
Важнейшим процессом, происходящим во всех клетках, является синтез белка. Информация о последовательности аминокислот, составляющих первичную структуру белка, заключена в ДНК. Молекулы ДНК в основном находятся в ядрах клеток (ядерная ДНК), небольшое количество ДНК содержится в митохондриях и пластидах (внеядерная ДНК).
ДНК — полинуклеотид. Каждый нуклеотид (мономер) ДНК содержит:
- пятиуглеродный сахар — дезоксирибозу,
- остаток фосфорной кислоты,
- одно из четырёх азотистых оснований: аденин, гуанин, цитозин и тимин.
Молекула дезоксирибонуклеиновой кислоты (ДНК) состоит из двух спирально закрученных цепей. Цепи в молекуле ДНК противоположно направлены. Остов цепей ДНК образован сахарофосфатными остатками, а азотистые основания одной цепи располагаются в строго определённом порядке напротив азотистых оснований другой (
Обрати внимание!
Напротив аденина одной цепи всегда располагается тимин другой цепи, напротив гуанина — цитозин.
Между аденином и тимином всегда возникают две, а гуанином и цитозином — три водородные связи.
Пара А — Т соединена двумя водородными связями, а пара Г — Ц — тремя.
Таким образом, пары нуклеотидов аденин и тимин, а также гуанин и цитозин строго соответствуют друг другу и являются комплементарными друг другу. Зная последовательность расположения нуклеотидов в одной цепи ДНК, по принципу комплементарности можно установить нуклеотиды другой (второй) цепи.
Соотношение количества нуклеотидов разных типов и азотистых оснований в молекуле ДНК определяет правило Чаргаффа (правило комплементарности).
В молекуле ДНК количество аденина равно количеству тимина, а количество гуанина — количеству цитозина: А = Т, Г = Ц.
Источники:
Каменский А. А., Криксунов Е. А., Пасечник В. В. Биология. 9 класс // ДРОФА.
Каменский А. А., Криксунов Е. А., Пасечник В. В. Биология. Общая биология (базовый уровень) 10–11 класс // ДРОФА.
Лернер Г. И. Биология: Полный справочник для подготовки к ЕГЭ: АСТ, Астрель.
http://distant-lessons.ru/nukleinovye-kisloty.html
К азотистым основаниям относят аденин (A), гуанин (G), цитозин (C) и тимин (T), который входит в состав только ДНК, а урацил (U) заменяет его в РНК. Они обладают схожими структурами и химическими свойствами. Это гетероциклические органические соединения, производные пиримидина и пурина, входящие в состав нуклеотидов. Аденин и гуанин — производные пурина, а цитозин, урацил и тимин — производные пиримидина.
Когда азотистые основания присоединяются ковалентной связью к 1′ атомам рибозы (в РНК) или дезоксирибозы (в ДНК), а к 5′-гидроксильной группе сахара присоединяется одна или несколько фосфатных групп, образуются нуклеотиды.
стандартные нуклеотиды ДНК составляют триплеты — участки ДНК, кодирующие одну аминокислоту. Например, с триплета АУГ (ему соответствует аминокислота метионин) обычно начинается синтез белка на рибосомах. Нуклеозиды, приведённые в таблице, входят в состав моно-, ди- и трифосфатов. Например, аденозин входит в состав АТФ — важнейшего энергетического ресурса организма.
Пурин C5N4h5 — гетероциклические соединения, имидазольные производные пиримидинов. Производные пурина играют важную роль в химии природных соединений (пуриновые основания ДНК и РНК; кофермент NAD; алкалоиды, кофеин и т. д.) и, благодаря этому, в фармацевтике — ядро пурина входит в состав некоторых антибиотиков. Пурин и ряд его производных обладают противоопухолевой, противовирусной и противоаллергической активностью. Аденин — азотистое основание, аминопроизводное пурина. Образует две водородных связи с урацилом (в РНК) и тимином (в ДНК) по правилу комплементарности. Представляет собой бесцветные кристаллы. Химическая формула С5H5N5. Аденин проявляет основные свойства. Аденин входит в состав многих жизненно важных для живых организмов соединений, таких как аденозин, аденозинфосфорные кислоты, нуклеиновые кислоты, адениновые нуклеотиды и др. В виде этих соединений аденин широко распространен в живой природе. Гуанин — азотистое основание, аминопроизводное пурина, является составной частью нуклеиновых кислот. Химическая формула — C5H5N5O. В ДНК и РНК образует три водородные связи с цитозином по правилу комплементарности. Производные гуанилового нуклеотида — ГДФ, ГТФ и цАМФ — участвуют во многих сигнальных путях клетки. Для некоторых процессов, происходящих в клетке — например, для сборки микротрубочек — ГТФ используется как источник энергии. Пиримидин C4N2h5 — гетероциклическое соединение, имеющее плоскую молекулу, простейший представитель диазинов. Пиримидин представляет собой бесцветные кристаллы с характерным запахом. Пиримидин проявляет свойства слабого двукислотного основания, так как атомы азота могут присоединять протоны. Производные пиримидина широко распространены в живой природе, где участвуют во многих важных биологических процессах. Его производные цитозин, тимин, урацил входят в состав нуклеотидов. Биологическая роль пиримидинов не ограничена нуклеиновыми кислотами. Некоторые пиримидиновые нуклеотиды играют важную роль в процессах обмена углеводов и липидов. Витамин В1 (тиамин) — пиримидиновое производное. Пиримидиновое ядро входит в состав некоторых коферментов и антибиотиков. Тимин — производное пиримидина. Формула C5H6N2O2. Присутствует во всех живых организмах, где вместе с дезоксирибозой входит в состав нуклеозида тимидина, который может фосфорилироваться 1-3 остатками фосфорной кислоты с образованием нуклеотидов тимидин моно-, ди- или трифосфорной кислоты (ТМФ, ТДФ и ТТФ). Дезоксирибонуклеотиды тимина входят в состав ДНК, в РНК на его месте располагается рибонуклеотид урацила. Тимин комплементарен аденину, образует с ним 2 водородные связи. Цитозин — азотистое основание, производное пиримидина. С рибозой образует нуклеозид цитидин, входит в состав нуклеотидов ДНК и РНК. Во время репликации и транскрипции по правилу комплементарности образует три водородных связи с гуанином. Представляет собой бесцветные кристаллы. Химическая формула C4H5N3O. Его производные цитозин, тимин, урацил входят в состав нуклеотидов, проявляет основные свойства. Урацил — пиримидиновое основание, которое является компонентом рибонуклеиновых кислот (РНК) и, как правило, отсутствует в дезоксирибонуклеиновых кислотах (ДНК). В составе РНК может комплементарно связываться с аденином, образуя две водородные связи. Эрвин Чаргафф открыл правила, описывающие количественные соотношения нуклеотидов. Правило Чаргаффа — биологический закон, в соответствии с которым установлены количественные соотношения между азотистыми основаниями разных типов. Для того, чтобы определить точные количественные соотношения нуклеотидов, Чаргафф разделил нуклеотиды ДНК методом бумажной хроматографии. Ему удалось выявить три закономерности:
Число аденинов равно числу тиминов, а число гуанинов — числу цитозинов: А=Т, Г=Ц
Число пуринов равно числу пиримидинов: А+Г=Т+Ц
Число аденина и цитозина равно числу гуанина и тимина: А+Ц=Г+Т
Состав ДНК разных организмов различается суммарным числом комплементарных оснований. Соотношение комплементарных нуклеотидов тоже может быть различным в разных молекулах ДНК. У одних оорганизмов в ДНК преобладают пары аденин-тимин, а у других — гуанин-цитозин. При этом правила Чаргаффа будут выполняться в любом случае.
Вопрос
Если в ДНК содержится 21 % аденина, то какова долю (%) остальных нуклеотидов, содержащихся в ДНК?
Данные рентгеноструктурного анализа показали, что молекулы ДНК имеют спиральную структуру. В 50-х годах 20 века многие химики и биологи пытались исследовать структуру ДНК. В Королевском колледже в Лондоне Морис Уилкинс и Розалинда Франклин пытались решить эту проблему методом рентгеноструктурного анализа солей ДНК. Но такой способ позволял выявить только общую структуру молекулы. Тем временем Джеймс Уотсон и Френсис Крик в Кавендишской лаборатории Кембриджского университета, используя данные, полученные М. Уилкинсом, стали строить пространственные 3-D модели ДНК. Они пытались создать структуру, которая согласовывалась бы со всеми данными рентгеноструктурного анализа. В итоге Уотсон и Крик пришли к выводу, что ДНК имеет спиральную структуру с периодичностью 0,34 нм вдоль оси. В 1953 году Дж. Уотсон и Ф. Крик открыли строение ДНК, предложив модель двойной спирали. Нуклеиновые кислоты, подобно белкам, обладают первичной структурой (под которой подразумевается их нуклеотидная последовательность) и трехмерной структурой. Интерес к структуре ДНК усилился, когда в начале XX века возникло предположение, что ДНК, возможно, представляет собой генетический материал. В начале 50-х годов американский химик, лауреат Нобелевской премии Лайнус Полинг, уже изучивший к тому времени α-спиральную структуру, характерную для многих фибриллярных белков, обратился к исследованию структуры ДНК, которая по имеющимся в то время сведениям также представлялась фибриллярной молекулой. Одновременно в Королевском колледже Морис Уилкинс и Розалинда Франклин пытались решить ту же проблему методом рентгеноструктурного анализа. Их исследования требовали долгой и трудоёмкой работы по приготовлению чистых препаратов солей ДНК, для которых удавалось получать сложные дифракционные картины. С помощью этих картин можно было, однако, выявить лишь общую структуру молекулы ДНК, не столь детализированную, как та, которую позволяли получить чистые кристаллы белка. Тем временем Фрэнсис Крик и Джеймс Уотсон в Кавендишской лаборатории Кембриджского университета избрали иной подход, который в конечном счёте и обеспечил успешное решение проблемы. Используя все физические и химические данные, какие оказались в их распоряжении, Уотсон и Крик стали строить пространственные модели ДНК в надежде на то, что рано или поздно им удастся получить достаточно убедительную структуру, согласующуюся со всеми этими данными. Истории их поисков увлекательно описаны Уотсоном в его книге «Двойная спираль». Два обстоятельства оказались для Уотсона и Крика решающими. Во-первых, они имели возможность регулярно знакомиться с результатами работ Уилкинса и, сопоставляя с его рентгенограммами свои модели, могли таким образом проверять эти модели. А рентгенограммы Уилкинса свидетельствовали в пользу спиральной структуры с периодичностью 0,34 нм вдоль оси. Во-вторых, Уотсон и Крик отдавали себе отчёт в важном значении закономерностей, касающихся соотношения различных оснований в ДНК. Обнаружил эти закономерности и сообщил о них в 1951 году Эрвин Чаргафф. Это открытие, однако, при всей своей важности не привлекло к себе должного внимания. Уотсон и Крик задались целью проверить предположение, что молекула ДНК состоит из двух спиральных полинуклеотидных цепей, удерживаемых вместе благодаря спариванию оснований, принадлежащих соседним цепям. Основания удерживаются вместе водородными связями. Аденин спаривается с тимином, гуанин — с цитозином; АТ-пара соединяется двумя водородными связями, а ГЦ-пара — тремя. Уотсон попытался представить себе такой порядок спаривания оснований и позже вспоминал об этом так: «От радости я почувствовал себя на седьмом небе, ибо тут я уловил возможный ответ на мучившую нас загадку: почему число остатков пуринов в точности равно числу остатков пиримидинов?» Уотсон увидел, что при таком сочетании основания оказываются очень точно подогнанными друг к другу, а общий размер и форма этих двух пар оснований одинаковы, так как обе пары содержат по три кольца. Водородные связи при других сочетаниях оснований в принципе возможны, но они гораздо слабее. После того как все эти обстоятельства выяснились, можно было наконец приступить к созданию достоверной модели ДНК. Уотсон и Крик показали, что ДНК состоит из двух антипараллельных (направленных в противоположные стороны) полинуклеотидных цепей. Каждая цепь закручена в спираль вправо, и обе они свиты вместе, то есть закручены вправо вокруг одной и той же оси, образуя двойную спираль. Каждая цепь состоит из сахарофосфатного остова, вдоль которого перпендикулярно длинной оси спирали располагаются азотистые основания. Находящиеся друг против друга основания двух противоположных цепей двойной спирали связаны между собой водородными связями. Расстояние между сахарофосфатными остовами двух цепей постоянно и равно расстоянию, занимаемому парой оснований, то есть одним пурином и одним пиримидином. Два пурина занимали бы слишком много места, а два пиримидина — слишком мало для того, чтобы заполнить промежутки между двумя цепями. Вдоль оси молекулы соседние пары оснований располагаются на расстоянии 0,34 нм одна от другой, чем и объясняется обнаруженная на рентгенограммах периодичность. Полный оборот спирали приходится на 3,4 нм, то есть на 10 пар оснований. Никаких ограничений относительно последовательности нуклеотидов в одной цепи не существует, но в силу правила спариваний оснований эта последовательность в одной цепи определяет собой последовательность нуклеотидов в другой цепи. Поэтому мы говорим, что две цепи двойной спирали комплементарны друг другу. Уотсон и Крик опубликовали сообщение о своей модели ДНК в журнале «Nature» в 1953 г., а в 1962 г. они вместе с Морисом Уилкинсом были удостоены за эту работу Нобелевской премии. Розалинду Франклин, умершую от рака ранее присуждения этой премии, не включили в число лауреатов, поскольку Нобелевская премия посмертно не присуждается. Для того чтобы признать, что имеющая предложенную структуру молекула может выполнять роль генетического материала, требовалось показать, что она способна: 1) нести в себе закодированную информацию и
2) точно воспроизводиться (реплицироваться). Уотсон и Крик отдавали себе отчет в том, что их модель удовлетворяет этим требованиям. В конце своей первой статьи они отметили: «От нашего внимания не ускользнуло, что постулированное нами специфическое спаривание оснований сразу же позволяет постулировать и возможный механизм копирования для генетического материала». Во второй статье, опубликованной в том же 1953 году, они обсудили выводы, которые следовали из их модели для возможного механизма передачи наследственной информации. Это открытие показало, сколь явно структура может быть связана с функцией уже на молекулярном уровне. Оно дало начало стремительному развитию молекулярной биологии.
Генетический код сложнее, разнообразнее и компактнее двоичного!
Во-первых, почему ваша формулировка «аденин = тимин, цитозин = гуанин» неверна — потому что аденин ≠ тимин и цитозин ≠ гуанин, это четыре разных азотистых основания. Да, они соответствуют друг другу по принципу комплементарности в соединяющихся цепочках, но это не значит, что они идентичны друг другу по структуре или функции. Комплементарность нужна для сохранения и передачи данных.
Цепочка ДНК двойная и на противоположных участках содержит соотнесённые азотистые основания для постоянного поддержания структуры и порядка кода, чтобы при любых повреждениях или мутациях одной цепи она могла восстановиться по второй.
К цепочке ДНК подходит только такая цепочка РНК, в которой все азотистые основания соответственно противопоставлены (аденин ДНК — урацил РНК, тимин ДНК — аденин РНК, цитозин ДНК — гуанин РНК, гуанин ДНК — цитозин РНК), потому что, с одной стороны, все мы знаем, что два одинаково заряженных магнита отталкиваются, и здесь работает примерно то же (поэтому должны быть противоположные основания, а не такие же), а с другой стороны, ключ должен идеально подходить к замку, чтобы провзаимодействовать с ним, и здесь работает примерно то же (поэтому должны быть именно противоположные основания, а не какие угодно другие).
Во-вторых, что вообще кодирует ДНК? ДНК кодирует синтез белка. После того как РНК считала информацию с неприкосновенной ДНК, эта РНК передаёт информацию другим РНК, которые, в свою очередь, плывут на рибосомы, где по этим данным синтезируются белки. Белки синтезируются из аминокислот. Мажорных аминокислот (самых распространённых) у человека всего 20, то есть белки сложены из 20 аминокислот, в очень-очень разном порядке, очень-очень разной длины, поэтому таково их разнообразие. Так, цепочка из 100 аминокислот (а это белок далеко не большой) может быть представлена более чем десятью в степени 130 (число со 130 нулями) вариантами.
Но азотистые основания — не совсем элементарные единицы кода. Тогда их было бы всего четыре и всё разнообразие кодируемых ДНК белков ограничивалось сложенными из четырёх аминокислот, но это не так, потому что аминокислот попросту больше. Четыре азотистых основания складываются в 64 варианта триплетов (по три азотистых основания), и уже они кодируют аминокислоты (некоторые триплеты кодируют одни и те же аминокислоты, а некоторые кодируют остановку считывания, поэтому триплетов больше, чем мажорных аминокислот).
Самое время сделать оговорку, что четыре (пять, учитывая урацил РНК, заменяющий тимин ДНК) выше обозначенных азотистых основания — не единственные в природе. Они, как и 20 аминокислот, мажорные. Существуют ещё и минорные азотистые основания. К тому же мы говорим только о мажорных аминокислотах! В природе насчитывается до 500 аминокислот. Представьте себе такое разнообразие белков.
Более того, в иммунных клетках возможны и более сложные варианты синтеза белка, когда данные считываются не по порядку, а во всех возможных вариациях, что порождает ещё огромное количество белков иммунной системы.
Генетический код гораздо сложнее двоичного — и гораздо более компактен! — за счёт системы последовательного кодирования:
4+ азотистых основания →
64+ триплета →
20+ аминокислот →
мириады мириад мириад… белков →
невообразимое разнообразие результатов!
Мономеры ДНК — дезоксирибонуклеотиды[править]
Дезоксирибонуклеотиды — мономеры ДНК. Каждый дезоксирибонуклеотид состоит из азотистого основания, остатка фосфорной кислоты и пятиуглеродного сахара (дезоксирибозы).
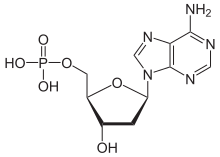 Дезоксиаденозинмонофосфат.
Дезоксиаденозинмонофосфат.Они отличаются от рибонуклеотидов строением пятиуглеродного сахара (дезоксирибоза или рибоза). Между собой дезоксирибонуклеотиды отличаются только азотистыми основаниями (аденин, гуанин, цитозин, тимин), которые присоединяются к 1′ атому дезоксирибозы ковалентной связью. Дезоксирибонуклеотиды одной цепи соединены между собой ковалентной связью, возникающей между фосфатной группой одного и 3′ атомом углерода другого дезоксирибонуклеотида. Между азотистыми основаниями дезоксирибонуклеотидов двух разных цепочек образуется три или две водородные связи (гуанин связывается с цитозином тремя связями, а аденин с тимином двумя).
Азотистые основания нуклеотидов ДНК — аденин, гуанин, тимин и цитозин[править]
К азотистым основаниям относят аденин (A), гуанин (G), цитозин (C) и тимин (T), который входит в состав только ДНК, а урацил (U) заменяет его в РНК. Они обладают схожими структурами и химическими свойствами. Это гетероциклические органические соединения, производные пиримидина и пурина, входящие в состав нуклеотидов. Аденин и гуанин — производные пурина, а цитозин, урацил и тимин — производные пиримидина.
Когда азотистые основания присоединяются ковалентной связью к 1′ атомам рибозы (в РНК) или дезоксирибозы (в ДНК), а к 5′-гидроксильной группе сахара присоединяется одна или несколько фосфатных групп, образуются нуклеотиды.
стандартные нуклеотиды ДНК составляют триплеты — участки ДНК, кодирующие одну аминокислоту. Например, с триплета АУГ (ему соответствует аминокислота метионин) обычно начинается синтез белка на рибосомах.
В таблице приведена структура главных азотистых оснований.
Нуклеозиды, приведённые в таблице, входят в состав моно-, ди- и трифосфатов. Например, аденозин входит в состав АТФ — важнейшего энергетического ресурса организма.
Пуринs C5N4H4 — гетероциклические соединения, имидазольные производные пиримидинов. Производные пурина играют важную роль в химии природных соединений (пуриновые основания ДНК и РНК; кофермент NAD; алкалоиды, кофеин и т. д.) и, благодаря этому, в фармацевтике — ядро пурина входит в состав некоторых антибиотиков. Пурин и ряд его производных обладают противоопухолевой, противовирусной и противоаллергической активностью.
В таблице приведены производные пурина.
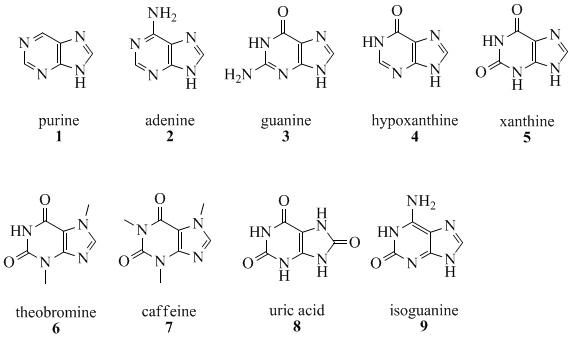
Аденин — азотистое основание, аминопроизводное пурина. Образует две водородных связи с урацилом (в РНК) и тимином (в ДНК) по правилу комплементарности.
Представляет собой бесцветные кристаллы. Химическая формула С5H5N5. Аденин проявляет основные свойства.
Аденин входит в состав многих жизненно важных для живых организмов соединений, таких как аденозин, аденозинфосфорные кислоты, нуклеиновые кислоты, адениновые нуклеотиды и др. В виде этих соединений аденин широко распространен в живой природе.
Гуанин — азотистое основание, аминопроизводное пурина, является составной частью нуклеиновых кислот. Химическая формула — C5H5N5O.
В ДНК и РНК образует три водородные связи с цитозином по правилу комплементарности. Производные гуанилового нуклеотида — ГДФ, ГТФ и цАМФ — участвуют во многих сигнальных путях клетки. Для некоторых процессов, происходящих в клетке — например, для сборки микротрубочек — ГТФ используется как источник энергии.
Пиримидин C4N2H4 — гетероциклическое соединение, имеющее плоскую молекулу, простейший представитель диазинов.
Пиримидин представляет собой бесцветные кристаллы с характерным запахом. Пиримидин проявляет свойства слабого двукислотного основания, так как атомы азота могут присоединять протоны. Производные пиримидина широко распространены в живой природе, где участвуют во многих важных биологических процессах. Его производные цитозин, тимин, урацил входят в состав нуклеотидов.
Биологическая роль пиримидинов не ограничена нуклеиновыми кислотами. Некоторые пиримидиновые нуклеотиды играют важную роль в процессах обмена углеводов и липидов. Витамин В1 (тиамин) — пиримидиновое производное. Пиримидиновое ядро входит в состав некоторых коферментов и антибиотиков.
Тимин — производное пиримидина. Формула C5H6N2O2.
Присутствует во всех живых организмах, где вместе с дезоксирибозой входит в состав нуклеозида тимидина, который может фосфорилироваться 1-3 остатками фосфорной кислоты с образованием нуклеотидов тимидин моно-, ди- или трифосфорной кислоты (ТМФ, ТДФ и ТТФ).
Дезоксирибонуклеотиды тимина входят в состав ДНК, в РНК на его месте располагается рибонуклеотид урацила. Тимин комплементарен аденину, образует с ним 2 водородные связи.
Цитозин — азотистое основание, производное пиримидина. С рибозой образует нуклеозид цитидин, входит в состав нуклеотидов ДНК и РНК. Во время репликации и транскрипции по правилу комплементарности образует три водородных связи с гуанином.
Представляет собой бесцветные кристаллы. Химическая формула C4H5N3O. Его производные цитозин, тимин, урацил входят в состав нуклеотидов, проявляет основные свойства.
Урацил — пиримидиновое основание, которое является компонентом рибонуклеиновых кислот (РНК) и, как правило, отсутствует в дезоксирибонуклеиновых кислотах (ДНК). В составе РНК может комплементарно связываться с аденином, образуя две водородные связи.
Эрвин Чаргафф открыл правила, описывающие количественные соотношения нуклеотидов[править]
Правило Чаргаффа — биологический закон, в соответствии с которым установлены количественные соотношения между азотистыми основаниями разных типов. Для того, чтобы определить точные количественные соотношения нуклеотидов, Чаргафф разделил нуклеотиды ДНК методом бумажной хроматографии. Ему удалось выявить три закономерности:
- Число аденинов равно числу тиминов, а число гуанинов — числу цитозинов: А=Т, Г=Ц
- Число пуринов равно числу пиримидинов: А+Г=Т+Ц
- Число аденина и цитозина равно числу гуанина и тимина: А+Ц=Г+Т
Состав ДНК разных организмов различается суммарным числом комплементарных оснований. Соотношение комплементарных нуклеотидов тоже может быть различным в разных молекулах ДНК. У одних оорганизмов в ДНК преобладают пары аденин-тимин, а у других — гуанин-цитозин. При этом правила Чаргаффа будут выполняться в любом случае.
- Вопрос
- Если в ДНК содержится 21 % аденина, то какова долю (%) остальных нуклеотидов, содержащихся в ДНК?
Данные рентгеноструктурного анализа показали, что молекулы ДНК имеют спиральную структуру[править]
В 50-х годах 20 века многие химики и биологи пытались исследовать структуру ДНК. В Королевском колледже в Лондоне Морис Уилкинс и Розалинда Франклин пытались решить эту проблему методом рентгеноструктурного анализа солей ДНК. Но такой способ позволял выявить только общую структуру молекулы. Тем временем Джеймс Уотсон и Френсис Крик в Кавендишской лаборатории Кембриджского университета, используя данные, полученные М. Уилкинсом, стали строить пространственные 3-D модели ДНК. Они пытались создать структуру, которая согласовывалась бы со всеми данными рентгеноструктурного анализа. В итоге Уотсон и Крик пришли к выводу, что ДНК имеет спиральную структуру с периодичностью 0,34 нм вдоль оси.
В 1953 году Дж. Уотсон и Ф. Крик открыли строение ДНК, предложив модель двойной спирали[править]
Нуклеиновые кислоты, подобно белкам, обладают первичной структурой (под которой подразумевается их нуклеотидная последовательность) и трехмерной структурой. Интерес к структуре ДНК усилился, когда в начале XX века возникло предположение, что ДНК, возможно, представляет собой генетический материал.
В начале 50-х годов американский химик, лауреат Нобелевской премии Лайнус Полинг, уже изучивший к тому времени α-спиральную структуру, характерную для многих фибриллярных белков, обратился к исследованию структуры ДНК, которая по имеющимся в то время сведениям также представлялась фибриллярной молекулой. Одновременно в Королевском колледже Морис Уилкинс и Розалинда Франклин пытались решить ту же проблему методом рентгеноструктурного анализа. Их исследования требовали долгой и трудоёмкой работы по приготовлению чистых препаратов солей ДНК, для которых удавалось получать сложные дифракционные картины. С помощью этих картин можно было, однако, выявить лишь общую структуру молекулы ДНК, не столь детализированную, как та, которую позволяли получить чистые кристаллы белка.
Тем временем Фрэнсис Крик и Джеймс Уотсон в Кавендишской лаборатории Кембриджского университета избрали иной подход, который в конечном счёте и обеспечил успешное решение проблемы. Используя все физические и химические данные, какие оказались в их распоряжении, Уотсон и Крик стали строить пространственные модели ДНК в надежде на то, что рано или поздно им удастся получить достаточно убедительную структуру, согласующуюся со всеми этими данными. Истории их поисков увлекательно описаны Уотсоном в его книге «Двойная спираль»[1].
Два обстоятельства оказались для Уотсона и Крика решающими. Во-первых, они имели возможность регулярно знакомиться с результатами работ Уилкинса и, сопоставляя с его рентгенограммами свои модели, могли таким образом проверять эти модели. А рентгенограммы Уилкинса свидетельствовали в пользу спиральной структуры с периодичностью 0,34 нм вдоль оси. Во-вторых, Уотсон и Крик отдавали себе отчёт в важном значении закономерностей, касающихся соотношения различных оснований в ДНК. Обнаружил эти закономерности и сообщил о них в 1951 году Эрвин Чаргафф. Это открытие, однако, при всей своей важности не привлекло к себе должного внимания.
Уотсон и Крик задались целью проверить предположение, что молекула ДНК состоит из двух спиральных полинуклеотидных цепей, удерживаемых вместе благодаря спариванию оснований, принадлежащих соседним цепям. Основания удерживаются вместе водородными связями. Аденин спаривается с тимином, гуанин — с цитозином; АТ-пара соединяется двумя водородными связями, а ГЦ-пара — тремя. Уотсон попытался представить себе такой порядок спаривания оснований и позже вспоминал об этом так: «От радости я почувствовал себя на седьмом небе, ибо тут я уловил возможный ответ на мучившую нас загадку: почему число остатков пуринов в точности равно числу остатков пиримидинов?»
Уотсон увидел, что при таком сочетании основания оказываются очень точно подогнанными друг к другу, а общий размер и форма этих двух пар оснований одинаковы, так как обе пары содержат по три кольца. Водородные связи при других сочетаниях оснований в принципе возможны, но они гораздо слабее. После того как все эти обстоятельства выяснились, можно было наконец приступить к созданию достоверной модели ДНК.
Уотсон и Крик показали, что ДНК состоит из двух антипараллельных (направленных в противоположные стороны) полинуклеотидных цепей. Каждая цепь закручена в спираль вправо, и обе они свиты вместе, то есть закручены вправо вокруг одной и той же оси, образуя двойную спираль. Каждая цепь состоит из сахарофосфатного остова, вдоль которого перпендикулярно длинной оси спирали располагаются азотистые основания. Находящиеся друг против друга основания двух противоположных цепей двойной спирали связаны между собой водородными связями. Расстояние между сахарофосфатными остовами двух цепей постоянно и равно расстоянию, занимаемому парой оснований, то есть одним пурином и одним пиримидином. Два пурина занимали бы слишком много места, а два пиримидина — слишком мало для того, чтобы заполнить промежутки между двумя цепями. Вдоль оси молекулы соседние пары оснований располагаются на расстоянии 0,34 нм одна от другой, чем и объясняется обнаруженная на рентгенограммах периодичность. Полный оборот спирали приходится на 3,4 нм, то есть на 10 пар оснований. Никаких ограничений относительно последовательности нуклеотидов в одной цепи не существует, но в силу правила спариваний оснований эта последовательность в одной цепи определяет собой последовательность нуклеотидов в другой цепи. Поэтому мы говорим, что две цепи двойной спирали комплементарны друг другу.
Уотсон и Крик опубликовали сообщение о своей модели ДНК в журнале «Nature» в 1953 г., а в 1962 г. они вместе с Морисом Уилкинсом были удостоены за эту работу Нобелевской премии. Розалинду Франклин, умершую от рака ранее присуждения этой премии, не включили в число лауреатов, поскольку Нобелевская премия посмертно не присуждается.
Для того чтобы признать, что имеющая предложенную структуру молекула может выполнять роль генетического материала, требовалось показать, что она способна: 1) нести в себе закодированную информацию и 2) точно воспроизводиться (реплицироваться). Уотсон и Крик отдавали себе отчет в том, что их модель удовлетворяет этим требованиям. В конце своей первой статьи они отметили: «От нашего внимания не ускользнуло, что постулированное нами специфическое спаривание оснований сразу же позволяет постулировать и возможный механизм копирования для генетического материала». Во второй статье, опубликованной в том же 1953 году, они обсудили выводы, которые следовали из их модели для возможного механизма передачи наследственной информации. Это открытие показало, сколь явно структура может быть связана с функцией уже на молекулярном уровне. Оно дало начало стремительному развитию молекулярной биологии.
[2]Дж. Уотсон. Двойная спираль. (русский перевод).
⇐ ПредыдущаяСтр 25 из 25
Нуклеотиды – это мономеры нуклеиновых кислот. Нуклеиновые кислоты в эукариотических клетках находятся в ядре. Они есть у всех живых организмов. Мономеры, из которых потом строятся нуклеиновые кислоты, состоят из азотистого основания, остатка сахара (дезоксирибоза или рибоза) и фосфата. Сахара вместе с азотистым основанием называются нуклеозидами (аденозин, гуанозин, тимидин, цитидин). Если к ним присоединены 1-, 2-, или 3-фосфорных остатка, то вся эта структура называется соответственно, нуклеотизид монофосфатом, дифосфатом или трифосфатом или нуклеотидом (аденин, гуанин, тимин, цитозин).
Рис. 20. Модель АТФ в пространстве
Азотистое основание, входящее в состав ДНК делится на две группы – пиримидиновую и пуриновую. В состав ДНК входит аденин, тимин, цитозин и гуанин, в РНК вместо тимина урацил. Как известно, ДНК – это большой архив, в котором хранится информация, а РНК – это молекула, которая переносит информацию из ядра в цитоплазму для синтеза белков. С различием в функциях связаны различия в строении. РНК более химически активно из-за того, что ее сахар — рибоза – имеет в своем составе гидроксильную группу, а в дезоксирибозе кислорода нет. Из-за отсутствия кислорода ДНК более инертно, что важно для ее функции хранения информации, чтобы она не вступала ни в какие реакции.
Нуклеотиды способны взаимодействовать друг с другом, при этом «выбрасывается» два фосфора, и между соседними нуклеотидами образуется связь. В молекуле фуранозы молекулы углерода пронумерованы. С первым связано азотистое основание. Когда образуется цепочка нуклеотидов, связь осуществляется между пятым углеродом одной и третьим углеродом другой фосфорной кислоты. Поэтому в цепочке нуклеиновых кислот выделяют разные неравнозначные концы, относительно которых молекула не симметрична.
Комплементарные друг другу одноцепочечные молекулы нуклеиновой кислоты способны образовывать двухцепочечную структуру. Внутри этой спирали аденин образует пару с тимином, а гуанин — с цитозином. Встречается утверждение, что нуклеотиды подходят друг другу, как осколки разбитого стекла, поэтому они и образуют пары. Но это утверждение неверно. Нуклеотиды способны образовывать пары как угодно. Единственная причина, по которой они соединяются так, и никак иначе, заключается в том, что угол между «хвостиками», которые идут к сахарам, совпадает только в этих парах, и, кроме того, совпадают их размеры. Никакая другая пара не образует такой конфигурации. А поскольку они совпадают, то их через сахаро-фосфатный остов можно связать друг с другом. Структуру двойной спирали открыли в 1953 году Джеймс Уотсон и Фрэнсис Крик.
Рис. 21. Модель ДНК
При соединении друг с другом против 5-конца одной нити находится 3-конец другой нити. То есть нити идут в противоположных направлениях – говорят, что нити в ДНК антипараллельны. На рисунке 21 видно, что аденин соединяется с тимином двумя водородными связями, а гуанин соединяется с цитозином тройной водородной связью. Если молекулу ДНК подогревать, то ясно, что две связи легче разорвать, чем три, это существенно для свойств ДНК. В силу пространственного расположения сахаро-фосфатного остова и нуклеотидов, когда нуклеотиды накладывают один на другой и «сшивают» через сахаро-фосфатный остов, цепочка начинает заворачиваться, тем самым, образуя знаменитую двойную спираль.
При нагревании ДНК водородные связи разрываются, и нити в двойной спирали расплетаются. Процесс нагревания называется плавлением ДНК, при этом разрушаются связи между парами А-Т и Г-Ц .Чем больше в ДНК пар А-Т, тем менее прочно нити друг с другом связаны, тем легче ДНК расплавить. Переход из двухспиральной ДНК в односпиральную измеряется на спектрофотометрах по поглощению света при 260 нм. Температура плавления ДНК зависит от А-Т/Г-Ц состава и размера фрагмента молекулы. Ясно, что если фрагмент состоит из нескольких десятков нуклеотидов, то его гораздо легче расплавить, чем более длинные фрагменты.
У человека в гаплоидном геноме, то есть единичном наборе хромосом, 3 млрд. пар нуклеотидов, и их длина составляет 1,7 м, а клетка гораздо меньше, как вы догадываетесь. Для того чтобы ДНК смогла в ней поместиться, она достаточно плотно свернута, и в эукариотической клетке свернуться ей помогают белки – гистоны. Гистоны имеют положительный заряд, а так как ДНК заряжена отрицательно, то гистоны обладают сродством к ДНК. Упакованная при помощи гистонов ДНК имеет вид бусин, называемых нуклеосомами. 200 пар нуклеотидов идет на одну нуклеосому, 146 пар накручиваются на гистоны, а остальные 54 висят в виде линкерных (связывающих нуклеосомы) ДНК. Это первый уровень компактизации ДНК. В хромосомах ДНК свернута еще несколько раз для того, чтобы образовались компактные структуры.
К нуклеиновым кислотам кроме ДНК относится также РНК. В клетке присутствуют разные типы РНК: рибосомные, матричные, транспортные. РНК синтезируется в виде одноцепочечной молекулы, но отдельные ее участки входят в состав двухцепочечных спиралей. Для РНК также говорят о первичной структуре (последовательности нуклеотидов) и вторичной структуре (образование двуспиральных участков).
Рис. 23. Вторичная структура тРНК
Липиды
В состав липидов входят жирные кислоты, имеющие длинные углевододородные цепи. Жирные кислоты гидрофобны, то есть, не растворимы в воде.
Таблица 8
Природные жирные кислоты
В клетке важную роль играют липиды, в которых к глицерину присоединен остаток фосфорной кислоты и 2 жирных кислоты. Они называются фосфолипидами. Молекулы фосфолипидов имеют полярную (то есть гидрофильную, хорошо растворимую) группу на одном конце молекулы и длинный гидрофобный хвост. К фосфолипидам относится фосфатидилхолин. В водном растворе фосфолипиды образуют мицеллы, в которых молекулы обращены полярными «головами» наружу, в сторону воды, а гидрофобные «хвосты» оказываются внутри мицеллы, спрятанными от воды. Клеточную мембрану образуют также липиды с полярными «головами», которые обращены наружу по обе стороны мембраны, а гидрофобные «хвосты» находятся внутри липидного бислоя.
Библиографический список
1. Ахметов, Н. С. Общая и неорганическая химия [Текст]: учеб. для химико-технол. специальностей вузов — М.: Высш. шк., 1998. — 743 с.
2. Гельфман, М.И. Химия [Текст] / М.И. Гельфман, В.П. Юстратов – СПб.: Лань, 2001. – 480 с.
3. Глинка, Н.Л. Общая химия [Текст]: учеб. пособие для вузов. – Л.: Химия, 1983. – 704 с.
4. Карапетьянц, М.Х. Общая и неорганическая химия [Текст]: учебник для вузов / М.Х. Карапетьянц, С.И. Дракин – М.: Химия, 1994. – 592 с.
5. Коровин, Н.В. Общая химия [Текст]: учебник для технических направ. и спец. вузов. – М.: Высш.школа, 2000. – 558 с.
6. Кудряшов И.В., Сборник примеров и задач по физической химии [Текст]: учеб. пособие для хим.-технол. спец. вузов. – М.: Высш. школа, 1991 – 527 с.
7. Общая химия. Биофизическая химия. Химия биогенных элементов [Текст]: учеб. для вузов по мед., биол., агрон., ветеринар., экол. специальностям/ Ю. А. Ершов, В. А. Попков, А. С. Берлянд, А. З. Книжник — М.: Высш. шк., 2000. — 560 с.
8. Пузаков, С.А. Сборник задач и упражнений по общей химии [Текст]: учеб. пособие / С.А. Пузаков, В.А. Попков, А.А. Филиппова. – М.: Высш. школа, 2004. – 255 с.
9. Суворов, А.В. Общая химия [Текст]: учеб. пособие для вузов / А.В. Суворов, А.Б. Никольский – СПб: Химия, 1995 – 624 с
10. Угай, Я. А. Общая и неорганическая химия [Текст]: учеб. для вузов по направлению и специальности «Химия» — М.: Высш. школа, 1997. — 527 с.
11. Харин, А.Н. Курс химии [Текст]: учебник для приборостроит. спец. вузов / А.Н. Харин, Н.А. Катаева, Л.Т. Харина. – М.: Высш. школа, 1983. – 511 с.
12. http://www.bio.fizten.ru
13. http://www.kazchem.ru
УЧЕБНОЕ ИЗДАНИЕ
Денисова Елена Сергеевна
Конспект лекций по дисциплине «Химия».
Учебное пособие
Ответственный за выпуск: И.А. Кибанова
Ответственный редактор: В.А. Филатова
Подписано в печать 18.05.2009
Формат 60х80/16. Бумага ксероксная
Гарнитура Times New Roman
Оперативный способ печати
Усл.печ.л. 10, уч.-изд.л. 10
Тираж 100 экз. Заказ № 31
Цена договорная
Омский институт водного транспорта (филиал)
ФГОУ ВПО «НГАВТ»
644099, г. Омск, ул. И. Алексеева, 4
Отпечатано в ООО «Технотранс»
644012, г. Омск, ул. 9 Дунайская, 20
Рекомендуемые страницы:
Согласно гипотезе Уотсона — Крика, четыре основания — аденин, ТИМИН, гуанин -и цитозин, обозначаемые соответственно буквами А, Т, О и С, — расположены в определенной последовательности в одной из [c.456]
В ДНК две спирали связаны друг с другом через определенные интервалы при помощи водородных связей. Изучение молекулярных моделей показало, что водородные связи могут образоваться лишь между аденином и тимином и между гуанином и цитозином водородные связи между другими парами оснований не позволят им разместиться в структуре двойной спирали. В соответствии с этим представлением отношения аденин тимин и гуанин цитозин равны 1 1. [c.1063]
РНК исследовались методом рентгеноструктурного анализа с целью установления того, имеют ли их полинуклеотидные цепи спиралевидную форму, аналогичную ДНК, но до настоящего времени определенных выводов сделать не удалось из-за отсутствия однородных образцов кристаллической РНК. Однако дрожжевую транспортную РНК удалось очистить и получить в кристаллическом состоянии. Получены хорошие рентгенограммы этого вещества, и они оказались чрезвычайно сходными с теми, которые дает ДНК таким образом, эти две структуры должны быть близкими. В соответствии с этим вероятное строение дрожжевой транспортной РНК должно быть таким, при котором каждая полинуклеотидная цепочка сложена вдвое по всей длине и скручена таким образом, что образует двойную спираль. Две половины цепи РНК соответствуют, следовательно, комплементарным цепям ДНК. Не вызывает сомнения, что спиральная структура поддерживается за счет водородных связей между парами оснований аденин — урацил и гуанин — цитозин (партнеры, образующие такую пару, находятся в разных половинах цепи). В месте перегиба цепи имеется несколько неспаренных оснований, и небольшой хвост неспаренных оснований имеется на одном из концов цепи. В этом состоит главное отличие спиральной конфигурации дрожжевой транспортной РНК от спиральной конфигурации ДНК. Дифракционная картина, полученная при рентгеноструктурном исследовании РНК из других источников, сходна с дифракционной картиной, полученной при исследовании дрожжевой транспортной РНК следовательно, спиральная конфигурация присуща, по-видимому, многим формам РНК. [c.142]
Полимерные молекулы ДНК состоят из расположенных в определенных последовательностях остатков четырех типов дезоксирибонуклеотидов, в которые в качестве углеводного компонента входит дезоксирибоза, а гетероциклическими основаниями являются аденин, гуанин, цитозин и тимин [c.443]
Нуклеиновые кислоты содержатся в каждой живой клетке. ДНК служит носителем генетической информации. Это обусловлено тесной связью между двумя витками спиралей нуклеиновых кислот, которая основана на очень специфических водородных связях между адениновым (А) остатком одного витка и тиминовым (Т) остатком другого витка, который расположен строго напротив первого, а также между цитозиновым (Ц) остатком одного витка и гуаниновым (Г) остатком другого. Такое образование пар абсолютно специфично аденин не может образовывать мультивалентные водородные связи с гуанином или цитозином, а цитозин не может образовывать связи с тимином или аденином. Изумительно, что вся наследственность и эволюция зависят от двух групп водородных связей Генетический код для синтеза определенной аминокислоты обус- [c.578]
Определенные основания в ДНК могут подвергаться изменениям под действием алкилирующих агентов например, диметилсульфат (рис. 30-5), обладающий высокой реакционной способностью, метилирует остатки гуанина. В результате образуется О-метилгуанин (рис. 30-7), который не способен спариваться с обычным партнером гуанина цитозином. Как у бактерии, так и в животных тканях имеются ферменты, которые специфически удаляют О-метилгуанин и заменяют его на нормальный гуанин. И в этом случае репарация осуществляется по механизму разрезал-залатал-зашил , сходному с тем, который показан на рис. 30-4. [c.968]
Здесь видно, как из одной молекулы ДНК могут образоваться две новые, совершенно тождественные по своей структуре молекулы. В самом деле, после расхождения цепей к основаниям одиночных цепей из окружающей среды присоединяются комплементарно соответствующие основания (или нуклеотиды) к аде-нину — тимин, к тимину — аденин, к гуанину — цитозин и, наконец, к цитозину — гуанин. В результате этого определяется расположение оснований во вто рой цепи в строгом соответствии с их расположением в первой цепи. Затем ферментные системы смыкают установленные определенным образом нуклеотиды во вторую комплементарную цепь, которая уже соединена с первой основной цепью водородными связями. Этот процесс специфического синтеза ДНК имеет место в период деления клетки, когда, как показывает химический анализ, действительно происходит удвоение количества ДНК. [c.60]
Последовательность нуклеотидов и генетический код. Методы определения последовательности аминокислот в полипептидной цепи были известны еще в 50-х гг. Теоретически это относительно легкая проблема, поскольку все 20 аминокислот, встречающиеся в природных белках, имеют разные свойства. С другой стороны, нуклеотидная последовательность ДНК относительно однородна по составу элементарных звеньев, так как содержит только четыре типа азотистых оснований-гуанин, цитозин, аденин и тимин. Когда еще в 60-х г. был расшифрован генетический код, появилась возможность восстанавливать (дедуцировать) нуклеотидную последовательность транскрибируемой ДНК по аминокислотной последовательности соответствующего белка. Однако генетический код является вырожденным, то есть одной и той же аминокислоте соответствуют несколько разных нуклеотидных триплетов. Следовательно, суждения о нуклеотидной последовательности, основанные на последовательности аминокислот в белке, не однозначны. Кроме того, последовательности аминокислот не содержат никакой информации о последовательности некодирующих участков ДНК. В настоящее время разработаны методы непосредственного секвенирования ДНК [117]. Принцип состоит в следующем длинную молекулу ДНК фрагментируют при помощи агентов, расщепляющих ее в специфических сайтах. Затем определяют последовательность нуклеотидов в каждом из этих фрагментов. Очередность фрагментов в целой молекуле восстанавливают, используя перекрывающиеся концы идентичные цепи разрезают повторно другой рестриктазой, а затем последовательности перекрывающихся фрагментов, образующихся при обработке двумя рестриктазами разной специфичности, сравнивают. Так может быть реконструирована полная последовательность. В пределах от
Урок 7. Нуклеиновые кислоты
В клетке имеются два типа нуклеиновых кислот: рибонуклеиновая кислота (РНК) и дезоксирибонуклеиновая кислота (ДНК). И ДНК, и РНК – полимерные цепочки, состоящие из отдельных звеньев – нуклеотидов. Сами нуклеотиды тоже имеют довольно сложное строение. Они содержат азотистое основание, моносахаридный остаток и остаток фосфорной кислоты.
Главная функция ДНК – хранение генетической информации: гены представляют собой участки молекул ДНК, в которых хранится информация обо всех белках и РНК организма. Изменения в структуре ДНК называются мутациями. РНК выполняет в клетке несколько важных функций, связанных с синтезом белка. Основные свойства молекул ДНК и РНК суммированы в табл. 1.
|
Таблица 1 |
|||||||||||||
|
Основу цепочек ДНК составляет «сахаро-фосфатный остов» – остатки дезоксирибозы, соединенные через остатки фосфорной кислоты. В этой цепочке азотистое основание присоединено к первому углеродному атому дезоксирибозы, «свой» фосфат – к пятому, а по третьему атому данное нуклеотидное звено соединяется с фосфатом другого звена (см. урок 3).
|
Рис. 1. Структура цепей ДНК |
Молекула ДНК представляет собой две цепочки нуклеотидов, спирально закрученные вокруг общей оси. Азотистые основания двух цепочек располагаются друг напротив друга перпендикулярно оси спирали. На один виток спирали приходится 10 пар азотистых оснований, диаметр спирали составляет 2,37 нанометра.
Цепочки ДНК удерживаются в двойной спирали благодаря водородным связям между азотистыми основаниями, в состав которых входят атомы кислорода и азота с частично отрицательным зарядом, а также атомы водорода, на которых имеется частичный положительный заряд. Благодаря электростатическому притяжению могут возникать связи между азотистыми основаниями, причем отнюдь не между любыми из них. Полностью подходящими будут пары «аденин–тимин» и «гуанин–цитозин». Такое строгое соответствие оснований одной и другой цепочки получило название комплементарности (дополнительности). Пара «аденин–тимин» соединяется двумя водородными связями, а пара «гуанин-цитозин» – тремя. При повышении температуры раствора происходит денатурация ДНК – двойная спираль расплетается на отдельные цепочки.
|
Модель 1. Комплементарность оснований в ДНК |
Если мысленно двигаться по двойной спирали, то по одной цепочке мы будем перемещаться от третьего гидроксила одной дезоксирибозы к пятому другой дезоксирибозы, а по второй цепочке будет происходить движение от пятого гидроксила одной дезоксирибозы к третьему другой. Иными словами, цепочки в ДНК противоположно направлены, или антипараллельны.
Двуспиральные молекулы ДНК выполняют роль генетического материала у всех клеточных организмов. У вирусов генетический материал более разнообразен. Кроме двухцепочечных молекул ДНК, им могут быть одноцепочечные ДНК, одно- или двухцепочечные РНК.
У эукариот большая часть ДНК сосредоточена в ядре – во время деления она конденсируется в хромосомах, а между делениями распределена в виде хроматина по всему пространству ядра. ДНК в хромосомах эукариот линейна: в состав каждой хромосомы только что поделившейся клетки входит одна очень длинная двойная спираль ДНК. У бактерий имеется кольцевая ДНК в единственной «хромосоме», а, кроме того, бывает несколько небольших кольцевых ДНК – так называемые плазмиды. В клетках эукариот ДНК содержится не только в ядре, но также в хлоропластах и митохондриях. Хлоропласты имеют кольцевые ДНК. ДНК митохондрий, как правило, кольцевая, но иногда бывает и линейной.
Молекулы ДНК имеют огромную длину. У человека одинарный набор из 23 хромосом, находящийся в гаметах, содержит около 3 миллиардов пар нуклеотидов. Если вытянуть все эти цепочки в одну нить, ее длина будет равна примерно 1 м. Между митозами ДНК в виде хроматина распределена по всему объему ядра. При митозе ДНК находится в виде очень коротких (около микрометра) хромосом; следовательно, при этом происходит сильная компактизация укладки этой молекулы.
У эукариот первый уровень компактизации составляет нуклеосомная укладка. В хромосомах, кроме ДНК, содержатся щелочные белки – гистоны, у человека имеется пять их видов: Н1, h3A, h3B, h4 и h5. По две молекулы гистонов h3A, h3B, h4 и h5 образуют комплекс-октамер, по форме напоминающий бусину. Двойная спираль ДНК намотана на эти бусины, образованная структура называется нуклеосомой. При этом ДНК, как и следовало ожидать от кислоты, образует электростатические нековалентные связи с гистонами. На нуклеосому навернуто 1,75 оборота ДНК – примерно 160–170 пар оснований.
|
Рис. 2. Нуклеосомая укладка ДНК. Сине-красная двойная спираль ДНК, намотанная на октамер гистонов (на рисунке видны их α–спиральные участки), показана с разных углов зрения. |
Накрученная на нуклеосому двойная спираль в 7 раз короче свободной. «Бусы» из нуклеосом соединяются друг с другом с помощью гистона Н1, что обеспечивает еще большую компактизацию. Дальнейшие уровни укладки ДНК пока изучены не достаточно, однако доказано, что цепочки ДНК по всей длине хромосомы нигде не прерываются. Укладка ДНК влияет на генетическую активность. В целом, чем компактнее уложена ДНК, тем менее она активна – в хромосомах во время митоза почти не происходит ни активной работы генов, ни синтеза новой ДНК.
У бактерий ДНК довольно короткая и на клетку приходится всего одна «хромосома» – а значит, нет и задачи все компактно упаковать, чтобы точнее «растащить» по дочерним клеткам. Гистоны у прокариот отсутствуют.
Количество генетического материала сильно отличается у разных организмов. Так, геном примитивных бактерий – микоплазм – содержит от нескольких сотен тысяч до двух миллионов нуклеотидных пар; у других бактерий эта величина составляет несколько миллионов. У разных грибов и водорослей геном достигает десятков миллионов пар оснований, у червей может превышать сто миллионов, у членистоногих, моллюсков и иглокожих лежит в пределах от ста миллионов до пяти миллиардов. Хрящевые рыбы имеют геномы размером несколько миллиардов пар оснований, у костных рыб он варьирует от сотен миллионов до нескольких миллиардов. У амфибий вариации размеров генома очень широки: от сотен миллионов до шестидесяти миллиардов нуклеотидных пар. Рептилии и млекопитающие ограничиваются геномом в несколько миллиардов пар оснований; геном птиц в среднем составляет около миллиарда. Наконец, цветковые растения имеют чрезвычайно разнообразный размер генома – от сотен миллионов до более сотни миллиардов нуклеотидных пар. В целом можно заметить существенное увеличение размеров генома при переходе от прокариот к эукариотам и от одноклеточных организмов к многоклеточным.
В хлоропластах и митохондриях содержится гораздо меньше ДНК, чем в ядре: обычный размер генома хлоропластов составляет 120–200 тысяч пар нуклеотидов, а в геноме митохондрий человека имеется всего 16569 нуклеотидных пар.
Для того чтобы клетка могла поделиться, должно произойти удвоение молекул ДНК – репликация – что позволит каждой дочерней клетке заиметь свою копию молекулы наследственности. В процессе репликации двойная спираль временно расплетается, и каждая цепочка служит матрицей для синтеза новой цепи. Отдельные нуклеотиды, имеющиеся в клетке, присоединяются к матрице по принципу комплементарности – напротив аденина встраивается тимин, напротив гуанина – цитозин. Новые нуклеотиды присоединяются только к 3′-концу (т. е. к гидроксилу третьего атома углерода дезоксирибозы) растущей цепочки, но не к 5′-концу (не к пятому атому). В результате образуются две одинаковые двойные спирали, и каждая из них при делении попадет в одну из дочерних клеток.
Заметим, что каждой клетке достанется одна цепь из состава «старой» молекулы ДНК и одна – вновь синтезированная. Такой способ удвоения был назван полуконсервативным.
|
Рис. 3. Схематичное изображение полуконсервативного механизма репликации ДНК |
Для синтеза ДНК используются не обычные нуклеотиды, а нуклеотидтрифосфаты. При образовании ковалентной связи между нуклеотидтрифосфатами два фосфата отщепляются, а третий образует связь с третьим атомом углерода рибозы. Образовавшийся пирофосфат потом расщепляется на фосфаты.
Синтез РНК или ДНК из нуклеотидов (не нуклеотидтрифосфатов!) – процесс слабо эндэргонический, он требует энергии и не может протекать самопроизвольно. Расщепление цепочки из нескольких фосфатов – процесс сильно экзэргонический, он сопровождается выделением энергии. Сильно экзэргонический процесс как бы «сопрягается» со слегка эндэргоническим, и в итоге синтез цепочки ДНК из нуклеотидтрифосфатов становится возможным. Разложение пирофосфата дополнительно сдвигает равновесие в сторону синтеза полинуклеотидной цепочки (см. урок 9).
|
Рис. 4. Удлинение цепочки ДНК в процессе синтеза |
Матричный синтез ДНК осуществляется специальными ферментами – ДНК-полимеразами. И у бактерий кишечной палочки, и у человека обнаружено несколько различных форм ДНК-полимераз. Репликация ДНК протекает с чрезвычайно высокой точностью: одна ошибка на несколько миллиардов правильно встроенных нуклеотидов.
|
Модель 2. Синтез комплементарной цепи ДНК |
При репликации ДНК у эукариот имеющиеся нуклеосомы случайным образом распределяются между двумя цепочками, а сразу же после репликации на обеих молекулах ДНК формируются новые нуклеосомы.
Удвоение ДНК в клетках многоклеточных организмов находится под строгим контролем – неконтролируемая репликация приводит к неконтролируемому делению клетки и перерождению ее из нормальной в раковую. Для удвоения нормальной животной клетки нужно соблюдение нескольких условий. Во-первых, нужно, что бы на ее рецепторы подействовали факторы роста (см. урок 6). Во-вторых, нужно, чтобы рядом было свободное место (чтобы ее не сдавливали соседи). Наконец, нужно, чтобы ее ДНК была неповрежденной. В раковых клетках из-за мутаций нарушаются все эти правила – они делятся без воздействия факторов роста извне, они расталкивают и даже разрушают соседние клетки, они удваивают поврежденную ДНК, что ведет к накоплению еще большего числа мутаций.
Итак, ДНК является носителем наследственной информации. Но возникает вопрос – что же это за информация? Еще в 30-е годы в результате исследования мутаций грибов и бактерий была сформулирована теория «один ген – один фермент». Согласно этой теории, каждый фермент, катализирующий определенную реакцию в организме, кодируется одним геном, и мутации в этом гене приводят к нарушению обмена веществ на стадии, катализируемой данным ферментом. Кроме ферментов, в организме содержатся и другие белки – структурные, транспортные, регуляторные и др. Было обнаружено, что и они кодируются определенными генами, мутации в которых вызывают наследственные болезни.
Так, у человека существует тяжелое наследственное заболевание – серповидноклеточная анемия. При этой болезни в белке β-глобине (входящем в белковую часть гемоглобина) на 6-м месте вместо глютаминовой кислоты стоит валин. В результате этот белок становится плохорастворимым, и красные кровяные тельца приобретают форму серпа. Оказалось, что причиной заболевания является мутация в гене, расположенном в 11-й хромосоме – в определенном участке этого гена аденин заменяется на тимин. Согласно современным представлениям, концепция формулируется как «один ген – одна полипептидная цепь», поскольку многие белки состоят не из одной, а из нескольких полипептидных цепочек.
Действительно, для обеспечения синтеза почти всех нужных веществ организму достаточно записи информации о его белках – ведь белки-ферменты направляют синтез и углеводов, и липидов, и других веществ. Последовательность нуклеотидов в определенном участке ДНК соответствует последовательности аминокислот в соответствующем белке. При этом три нуклеотида ДНК соответствуют одной аминокислоте белка. В самом деле, нуклеотидов всего 4, а аминокислот, из которых строится белок, – 20. Если бы один нуклеотид соответствовал одной аминокислоте, то можно было бы закодировать всего 4 разных аминокислоты, а если бы одной аминокислоте соответствовали два нуклеотида – то только 4 × 4 = 16 аминокислот. При существующем триплетном генетическом коде число возможных троек нуклеотидов составляет 4 × 4 × 4 = 64, что с избытком хватает для кодирования 20 аминокислот (на самом деле, нужно еще обозначать сигнал «стоп», чтобы закончить синтез белка в нужном месте, но 64 триплетов все равно более чем достаточно).
При считывании генетической информации сама ДНК не используется в качестве матрицы для синтеза белка – ведь любое изменение в ее нуклеотидах представляет собой мутацию, и в дальнейшем оно будет передано всем потомкам этой клетки. С матрицы ДНК снимается «рабочая копия»: если она и подпортится, то дефектными будут лишь несколько молекул белка. Использование «рабочей копии» дает еще одно важное преимущество: такие молекулы можно в большом количестве считать с одного гена, а затем с каждой «рабочей копии» считать много молекул белка, что позволяет существенно ускорить его синтез.
Функцию такой «рабочей копии» выполняет информационная РНК; ее называют также матричной РНК (мРНК). Считывание РНК с матрицы ДНК называется транскрипцией. Она осуществляется специальным ферментом – РНК-полимеразой, которая присоединяет новые нуклеотиды к 3′-концу растущей цепочки, т. е. цепочка РНК удлиняется в направлении от 5′- к 3′-концу. Для синтеза РНК используются не просто нуклеотиды, а нуклеотидтрифосфаты – подобно тому, как происходит при синтезе ДНК. Только молекулы нуклеотидтрифосфатов, используемые для синтеза ДНК, содержат дезоксирибозу, а для синтеза РНК – рибозу.
|
Рис. 5. Строение РНК |
мРНК считывается с участка ДНК, кодирующего данный белок, в соответствии с принципом комплементарности: напротив аденина ДНК становится урацил РНК, напротив гуанина – цитозин, напротив тимина – аденин и напротив цитозина – гуанин. Может возникнуть вопрос: а зачем нужно иметь два различных основания, урацил и тимин, если все равно они комплементарны аденину? Не лучше ли было ограничиться урацилом, ведь он проще по строению? Дело в том, что в водной среде азотистое основание цитозин самопроизвольно подвергается дезаминированию и превращается в урацил. В каждой клетке человека за сутки происходит около сотни таких замен. Если бы урацил был нормальным, природным компонентом ДНК, то никакие системы не могли бы отличить «законный» урацил от «незаконного». А поскольку в нормальной ДНК не должно быть урацила, то специальные ферменты узнают его и исправляют повреждение. Что же касается молекул РНК, то их повреждение не имеет столь большого значения: время их жизни ограничено, и если имеется неповрежденная матрица ДНК, то с нее вскоре считаются новые правильные РНК.
Итак, реализация генетической информации в живой клетке происходит по схеме:
ДНК → РНК → белок. Участок молекулы ДНК, в котором закодирована одна полипептидная цепь, или одна рибосомная РНК, или одна транспортная РНК (о них мы расскажем позже) – это и есть ген (правда, есть еще регуляторные участки, которые сами ничего не кодируют, но входят в состав генов). Точное число генов сейчас известно только для некоторых организмов. У вирусов имеется от нескольких штук до нескольких сотен генов, у бактерий обычное число генов составляет несколько тысяч, у дрожжей – несколько больше, у нематоды Caenorhabditis – более 10 тыс. По последним данным у человека имеется более 20 тыс. генов.
У высших эукариот (например, человека) свыше 90 % всей ДНК вообще не кодирует ни белков, ни РНК; функции этих «молчащих» участков еще плохо изучены.
Молекула РНК менее стабильна, чем ДНК – наличие гидроксильной группы при втором атоме углерода рибозы делает ее менее стойкой по сравнению с дезоксирибозой. Поэтому молекула ДНК лучше подходит на роль носителя наследственной информации.
И у прокариот, и у эукариот, как правило, на каждом отдельном участке ДНК только одна цепочка является кодирующей; на соседнем участке кодирующей может быть другая цепочка, но тоже одна. Участки кодирования в подавляющем большинстве случаев не перекрываются. Исключение составляют вирусы: у них часто на одном участке ДНК кодирующими являются обе цепочки.
|
Рис. 6. Короткий участок типичной хромосомы. Стрелки показывают направление считывания с соответствующей цепи ДНК |
Считывание гена начинается с того, что РНК-полимераза садится на промотор – особую последовательность нуклеотидов, имеющуюся в начале каждого гена. Промоторы разных генов похожи друг на друга, но все же имеют некоторые отличия. У бактерии кишечной палочки типичный промотор содержит в транскрибируемой цепи ДНК последовательность АТАТТА в 15 нуклеотидах перед точкой начала транскрипции и последовательность ААЦТГА в 35 нуклеотидах перед этой точкой. Гены с сильно отличающимися от этой последовательности промоторами обычно считываются плохо. Различия в последовательности нуклеотидов в промоторе определяют, какой ген будет считываться интенсивно, а какой слабо.
После промотора в гене идет транскрибируемая часть. Именно в ней закодирована последовательность аминокислот соответствующего белка. Транскрипцию нужно не только начать, но и закончить, поэтому в конце гена есть небольшой участок – терминатор транскрипции.
После того как РНК-полимераза садится на промотор, двойная спираль ДНК частично расплетается. Фермент начинает двигаться по матричной цепи; за ним тянется цепочка синтезируемой РНК. По мере продвижения РНК-полимеразы эта РНК освобождается из комплекса с ДНК, и двойная спираль ДНК восстанавливается.
Дойдя до терминатора, РНК-полимераза отваливается от ДНК, и образовавшаяся РНК тоже освобождается. Скорость транскрипции составляет около 3000 нуклеотидов в минуту. Частота ошибок транскрипции – примерно один неправильно встроенный нуклеотид на десятки тысяч «правильных» нуклеотидов. Точность этого процесса значительно ниже, чем в случае репликации, но поскольку с одного гена считывается много РНК, да и живут они ограниченное время, эти ошибки не наносят клетке большого вреда.
|
Модель 3. Транскрипция РНК |
У прокариот имеется одна форма РНК-полимеразы, у эукариот – 3. Для синтеза мРНК (т. е. для считывания генов, кодирующих белки) используется РНК-полимераза II. Синтез мРНК играет важнейшую роль в жизнедеятельности клетки. Что будет с организмом, если его заблокировать, видно из следующего примера. Самый опасный ядовитый гриб, растущий в средней полосе, – бледная поганка (для смертельного отравления достаточно съесть 1/3 гриба). Она содержит несколько различных токсинов, самый сильный из них – α-аманитин. Этот яд ингибирует РНК-полимеразу II и препятствует синтезу мРНК. Впрочем, ингибиторы синтеза РНК используются и в лечебных целях. Так, антибиотик актиномицин D, подавляющий транскрипцию, используется для лечения некоторых видов раковых опухолей.
Молекулы РНК расщепляются специальными ферментами РНКазами. В клетках прокариот среднее время жизни мРНК составляет всего около 3 мин. Такой быстрый оборот позволяет им быстро изменять содержание этих информационных молекул в ответ на изменение условий среды.
В эукариотических клетках мРНК более стабильны. Так, молекула, кодирующая β-цепь глобина, существует в среднем около 10 ч. Однако мРНК некоторых белков – регуляторов клеточного цикла живут меньше 10 мин. Время жизни гистоновых мРНК регулируется: если в клетке происходит синтез ДНК, их период полураспада составляет около часа, а если синтеза нет, – около 10 мин.
Помимо информационной РНК в клетке имеются рибосомальные и транспортные РНК, они не кодируют белков. Рибосомальная РНК (рРНК) входит в состав рибосом (вместе с определенными белками). В клетках эукариот рРНК синтезируется в специальном участке ядра – ядрышке. Там же она связывается с рибосомальными белками и образует субъединицы рибосом, которые потом выходят в цитоплазму и приступают к синтезу белка.
Недавно были проделаны эксперименты, доказавшие что именно рРНК, а не рибосомальные белки, как считалось раньше (см. урок 4), катализируют образование новых пептидных связей при синтезе полипептидных цепочек. Таким образом, не только молекулы белков, но и молекулы РНК могут служить биологическими катализаторами. Каталитическая активность молекул РНК была открыта еще в начале 80-х годов ХХ-го века, однако известные до последнего времени немногочисленные реакции, катализировавшиеся РНК, не имели столь уж важного общебиологического значения. Теперь ситуация изменилась, доказано, что РНК необходима для синтеза белка, который является важнейшим биохимическим процессом, необходимым для жизнедеятельности абсолютно всех клеток.
Открытие каталитической активности РНК позволило разрешить великую загадку эволюционной биологии: что появилось первым в процессе самозарождения жизни на Земле – белки или нуклеиновые кислоты? С одной стороны, все генетические функции в клетке выполняют нуклеиновые кислоты, но с другой стороны, все катализаторы (как считалось еще совсем недавно) являются белками. В настоящее время полагают, что на заре эволюции, в эпоху возникновения первых живых систем, существовал особый «РНКовый мир». Тогда молекулы РНК выполняли обе перечисленные выше важнейшие функции. Постепенно белки взяли на себя каталитическую функцию, почти полностью вытеснив РНК (известно не более десятка реакций живой клетки, которые катализируется молекулами РНК, тогда как белки ускоряют многие тысячи реакций). Функцию же носителя генетической информации взяла на себя ДНК.
Транспортная РНК (тРНК) играет важную роль в процессе синтеза белка, доставляя нужные аминокислоты в рибосому.
Около 80 % и более от общего количества РНК в клетке (по массе) составляет рРНК, около 15 % – тРНК, и лишь очень небольшую часть – обычно менее 5 % – составляет мРНК. Синтез рРНК и тРНК происходит подобно синтезу мРНК, но рРНК и тРНК не несут информации о строении белка, в отличие от мРНК.
Краткое содержание урока
|
ISTOCK, BEHOLDINGEYEБазовое редактирование — это относительно недавняя методика редактирования генома, которая меняет одну пару оснований ДНК на другую, предлагая исследователям потенциал для исправления вредных однонуклеотидных мутаций в геноме человека. Но современные классы базовых редакторов допускают только преобразование пар оснований цитозин-гуанин (C-G) в пары оснований тимин-аденин (T-A), что делает значительную долю таких мутаций нецелевой с этим подходом.
Теперь исследователи из Гарвардского университета разработали новый класс редакторов адениновой основы (ABE), которые могут эффективно превращать A-T в G-C, открывая большинство патогенных точечных мутаций для редактирования. Команда сообщила о своих результатах сегодня (25 октября) в Nature .
«Это очень элегантное исследование», — говорит Эндрю Бассетт, руководитель отдела исследований в области сотовой связи в институте Wellcome Trust Sanger, который не принимал участия в работе. «Возможность расширить [базовое редактирование] на другие типы действительно очень важна.”
Традиционное редактирование генома CRISPR-Cas9 делает двухцепочечный разрыв в ДНК с целью введения вставок или делеций в целевые сайты. Но создание этого разрыва может привести к значительному количеству ошибок в сайте-мишени, таких как случайная вставка или делеция нуклеотидов (инделя).
Не существует известного фермента, нацеленного на ДНК, который бы переключился с аденина на гуанин.
Напротив, редактирование оснований непосредственно преобразует одну пару оснований ДНК в другую, не разрезая нуклеиновую кислоту.Техника, впервые разработанная химическим биологом Дэвидом Лиу и его коллегами из Гарварда, связана с меньшим количеством идентификаторов и более высокой эффективностью редактирования.
См. «Достижения в редактировании генома»
Современные редакторы состоят из конвертирующего основание фермента, привязанного к специально разработанной, не режущей версии эндонуклеазы Cas9. После конверсии оснований на одной цепи ДНК этот белок Cas9 делает метку, называемую «ник» на противоположной, неотредактированной цепи, побуждая механизмы репарации ДНК клетки заменять оригинальный, теперь несоответствующий нуклеотид и, таким образом, завершая замену пары оснований.
Но конвертирующий основание фермент, цитидин-деаминаза, может превращать только цитозин ДНК в урацил (U) — функциональный эквивалент основания тимина. Не существует известного белка, нацеленного на ДНК, который может перейти с аденина на гуанин.
«Природа удобно снабжает нас ферментами цитозин-деаминазы, которые действуют на ДНК», — говорит Лю The Scientist . «Самая большая проблема с разработкой АБЕ не заключалась в определении потребности — это ясно, когда вы смотрите на диспропорциональное изобилие патогенных человеческих мутаций, которые могут быть исправлены АБЕ — это тот факт, что природа не обеспечивает фермент, который дезаминирует аденин в ДНК ,”
См. «CRISPR: резка не требуется»
Как построить аденин-редактор
Однако существует несколько известных ферментов деаминазы, которые действуют на РНК. Одним из них является фермент адениндеаминаза в Escherichia coli , называемый TadA, который превращает аденин в инозин (I) — основание, функционально эквивалентное G — в транспортной РНК (тРНК). По словам Лю, предыдущие попытки лаборатории Лю и других связать этот фермент с Cas9 и добиться конверсии основания в ДНК оказались безуспешными.
Так, для своего последнего проекта исследователь Лю и Гарвард Николь Гауделли решила создать свою собственную версию TadA для редактирования ДНК путем сочетания направленной эволюции и белковой инженерии в E. coli .
Им удалось получить что-то, что на самом деле работает невероятно хорошо.
Эндрю Бассетт,
Wellcome Trust Sanger Institute
Во-первых, исследователи предоставили геномам бактерий изготовленную на заказ дефектную версию гена устойчивости к антибиотикам.«Мы установили мутации, которые требовали дезаминирования аденина в ДНК, чтобы восстановить активность устойчивости к антибиотикам», — объясняет Лю. Эти бактерии погибнут при лечении антибиотиком хлорамфениколом — если только они не смогут каким-то образом преобразовать А в I.
Чтобы спроектировать эту способность к преобразованию А-Я, команда создала библиотеку слияний TadA-Cas9, содержащую различные мутации в части аденин-деаминазы механизма. «Мы снабдили каждую из этих клеток E.coli своей мутантной формой [TadA]», — говорит Лю.«Николь сделала миллионы вариантов в каждом поколении этих мутантов TadA, а затем потребовала, чтобы бактерии выжили в присутствии растущих концентраций антибиотика».
Бактерии, которые прошли через это испытание, то есть те, которые исправили свои дефектные гены устойчивости к антибиотикам, должны были содержать мутантов TadA, которые могли действовать на ДНК, рассуждал Лю и его коллеги. Конечно, когда исследователи высказали версии этих конкретных мутантов TadA-Cas9 из выживших бактерий в клетках человека, они смогли заменить AT на GC в ДНК, хотя и с очень низкой эффективностью (только около 3 процентов секвенированных клеток показали изменение ).
В последующих раундах исследований исследователи использовали свою бактериальную систему для дальнейшего повышения эффективности АБЕ и обеспечения возможности редактирования баз в различных контекстах, например, перед предшествующими A, T, C или G. Раундом В-седьмых, у Лю и его коллег осталась АБЕ, которая могла бы преобразовать А в I с минимальными ошибками с эффективностью более 50% — показатель, типичный для современных редакторов CG и TA.
«Это огромные усилия, чтобы сделать это», — говорит Бассетт, добавляя, что само исследование является впечатляющим примером того, как белки могут быть изменены путем направленной эволюции.«Им удалось получить что-то, что действительно работает невероятно хорошо. Это довольно большое дело с точки зрения расширения всей области редактирования базы ».
Терапевтические возможности базового редактирования
Чтобы продемонстрировать терапевтический потенциал редактора, команда исправила точечную мутацию, связанную с потенциально опасным для жизни нарушением всасывания железа, называемым наследственным гемохроматозом. Исследователи обнаружили, что нарушающий А нуклеотид был заменен с эффективностью почти 30 процентов в клеточной линии человека без каких-либо обнаруживаемых признаков.
Биоинженер Фенг Чжан из Института широкого профиля MIT отмечает в электронном письме, что команда использовала «комплексный и творческий подход» для достижения такой точности. «Как область, мы искали способы точного переписывания частей генетического кода», — пишет Фэн, чей собственный метод на основе CRISPR для редактирования отдельных баз в РНК был опубликован сегодня в Science . «Базовые редакторы приближают нас к этой цели».
См. «Редактирование РНК с помощью CRISPR-Cas13»
Конечно, новый редактор не завершает набор — все еще невозможно преобразовать, например, G в C или A в T.Группа Лю работает над созданием новых классов базовых редакторов для внесения этих изменений.
Существуют возможности для улучшения и с нынешним ABE. Бассетт отмечает, что окно редактирования системы — шириной около шести нуклеотидов — достаточно велико, чтобы основания А рядом с мишенью А могли быть отредактированы непреднамеренно. Кроме того, в настоящее время на некоторые последовательности ДНК легче нацеливаться, чем на другие.
«Мы работаем над созданием новых форм ABE, которые максимизируют их охват таргетинга и их полезность», — говорит Лю.«И мы, конечно, используем наших редакторов в сотрудничестве с экспертами по биологии болезней, чтобы попытаться скорректировать модели генетических заболеваний человека на животных».
Н. Гауделли и др., «Программируемое базовое редактирование A • T — G • C в геномной ДНК без расщепления ДНК», Nature , doi: 10.1038 / nature24644 , 2017.
, DNA Base Pair Molecule Jewellery
Этот список предназначен для ОДНОЙ ДНК-подвески (купите две, если хотите подходящую пару)
Молекулярная структура оснований ДНК:
Тимин
Аденин
Гуанин
Цитозин
Соответствующие пары:
Тимин & Adenine
Гуанин и цитозин
Их можно превратить в соответствующие наборы серег, брелков, браслетов и брошей / значков.
Подвеска имеет длину около 1 и 1,5 дюйма.
Это прекрасно детализированный и прекрасный подарок для всех любителей науки!
В НАЛИЧИИ:
ОЖЕРЕЛЬЕ 45см (цепочка тонов серебро / золото / розовое золото ИЛИ черный / коричневый / белый шнур — ПОЖАЛУЙСТА, ВЫБЕРИТЕ СООБЩЕНИЕ)
ОЖЕРЕЛЬЕ 75см
ЧОКЕР 1 (эластичное колье с татуировкой)
ЧОКЕР 2 (простая черная полоса Ширина 5 мм)
CHOKER 3 (черный шнур тонкий)
Брелок
СЕРЬГИ (крючки)
СЕРЬГИ (шпильки) — подвески с подвесками из круглого стержня — вы можете выбрать один из различных цветов)
БРАСЛЕТ 1 (цепочка тонов серебро / золото / розовое золото) )
БРАСЛЕТ 2 (Слип-узел черный / коричневый шнур — ПОЖАЛУЙСТА, ВЫБЕРИТЕ СООБЩЕНИЕ)
(может иметь более одного очка, пожалуйста, сообщение)
Браслет (серебряный / золотой браслет с брелоком)
ANKLET 1 (серебро / золото / розовое золото ИЛИ черная / коричневая цепочка — ПОЖАЛУЙСТА, ВЫБОР)
ANKLET 2 (Slip Knot черная / коричневая цепочка — ПОЖАЛУЙСТА, ВЫБОР)
ЗНАЧОК (шарм болтается на круговом значке — вы можете выбрать один из нескольких цветов)
КОЛЛАРНЫЕ ПИНС-цепочка (один с каждой стороны с цепочкой) шарм болтается на круговой булавке — можно oose из разных цветов)
КОЛЛЕКЦИОННЫЕ ШПИЛЬКИ — без цепочки
CHARM (можно использовать для подвешивания сумки, одежды и молний!)
PLANNER CHARM (Выбор подвесок и цветных бусин, висящих на цепочке для корешка книги)
BOOKMARK METAL (Dangling брелок с металлической закладкой)
BOOKMARK RIBBON (закладка с оборванной подвеской на ленте — сообщение для выбора цвета)
ORNAMENT (чтобы повесить на елку и т. д. — пожалуйста, укажите выбор цвета ленты — или отправьте случайно)
MAGNET
HAT PIN-код (длинная иголка) с висящим оберегом, используйте, чтобы прикрепить шляпы к голове) PIN-код ВОЛОСЫ ДЛЯ ВОЛОС (очарование спирали закручивает в ваши волосы, так что только очарование видимо)
ПОТРЯСКА С шарфом (Подвеска с подвесками, которая используется для прорезания шарфа)
ОБУВЬ ОБУВИ (Очарование врезается в шнурок для обуви)
ТЕЛЕФОН ЧАРМ (болтается с телефона)
АКСЕССУАР ДЛЯ АВТОМОБИЛЯ (Повесьте на зеркало заднего вида)
БРАСЛЕТ ДЛЯ ОБУВИ (больше, чем ножной браслет, который сидит вокруг ботинка)
ПЫЛЬНИК (сидит в телефоне с шармом болтался)
K ILT PIN (3-5 подвесок могут висеть на килте)
ЗАРЯДКА ПЛАНЕРА (может иметь несколько подвесок на цепочке, которые могут защелкнуться на планировщике или вяжущем)
ЗАДВИЖКА ДЛЯ ВОЛОС
GARLAND (нанизывается для подвешивания в комнате)
ЗАЖИГАТЕЛЬ ДЛЯ ЧАСОВ — (Брелок прикреплен к застежке, чтобы добавить к воротнику)
ДОБАВЛЕННЫЕ ШАРМЫ:
Чтобы добавить письмо или бусину, перейдите сюда и добавьте в корзину:
https: // www.etsy.com/au/listing/672538195
Можно добавить бусину с цветными бусинами к любому ювелирному изделию (выбор: прозрачный, черный, светло-красный, темно-красный, светло-желтый, темно-желтый, светло-синий, темно-синий, светло-зеленый, темный зеленый).
Можно добавить букву алфавита (AZ)
Пожалуйста, СООБЩЕНИЕ, если вы хотите какие-либо индивидуальные настройки или комбинацию подвесок для одного ювелирного изделия
_________________________________________________________________
**** БЕСПЛАТНАЯ ДОСТАВКА на любой дополнительный приобретенный предмет ****
Все украшения поставляются в подарочных пакетах или конвертах.
_________________________________________________________________
Доставка по международным заказам занимает в среднем от 10 дней до 3 недель.
Доставка в Австралию зависит от штата 2 дня — 10 дней
_________________________________________________________________
Следите за новостями и конкурсами в Instagram @curioscollection!
_________________________________________________________________
,
Leave A Comment