
Хемосинтезирующие бактерии окислили марганец
Ученые обнаружили бактерий, которые умеют окислять марганец и использовать энергию этой химической реакции для фиксации углекислого газа. Два вида бактерий в кокультуре превращали суспензию карбоната марганца в бляшки оксида марганца; в их транскриптоме нашли гены белков, которые могут осуществлять биохимические каскады хемосинтеза. Найденные микроорганизмы, считают авторы статьи в Nature, могут замыкать природные циклы марганца и влиять на круговорот других элементов.
Автотрофы — организмы, которые умеют фиксировать атмосферный углерод и синтезировать органические вещества из неорганических. Энергию для этого они берут либо из солнечного света и тогда называются фотоавтотрофами, либо из химических реакций неорганических веществ, тогда они — хемолитоавтотрофы.
Бактерии, которые используют для хемосинтеза соединения азота, серы и железа, известны уже более века. Известен также ряд микроорганизмов, которые окисляют марганец, но зачем они это делают, остается непонятным. Ученые предполагают, что из таких химических реакций микроорганизмы умеют получать энергию. Однако до сих пор это не удавалось подтвердить экспериментально, и хемоавтотрофы, растущие на марганцевых средах, не были найдены.
Ученые предполагают, что из таких химических реакций микроорганизмы умеют получать энергию. Однако до сих пор это не удавалось подтвердить экспериментально, и хемоавтотрофы, растущие на марганцевых средах, не были найдены.
Джареду Ледбеттеру (Jared Leadbetter) из Калифорнийского технологического института повезло — он на несколько месяцев оставил в раковине грязную посуду с остатками карбоната марганца (II), залитую водой из под крана. Вернувшись в лабораторию, исследователь обнаружил, что вместо светлой суспензии карбоната колбы покрыты темным налетом — оксидом марганца. Ледбеттер предположил, что марганец окислили бактерии. С коллегой он перенес материал в среду со строго определенным составом, а в качестве контроля использовал аналогичные стерильные среды.
На стерильных чашках марганец не окислился даже через год, зато в среде с биологическим материалом уже через четыре месяца образовались темные оксиды марганца. Окисление требовало наличия кислорода и происходило при температуре до 42 градусов Цельсия, оптимально — при 34-40 градусах, что предполагает ферментативный характер катализа. Образование оксидов не происходило после добавления антибиотиков или пастеризации.
Образование оксидов не происходило после добавления антибиотиков или пастеризации.
Ученые исследовали налет оксида марганца, который вырабатывали бактерии. Оказалось, что он состоит из бляшек диаметром 20-500 микрометров, темно-коричневых с темно-оранжевыми впячиваниями — в них и находилась большая часть бактерий.
Анализ рибосомной РНК показал, что в среде содержалось около 70 видов бактерий, однако после последовательного разбавления в растворе карбоната марганца выжило только два вида: один относится к типу Nitrospirae, а второй — к бета-протеобактериям. На момент написания статьи в отдельную культуру удалось выделить только второй вид, и он не мог в одиночку окислять марганец — значит, либо первый вид делает это самостоятельно, либо окисление марганца требует кооперации.
Количество окисленного марганца в кокультуре двух видов сначала возрастало экспоненциально, а затем замедлялось, и так же себя вела биомасса. Это соответствует представлениям о росте бактерий в культуре, а скорость роста колоний была сопоставима с таковой для известных хемолитотрофов.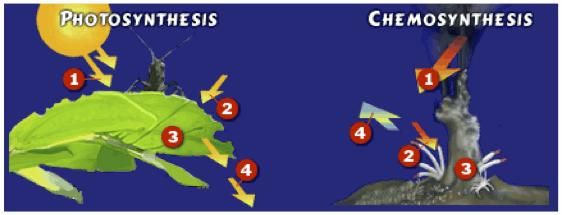
Чтобы проверить, могут ли бактерии использовать энергию окисления марганца для фиксации углерода, исследователи вырастили их в культуре с меченым карбонатом марганца. В результате оба вида включили в себя тяжелый изотоп углерода — значит, что бактерии не просто умеют окислять марганец, но и используют энергию этой реакции для усвоения неорганического углерода, то есть являются автотрофами. С помощью анализа транскриптома ученые нашли активные гены белков, которые могли бы осуществлять биохимические каскады переноса электронов от внеклеточного марганца на кислород и автотрофную фиксацию углекислого газа.
Ученые рассчитали, что колония, выросшая из двух клеток (по одной каждого из двух видов), может за два года создать количество оксида, равное общему мировому запасу марганца. Найденные бактерии могут замыкать неизвестные до сих пор циклы марганца в природе, окисляя металл, который восстанавливают другие хемотрофы. Эти циклы, в свою очередь, могут взаимодействовать с круговоротом углерода, азота, водорода и кислорода.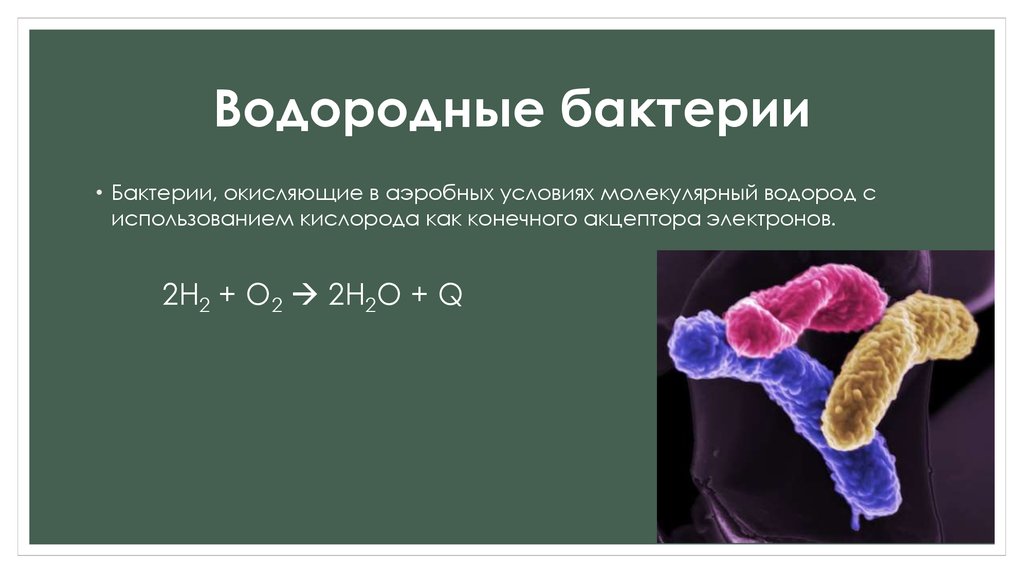
Для хемосинтеза открытым бактериям необходим не только марганец, но и кислород, как и большинству современных автотрофов. Но до кислородной катастрофы, когда все организмы были анаэробами, приходилось обходиться без кислорода. Так действовали древнейшие найденные сероокисляющие бактерии, которых геологи обнаружили несколько лет назад.
Алиса Бахарева
Нашли опечатку? Выделите фрагмент и нажмите Ctrl+Enter.
Хемосинтезирующие бактерии и их экологическое значение
Хемосинтезирующие бактерии — бактерии, которые используют в качестве источника углерода CO2, но энергию получают не от солнца, а от химических реакций. Энергия может выделяться при окислении водорода, сероводорода, серы, железа, аммиака и др неорганических соединений. Хемосинтезирующие бактерии играют важную роль в биосфере; в основном они участвуют в круговороте азота и таким образом поддерживают плодородие почвы.
Существует несколько
групп хемосинтезирующих бактерий, из
которых наибольшее значение имеют
нитрифицирующие (получают энергию
для синтеза органических веществ,
окисляя аммиак до азотистой, а затем до
азотной кислоты), водородные (окисляя
водород до воды), серобактерии (окисляя
сероводород до сульфатов) и железобактерии
(превращая закисные соли железа в
окисные).
Процесс хемосинтеза, при котором из СO2 образуется органическое вещество, протекает аналогично темновой фазе фотосинтеза. Благодаря жизнедеятельности бактерий-хемосинтетиков в природе накапливаются большие запасы селитры и болотной руды.
Молекулярная природа и виды изменчивости
Наследственность — свойство живых организмов воспроизводить себе подобное, или передавать в ряду поколений определенный тип индивидуального развития, в ходе которого у потомков формируется комплекс видовых признаков.
Изменчивость – в противоположность наследственности – обеспечивает разнообразие признаков и свойств у сособей одного вида . Различают изменчивость модификационную, или фенотипическую, и мутационную, или генотипическую
Формы изменчивости :
Ненаследственная модификационная (фенотипическая)
Причины : Изменение условий среды, в результате чего организм изменяется в пределах нормы реакции, заданной генотипом
Значение : Адаптация – приспособление к данным условиям среды, выживание, сохранение потомства
Наследственная (генотипическая), мутационная
Причины
: Влияние внешних и внутренних мутагенных
факторов, в результате чего происходит
изменение в генах и хромосомах.
Значение : Материал для естественного и искусственного отбора, так как мутации могут быть полезные, вредные и безразличные, доминантные и рецессивные
Наследственная (генотипическая) , комбинатнвная
Причины : Возникает стихийно в рамках популяции при скрещивании, когда у потомков появляются новые комбинации генов
Значение : Распространение в популяции новых наследственных изменений, которые служат материалом для отбора
Наследственная (генотипическая) , соотносительная (коррелятивная)
Причины : Возникает в результате свойства генов влиять на формирование не одного, а двух и более признаков
Значение : Постоянство взаимосвязанных признаков, целостность организма как системы
Строение , образ жизни и значение вирусов.
Вирусы
(лат. virus
– яд) – субмикроскопические
неклеточные организмы, способные
размножаться только внутри живых клеток
(облигатные паразиты).
Размеры вирусов колеблются от 15 до 350 нм (длина некоторых нитевидных достигает 3 000 нм; 1 нм = 1·10–9м), т.е. большинство из них не видны в световой микроскоп (субмикроскопические) и их изучение стало возможным только после изобретения электронного микроскопа.
Зрелая
вирусная частица (т.е. внеклеточная,
покоящаяся – вирион)
устроена очень
просто: она состоит из одной или нескольких
молекул нуклеиновых кислот, составляющих
сердцевину вируса,
и белковой оболочки (капсид)
– это так называемые простые
вирусы. Сложные
вирусы (например,
герпеса
или гриппа)
кроме, белков капсида и нуклеиновой
кислоты содержат дополнительную
липопротеидную
мембрану
(оболочку, суперкапсид образуемый из
плазматической мембраны клетки хозяина),
различные углеводы
и ферменты
(рис.3.1). Ферменты способствуют
проникновению вирусной НК в клетку
и выходу образовавшихся вирионов
в среду (нейраминидаза
миксовирусов, АТФ-аза
и лизоцим
некоторых фагов и др.
Белковая оболочка защищает нуклеиновую кислоту от различных физических и химических воздействий, а также препятствует проникновению к ней клеточных ферментов, предотвращая тем самым ее расщепление (защитная функция). Также, в составе капсида имеется рецептор, комплементарный рецептору заражаемой клетки – вирусы поражают строго определенный круг хозяев (определительная функция). Вирионы многих вирусов растений и ряда фагов имеют спиральный капсид, в котором белковые субъединицы (капсомеры) уложены по спирали вокруг оси. Например, ВТМ (вирус табачной мозаики) имеет форму палочек диаметром 15 – 17 нм и длиной до 300 нм (рис. 3.2.). Внутри его капсида имеется полый канал диаметром 4 нм. Генетическим
материалом ВТМ является одноцепочечная РНК, плотно уложенная в желобке спирального капсида. Для вирионов со спиральным капсидом характерно высокое содержание белка (90 – 98%) по отношению к
Рис. 3.2. Строение
вируса табачной мозаики.
3.2. Строение
вируса табачной мозаики.
нуклеиновой кислоте.
Капсиды вирионов многих вирусов (например, аденовирус, вирус герпеса, вирус желтой мозаики турнепса – ВЖМТ) имеют форму симметричного многогранника, чаще всего икосаэдра (многогранник с 12 вершинами, 20 треугольными гранями и 30 ребрами). Такие капсиды называют изометрическими (рис. 3.3.). В таких вирионах содержание белка составляет около 50% по отношению к НК. В вирусе присутствует всегда один тип нуклеиновой кислоты (либо ДНК, либо РНК), поэтому все вирусы делят на ДНК-содержащие и РНК-содержащие. Молекулы нуклеиновой кислоты в вирионе могут быть линейными (РНК, ДНК) или иметь форму кольца (ДНК). Причем эти нуклеиновые кислоты могут состоять из одной цепочки или из двух. Вирусная НК имеет от 3 до 200 генов.
ДНК-содержащие
вирусы (роды) – поксвирус,
иридовирус, герпесвирус, аденовирус
и др. РНК-содержащие
вирусы – парамиксовирус,
ортомиксовирус, коронавирус, аренавирус,
лейковирус, рабдовирус
и др. Нуклеиновая кислота вируса совмещает
в себе функции обеих кислот (ДНК и РНК)
– это хранение и передача наследственной
информации, а также управление синтезом
белков.
Как уже было сказано выше, вне живой клетки вирусы размножаться не могут. Вирус попадает в клетку, либо впрыскивая в нее свою нуклеиновую кислоту оставляя при этом белковую оболочку снаружи клетки (как это делают бактериофаги), либо при фагоцитозе (пиноцитозе) вместе с белковой оболочкой (вирусы животных), либо через нарушенную клеточную оболочку (вирусы растений).
Вирусные
РНК или ДНК, попав в клетку-хозяина,
начинают самовоспроизводиться
(удваиваться).
Хемосинтетические бактерии – определение, примеры, пути и процессы
Определение, примеры, пути и процессы
Определение: Что такое хемосинтетические бактерии?
По сути, хемосинтетические бактерии включают группу автотрофных бактерий, которые используют химическую энергию для производства своей пищи. Как и фотосинтезирующие бактерии, хемосинтезирующие бактерии нуждаются в источнике углерода (например, двуокиси углерода), а также в источнике энергии для производства собственной пищи.
Химические реакции в генераторе биогаза…
Пожалуйста, включите JavaScript
Химические реакции в генераторе биогаза, биогазовая установка.
По большей части эти бактерии являются аэробными и поэтому для успешного завершения этого процесса полагаются на кислород. Однако некоторые виды (например, Sulfuricurvum kujiense) связаны с анаэробным хемосинтезом.
Благодаря своей способности производить себе пищу с использованием химической энергии, эти организмы способны выживать в различных средах обитания/окружающей среде, включая суровые условия с экстремальными условиями, в качестве свободноживущих организмов или в ассоциации с другими организмами посредством симбиоза с другими организмами .
* Unlike photosynthesis which is common in eukaryotic organisms and cyanobacteria, chemosynthetic reactions are mostly carried out by prokaryotic microorganisms (particularly bacteria and archaea)
Examples of chemosynthetic bacteria include:
- Venenivibrio stagnispumantis
- Beggiatoa
- T.
 neapolitanus
neapolitanus - T. novelus
- ferrooxidans
 neapolitanus
neapolitanusТипы хемосинтезирующих бактерий
Как уже упоминалось, хемосинтез позволяет различным типам бактерий (хемосинтезирующим бактериям) выживать, не полагаясь на световую энергию или другие организмы в качестве пищи.
Здесь энергия, используемая для производства пищевых материалов, получается из различных неорганических химических веществ и, следовательно, различных химических реакций. По этой причине существуют различные типы хемосинтетических бактерий в зависимости от типа соединений, которые они используют в качестве источника энергии.
* Некоторые хемосинтетические бактерии живут в солнечной среде и поэтому подвергаются воздействию солнечного света. Однако они не полагаются на солнечный свет как на источник энергии
Серобактерии — Эти бактерии (например, Paracoccus) окисляют такие соединения серы, как сероводород (сульфиды), тиосульфаты и неорганическую серу и т.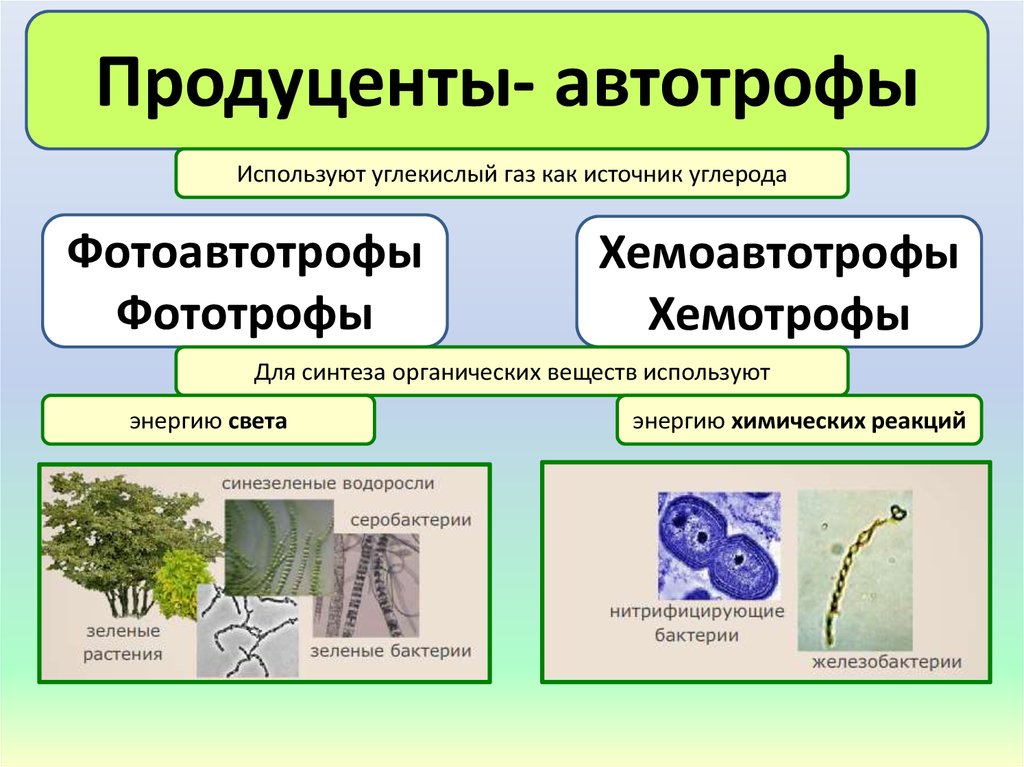 д. В зависимости от организма или В зависимости от типа используемого соединения серы процесс окисления протекает в несколько стадий.
д. В зависимости от организма или В зависимости от типа используемого соединения серы процесс окисления протекает в несколько стадий.
В некоторых организмах, например, неорганическая сера будет храниться до тех пор, пока она не потребуется для использования.
Азотные бактерии – делятся на три группы, включающие нитрифицирующие бактерии, денитрифицирующие бактерии и азотфиксирующие бактерии. У нитрифицирующих бактерий аммиак сначала окисляется до гидроксиламина в цитоплазме (монооксигеназой аммония).
Затем гидроксиламин окисляется с образованием нитрита в периплазме под действием гидроксиламиноксидоредуктазы. Этот процесс производит протон (один протон на каждую молекулу аммония). По сравнению с нитрифицирующими бактериями денитрифицирующие бактерии окисляют нитратные соединения как источник энергии.
Метанобактерии/метановые бактерии — Хотя некоторые ученые предполагают, что некоторые бактерии используют метан в качестве источника энергии для хемосинтеза, это особенно распространено среди хемосинтетических архебактерий.
Водородные бактерии — Такие бактерии, как Hydrogenovibrio marinus и Helicobacter pylori, окисляют водород как источник энергии в микроаэрофильных условиях.
Было показано, что по большей части эти бактерии являются анаэробными и поэтому процветают в областях с очень низким содержанием кислорода или вообще без него. Во многом это связано с тем, что фермент, используемый для целей окисления (гидрогеназа), эффективно функционирует в анаэробных условиях.
Железобактерии – Acidithiobacillus ferrooxidans и Leptospirillum ferrooxidans – некоторые из бактерий, окисляющих железо. Было показано, что этот процесс происходит в разных условиях в зависимости от организма (например, при низком pH и кислородно-аноксическом воздействии).
Во время хемосинтеза хемосинтетические бактерии, будучи нефотосинтезирующими, должны полагаться на энергию, вырабатываемую при окислении этих соединений (неорганических), чтобы производить пищу (сахара), в то время как азотфиксирующие бактерии превращают газообразный азот в нитрат. Все эти процессы служат для производства протона, используемого для фиксации углекислого газа.
Все эти процессы служат для производства протона, используемого для фиксации углекислого газа.
В норме эти реакции протекают в цитоплазме в присутствии мембраносвязанных дыхательных ферментов. Например, в случае окисления водорода гидрогеназы NiFe группы 1, обнаруженные в цитоплазме, катализируют реакцию образования 2 электронов и протонов (водород с положительным зарядом) из молекулы водорода (h3 <> 2H+ и 2e-). Затем эти электроны направляются в пул хинонов в цепи переноса электронов.
В случае сероводорода соединение подвергается окислению с высвобождением электронов и ионов водорода (называемых протонами, поскольку они отделяются от соединения и электронов и приобретают положительный заряд). Таким образом, продуктами этой реакции являются сера, электроны, а также протоны. Затем электроны и протоны входят в цепь переноса электронов (на мембрану).
Когда электроны входят в эту цепь, протоны выкачиваются из клетки. Электроны, с другой стороны, принимаются кислородом и притягивают протоны (ионы водорода), тем самым образуя молекулы воды. С помощью фермента, известного как АТФ-синтаза, протоны, которые ранее были выкачаны из клетки, направляются обратно в клетку, а их энергия (кинетическая энергия) сохраняется в виде АТФ и используется для синтеза сахара.
С помощью фермента, известного как АТФ-синтаза, протоны, которые ранее были выкачаны из клетки, направляются обратно в клетку, а их энергия (кинетическая энергия) сохраняется в виде АТФ и используется для синтеза сахара.
Ассимиляция углерода в хемосинтетических бактериях (фиксация)
В зависимости от типа бактерий, среды их обитания и источника углерода существует ряд метаболических путей, используемых для фиксации.
Некоторые из наиболее распространенных путей: , 5-бисфосфат. В результате этого процесса образуется шестиуглеродное соединение, которое, в свою очередь, превращается в две молекулы 3-PGA (3-фосфоглицерат). Этот процесс называется фиксацией углерода, так как он включает превращение углекислого газа в органические молекулы.
За счет энергии, запасенной в АТФ и НАДФН (вырабатываемой в процессе окисления), углеродное соединение (3-PGA) снова превращается в другое углеродное соединение с образованием G3P (глицеральдегид-3-фосфат) в фазе восстановления.
Поскольку одна из этих молекул покидает цепь Кальвина (с образованием молекулы углевода/сахара), другая участвует в образовании RuBP.
Обратный цикл Кребса — По сравнению с циклом Кальвина фиксация углерода в обратном цикле Кребса приводит к образованию пирувата. Этот цикл, также известный как восстановительный цикл трикарбоновых кислот, начинается с фиксации двух молекул углекислого газа. Это приводит к образованию ацетилкофермента А (ацетил-КоА), который, в свою очередь, редуктивно карбоксилируется с образованием пирувата.
Пируват, полученный в процессе, затем используется для синтеза органических клеточных материалов.
Некоторые из других процессов, используемых этими бактериями, включают:
· наличие ацетил-КоА- и пропионил-КоА-карбоксилаз. Затем он расщепляется с образованием ацетил-КоА и глиоксилата. В конечном итоге этот путь приводит к производству пирувата, который используется для синтеза различных органических материалов, необходимых клетке.
· Восстановительный путь ацетил-КоА – В этом пути две молекулы диоксида углерода фиксируются с образованием ацетил-КоА. Как правило, водород выступает в качестве донора электронов в этой реакции, а углекислый газ является акцептором электронов.
· Цикл дикарбоксилата/4-гидроксибутирата — Этот цикл распространен среди бактерий, обитающих в анаэробных и микроаэробных средах (например, Desulfurococcales). Подобно циклу 3-гидроксипропионат/4-гидроксибутират, этот цикл превращает цетил-КоА и две молекулы углерода в сукцинил-коэнзим (КоА). Некоторые из ферментов, участвующих в этом цикле, включают пируватсинтазу и фосфоенолпируват (ФЕП) карбоксилазу.
Важность хемосинтезирующих бактерий
По сути, хемосинтеза относится к процессу, посредством которого хемосинтетические бактерии перерабатывают пищу с использованием химической энергии. Следовательно, по сравнению с фотосинтезом, эти организмы не зависят от световой энергии для производства. Это делает их важными первичными продуцентами в различных средах обитания, содержащих такие окислители, как нитраты и сульфаты.
Это делает их важными первичными продуцентами в различных средах обитания, содержащих такие окислители, как нитраты и сульфаты.
Например, в экосистемах глубоководных жерл отсутствие солнечного света означает, что фотосинтез невозможен. Из-за способности некоторых бактерий производить пищу посредством хемосинтеза они играют важную роль производителей в этой экосистеме.
Было также показано, что такое поведение приносит пользу другим организмам посредством симбиотических отношений. Например, было показано, что в различных средах азотфиксирующие бактерии образуют симбиотические отношения, которые приносят пользу различным организмам (водорослям, диатомовым водорослям, бобовым, губкам и т. д.). Здесь они способны преобразовывать азот (в изобилии в природе) в пригодные для использования формы.
Здесь эти бактерии могут катализировать атмосферный азот для производства аммиака (используя фермент, известный как нитрогеназа), который затем используется растениями для синтеза азотсодержащих биомолекул.
Одним из других симбиотических отношений, которым уделяется значительное внимание, являются отношения между трубчатыми червями (Riftia pachyptila) и хемосинтезирующими бактериями в гидротермальных источниках. В этой среде температура воды чрезвычайно высока из-за геотермального тепла. Более того, эти черви живут на морском дне (в среде, лишенной световой энергии).
Несмотря на неблагоприятные условия в этой среде (экстремально высокие температуры и недостаток света), наличие сероводорода позволяет бактериям осуществлять хемосинтез.
Используя сильно васкуляризированное жаберное перо, червь способен поглощать растворенный углекислый газ, кислород и сероводород (гемоглобин этих организмов способен связывать кислород и сульфиды). Затем они транспортируются в специализированные клетки, известные как бактериоциты, где обитают хемосинтетические бактерии.
Используя сульфид и кислород, бактерии производят энергию (АТФ), которая затем используется для преобразования углекислого газа в сахара.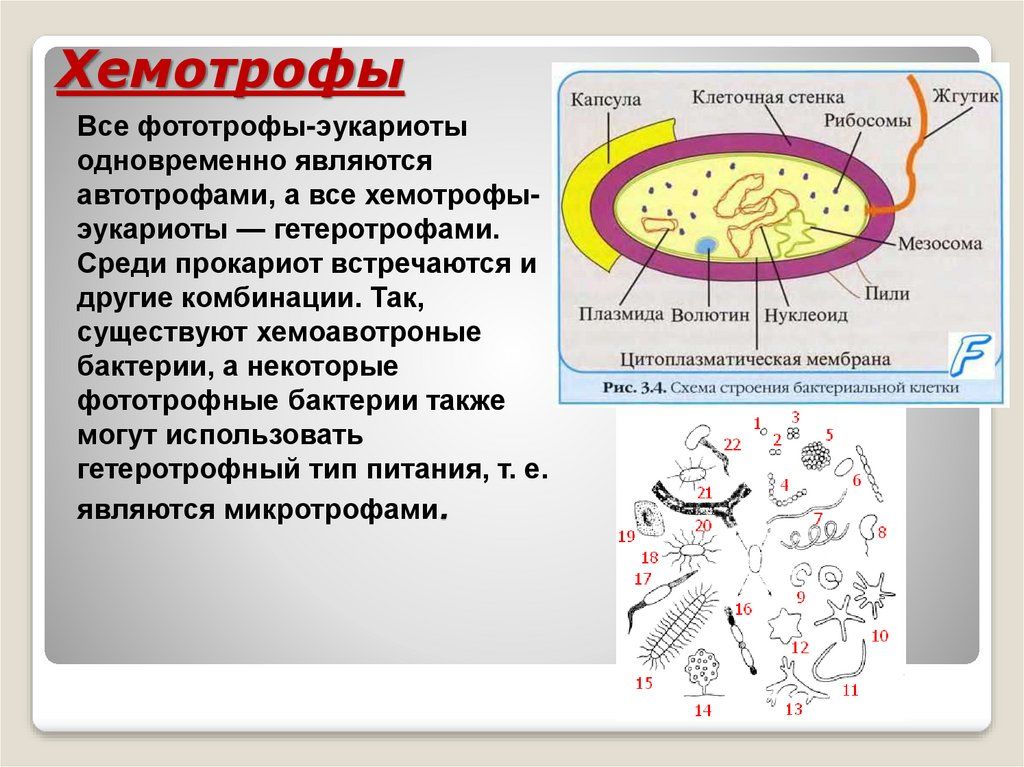 Эти сахара затем используются моллюском в качестве источника пищи.
Эти сахара затем используются моллюском в качестве источника пищи.
Such symbiotic relationships have also been identified with:
- Solemyid and lucinid bivalves
- Achinoids
- Ciliate protists
- Marine sponges
- Mussels
Some of the characteristics that have были связаны с симбионтом (хемосинтетическими бактериями), включают:
· Имеют грамотрицательную оболочку
· Различаются по форме от мелких коккоидных эндосимбионтов диаметром около 0,25 мкм до относительно крупных (около 10 мкм в длину) палочковидных хемотрофных бактерий
· В зависимости от вида они могут быть эндосимбионтами или просто поверхность тела хозяев
Возврат из Chemosynthetic Bacteria в MicroscopeMaster домой
сообщить об этом объявлении
Ссылки
Коллин М. Кавано, Зои П. Маккинс, Ирен Л.Г. Ньютон и Фрэнк Дж. Стюарт. (2006). Морские хемосинтетические симбиозы.
Кавано, Зои П. Маккинс, Ирен Л.Г. Ньютон и Фрэнк Дж. Стюарт. (2006). Морские хемосинтетические симбиозы.
Х.В. Дж аннаш. (1985). Химикосинтетическая поддержка жизни и микробное разнообразие в глубоководных гидротермальных источниках.
Дженнифер Дж. Вернегрин. (2013). Эндосимбиоз.
Зоран Минич и Премила Д. Тонгбам. (2011). Биологическое глубоководное гидротермальное отверстие как модель для изучения ферментов, улавливающих углекислый газ.
Ссылки
https://ocw.mit.edu/high-school/biology/exam-prep/клеточная-энергетика/фотосинтез/хемосинтез/
Узнайте, как размещать рекламу на MicroscopeMaster!
Хемосинтез — Образовательный проект Deep Ocean
Хемосинтез — это синтез органических соединений бактериями или другими живыми организмами с использованием энергии, полученной в результате реакций с участием неорганических химических веществ, обычно в отсутствие солнечного света.
Большая часть жизни на планете основана на пищевой цепи, которая вращается вокруг солнечного света, поскольку растения производят пищу посредством фотосинтеза. Однако в глубинах океана нет
свет, поэтому растений нет. Вместо того, чтобы солнечный свет был основной формой энергии, химическая энергия производится в процессе, называемом хемосинтезом. Места с хемосинтезирующими организмами, такие как гидротермальные источники, могут стать невероятными оазисами жизни в морских глубинах.
Хемосинтез был впервые идентифицирован в 1977 году
, когда группа ученых, участвовавших в экспедиции по исследованию океана недалеко от Галапагосских островов у побережья Эквадора, обнаружила на дне океана горячие источники, извергающие химический суп из горячей жидкости. Вокруг этих гидротермальных источников было сообщество нескольких новых видов животных, которые процветали, несмотря на то, что жили в полной темноте без доступа к солнечному свету! С тех пор эти невероятные сообщества были обнаружены в центрах распространения и зонах субдукции по всему миру.
На этой диаграмме сравниваются примеры этих двух процессов — хемосинтеза в бактериях гидротермальных источников на морском дне и фотосинтеза в наземных растениях.
Рисунок адаптирован из исследования океана NOAA.Фотосинтез
Все фотосинтезирующие организмы используют солнечную энергию для превращения углекислого газа (CO2) и воды (h3O) в сахар (C6h22O6) и кислород (O2). Существует только одна формула фотосинтеза:
.Фотосинтез происходит в растениях и некоторых бактериях везде, где достаточно солнечного света — на суше, на мелководье, даже внутри и подо льдом.
Хемосинтез ( на гидротермальных источниках) Хемосинтетические организмы используют энергию, высвобождаемую в результате химических реакций, для производства сахара или пищи. Сероводород в изобилии содержится в чрезвычайно горячей воде, извергающейся из гидротермальных источников.
Leave A Comment