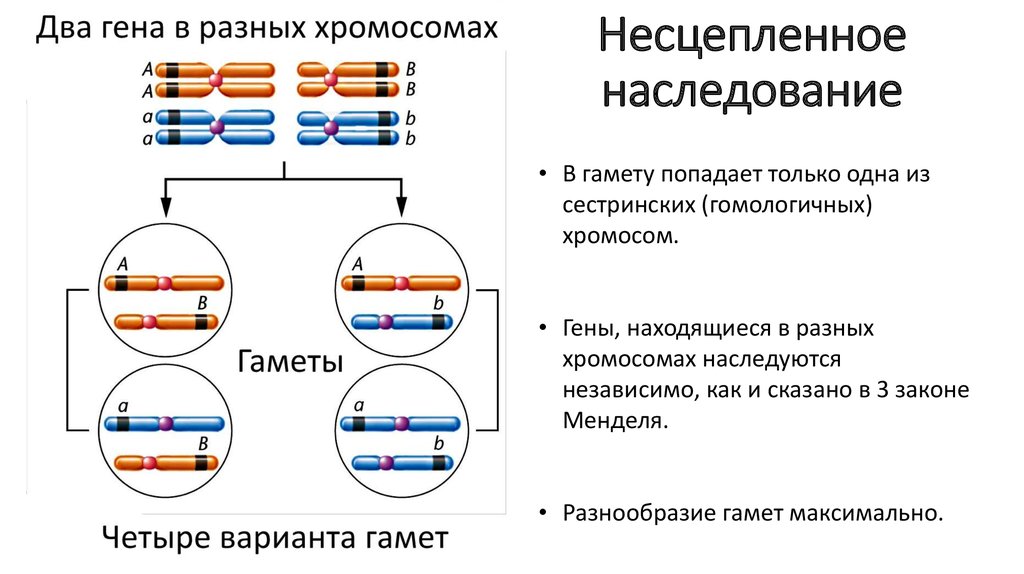 Прояв ознаки є проміжним порівняно з
Прояв ознаки є проміжним порівняно з
батьківськими формами.
Наприклад, при схрещуванні гомозиготних рослин із червоними (АА) та білими (аа) квітками у гібридів першого покоління квітки виявляються рожевими (Аа).
У людини за типом неповного домінування успадковується ознака, що визначає форму волосся: ген кучерявого волосся (А) неповністю домінує над геном прямого волосся (а), хвилясте волосся визначається генотипом — Аа.
3. Кодомінування- Це взаємодія двох домінантних алельних генів. Наприклад, кожен з алельних генів кодує певний білок і у гетерозиготного організму синтезуються два види білка. За типом кодомінування у людини успадковується четверта група крові (I I).
4. Наддомінування — в гетерозиготному стані (Аа) домінантний аллель проявляється більшою мірою, ніж у гомозиготному (АА). Наприклад, гібриди кукурудзи відрізняються вищим зростанням, урожайністю зерна порівняно з гомозиготними рослинами. Таке явище називається гетерозисом або гібридною силою. У людини але типу наддомінування проявляється акселерація.
У людини але типу наддомінування проявляється акселерація.
5. Плейотролія – один ген впливає на прояв кількох ознак, таке явище називається множинною дією одного гена. Наприклад, у людини відома хвороба – синдром Марфана – арахнодактилія («павучі пальці») детермінується домінантним геном, який відповідає за патологічний розвиток сполучної тканини, внаслідок цього проявляється комплекс патологічних ознак – довгі, тонкі («павучі») пальці, дефекти розвитку серцево- судинної системи та підвивих кришталика (порушення зору). В основі таких патологічних ознак лежить дефект розвитку сполучної тканини, зумовлений патологічним геном.
Взаємодія неалельних генів
III — I B I B l B i
IV — I A 1 B
Проте, існує рідкісний епістатичний ген (який у гомозиготному рецесивному () стан пригнічує всі домінантні алелі, що визначають групи крові. Внаслідок цього у людей із генотипом срср,фенотипно проявляється перша лише група крові.
Наприклад, у людей із генотипом \ А 1 Авиявлятиметься I група крові, т.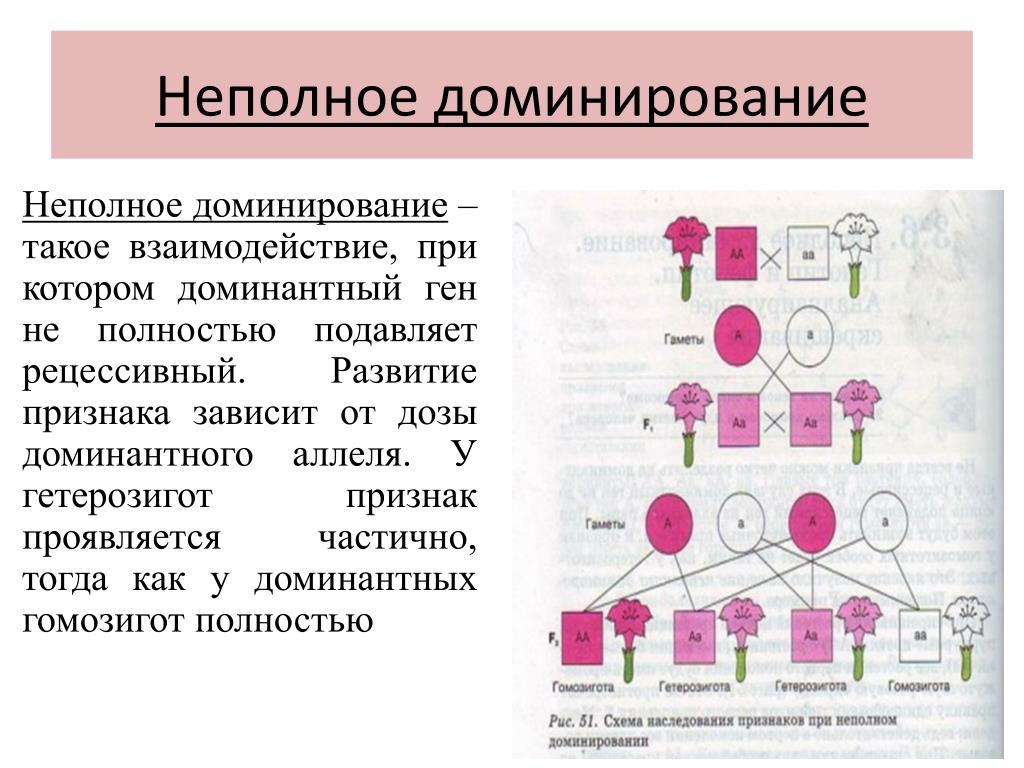 АгАг — темна aia, A 2 A 2 — смаглява а! а | А 2 а 2 — світла а!
АгАг — темна aia, A 2 A 2 — смаглява а! а | А 2 а 2 — світла а!
Крім успадкування кольору шкіри, полімерними генами у людини визначаються більшість кількісних ознак, таких як ріст, маса тіла, інтелектуальні особливості, схильність до підвищення артеріального тиску, стійкість до інфекційних захворювань та інші.
Ознаки, що визначаються кількома парами неалельних генів, називаються полігенними.
Завдання 1. Трансформуйте активні конструкції на пасивні:
щозаповнює що щоприховує що/Присутність/ щопригнічує дію чого щопригнічує що щоблокує активність чого.
Завдання 2. Напишіть речення, розкриваючи дужки:
1. Кодомінування – це явище, коли (Гетерозиготний стан)виявляються ознаки обох генів.
2. Комплементарність виявляється тоді, коли дія одного гена доповнюється (Дія іншого гена).
3. Існує рідкісний епістатичний ген (ф), який у гомозиготному рецесивному стані пригнічує (Всі домінантні алелі груп крові).
4. Активність генів блокується (Епістатичний ген у гомозиготному стані).
щообумовлено чим(тим що…)
Завдання 3. Трансформуйте прості пропозиції на складні.
1. Нормальний слух обумовлений присутністю двох неалельних генів ДіЕ.
2. Забарвлення віночків квітки обумовлено наявністю двох домінантних генів А та В.
3. Групи крові за системою АВО обумовлені успадкуванням трьох алелей одного типу (1А, 1В, Ю).
Завдання 4. Прочитайте інформацію. Скажіть, за якими ознаками розрізняються аллельні та неалельні гени:
а) аллельні гени — це гени, розташовані в однакових місцях гомологічних хромосом. Взаємодія відбувається лише у гетерозиготному стані;
б) неалельні гени — це гени, які розташовані в негомологічних хромосомах. Взаємодія між неаллельними генами відбувається у тому випадку, якщо вони відповідають за розвиток однієї ознаки.
Завдання 5. Прочитайте текст 1 «Взаємодія аллельних генів» та дайте визначення кожного з варіантів взаємодії аллельних генів.
Завдання 6. Прочитайте частину тексту «Епістаз» та дайте відповідь на запитання: «Що розуміють під…?»
а) епістаз;
б) домінантним епістазом;
в) «бомбейським феноменом»?
Завдання 7. Розкажіть про полімерію за таким планом.
1. Визначення полімерії.
2. Залежність ступеня прояви ознаки кількості домінантних генів.
3. Ознаки, що визначаються полімерними генами.
4. Приклад полімерного впливу генів.
Вирішення типових завдань
Взаємодія аллельних генів (неповне домінування)
I. У людини пряме волосся — рецесивна ознака, а кучеряве -неповністю домінують над прямими, у гетерозигот проявляються хвилясте волосся. Які діти можуть бути у батьків з хвилястим волоссям?
Позначимо гени:
А — кучеряве волосся
а — пряме волосся
Аа — генотип людини з хвилястим волоссям.
Схема шлюбу:
Р: $Аа х с?Аа
Гамети: А, а А, а
F| : АА; Аа, Аа; аа
кучеряве хвилясте пряме волосся волосся волосся
Відповідь: 25% дітей матимуть кучеряве волосся (ЛА), 25% — пряме волосся (аа) і 50% — хвилясте волосся (Аа).
Взаємодія алельних генів (кодомінування)
2. Чоловік з ІІ групою крові (гомозиготний) одружився з жінкою з ІІІ групою крові (гомозигота). Позначте:
б) генотипи батьків;
в) напишіть схему шлюбу;
д) визначте, які групи крові може бути дітей;
е) яке взаємодія алельних генів виявилося у цій ситуації?
Численні досліди підтвердили правильність встановлених Менделем закономірностей. Разом з тим, з’явилися факти, що показують, що отримані Менделем числові співвідношення при розщепленні гібридного покоління не завжди дотримувалися. Це вказувало на те, що взаємини між генами та ознаками мають більш складний характер. З’ясувалося: той самий ген може впливати на розвиток кількох ознак; один і той самий ознака може розвиватися під впливом багатьох генів.
Слід зазначити, що взаємодія генів має біохімічну природу, тобто взаємодіють друг з одним гени, які продукти. Продуктом еукаріотичного гена може бути поліпептид, або тРНК, або рРНК.
ВИДИ ВЗАЄМОДІЇ АЛЕЛЬНИХ ГЕНІВ
Розрізняють повне домінування, неповне домінування, кодомінування, алельний виняток.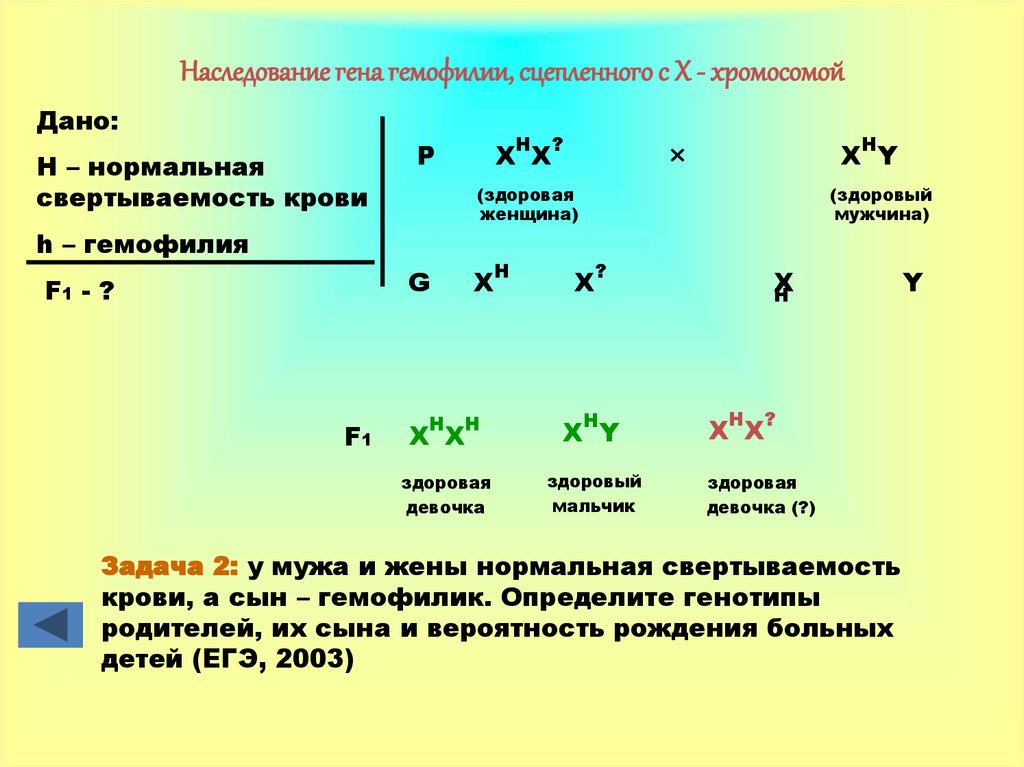
Алельними генаминазиваються гени, розташовані в ідентичних локусах гомологічних хромосом. Ген може мати одну, дві та більше молекулярних форм. Поява другої та наступних молекулярних форм є наслідком мутації гена. Якщо ген має три і більше молекулярних форм, говорять про множинному алелізмі. З усієї множини молекулярних форм в одного організму можуть бути тільки дві, що пояснюється парністю хромосом.
Повне домінування
Повне домінування— це вид взаємодії алельних генів, при якому фенотип гетерозигот не відрізняється від фенотипу гомозигот за домінантою, тобто у фенотипі гетерозигот є продукт домінантного гена. Повне домінування широко поширене в природі, має місце при успадкування, наприклад, забарвлення та форми насіння гороху, кольору очей та кольору волосся у людини, резус-антигена та багато інших. ін.
Наявність резус-антигена (резус-фактора) еритроцитів визначається домінантним геном Rh. Тобто генотип резус-позитивної людини може бути двох видів: або RhRh, або Rhrh; генотип резус-негативної людини — rhrh. Якщо, наприклад, мати — резус-негативна, а батько резус-позитивний і гетерозиготний за цією ознакою, то при даному типі шлюбу з однаковою ймовірністю може народитися як резус-позитивна, так і резус-негативна дитина.
Якщо, наприклад, мати — резус-негативна, а батько резус-позитивний і гетерозиготний за цією ознакою, то при даному типі шлюбу з однаковою ймовірністю може народитися як резус-позитивна, так і резус-негативна дитина.
Між резус-позитивним плодом та резус-негативною матір’ю може виникнути резус-конфлікт.
Так називається вид взаємодії алельних генів, при якому фенотип гетерозигот відрізняється як від фенотипу гомозигот за домінантою, так і від фенотипу гомозигот за рецесивом і має середнє (проміжне) значення між ними. Має місце при спадкуванні забарвлення оцвітини нічної красуні, левового зіву, забарвлення вовни морських свинок та ін.
Сам Мендель зіткнувся з неповним домінуванням, коли схрещував крупнолистий сорт гороху з дрібнолистим. Гібриди першого покоління не повторювали ознаки жодної з батьківських рослин, вони мали листя середньої величини.
При схрещуванні гомозиготних червоноплідних та білоплідних сортів суниці все перше покоління гібридів має рожеві плоди.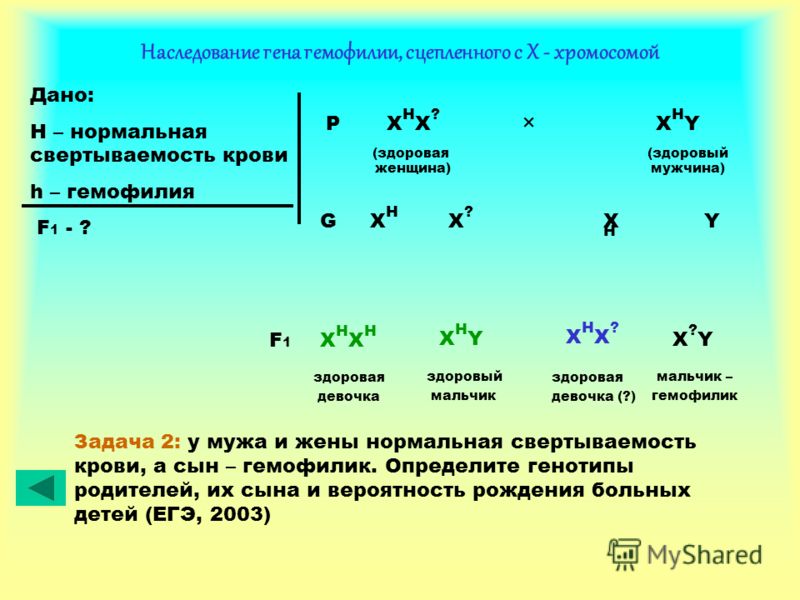 При схрещуванні цих гібридів один з одним отримуємо: по фенотипу — 1/4 червоноплідних, 2/4 рожевоплодних і 1/4 білоплідних рослин, по генотипу — 1/4 АА, 1/2 Аа, 1/4 аа (і за фенотипом, і за генотипом співвідношення 1:2:1). Відповідність розщеплення за генотипом розщеплення за фенотипом є характерною при неповному домінуванні, так як гетерозиготи фенотипно відрізняються від гомозигот.
При схрещуванні цих гібридів один з одним отримуємо: по фенотипу — 1/4 червоноплідних, 2/4 рожевоплодних і 1/4 білоплідних рослин, по генотипу — 1/4 АА, 1/2 Аа, 1/4 аа (і за фенотипом, і за генотипом співвідношення 1:2:1). Відповідність розщеплення за генотипом розщеплення за фенотипом є характерною при неповному домінуванні, так як гетерозиготи фенотипно відрізняються від гомозигот.
Кодомінування
Кодомінуваннявид взаємодії алельних генів, при якому фенотип гетерозигот відрізняється як від фенотипу гомозигот по домінанті, так і від фенотипу гомозигот за рецесивом, і у фенотипі гетерозигот присутні продукти обох генів. Має місце для формування, наприклад, IV групи крові системи (АВ0) в людини.
Для того щоб уявити, як відбувається успадкування груп крові у людини, можна подивитися, народження дітей з якою групою крові можливе у батьків, які мають один – другу, інший – третю групу крові та є гетерозиготними за цією ознакою.
| Р | ♀I A i 0 II (A) | × | ♂I B i 0 III (B) | ||
| Типи гамет | I A | i 0 | I B | i 0 | |
| F | i 0 i 0 I (0) 25% | I A i 0 II (A) 25% | I B i 0 III (B) 25% | I A I B IV (AB) 25% | |
Алельний виняток
Алельним виняткомназивається відсутність чи інактивація одного з пари генів; у цьому випадку у фенотипі присутній продукт іншого гена (гемізиготність, делеція, гетерохроматизація ділянки хромосоми, в якій міститься необхідний ген).
ВИДИ ВЗАЄМОДІЇ НЕАЛЕЛЬНИХ ГЕНІВ
Комплементарність, епістаз, полімерія.
Неалельні гени— гени, розташовані або в неідентичних локусах гомологічних хромосом, або в різних парах гомологічних хромосом.
Взаємодія генів – Медичні статті
Взаємодія алельних генів
До складу генотипу входить велика кількість генів, які функціонують і взаємодіють як цілісна система. Г. Мендель у своїх дослідах виявив тільки одну форму взаємодії між алельними генами — повне домінування одного алеля і повну рецесивність іншого. Генотип організму не можна розглядати як просту суму незалежних генів, кожен з яких функціонує поза зв’язком з іншими. Фснотипні прояви тієї або іншої ознаки є результатом взаємодії багатьох генів.
Розрізняють дві основних групи взаємодії генів: взаємодія між алельними генами і взаємодія між неалельними генами. Проте слід розуміти, що це не фізична взаємодія самих генів, а взаємодія первинних і вторинних продуктів, які зумовлять ту чи іншу ознаку.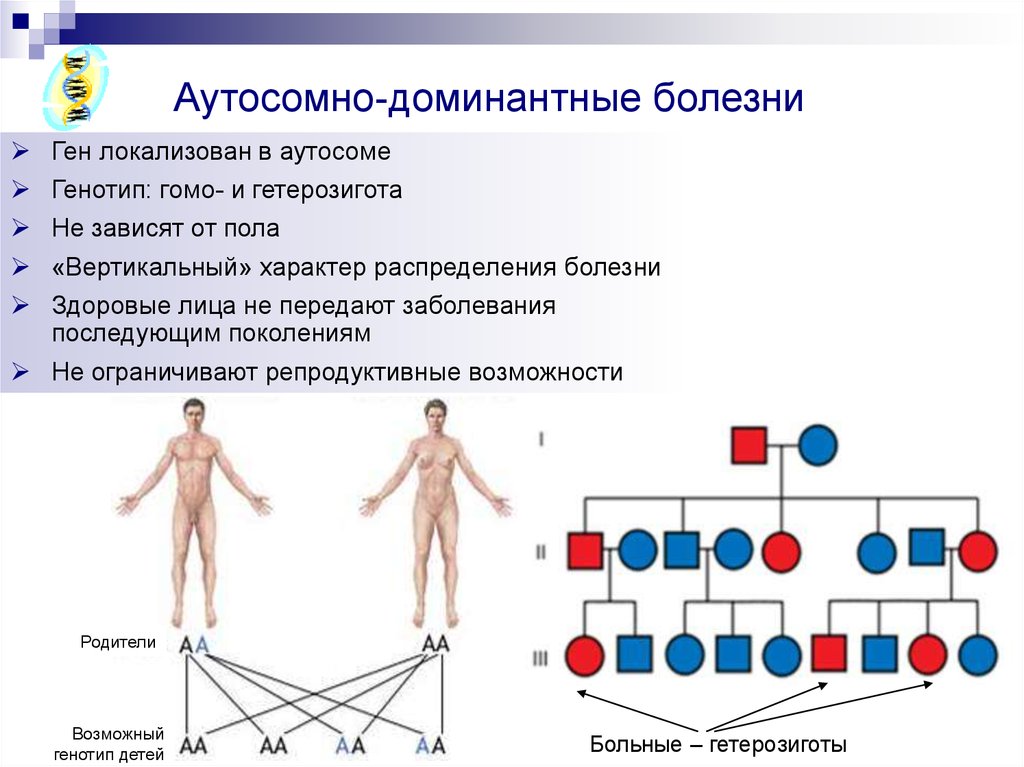 У цитоплазмі відбувається взаємодія між білками – ферментами, синтез яких визначається генами, або між речовинами, які утворюються під впливом цих ферментів.
У цитоплазмі відбувається взаємодія між білками – ферментами, синтез яких визначається генами, або між речовинами, які утворюються під впливом цих ферментів.
Можливі такі типи взаємодії:
1) для утворення певної ознаки необхідна взаємодія двох ферментів, синтез яких визначається двома неалельними генами;
2) фермент, який синтезувався за участю одного гена, повністю пригнічує або інактивує дію ферменту, утвореного іншим неалельиим геном;
3) два ферменти, утворення яких контролюється двома неалельними генами, впливають на одну ознаку або на один процес так, що їх сумісна дія призводить до виникнення і підсилення прояву ознаки.
Взаємодія алельних генів. Гени, які займають ідентичні (гомологічні) локуси в гомологічних хромосомах, називаються алельними. У кожного організму є лише по два алельних гени.
Відомі такі форми взаємодії між алельними генами: повне домінування, неповне домінування, кодомінування і наддомінування.
Основна форма взаємодії – повне домінування, яке вперше описано Г. Менделем . Суть його полягає в тому, що в гетерозиготного організму прояв одного з алелів домінує над проявом іншого. За повного домінування розщеплення за генотипом 1:2:1 не співпадає з розщепленням за фенотипом — 3:1. У медичній практиці з двох тисяч моногенних спадкових хвороб у майже половини має місце домінування прояву патологічних генів над нормальними. У гетерозигот патологічний алель проявляється у більшості випадків ознаками захворювання (домінантний фенотип).
Менделем . Суть його полягає в тому, що в гетерозиготного організму прояв одного з алелів домінує над проявом іншого. За повного домінування розщеплення за генотипом 1:2:1 не співпадає з розщепленням за фенотипом — 3:1. У медичній практиці з двох тисяч моногенних спадкових хвороб у майже половини має місце домінування прояву патологічних генів над нормальними. У гетерозигот патологічний алель проявляється у більшості випадків ознаками захворювання (домінантний фенотип).
Неповне домінування — така форма взаємодії, коли у гетерозиготного організму (Аа) домінантний ген (А) не повністю пригнічує рецесивний ген (а), внаслідок чого проявляється проміжна між батьківськими ознака. Тут розщеплення за генотипом і фенотипом співпадає і складає 1:2:1
При кодомінуванні в гетерозиготних організмів кожний з алельних генів викликає формування залежного від нього продукту, тобто виявляються продукти обох алелів. Класичним прикладом такого прояву є система груп крові, зокрема система АBО, коли еритроцити людини несуть на поверхні антигени, що контролюються обома алелями, така форма прояву носить назву кодомінування.
Наддомінування — коли домінантний ген у гетерозиготному стані виявляється сильніше, ніж у гомозиготному. Так, у дрозофіли при генотипі АА -нормальна тривалість життя; Аа – подовжена триватість життя; аа – летальний наслідок
Множинний алелізм
У кожного організму є лише по два алельних гени. Разом з тим нерідко в природі кількість алелів може бути більше двох, коли якийсь локус може перебувати в різних станах. У таких випадках говорять про множинні алелі або множинний алеломорфізм.
Множинні алелі позначаються однією літерою з різними індексами, наприклад : А , А1, А3… Алельні гени локалізуються в однакових ділянках гомологічних хромосом. Оскільки в каріотипі завжди присутні по дві гомологічних хромосоми, то і при множинних алелях кожний організм може мати одночасно лише по два однакових або різних алелі. У статеву клітину (разом із розходженням гомологічних хромосом) попадає тільки по одному із них. Для множинних алелів характерний вплив всіх алелів на одну і ту ж ознаку. Відмінність між ними полягає лише в ступені розвитку ознаки.
Відмінність між ними полягає лише в ступені розвитку ознаки.
Другою особливістю є те, що в соматичних клітинах або в клітинах диплоїдних організмів міститься максимум по два алелі з декількох, оскільки вони розташовані в одному і тому ж локусі хромосоми. Ще одна особливість властива множинним алелям. За характером домінування алеломорфні ознаки розміщуються в послідовному ряду: частіше нормальна, незмінена ознака домінує над іншими; другий ген ряду рецесивний щодо першого, проте домінує над наступними і т.д. Одним з прикладів прояву множинних алелів у людини є групи крові системи АВО. Множинний алелізм має важливе біологічне і практичне значення, оскільки підсилює комбінативну мінливість, особливо генотипну.
Взаємодія неалельних генів
Відомо багато випадків, коли ознака або властивості детермінуються двома або більше неалельними генами, які взаємодіють між собою. Хоча і тут взаємодія умовна, бо взаємодіють не гени, а контрольовані ними продукти. При цьому має місце відхилення від менделівських закономірностей розщеплення.
Розрізняють чотири основних типи взаємодії генів: комплементарність, епістаз, полімерію і модифікуючу дію (плейотропія).
Комплементарність — такий тип взаємодії неалельних генів, коли один домінантний ген доповнює дію іншого не алельного домінантного гена, і вони разом визначають нову ознаку, яка відсутня у батьків. Причому відпо¬відна ознака розвивається тільки в присутності обох неалельних генів. Наприклад, сіре забарвлення шерсті у мишей контролюється двома генами (А і В). Ген А детермінує синтез пігменту, проте як гомо-зиготи (АА), так і гетерозиготи (Аа) – альбіноси. Інший ген В забезпечує скупчення пігменту пере¬важно біля основи і на кінчиках волосся. Схрещування дигетерозигот (АаВЬ х АаВЬ) призводить до розщеплення гібридів у співвідношені 9:3:4. Числові співвідношення при комплементарній взаємодії можуть бути як 9:7; 9:6:1 (видозміна менделівського розщеплення).
Прикладом комплементарної взаємодії генів у людини може бути синтез захисного білка – інтер¬ферону. Його утворення в організмі пов’язано з комп¬лементарною взаємодією двох неалельних генів, розташованих у різних хромосомах.
Його утворення в організмі пов’язано з комп¬лементарною взаємодією двох неалельних генів, розташованих у різних хромосомах.
Епістаз – це така взаємодія неалеьних генів, за якої один ген пригнічує дію іншого неалельного гена. Пригнічення можуть викликати як домінантні, так і рецесивні гени (А>В, а>В, В>А, В>А), і залежно від цього розрізня¬ють епістаз домінантний і рецесивний. Пригнічуючий ген отримав назву інгібітора, або супресора. Гени-інгібітори в основному не детермінують розвиток певної ознаки, а лише пригнічують дію іншого гена.
Ген, ефект якого пригнічується, отримав назву гіпостатичного. При епістатичній взаємодії генів розщеплення за фенотипом в F2 складає 13:3; 12:3:1 або 9:3:4 та ін. Забарвлення плодів гарбузів, масть коней визначаються цим типом взаємодії.
Якщо ген-супресор рецесивний, то виникає криптомерія (від грец. хріштад – таємний, прихований). У людини таким прикладом може бути «бомбейський феномен». У цьому випадку рідкісний рецесивний алель «х» у гомозиготному стані (хх) пригнічує активність гена jB (який визначає В (III) групу крові системи АВО). Тому жінка з генотипом jв_хх, фенотипно має І групу крові – 0 (І).
Тому жінка з генотипом jв_хх, фенотипно має І групу крові – 0 (І).
Полігенне успадкування кількісних ознак
– плейотропія
– експресивність та пенетрантність генів.
Більшість кількісних ознак організмів визначається декількома неалельними генами (полігенами). Взаємодія таких генів у процесі формування ознаки називається полімерною. У цьому випадку два або більше домінантних алелі однаковою мірою впливають на розвиток однієї і тієїж ознаки. Тому полімерні гени прийнято позначати однією літерою латинського алфавіту з цифровим індексом, наприклад: А1А1 і а1а1. Вперше однозначні фактори були виявлені шведським генетиком Нільсоном-Еле (1908 р.) при вивченні успадкування кольору у пшениці. Було встановлено, що ця ознака залежить від двох полімерних генів, тому при схрещуванні домінантних і рецесивних дигомозигот – забарвленої (А1А1 , А2 А2) з безколірною (а1а1, а2а2) – у F, всі рослини дають забарвлене насіння, хоча вони помітно світліші, ніж батьківські екземпляри, які мають червоне насіння. У F, при схрещуванні особин першого покоління виявляється розщеплення за фенотипом у співвідношенні 15 : 1, бо безколірними є лише рецесивні дигомозиготи (аІа1 а2а2.). У пігментованих екземплярів інтенсивність кольору дуже коливається залежно від кількості отриманих ними домінантних алелів: максимальна в домінантних дигомозигот (А1А1 А2 А2 і мінімальна в носіїв одного з домінантних алелів).
У F, при схрещуванні особин першого покоління виявляється розщеплення за фенотипом у співвідношенні 15 : 1, бо безколірними є лише рецесивні дигомозиготи (аІа1 а2а2.). У пігментованих екземплярів інтенсивність кольору дуже коливається залежно від кількості отриманих ними домінантних алелів: максимальна в домінантних дигомозигот (А1А1 А2 А2 і мінімальна в носіїв одного з домінантних алелів).
Важлива особливість полімерії – сумація дії неалельних генів на розвиток кількісних ознак. Якщо при моногенному успадкуванні ознаки можливі три варіанти «доз» гена в генотипі: АА, Аа, аа, то при полігенному кількість їх зростає до чотирьох і більше. Сумація «доз» полімерних генів забез¬печує існування неперервних рядів кількісних змін.
Біологічне значення полімерії полягає ще й в тому, що ознаки, які кодуються цими генами, більш стабільні, ніж ті, що кодуються одним геном. Організм без полімерних генів був би дуже нестійким: будь-яка мутація або рекомбінація призводила б до різкої мінливості, а це в більшості випадків має несприятливий характер.
У тварин і рослин є багато полігенних ознак, серед них і цінні для господарства: інтенсивність росту, скороспілість, несучість, кількість молока, вміст цукристих речовин і вітамінів тощо. Пігментація шкіри у людини визначається п’ятьма або шістьма полімерними генами. У корінних жителів Африки (негроїдної раси) переважають до¬мінантні алелі, у представників європеоїдної раси – рецесивні. Тому мулати мають проміжну пігментацію, але при шлюбах мулатів у них можлива поява як більш, так і менш інтенсивно пігментованих дітей .
Багато морфологічних, фізіологічних і патологічних особливостей людини визначається полімерними генами: зріст, маса тіла, величина артеріального тиску та ін. Розвиток таких ознак у людини підпорядковується загальним законам полігенного успадкування і залежить від умов середовища. У цих випадках спостерігається, наприклад, схильність до гіпертонічної хвороби, ожиріння тощо. Дані ознаки за сприятливих умов середовища можуть не проявитися або проявитися незначною мірою. Цим полігенноспадкові ознаки відрізняються від моногенних. Змінюючи умови середовища, можна значною мірою забезпечити профілактику ряду полігенних захворювань.
Цим полігенноспадкові ознаки відрізняються від моногенних. Змінюючи умови середовища, можна значною мірою забезпечити профілактику ряду полігенних захворювань.
Плейотропія.
Плейотропна дія генів – це залежність кількох ознак від одного гена, тобто множинна дія одного гена. У дрозофіли ген білого кольору очей воднораз впливає на колір тіла, довжи-ну крил, будову статевого апарату, знижує плодю¬чість, зменшує тривалість життя. У людини відома спадкова хвороба – арахнодактилія («павучі пальці» -дуже тонкі і довгі пальці), або хвороба Марфана. Ген, який відповідає за цю хворобу, викликає порушення розвитку сполучної тканини й одночасно впли¬ває на розвиток кількох ознак: порушення будови кришталика ока, аномалії у серцево-судинній системі.
Плейотропна дія гена може бути первинною і вторинною. При первинній плейотропії ген проявляє свій множинний ефект. Наприклад, при хворобі Хартнупа мутація гена призводить до порушення всмоктування амінокислоти триптофану в кишках і його реабсорбції у ниркових канальцях.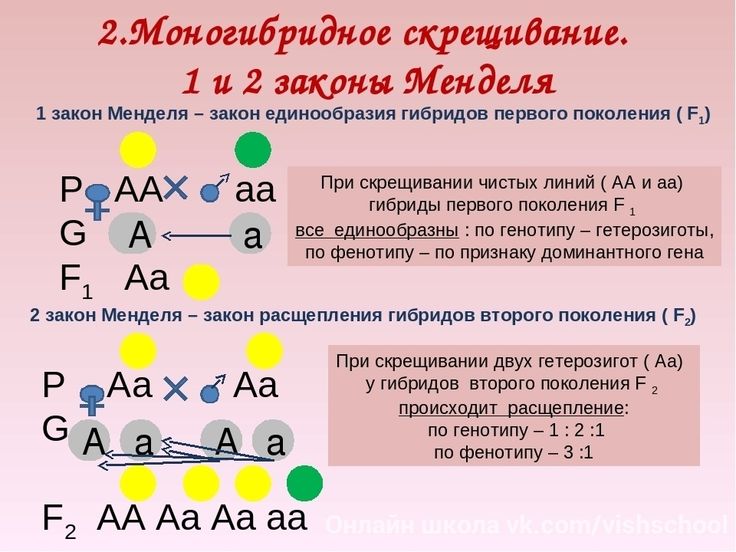 При цьому вражаються одночасно мембрани епітеліальних клітин кишок і ниркових канальців з розладами травної і видільної систем. При вторинній плейотропії є один первинний фенотипний прояв гена, услід за яким роз-вивається ступінчастий процес вторинних змін, які призводять до множинних ефектів. Так, при серпоклітинній анемії у гомозигот спостерігається кілька патологічних ознак: анемія, збільшена селезінка, ураження шкіри, серця, нирок і мозку. Тому гомозиготи за геном серпоподібноклітинної анемії гинуть, як правило, в дитячому віці. Всі ці фенотипні прояви гена складають ієрархію вторинних проявів. Першопричиною, безпосереднім фенотипним проявом дефектного гена є аномальний гемоглобін і еритроцити серпоподібної форми. Внаслідок цього відбуваються послідовно інші патологічні процеси: злипання і руйнування еритроцитів, анемія, дефекти в нирках, серці, мозку Ці патологічні ознаки є вторинними. При плейотропії ген, впливаючи на якусь одну основну ознаку, може також змінювати, модифікувати прояв інших генів, у зв’язку з чим введено поняття про гени-модифікатори.
При цьому вражаються одночасно мембрани епітеліальних клітин кишок і ниркових канальців з розладами травної і видільної систем. При вторинній плейотропії є один первинний фенотипний прояв гена, услід за яким роз-вивається ступінчастий процес вторинних змін, які призводять до множинних ефектів. Так, при серпоклітинній анемії у гомозигот спостерігається кілька патологічних ознак: анемія, збільшена селезінка, ураження шкіри, серця, нирок і мозку. Тому гомозиготи за геном серпоподібноклітинної анемії гинуть, як правило, в дитячому віці. Всі ці фенотипні прояви гена складають ієрархію вторинних проявів. Першопричиною, безпосереднім фенотипним проявом дефектного гена є аномальний гемоглобін і еритроцити серпоподібної форми. Внаслідок цього відбуваються послідовно інші патологічні процеси: злипання і руйнування еритроцитів, анемія, дефекти в нирках, серці, мозку Ці патологічні ознаки є вторинними. При плейотропії ген, впливаючи на якусь одну основну ознаку, може також змінювати, модифікувати прояв інших генів, у зв’язку з чим введено поняття про гени-модифікатори. Останні підсилюють або послаблюють розвиток ознак, які кодуються «основним» геном. Показниками залежності функціонування спадкових задатків від характеристик генотипу є пенетрантність і експресивність.
Останні підсилюють або послаблюють розвиток ознак, які кодуються «основним» геном. Показниками залежності функціонування спадкових задатків від характеристик генотипу є пенетрантність і експресивність.
Розглядаючи дію генів, їх алелів необхідно врахувати і модифікуючий вплив середовища, в якому розвивається організм. Якщо рослини примули схрещувати при температурі 15-20 °С, то в F1 згідно з менделівською схемою, все покоління матиме рожеві квіти. Але коли таке схрещування проводити за температури 35 °С, то всі гібриди матимуть квіти білого кольору. Якщо ж здійснювати схрещування при температурі близько 30 °С, то виникає різне співвідношення (від 3:1 до 100 відсотків) рослин з білими квітами.
Таке коливання класів при розщепленні залежно від умов середовища отримало назву пенетрантність – сила фенотипного прояву. Отже, пенетрантність — це частота прояву гена, явище появи або відсутності ознаки у організмів, однакових за генотипом.
Пенетрантність значно коливається як серед домінантних, так і серед рецесивних генів.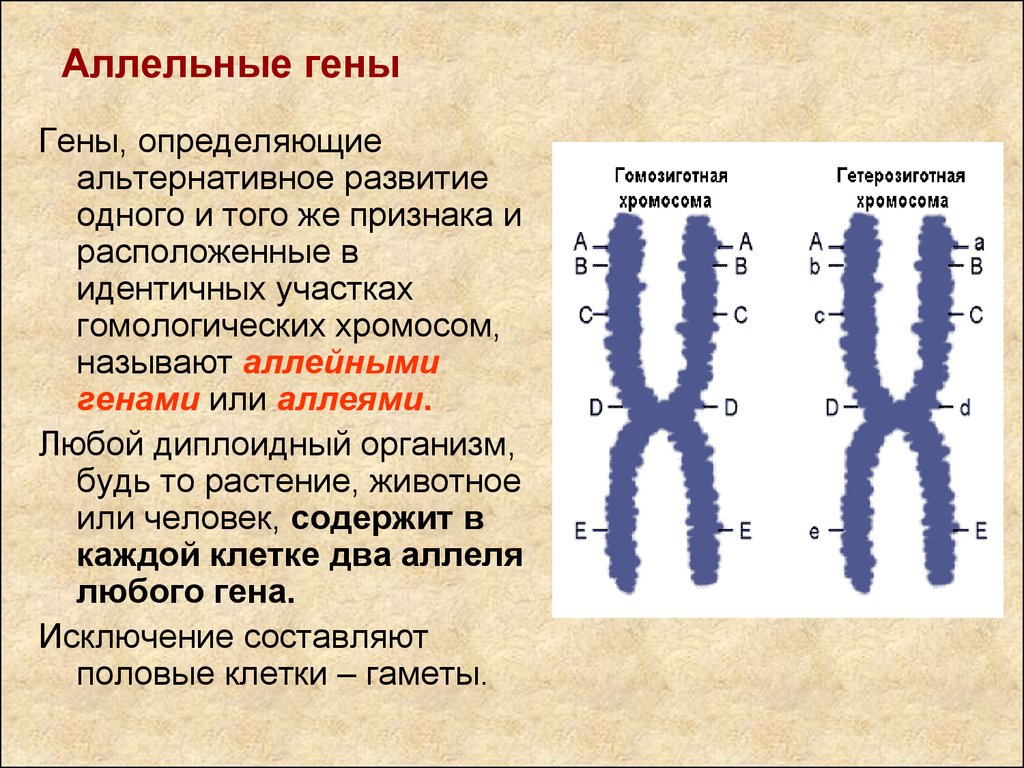 Поряд з генами, фенотип яких з’являється тільки за поєднання певних умов і досить рідкісних зовнішніх умов (висока пенетрантність), у людини є гени, фснотипний прояв яких відбувається за будь-яких поєднань зовнішніх умов (низька пенетрантність). Пенетрантність вимірюється відсотком організмів з фенотипною ознакою від загальної кількості обстежених носіїв відповідного алеля.
Поряд з генами, фенотип яких з’являється тільки за поєднання певних умов і досить рідкісних зовнішніх умов (висока пенетрантність), у людини є гени, фснотипний прояв яких відбувається за будь-яких поєднань зовнішніх умов (низька пенетрантність). Пенетрантність вимірюється відсотком організмів з фенотипною ознакою від загальної кількості обстежених носіїв відповідного алеля.
Якщо ген повністю , незалежно від навколишнього середовища , визначає фенотиповий прояв, то він має пенетрантність 100 відсотків. Проте деякі домінантні гени проявляються менш регулярно. Так, полідактилія має чітке вертикальне успадкування, але бувають пропуски поколінь. Домінантна аномалія – передчасне статеве дозрівання – властиве тільки чоловікам, проте іноді може передатися захворювання від чоловіка, який не страждав цією патологією. Пенетрантність показує, у якому відсотку носіїв гена виявляється відповідний фенотип. Отже, пенетрантність залежить від генів, від середовища, від того й іншого. Таким чином, це не константна властивість гена, а функція генів у певних умовах середовища.
Таким чином, це не константна властивість гена, а функція генів у певних умовах середовища.
Експресивність (від лат. ехргеssіо — вираз) -це зміна кількісного прояву ознаки у різних особин -носіїв відповідного алеля.
При домінантних спадкових захворюваннях експресивність може коливатися. В одній і тій самій родині можуть проявлятися спадкові хвороби за перебігом від легких, ледь помітних до тяжких: різні форми гіпертонії, шизофренії, цукрового діабету тощо. Рецесивні спадкові захворювання в межах сім’ї проявляються однотипно і мають незначні коливання експресивності.
Что такое доминантный и рецессивный?
Типы наследования
Серповидноклеточная анемия — наследственное заболевание, вызывающее боль и повреждение органов и
мышцы. Вместо уплощенных круглых эритроцитов у людей с этим заболеванием
жесткие, серповидные клетки. Длинные заостренные клетки крови попадают в капилляры, где
они блокируют кровоток. Мышцы и клетки органов не получают достаточного количества кислорода и питательных веществ, и
они начинают умирать.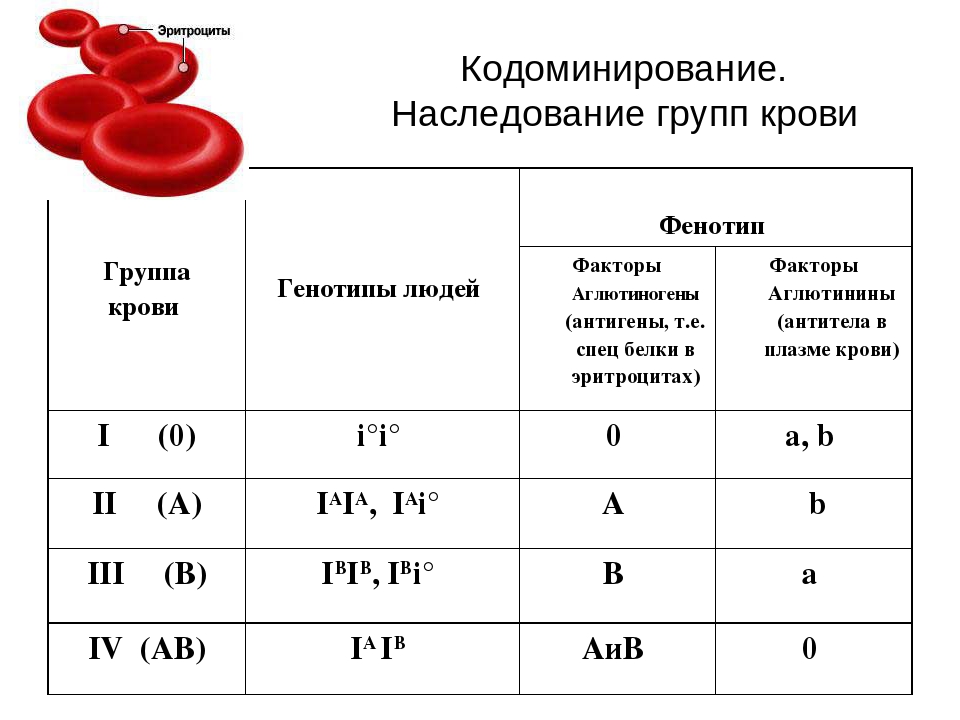
Заболевание имеет рецессивный тип наследования: только лица с двумя копиями серповидноклеточный аллель имеет заболевание. Люди с одной копией здоровы.
Аллель серповидноклеточной анемии не только вызывает заболевание, но и делает носителей устойчивыми к малярии, тяжелому заболеванию, переносимому комарами. Устойчивость к малярии имеет доминирующее характер наследования: всего одной копии аллеля серповидноклеточной анемии достаточно для защиты от инфекционное заболевание. Это тот самый аллель, который при рецессивном типе наследования вызывает серповидно-клеточная анемия!
Теперь давайте снова посмотрим на форму клеток крови. Люди с двумя копиями серповидноклеточной анемии
аллель имеют много серповидных эритроцитов. Люди с двумя копиями «нормального» аллеля имеют
дисковидные эритроциты. Люди с одним серповидно-клеточным аллелем и одним нормальным аллелем имеют
небольшое количество серповидных клеток, и их клетки легче серповидны при определенных условиях.
Таким образом, мы можем сказать, что форма эритроцитов имеет кодоминантный тип наследования. То есть,
люди с одной копией каждого аллеля имеют промежуточный фенотип.
То есть,
люди с одной копией каждого аллеля имеют промежуточный фенотип.
Так является ли аллель серповидноклеточной анемии доминантным, рецессивным или кодоминантным? Это зависит от того, как вы на это смотрите.
Функция белка
Если мы посмотрим на белки, которые кодируются двумя аллелями, картина станет немного более ясной. Пораженным белком является гемоглобин, молекула, несущая кислород, которая наполняет эритроциты. Аллель серповидноклеточной анемии кодирует слегка модифицированную версию белка гемоглобина. Модифицированный белок гемоглобина по-прежнему переносит кислород, но в условиях низкого содержания кислорода белки слипаются.
Когда у человека есть два аллеля серповидноклеточной анемии, весь его гемоглобин представляет собой липкую форму, а
белки образуют очень длинные жесткие волокна, которые искажают эритроциты. Когда у кого-то есть
один серповидноклеточный аллель и один нормальный аллель, только часть гемоглобина липкая.
Нелипкий гемоглобин образуется из нормального аллеля, а липкий гемоглобин образуется из
серповидноклеточный аллель (каждая клетка имеет копию обоих аллелей). Склеивание
эффект разбавлен, и в большинстве клеток белки не образуют волокон.
Склеивание
эффект разбавлен, и в большинстве клеток белки не образуют волокон.
Протисты, вызывающие малярию, растут и размножаются в эритроцитах. Как именно Аллель серповидноклеточной анемии приводит к устойчивости к малярии, является сложным и не до конца изученным. Однако оказывается, что паразит размножается медленнее в клетках крови, которые немного модифицированного гемоглобина. А инфицированные клетки, поскольку они легко деформируются, быстрее изымаются из обращения и уничтожаются.
Чтобы увидеть больше примеров того, как вариации генов влияют на черты, посетите The Outcome of Mutation.
Неполное доминирование – определение и примеры
Неполное доминирование Определение
Неполное доминирование – это когда доминантный аллель или форма гена не полностью маскирует эффекты рецессивного аллеля, и в результате внешний вид организма демонстрирует смешение оба аллеля. Его также называют полудоминированием или частичным доминированием. Один пример показан в розах. Аллель красного цвета доминирует над аллелем белого цвета, но гетерозиготные розы, имеющие оба аллеля, розовые. Обратите внимание, что это отличается от кодоминирования, когда оба аллеля экспрессируются одновременно.
Аллель красного цвета доминирует над аллелем белого цвета, но гетерозиготные розы, имеющие оба аллеля, розовые. Обратите внимание, что это отличается от кодоминирования, когда оба аллеля экспрессируются одновременно.
Механизмы неполного доминирования
Многие гены демонстрируют полное доминирование. Это означает, что если человек гетерозиготен по определенному гену, доминантный аллель будет полностью маскировать рецессивный аллель. Многие из свойств, которые австрийский монах Грегор Мендель изучал в своих знаменитых растениях гороха, контролировались генами, проявлявшими полное доминирование. Например, доминирующий цвет цветка был фиолетовым, а рецессивный — белым. Растения, которые были гетерозиготными, также были фиолетовыми, поскольку фиолетовый был доминирующим аллелем, хотя у них также был белый аллель. У растения были белые цветы только в том случае, если оно было гомозиготным по рецессивному аллелю, что означает наличие двух копий этого аллеля. (Вот почему два пурпурных растения иногда давали белые; часть потомства получала два рецессивных аллеля. )
)
Почему возникает неполное доминирование? Как мы видели, это не всегда происходит с окраской цветка; розы (а также тюльпаны, гвоздики и львиный зев среди прочих) показывают неполное доминирование, но растения гороха Менделя показали полное доминирование. Неполное доминирование может происходить из-за того, что ни один из двух аллелей не полностью доминирует над другим, или из-за того, что доминантный аллель не полностью доминирует над рецессивным аллелем. Это приводит к фенотипу, который отличается как от доминантного, так и от рецессивного аллелей и, по-видимому, представляет собой смесь обоих.
Этот квадрат Пеннета показывает неполное доминирование. Гомозиготный красный цветок имеет два доминантных аллеля красного цвета, которые обозначаются буквами RR. Гомозиготный белый цветок представлен rr. Все их потомки гетерозиготны по Rr и имеют розовые цветки. Это первое дочернее поколение, или F 1 . При перекрестном опылении поколения F 1 их потомство будет RR, Rr и rr в соотношении 1:2:1. Некоторые из их потомков (F 2 ) унаследуют два аллеля R, некоторые унаследуют два аллеля r, а некоторые унаследуют оба аллеля.
Некоторые из их потомков (F 2 ) унаследуют два аллеля R, некоторые унаследуют два аллеля r, а некоторые унаследуют оба аллеля.
Неполное доминирование и кодоминирование
Неполное доминирование — это не то же самое, что кодоминирование. При кодоминантности оба аллеля могут проявляться в фенотипе одновременно. Вместо того, чтобы быть однородно розовым, цветок с красными и белыми аллелями, которые демонстрируют кодоминирование, будет иметь пятна красного и белого цветов. Как и при неполном доминировании, поколение F 2 от гетерозиготных растений будет иметь соотношение красных, пятнистых и белых цветков 1:2:1. Кодоминирование также показано у людей с группой крови AB; экспрессируются аллели групп крови А и В.
Примеры неполного доминирования
У человека
Ребенок, рожденный от родителей с прямыми волосами и родителей с кудрявыми волосами, обычно имеет волнистые волосы или волосы, которые немного завиты, из-за выражения как кудрявых, так и прямых аллели. Неполное доминирование можно увидеть во многих других физических характеристиках, таких как цвет кожи, рост, размер руки и высота голоса.
Неполное доминирование можно увидеть во многих других физических характеристиках, таких как цвет кожи, рост, размер руки и высота голоса.
У носителей болезни Тея-Сакса также наблюдается неполное доминирование. У людей с болезнью Тея-Сакса отсутствует фермент, расщепляющий липиды, что приводит к накоплению слишком большого количества липидов в головном мозге и других частях нервной системы. Это приводит к ухудшению состояния нервов и потере физических и умственных способностей. Болезнь Тея-Сакса возникает у людей с двумя рецессивными аллелями болезни, а люди с одним аллелем являются носителями, но не проявляют симптомов. Однако они продуцируют половину нормального количества фермента, демонстрируя промежуточный фенотип между теми, у кого есть расстройство, и теми, у кого нет рецессивных аллелей Тея-Сакса.
У других животных
Андалузский цыпленок, тип цыпленка, произрастающий в регионе Андалусия в Испании, демонстрирует неполное преобладание цвета оперения. Белый самец и черная самка часто производят потомство с голубоватыми перьями. Это вызвано геном разбавления, который частично разбавляет пигмент меланин и делает перья светлее.
Это вызвано геном разбавления, который частично разбавляет пигмент меланин и делает перья светлее.
При разведении определенных видов длинношерстных и короткошерстных кроликов их потомство будет иметь мех средней длины. Это явление можно увидеть и по длине хвостов собак. Кроме того, животное с большим количеством пятен будет иметь потомство с несколькими пятнами, если его скрестить с животным без пятен. Это часто наблюдается у собак, кошек и лошадей.
- Аллель – определенная форма гена.
- Доминантный – Аллель, маскирующий фенотип рецессивного аллеля того же гена.
- Фенотип – Наблюдаемые физические характеристики генов и окружающей среды.
- Квадрат Пеннета – Диаграмма, показывающая возможные результаты размножения двух особей.
Тест
1. Что НЕ является примером неполного доминирования?
A. Розовый цветок, полученный из красных и белых цветов
B. Красный и белый цветок, полученный из красных и белых цветов потомство с шерстью
Красный и белый цветок, полученный из красных и белых цветов потомство с шерстью
D. Собака с сильными пятнами и собака без пятен, приносящие щенков с несколькими пятнами
Ответ на вопрос №1
B верно. Это пример кодоминирования, а не неполного доминирования, потому что показаны оба фенотипа вместо одного промежуточного фенотипа.
2. Роза демонстрирует неполное доминирование; при скрещивании растения с красными цветками с растением с белыми цветками потомство будет розовым с фенотипом Rr. Если этих розовых особей скрестить, каково будет соотношение их потомства?
A. 1:2:1 красный, розовый, белый
B. 1:2:1 красный, пятнистый, белый
C. 3:1 красный, белый
D. 2:1 розовый, красный
Ответ на вопрос № 2
Правильно . Четверть потомства будет RR и иметь красный фенотип, а еще четверть будет rr и белым.
Leave A Comment