
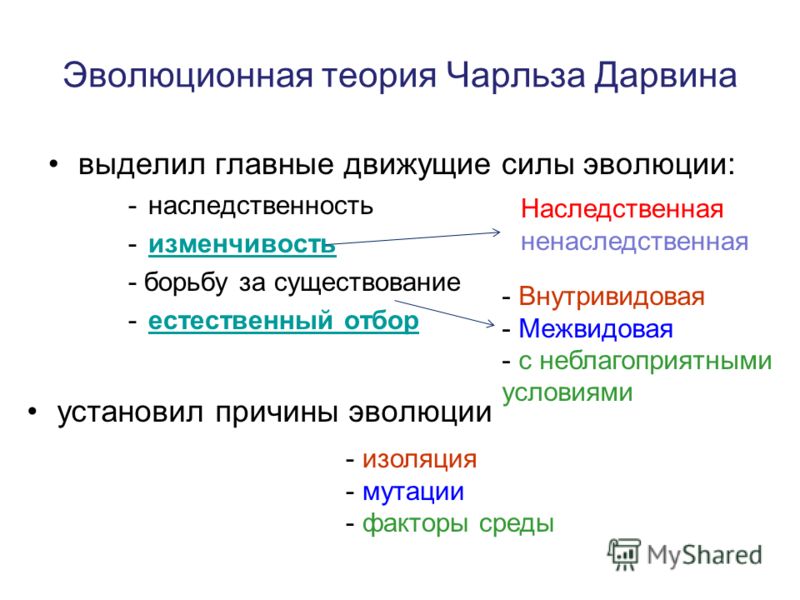
«Основы учения об эволюции». Тест
Категория: Биология.
«Основы учения об эволюции». Тест
Уровень А.
1. Ведущую роль в эволюции играет следующий вид изменчивости:
а. Определенная; б) Модификационная в) Групповая г) Мутационная
2. Способность живых существ производить большое количество потомков и ограниченность мест обитания и жизненных ресурсов являются непосредственными причинами:
а. Наследственной изменчивости;
б. Борьбы за существование;
в. Вымирания;
г. Видообразования.
3. Наиболее напряженной формой борьбы за существование Ч.Дарвин считал:
а. Борьбу с неблагоприятными условиями;
б. Внутривидовую;
в. Межвидовую;
г. Все перечисленные формы в равной степени.
4. Наследственная изменчивость в процессе эволюции:
а. Создает новые виды;
б. Поставляет материал для эволюции
в. Закрепляет созданный в процессе эволюции материал
г.
5. Естественный отбор действует на уровне:
а. Отдельного организма
б. Популяции
в. Вида
г. Биоценоза
6. Движущая форма отбора обычно приводит:
а. К уничтожению особей с отклонениями от прежней нормы реакции
б. Сужению прежней нормы реакции
в. Расширению прежней нормы реакции
г. Сдвигу прежней нормы реакции
7. Результатом действия естественного отбора не является:
а. Приспособленность организмов к среде обитания
б. Многообразие органического мира
в. Борьбе за существование
г. Совершенствование организации живых существ.
8. Изоляция – это фактор эволюции, который:
б. Замедляет процесс формирования приспособленности
в. Не препятствует смешиванию популяции внутри вида
г. Ускоряет эволюционный процесс.
9. Микроэволюция приводит к образованию новых:
а. Семейных групп
б.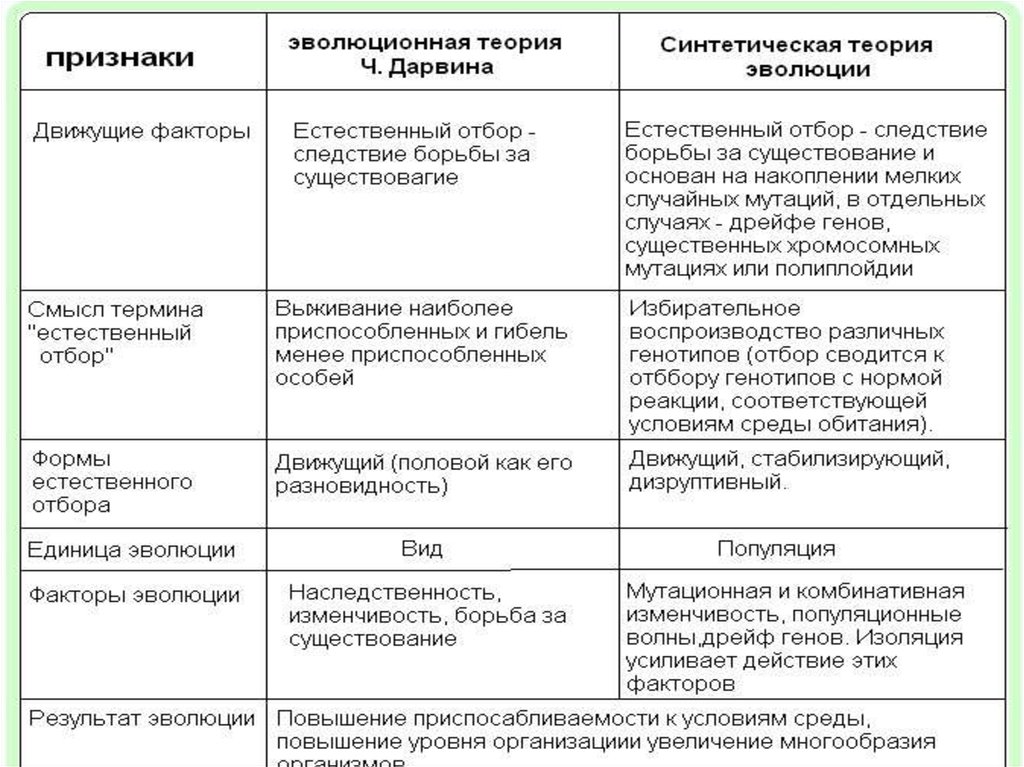 Подвидов и видов
Подвидов и видов
в. Родов
г. Отрядов и семейств.
10. Крупные систематические группы в процессе эволюции возникают, как правило, путем:
а. Ароморфоза
б. Идиоадаптации
в. Общей дегенерации
г. Направленной эволюции
11. К идиоадаптации следует отнести:
а. Многоклеточность
б. Четырехкамерное сердце млекопитающих
в. Игольчатые листья растений пустынь
г. Все перечисленные эволюционные изменения.
12. Примером ароморфоза является:
а. Уплощение тела у донных рыб.
б. Покровительственная окраска.
в. Отсутствие кишечника у паразитических червей.
г. Возникновение полового процесса.
13. Естественный отбор – это:
а. Сложные отношения между организмами и неживой природой.
б. Процесс сохранения особей с полезными им наследственными изменениями.
в. Процесс образования новых видов в природе.
г. Процесс роста численности популяций.
14. Борьба за существование играет большую роль в эволюции:
а. Сохраняет особей преимущественно с полезными изменениями.
Сохраняет особей преимущественно с полезными изменениями.
б. Сохраняет особей с любыми наследственными изменениями.
в. Поставляет материал для отбора.
г. Обостряет взаимоотношения между особями.
15. В результате взаимодействия движущих сил эволюции происходит:
а. Размножение организмов.
б. Образование новых видов в природе.
в. Мутационный процесс.
г. Изоляция популяций.
16. Укажите неверное утверждение: «В процессе эволюции борьба с неблагоприятными условиями приводит…»
а. К повышению сопротивляемости организмов.
б. Выживанию наиболее жизненных особей.
в. Вымиранию вида.
г. Совершенствованию вида.
17. К движущим силам эволюции относят:
а. Многообразие видов.
б. Борьбу уза существование.
в. Видообразование.
г. Приспособленность.
18. Фактор эволюции, основу которого составляет возникновение преград к свободному скрещиванию особей, называют:
а. Модификацией.
б. Естественным отбором.
в. Изоляцией.
г. Популяционными волнами.
19. Среди движущих сил эволюции, ведущих к возникновению приспособлений у особей к среде обитания, направляющий характер имеет:
а. Естественный отбор.
б. Искусственный отбор.
в. Изоляция.
г. Борьба за существование.
20. Вид отбора, действующий в популяциях, обитающих в относительно постоянных условиях среды, — …:
а. Стихийный.
б. Стабилизирующий.
в. Движущий.
г. Половой.
УРОВЕНЬ Б
1. Приспособления к жизни в воде, сформировавшиеся в процессе эволюции у китов:
а. Превращение передних конечностей в ласты.
б. Дыхание кислородом, растворенным в воде.
в. Дыхание кислородом воздуха.
г. Обтекаемая форма тела.
д. Развитый толстый подкожный слой жира.
е. Постоянная температура тела.
2. Установите соответствие между парами животных и типом их взаимоотношений.
Пары животных тип взаимодействия
1. Гидра – дафния. А) паразит-хозяин
Гидра – дафния. А) паразит-хозяин
2. Рысь – заяц-беляк Б) хищник-жертва.
3. Аскарида – человек
4. Черный коршун – лесная полевка.
5. Таежный клещ – лесная мышь
6. Бычий цепень – копытное животное.
Н. С. Петухова, МОУ «СОШ №76», г. Котлас, Архангельская область
Метки: Биология
Тема №8 «эволюционное учение. Антропогенез»
АРОГЕНЕЗ – ПУТЬ БИОЛОГИЧЕСКОГО ПРОГРЕССА, СВЯЗАННЫЙ С: усложнением морфо-физиологической организации с ароморфозом |
БИОЛОГИЧЕСКАЯ (РЕПРОДУКТИВНАЯ) ИЗОЛЯЦИЯ ХАРАКТЕРИЗУЕТСЯ: нескрещиваемостью совместно обитающих организмов одного вида |
БИОЛОГИЧЕСКИЙ ПРОГРЕСС – ЭТО: возрастание приспособленности потомков по сравнению с предками |
В МАЛЫХ ПО ЧИСЛЕННОСТИ ПОПУЛЯЦИЯХ: закон Харди-Вайнберга нарушается |
В СОВРЕМЕННОЙ БИОЛОГИИ ПОПУЛЯЦИЯ РАССМАТРИВАЕТСЯ КАК: единица процесса микроэволюции |
В
ТЕОРИИ ЭВОЛЮЦИИ Ж. |
ВИД ЕСТЕСТВЕННОГО ОТБОРА, БЛАГОПРИЯТСТВУЮЩИЙ ДВУМ ИЛИ НЕСКОЛЬКИМ ПОЛЕЗНЫМ НАПРАВЛЕНИЯМ ИЗМЕЧИВОСТИ, ИСКЛЮЧАЯ ПРОМЕЖУТОЧНЫЕ СОСТОЯНИЯ ФЕНОТИПОВ, НАЗЫВАЕТСЯ: дизруптивным |
ВОЗНИКНОВЕНИЕ ИНДУСТРИАЛЬНОГО МЕЛАНИЗМА ЯВЛЯЕТСЯ ПРИМЕРОМ: преадаптивной адаптации |
ВОЗНИКНОВЕНИЕ ПЛАЦЕНТЫ У МЛЕКОПИТАЮЩИХ МОЖНО РАССМАТРИВАТЬ КАК: ценогенез |
ГЕНЕТИЧЕСКАЯ СУЩНОСТЬ ЕСТЕСТВЕННОГО ОТБОРА ЗАКЛЮЧАЕТСЯ В: избирательном сохранении в популяции определенных генотипов |
ГЕНЕТИЧЕСКИ ПОПУЛЯЦИЯ ХАРАКТЕРИЗУЕТСЯ: генофондом |
ГЕНЫ, У КОТОРЫХ В ПОПУЛЯЦИИ ОБНАРУЖИВАЮТСЯ ДВА И БОЛЕЕ АЛЛЕЛЬНЫХ ВАРИАНТА, НАЗЫВАЮТСЯ: полиморфными |
ГЕОГРАФИЧЕСКАЯ ИЗОЛЯЦИЯ СВЯЗАНА С: аллопатрическим видообразованием |
ГЕТЕРОБАТМИЯ – ЭТО: неодинаковый эволюционный уровень развития частей органа или организма |
ГИБРИДОГЕННОЕ ВИДООБРАЗОВАНИЕ ПРИВОДИТ К: слиянию двух видов с образованием нового вида |
ДАРВИН
ПРИШЕЛ К ИДЕЕ О ЕСТЕСТВННОМ ОТБОРЕ
ПОД ВЛИЯНИЕМ:
теории Т. |
ДИВЕРГЕНЦИЯ – ЭТО: возникновение различий на основе одной и той же организации |
ДИЗРУПТИВНЫЙ ОТБОР (РАЗРЫВАЮЩИЙ): отбраковывает особи со средними значениями признака |
ДЛЯ БИОЛОГИЧЕСКОГО ПРОГРЕССА ВИДА ХАРАКТЕРНО: расширение занимаемой территории |
ДЛЯ БИОЛОГИЧЕСКОГО РЕГРЕССА ВИДА ХАРАКТЕРНО: сокращение ареала |
ДЛЯ ИДЕАЛЬНОЙ ПОПУЛЯЦИИ ХАРАКТЕРНО: отсутствие эволюции |
ДРЕЙФ ГЕНОВ – ЭТО: спонтанное изменение частоты аллелей в большую или меньшую сторону |
ЕСТЕСТВЕННЫЙ ОТБОР ПО ПОЛОВЫМ ПРИЗНАКАМ ОТНОСИТСЯ К: половому |
ЖИЗНЬ
НА ЗЕМЛЕ ПОЯВИЛАСЬ ОКОЛО:
3-4 млр. |
ЗАКОН, ПОЗВОЛЯЮЩИЙ ИССЛЕДОВАТЬ ГЕНЕТИЧЕСКУЮ СТРУКТУРУ ПОПУЛЯЦИИ, БЫЛ СФОРМУЛИРОВАН: Г.Харди и В.Вайнбергом |
Истинный путь видообразованиЯ, благодаря которому появилось большинство видов организмов связан с: дивергенцией исходного вида |
ИСЧЕЗНОВЕНИЕ У ЛЕНТОЧНЫХ ЧЕРВЕЙ ПИЩЕВАРИТЕЛЬНОЙ И НЕРВНОЙ СИСТЕМ – ПРИМЕР: биологического прогресса |
К КОЛЕБАНИЯМ ЧИСЛЕННОСТИ ОСОБЕЙ В ПРИРОДНЫХ ПОПУЛЯЦИЯХ ПРИВОДЯТ: популяционные волны |
К ПРИЗНАКАМ, ХАРАКТЕРНЫМ ДЛЯ АРХАНТРОПОВ ОТНОСЯТСЯ поддержание огня, примитивная речь, состоящая из отдельных выкриков, простые формы коллективной деятельности |
К ПРИЗНАКАМ, ХАРАКТЕРНЫМ ДЛЯ НЕАНТРОПОВ ОТНОСЯТСЯ настоящая речь, мышление, искусство |
К ПРИЗНАКАМ, ХАРАКТЕРНЫМ ДЛЯ ПАЛЕАНТРОПОВ ОТНОСЯТСЯ добывание огня, сложные формы коллективной деятельности, забота о ближних, речь- продвинутая форма лепета |
К ПРИЗНАКАМ, ХАРАКТЕРНЫМ ДЛЯ ПРОАНТРОПОВ ОТНОСЯТСЯ прямохождение, изготовление орудий |
К ФОРМЕ ЕСТЕСТВЕННОГО ОТБОРА, НАПРАВЛЕННОГО НА ПОДДЕРЖАНИЕ В ПОПУЛЯЦИИ СРЕДНЕГО ЗНАЧЕНИЯ ПРИЗНАКА, ОТНОСИТСЯ: стабилизирующий |
КАКОЕ ИЗ СЛЕДУЮЩИХ ПОЛОЖЕНИЙ НЕ ВХОДИТ В СОВРЕМЕННОЕ ОПРЕДЕЛЕНИЕ ПОНЯТИЯ «ВИД»? популяция организмов обладающих внешним сходством |
КАКОЕ ПРАВИЛО ИЛЛЮСТРИРУЕТ ПРОИСХОЖДЕНИЕ МЛЕКОПИТАЮЩИХ ОТ ПРИМИТИВНЫХ ФОРМ РЕПТИЛИЙ? правило происхождения от неспециализированных предков |
КОНВЕРГЕНЦИЯ
– ЭТО:
возникновение сходных черт организации
на изначально различной основе т. |
КРИТЕРИИ БИОЛОГИЧЕСКОГО ПРОГРЕССА БЫЛИ ПРЕДЛОЖЕНЫ: А.Н. Северцовым |
КРОМАНЬОНЦЫ ОТНОСЯТСЯ К неоантропам |
КРОМАНЬОНЦЫ ПОЯВИЛИСЬ менее 40 тыс. лет назад |
МУТАЦИИ, ОБЕСПЕЧИВАЮЩИЕ ИЗМЕНЕНИЕ ФЕНОТИПА В ОПРЕДЕЛЕННОМ НАПРАВЛЕНИИ ОПРЕДЕЛЯЕТ: движущий отбор |
МУТАЦИОННЫЙ ГРУЗ ПРЕДСТАВЛЯЕТ СОБОЙ: доминантные мутации, появляющиеся в каждом поколении заново |
НА ПЕРВОЙ СТАДИИ ВИДООБРАЗОВАНИЯ: две вновь образовавшиеся популяции еще могут слиться и образовать общий генофонд |
НАИБОЛЕЕ АКТИВНО БОРЬБА ЗА СУЩЕСТВОВАНИЕ ИДЕТ: внутри популяции |
НАСЛЕДСТВЕННЫЙ ПОЛИМОРФИЗМ ЧЕЛОВЕЧЕСКИХ ПОПУЛЯЦИЙ ОПРЕДЕЛЯЕТСЯ: генетической гетерогенностью |
НЕАНДЕРТАЛЬЦЫ ОТНОСЯТСЯ К древним людям |
НЕАНДЕРТАЛЬЦЫ
ПОЯВИЛИСЬ
менее 1, 5 млн. |
ОГРАНИЧЕНИЕ ИЛИ ПОЛНОЕ ИСКЛЮЧЕНИЕ СВОБОДНОГО СКРЕЩИВАНИЯ МЕЖДУ ОСОБЯМИ ОДНОГО ВИДА изоляция |
ОРГАНИЗМЫ, ПРЕЖДЕ СПОСОБНЫЕ К ВЗАИМНОМУ СКРЕЩИВАНИЮ И ОБРАЗОВАНИЮ ПЛОДОВИТОГО ПОТОМСТВА УТРАЧИВАЮТ ЭТУ СПОСОБНОСТЬ. ПРОИЗОШЛА: репродуктивная изоляция |
Основной труд Ч. Дарвина называется: «Происхождение видов путем естественного отбора» |
ОСНОВНЫЕ СИСТЕМАТИЧЕСКИЕ ЕДИНИЦЫ РАСПОЛАГАЮТСЯ В СЛЕДУЮЩЕМ ПОРЯДКЕ: империя – царство – тип – класс – отряд – семейство — род — вид |
ОСОБЕННОСТИ СТРОЕНИЯ КОНЕЧНОСТЕЙ У ЖИВОТНЫХ, ОСОБЕННОСТИ КЛЮВА У ПТИЦ – ПРИМЕРЫ: идиоадаптации |
ОТБОР, ПРИВОДЯЩИЙ К ПОЛИМОРФИЗМУ: дизруптивный |
ОТДЕЛЕНИЕ
ВЕТВИ ГОМИНИД И ПОЯВЛЕНИЕ АВТРАЛОПИТЕКА
ПРОИЗОШЛО
более 10 млн. |
ПИТЕКАНТРОП (ЧЕЛОВЕК ПРЯМОХОДЯЩИЙ) ОТНОСЯТСЯ К древнейшим людям |
ПИТЕКАНТРОПЫ ПОЯВИЛИСЬ менее 2, 6 млн. лет назад |
ПОПУЛЯЦИИ МОГУТ ЭВОЛЮЦИОНИРОВАТЬ ПУТЕМ: все вышесказанное |
ПОПУЛЯЦИЯ – ИСТОЧНИК ВОЗНИКНОВЕНИЯ: новых видов |
ПОПУЛЯЦИЯ — ЭТО: элементарная единица эволюции |
ПРАВИЛО НЕОБРАТИМОСТИ ЭВОЛЮЦИИ ГЛАСИТ: организмы не могут вернуться к прежнему состоянию, уже осуществленному в ряду его предков |
ПРЕЗИГОТИЧЕСКИЕ И ПОСТЗИГОТИЧЕСКИЕ МЕХАНИЗМЫ У РАЗНЫХ ВИДОВ ОТНОСЯТСЯ К: генетической изоляции |
ПРИСПОСОБЛЕННОСТЬ ОРГАНИЗМОВ К СРЕДЕ ОБИТАНИЯ — РЕЗУЛЬТАТ: взаимодействия движущих сил эволюции |
Происхождение потомков от одного общего исторического предка-это явление: монофилии |
ПУТЬ ВОЗНИКНОВЕНИЯ АДАПТАЦИИ ПУТЕМ ВЗАИМОДЕЙСТВИЯ ГЕНОВ: комбинативный |
ПУТЬ ДОСТИЖЕНИЯ БИОЛОГИЧЕСКОГО ПРОГРЕССА, СВЯЗАННЫЙ С УПРОЩЕНИЕМ СТРОЕНИЯ И ФИЗИОЛОГИИ ОРГАНИЗМА: катагенез |
ПУТЬ ДОСТИЖЕНИЯ БИОЛОГИЧЕСКОГО ПРОГРЕССА, СВЯЗАННЫЙ С ЭМБРИОАДАПТАЦИЕЙ – ценогенез |
РЕЗЕРВ НАСЛЕДСТВЕННОЙ ИЗМЕНЧИВОСТИ СОСТАВЛЯЮТ: рецессивные аллели в гетерозиготном состоянии |
РЕЗУЛЬТАТ МИКРОЭВОЛЮЦИИ — ОБРАЗОВАНИЕ НОВЫХ: подвидов и видов |
СЕГРЕГАЦИОННЫЙ ГРУЗ В ПОПУЛЯЦИИ ЧЕЛОВЕКА — ЭТО: рецессивно передающиеся мутантные признаки |
СОГЛАСОВАННОСТЬ РАЗВИТИЯ РАЗНЫХ ПРИЗНАКОВ У ОСОБИ — ЭТО: коадаптация |
СООТВЕТСТВИЕ СТРОЕНИЯ АППАРАТОВ РАЗМНОЖЕНИЯ САМЦОВ И САМОК –ЭТО: конгруэнция |
СТУПЕНИ
ВОСХОЖДЕНИЯ ОТ ПРОСТОГО К СЛОЖНОМУ
(«ЗАКОН ГРАДАЦИИ») ПЕРВЫМ ПРЕДЛОЖИЛ:
Ж. |
тип видообразования основанНЫЙ на экологической изоляции: симпатрический |
ТИП ВИДООБРАЗОВАНИЯ, ХАРАКТЕРИЗУЮЩИЙСЯ ОБРАЗОВАНИЕМ ГЕОГРАФИЧЕСКИХ БАРЬЕРОВ, РАЗДЕЛЯЮЩИХ ДВЕ ПОПУЛЯЦИИ: аллопатрическое |
УСЛОВИЕМ ВЫПОЛНЕНИЯ ЗАКОНА ХАРДИ – ВАЙНБЕРГА ЯВЛЯЕТСЯ: постоянство генофонда |
ФАКТОР ЭВОЛЮЦИИ, ОСНОВУ КОТОРОГО СОСТАВЛЯЕТ ВОЗНИКНОВЕНИЕ ПРЕГРАД К СВОБОДНОМУ СКРЕЩИВАНИЮ ОСОБЕЙ, НАЗЫВАЮТ: изоляцией. |
ФАКТОРАМИ ЭВОЛЮЦИИ НЕ ЯВЛЯЮТСЯ: зародышевое сходство позвоночных |
ФИЛЕТИЧЕСКОЕ ВИДООБРАЗОВАНИЕ ПОДРАЗУМЕВАЕТ: прямое преобразование одного вида в другой |
ФОРМА ЭВОЛЮЦИИ ГРУПП, КОТОРАЯ ПРЕДСТАВЛЯЕТ СОБОЙ КОНВЕРГЕНТНОЕ РАЗВИТИЕ ГЕНЕТИЧЕСКИ БЛИЗКИХ ГРУПП ОРГАНИЗМОВ, НАЗЫВАЕТСЯ: паралелизм |
ЧТО ХАРАКТЕРНО ДЛЯ ВТОРОЙ СТАДИИ ВИДООБРАЗОВАНИЯ? появление репродуктивной изоляции в форме презиготических репродуктивно-изолирующих механизмов |
ЭЛЕМЕНТАРНАЯ ЕДИНИЦА ВИДА: популяция |
Элементарным явлением на популяционно- видовом уровне организации живой материи является: действие элементарных эволюционных факторов |
 Б.ЛАМАРКА ВЫДВИНУТА
ИДЕЯ О ТОМ ЧТО:
полезные признаки, приобретённые
особями передаются потомству
Б.ЛАМАРКА ВЫДВИНУТА
ИДЕЯ О ТОМ ЧТО:
полезные признаки, приобретённые
особями передаются потомству Р. Мальтуса о народонаселении
Р. Мальтуса о народонаселении лет назад
лет назад е.
путь развития аналогичных приспособлений
е.
путь развития аналогичных приспособлений лет назад
лет назад лет назад
лет назад Б.Ламарк
Б.Ламарк15
Механизмы эволюции
Популяции эволюционируют, а отдельные организмы — нет. Популяция — это скрещивающаяся группа особей одного вида в данной географической области в одно и то же время. Популяция развивается, потому что популяция содержит набор генов, называемый генофондом . По мере того, как происходят изменения в генофонде, популяция эволюционирует.
Популяция — это скрещивающаяся группа особей одного вида в данной географической области в одно и то же время. Популяция развивается, потому что популяция содержит набор генов, называемый генофондом . По мере того, как происходят изменения в генофонде, популяция эволюционирует.
Мутация
Мутация, движущая сила эволюции — это случайное изменение в генетическом составе организма, влияющее на генофонд популяции. Это изменение характера ДНК в одной или нескольких хромосомах. Мутации порождают новые аллели; следовательно, они являются источником генетической изменчивости в популяции.
Мутации могут быть вредными или полезными, но они также могут быть и полезными. Например, мутация может позволить организмам в популяции производить ферменты, которые позволят им использовать определенные пищевые материалы. Со временем эти типы людей выживают, в то время как те, у кого нет мутаций, с большей вероятностью погибнут. Следовательно, естественный отбор имеет тенденцию удалять менее приспособленных особей, позволяя более приспособленным особям выживать и формировать популяцию.
Следовательно, естественный отбор имеет тенденцию удалять менее приспособленных особей, позволяя более приспособленным особям выживать и формировать популяцию.
Генный поток
Другой механизм эволюции может иметь место при миграции особей из одной группы или локации в другую. Когда мигрирующие особи скрещиваются с новой популяцией, они вносят свои гены в генофонд местной популяции. Это устанавливает поток генов в популяции.
Поток генов возникает, например, когда ветер разносит семена далеко за пределы популяции родительских растений. Другой пример: животные могут быть изгнаны из стада. Это вынуждает их мигрировать в новую популяцию, тем самым внося новые гены в генофонд. Поток генов имеет тенденцию увеличивать сходство между оставшимися популяциями одного и того же вида, потому что он делает генофонды более похожими друг на друга.
Генетический дрейф
Другим механизмом эволюции является генетический дрейф, который может произойти, когда небольшая группа особей покидает популяцию и создает новую в географически изолированном регионе. Например, если в озеро поместить небольшую популяцию рыб, популяция рыб превратится в другую, отличную от исходной. Приспособленность популяции не учитывается при генетическом дрейфе, и генетический дрейф не происходит в очень большой популяции.
Например, если в озеро поместить небольшую популяцию рыб, популяция рыб превратится в другую, отличную от исходной. Приспособленность популяции не учитывается при генетическом дрейфе, и генетический дрейф не происходит в очень большой популяции.
Естественный отбор
Другим механизмом эволюции является естественный отбор, который происходит, когда популяции организмов подвергаются воздействию окружающей среды. У наиболее приспособленных существ больше шансов выжить и передать свои гены своим потомкам, создавая популяцию, которая лучше приспособлена к окружающей среде. Гены менее приспособленных людей с меньшей вероятностью будут переданы следующему поколению. Важным фактором отбора в естественном отборе является окружающая среда.
Экологичность может быть выражен несколькими способами. Например, это может включать в себя способность человека избегать хищников, это может подразумевать большую устойчивость к болезням, это может повышать способность добывать пищу или это может означать устойчивость к засухе. Пригодность также может быть измерена как повышенная репродуктивная способность, например, способность привлекать партнера. Лучше адаптированные особи производят относительно больше потомства и более успешно передают свои гены, чем менее адаптированные особи.
Пригодность также может быть измерена как повышенная репродуктивная способность, например, способность привлекать партнера. Лучше адаптированные особи производят относительно больше потомства и более успешно передают свои гены, чем менее адаптированные особи.
По-видимому, на популяции влияет несколько типов естественного отбора. Один тип, стабилизирующий отбор, происходит, когда окружающая среда отбирает организмы популяции с экстремальными версиями признака. Другой вид естественного отбора — подрывной отбор. Здесь среда благоприятствует крайним типам в популяции за счет промежуточных форм, тем самым разделяя популяцию на две или более субпопуляции. Третий тип естественного отбора — это направленный отбор. В этом случае среда выбирает экстремальную характеристику. Развитие устойчивых к антибиотикам бактерий в современную эпоху является примером направленного отбора.
Развитие видов
вида — это группа особей, которые имеют ряд общих признаков и способны скрещиваться друг с другом, производя плодовитое (нестерильное) потомство. (Когда особи одного вида спариваются с особями другого вида, любое потомство обычно бесплодно.) Вид также определяется как популяция, представители которой имеют общий генофонд.
(Когда особи одного вида спариваются с особями другого вида, любое потомство обычно бесплодно.) Вид также определяется как популяция, представители которой имеют общий генофонд.
Эволюция вида — это видообразование, которое может произойти, когда популяция изолирована географическими барьерами, как это произошло в изоляции Австралии, Новой Зеландии и Галапагосских островов. Разнообразие форм жизни, встречающихся только в Австралии, но нигде больше, является примером видообразования за счет географических барьеров.
Видообразование также может происходить при развитии репродуктивных барьеров. Например, когда у членов популяции развиваются анатомические барьеры, которые затрудняют спаривание с другими членами популяции, может развиться новый вид. Время начала половой жизни — еще один пример репродуктивного барьера. Пространственные различия, например, когда один вид обитает на верхушках деревьев, а другой вид живет на уровне земли, являются еще одной причиной развития видов.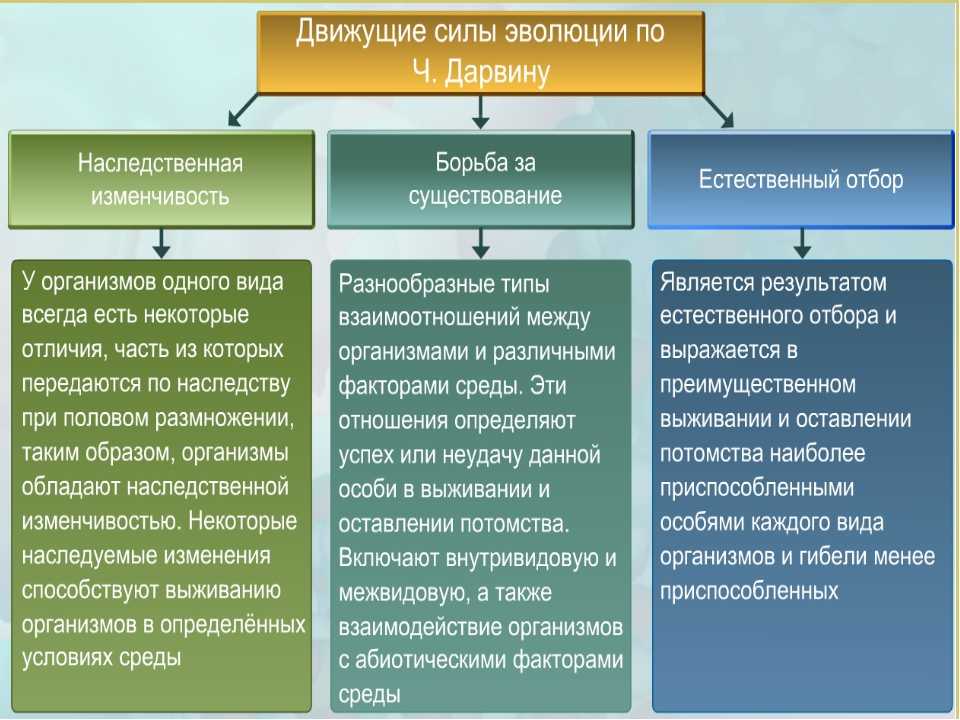
Постепенное или быстрое изменение
Теория Дарвина включала в себя наблюдение, что эволюционные изменения происходят медленно. Во многих случаях летопись окаменелостей показывает, что виды менялись постепенно с течением времени. Теория о том, что эволюция происходит постепенно, известна как градуализм .
Противоположностью градуализму является теория прерывистого равновесия, которая является предметом обсуждения среди ученых. Согласно теории прерывистого равновесия, некоторые виды имеют длительные стабильные периоды существования, прерываемые относительно короткими периодами быстрых изменений.
Обе группы ученых согласны с тем, что естественный отбор является самым важным фактором эволюционных изменений видов. Независимо от того, являются ли изменения медленными и постепенными или точечными и быстрыми, одно можно сказать наверняка: организмы эволюционировали с течением времени.
География эволюции и эволюция географии | Evolution: Education and Outreach
Volume 5 Supplement 4
Biogeography: The Geography of Evolution
- Geography and Evolution
- Открытый доступ
- Опубликовано:
- Брюс С. Либерман 1
Эволюция: образование и распространение информации том 5 , страницы 521–525 (2012 г.)Процитировать эту статью
18k обращений
2 Цитаты
Сведения о показателях
Abstract
Понимание географии жизни сыграло фундаментальную роль в мотивации крупных разработок в области эволюционной биологии. Основное внимание здесь уделяется изложению некоторых из этих основных событий, особенно в контексте палеонтологии, путем подчеркивания значения географической изоляции и аллопатрического видообразования, прерывистого равновесия и гипотезы пульса оборота для эволюционной теории. Один из основных споров в области эволюции касается относительного вклада абиотических и биотических факторов в макроэволюцию, и каждое из этих событий все больше предполагало, что именно климатические и геологические факторы, а не конкуренция, играли основную роль в мотивации макроэволюции. Новые технические разработки, в том числе в области географических информационных систем, позволяют продолжать детальное тестирование относительных ролей, которые биотические факторы играют в противоположность абиотическим факторам в процессе эволюции, и будут также описаны некоторые работы в этой области.
Один из основных споров в области эволюции касается относительного вклада абиотических и биотических факторов в макроэволюцию, и каждое из этих событий все больше предполагало, что именно климатические и геологические факторы, а не конкуренция, играли основную роль в мотивации макроэволюции. Новые технические разработки, в том числе в области географических информационных систем, позволяют продолжать детальное тестирование относительных ролей, которые биотические факторы играют в противоположность абиотическим факторам в процессе эволюции, и будут также описаны некоторые работы в этой области.
Введение
Чарльз Лайель (1832 г.) утверждал актуальность информации о распределении организмов для областей биологии и геологии, и он сослался на известное изречение «Как в пространстве, так и во времени». Здесь представлены аргументы в пользу расширения этого изречения, чтобы охватить как биогеографию, так и палеонтологию, «Пространство и время». Биогеография — это дисциплина, занимающаяся реконструкцией истории биоты, и под биотами понимаются группы видов, встречающихся в определенных регионах. Эта дисциплина имеет долгую и обширную историю, уходящую корнями далеко в восемнадцатый век, и биогеографические исследования сыграли ключевую роль в развитии эволюции, особенно в работах Уоллеса и Дарвина (Брукс и МакЛеннан 19).91; Либерман 2000; Морроне 2008; Уайли и Либерман, 2011). Это исследование было особенно актуальным, поскольку в нем рассматривалось, как дифференциация видов в географическом пространстве трансформировалась в эволюционную дифференциацию во времени. Сам Дарвин во введении к Происхождению видов утверждает, что именно биогеографические закономерности и закономерности в палеонтологической летописи убедили его в том, что эволюция имела место, а роль географии и палеонтологии в развитии идей Дарвина была подробно рассмотрена. на страницах этого журнала (например, Eldredge 2009а, б), так что это не будет здесь основным акцентом. Вместо этого основное внимание будет уделено некоторым ключевым достижениям эволюционной теории после Дарвина (1859 г.), особенно в области макроэволюции, а также тому, как они включали более тщательную и правильную интеграцию географии в эволюцию посредством анализа закономерностей, сохранившихся в окаменелостях.
Эта дисциплина имеет долгую и обширную историю, уходящую корнями далеко в восемнадцатый век, и биогеографические исследования сыграли ключевую роль в развитии эволюции, особенно в работах Уоллеса и Дарвина (Брукс и МакЛеннан 19).91; Либерман 2000; Морроне 2008; Уайли и Либерман, 2011). Это исследование было особенно актуальным, поскольку в нем рассматривалось, как дифференциация видов в географическом пространстве трансформировалась в эволюционную дифференциацию во времени. Сам Дарвин во введении к Происхождению видов утверждает, что именно биогеографические закономерности и закономерности в палеонтологической летописи убедили его в том, что эволюция имела место, а роль географии и палеонтологии в развитии идей Дарвина была подробно рассмотрена. на страницах этого журнала (например, Eldredge 2009а, б), так что это не будет здесь основным акцентом. Вместо этого основное внимание будет уделено некоторым ключевым достижениям эволюционной теории после Дарвина (1859 г.), особенно в области макроэволюции, а также тому, как они включали более тщательную и правильную интеграцию географии в эволюцию посредством анализа закономерностей, сохранившихся в окаменелостях. рекорд (Либерман 2008). Тем не менее, некоторые краткие сведения о роли биогеографических паттернов в убеждении Дарвина в том, что эволюция имела место, заслуживают внимания. В частности, во время путешествия Дарвина на борту HMS Beagle , он признал существование очень похожих видов, разделенных географическими барьерами. Например, в Южной Америке есть два вида крупных нелетающих птиц, или нанду. Один встречается к северу от большой реки в Аргентине, Рио-Негро, тогда как другой вид встречается к югу от реки. Дарвин узнал о других примерах на Галапагосских островах, включая пересмешников и черепах. Хотя он не утверждал эволюционного значения этого в своей книге «Путешествие на «Бигле» 9».0096 (Дарвин, 1839 г.), он сделал это в дарвиновских записных книжках (Саллоуэй, 1979; Барретт и др., 1987), где утверждал, что географические барьеры могут служить для изоляции популяций видов, что ведет к дифференциации.
рекорд (Либерман 2008). Тем не менее, некоторые краткие сведения о роли биогеографических паттернов в убеждении Дарвина в том, что эволюция имела место, заслуживают внимания. В частности, во время путешествия Дарвина на борту HMS Beagle , он признал существование очень похожих видов, разделенных географическими барьерами. Например, в Южной Америке есть два вида крупных нелетающих птиц, или нанду. Один встречается к северу от большой реки в Аргентине, Рио-Негро, тогда как другой вид встречается к югу от реки. Дарвин узнал о других примерах на Галапагосских островах, включая пересмешников и черепах. Хотя он не утверждал эволюционного значения этого в своей книге «Путешествие на «Бигле» 9».0096 (Дарвин, 1839 г.), он сделал это в дарвиновских записных книжках (Саллоуэй, 1979; Барретт и др., 1987), где утверждал, что географические барьеры могут служить для изоляции популяций видов, что ведет к дифференциации.
Сегодня мы называем этот вид видообразования географическим или аллопатрическим видообразованием. Этот стиль видообразования особенно важен для эволюционной теории, потому что если видообразование преимущественно аллопатрическое, то силы, создающие географические барьеры, играют большую роль в эволюции. Эти силы в основном связаны с эпизодами геологических и климатических изменений. Примечательно, что явления, сходные с аллопатрическим видообразованием, были описаны еще до путешествия Дарвина на 9-м0121 Бигль ; например, Леопольдом фон Бухом в 1825 году на основе его работы на Канарских островах. Тем не менее, Дарвин, наряду с Уоллесом, был одним из первых, кто синтезировал эту закономерность и осознал ее значение (Lieberman 2000). Интересно, что, хотя эта закономерность помогла Дарвину убедить в том, что эволюция имела место, позже он стал приуменьшать значение аллопатрического видообразования, особенно у Дарвина (1859). Там он считал, что к образованию новых видов привела не географическая изоляция, а прежде всего конкуренция. Его акцент сместился на опору на то, что сегодня называется симпатрическим видообразованием для объяснения диверсификации, и, таким образом, на представление о том, что эволюция управляется конкуренцией, а не изменением климата и геологией (Lieberman 2008).
Этот стиль видообразования особенно важен для эволюционной теории, потому что если видообразование преимущественно аллопатрическое, то силы, создающие географические барьеры, играют большую роль в эволюции. Эти силы в основном связаны с эпизодами геологических и климатических изменений. Примечательно, что явления, сходные с аллопатрическим видообразованием, были описаны еще до путешествия Дарвина на 9-м0121 Бигль ; например, Леопольдом фон Бухом в 1825 году на основе его работы на Канарских островах. Тем не менее, Дарвин, наряду с Уоллесом, был одним из первых, кто синтезировал эту закономерность и осознал ее значение (Lieberman 2000). Интересно, что, хотя эта закономерность помогла Дарвину убедить в том, что эволюция имела место, позже он стал приуменьшать значение аллопатрического видообразования, особенно у Дарвина (1859). Там он считал, что к образованию новых видов привела не географическая изоляция, а прежде всего конкуренция. Его акцент сместился на опору на то, что сегодня называется симпатрическим видообразованием для объяснения диверсификации, и, таким образом, на представление о том, что эволюция управляется конкуренцией, а не изменением климата и геологией (Lieberman 2008).
Возвращение географии в эволюцию
После публикации Дарвина (1859 г.) в течение длительного времени большинство ученых были склонны приуменьшать значение роли аллопатрического видообразования, хотя некоторые, включая Гулика (1872 г.), Вагнера (1873 г.) и Джордана (1908) продолжали настаивать на его значении, так что климатические и геологические факторы обычно не рассматривались в качестве основных факторов, побуждающих к видообразованию. Так продолжалось до тех пор, пока Эрнст Майр не задокументировал вездесущность аллопатрического видообразования, например, Майр (1942, 1963), что аллопатрическое видообразование вернулось на первый план. В настоящее время большинство неонтологических и палеонтологических исследований продолжают повторять представление о том, что видообразование является трансцендентально аллопатрическим (например, Eldredge, 1971; Eldredge and Gould, 1972; Wiley and Mayden, 1985; Eldredge, 1989; Brooks and McLennan, 1991; Lieberman, 2000; Coyne and Orr, 2004; Rode). и Либерман, 2005; Морроне, 2008; Стигал, 2010). Хотя Майр действительно осознал центральное значение аллопатрического видообразования, потребовалось некоторое время, прежде чем его полное макроэволюционное значение было признано.
и Либерман, 2005; Морроне, 2008; Стигал, 2010). Хотя Майр действительно осознал центральное значение аллопатрического видообразования, потребовалось некоторое время, прежде чем его полное макроэволюционное значение было признано.
Прерывистое равновесие
Элдридж (1971), а затем Элдридж и Гулд (1972) провели исследование и сделали теоретические выводы, которые привели к признанию значения аллопатрического видообразования для палеонтологии и макроэволюционной теории. В каком-то смысле именно реинтеграция надлежащей роли географии в эволюционную теорию в целом и видообразование в частности позволила им сделать ключевые предсказания о природе видов и видообразовании в летописи окаменелостей. Они признали, что большинство видов на протяжении всей своей истории, часто на протяжении многих миллионов лет, были стабильными и демонстрировали то, что они назвали застоем (Элдридж 19).85). Этот застой не обязательно должен быть стойким, в морфологии могут быть тонкие колебания (Lieberman et al. , 1995; Eldredge et al., 2005), но превалирует общая стабильность сети. Кроме того, эволюционные изменения были сосредоточены в относительно коротких с геологической точки зрения интервалах, которые могли составлять от 5000 до 50 000 лет. Основываясь на применении аллопатрической модели, они пришли к выводу, что изменения в летописи окаменелостей будут казаться относительно внезапными и, кроме того, обычно будут происходить в узких периферийных средах вдоль границ ареала вида. Таким образом, обычно было бы трудно зафиксировать видообразование в действии в летописи окаменелостей. Вместо этого появление нового вида в летописи окаменелостей обычно представляло собой миграцию популяции, развившейся в результате аллопатрии. Напротив, широко распространенные, многочисленные виды, виды, которые, скорее всего, сохранятся в летописи окаменелостей, должны демонстрировать незначительные согласованные изменения. Паттерн прерывистого равновесия не только характеризовал большую часть видов, сохранившихся в летописи окаменелостей, но также сильно противоречил дарвиновскому представлению о том, что виды должны изменяться медленно и постепенно в течение миллионов лет во всем своем ареале (Элдридж и Гулд 19).
, 1995; Eldredge et al., 2005), но превалирует общая стабильность сети. Кроме того, эволюционные изменения были сосредоточены в относительно коротких с геологической точки зрения интервалах, которые могли составлять от 5000 до 50 000 лет. Основываясь на применении аллопатрической модели, они пришли к выводу, что изменения в летописи окаменелостей будут казаться относительно внезапными и, кроме того, обычно будут происходить в узких периферийных средах вдоль границ ареала вида. Таким образом, обычно было бы трудно зафиксировать видообразование в действии в летописи окаменелостей. Вместо этого появление нового вида в летописи окаменелостей обычно представляло собой миграцию популяции, развившейся в результате аллопатрии. Напротив, широко распространенные, многочисленные виды, виды, которые, скорее всего, сохранятся в летописи окаменелостей, должны демонстрировать незначительные согласованные изменения. Паттерн прерывистого равновесия не только характеризовал большую часть видов, сохранившихся в летописи окаменелостей, но также сильно противоречил дарвиновскому представлению о том, что виды должны изменяться медленно и постепенно в течение миллионов лет во всем своем ареале (Элдридж и Гулд 19).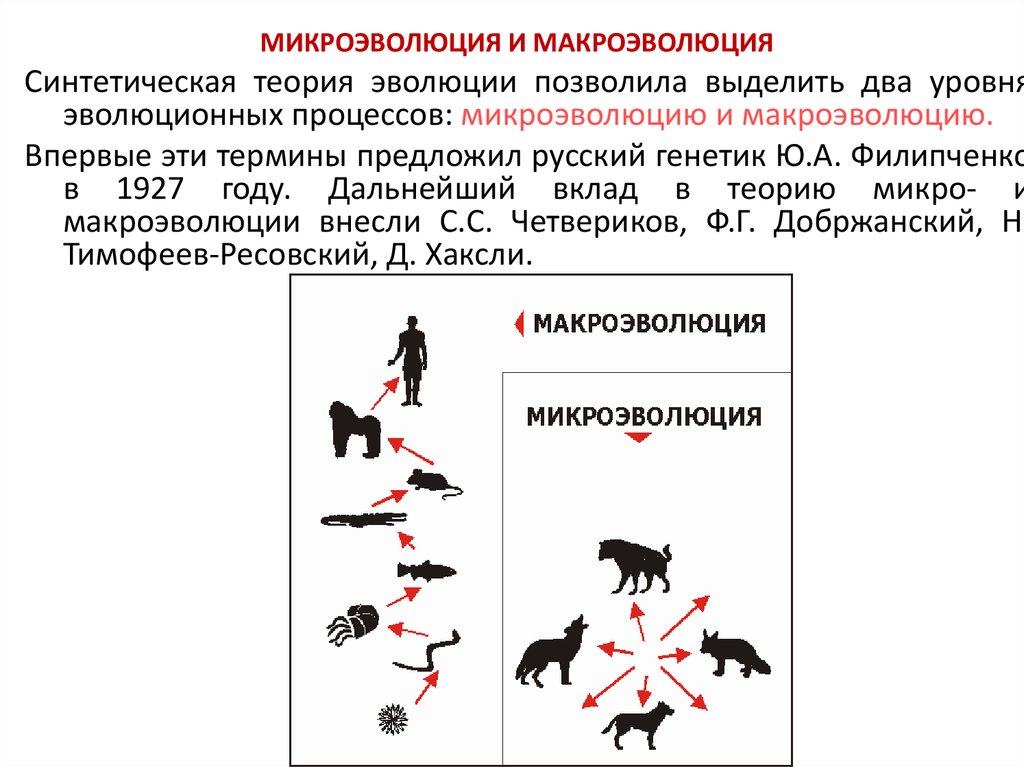 72; Элдридж, 1985). С одной стороны, развитие прерывистого равновесия, безусловно, во многом зависело от правильного включения географии в эволюцию, но, конечно же, оно также зависело от более буквального прочтения летописи окаменелостей, чем пропагандировал Дарвин. Одним из основных моментов Дарвина (1859 г.) является то, что летопись окаменелостей слишком неполна, чтобы подробно изучать эволюцию. Симпсон (1944) и Элдридж и Гулд (1972) справедливо отвергли эту точку зрения.
72; Элдридж, 1985). С одной стороны, развитие прерывистого равновесия, безусловно, во многом зависело от правильного включения географии в эволюцию, но, конечно же, оно также зависело от более буквального прочтения летописи окаменелостей, чем пропагандировал Дарвин. Одним из основных моментов Дарвина (1859 г.) является то, что летопись окаменелостей слишком неполна, чтобы подробно изучать эволюцию. Симпсон (1944) и Элдридж и Гулд (1972) справедливо отвергли эту точку зрения.
Гипотеза пульса товарооборота
Другим крупным нововведением в макроэволюционной теории стала разработка Элизабет Врба гипотезы пульса оборота (Врба, 1980, 1985, 1992). Элдридж (1971) и Элдридж и Гулд (1972) в своей формулировке прерывистого равновесия действительно опирались на аллопатрическое видообразование, но они специально сосредоточились на обсуждении Майром (1942, 1963) концепции, согласно которой географическая изоляция возникает, когда популяция вида рассеивается по территории. ранее существовавший географический барьер. Это явно важный способ аллопатрического видообразования (например, Wiley and Mayden 19).85; Брукс и Макленнан, 1991 г.; Либерман 2000; Стигал 2010; Уайли и Либерман, 2011). Однако, когда аллопатрическое видообразование происходит таким образом, оно обычно одновременно включает только один вид; и далее, произойдет видообразование или нет, зависит в значительной степени от индивидуальных экологических характеристик организмов вида. Новаторство Врбы (1980, 1985, 1992) с гипотезой пульса оборота заключалось в том, что она признала важность того факта, что в периоды изменения климата географические ареалы многих видов будут затронуты одновременно. Особое внимание она уделила неогеновым видам тропических млекопитающих, которые в теплое время могли иметь широкий географический ареал. Когда температура понизится, ареалы станут более узкими, а популяции многих видов изолируются в небольших рефугиумах. Некоторые из этих видов столкнулись бы с таким резким сокращением ареалов, что вымерли бы (фаза оборота ее гипотезы).
Это явно важный способ аллопатрического видообразования (например, Wiley and Mayden 19).85; Брукс и Макленнан, 1991 г.; Либерман 2000; Стигал 2010; Уайли и Либерман, 2011). Однако, когда аллопатрическое видообразование происходит таким образом, оно обычно одновременно включает только один вид; и далее, произойдет видообразование или нет, зависит в значительной степени от индивидуальных экологических характеристик организмов вида. Новаторство Врбы (1980, 1985, 1992) с гипотезой пульса оборота заключалось в том, что она признала важность того факта, что в периоды изменения климата географические ареалы многих видов будут затронуты одновременно. Особое внимание она уделила неогеновым видам тропических млекопитающих, которые в теплое время могли иметь широкий географический ареал. Когда температура понизится, ареалы станут более узкими, а популяции многих видов изолируются в небольших рефугиумах. Некоторые из этих видов столкнулись бы с таким резким сокращением ареалов, что вымерли бы (фаза оборота ее гипотезы). Однако те популяции, которые сохранялись и оставались изолированными в этих убежищах достаточно долго, дивергировали и в конечном итоге видообразовали (Врба 19).80, 1985, 1992) (импульсная фаза ее гипотезы). Таким образом, несколько новых видов в одном и том же регионе возникнут примерно в одно и то же время. Кроме того, эти события видообразования будут вызваны непосредственно изменением климата. Гипотеза импульса товарооборота предоставила средства расширения прерывистого равновесия от случая одного вида до случая нескольких видов.
Однако те популяции, которые сохранялись и оставались изолированными в этих убежищах достаточно долго, дивергировали и в конечном итоге видообразовали (Врба 19).80, 1985, 1992) (импульсная фаза ее гипотезы). Таким образом, несколько новых видов в одном и том же регионе возникнут примерно в одно и то же время. Кроме того, эти события видообразования будут вызваны непосредственно изменением климата. Гипотеза импульса товарооборота предоставила средства расширения прерывистого равновесия от случая одного вида до случая нескольких видов.
Работа Врбы тесно связана с разработками в области биогеографии наместничества. В викариантной биогеографии акцент делается на использовании филогении, чтобы увидеть, демонстрируют ли группы видов сходные или «конгруэнтные» модели эволюции в географическом пространстве (Wiley 19).81; Брукс и Макленнан, 1991 г.; Либерман 2000; Морроне 2008; Уайли и Либерман, 2011). Ценность этого подхода заключается в том, что он делал упор на разработку методов строгого анализа биогеографических закономерностей с использованием филогении. Затем, если сходные модели эволюции обнаруживаются в географическом пространстве, это предполагает, что изменения в геологии или климате мотивировали макроэволюционные модели в группе. Если, напротив, разные группы видов, встречающихся в одном и том же регионе, демонстрируют разные модели эволюции в географическом пространстве, это предполагает, что индивидуальные экологические характеристики видов сыграли основную роль в мотивации диверсификации. Таким образом, викариантная биогеография предоставляет средства для непосредственного рассмотрения и проверки относительных ролей, которые геология и климат играют как процессы, мотивирующие макроэволюцию. Врба (1980, 1985, 1992) существенно расширил викариантную биогеографию непосредственно в область эволюционной теории, сосредоточив внимание на значительной роли, которую изменение климата играет в возникновении эволюции посредством механизмов географической изоляции и аллопатрического видообразования.
Затем, если сходные модели эволюции обнаруживаются в географическом пространстве, это предполагает, что изменения в геологии или климате мотивировали макроэволюционные модели в группе. Если, напротив, разные группы видов, встречающихся в одном и том же регионе, демонстрируют разные модели эволюции в географическом пространстве, это предполагает, что индивидуальные экологические характеристики видов сыграли основную роль в мотивации диверсификации. Таким образом, викариантная биогеография предоставляет средства для непосредственного рассмотрения и проверки относительных ролей, которые геология и климат играют как процессы, мотивирующие макроэволюцию. Врба (1980, 1985, 1992) существенно расширил викариантную биогеографию непосредственно в область эволюционной теории, сосредоточив внимание на значительной роли, которую изменение климата играет в возникновении эволюции посредством механизмов географической изоляции и аллопатрического видообразования.
Количественная оценка географического диапазона для получения информации о макроэволюции
Новое понимание макроэволюции, основанное на анализе биогеографических закономерностей, сохранившихся в летописи окаменелостей, также стало возможным благодаря применению географических информационных систем (ГИС). Эти методы позволяют точно и многократно вычислять географический ареал отдельных видов, используя информацию о местонахождении. Этот подход может быть применен к современным видам, а также к ископаемым видам. Что касается ископаемых видов, первые распределения окаменелостей из различных местонахождений наносятся на современную карту в ГИС. Затем с помощью программы PaleoGIS (Ross and Scotese 2000) континенты можно повернуть обратно в положение, в котором они когда-то были, когда эти ископаемые таксоны действительно жили (рис. 1). Одним из примеров, когда такие методы доказали свою ценность, является демонстрация роли, которую инвазивные виды сыграли в возникновении кризиса биоразнообразия в позднем девоне (Роде и Либерман, 2004, 2005; Стигал, 2010). Другой пример, когда ГИС доказала свою ценность в анализе географических данных по окаменелостям, — это исследования роли конкуренции в макроэволюции (Майерс и Либерман, 2011). Дарвин (1859 г.), Докинз (1976) и многие другие утверждали, что конкуренция представляет собой значительную эволюционную силу, однако свидетельства палеонтологической летописи всегда были ограничены (Гулд и Кэллоуэй, 1980; Бентон, 1996).
Эти методы позволяют точно и многократно вычислять географический ареал отдельных видов, используя информацию о местонахождении. Этот подход может быть применен к современным видам, а также к ископаемым видам. Что касается ископаемых видов, первые распределения окаменелостей из различных местонахождений наносятся на современную карту в ГИС. Затем с помощью программы PaleoGIS (Ross and Scotese 2000) континенты можно повернуть обратно в положение, в котором они когда-то были, когда эти ископаемые таксоны действительно жили (рис. 1). Одним из примеров, когда такие методы доказали свою ценность, является демонстрация роли, которую инвазивные виды сыграли в возникновении кризиса биоразнообразия в позднем девоне (Роде и Либерман, 2004, 2005; Стигал, 2010). Другой пример, когда ГИС доказала свою ценность в анализе географических данных по окаменелостям, — это исследования роли конкуренции в макроэволюции (Майерс и Либерман, 2011). Дарвин (1859 г.), Докинз (1976) и многие другие утверждали, что конкуренция представляет собой значительную эволюционную силу, однако свидетельства палеонтологической летописи всегда были ограничены (Гулд и Кэллоуэй, 1980; Бентон, 1996). ГИС предоставляет отличные средства для рассмотрения того, как географические ареалы видов взаимодействуют друг с другом во времени. Майерс и Либерман (2011) использовали ГИС для рассмотрения географических ареалов нескольких ископаемых видов морских позвоночных из мелового Западного внутреннего морского пути, большого, в основном тропического морского пути, проходящего через центр Северной Америки (рис. 1), который является источником многочисленных обширных ископаемых морских месторождений. Майерс и Либерман (2011) специально сосредоточились на том, были ли какие-либо доказательства конкурентного замещения: явления, когда один вид появляется в пределах географического ареала другого вида, его ареал расширяется, и это медленно и постепенно приводит к исчезновению других видов. Они не нашли статистических доказательств конкурентной замены. Вместо этого в меловом периоде Западного внутреннего морского пути изменения в физической среде, по-видимому, были основными факторами, контролирующими изменения в географическом распространении во времени, а также основными факторами, вызывающими вымирание видов (рис.
ГИС предоставляет отличные средства для рассмотрения того, как географические ареалы видов взаимодействуют друг с другом во времени. Майерс и Либерман (2011) использовали ГИС для рассмотрения географических ареалов нескольких ископаемых видов морских позвоночных из мелового Западного внутреннего морского пути, большого, в основном тропического морского пути, проходящего через центр Северной Америки (рис. 1), который является источником многочисленных обширных ископаемых морских месторождений. Майерс и Либерман (2011) специально сосредоточились на том, были ли какие-либо доказательства конкурентного замещения: явления, когда один вид появляется в пределах географического ареала другого вида, его ареал расширяется, и это медленно и постепенно приводит к исчезновению других видов. Они не нашли статистических доказательств конкурентной замены. Вместо этого в меловом периоде Западного внутреннего морского пути изменения в физической среде, по-видимому, были основными факторами, контролирующими изменения в географическом распространении во времени, а также основными факторами, вызывающими вымирание видов (рис.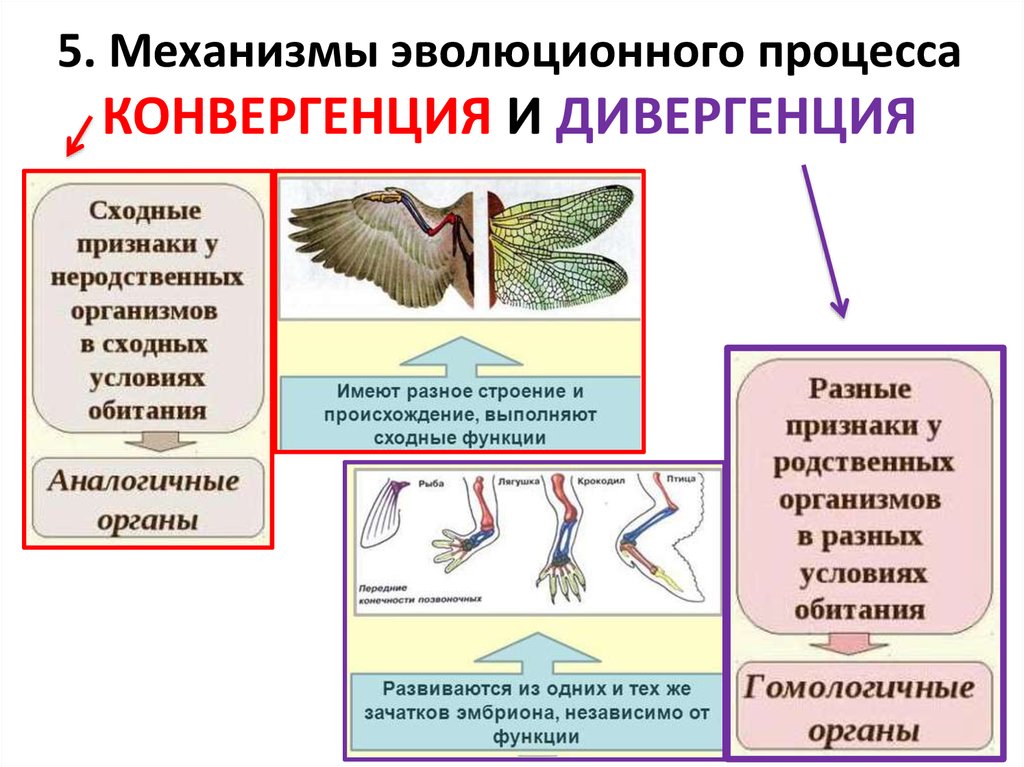 2). Здесь ГИС подтвердила важную роль окружающей среды во влиянии на макроэволюцию и, соответственно, более ограниченную роль конкуренции на этой арене.
2). Здесь ГИС подтвердила важную роль окружающей среды во влиянии на макроэволюцию и, соответственно, более ограниченную роль конкуренции на этой арене.
Записи о встречаемости некоторых позднемеловых морских позвоночных, акулы ( светло-серый ) и мозазавра ( темно-серый ), показанные на карте современного мира ( a ) и мелового периода world ( b ), из Myers and Lieberman (2011), используется с разрешения
Изображение в полный размер
Рис. 2Реконструкция ареалов морских позвоночных позднемелового периода, показывающая, как распределение видов обычно меняется во времени. Более темные многоугольники показывают ареалы мозазавра Platecarpus для разных стадий ( a – d ) позднего мелового периода. Более светлые полигоны показывают диапазоны для мозазавра Тилозавра . Эти диапазоны меняются со временем, но не показывают статистической связи друг с другом, а вместо этого кажутся связанными с изменениями в окружающей среде, из Майерса и Либермана (2011), используется с разрешения
Полноразмерное изображение
Выводы
Закономерности, относящиеся к географическому распространению организмов, и закономерности в летописи окаменелостей сыграли фундаментальную роль в убеждении Дарвина и других ученых в том, что эволюция имела место, и они до сих пор играют важную роль в обеспечении понимания эволюционного процесса. Выдающиеся достижения, способствующие прогрессу в области биогеографии со времен Дарвина, включают более глубокое понимание видообразования, открытия в области палеонтологии и макроэволюционной теории, а также разработку новых методов анализа биогеографических данных, включая филогенетические подходы и подходы с использованием ГИС. Одной из методологий, основанных на ГИС, которая считается особенно важной для будущих биогеографических открытий в области палеонтологии, является моделирование экологических ниш. Этот подход можно использовать для рассмотрения множества интересных биогеографических вопросов, включая экологические изменения при видообразовании (Петерсон и др. 19).99), какие абиотические и биотические факторы контролируют ареалы видов (Stigall and Lieberman 2006; Maguire and Stigall 2009), а также как видовые ниши меняются с течением времени (Dudei and Stigall 2010). Интеграция этих новых методологических разработок с важными теоретическими открытиями открывает большие перспективы для будущего биогеографии и намечает путь, соединяющий прошлые и будущие открытия.
Выдающиеся достижения, способствующие прогрессу в области биогеографии со времен Дарвина, включают более глубокое понимание видообразования, открытия в области палеонтологии и макроэволюционной теории, а также разработку новых методов анализа биогеографических данных, включая филогенетические подходы и подходы с использованием ГИС. Одной из методологий, основанных на ГИС, которая считается особенно важной для будущих биогеографических открытий в области палеонтологии, является моделирование экологических ниш. Этот подход можно использовать для рассмотрения множества интересных биогеографических вопросов, включая экологические изменения при видообразовании (Петерсон и др. 19).99), какие абиотические и биотические факторы контролируют ареалы видов (Stigall and Lieberman 2006; Maguire and Stigall 2009), а также как видовые ниши меняются с течением времени (Dudei and Stigall 2010). Интеграция этих новых методологических разработок с важными теоретическими открытиями открывает большие перспективы для будущего биогеографии и намечает путь, соединяющий прошлые и будущие открытия.
Ссылки
Барретт П.Х., Готри П.Дж., Герберт С., Кон Д., Смит С. Записные книжки Чарльза Дарвина, 1836–1844 гг. Итака: Издательство Корнельского университета; 1987.
Google Scholar
Бентон М.Дж. Проверка роли конкуренции и экспансии в эволюции четвероногих. Proc R Soc Lond B Biol Sci. 1996; 263: 641–6.
Артикул Google Scholar
Брукс Д.Р., Макленнан Д.А. Филогения, экология и поведение. Чикаго: Издательство Чикагского университета; 1991.
Google Scholar
Coyne JA, Orr HA. Спецификация. Сандерленд: Синауэр; 2004.
Google Scholar
Дарвин К. Путешествие на «Бигле». Нью-Йорк: Пингвин; 1839.
Google Scholar
Дарвин К.
 О происхождении видов. Лондон: Джон Мюррей; 1859.
О происхождении видов. Лондон: Джон Мюррей; 1859.Google Scholar
Докинз Р. Эгоистичный ген. Нью-Йорк: Издательство Оксфордского университета; 1976.
Google Scholar
Dudei NL, Stigall AL. Подход к моделированию экологической ниши для оценки биогеографической реакции видов брахиопод во время ричмондского (позднего ордовика) вторжения в арку Цинциннати. Палеогеогр Палеоклиматол Палеоэкол. 2010; 296:28–43.
Артикул Google Scholar
Элдридж Н. Аллопатрическая модель и филогения палеозойских беспозвоночных. Эволюция. 1971;25:156–67.
Артикул Google Scholar
Элдридж Н. Временные рамки. Принстон: Издательство Принстонского университета; 1985.
Google Scholar
Элдридж Н.
Макроэволюционная динамика. Нью-Йорк: Макгроу Хилл; 1989.Google Scholar
Элдридж Н. Эксперименты с трансмутацией: Дарвин, бигль и эволюция. Evo Edu Outreach. 2009 г.а; 2:35–54.
Артикул Google Scholar
Элдридж Н. Вопрос индивидуальности: Чарльз Дарвин, Джордж Гейлорд Симпсон и переходные окаменелости. Evo Edu Outreach. 2009б; 2:150–5.
Артикул Google Scholar
Элдридж Н., Гулд С.Дж. Прерывистое равновесие: альтернатива филетическому градуализму. В: Schopf TJM, редактор. Модели в палеобиологии. Сан-Франциско: Фримен; 1972. с. 82–115.
Google Scholar
Элдридж Н., Томпсон Дж., Брейкфилд П., Гаврилец С., Яблонски Д., Джексон Дж., Ленски Р., Либерман Б.С., Макпик М., Миллер III В. Динамика эволюционного застоя. Палеобиология. 2005; 31: 133–45.
Артикул Google Scholar
Гулд С.Дж., Кэллоуэй С.Б. Моллюски и брахиоподы: корабли, проплывающие ночью. Палеобиология. 1980; 6: 383–9.6.
Google Scholar
Гулик Дж.Т. Об изменчивости видов в зависимости от их географического распространения на примере ахатинеллид. Природа. 1872; 6: 222–4.
Артикул Google Scholar
Иордания ДС. Закон близнецовых видов. Я Нат. 1908; 42: 73–80.
Артикул Google Scholar
Либерман Б.С. Палеобиогеография: использование окаменелостей для изучения глобальных изменений, тектоники плит и эволюции. Нью-Йорк: Kluwer Academic Press; 2000.
Книга Google Scholar
Либерман Б.С. Новые синтезы между палеобиогеографией и макроэволюционной теорией.
Прок Рой Сок Виктория. 2008;120(1):51–7.Google Scholar
Либерман Б.С., Бретт К.Э., Элдридж Н. Паттерны и процессы застоя в линиях двух видов среднего девона штата Нью-Йорк. Палеобиология. 1995;21:15–27.
Google Scholar
Лайель К. Основы геологии, том. 2. 2-е изд. Чикаго: Издательство Чикагского университета; 1832.
Google Scholar
Магуайр К.С., Стигал А.Л. Распространение ископаемых лошадей на Великих равнинах в миоцене и плиоцене: подход к моделированию экологической ниши. Палеобиология. 2009; 35: 597–611.
Артикул Google Scholar
Майр Э. Систематика и происхождение видов. Нью-Йорк: издательство Колумбийского университета; 1942 г. репринтное издание.
Google Scholar
Майр Э.
Виды животных и эволюция. Кембридж: Издательство Гарвардского университета; 1963.Книга Google Scholar
Морроне Дж.Дж. Эволюционная биогеография: комплексный подход с изучением конкретных случаев. Нью-Йорк: издательство Колумбийского университета; 2008.
Google Scholar
Майерс С., Либерман Б.С. Акулы, проплывающие ночью: использование ГИС для исследования конкуренции в меловом Западном внутреннем морском пути. Proc R Soc Lond B Biol Sci. 2011; 278 (1706): 681–9.
Артикул Google Scholar
Петерсон А.Т., Соберон Дж., Санчес-Кордеро В. Консерватизм экологических ниш в эволюционном времени. Наука. 1999; 285:1265–7.
Артикул КАС пабмед Google Scholar
Роде А.Л., Либерман Б.С. Использование ГИС для изучения биогеографии позднедевонского кризиса биоразнообразия.
Палеогеогр Палеоклиматол Палеоэкол. 2004; 211:345–59.Артикул Google Scholar
Роде А.Л., Либерман Б.С. Интеграция биогеографии и эволюции с использованием филогенетики и ПалеоГИС: тематическое исследование девонских ракообразных. Дж Палеонтол. 2005;79: 267–76.
Артикул Google Scholar
Росс М.И., Скотез ЧР. ПалеоГИС/Arcview 3.5, проект PALEOMAP. Арлингтон: Техасский университет; 2000.
Google Scholar
Симпсон Г.Г. Темп и лад в эволюции. переиздание изд. Нью-Йорк: издательство Колумбийского университета; 1944.
Google Scholar
Стигал А.Л. Сокращение видообразования во время позднедевонского кризиса биоразнообразия, связанного с инвазиями видов. ПЛОС Один. 2010;5(12):e15584.
Артикул КАС пабмед Google Scholar
Стигал А.
Л., Либерман Б.С. Количественная палеобиогеография: ГИС, филогенетический биогеографический анализ и идеи сохранения. J Биогеогр. 2006;33:2051–60.Артикул Google Scholar
Саллоуэй Ф.Дж. Географическая изоляция в мышлении Дарвина: превратности ключевой идеи. В: Coleman W, Limoges C, редакторы. Исследования по истории биологии. Балтимор: Издательство Университета Джона Хопкинса; 1979. с. 23–65.
Google Scholar
Врба ЭС. Эволюция, виды и окаменелости: как развивается жизнь? S Afr J Sci. 1980; 76: 61–84.
Google Scholar
Врба ЭС. Окружающая среда и эволюция: альтернативные причины временного распределения эволюционных событий. S Afr J Sci. 1985; 81: 229–36.
Google Scholar
Врба ЭС. Млекопитающие как ключ к эволюционной теории. J Млекопитающее.
1992; 73:1–28.Артикул Google Scholar
Вагнер М. Дарвиновская теория и закон миграции организмов. Лондон: Э. Стэнфорд; 1873 г. (перевод с немецкой версии 1868 г., опубликованной Duncker and Humblot, Лейпциг).
Google Scholar
Wiley EO. Филогенетика: теория и практика филогенетической систематики. Нью-Йорк: Уайли; 1981.
Google Scholar
Wiley EO, Lieberman BS. Филогенетика: теория и практика филогенетической систематики. 2-е изд. Нью-Йорк: Уайли; 2011.
Книга Google Scholar
Wiley EO, Mayden RL. Виды и видообразование в филогенетической систематике на примере ихтиофауны Северной Америки. Энн Мо Бот Гард. 1985; 72: 596–635.
Артикул Google Scholar
 О происхождении видов. Лондон: Джон Мюррей; 1859.
О происхождении видов. Лондон: Джон Мюррей; 1859. Макроэволюционная динамика. Нью-Йорк: Макгроу Хилл; 1989.
Макроэволюционная динамика. Нью-Йорк: Макгроу Хилл; 1989.
 Прок Рой Сок Виктория. 2008;120(1):51–7.
Прок Рой Сок Виктория. 2008;120(1):51–7. Виды животных и эволюция. Кембридж: Издательство Гарвардского университета; 1963.
Виды животных и эволюция. Кембридж: Издательство Гарвардского университета; 1963. Палеогеогр Палеоклиматол Палеоэкол. 2004; 211:345–59.
Палеогеогр Палеоклиматол Палеоэкол. 2004; 211:345–59. Л., Либерман Б.С. Количественная палеобиогеография: ГИС, филогенетический биогеографический анализ и идеи сохранения. J Биогеогр. 2006;33:2051–60.
Л., Либерман Б.С. Количественная палеобиогеография: ГИС, филогенетический биогеографический анализ и идеи сохранения. J Биогеогр. 2006;33:2051–60. 1992; 73:1–28.
1992; 73:1–28.Скачать ссылки
Благодарности
Я благодарю Найлса Элдреджа за приглашение подготовить специальный выпуск «География и эволюция» и Коринн Майерс за помощь с цифрами.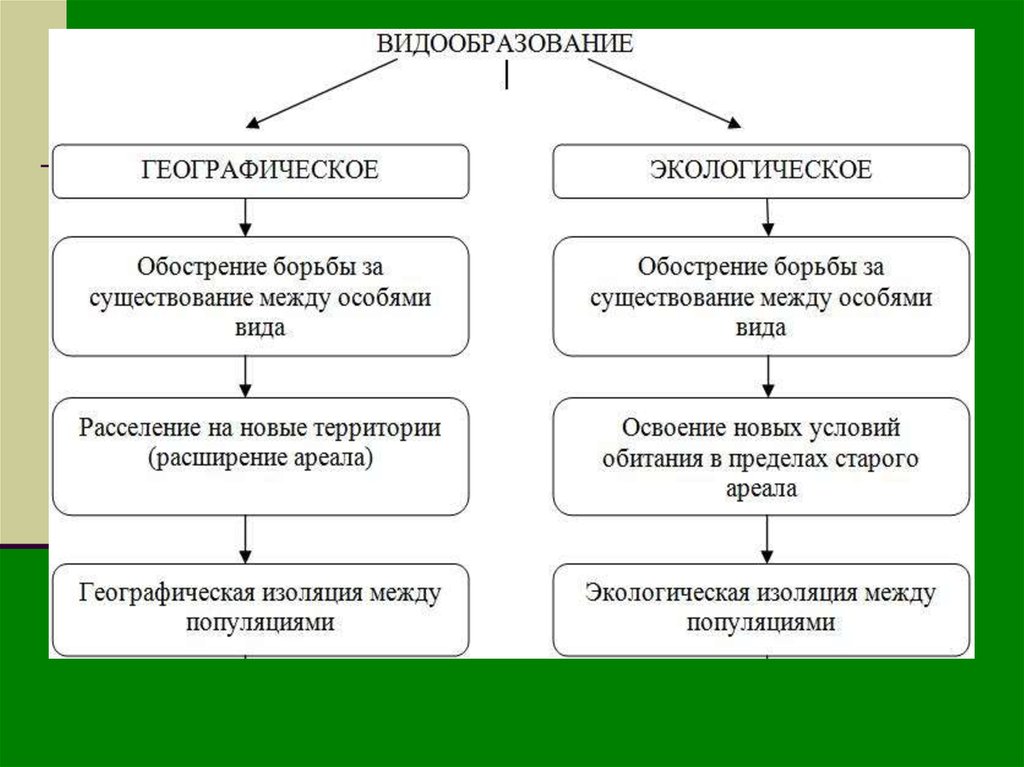
Leave A Comment