
7 класс. Биология. Животные.Кишечнополостные . Классы Кишечнополостных — Кишечнополостные
Комментарии преподавателяПредставители животного мира, которые принадлежат к типу кишечнополостных, являются довольно древними жителями нашей планеты. Их можно легко охарактеризовать наличием радиальной симметрии, кишечной полости и ротового отверстия.
Большинство представителей этого типа выбирают для себя водную сферу обитания. Необходимо обратить внимание, что кишечнополостный тип разделяется на:
- сидячие формы или бентос;
- плавающие формы или планктон.
Говоря о представителях кишечнополостных, идет речь о тех хищниках, которые питаются мелкими рачками, мальками рыб, а также водными насекомыми, хотя среди них также встречаются и паразитические виды.
Благодаря тому, что этот тип не имеет настоящих органов, он позволяет изучить и проанализировать весь процесс эволюции, который произошел в животном мире. Кишечнополостные прародители всех высших многоклеточных животных.
Кишечнополостные прародители всех высших многоклеточных животных.
На сегодняшний день существует приблизительно 900 видов кишечнополостных животных, которые прикрепляются к различным предметам, находящиеся под водой, или двигаются медленно в водном пространстве. Они обладают бокаловидной формой (идет речь о полипах). Что же касается плавающих животных этого типа, то они имеют форму колокола или зонтика (идет речь о медузах).
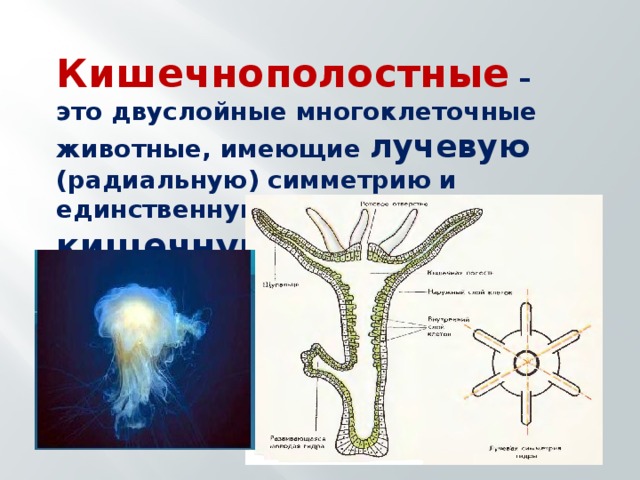
Внутреннее строение кишечнополостных животных
Кишечнополостные животные обладают радиальной или лучевой симметрией. Таким образом, через все их тело можно провести до восьми плоскостей, которые смогут поделить весь организм на полностью одинаковые части. Если сравнивать тело выше названных животных, то оно скорее похоже на двухслойный мешок.
Во внутреннем строении организма кишечнополостных развита исключительно гастральная часть, которая выполняет роль первичного кишечника. Этот так называемый кишечник имеет одно-единственное отверстие, которое играет роль и ротового, и анального отверстия.
Большинство представителей этого типа обладает довольно большим количеством специальных каналов, которые, отходя от гастральной полости образуют с желудком сложную систему (такую систему ученые называют гастроваскулярной или желудочно-сосудистой).
Какую роль выполняют кишечнополостные?
Довольно яркую роль кишечнополостные играют в судоходстве, где они снижают скорость судна благодаря тому, что способны прикрепляться к различным предметам под водой, создавая мохнатую «шубку». Иногда даже они становились препятствием для нормальной подачи воды.
Если говорить о тех представителях типа кишечнополостных, которые обитают на глубине, то они приводят к следующим последствиям:
- обеспечивают человеку сильные и болезненные ожоги;
- вызывают серьезные заболевания;
- образуют коралловые рифы и целые острова;
- паразитируют на других животных.
- ***************************************************************************8
На предыдущем уроке мы разобрали губок. Вы уже знаете, что они не имеют настоящего тканевого строения. Принадлежность губок к многоклеточным животным вызывает споры среди ученых. Сегодня мы приступаем к изучению кишечнополостных, которые, несомненно, являются настоящими многоклеточными животными. Их клетки формируют ткани.
Вы уже знаете, что они не имеют настоящего тканевого строения. Принадлежность губок к многоклеточным животным вызывает споры среди ученых. Сегодня мы приступаем к изучению кишечнополостных, которые, несомненно, являются настоящими многоклеточными животными. Их клетки формируют ткани.
Тело кишечнополостных представляет собой мешок, стенка которого имеет два слоя. Между ними находится студенистое неклеточное вещество. Полость мешка – кишечная полость, там происходит переваривание пищи. Она имеет одно отверстие – рот. Наружный слой клеток – эктодерма. В ней расположены разные клетки, в том числе нервные и стрекательные. Все нервные клетки соединены между собой в сетевидное нервное сплетение. Нервные, стрекательные и половые клетки формируются из промежуточных клеток.
Промежуточные клетки как бы запасные, они обеспечивают замену старым, отмирающим клеткам. Второе название кишечнополостных – стрекающие, получено ими за наличие стрекательных клеток. Обычно такая клетка снабжена чувствительным волоском, который отвечает за выброс стрекательной нити. Через неё в тело жертвы или хищника попадает яд.
Второе название кишечнополостных – стрекающие, получено ими за наличие стрекательных клеток. Обычно такая клетка снабжена чувствительным волоском, который отвечает за выброс стрекательной нити. Через неё в тело жертвы или хищника попадает яд.
Энтодерма – внутренний слой, состоящий из железистых и эпителиально-мускульных клеток. Железистые клетки выделяют пищеварительный секрет. Эпителиально-мускульные клетки обеспечивают сокращение или растяжение всего тела и отдельных его частей. Клетки энтодермы имеют жгутики.
Пищевые частицы захватываются и перевариваются внутри клеток. Почти все клетки тела кишечнополостных контактируют с водой. Вода обеспечивает их газообмен и выделение.
Кишечнополостные имеют лучевую симметрию, которая позволяет провести несколько плоскостей через тело животного, поделив его каждой плоскостью на две равные части.
Один из характернейших признаков кишечнополостных – щупальца. Это длинные и тонкие придатки, способные сильно вытягиваться и сокращаться. Они служат для ловли добычи и защиты. Почти все кишечнополостные – хищники: питаются мелкими животными.
Для кишечнополостных характерно чередование поколений – сидячего (полипы) и свободноплавающего (медузы). Внешне медуза и полип мало похожи друг на друга. Размножение кишечнополостных происходит половым и бесполым путем.
Типичный жизненный цикл выглядит следующим образом. После оплодотворения яйцеклетки образуется подвижная личинка. Она перемещается в толще воды, а затем, прикрепившись ко дну, превращается в полип. Полип размножается бесполым путём, иногда образуя колонию. В дальнейшем полипы отпочковывают медуз, которые размножаются половым путем.
Известно около 11 тыс. видов кишечнополостных. Большинство обитает в морях и океанах и лишь немногие в пресных водах. Кишечнополостные могут вести колониальный или одиночный образ жизни. Колонии организмов, имеющих известковый скелет, образуют рифы.
Другие колониальные кишечнополостные могут плавать: например, португальский кораблик. Одиночные особи существуют в форме полипа или медузы. Полипы малоподвижны, часто ведут сидячий образ жизни. Медузы обычно свободно плавают в толще воды.
В типе кишечнополостных три класса. О них мы поговорим на следующем уроке.
Дополнительный материал
Симбионты кишечнополостных
Для кишечнополостных характерны различные типы симбиоза – комменсализм, мутуализм и паразитизм.
Многие кишечнополостные ведут прикреплённый образ жизни и имеют прочный скелет. Хорошим примером тому служат коралловые полипы. Такие кишечнополостные часто служат субстратом для организмов-обрастателей, в том числе для других представителей типа. Кишечнополостные могут обрастать поверхность тела асцидий, губок, слоевища водорослей.
Сожительство некоторых гидроидных полипов и раков-отшельников выгодно обоим организмам. Постоянное передвижение рака способствует лучшему газообмену гидроида, а остатки пищи, пожираемой раком, поедаются полипом. Вместе с тем поросль гидроида маскирует раковину.
Яркий пример мутуализма – сожительство актинии адамсии и рака-отшельника. Защитить себя рак может, только вступив в такой симбиоз. Он получает надёжную защиту стрекательных клеток актинии, а актиния получает возможность путешествовать по водоему и питаться остатками пищи рака.
Многие кишечнополостные питаются в основном за счет одноклеточных водорослей, живущих внутри их тела.
Существует любопытный симбиоз актиний и рыб-клоунов. Эти рыбки при любой опасности прячутся среди щупалец. Актиния таким образом защищает их и в свою очередь получает питание из остатков пищи рыбок. Слизь, покрывающая тело рыб-клоунов, препятствует срабатыванию стрекательных клеток актинии против них.
Среди кишечнополостных имеется небольшое число настоящих паразитов. Так, некоторые гидроидные медузы способны питаться за счет других медуз, а гидроидный полип-полиподиум развивается внутри икринок осетровых рыб.
Рекорды кишечнополостных
Самые мелкие представители этого типа имеют длину около 1 мм, а самые крупные, такие как медуза цианея, имеют щупальца длиной до 30 м.
Кишечнополостные могут обитать на прибрежных мелководьях, на совсем небольшой глубине. Однако некоторые актинии обитают на максимальных глубинах, до 10 км.
Однако некоторые актинии обитают на максимальных глубинах, до 10 км.
Площадь коралловых островов и обнажающихся при отливе рифов равна 8 млн км2, что немного больше, например, площади материка Австралия.
Гребневики
Ближайшими родственниками кишечнополостных являются гребневики. Это морские, преимущественно планктонные животные. Их название связано с наличием характерных «гребней» – рядов гребных пластинок, образованных сросшимися ресничками. Размеры колеблются от 2–3 мм до 3 метров. Известно около 150 видов.
Гребневики обладают радиальной двулучевой симметрией. Единственное, что отличает их в этом смысле от двусторонне-симметричных животных, – отсутствие выраженных спинной и брюшной сторон тела.
Тело гребневиков снаружи покрыто однослойным эпителием. Имеется восемь рядов ресничных клеток, образующих гребные пластинки – гребни из сросшихся ресничек.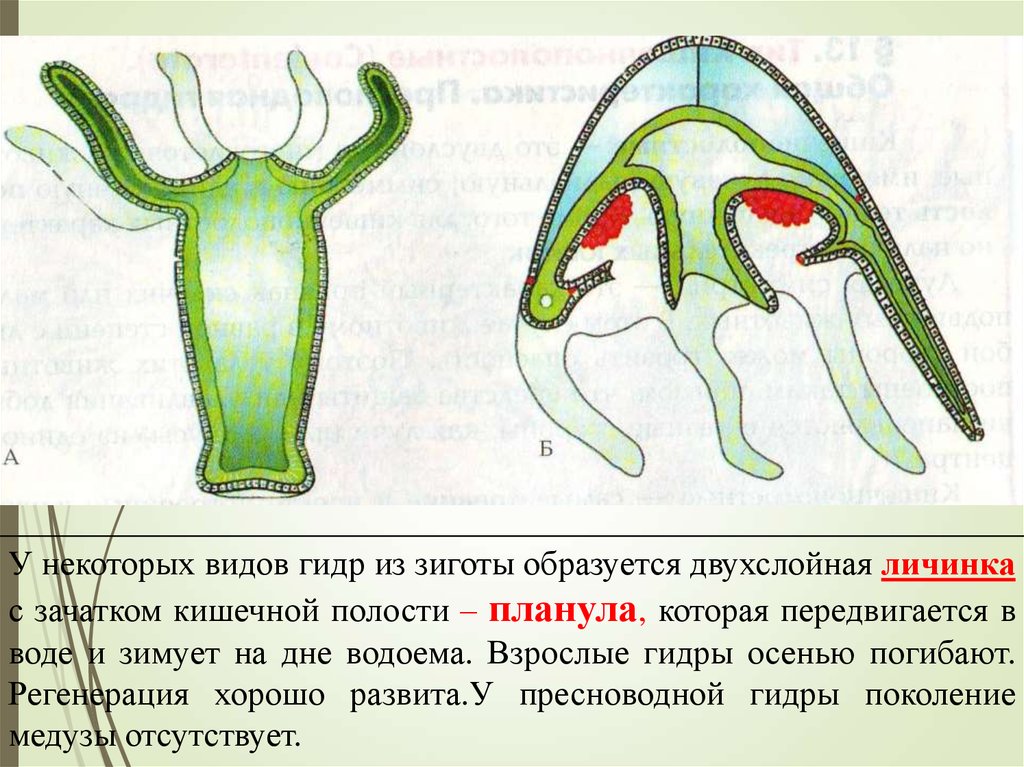 У щупальцевых гребневиков из специальных углублений растут два щупальца. В покровном эпителии щупалец имеются клетки, снабжённые клейкими капсулами. Они позволяют гребневикам ловить мелкие планктонные организмы.
У щупальцевых гребневиков из специальных углублений растут два щупальца. В покровном эпителии щупалец имеются клетки, снабжённые клейкими капсулами. Они позволяют гребневикам ловить мелкие планктонные организмы.
Кишечная полость открывается единственным отверстием – ртом.
Между покровным эпителием и полостью кишечника расположен толстый слой студенистого межклеточного вещества.
Нервная система представлена нервным сплетением, расположенным под покровным эпителием.
Большинство гребневиков – хищники. Замечательна способность некоторых из них к свечению и радужному преломлению света на гребных пластинках.
источник конспекта -http://interneturok.ru/ru/school/biology/7-klass/zhivotnye-kishechnopolostnye/kishechnopolostnye?seconds=0&chapter_id=78
источник видео — http://www.youtube.com/watch?v=dBP40d0sG8w
источник видео — http://www. youtube.com/watch?v=Z_HAvMAPOM4
youtube.com/watch?v=Z_HAvMAPOM4
источник видео — http://www.youtube.com/watch?v=sHqse68IwkU
источник видео — http://www.youtube.com/watch?v=hHbKB7R3nk8
источник видео — http://www.youtube.com/watch?v=fObn7iA3OJU
источник презентации — http://ppt4web.ru/biologija/kishechnopolostnye1.html
http://www.animals-wild.ru/964-kishechnopolostnye-zhivotnye.html
Двусторонняя и лучевая симметрия тела. Симметрия животных
Кишечнополостные
— древние животные, обитавшие еще в кембрийском море. Отсутствие настоящих органов и тканей дает основание считать их (наряду с губками — первейшими многоклеточными организмами) наиболее примитивными многоклеточными животными. Большинство видов обитает в морях и океанах, лишь немногие живут в пресных водоемах.Класс гидроидные
Гидра — пресноводный полип («полип» означает «многоног»), обитающий в чистой проточной воде. Тело гидры цилиндрической формы размером от 1 до 1,5 см (причем тело обычно не превышает в длину 5-7 мм, зато щупальца способны вытягиваться на несколько сантиметров) .На одном конце находится подошва, служащая для прикрепления к подводным предметам, на противоположном — ротовое отверстие, окруженное длинными щупальцами (5-12). Гидра ведет малоподвижный образ жизни. Стенки тела гидры двухслойны и представлены эктодермой и энтодермой, между которыми располагается мезоглея. Тело гидры обладает радиальной симметрией, или лучевой симметрией. Лучевая симметрия — особый порядок расположения частей тела животного (у гидры — щупалец) по отношению к оси его симметрии, при котором они расходятся от нее подобно лучам от источника света. В нем можно различить главную продольную ось, вокруг которой в радиальном порядке размещения различные органы. Через тело можно провести несколько (2-4-6-8- и т.д.) плоскостей симметрии. Радиальная симметрия тела возникла в процессе эволюции у животных, которые вели прикрепленный образ жизни, т.к. жертва может появиться с любой стороны, лучеобразно расставленные щупальца лучше всего соответствуют такому способу охоты. Сидячий образ жизни вели предки кишечнополостных.
Особенности строения клетки многоклеточного животного организма.
Тело многоклеточных животных состоит из множества клеток и их производных. Клетки дифференцированы по строению и функции, они утратили самостоятельность, поскольку представляют собой лишь составные части целостного организма. Для жизненного цикла многоклеточных характерно сложное индивидуальное развитие (онтогенез), в процессе которого оплодотворенное яйцо дробится на множество клеток (бластомеров), затем дифференцирующихся на зародышевые листки и зачатки органов. В дальнейшем из зародыша развивается взрослый организм. (При партеногенезе — из неоплодотворенного яйца формируется взрослый организм).Всех многоклеточных можно разделить на 2 группы:
а) лучистые
(радиально-симметричные), или двухслойные. Им свойственно наличие нескольких плоскостей симметрии и радиальное расположение органов вокруг главной оси тела. В процессе онтогенеза у них образуется только 2 зародышевых листка — эктодерма и энтодерма.б) трехслойные, или двусторонне симметричные,
в отличие от лучистых, имеют одну плоскость симметрии, которая делит их тело на 2 зеркально одинаковые половинки (левую и правую). У них, кроме эктодермы и энтодермы, образуется и 3-ий зародышевый листок — мезодерма. Из него формируются многие внутренние органы.
 На одном конце находится подошва, служащая для прикрепления к подводным предметам, на противоположном — ротовое отверстие, окруженное длинными щупальцами (5-12). Гидра ведет малоподвижный образ жизни. Стенки тела гидры двухслойны и представлены эктодермой и энтодермой, между которыми располагается мезоглея. Тело гидры обладает радиальной симметрией, или лучевой симметрией. Лучевая симметрия — особый порядок расположения частей тела животного (у гидры — щупалец) по отношению к оси его симметрии, при котором они расходятся от нее подобно лучам от источника света. В нем можно различить главную продольную ось, вокруг которой в радиальном порядке размещения различные органы. Через тело можно провести несколько (2-4-6-8- и т.д.) плоскостей симметрии. Радиальная симметрия тела возникла в процессе эволюции у животных, которые вели прикрепленный образ жизни, т.к. жертва может появиться с любой стороны, лучеобразно расставленные щупальца лучше всего соответствуют такому способу охоты. Сидячий образ жизни вели предки кишечнополостных.
На одном конце находится подошва, служащая для прикрепления к подводным предметам, на противоположном — ротовое отверстие, окруженное длинными щупальцами (5-12). Гидра ведет малоподвижный образ жизни. Стенки тела гидры двухслойны и представлены эктодермой и энтодермой, между которыми располагается мезоглея. Тело гидры обладает радиальной симметрией, или лучевой симметрией. Лучевая симметрия — особый порядок расположения частей тела животного (у гидры — щупалец) по отношению к оси его симметрии, при котором они расходятся от нее подобно лучам от источника света. В нем можно различить главную продольную ось, вокруг которой в радиальном порядке размещения различные органы. Через тело можно провести несколько (2-4-6-8- и т.д.) плоскостей симметрии. Радиальная симметрия тела возникла в процессе эволюции у животных, которые вели прикрепленный образ жизни, т.к. жертва может появиться с любой стороны, лучеобразно расставленные щупальца лучше всего соответствуют такому способу охоты. Сидячий образ жизни вели предки кишечнополостных.
 Сюда относятся все представители типа кишечнополостных;
Сюда относятся все представители типа кишечнополостных;симметрия подобия;
радиально-лучевая симметрия
Отражение – это наиболее известная и чаще других встречающаяся в природе разновидность симметрии. Зеркало в точности воспроизводит то, что оно «видит», но рассмотренный порядок является обращенным: правая рука у вашего двойника в действительности окажется левой, так как пальцы расположены на ней в обратном порядке.
Зеркальную симметрию
можно обнаружить повсюду: в листьях и цветах растений, архитектуре, орнаментах. Человеческое тело, если говорить лишь о наружном виде, обладает зеркальной симметрией, хотя и не вполне строгой. Более того, зеркальная симметрия свойственна телам почти всех живых существ, и такое совпадение отнюдь не случайно.
Зеркальной симметрией обладает все, допускающее разбиение на две зеркально равные половинки. Каждая из половинок служит зеркальным отражением другой, а разделяющая их плоскость называется плоскостью зеркального отражения, или зеркальной плоскостью. Эту плоскость можно назвать элементом симметрии, а соответствующую операцию – операцией симметрии.
Поворотная симметрия.
Внешний вид узора не изменится, если его повернуть на некоторый угол вокруг оси. Симметрия, возникающая при этом, называется поворотной симметрией. Во многих танцах фигуры основаны на вращательных движениях, нередко совершаемых только в одну сторону (т.е. без отражения), например, хороводы.
Листья и цветы многих растений обнаруживают радиальную симметрию. Это такая симметрия, при которой лист или цветок, поворачиваясь вокруг оси симметрии, переходит в себя. На поперечных сечениях тканей, образующих корень или стебель растения, отчетливо бывает видна радиальная симметрия. Соцветия многих цветков также обладают радиальной симметрией.
Отражение в центре симметрии.
Примером объекта наивысшей симметрии, характеризующим эту операцию симметрии, является шар. Шаровые формы распространены в природе достаточно широко. Они обычны в атмосфере (капли тумана, облака), гидросфере (различные микроорганизмы), литосфере и космосе. Шаровую форму имеют споры и пыльца растений, капли воды, выпущенной в состоянии невесомости на космическом корабле. На метагалактическом уровне наиболее крупными шаровыми структурами являются галактики шаровой формы. Чем плотнее скопление галактик, тем ближе оно к шаровой форме. Звездные скопления – тоже шаровые формы.
Трансляция, или перенос фигуры на расстояние.
Трансляция, или параллельный перенос фигуры на расстояние – это любой неограниченно повторяющийся узор. Она может быть одномерной, двумерной, трехмерной. Трансляция в одном и том же или противоположных направлениях образует одномерный узор. Трансляция по двум непараллельным направлениям образует двумерный узор. Паркетные полы, узоры на обоях, кружевные ленты, дорожки, вымощенные кирпичом или плитками, кристаллические фигуры образуют узоры, которые не имеют естественных границ.
Винтовые повороты.
Трансляцию можно комбинировать с отражением или поворотом, при этом возникают новые операции симметрии. Поворот на определенное число градусов, сопровождаемый трансляцией на расстояние вдоль оси поворота, порождает винтовую симметрию – симметрию винтовой лестницы. Пример винтовой симметрии – расположение листьев на стебле многих растений.
Головка подсолнечника имеет отростки, расположенные по геометрическим спиралям, раскручивающимся от центра наружу. Самые молодые члены спирали находятся в центре.
В таких системах можно заметить два семейства спиралей, раскручивающихся в противоположные стороны и пересекающихся под углами, близкими к прямым.
Вслед за Гете, который говорил о стремлении природы к спирали, можно предположить, что движение это осуществляется по логарифмической спирали, начиная всякий раз с центральной, неподвижной точки и сочетая поступательное движение (растяжение) с поворотом вращения.
Симметрия подобия.
К перечисленным выше операциям симметрии можно добавить операцию симметрии подобия, представляющую собой своеобразные аналогии трансляций, отражений в плоскостях, повороты вокруг осей с той только разницей, что они связаны с одновременным увеличением или уменьшением подобных частей фигуры и расстояний между ними.
Симметрия подобия, осуществляющаяся в пространстве и во времени, повсеместно проявляется в природе на всем, что растет. Именно к растущим формам относятся бесчисленные фигуры растений, животных и кристаллов. Форма древесного ствола – коническая, сильно вытянутая. Ветви обычно располагаются вокруг ствола по винтовой линии. Это не простая винтовая линия: она постепенно суживается к вершине. Да и сами ветви уменьшаются по мере приближения к вершине дерева. Следовательно, здесь мы имеем дело с винтовой осью симметрии подобия.
Живая природа в любых ее проявлениях обнаруживает одну и ту же цель: всякий живой предмет повторяет себя в себе подобном. Главной задачей жизни является Жизнь, а доступная форма бытия заключается в существовании отдельных целостных организмов.
Радиально-лучевая симметрия в природе.
Внимательно приглядевшись к обступающей нас природе, можно увидеть общее даже в самых незначительных вещах и деталях. Форма листа дерева не является случайной: она строго закономерна.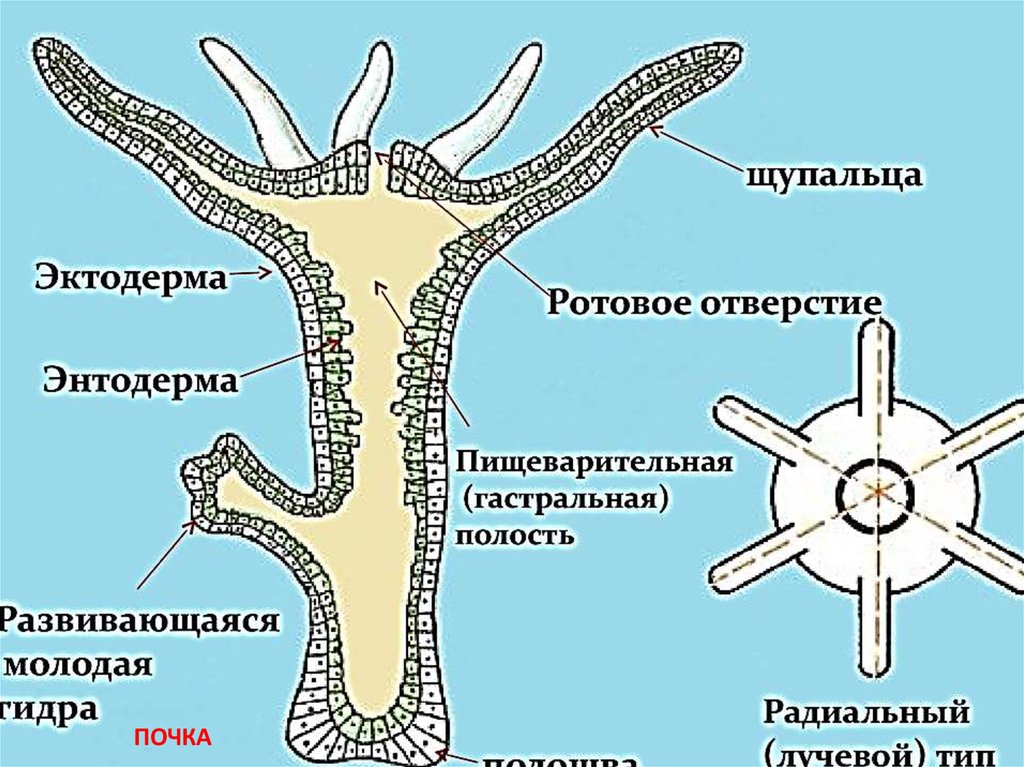 Листок как бы склеен из двух более или менее одинаковых половинок, одна из которых расположена зеркально относительно другой. Симметрия листка упорно повторяется, будь то гусеница, бабочка, жучок и т.п.
Листок как бы склеен из двух более или менее одинаковых половинок, одна из которых расположена зеркально относительно другой. Симметрия листка упорно повторяется, будь то гусеница, бабочка, жучок и т.п.
Радиально-лучевой симметрией обладают цветы, грибы, деревья, фонтаны. Здесь можно отметить, что на не сорванных цветах и грибах, растущих деревьях, бьющем фонтане или столбе паров плоскости симметрии ориентированы всегда вертикально.
Таким образом, можно сформулировать в несколько упрощенном и схематизированном виде общий закон, ярко и повсеместно проявляющийся в природе: все, что растет или движется по вертикали, т.е. вверх или вниз относительно земной поверхности, подчиняется радиально-лучевой симметрии в виде веера пересекающихся плоскостей симметрии. Все то, что растет и движется горизонтально или наклонно по отношению к земной поверхности, подчиняется билатеральной симметрии, симметрии листка. Этому всеобщему закону подчиняются не только цветы, животные, легкоподвижные жидкости и газы, но и камни. Этот закон влияет на изменчивые формы облаков. В безветренный день они имеют куполовидную форму с более или менее ясно выраженной радиально-лучевой симметрией.
Этот закон влияет на изменчивые формы облаков. В безветренный день они имеют куполовидную форму с более или менее ясно выраженной радиально-лучевой симметрией.
На вопрос Что такое лучевая симметрия? заданный автором Катя Черных лучший ответ это Лучевая (радиальная) симметрия — форма симметрии, при которой тело (или фигура) совпадает само с собой при вращении объекта вокруг определённой точки или прямой.
Как правило, у многоклеточных животных два конца (полюса) единственной оси симметрии неравноценны (например, у медуз на одном полюсе (оральном) находится рот, а на противоположном (аборальном) — верхушка колокола. Такая симметрия (вариант радиальной симметрии) в сравнительной анатомии называется одноосно-гетеропольной. В двухмерной проекции радиальная симметрия может сохраняться, если ось симметрии направлена перпендикулярно к проекционной плоскости. Иными словами, сохранение радиальной симметрии зависит от угла наблюдения.
Лучевая симметрия характерна, в основном, для кишечнополостных животных. Кишечнополостным, как сидячим, так и пелагическим (медузы) , свойственна радиально-осевая симметрия, при которой сходственные части расположены вокруг оси вращения, причем эта симметрия может быть самого различного порядка в зависимости от того, на какой угол следует повернуть тело животного, чтобы новое положение совпало с исходным. Таким образом, может получаться 4-, 6-, 8лучевая симметрия и более, до симметрии порядка бесконечности. У радиолярий встречается радиально-осевая симметрия с одинаковыми полюсами, или, как говорят, гомополярная. У кишечнополостных — гетерополярная осевая симметрия: один полюс симметрии несет рот и щупальца (оральный) , другой (а б о р а л ь н ы й) служит для прикрепления (стадия полипа) , или у плавающих форм несет орган чувств (ктенофоры) , или ничем не вооружен (медузы) .
Кишечнополостным, как сидячим, так и пелагическим (медузы) , свойственна радиально-осевая симметрия, при которой сходственные части расположены вокруг оси вращения, причем эта симметрия может быть самого различного порядка в зависимости от того, на какой угол следует повернуть тело животного, чтобы новое положение совпало с исходным. Таким образом, может получаться 4-, 6-, 8лучевая симметрия и более, до симметрии порядка бесконечности. У радиолярий встречается радиально-осевая симметрия с одинаковыми полюсами, или, как говорят, гомополярная. У кишечнополостных — гетерополярная осевая симметрия: один полюс симметрии несет рот и щупальца (оральный) , другой (а б о р а л ь н ы й) служит для прикрепления (стадия полипа) , или у плавающих форм несет орган чувств (ктенофоры) , или ничем не вооружен (медузы) .
У некоторых медуз на этой аборальной стороне образуется стебелек для прикрепления к подводным предметам (Lucernariida). Нарушение радиал ьно-осевой симметрии возникает при уменьшении числа шупалец или изменении формы ротовой щели, пищевода и разветвлений пищеварительной системы. Количество щупалец может уменьшаться до одного (Мопоbrachium), и тогда их радиальное расположение сменяется двубоковым. Глотка может сплющиваться, и тогда тоже получается двубоковая симметрия, этому способствует и образование в глотке сифоноглифов (желобок вдоль глотки) .
Количество щупалец может уменьшаться до одного (Мопоbrachium), и тогда их радиальное расположение сменяется двубоковым. Глотка может сплющиваться, и тогда тоже получается двубоковая симметрия, этому способствует и образование в глотке сифоноглифов (желобок вдоль глотки) .
Наибольшее усложнение радиально-осевой симметрии наблюдается у ктенофор, где, помимо 8-лучевой симметрии, в расположении отдельных частей тела и органов наблюдается 4-лучевая и двубоковая симметрия. Это весьма существенный момент, так как большинство зоологов именно от ктенофорообразных предков выводит оба ствола высших животных, как первично-, так и вторичноротых.
Гетерополярная радиальноосевая симметрия вполне соответствует образу жизни кишечнополостных — неподвижному существованию в прикрепленном положении или медленному плаванию при помощи реактивного движения.
С другой стороны, от сложного типа радиально-осевой симметрии ктенофор можно перейти к двусторонней симметрии, или, как говорят, симметрии зеркального изображения, единственного плана симметрии трехслойных животных, симметрии быстрого движения, с выработкой переднего по движению конца тела, с центральным мозговым скоплением и основными органами чувств, спинной и брюшной, правой и левой сторонами тела.
..подробнее — . berl. ru/article/ nauka/cimmetria_u_givotnyh.htm здесь (уберите про)
При сравнении представителей разных систематических групп создается впечатление, что они необычайно разнообразны. Тем не менее различия животных не бесконечны.
Как было показано Ч. Дарвином, множество родственных групп животных произошло от одной предковой линии. «Спускаясь» от кончиков ветвей родословного древа животных к узлам ветвления и в конечном итоге к стволам, мы улавливаем общность многих организмов в их планах строения. Ученые установили несколько таких планов, в которые укладывается большое число вариантов. Следует помнить, что план строения — это то общее, что характерно для множества групп. Варианты же — это частности, детали, которые первыми бросаются в глаза и нередко маскируют принадлежность животного к определенному типу. Общность планов строения указывает на гомологию — сходство, основанное на родстве организмов.
За немногими исключениями, животные отличаются симметричным строением. Различают два типа симметрии — радиальную, или лучевую, и билатеральную, или двустороннюю. Оба этих типа одновременно встречаются только у беспозвоночных животных. Позвоночные — всегда билатеральны.
Различают два типа симметрии — радиальную, или лучевую, и билатеральную, или двустороннюю. Оба этих типа одновременно встречаются только у беспозвоночных животных. Позвоночные — всегда билатеральны.
В теле радиально-симметричного животного (рис. 1) можно различить главную продольную ось, вокруг которой в радиальном (лучевом) порядке расположены органы.
Порядок радиальной симметрии зависит от числа повторяющихся органов. Если вокруг этой воображаемой главной оси расположено 5 одинаковых органов, то симметрия называется пятилучевой, если 4 — четырехлучевой и т. д. Вследствие этого через тело животного (его центр) можно провести строго определенное
число плоскостей симметрии, которыми тело делится на две половины, зеркально отображающие друг друга. Радиальная симметрия имеет две разновидности: радиально-лучевую и радиаль-но-осевую симметрии.Радиально-лучевая симметрия наблюдается у многих организмов, взвешенных в воде (ряда одноклеточных, а также колониальных одноклеточных и некоторых колоний многоклеточных), у которых со всех сторон среда обитания одинакова.
Радиально-осевая симметрия наблюдается у нескольких групп беспозвоночных (кишечнополостных, иглокожих и др.), которые характеризуются тем, что ведут (или их предковые формы вели) прикрепленный образ жизни. Значит, сидячий образ жизни способствует развитию лучевой симметрии (Догель, 1981). Биологическое объяснение такого строения заключается в следующем. Сидячие животные одним полюсом (аборальным) прикрепляются к субстрату, другой же полюс (оральный), на котором находится ротовое отверстие, свободен. Этот полюс со всех сторон поставлен в идентичные условия по отношению к факторам окружающей среды. Поэтому различные органы одинаково развиваются на радиально расположенных частях тела, а главная ось соединяет оба полюса.
Билатеральная симметрия тела животного характерна тем, что через его тело можно провести только одну плоскость симметрии, делящую его на две равные (зеркально отражающие друг друга) половины — левую и правую. Двусторонняя симметрия возникла у животных при переходе их планктонных предков к жизни и передвижению на дне.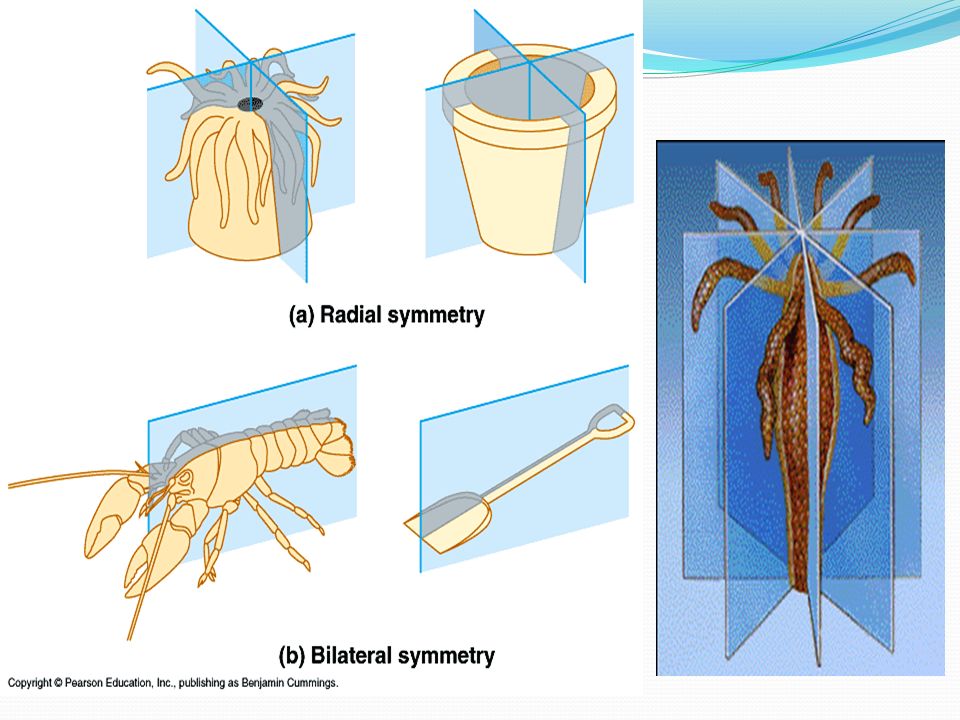 При этом кроме переднего и заднего конца тела у них стали различаться спинная (дорсальная) и брюшная (вентральная) стороны. Примерами билатерально-симметричных животных могут служить черви, членистоногие, все хордовые животные, в том числе и человек.
При этом кроме переднего и заднего конца тела у них стали различаться спинная (дорсальная) и брюшная (вентральная) стороны. Примерами билатерально-симметричных животных могут служить черви, членистоногие, все хордовые животные, в том числе и человек.
Биологическое объяснение билатеральности заключается в следующем.
При переходе к ползающему (на дне) образу жизни две стороны животного — брюшная и спинная — попадают в разные условия по отношению к факторам среды. Один конец тела становится передним и к нему сдвигается ротовое отверстие, а также органы чувств. Это и понятно, поскольку при движении этот конец первым встречает источники раздражения. Главная ось тела проходит от переднего полюса, на котором оказывается рот, до заднего, где расположено анальное отверстие. Боковые части находятся в равном положении. Единственную плоскость симметрии можно провести, только «разрезав» животное на левую и правую половины вдоль главной оси тела.
Что такое лучевая симметрия?
- Многоклеточные животные образуют самую многочисленную группу живых организмов планеты, насчитывающую более 1,5 млн. видов. Ведя свое происхождение от простейших, они претерпели в процессе эволюции существенные преобразования, связанные с усложнением организации.
Одной из важнейших черт организации многоклеточных является морфологическое и функциональное различие клеток их тела. В ходе эволюции сходные клетки в теле многоклеточных животных специализировались на выполнении определенных функций, что привело к формированию тканей.
Разные ткани объединились ворганы, а органы асистемы органов. Для осуществления взаимосвязи между ними и координации их работы образовалисьрегуляторные системы нервная и эндокринная. Благодаря нервной и гуморальной регуляции деятельности всех систем, многоклеточный организм функционирует как целостная биологическая система.
Процветание группы многоклеточных животных связано с усложнением анатомического строения и физиологических функций. Так, увеличение размеров тела привело к развитию пищеварительного канала, что позволило им питаться крупным пищевым материалом, поставляющим большое количество энергии для осуществления всех процессов жизнедеятельности. Развившиеся мышечная и скелетная системы обеспечили передвижение организмов, поддержание определенной формы тела, защиту и опору для органов. Способность к активному передвижению позволила животным осуществлять поиск пищи, находить укрытия и расселяться.
С увеличением размеров тела животных возникла необходимость в появлении внутритранспортных циркуляторных систем, доставляющих удаленным от поверхности тела тканям и органам средства жизнеобеспечения питательные вещества, кислород, а также удаляющих конечные продукты обмена веществ.
Такой циркуляторной транспортной системой стала жидкая ткань кровь.
Интенсификация дыхательной активности шла параллельно с прогрессивным развитием нервной системы и органов чувств. Произошло перемещение центральных отделов нервной системы в передний конец тела животного, в результате чего обособился головной отдел. Такое строение передней части тела животного позволило ему получать информацию об изменениях в окружающей среде и адекватно реагировать на них.
По наличию или отсутствию внутреннего скелета животные подразделяются на две группы беспозвоночные (все типы, кроме Хордовых) и позвоночные (тип Хордовые) .
В зависимости от происхождения ротового отверстия у взрослого организма выделяют две группы животных: первично- и вто-ричноротые. Первичноротые объединяют животных, у которых первичный рот зародыша на стадии гаструлы бластопор остается ртом взрослого организма. К ним относятся животные всех типов, кроме Иглокожих и Хордовых. У последних первичный рот зародыша превращается в анальное отверстие, а истинный рот закладывается вторично в виде эктодермального кармана. По этой причине их называют вторичноротыми животными.
По типу симметрии тела выделяют группу лучистых, или радиально-симметричных, животных (типы Губки, Кишечнополостные и Иглокожие) и группу двусторонне-симметричных (все остальные типы животных) . Лучевая симметрия формируется под влиянием сидячего образа жизни животных, при котором весь организм поставлен по отношению к факторам среды в совершенно одинаковые условия. Эти условия и формируют расположение одинаковых органов вокруг главной оси, проходящей через рот до противоположного ему прикрепленного полюса.
Двусторонне-симметричные животные подвижны, обладают одной плоскостью симметрии, по обе стороны которой располагаются различные парные органы. У них различают левую и правую, спинную и брюшную стороны, передний и задний концы тела.
Многоклеточные животные чрезвычайно разнообразны по строению, особенностям жизнедеятельности, различны по размерам, массе тела и т. д. На основе наиболее существенных общих черт строения они подразделяются на 14 типов, часть из которых рассматривается в данном пособии. - Лучевая (радиальная) симметрия форма симметрии, при которой тело (или фигура) совпадает само с собой при вращении объекта вокруг определнной точки или прямой.
Как правило, у многоклеточных животных два конца (полюса) единственной оси симметрии неравноценны (например, у медуз на одном полюсе (оральном) находится рот, а на противоположном (аборальном) — верхушка колокола. Такая симметрия (вариант радиальной симметрии) в сравнительной анатомии называется одноосно-гетеропольной. В двухмерной проекции радиальная симметрия может сохраняться, если ось симметрии направлена перпендикулярно к проекционной плоскости. Иными словами, сохранение радиальной симметрии зависит от угла наблюдения.
Лучевая симметрия характерна, в основном, для кишечнополостных животных. Кишечнополостным, как сидячим, так и пелагическим (медузы) , свойственна радиально-осевая симметрия, при которой сходственные части расположены вокруг оси вращения, причем эта симметрия может быть самого различного порядка в зависимости от того, на какой угол следует повернуть тело животного, чтобы новое положение совпало с исходным. Таким образом, может получаться 4-, 6-, 8лучевая симметрия и более, до симметрии порядка бесконечности. У радиолярий встречается радиально-осевая симметрия с одинаковыми полюсами, или, как говорят, гомополярная. У кишечнополостных гетерополярная осевая симметрия: один полюс симметрии несет рот и щупальца (оральный) , другой (а б о р а л ь н ы й) служит для прикрепления (стадия полипа) , или у плавающих форм несет орган чувств (ктенофоры) , или ничем не вооружен (медузы) .
У некоторых медуз на этой аборальной стороне образуется стебелек для прикрепления к подводным предметам (Lucernariida). Нарушение радиал ьно-осевой симметрии возникает при уменьшении числа шупалец или изменении формы ротовой щели, пищевода и разветвлений пищеварительной системы. Количество щупалец может уменьшаться до одного (Мопоbrachium), и тогда их радиальное расположение сменяется двубоковым. Глотка может сплющиваться, и тогда тоже получается двубоковая симметрия, этому способствует и образование в глотке сифоноглифов (желобок вдоль глотки) .
Наибольшее усложнение радиально-осевой симметрии наблюдается у ктенофор, где, помимо 8-лучевой симметрии, в расположении отдельных частей тела и органов наблюдается 4-лучевая и двубоковая симметрия. Это весьма существенный момент, так как большинство зоологов именно от ктенофорообразных предков выводит оба ствола высших животных, как первично-, так и вторичноротых.
Гетерополярная радиальноосевая симметрия вполне соответствует образу жизни кишечнополостных неподвижному существованию в прикрепленном положении или медленному плаванию при помощи реактивного движения.
С другой стороны, от сложного типа радиально-осевой симметрии ктенофор можно перейти к двусторонней симметрии, или, как говорят, симметрии зеркального изображения, единственного плана симметрии трехслойных животных, симметрии быстрого движения, с выработкой переднего по движению конца тела, с центральным мозговым скоплением и основными органами чувств, спинной и брюшной, правой и левой сторонами тела.
..подробнее — ссылка заблокирована по решению администрации проекта. berl. ru/article/ nauka/cimmetria_u_givotnyh.htm здесь (уберите про)
 видов. Ведя свое происхождение от простейших, они претерпели в процессе эволюции существенные преобразования, связанные с усложнением организации.
видов. Ведя свое происхождение от простейших, они претерпели в процессе эволюции существенные преобразования, связанные с усложнением организации.  Развившиеся мышечная и скелетная системы обеспечили передвижение организмов, поддержание определенной формы тела, защиту и опору для органов. Способность к активному передвижению позволила животным осуществлять поиск пищи, находить укрытия и расселяться.
Развившиеся мышечная и скелетная системы обеспечили передвижение организмов, поддержание определенной формы тела, защиту и опору для органов. Способность к активному передвижению позволила животным осуществлять поиск пищи, находить укрытия и расселяться. 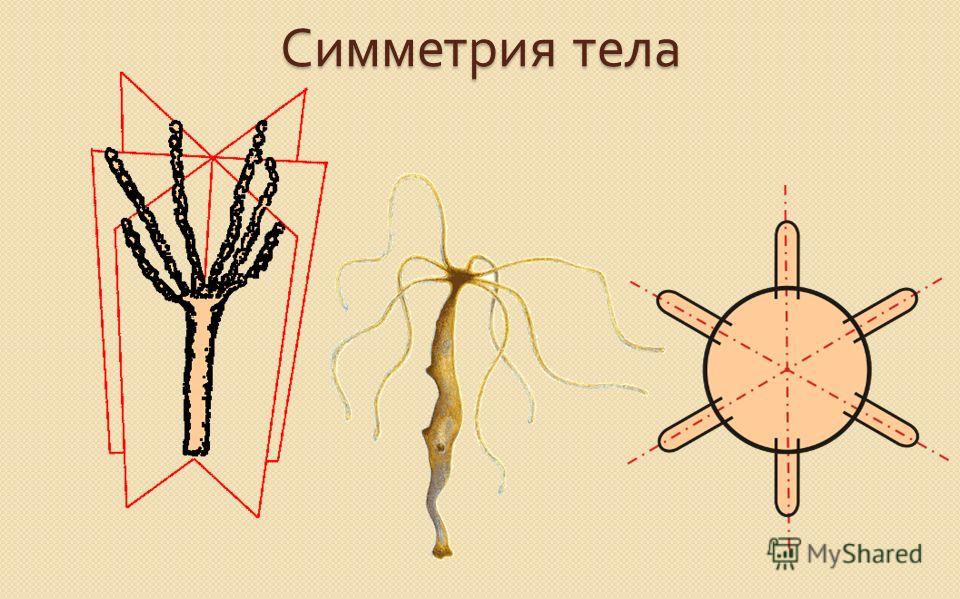
 Эти условия и формируют расположение одинаковых органов вокруг главной оси, проходящей через рот до противоположного ему прикрепленного полюса.
Эти условия и формируют расположение одинаковых органов вокруг главной оси, проходящей через рот до противоположного ему прикрепленного полюса.  Такая симметрия (вариант радиальной симметрии) в сравнительной анатомии называется одноосно-гетеропольной. В двухмерной проекции радиальная симметрия может сохраняться, если ось симметрии направлена перпендикулярно к проекционной плоскости. Иными словами, сохранение радиальной симметрии зависит от угла наблюдения.
Такая симметрия (вариант радиальной симметрии) в сравнительной анатомии называется одноосно-гетеропольной. В двухмерной проекции радиальная симметрия может сохраняться, если ось симметрии направлена перпендикулярно к проекционной плоскости. Иными словами, сохранение радиальной симметрии зависит от угла наблюдения. 

Симметрия и щупальцевой аппарат у Cnidaria
Алешин В.В. и Петров Н.Б. Молекулярные доказательства регрессии в эволюции Metazoa // Ж. общ. биол. , 2002, том. 63, стр. 195–208.
КАС пабмед Google Scholar
Беклемишев В.Н., Основы сравнительной анатомии беспозвоночных, Том 1.
Проморфология . Москва: Наука, 1964.Google Scholar
Владыченская Н.С., Кедрова О.С., Милютина И.А. и др. Положение филы Placozoa в системе Metazoa на основе сравнения последовательностей генов 18S рибосомной РНК // Докл. акад. Наук , 1995, т. 1, с. 344, нет. 1, стр. 1–3.
Google Scholar
Ивановский А.Б., Этюды оранжекаменноугольных ругозов , Москва: Наука, 1967.
Google Scholar
Ивановский А.Б., Ругозы (Ругоса), Москва: Наука, 1975. , стр. 130–140.
Google Scholar
Иванцов А.Ю., Вендия и другие докембрийские «членистоногие», Палеонтол. ж. , 2001, вып. 4, стр. 3–10.
Google Scholar
Иванцов А.Ю. и Федонкин М.А. Следы самоподдерживающегося движения — окончательное свидетельство животной природы эдиакарских организмов // Матер. II межд. дурачок. «Эволюция жизни на Земле», Томск, 12–15 ноября 2001 г. . (Труды II Междунар. симп. «Эволюция жизни на Земле», Томск, 12–15 ноября 2001 г.). Томск, 2001. С. 133–137.
Google Scholar
Малахов В.В., Новые представления о происхождении билатеральных животных, рус. Дж. Мар. Биол. , 2004, том. 30, нет. 1 (дополнение), стр. S22–S33.
Артикул Google Scholar
Молодцова Т.Н. Малахов В.В., К биологии Cerianthus lloydii Gosse (Anthozoa, Ceriantharia) из Кратерной бухты (Курильские острова), Тр.
Зоол. Ин-т, Росс. акад. Наук , 1995, т. 1, с. 261, стр. 90–94.Google Scholar
Молодцова Т.Н. и Малахов В.В. Cerianthus lloydii (Anthozoa, Ceriantharia) из вулканической экосистемы бухты Кратерная. 2. Развитие личинок, Зоол. ж. , 1995, том. 74, нет. 11, стр. 4–11.
Google Scholar
Наумов Д.В., Гидроиды и гидромедузы морских, солоноватоводных и пресноводных бассейнов СССР . Гидроиды и гидромедузы морского, солоноватого и пресноводного бассейнов Советского Союза. М.: Акад. Наук СССР, 1960.
Google Scholar
Рич., П.В., Рич, Т.Х., и Фентон, Массачусетс, Книга окаменелостей: летопись доисторической жизни , Нью-Йорк: Дувр, 1997.
Google Scholar
Федонкин М.А. Колодец в прошлое глубиной 600 млн лет // Наука в России. 6, стр. 6–15.
Google Scholar
Оллман, Г.Дж., Монография гимнобластических или трубчатых гидроидов , Лондон: Ray Society, 1872.
Google Scholar
Андрес, А., Le Actinie, Fauna und Flora des Golfes von Neapel , 1884, vol. 13, стр. 1–459.
Google Scholar
Baguna, J., Martinez, P., Paps, J. и Riutort, M., Назад во времени: новое систематическое предложение для Bilateria, Philos. Транс. R. Soc., B , 2008, vol. 363, стр. 1481–1491.
Артикул КАС Google Scholar
Brinkmann-Voss, A., Anthomedusae/Athecatae (Hydrozoa, Cnidaria) Средиземноморья, Fauna Flora Golfo Napoli , 1970, no. 39, стр. 1–96.
Google Scholar
Карлгрен О., Zur Kenntnis der Septenmuskulatur bei Ceriantheen und der Schlundrinnen bei Anthozoen, Öfv. К. Свен. Ветенск.-Акад. Форхандль, Стокгольм , 1893 г., том. 50, стр. 239–247.
Google Scholar
Carlgren, O., Zoantarien, Ergebnisse der Hamburger Magalhaensische Sammelreise , 1899, vol. 1, стр. 1–47.
Google Scholar
Carlgren, O., Ceriantharia, Danish Ingolf-Exped.
, 1912, том. 1, пт. 3, стр. 1–78.Google Scholar
Карлгрен, О., Ueber Ceriantharien des Mittelmeeres, Mitt. Зоол. Стат. Неапель , 1912, том. 20, стр. 356–394.
Google Scholar
Carlgren, O., Ceriantharia and Zoantharia, Wiss. Эргеб. Немецкий. Тифзее-Exped. Дампфер «Вальдивия» 1898–1899 , 1923, том. 19, нет. 7, стр. 242–337.
Google Scholar
Карлгрен, О., Die Larven der Ceriantharien, Zoantarien und Actiniarien, Висс. Эргеб. Немецкий. Тифзее-Exped. Дампфер «Вальдивия» 1898–1899 , 1924, том. 19, стр. 339–476.
Google Scholar
Carlgren, O., South African Actiniaria and Zoantharia, K. Sven. Ветенск.-Акад. Ручка , 1938, том. 17, нет.
3, стр. 1–148.Google Scholar
Шевалье, Ж.П. и Бове, Л., Ordre des Scléractinaires, in Traitéde zoologie, Cnidaires, Anthozoaires , Париж: Массон, 1987, том. 3, фас. 3, стр. 403–764.
Google Scholar
Коллинз, А.Г., Оценка нескольких альтернативных гипотез происхождения Bilateria: анализ молекулярных доказательств 18S рРНК, Proc. Натл. акад. науч. США , 1998, том. 95, стр. 15458–15463.
Артикул КАС пабмед ПабМед Центральный Google Scholar
Коллинз, А.Г. и Валентайн, Дж.В., Определение типов: эволюционные пути к многоклеточным телам, Evol. Дев. , 2001, том. 3, стр. 432–442.
Артикул КАС пабмед Google Scholar
Collins, A.
G., Winkelmann, S., Hadrys, H., и Schierwater, B., Филогенез Capitata и Corynidae (Cnidaria, Hydrozoa) в свете данных митохондриальной 16S рДНК, Zool. Скр. , 2005, том. 34, стр. 91–99.Артикул Google Scholar
Дейли М., Брюглер М.Р., Картрайт П. и др. Тип Cnidaria: обзор филогенетических моделей и разнообразия через 300 лет после Линнея, Zootaxa , 2007, vol. 1668, стр. 127–182.
Google Scholar
Делаж Ю. и Эруар Э., Traité de zoologie бетон, Les Coelentérés , Paris: Reinwald, 1901, том. 2, фас. 2, стр. 1–848.
Google Scholar
Duerden, J.E., Edwardsia-стадия актиний Lebrunia и формирование желудочно-целомической полости, J. Linn. соц. Лонд. , 1899, т. 1, с. 27, стр. 269–316.
Артикул Google Scholar
Дьюрден, Дж. Э., Отчет об актиниях Порто-Рико, Bull. US Fish Comm. , 1902, том. 20, стр. 321–374.
Google Scholar
Duerden, J.E., West Indian Madreporarian polyps, Mem. Натл. акад. науч. США , 1902, том. 8, пт. 7, стр. 399–648.
Google Scholar
Дюрден, Дж. Э., Морфология Madreporaria. V. Перегородочная последовательность, Biol. Бык. , 1904, т. 1, с. 7, стр. 79–104.
Артикул Google Scholar
Дьюрден, Дж. Э., Коралл Siderastrea radians и его постличиночное развитие, Carnegie Inst.
Вашингтон Паблик. , 1904, вып. 20, стр. 1–130.Google Scholar
Дьюэрден, Дж. Э., Недавние результаты по морфологии и развитию коралловых полипов, стр. 9.0005 Смитсон. Разное Сб. , 1905, том. 47, стр. 93–111.
Google Scholar
Дюрден, Дж. Э., Морфология Madreporaria. 8. Первичные перегородки ругозы, Ann. Маг. Нац. История , 1906, вып. 105, стр. 226–242.
Артикул Google Scholar
Dunn, C.W., Hejnol, A., Matus, D.Q., et al., Широкая филогеномная выборка улучшает разрешение дерева жизни животных, Природа , 2008, том. 452, стр. 745–750.
Артикул КАС пабмед Google Scholar
Дзик Ю., Иванцов А.Ю. Асимметрично-сегментарный организм из венда России и статус Dipleurozoa, Hist.
биол. , 1999, том. 13, стр. 255–268.Артикул Google Scholar
Дзик Ю., Иванцов А.Ю. Внутренняя анатомия нового докембрийского dickinsoniid dipleurozoan из северной России, Новый Ярб. геол. Палеонтол., Монач. , 2002, вып. 7, стр. 385–396.
Google Scholar
Faurot, L., Études sur l’anatomie, l’histologie et le développement des actinides, Arch. Зоол. Эксп. Ген., сер. 4 , 1895, т. 1, с. 3, стр. 43–262.
Google Scholar
Федонкин М.А., Ваггонер Б.М. Позднедокембрийская ископаемая кимберелла — моллюскоподобный двусторонний организм, Природа , 1997, том. 338, 28 августа, стр. 868–871.
Артикул КАС Google Scholar
Федоровски Дж.
, Принципы раннего онтогенеза морщинистых кораллов: критический обзор, Hydrobiologia , 1991, vol. 216–217, вып. 1, стр. 413–418.Артикул Google Scholar
Finnerty, J.R., Книдарийцы раскрывают промежуточные этапы эволюции Hox-кластеров и осевой сложности, 900:05 утра. Зоол. , 2001, том. 41, стр. 608–620.
КАС Google Scholar
Финнерти, Дж. Р., Происхождение осевого паттерна у метазоа: сколько лет билатеральной симметрии? Междунар. Дж. Дев. биол. , 2003, том. 47, стр. 523–529.
ПабМед Google Scholar
Финнерти, Дж. Р. и Мартиндейл, М. К., Гомеобоксы морских анемонов (Cnidaria, Anthozoa): исследование Nematostella vectensis и Metridium senile на основе ПЦР, биол. Бык. , 1997, том. 193, стр. 62–76.
Артикул КАС пабмед Google Scholar
Финнерти Дж. Р., Полсон Д., Бертон П. и др. Ранняя эволюция гомеобоксного гена: ген paraHox Gsx у Cnidaria и Bilateria, Эволюция. Дев. , 2003, том. 5, стр. 331–345.
Артикул КАС пабмед Google Scholar
Геммилл, Дж. Ф., Развитие актиний Metridium dianthus и Adamsia palliate, Philos. Транс. R. Soc., B , 1920, vol. 209, стр. 351–375.
Артикул Google Scholar
Геммил, Дж. Ф., Развитие актинии Boloceratiediae, кв. Дж. Микроск. науч. , 1921, том. 65, стр. 577–587.
Google Scholar
Greef, R., Protohydra leuckarti. Eine Marine Stammform der Coelenteraten, Z. Wiss. Зоол. , 1869, том. 20, стр. 37–54.
Google Scholar
Галаныч К.М. Новый взгляд на филогению животных // Annu. Преподобный Экол. Эвол. Сист. , 2004, том. 35, стр. 229–256.
Артикул Google Scholar
Галаныч К.М. и Пассаманек, Ю., Краткий обзор филогении многоклеточных животных и будущих перспектив в Hoxresearch, Am. Зоол. , 2001, том. 41, стр. 629–639.
КАС Google Scholar
Хартог Дж. К.
и Гребельный С. Д. Соотношение дисковых и краевых щупалец как признак видов, характерных для Corallimorphus (Anthozoa, Corallimorpharia), Proc. Зоол. ин-т, рус. акад. науч. , 2004, том. 300, стр. 109–120.Google Scholar
Хайдер, А.Р. фон, Cerianthus membranaceus Haime. Ein Beitrag zur Anatomie der Actinien, Зитцунгсбер. Кайзер. акад. Висс. Wien, Math.-Naturwiss. Кл. , 1879, т. 1, с. 79, стр. 21–38.
Google Scholar
Hejnol, A., Obst, M., Stamatakis, A., et al., Оценка корня билатеральных животных с помощью масштабируемых филогеномных методов, Proc. Р. Соц., сер. Б. , 2009, вып. 276, стр. 4261–4270.
Артикул Google Scholar
Хертвиг, О., Отчет об актиниарии, вычерпанной H.M.S. Челленджер в 1873–1876 гг., Респ. Результаты Voyage H.
M.S. Челленджер , 1882 г., т. 1, с. 6, стр. 1–136.Google Scholar
Хертвиг, О. и Хертвиг, Р., Die Actinien, anatomisch und histologisch mit besonderer Berücksichtigung des Nervenmuskelsystems untersucht, Jena. З. Натурвисс. , 1879, вып. 1, стр. 1–224.
Google Scholar
Hill, D., Британская терминология морщинистых кораллов, Геол. Маг. , 1935, том. 72, стр. 481–519.
Артикул Google Scholar
Hill, D., Rugosa, in Treatise on Invertebrate Paleontology, Part F: Coelenterata, Lawrence, KS: Geol. соц. Являюсь. , 1956, стр. 233–324.
Google Scholar
Хилл, Д., Ругоза и Табулата, в «Трактате о палеонтологии беспозвоночных», часть F: Coelenterata, Приложение 1, Лоуренс, КС: Геол.
соц. Являюсь. , 1981, стр. 1–762.Google Scholar
Хайман, Л.Х., Беспозвоночные, Том. 1: Protozoa Trough Ctenophora , New York: McGraw-Hill, 1940.
Google Scholar
Ким, Дж., Ким, В., и Каннингем, К.В., Новый взгляд на взаимоотношения низших многоклеточных животных из последовательностей 18S рДНК, Mol. биол. Эвол. , 1999, том. 16, стр. 423–427.
Артикул КАС пабмед Google Scholar
Крамп П.Л. Гидромедузы Атлантического океана и прилегающих вод. 46, стр. 1–283.
Google Scholar
Крамп, П.Л., Краткий обзор медуз мира, J. Mar. Biol. доц. Великобритания , 1961, том. 40, стр. 1–469.
Google Scholar
Крамп, П.Л., Гидромедузы Тихого и Индийского океанов, Sect. II и III, Dana Rep. , 1968, вып. 72, стр. 1–200.
Google Scholar
Кунт, А., Окаменелость Beiträge zur Kenntnis. Кораллен. 2. Das Wachstumsgesetz der Zoantharia Rugosa und über Calceola sandalina, З. Дойч. геол. Гэс. , 1869, т. 1, с. 21, стр. 1–284.
Google Scholar
Куссеров А., Панг К., Штурм С. и др., Неожиданная сложность семейства генов Wnt у морских актиний, Nature , 2005, vol. 433, стр. 156–160.
Артикул КАС пабмед Google Scholar
Ли, Х.Е.
, Юн, В.Д., и Лим, Д., Описание аппарата и механизма питания у Nemopilema nomurai Kishinouye (Scyphozoa: Rhizostomeae), Науки об океане. J. , 2008, том. 43, стр. 61–65.Артикул Google Scholar
Leloup, E., Cérianthaires de l’Ocean Atlantique, Bull. Мус. Р. Хист. Нац. бельг. , 1932, том. 8, стр. 1–19.
Google Scholar
Leloup, E., Larves de Cérianthaires appartenant a trois especes nouvelles, Bull. Мус. Р. Хист. Нац. бельг. , 1942, том. 18, стр. 1–16.
Google Scholar
Leloup, E., Coelenteres, Faune de Belgique, Bruxelles: Inst. Р. Наук. Нац. бельг. , 1952, стр. 1–283.
Google Scholar
Leloup, E., Larves de Cérianthaires de Monaco et de Villefranche-sur-Mer, Bull.
Инст. Океаногр. , 1960, том. 57, стр. 1–19.Google Scholar
Leloup, E., Larves de Cerianthaires, Discovery Rep. , 1964, vol. 33, стр. 251–307.
Google Scholar
Марлоу, Х.К., Сривастава, М., Матус, Д.К., и др., Анатомия и развитие нервной системы Nematostella vectensis, корифея книдарий, Dev. Нейробиол. , 2009, том. 69, стр. 235–254.
Артикул КАС пабмед Google Scholar
Martindale, M.Q., Pang, K., and Finnerty, J.R., Изучение происхождения триплообластии: «мезодермальная» экспрессия гена у диплобластного животного, актинии Nematostella vectensis (тип Cnidaria, класс Anthozoa), Development , 2004, вып. 131, стр. 2463–2475.
КАС Google Scholar
Matus, D.Q., Magie, C.R., Pang, K., et al., Семейство генов Hedgehog книдарий, Nematostella vectensis, и последствия для понимания эволюции путей многоклеточных Hedgehog, Dev. биол. , 2008, том. 313, стр. 501–518.
Артикул КАС пабмед Google Scholar
Matus, D.Q., Thomsen, G.H., и Martindale, M.Q., Дорсо/вентральные гены экспрессируются асимметрично и участвуют в демаркации зародышевого слоя во время кишечной гаструляции, Curr. биол. , 2006, том. 16, стр. 499–505.
Артикул КАС пабмед Google Scholar
Матус Д.К.
, Панг К., Марлоу Х. и др. Молекулярные доказательства глубоких эволюционных корней билатеральности в развитии животных, стр. Проц. Натл. акад. науч. США , 2006, том. 103, стр. 11195–11200.Артикул КАС пабмед ПабМед Центральный Google Scholar
Matus, D.Q., Pang, K., Daly, M., и Martindale, M.Q., Экспрессия членов семейства генов Pax у антозойных книдарий, Nematostella vectensis, Evol. Дев. , 2007, том. 9, стр. 25–38.
Артикул КАС пабмед Google Scholar
Матус, Д.К., Томсен, Г.Х., и Мартиндейл, М.К., Передача сигналов FGF в гаструляции и развитии нервной системы у Nematostella vectensis, корифея книдарий, Dev. Гены Эвол. , 2007, том. 217, стр. 137–148.
Артикул КАС пабмед ПабМед Центральный Google Scholar
Макмеррич, Дж. П., Вклад в морфологию Actinozoa. II. О развитии Hexactiniae, J. Morphol. , 1891, том. 4, стр. 303–330.
Артикул Google Scholar
McMurrich, J.P., Род Arachnactis, J. Exp. Зоол. , 1910, том. 9, стр. 159–168.
Артикул Google Scholar
Миллард, Н.А.Х., Монография о гидроиде южной Африки, Ann. С. Афр. Мус. , 1975. Том. 68, стр. 1–513.
Google Scholar
Милн-Эдвардс, Х. и Хайме, Дж., Монография окаменелостей полипов палеозойских ландшафтов, Arch.
Мус. Натл. История Нац. , 1851, том. 5, стр. 1–502.Google Scholar
Nyholm, K.G., Zur Entwicklung und Entwicklungsbiologie der Ceriantharien und Aktinien, Zool. Бидр. Уппсала , 1943, том. 22, стр. 87–248.
Google Scholar
Pax, F., Die Actinien, Ergeb. Фортшр. Зоол. , 1914, том. 4, стр. 339–642.
Google Scholar
Pax, F., Unterklasse: Hexacorallia, in Handbuch der Zoologie , Berlin: Walter de Gruyter, 1925, vol. 1, стр. 770–901.
Google Scholar
Pax, F., Anthozoa, Bronn’s Klassen und Ordnungen des Theyreichs , 1940, vol. 2, пт. 2, фас. 3, стр. 177–336.
Google Scholar
Philippe, H., Lartillot, N., and Brinkmann, H., Мультигенный анализ билатеральных животных подтверждает монофилию Ecdysozoa, Lophotrochozoa и Protostomia, Mol. биол. Эвол. , 2005, том. 22, стр. 1246–1253.
Артикул КАС пабмед Google Scholar
Рупперт Э.Э., Фокс Р.С. и Барнс Р.Д., Зоология беспозвоночных, функциональный эволюционный подход , Белмонт, Калифорния: Brooks/Cole-Thomson Learning, 2004.
Google Scholar
Рассел, Ф.С., Медузы Британских островов, Vol.
1: Anthomedusae, Leptomedusae, Limnomedusae, Trachymedusae и Narcomedusae , Кембридж: Cambridge Univ. Пресса, 1953.Google Scholar
Russell, FS, Pelagic Scyphozoa с дополнением к первому тому по Hydromedusae , Vol. 2: Медузы Британских островов, Кембридж: Кембриджский унив. Пресс, 1970.
Google Scholar
Саина М., Геничович Э., Ренфер Э. и Технау У., BMPs и Chordin регулируют формирование направляющей оси у морского анемона, Proc. Натл. акад. науч. США , 2009, том. 106, стр. 18592–18597.
Артикул КАС пабмед ПабМед Центральный Google Scholar
Scholz, CB и Technau, U., Роль предков Brachyury: экспрессия NemBra1 в базальной книдарии Nematostella vectensis (Anthozoa), Dev. Гены Эвол. , 2003, том. 212, стр.
563–570.КАС пабмед Google Scholar
Thiel, H., Cnidaria, в Введение в изучение мейофауны , Вашингтон, округ Колумбия: Smithsonian Inst. Press, 1988, стр. 266–272.
Google Scholar
Van Beneden, É., Recherches sur le développement des Arachnactis. Вклад в морфологию Cérianthides, Arch. биол. (Париж) , 1891, том. 11, стр. 114–146.
Google Scholar
Van Beneden, É., Les Anthozoaires de la Plankton-Expedition; Die Anthozoen der Plankton-Expedition, Ergebnisse der in dem Atlantischen Ocean von Mitte Juli bis Anfang, ноябрь 1889 г.ausgeführten Plankton-Expedition der Humboldt-Stiftung , Киль: Липсиус и Тишер, 1897.
Google Scholar
Van Beneden, É.
, Travaux posthumes d’Edouard Van Beneden sur les Cérianthaires, Arch. Biol., Hors-Ser. , 1923, стр. 1–242.Google Scholar
Werner, B., Stamm Cnidaria, in Lehrbuch der spezielle Zologie , Vol. 1: Wirbellose Tiere, 2. Cnidaria, Ctenophora, Mesozoa, Plathelminthes, Nemertini, Entoprocta, Nemathelminthes, Priapuluda, Jena: Gustav Fischer Verlag, 1984, стр. 11–305.
Google Scholar
Уиллмер, П., Взаимоотношения беспозвоночных: закономерности в эволюции животных , Кембридж: Кембриджский унив. Пресс, 1990.
Книга Google Scholar
Wilson, H.V., О развитии Manicina areolata, J. Morphol. , 1888 г., том. 2, нет. 2, стр. 191–252.
Артикул Google Scholar
 Проморфология . Москва: Наука, 1964.
Проморфология . Москва: Наука, 1964. «>
«>Иванцов А.Ю. и Малаховская Я.Е. Гигантские следы вендских животных. Докл. наук о Земле. , 2002, том. 385А, нет. 6, стр. 618–622.
КАС Google Scholar
 Зоол. Ин-т, Росс. акад. Наук , 1995, т. 1, с. 261, стр. 90–94.
Зоол. Ин-т, Росс. акад. Наук , 1995, т. 1, с. 261, стр. 90–94. «>
«>Федонкин М.А. Второе рождение Кимбереллы, Природа , 1998, вып. 1, стр. 3–10.
Google Scholar
 «>
«>Bouillon, J., Gravili, C., Pagès, F., et al., Введение в Hydrozoa, Mem. Мус. Натл. История Нац. , 2006, том. 194, стр. 1–591.
Google Scholar
 , 1912, том. 1, пт. 3, стр. 1–78.
, 1912, том. 1, пт. 3, стр. 1–78. 3, стр. 1–148.
3, стр. 1–148. G., Winkelmann, S., Hadrys, H., и Schierwater, B., Филогенез Capitata и Corynidae (Cnidaria, Hydrozoa) в свете данных митохондриальной 16S рДНК, Zool. Скр. , 2005, том. 34, стр. 91–99.
G., Winkelmann, S., Hadrys, H., и Schierwater, B., Филогенез Capitata и Corynidae (Cnidaria, Hydrozoa) в свете данных митохондриальной 16S рДНК, Zool. Скр. , 2005, том. 34, стр. 91–99.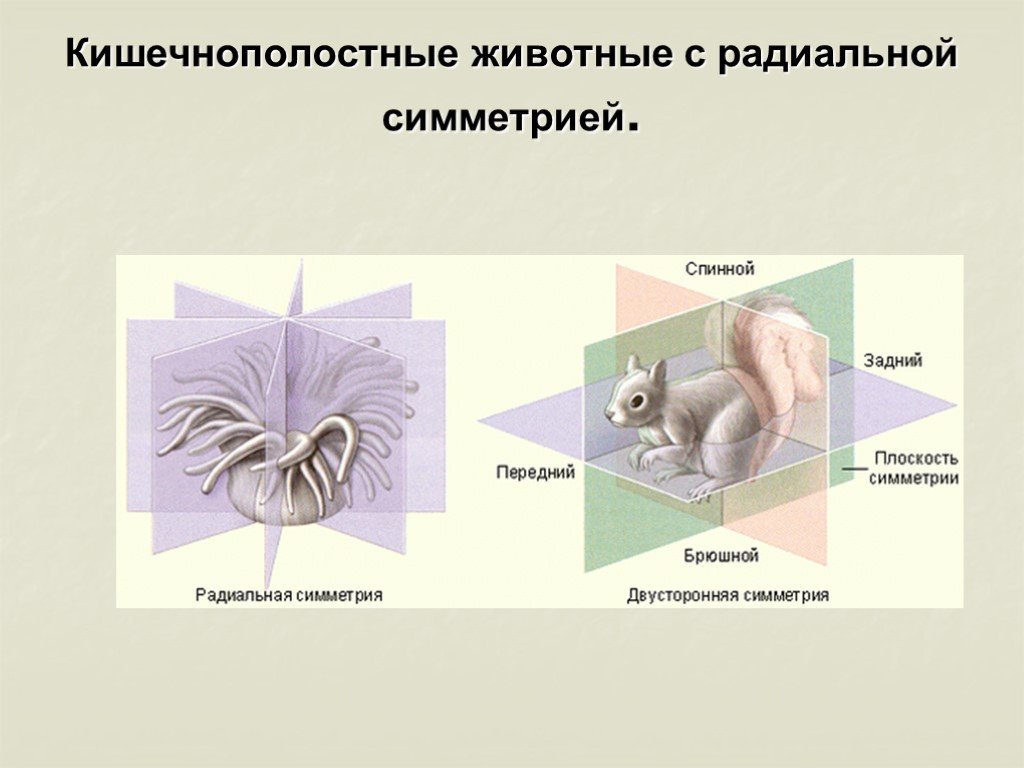 «>
«>Duerden, J.E. Порядок появления брыжейки и перегородок в Madreporaria, John Hopkins Univ. Цирк. , 1900, том. 19, стр. 47–53.
Google Scholar
 Вашингтон Паблик. , 1904, вып. 20, стр. 1–130.
Вашингтон Паблик. , 1904, вып. 20, стр. 1–130. биол. , 1999, том. 13, стр. 255–268.
биол. , 1999, том. 13, стр. 255–268. , Принципы раннего онтогенеза морщинистых кораллов: критический обзор, Hydrobiologia , 1991, vol. 216–217, вып. 1, стр. 413–418.
, Принципы раннего онтогенеза морщинистых кораллов: критический обзор, Hydrobiologia , 1991, vol. 216–217, вып. 1, стр. 413–418. «>
«>Финнерти Дж. Р., Панг К., Бертон П. и др., Происхождение билатеральной симметрии: экспрессия Hox и dpp у морских актиний, Science , 2004, vol. 304, стр. 1335–1337.
Артикул КАС пабмед Google Scholar
 «>
«>Гирибет, Г., Данн, К.В., Эджкомб, Г.Д., и Роуз, Г.В., Современный взгляд на древо жизни животных, Zootaxa , 2007, №. 1668, стр. 61–79.
Google Scholar
 и Гребельный С. Д. Соотношение дисковых и краевых щупалец как признак видов, характерных для Corallimorphus (Anthozoa, Corallimorpharia), Proc. Зоол. ин-т, рус. акад. науч. , 2004, том. 300, стр. 109–120.
и Гребельный С. Д. Соотношение дисковых и краевых щупалец как признак видов, характерных для Corallimorphus (Anthozoa, Corallimorpharia), Proc. Зоол. ин-т, рус. акад. науч. , 2004, том. 300, стр. 109–120. M.S. Челленджер , 1882 г., т. 1, с. 6, стр. 1–136.
M.S. Челленджер , 1882 г., т. 1, с. 6, стр. 1–136. соц. Являюсь. , 1981, стр. 1–762.
соц. Являюсь. , 1981, стр. 1–762. «>
«>Крамп, П.Л., Гидромедузы Тихого и Индийского океанов, Dana-Rep. , 1965, вып. 63, стр. 1–162.
Google Scholar
 , Юн, В.Д., и Лим, Д., Описание аппарата и механизма питания у Nemopilema nomurai Kishinouye (Scyphozoa: Rhizostomeae), Науки об океане. J. , 2008, том. 43, стр. 61–65.
, Юн, В.Д., и Лим, Д., Описание аппарата и механизма питания у Nemopilema nomurai Kishinouye (Scyphozoa: Rhizostomeae), Науки об океане. J. , 2008, том. 43, стр. 61–65. Инст. Океаногр. , 1960, том. 57, стр. 1–19.
Инст. Океаногр. , 1960, том. 57, стр. 1–19. «>
«>Мартинес Д.Э., Бридж Д., Масуданакагава Л.М. и Картрайт П., Книдарийские гомеобоксы и зоотип, Природа , 1998, вып. 393, стр. 748–749.
Артикул КАС пабмед Google Scholar
 , Панг К., Марлоу Х. и др. Молекулярные доказательства глубоких эволюционных корней билатеральности в развитии животных, стр. Проц. Натл. акад. науч. США , 2006, том. 103, стр. 11195–11200.
, Панг К., Марлоу Х. и др. Молекулярные доказательства глубоких эволюционных корней билатеральности в развитии животных, стр. Проц. Натл. акад. науч. США , 2006, том. 103, стр. 11195–11200. «>
«>Макмеррич, Дж. П., Вклад в морфологию Actinozoa. I. Строение Cerianthus americanus, Дж. Морфол. , 1890, т. 1, с. 4, стр. 131–150.
Артикул Google Scholar
 Мус. Натл. История Нац. , 1851, том. 5, стр. 1–502.
Мус. Натл. История Нац. , 1851, том. 5, стр. 1–502.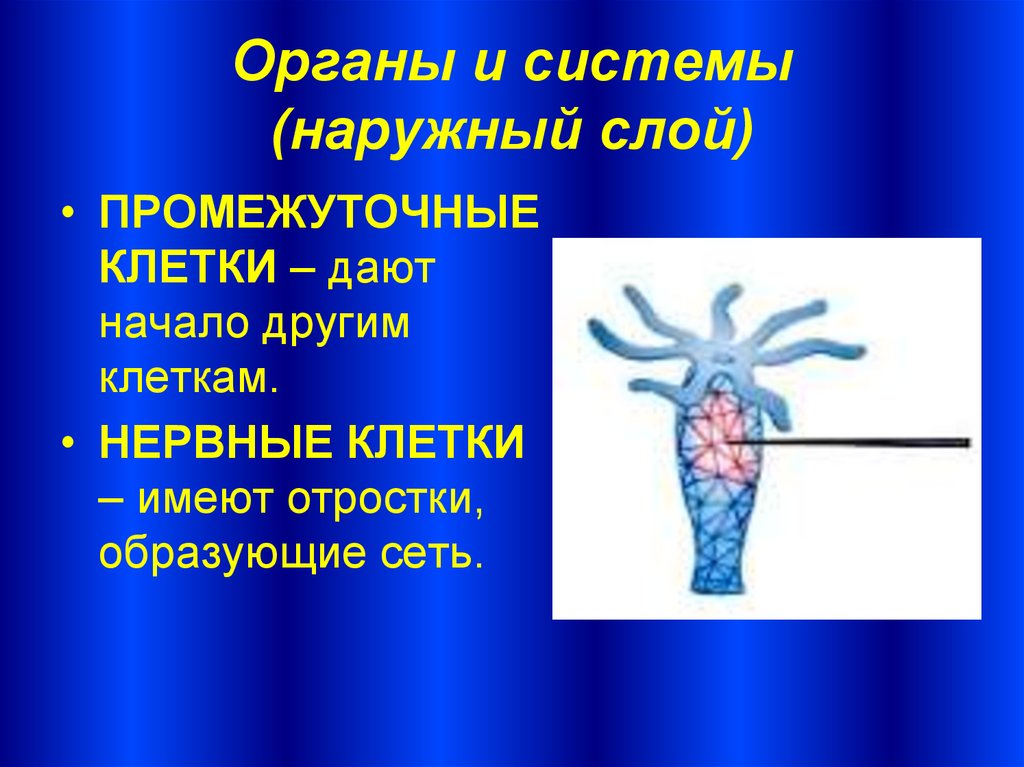 «>
«>Петерсон, К.Дж., Коттон, Дж.А., Гелинг, Дж.Г., и Пизани, Д., Эдиакарское появление билатерий: соответствие между генетическими и геологическими окаменелостями, Филос. Транс. R. Soc., B , 2008, vol. 363, стр. 1435–1443.
Артикул Google Scholar
 1: Anthomedusae, Leptomedusae, Limnomedusae, Trachymedusae и Narcomedusae , Кембридж: Cambridge Univ. Пресса, 1953.
1: Anthomedusae, Leptomedusae, Limnomedusae, Trachymedusae и Narcomedusae , Кембридж: Cambridge Univ. Пресса, 1953.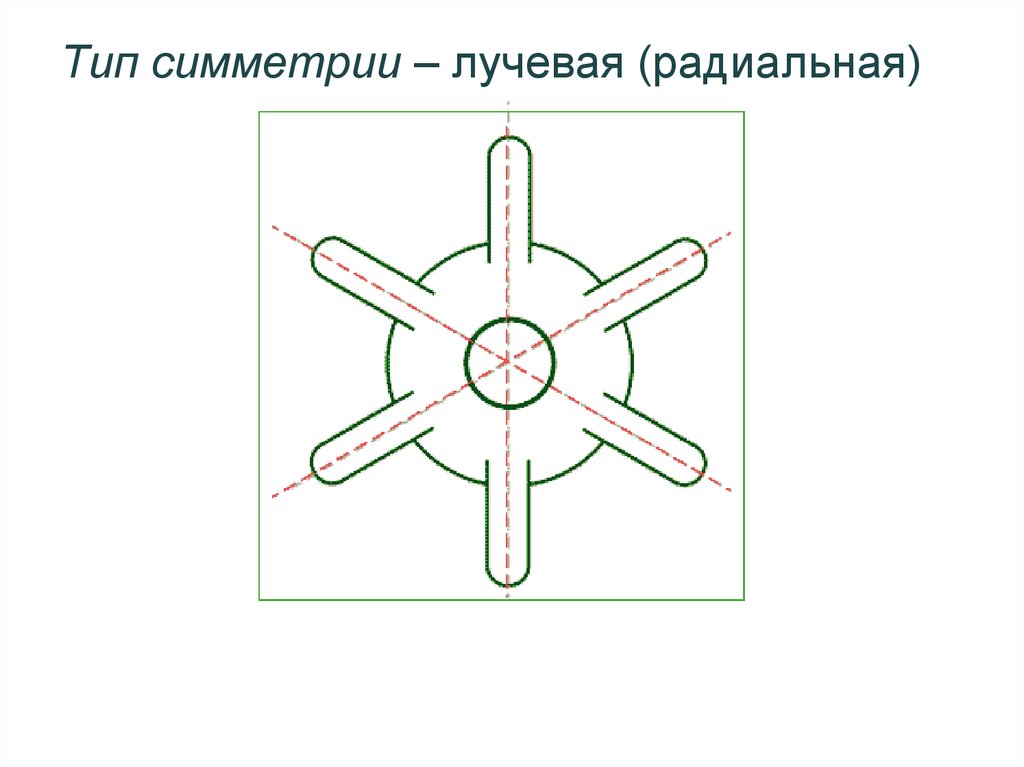 563–570.
563–570. , Travaux posthumes d’Edouard Van Beneden sur les Cérianthaires, Arch. Biol., Hors-Ser. , 1923, стр. 1–242.
, Travaux posthumes d’Edouard Van Beneden sur les Cérianthaires, Arch. Biol., Hors-Ser. , 1923, стр. 1–242. «>
«>Zapata, F., Goetz, F.E., Smith, S.A., et al., Филогеномный анализ поддерживает традиционные отношения внутри Cnidaria, PLoS One , 2015, vol. 10, нет. 10, с. e0139068. doi 10.1371/journal.pone.0139068
Статья пабмед КАС ПабМед Центральный Google Scholar
Загрузка ссылок
Признаки, используемые для классификации животных
Цели обучения
К концу этого раздела вы должны выполнить следующие задачи:
- Объясните различия в строении тела животных, которые поддерживают основную классификацию животных
- Сравните и сопоставьте эмбриональное развитие первичноротых и вторичноротых
Ученые разработали схему классификации, которая классифицирует всех представителей животного мира, хотя есть исключения из большинства «правил», регулирующих классификацию животных (рис. 1). Животные в первую очередь классифицируются в соответствии с морфологическими характеристиками и особенностями развития, такими как строение тела.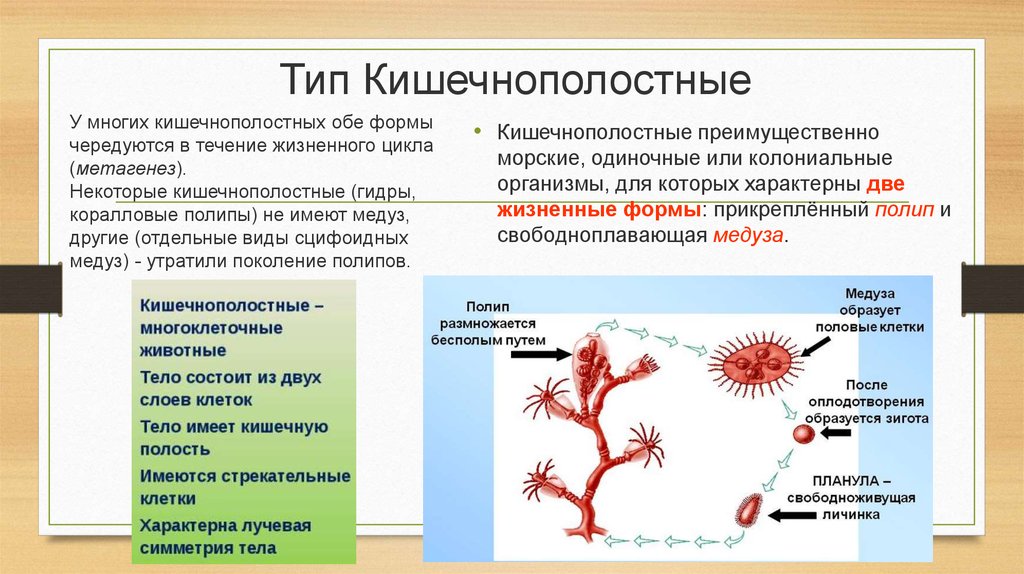 Одной из самых выдающихся особенностей строения тела настоящих животных является то, что они морфологически симметричны. Это означает, что их распределение частей тела сбалансировано по оси. Дополнительные характеристики включают количество слоев ткани, образующихся в процессе развития, наличие или отсутствие внутренней полости тела и другие особенности эмбрионального развития, такие как происхождение рта и ануса.
Одной из самых выдающихся особенностей строения тела настоящих животных является то, что они морфологически симметричны. Это означает, что их распределение частей тела сбалансировано по оси. Дополнительные характеристики включают количество слоев ткани, образующихся в процессе развития, наличие или отсутствие внутренней полости тела и другие особенности эмбрионального развития, такие как происхождение рта и ануса.
Art Connection
Рисунок 1. Филогенетическое древо животных основано на морфологических, ископаемых и генетических данных.
Какое из следующих утверждений неверно?
- Эвметазои имеют специализированные ткани, а паразои — нет.
- Lophotrochozoa и Ecdysozoa относятся к Bilataria.
- Acoela и Cnidaria обладают радиальной симметрией.
- Членистоногие более тесно связаны с нематодами, чем с кольчатыми червями.
Утверждение 3 неверно.
Характеристика животных на основе симметрии тела
На самом базовом уровне классификации настоящих животных можно разделить на три группы в зависимости от типа симметрии их тела: радиально-симметричные, двусторонне-симметричные и асимметричные.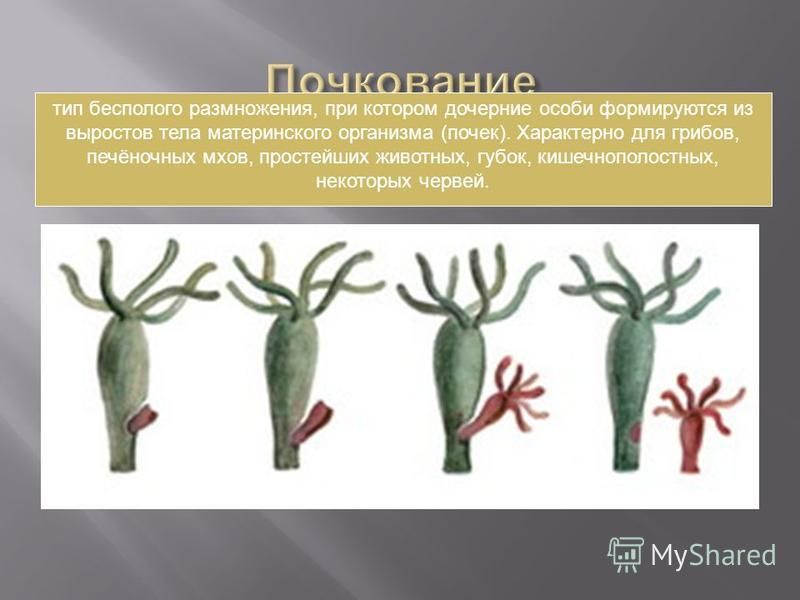 Асимметрия — уникальная черта Parazoa (рис. 2а). Лишь немногие группы животных демонстрируют радиальную симметрию. Все типы симметрии хорошо подходят для удовлетворения уникальных требований образа жизни конкретного животного.
Асимметрия — уникальная черта Parazoa (рис. 2а). Лишь немногие группы животных демонстрируют радиальную симметрию. Все типы симметрии хорошо подходят для удовлетворения уникальных требований образа жизни конкретного животного.
Радиальная симметрия — расположение частей тела вокруг центральной оси, как в стакане или пироге. Это приводит к тому, что животные имеют верхнюю и нижнюю поверхности, но не имеют левой и правой сторон, передней или задней части. Две половины радиально-симметричного животного могут быть описаны как сторона со ртом или «оральная сторона» и сторона без рта («аборальная сторона»). Эта форма симметрии отмечает планы тела животных из типов Ctenophora и Cnidaria, включая медуз и взрослых актиний (рис. 2b и 2c). Радиальная симметрия позволяет этим морским существам (которые могут вести малоподвижный образ жизни или способны только к медленному движению или плаванию) одинаково ощущать окружающую среду со всех сторон.
Рисунок 2. Губка (а) асимметрична. Медуза (б) и анемон (в) радиально-симметричны, а бабочка (г) двусторонне-симметрична. (кредит a: модификация работы Эндрю Тернера; кредит b: модификация работы Роберта Фрейбургера; кредит c: модификация работы Сэмюэля Чоу; кредит d: модификация работы Кори Занкера)
Медуза (б) и анемон (в) радиально-симметричны, а бабочка (г) двусторонне-симметрична. (кредит a: модификация работы Эндрю Тернера; кредит b: модификация работы Роберта Фрейбургера; кредит c: модификация работы Сэмюэля Чоу; кредит d: модификация работы Кори Занкера)
Рисунок 3. Двусторонне-симметричное человеческое тело можно разделить на плоскости.
Двусторонняя симметрия включает в себя деление животного в сагиттальной плоскости, в результате чего образуются две зеркальные копии, правая и левая половины, например, у бабочки (рис. 2d), краба или человеческого тела. У животных с билатеральной симметрией есть «голова» и «хвост» (передняя и задняя), передняя и задняя (дорсальная и вентральная), а также правая и левая стороны (рис. 3). Все настоящие животные, кроме обладающих радиальной симметрией, билатерально-симметричны. Эволюция билатеральной симметрии, которая позволила сформировать передний и задний (головной и хвостовой) концы, способствовала феномену, называемому цефализацией, который относится к скоплению организованной нервной системы на переднем конце животного.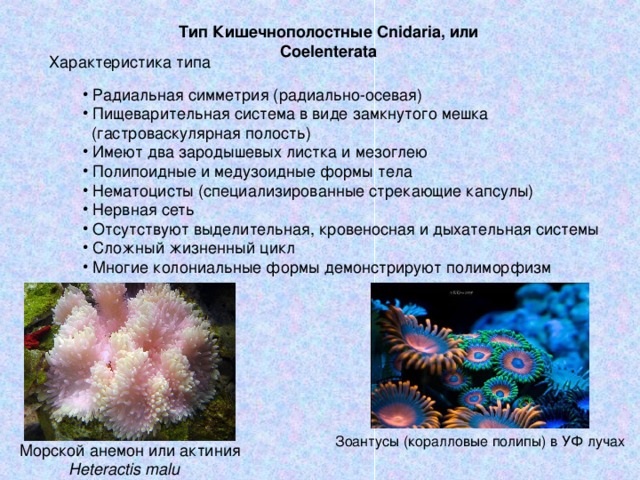 В отличие от радиальной симметрии, которая лучше всего подходит для стационарного образа жизни или образа жизни с ограниченным движением, билатеральная симметрия обеспечивает обтекаемое и направленное движение. С точки зрения эволюции, эта простая форма симметрии способствовала активной мобильности и усложнению поиска ресурсов и взаимоотношений хищник-жертва.
В отличие от радиальной симметрии, которая лучше всего подходит для стационарного образа жизни или образа жизни с ограниченным движением, билатеральная симметрия обеспечивает обтекаемое и направленное движение. С точки зрения эволюции, эта простая форма симметрии способствовала активной мобильности и усложнению поиска ресурсов и взаимоотношений хищник-жертва.
Животные типа иглокожих (такие как морские звезды, морские ежи и морские ежи) во взрослом состоянии демонстрируют радиальную симметрию, но их личиночные стадии проявляют двустороннюю симметрию. Это называется вторичной радиальной симметрией. Считается, что они произошли от билатерально-симметричных животных; таким образом, они классифицируются как двусторонне-симметричные.
Ссылка на обучение
Посмотрите это видео, чтобы увидеть краткий обзор различных типов симметрии тела.
Характеристика животных на основе особенностей эмбрионального развития
У большинства видов животных происходит разделение тканей на зародышевые листки во время эмбрионального развития. Напомним, что эти зародышевые листки формируются во время гаструляции и предопределены для развития в специализированные ткани и органы животного. У животных развиваются два или три слоя эмбриональных зародышей (рис. 4). У животных с радиальной симметрией развиваются два зародышевых листка: внутренний (энтодерма) и наружный (эктодерма). Эти животные называются диплобластов . Диплобласты имеют неживой слой между энтодермой и эктодермой. У более сложных животных (с двусторонней симметрией) развиваются три слоя ткани: внутренний слой (энтодерма), внешний слой (эктодерма) и средний слой (мезодерма). Животные с тремя слоями ткани называются триплобластами .
Напомним, что эти зародышевые листки формируются во время гаструляции и предопределены для развития в специализированные ткани и органы животного. У животных развиваются два или три слоя эмбриональных зародышей (рис. 4). У животных с радиальной симметрией развиваются два зародышевых листка: внутренний (энтодерма) и наружный (эктодерма). Эти животные называются диплобластов . Диплобласты имеют неживой слой между энтодермой и эктодермой. У более сложных животных (с двусторонней симметрией) развиваются три слоя ткани: внутренний слой (энтодерма), внешний слой (эктодерма) и средний слой (мезодерма). Животные с тремя слоями ткани называются триплобластами .
Art Connection
Рисунок 4. Во время эмбриогенеза в диплобластах развиваются два эмбриональных зародышевых листка: эктодерма и энтодерма. Триплобласты образуют третий слой — мезодерму — между энтодермой и эктодермой.
Какое из следующих утверждений о диплобластах и триплобластах неверно?
- Животные, демонстрирующие радиальную симметрию, относятся к диплобластам.
- Животные, демонстрирующие билатеральную симметрию, являются триплобластами.
- Энтодерма дает начало слизистой оболочке пищеварительного тракта и дыхательных путей.
- Мезодерма дает начало центральной нервной системе.

Утверждение 4 неверно.
Каждый из трех зародышевых слоев запрограммирован дать начало определенным тканям и органам тела. Энтодерма образует слизистую оболочку пищеварительного тракта (включая желудок, кишечник, печень и поджелудочную железу), а также слизистую оболочку трахеи, бронхов и легких дыхательных путей, а также некоторые другие структуры. Эктодерма развивается во внешний эпителиальный покров поверхности тела, центральную нервную систему и некоторые другие структуры. Мезодерма — третий зародышевый листок; он образуется между энтодермой и эктодермой в триплобластах. Этот зародышевый слой дает начало всем мышечным тканям (включая ткани сердца и мышцы кишечника), соединительным тканям, таким как скелет и клетки крови, и большинству других внутренних органов, таких как почки и селезенка.
Наличие или отсутствие целома
Дальнейшее подразделение животных с тремя зародышевыми листками (триплобластами) приводит к разделению животных, у которых может развиться внутренняя полость тела, происходящая из мезодермы, называемая целомом , и тех, у которых нет. Эта выстланная эпителиальными клетками целомическая полость представляет собой обычно заполненное жидкостью пространство, лежащее между висцеральными органами и стенкой тела. В нем находится множество органов, таких как пищеварительная система, почки, репродуктивные органы и сердце, а также система кровообращения. У некоторых животных, таких как млекопитающие, часть целома, называемая плевральной полостью, обеспечивает пространство для расширения легких во время дыхания. Эволюция целома связана со многими функциональными преимуществами. Прежде всего, целом обеспечивает амортизацию и амортизацию для основных систем органов. Органы, находящиеся внутри целома, могут свободно расти и двигаться, что способствует оптимальному развитию и размещению органов.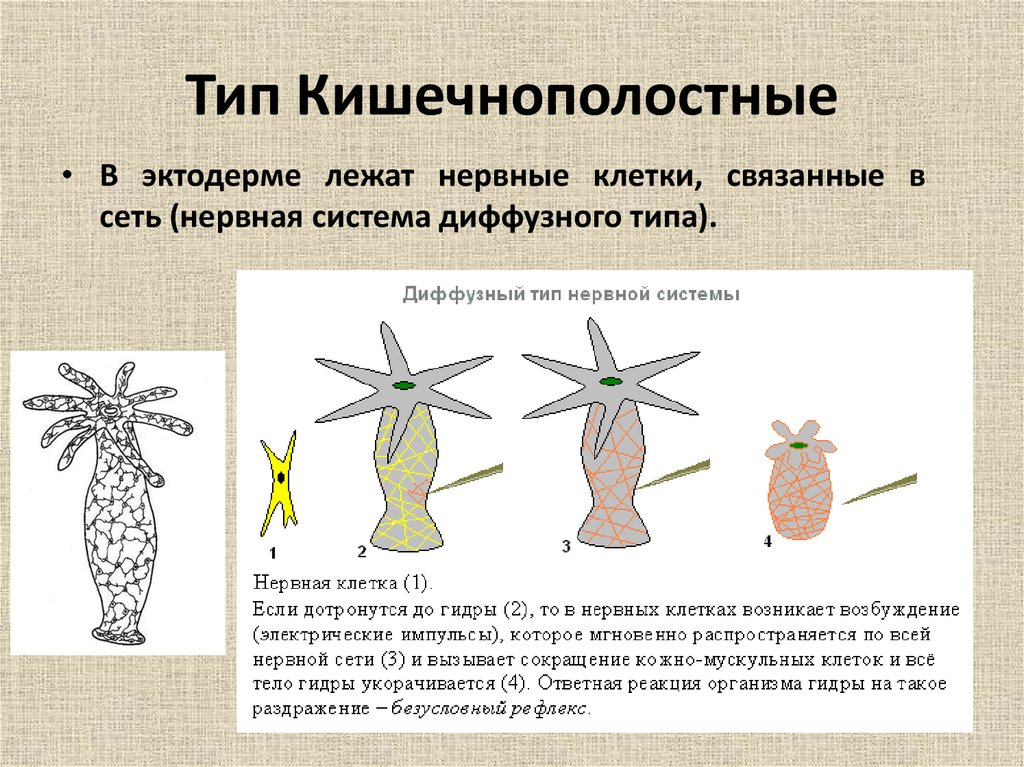 Целом также обеспечивает пространство для диффузии газов и питательных веществ, а также обеспечивает гибкость тела, способствуя улучшению подвижности животных.
Целом также обеспечивает пространство для диффузии газов и питательных веществ, а также обеспечивает гибкость тела, способствуя улучшению подвижности животных.
Триплобласты, у которых не развивается целом, называются ацеломатами , и их область мезодермы полностью заполнена тканью, хотя у них все еще есть кишечная полость. Примеры акоеломатов включают животных типа Platyhelminthes, также известных как плоские черви. Животных с истинным целомом называют эуцеломатами (или целоматами) (рис. 5). Истинный целом возникает полностью внутри зародышевого слоя мезодермы и выстлан эпителиальной мембраной. Эта мембрана также выстилает органы внутри целома, соединяя и удерживая их на месте, позволяя им свободно двигаться. Аннелиды, моллюски, членистоногие, иглокожие и хордовые — все это эуцеломаты. Третья группа триплобластов имеет немного другой целом, полученный частично из мезодермы и частично из энтодермы, который находится между двумя слоями. Хотя они все еще функционируют, они считаются ложными целомами, и этих животных называют псевдоцеломаты . Тип Nematoda (круглые черви) является примером псевдоцеломата. Истинные целоматы могут быть дополнительно охарактеризованы на основе некоторых особенностей их раннего эмбриологического развития.
Тип Nematoda (круглые черви) является примером псевдоцеломата. Истинные целоматы могут быть дополнительно охарактеризованы на основе некоторых особенностей их раннего эмбриологического развития.
Рисунок 5. Триплобласты могут быть (а) ацеломатами, (б) эуцеломатами или (в) псевдоцеломатами. Акоеломаты не имеют полости тела. Эвцеломаты имеют полость тела внутри мезодермы, называемую целомом, которая выстлана мезодермой. Псевдоцеломаты также имеют полость тела, но она зажата между энтодермой и мезодермой. (кредит a: модификация работы Яна Дерка; кредит b: модификация работы NOAA; кредит c: модификация работы USDA, ARS)
Эмбриональное развитие рта
Рисунок 6. Эуцеломатов можно разделить на две группы в зависимости от их раннего эмбрионального развития. У первичноротых часть мезодермы отделяется, образуя целом, в процессе, называемом шизоцелией. У вторичноротых мезодерма отщипывается с образованием целома в процессе, называемом энтероцелия. Долгое время считалось, что бластопор превратился в рот у первичноротых и в задний проход у вторичноротых, но недавние данные оспаривают это мнение.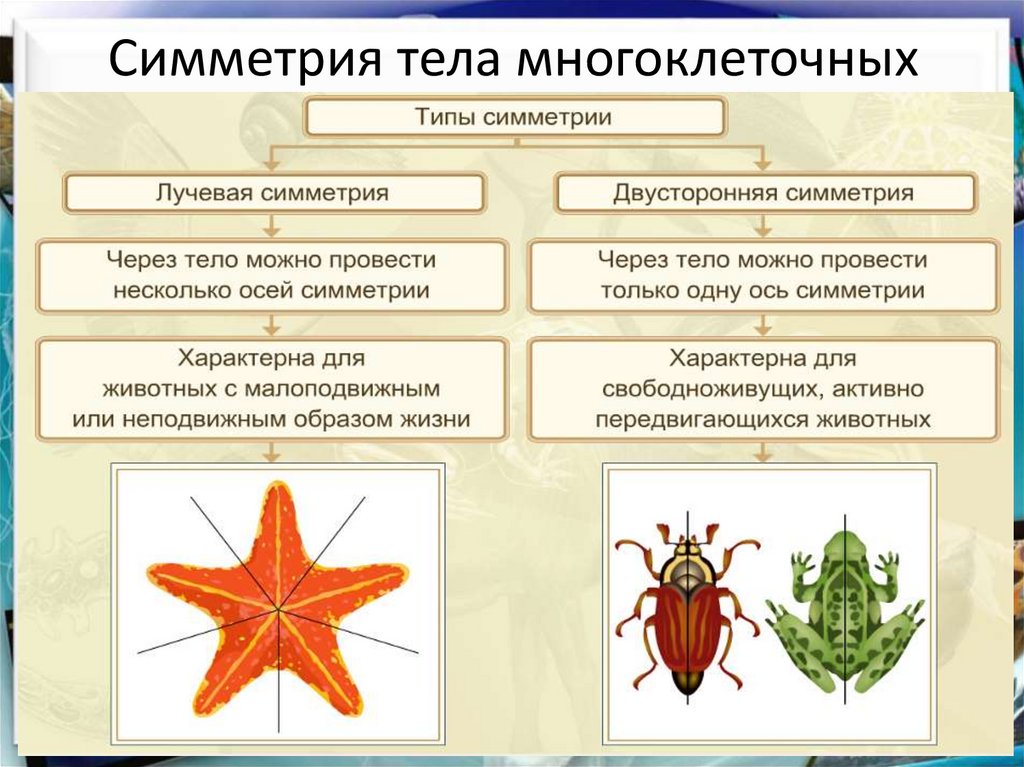
Двусторонне-симметричные триблобластные эуцеломаты можно разделить на две группы на основании различий в их раннем эмбриональном развитии. Первичноротые включают членистоногих, моллюсков и кольчатых червей. Вторичноротые включают более сложных животных, таких как хордовые, но также и некоторых простых животных, таких как иглокожие. Эти две группы разделяются в зависимости от того, какое отверстие пищеварительной полости развивается раньше: ротовое или анальное. Слово протостом происходит от греческого слова, означающего «сначала рот», а вторичноротое происходит от слова, означающего «второй рот» (в этом случае анус развивается первым). Рот или задний проход развивается из структуры, называемой бластопором (рис. 6). бластопор представляет собой углубление, образующееся на начальных стадиях гаструляции. На более поздних стадиях формируется второе отверстие, и эти два отверстия в конечном итоге дадут начало рту и анусу (рис. 6). Долгое время считалось, что бластопор развивается в рот первичноротых, а второе отверстие — в задний проход; обратное верно для вторичноротых.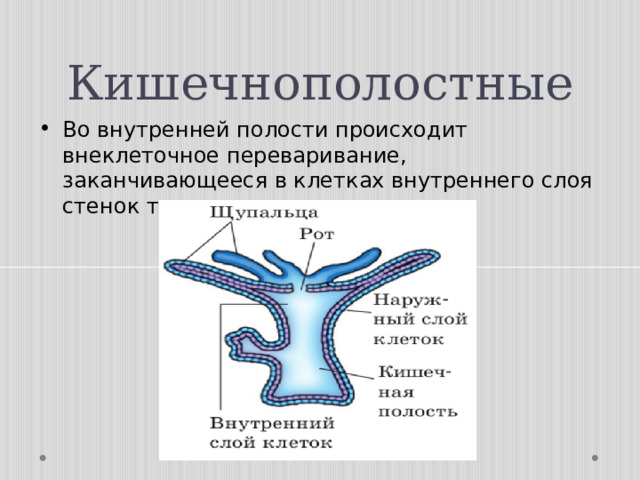 Однако недавние данные поставили под сомнение эту точку зрения на развитие бластопора первичноротых, и эта теория остается предметом споров.
Однако недавние данные поставили под сомнение эту точку зрения на развитие бластопора первичноротых, и эта теория остается предметом споров.
Еще одно различие между первичноротыми и вторичноротыми заключается в способе образования целома, начиная со стадии гаструлы. Целом большинства первичноротых формируется в результате процесса, называемого schizocoely , что означает, что во время развития твердая масса мезодермы расщепляется и образует полое отверстие целома. Вторичноротые отличаются тем, что их целом образуется в результате процесса, называемого энтероцелия . Здесь мезодерма развивается в виде мешочков, отщипываемых от ткани энтодермы. Эти мешочки в конечном итоге сливаются, образуя мезодерму, которая затем дает начало целому.
Самое раннее различие между первичноротыми и вторичноротыми заключается в типе дробления зиготы. Первичноротые подвергаются спиральному расщеплению , что означает, что клетки одного полюса эмбриона повернуты и, таким образом, смещены по отношению к клеткам противоположного полюса.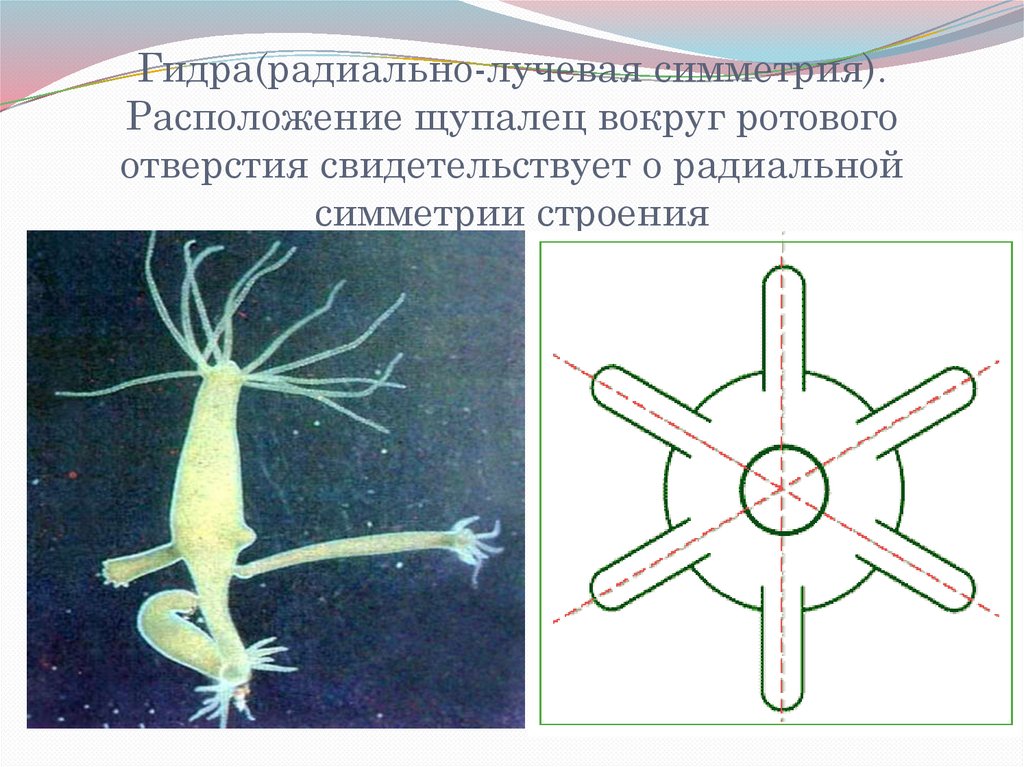 Это связано с косым углом спайности. Вторичноротые претерпевают радиальное дробление , где оси дробления либо параллельны, либо перпендикулярны полярной оси, что приводит к выравниванию клеток между двумя полюсами.
Это связано с косым углом спайности. Вторичноротые претерпевают радиальное дробление , где оси дробления либо параллельны, либо перпендикулярны полярной оси, что приводит к выравниванию клеток между двумя полюсами.
Существует второе различие между типами дробления у первичноротых и вторичноротых. В дополнение к спиральному дроблению первичноротые также подвергаются детерминированному дроблению . Это означает, что уже на этой ранней стадии судьба развития каждой эмбриональной клетки уже определена. Клетка не имеет способности развиваться в любой тип клеток. Напротив, вторичноротые претерпевают неопределенное дробление , при котором клетки еще не предопределены на этой ранней стадии для развития в специфические типы клеток. Эти клетки называются недифференцированными клетками. Эта характеристика вторичноротых отражается в существовании известных эмбриональных стволовых клеток, которые обладают способностью развиваться в клетки любого типа до тех пор, пока их судьба не будет запрограммирована на более поздней стадии развития.
Evolution Connection
Эволюция Целома
Одним из первых шагов в классификации животных является исследование тела животного. Изучение частей тела говорит нам не только о роли рассматриваемых органов, но и о том, как мог развиваться вид. Одной из таких структур, которая используется в классификации животных, является целом. Целом представляет собой полость тела, которая формируется в ходе раннего эмбрионального развития. Целом позволяет разделить части тела, так что могут развиваться различные системы органов и возможен транспорт питательных веществ. Кроме того, поскольку целом целом представляет собой полость, заполненную жидкостью, он защищает органы от ударов и сдавления. Простые животные, такие как черви и медузы, не имеют целома. У всех позвоночных есть целом, который помог им развить сложные системы органов.
Животные, не имеющие целома, называются акоеломатами. Плоские черви и ленточные черви являются примерами акоеломатов. Они полагаются на пассивную диффузию для транспортировки питательных веществ по всему телу.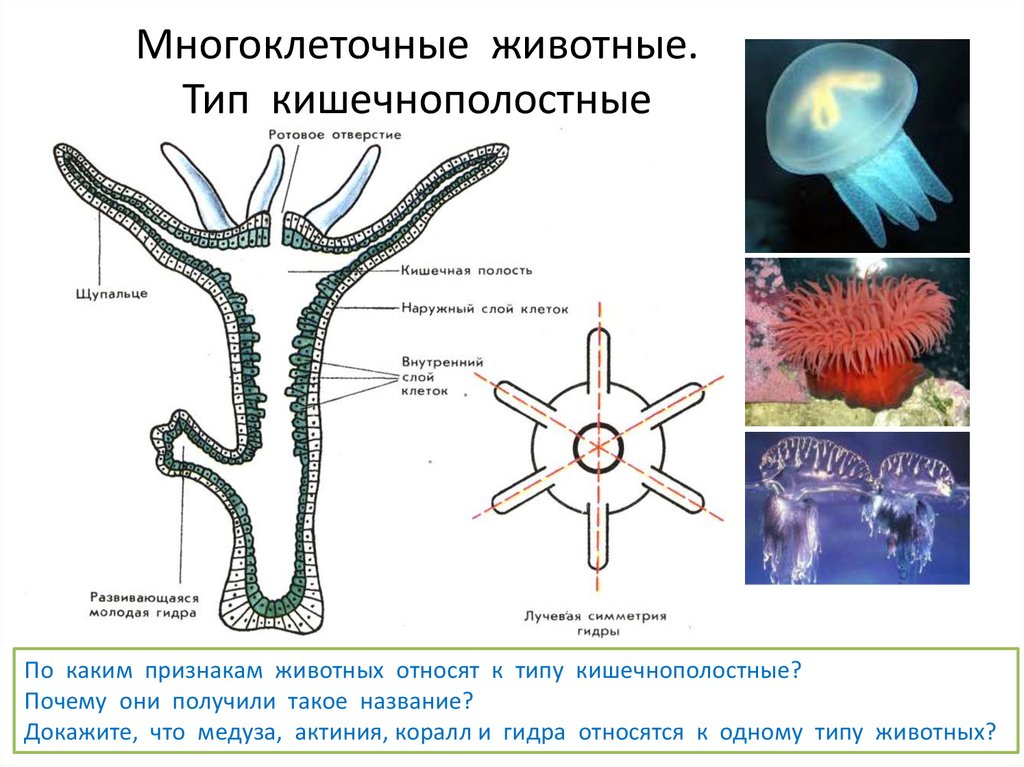 Кроме того, внутренние органы акоеломатов не защищены от раздавливания.
Кроме того, внутренние органы акоеломатов не защищены от раздавливания.
Животных, имеющих настоящий целом, называют эуцеломатами; все позвоночные — эуцеломаты. Целом развивается из мезодермы во время эмбриогенеза. Брюшная полость содержит желудок, печень, желчный пузырь и другие органы пищеварения. Другая категория беспозвоночных животных по признаку полости тела – псевдоцеломаты. Эти животные имеют псевдополость, которая не полностью выстлана мезодермой. Примеры включают паразитов-нематод и мелких червей. Считается, что эти животные произошли от целоматов и, возможно, утратили способность образовывать целом из-за генетических мутаций. Таким образом, этот этап раннего эмбриогенеза — образование целома — оказал большое эволюционное влияние на различные виды животного мира.
Резюме раздела
Организмы в животном мире классифицируются на основе их морфологии тела и развития. Настоящие животные делятся на животных с радиальной и двусторонней симметрией.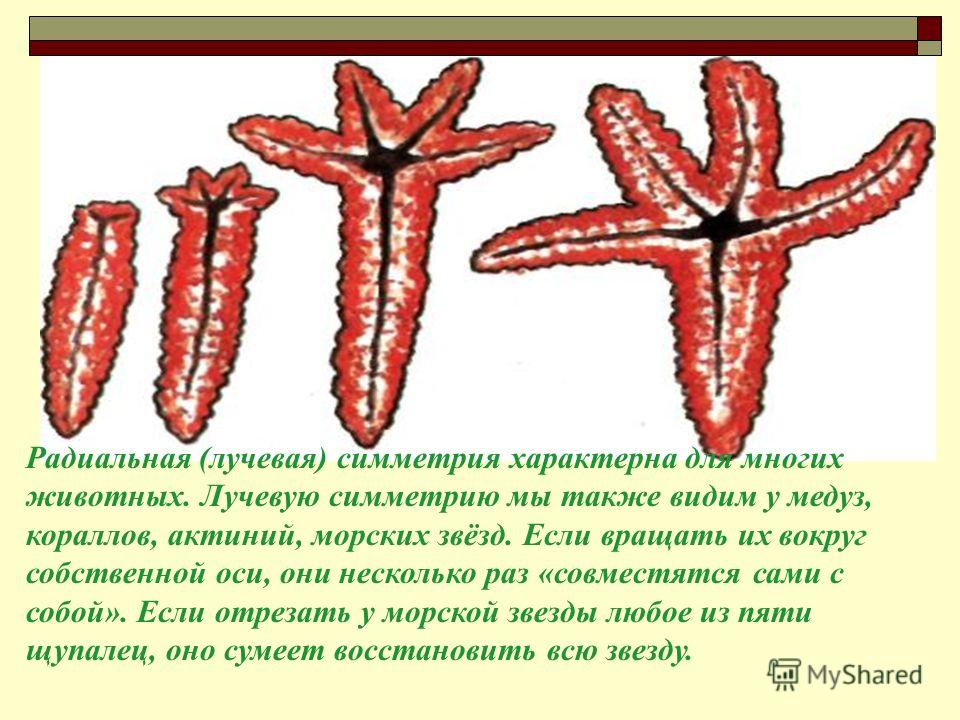
Leave A Comment