
Дендриты важнее для мозга, чем ранее считалось — Наука
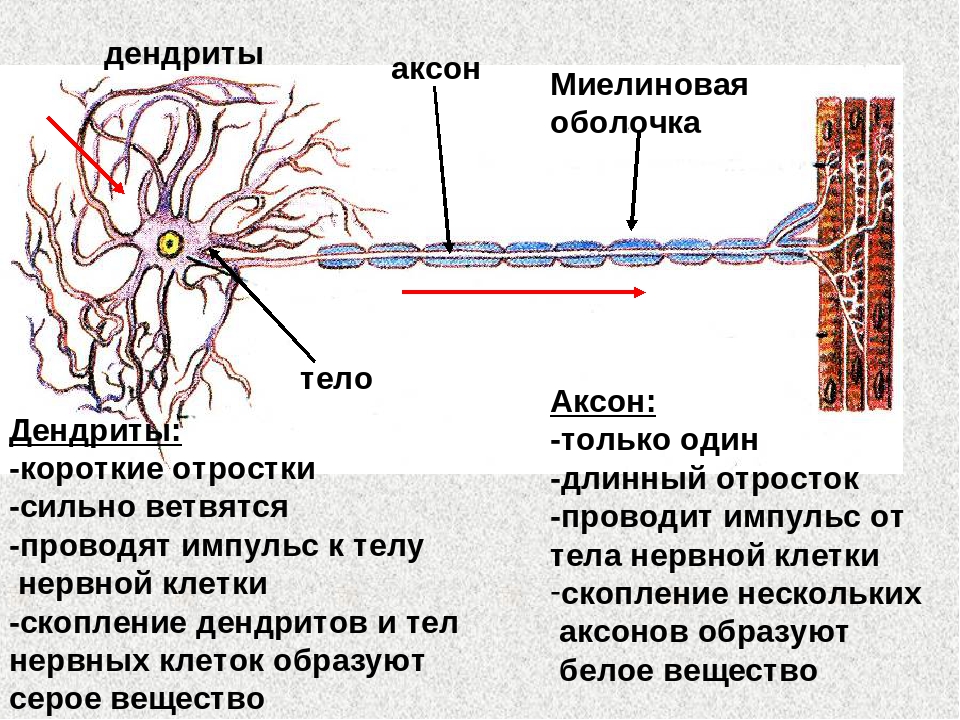
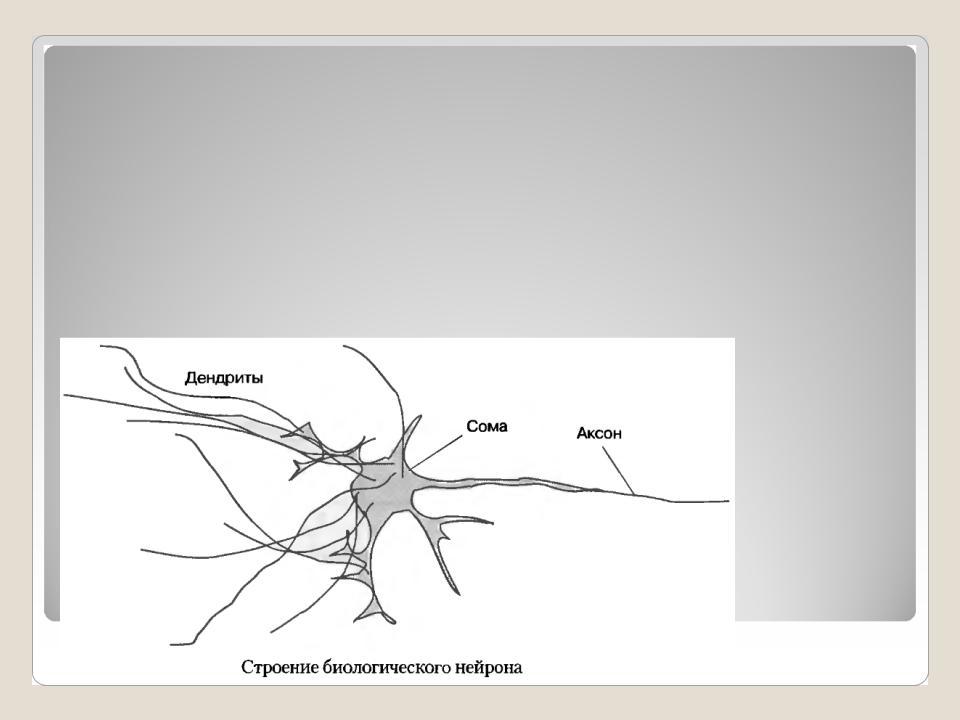
Нервная клетка — нейрон — имеет сложную структуру и состоит из тела (сомы) и отростков — аксона и нескольких дендритов. Аксон — передающий отросток, через него импульс идет от тела клетки к другому нейрону. Дендриты — принимающие отростки, они собирают импульсы от других нейронов и передают их телу нейрона. Как сегодня известно, дендриты составляют почти 90% нервной ткани. И новое исследование показывает, какую большую роль на самом деле играют дендриты в работе мозга и нервной системы.
Ранее ученые предполагали, что главную роль в генерировании и обработке импульсов играет именно тело нервной клетки. Дендритам ранее отводилась функция простых проводников. Новая работа американских ученых показала, что дендриты сами могут генерировать импульс. Это первое исследование такого рода. До этого считалось, что электрическая активность дендритов зависела от электрической активности тела нейрона-передатчика: какой импульс вышел из нейрона, такой и пройдет по дендриту.
Нейробиологи использовали современные технологии, позволяющие ввести микроэлектроды, показывающие электрическую активность, не в сам дендрит, а рядом с ним. Ранее электроды могли вводить лишь в сам дендрит, после чего нейрон погибал. Новая технология позволила наконец измерить электрическую активность самих дендритов. Ученые в течение нескольких дней наблюдали за дендритами задней теменной коры крыс, которые двигались, гуляя по клетке. Именно задняя теменная кора была выбрана не случайно, ее основная функция — это координация движений.
Сравнение активности нейронных тел и дендритов показало, что отростки работают намного интенсивнее: по сравнению с нейронными телами они генерировали в десять раз больше импульсов во время бодрствования и в пять раз больше — во время сна. Более того, как говорят ученые, дендрит непосредственным образом участвует в начальном зарождении импульса. И если нейрон генерирует импульс по принципу «есть-нет» (то есть импульс либо есть, либо нет), что похоже на цифровую кодировку информации, то дендрит, как оказалось, работает несколько иначе — по аналоговому принципу, то есть сигнал в нем может меняться в определенном диапазоне.
«Фундаментальное представление нейробиологии заключалась в том, что нейроны работают по цифровому принципу. Они либо генерируют импульс, либо нет. Наши результаты показывают, что дендриты не ведут себя исключительно как цифровое устройство. Они могут генерировать цифровые, «да-нет»-импульсы, но они также показывают большие аналоговые колебания. Это серьезный отход от того, что неврологи полагают уже около 60 лет», — резюмировали свою работу исследователи.
Исследование опубликовано в журнале Science.
Ранее ученые обнаружили, какие нарушения в строении нейронов ведут к рассеянному склерозу.
Евгения Щербина
Самостоятельные дендриты
Недавно мы писали, как новые методы детекции электрической активности нейронов позволили расширить функции мозжечка. В новом исследовании новые методы используются шире – чтобы пересмотреть в целом в какой части нейрона возникает (или не возникает) возбуждение.
В новом исследовании новые методы используются шире – чтобы пересмотреть в целом в какой части нейрона возникает (или не возникает) возбуждение.
В нынешнем исследовании ученые
использовали тетрод, специальный вид электродов, который измеряет
изменение напряжения на расстоянии, без проникновения в дендрит.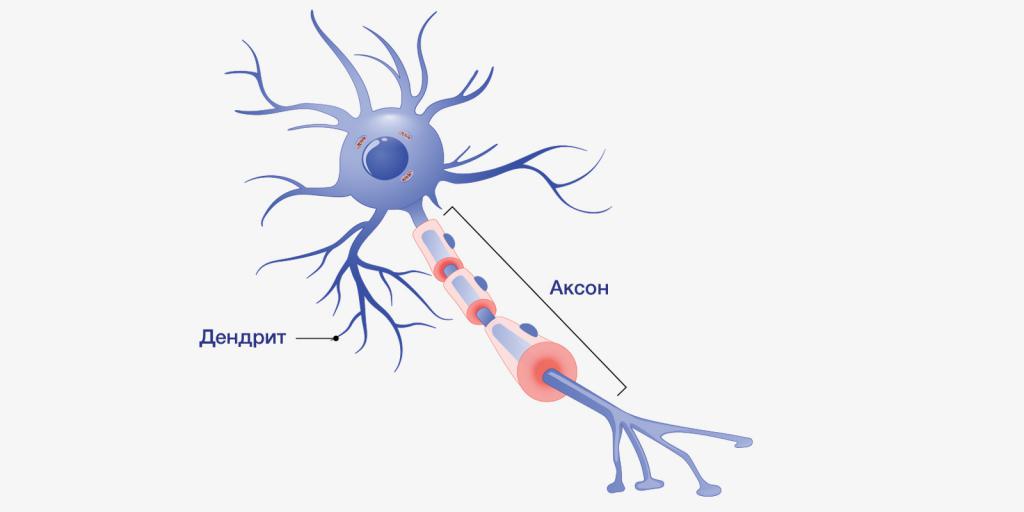 Естественно, если разместить этот тетрод у головы животного, то
измерение активности отдельных нейронов, не говоря уже о дендритах,
получить не получится. Необходимо с ювелирной точностью внедрить тетрод к
отдельных дедритам хирургическим путем. При этом действовать надо
быстро — тетроды вызывают иммунную реакцию, и соответственно быстро
обволакиваются глией, делая измерения невозможными.
Естественно, если разместить этот тетрод у головы животного, то
измерение активности отдельных нейронов, не говоря уже о дендритах,
получить не получится. Необходимо с ювелирной точностью внедрить тетрод к
отдельных дедритам хирургическим путем. При этом действовать надо
быстро — тетроды вызывают иммунную реакцию, и соответственно быстро
обволакиваются глией, делая измерения невозможными.
Всё же, героическими усилиями группа из Университета Калифорнии Лос-Анжелес смогла получить измерения поведения отдельных дендритов у мышей на протяжении четырех дней. Удивительным образом они зафиксировали, что дендриты возбуждались чаще, чем тела нейронов, с которыми они связывались: частота возбуждения дендритов была в 5 раз выше во сне и в 10 раз выше при бодрствовании чем частота возбуждения тел нейронов. Логичное объяснение этому — это то, что дендриты не пассивны, а могут сами генерировать возбуждение.
Этот вывод приглашает как к глобальному
пересмотру учебников нейробиологии, так и повсеместному пересчету многих
данных о функционировании нейронов — очевидно, в формулах нужно ввести
новую переменную: самостоятельное возбуждение дендритов.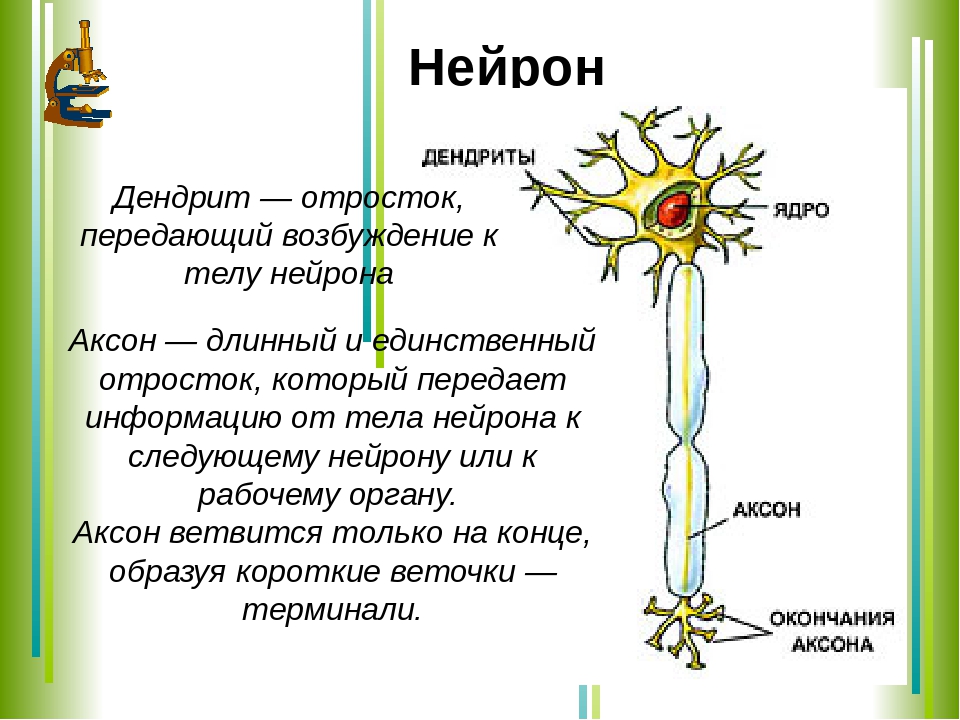
Оригинал текста на сайте Нейроновости
Нейронные сети — Библиотека разработчика Programming Store
Нейронные сети – это вычислительная эмуляция работы нейронов головного мозга. Суть состоит в том, что эмулируемый нейрон – это некая математическая функция, на вход которой подаются данные (первичные или с выходов других нейронов), а выходные данные функции могут подаваться на другие искусственные (эмулируемые) нейроны и тоже обрабатываться. Таким образом, связав в сеть несколько нейронов, мы можем находить закономерности в различных набора данных, например, предсказывать числовые ряды (погода, биржевые котировки и так далее). Однако, этим не ограничивается применение нейронных сетей. НС можно также использовать для распознавания образов, в задачах управления и даже для сжатия данных.
Следует, правда, четко осознавать, что могу нейронные сети и чего не могут. Если задача сводиться к тому, что на входе некий данные, которым должен каким-то волшебным образом соответствовать выходной набор данных по принципу Ключ->Значение, и требуется как раз найти Значение, то эти задачи решаются нейронными сетями.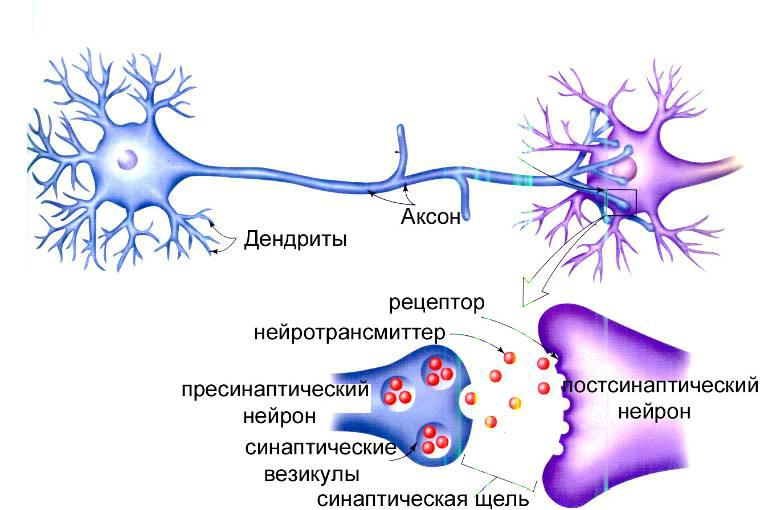 Например, распознавание образов, предсказание погоды или биржевых котировок. В данном случае тут как раз мы имеем дело с таким соответствием Ключ->Значение, и нам известен Ключ, но не известно значение. В случае, например, биржевых котировок в качестве Ключа выступают несколько предыдущих значений, а в качестве Значения — то что мы по ним предсказываем. Но совсем другое дело, когда нам, например, надо вычислить квадратный корень или составить план действий в какой-то определенной ситуации. Такие задачи нейросети не решает. То есть, нейросети не эффективны там, где нужен точный результат или где необходимы процедурные знания.
Например, распознавание образов, предсказание погоды или биржевых котировок. В данном случае тут как раз мы имеем дело с таким соответствием Ключ->Значение, и нам известен Ключ, но не известно значение. В случае, например, биржевых котировок в качестве Ключа выступают несколько предыдущих значений, а в качестве Значения — то что мы по ним предсказываем. Но совсем другое дело, когда нам, например, надо вычислить квадратный корень или составить план действий в какой-то определенной ситуации. Такие задачи нейросети не решает. То есть, нейросети не эффективны там, где нужен точный результат или где необходимы процедурные знания.
Нейронные сети одно из перспективных направлений развития искусственного интеллекта. Здесь за основу взят принцип организации нервной системы живого существа. Теоретически, можно смоделировать даже мозг человека. Правда, практически это сделать в настоящее время не реально, так как мозг человека содержит 10
Дендриты — это ветвеобразные отростки, которые обеспечивают сбор информации от других нейронов или рецепторов. Тело нервной клетки называется сома. В ней происходят сложные биохимические процессы, благодаря которым происходят сложные нелинейные преобразования сигналов.
Сигналы, распространяющиеся в биологической нейронной сети, представляют собой короткие электрические импульсы. Под их воздействием изменяется потенциал самих нейронов. В момент достижения им некоторой пороговой величины вырабатывается импульс, который распространяется вдоль аксона. Потенциал сомы снижается, нейрон разряжается. Через некоторое время нейрон может снова сформировать импульс. Если импульсы, попадая на синапс повышают потенциал сомы, то это 
При построении искусственной нейронной сети (ИНС), данную модель, как правило упрощают. В настоящее время существуют несколько моделей нейронов. И так, рассмотрим модель Маккаллоха и Питтса:
которая выражается формулой:
где g() — функция преобразования нейрона, θi — пороговое значение, n — количество входов нейрона. Для модели нейронной сети Маккаллоха — Питтса функцию g() можно выразить формулой:
Эта функция так же называется функцией Хэвисайда H(u).
Таким образом, если сумма входов, помноженных на коэффициенты, превышает θ, то на выходе нейрона мы получим 1, в противном случае 0. Стоит заметить, что выходной сигнал нейрона может меняться только в дискретные моменты времени.
Функция, определяющая способ объединения входных признаков называется сетевой или базовой функцией. Она обозначается net. Сейчас мы разобрали лишь частный случай такой функции:
На самом деле, сетевые функции и функции преобразования нейрона могут быть разные, например, квадратичная сетевая
Радиальная:
Могут быть и вообще экзотические, типа произведения.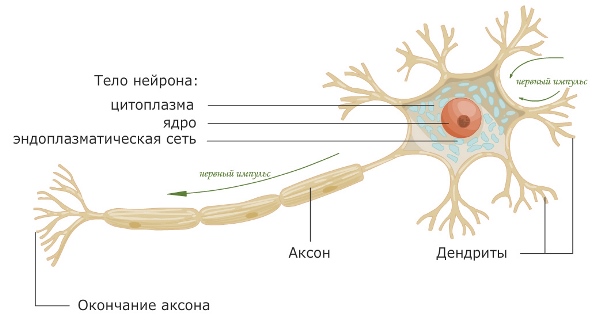
Теперь поговорим о передаточных функциях. Их еще называют активаторными функциями. В качестве функции преобразование может быть использована ступенчатая функция:
линейная функция с насыщением:
Сигмоидная функция:
и, наконец, гауссова функция:
где c и σ — постоянные коэффициенты.
Если сетевая функция линейная, а функция преобразования ступенчатая или знаковая, то такие нейроны называются линейные, а нейросети на их основе нейронными сетями с линейными пороговыми элементами
Рассмотрим пример. Пусть у нас есть пороговый нейрон с порогом равным 2, а весовые коэффициенты 1. Такой нейрон будет выполнять функцию «И» (для случая ступенчатой функции преобразования)
| X1 | 0 | 0 | 1 | 1 |
| X2 | 0 | 1 | 0 | 1 |
| Y | 0 | 0 | 0 | 1 |
А если у того же нейрона пороговое значение поставить 1, то это будет уже функция ИЛИ:
| X1 | 0 | 0 | 1 | 1 |
| X2 | 0 | 1 | 0 | 1 |
| Y | 0 | 1 | 1 | 1 |
А вот функцию XOR один нейрон вычислить не может, так как она не является линейно разделимой. Тут уже потребуется как минимум, два нейрона.
Нейронные дендриты манипулируют информацией
Дендриты, как и аксоны, способны генерировать собственный импульс, с помощью которого они могут влиять на избирательность аналитических центров коры по отношению к внешнему раздражителю.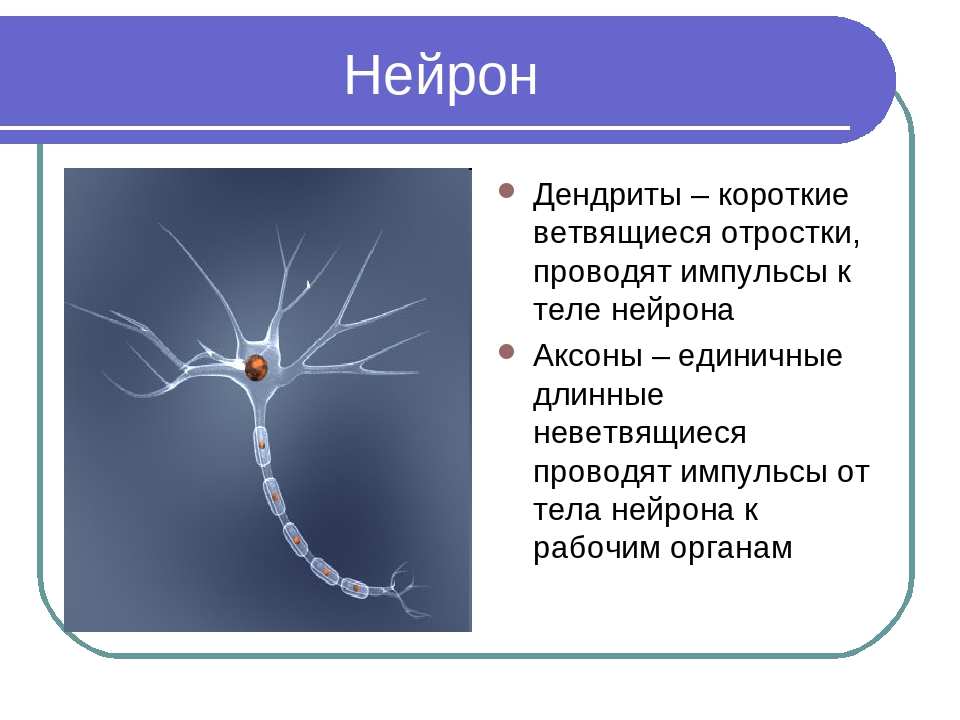
У нервной клетки, как известно, есть два вида отростков-проводников: аксон, проводящий сигнал «на выход», то есть к следующему нейрону или какой другой клетке, и сильно разветвлённые дендриты, которые работают «на вход», то есть принимают сигнал от аксонов других нейронов.
Долгое время считалось, что дендриты — это просто пассивные коллекторы сигнала, что они могут его только проводить, но никак не изменять или генерировать. Но, по-видимому, это не совсем так — учёные обнаружили, что в ответ на принятый сигнал дендриты способны сами реагировать на него и как-то модулировать обработку этого импульса в аналитических центрах.
То, что дендриты располагают молекулами, подобными тем, что есть в аксонах, и что благодаря им дендриты могут генерировать собственный электрический импульс, исследователи знали давно, но опыты на эту тему ставились на препарированных тканях и клетках. То, как обстоят дела в живых мозгах, проверили Спенсер Смит и его коллеги по Медицинской школе Университета Северной Каролины (США) и Университетскому колледжу Лондона (Великобритания). Им удалось напрямую «подслушать», что происходит в отдельных дендритах пирамидальных нейронов визуальной коры крыс.
Им удалось напрямую «подслушать», что происходит в отдельных дендритах пирамидальных нейронов визуальной коры крыс.
Животным предъявляли некий визуальный стимул и следили за импульсами в нейронных отростках. Как пишут авторы работы в Nature, при визуальной стимуляции в дендритах появлялись такие импульсы, которые отличались от обычных импульсов проведения принятого сигнала. Дальнейшие эксперименты лишь подтвердили, что дендриты нейронов могли возбуждаться в то время, когда все остальные части нейрона молчали.
Исследователи подчёркивают, что эти нестандартные импульсы появлялись как раз тогда, когда внимание животного захватывало нечто, попадающее в поле зрения, и такая электрическая активность дендритов действительно играла роль в ориентации и внимании по отношению к стимулу.
Дендрит и стеклянная игла, воткнутая в нейронный отросток, чтобы учёные могли следить за его электрической активностью
То есть, делают вывод учёные, добавочная активность дендритов повышает избирательность нейронного ответа на раздражение извне и помогает мозгу сконцентрировать внимание.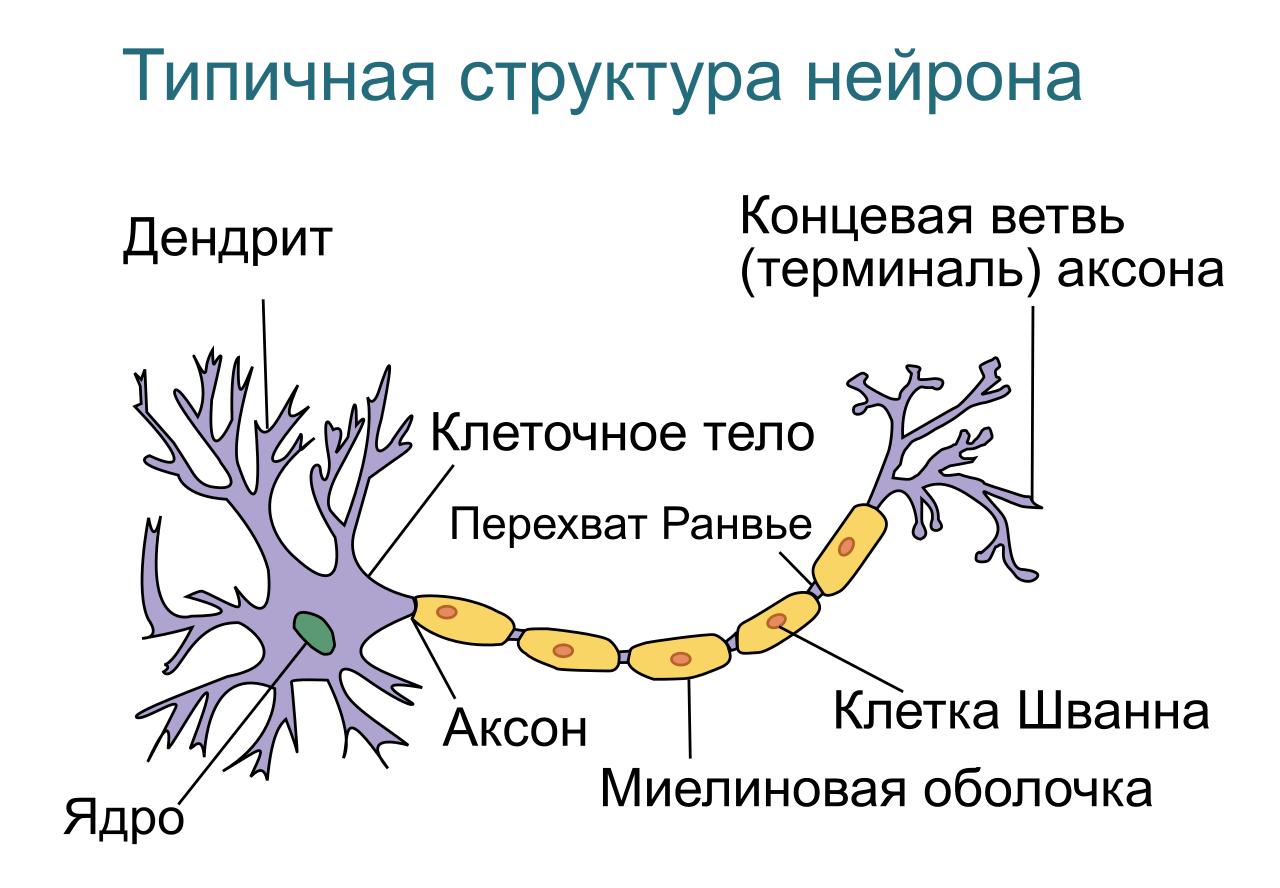
Дендриты оказались не просто собирателями и проводниками импульсов, но и своеобразными «вычислительными элементами». Об этой способности исследователи, может, и подозревали, но никогда не видели её воочию.
Нейронная трансформация пломбира
Наш мозг способен на многие вещи, и некоторые из них совсем уж странные: например, он может связать в нашем восприятии пломбир с углём, а восьмерку с известью или лишить нас возможности что-либо забывать. Звучит едва ли не магически, но никакой магии тут нет — только действие нейронов, синапсов, аксонов и дендритов. Писатель и программист Владимир Губайловский в книге «Искусственный интеллект и мозг человека», вошедшей в лонг-лист премии «Просветитель», подробно описал, как все это работает и к чему приводит.
Владимир Губайловский. Искусственный интеллект и мозг человека. М.: Наука, 2019. Содержание
Мозг как биокомпьютер
Производительность компьютеров IBM, используемых NASA при реализации проекта «Apollo», была примерно в сто тысяч раз меньше «оперативки» средненького современного смартфона.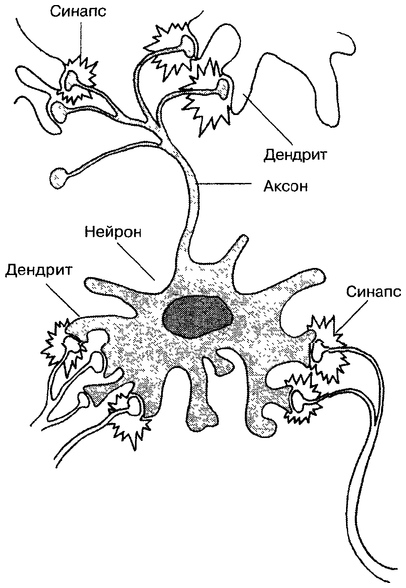 Темпы роста производительности техники в последние десятилетия беспрецедентны, но динамика этого разгона имеет свой предел.
Темпы роста производительности техники в последние десятилетия беспрецедентны, но динамика этого разгона имеет свой предел.
«Достигнута максимальная не только на сегодняшний день, но и вообще возможная частота процессора того типа, который мы сегодня массово используем, — это 3–5 ГГц (<…> мы близко подошли к „тепловому барьеру” Ландауэра), практически достигнута предельная плотность размещения вентилей на кремниевой подложке) <…>. Мы перестали ускорять процессоры, делая упор на распараллеливание процессов (в частности, увеличивать количество ядер)», — отмечает Владимир Губайловский.
Несравнимо большими показателями отличается человеческий мозг, который в каком-то смысле можно назвать «биологическим аналоговым компьютером». Как у всякого компьютера, у мозга есть «порты ввода / вывода», через которые он получает данные из внешнего мира и выводит преобразованную информацию. «Порты ввода» — это органы чувств. Если человек потеряет их (например, из-за тяжелой травмы), он окажется замкнутым внутри самого себя и утратит большую часть возможностей коммуникации с внешним миром.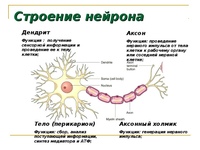
На сегодняшний день наиболее реалистичная модель мозга — искусственная импульсная нейронная сеть (Spiking Neural Network — SNT). В ней нейроны обмениваются короткими импульсными сигналами (1–2 мс), но это лишь малая толика многообразия их «взаимоотношений». Чтобы представить, насколько эта модель далека от более-менее обстоятельного и тем более полного «картографирования», представим, какие можно было бы сделать выводы о движении на оживленных улицах, основываясь только на свете автомобильных фар. Детали (такие, например, как особенности дорожного полотна) ускользают, но кое-что мы все-таки узнаем: «один район может быть финансовым центром, в другом <…> находится аэропорт. По выходным движение потоков наверняка изменится. В плохую погоду больше людей останутся дома <…> однако все равно будут использоваться постоянно существующие улицы и дороги».
Правила дорожного движения
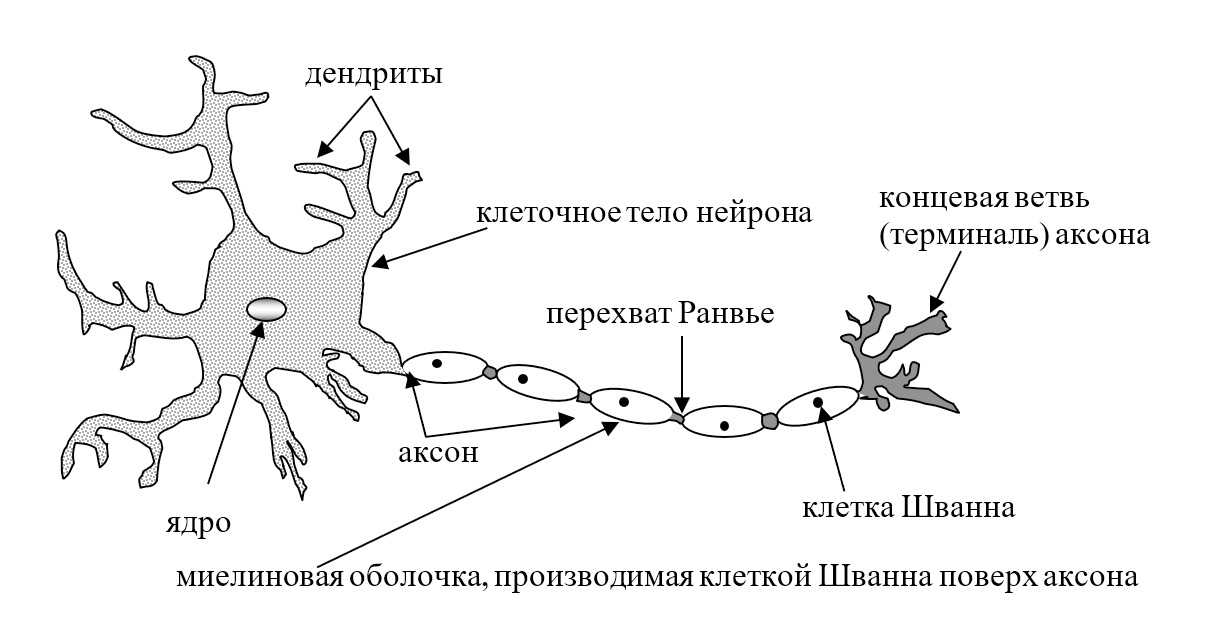
Главные жители этого «большого города» — нейроны, клетки нервной системы размером от 3 до 130 микрон. У каждого нейрона есть тело с ядром, дендриты («входы», нервные окончания, по которым поступают сигналы из внешнего мира или от других нейронов) и аксоны («выходы»). Аксон обычно один (но может ветвиться на конце), а вот количество дендритных ветвей может достигать сотен тысяч.
У каждого нейрона есть тело с ядром, дендриты («входы», нервные окончания, по которым поступают сигналы из внешнего мира или от других нейронов) и аксоны («выходы»). Аксон обычно один (но может ветвиться на конце), а вот количество дендритных ветвей может достигать сотен тысяч.
Места контактов между нейронами называются синапсами — они бывают электрическими и химическими. Электрические обеспечивают прямой контакт — они нужны, например, когда необходима скорость реакции (обжег руку — отдернул). В химических синапсах задействован нейромедиатор: аксон одного нейрона выбрасывает вещество, которое принимает дендрит другого. Это входные данные.
«Нейрон анализирует данные и решает: формировать ему потенциал действия или погодить. Информация на входе весьма разнообразна — одни аксоны передают возбуждающие медиаторы, другие — „молчат”, третьи передают тормозящие медиаторы. Анализ, который проводит нейрон, зависит не только от входных данных, но от его собственной структуры — например, наличия некоторых важных белков».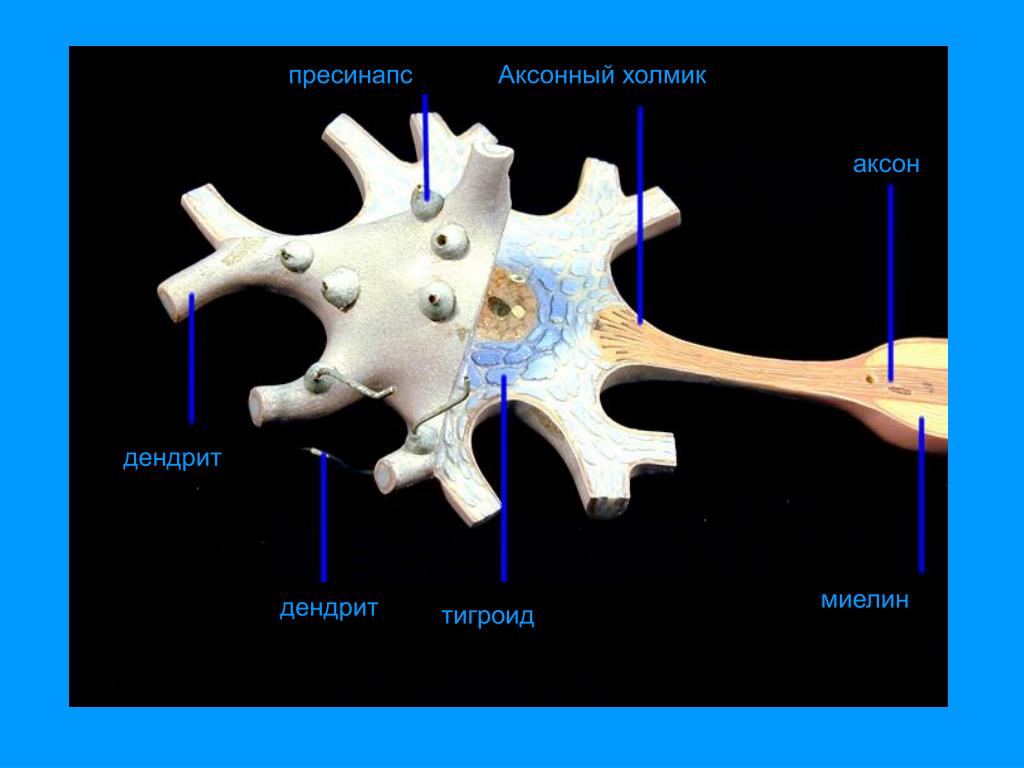
То есть нейрон в рабочем состоянии — это такой заряженный конденсатор. На внешнем слое мембраны концентрируются положительные ионы (ионы натрия — Na+), на внутреннем — возникает отрицательный заряд.
Если нейрон решает действовать, открываются натриевые канальцы, ионы проникают внутрь и начинается деполяризация — заряды нейтрализуются.
Сигнал деполяризации бежит по аксону до конца и дает команду выбросить нейромедиатор в синапсическую щель — передать сигнал дендритам следующего нейрона. После этого нейрон некоторое время «заряжается» (помогают окружающие его глиальные клетки, которые кормят нейрон глюкозой и защищают — например, от микроорганизмов). После подпитки нейрон снова готов к действию. Таким образом нейроны образуют пути, по которым передаются химические сигналы.
Нейроны и синапсы — химический синапс. 1 — тело нейрона, 2 — дендриты, 3 — аксон, 4 — передаваемые нейроном электрические импульсы, 5 — молекулы нейротрансмиттеров, 6 — синаптическая щель, 7 — постсинаптический рецептор
Анатомия памяти
Информация, поступающая в мозг по сенсорным каналам, сначала попадает в кратковременную рабочую память (КРП).
«Здесь мы не будем касаться вопроса, а существует ли такая память „физически” или есть только долговременная <…> и мозг просто выделяет из нее специфический фрагмент в виде КРП для своих операционных нужд (такая точка зрения вовсе небезосновательна). Мы будем исходить из того, что КРП — память, в которую информация попадает из входных каналов, и эта память характеризуется двумя параметрами — емкостью и временем хранения».
При этом емкость КРП очень мала (3–7 объектов), а время «хранения» — крайне ограничено (5–15 секунд). Оба параметра существенно зависят от концентрации внимания. Чем мы сосредоточеннее, тем КРП больше и тем дольше она удерживает объекты.
Человек — это не tabula rasa, как полагал Джон Локк. Когда мы рождаемся, наш мозг уже многое «знает». Структура мозга (его главные области и некоторые проводящие пути) уже сформированы. Структура будет меняться (усложняться или упрощаться), но основная топология остается неизменной. Главная перестройка происходит в синапсах.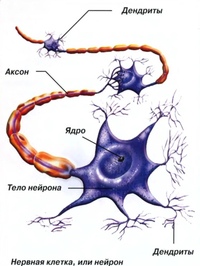 Нейроны меняют частоту импульсов, синапсы — пропускную способность. Так возникают новые, быстро проводящие сигнал нейронные контуры.
Нейроны меняют частоту импульсов, синапсы — пропускную способность. Так возникают новые, быстро проводящие сигнал нейронные контуры.
«Так и реализуется долговременная память. Она распределена по всему мозгу, но представляет собой не нули и единички — намагниченные домены на пластине жесткого диска или проколы на плоскости CD, — а долговременные структурные перестройки нейронной сети. Дендритные шипики, если обучение заканчивается, могут рассасываться — это забывание».
Память подразделяется на имплицитную (информация сохраняется в мозге без нашего активного участия, мы не можем ее вспомнить сознательным усилием) и эксплицитную. А последняя, в свою очередь, — на семантическую (что мы знаем) и эпизодическую (что мы помним).
Как отмечает Губайловский, именно эпизодическая память, формирующаяся в возрасте трех-четырех лет, и есть основа идентичности человека: если эпизодическая память разрушается при болезни Альцгеймера, человек уже не может ничего вспомнить, хотя еще многое знает.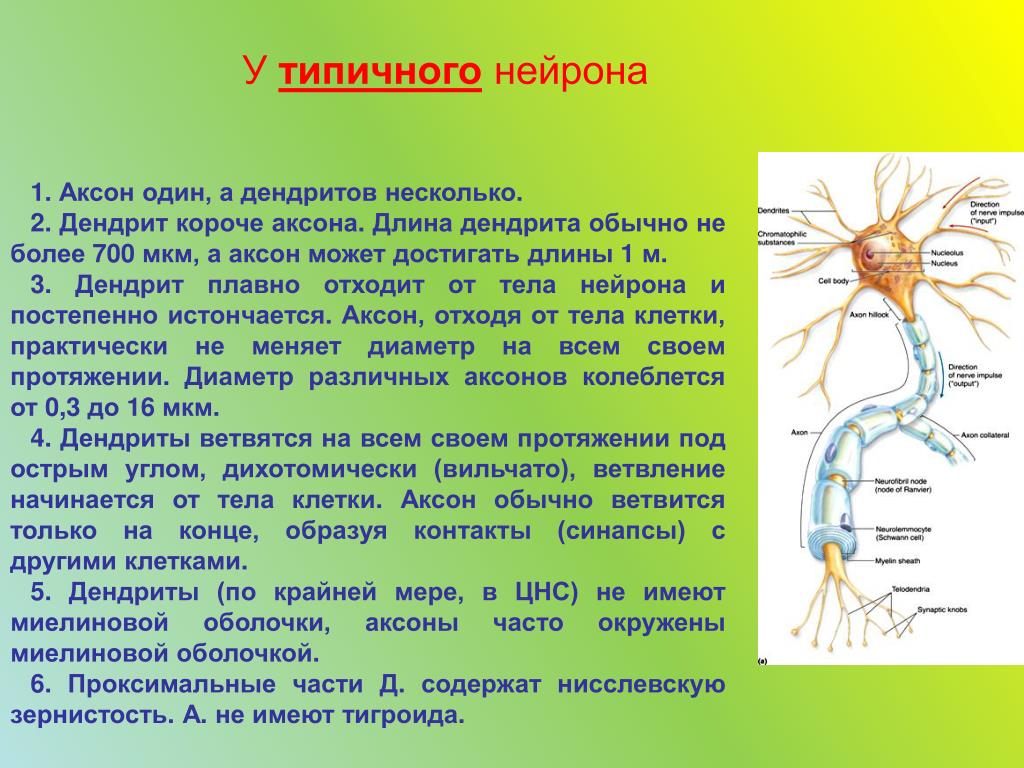
Эпизодическая память активно взаимодействует с семантической памятью. То, что мы знаем, своеобразно коррелирует с тем, что мы можем вспомнить. Семантическая память первоначально формируется именно как эпизодическая: сначала мы помним не только, что мы узнали, но и когда и как мы это узнали — сохраняется весь комплекс восприятия, который сопровождал процесс узнавания.
Пока смерть не разлучит нас
Владимир Губайловский вспоминает миф о том, что каждые семь лет все клетки нашего тела обновляются. И развенчивает его: в нашем организме есть клетки, которые вместе с нами рождаются и умирают, — это клетки сердечной мышцы и нервной системы.
Они не делятся, как клетки кожи или внутренних органов. Если клетки сердца (например, после инфаркта) погибают, им нет замены. Аналогичная история с нейронами: после тяжелой травмы спинного мозга человек может остаться полностью или частично парализованным.
Количество нейронов коры головного мозга достигает максимума на 28–32 неделе пренатального периода, а к моменту рождения уменьшается на 70 %. Нейрогенез (рождение нейронов) и апоптоз (умирание) в последние недели беременности идет невероятно интенсивно. Погибшие нервные клетки уничтожаются макрофагами, попадающими в нервную систему из крови. Не менее интенсивно идет и процесс образования синапсов. К моменту рождения наиболее развиты синапсы сенсорно-моторной коры — количество соединений между ними быстро убывает до двух лет.
Нейрогенез (рождение нейронов) и апоптоз (умирание) в последние недели беременности идет невероятно интенсивно. Погибшие нервные клетки уничтожаются макрофагами, попадающими в нервную систему из крови. Не менее интенсивно идет и процесс образования синапсов. К моменту рождения наиболее развиты синапсы сенсорно-моторной коры — количество соединений между ними быстро убывает до двух лет.
Потом развивается префронтальная кора, отвечающая, в частности, за социальные функции: на 3–6 лет приходится максимум синапсов (это что-то вроде чернового наброска, который потом «уточняется» и «переписывается»).
Полностью развившийся мозг содержит примерно 86,1±8,1 млрд нейронов, из которых 19 % — нейроны коры. Мозг постоянно перестраивается в зависимости от решаемых им задач: невостребованные нейроны отмирают, неработающие синапсы — стираются.
«Объем человеческого мозга достигает своего максимума к 30–40 годам (именно на этот возраст приходится максимум белого вещества — аксонов <. ..>), а потом <…> аксоны, которые отвечают за межнейронные связи, редуцируются, годам к восьмидесяти мозг сокращается и становится примерно таким же, как у семилетнего ребенка (по массе, но не по структуре)».
..>), а потом <…> аксоны, которые отвечают за межнейронные связи, редуцируются, годам к восьмидесяти мозг сокращается и становится примерно таким же, как у семилетнего ребенка (по массе, но не по структуре)».
Ворох черного шлака
В рассказе Борхеса «Фунес памятливый» говорится о человеке, который помнил абсолютно все, что видел:
«Форму облаков в южной части неба на рассвете <…> он мог сравнить по памяти и с искусным узором кожаного переплета книги, который он видел только раз, и с воспоминаниями об очертаниях брызг, которые поднял гребец в Рио-Негро во время битвы Квебрахо. Эти воспоминания не были простыми: каждый зрительный образ был связан с мускульными ощущениями, тепловыми ощущениями и т. д. Он мог восстановить все свои мечты и фантазии. Два или три раза он воссоздал целый день».
В первой половине прошлого века жил очень похожий на Фунеса человек — Соломон Вениаминович Шерешевский, которого описал в своей работе «Маленькая книжка о большой памяти: Ум мнемониста» нейропсихолог Александр Романович Лурия (книга вышла спустя 25 лет после публикации рассказа Борхеса).
Шерешевский запоминал грандиозные таблицы цифр и букв (50 и более порядков), мог прочитать их по памяти подряд или чередуя как угодно. Он помнил всякую таблицу даже спустя 10–15 лет после каждого «мнемонического» сеанса. Он просто не мог их забыть.
Субъективно Шерешевский воспринимал запоминаемые буквы и цифры так: «они имеют форму. 1 — это острое число, независимо от его графического изображения это что-то законченное, твердое; 2 — более плоское, четырехугольное, беловатое, бывает чуть серое; 3 — отрезок заостренный и вращается; 4 — опять квадратное, тупое, похожее на 2, но более значительное, толстое; 5 — полная законченность в виде конуса, башни, фундаментальное; 6 — это первая за „5”, беловатая; 8 — невинное, голубовато-молочное, похожее на известь».
Как поясняет Владимир Губайловский, это феномен обостренной формы синестезии — когда воспринимаемые звуки окрашены, цвета звучат, тактильные ощущения дают зрительные образы, а увиденное может воздействовать тактильно: например, «уколоть».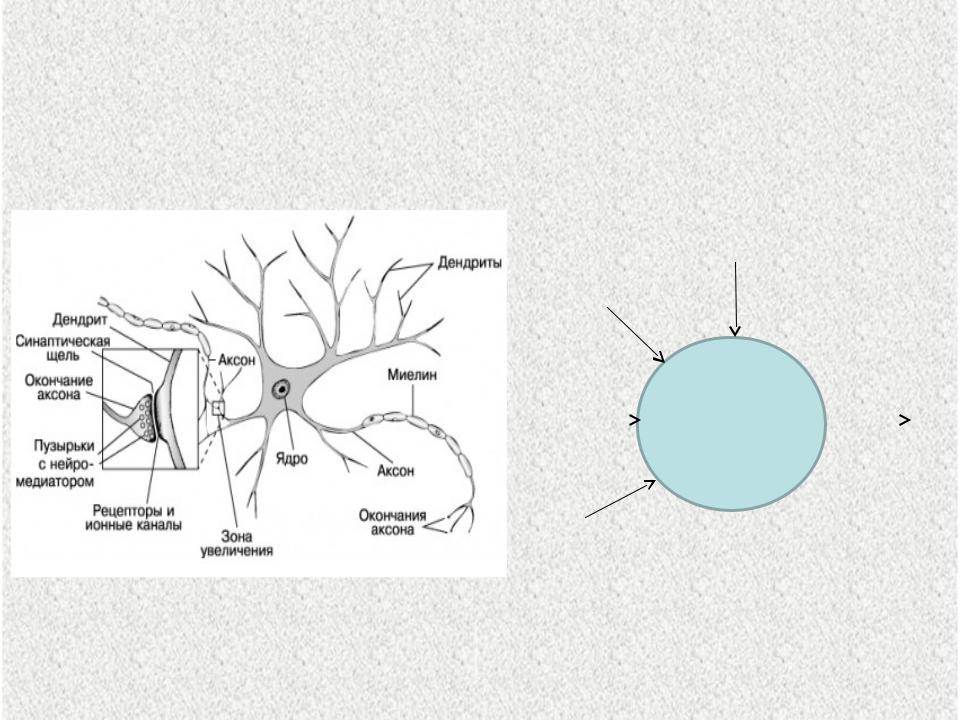 Объясняя, как он запоминает таблицы цифр, букв или слов, Шерешевский говорил, что выстраивает их вдоль длинной улицы, как ставил бы вещи, — а потом «идет мимо», видит их и перечисляет.
Объясняя, как он запоминает таблицы цифр, букв или слов, Шерешевский говорил, что выстраивает их вдоль длинной улицы, как ставил бы вещи, — а потом «идет мимо», видит их и перечисляет.
Соломон Шерешевский вспоминал любопытный случай из своей жизни: «Я подошел к мороженщице, спросил, что у нее есть. „Пломбир”, — она ответила таким голосом, что целый ворох углей, черного шлака выскочил у нее изо рта, — и я уже не мог купить мороженое».
Чтобы научиться забывать, мнемонисту приходилось стараться. Он нашел странное решение: фокусировался на воспоминании и давал себе команду: «Я не хочу это помнить, мне мучительно это помнить», — и это почему-то помогало.
Соломон Шерешевский
Лурия пишет о Шерешевском:
«Так он и оставался неустроенным <…> менявшим десятки профессий, из которых все были „временными”. Он выполнял поручения редактора, он поступал в музыкальную школу, он играл на эстраде, был рационализатором, затем мнемонистом, вспомнил, что он знает древнееврейский и арамейский языки, и стал лечить людей травами, пользуясь этими древними источниками. .. У него была семья: хорошая жена, способный сын, но и это все он воспринимал сквозь дымку. И трудно сказать, что было реальнее — мир воображения, в котором он жил, или мир реальности, в котором он оставался временным гостем…»
.. У него была семья: хорошая жена, способный сын, но и это все он воспринимал сквозь дымку. И трудно сказать, что было реальнее — мир воображения, в котором он жил, или мир реальности, в котором он оставался временным гостем…»
Логично; ведь способность забывать напрямую связана с умением быстрой адаптации в быстро меняющихся средах — в каковых мы и живем.
Нейронет и экзокортекс
Из книги Владимира Губайловского «Искусственный интеллект и мозг человека» мы также узнаем:
- о том, почему человек не рождается, чтобы читать, а «печатное слово приходится тщательно прикручивать болтами»;
- о соединении мозга и компьютерной сети (Нейронета), через которую сигналы одного мозга передаются в другой;
- о попытках создания экзокортекса (цифровой копии или дополнения коры головного мозга).
А еще о том, почему Илон Маск (и не только он) не исключает, что все мы живем внутри компьютерной симуляции.
Аксон — Axon — qaz.wiki
Длинная проекция на нейрон, отводящий сигналы
Аксон (от греческого ἄξων аксона , осей), или нервного волокна (или нервного волокна : см буквам различия ), длинный, тонкий выступ нервной клетки, или нейроны , у позвоночных, которые обычно проводит электрические импульсы , известные как потенциалы действия от тела нервной клетки . Функция аксона — передавать информацию различным нейронам, мышцам и железам. В некоторых сенсорных нейронах ( псевдоуниполярных нейронах ), таких как нейроны прикосновения и тепла, аксоны называются афферентными нервными волокнами, и электрический импульс проходит по ним от периферии к телу клетки и от тела клетки к спинному мозгу вдоль другого. ветвь того же аксона. Дисфункция аксонов является причиной многих наследственных и приобретенных неврологических расстройств, которые могут поражать как периферические, так и центральные нейроны. Нервные волокна классифицируются на три типа — группа А нервные волокна , группа B нервных волокон , и группа С нервных волокон . Группы A и B миелинизированы , а группа C немиелинизирована. Эти группы включают как сенсорные волокна, так и двигательные волокна. Другая классификация группирует только сенсорные волокна как Тип I, Тип II, Тип III и Тип IV.
Функция аксона — передавать информацию различным нейронам, мышцам и железам. В некоторых сенсорных нейронах ( псевдоуниполярных нейронах ), таких как нейроны прикосновения и тепла, аксоны называются афферентными нервными волокнами, и электрический импульс проходит по ним от периферии к телу клетки и от тела клетки к спинному мозгу вдоль другого. ветвь того же аксона. Дисфункция аксонов является причиной многих наследственных и приобретенных неврологических расстройств, которые могут поражать как периферические, так и центральные нейроны. Нервные волокна классифицируются на три типа — группа А нервные волокна , группа B нервных волокон , и группа С нервных волокон . Группы A и B миелинизированы , а группа C немиелинизирована. Эти группы включают как сенсорные волокна, так и двигательные волокна. Другая классификация группирует только сенсорные волокна как Тип I, Тип II, Тип III и Тип IV.
Аксон — это один из двух типов цитоплазматических выступов из тела клетки нейрона; другой тип — дендрит . Аксоны отличаются от дендритов несколькими особенностями, включая форму (дендриты часто сужаются, в то время как аксоны обычно поддерживают постоянный радиус), длину (дендриты ограничены небольшой областью вокруг тела клетки, в то время как аксоны могут быть намного длиннее) и функцию (дендриты получают сигналы, тогда как аксоны передают их). Некоторые типы нейронов не имеют аксона и передают сигналы от своих дендритов. У некоторых видов аксоны могут исходить из дендритов, известных как дендриты, несущие аксоны. Ни у одного нейрона никогда не бывает более одного аксона; однако у беспозвоночных, таких как насекомые или пиявки, аксон иногда состоит из нескольких областей, которые функционируют более или менее независимо друг от друга.
Аксоны отличаются от дендритов несколькими особенностями, включая форму (дендриты часто сужаются, в то время как аксоны обычно поддерживают постоянный радиус), длину (дендриты ограничены небольшой областью вокруг тела клетки, в то время как аксоны могут быть намного длиннее) и функцию (дендриты получают сигналы, тогда как аксоны передают их). Некоторые типы нейронов не имеют аксона и передают сигналы от своих дендритов. У некоторых видов аксоны могут исходить из дендритов, известных как дендриты, несущие аксоны. Ни у одного нейрона никогда не бывает более одного аксона; однако у беспозвоночных, таких как насекомые или пиявки, аксон иногда состоит из нескольких областей, которые функционируют более или менее независимо друг от друга.
Аксоны покрыты мембраной, известной как аксолемма ; цитоплазма аксона называется аксоплазмой . Большинство аксонов разветвляются, в некоторых случаях очень обильно. Концевые ветви аксона называются телодендриями . Набухший конец телодендрона известен как окончание аксона, которое соединяется с дендроном или телом клетки другого нейрона, образуя синаптическое соединение.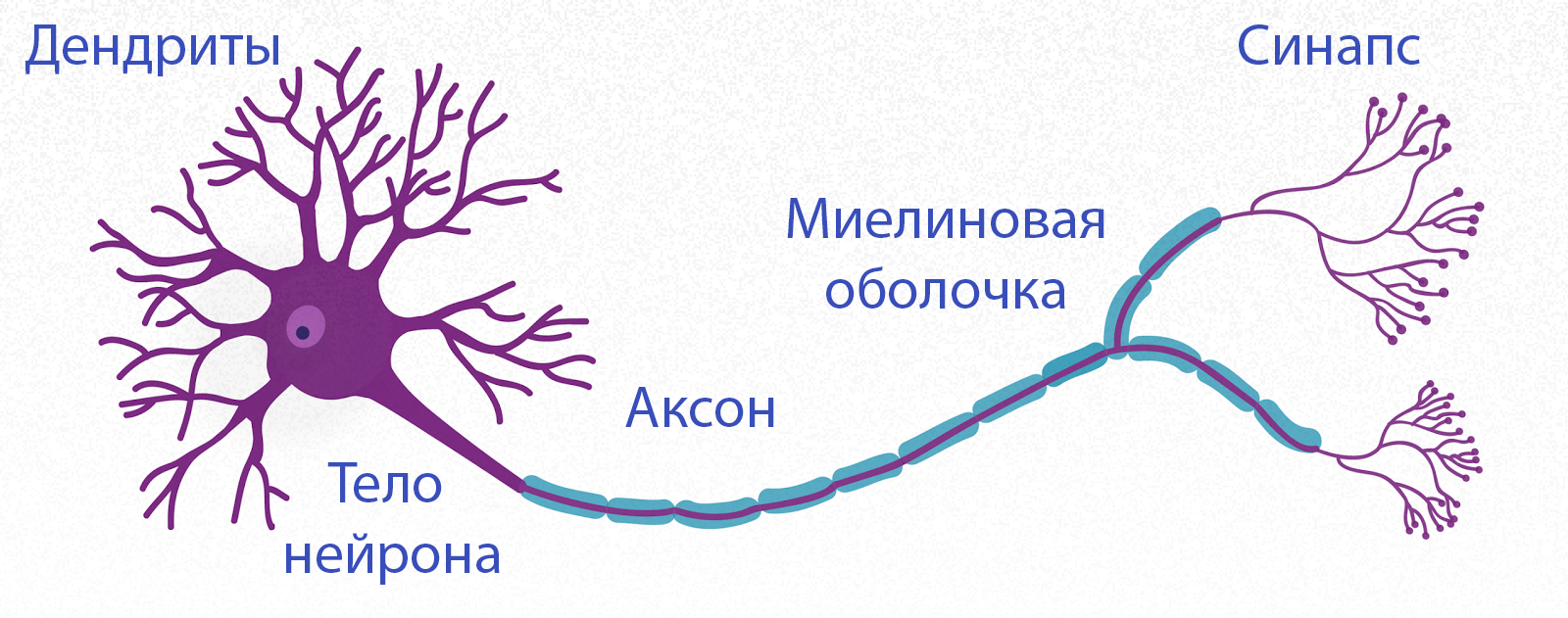 Аксоны контактируют с другими клетками — обычно с другими нейронами, но иногда с клетками мышц или желез — в соединениях, называемых синапсами . В некоторых случаях аксон одного нейрона может образовывать синапс с дендритами того же нейрона, что приводит к аутапсу . В синапсе мембрана аксона плотно прилегает к мембране клетки-мишени, а специальные молекулярные структуры служат для передачи электрических или электрохимических сигналов через промежуток. Некоторые синаптической щели появляются вдоль аксона , как она проходит, они называются Мимоходом ( «попутно») синапсов и может быть в сотни или даже тысячи вдоль одного аксона. Другие синапсы выглядят как терминалы на концах аксональных ветвей.
Аксоны контактируют с другими клетками — обычно с другими нейронами, но иногда с клетками мышц или желез — в соединениях, называемых синапсами . В некоторых случаях аксон одного нейрона может образовывать синапс с дендритами того же нейрона, что приводит к аутапсу . В синапсе мембрана аксона плотно прилегает к мембране клетки-мишени, а специальные молекулярные структуры служат для передачи электрических или электрохимических сигналов через промежуток. Некоторые синаптической щели появляются вдоль аксона , как она проходит, они называются Мимоходом ( «попутно») синапсов и может быть в сотни или даже тысячи вдоль одного аксона. Другие синапсы выглядят как терминалы на концах аксональных ветвей.
Один аксон со всеми его ветвями, взятыми вместе, может иннервировать несколько частей мозга и генерировать тысячи синаптических окончаний. Связка аксонов образует нервный тракт в центральной нервной системе и пучок в периферической нервной системе .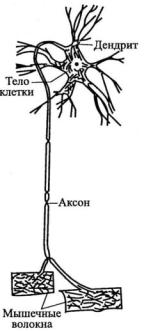 У плацентарных млекопитающих самый большой тракт белого вещества в головном мозге — мозолистое тело , образованное примерно из 200 миллионов аксонов в человеческом мозге .
У плацентарных млекопитающих самый большой тракт белого вещества в головном мозге — мозолистое тело , образованное примерно из 200 миллионов аксонов в человеческом мозге .
Анатомия
Типичный миелинизированный аксон Аксоны являются первичными линиями передачи нервной системы , и как пучки они образуют нервы . Некоторые аксоны могут достигать одного метра и более, а другие — всего лишь один миллиметр. Самые длинные аксоны в человеческом теле — это аксоны седалищного нерва , которые проходят от основания спинного мозга до большого пальца каждой ноги. Диаметр аксонов также варьируется. Большинство отдельных аксонов имеют микроскопический диаметр (обычно около одного микрометра (мкм) в поперечнике). Самые большие аксоны млекопитающих могут достигать в диаметре до 20 мкм. Кальмара гигантский аксон , который специализируется на проведение сигналы очень быстро, близок к 1 миллиметру в диаметре, размер с небольшим карандашом свинца. Количество аксональных телодендрий (разветвленных структур на конце аксона) также может отличаться от одного нервного волокна к другому. Аксоны в центральной нервной системе (ЦНС) обычно имеют множественные телодендрии с множеством синаптических конечных точек. Для сравнения, аксон гранулярных клеток мозжечка характеризуется одним Т-образным узлом ветви, от которого отходят два параллельных волокна . Продуманное ветвление позволяет одновременно передавать сообщения большому количеству целевых нейронов в одной области мозга.
Количество аксональных телодендрий (разветвленных структур на конце аксона) также может отличаться от одного нервного волокна к другому. Аксоны в центральной нервной системе (ЦНС) обычно имеют множественные телодендрии с множеством синаптических конечных точек. Для сравнения, аксон гранулярных клеток мозжечка характеризуется одним Т-образным узлом ветви, от которого отходят два параллельных волокна . Продуманное ветвление позволяет одновременно передавать сообщения большому количеству целевых нейронов в одной области мозга.
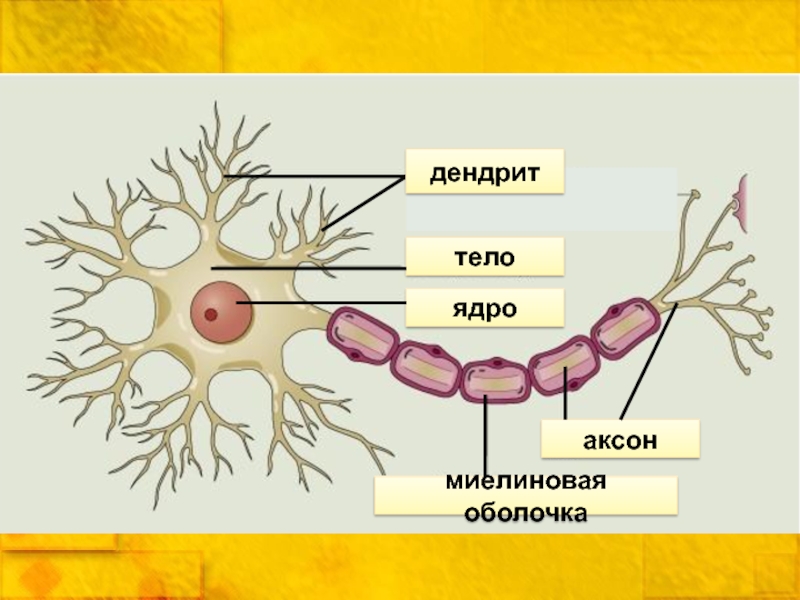
В нервной системе есть два типа аксонов : миелинизированные и немиелинизированные аксоны. Миелин — это слой изолирующего жирового вещества, который образован двумя типами глиальных клеток : шванновскими клетками и олигодендроцитами . В периферической нервной системе шванновские клетки образуют миелиновую оболочку миелинизированного аксона. В центральной нервной системе олигодендроциты образуют изолирующий миелин.![]() Вдоль миелинизированных нервных волокон через равные промежутки времени возникают промежутки в миелиновой оболочке, известные как узлы Ранвье . Миелинизация обеспечивает особенно быстрый способ распространения электрических импульсов, называемый скачкообразной проводимостью .
Вдоль миелинизированных нервных волокон через равные промежутки времени возникают промежутки в миелиновой оболочке, известные как узлы Ранвье . Миелинизация обеспечивает особенно быстрый способ распространения электрических импульсов, называемый скачкообразной проводимостью .
Миелинизированные аксоны нейронов коры образуют основную часть нервной ткани, называемой белым веществом головного мозга. Миелин придает ткани белый цвет в отличие от серого вещества коры головного мозга, которое содержит тела нейрональных клеток. Аналогичное расположение наблюдается в мозжечке . Связки миелинизированных аксонов составляют нервные пути в ЦНС. Там, где эти тракты пересекают среднюю линию мозга и соединяют противоположные области, они называются комиссурами . Самым крупным из них является мозолистое тело, которое соединяет два полушария головного мозга , и у него около 20 миллионов аксонов.
Видно, что структура нейрона состоит из двух отдельных функциональных областей или компартментов — тела клетки вместе с дендритами в качестве одной области и аксональной области в качестве другой.
Аксональная область
Аксональная область или компартмент включает бугорок аксона, начальный сегмент, остальную часть аксона и телодендрии аксона, а также терминалы аксона. Он также включает миелиновую оболочку. В органы Ниссля , которые производят нейронные белки отсутствуют в аксонов регионе. Белки, необходимые для роста аксона и удаления отходов жизнедеятельности, нуждаются в транспортном каркасе. Этот аксональный транспорт обеспечивается в аксоплазме за счет расположения микротрубочек и промежуточных филаментов, известных как нейрофиламенты .
Аксонный бугорок
Деталь, показывающая микротрубочки на бугорке аксона и начальном сегменте. Аксона бугор является областью образована из тела клетки нейрона , как он проходит , чтобы стать аксоном. Он предшествует начальному сегменту. Полученные потенциалы действия, которые суммируются в нейроне, передаются на бугорок аксона для генерации потенциала действия из начального сегмента.
Начальный сегмент
Аксонов начальный сегмент (АИС) представляет собой структурно и функционально отдельный микродоменной аксона. Одна из функций начального сегмента — отделить основную часть аксона от остальной части нейрона; другая функция — помочь инициировать потенциалы действия . Обе эти функции поддерживают полярность нейронных клеток , при которой дендриты (и, в некоторых случаях, сома ) нейрона получают входные сигналы в базальной области, а в апикальной области аксон нейрона обеспечивает выходные сигналы.
Начальный сегмент аксона немиелинизирован и содержит специализированный комплекс белков. Его длина составляет примерно от 20 до 60 мкм, и он функционирует как место инициации потенциала действия. Как положение на аксоне, так и длина AIS могут меняться, показывая степень пластичности, которая может точно настраивать нейрональный выход. Более длинный AIS связан с большей возбудимостью. Пластичность также проявляется в способности AIS изменять свое распределение и поддерживать активность нейронных схем на постоянном уровне.
AIS узкоспециализирован для быстрой передачи нервных импульсов . Это достигается за счет высокой концентрации потенциал-управляемых натриевых каналов в начальном сегменте, где возникает потенциал действия. Ионные каналы сопровождаются большим количеством молекул клеточной адгезии и каркасных белков, которые прикрепляют их к цитоскелету. Взаимодействие с анкирином G важно, так как он является основным организатором в AIS.
Аксональный транспорт
Аксоплазме является эквивалентом цитоплазмы в клетке . В аксоплазме на бугорке аксона образуются микротрубочки . Они расположены по длине аксона в перекрывающихся участках и все направлены в одном направлении — к окончанию аксона. Об этом говорят положительные окончания микротрубочек. Это перекрывающееся расположение обеспечивает маршруты транспортировки различных материалов из тела клетки. Исследования аксоплазмы показали движение многочисленных пузырьков всех размеров, которые можно увидеть вдоль филаментов цитоскелета — микротрубочек и нейрофиламентов , в обоих направлениях между аксоном и его окончаниями и телом клетки.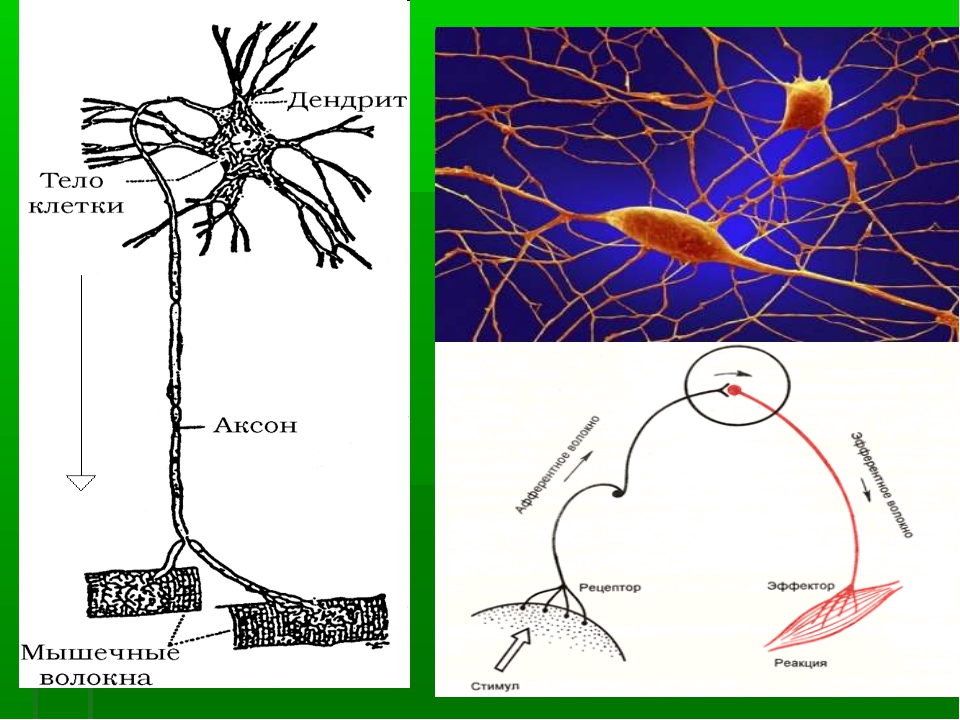
Исходящий антероградный транспорт из тела клетки по аксону переносит митохондрии и мембранные белки, необходимые для роста, к концу аксона. Ретроградный транспорт переносит отходы клетки от терминала аксона к телу клетки. Исходящие и входящие треки используют разные наборы моторных белков . Исходящий транспорт обеспечивается кинезином , а входящий обратный трафик обеспечивается динеином . Динеин направлен на минус-конец. Существует множество форм моторных белков кинезина и динеина, и считается, что каждая из них несет свой груз. Исследования транспорта в аксоне привели к названию кинезина.
Миелинизация
ПЭМ миелинизированного аксона в поперечном сечении.В нервной системе аксоны могут быть миелинизированными или немиелинизированными. Это обеспечение изолирующего слоя, называемого миелиновой оболочкой. Миелиновая мембрана уникальна своим относительно высоким соотношением липидов к белку.
В периферической нервной системе аксоны миелинизируются глиальными клетками, известными как клетки Шванна . В центральной нервной системе миелиновую оболочку составляют глиальные клетки другого типа — олигодендроциты . Клетки Шванна миелинизируют единственный аксон. Олигодендроцит может миелинизировать до 50 аксонов.
В центральной нервной системе миелиновую оболочку составляют глиальные клетки другого типа — олигодендроциты . Клетки Шванна миелинизируют единственный аксон. Олигодендроцит может миелинизировать до 50 аксонов.
Состав миелина у этих двух типов разный. В ЦНС основным белком миелина является протеолипидный белок , а в ПНС — основным белком миелина .
Узлы Ранвье
Узлы Ранвье (также известные как щели миелиновой оболочки ) представляют собой короткие немиелинизированные сегменты миелинизированного аксона , которые периодически встречаются между сегментами миелиновой оболочки. Следовательно, в точке узла Ранвье аксон уменьшается в диаметре. Эти узлы являются областями, где могут быть созданы потенциалы действия. При скачкообразной проводимости электрические токи, возникающие в каждом узле Ранвье, проходят с небольшим затуханием к следующему узлу в линии, где они остаются достаточно сильными, чтобы генерировать другой потенциал действия. Таким образом, в миелинизированном аксоне потенциалы действия эффективно «прыгают» от узла к узлу, минуя миелинизированные участки между ними, в результате чего скорость распространения намного выше, чем может выдержать даже самый быстрый немиелинизированный аксон.
Таким образом, в миелинизированном аксоне потенциалы действия эффективно «прыгают» от узла к узлу, минуя миелинизированные участки между ними, в результате чего скорость распространения намного выше, чем может выдержать даже самый быстрый немиелинизированный аксон.
Терминалы Axon
Аксон может делиться на множество ветвей, называемых телодендриями (греч. Конец дерева). В конце каждого телодендрона находится терминал аксона (также называемый синаптическим бутоном или терминальным бутоном). Терминалы аксонов содержат синаптические пузырьки , в которых хранится нейромедиатор для высвобождения в синапсе . Это делает возможными множественные синаптические связи с другими нейронами. Иногда аксон нейрона может синапсировать с дендритами того же нейрона, что называется аутапсом .
Потенциалы действия
Большинство аксонов несут сигналы в виде потенциалов действия , которые представляют собой дискретные электрохимические импульсы, которые быстро проходят по аксону, начиная с тела клетки и заканчиваясь в точках, где аксон устанавливает синаптический контакт с клетками-мишенями. Определяющей характеристикой потенциала действия является то, что он действует по принципу «все или ничего» — каждый потенциал действия, который генерирует аксон, по существу имеет одинаковый размер и форму. Эта характеристика «все или ничего» позволяет передавать потенциалы действия от одного конца длинного аксона к другому без какого-либо уменьшения размера. Однако есть некоторые типы нейронов с короткими аксонами, которые несут ступенчатые электрохимические сигналы переменной амплитуды.
Определяющей характеристикой потенциала действия является то, что он действует по принципу «все или ничего» — каждый потенциал действия, который генерирует аксон, по существу имеет одинаковый размер и форму. Эта характеристика «все или ничего» позволяет передавать потенциалы действия от одного конца длинного аксона к другому без какого-либо уменьшения размера. Однако есть некоторые типы нейронов с короткими аксонами, которые несут ступенчатые электрохимические сигналы переменной амплитуды.
Когда потенциал действия достигает пресинаптического терминала, он активирует процесс синаптической передачи. Первый шаг — это быстрое открытие каналов для ионов кальция в мембране аксона, позволяя ионам кальция проходить внутрь через мембрану. Результирующее увеличение внутриклеточной концентрации кальция приводит к тому, что синаптические везикулы (крошечные контейнеры, окруженные липидной мембраной), заполненные химическим нейромедиатором, сливаются с мембраной аксона и выводят свое содержимое во внеклеточное пространство.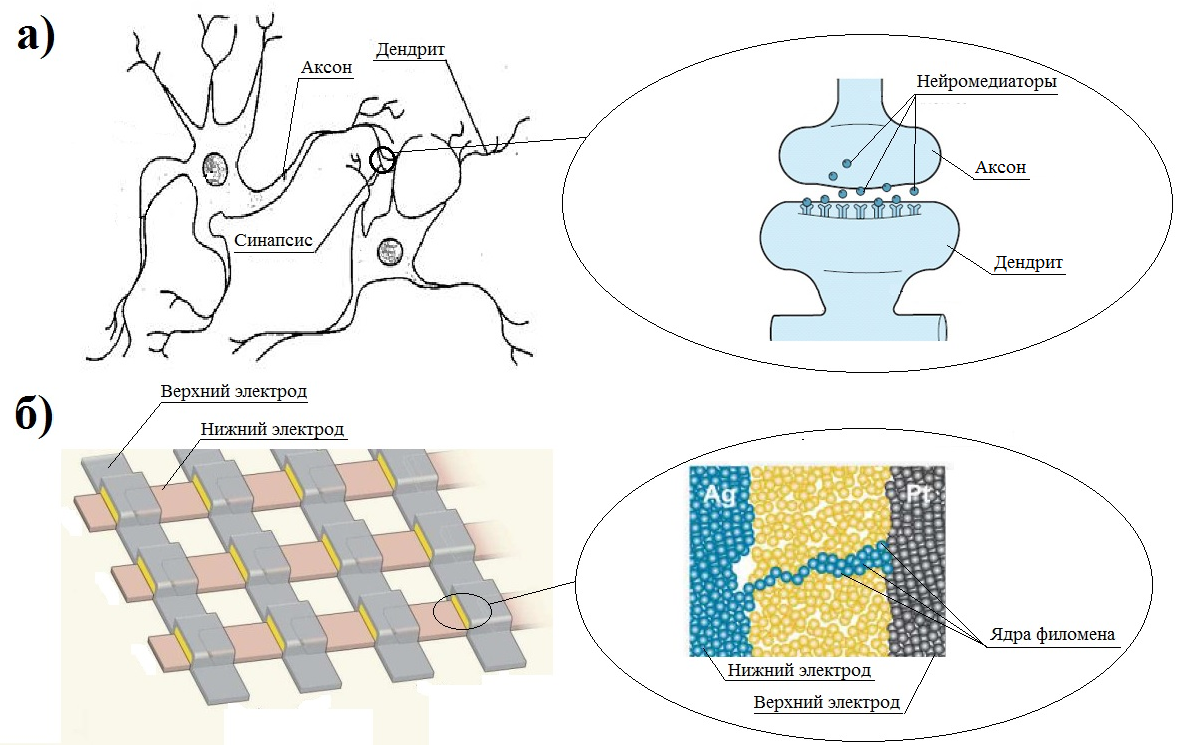 Нейромедиатор высвобождается из пресинаптического нерва посредством экзоцитоза . Затем химический нейротрансмиттер диффундирует к рецепторам, расположенным на мембране клетки-мишени. Нейромедиатор связывается с этими рецепторами и активирует их. В зависимости от типа активируемых рецепторов действие на клетку-мишень может заключаться в возбуждении клетки-мишени, ее подавлении или изменении ее метаболизма каким-либо образом. Вся эта последовательность событий часто происходит менее чем за тысячную долю секунды. После этого внутри пресинаптического терминала новый набор везикул перемещается в положение рядом с мембраной, готовых к высвобождению при достижении следующего потенциала действия. Потенциал действия — это последний электрический шаг в интеграции синаптических сообщений в масштабе нейрона.
Нейромедиатор высвобождается из пресинаптического нерва посредством экзоцитоза . Затем химический нейротрансмиттер диффундирует к рецепторам, расположенным на мембране клетки-мишени. Нейромедиатор связывается с этими рецепторами и активирует их. В зависимости от типа активируемых рецепторов действие на клетку-мишень может заключаться в возбуждении клетки-мишени, ее подавлении или изменении ее метаболизма каким-либо образом. Вся эта последовательность событий часто происходит менее чем за тысячную долю секунды. После этого внутри пресинаптического терминала новый набор везикул перемещается в положение рядом с мембраной, готовых к высвобождению при достижении следующего потенциала действия. Потенциал действия — это последний электрический шаг в интеграции синаптических сообщений в масштабе нейрона.
(B) Средняя и стандартная ошибка времени прохождения пика для интернейронов пирамидных клеток и предполагаемых аксонов;
(C) График разброса отношения сигнала к шуму для отдельных единиц в зависимости от времени пика для аксонов, пирамидных клеток (PYR) и интернейронов (INT).
Внеклеточные записи распространения потенциала действия в аксонах были продемонстрированы у свободно движущихся животных. В то время как внеклеточные соматические потенциалы действия использовались для изучения клеточной активности у свободно перемещающихся животных, таких как клетки места , также можно регистрировать аксональную активность как в белом, так и в сером веществе . Внеклеточные записи распространения потенциала действия аксона отличаются от соматических потенциалов действия по трем причинам: 1. Сигнал имеет более короткую продолжительность пика и спада (~ 150 мкс), чем у пирамидных клеток (~ 500 мкс) или интернейронов (~ 250 мкс). 2. Изменение напряжения трехфазное. 3. Активность, записанная на тетроде, видна только на одном из четырех проводов записи. В записях от свободно движущихся крыс аксональные сигналы были изолированы в трактах белого вещества, включая альвеус и мозолистое тело, а также серое вещество гиппокампа.
Фактически, генерация потенциалов действия in vivo носит последовательный характер, и эти последовательные всплески образуют цифровые коды в нейронах . Хотя предыдущие исследования указывают на аксональное происхождение одиночного спайка, вызванного кратковременными импульсами, физиологические сигналы in vivo запускают инициирование последовательных спайков в телах клеток нейронов.
Хотя предыдущие исследования указывают на аксональное происхождение одиночного спайка, вызванного кратковременными импульсами, физиологические сигналы in vivo запускают инициирование последовательных спайков в телах клеток нейронов.
Помимо распространения потенциалов действия к окончанию аксона, аксон способен усиливать потенциалы действия, что обеспечивает безопасное распространение последовательных потенциалов действия к окончанию аксона. Что касается молекулярных механизмов, потенциал-управляемые натриевые каналы в аксонах обладают более низким порогом и более коротким рефрактерным периодом в ответ на краткосрочные импульсы.
Развитие и рост
Разработка
Развитие аксона до его мишени является одним из шести основных этапов общего развития нервной системы . Исследования, проведенные на культивируемых нейронах гиппокампа, предполагают, что нейроны изначально производят несколько эквивалентных нейритов , но только одному из этих нейритов суждено стать аксоном. Неясно, предшествует ли спецификация аксона удлинению аксона или наоборот, хотя недавние данные указывают на последнее. Если разрезать не полностью развитый аксон, полярность может измениться, и другие нейриты потенциально могут стать аксоном. Это изменение полярности происходит только в том случае, если аксон обрезан как минимум на 10 мкм короче, чем другие нейриты. После того, как разрез будет сделан, самый длинный нейрит станет будущим аксоном, а все остальные нейриты, включая исходный аксон, превратятся в дендриты. Воздействие внешней силы на нейрит, заставляющее его удлиниться, превращает его в аксон. Тем не менее, развитие аксонов достигается за счет сложного взаимодействия между внеклеточной передачей сигналов, внутриклеточной передачей сигналов и динамикой цитоскелета .
Неясно, предшествует ли спецификация аксона удлинению аксона или наоборот, хотя недавние данные указывают на последнее. Если разрезать не полностью развитый аксон, полярность может измениться, и другие нейриты потенциально могут стать аксоном. Это изменение полярности происходит только в том случае, если аксон обрезан как минимум на 10 мкм короче, чем другие нейриты. После того, как разрез будет сделан, самый длинный нейрит станет будущим аксоном, а все остальные нейриты, включая исходный аксон, превратятся в дендриты. Воздействие внешней силы на нейрит, заставляющее его удлиниться, превращает его в аксон. Тем не менее, развитие аксонов достигается за счет сложного взаимодействия между внеклеточной передачей сигналов, внутриклеточной передачей сигналов и динамикой цитоскелета .
Внеклеточные сигналы, которые распространяются через внеклеточный матрикс, окружающий нейроны, играют важную роль в развитии аксонов. Эти сигнальные молекулы включают белки, нейротрофические факторы , внеклеточный матрикс и молекулы адгезии. Нетрин (также известный как UNC-6), секретируемый белок, участвует в образовании аксонов. Когда рецептор нетрина UNC-5 мутирует, несколько нейритов нерегулярно проецируются из нейронов и, наконец, один аксон вытягивается вперед. Нейротрофические факторы — фактор роста нервов (NGF), нейротрофический фактор головного мозга (BDNF) и нейротрофин-3 (NTF3) также участвуют в развитии аксонов и связываются с рецепторами Trk .
Нетрин (также известный как UNC-6), секретируемый белок, участвует в образовании аксонов. Когда рецептор нетрина UNC-5 мутирует, несколько нейритов нерегулярно проецируются из нейронов и, наконец, один аксон вытягивается вперед. Нейротрофические факторы — фактор роста нервов (NGF), нейротрофический фактор головного мозга (BDNF) и нейротрофин-3 (NTF3) также участвуют в развитии аксонов и связываются с рецепторами Trk .
Ганглиозидов -converting фермента плазматической мембраны ганглиозидов сиалидазы (PMGS), который участвует в активации TrkA на кончике neutrites, требуется для удлинения аксонов. PMGS асимметрично распространяется на кончик нейрита, которому суждено стать будущим аксоном.
Внутриклеточная передача сигналов
Во время развития аксона активность PI3K увеличивается на конце предназначенного аксона. Нарушение активности PI3K тормозит развитие аксонов. Активация PI3K приводит к выработке фосфатидилинозитол (3,4,5) -трисфосфата (PtdIns), который может вызывать значительное удлинение нейрита, превращая его в аксон. Таким образом, сверхэкспрессия фосфатаз , дефосфорилирующих PtdIns, приводит к нарушению поляризации.
Таким образом, сверхэкспрессия фосфатаз , дефосфорилирующих PtdIns, приводит к нарушению поляризации.
Цитоскелетная динамика
Нейрит с наименьшим содержанием актиновых филаментов станет аксоном. Концентрация PGMS и содержание f-актина обратно коррелированы; когда PGMS становится обогащенным на кончике нейрита, содержание в нем f-актина существенно снижается. Кроме того, воздействие препаратов, деполимеризующих актин, и токсина B (который инактивирует передачу сигналов Rho ) вызывает образование множественных аксонов. Следовательно, разрыв актиновой сети в конусе роста будет способствовать превращению ее нейрита в аксон.
Рост
Аксон девятидневной мыши с видимым конусом роста Растущие аксоны перемещаются в окружающей среде через конус роста , который находится на кончике аксона. Конус роста имеет широкое пластинчатое расширение, называемое ламеллиподиумом, которое содержит выступы, называемые филоподиями .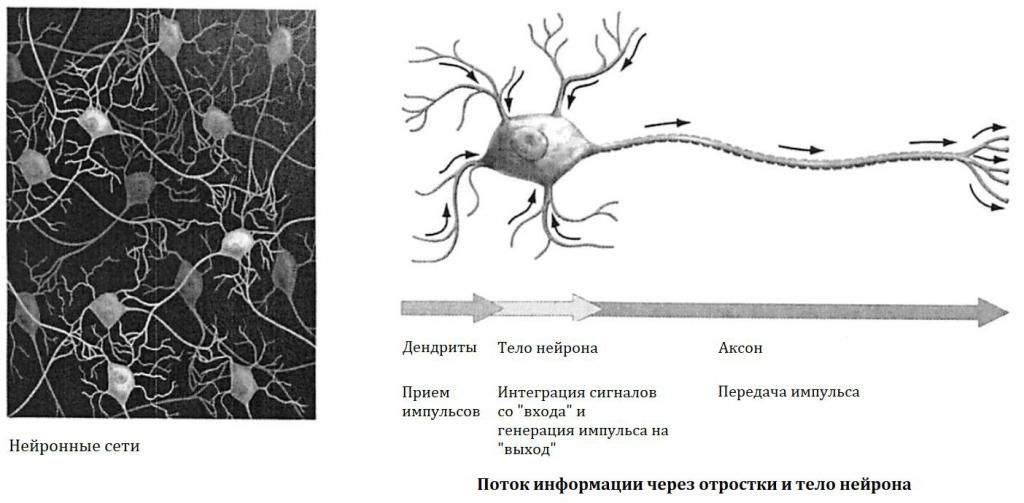 Филоподии — это механизм, с помощью которого весь процесс прикрепляется к поверхностям и исследует окружающую среду. Актин играет важную роль в подвижности этой системы. Среды с высоким уровнем молекул клеточной адгезии (CAM) создают идеальную среду для роста аксонов. Похоже, что это обеспечивает «липкую» поверхность для роста аксонов. Примеры CAM, специфичных для нервных систем, включают N-CAM , TAG-1 — аксональный гликопротеин — и MAG , все из которых являются частью суперсемейства иммуноглобулинов . Другой набор молекул, называемый внеклеточным матриксом, — молекулы адгезии также обеспечивают липкий субстрат для роста аксонов. Примеры этих молекул включают ламинин , фибронектин , тенасцин и перлекан . Некоторые из них поверхностно связаны с клетками и, таким образом, действуют как аттрактанты или репелленты ближнего действия. Другие являются диффундирующими лигандами и, следовательно, могут иметь эффекты длительного действия.
Филоподии — это механизм, с помощью которого весь процесс прикрепляется к поверхностям и исследует окружающую среду. Актин играет важную роль в подвижности этой системы. Среды с высоким уровнем молекул клеточной адгезии (CAM) создают идеальную среду для роста аксонов. Похоже, что это обеспечивает «липкую» поверхность для роста аксонов. Примеры CAM, специфичных для нервных систем, включают N-CAM , TAG-1 — аксональный гликопротеин — и MAG , все из которых являются частью суперсемейства иммуноглобулинов . Другой набор молекул, называемый внеклеточным матриксом, — молекулы адгезии также обеспечивают липкий субстрат для роста аксонов. Примеры этих молекул включают ламинин , фибронектин , тенасцин и перлекан . Некоторые из них поверхностно связаны с клетками и, таким образом, действуют как аттрактанты или репелленты ближнего действия. Другие являются диффундирующими лигандами и, следовательно, могут иметь эффекты длительного действия.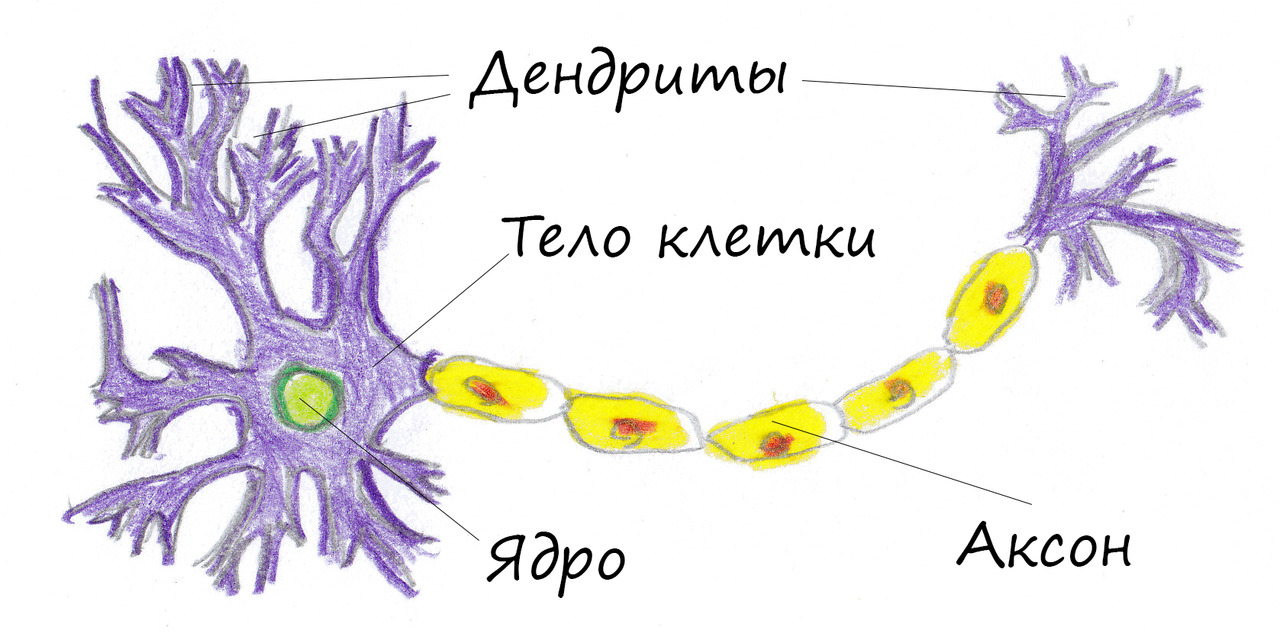
Клетки называемой Вехи клетки помочь в руководстве нейронального роста аксонов. Эти клетки, которые помогают вести аксон , обычно представляют собой другие нейроны, которые иногда незрелы. Когда аксон завершает свой рост в месте его соединения с мишенью, диаметр аксона может увеличиваться до пяти раз, в зависимости от требуемой скорости проводимости .
В ходе исследований также было обнаружено, что если аксоны нейрона были повреждены, то до тех пор, пока сома (тело клетки нейрона ) не повреждено, аксоны будут регенерировать и воссоздавать синаптические связи с нейронами с помощью направляющего столба. клетки . Это также называется нейрорегенерацией .
Nogo-A — это тип компонента, подавляющего рост нейритов, который присутствует в миелиновых мембранах центральной нервной системы (обнаружен в аксоне). Он играет решающую роль в ограничении регенерации аксонов в центральной нервной системе взрослых млекопитающих. В недавних исследованиях, если Nogo-A заблокирован и нейтрализован, можно вызвать регенерацию аксонов на большом расстоянии, что приводит к усилению функционального восстановления у крыс и спинного мозга мыши.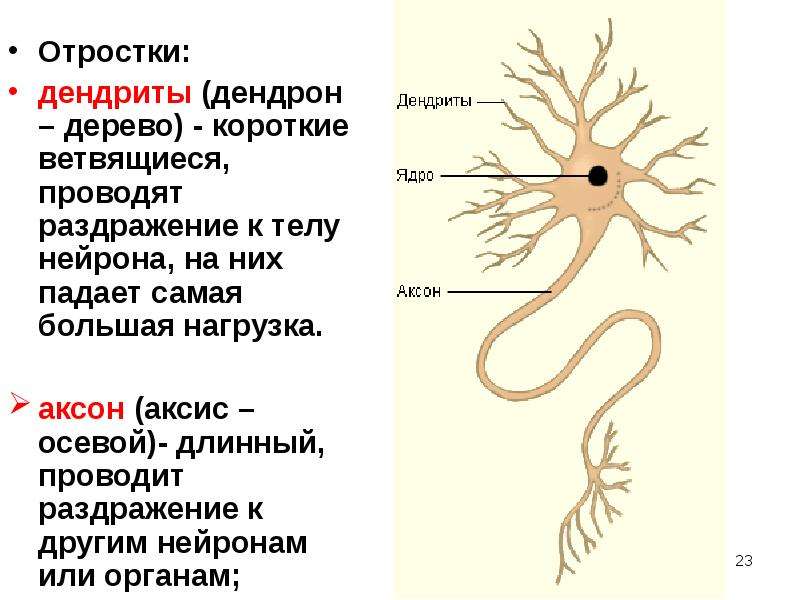 Этого еще предстоит сделать на людях. Недавнее исследование также показало, что макрофаги, активируемые специфическим воспалительным путем, активируемым рецептором Dectin-1 , способны способствовать восстановлению аксонов, однако также вызывают нейротоксичность в нейроне.
Этого еще предстоит сделать на людях. Недавнее исследование также показало, что макрофаги, активируемые специфическим воспалительным путем, активируемым рецептором Dectin-1 , способны способствовать восстановлению аксонов, однако также вызывают нейротоксичность в нейроне.
Регулировка длины
Аксоны в значительной степени различаются по длине от нескольких микрометров до метров у некоторых животных. Это подчеркивает, что должен существовать механизм регулирования длины клетки, позволяющий нейронам ощущать длину своих аксонов и соответственно контролировать их рост. Было обнаружено, что моторные белки играют важную роль в регулировании длины аксонов. Основываясь на этом наблюдении, исследователи разработали четкую модель роста аксонов, описывающую, как моторные белки могут влиять на длину аксона на молекулярном уровне. Эти исследования предполагают, что моторные белки переносят сигнальные молекулы от сомы к конусу роста и наоборот, концентрация которых колеблется во времени с частотой, зависящей от длины.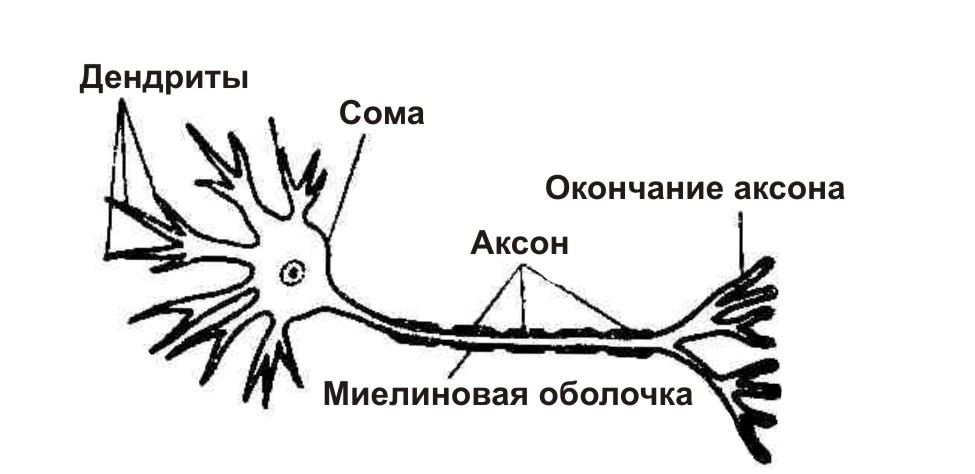
Классификация
Аксоны нейронов периферической нервной системы человека можно классифицировать по их физическим характеристикам и свойствам проводимости сигнала. Было известно, что аксоны имеют разную толщину (от 0,1 до 20 мкм), и считалось, что эти различия связаны со скоростью, с которой потенциал действия может перемещаться по аксону — его проводимостью . Эрлангер и Гассер доказали эту гипотезу и идентифицировали несколько типов нервных волокон, установив связь между диаметром аксона и скоростью проводимости по нему . Они опубликовали свои результаты в 1941 году, дав первую классификацию аксонов.
Аксоны подразделяются на две системы. Первый, введенный Эрлангером и Гассером, сгруппировал волокна в три основные группы, используя буквы A, B и C. Эти группы, группа A , группа B и группа C, включают как сенсорные волокна ( афференты ), так и двигательные волокна. ( эфференты ). Первая группа A была разделена на альфа, бета, гамма и дельта волокна — Aα, Aβ, Aγ и Aδ. Моторными нейронами различных моторных волокон были нижние моторные нейроны — альфа-мотонейрон , бета-мотонейрон и гамма-мотонейрон, имеющие нервные волокна Aα, Aβ и Aγ соответственно.
Моторными нейронами различных моторных волокон были нижние моторные нейроны — альфа-мотонейрон , бета-мотонейрон и гамма-мотонейрон, имеющие нервные волокна Aα, Aβ и Aγ соответственно.
Позже другие исследователи обнаружили две группы волокон Aa, которые были сенсорными волокнами. Затем они были введены в систему, которая включала только сенсорные волокна (хотя некоторые из них были смешанными нервами, а также двигательными волокнами). В этой системе сенсорные группы называются Типами и используются римские цифры: Тип Ia, Тип Ib, Тип II, Тип III и Тип IV.
Мотор
Нижние мотонейроны имеют два типа волокон:
Сенсорный
Разные сенсорные рецепторы иннервируют разные типы нервных волокон. Проприоцепторы иннервируются сенсорными волокнами типа Ia, Ib и II, механорецепторы — сенсорными волокнами типа II и III, а ноцицепторы и терморецепторы — сенсорными волокнами типа III и IV.
Автономный
Вегетативная нервная система имеет два вида периферических волокон:
Клиническое значение
По степени тяжести повреждение нерва может быть описано как нейрапраксия , аксонотмезис или нейротмезис .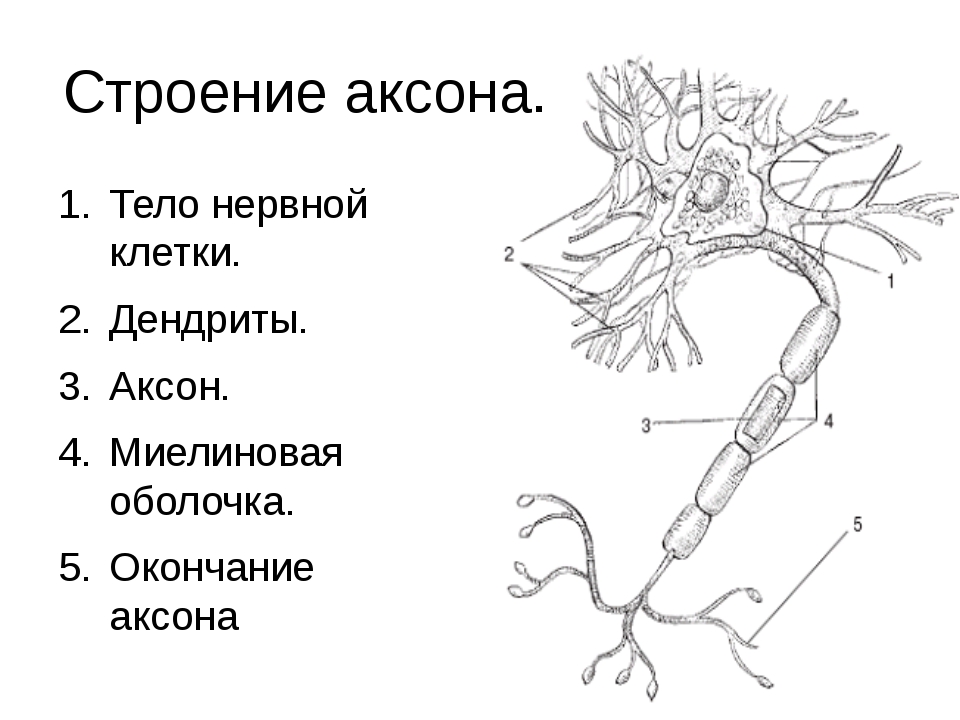 Сотрясение мозга считается легкой формой диффузного повреждения аксонов . Поражение аксонов также может вызвать центральный хроматолиз . Дисфункция аксонов нервной системы является одной из основных причин многих наследственных неврологических расстройств , поражающих как периферические, так и центральные нейроны.
Сотрясение мозга считается легкой формой диффузного повреждения аксонов . Поражение аксонов также может вызвать центральный хроматолиз . Дисфункция аксонов нервной системы является одной из основных причин многих наследственных неврологических расстройств , поражающих как периферические, так и центральные нейроны.
Когда аксон раздавлен, активный процесс дегенерации аксона происходит в части аксона, наиболее удаленной от тела клетки. Эта дегенерация происходит быстро после травмы, когда часть аксона блокируется мембранами и разрушается макрофагами. Это известно как валлеровское вырождение . Отмирание аксона также может иметь место при многих нейродегенеративных заболеваниях, особенно при нарушении транспорта аксонов, это известно как дегенерация, подобная валлеровской. Исследования показывают, что дегенерация происходит в результате того, что аксональный белок NMNAT2 не может достичь всего аксона.
Демиелинизация аксонов вызывает множество неврологических симптомов, обнаруживаемых при заболевании рассеянным склерозом .
Дисмиелинизация — это аномальное образование миелиновой оболочки. Это связано с несколькими лейкодистрофиями , а также с шизофренией .
Тяжелая черепно-мозговая травма может привести к обширным поражениям нервных путей, повреждая аксоны в состоянии, известном как диффузное повреждение аксонов . Это может привести к стойкому вегетативному состоянию . В исследованиях на крысах было показано, что повреждение аксонов в результате однократного легкого черепно-мозгового повреждения может привести к дальнейшему повреждению после повторных легких черепно-мозговых травм.
Канал наведения нервов — это искусственное средство направления роста аксонов, обеспечивающее нейрорегенерацию , и одно из многих методов лечения, используемых при различных видах повреждения нервов .
История
Немецкому анатому Отто Фридриху Карлу Дейтерсу обычно приписывают открытие аксона, отличив его от дендритов. Швейцарский Рюдольф Альберт фон Келликер и немец Роберт Ремак были первыми, кто идентифицировал и охарактеризовал начальный сегмент аксона. Кёлликер назвал аксон в 1896 году. Луи-Антуан Ранвье был первым, кто описал промежутки или узлы, обнаруженные на аксонах, и из-за этого вклада эти аксональные особенности теперь обычно называют узлами Ранвье . Сантьяго Рамон-и-Кахаль , испанский анатом, предположил, что аксоны были выходными компонентами нейронов, описывая их функции. Джозеф Эрлангер и Герберт Гассер ранее разработали систему классификации периферических нервных волокон, основанную на скорости проводимости аксонов, миелинизации , размере волокна и т. Д. Алан Ходжкин и Эндрю Хаксли также использовали гигантский аксон кальмара (1939), и к 1952 году они получили полную количественную оценку. описание ионной основы потенциала действия , приведшее к формулировке модели Ходжкина – Хаксли . В 1963 году Ходжкин и Хаксли были совместно удостоены Нобелевской премии за эту работу. Формулы, описывающие аксональную проводимость, были распространены на позвоночных в уравнениях Франкенхейзера-Хаксли.
Кёлликер назвал аксон в 1896 году. Луи-Антуан Ранвье был первым, кто описал промежутки или узлы, обнаруженные на аксонах, и из-за этого вклада эти аксональные особенности теперь обычно называют узлами Ранвье . Сантьяго Рамон-и-Кахаль , испанский анатом, предположил, что аксоны были выходными компонентами нейронов, описывая их функции. Джозеф Эрлангер и Герберт Гассер ранее разработали систему классификации периферических нервных волокон, основанную на скорости проводимости аксонов, миелинизации , размере волокна и т. Д. Алан Ходжкин и Эндрю Хаксли также использовали гигантский аксон кальмара (1939), и к 1952 году они получили полную количественную оценку. описание ионной основы потенциала действия , приведшее к формулировке модели Ходжкина – Хаксли . В 1963 году Ходжкин и Хаксли были совместно удостоены Нобелевской премии за эту работу. Формулы, описывающие аксональную проводимость, были распространены на позвоночных в уравнениях Франкенхейзера-Хаксли. Понимание биохимической основы распространения потенциала действия продвинулось дальше и включает множество деталей об отдельных ионных каналах .
Понимание биохимической основы распространения потенциала действия продвинулось дальше и включает множество деталей об отдельных ионных каналах .
Другие животные
Аксоны у беспозвоночных широко изучены. Longfin Прибрежная кальмара , часто используется в качестве модельного организма имеет самый длинный известный аксон. У гигантского кальмара самый большой из известных аксонов . Его размер колеблется от половины (обычно) до одного миллиметра в диаметре и используется для управления его реактивной двигательной установкой. Самая быстрая зарегистрированная скорость проводимости 210 м / с обнаружена в заключенных в оболочку аксонах некоторых пелагических креветок Penaeid, и обычный диапазон составляет от 90 до 200 м / с ( ср. 100–120 м / с для самого быстрого аксона миелинизированного позвоночного).
В других случаях, как показали исследования на крысах, аксон происходит от дендрита; такие аксоны, как говорят, имеют «дендритное происхождение».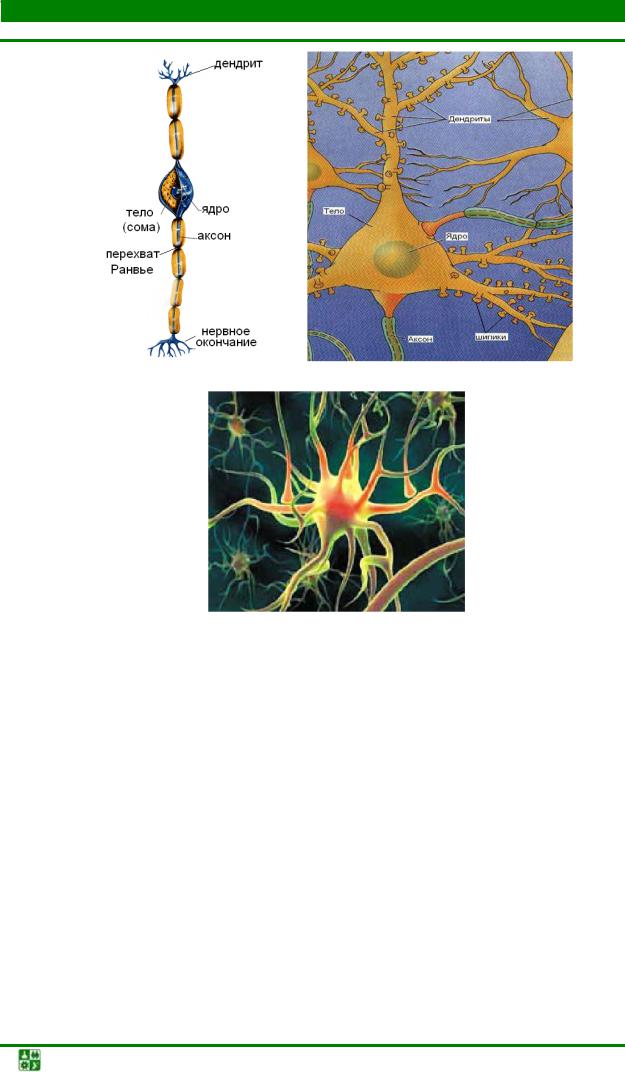 Некоторые аксоны с дендритным происхождением аналогичным образом имеют «проксимальный» начальный сегмент, который начинается непосредственно в источнике аксона, в то время как другие имеют «дистальный» начальный сегмент, заметно отделенный от источника аксона. У многих видов некоторые из нейронов имеют аксоны, которые исходят из дендрита, а не из тела клетки, и они известны как дендриты, несущие аксоны. Во многих случаях аксон берет свое начало от бугорка аксона на соме; такие аксоны, как говорят, имеют «соматическое происхождение». Некоторые аксоны соматического происхождения имеют «проксимальный» начальный сегмент, прилегающий к бугорку аксона, в то время как другие имеют «дистальный» начальный сегмент, отделенный от сомы протяженным бугорком аксона.
Некоторые аксоны с дендритным происхождением аналогичным образом имеют «проксимальный» начальный сегмент, который начинается непосредственно в источнике аксона, в то время как другие имеют «дистальный» начальный сегмент, заметно отделенный от источника аксона. У многих видов некоторые из нейронов имеют аксоны, которые исходят из дендрита, а не из тела клетки, и они известны как дендриты, несущие аксоны. Во многих случаях аксон берет свое начало от бугорка аксона на соме; такие аксоны, как говорят, имеют «соматическое происхождение». Некоторые аксоны соматического происхождения имеют «проксимальный» начальный сегмент, прилегающий к бугорку аксона, в то время как другие имеют «дистальный» начальный сегмент, отделенный от сомы протяженным бугорком аксона.
Смотрите также
Рекомендации
внешняя ссылка
Логика сознания. Часть 2. Дендритные волны
В предыдущей части мы показали, что в клеточном автомате могут возникать волны, имеющие специфический внутренний узор. Такие волны могут запускаться из любого места клеточного автомата и распространяться по всему пространству клеток автомата, перенося информацию. Соблазнительно предположить, что реальный мозг может использовать схожие принципы. Чтобы понять возможность аналогии, немного разберемся с тем, как работают нейроны реального мозга.
Такие волны могут запускаться из любого места клеточного автомата и распространяться по всему пространству клеток автомата, перенося информацию. Соблазнительно предположить, что реальный мозг может использовать схожие принципы. Чтобы понять возможность аналогии, немного разберемся с тем, как работают нейроны реального мозга.Головной мозг состоит из серого и белого вещества. Серое вещество — это мозговые структуры, состоящие из нейронов и глиальных клеток. Белое вещество — это аксоны нейронов, они же нервные волокна. Эти волокна образуют связи одних структур мозга с другими.
Распределение белого и серого вещества на фронтальном срезе мозга
Структуры, которые находятся ближе к центру мозга, принято относить к древнему мозгу. Древний мозг роднит нас с животными и реализует механизмы, отточенные эволюцией и более-менее общие для многих живых существ. Основной объем серого вещества человека приходится на кору. Кора — это слой серого вещества толщиной от 1.3 до 4.5 мм, составляющий наружную поверхность мозга.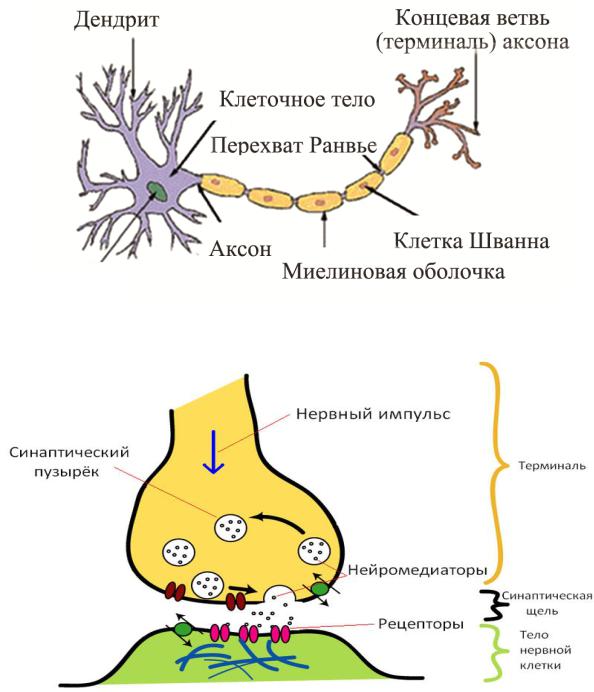 Есть множество доводов в пользу того, что кора, в отличие от древнего мозга, реализует не генетически заложенные алгоритмы, а способна к обучению и самоорганизации.
Есть множество доводов в пользу того, что кора, в отличие от древнего мозга, реализует не генетически заложенные алгоритмы, а способна к обучению и самоорганизации.
Основные клетки мозга — это нейроны и глиальные клетки. Похоже, что и те, и другие играют существенную роль в информационных процессах. Для упрощения повествования, пока, мы будем говорить только о нейронах. Разговор же о глиальных клетках отложим на некоторое время.
Нейроны бывают разных типов. Самые массовые нейроны коры — это пирамидальные нейроны. На их долю в коре приходится 75% от всех нейронов. На рисунке ниже показаны именно они.
Структура пирамидального нейрона, черный — дендрит, серый — аксон, линейка — 0.1 мм (Braitenberg, 1978)
У большинства нейронов есть тело, дендритное дерево и аксон. И аксон, и дендрит сильно ветвятся и образуют запутанную структуру со множеством переплетений с дендритами и аксонами других нейронов. Составить общее представление о сложности и запутанности переплетений аксонов и дендритов можно, например, по видео.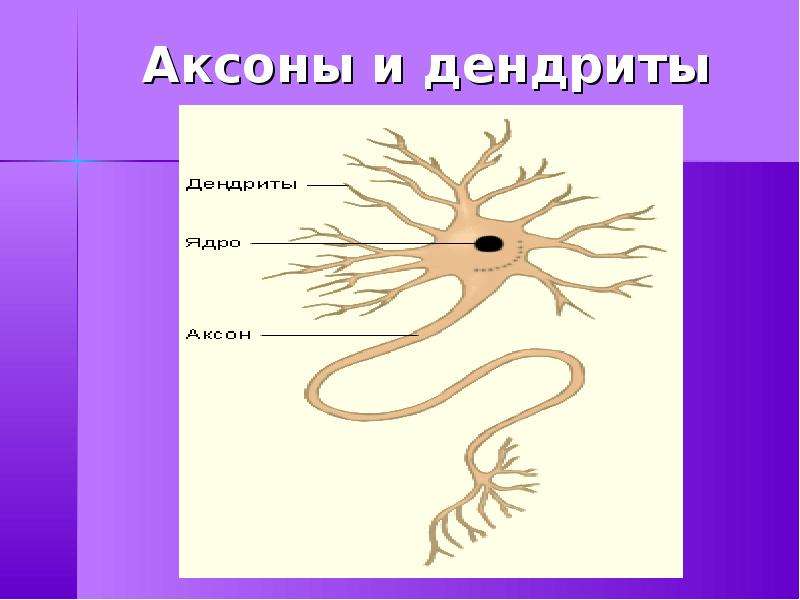
Общую конфигурацию нейрона хорошо представить по классической картинке из википедии.
Тела нейронов, их дендриты и аксоны, окружающие глиальные клетки все они тесно упакованы между собой, оставляя свободными только узкие щели. Эти щели заполнены сложным раствором существенную часть которого составляют электролиты (в основном, ионы калия, кальция, натрия и хлора). Плотность упаковки можно увидеть и оценить на реконструкции небольшого объема коры, приведенной ниже.
(connectomethebook.com)
Поверхность нейрона называется мембраной. Задача мембраны оградить внутреннюю среду нейрона от внешней. При этом в мембрану встроено огромное количество белков. Некоторые из них пронзают мембрану насквозь и контактируют, как с внешней, так и внутренней средой нейрона. Такие белки называют трансмембранными (рисунок ниже).
Трансмембранные белки
Трансмембранные белки выполняют разные функции. Если белки занимаются транспортировкой ионов в клетку или из клетки и делают это постоянно, то это ионные насосы. Чтобы осуществлять транспорт ионов белки создают ионные каналы. Ионные каналы могут иметь внешнее управление, то есть открываться и закрываться при определенных сигналах. Если канал управляется мембранным потенциалом, то говорят о потенциал-зависимых ионных каналах.
Если белок реагирует на какое-либо вещество снаружи нейрона и передает эту реакцию каким-либо способом внутрь нейрона, то такие белки называют рецепторами. Вещество, которые воздействуют на конкретный рецептор называют его лигандом. Если рецептор имеет ионный канал, который открывается под воздействием лиганда, то такой рецептор называется ионотропным. Если у рецептора нет ионного канала и он воздействует на состояние нейрона окольным путем, то это метаботропный рецептор.
Рецепторы и другие белки не сосредоточены где-то в одном месте, а распределены по всей поверхности нейрона.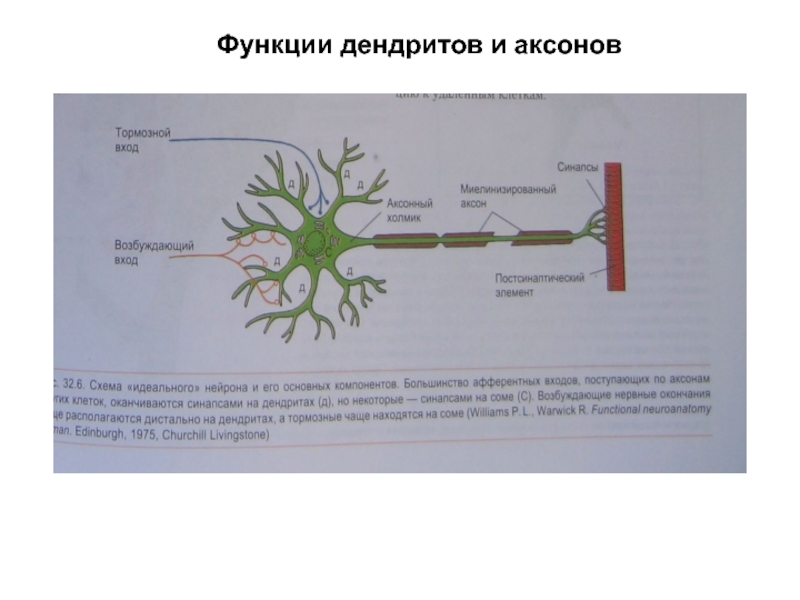 Средний нейрон коры имеет порядка 10 000 синапсов, распределенных по его дендриту и телу. На каждый синапс приходится несколько сотен рецепторов.
Средний нейрон коры имеет порядка 10 000 синапсов, распределенных по его дендриту и телу. На каждый синапс приходится несколько сотен рецепторов.
В состоянии покоя между внутренней и внешней средой нейрона существует разность потенциалов — мембранный потенциал, составляющий около 70 милливольт. Он образуется за счет работы белковых молекул, работающих как ионные насосы. В зависимости от своего типа ионные насосы меняют соотношение определенных ионов снаружи и внутри клетки. Насосы первого типа меняют соотношение ионов калия и натрия, второго типа — выводят из клетки ионы кальция, третьего типа — транспортируют наружу протоны. В результате мембрана приобретает поляризацию, при которой отрицательный заряд накапливается внутри клетки, а положительный снаружи.
Места контактов аксонов с дендритами или телами нейронов называются синапсами. Основной тип синапсов — это химический синапс.
Когда нервный импульс по аксону поступает в синапс, он высвобождает из специальных пузырьков молекулы нейромедиаторов, характерные для этого синапса.
 На мембране нейрона, получающего сигнал, есть белковые молекулы — рецепторы. Рецепторы взаимодействуют с нейромедиаторами. Рецепторы, расположенные в синаптической щели, являются ионотропными, то есть они же являются ионными каналами, способными перемещать ионы. Нейромедиаторы так воздействуют на рецепторы, что их ионные каналы открываются. Соответственно, мембрана либо деполяризуется, либо гиперполяризуется — в зависимости от того, какие каналы затронуты и, соответственно, какого типа этот синапс. В возбуждающих синапсах открываются каналы, преимущественно пропускающие катионы внутрь клетки, — мембрана деполяризуется. В тормозных синапсах открываются каналы, выводящие катионы из клетки, что приводит к гиперполяризации мембраны.
На мембране нейрона, получающего сигнал, есть белковые молекулы — рецепторы. Рецепторы взаимодействуют с нейромедиаторами. Рецепторы, расположенные в синаптической щели, являются ионотропными, то есть они же являются ионными каналами, способными перемещать ионы. Нейромедиаторы так воздействуют на рецепторы, что их ионные каналы открываются. Соответственно, мембрана либо деполяризуется, либо гиперполяризуется — в зависимости от того, какие каналы затронуты и, соответственно, какого типа этот синапс. В возбуждающих синапсах открываются каналы, преимущественно пропускающие катионы внутрь клетки, — мембрана деполяризуется. В тормозных синапсах открываются каналы, выводящие катионы из клетки, что приводит к гиперполяризации мембраны.Поляризация мембраны нейрона выглядит, как скопление ионов в относительной близости от мембраны (рисунок ниже).
Когда открываются ионные каналы рецепторов и начинается обмен ионами с внешней средой, то только в том месте поверхности нейрона где расположены рецепторы и происходит изменение поляризации.
 Маленький участок мембраны оказывается заряжен иначе чем все его окружение.
Маленький участок мембраны оказывается заряжен иначе чем все его окружение.Если сработали возбуждающие рецепторы, то участок в соответствующем месте окажется деполяризован, то есть его потенциал будет выше чем в среднем по мембране нейрона. Если эта деполяризация достигнет критического значения, то возникнет спайк, который начнет распространяться по мембране.
За возникновение и распространение спайка отвечают потенциал-зависимые ионные каналы. Они управляются не нейромедиаторами, а величиной мембранного потенциала. Например, для аксона их работа выглядит следующим образом.
Когда потенциал поднимается до критического значения открываются натриевые каналы, которые начинают гнать положительно заряженные ионы натрия внутрь клетки. Соответственно, потенциал в этом месте лавинообразно возрастает. Но в определенный момент включаются калиевые потенциал-зависимые каналы. Они начинают выводить положительно заряженные ионы калия из клетки наружу, понижая тем самым мембранный потенциал. В результате происходит кратковременный локальный всплеск потенциала. Затем наступает рефрактерный период, когда это место оказывается нечувствительно к изменению потенциала. Но сильный всплеск в одном месте приводит к менее сильному подъему потенциала в соседних с ним местах. Там происходит превышение порогового значения и рождается свой всплеск. В результате потенциал действия или иначе спайк распространяется по всей длине аксона.
В результате происходит кратковременный локальный всплеск потенциала. Затем наступает рефрактерный период, когда это место оказывается нечувствительно к изменению потенциала. Но сильный всплеск в одном месте приводит к менее сильному подъему потенциала в соседних с ним местах. Там происходит превышение порогового значения и рождается свой всплеск. В результате потенциал действия или иначе спайк распространяется по всей длине аксона.
Распространение спайка — самовоспроизводящийся процесс. Спайк, возникнув в одном месте, заставляет соседние с ним места сгенерировать свой спайк и так далее. Это, кстати, напоминает простейший клеточный автомат, наподобие того, что мы описывали в предыдущей части. Возникнув в одном месте спайк распространяется во все стороны от этого места. Но если спайк не возник в этом месте, а пришел со стороны, то за счет того, что есть рефракторный период, он может распространить только туда, где его еще не было.
В аксонах покрытых миелиновой оболочкой потенциал действия распространяется несколько иначе. Миелиновая оболочка не дает распространяться спайку, но, зато, хорошо изолирует нервное волокно. В результате, внутри изолированной части происходит передача электрического сигнала, как по кабелю. Затем в неизолированном участке, перехвате, происходит генерация нового спайка. За счет таких «прыжков» скорость передачи нервного импульса в толстых, покрытых миелином, аксонах оказывается много выше, чем в нервных волокнах без такой оболочки.
Миелиновая оболочка не дает распространяться спайку, но, зато, хорошо изолирует нервное волокно. В результате, внутри изолированной части происходит передача электрического сигнала, как по кабелю. Затем в неизолированном участке, перехвате, происходит генерация нового спайка. За счет таких «прыжков» скорость передачи нервного импульса в толстых, покрытых миелином, аксонах оказывается много выше, чем в нервных волокнах без такой оболочки.
В дендритах также есть потенциал-зависимые ионные каналы и по ним, как и по аксону может распространяться потенциал действия. Аксонные спайки имеют амплитуду порядка 100 мВ, амплитуда дендритных спайков несколько ниже. Аксонные спайки возникают на теле нейрона в дендритном холмике. Оттуда они распространяются далее по аксону. Возбуждение в аксонном холмике может распространяться и на дендрит, в этом случае возникают дендритные спайки, которые являются сигналом обратного распространения по отношению к аксонному потенциалу действия.
Дендритные спайки могут возникать и непосредственно в дендрите.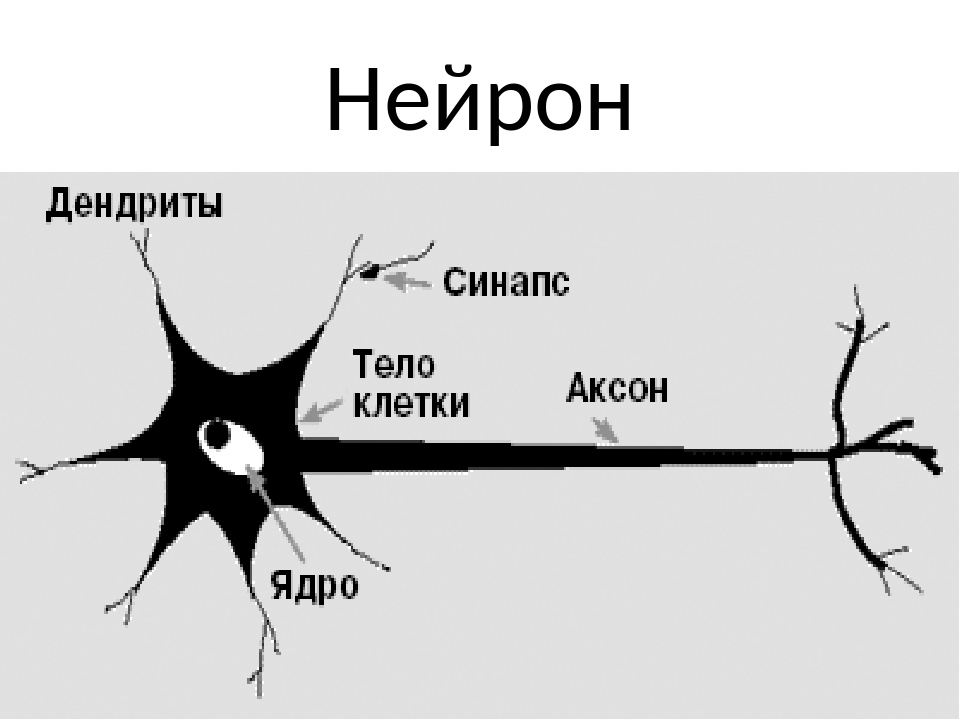 Для этого требуется, чтобы в течении короткого временного интервала (порядка 3–10 мс) на небольшом участке дендрита случилось достаточно много синаптических возбуждений. Например, если длина участка 100 мкм и временной интервал 3 мс, то потребуется, чтобы сработало около 50 синапсов, чтобы появился дендритный спайк. Стоит учесть, что на таком участке располагается порядка 200 синапсов. Добиться синхронной активности четверти всех синапсов можно при искусственном возбуждении «в пробирке». Трудно сказать возможно ли такое в живой ткани.
Для этого требуется, чтобы в течении короткого временного интервала (порядка 3–10 мс) на небольшом участке дендрита случилось достаточно много синаптических возбуждений. Например, если длина участка 100 мкм и временной интервал 3 мс, то потребуется, чтобы сработало около 50 синапсов, чтобы появился дендритный спайк. Стоит учесть, что на таком участке располагается порядка 200 синапсов. Добиться синхронной активности четверти всех синапсов можно при искусственном возбуждении «в пробирке». Трудно сказать возможно ли такое в живой ткани.
Распространение спайков не единственный механизм передачи информации, свойственный дендриту. Показано, что дендриты обладают кабельными свойствами. Ветку дендрита можно сопоставить с кабелем, у которого есть внутреннее сопротивление, сопротивление утечки и поверхностная емкость. Хотя сопротивление дендрита и очень велико, а утечки существенны, тем не менее, токи, которые возникают от возбуждающих постсинаптических потенциалов могут оказывать существенное влияние на общее состояние нейрона.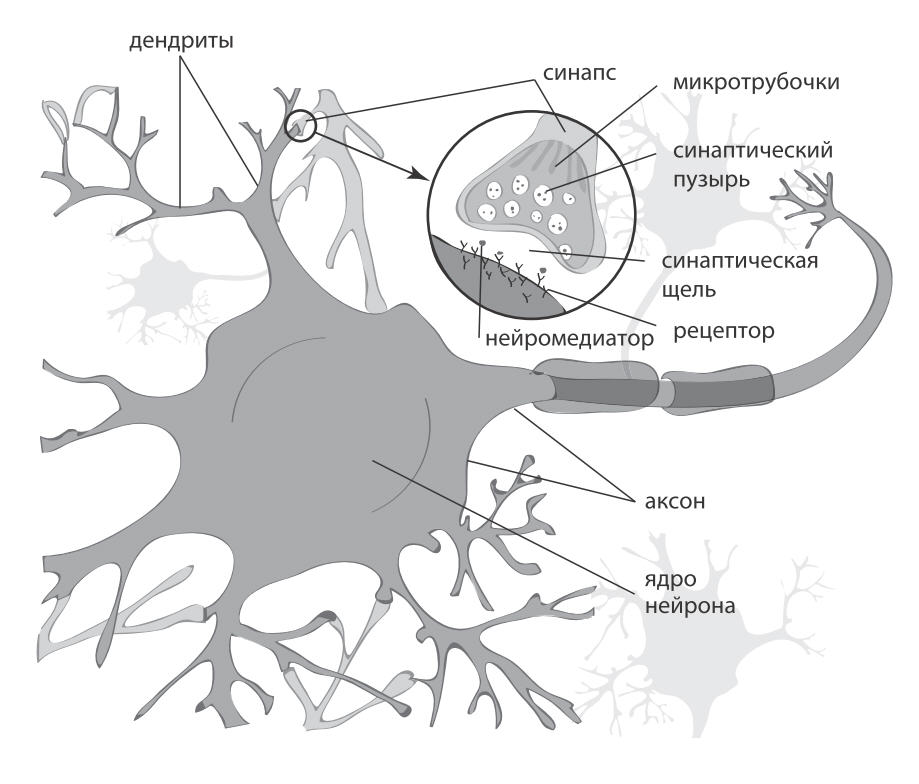 Можно предположить, что особенно роль этих токов велика на коротких расстояниях, например, в пределах одной веточки дендритного дерева.
Можно предположить, что особенно роль этих токов велика на коротких расстояниях, например, в пределах одной веточки дендритного дерева.
И аксон, и дендритные ветки представляют собой тонкие трубочки. Распространение спайка по ним — это перемещение кольцевой области деполяризации. Но спайки — это достаточно энергетически затратные явления. Кроме них есть более слабые, но зато и более массовые сигналы. Нейробиологи, порою, говорят, что нейроны, в основном, не кричат друг другу (подразумевая спайки), а шепчут.
Вернемся к химическому синапсу. Нервный импульс, распространяясь по аксону, доходит до многочисленных аксонных терминалей. Большая часть терминалей образует контакты с дендритами. Это и есть химические синапсы. Дойдя до терминали, спайк вызывает массовый выброс нейромедиаторов в синаптическую щель. Нейромедиаторы упакованы в специальные пузырьки (везикулы). В одной везикуле содержится несколько тысяч молекул.
Приход спайка вызывает массовый выброс нейромедиаторов, состоящий из десятка пузырьков.
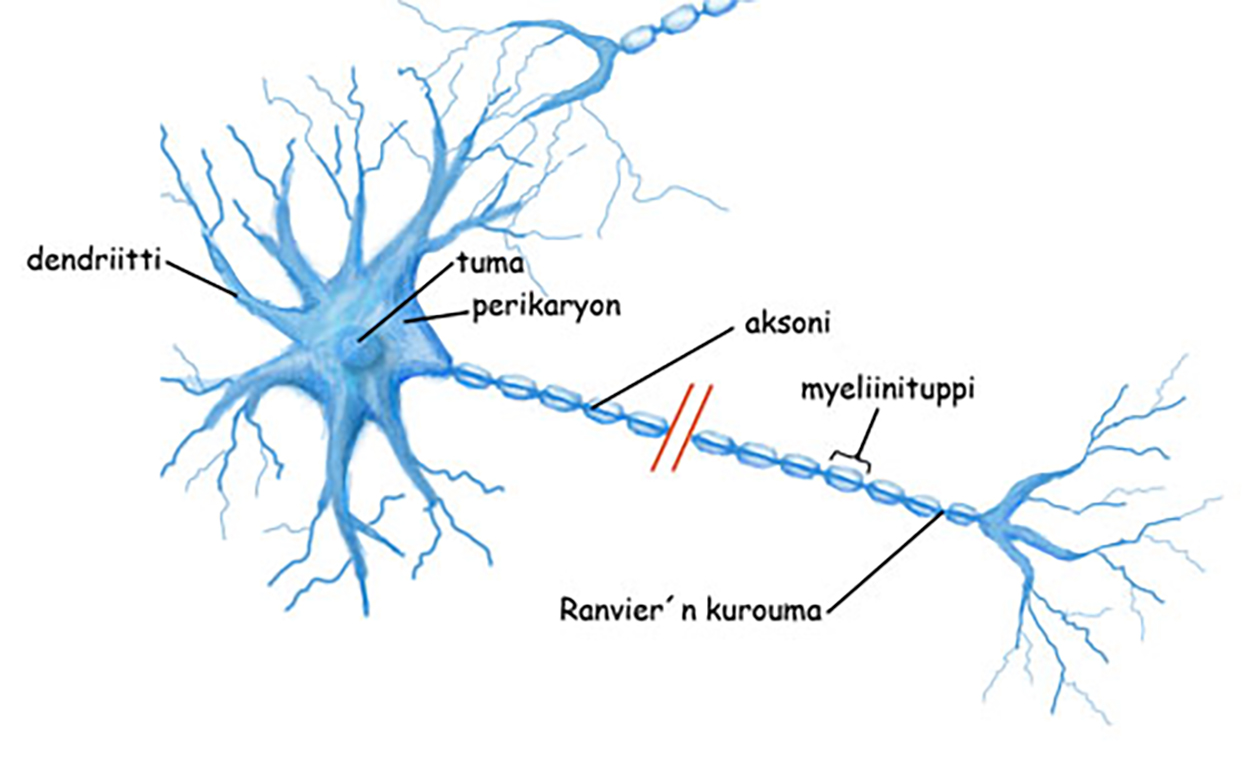 Дозу нейромедиаторов, содержащуюся в одном синаптическом пузырьке принято называть квантом нейромедиатора.
Дозу нейромедиаторов, содержащуюся в одном синаптическом пузырьке принято называть квантом нейромедиатора.Кроме массового выброса нейромедиаторов, происходящего в момент прихода аксонного спайка, есть еще, так называемое, квантовое испускание, когда выделяется только одна везикула с нейромедиаторами. Причем, квантовая активность может быть не связана с вызванной активностью нейронов к которым относится синапс и происходить независимо от нее.
Измерения, проделанные рядом с синапсами, показывают, что на мембране рядом с каждым синапсом время от времени фиксируются возбуждающие постсинаптические потенциалы, имеющие амплитуду порядка 1 мВ или кратную. Считается, что такие миниатюрные постсинаптические потенциалы связаны именно с квантовым испусканием нейромедиаторов.
Когда нейромедиаторы выбрасываются в синаптическую щель, то часть медиаторов попадает за пределы синаптической щели и растекается по пространству, образованному нейронами и окружающими их глиальными клетками. Это явление называется спиловером (spillover). Кроме того, медиаторы испускаются несинаптическими терминалями аксонов и глиальными клетками (рисунок ниже).
Кроме того, медиаторы испускаются несинаптическими терминалями аксонов и глиальными клетками (рисунок ниже).
Источники медиаторов за пределами синаптической щели (Sykova E., Mazel T., Vagrova L., Vorisek I., Prokopova-Kubinova S., 2000)
Когда на дендрите нейрона что-то происходит это сопровождается выделением нейромедиаторов. Внутри синапса нейромедиаторы влияют на ионотропные рецепторы и как следствие на локальное изменение мембранного потенциала дендрита. Когда же нейромедиаторы попадают за пределы синапса они начинают влиять на все, что находится в непосредственной близости. При этом не важно есть ли между этими элементами непосредственный контакт. Это можно сравнить с толпой людей. Люди в толпе могут разбиться на пары и вести между собой беседы, но слышать эти беседы будут не только сами собеседники, но их ближайшие соседи.
Еще про синапсы надо сказать, что в их синаптических пузырьках хранится не один нейромедиатор, а определенный коктейль. Как правило, это смесь из одного основного нейромедиатора и нескольких дополнительных нейропептидов, которые называют нейромодуляторами. Таким образом, спиловер выбрасывает из синапса наружу целый набор сигнальный веществ. Разные нейроны одного типа могут иметь общий основной нейромедиатор, но при этом различаться составом дополнительных.
Таким образом, спиловер выбрасывает из синапса наружу целый набор сигнальный веществ. Разные нейроны одного типа могут иметь общий основной нейромедиатор, но при этом различаться составом дополнительных.
Большинство синапсов, где-то три четверти их числа, располагаются на небольших отростках, называемых шипиками. Шипики отодвигают синапс от дендрита и создают в пространстве такое распределение синапсов, что синапсы от разных дендритов оказываются перемешаны между собой.
Реконструкция участка дендрита пирамидальной клетки. Красным отмечены синапсы на шипиках, синим — на дендритном стволе (Dr. Kristen M. Harris)
Если взять участок дендрита длиною 5 мкм (рисунок ниже), то на нем окажется порядка десяти синапсов. Но дендритные ветки одних нейронов тесно переплетаются с ветками других нейронов. Все они проходят друг от друга в самой непосредственной близости. В цилиндрический объем высотою 5 мкм и диаметром тоже 5 мкм попадает около 100 синапсов. То есть в 10 раз больше того количества, что непосредственно располагается на самой дендритной ветке.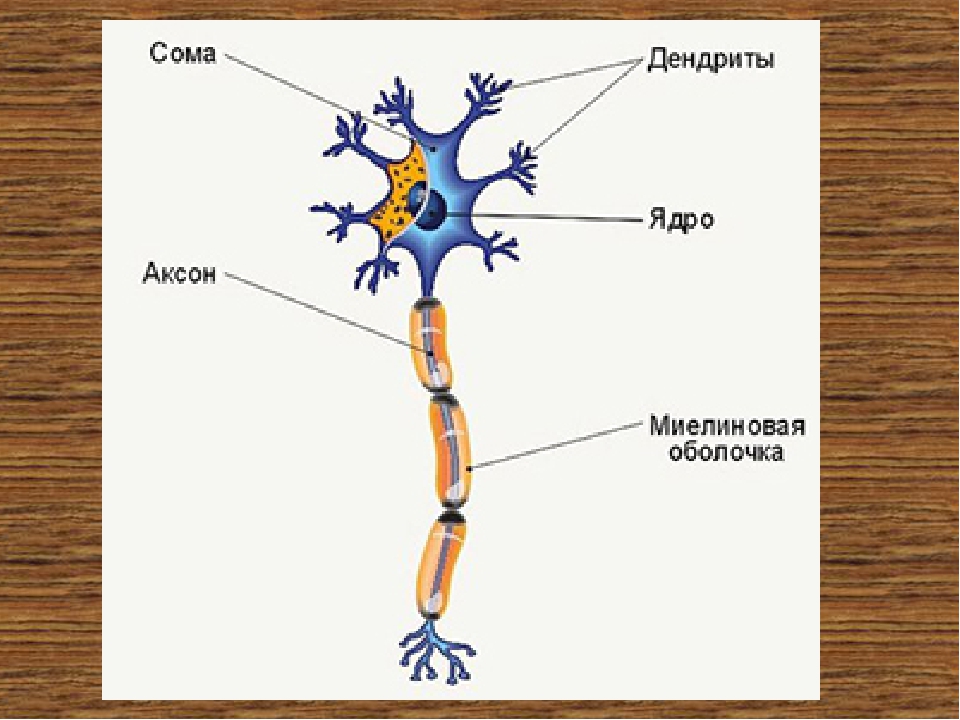
Участок дендрита (Braitenberg V., Schuz A., 1998)
В результате, синапсы образуют систему распределенных случайным образом источников нейромедиаторов для внесинаптической среды. Любая активность в синапсах вызывает появление нейромедиаторов в окружающем их пространстве. Если где-то одновременно становятся активны несколько соседних синапсов, то в таком месте возникает коктейль из тех медиаторов, что выделились из этих синапсов.
Если взять любое место, то поблизости друг от друга в радиусе полутора микрометров окажется около 10 синапсов. Большинство из них будет принадлежать разным дендритам. Если пронаблюдать за тем, какие сочетания нейромедиаторов будут появляться в этом месте, то окажется, что по составу «коктейля» можно каждый раз достаточно точно сказать какие именно синапсы были активны.
Можно привести такой пример. Представьте, что в округе есть 10 баров. Всего существует сотня сортов пива. В каждом разливают только по 3 сорта пива. Эти сорта в каждом баре бармен выбрал однажды случайным образом и теперь разливает только их. Вы обходите несколько баров, выпиваете в каждом по три пива разного сорта и забираете с собой картонные подставки под стаканы с названием выпитого пива. В итоге, по сочетанию картонок, почти всегда, вашей жене удастся определить в каких барах вы побывали.
Вы обходите несколько баров, выпиваете в каждом по три пива разного сорта и забираете с собой картонные подставки под стаканы с названием выпитого пива. В итоге, по сочетанию картонок, почти всегда, вашей жене удастся определить в каких барах вы побывали.
Нейромедиаторы, оказавшиеся вне синапсов имеют свой специфический механизм, позволяющий влиять на работу нейронов. На поверхности дендрита и тела нейрона в больших количествах располагаются метаботропные рецепторы. Эти рецепторы не имеют ионных каналов и не могут напрямую влиять на мембранный потенциал нейрона. С внутренней стороны мембраны эти рецепторы связаны с так называемым G-белком. За это их часто так и называют — рецепторы, сопряженные с G-белком (G-protein-coupled receptors, GPCRs). Когда метаботропные рецепторы активируются своим лигандом они отпускают G-белок и тот начинает оказывать влияние на внутреннее состояние клетки.
Различают два типа воздействия G-белков на клетку (рисунок ниже). В первом случае G-белки напрямую связываются с ближайшими ионными каналами и открывают либо закрывают их, что соответствующим образом меняет мембранный потенциал. Во втором случае G-белки связываются с энзимами, которые запускают работу вторичных посредников. Задействование вторичных внутриклеточных посредников ведет к многократному усилению эффективности рецепторов. Вызванные вторичными посредниками изменения являются медленными, но при этом могут глобально менять состояние всей клетки.
Во втором случае G-белки связываются с энзимами, которые запускают работу вторичных посредников. Задействование вторичных внутриклеточных посредников ведет к многократному усилению эффективности рецепторов. Вызванные вторичными посредниками изменения являются медленными, но при этом могут глобально менять состояние всей клетки.
Работу ионотропных рецепторов принято называть быстрым взаимодействием. Для изменения мембранного потенциала требуется время порядка всего одной миллисекунды. Работу метаботропных рецепторов принято относить к медленным взаимодействиям. При вовлечении вторичных посредников изменения в клетке могут длиться от секунд до часов. Прямое управление метаботропными рецепторами работы ионных каналов протекает значительно быстрее и сопоставимо по времени с быстрым взаимодействием.
Если посмотреть на метаботропный рецептор ближе, то окажется, что у него есть семь трансмембранных доменов и два свободных конца (рисунок ниже).
Структура метаботропного рецептора
За счет свободных концов соседние рецепторы могут соединяться, создавая димеры (рисунок ниже).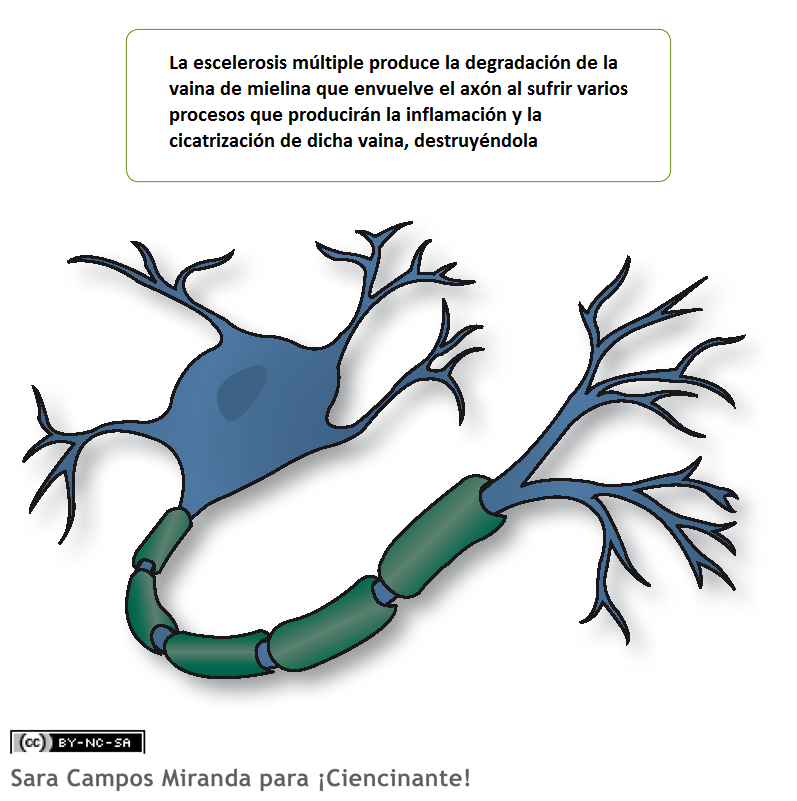 Димеры, в свою очередь, объединяясь, формируют рецептивные кластеры. Аминный и карбоксильный концы рецепторов выполняют роль своеобразных «застежек-липучек», которые за счет электростатического «прилипания» могут формировать различные по составу кластеры рецепторов. Так как в биологических системах нет ничего случайного, то можно предположить, что образование кластеров из различных метаботропных рецепторов несет определенный смысл. Если допустить, что кластер рецепторов реагирует на нейромедиаторы не по отдельности, каждым рецептором самостоятельно, а как единый механизм, то такая реакция может быть сопоставлена с детектированием определенных комбинаций веществ, образующихся при спиловере соседних синапсов.
Димеры, в свою очередь, объединяясь, формируют рецептивные кластеры. Аминный и карбоксильный концы рецепторов выполняют роль своеобразных «застежек-липучек», которые за счет электростатического «прилипания» могут формировать различные по составу кластеры рецепторов. Так как в биологических системах нет ничего случайного, то можно предположить, что образование кластеров из различных метаботропных рецепторов несет определенный смысл. Если допустить, что кластер рецепторов реагирует на нейромедиаторы не по отдельности, каждым рецептором самостоятельно, а как единый механизм, то такая реакция может быть сопоставлена с детектированием определенных комбинаций веществ, образующихся при спиловере соседних синапсов.
Кластеризация рецепторов. A — одиночный рецептор и его взаимодействие с окружающими рецепторами. B — мономерная рецептивная молекула. C — рецептивный димер. D — объединение двух мономеров в контактный (E) и комбинационный (F) димеры. (Радченко, 2007)
Выше описаны далеко не все механизмы, участвующие в работе нейрона.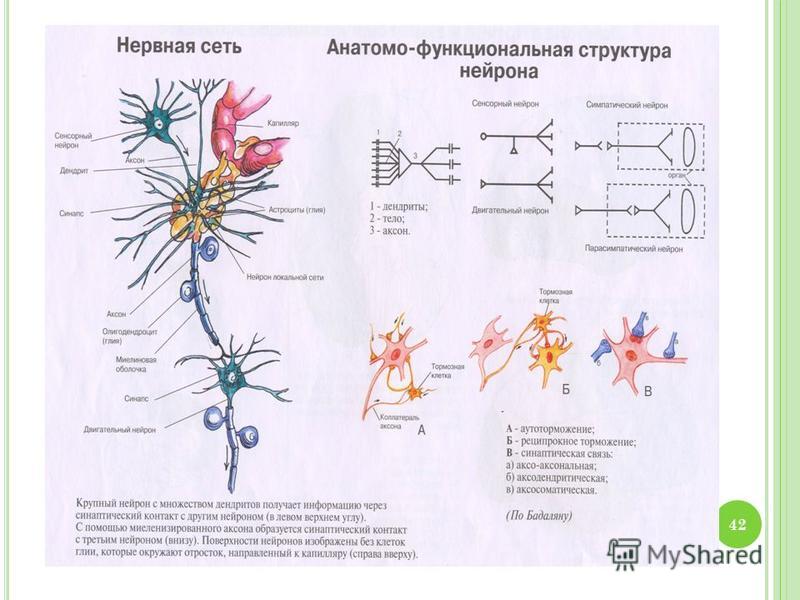 Но уже этого достаточно, чтобы осознать, что реальный нейрон не просто значительно сложнее, чем его формальный аналог. Реальный нейрон -это нечто совсем другое. Похоже, что нейронные сети — это изобретение человека, не имеющее прямых аналогов в природе. Когда искусственной нейронной сети удается решать какие-либо практические задачи, то, похоже, что аналогии с мозгом стоит проводить не на уровне нейронов и связей, а на уровне самих алгоритмических принципов, которые реализует эта сеть.
Но уже этого достаточно, чтобы осознать, что реальный нейрон не просто значительно сложнее, чем его формальный аналог. Реальный нейрон -это нечто совсем другое. Похоже, что нейронные сети — это изобретение человека, не имеющее прямых аналогов в природе. Когда искусственной нейронной сети удается решать какие-либо практические задачи, то, похоже, что аналогии с мозгом стоит проводить не на уровне нейронов и связей, а на уровне самих алгоритмических принципов, которые реализует эта сеть.
Вернемся к клеточным автоматам и вопросу о возможной биологической аналогии. Чтобы претендовать на роль элемента, участвующего в передаче информации, кандидат должен удовлетворять нескольким требованиям:
- У кандидата должны быть различимы, как минимум, два состояния;
- Должна присутствовать возможность передавать информацию о своем состоянии соседям;
- Должен существовать механизм, позволяющий кандидату под воздействием узора, созданного ативностью соседей, менять свое состояние;
- Должен существовать механизм, позволяющий избирательно реагировать на различные окружающие узоры;
- Передача информации должна быть достаточно быстрой, чтобы соответствовать ритмам работы мозга;
- Так как предполагается, что паттерно-волновой механизм должен каждый раз вовлекать в передачу большое количество элементов, то энергетические затраты каждого элемента должны быть минимальны.

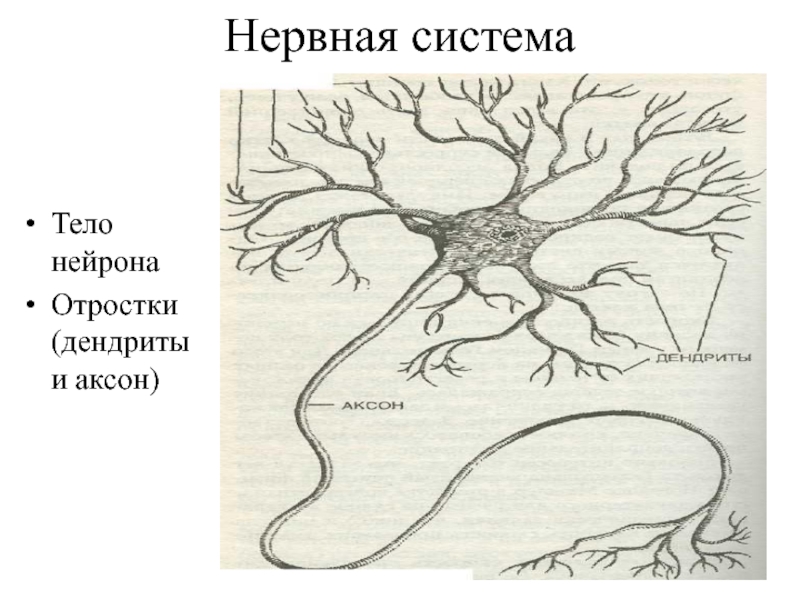
В разное время я рассматривал разных кандидатов на роль биологических аналогов. Обилие механизмов, свойственных мозгу, позволяет почти для всего, что есть в коре придумать гипотетическое обоснование почему именно это может быть аналогом элементов клеточного автомата. Сейчас я склоняюсь к тому, что наиболее подходящий кандидат — это тонкие веточки дендритных деревьев.
Веточки дендритных деревьев, безусловно, являются составной частью нейронов и участвуют в общем механизме его работы. Но это не мешает им проявлять и индивидуальные свойства и быть в каких-то ситуациях автономными элементами.
Когда на веточке возникает миниатюрный возбуждающий постсинаптический потенциал, он как по кабелю распространяется в пределах длины этой веточки. Можно предположить, что распространение электрического сигнала провоцирует минимальное испускание нейромедиаторов из каждого синапса, принадлежащего этой веточке. При этом испускание не затрагивает мембранный потенциал дендрита, а в основном распространяется за пределы синапса. Со стороны это будет выглядеть, как постоянная утечка нейромедиаторов. Состояние, в котором по дендриту пробегает электрический сигнал можно назвать активным состоянием элемента. В момент активности веточки дендрита вокруг нее, предположительно, создается облако нейромедиаторов. В каждом месте этого облака состав коктейля индивидуален и определяется ближайшими синапсами.
В каждом месте коры соседствуют порядка десятка синапсов от различных дендритов. Если несколько дендритов оказываются активны одновременно, то в определенных местах возникает специфический для этого сочетания нейромедиаторов коктейль. Если в таком месте окажется дендрит с метаботропным рецептором чувствительным именно к этому коктейлю, то такой дендрит может получит возбуждающий потенциал и перейти в активное состояние.
В принципе, из такого конструктора несложно собрать биологический аналог нашего клеточного автомата. За счет того, что речь идет о миниатюрных постсинаптических потенциалах и квантовом испускании нейромедиаторов энергетика такой передачи будет крайне невысока.
В клеточном автомате для создания уникальных узоров требовался случайный начальный выбор состояний и память элементов автомата относительно знакомых им узоров. Это проистекало из того, что изначально клеточный автомат был чист и однороден. Для возникновения любой повторяемой неоднородности автомату требовалась случайность и память. С дендритами ситуация несколько интереснее. Дендритные ветки изначально сильно переплетены, причем совершенно случайным образом. Такая неоднородность, по сути, уже является готовой памятью. Эта память позволяет воспринять любой сигнал и выдать предопределенный структурой переплетений ответ. Причем, ответ повторяемый. Это как хеш-функция, которая выдает результат, возможно, не очень понятный, но зато всегда один и тот же для одного и того же входного сигнала.
Упрощенно это выглядит так. Создадим в локальной области узор из нескольких веточек. Где-то в объеме этой локальной области найдутся места, в которых эти веточки пройдут рядом друг с другом. Нейромедиаторы испущенные из них создадут «коктейли». Если рядом с «коктейлями» окажутся дендритные веточки, на которых в этом месте окажется соответствующий рецептор, то такая ветка активируется.
Нейромедиаторы испущенные из них создадут «коктейли». Если рядом с «коктейлями» окажутся дендритные веточки, на которых в этом месте окажется соответствующий рецептор, то такая ветка активируется.
То есть сама система случайных переплетений уже содержит внутри себя механизм создания продолжении для любых комбинаций активности. Это удобно, так как, потенциально, не требует дополнительной памяти кроме того, что уже заложено в хаос переплетений. Но такая конструкция обладает только локальной временной устойчивостью. Если конфигурация дендритов или шипиков изменится, то могут помяться и все получающиеся узоры. Можно предположить, что если мозг, действительно, выбрал такой механизм, то должны существовать системы, обеспечивающие стабильность используемых узоров, оптимизацию их распространения и минимизацию вероятности ошибок распространения. Не исключено, что изменения в дендритных деревьях и изменения, происходящие с количеством и формой шипиков — это отголоски такой оптимизации.
Чтобы проиллюстрировать описанные предположения Антон Морозов сделал 3D масштабную модель в которой воспроизвел волны, основанные на узорах из дендритных веточек.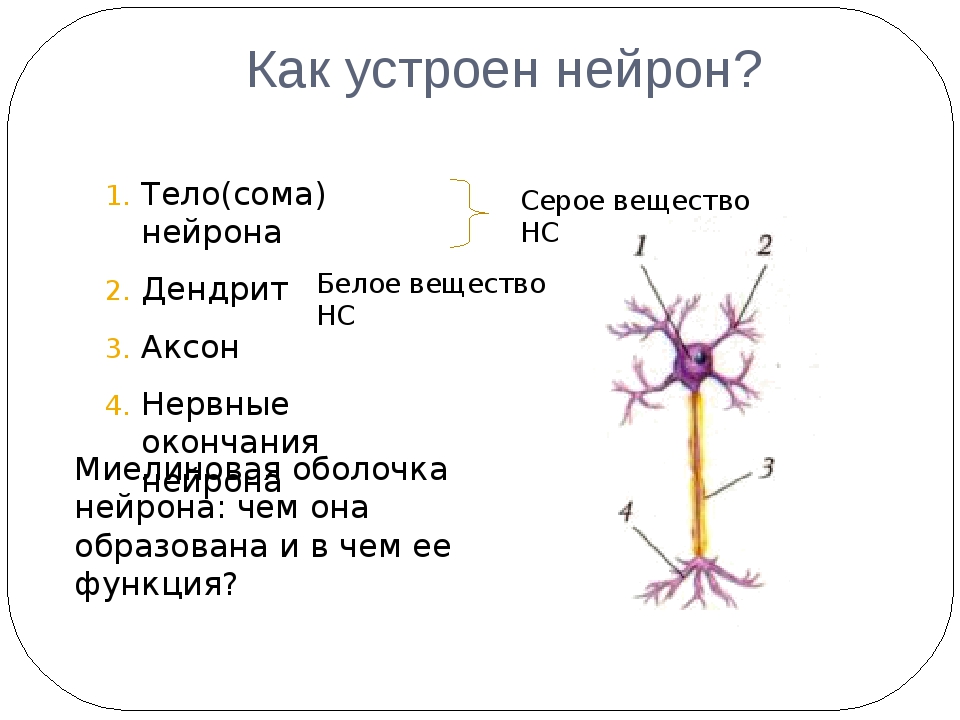 В модели веточки были заменены тонкими трубочками длиной по 50 мкм, что соответствует средней длине дендритной веточки. При той же плотности укладки дендритов, что и в реальной коре получилось нечто, изображенное на рисунке ниже.
В модели веточки были заменены тонкими трубочками длиной по 50 мкм, что соответствует средней длине дендритной веточки. При той же плотности укладки дендритов, что и в реальной коре получилось нечто, изображенное на рисунке ниже.
Задается начальный компактный узор из веточек. В модели у веточек нет собственной памяти. Активируются те веточки, для которых это диктует геометрия случайных связей. Соответственно, любой случайный узор из активных веточек порождает предопределенный геометрией узор продолжения. Новый узор порождает следующий и так далее. Ниже приведено несколько шагов моделирования.
Не стоит искать в описанном механизме распространения дендритных волн какого-либо глубокого смысла, связанного с обработкой информации. По сути, мы просто показали возможный механизм передачи дискретной информации по пространству коры и между структурами мозга. Он, кстати, по своей идее аналогичен механизму передачи цифровой информации по шинам данных, используемым в компьютерах.
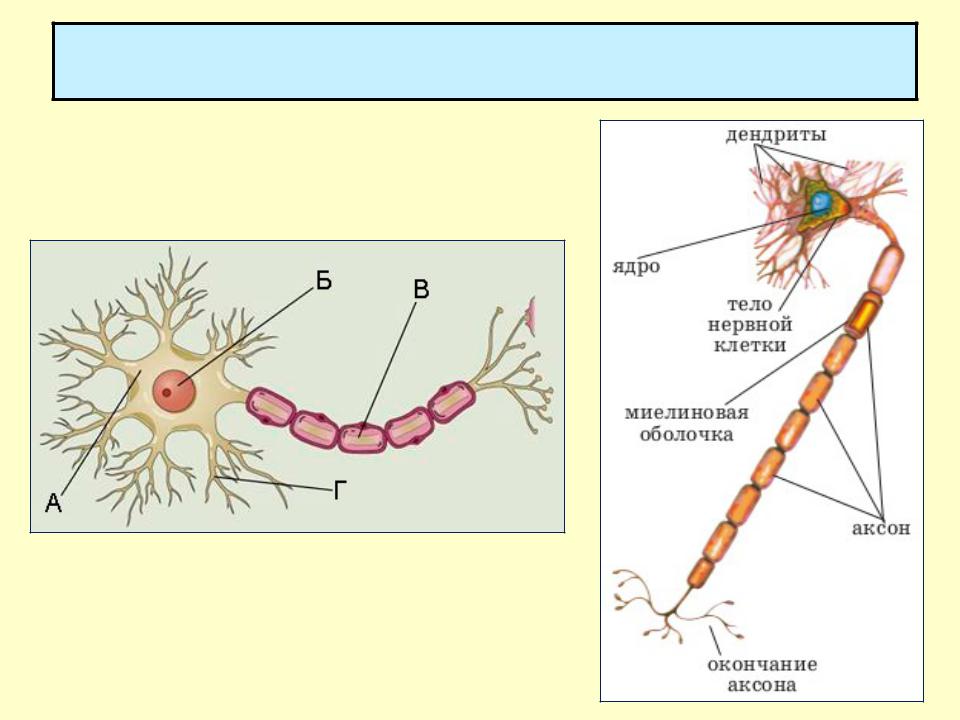 Функция шины данных — передать узор, составленный из нулей и единиц, всем узлам компьютера. С шиной данных несколько проще, ее узор одинаково выглядит в любом месте шины. Но, теоретически, можно представить компьютер, в котором битовый сигнал на шине данных будет меняться по мере движения от одного узла к другому. Если при этом будет соблюдаться однозначное соответствие получаемых кодов, то несложно приспособить узлы компьютера для работы с такой информацией. Но не стоит и недооценивать полученную модель. Далее мы покажем, что развитие этой модели дает удивительные результаты.
Функция шины данных — передать узор, составленный из нулей и единиц, всем узлам компьютера. С шиной данных несколько проще, ее узор одинаково выглядит в любом месте шины. Но, теоретически, можно представить компьютер, в котором битовый сигнал на шине данных будет меняться по мере движения от одного узла к другому. Если при этом будет соблюдаться однозначное соответствие получаемых кодов, то несложно приспособить узлы компьютера для работы с такой информацией. Но не стоит и недооценивать полученную модель. Далее мы покажем, что развитие этой модели дает удивительные результаты.Алексей Редозубов
Логика сознания. Вступление
Что такое нейрон? — Queensland Brain Institute
Нейроны (также называемые нейронами или нервными клетками) — это фундаментальные единицы мозга и нервной системы, клетки, отвечающие за получение сенсорной информации из внешнего мира, отправку двигательных команд нашим мышцам, а также за преобразование и ретрансляция электрических сигналов на каждом этапе между ними. Более того, их взаимодействие определяет, кем мы являемся как люди. При этом наши примерно 100 миллиардов нейронов действительно тесно взаимодействуют с другими типами клеток, широко классифицируемыми как глия (на самом деле они могут превосходить количество нейронов, хотя на самом деле это не известно).
Более того, их взаимодействие определяет, кем мы являемся как люди. При этом наши примерно 100 миллиардов нейронов действительно тесно взаимодействуют с другими типами клеток, широко классифицируемыми как глия (на самом деле они могут превосходить количество нейронов, хотя на самом деле это не известно).
Создание новых нейронов в головном мозге называется нейрогенезом, и это может происходить даже у взрослых.
Как выглядит нейрон?
Полезная аналогия — думать о нейроне как о дереве. Нейрон состоит из трех основных частей: дендритов , аксона и тела клетки или сомы (см. Изображение ниже), которые могут быть представлены как ветви, корни и ствол дерева соответственно. Дендрит (ветвь дерева) — это место, где нейрон получает входные данные от других клеток.Дендриты разветвляются по мере продвижения к своим кончикам, точно так же, как ветки деревьев, и на них даже есть листообразные структуры, называемые шипами .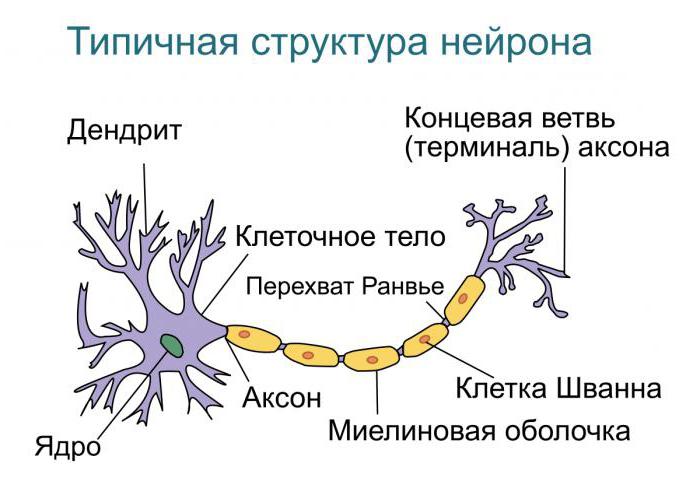
Аксон (корни дерева) является выходной структурой нейрона; когда нейрон хочет поговорить с другим нейроном, он посылает электрическое сообщение, называемое потенциалом действия , по всему аксону. Сома (ствол дерева) — это то место, где лежит ядро, где размещается ДНК нейрона и где белки транспортируются по аксону и дендритам.
Древовидная структура нейрона. Дендритные шипы — это небольшие структуры, которые получают сигналы от аксонов других нейронов. Нижнее правое изображение: сегмент дендрита, от которого отходят колючки, как листья на ветке дерева. Обратите внимание на очень маленький размер (~ 0,001 мм). (Изображение: Алан Вудрафф; Де Ру и др. / CC BY-SA 3.0 через Commons)Существуют разные типы нейронов как в головном, так и в спинном мозге. Обычно они делятся в зависимости от того, где они возникают, куда они проецируются и какие нейротрансмиттеры они используют.
Понятия и определения
Axon — длинная тонкая структура, в которой генерируются потенциалы действия; передающая часть нейрона. После инициации потенциалы действия перемещаются по аксонам, вызывая высвобождение нейромедиатора.
После инициации потенциалы действия перемещаются по аксонам, вызывая высвобождение нейромедиатора.
Дендрит — Приемная часть нейрона. Дендриты получают синаптические входы от аксонов, при этом общая сумма дендритных входов определяет, будет ли нейрон запускать потенциал действия.
Позвоночник — Небольшие выступы на дендритах, которые для многих синапсов являются местом постсинаптического контакта.
Потенциал действия — Кратковременное электрическое событие, обычно генерируемое в аксоне, которое сигнализирует нейрону как «активный». Потенциал действия проходит по длине аксона и вызывает высвобождение нейромедиатора в синапс . Потенциал действия и последующее высвобождение медиатора позволяют нейрону общаться с другими нейронами.
Автор: Д-р Алан Вудрафф
Нейробиология для детей — синапс
Синапс
Нейронов имеют
специализированные проекции под названием дендритов и аксонов .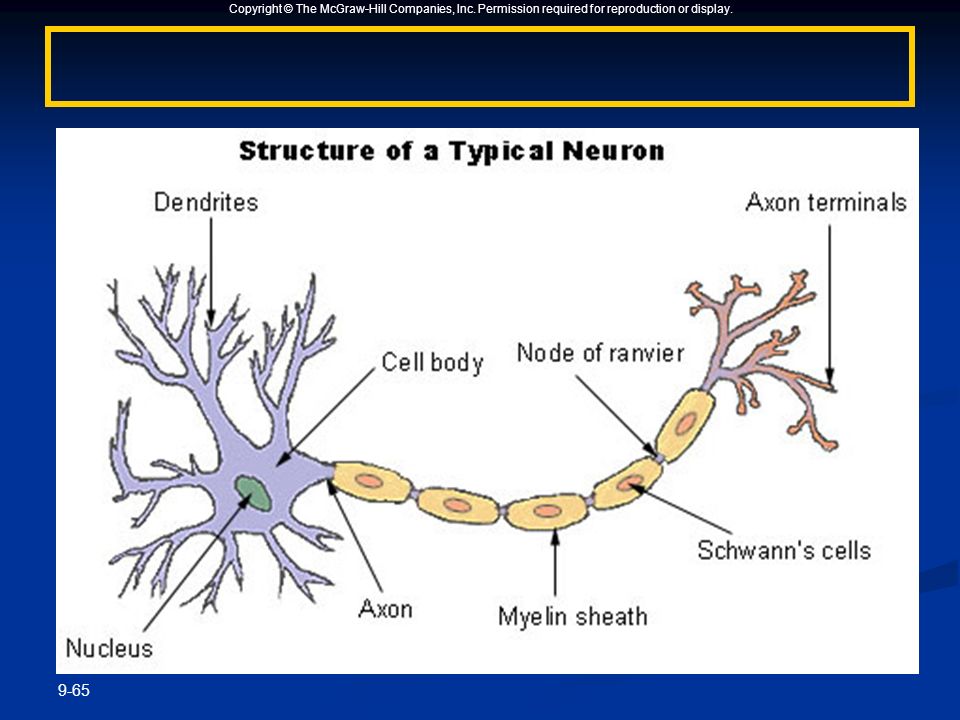 Дендриты несут информацию в тело клетки и
аксоны забирают информацию от тела клетки.
Дендриты несут информацию в тело клетки и
аксоны забирают информацию от тела клетки.
Информация от одного нейрона течет к другому нейрону через синапс . В синапсе есть небольшая щель разделяющие нейроны. Синапс состоит из:
- пресинаптическое окончание, которое содержит нейротрансмиттеры, митохондрии и другие клетки органеллы
- постсинаптическое окончание, которое содержит рецепторные сайты для нейротрансмиттеры
- синаптическая щель или пространство между пресинаптическим и постсинаптическим концовки.
Услышь ЭТО !: Аксон | Дендрит | Миелин | Нейрон | Нейротрансмиттер | Сома | Synapse | Везикула
Электрический триггер для нейротрансмиссии
Для установления связи между нейронами используется электрический импульс. должен пройти по аксону к синаптическому окончанию.
Мобилизация и высвобождение нейротрансмиттеров
В синаптике
терминал (пресинаптическое окончание), электрический импульс вызовет
миграция везикул (красные точки на рисунке в
слева), содержащий нейротрансмиттеры к пресинаптической мембране.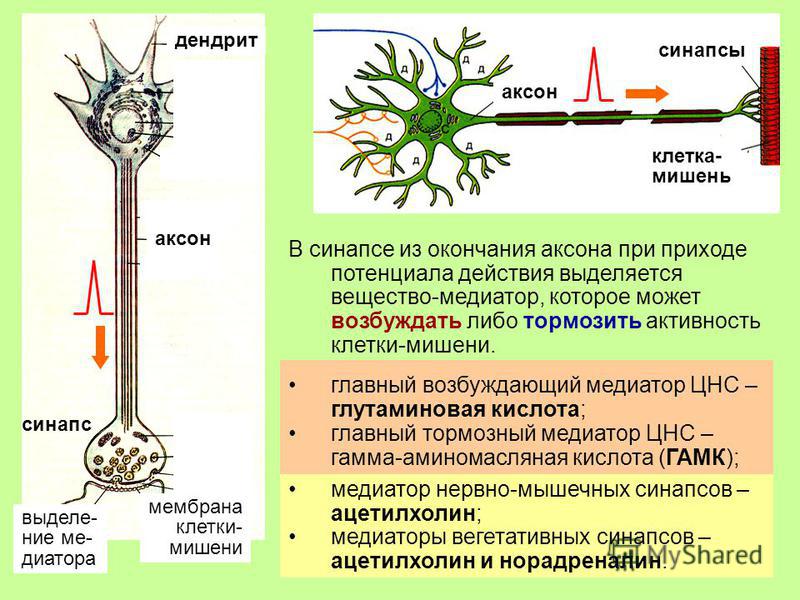 В
мембрана везикул будет сливаться с пресинаптической мембраной, высвобождая
нейротрансмиттеры в синаптическую щель. До недавнего времени считалось
что нейрон производит и высвобождает только один тип нейромедиатора.
Это называлось «Законом Дейла». Однако теперь есть доказательства того, что нейроны
может содержать и выделять более одного вида нейромедиатора.
В
мембрана везикул будет сливаться с пресинаптической мембраной, высвобождая
нейротрансмиттеры в синаптическую щель. До недавнего времени считалось
что нейрон производит и высвобождает только один тип нейромедиатора.
Это называлось «Законом Дейла». Однако теперь есть доказательства того, что нейроны
может содержать и выделять более одного вида нейромедиатора.
Диффузия нейротрансмиттеров по синапсу Расщелина
Затем молекулы нейромедиатора диффундируют через синаптическую щель. где они могут связываться с рецепторными сайтами на постсинаптическом окончании влияют на электрический ответ постсинаптического нейрона.В рисунок справа, постсинаптическое окончание — дендрит (аксодендритный синапс), но синапсы могут возникать на аксонах (аксоаксонический синапс) и тела клеток (аксосоматический синапс).
Когда нейротрансмиттер связывается с рецептором на постсинаптической стороне
синапс, он изменяет возбудимость постсинаптической клетки:
постсинаптическая клетка с большей или меньшей вероятностью инициирует действие
потенциал.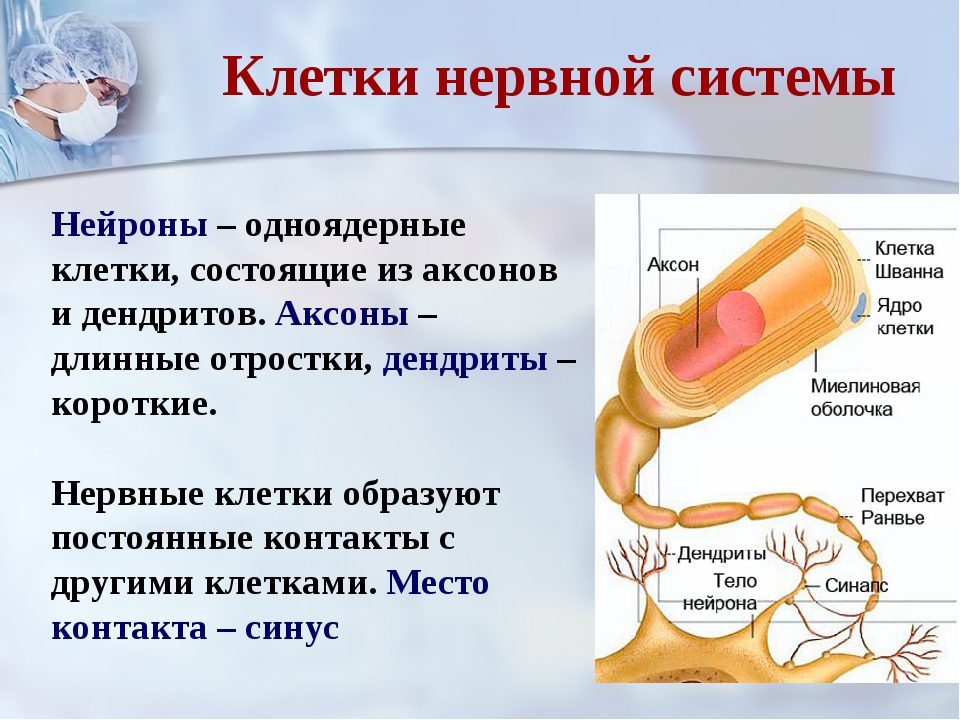 Если количество возбуждающих постсинаптических событий велико
достаточно, они добавят, чтобы вызвать потенциал действия в постсинаптическом
ячейка и продолжение сообщения.«
Если количество возбуждающих постсинаптических событий велико
достаточно, они добавят, чтобы вызвать потенциал действия в постсинаптическом
ячейка и продолжение сообщения.«
Многие психоактивные препараты и нейротоксины могут изменять свойства высвобождение нейротрансмиттера, обратный захват нейротрансмиттера и доступность сайтов связывания рецепторов.
Типы синапсов
Поздравляем со 123-м Днём Рождения! слово «СИНАПС». В 2020 году слову «синапс» исполнилось 123 года. В слово «синапс» впервые было использовано в книге под названием « Учебник физиологии», часть третья: Центральная нервная система , Майкл Фостер и при поддержке Чарльза С.Шеррингтоном в 1897 году. Вероятно, это был Чарльз С. Шеррингтон, который ввел термин синапс. Слово «синапс» происходит от от греческих слов «син» и «гаптейн», которые означают «вместе» и «к застежка, соответственно.
«Вы — ваши синапсы. Они
кто вы есть «.
— Джозеф Леду, 2002 (в Synaptic Self )
Посмотреть некоторые синапсы
«Близко и лично».
Играйте в интерактивный поиск слов на нейрон и нейротрансмиттеры. Играйте во внешнюю игру, чтобы закрепить то, что у вас есть узнал о синапсе.Раскрасьте синапс онлайн: Рисунок 1 | Картинка 2
Copyright © 1996-2020, Эрик Х. Чудлер. Все права. Зарезервированный.
Нейрон
Клетки нервной системы, называемые нейронами, взаимодействуют друг с другом уникальными способами. Нейрон — это основная рабочая единица мозга, специализированная клетка, предназначенная для передачи информации другим нервным клеткам, мышцам или клеткам железы.
Нейроны — это клетки нервной системы, которые передают информацию другим нервным клеткам, мышцам или клеткам железы.У большинства нейронов есть тело клетки, аксон и дендриты. Тело клетки содержит ядро и цитоплазму. Аксон идет от тела клетки и часто дает начало множеству более мелких ветвей, прежде чем оканчивается на нервных окончаниях. Дендриты выходят из тела клетки нейрона и получают сообщения от других нейронов. Синапсы — это точки контакта, где один нейрон взаимодействует с другим. Дендриты покрыты синапсами, образованными концами аксонов других нейронов.
Синапсы — это точки контакта, где один нейрон взаимодействует с другим. Дендриты покрыты синапсами, образованными концами аксонов других нейронов.
Иллюстрация Лидии В.Кибюк, Балтимор, Мэриленд; Девон Стюарт, Гаррисбург, Пенсильвания
Нейроны — это клетки нервной системы, которые передают информацию другим нервным клеткам, мышцам или клеткам железы. У большинства нейронов есть тело клетки, аксон и дендриты. Тело клетки содержит ядро и цитоплазму. Аксон идет от тела клетки и часто дает начало множеству более мелких ветвей, прежде чем оканчивается на нервных окончаниях. Дендриты выходят из тела клетки нейрона и получают сообщения от других нейронов. Синапсы — это точки контакта, где один нейрон взаимодействует с другим.Дендриты покрыты синапсами, образованными концами аксонов других нейронов.
Мозг стал таким, какой он есть, благодаря структурным и функциональным свойствам взаимосвязанных нейронов. Мозг млекопитающих содержит от 100 до 100 миллиардов нейронов, в зависимости от вида. Каждый нейрон млекопитающего состоит из тела клетки, дендритов и аксона.
Каждый нейрон млекопитающего состоит из тела клетки, дендритов и аксона.
Тело клетки содержит ядро и цитоплазму. Аксон идет от тела клетки и часто дает начало множеству более мелких ветвей, прежде чем оканчивается на нервных окончаниях.
Дендриты отходят от тела клетки нейрона и получают сообщения от других нейронов. Синапсы — это точки контакта, где один нейрон взаимодействует с другим. Дендриты покрыты синапсами, образованными концами аксонов других нейронов.
Когда нейроны получают или отправляют сообщения, они передают электрические импульсы по своим аксонам, длина которых может варьироваться от крошечной доли дюйма (или сантиметра) до трех футов (около одного метра) или более. Многие аксоны покрыты слоистой миелиновой оболочкой, которая ускоряет передачу электрических сигналов по аксону.Эта оболочка состоит из специализированных клеток, называемых глией. В головном мозге глии, образующие оболочку, называются олигодендроцитами, а в периферической нервной системе они известны как клетки Шванна.
В мозге содержится по крайней мере в десять раз больше глии, чем нейронов. Глия выполняет множество работ. Некоторое время исследователи знали, что глия переносит питательные вещества к нейронам, очищает мозг от мусора, переваривает части мертвых нейронов и помогает удерживать нейроны на месте. Текущие исследования открывают новые важные роли глии в работе мозга.
дендритов — обзор | Темы ScienceDirect
Ремоделирование клеток мозга с помощью глюкокортикоидов
Дендриты (проекции, исходящие из тела клетки нейрона) позволяют нейронам взаимодействовать с другими нейронами (см. Рис. 15.5). Больше и / или более длинные дендриты увеличивают возможности для общения.Дендриты — очень пластичные структуры, и изменения дендритов и синаптических контактов между нейронами также являются частью физической основы памяти.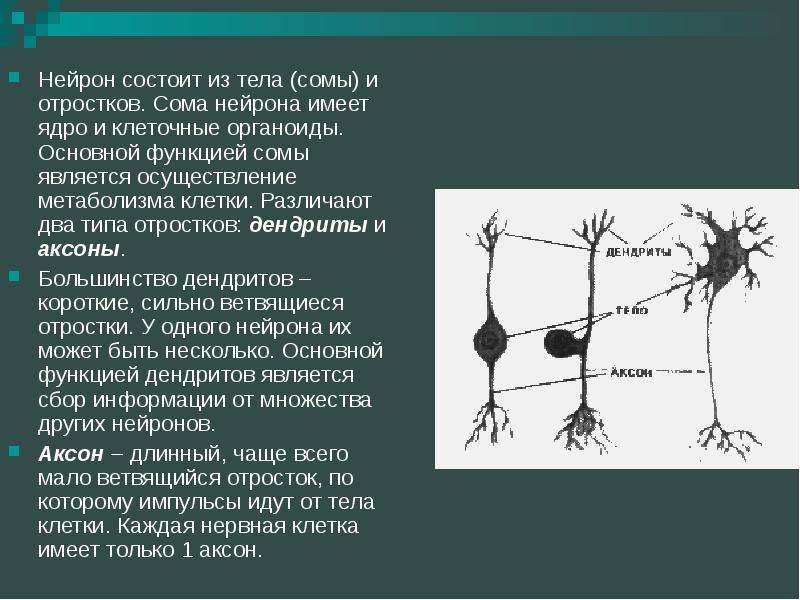 Глюкокортикоиды играют роль в ремоделировании дендритов. Хотя стрессовый опыт, который снижает производительность при тестировании памяти, может привести к сокращению дендритов, острый стресс, который улучшает производительность памяти, способствует их росту. Исследователи из Принстонского университета изучили выполнение задания на память, зависящее от гиппокампа.Они показали, что острый стресс улучшает выполнение этой задачи. Кроме того, у животных, которые хорошо справлялись с этой задачей, также наблюдалось увеличение количества дендритных шипов в гиппокампе. В свою очередь, исследователи обнаружили прямую связь между переживанием стресса (легкие электрические разряды ног) и увеличением дендритных шипов у самцов крыс. В целом, исследования на животных показали, что острый стресс и повышенный уровень глюкокортикоидов влияют на выполнение задач с памятью, улучшая некоторые аспекты обработки памяти и затрудняя другие, в зависимости от дозы глюкокортикоидов и типа задачи с памятью.Кроме того, стресс и глюкокортикоиды влияют на функцию и структуру нервных областей, которые имеют решающее значение для памяти.
Глюкокортикоиды играют роль в ремоделировании дендритов. Хотя стрессовый опыт, который снижает производительность при тестировании памяти, может привести к сокращению дендритов, острый стресс, который улучшает производительность памяти, способствует их росту. Исследователи из Принстонского университета изучили выполнение задания на память, зависящее от гиппокампа.Они показали, что острый стресс улучшает выполнение этой задачи. Кроме того, у животных, которые хорошо справлялись с этой задачей, также наблюдалось увеличение количества дендритных шипов в гиппокампе. В свою очередь, исследователи обнаружили прямую связь между переживанием стресса (легкие электрические разряды ног) и увеличением дендритных шипов у самцов крыс. В целом, исследования на животных показали, что острый стресс и повышенный уровень глюкокортикоидов влияют на выполнение задач с памятью, улучшая некоторые аспекты обработки памяти и затрудняя другие, в зависимости от дозы глюкокортикоидов и типа задачи с памятью.Кроме того, стресс и глюкокортикоиды влияют на функцию и структуру нервных областей, которые имеют решающее значение для памяти.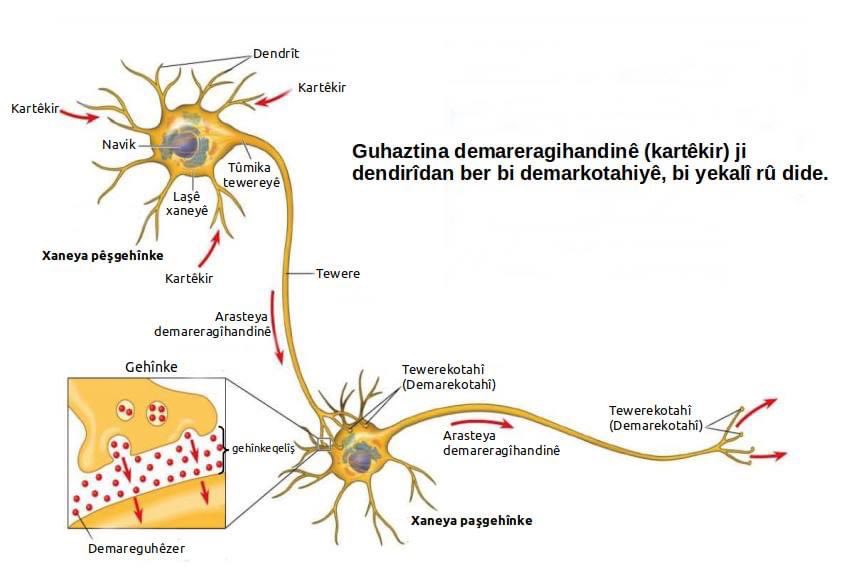
В литературе, посвященной исследованию стресса и памяти у людей, есть некоторые несоответствия, которые, вероятно, связаны с такими факторами, как исследуемая стадия обработки памяти, фармакологическое введение в сравнении с вызванным стрессом повышением глюкокортикоидов, типом теста памяти, характеристиками субъектов ( молодой, пожилой, мужской, женский) и так далее. Однако, как правило, влияние стресса на память, обнаруженное у людей, согласуется с результатами экспериментов на животных.Например, недавнее исследование в Университете Висконсина обнаружило перевернутую U-образную зависимость между выполнением задачи на распознавание памяти, включающей как эмоционально отрицательные, так и нейтральные стимулы и уровни глюкокортикоидов. Точно так же исследователи из Университета Трира в Германии заметили, что либо психосоциальный стресс, либо введение кортизола субъектам ухудшали производительность при выполнении декларативной задачи памяти, которая очень чувствительна к функции гиппокампа, но не к задаче процедурной памяти, которая зависит от структур, отличных от гиппокамп.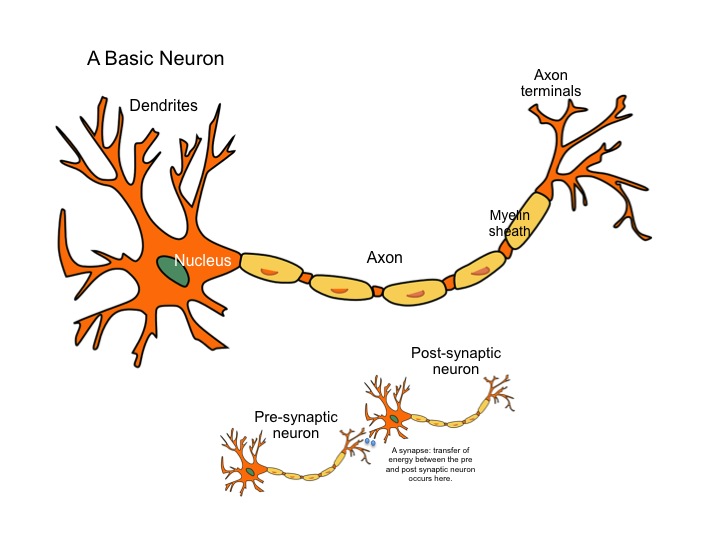 Результаты исследователей из Университета Макгилла предполагают, что префронтальная кора и рабочая память могут быть особенно чувствительны к резкому повышению уровня кортизола. Криволинейная зависимость была обнаружена между характеристиками рабочей памяти и уровнем кортизола после введения различных доз гидрокортизона молодым мужчинам. Таким образом, хотя не все проблемы решены, у людей, как и у экспериментальных животных, есть свидетельства того, что резкие колебания гормонов стресса влияют на обучение и память.В некоторых случаях резкие изменения уровня глюкокортикоидов могут улучшить функцию памяти, в то время как в других такие изменения могут препятствовать функции памяти. Перед исследователями стоит серьезная задача — лучше определить и предсказать обстоятельства, при которых острый стресс повысит или снизит продуктивность как людей, так и лабораторных животных.
Результаты исследователей из Университета Макгилла предполагают, что префронтальная кора и рабочая память могут быть особенно чувствительны к резкому повышению уровня кортизола. Криволинейная зависимость была обнаружена между характеристиками рабочей памяти и уровнем кортизола после введения различных доз гидрокортизона молодым мужчинам. Таким образом, хотя не все проблемы решены, у людей, как и у экспериментальных животных, есть свидетельства того, что резкие колебания гормонов стресса влияют на обучение и память.В некоторых случаях резкие изменения уровня глюкокортикоидов могут улучшить функцию памяти, в то время как в других такие изменения могут препятствовать функции памяти. Перед исследователями стоит серьезная задача — лучше определить и предсказать обстоятельства, при которых острый стресс повысит или снизит продуктивность как людей, так и лабораторных животных.
Ключевые молекулярные детали распространенного типа черепно-мозговой травмы и возможная новая стратегия лечения — ScienceDaily
Среди всего плохого, что может случиться с мозгом при сильной толчке — например, в автомобильной аварии — одним из наиболее распространенных и вызывающих беспокойство является повреждение аксонов. Аксоны — это длинные ножки, которые вырастают из тел нейронов и передают сигналы другим нейронам. Они являются частью «проводов» головного мозга, и иногда они достигают поразительной длины — от головного мозга до спинного мозга. Но аксоны тонкие и хрупкие. Когда мозг получает сильный удар, аксоны часто выходят за пределы своих структурных ограничений. Они либо ломаются, либо стремительно деградируют.
Аксоны — это длинные ножки, которые вырастают из тел нейронов и передают сигналы другим нейронам. Они являются частью «проводов» головного мозга, и иногда они достигают поразительной длины — от головного мозга до спинного мозга. Но аксоны тонкие и хрупкие. Когда мозг получает сильный удар, аксоны часто выходят за пределы своих структурных ограничений. Они либо ломаются, либо стремительно деградируют.
Это мы знаем. Но ученые не поняли, что будет дальше. Что происходит с нейроном, когда его аксон движется на юг?
«Разобраться в точных механизмах того, что происходит после повреждения аксонов, было действительно сложно», — говорит Энн Марион Тейлор, доктор философии, доцент Объединенного департамента биомедицинской инженерии UNC / NC.«Но мы думаем, что наконец-то выяснили ключевую часть того, что происходит и почему».
В статье Nature Communications Тейлор и его коллеги раскрыли новые молекулярные детали аксотомии — когда нейроны повреждены или полностью оторваны.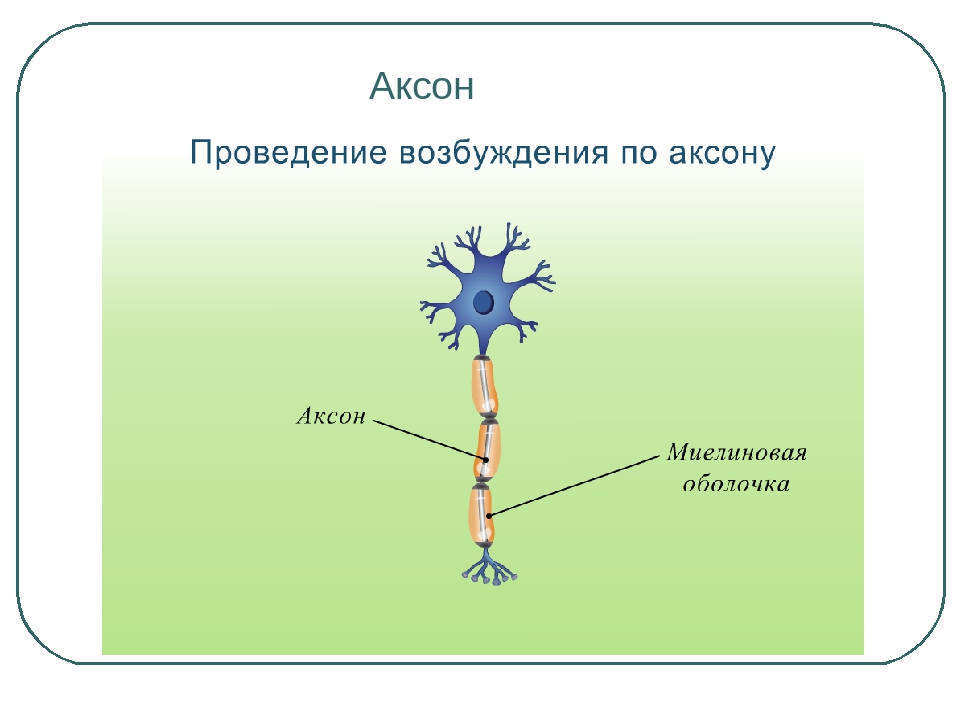
Сокращение дендритных шипов, повышение возбудимости
Ученые действительно знают, что оторванный аксон заставляет нейрон быстро терять некоторые входящие связи от других нейронов. Эти связи происходят на коротких, похожих на корень усиках, называемых дендритами, которые вырастают из тела клетки нейрона или сомы.Сами дендриты вырастают крошечные выступы, называемые шипами, для создания реальных связей или синапсов с входящими аксонами. Число этих дендритных шипов уменьшается после аксотомии.
По мере того, как он теряет входные соединения, раненый нейрон также становится более возбудимым: нейрон с большей вероятностью посылает сигналы по усеченному аксону, когда это стимулируется другими нейронами. Нейроны обычно имеют несколько входов. Некоторые из них возбуждающие, заставляя нейрон активироваться; другие являются тормозящими, удерживая нейрон от возбуждения.Нейроны с усеченными аксонами показывают нарушение нормального баланса возбуждения / торможения в пользу возбудимости.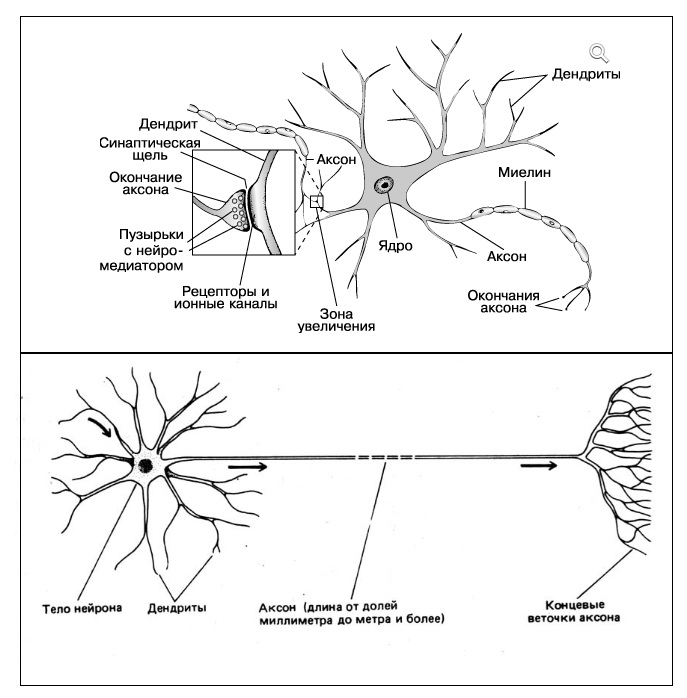
Считается, что эта повышенная возбудимость в течение недель и месяцев после травмы в значительной степени является адаптивной, полезной реакцией — переключением на нейронный «режим поиска», подобный тому, который наблюдается в развивающемся мозге. Этот полезный переключатель увеличивает шанс того, что нейрон с усеченным аксоном сможет соединиться с новым партнером и продолжать оставаться продуктивным членом нейронного общества.
«Неврологи это знают», — сказал Тейлор, член Центра нейробиологии UNC. «Вот почему они продвигают физиотерапию и переподготовку для людей, страдающих травмой головы. В течение этого длительного периода возбудимости физическая нагрузка и переподготовка могут помочь направить поврежденные нейроны по полезным путям».
Но возбудимость нейрона, вызванная травмой, тоже может вызвать проблемы. Нейрон может погибнуть от перевозбуждения (нейробиологи называют это эксайтотоксичностью). Гиперактивность нейронов после травмы также может привести к не поддающейся лечению боли, мышечным спазмам или возбуждению у пациента. В первые дни сразу после травмы врачи часто лечат пациентов с черепно-мозговой травмой лекарствами, такими как габапентин, специально разработанными для подавления гипервозбудимости нейронов.
В первые дни сразу после травмы врачи часто лечат пациентов с черепно-мозговой травмой лекарствами, такими как габапентин, специально разработанными для подавления гипервозбудимости нейронов.
То, что ученые не очень хорошо понимают, — это биологические детали, причины и причины потери дендритных шипов и гипервозбудимости. Эти детали были неуловимы из-за сложности мозга, похожей на спагетти, что делает чрезвычайно трудным для ученого изолировать нейрон и его аксон для манипуляций и анализа, будь то в лабораторной посуде или лабораторном животном.
Несколько лет назад, будучи аспирантом по биомедицинской инженерии Калифорнийского университета в Ирвине, Тейлор изобрел устройство, помогающее решить эту проблему. Это микрофлюидная камера с крошечными бороздками, которые захватывают отдельные аксоны культивируемых нейронов по мере их роста.
«Аксоны не могут вращаться, поэтому они просто продолжают расти прямо, пока не достигнут отдельного отсека», — говорит Тейлор. «Мы можем разрезать аксон в его компартменте, а затем посмотреть на ответы в ассоциированной соме или дендритах, не затрагивая аксоны в других компартментах.«
«Мы можем разрезать аксон в его компартменте, а затем посмотреть на ответы в ассоциированной соме или дендритах, не затрагивая аксоны в других компартментах.«
Потеря торможения
Тейлор и ее коллеги использовали устройство в новом исследовании, чтобы проанализировать, что происходит, когда аксон разрывается. Они обнаружили, что события внутри самого нейрона приводят к потере дендритных шипов и повышенной возбудимости. Сигналы, возникающие в месте повреждения, быстро возвращаются вдоль оставшейся части аксона к соме и ядру нейрона, запуская новый паттерн активности генов. Команде Тейлора удалось заблокировать активность гена нейрона, чтобы предотвратить потерю дендритного шипа и повышенную возбудимость.
Тейлор и его коллеги проанализировали, как изменялась активность генов до и после аксотомии. Множественные гены были изменены после аксотомии. Оказалось, что активность одного из этих генов, кодирующего белок нетрин-1, резко снизилась. Отдельный анализ показал аналогичное падение уровня нетрина-1 в пораженных нейронах у крыс, аксоны которых от головного до спинного мозга были разрезаны.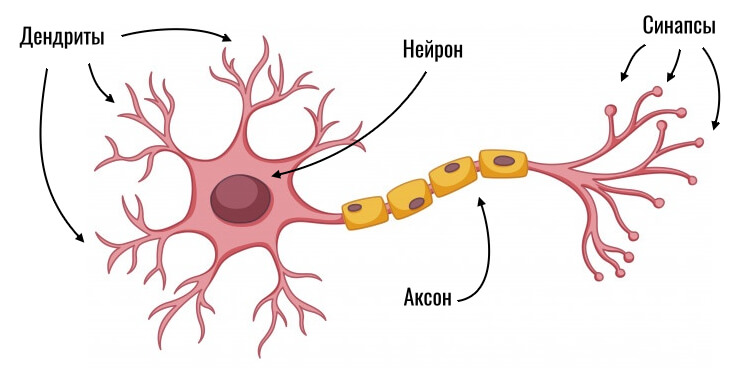 Вместе эти результаты указывают на то, что отсутствие нетрина-1 может быть основным фактором, приводящим к изменениям нейронов после аксотомии.
Вместе эти результаты указывают на то, что отсутствие нетрина-1 может быть основным фактором, приводящим к изменениям нейронов после аксотомии.
Когда Тейлор и его коллеги добавили нетрин-1 к аксотомированным нейронам, чтобы восстановить белок до нормального уровня — даже через два полных дня после отделения аксона — они обнаружили, что лечение быстро обратило вспять всю потерю дендритного шипа и большую часть гипер- возбудимость.
«Обработанные нейроны больше напоминали неповрежденные контрольные», — говорит Тейлор. «Это было поразительное открытие, и мы были удивлены, обнаружив, что нетрин-1 нормализует как количество синапсов, так и возбудимость, даже при применении через несколько дней после травмы.«
Она добавила: «Нам еще очень далеко, но мы действительно надеемся превратить открытие нетрина-1 в новую терапию. В идеале, он будет делать то, на что нацелены габапентин и родственные препараты для травм головы, только лучше и точнее.»
СТРУКТУР НЕЙРОНА — Ричардс в мозге
Аксон : один из трех основных (компонентов) нейрона.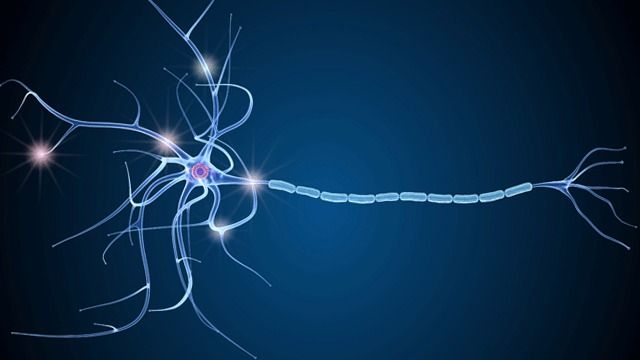 Проекция нейрона «клеточного тела», которая может достигать больших расстояний в мозгу. Функция аксона заключается в отправке данных из клетки.(Рамачандран, 9) Аксоны — это «выходные» каналы, а дендриты — «входные» каналы. (LeDoux, 40) Выступ, исходящий из тела клетки, который вместе с «дендритом» другого нейрона делает возможными связи между нейронами. (Голдберг, 39) Длинное волокно нейрона, часто с тысячами точек контакта. (The Brain-Francis Crick, 132) Возникает на одном конце тела клетки и может достигать нескольких футов. Часто по длине распадается на одну или несколько ветвей. (Кандел, 64) У большинства нейронов есть только аксон.Однако каждый аксон много раз разветвляется, прежде чем закончится, позволяя одному нейрону порождать множество терминалов. В результате сообщения, отправленные из одной ячейки, могут повлиять на многие другие. (LeDoux, 42) Критическая часть нейрона. Отходит от тела нейрона. Обеспечивает путь, по которому сигналы могут проходить от тела клетки, иногда на большие расстояния, к нейронам в других частях мозга и в нервной системе.
Проекция нейрона «клеточного тела», которая может достигать больших расстояний в мозгу. Функция аксона заключается в отправке данных из клетки.(Рамачандран, 9) Аксоны — это «выходные» каналы, а дендриты — «входные» каналы. (LeDoux, 40) Выступ, исходящий из тела клетки, который вместе с «дендритом» другого нейрона делает возможными связи между нейронами. (Голдберг, 39) Длинное волокно нейрона, часто с тысячами точек контакта. (The Brain-Francis Crick, 132) Возникает на одном конце тела клетки и может достигать нескольких футов. Часто по длине распадается на одну или несколько ветвей. (Кандел, 64) У большинства нейронов есть только аксон.Однако каждый аксон много раз разветвляется, прежде чем закончится, позволяя одному нейрону порождать множество терминалов. В результате сообщения, отправленные из одной ячейки, могут повлиять на многие другие. (LeDoux, 42) Критическая часть нейрона. Отходит от тела нейрона. Обеспечивает путь, по которому сигналы могут проходить от тела клетки, иногда на большие расстояния, к нейронам в других частях мозга и в нервной системе.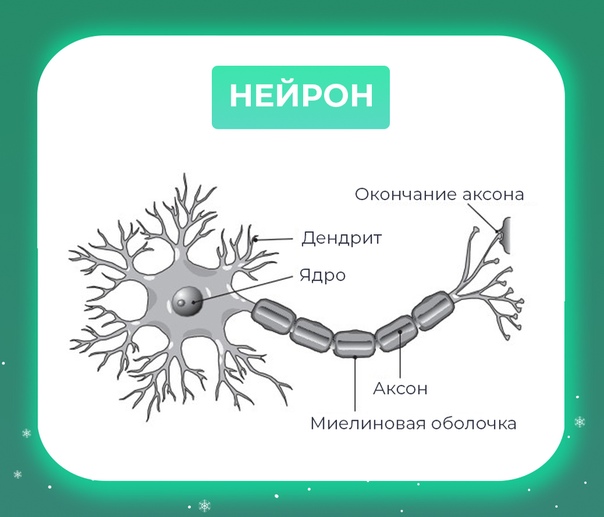 (The Brain-Charles Stevens, 15). Живой кабель различной длины (от микроскопической до шести футов).Их часто сравнивают с проводами, потому что они переносят электрические импульсы с очень высокой скоростью (от 2 до 200 миль в час) к дендритам соседних нейронов. (Doidge, 53) Также называется «волокно», «нервное волокно», «процесс» и «клеточный процесс».
(The Brain-Charles Stevens, 15). Живой кабель различной длины (от микроскопической до шести футов).Их часто сравнивают с проводами, потому что они переносят электрические импульсы с очень высокой скоростью (от 2 до 200 миль в час) к дендритам соседних нейронов. (Doidge, 53) Также называется «волокно», «нервное волокно», «процесс» и «клеточный процесс».
Коллатерали аксона : ветви, которые расходятся от главного аксона под прямым углом. (Патестас, 30) Также называется «коллатералями».
Axon Hillock : место соединения (тела нейронной клетки) и аксона. Где начинается потенциал действия.(Kolb, 79)
Мембрана аксона : мембрана, которая окружает аксон. Содержит специальные отверстия, известные как «ионные каналы», которые позволяют «ионам» калия выходить изнутри клетки, где они присутствуют в высоких концентрациях, наружу, где они присутствуют в низких концентрациях. Поскольку калий является положительно заряженным ионом, его выход из клетки оставляет внутреннюю поверхность мембраны с небольшим избытком отрицательного заряда. Наружная поверхность клеточной мембраны покрывается положительными зарядами ионов калия, которые диффундировали из клетки.Внутренняя часть мембраны покрывается отрицательными зарядами от «белков» (внутри клетки), пытающихся втянуть ионы калия обратно в клетку. Этот баланс ионов поддерживает стабильный «мембранный потенциал покоя» 70 милливольт. (Кандел, 80)
Наружная поверхность клеточной мембраны покрывается положительными зарядами ионов калия, которые диффундировали из клетки.Внутренняя часть мембраны покрывается отрицательными зарядами от «белков» (внутри клетки), пытающихся втянуть ионы калия обратно в клетку. Этот баланс ионов поддерживает стабильный «мембранный потенциал покоя» 70 милливольт. (Кандел, 80)
Axon Terminal : крошечный конец каждой ветви аксона. (Кандел, 64) Обеспечивает связь с другими нейронами. (Рамачандран, 9) Содержит нейротрансмиттеры в синаптических пузырьках. (NCIt) Точка, в которой передающий нейрон связывается с принимающими нейронами.(LeDoux, 40) На своем конце аксон может «ветвиться», образуя множество окончаний аксона, что позволяет одному аксону вступать в синаптический контакт с множеством других нейронов, мышечных клеток или клеток железы. (Patestas, 30) Также упоминается как «терминал», «нервный терминал», «синаптический терминал», «пресинаптический терминал», «пресинаптический узел», «конечный конец», «терминальный бутон» и «терминальный конец бутона».
Пресинаптическая мембрана : мембрана на стороне выхода передатчика синапса. Образует окончание аксона.(Kolb, 153)
Синаптические пузырьки : небольшие секреторные пузырьки, содержащие нейротрансмиттер, находятся внутри аксона рядом с пресинаптической мембраной и выпускают свое содержимое в «синаптическую щель» после слияния с мембраной. (GHR) Содержат от 10 000 до 100 000 молекул определенного типа нейромедиатора. В одном терминале могут быть тысячи синаптических пузырьков. Везикулы служат для защиты передающих молекул от «ферментов» внутри терминала, которые в противном случае разрушили бы их.(The Brain-Leslie Iversen, 76) Содержат нейротрансмиттеры. В синапсе электрический импульс запускает «миграцию» пузырьков. Мембрана пузырька сливается с мембраной терминала. Это действие высвобождает нейротрансмиттеры в синаптику (щель). (Чудлер, 15) Характеризуется единственной «фосфолипидной» мембраной. Они переносят вещества через клетку, внутрь и из клетки. Производится «эндоплазматическим ретикулумом», «аппаратом Гольджи» и клеточной мембраной. Везикулы передающей клетки сливаются с ее мембраной, высвобождая химические регуляторы посредством «экзоцитоза» во «внеклеточную жидкость».»(Norton Lectures, 6/2/09) Клеточная структура нервной системы настолько миниатюризирована, что длина волны видимого света слишком мала, чтобы ее исследовать. Длина волны зеленого света в десять раз больше, чем размер синаптических пузырьков (Fields, 17). Их также называют «пузырьками» и «транспортными пузырьками».
Производится «эндоплазматическим ретикулумом», «аппаратом Гольджи» и клеточной мембраной. Везикулы передающей клетки сливаются с ее мембраной, высвобождая химические регуляторы посредством «экзоцитоза» во «внеклеточную жидкость».»(Norton Lectures, 6/2/09) Клеточная структура нервной системы настолько миниатюризирована, что длина волны видимого света слишком мала, чтобы ее исследовать. Длина волны зеленого света в десять раз больше, чем размер синаптических пузырьков (Fields, 17). Их также называют «пузырьками» и «транспортными пузырьками».
Тело клетки : увеличенная часть нейрона, содержащая «ядро». (OxfordMed) Участвует в важных хозяйственных функциях, таких как хранение генетического материала и создание белков и других молекул, необходимых для выживания клетки.(LeDoux, 40) Наиболее заметной особенностью является ядро с тонкой «хроматиновой» сеткой и четко выраженным «ядрышком». «Цитоплазма» богата свободными «рибосомами» и «грубым эндоплазматическим ретикулумом». Синтез белка происходит на рибосомах для использования в «цитозоле» и на шероховатом эндоплазматическом ретикулуме для окончательной упаковки. «Комплекс Гольджи» отвечает за модификацию и упаковку различных белков, ферментов и молекул химических посредников, производимых на шероховатом эндоплазматическом ретикулуме.Энергетическая потребность нейрона удовлетворяется наличием множества «митохондрий», которые распределены по всему телу клетки. Таким образом, в этой области происходят «синтез белка», «дыхание» и многие важные клеточные функции. (Патестас, 29) Произведенный материал (здесь) транспортируется с помощью «микротрубочек» к аксону для его использования. Материал может (также) перемещаться в противоположном направлении к телу клетки нейрона. (Patestas, 30-31) Также упоминается как «сома», «нервное тело», «тело нейрона» и «тело нейрона».’
Синтез белка происходит на рибосомах для использования в «цитозоле» и на шероховатом эндоплазматическом ретикулуме для окончательной упаковки. «Комплекс Гольджи» отвечает за модификацию и упаковку различных белков, ферментов и молекул химических посредников, производимых на шероховатом эндоплазматическом ретикулуме.Энергетическая потребность нейрона удовлетворяется наличием множества «митохондрий», которые распределены по всему телу клетки. Таким образом, в этой области происходят «синтез белка», «дыхание» и многие важные клеточные функции. (Патестас, 29) Произведенный материал (здесь) транспортируется с помощью «микротрубочек» к аксону для его использования. Материал может (также) перемещаться в противоположном направлении к телу клетки нейрона. (Patestas, 30-31) Также упоминается как «сома», «нервное тело», «тело нейрона» и «тело нейрона».’
Дендриты : напоминают густые ветвистые заросли. (RamachandranTTB, 14) Широко разветвляется, образуя древовидную структуру, которая вырастает из тела клетки и распространяется по большой площади.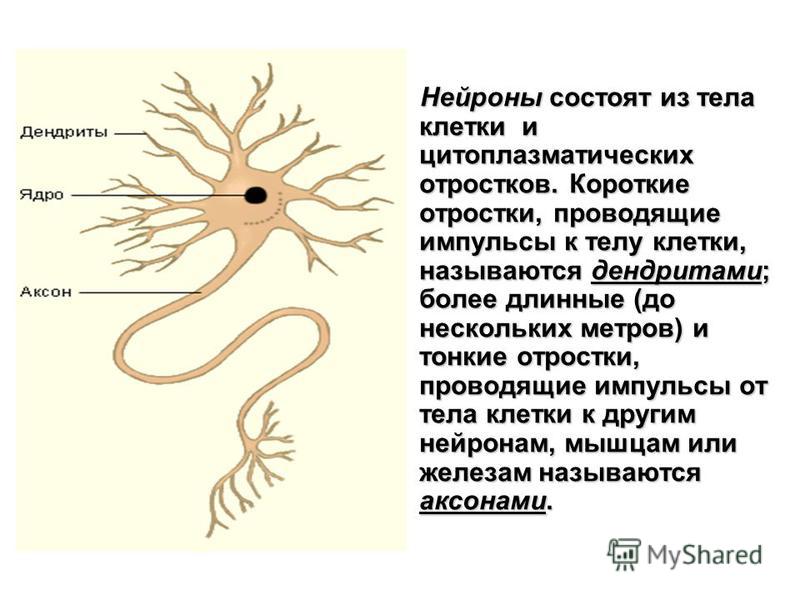 Обычно возникает на противоположной от аксона стороне тела клетки. (Кандел, 64) Часто обнаруживается, что дендриты замысловато переплетаются с другими дендритами, хотя обычно они не соприкасаются друг с другом. (The Brain-Francis Crick, 132). Самые тонкие ветви нейронов имеют диаметр менее одной десятой «микрона», что меньше длины волны видимого света.(Cerebrum2009, 70) Получает входные данные от других нейронов. (Doidge, 52) Поскольку дендриты обычно представляют собой сильно разветвленные структуры, они могут одновременно получать информацию из множества различных источников. (Патестас, 30) Они также участвуют в «петлях обратной связи». (CampbellVA, 144) Иногда общаются друг с другом. (LeDoux, 41)
Обычно возникает на противоположной от аксона стороне тела клетки. (Кандел, 64) Часто обнаруживается, что дендриты замысловато переплетаются с другими дендритами, хотя обычно они не соприкасаются друг с другом. (The Brain-Francis Crick, 132). Самые тонкие ветви нейронов имеют диаметр менее одной десятой «микрона», что меньше длины волны видимого света.(Cerebrum2009, 70) Получает входные данные от других нейронов. (Doidge, 52) Поскольку дендриты обычно представляют собой сильно разветвленные структуры, они могут одновременно получать информацию из множества различных источников. (Патестас, 30) Они также участвуют в «петлях обратной связи». (CampbellVA, 144) Иногда общаются друг с другом. (LeDoux, 41)
Дендритный шип : маленькие выступы, отходящие от дендритов. Особенно важно как получатели сообщений от аксонов. (LeDoux, 41-42) Тонкое трубчатое расширение нейрона.Дендриты и аксоны начинают развиваться во время (беременности). Дендриты начинают прорастать в процессе, называемом «ветвлением». (Goldberg, 40)
(Goldberg, 40)
Постсинаптическая мембрана : мембрана на входной стороне передатчика синапса. (Kolb, 153) Мембрана постсинаптической клетки, обогащенная рецепторами нейротрансмиттеров. (NCIt)
Клеточные составляющие мозга
Мозг состоит из нейронов и глии. Хотя большая часть нашего обсуждения будет касаться функций нейронов, растет признание роли глиальных клеток в поддержании важнейших функций мозга.Следовательно, нам нужно будет сказать несколько слов и о глии.
Нейрон
Основным строительным блоком нервной системы является нейрон. Каждый нейрон имеет тело клетки, дендриты и один или несколько аксонов. Тело клетки, также называемое сомой или перикарионом, содержит метаболический аппарат и генетический материал для нейрона. Дентриты являются основным местом синапсов с другими нейронами, часто в выступах, называемых дендритными шипами. Аксон, начинающийся от бугорка аксона, представляет собой процесс, который передает электрический сигнал от нейрона, и его часто называют нервным волокном. Дендриты и аксоны обычно имеют много ответвлений, а аксо-дендритные синапсы являются наиболее частым местом связи между нейронами.
Дендриты и аксоны обычно имеют много ответвлений, а аксо-дендритные синапсы являются наиболее частым местом связи между нейронами.
Мембранный потенциал покоя
Нейрон поддерживает мембранный потенциал покоя. Этот потенциал основан на градиенте концентрации ионов, который создается зависящим от энергии насосом натрий-калиевой АТФазы. Этот насос перемещает 2 иона калия в нейрон в обмен на 3 иона натрия, выходящих из клетки. Таким образом, этот насос непосредственно создает небольшой электрический потенциал, при этом внешняя часть ячейки является слегка положительной.Насос считается электрогенным. Однако большая часть мембранного потенциала покоя возникает не напрямую из этого эффекта самого насоса, а скорее из силы градиента концентрации, создаваемого насосом. Поскольку калий находится в высокой концентрации внутри нейрона, существует градиент концентрации, который пытается вытеснить калий из клетки. Подобные силы будут пытаться переместить ионы натрия в нейрон.
Мембрана нейрона представляет собой липидный бислой со встроенными белками. Некоторые из этих белков являются насосами (например, натрий-калиевый насос АТФазы), но многие из них являются ионными каналами. Эти ионные каналы представляют собой трансмембранные белки со сложной конфигурацией, которая включает в себя поры, через которые могут проходить ионы. Некоторые каналы позволяют постоянно проходить ионам («каналы утечки»), в то время как некоторые открываются при изменении напряжения («управляемые по напряжению») или химическими веществами («управляемые лигандом»). Большинство каналов обладают избирательностью, позволяя проходить только определенным ионам, когда они открыты. Эта селективность обусловлена особой конформацией белка или расположением зарядов в ионной поре.
Некоторые из этих белков являются насосами (например, натрий-калиевый насос АТФазы), но многие из них являются ионными каналами. Эти ионные каналы представляют собой трансмембранные белки со сложной конфигурацией, которая включает в себя поры, через которые могут проходить ионы. Некоторые каналы позволяют постоянно проходить ионам («каналы утечки»), в то время как некоторые открываются при изменении напряжения («управляемые по напряжению») или химическими веществами («управляемые лигандом»). Большинство каналов обладают избирательностью, позволяя проходить только определенным ионам, когда они открыты. Эта селективность обусловлена особой конформацией белка или расположением зарядов в ионной поре.
Мембрана в состоянии покоя является полупроницаемой с гораздо большей проницаемостью для калия, чем для натрия (она содержит «каналы утечки» калия). Следовательно, градиент концентрации калия заставит некоторые положительно заряженные ионы калия покидать клетку. Это первичный генератор разности напряжений на мембране, называемой «мембранным потенциалом покоя». Этот потенциал обычно находится между -70 и -90 милливольт, при этом внутренняя часть ячейки отрицательна, потому что положительные ионы калия покинули ячейку.Конечно, этот потенциал будет зависеть от силы, оказываемой градиентом концентрации. По мере того, как калий покидает нейрон, внутренняя часть клетки становится все более отрицательной, что будет притягивать положительные ионы калия, предотвращая дальнейший отток. Если эта электрическая сила достаточно велика, она действительно будет втягивать ионы калия снаружи клетки обратно внутрь.
Этот потенциал обычно находится между -70 и -90 милливольт, при этом внутренняя часть ячейки отрицательна, потому что положительные ионы калия покинули ячейку.Конечно, этот потенциал будет зависеть от силы, оказываемой градиентом концентрации. По мере того, как калий покидает нейрон, внутренняя часть клетки становится все более отрицательной, что будет притягивать положительные ионы калия, предотвращая дальнейший отток. Если эта электрическая сила достаточно велика, она действительно будет втягивать ионы калия снаружи клетки обратно внутрь.
Ионы калия будут покидать нейрон до тех пор, пока сила электрического притяжения развивающегося отрицательного заряда внутри клетки не сравняется с силой, создаваемой градиентом концентрации, вытесняющим калий.Имейте в виду, что для создания значительной электрической силы не требуется движения многих положительно заряженных ионов калия, поэтому общий градиент концентрации не сильно меняется при этом движении. В точке, где внутренние и внешние силы уравновешиваются, нейрон находится в равновесии, и один ион калия перемещается в нейрон на каждый уходящий.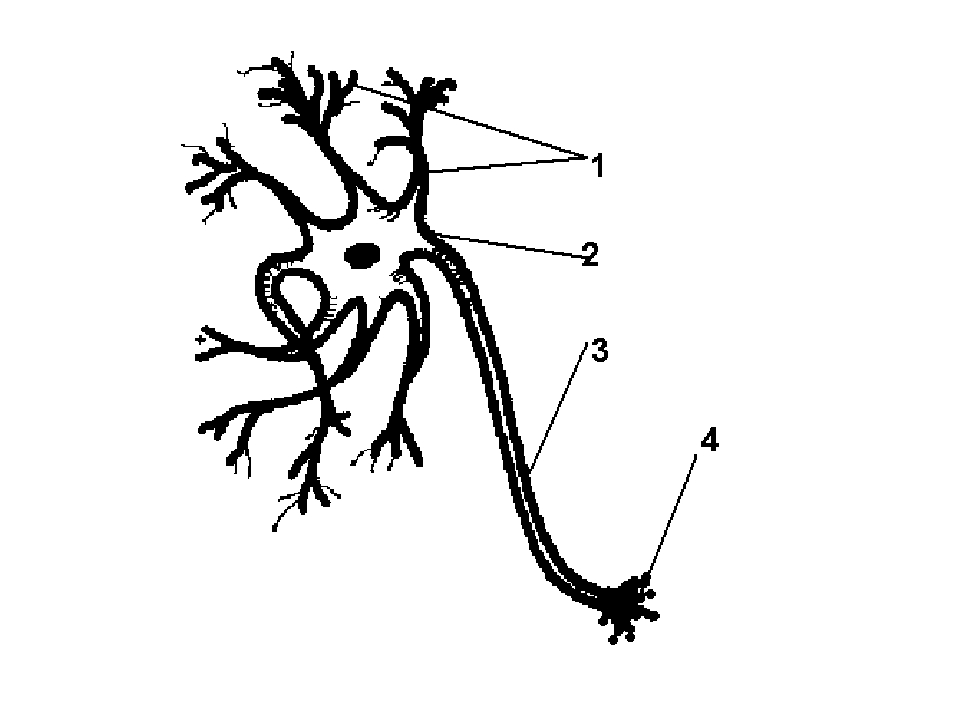 Трансмембранное напряжение, при котором возникает этот баланс, называется «равновесным потенциалом калия» и может быть рассчитано по уравнению Нернста (рисунок 1).Это уравнение в основном говорит о том, что равновесный потенциал будет произведением константы, умноженной на натуральный логарифм соотношений концентраций вне и внутри клетки. Мембранный потенциал может быть создан только ионами, которые действительно могут пересечь мембрану. Поскольку в случае нейронов в состоянии покоя это калий, равновесный потенциал калия почти равен измеренному мембранному потенциалу покоя. Однако следует иметь в виду, что движение других ионов дает очень небольшой вклад в мембранный потенциал.Этот вклад пропорционален их очень низкой проницаемости. Уравнение Гольдмана-Ходжкина-Каца (рис. 1) принимает во внимание эти другие ионы, хотя, если проницаемость для иона очень мала по сравнению с калием, ион можно в основном игнорировать при рассмотрении мембранного потенциала покоя. Важно понимать мембранный потенциал покоя, поскольку это необходимо для понимания того, что происходит, когда нейрон возбуждается.
Трансмембранное напряжение, при котором возникает этот баланс, называется «равновесным потенциалом калия» и может быть рассчитано по уравнению Нернста (рисунок 1).Это уравнение в основном говорит о том, что равновесный потенциал будет произведением константы, умноженной на натуральный логарифм соотношений концентраций вне и внутри клетки. Мембранный потенциал может быть создан только ионами, которые действительно могут пересечь мембрану. Поскольку в случае нейронов в состоянии покоя это калий, равновесный потенциал калия почти равен измеренному мембранному потенциалу покоя. Однако следует иметь в виду, что движение других ионов дает очень небольшой вклад в мембранный потенциал.Этот вклад пропорционален их очень низкой проницаемости. Уравнение Гольдмана-Ходжкина-Каца (рис. 1) принимает во внимание эти другие ионы, хотя, если проницаемость для иона очень мала по сравнению с калием, ион можно в основном игнорировать при рассмотрении мембранного потенциала покоя. Важно понимать мембранный потенциал покоя, поскольку это необходимо для понимания того, что происходит, когда нейрон возбуждается.
Конечно, все клетки тела обладают мембранным потенциалом покоя.Этот потенциал покоя позволяет клеткам выполнять такую работу, как перенос молекул через свои мембраны. Однако нервные и мышечные клетки в некоторой степени уникальны тем, что обладают свойством возбудимости. Эта возбудимость означает, что они могут изменять свой потенциал из-за внешних химических или механических факторов.
Потенциал действия
Когда мы обсуждаем электрические изменения в нейрональной мембране, это изменение происходит от начальной точки мембранного потенциала покоя, когда внутренняя часть клетки отрицательна, а внешняя — положительна.В состоянии покоя нейроны считаются «поляризованными», а «деполяризация» делает внутреннюю часть клетки менее отрицательной, а внешнюю — менее положительной. «Гиперполяризация» — это наоборот. Открытие любого ионного канала будет стремиться приблизить трансмембранное напряжение к равновесному потенциалу любых ионов, проницаемых через этот канал. Например, если натриевой канал был открыт, мембранный потенциал сдвинулся бы к равновесному потенциалу натрия, при этом внутренняя часть клетки была слегка положительной по отношению к внешней.Опять же, это не требует перемещения очень большого количества положительно заряженных ионов в ячейку и может происходить довольно быстро, когда канал открыт. Термин «реполяризация» применяется к процессу восстановления мембранного потенциала покоя после события, которое деполяризует клетку.
Например, если натриевой канал был открыт, мембранный потенциал сдвинулся бы к равновесному потенциалу натрия, при этом внутренняя часть клетки была слегка положительной по отношению к внешней.Опять же, это не требует перемещения очень большого количества положительно заряженных ионов в ячейку и может происходить довольно быстро, когда канал открыт. Термин «реполяризация» применяется к процессу восстановления мембранного потенциала покоя после события, которое деполяризует клетку.
Определенные ионные каналы в нейрональной мембране быстро открываются, когда мембранный потенциал достигает определенного напряжения. Эти каналы называются стробируемыми по напряжению. Конкретное напряжение, при котором открывается канал, называется «пороговым потенциалом», при котором конформация трансмембранных белков, составляющих ионный канал, внезапно меняет свою конформацию.Затем канал становится проницаемым для всех ионов, проводимых этим конкретным каналом с регулируемым напряжением. Поскольку многие из этих потенциалзависимых каналов проницаемы для натрия, который находится в высокой концентрации за пределами нейрона, происходит быстрое перемещение нескольких ионов натрия в нейрон, которое меняет обычно отрицательный потенциал внутри клетки на небольшой положительный. . Эта быстрая деполяризация называется «потенциалом действия» (см. Рисунок 2).
. Эта быстрая деполяризация называется «потенциалом действия» (см. Рисунок 2).
Открытые потенциалозависимые натриевые каналы быстро закрываются — процесс, называемый инактивацией.Это завершает фазу деполяризации или нарастания потенциала действия. Кроме того, открываются калиевые каналы, управляемые напряжением, что вызывает реполяризацию к равновесному потенциалу калия за счет перемещения калия из клетки. Фактически это вызывает небольшую гиперполяризацию. В течение короткого периода инактивации натриевых каналов эти каналы не могут быть снова открыты и мембрана не может быть деполяризована. Это называется периодом абсолютной рефрактерности. В течение периода гиперполяризации, который следует за потенциалом действия, требуется большой стимул для генерации другого потенциала действия, и этот период называется относительным рефрактерным периодом.
Потенциал действия вызывает большое изменение напряжения части нейрона. Поскольку ионы распространяются через цитоплазму и внеклеточную жидкость, потенциал действия также деполяризует соседние области мембраны. Эта деполяризация открывает там ионные каналы, управляемые напряжением, и потенциал действия восстанавливается в этих соседних участках. Этот потенциал действия затем распространяется по нервной клетке и восстанавливается во всех областях, где есть потенциалзависимые ионные каналы. Это включает распространение по аксону.Поскольку потенциал регенерируется на каждом уровне нейрона, потенциал действия может увеличивать длину аксона и распространяться на все ветви без уменьшения размера. Это называется «распространением» потенциала действия. Повреждение мембраны аксона или токсины, которые влияют на ионные каналы, будут влиять на генерацию потенциала действия или проведение импульса.
Эта деполяризация открывает там ионные каналы, управляемые напряжением, и потенциал действия восстанавливается в этих соседних участках. Этот потенциал действия затем распространяется по нервной клетке и восстанавливается во всех областях, где есть потенциалзависимые ионные каналы. Это включает распространение по аксону.Поскольку потенциал регенерируется на каждом уровне нейрона, потенциал действия может увеличивать длину аксона и распространяться на все ветви без уменьшения размера. Это называется «распространением» потенциала действия. Повреждение мембраны аксона или токсины, которые влияют на ионные каналы, будут влиять на генерацию потенциала действия или проведение импульса.
Распространение потенциала действия по аксону с регенерацией на каждом соседнем участке происходит довольно медленно, обычно менее метра в секунду.Это немного быстрее в аксоне большего диаметра, но все же довольно медленно. Миелинизация — это метод увеличения скорости проводимости. Миелинизация — это процесс плотного обертывания слоев клеточной мембраны вокруг аксона.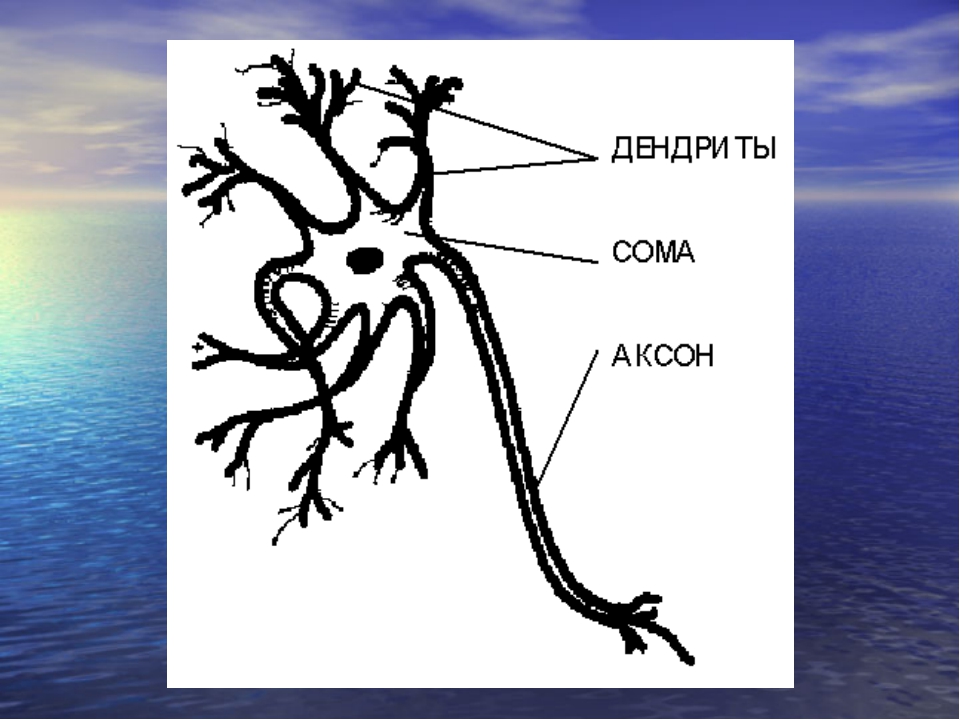 Это имеет функцию предотвращения прохождения ионов через эти части аксональных мембран, а также уменьшения емкости мембраны за счет большего разделения отрицательных электрических зарядов внутри от положительных зарядов снаружи мембраны.
Это имеет функцию предотвращения прохождения ионов через эти части аксональных мембран, а также уменьшения емкости мембраны за счет большего разделения отрицательных электрических зарядов внутри от положительных зарядов снаружи мембраны.
В центральной нервной системе миелиновая оболочка развивается из олигодендроглиальных отростков.В периферической нервной системе каждый сегмент миелина является результатом обертывания отдельной клетки, называемой шванновской клеткой. Между сегментами миелина, узлами Ранвье, есть промежутки. Это места высокой концентрации потенциалзависимых ионных каналов. Поскольку движение ионов через мембрану там, где присутствует миелиновая оболочка, невелико, и из-за низкой концентрации заряда вдоль мембраны в межузловом сегменте (из-за низкой емкости) ионы могут относительно свободно перемещаться по аксону.Это вызывает достаточную деполяризацию в следующем узле Ранвье без необходимости регенерировать потенциал действия в каждой последующей части аксона. Это заметно увеличивает скорость проводимости аксона, при этом расстояние между узлами прямо пропорционально степени увеличения скорости. Повреждение миелина значительно замедляет или даже может блокировать аксональную проводимость. Самые большие моторные аксоны — это те, которые соединяют нервную систему со скелетными мышцами и проводят от 50 до 100 метров в секунду.Вегетативные постганглионарные моторные аксоны, ведущие к железам, органам и кровеносным сосудам, немиелинизированы и проводят очень медленно (обычно менее 1 метра в секунду). Ощущение прикосновения, давления и растяжения мышц передается через крупные миелинизированные волокна, а ощущение боли и температуры — через небольшие миелинизированные или немиелинизированные сенсорные волокна.
Это заметно увеличивает скорость проводимости аксона, при этом расстояние между узлами прямо пропорционально степени увеличения скорости. Повреждение миелина значительно замедляет или даже может блокировать аксональную проводимость. Самые большие моторные аксоны — это те, которые соединяют нервную систему со скелетными мышцами и проводят от 50 до 100 метров в секунду.Вегетативные постганглионарные моторные аксоны, ведущие к железам, органам и кровеносным сосудам, немиелинизированы и проводят очень медленно (обычно менее 1 метра в секунду). Ощущение прикосновения, давления и растяжения мышц передается через крупные миелинизированные волокна, а ощущение боли и температуры — через небольшие миелинизированные или немиелинизированные сенсорные волокна.
Синапс
Большинство аксонов оканчиваются «синапсом» на дендритах других нейронов, часто на небольших дендритных шипах. Синапс — это место передачи между нейронами.Хотя существуют редкие синапсы типа щелевых соединений, в которых существует реальный поток ионов между нейронами, подавляющее большинство синапсов являются «химическими синапсами». Они включают высвобождение нейромедиатора окончанием аксона, когда потенциал действия достигает его. Терминал аксона содержит пузырьки или «кванты» нейромедиатора, а высвобождение медиатора из синаптических пузырьков происходит в многоэтапном процессе. Потенциал действия открывает управляемые напряжением кальциевые каналы на нервном окончании, что приводит к притоку кальция, потому что кальций находится в более высокой концентрации вне клеток.Кальций в нервном окончании приводит к серии химических взаимодействий между белками в нервном окончании, что в конечном итоге приводит к мобилизации, связыванию и последующему слиянию везикул нейротрансмиттера с пресинаптической мембраной аксона. Это слияние приводит к высвобождению содержимого в синаптическую щель. Некоторые нервные окончания высвобождают содержимое сотен пузырьков каждый раз, когда потенциал действия достигает нервного окончания. Это верно в отношении нервно-мышечного соединения. Некоторые нервные окончания высвобождают только один пузырек с деполяризацией.
Они включают высвобождение нейромедиатора окончанием аксона, когда потенциал действия достигает его. Терминал аксона содержит пузырьки или «кванты» нейромедиатора, а высвобождение медиатора из синаптических пузырьков происходит в многоэтапном процессе. Потенциал действия открывает управляемые напряжением кальциевые каналы на нервном окончании, что приводит к притоку кальция, потому что кальций находится в более высокой концентрации вне клеток.Кальций в нервном окончании приводит к серии химических взаимодействий между белками в нервном окончании, что в конечном итоге приводит к мобилизации, связыванию и последующему слиянию везикул нейротрансмиттера с пресинаптической мембраной аксона. Это слияние приводит к высвобождению содержимого в синаптическую щель. Некоторые нервные окончания высвобождают содержимое сотен пузырьков каждый раз, когда потенциал действия достигает нервного окончания. Это верно в отношении нервно-мышечного соединения. Некоторые нервные окончания высвобождают только один пузырек с деполяризацией.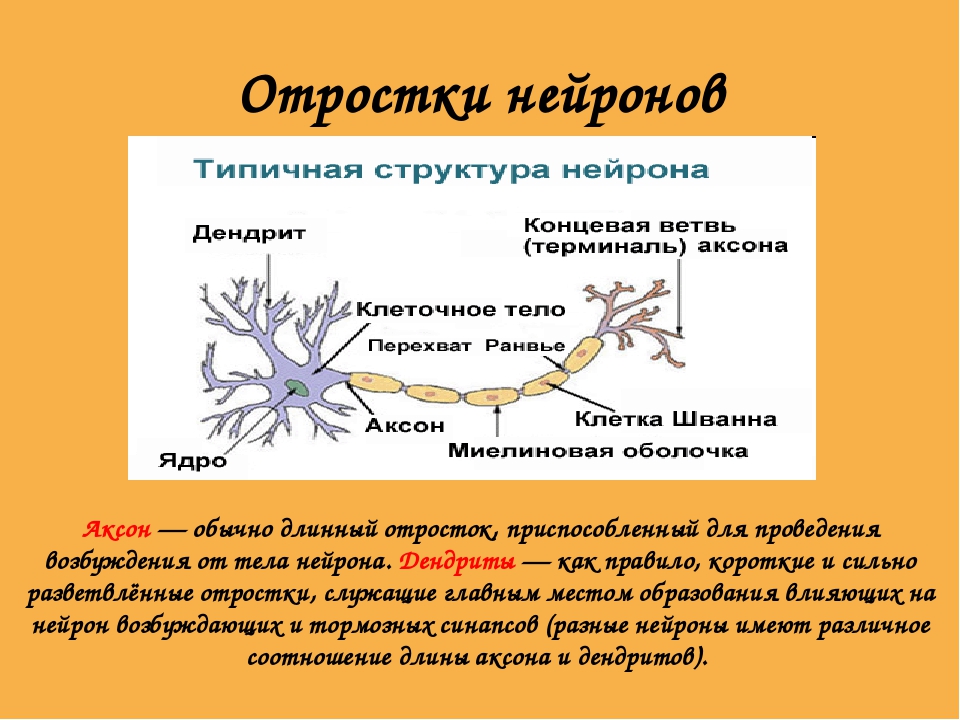 Это происходит на многих терминалах аксонов центральной нервной системы. В некоторых синапсах только каждый второй или третий потенциал действия фактически приводит к высвобождению везикулы нейромедиатора.
Это происходит на многих терминалах аксонов центральной нервной системы. В некоторых синапсах только каждый второй или третий потенциал действия фактически приводит к высвобождению везикулы нейромедиатора.
Нейромедиатор диффундирует через «синаптическую щель» на постсинаптическую сторону (обычно на дендрите), где передатчик связывается со специфическими рецепторами. Многие из этих рецепторов являются частью ионного канала, называемого ионным каналом, управляемым лигандом. Эти ионные каналы обладают очень специфической проницаемостью для определенных ионов.В зависимости от конкретной ионной проницаемости каналов рецептор может возбуждать или ингибировать постсинаптический нейрон. Есть и другие рецепторы, не являющиеся ионными каналами. Эти рецепторы, называемые «рецепторами вторичных мессенджеров», связаны с внутренними процессами внутри постсинаптического нейрона, такими как ферменты, которые могут изменять функцию нейрона. Многие из них связаны с g-белками, которые могут увеличивать внутриклеточную концентрацию таких соединений, как циклический АМФ, инозитолтрифосфат или диацилглицерин.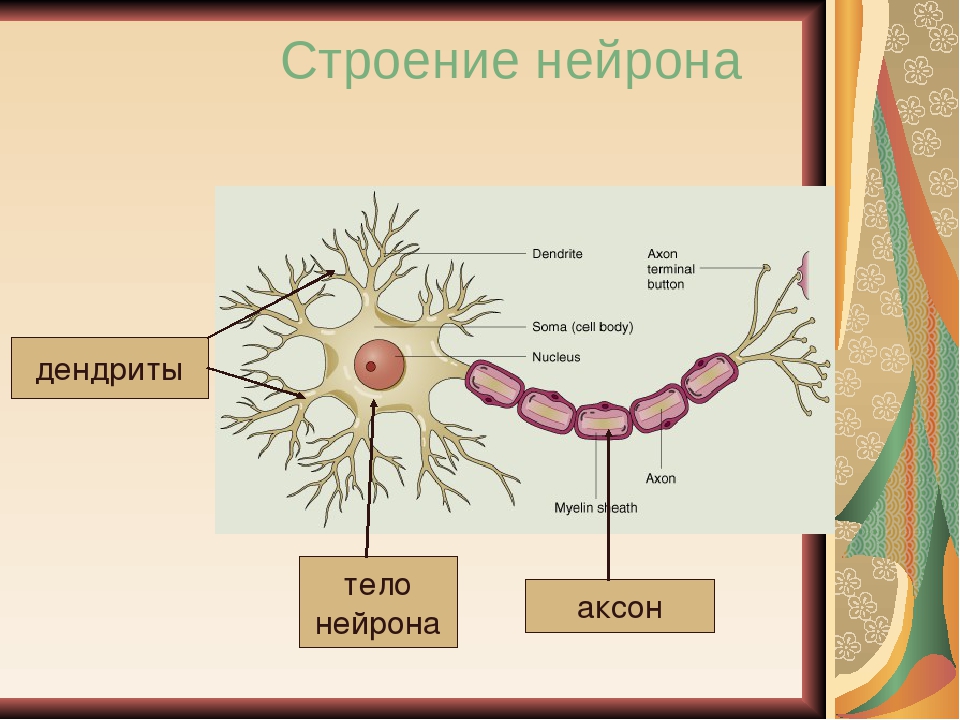 Поскольку эти рецепторы часто не влияют напрямую на мембранный потенциал, их часто описывают как модулирующие, а передатчики, которые связываются с этими рецепторами, часто называют «нейромодуляторами». Поскольку они могут изменять многие функции внутри клетки, их воздействие может быть очень длительным.
Поскольку эти рецепторы часто не влияют напрямую на мембранный потенциал, их часто описывают как модулирующие, а передатчики, которые связываются с этими рецепторами, часто называют «нейромодуляторами». Поскольку они могут изменять многие функции внутри клетки, их воздействие может быть очень длительным.
Действие нейромедиатора прекращается одним из нескольких механизмов. Могут присутствовать ферменты, расщепляющие нейротрансмиттер, например, действие ацетилхолинэстеразы на ацетилхолин.Блокада ацетилхолинэстеразы лекарствами или токсинами продлит эффект высвобождения ацетилхолина. Другая возможность заключается в том, что передатчик может диффундировать от синаптической щели, обычно для поглощения глиальными клетками вокруг этой области. Третья возможность состоит в том, что передатчик может быть доставлен обратно в нервный конец через определенные транспортные механизмы. В этом случае передатчик может быть переупакован для более позднего выпуска. На этот процесс влияют такие препараты, как селективные ингибиторы обратного захвата серотонина.
В случае рецептора типа ионного канала, управляемого лигандом, эффект открытия канала зависит от конкретных ионов, которые могут проходить через него. В любом случае открытие этих каналов будет приводить мембранный потенциал к равновесному потенциалу для ионов, для которых он проницаем. Это было названо «обратным потенциалом» рецептора. Если этот реверсивный потенциал находится на уровне, который может привести к достижению мембраной порогового значения, он называется возбуждающим рецептором, поскольку при открытии достаточного количества этих каналов будет генерироваться потенциал действия.Например, любой рецептор, управляемый лигандом, который в первую очередь проницаем для натрия или кальция, будет возбуждающим. Если обратный потенциал рецептора находится на уровне, не достигающем порогового значения, это будет рецептор ингибирования. Открытие этого типа канала предотвратит достижение нейроном порога и, следовательно, не позволит ему генерировать потенциал действия. Например, открытие рецептора, проницаемого для калия, будет тормозящим.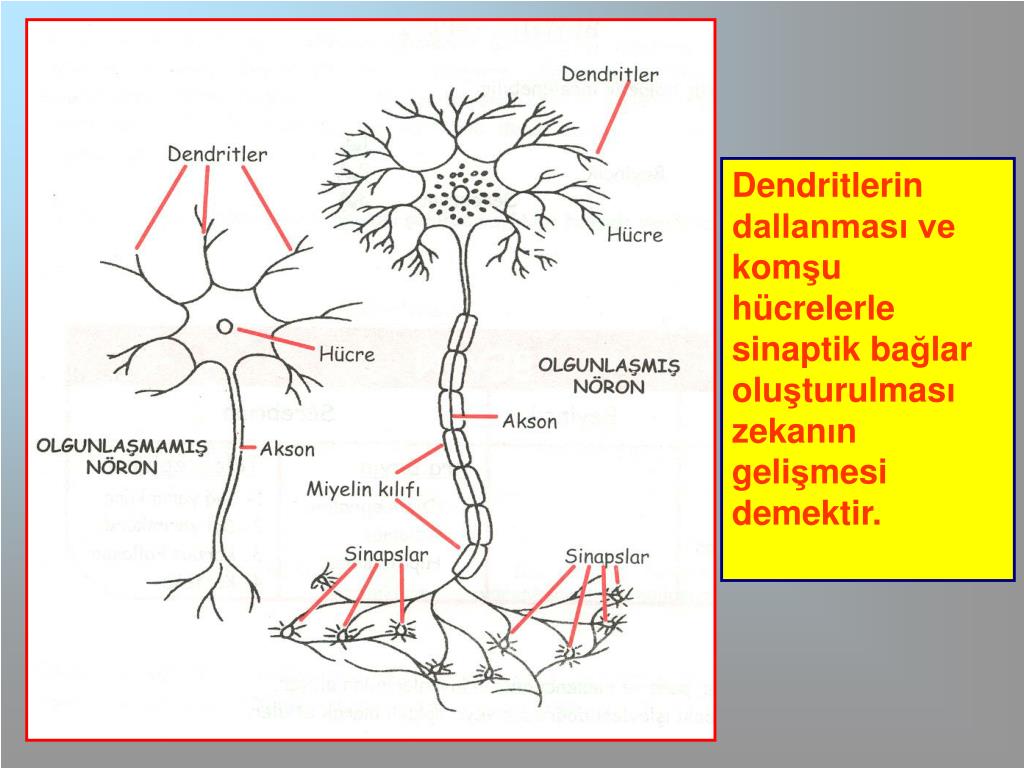
Одна концепция, которую трудно понять поначалу, заключается в том, что передатчик может приводить к деполяризации мембраны зрения, но все же быть ингибирующим рецептором.Это могло бы произойти, если бы реверсивный потенциал рецептора был более положительным, чем мембранный потенциал покоя, но все же более отрицательным, чем пороговое значение для нейрона. Хотя открытие этого типа рецептора слегка деполяризует нейрон, оно будет иметь тенденцию удерживать нейрон на этом потенциале и сопротивляться дальнейшей деполяризации к порогу. Следовательно, это ингибитор генерации потенциала действия. Так обстоит дело с открытием хлоридных каналов в нервной системе взрослого человека. Наиболее распространенным ингибирующим рецептором в головном мозге является рецептор ГАМКА, который связывает гамма-аминомасляную кислоту и проницаем для хлоридов.Этот рецептор особенно важен, поскольку многие ингибирующие препараты модулируют активность этого рецептора, включая алкоголь, бензодиазепины и барбитураты.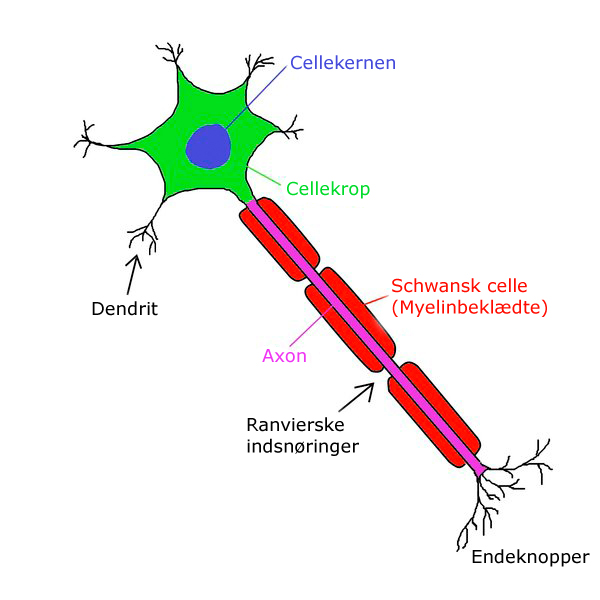
Если совокупная активность тысяч синапсов приводит нейрон к пороговому значению в процессе суммирования, генерируется потенциал действия, и процесс передачи нервного импульса начинается снова. На дендритах очень мало каналов с регулируемым напряжением, и поэтому потенциалы действия обычно не начинаются там.Управляемые напряжением каналы сгруппированы в области холмика аксона в начале аксона, который является типичным местом генерации потенциала действия. Деполяризация дендритов должна распространяться на бугорок аксона, чтобы суммироваться в потенциал действия. Поскольку эффект этой деполяризации должен распространяться по нейрону, он становится все меньше и меньше, чем дальше он должен пройти. Это называется «электротоническим распространением» деполяризации и означает, что синапс, расположенный дальше на дендрите, будет производить меньшие изменения на бугорке аксона, чем аналогичный синапс рядом с сомой.
Глия
Глиальные клетки важны для мозга. Мы уже видели, как олигодендроциты составляют миелин центральной нервной системы. Астроциты окружают нейроны и кровеносные сосуды. Они отвечают за критическую функцию поддержания нормальной ионной и питательной среды вокруг нейронов. Они удаляют ионы и передатчики, которые накапливаются из-за активности нейронов, и обеспечивают идеальную внеклеточную среду для нейрона. Они выделяют факторы роста и цитокины, важные для здоровья нейронов.Наконец, они стимулируют капилляры головного мозга к развитию плотных контактов и других специализаций эндотелиальных клеток, которые вносят вклад в гематоэнцефалический барьер (см. Раздел XIIb).
Астроциты окружают нейроны и кровеносные сосуды. Они отвечают за критическую функцию поддержания нормальной ионной и питательной среды вокруг нейронов. Они удаляют ионы и передатчики, которые накапливаются из-за активности нейронов, и обеспечивают идеальную внеклеточную среду для нейрона. Они выделяют факторы роста и цитокины, важные для здоровья нейронов.Наконец, они стимулируют капилляры головного мозга к развитию плотных контактов и других специализаций эндотелиальных клеток, которые вносят вклад в гематоэнцефалический барьер (см. Раздел XIIb).
Эпендимные клетки — это тип глиальных клеток, выстилающих желудочковую систему. Эти клетки не создают барьер между спинномозговой жидкостью и мозгом. Следовательно, вещества, доставленные в спинномозговую жидкость, имеют прямой доступ к головному и спинному мозгу.
.
Leave A Comment