
Споры грибов
Споры грибов, как правило, являются наименее изменяющимися структурами гриба. Споры по форме, размерам и целому ряду других признаков оказываются для каждого вида относительно постоянными, поэтому при классификации грибных организмов постоянно обращается внимание на строение спор. Известно много различных видов спор. К их числу относятся зооспоры, спорангиоспоры, конидии, хламидоспоры, цисты, зигоспоры, ооспоры, аскоспоры, уредоспоры, эцидиоспоры, телейтоспоры, базидиоспоры и др. Не все споры служат для размножения. Некоторые из них являются покоящимися, с их помощью грибы переносят неблагоприятные условия, например зиму. К ним относятся цисты, зигоспоры, ооспоры, телейтоспоры и др. Однако споры некоторых видов головневых грибов служат как для распространения, так и для перезимовки (например, хламидоспоры пыльной головни овса — Ustilago avenae).
Грибы образуют огромное количество спор. Проводилось много подсчетов количества спор, образующихся разными видами грибов. Так, один экземпляр гигантского дождевика Calvatia gigatitea содержит 7Х1012 спор. Трутовый гриб Ganoderma applanatum может освобождать до 30X109 спор в день, причем такая продуктивность спор может сохраняться в течение всего периода спорообразования, длившегося до 6 месяцев. Колония плесневого гриба Penicillium, с диаметром в 2,5 см, может образовывать до 400X106 конидий. Но это, однако, не означает, что все образующиеся споры грибов воспроизводят потомство. При наличии огромного количества спор у грибов потери репродуктивных элементов также достигают огромных размеров, ибо только отдельным спорам одного экземпляра гриба удается выполнить свою функцию, т. е. прорасти и дать новое потомство.
Так, один экземпляр гигантского дождевика Calvatia gigatitea содержит 7Х1012 спор. Трутовый гриб Ganoderma applanatum может освобождать до 30X109 спор в день, причем такая продуктивность спор может сохраняться в течение всего периода спорообразования, длившегося до 6 месяцев. Колония плесневого гриба Penicillium, с диаметром в 2,5 см, может образовывать до 400X106 конидий. Но это, однако, не означает, что все образующиеся споры грибов воспроизводят потомство. При наличии огромного количества спор у грибов потери репродуктивных элементов также достигают огромных размеров, ибо только отдельным спорам одного экземпляра гриба удается выполнить свою функцию, т. е. прорасти и дать новое потомство.
Споры грибов по строению и внешнему облику можно разделить на две большие группы — подвижные и неподвижные споры. Подвижные споры — зооспоры — имеют вид голых комочков протоплазмы, лишенных оболочки, снабженных двигательным аппаратом, жгутиками. Зооспоры встречаются преимущественно у водных организмов, что вполне естественно, имея в виду приспособленность этих спор к передвижению в водной среде.
В силу этих обстоятельств и в силу того, что зооспоры встречаются у наименее организованных представителей грибов, они относительно мало разнообразны. Различия могут заключаться в форме зооспор, либо симметричных, обычно грушевидных, снабженных одним или двумя жгутиками, либо не симметричных — бобовидных. Как правило, жгутики грушевидных зооспор располагаются у заднего конца, передний закругленный конец у некоторых представителей может быть не закругленным, а амебообразным. У несимметричных (бобовидных) зооспор жгутики располагаются сбоку на вогнутой стороне. К этому и сводится все разнообразие подвижных спор — зооспор. Однако указанные в строении зооспор признаки являются более или менее постоянными и именно по наличию одного или двух жгутиков выделяют у низших грибов различные эволюционные ряды (одножгутиковый и двужгутиковый).
Неподвижные споры с самого начала их возникновения имеют оболочку и лишены двигательного аппарата. По форме, структуре и целому ряду других признаков неподвижные споры очень разнообразны. Любая неподвижная спора закладывается в самом начале как одноклеточное образование. Это простая клетка, в большинстве случаев либо правильно шаровидная, либо яйцевидная или эллипсоидальная. Очень часто такая форма споры сохраняется вплоть до ее вызревания. В процессе роста спора увеличивается в своих размерах, нередко сохраняя ту форму, которую она получила в самый момент возникновения. Однако во многих случаях в процессе роста форма споры может резко изменяться: будучи в самом начале эллипсоидальной или шаровидной, при полном вызревании спора может становиться цилиндрической или нитевидной, игловидной.
Любая неподвижная спора закладывается в самом начале как одноклеточное образование. Это простая клетка, в большинстве случаев либо правильно шаровидная, либо яйцевидная или эллипсоидальная. Очень часто такая форма споры сохраняется вплоть до ее вызревания. В процессе роста спора увеличивается в своих размерах, нередко сохраняя ту форму, которую она получила в самый момент возникновения. Однако во многих случаях в процессе роста форма споры может резко изменяться: будучи в самом начале эллипсоидальной или шаровидной, при полном вызревании спора может становиться цилиндрической или нитевидной, игловидной.
В ряде случаев споры приобретают сложную структуру. У некоторых несовершенных грибов мы наблюдаем споры — типичные трехлучевые или иногда четырех-пятилучевые. В других случаях споры грибов закручиваются в крутую спираль в несколько завитков. Такая спираль может быть с приподнятой вершиной и т. д.
Будучи вначале одноклеточным образованием, споры грибов в процессе созревания могут оказаться более сложными в своем строении. Так, вначале в спорах появляются поперечные перегородки, делящие споры на отдельные клетки. Таким образом, кроме одноклеточных спор имеются двухклеточные и многоклеточные с наличием поперечных и продольных перегородок.
Так, вначале в спорах появляются поперечные перегородки, делящие споры на отдельные клетки. Таким образом, кроме одноклеточных спор имеются двухклеточные и многоклеточные с наличием поперечных и продольных перегородок.
Для целого ряда спор характерно образование только поперечных перегородок. Однако в последующем у многих грибов как сумчатых, так и несовершенных появляются и продольные перегородки. В этих случаях получается структура, очень сходная с кирпичной кладкой, почему такие споры и носят название муральных спор.
Многоклеточные споры характерны только для сумчатых грибов и связанных с ними несовершенных грибов, а также наблюдаются у небольшой группы базидиомицетов. У всех остальных представителей грибов, — как правило, одноклеточные споры. Размеры спор различны — от 1—2 мкм в диаметре до 100 мкм и более. Однако огромное большинство спор имеет размеры в пределах от 5 до 50 мкм.
По химическому составу оболочка спор, по крайней мере у некоторых, но далеко не у всех низших грибов, представляет собой целлюлозу. Однако в последующем у более высокоорганизованных грибов к этой целлюлозе примешиваются и жироподобные вещества. У очень многих спор на поверхности оболочки имеется воск, что обусловливает несмачиваемость таких спор и т. д. В этом случае споровые оболочки довольно прочны по своему химическому составу и даже более или менее крепкие кислоты и щелочи с трудом воздействуют на споровую оболочку.
Однако в последующем у более высокоорганизованных грибов к этой целлюлозе примешиваются и жироподобные вещества. У очень многих спор на поверхности оболочки имеется воск, что обусловливает несмачиваемость таких спор и т. д. В этом случае споровые оболочки довольно прочны по своему химическому составу и даже более или менее крепкие кислоты и щелочи с трудом воздействуют на споровую оболочку.
Как правило, споровые оболочки являются по меньшей мере двухслойными. Здесь мы различаем так называемый эндоспорий, т. е. внутреннюю оболочку, и экзоспорий — поверхностную оболочку. В большинстве случаев они неясно отграничены друг от друга, и экзоспорий при обычном наблюдении сливается с эндоспорием. Однако при проращивании спор грибов или при воздействии на оболочку слабым раствором кислоты или щелочи можно обнаружить расслоение их на эндоспории и экзоспории.
Эндоспорий — в большинстве случаев тонкий, чаще всего бесцветный, экзоспорий может быть различным. У ряда грибов, имеющих бесцветные и тонкие оболочки спор, экзоспорий почти такого же строения, как и эндоспорий. В окрашенных спорах эта окраска сосредоточена в наружных слоях оболочки, что вполне естественно, если иметь в виду защитную роль таких окрасок: темные пигменты поглощают короткие лучи света и, таким образом, предохраняют споры от проникновения этих лучей внутрь протоплазмы живой клетки. На поверхности экзоспория часто наблюдаются различного рода щетинки, покрывающие всю поверхность споры, либо расположенные в отдельных местах споры; на поверхности споры могут возникать шипы, бугорки, бородавочки и т. д. В ряде случаев мы наблюдаем очень сложную скульптуру на поверхности спор. У некоторых представителей сумчатых грибов, и особенно часто у головневых грибов, имеется так называемая сетчатая скульптура поверхности оболочки, когда на оптическом разрезе такой споры видны ребра, довольно высоко выступающие за пределы споры, и углубленные ячейки, ограниченные по краям выступающими ребрами. Как форма, так и строение поверхности спор у разных представителей является чрезвычайно пестрой и разнообразной, однако для определенного вида грибов форма и строение спор являются постоянными.
В окрашенных спорах эта окраска сосредоточена в наружных слоях оболочки, что вполне естественно, если иметь в виду защитную роль таких окрасок: темные пигменты поглощают короткие лучи света и, таким образом, предохраняют споры от проникновения этих лучей внутрь протоплазмы живой клетки. На поверхности экзоспория часто наблюдаются различного рода щетинки, покрывающие всю поверхность споры, либо расположенные в отдельных местах споры; на поверхности споры могут возникать шипы, бугорки, бородавочки и т. д. В ряде случаев мы наблюдаем очень сложную скульптуру на поверхности спор. У некоторых представителей сумчатых грибов, и особенно часто у головневых грибов, имеется так называемая сетчатая скульптура поверхности оболочки, когда на оптическом разрезе такой споры видны ребра, довольно высоко выступающие за пределы споры, и углубленные ячейки, ограниченные по краям выступающими ребрами. Как форма, так и строение поверхности спор у разных представителей является чрезвычайно пестрой и разнообразной, однако для определенного вида грибов форма и строение спор являются постоянными.
В двухслойной оболочке спор иногда обнаруживаются утонченные места в виде ростковых пор. Ростковые поры особенно хорошо видны в уредоспорах и в телейтоспорах ржавчинных грибов, в спорах некоторых сумчатых грибов. Ростковые поры могут образоваться как за счет эндоспория, что особенно характерно для уредоспор ржавчинных грибов, так и за счет экзоспория. Количество ростковых пор в ряде случаев бывает совершенно определенным. Нередко близкие виды ржавчинных грибов отличаются друг от друга по уредоспорам, если учитывать количество ростковых пор и характер их распределения в оболочке. В телейтоспорах ржавчинных грибов в большинстве случаев ростковые споры располагаются за счет утонченных мест экзоспория. Утонченные места или поры иногда сверху прикрыты легко взбухающими веществами.
Внутреннее содержимое споры представлено протоплазменной массой. В молодом состоянии протоплазма заполняет всю полость споры, в более зрелом состоянии в протоплазменной массе мы обнаруживаем вакуоли — отдельные пустоты, заполненные клеточным соком.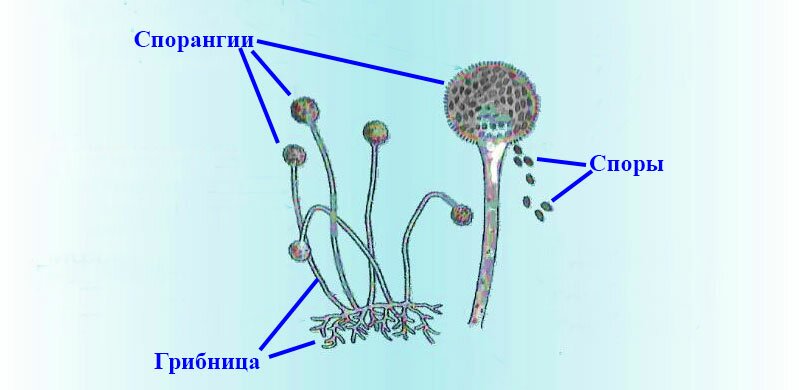 Внутри споры имеется запас питательных веществ. Из них наиболее часто в споре бывает гликоген, углевод, очень близкий по составу к крахмалу. Гликоген очень мелко распылен в протоплазменной массе. Кроме гликогена в спорах содержатся жиры, в одних случаях рассеянные мелкими каплями, в других — сосредоточенные в виде более или менее крупных капель. Расположение жира в спорах нередко более или менее постоянно: в эллипсоидальных спорах часто наблюдаются две капельки жира у полюсов споры.
Внутри споры имеется запас питательных веществ. Из них наиболее часто в споре бывает гликоген, углевод, очень близкий по составу к крахмалу. Гликоген очень мелко распылен в протоплазменной массе. Кроме гликогена в спорах содержатся жиры, в одних случаях рассеянные мелкими каплями, в других — сосредоточенные в виде более или менее крупных капель. Расположение жира в спорах нередко более или менее постоянно: в эллипсоидальных спорах часто наблюдаются две капельки жира у полюсов споры.
Запас питательных веществ в разных спорах может быть очень разнообразным. Наиболее богаты запасными питательными веществами споры, претерпевшие покой. В хламидоспорах внутренняя полость заполнена жировыми включениями наиболее обильно. Капельки жира в спорах либо совершенно бесцветны, либо в ряде случаев ярко окрашены, как, например, в спорах ржавчинных грибов.
Количество ядер в спорах также может быть очень различным. В большинстве случаев споры закладываются одноядерными. В спорангиях зигомицентов, которые развиваются на вегетативном мицелии, спорангноспоры всегда закладываются вокруг нескольких ядер. В то же время в спорангиях, вырастающих из зигоспор, споры закладываются как одноядерные и затем при созревании бывают многоядерными.
В то же время в спорангиях, вырастающих из зигоспор, споры закладываются как одноядерные и затем при созревании бывают многоядерными.
В сумках споры закладываются, как правило, вокруг одного ядра. Однако известны сумчатые грибы, у которых споры с самого начала развития бывают двухъядерными (Neurospora). В таком случае спора формируется вокруг двух ядер и является диплоидной.
У базидиальных грибов базидиоспоры, как правило, одноядерные; повторно они становятся иногда многоядерными, Что касается большинства сумчатых и связанных с ними несовершенных грибов, то здесь количество ядер в клетках может быть самым неопределенным — от одного до очень многих (иногда в одной клетке содержится до нескольких десятков ядер).
По биологическому значению мы различаем два типа спор: так называемые пропагативные, т. е. споры, предназначенные для массового размножения, и покоящиеся. К первой группе спор относятся зооспоры, спорангиоспоры, конидии, аскоспоры, базидиоспоры и др. Покоящимися спорами являются цисты, ооспоры, зигоспоры, телейтоспоры, хламидоспоры многих грибов и др. Эти две группы спор отличаются друг от друга рядом признаков. Прежде всего споры, предназначенные для размножения, характеризуются тем, что они всегда возникают в очень большом количестве. Кроме того, пропагативные споры при прорастании могут вновь давать споры. Это часто наблюдается у представителей рода Fusarium, когда многоклеточная спора, вырастая в вегетативный росток, затем сейчас же дает начало такой же конидии. Такая же картина наблюдается и при прорастании аскоспор Claviceps purpurea.
Эти две группы спор отличаются друг от друга рядом признаков. Прежде всего споры, предназначенные для размножения, характеризуются тем, что они всегда возникают в очень большом количестве. Кроме того, пропагативные споры при прорастании могут вновь давать споры. Это часто наблюдается у представителей рода Fusarium, когда многоклеточная спора, вырастая в вегетативный росток, затем сейчас же дает начало такой же конидии. Такая же картина наблюдается и при прорастании аскоспор Claviceps purpurea.
Для такого спорообразования необходим запас питательных веществ внутри самой споры. Как правило, пропагативные споры являются спорами гаплоидными. Они начинают гаплоидную генерацию и повторяются несколько раз в течение вегетационного периода.
Покоящиеся споры обладают, как правило, прямо противоположными свойствами: они образуются в малом количестве, в большинстве случаев возникают внутри субстрата и не имеют каких-либо приспособлений для распространения. Покоящиеся споры имеют назначение сохранения вида в неблагоприятный период (период перезимовки и т.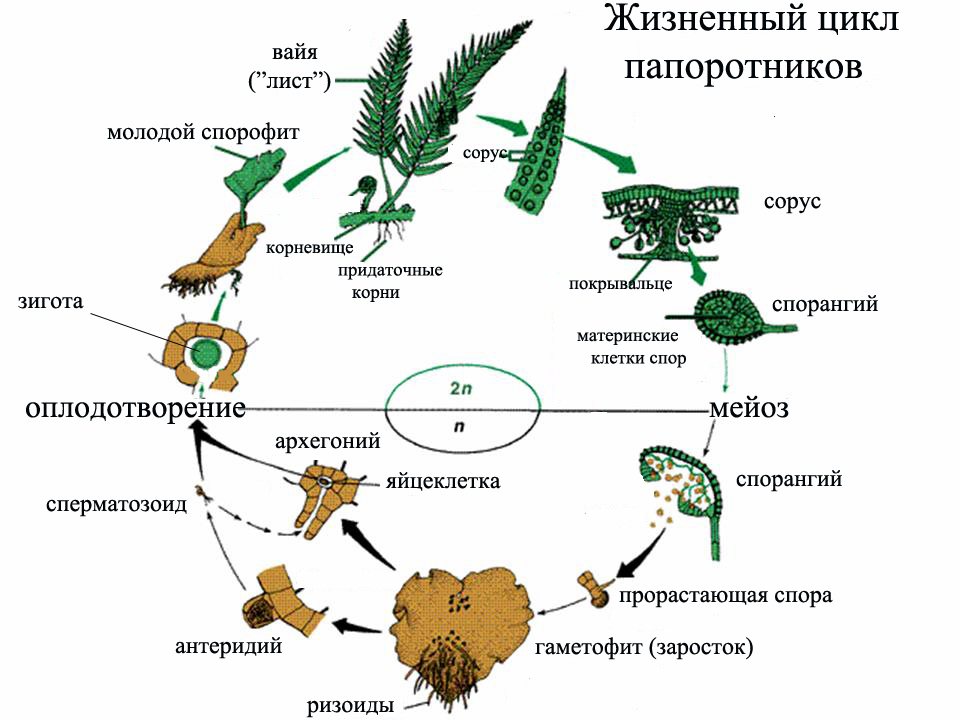 п.). Для них характерна способность сохраняться в покое довольно длительный промежуток времени. Так, некоторые хламидоспоры головневых грибов могут храниться десятки лет в условиях лаборатории. Мало того, для подавляющего большинства таких спор покой является необходимым, обязательным, и без него они, как правило, не прорастают.
п.). Для них характерна способность сохраняться в покое довольно длительный промежуток времени. Так, некоторые хламидоспоры головневых грибов могут храниться десятки лет в условиях лаборатории. Мало того, для подавляющего большинства таких спор покой является необходимым, обязательным, и без него они, как правило, не прорастают.
Хорошим примером являются телейтоспоры ржавчинных грибов. Собранные осенью вполне зрелые на вид телейтоспоры не удается прорастить, даже создав этим спорам благоприятные условия. Для телейтоспор ржавчинных грибов состояние покоя в естественной обстановке является обязательным. При этом на них воздействуют смены температур и влажности, которые в течение осени, зимы и весны мы наблюдаем в природе. При этих условиях телейтоспоры следующей весной оказываются способными прорастать.
Покоящиеся споры в силу этих особенностей заключают внутри себя, как правило, большой запас питательных веществ. Кроме того, покоящиеся споры являются диплоидными или иногда дикариотическими.
Одним из свойств пропагативных спор является способность прорастать сейчас же после образования. Для этих спор состояние покоя не требуется, они способны прорастать непосредственно вслед за созреванием. В ряде случаев у многих сумчатых грибов и у ряда несовершенных грибов пропагативные споры способны прорастать еще до полного созревания.
Пропагативные споры, как правило, не способны к длительному хранению. В большинстве случаев их способность прорастать сохраняется в течение небольшого промежутка времени. Это связано с запасом питательных веществ, с наличием более или менее плотной оболочки и с рядом других свойств. Споры, имеющие тонкую, прозрачную оболочку и не имеющие достаточного запаса питательных веществ, обычно способны сохраняться в течение нескольких дней. Например, зооспоры при отсутствии влаги погибают уже через несколько часов, неподвижные споры сохраняются до 10—12 дней.
Для пропагативных спор характерно, что их способность к прорастанию возрастает по мере созревания спор, достигает кульминационного пункта в период полной зрелости и затем постепенно падает.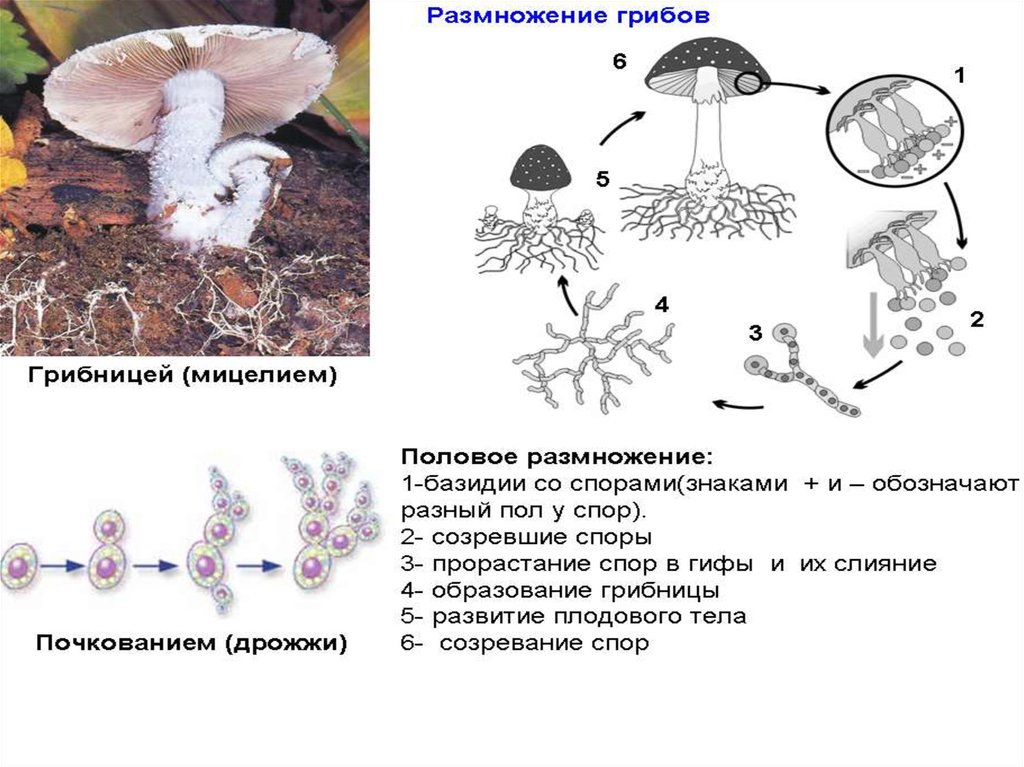 Зооспоры Phytophthora infestans, например, через 4—5 дней после их возникновения прорастают наиболее интенсивно, но уже через 8—10 дней способность их к прорастанию резко снижается.
Зооспоры Phytophthora infestans, например, через 4—5 дней после их возникновения прорастают наиболее интенсивно, но уже через 8—10 дней способность их к прорастанию резко снижается.
Пропагативные споры, как правило, прорастают путем образования ростков грибницы, т. е. вегетативно. Однако здесь имеются исключения. Так, споры Plasmopara или Phytophthora и других грибов прорастают с образованием зооспор.
В отличие от пропагативных спор, прорастающих вегетативно, покоящиеся споры, как правило, прорастают с образованием органов спороношений, т. е. фруктификативно. Покоящиеся споры в виде зиготы у низших грибов прорастают с образованием зооспорангия (например, циста) или спорангия (например, зигоспора мукоровых грибов).
У ржавчинных грибов покоящиеся споры типа телейтоспор прорастают с образованием базидии. Многие хламидоспоры, покоящиеся споры несовершенных, сумчатых грибов прорастают с образованием конидиальных спороношений.
У большинства грибов распространение спор в природе распадается на 2 фазы: освобождение спор от непосредственного контакта с материнской тканью, их образовавшей, и распространение уже освободившихся спор.
У большинства грибов споры отбрасываются, т. е. освобождаются от спорообразующих элементов активно. Однако у некоторых грибов отделение спор происходит пассивно при помощи ветра, дождя,- животных. И чаще всего эти факторы способствуют как освобождению спор, так и их распространению. Примером пассивного освобождения спор могут служить дождевики (Bovista, Lycoperdon). Пассивным; т. е. с помощью ветра, является распространение спор у так называемых сухоспоровых грибов (Aspergillus, Penicillium, Cladosporium и др.). У некоторых грибов сам по себе ветер не может отделить споры, но в присутствии воды они легко отделяются от конидиеносцев, и их распространение происходит при порывах ветра с дождем. К ним относятся, например, Fusicladium dendriticum, Tubercularia vulgaris и т. д. Но все же у грибов чаще мы встречаем различные приспособления и способы освобождения спор. На отдельных примерах из различных систематических групп рассмотрим, как это происходит.
Среди класса Zygomycetes очень интересно осуществляется активное выбрасывание спор у одного из представителей мукоровых грибов — Pilobolus. Виды его развиваются в большом количестве на навозе во влажных условиях, для этого свежий конский навоз нужно поместить на свет под стеклянный колпак. Спорангиеносец Pilobolus состоит из базальной расширенной части, переходящей в собственно спорангиеносец, и подспорангиального вздутия, расположенного непосредственно под спорангием, причем верхняя часть подспорангиального вздутия (колюмелла) вдается в спорангий. В протоплазменном слое спорангиального вздутия спорангиеносца находится вакуоль, заполненная клеточным соком, осмотическое давление которого достигает 5 атм. Оболочка спорангия этого гриба утолщена неравномерно — верхняя часть спорангия имеет более толстую оболочку, а его нижняя часть (непосредственно примыкающая к подспорангиальному вздутию) имеет очень тонкую оболочку. Благодаря большому давлению происходит разрыв оболочки спорангия, и наружу выбрасывается капля клеточного сока, которая выносит спорангий на высоту 0,5—2 см. Спорангий ударяется о новый субстрат, прикрепляется к нему наружной оболочкой.
Виды его развиваются в большом количестве на навозе во влажных условиях, для этого свежий конский навоз нужно поместить на свет под стеклянный колпак. Спорангиеносец Pilobolus состоит из базальной расширенной части, переходящей в собственно спорангиеносец, и подспорангиального вздутия, расположенного непосредственно под спорангием, причем верхняя часть подспорангиального вздутия (колюмелла) вдается в спорангий. В протоплазменном слое спорангиального вздутия спорангиеносца находится вакуоль, заполненная клеточным соком, осмотическое давление которого достигает 5 атм. Оболочка спорангия этого гриба утолщена неравномерно — верхняя часть спорангия имеет более толстую оболочку, а его нижняя часть (непосредственно примыкающая к подспорангиальному вздутию) имеет очень тонкую оболочку. Благодаря большому давлению происходит разрыв оболочки спорангия, и наружу выбрасывается капля клеточного сока, которая выносит спорангий на высоту 0,5—2 см. Спорангий ударяется о новый субстрат, прикрепляется к нему наружной оболочкой.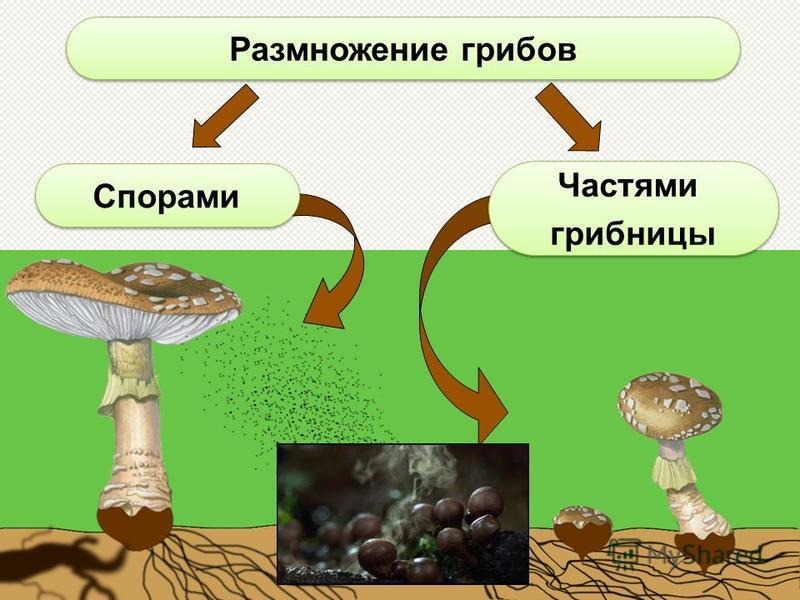 В этом случае мы встречаемся с «летательным снарядом», обеспечивающим активное отделение спорангиоспор от спорангиеносца.
В этом случае мы встречаемся с «летательным снарядом», обеспечивающим активное отделение спорангиоспор от спорангиеносца.
У многих представителей класса Oomycetes (Peronospora, Plasmopara, Premia и др.) конидии освобождаются вследствие гигроскопических движении основной ветви конидиеносцев. Дихотомически разветвленные конидиеносцы, в частности Premia и Peronospora, во влажных условиях имеют вид цилиндрических нитей, которые при высыхании теряют тургор и приобретают вид плоской ленты. И чем больше они высыхают, тем сильнее закручиваются вокруг своей продольной оси. Эти конидиеносцы обладают высокой гигроскопичностью, и поэтому даже при незначительном изменении влажности окружающего воздуха их тугор изменяется. При этом конидиеносец приобретает вращательное движение, и в результате такого движения зрелые конидии разбрасываются во все стороны.
Освобождение спор у класса Ascomycetes может происходить в различных группах по-разному. У клейстомицетов, характеризующихся замкнутыми плодовыми телами типа клейстотециев, внутри этих плодовых тел развивается одна или несколько сумок. Так, у Sphaerotheca mors-uvae, в клейстотециях которого находится одна сумка, разрыв плодового тела и разбрасывание аскоспор происходят благодаря ферментативному процессу, ведущему к сильному разбуханию сумки, что в свою очередь приводит к разрыву оболочки клейстотеция в виде щели. Через образовавшуюся щель высовывается сумка, которая продолжает разбухать, что приводит к тому, что оболочка сумки лопается и споры разбрасываются на некоторое расстояние.
Так, у Sphaerotheca mors-uvae, в клейстотециях которого находится одна сумка, разрыв плодового тела и разбрасывание аскоспор происходят благодаря ферментативному процессу, ведущему к сильному разбуханию сумки, что в свою очередь приводит к разрыву оболочки клейстотеция в виде щели. Через образовавшуюся щель высовывается сумка, которая продолжает разбухать, что приводит к тому, что оболочка сумки лопается и споры разбрасываются на некоторое расстояние.
У других видов мучнисто-росяных грибов (Podosphaera leucotricha) выбрасывание аскоспор осуществляется в два этапа. Из образовавшегося аналогичным образом разрыва клейстотеция высовывается сумка, которая благодаря увеличивающемуся давлению внутри клейстотеция выбрасывается в воздух на расстояние в несколько сантиметров. Далее, попав во влажные условия, сумка продолжает разбухать, через некоторое время (1—2 мин) ее оболочка разрывается, и аскоспоры разбрасываются во всех направлениях.
У пиреномицетов — наиболее крупной группы сумчатых грибов — сумки находятся в перитециях, снабженных узким отверстием, через которое и осуществляется выход сумок и аскоспор. При этом у некоторых видов это отверстие находится на вершине вытянутого хоботка, имеющего в зависимости от вида гриба различную длину. Для успешного выбрасывания спор верхушка сумки должна высунуться из отверстия прежде, чем ее оболочка лопнет. Среди пиреномицетов различают несколько наиболее изученных типов выбрасывания спор.
При этом у некоторых видов это отверстие находится на вершине вытянутого хоботка, имеющего в зависимости от вида гриба различную длину. Для успешного выбрасывания спор верхушка сумки должна высунуться из отверстия прежде, чем ее оболочка лопнет. Среди пиреномицетов различают несколько наиболее изученных типов выбрасывания спор.
Для изучения процесса выбрасывания спор особенно удобны пиреномицеты с одиночными перитециями. Большой интерес в этом плане представляют виды Sordaria, оболочка перитециев у которых полупрозрачна, что и позволяет легко наблюдать весь этот процесс.
Виды Sordaria относятся к числу копрофилов, развивающихся на навозе травоядных животных, а также на растительных остатках, и имеют перитеции грушевидной формы. Перед выходом аскоспор, одна из созревших сумок обычно удлиняется, верхушка ее достигает отверстия перитеция, и сумка высовывается из него. Затем она лопается, выбрасывая аскоспоры с некоторым количеством клеточного сока на расстояние до 20 см. После выбрасывания спор эластичная оболочка сумки втягивается в перитеций, поскольку сумка оставалась прикрепленной к основанию перитеция, и только после этого она вскоре ослизняется. Как только первая сумка выбросила находящиеся в ней аскоспоры, начинает удлиняться следующая сумка, и этот процесс повторяется и т. д.
Как только первая сумка выбросила находящиеся в ней аскоспоры, начинает удлиняться следующая сумка, и этот процесс повторяется и т. д.
Особенность выхода аскоспор у видов Sporormia заключается в том, что сумка имеет двойную оболочку и состоит из наружной (более толстой и жесткой) и внутренней (более тонкой и эластичной). Сумки обычно прикреплены ножками нижней частью к ткани основания перитеция. Незадолго до выбрасывания аскоспор наружная оболочка сумки разрывается в верхней части, и сумка, окруженная внутренней тонкой оболочкой, начинает быстро удлиняться в направлении к устьицу и вскоре высовывается через отверстие, выстреливая аскоспоры. После этого сумка занимает свое прежнее положение и разрушается.
У видов Gnomonia и других мы встречаем иной способ освобождения аскоспор. Перитеции у этой группы видов погружены в гниющую древесину, а над субстратом виден только выводной канал с отверстием на его вершине. Внутри перитеция развиваются многочисленные сумки, которые при созревании отделяются от ткани перитеция и лежат свободно в его полости (тип отделяющихся сумок). Ко времени созревания сумок внутри перитеция создается высокое давление, благодаря которому сумки продвигаются в канал по одной друг за другом. Сумка высовывается через отверстие, затем оболочка ее лопается, и споры разбрасываются. Сразу же опустевшая сумка выталкивается нижележащей сумкой, которая занимает ее место и т. д. Этот процесс осуществляется очень быстро, и от одного выбрасывания аскоспор до другого проходит всего несколько секунд.
Ко времени созревания сумок внутри перитеция создается высокое давление, благодаря которому сумки продвигаются в канал по одной друг за другом. Сумка высовывается через отверстие, затем оболочка ее лопается, и споры разбрасываются. Сразу же опустевшая сумка выталкивается нижележащей сумкой, которая занимает ее место и т. д. Этот процесс осуществляется очень быстро, и от одного выбрасывания аскоспор до другого проходит всего несколько секунд.
У ряда пиреномицетов сумки не лопаются, а их оболочка ослизняется внутри перитеция, и освобожденные аскоспоры образуют массу, смешанную со слизью. Эта масса поглощает воду, набухает и выдавливается через отверстие перитеция, подобно зубной пасте из тюбика, часто образуя или нити, или облако из спор. Этот тип освобождения спор характерен для видов Chaetomium и назван типом ослизняющихся сумок.
У дискомицетов, как известно, созревание сумок в апотециях происходит не одновременно, а периодами, примерно по 20—40 штук в день, при этом все зрелые сумки отстреливают зрелые споры один раз в день. У разных представителей этой группы аскомицетов имеются и различные приспособления для выхода аскоспор из сумок. Например, у широко распространенного копрофильного гриба из числа дискомицетов р. Ascobolus, апотеций которого функционирует в течение нескольких дней, при освобождении аскоспор сумка на вершине открывается маленькой крышечкой, растянутая оболочка сумки сокращается в продольном и поперечном направлениях, и споры с некоторым количеством клеточного сока выбрасываются в воздух на несколько сантиметров.
У разных представителей этой группы аскомицетов имеются и различные приспособления для выхода аскоспор из сумок. Например, у широко распространенного копрофильного гриба из числа дискомицетов р. Ascobolus, апотеций которого функционирует в течение нескольких дней, при освобождении аскоспор сумка на вершине открывается маленькой крышечкой, растянутая оболочка сумки сокращается в продольном и поперечном направлениях, и споры с некоторым количеством клеточного сока выбрасываются в воздух на несколько сантиметров.
Очень своеобразен выбрасывающий споры аппарат у видов Coprinus (класс Basidiomycetes). Его виды встречаются на гнилой древесине, находящейся в почве. Гименофор его пластинчатый, пластинки очень тонкие, многочисленные и расположены близко друг к другу. Созревание базидий обычно начинается от свободного края каждой пластинки, и постепенно этот процесс продвигается по направлению к ткани шляпки. Молодые, еще незрелые базидиоспоры вначале бесцветные. По мере созревания они становятся черно-коричневого или даже черного цвета.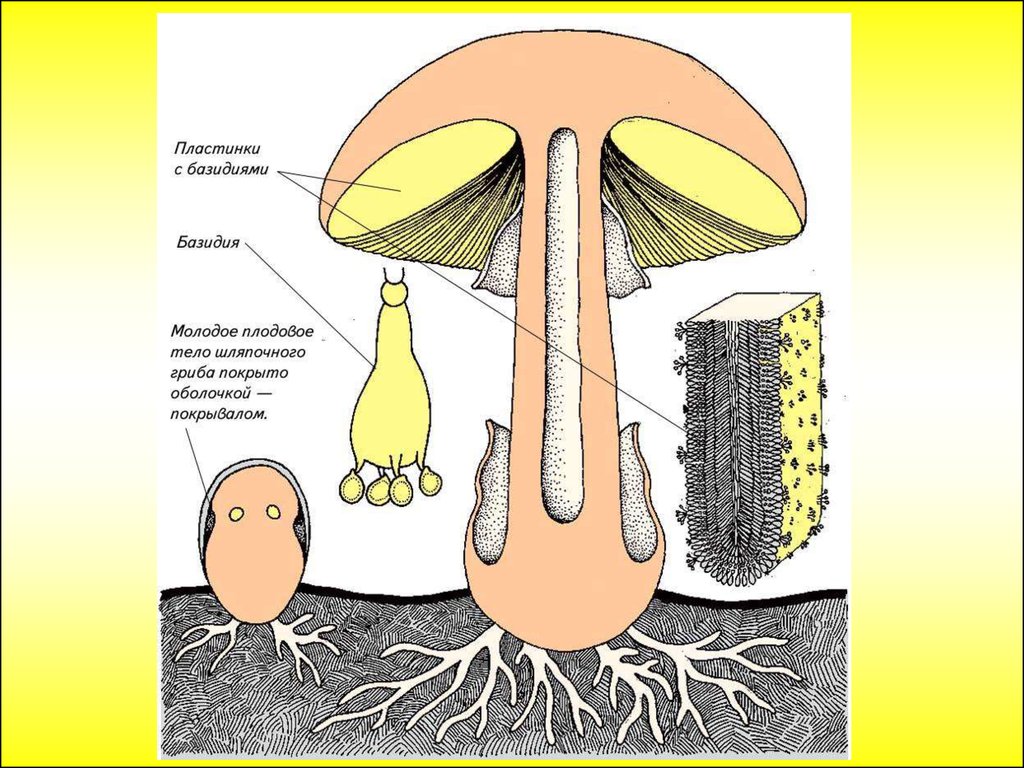 Зона возле края пластинки вскоре начинает подвергаться автолизу (саморастворению под влиянием ферментов), превращаясь в жидкость. Эта жидкость при обилии черных базидиоспор чернеет и стекает вниз по наклонному краю пластинки в виде чернильных капель на краю шляпки (отсюда название этих грибов — «чернильные» грибы).
Зона возле края пластинки вскоре начинает подвергаться автолизу (саморастворению под влиянием ферментов), превращаясь в жидкость. Эта жидкость при обилии черных базидиоспор чернеет и стекает вниз по наклонному краю пластинки в виде чернильных капель на краю шляпки (отсюда название этих грибов — «чернильные» грибы).
Вторая фаза расселения спор грибов представляет собой непосредственное распространение уже освободившихся спор. Освобожденные тем или иным путем споры могут распространяться воздушными течениями, насекомыми, водой, семенами растений, животными и человеком.
Как известно, перенесение пыльцы покрытосеменных растений с цветка на цветок насекомыми — одно из самых любопытных и поразительных явлений в ботанике. Однако и среди грибов мы находим удивительные приспособления к распространению их спор с помощью насекомых. Так, у базидиальных грибов пор. Phallales строение плодовых тел таково, что они привлекают насекомых. Эта группа в основном тропических видов, но отдельные его представители встречаются и в наших широтах.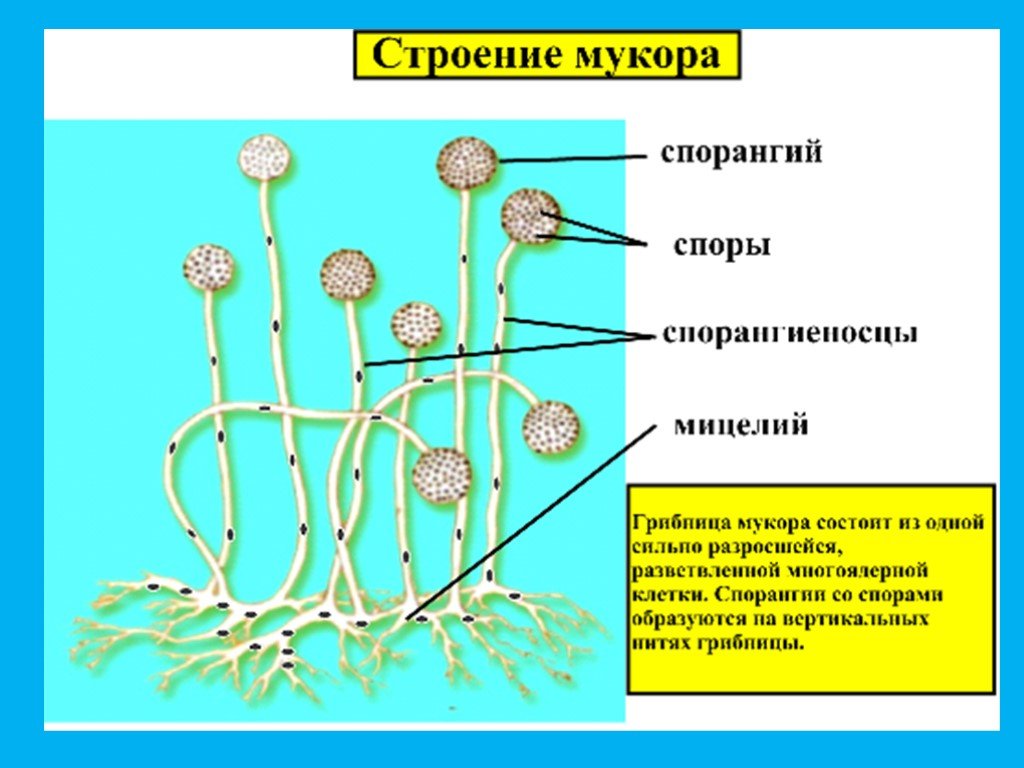 Так, например, некоторые виды родов Mutinus, Dictyophora и другие известны и у нас. Плодовые тела их вначале замкнутые, яйцевидные, затем наружная оболочка их разрывается, и из ее разрывов вырастает губчатая ткань в виде ножки (рецептакула), выносящей на вершине плодоносящую ткань — Глебу, которая у одних ярко-оранжевого, у других зеленоватого цвета. Одновременно в результате ослизнения трамы образуется стекающая сахаристая жидкость, сопровождающаяся трупным запахом. Мухи и другие насекомые, привлеченные запахом и окраской, поедают слизь, в которой содержится масса базидиоспор. Базидиоспоры переносятся на теле и лапках насекомых, а часто проходят через их пищеварительный тракт, не теряя при этом своей жизнеспособности.
Так, например, некоторые виды родов Mutinus, Dictyophora и другие известны и у нас. Плодовые тела их вначале замкнутые, яйцевидные, затем наружная оболочка их разрывается, и из ее разрывов вырастает губчатая ткань в виде ножки (рецептакула), выносящей на вершине плодоносящую ткань — Глебу, которая у одних ярко-оранжевого, у других зеленоватого цвета. Одновременно в результате ослизнения трамы образуется стекающая сахаристая жидкость, сопровождающаяся трупным запахом. Мухи и другие насекомые, привлеченные запахом и окраской, поедают слизь, в которой содержится масса базидиоспор. Базидиоспоры переносятся на теле и лапках насекомых, а часто проходят через их пищеварительный тракт, не теряя при этом своей жизнеспособности.
С помощью насекомых переносятся конидии возбудителя спорыньи злаков — Claviceps purpurea. Насекомые, привлеченные медвяной росой, сладковатой жидкостью, выделяемой конидиями гриба, переносят споры на поверхности своего тела и заражают новые растения злаков. Очень многие грибы распространяются также семенами растения-хозяина.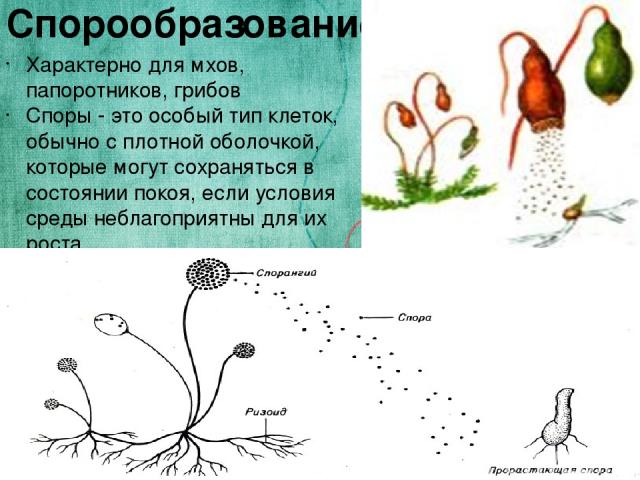 К их числу относятся многие фитопатогенные грибы — возбудители болезней растений (Helminthosporium avenae, Tilletia tritici, Uslilago tritici, Ustilago nuda и многие другие).
К их числу относятся многие фитопатогенные грибы — возбудители болезней растений (Helminthosporium avenae, Tilletia tritici, Uslilago tritici, Ustilago nuda и многие другие).
Источник: Н.П. Черепанова. Морфология и размножение грибов. Учебное пособие. Изд-во Ленинградского университета. Ленинград. 1981
Споры | это… Что такое Споры?
Не следует путать с Спор.
Спо́ры (греч. σπορά, σπόρος — сеяние, посев, семя) — особый тип клеток с плотной оболочкой.
В биологии понятие «спора» может относиться к нескольким категориям организмов:
- споры бактерий, служащие для пережидания неблагоприятных условий;
- споры растений, споровиков и грибов, служащие для размножения.
Содержание
|
Споры бактерий
Прокариотические организмы — бактерии обладают способностью к спорообразованию, которая заключается в том, что при наступлении условий, неблагоприятных для жизни, клетка частично теряет воду, объём и форму; под внешней мембраной образуется плотная сферическая оболочка.
В виде споры бактерия может выдерживать огромные механические, температурные и химические нагрузки. Например, некоторые споры выдерживают трёхчасовое кипячение или температуру жидкого азота.
Также в виде споры более эффективно проходит расселение, потому что частично обезвоженная клетка имеет меньшую массу.
Споры неустойчивы к ультрафиолету, как и вообще бактерии, и быстро погибают под таким излучением. Поэтому меньше всего бактерий — в высокогорной местности, а некоторые методы лечения инфекционных заболеваний, вызванных бактериальным возбудителем, предусматривают облучение пациента.
Следует помнить, что споры у бактерий, в отличие от растений и грибов, служат не для размножения.
Споры бактерий окрашивают по методу Пешкова или по методу Марцелли.
Споры растений
Споры растений
Споры растений — микроскопические зачатки низших и высших растений, имеющие разное происхождение и служащие для их размножения и (или) сохранения при неблагоприятных условиях. Представляют собой одноклеточные, реже двуклеточные или состоящие из нескольких клеток образования. Обычно более или менее шарообразной, эллипсоидальной формы, реже — цилиндрической и др.
Представляют собой одноклеточные, реже двуклеточные или состоящие из нескольких клеток образования. Обычно более или менее шарообразной, эллипсоидальной формы, реже — цилиндрической и др.
Спородерма
Совокупность оболочек споры, спородерма (от sporá + dérma — «кожа, оболочка»), имеет сложное строение, дифференцируясь на наружный толстый слой, экзину (англ. exine), и внутренний — интину. Экзина состоит из спорополленина, одного из самых стойких органических веществ, способного выдерживать длительные температурные и химические воздействия[1]. Благодаря стойкости экзины, споры обычно длительное время сохраняют способность к прорастанию и могут сохраняться в отложениях на протяжении геологических эпох. Спородерма часто имеет скульптурированные оболочки, покрытые гребешками, бугорками, шипами и т. д.[2]
Размножение спорами
В цикле развития растений, начиная с водорослей и заканчивая покрытосеменными, последовательно чередуются стадии спорофита (растения, образующего споры) и гаметофита (растения, образующего гаметы). Так, у папоротника спорофитом является взрослое растение, распространяющее споры; из каждой такой споры вырастает заросток, являющийся гаметофитом: он образует женские гаметангии архегонии и мужские гаметангии антеридии, в которых развиваются половые гаметы и, сливаясь (обычно перекрёстно с разных растений), образуют зиготу, развивающуюся на заростке и вырастающую во взрослое растение.
Так, у папоротника спорофитом является взрослое растение, распространяющее споры; из каждой такой споры вырастает заросток, являющийся гаметофитом: он образует женские гаметангии архегонии и мужские гаметангии антеридии, в которых развиваются половые гаметы и, сливаясь (обычно перекрёстно с разных растений), образуют зиготу, развивающуюся на заростке и вырастающую во взрослое растение.
Размножение спорами — бесполое. Однако споры участвуют в процессе полового размножения у голосеменных и покрытосеменных, или цветковых, растений. Взрослое растение здесь является спорофитом, образующим макро- (женские) и микроспоры (мужские), которые развиваются соответственно в зародышевый мешок и зрелое пыльцевое зерно, являющиеся гаметофитами.
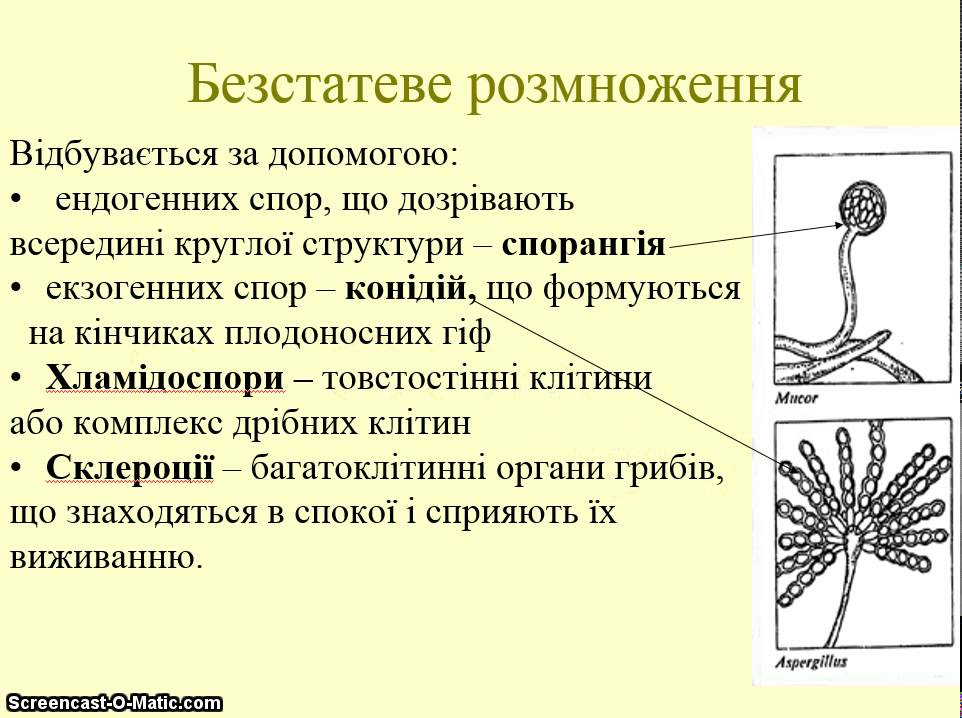
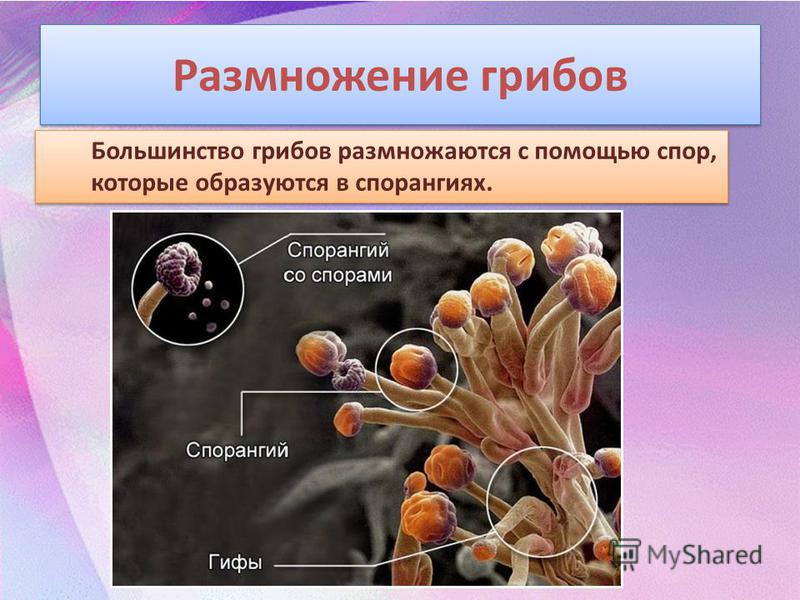
Споры грибов
У грибов споры могут развиваться внутри специальных споровместилищ (эндогенно) или на конце особых выростов мицелия — конидиеносцах (экзогенно).
См. также
- Размножение
- Цисты
Примечания
Ссылки
- Микроспорогенез и микрогаметогенез семенных растений
Британника
- Развлечения и поп-культура
- География и путешествия
- Здоровье и медицина
- Образ жизни и социальные вопросы
- Литература
- Философия и религия
- Наука
- Спорт и отдых
- Технология
- Изобразительное искусство
- Всемирная история
- Этот день в истории
- Викторины
- Подкасты
- Словарь
- Биографии
- Резюме
- Популярные вопросы
- Обзор недели
- Инфографика
- Демистификация
- Списки
- #WTFact
- Галереи изображений
- Прожектор
- Форум
- Один хороший факт
- Развлечения и поп-культура
- География и путешествия
- Здоровье и медицина
- Образ жизни и социальные вопросы
- Литература
- Философия и религия
- Политика, право и правительство
- Наука
- Спорт и отдых
- Технология
- Изобразительное искусство
- Всемирная история
- Britannica объясняет
В этих видеороликах Britannica объясняет различные темы и отвечает на часто задаваемые вопросы.
- Britannica Classics
Посмотрите эти ретро-видео из архивов Encyclopedia Britannica. - #WTFact Видео
В #WTFact Britannica делится некоторыми из самых странных фактов, которые мы можем найти. - На этот раз в истории
В этих видеороликах узнайте, что произошло в этом месяце (или любом другом месяце!) в истории. - Demystified Videos
В Demystified у Britannica есть все ответы на ваши животрепещущие вопросы.

- Студенческий портал
Britannica — это главный ресурс для учащихся по ключевым школьным предметам, таким как история, государственное управление, литература и т. д. - Портал COVID-19
Хотя этот глобальный кризис в области здравоохранения продолжает развиваться, может быть полезно обратиться к прошлым пандемиям, чтобы лучше понять, как реагировать сегодня. - 100 женщин
Britannica празднует столетие Девятнадцатой поправки, выделяя суфражисток и политиков, творящих историю. - Britannica Beyond
Мы создали новое место, где вопросы находятся в центре обучения. Вперед, продолжать. Просить. Мы не будем возражать. - Спасение Земли
Британника представляет список дел Земли на 21 век. Узнайте об основных экологических проблемах, стоящих перед нашей планетой, и о том, что с ними можно сделать! - SpaceNext50
Britannica представляет SpaceNext50. От полёта на Луну до управления космосом — мы изучаем широкий спектр тем, которые подпитывают наше любопытство к космосу!
Страница не найдена
Приносим свои извинения! Этот контент недоступен. Посетите домашнюю страницу Britannica или воспользуйтесь полем поиска ниже.
Поиск в Британике
От латинского
Месяц Американского Красного Креста 2016 г.
Споры грибов: критический обзор токсикологических и эпидемиологических данных в качестве основы для установления пределов воздействия на рабочем месте
Обзор
. 2009 г.;39(10):799-864.
дои: 10.3109/10408440903307333.
Вейнанд Эдуард 1
принадлежность
- 1 Национальный институт гигиены труда, Осло, Норвегия. wijand.eduard@stami.no
- PMID: 19863384
- DOI: 10.3109/10408440903307333
Обзор
Вейнанд Эдуард. Критический преподобный Toxicol.
2009.
Критический преподобный Toxicol.
2009.
. 2009;39(10):799-864.
дои: 10.3109/10408440903307333.
Автор
Вейнанд Эдуард 1
принадлежность
- 1 Национальный институт гигиены труда, Осло, Норвегия. wijand.eduard@stami.no
- PMID: 19863384
- DOI: 10.3109/10408440903307333
Абстрактный
Споры грибов широко распространены в окружающей среде. Однако уровни воздействия на рабочих местах, где обрабатываются материалы с плесенью, намного выше, чем в обычных помещениях и на открытом воздухе. Споры всех протестированных видов вызывали воспаление в экспериментальных исследованиях. Реакция на продуцирующие микотоксины и патогенные виды была намного сильнее. В исследованиях на животных после однократного введения преобладали неаллергические реакции. Возникали также аллергические реакции, особенно на продуцирующие микотоксины и патогенные виды, а также после многократного воздействия. Вдыхание разовой дозы спор субъектами с синдромом больного здания не выявило наблюдаемых уровней воздействия 4 x 10(3) спор Trichoderma harzianum/м(3) и 8 x 10(3) спор Penicillium chrysogenum/м(3) на функцию легких. , респираторные симптомы и воспалительные клетки в крови. У больных астмой с аллергией на Penicillium sp. или Alternaria alternata, самые низкие наблюдаемые уровни воздействия (LOEL) для снижения проводимости дыхательных путей составляли 1 x 10(4) и 2 x 10(4) спор/м(3) соответственно.
Однако уровни воздействия на рабочих местах, где обрабатываются материалы с плесенью, намного выше, чем в обычных помещениях и на открытом воздухе. Споры всех протестированных видов вызывали воспаление в экспериментальных исследованиях. Реакция на продуцирующие микотоксины и патогенные виды была намного сильнее. В исследованиях на животных после однократного введения преобладали неаллергические реакции. Возникали также аллергические реакции, особенно на продуцирующие микотоксины и патогенные виды, а также после многократного воздействия. Вдыхание разовой дозы спор субъектами с синдромом больного здания не выявило наблюдаемых уровней воздействия 4 x 10(3) спор Trichoderma harzianum/м(3) и 8 x 10(3) спор Penicillium chrysogenum/м(3) на функцию легких. , респираторные симптомы и воспалительные клетки в крови. У больных астмой с аллергией на Penicillium sp. или Alternaria alternata, самые низкие наблюдаемые уровни воздействия (LOEL) для снижения проводимости дыхательных путей составляли 1 x 10(4) и 2 x 10(4) спор/м(3) соответственно.
Похожие статьи
Влияние концентрированных частиц окружающей среды на нормальные и гиперсекреторные дыхательные пути у крыс.
Харкема Дж.Р., Киллер Г., Вагнер Дж., Моришита М., Тимм Э., Хотчкисс Дж., Марсик Ф., Двонч Т., Камински Н., Барр Э. Харкема Дж. Р. и соавт. Res Rep Health Eff Inst. 2004 г., август; (120): 1-68; обсуждение 69-79.
Res Rep Health Eff Inst. 2004.
PMID: 15543855Критический обзор данных о кратковременном воздействии двуокиси азота (NO2) на человека: данные о неэффективных уровнях NO2.
Хестерберг Т.В., Банн В.Б., Макклеллан Р.О., Хамаде А.К., Лонг С.М., Валберг П.А. Хестерберг Т.В. и соавт. Критический преподобный Toxicol. 2009;39(9):743-81. дои: 10.3109/10408440903294945. Критический преподобный Toxicol. 2009. PMID: 19852560 Обзор.
Переносимые по воздуху грибковые споры при поперечном исследовании офисных зданий.
MacIntosh DL, Brightman HS, Baker BJ, Myatt TA, Stewart JH, McCarthy JF. MacIntosh DL и др. J Occup Environ Hyg. 2006 г., июль; 3(7):379-89. дои: 10.1080/10543400600760438. J Occup Environ Hyg.
2006.
PMID: 16835164Влияние высоты отбора проб на концентрацию переносимых по воздуху спор грибов.
Хаттаб А., Леветин Э. Хаттаб А. и др. Энн Аллергия Астма Иммунол. 2008 ноябрь; 101 (5): 529-34. doi: 10.1016/S1081-1206(10)60293-1. Энн Аллергия Астма Иммунол. 2008. PMID: 19055208
Профессиональный контакт с микотоксинами в различных отраслях промышленности.
Сорока П.М., Кипровский М., Шадковска-Станчик И. Сорока П.М. и др. Мед пр. 2008;59(4):333-45. Мед пр. 2008. PMID: 19143225 Обзор. польский.
 Res Rep Health Eff Inst. 2004.
PMID: 15543855
Res Rep Health Eff Inst. 2004.
PMID: 15543855 2006.
PMID: 16835164
2006.
PMID: 16835164Посмотреть все похожие статьи
Цитируется
Оценка качества воздуха в помещении для групповых макак ( Macaca spp.
).Мааскант А., Янссен И., Воутерс И.М., Эрденбург FJCMV, Ремарк Э.Дж., Лангерманс Я.М., Баккер Дж. Мааскант А. и др. Животные (Базель). 2022 7 июля; 12 (14): 1750. дои: 10.3390/ани12141750. Животные (Базель). 2022. PMID: 35883296 Бесплатная статья ЧВК.
Последствия иммунного ландшафта при ХОБЛ и раке легких: курение в сравнении с другими причинами.
Таучер Э., Миколюк И., Линденманн Дж., Смолле-Юеттнер Ф.М. Таучер Э. и др. Фронт Иммунол. 2022 21 марта; 13:846605. doi: 10.3389/fimmu.2022.846605. Электронная коллекция 2022. Фронт Иммунол. 2022. PMID: 35386685 Бесплатная статья ЧВК. Обзор.
Воздушный микобиом и ассоциации с микотоксинами и воспалительными маркерами в зерновой промышленности Норвегии.
Straumfors A, Mundra S, Foss OAH, Mollerup SK, Kauserud H. Страумфорс А. и др. Научный представитель 2021 г., 30 апреля; 11 (1): 9357. doi: 10.1038/s41598-021-88252-1. Научный представитель 2021. PMID: 33931660 Бесплатная статья ЧВК.
Влияние длины среза листовых пластинок многолетнего райграса на ферментацию in vitro анаэробным грибком Neocallimastix frontalis .
Хименес Х.Р., Эдвардс Дж.Э., Сандерсон Р., Кингстон-Смит А.Х., Макьюэн Н.Р., Теодору М.К. Хименес Х.Р. и др. Микроорганизмы. 2020 11 ноября; 8 (11): 1774. doi: 10.3390/microorganisms8111774. Микроорганизмы. 2020. PMID: 33187375 Бесплатная статья ЧВК.
Респираторные заболевания и аллергия у фермеров, работающих с домашним скотом: документ с изложением позиции EAACI.
 ).
).
Leave A Comment