
Семенные растения — Умскул Учебник
На этой странице вы узнаете- Семя vs спора: какое размножение эффективнее и почему?
- Почему название отдела Голосеменные имеет тот же корень, что и слово “голый”?
- Как семенные растения смогли стать практически независимыми от влажности среды?
Вспомните любое культурное растение: картофель, пшеница, морковь, лук… Все они относятся к семенным. Хорошо, а какие деревья мы увидим, если пойдём по грибы в лес? Ель, сосну, березу, — и это тоже семенные растения. Нигде от них не укрыться: эта группа занимает господствующее положение в современной флоре Земли. А почему — вы узнаете при прочтении этой статьи.
Общая характеристикаК семенным растениям относятся представители отделов
- Голосеменные,
- Покрытосеменные.
Для них характерен ряд признаков:
- Размножение семенами
Это намного эффективнее, чем размножение спорами.
| Семя vs спора: какое размножение эффективнее и почему? Семя — многоклеточный орган, который имеет в составе зародыш, эндосперм и семенную кожуру. Эндосперм — запас питательных веществ и воды для зародыша. Для того, чтобы начать получать эти вещества из почвы, зародышу необходимо сначала “отрастить” корешок и листья, но для этого тоже необходима энергия. Её зародыш получает из эндосперма. Тот играет роль своеобразного “холодильника с продуктами”, который заботливо оставила родительская особь. Семенная кожура защищает зародыш от воздействия неблагоприятных факторов среды. Даже если семя съест какое-нибудь животное, то оно вполне может беспрепятственно пройти через пищеварительные пути и остаться жизнеспособным! Зато спор у растения обычно образуется гораздо больше, чем семян. А еще за счёт одноклеточности споры маленькие и лёгкие, а значит, быстрее распространяются ветром на большие расстояния. |

- Преобладающее поколение — спорофит
Гаметофит (половое поколение) созревает на спорофите (бесполом поколении) в период полового размножения. Чтобы лучше разобраться с понятиями “гаметофит” и “спорофит”, читайте статью «Споровые растения».
Женский гаметофит семенных называется “зародышевым мешком”, а мужской — “пыльцевым зерном”.
- Разноспоровость
Все семенные растения разноспоровые. Это означает, что существуют женская и мужская споры. Женская называется макроспорой, так как она очень крупная. Мужская спора маленькая, поэтому её называют микроспорой.
- Половой процесс не зависит от воды
| Как семенные растения смогли стать практически независимыми от влажности среды? У них есть особый артефакт, который помогает им проходить уровень игры “размножение” с бОльшим комфортом, чем другим растениям. Теперь гаметам необязательно попадать в воду, чтобы встретиться — оплодотворение происходит в женском гаметофите — зародышевом мешке! |
 Это пыльцевая трубка.
Это пыльцевая трубка.Пыльцевое зерно состоит из вегетативной и генеративной клеток. Всё, что называется “генеративным”, в биологии отвечает за половое размножение. Так и запомним: вегетативная клетка прорастает в пыльцевую трубку, а генеративная делится с образованием двух спермиев.
Как же все эти ароморфозы повлияли на жизнь семенных растений?
Все эти признаки позволяют голо- и покрытосеменным растениям произрастать даже на тех территориях, где обитание споровых невозможно!
Например, они населяют холодные и аридные районы. Семенные растения представлены всем многообразием форм: среди них есть травы, кустарники, лианы, деревья.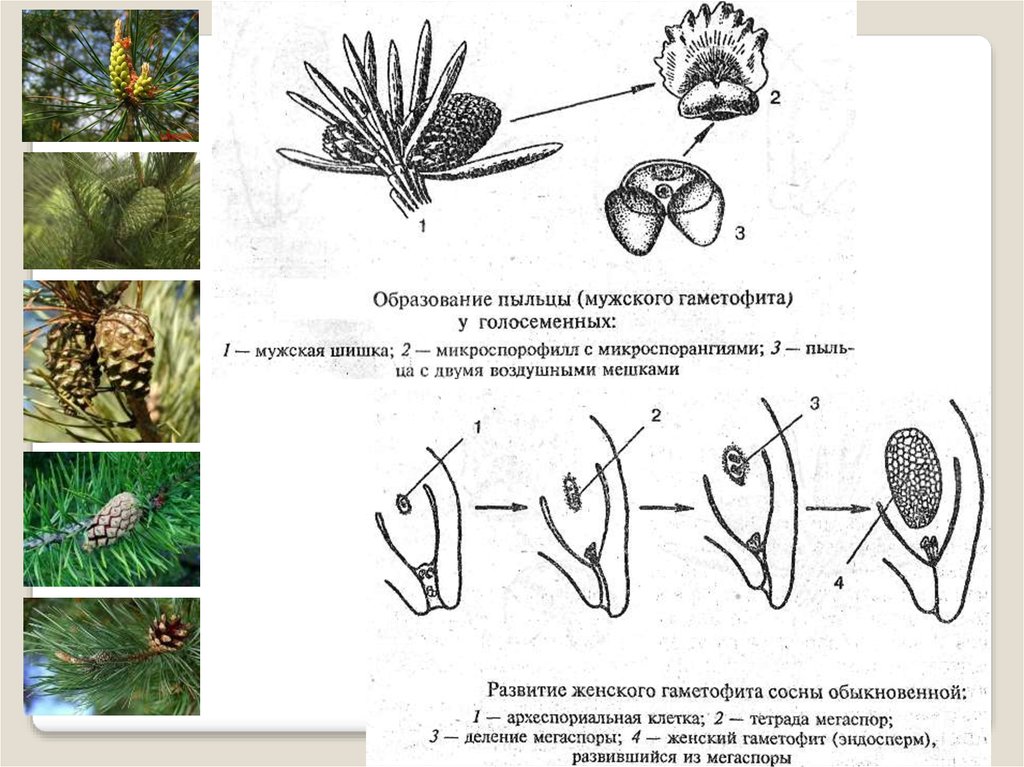
Семенные папоротники — это очень древние растения. Именно ими питались в основном динозавры палеозоя. Современные семенные растения произошли именно от них.
Семенные папоротники — предки современных семенных растений.
Это группа вымерших в Палеозойской эре растений, которые обладали признаками как папоротников, так и голосеменных.
Представители: медуллоза Ноэ, алетоптерис, калиматотека.
Признаки папоротников
- Вайи — видоизмененные перисто-рассеченные листья.
- Отсутствие шишек. Семена у семенных папоротников формировались на нижней стороне листа, как споры у папоротников.
Признаки голосеменных
- Размножение семенами, в которых до сих пор не найдены зародыши. Отсутствие зародыша, констатированное при изучении многих сотен ископаемых семян этих растений, до сих пор представляется загадкой.

- Многообразие форм. Семенные папоротники были представлены в основном древесными и лиановидными растениями.

Голосеменные — отдел современных семенных растений, включающий более 1100 видов.
| Почему название отдела Голосеменные имеет тот же корень, что и слово “голый”? Группа названа так, потому что семена этих растений лежат открыто (“голо”) на шишках и не защищены цветком или плодом, в отличие от покрытосеменных. |
Общая характеристика
Голосеменные не имеют настоящих сосудов. Их ксилема (проводящая ткань) представлена только трахеидами, что осложняет восходящий ток минеральных веществ по растению. Но несмотря на это, большая часть голосеменных — высокие древесные растения.
Большинство представителей — многолетние вечнозелёные растения. Ель, сосна, лиственница — типичные представители этого отдела. Они не теряют листья зимой и постоянно остаются зелёными. Кстати для самого растения это не очень хорошо: в листьях за год накапливается большое количество токсинов, которые приходится перерабатывать и обезвреживать.
Они не теряют листья зимой и постоянно остаются зелёными. Кстати для самого растения это не очень хорошо: в листьях за год накапливается большое количество токсинов, которые приходится перерабатывать и обезвреживать.
Классификация и представители
Отдел Голосеменные растения включает классы
- Хвойные,
- Гинкговые,
- Гнетовые
- и Саговниковые.
- Класс Хвойные
Представители: ель, сосна, лиственница, кипарис.
Этот класс наиболее многочислен. Представители группы имеют видоизмененные игольчатые (как у ели и сосны) или чешуевидные (как у кипариса) листья.
| Важно запомнить! Хвойные растения не бывают травянистыми, они представлены древесными и кустарниковыми формами. Видимо, когда-то травянистые хвойные растения (или их предки) существовали, но их вытеснила группа покрытосеменных растений, у которых гораздо больше приспособлений к жизни в суровом первом ярусе растительного сообщества. |

Древесина развита очень хорошо, в ней присутствует густая сеть смоляных ходов. Хвойные растения выделяют токсические соединения — фитонциды.
Фитонциды — вещества, обладающие лекарственными свойствами для человека.
Фитонцидные лекарственные препараты обладают антибактериальными, противовирусными и противогрибковыми свойствами и оказывают иммуностимулирующее действие. Например, популярное средство от насморка «Пиносол» имеет в составе пихтовое масло.
- Класс Гинкговые
Единственным представителем этого класса в современной флоре Земли является Гинкго двулопастной, который растет только в некоторых районах Восточной Азии. Такие виды, как Гинкго двулопастной, считаются реликтовыми.
Реликты — живые свидетельства существования почти вымершей группы организмов.
Листья Гинкго имеют необычную форму и напоминают вывернутый зонтик:
Экстракт гинкго применяется в медицине как биологически активная добавка (БАД).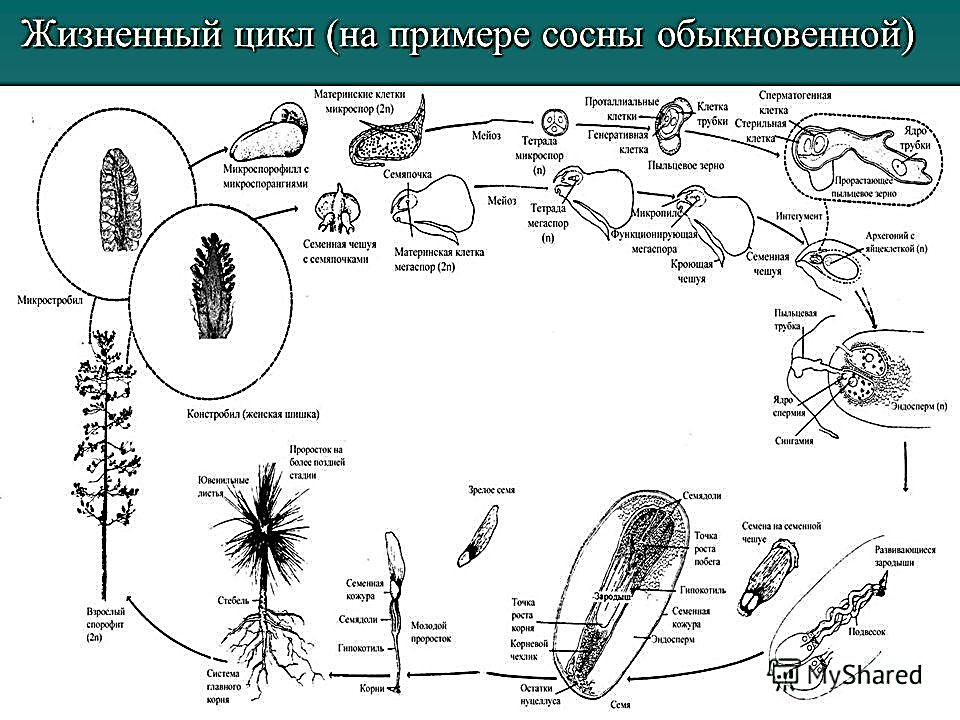 Препарат, получаемый из него используют для улучшения мозгового кровообращения.
Препарат, получаемый из него используют для улучшения мозгового кровообращения.
- Класс Гнетовые
К группе относятся вельвичия, гнетум и эфедра.
Гнетум — тропическая лиана.
Эфедра — кустарник, имеющий чешуевидные листья и произрастающий в пустынях.
Вельвичия — единственный представитель семейства Вельвичиевые. Растение выглядит как подземный побег с выходящими на поверхность длинными массивными листьями.
| Интересный факт! Одно растение вельвичии может расти больше 1000 лет. |
- Класс Саговниковые
Представители этой группы — растения с мощной сердцевиной и слабо развитой древесиной. Для саговниковых характерны неветвящиеся стебли. Некоторые роды саговниковых считаются эндемиками.
Эндемики — организмы, живущие только в определённых ареалах и не встречающиеся на других территориях.
Например, представители рода Саговники обитают только в Африке.
Размножение и развитие
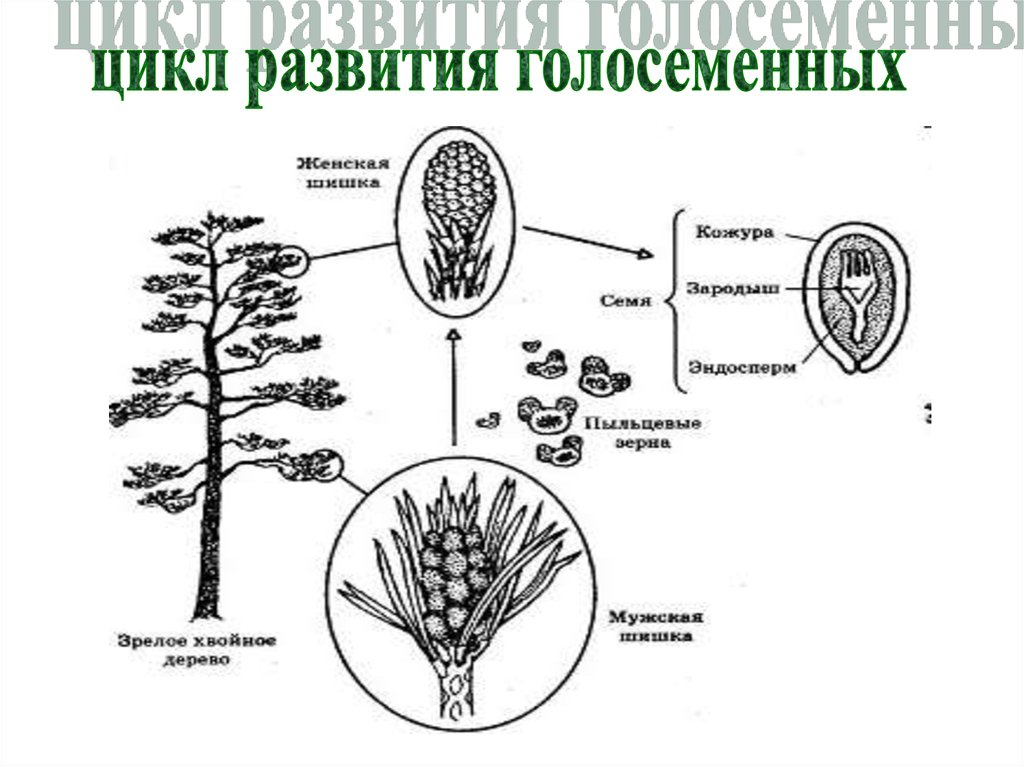
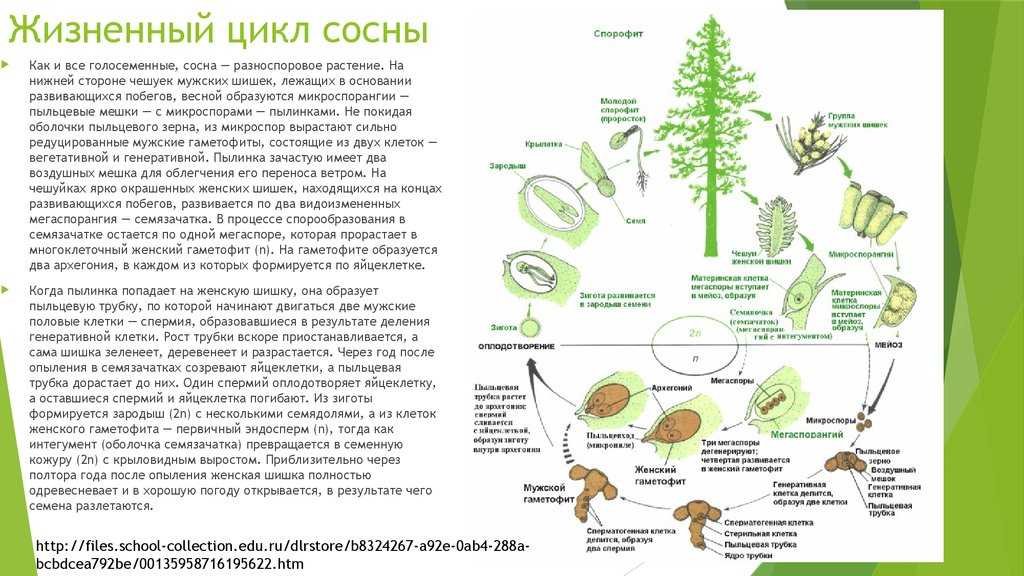
Взрослое листостебельное растение сосны (представителя отдела Голосеменные) представлено спорофитом.
- На нём в период полового созревания вырастают шишки.
- В них формируются спорангии. Подробнее об этих органах вы можете прочитать в статье «Споровые растения».
- Женская спора делится митозом и образует зародышевый мешок, мужская — пыльцевое зерно.
- При опылении ветром пыльцевое зерно попадает на женскую шишку.
- Из его вегетативной клетки прорастает пыльцевая трубка. Генеративная клетка делится митозом на 2 спермия.
- По пыльцевой трубке спермии попадают в зародышевый мешок, в котором уже сформировались 2 женские гаметы — яйцеклетки. Происходит оплодотворение, формируется зигота.
| Важно знать! У голосеменных сливается только одна пара гамет (спермий + яйцеклетка), вторая пара погибает — она лизируется, и содержащиеся в ней питательные вещества остаются в семязачатке. |

- Из зиготы путем нескольких митотических делений образуется зародыш.
- Семена распространяются ветром, попадают в субстрат и прорастают во взрослое растение — спорофит.
Анемохория — распространение семян ветром.
Значение для экосистем
- производят намного больше кислорода, чем покрытосеменные, за счёт увеличенной суммарной площади поверхности листьев;
- вытесняют лиственные деревья из лесов.
Значение для человека
- используются для изготовления бумаги и ценной древесины,
- производства эфирных масел;
- в медицине для получения лекарственных препаратов.
Покрытосеменные — отдел современных семенных растений, насчитывающий около 272 тысяч видов.
Среди цветковых есть и травы, и кустарники, и деревья, и лианы.
Отличительные признаки покрытосеменных растений
- Наличие цветка и плода
Цветок и плод защищают зародыш, “покрывая” его, именно за счет этого покрытосеменные получили такое название.
Мужские цветки имеют тычинки, женские — пестики.
Пестики и тычинки — это генеративные органы покрытосеменных.
Пестик состоит из завязи, столбика и рыльца.
Тычинка — из тычиночной нити и пыльника.
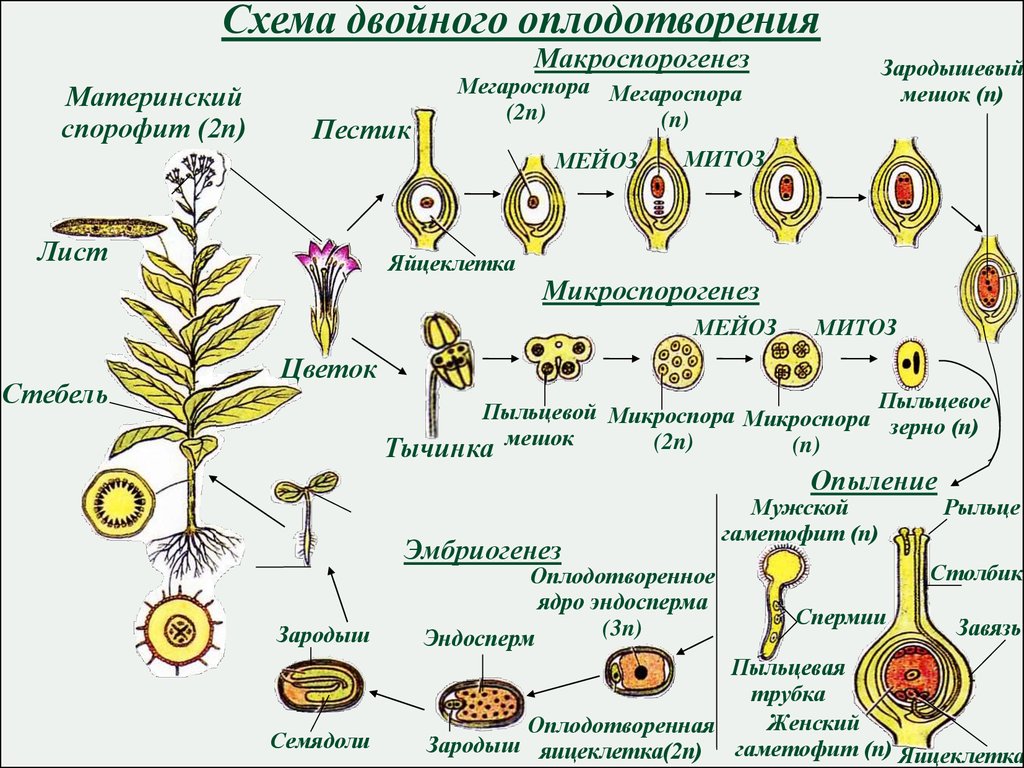
- Двойное оплодотворение
При оплодотворении голосеменных сливается только одна пара гамет (спермий + яйцеклетка).
У покрытосеменных один спермий сливается с гаплоидной яйцеклеткой, образуя зиготу, а второй — с диплоидной центральной клеткой, формируя триплоидный (2n + n = 3n) эндосперм.
- Триплоидный эндосперм
Это усложнение строения семени очень выгодно. В три раза увеличенный набор хромосом позволяет синтезу белка в эндосперме происходить быстрее. А это значит, что у зародыша будет гораздо больше питательных веществ в запасе, и он сможет переживать неблагоприятные условия дольше.
В три раза увеличенный набор хромосом позволяет синтезу белка в эндосперме происходить быстрее. А это значит, что у зародыша будет гораздо больше питательных веществ в запасе, и он сможет переживать неблагоприятные условия дольше.
Значение для человека
- Розы, тюльпаны, нарциссы используются людьми как декоративные растения. Их мы дарим на праздники близким, выращиваем в садах и огородах.
- Многие цветковые, например, ромашка, лаванда используются в качестве сырья для производства лекарственных препаратов и бытовой химии (гелей для душа, шампуней).
- Картофель, томат, лук — эти и многие другие цветковые растения люди употребляют в пищу.
Подробнее о представителях различных классов покрытосеменных вы можете прочитать в статье «Классы и семейства покрытосеменных».
Фактчек- К семенным растениям относятся представители отделов Голосеменные и Покрытосеменные.
- Размножаются семенные растения семенами, что намного эффективнее размножения спорами.
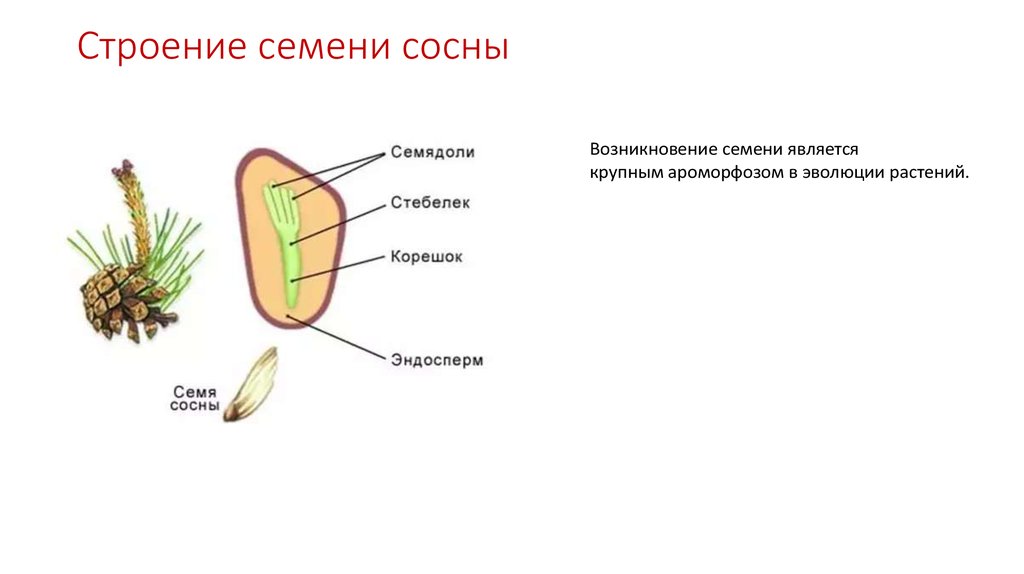
- Семя — многоклеточный орган, имеющий в составе зародыш, эндосперм и семенную кожуру.
- Преобладающее поколение семенных — спорофит.
- Женский гаметофит семенных называется “зародышевым мешком”, а мужской — “пыльцевым зерном”.
Задание 1.
Чем цветковые растения отличаются от голосеменных?
- они являются разноспоровыми
- преобладающее поколение — спорофит
- они имеют цветки и плоды
- при оплодотворении сливается только одна пара гамет
Задание 2.
Семенные папоротники…
- широко распространены на планете
- обитают в северных районах
- являются вымершей группой растений
- на данный момент представлены только одним родом
Задание 3.
Класс Гинкговые насчитывает…
- один вид
- три рода
- большое количество видов
- около десяти родов растений
Задание 4.
Кипарис относится к…
- покрытосеменным
- голосеменным
- папоротниковидным
- моховидным
Задание 5.
Какой признак папоротниковидных характерен для медуллозы Ноэ?
- наличие корня
- размножение семенами
- размножение спорами
- наличие видоизмененных листьев — вай
Задание 6.
Какой набор хромосом у эндосперма лилии?
- гаплоидный
- диплоидный
- триплоидный
- полиплоидный
Ответы: 1 — 3; 2 — 3; 3 — 1; 4 — 2; 5 — 4; 6 — 3.
ТерминыАридные районы — засушливые территории пустынь и полупустынь.
Решение задач ЕГЭ по жизненному циклу растений
Цели занятия: повторение и обобщение материала по разделу “Жизненные циклы растений”; обучение решению задач части С5 ЕГЭ по жизненным циклам растений разных отделов.
Форма занятия: лекционно-практическая.
Оборудование: проектор, слайды, набор карточек с задачами.
Ход занятия
Лекция
Понятие о жизненном цикле растений
В жизненном цикле растений происходит чередование бесполого и полового размножения и связанное с этим чередований поколений.
Гаплоидный (n) растительный организм, образующий гаметы, называется гаметофитом (n). Он представляет половое поколение. Гаметы формируются в половых органах путём митоза: сперматозоиды (n) — в антеридиях (n), яйцеклетки (n) – в архегониях (n) .
Гаметофиты бывают обоеполые (на нём развиваются антеридии и архегонии) и раздельнополые (антеридии и архегонии развиваются на разных растениях).
После слияния гамет (n) образуется зигота с
диплоидным набором хромосом (2n), а из неё
развивается путём митоза бесполое поколение –
спорофит (2n). В специальных органах — спорангиях
(2n) спорофита (2n) после мейоза образуются
гаплоидные споры (n), при делении которых митозом
развиваются новые гаметофиты (n).
Жизненный цикл зелёных водорослей
В жизненном цикле зелёных водорослей преобладает гаметофит (n), то есть клетки их слоевища гаплоидны (n). При наступлении неблагоприятных условий (похолодание, пересыхание водоёма) происходит половое размножение – образуются гаметы (n), которые попарно сливаются в зиготу (2n). Зигота (2n), покрытая оболочкой зимует, после чего при наступлении благоприятных условий делится мейозом с образованием гаплоидных спор (n), из которых развиваются новые особи (n). (Демонстрация слайдов).
Схема 1. Жизненный цикл зелёных водорослей. (Приложение)
Практикум
Задача 1. Какой набор хромосом характерен для клеток слоевища улотрикса и для его гамет? Объясните, из каких исходных клеток и в результате, какого деления они образуются.
Ответ:
1. В клетках слоевища гаплоидный набор хромосом
(n), они развиваются из споры с гаплоидным набором
хромосом (n) путём митоза.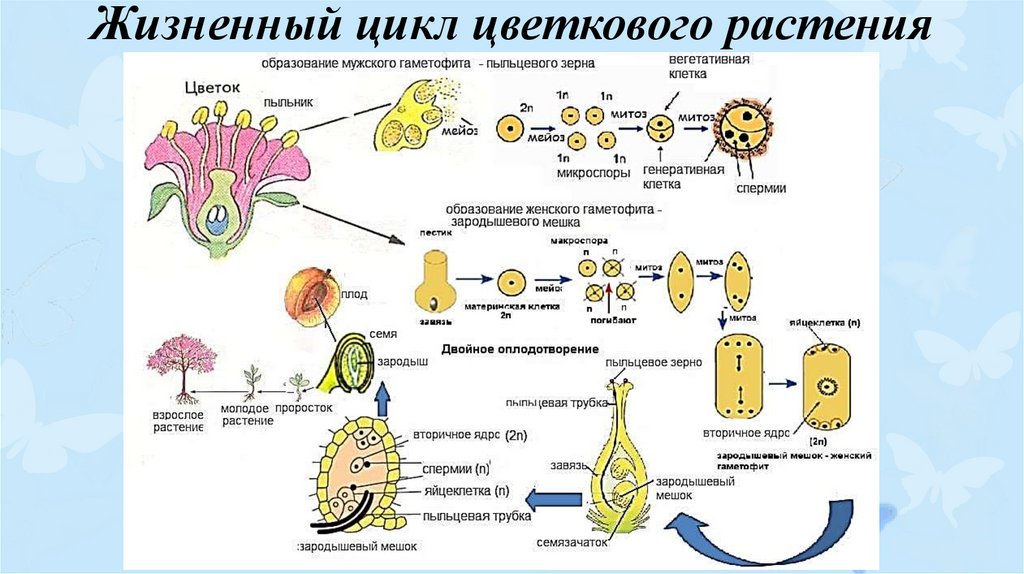
2. В гаметах гаплоидный набор хромосом (n), они образуются из клеток слоевища с гаплоидным набором хромосом (n) путём митоза.
Задача 2. Какой набор хромосом характерен для зиготы и для спор зелёных водорослей? Объясните, из каких исходных клеток и как они образуются.
Ответ:
1. В зиготе диплоидный набор хромосом (2n), она образуется при слиянии гамет с гаплоидным набором хромосом (n).
2. В спорах гаплоидный набор хромосом (n), они образуются из зиготы с диплоидным набором хромосом (2n) путём мейоза.
Лекция
Жизненный цикл мхов (кукушкин лён)
У мхов в цикле развития преобладает половое
поколение (n). Листостебельные растения мхов –
раздельнополые гаметофиты (n). На мужских
растениях (n) формируются антеридии (n) со
сперматозоидами (n), на женских (n) – архегонии (n) с
яйцеклетками (n). С помощью воды (во время дождя)
сперматозоиды (n) попадают к яйцеклеткам (n),
происходит оплодотворение, возникает зигота (2n).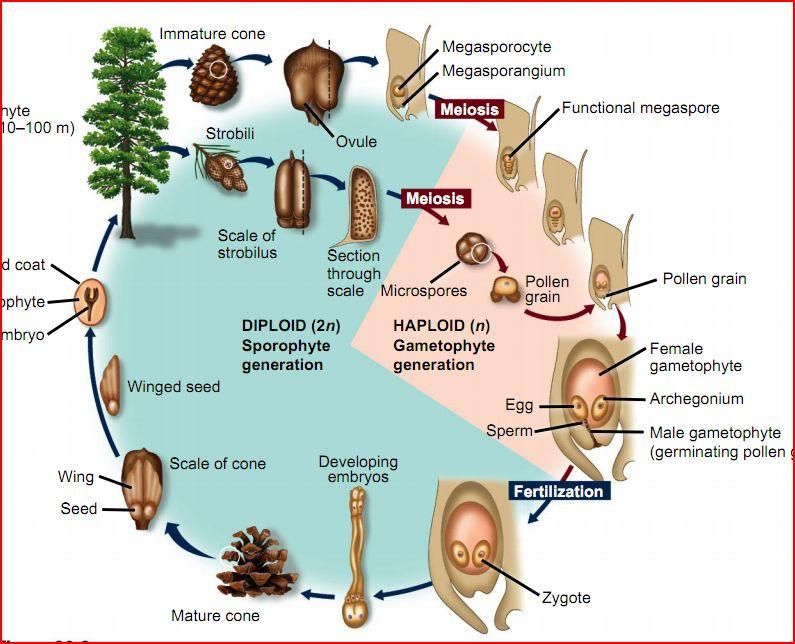
В коробочке спорофита (2n) путём мейоза образуются споры (n). Мхи – разноспоровые растения, различают микроспоры – мужские и макроспоры – женские. Из спор (n) путём митоза развиваются сначала предростки, а затем взрослые растения (n). (Демонстрация слайдов).
Схема 2. Жизненный цикл мха (кукушкин лён)
Практикум
Задача 3. Какой хромосомный набор характерен для гамет и спор кукушкина льна? Объясните, из каких исходных клеток и в результате, какого деления они образуются.
Ответ:
1. В гаметах мха кукушкина льна гаплоидный набор
хромосом (n), они образуются из антеридиев (n) и
архегониев (n) мужского и женского гаметофитов с
гаплоидным набором хромосом (n) путём митоза.
2. В спорах гаплоидный набор хромосом (n), они образуются из клеток спорофита — коробочки на ножке с диплоидным набором хромосом (2n) путём мейоза.
Задача 4. Какой хромосомный набор характерен для клеток листьев и коробочки на ножке кукушкина льна? Объясните, из каких исходных клеток и в результате, какого деления они образуются.
Ответ:
1. В клетках листьев кукушкина льна гаплоидный набор хромосом (n), они, как и всё растение, развиваются из споры с гаплоидным набором хромосом (n) путём митоза.
2. В клетках коробочки на ножке диплоидный набор хромосом (2n), она развивается из зиготы с диплоидным набором хромосом (2n) путём митоза.
Лекция
Жизненный цикл папоротников
У папоротников (также хвощей, плаунов) в
жизненном цикле преобладает спорофит (2n). На
нижней стороне листьев растения (2n) развиваются
спорангии (2n), в которых путём мейоза образуются
споры (n). Из споры (n), попавшей во влажную почву,
прорастает заросток (n) – обоеполый гаметофит. На
его нижней стороне развиваются антеридии (n) и
архегонии (n), а в них путём митоза образуются
сперматозоиды (n) и яйцеклетки (n). С капельками
росы или дождевой воды сперматозоиды (n) попадают
к яйцеклеткам (n), образуется зигота (2n), а из нее –
зародыш нового растения (2n). (Демонстрация
слайдов).
Из споры (n), попавшей во влажную почву,
прорастает заросток (n) – обоеполый гаметофит. На
его нижней стороне развиваются антеридии (n) и
архегонии (n), а в них путём митоза образуются
сперматозоиды (n) и яйцеклетки (n). С капельками
росы или дождевой воды сперматозоиды (n) попадают
к яйцеклеткам (n), образуется зигота (2n), а из нее –
зародыш нового растения (2n). (Демонстрация
слайдов).
Схема 3. Жизненный цикл папоротников
Практикум
Задача 5. Какой хромосомный набор характерен для листьев (вай) и заростка папоротника? Объясните, из каких исходных клеток и в результате, какого деления образуются эти клетки.
Ответ:
1. В клетках листьев папоротника диплоидный набор хромосом (2n), так они, как и всё растение, развиваются из зиготы с диплоидным набором хромосом (2n) путём митоза.
2. В клетках заростка гаплоидный набор хромосом
(n), так как заросток образуется из гаплоидной
споры (n) путём митоза.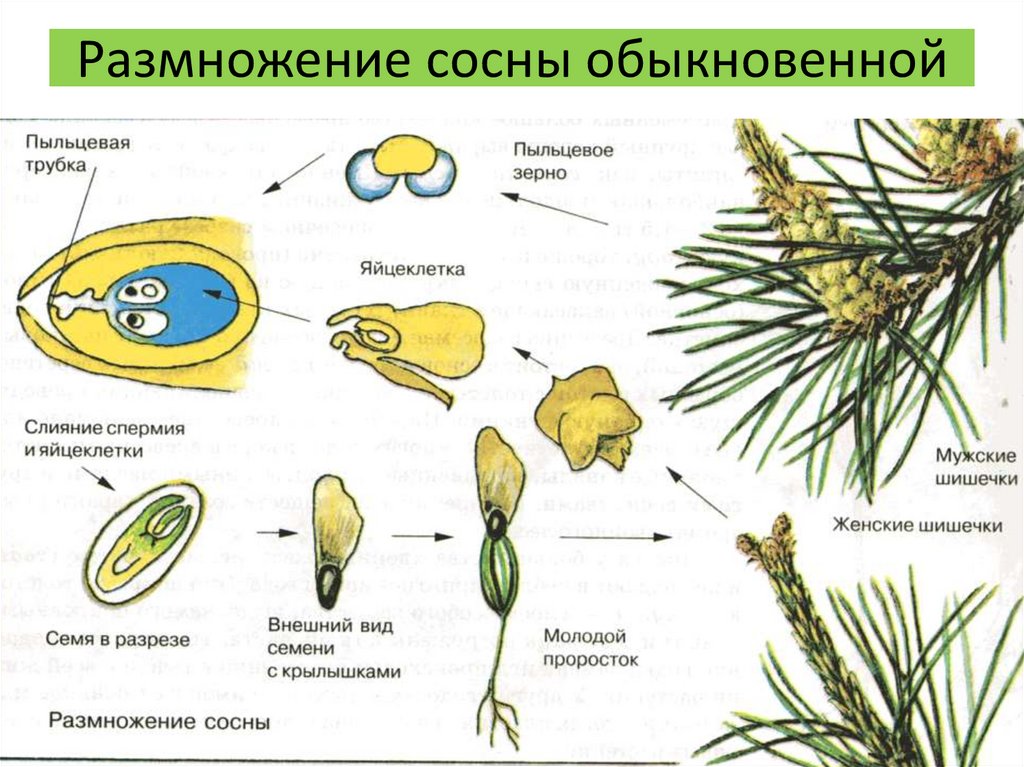
Лекция
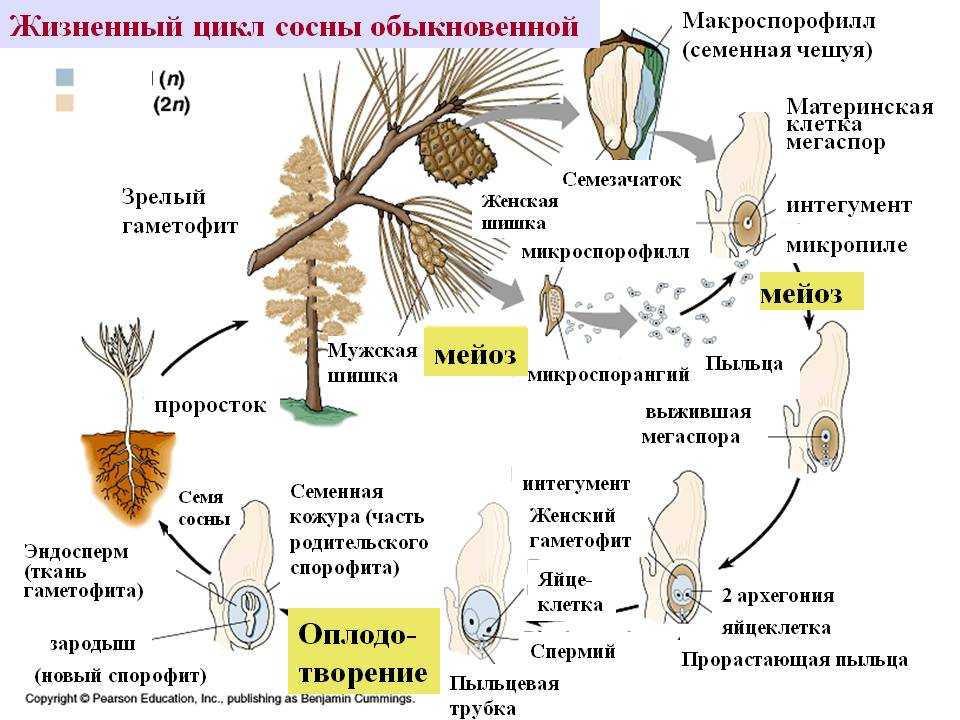
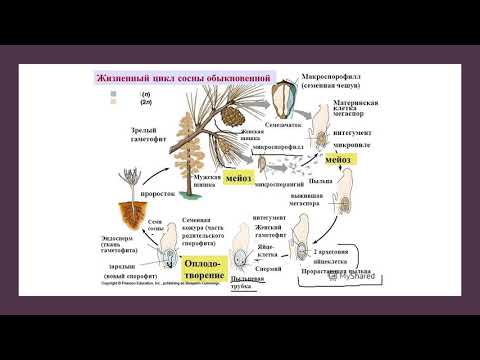
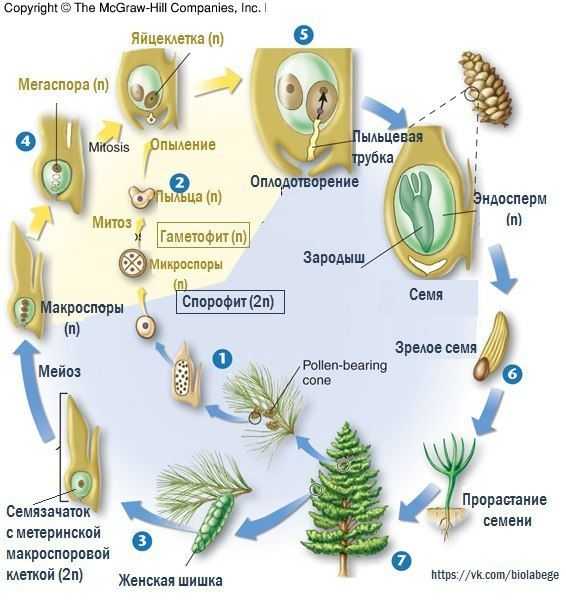
Жизненный цикл голосеменных растений (сосна)
Листостебельное растение голосеменных растений – спорофит (2n), на котором развиваются женские и мужские шишки (2n).
На чешуйках женских шишек расположены семязачатки – мегаспорангии (2n), в которых путём мейоза образуются 4 мегаспоры (n), 3 из них погибают, а из оставшейся – развивается женский гаметофит – эндосперм (n) с двумя архегониями (n). В архегониях образуются 2 яйцеклетки (n), одна погибает.
На чешуйках мужских шишек располагаются пыльцевые мешки – микроспорангии (2n), в которых путём мейоза образуются микроспоры (n), из них развиваются мужские гаметофиты – пыльцевые зёрна (n), состоящие из двух гаплоидных клеток (вегетативной и генеративной) и двух воздушных камер.
Пыльцевые зёрна (n) (пыльца) ветром переносятся
на женские шишки, где митозом из генеративной
клетки (n) образуются 2 спермия (n), а из
вегетативной (n) – пыльцевая трубка (n), врастающая
внутрь семязачатка и доставляющая спермии (n) к
яйцеклетке (n).
В результате из семязачатка формируется семя, покрытое кожурой и содержащее внутри зародыш (2n) и эндосперм (n).
Схема 4. Жизненный цикл голосеменных растений (сосна)
Практикум
Задача 6. Какой хромосомный набор характерен для клеток пыльцевого зерна и спермиев сосны? Объясните, из каких исходных клеток и в результате, какого деления образуются эти клетки.
Ответ:
1. В клетках пыльцевого зерна гаплоидный набор хромосом (n), так как оно образуется из гаплоидной микроспоры (n) путём митоза.
2. В спермиях гаплоидный набор хромосом (n), так как они образуются из генеративной клетки пыльцевого зерна с гаплоидным набором хромосом (n) путём митоза.
Задача 7.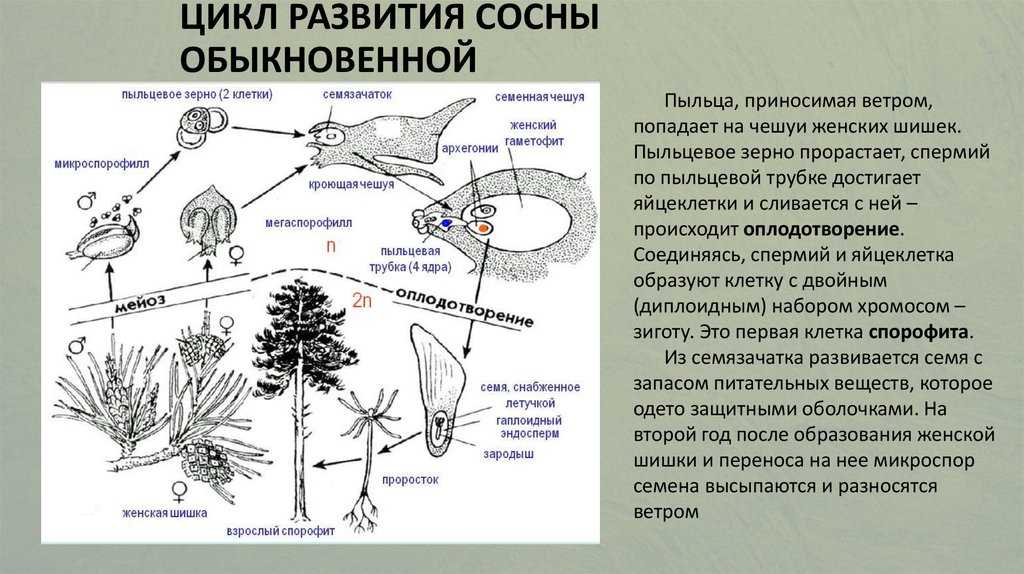 Какой хромосомный набор характерен
для мегаспоры и клеток эндосперма сосны?
Объясните, из каких исходных клеток и в
результате, какого деления образуются эти
клетки.
Какой хромосомный набор характерен
для мегаспоры и клеток эндосперма сосны?
Объясните, из каких исходных клеток и в
результате, какого деления образуются эти
клетки.
Ответ:
1. В мегаспорах гаплоидный набор хромосом (n), так как они образуются из клеток семязачатка (мегаспорангия) с диплоидным набором хромосом (2n) путём мейоза.
2. В клетках эндосперма гаплоидный набор хромосом (n), так как эндосперм формируется из гаплоидных мегаспор (n) путём митоза.
Лекция
Жизненный цикл покрытосеменных растений
Покрытосеменные растения являются спорофитами (2n). Органом их полового размножения является цветок.
В завязи пестиков цветка находятся семязачатки
– мегаспорангии (2n), где происходит мейоз и
образуются 4 мегаспоры (n), 3 из них погибают, а из
оставшейся – развивается женский гаметофит –
зародышевый мешок из 8 клеток (n), одна из них –
яйцеклетка (n), а две сливаются в одну – крупную
(центральную) клетку с диплоидным набором
хромосом (2n).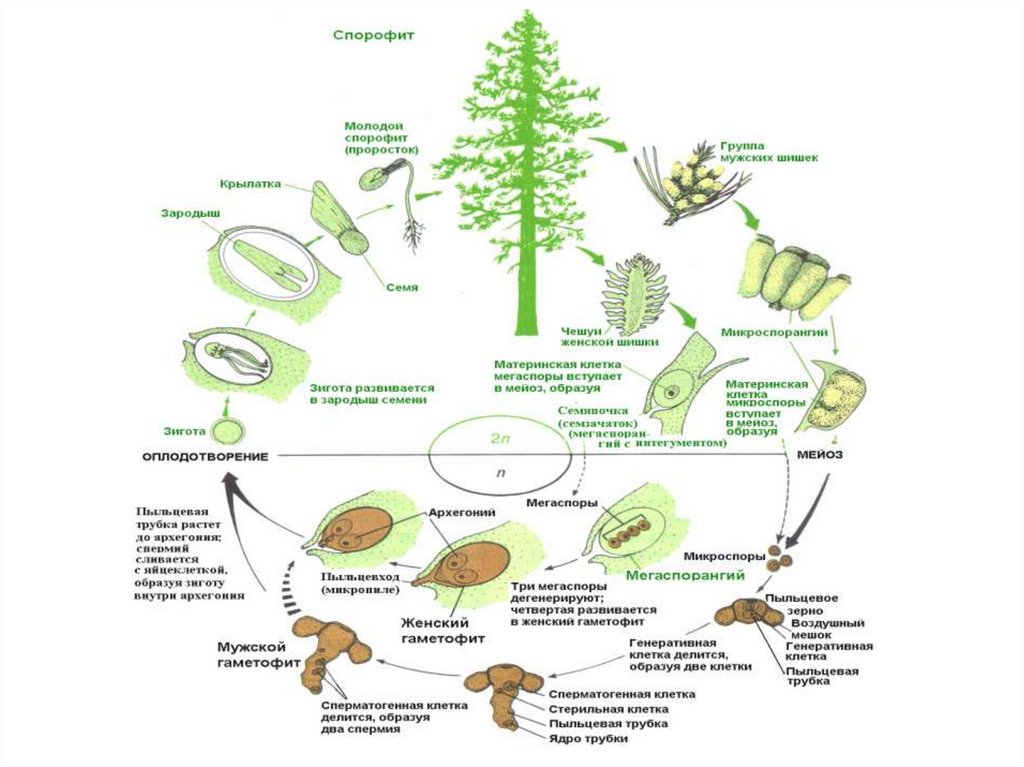
В микроспорангиях (2n) пыльников тычинок путём мейоза образуются микроспоры (n), из которых развиваются мужские гаметофиты – пыльцевые зёрна (n), состоящие из двух гаплоидных клеток (вегетативной и генеративной).
После опыления из генеративной клетки (n) образуются 2 спермия (n), а из вегетативной (n) – пыльцевая трубка (n), врастающая внутрь семязачатка и доставляющая спермии (n) к яйцеклетке (n) и центральной клетке (2n) . Один спермий (n) сливается с яйцеклеткой (n) и образуется зигота (2n), из которой митозом формируется зародыш растения (2n). Второй спермий (n) сливается центральной клеткой (2n) с образованием триплоидного эндосперма (3n). Такое оплодотворение у покрытосеменных растений называется двойным.
В результате из семязачатка формируется семя, покрытое кожурой и содержащее внутри зародыш (2n) и эндосперм (3n).
Схема 5. Жизненный цикл покрытосеменных растений
Практикум
Задача 8.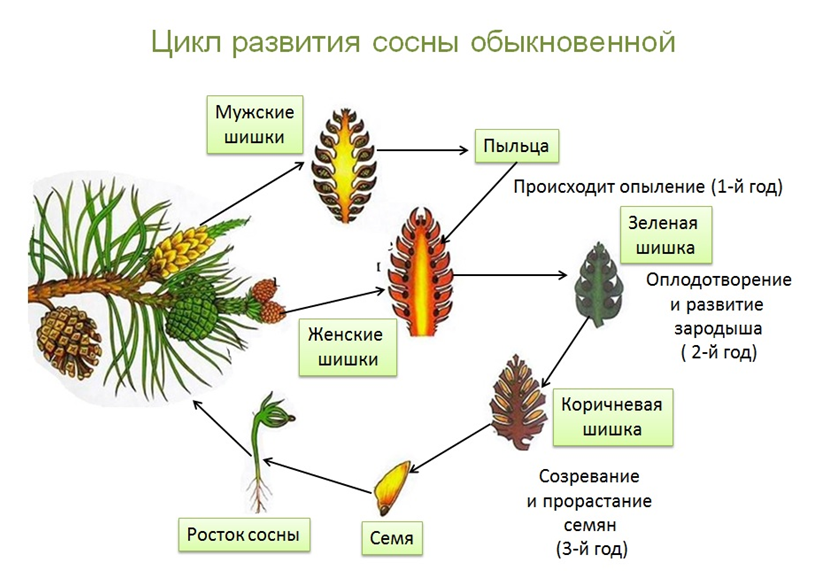 Какой хромосомный набор характерен
для микроспоры, которая образуется в пыльнике, и
клеток эндосперма семени цветкового растения?
Объясните, из каких исходных клеток и как они
образуются.
Какой хромосомный набор характерен
для микроспоры, которая образуется в пыльнике, и
клеток эндосперма семени цветкового растения?
Объясните, из каких исходных клеток и как они
образуются.
Ответ:
1. В микроспорах гаплоидный набор хромосом (n), так как они образуются из клеток микроспорангиев с диплоидным набором хромосом (2n) путём мейоза.
2. В клетках эндосперма триплоидный набор хромосом (3n), так как эндосперм образуется при слиянии гаплоидного спермия (n) с диплоидной центральной клеткой (2n).
Общие выводы
1. В процессе эволюции растений происходила постепенная редукция гаметофита и развитие спорофита.
2. В гаметах растений гаплоидный набор (n) хромосом, они образуются путём митоза.
3. В спорах растений гаплоидный набор (n) хромосом, они образуются путём мейоза.
Эмбриогенез сосны — PMC
1. Сарвас Р. Исследования цветения и семенного материала Pinus silvestris . Комм Инст Форест Фенн. 1962; 53: 1–198. [Google Scholar]
Комм Инст Форест Фенн. 1962; 53: 1–198. [Google Scholar]
2. Филонова Л.Х., фон Арнольд С., Даниэль Г., Божков П.В. Запрограммированная гибель клеток уничтожает все зародыши, кроме одного, в семени полиэмбрионального растения. Смерть клеток 2002; 9: 1057–1062. [PubMed] [Google Scholar]
3. Догра П.Д. Стерильность семян и нарушения эмбриогенеза у хвойных с особым упором на тестирование семян и селекцию деревьев Pinaceae. Стад для Suec. 1967;45:1–92. [Google Scholar]
4. Yeung EC, Meinke DW. Эмбриогенез у покрытосеменных – развитие суспензора. Растительная клетка. 1993; 5: 1371–1381. [Бесплатная статья PMC] [PubMed] [Google Scholar]
5. Оуэнс Дж. Н., Симпсон С. Дж., Молдер М. Половое размножение Pinus contorta II. Постпокойная семяпочка, зародыш и развитие семени. Может Джей Бот. 1982; 60: 2071–2083. [Google Scholar]
6. Tillman-Sutela E, Kauppi A, Karppinen K, Tomback DF. Вариант зрелости в семенных структурах Pinus albicaulis (Engelm.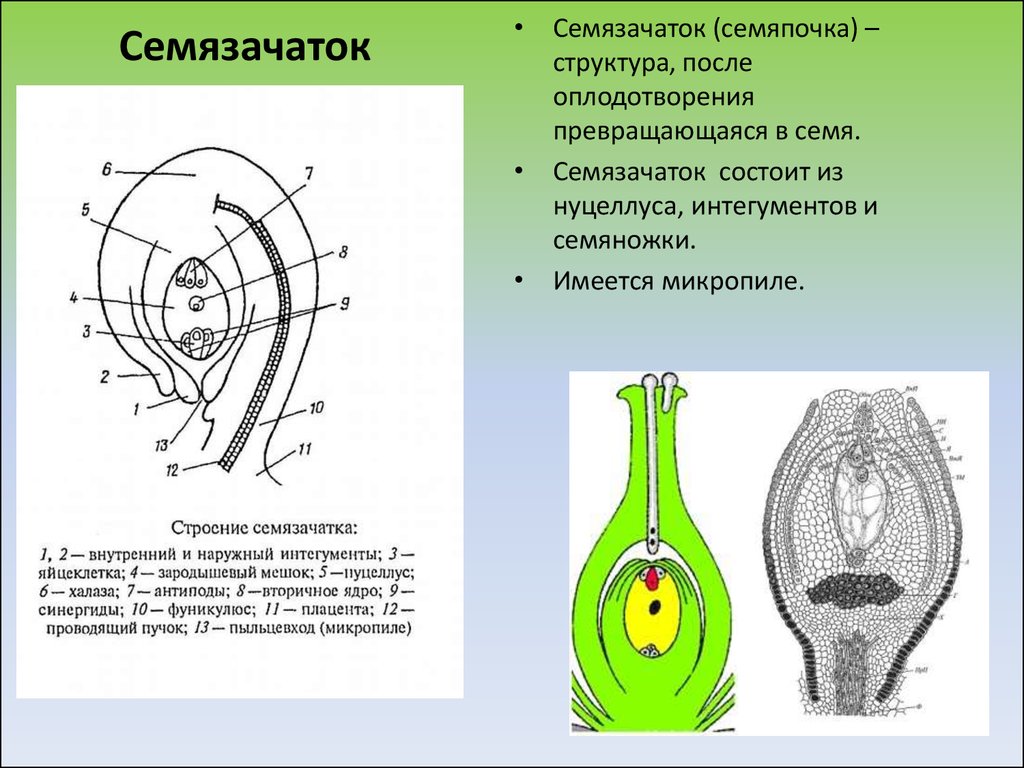 ) и Pinus sibirica (Du Tour): ключ к почвенному банку семян, необычный среди хвойных? Деревья-Структура Функц. 2008; 22: 225–236. [Google Scholar]
) и Pinus sibirica (Du Tour): ключ к почвенному банку семян, необычный среди хвойных? Деревья-Структура Функц. 2008; 22: 225–236. [Google Scholar]
7. Vuosku J, Sarjala T, Jokela A, Sutela S, Sääskilahti M, Suorsa M, et al. Одна ткань, две судьбы: разные роли клеток мегагаметофитов в эмбриогенезе сосны обыкновенной. J Опытный бот. 2009; 60: 1375–1386. [Бесплатная статья PMC] [PubMed] [Google Scholar]
8. Олсен О-А. Развитие ядерного эндосперма у злаков и Arabidopsis thaliana . Растительная клетка. 2004; 16: 214–227. [Бесплатная статья PMC] [PubMed] [Google Scholar]
9. Stone SL, Gifford DJ. Структурно-биохимические изменения семян сосны обыкновенной ( Pinus taeda L.) при прорастании и раннем росте проростков. II Хранение триацилглицеролов и углеводов. Int J Plant Sci. 1999; 158:727–737. [Google Scholar]
10. Хирацука Р., Ямада Ю., Терасака О. Запрограммированная гибель клеток Pinus nucellus в ответ на проникновение пыльцевой трубки. J Завод Res. 2002; 115:141–148. [PubMed] [Академия Google]
J Завод Res. 2002; 115:141–148. [PubMed] [Академия Google]
11. Tillman-Sutela E, Kauppi A, Sahlen K. Влияние нарушенного фотопериода на структуру поверхности созревающих семян сосны обыкновенной ( Pinus sylvestris L.). Деревья. 1998; 12: 499–506. [Google Scholar]
12. Tillman-Sutela E, Kauppi A. Морфологические предпосылки набухания семян Pinus sylvestris L. различного происхождения. Деревья. 1995; 9: 123–133. [Google Scholar]
13. Tillman-Sutela E, Kauppi A. Значение структуры для впитывания семян ели европейской, Picea abies (L.) Карст. Деревья. 1995; 9: 269–278. [Google Scholar]
14. Коста Л.М., Гутьеррес-Маркос Дж.Ф., Дикинсон Х.Г. Больше, чем желток: короткая жизнь и сложные времена эндосперма растений. Тенденции Растениевод. 2004; 9: 507–514. [PubMed] [Google Scholar]
15. Young TE, Gallie DR. Анализ запрограммированной гибели клеток в эндосперме пшеницы выявляет различия в развитии эндосперма между злаками.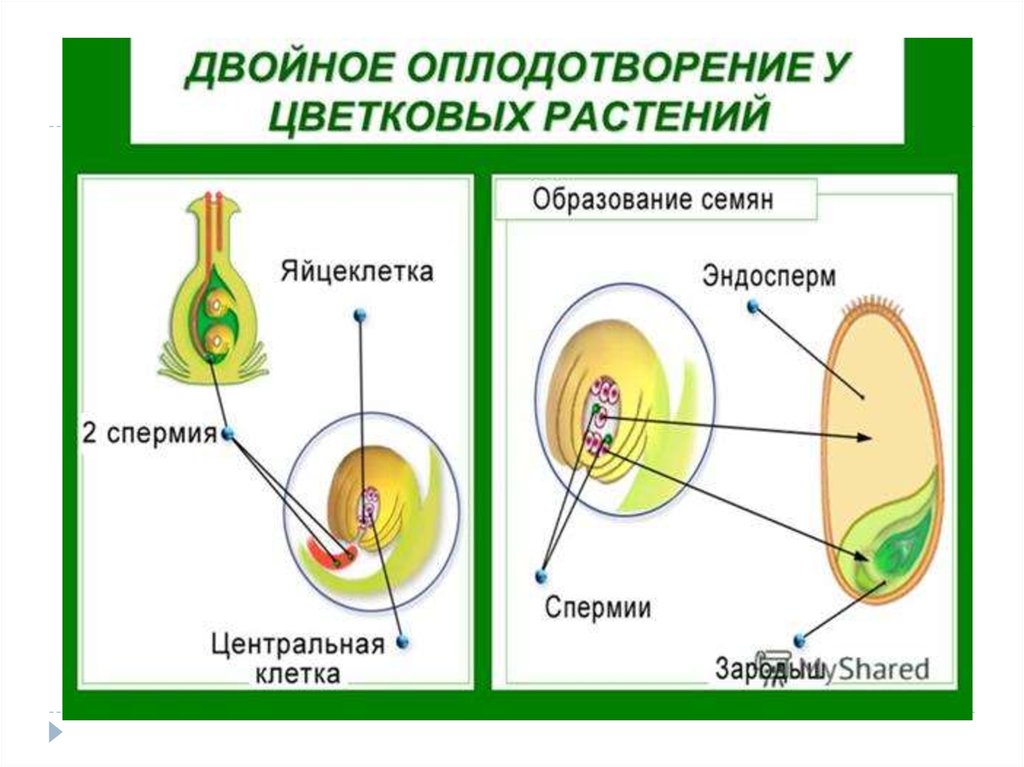 Завод Мол Биол. 1999; 39: 915–926. [PubMed] [Google Scholar]
Завод Мол Биол. 1999; 39: 915–926. [PubMed] [Google Scholar]
16. Tillman-Sutela E, Kauppi A. Структуры, способствующие завершению прорастания семян хвойных. Деревья. 2000;14:191–197. [Google Scholar]
17. Сингх Х. Эмбриология голосеменных растений. Берлин: Борнтрагер; 1978. [Google Scholar]
18. Krasowski MJ, Owens JN. Ультраструктурное и гистохимическое развитие мегагаметофитов и зиготических зародышей ели белой ( Picea glauca ) после оплодотворения с акцентом на отложение продуктов хранения семян. Может Джей Бот. 1993; 71: 98–112. [Google Scholar]
19. Оуэнс Дж. Н., Моррис С. Дж., Мисра С. Ультраструктурное, гистохимическое и биохимическое развитие мегагаметофита после оплодотворения и зиготического эмбриона Псевдоцуга мензиесии . Can J Forest Res. 1993; 23: 816–827. [Google Scholar]
20. Оуэнс Дж. Н., Коланджели А. М., Моррис С. Дж. Факторы, влияющие на завязывание семян пихты Дугласа ( Pseudotsuga menziesii ) Can J Bot. 1991; 69: 229–238. [Google Scholar]
1991; 69: 229–238. [Google Scholar]
21. von Aderkas P, Rouault G, Wagner R, Chiwocha S, Roques A. Многоядерные запасающие клетки пихты Дугласа ( Pseudotsuga menziesii (Mirbel) Franco) и влияние паразитизма семян халцидом Мегастигмус спермотроф Часы. Наследственность. 2005; 94: 616–622. [PubMed] [Google Scholar]
22. Chiwocha S, Rouault G, Abrams S, von Aderkas P. Паразитизм семян пихты Дугласа ( Pseudotsuga menziesii ) семенами халцида, Megastigmus спермотрофа , и его влияние на Физиология семенных гормонов. Половое растение Репрод. 2007; 20:19–25. [Google Scholar]
23. Капик Р.Х., Динус Р.Дж., Дин Д.Ф.Д. Абсцизовая кислота и зиготический эмбриогенез у Pinus taeda . Физиол дерева. 1995; 15: 485–49.0. [PubMed] [Google Scholar]
24. Kong LS, Attree SM, Fowke LC. Изменения уровня эндогенных гормонов в развивающихся семенах, зиготических зародышах и мегагаметофитах у Picea glauca . Завод Физиол. 1997; 101: 23–30.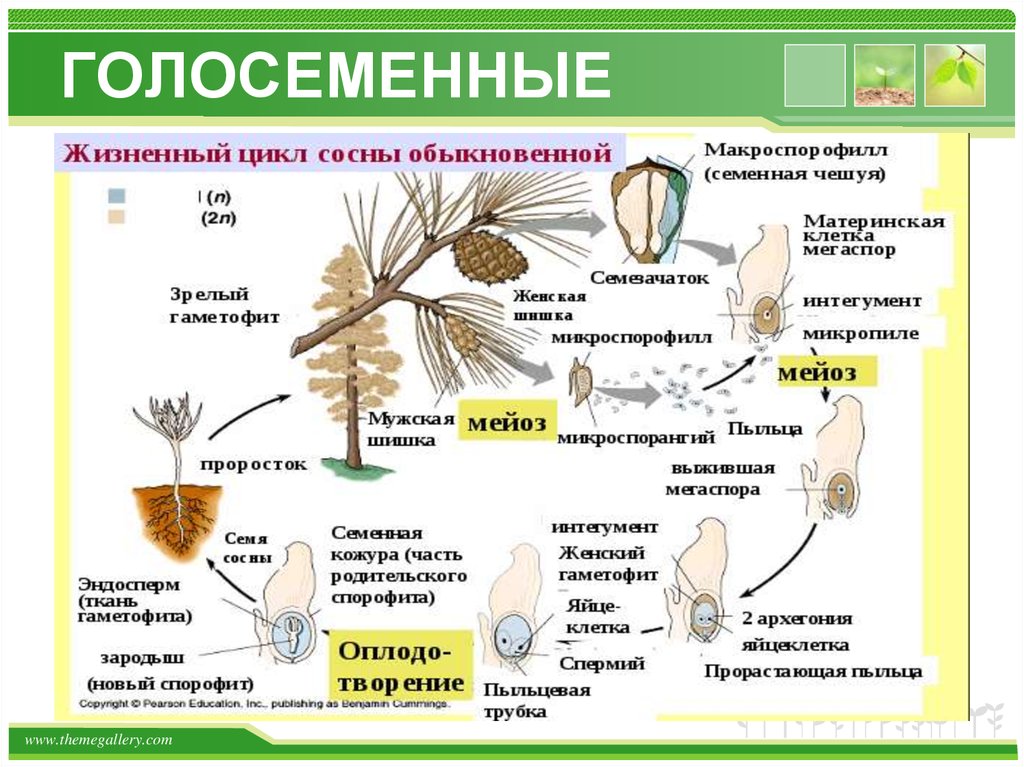 [Google Scholar]
[Google Scholar]
25. Carrier DJ, Kendall EJ, Bock CA, Cunningham JE, Dunstan DI. Содержание воды, отложение липидов и содержание (+)-абсцизовой кислоты в развивающихся семенах ели белой. J Опытный бот. 1999;50:1359–1364. [Google Scholar]
26. Carman JG, Reese G, Fuller RJ, Ghermay T, Timmis R. Уровни питательных веществ и гормонов в коррозионных полостях дугласовой пихты, мегагаметофитах и эмбрионах во время эмбрионального развития. Может J для Res. 2005; 35: 2447–2456. [Академия Google]
27. Конг Л.С., Юнг Э.К. Влияние нитрата серебра и полиэтиленгликоля на соматическое развитие зародыша ели белой ( Picea glauca ): усиление образования семядольных зародышей и содержания эндогенной АБК. Физиол Плантарум. 1995; 93: 298–304. [Google Scholar]
28. Hakman I, Stabel P, Engström P, Eriksson T. Накопление запасных белков во время развития зиготических и соматических эмбрионов у Picea abies (ель обыкновенная) Physiol Plant. 1990; 80: 441–445. [Академия Google]
[Академия Google]
29. Флинн Б.С., Робертс Д.Р., Уэбб Д.Т., Саттон Б.С. Изменения запасного белка во время зиготического эмбриогенеза у интерьерной ели. Физиол дерева. 1991; 8: 71–81. [PubMed] [Google Scholar]
30. Misra S, Attree SM, Leal I, Fowke LC. Влияние абсцизовой кислоты, осмотика и высушивания на синтез запасных белков при развитии соматических зародышей ели белой. Энн Бот. 1993; 71:11–22. [Google Scholar]
31. Леал И., Мисра С., Аттри С.М., Фоук Л.С. Влияние абсцизовой кислоты, осмотика и высушивания на экспрессию гена запасного белка 11S в соматических зародышах ели белой. Растениевод. 1995;106:121–128. [Google Scholar]
32. Häggman HM, Ryynänen LA, Aronen TS, Krajnakova J. Криоконсервация эмбриогенных культур сосны обыкновенной. Культ органов растительных клеток. 1998; 54:45–53. [Google Scholar]
33. Häggman HM, Vuosku J, Sarjala T, Jokela A, Niemi K. Соматический эмбриогенез видов сосны — от функциональной геномики до плантационного лесоводства.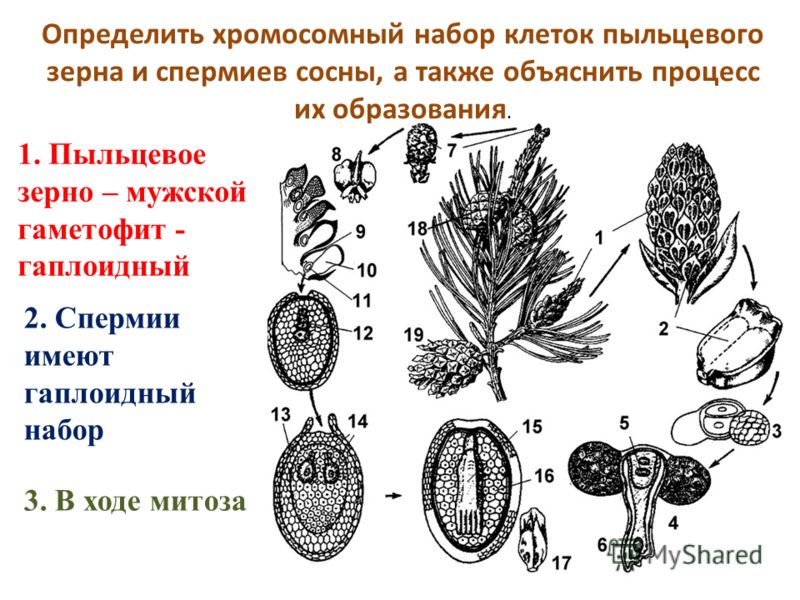 В: Муджиб А., Самадж Дж., редакторы. Соматический эмбриогенез. Том. 2. Берлин: Спрингер; 2005. С. 119–140. [Google Scholar]
В: Муджиб А., Самадж Дж., редакторы. Соматический эмбриогенез. Том. 2. Берлин: Спрингер; 2005. С. 119–140. [Google Scholar]
34. Häggman H, Jokela A, Krajnakova J, Kauppi A, Niemi K, Aronen T. Соматический эмбриогенез сосны обыкновенной: обработка холодом и характеристики эксплантов, влияющих на индукцию. J Опытный бот. 1999;50:1769–1778. [Google Scholar]
35. Ungru A, Nowack MK, Reymond M, Shirzadi R, Kumar M, Biewers S, et al. Естественная изменчивость степени автономного образования эндосперма выявляет независимость и ограничения роста зародыша во время развития семян у Arabidopsis thaliana . Генетика. 2008; 179: 829–841. [Бесплатная статья PMC] [PubMed] [Google Scholar]
36. Cairney J, Pullman GS. Клеточная и молекулярная биология эмбриогенеза хвойных. Новый Фитол. 2007; 176: 511–536. [PubMed] [Академия Google]
37. О Т.Дж., Уартелл Р.М., Кэрни Дж., Пуллман Г.С. Доказательства стадийно-специфической модуляции специфических микроРНК (миРНК) и компонентов процессинга микроРНК в зиготических эмбрионах и женских гаметофитах сосны лоблолли ( Pinus taeda ) New Phytol.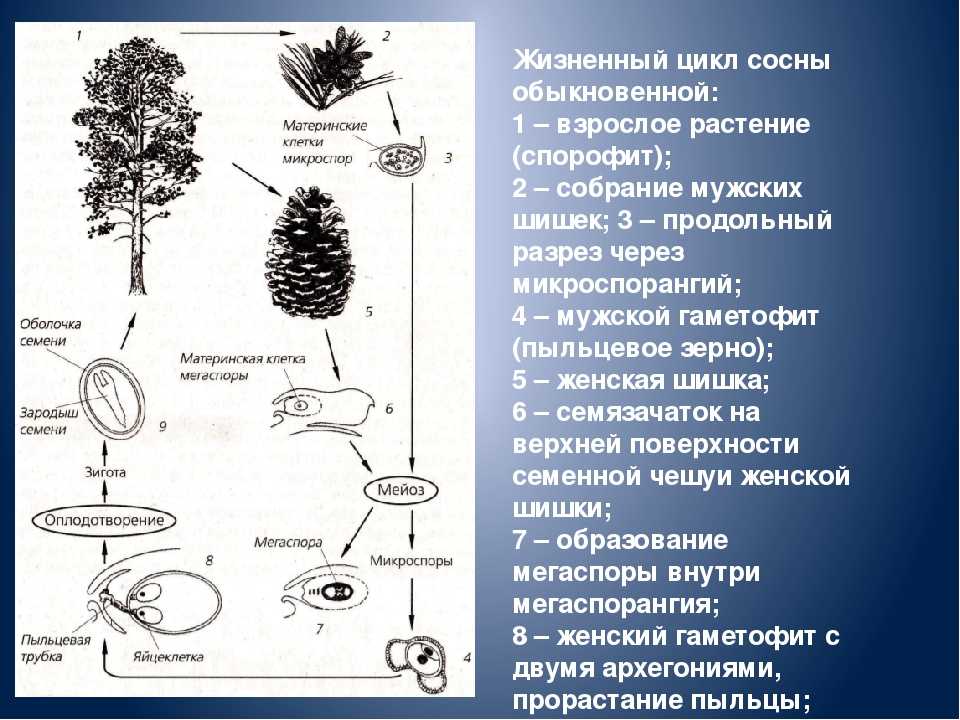 2008; 179: 67–80. [PubMed] [Google Scholar]
2008; 179: 67–80. [PubMed] [Google Scholar]
38. Earnshaw WC. Ядерные изменения при апоптозе. Curr Opin Cell Biol. 1995; 7: 337–343. [PubMed] [Google Scholar]
39. Balk J, Chew SK, Leaver CJ, McCabe PF. Межмембранное пространство митохондрий растений содержит ДНКазную активность, которая может быть вовлечена в запрограммированную гибель клеток. Плант Дж. 2003; 34: 573–583. [PubMed] [Академия Google]
40. Хе X, Кермоуд АР. Нуклеазная активность и фрагментация ДНК при запрограммированной гибели клеток мегагаметофитов семян белой ели ( Picea glauca ). Завод Мол Биол. 2003; 51: 509–521. [PubMed] [Google Scholar]
41. Уолтеринг Э.Дж. Протеазы смерти оживают. Тенденции Растениевод. 2004; 9: 469–472. [PubMed] [Google Scholar]
42. Божков П., Янссон С. Аутофагия и протеазы клеточной гибели у растений. Два колеса похоронной тележки. Аутофагия. 2007; 3: 136–138. [PubMed] [Академия Google]
43. Bonneau L, Ge Y, Drury GE, Gallois P. Что случилось с каспазами растений? J Опытный бот. 2008; 59: 491–499. [PubMed] [Google Scholar]
2008; 59: 491–499. [PubMed] [Google Scholar]
44. Qiu JZ, Yoon JH, Shen BH. Поиск апоптотических нуклеаз у дрожжей — роль нуклеазы Tat-D в апоптотической деградации ДНК. Дж. Биол. Хим. 2005; 280:15370–15379. [PubMed] [Google Scholar]
45. He X, Kermode AR. Протеазы, ассоциированные с запрограммированной клеточной гибелью клеток мегагаметофитов после прорастания ели белой ( Picea glauca ) семена. Завод Мол Биол. 2003; 52: 729–744. [PubMed] [Google Scholar]
46. Божков П.В., Суарес М.Ф., Филонова Л.Х., Даниэль Г., Замятнин А.А., Родригес-Ньето С. и соавт. Цистеиновая протеаза mcll-Pa осуществляет запрограммированную гибель клеток во время эмбриогенеза растений. Proc Natl Acad Sci USA. 2005; 102:14463–14468. [PMC free article] [PubMed] [Google Scholar]
47. Божков П.В., Филонова Л.Х., Суарес М.Ф., Хельмерссон А., Смертенко А.П., Животовский Б., фон Арнольд С. гибели клеток и необходим для формирования эмбрионального паттерна. Смерть клеток 2004; 11: 175–182. [PubMed] [Академия Google]
[PubMed] [Академия Google]
48. Хельмерссон А., фон Арнольд С., Божков П.В. Уровень свободного внутриклеточного цинка опосредует запрограммированные решения о гибели клеток/выживании клеток у эмбрионов растений. Завод Физиол. 2008; 147:1158–1167. [Бесплатная статья PMC] [PubMed] [Google Scholar]
49. Sorensen FC. Роль полиэмбрионии и жизнеспособности зародышей в генетической системе хвойных. Эволюция. 1982; 36: 725–733. [PubMed] [Google Scholar]
50. Williams CG. Гибель самоопыляющихся эмбрионов у Pinus taeda : фенотипический профиль. Новый Фитол. 2008; 178: 210–222. [PubMed] [Академия Google]
51. Нил Д.Б., Седерофф Р.Р. Отцовское наследование хлоропластной ДНК и материнское наследование митохондриальной ДНК у сосны лоблолли. Теория Appl Genet. 1989; 77: 212–216. [PubMed] [Google Scholar]
52. Bruns D, Owens JN. Сосна западная белая ( Pinus monticola Dougl.) репродукция: II. Оплодотворение и цитоплазматическая наследственность. Половое растение Репрод. 2000; 13:75–84. [Google Scholar]
Половое растение Репрод. 2000; 13:75–84. [Google Scholar]
53. Пюхяярви Т., Салмела М.Ю., Саволайнен О. Пути колонизации Pinus sylvestris выведено из распределения вариаций митохондриальной ДНК. Генетические геномы деревьев. 2008; 4: 247–254. [Google Scholar]
54. Журов В., Терзин Т., Грбич М. (Ин)дискретное очарование полиэмбрионии: эволюция клонирования эмбрионов. Cell Mol Life Sci. 2007; 64: 2790–2798. [PubMed] [Google Scholar]
55. Buchholz JT. Происхождение полиэмбрионии дробления у хвойных. Бот Газ. 1926; 81: 55–71. [Google Scholar]
56. О’Коннелл Л.М., Ритланд К. Механизмы постопыления, способствующие ауткроссингу у самоплодных хвойных деревьев, Thuja plicata (Cupressaceae) Can J Bot. 2005; 83: 335–342. [Google Scholar]
57. Орр-Юинг А.Л. Цитологическое исследование влияния самоопыления на Pseudotsuga menziesii (Mirb.) Franco. Сильве Жене. 1957; 6: 179–185. [Google Scholar]
58. Стивс Т.А. Эволюция и биологическое значение семян.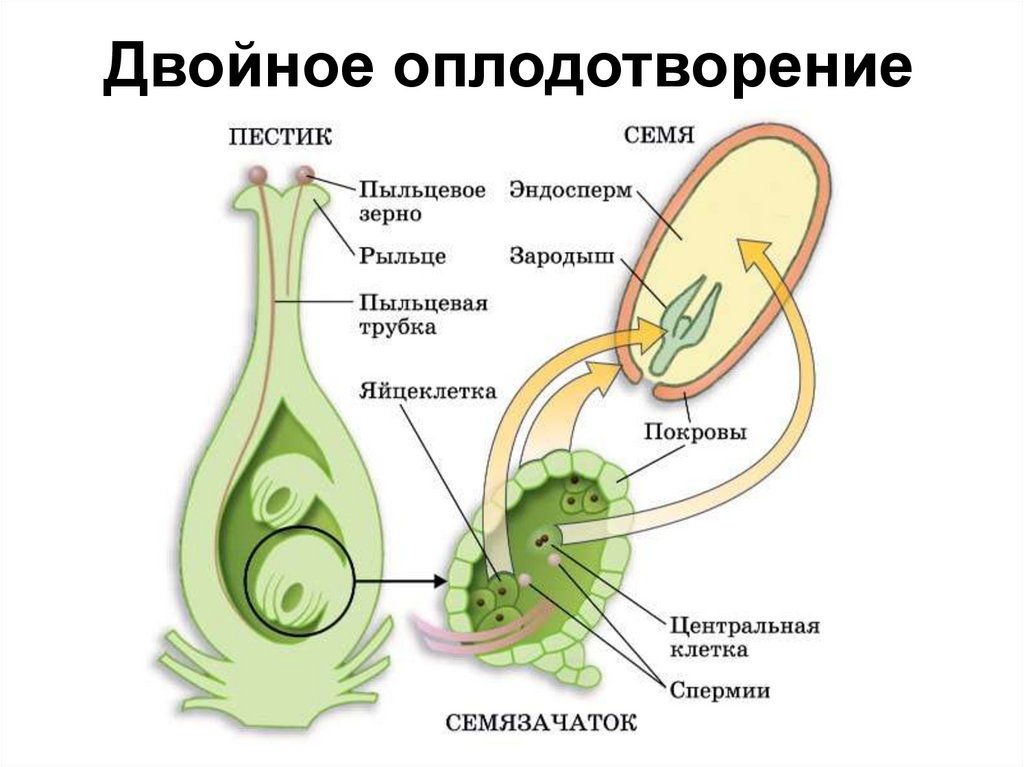 Может Джей Бот. 1983; 61: 3550–3560. [Google Scholar]
Может Джей Бот. 1983; 61: 3550–3560. [Google Scholar]
59. Ю С-М. Клеточные и генетические реакции растений на сахарное голодание. Завод Физиол. 1999;121:687–693. [Бесплатная статья PMC] [PubMed] [Google Scholar]
60. Green DR. Пути апоптоза: десять минут до смерти. Клетка. 2005; 121: 671–674. [PubMed] [Google Scholar]
61. Laporte C, Kosta A, Klein G, Aubry L, Lam D, Tresse E, et al. Модель гибели некротических клеток у протистов. Смерть клеток 2007; 14: 266–274. [PubMed] [Google Scholar]
62. Кришнамурти К.В., Кришнарадж Р., Чожавендан Р., Кристофер Ф.С. Программа клеточной гибели растений и животных. Сравнение. Curr Sci. 2000;79: 1169–1181. [Google Scholar]
63. Ameisen JC. О происхождении, эволюции и природе запрограммированной клеточной смерти: хронология в четыре миллиарда лет. Смерть клеток 2002; 9: 367–393. [PubMed] [Google Scholar]
64. Животовский Б. От нематоды и млекопитающих обратно к сосне: о разнообразии и эволюции программируемой клеточной смерти. Смерть клеток 2002; 9: 867–869. [PubMed] [Google Scholar]
Смерть клеток 2002; 9: 867–869. [PubMed] [Google Scholar]
Состав жирных кислот в липидах эндосперма и зародыша семян Pinus sibirica и P. sylvestris
Веселаке, Р.Дж. и Тейлор, Д.К., Исследование биосинтеза запасных липидов с использованием культур, полученных из семян масличного рапса, полученных из микроспор, Prog. Липид Рез. , 1999, том. 38, стр. 401–460.
Артикул пабмед КАС Google Scholar
Вольф Р.Л., Педроно Ф., Паскье Э. и Марпо А.М., Общие характеристики Pinus spp. Составы жирных кислот семян и значение 5 -Олефиновые кислоты в таксономии и филогении рода, Lipids , 2000, vol. 35, стр. 1–22.
Артикул пабмед КАС Google Scholar
Бейссон Ф., Ку А.Дж.К., Рууска С., Швендер Дж., Поллард М.
, Телен Дж.Дж., Пэддок Т., Салас Дж.Дж., Сэвидж Л., Милкампс А. , Mhaske, V.B., Cho, Y., and Ohlrogge, J.B., Гены арабидопсиса, участвующие в метаболизме ациллипидов. Перепись кандидатов 2003 г., исследование распределения тегов экспрессируемых последовательностей в органах и база данных в Интернете, Физиол растений. , 2003, том. 132, стр. 681–697.Артикул пабмед КАС Google Scholar
Шанклин, Дж. и Кахун, Э., Десатурация и родственные модификации жирных кислот, Annu. Преподобный Завод Физиол. Завод Мол. биол. , 1998, том. 49, стр. 611–649.
Артикул пабмед КАС Google Scholar
Мехедов С., де Илардуя О.М. и Олрогге Дж. К функциональному каталогу генома растений. Обзор генов биосинтеза липидов, Физиол растений. , 2000, том. 122, стр. 389–401.
Артикул пабмед КАС Google Scholar
Лос, Д.А., Десатуразы жирных кислот: структура, регуляция экспрессии и функционирование, Усп. биол. Наук , 2001, т. 1, с. 41, стр. 163–198.
КАС Google Scholar
Окулей Дж., Лайтнер Дж., Фельдман К., Ядов Н. и Ларк Э., Arabidopsis fad2 Ген кодирует фермент, необходимый для синтеза полиненасыщенных липидов, Растительная клетка , 1994, вып. 6, стр. 147–158.
Артикул пабмед КАС Google Scholar
Точер, Д.Р., Ливер, М.Дж., и Ходжсон, П.
А., Последние достижения в области биохимии и молекулярной биологии жирных ацилдесатураз, Prog. Липид Рез. , 1998, том. 37, стр. 73–117.Артикул пабмед КАС Google Scholar
Петрини Г.А., Альтабе С.Г. и Уттаро А.Д., Trypansoma brucei Олеатдесатураза может использовать цитохром b 5 -подобный домен в другой десатуразе в качестве донора электронов, Eur. Дж. Биохим. , 2004, том. 271, стр. 1079–1086.
Артикул пабмед КАС Google Scholar
Заборовска З., Старжицки М., Фемяк И., Свидерски М. и Легоцкий А., Ген желтого люпина, кодирующий стеароил-АСР-десатуразу — организация, экспрессия и потенциальное применение, Акта Биохим. пол. , 2002, том. 49, стр. 29–42.
КАС Google Scholar
Hu, X., Sullivan-Gilbert, M., Gupta, M., and Thompson, S.A. Картирование локусов, контролирующих содержание олеиновой и линоленовой кислот и развитие fad2 и fad3 аллель-специфических маркеров у канолы ( Brassica napus L.), Theor. заявл. Жене. , 2006, том. 113, стр. 497–507.
Артикул пабмед КАС Google Scholar
Кэрриер, Д.-Дж., Каннингем, Дж.Э., Хогге, Л.Р., Тейлор, Д.К., и Данстан, Д.И., Газохромато-масс-спектрометрическая характеристика некоторых кислот из белой и внутренней ели, Ж. Хроматогр.
, 1995, том. 715, стр. 317–324.Артикул КАС Google Scholar
Танака, Т., Хаттори, Т., Коучи, М., Хирано, К., и Сатоучи, К., Двойная связь с прерыванием метилена в полиненасыщенных жирных кислотах является важной структурой для метаболизма в цепи жирных кислот Система удлинения печени крыс, Биохим. Биофиз. Acta , 1998, vol. 1393, стр. 299–306.
ПабМед КАС Google Scholar
Wolff, R.L., Christie, W.W., Pedrone, F., Marpeau, A.M., Tsevegsuren, N., Aitzetmuller, K., and Gunstoune, F., 5 -Олефиновая кислота в липидах семян из Four Виды эфедры и их распределение между позициями a и b триацилглицеролов. Характеристики, общие для Coniferophytes и Cycadophytes, Lipids , 1999, vol. 34, стр. 855–864.
Артикул пабмед КАС Google Scholar
Айцетмюллер, К. и Цевегсурен, Н., Жирные кислоты семян, десатуразы «переднего конца» и хемотаксономия — тематическое исследование лютиковых, J. Физиол растений. , 1994, том. 143, стр. 538–543.
Google Scholar
Кнутзон Д.С., Турмонд Дж.М., Хуанг Ю.-С., Чаудхари С., Бобик Э.Г., Чан Г.М., Киршнер С.Дж. и Мукерджи П. Идентификация 5 -десатураза из Mortierella alpina путем гетерологичной экспрессии в хлебопекарных дрожжах и каноле, J. Biol. хим. , 1998, том. 273, с. 29 360–29 366.
Статья КАС Google Scholar
Тонон, Т., Саянова, О., Майклсон, Л.В., Цин, Р., Харви, Д., Ларсон, Т.Р., Ли, Ю., Нейпир, Дж.А., и Грэм, И.А., Десатуразы жирных кислот из микроводоросли Thalassisira pseudonana, FEBS J. , 2005, vol. 272, стр. 3401–3412.
Артикул КАС Google Scholar
Танака Т., Моришиге Дж., Такимото Т., Такай Ю. и Сатоути К. Метаболическая характеристика сциадоновой кислоты (5с, 11с, 14с-эйкозатриеновая кислота) как эффективной замены Арахидонат фосфатидилинозитола, евро. Дж. Биохим. , 2001, том. 268, стр. 4928–4939.
Артикул пабмед КАС Google Scholar
Сугано М., Икеда И., Вакамацу К. и Ока Т. Влияние кедра корейского ( Pinus koraiensis ) Масло семян цис -5, цис -9, цис--12-октадекатриеновая кислота на метаболизм полиненасыщенных жирных кислот, производство эйкозаноидов и кровяное давление у крыс, Br. Дж. Нутр. , 1994, том. 72, стр. 775–783.
Артикул пабмед КАС Google Scholar
Вольф Р.Л., Компс Б., Марпо А. и Делюк Л.Г., Таксономия Pinus Виды на основе состава жирных кислот масла семян, Trees , 1997, vol. 12, стр. 113–118.
Google Scholar
Вольф, Р.Л., Лавиаль, О., Педроно, Ф., Паскье, Э., Делус, Л., Марпо, А.М., и Айцетмюллер, К., Композиции жирных кислот Pinaceae как таксономические маркеры, Липиды , 2001, вып.
36, стр. 439–451.Артикул пабмед КАС Google Scholar
Нейпир, Дж. А., Саянова, О., Сперлинг, П., и Хайнц, Э. А., Растущее семейство цитохромов b 5 Fusion Proteins, Trends Plant Sci. , 1999, том. 4, стр. 2–5.
Артикул Google Scholar
Саянова О. и Напье Дж. А., Эйкозапентаеновая кислота: пути биосинтеза и возможности синтеза в трансгенных растениях, Фитохимия , 2004, том. 65, стр. 147–158.
Артикул пабмед КАС Google Scholar
Asset, G., Bauge, E., Wolff, R.L., Fruchart, J.C., and Dallongeville, J., Pinus pinaster Масло влияет на метаболизм липопротеинов у мышей с дефицитом аполипопротеина E, J. Nutr. , 1999, том. 129, стр. 972–978.
Google Scholar
Bligh, E.C. и Dyer, WJ, A Rapid Method of Total Lipid Extraction and Purification, Can. Дж. Биохим. Физиол. , 1959, том. 37, стр. 911–917.
ПабМед КАС Google Scholar
Карро, В.П. и Dubacq, J.P., Адаптация метода макромасштаба к микромасштабу для метиловой переэтерификации жирных кислот биологических липидных экстрактов, J. Chromatogr. , 1978, том. 151, стр. 384–390.
Артикул КАС Google Scholar
Christie, W.
W., Эквивалентные длины цепей производных метилового эфира жирных кислот в газовой хроматографии, J. Chromatogr. , 1988, том. 447, стр. 305–314.Артикул КАС Google Scholar
Лайонс Дж. М., Уитон Т. А. и Пратт Ю. К., Связь между физической природой митохондриальных мембран и чувствительностью растений к охлаждению, Физиол. растений. , 1964, том. 39, стр. 262–268.
Артикул пабмед КАС Google Scholar
Вонгватанарат, П., Майклсон, Л.В., Картер, А.В., Лазарус, К.М., Гриффитс, Г., Стобарт, А.К., Арчер, Д.Б., и Маккензи, Д.А., Две жирные кислоты 9 -Гены десатуразы ole1 и ole2 из Mortirella alpine Комплемент дрожжей ole1 Mutation, Microbiology , 1998, vol. 145, стр. 2939–2946.
Google Scholar
Макаренко С.П., Константинов Ю.М., Шмаков В.Н., Хотимченко С.В., Коненкина Т.А. Жирнокислотный состав липидов лиственницы ( Larix gmelinii (Rupr.) Rupr.), биол. член , 2006, том. 23, стр. 470–475.
КАС Google Scholar
Фукучи-Мизутани М., Мизутани М., Танака Ю., Кусуми Т. и Охта Д., Микросомальный перенос электрона у высших растений: клонирование и гетерологичная экспрессия НАДН-цитохрома b 5 Редуктаза из Arabidopsis, Plant Physiol. , 1999, том. 119, стр.
 , Телен Дж.Дж., Пэддок Т., Салас Дж.Дж., Сэвидж Л., Милкампс А. , Mhaske, V.B., Cho, Y., and Ohlrogge, J.B., Гены арабидопсиса, участвующие в метаболизме ациллипидов. Перепись кандидатов 2003 г., исследование распределения тегов экспрессируемых последовательностей в органах и база данных в Интернете, Физиол растений. , 2003, том. 132, стр. 681–697.
, Телен Дж.Дж., Пэддок Т., Салас Дж.Дж., Сэвидж Л., Милкампс А. , Mhaske, V.B., Cho, Y., and Ohlrogge, J.B., Гены арабидопсиса, участвующие в метаболизме ациллипидов. Перепись кандидатов 2003 г., исследование распределения тегов экспрессируемых последовательностей в органах и база данных в Интернете, Физиол растений. , 2003, том. 132, стр. 681–697.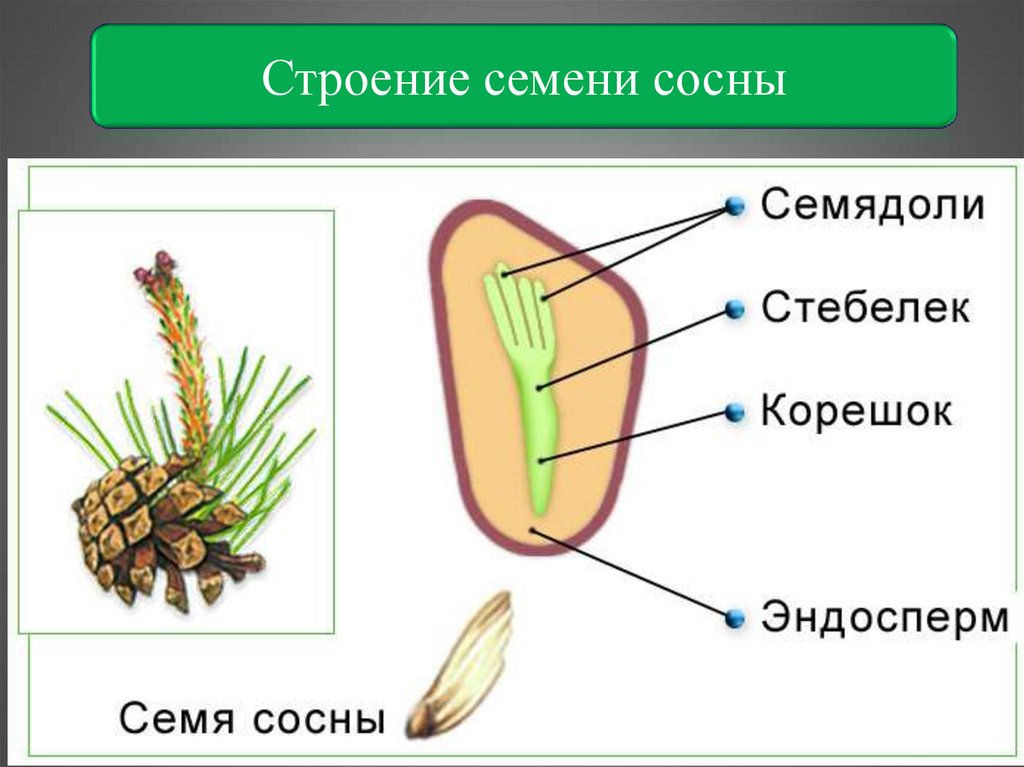 «>
«>Schultz, D.J., Suh, M.C., and Ohlrogge, J., Стеароил-ацил-носитель белка и необычный ацил-ацил-носитель белка-десатуразы по-разному влияют на активность ферредоксина, Plant Physiol. , 2000, том. 124, стр. 681–692.
Артикул пабмед КАС Google Scholar
 А., Последние достижения в области биохимии и молекулярной биологии жирных ацилдесатураз, Prog. Липид Рез. , 1998, том. 37, стр. 73–117.
А., Последние достижения в области биохимии и молекулярной биологии жирных ацилдесатураз, Prog. Липид Рез. , 1998, том. 37, стр. 73–117.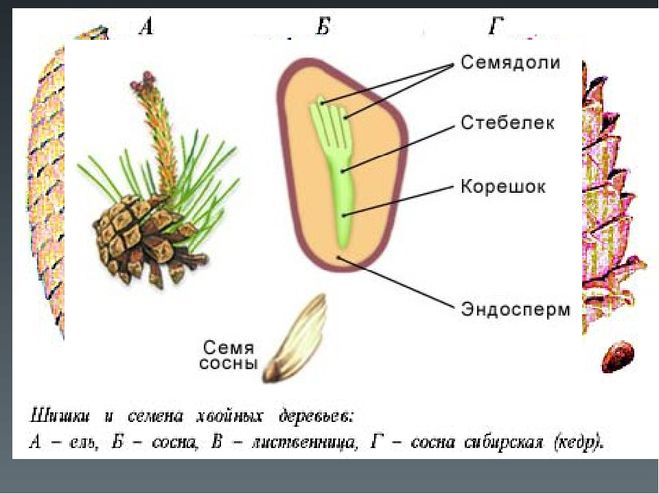 «>
«>Хамада, Т., Иба, К., и Шимада, Т., Снижение содержания триеновых жирных кислот путем экспрессии двухцепочечной РНК пластидного гена десатуразы жирных кислот ω3 в трансгенном табаке, Biotechnol. лат. , 2006, том. 28, стр. 779–785.
Артикул пабмед КАС Google Scholar
 , 1995, том. 715, стр. 317–324.
, 1995, том. 715, стр. 317–324.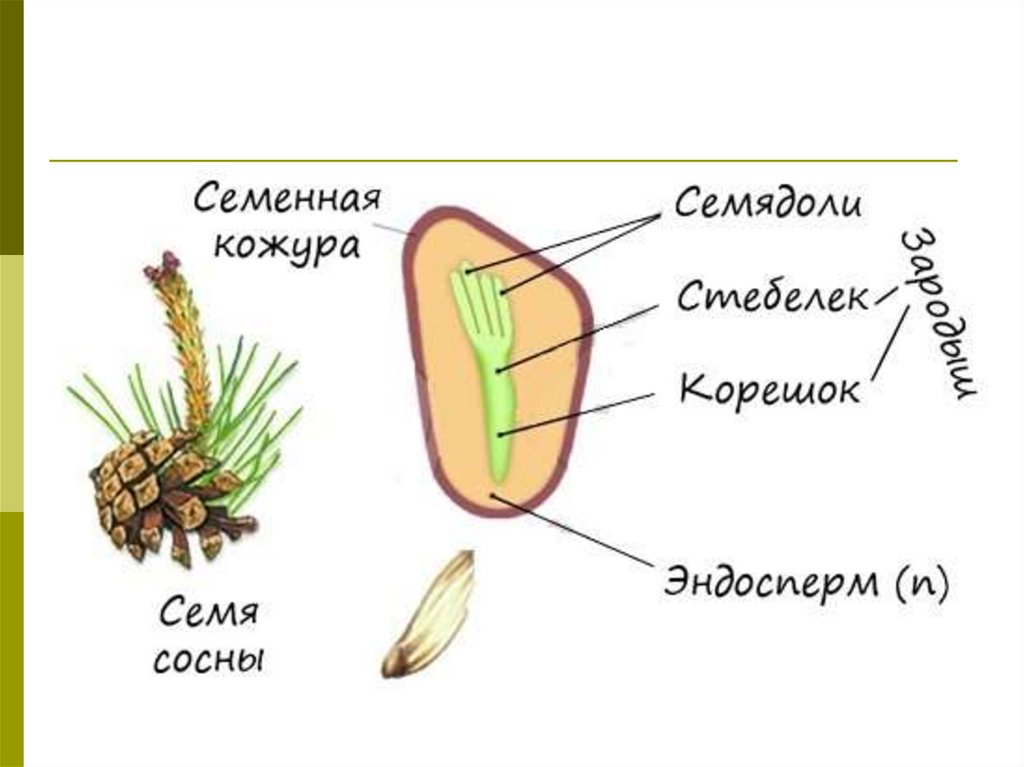 «>
«>Монгран, С., Бадок, А., Патуй, Б., Лакомблез, К., Шаван, М., Кассань, К., и Бессуле, Ж.-Ж., Таксономия голосеменных: многофакторный анализ Состав жирных кислот листьев, Фитохимия , 2001, vol. 58, стр. 101–115.
Артикул пабмед КАС Google Scholar
 «>
«>Domergque, F., Lerchl, J., Zahringer, U., и Heinz, E., Клонирование и функциональная характеристика Phaeodactylum tricornutum Передняя десатураза, участвующая в биосинтезе эйкозапентаеновой кислоты, Eur. Дж. Биохим. , 2002, том. 269, стр. 4105–4113.
Артикул КАС Google Scholar

 36, стр. 439–451.
36, стр. 439–451.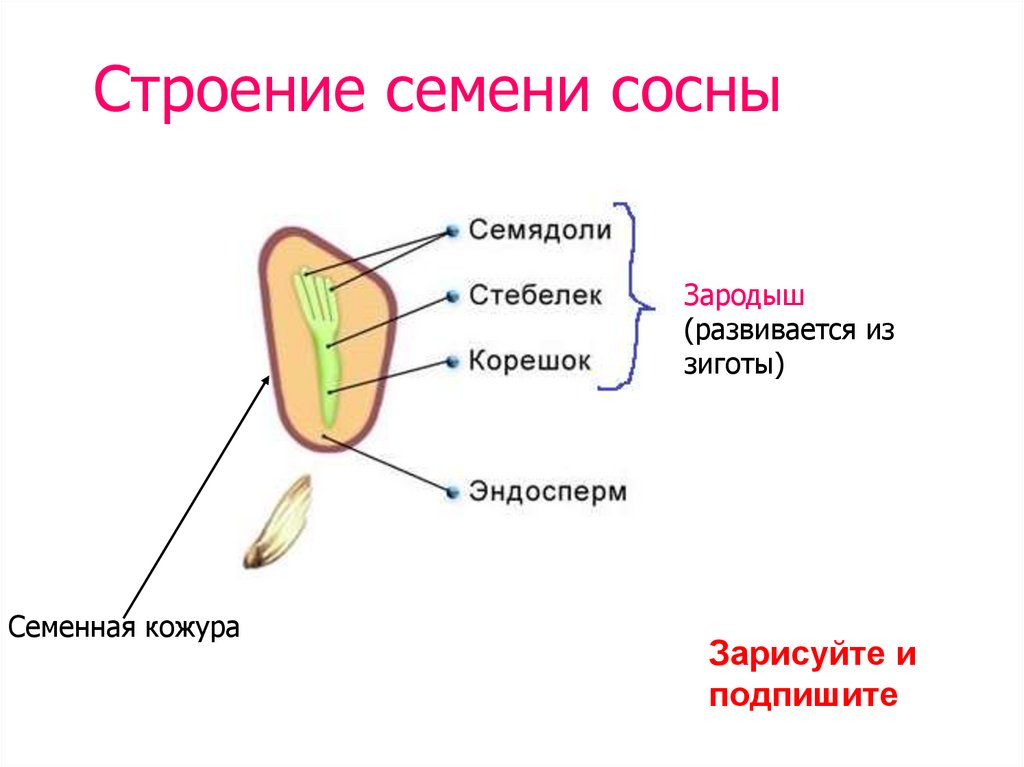 «>
«>Бергер А., Моннар И., Баур М., Шарбоннет С., Сафонова И. и Жомар А. Противовоспалительные свойства эпидермиса 5,11,14 20:3: Воздействие на эпидерму уха мыши, PGE 2 Уровни в культивируемых кератиноцитах и активация PPAR, Здоровье липидов , 2002, vol. 1, стр. 1–12.
Артикул Google Scholar
 W., Эквивалентные длины цепей производных метилового эфира жирных кислот в газовой хроматографии, J. Chromatogr. , 1988, том. 447, стр. 305–314.
W., Эквивалентные длины цепей производных метилового эфира жирных кислот в газовой хроматографии, J. Chromatogr. , 1988, том. 447, стр. 305–314.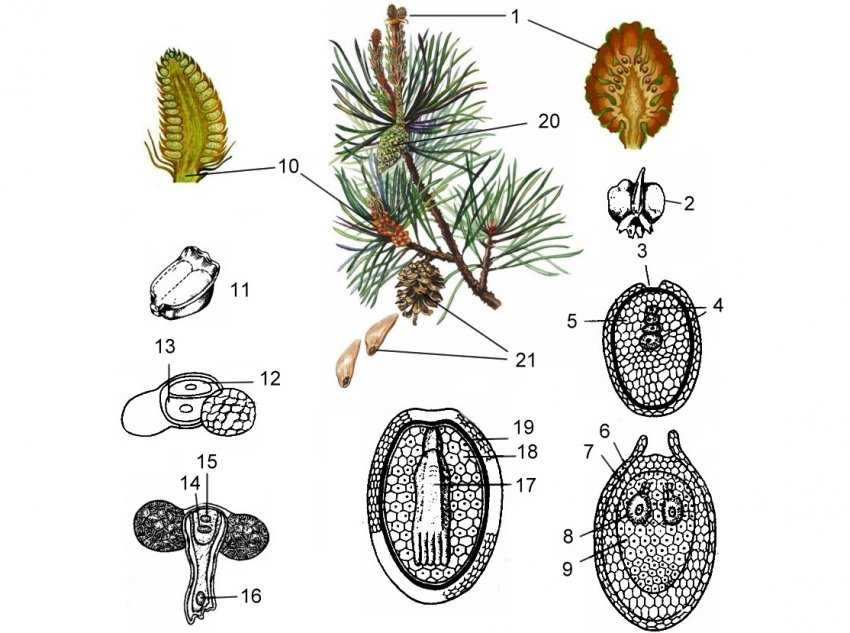 «>
«>O’Hara, P., Skabas, A.R., and Fawcett, T., Гены биосинтеза жирных кислот и липидов экспрессируются в постоянных молярных соотношениях, но на разных абсолютных уровнях во время эмбриогенеза, Plant Physiol. , 2002, том. 129, стр. 310–320.
Артикул пабмед КАС Google Scholar
Leave A Comment