
Азотистые основания нуклеотидов ДНК — аденин, гуанин, тимин и цитозин
К азотистым основаниям относят аденин (A), гуанин (G), цитозин (C) и тимин (T), который входит в состав только ДНК, а урацил (U) заменяет его в РНК. Они обладают схожими структурами и химическими свойствами. Это гетероциклические органические соединения, производные пиримидина и пурина, входящие в состав нуклеотидов. Аденин и гуанин — производные пурина, а цитозин, урацил и тимин — производные пиримидина.
Когда азотистые основания присоединяются ковалентной связью к 1′ атомам рибозы (в РНК) или дезоксирибозы (в ДНК), а к 5′-гидроксильной группе сахара присоединяется одна или несколько фосфатных групп, образуются нуклеотиды.
стандартные нуклеотиды ДНК составляют триплеты — участки ДНК, кодирующие одну аминокислоту. Например, с триплета АУГ (ему соответствует аминокислота метионин) обычно начинается синтез белка на рибосомах. Нуклеозиды, приведённые в таблице, входят в состав моно-, ди- и трифосфатов. Например, аденозин входит в состав АТФ — важнейшего энергетического ресурса организма.
Пурин C5N4h5 — гетероциклические соединения, имидазольные производные пиримидинов. Производные пурина играют важную роль в химии природных соединений (пуриновые основания ДНК и РНК; кофермент NAD; алкалоиды, кофеин и т. д.) и, благодаря этому, в фармацевтике — ядро пурина входит в состав некоторых антибиотиков. Пурин и ряд его производных обладают противоопухолевой, противовирусной и противоаллергической активностью. Аденин — азотистое основание, аминопроизводное пурина. Образует две водородных связи с урацилом (в РНК) и тимином (в ДНК) по правилу комплементарности. Представляет собой бесцветные кристаллы. Химическая формула С5H5N5. Аденин проявляет основные свойства. Аденин входит в состав многих жизненно важных для живых организмов соединений, таких как аденозин, аденозинфосфорные кислоты, нуклеиновые кислоты, адениновые нуклеотиды и др. В виде этих соединений аденин широко распространен в живой природе. Гуанин — азотистое основание, аминопроизводное пурина, является составной частью нуклеиновых кислот. Химическая формула — C5H5N5O. В ДНК и РНК образует три водородные связи с цитозином по правилу комплементарности. Производные гуанилового нуклеотида — ГДФ, ГТФ и цАМФ — участвуют во многих сигнальных путях клетки. Для некоторых процессов, происходящих в клетке — например, для сборки микротрубочек — ГТФ используется как источник энергии. Пиримидин C4N2h5 — гетероциклическое соединение, имеющее плоскую молекулу, простейший представитель диазинов. Пиримидин представляет собой бесцветные кристаллы с характерным запахом. Пиримидин проявляет свойства слабого двукислотного основания, так как атомы азота могут присоединять протоны. Производные пиримидина широко распространены в живой природе, где участвуют во многих важных биологических процессах. Его производные цитозин, тимин, урацил входят в состав нуклеотидов. Биологическая роль пиримидинов не ограничена нуклеиновыми кислотами. Некоторые пиримидиновые нуклеотиды играют важную роль в процессах обмена углеводов и липидов. Витамин В1 (тиамин) — пиримидиновое производное. Пиримидиновое ядро входит в состав некоторых коферментов и антибиотиков. Тимин — производное пиримидина. Формула C5H6N2O2. Присутствует во всех живых организмах, где вместе с дезоксирибозой входит в состав нуклеозида тимидина, который может фосфорилироваться 1-3 остатками фосфорной кислоты с образованием нуклеотидов тимидин моно-, ди- или трифосфорной кислоты (ТМФ, ТДФ и ТТФ). Дезоксирибонуклеотиды тимина входят в состав ДНК, в РНК на его месте располагается рибонуклеотид урацила. Тимин комплементарен аденину, образует с ним 2 водородные связи. Цитозин — азотистое основание, производное пиримидина. С рибозой образует нуклеозид цитидин, входит в состав нуклеотидов ДНК и РНК. Во время репликации и транскрипции по правилу комплементарности образует три водородных связи с гуанином. Представляет собой бесцветные кристаллы. Химическая формула C4H5N3O. Его производные цитозин, тимин, урацил входят в состав нуклеотидов, проявляет основные свойства. Урацил — пиримидиновое основание, которое является компонентом рибонуклеиновых кислот (РНК) и, как правило, отсутствует в дезоксирибонуклеиновых кислотах (ДНК). В составе РНК может комплементарно связываться с аденином, образуя две водородные связи. Эрвин Чаргафф открыл правила, описывающие количественные соотношения нуклеотидов. Правило Чаргаффа — биологический закон, в соответствии с которым установлены количественные соотношения между азотистыми основаниями разных типов. Для того, чтобы определить точные количественные соотношения нуклеотидов, Чаргафф разделил нуклеотиды ДНК методом бумажной хроматографии. Ему удалось выявить три закономерности:
Число аденинов равно числу тиминов, а число гуанинов — числу цитозинов: А=Т, Г=Ц
Число пуринов равно числу пиримидинов: А+Г=Т+Ц
Число аденина и цитозина равно числу гуанина и тимина: А+Ц=Г+Т
Состав ДНК разных организмов различается суммарным числом комплементарных оснований. Соотношение комплементарных нуклеотидов тоже может быть различным в разных молекулах ДНК. У одних оорганизмов в ДНК преобладают пары аденин-тимин, а у других — гуанин-цитозин. При этом правила Чаргаффа будут выполняться в любом случае.
Вопрос
Если в ДНК содержится 21 % аденина, то какова долю (%) остальных нуклеотидов, содержащихся в ДНК?
Данные рентгеноструктурного анализа показали, что молекулы ДНК имеют спиральную структуру. В 50-х годах 20 века многие химики и биологи пытались исследовать структуру ДНК. В Королевском колледже в Лондоне Морис Уилкинс и Розалинда Франклин пытались решить эту проблему методом рентгеноструктурного анализа солей ДНК. Но такой способ позволял выявить только общую структуру молекулы. Тем временем Джеймс Уотсон и Френсис Крик в Кавендишской лаборатории Кембриджского университета, используя данные, полученные М. Уилкинсом, стали строить пространственные 3-D модели ДНК. Они пытались создать структуру, которая согласовывалась бы со всеми данными рентгеноструктурного анализа. В итоге Уотсон и Крик пришли к выводу, что ДНК имеет спиральную структуру с периодичностью 0,34 нм вдоль оси. В 1953 году Дж. Уотсон и Ф. Крик открыли строение ДНК, предложив модель двойной спирали. Нуклеиновые кислоты, подобно белкам, обладают первичной структурой (под которой подразумевается их нуклеотидная последовательность) и трехмерной структурой. Интерес к структуре ДНК усилился, когда в начале XX века возникло предположение, что ДНК, возможно, представляет собой генетический материал. В начале 50-х годов американский химик, лауреат Нобелевской премии Лайнус Полинг, уже изучивший к тому времени α-спиральную структуру, характерную для многих фибриллярных белков, обратился к исследованию структуры ДНК, которая по имеющимся в то время сведениям также представлялась фибриллярной молекулой. Одновременно в Королевском колледже Морис Уилкинс и Розалинда Франклин пытались решить ту же проблему методом рентгеноструктурного анализа. Их исследования требовали долгой и трудоёмкой работы по приготовлению чистых препаратов солей ДНК, для которых удавалось получать сложные дифракционные картины. С помощью этих картин можно было, однако, выявить лишь общую структуру молекулы ДНК, не столь детализированную, как та, которую позволяли получить чистые кристаллы белка. Тем временем Фрэнсис Крик и Джеймс Уотсон в Кавендишской лаборатории Кембриджского университета избрали иной подход, который в конечном счёте и обеспечил успешное решение проблемы. Используя все физические и химические данные, какие оказались в их распоряжении, Уотсон и Крик стали строить пространственные модели ДНК в надежде на то, что рано или поздно им удастся получить достаточно убедительную структуру, согласующуюся со всеми этими данными. Истории их поисков увлекательно описаны Уотсоном в его книге «Двойная спираль». Два обстоятельства оказались для Уотсона и Крика решающими. Во-первых, они имели возможность регулярно знакомиться с результатами работ Уилкинса и, сопоставляя с его рентгенограммами свои модели, могли таким образом проверять эти модели. А рентгенограммы Уилкинса свидетельствовали в пользу спиральной структуры с периодичностью 0,34 нм вдоль оси. Во-вторых, Уотсон и Крик отдавали себе отчёт в важном значении закономерностей, касающихся соотношения различных оснований в ДНК. Обнаружил эти закономерности и сообщил о них в 1951 году Эрвин Чаргафф. Это открытие, однако, при всей своей важности не привлекло к себе должного внимания. Уотсон и Крик задались целью проверить предположение, что молекула ДНК состоит из двух спиральных полинуклеотидных цепей, удерживаемых вместе благодаря спариванию оснований, принадлежащих соседним цепям. Основания удерживаются вместе водородными связями. Аденин спаривается с тимином, гуанин — с цитозином; АТ-пара соединяется двумя водородными связями, а ГЦ-пара — тремя. Уотсон попытался представить себе такой порядок спаривания оснований и позже вспоминал об этом так: «От радости я почувствовал себя на седьмом небе, ибо тут я уловил возможный ответ на мучившую нас загадку: почему число остатков пуринов в точности равно числу остатков пиримидинов?» Уотсон увидел, что при таком сочетании основания оказываются очень точно подогнанными друг к другу, а общий размер и форма этих двух пар оснований одинаковы, так как обе пары содержат по три кольца. Водородные связи при других сочетаниях оснований в принципе возможны, но они гораздо слабее. После того как все эти обстоятельства выяснились, можно было наконец приступить к созданию достоверной модели ДНК. Уотсон и Крик показали, что ДНК состоит из двух антипараллельных (направленных в противоположные стороны) полинуклеотидных цепей. Каждая цепь закручена в спираль вправо, и обе они свиты вместе, то есть закручены вправо вокруг одной и той же оси, образуя двойную спираль. Каждая цепь состоит из сахарофосфатного остова, вдоль которого перпендикулярно длинной оси спирали располагаются азотистые основания. Находящиеся друг против друга основания двух противоположных цепей двойной спирали связаны между собой водородными связями. Расстояние между сахарофосфатными остовами двух цепей постоянно и равно расстоянию, занимаемому парой оснований, то есть одним пурином и одним пиримидином. Два пурина занимали бы слишком много места, а два пиримидина — слишком мало для того, чтобы заполнить промежутки между двумя цепями. Вдоль оси молекулы соседние пары оснований располагаются на расстоянии 0,34 нм одна от другой, чем и объясняется обнаруженная на рентгенограммах периодичность. Полный оборот спирали приходится на 3,4 нм, то есть на 10 пар оснований. Никаких ограничений относительно последовательности нуклеотидов в одной цепи не существует, но в силу правила спариваний оснований эта последовательность в одной цепи определяет собой последовательность нуклеотидов в другой цепи. Поэтому мы говорим, что две цепи двойной спирали комплементарны друг другу. Уотсон и Крик опубликовали сообщение о своей модели ДНК в журнале «Nature» в 1953 г., а в 1962 г. они вместе с Морисом Уилкинсом были удостоены за эту работу Нобелевской премии. Розалинду Франклин, умершую от рака ранее присуждения этой премии, не включили в число лауреатов, поскольку Нобелевская премия посмертно не присуждается. Для того чтобы признать, что имеющая предложенную структуру молекула может выполнять роль генетического материала, требовалось показать, что она способна: 1) нести в себе закодированную информацию и
2) точно воспроизводиться (реплицироваться). Уотсон и Крик отдавали себе отчет в том, что их модель удовлетворяет этим требованиям. В конце своей первой статьи они отметили: «От нашего внимания не ускользнуло, что постулированное нами специфическое спаривание оснований сразу же позволяет постулировать и возможный механизм копирования для генетического материала». Во второй статье, опубликованной в том же 1953 году, они обсудили выводы, которые следовали из их модели для возможного механизма передачи наследственной информации. Это открытие показало, сколь явно структура может быть связана с функцией уже на молекулярном уровне. Оно дало начало стремительному развитию молекулярной биологии.
cyberpedia.su
Известно четыре нуклеотида, которые называются аденин, гуанин, тимин, цитозин и урацил, они являются азотистыми основаниями.
Нуклеотиды – это мономеры нуклеиновых кислот. Нуклеиновые кислоты в эукариотических клетках находятся в ядре. Они есть у всех живых организмов. Мономеры, из которых потом строятся нуклеиновые кислоты, состоят из азотистого основания, остатка сахара (дезоксирибоза или рибоза) и фосфата. Сахара вместе с азотистым основанием называются нуклеозидами (аденозин, гуанозин, тимидин, цитидин). Если к ним присоединены 1-, 2-, или 3-фосфорных остатка, то вся эта структура называется соответственно, нуклеотизид монофосфатом, дифосфатом или трифосфатом или нуклеотидом (аденин, гуанин, тимин, цитозин).
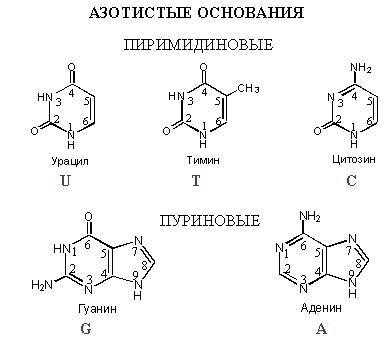
Рис. 20. Модель АТФ в пространстве
Азотистое основание, входящее в состав ДНК делится на две группы – пиримидиновую и пуриновую. В состав ДНК входит аденин, тимин, цитозин и гуанин, в РНК вместо тимина урацил. Как известно, ДНК – это большой архив, в котором хранится информация, а РНК – это молекула, которая переносит информацию из ядра в цитоплазму для синтеза белков. С различием в функциях связаны различия в строении. РНК более химически активно из-за того, что ее сахар — рибоза – имеет в своем составе гидроксильную группу, а в дезоксирибозе кислорода нет. Из-за отсутствия кислорода ДНК более инертно, что важно для ее функции хранения информации, чтобы она не вступала ни в какие реакции.
Нуклеотиды способны взаимодействовать друг с другом, при этом «выбрасывается» два фосфора, и между соседними нуклеотидами образуется связь. В молекуле фуранозы молекулы углерода пронумерованы. С первым связано азотистое основание. Когда образуется цепочка нуклеотидов, связь осуществляется между пятым углеродом одной и третьим углеродом другой фосфорной кислоты. Поэтому в цепочке нуклеиновых кислот выделяют разные неравнозначные концы, относительно которых молекула не симметрична.
Комплементарные друг другу одноцепочечные молекулы нуклеиновой кислоты способны образовывать двухцепочечную структуру. Внутри этой спирали аденин образует пару с тимином, а гуанин — с цитозином. Встречается утверждение, что нуклеотиды подходят друг другу, как осколки разбитого стекла, поэтому они и образуют пары. Но это утверждение неверно. Нуклеотиды способны образовывать пары как угодно. Единственная причина, по которой они соединяются так, и никак иначе, заключается в том, что угол между «хвостиками», которые идут к сахарам, совпадает только в этих парах, и, кроме того, совпадают их размеры. Никакая другая пара не образует такой конфигурации. А поскольку они совпадают, то их через сахаро-фосфатный остов можно связать друг с другом. Структуру двойной спирали открыли в 1953 году Джеймс Уотсон и Фрэнсис Крик.
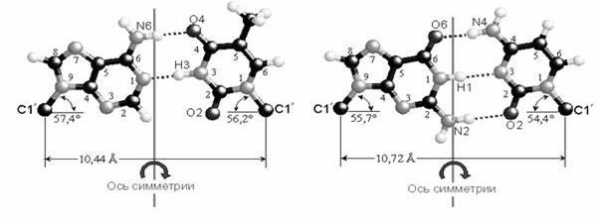
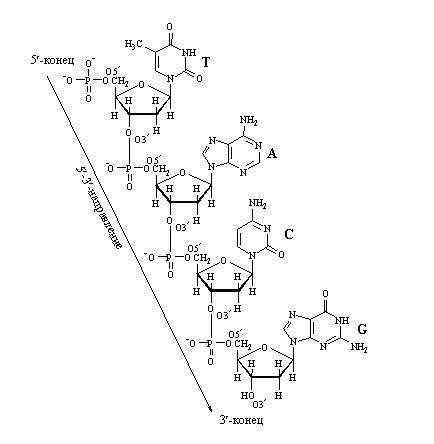
Рис. 21. Модель ДНК
При соединении друг с другом против 5-конца одной нити находится 3-конец другой нити. То есть нити идут в противоположных направлениях – говорят, что нити в ДНК антипараллельны. На рисунке 21 видно, что аденин соединяется с тимином двумя водородными связями, а гуанин соединяется с цитозином тройной водородной связью. Если молекулу ДНК подогревать, то ясно, что две связи легче разорвать, чем три, это существенно для свойств ДНК. В силу пространственного расположения сахаро-фосфатного остова и нуклеотидов, когда нуклеотиды накладывают один на другой и «сшивают» через сахаро-фосфатный остов, цепочка начинает заворачиваться, тем самым, образуя знаменитую двойную спираль.
При нагревании ДНК водородные связи разрываются, и нити в двойной спирали расплетаются. Процесс нагревания называется плавлением ДНК, при этом разрушаются связи между парами А-Т и Г-Ц .Чем больше в ДНК пар А-Т, тем менее прочно нити друг с другом связаны, тем легче ДНК расплавить. Переход из двухспиральной ДНК в односпиральную измеряется на спектрофотометрах по поглощению света при 260 нм. Температура плавления ДНК зависит от А-Т/Г-Ц состава и размера фрагмента молекулы. Ясно, что если фрагмент состоит из нескольких десятков нуклеотидов, то его гораздо легче расплавить, чем более длинные фрагменты.
У человека в гаплоидном геноме, то есть единичном наборе хромосом, 3 млрд. пар нуклеотидов, и их длина составляет 1,7 м, а клетка гораздо меньше, как вы догадываетесь. Для того чтобы ДНК смогла в ней поместиться, она достаточно плотно свернута, и в эукариотической клетке свернуться ей помогают белки – гистоны. Гистоны имеют положительный заряд, а так как ДНК заряжена отрицательно, то гистоны обладают сродством к ДНК. Упакованная при помощи гистонов ДНК имеет вид бусин, называемых нуклеосомами. 200 пар нуклеотидов идет на одну нуклеосому, 146 пар накручиваются на гистоны, а остальные 54 висят в виде линкерных (связывающих нуклеосомы) ДНК. Это первый уровень компактизации ДНК. В хромосомах ДНК свернута еще несколько раз для того, чтобы образовались компактные структуры.
К нуклеиновым кислотам кроме ДНК относится также РНК. В клетке присутствуют разные типы РНК: рибосомные, матричные, транспортные. РНК синтезируется в виде одноцепочечной молекулы, но отдельные ее участки входят в состав двухцепочечных спиралей. Для РНК также говорят о первичной структуре (последовательности нуклеотидов) и вторичной структуре (образование двуспиральных участков).
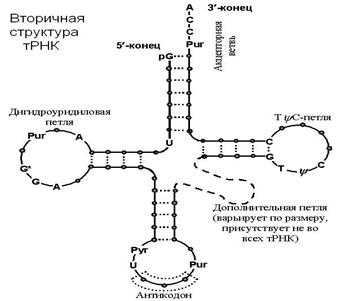
Рис. 23. Вторичная структура тРНК
Липиды
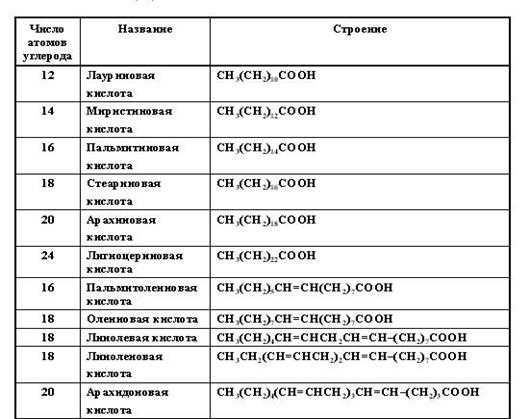
В состав липидов входят жирные кислоты, имеющие длинные углевододородные цепи. Жирные кислоты гидрофобны, то есть, не растворимы в воде.
Таблица 8
Природные жирные кислоты
В клетке важную роль играют липиды, в которых к глицерину присоединен остаток фосфорной кислоты и 2 жирных кислоты. Они называются фосфолипидами. Молекулы фосфолипидов имеют полярную (то есть гидрофильную, хорошо растворимую) группу на одном конце молекулы и длинный гидрофобный хвост. К фосфолипидам относится фосфатидилхолин. В водном растворе фосфолипиды образуют мицеллы, в которых молекулы обращены полярными «головами» наружу, в сторону воды, а гидрофобные «хвосты» оказываются внутри мицеллы, спрятанными от воды. Клеточную мембрану образуют также липиды с полярными «головами», которые обращены наружу по обе стороны мембраны, а гидрофобные «хвосты» находятся внутри липидного бислоя.
Библиографический список
1. Ахметов, Н. С. Общая и неорганическая химия [Текст]: учеб. для химико-технол. специальностей вузов — М.: Высш. шк., 1998. — 743 с.
2. Гельфман, М.И. Химия [Текст] / М.И. Гельфман, В.П. Юстратов – СПб.: Лань, 2001. – 480 с.
3. Глинка, Н.Л. Общая химия [Текст]: учеб. пособие для вузов. – Л.: Химия, 1983. – 704 с.
4. Карапетьянц, М.Х. Общая и неорганическая химия [Текст]: учебник для вузов / М.Х. Карапетьянц, С.И. Дракин – М.: Химия, 1994. – 592 с.
5. Коровин, Н.В. Общая химия [Текст]: учебник для технических направ. и спец. вузов. – М.: Высш.школа, 2000. – 558 с.
6. Кудряшов И.В., Сборник примеров и задач по физической химии [Текст]: учеб. пособие для хим.-технол. спец. вузов. – М.: Высш. школа, 1991 – 527 с.
7. Общая химия. Биофизическая химия. Химия биогенных элементов [Текст]: учеб. для вузов по мед., биол., агрон., ветеринар., экол. специальностям/ Ю. А. Ершов, В. А. Попков, А. С. Берлянд, А. З. Книжник — М.: Высш. шк., 2000. — 560 с.
8. Пузаков, С.А. Сборник задач и упражнений по общей химии [Текст]: учеб. пособие / С.А. Пузаков, В.А. Попков, А.А. Филиппова. – М.: Высш. школа, 2004. – 255 с.
9. Суворов, А.В. Общая химия [Текст]: учеб. пособие для вузов / А.В. Суворов, А.Б. Никольский – СПб: Химия, 1995 – 624 с
10. Угай, Я. А. Общая и неорганическая химия [Текст]: учеб. для вузов по направлению и специальности «Химия» — М.: Высш. школа, 1997. — 527 с.
11. Харин, А.Н. Курс химии [Текст]: учебник для приборостроит. спец. вузов / А.Н. Харин, Н.А. Катаева, Л.Т. Харина. – М.: Высш. школа, 1983. – 511 с.
12. http://www.bio.fizten.ru
13. http://www.kazchem.ru
УЧЕБНОЕ ИЗДАНИЕ
Денисова Елена Сергеевна
Конспект лекций по дисциплине «Химия».
Учебное пособие
Ответственный за выпуск: И.А. Кибанова
Ответственный редактор: В.А. Филатова
Подписано в печать 18.05.2009
Формат 60х80/16. Бумага ксероксная
Гарнитура Times New Roman
Оперативный способ печати
Усл.печ.л. 10, уч.-изд.л. 10
Тираж 100 экз. Заказ № 31
Цена договорная
Омский институт водного транспорта (филиал)
ФГОУ ВПО «НГАВТ»
644099, г. Омск, ул. И. Алексеева, 4
Отпечатано в ООО «Технотранс»
644012, г. Омск, ул. 9 Дунайская, 20
Рекомендуемые страницы:
lektsia.com
Гуанин и цитозин, определение — Справочник химика 21
Межмолекулярные водородные связи образуются только между определенными парами производных пиримидина и пурина, которые называются комплементарными парами. Такими комплементарными парами являются урацил—аденин, тимин—аденин и цитозин—гуанин (рис. 103). [c.716]Согласно гипотезе Уотсона — Крика, четыре основания — аденин, ТИМИН, гуанин -и цитозин, обозначаемые соответственно буквами А, Т, О и С, — расположены в определенной последовательности в одной из [c.456]
В ДНК две спирали связаны друг с другом через определенные интервалы при помощи водородных связей. Изучение молекулярных моделей показало, что водородные связи могут образоваться лишь между аденином и тимином и между гуанином и цитозином водородные связи между другими парами оснований не позволят им разместиться в структуре двойной спирали. В соответствии с этим представлением отношения аденин тимин и гуанин цитозин равны 1 1. [c.1063]
РНК исследовались методом рентгеноструктурного анализа с целью установления того, имеют ли их полинуклеотидные цепи спиралевидную форму, аналогичную ДНК, но до настоящего времени определенных выводов сделать не удалось из-за отсутствия однородных образцов кристаллической РНК. Однако дрожжевую транспортную РНК удалось очистить и получить в кристаллическом состоянии. Получены хорошие рентгенограммы этого вещества, и они оказались чрезвычайно сходными с теми, которые дает ДНК таким образом, эти две структуры должны быть близкими. В соответствии с этим вероятное строение дрожжевой транспортной РНК должно быть таким, при котором каждая полинуклеотидная цепочка сложена вдвое по всей длине и скручена таким образом, что образует двойную спираль. Две половины цепи РНК соответствуют, следовательно, комплементарным цепям ДНК. Не вызывает сомнения, что спиральная структура поддерживается за счет водородных связей между парами оснований аденин — урацил и гуанин — цитозин (партнеры, образующие такую пару, находятся в разных половинах цепи). В месте перегиба цепи имеется несколько неспаренных оснований, и небольшой хвост неспаренных оснований имеется на одном из концов цепи. В этом состоит главное отличие спиральной конфигурации дрожжевой транспортной РНК от спиральной конфигурации ДНК. Дифракционная картина, полученная при рентгеноструктурном исследовании РНК из других источников, сходна с дифракционной картиной, полученной при исследовании дрожжевой транспортной РНК следовательно, спиральная конфигурация присуща, по-видимому, многим формам РНК. [c.142]
Полимерные молекулы ДНК состоят из расположенных в определенных последовательностях остатков четырех типов дезоксирибонуклеотидов, в которые в качестве углеводного компонента входит дезоксирибоза, а гетероциклическими основаниями являются аденин, гуанин, цитозин и тимин [c.443]
Нуклеиновые кислоты содержатся в каждой живой клетке. ДНК служит носителем генетической информации. Это обусловлено тесной связью между двумя витками спиралей нуклеиновых кислот, которая основана на очень специфических водородных связях между адениновым (А) остатком одного витка и тиминовым (Т) остатком другого витка, который расположен строго напротив первого, а также между цитозиновым (Ц) остатком одного витка и гуаниновым (Г) остатком другого. Такое образование пар абсолютно специфично аденин не может образовывать мультивалентные водородные связи с гуанином или цитозином, а цитозин не может образовывать связи с тимином или аденином. Изумительно, что вся наследственность и эволюция зависят от двух групп водородных связей Генетический код для синтеза определенной аминокислоты обус- [c.578]
Определенные основания в ДНК могут подвергаться изменениям под действием алкилирующих агентов например, диметилсульфат (рис. 30-5), обладающий высокой реакционной способностью, метилирует остатки гуанина. В результате образуется О-метилгуанин (рис. 30-7), который не способен спариваться с обычным партнером гуанина цитозином. Как у бактерии, так и в животных тканях имеются ферменты, которые специфически удаляют О-метилгуанин и заменяют его на нормальный гуанин. И в этом случае репарация осуществляется по механизму разрезал-залатал-зашил , сходному с тем, который показан на рис. 30-4. [c.968]
Здесь видно, как из одной молекулы ДНК могут образоваться две новые, совершенно тождественные по своей структуре молекулы. В самом деле, после расхождения цепей к основаниям одиночных цепей из окружающей среды присоединяются комплементарно соответствующие основания (или нуклеотиды) к аде-нину — тимин, к тимину — аденин, к гуанину — цитозин и, наконец, к цитозину — гуанин. В результате этого определяется расположение оснований во вто рой цепи в строгом соответствии с их расположением в первой цепи. Затем ферментные системы смыкают установленные определенным образом нуклеотиды во вторую комплементарную цепь, которая уже соединена с первой основной цепью водородными связями. Этот процесс специфического синтеза ДНК имеет место в период деления клетки, когда, как показывает химический анализ, действительно происходит удвоение количества ДНК. [c.60]
Последовательность нуклеотидов и генетический код. Методы определения последовательности аминокислот в полипептидной цепи были известны еще в 50-х гг. Теоретически это относительно легкая проблема, поскольку все 20 аминокислот, встречающиеся в природных белках, имеют разные свойства. С другой стороны, нуклеотидная последовательность ДНК относительно однородна по составу элементарных звеньев, так как содержит только четыре типа азотистых оснований-гуанин, цитозин, аденин и тимин. Когда еще в 60-х г. был расшифрован генетический код, появилась возможность восстанавливать (дедуцировать) нуклеотидную последовательность транскрибируемой ДНК по аминокислотной последовательности соответствующего белка. Однако генетический код является вырожденным, то есть одной и той же аминокислоте соответствуют несколько разных нуклеотидных триплетов. Следовательно, суждения о нуклеотидной последовательности, основанные на последовательности аминокислот в белке, не однозначны. Кроме того, последовательности аминокислот не содержат никакой информации о последовательности некодирующих участков ДНК. В настоящее время разработаны методы непосредственного секвенирования ДНК [117]. Принцип состоит в следующем длинную молекулу ДНК фрагментируют при помощи агентов, расщепляющих ее в специфических сайтах. Затем определяют последовательность нуклеотидов в каждом из этих фрагментов. Очередность фрагментов в целой молекуле восстанавливают, используя перекрывающиеся концы идентичные цепи разрезают повторно другой рестриктазой, а затем последовательности перекрывающихся фрагментов, образующихся при обработке двумя рестриктазами разной специфичности, сравнивают. Так может быть реконструирована полная последовательность. В пределах отдельных фрагментов порядок нуклеотидов определяют с помощью специальных методов. Раньше секвенирование ДНК было весьма трудным делом, теперь же оно [c.131]
&
www.chem21.info
|
Поиск Лекций
К азотистым основаниям относят аденин (A), гуанин (G), цитозин (C) и тимин (T), который входит в состав только ДНК, а урацил (U) заменяет его в РНК. Они обладают схожими структурами и химическими свойствами. Это гетероциклические органические соединения, производные пиримидина и пурина, входящие в состав нуклеотидов. Аденин и гуанин — производные пурина, а цитозин, урацил и тимин — производные пиримидина. Когда азотистые основания присоединяются ковалентной связью к 1′ атомам рибозы (в РНК) или дезоксирибозы (в ДНК), а к 5′-гидроксильной группе сахара присоединяется одна или несколько фосфатных групп, образуются нуклеотиды. стандартные нуклеотиды ДНК составляют триплеты — участки ДНК, кодирующие одну аминокислоту. Например, с триплета АУГ (ему соответствует аминокислота метионин) обычно начинается синтез белка на рибосомах. Нуклеозиды, приведённые в таблице, входят в состав моно-, ди- и трифосфатов. Например, аденозин входит в состав АТФ — важнейшего энергетического ресурса организма. Пурин C5N4h5 — гетероциклические соединения, имидазольные производные пиримидинов. Производные пурина играют важную роль в химии природных соединений (пуриновые основания ДНК и РНК; кофермент NAD; алкалоиды, кофеин и т. д.) и, благодаря этому, в фармацевтике — ядро пурина входит в состав некоторых антибиотиков. Пурин и ряд его производных обладают противоопухолевой, противовирусной и противоаллергической активностью. Аденин — азотистое основание, аминопроизводное пурина. Образует две водородных связи с урацилом (в РНК) и тимином (в ДНК) по правилу комплементарности. Представляет собой бесцветные кристаллы. Химическая формула С5H5N5. Аденин проявляет основные свойства. Аденин входит в состав многих жизненно важных для живых организмов соединений, таких как аденозин, аденозинфосфорные кислоты, нуклеиновые кислоты, адениновые нуклеотиды и др. В виде этих соединений аденин широко распространен в живой природе. Гуанин — азотистое основание, аминопроизводное пурина, является составной частью нуклеиновых кислот. Химическая формула — C5H5N5O. В ДНК и РНК образует три водородные связи с цитозином по правилу комплементарности. Производные гуанилового нуклеотида — ГДФ, ГТФ и цАМФ — участвуют во многих сигнальных путях клетки. Для некоторых процессов, происходящих в клетке — например, для сборки микротрубочек — ГТФ используется как источник энергии. Пиримидин C4N2h5 — гетероциклическое соединение, имеющее плоскую молекулу, простейший представитель диазинов. Пиримидин представляет собой бесцветные кристаллы с характерным запахом. Пиримидин проявляет свойства слабого двукислотного основания, так как атомы азота могут присоединять протоны. Производные пиримидина широко распространены в живой природе, где участвуют во многих важных биологических процессах. Его производные цитозин, тимин, урацил входят в состав нуклеотидов. Биологическая роль пиримидинов не ограничена нуклеиновыми кислотами. Некоторые пиримидиновые нуклеотиды играют важную роль в процессах обмена углеводов и липидов. Витамин В1 (тиамин) — пиримидиновое производное. Пиримидиновое ядро входит в состав некоторых коферментов и антибиотиков. Тимин — производное пиримидина. Формула C5H6N2O2. Присутствует во всех живых организмах, где вместе с дезоксирибозой входит в состав нуклеозида тимидина, который может фосфорилироваться 1-3 остатками фосфорной кислоты с образованием нуклеотидов тимидин моно-, ди- или трифосфорной кислоты (ТМФ, ТДФ и ТТФ). Дезоксирибонуклеотиды тимина входят в состав ДНК, в РНК на его месте располагается рибонуклеотид урацила. Тимин комплементарен аденину, образует с ним 2 водородные связи. Цитозин — азотистое основание, производное пиримидина. С рибозой образует нуклеозид цитидин, входит в состав нуклеотидов ДНК и РНК. Во время репликации и транскрипции по правилу комплементарности образует три водородных связи с гуанином. Представляет собой бесцветные кристаллы. Химическая формула C4H5N3O. Его производные цитозин, тимин, урацил входят в состав нуклеотидов, проявляет основные свойства. Урацил — пиримидиновое основание, которое является компонентом рибонуклеиновых кислот (РНК) и, как правило, отсутствует в дезоксирибонуклеиновых кислотах (ДНК). В составе РНК может комплементарно связываться с аденином, образуя две водородные связи. Эрвин Чаргафф открыл правила, описывающие количественные соотношения нуклеотидов. Правило Чаргаффа — биологический закон, в соответствии с которым установлены количественные соотношения между азотистыми основаниями разных типов. Для того, чтобы определить точные количественные соотношения нуклеотидов, Чаргафф разделил нуклеотиды ДНК методом бумажной хроматографии. Ему удалось выявить три закономерности: Число аденинов равно числу тиминов, а число гуанинов — числу цитозинов: А=Т, Г=Ц Число пуринов равно числу пиримидинов: А+Г=Т+Ц Число аденина и цитозина равно числу гуанина и тимина: А+Ц=Г+Т Состав ДНК разных организмов различается суммарным числом комплементарных оснований. Соотношение комплементарных нуклеотидов тоже может быть различным в разных молекулах ДНК. У одних оорганизмов в ДНК преобладают пары аденин-тимин, а у других — гуанин-цитозин. При этом правила Чаргаффа будут выполняться в любом случае. Вопрос Если в ДНК содержится 21 % аденина, то какова долю (%) остальных нуклеотидов, содержащихся в ДНК? Данные рентгеноструктурного анализа показали, что молекулы ДНК имеют спиральную структуру. В 50-х годах 20 века многие химики и биологи пытались исследовать структуру ДНК. В Королевском колледже в Лондоне Морис Уилкинс и Розалинда Франклин пытались решить эту проблему методом рентгеноструктурного анализа солей ДНК. Но такой способ позволял выявить только общую структуру молекулы. Тем временем Джеймс Уотсон и Френсис Крик в Кавендишской лаборатории Кембриджского университета, используя данные, полученные М. Уилкинсом, стали строить пространственные 3-D модели ДНК. Они пытались создать структуру, которая согласовывалась бы со всеми данными рентгеноструктурного анализа. В итоге Уотсон и Крик пришли к выводу, что ДНК имеет спиральную структуру с периодичностью 0,34 нм вдоль оси. В 1953 году Дж. Уотсон и Ф. Крик открыли строение ДНК, предложив модель двойной спирали. Нуклеиновые кислоты, подобно белкам, обладают первичной структурой (под которой подразумевается их нуклеотидная последовательность) и трехмерной структурой. Интерес к структуре ДНК усилился, когда в начале XX века возникло предположение, что ДНК, возможно, представляет собой генетический материал. В начале 50-х годов американский химик, лауреат Нобелевской премии Лайнус Полинг, уже изучивший к тому времени α-спиральную структуру, характерную для многих фибриллярных белков, обратился к исследованию структуры ДНК, которая по имеющимся в то время сведениям также представлялась фибриллярной молекулой. Одновременно в Королевском колледже Морис Уилкинс и Розалинда Франклин пытались решить ту же проблему методом рентгеноструктурного анализа. Их исследования требовали долгой и трудоёмкой работы по приготовлению чистых препаратов солей ДНК, для которых удавалось получать сложные дифракционные картины. С помощью этих картин можно было, однако, выявить лишь общую структуру молекулы ДНК, не столь детализированную, как та, которую позволяли получить чистые кристаллы белка. Тем временем Фрэнсис Крик и Джеймс Уотсон в Кавендишской лаборатории Кембриджского университета избрали иной подход, который в конечном счёте и обеспечил успешное решение проблемы. Используя все физические и химические данные, какие оказались в их распоряжении, Уотсон и Крик стали строить пространственные модели ДНК в надежде на то, что рано или поздно им удастся получить достаточно убедительную структуру, согласующуюся со всеми этими данными. Истории их поисков увлекательно описаны Уотсоном в его книге «Двойная спираль». Два обстоятельства оказались для Уотсона и Крика решающими. Во-первых, они имели возможность регулярно знакомиться с результатами работ Уилкинса и, сопоставляя с его рентгенограммами свои модели, могли таким образом проверять эти модели. А рентгенограммы Уилкинса свидетельствовали в пользу спиральной структуры с периодичностью 0,34 нм вдоль оси. Во-вторых, Уотсон и Крик отдавали себе отчёт в важном значении закономерностей, касающихся соотношения различных оснований в ДНК. Обнаружил эти закономерности и сообщил о них в 1951 году Эрвин Чаргафф. Это открытие, однако, при всей своей важности не привлекло к себе должного внимания. Уотсон и Крик задались целью проверить предположение, что молекула ДНК состоит из двух спиральных полинуклеотидных цепей, удерживаемых вместе благодаря спариванию оснований, принадлежащих соседним цепям. Основания удерживаются вместе водородными связями. Аденин спаривается с тимином, гуанин — с цитозином; АТ-пара соединяется двумя водородными связями, а ГЦ-пара — тремя. Уотсон попытался представить себе такой порядок спаривания оснований и позже вспоминал об этом так: «От радости я почувствовал себя на седьмом небе, ибо тут я уловил возможный ответ на мучившую нас загадку: почему число остатков пуринов в точности равно числу остатков пиримидинов?» Уотсон увидел, что при таком сочетании основания оказываются очень точно подогнанными друг к другу, а общий размер и форма этих двух пар оснований одинаковы, так как обе пары содержат по три кольца. Водородные связи при других сочетаниях оснований в принципе возможны, но они гораздо слабее. После того как все эти обстоятельства выяснились, можно было наконец приступить к созданию достоверной модели ДНК. Уотсон и Крик показали, что ДНК состоит из двух антипараллельных (направленных в противоположные стороны) полинуклеотидных цепей. Каждая цепь закручена в спираль вправо, и обе они свиты вместе, то есть закручены вправо вокруг одной и той же оси, образуя двойную спираль. Каждая цепь состоит из сахарофосфатного остова, вдоль которого перпендикулярно длинной оси спирали располагаются азотистые основания. Находящиеся друг против друга основания двух противоположных цепей двойной спирали связаны между собой водородными связями. Расстояние между сахарофосфатными остовами двух цепей постоянно и равно расстоянию, занимаемому парой оснований, то есть одним пурином и одним пиримидином. Два пурина занимали бы слишком много места, а два пиримидина — слишком мало для того, чтобы заполнить промежутки между двумя цепями. Вдоль оси молекулы соседние пары оснований располагаются на расстоянии 0,34 нм одна от другой, чем и объясняется обнаруженная на рентгенограммах периодичность. Полный оборот спирали приходится на 3,4 нм, то есть на 10 пар оснований. Никаких ограничений относительно последовательности нуклеотидов в одной цепи не существует, но в силу правила спариваний оснований эта последовательность в одной цепи определяет собой последовательность нуклеотидов в другой цепи. Поэтому мы говорим, что две цепи двойной спирали комплементарны друг другу. Уотсон и Крик опубликовали сообщение о своей модели ДНК в журнале «Nature» в 1953 г., а в 1962 г. они вместе с Морисом Уилкинсом были удостоены за эту работу Нобелевской премии. Розалинду Франклин, умершую от рака ранее присуждения этой премии, не включили в число лауреатов, поскольку Нобелевская премия посмертно не присуждается. Для того чтобы признать, что имеющая предложенную структуру молекула может выполнять роль генетического материала, требовалось показать, что она способна: 1) нести в себе закодированную информацию и 2) точно воспроизводиться (реплицироваться). Уотсон и Крик отдавали себе отчет в том, что их модель удовлетворяет этим требованиям. В конце своей первой статьи они отметили: «От нашего внимания не ускользнуло, что постулированное нами специфическое спаривание оснований сразу же позволяет постулировать и возможный механизм копирования для генетического материала». Во второй статье, опубликованной в том же 1953 году, они обсудили выводы, которые следовали из их модели для возможного механизма передачи наследственной информации. Это открытие показало, сколь явно структура может быть связана с функцией уже на молекулярном уровне. Оно дало начало стремительному развитию молекулярной биологии. Рекомендуемые страницы: |
|
poisk-ru.ru
Нуклеотидная пара аденин—тимин — Справочник химика 21
Водородные связи возникают между пуриновым основанием одной цепи и пиримидиновым основанием другой цепи в результате избирательного спаривания оснований. Аденин всегда образует водородные связи с тимином (А-Т), а гуанин с цитозином (Г-Ц). Правило образования двунитевой структуры ДНК (А-Т, Г-Ц) называется правилом комплементарности. (лат. сотр1етепШт — дополнение). Образование водородных связей между комплементарными парами обусловлено их пространственным соответствием (рис. 1У.8). Это приводит к тому, что нуклеотидные последовательности двух антипараллельньЕХ цепей ДНК всегда строго комплементарны друг к другу, а порядок чередования нуклеотидов в обеих цепях ДНК оказывается взаимообусловленным. Именно комплемен-тарностью определяется точное воспроизведение последовательности оснований при копировании (репликации) молекул ДНК. [c.62]Избыточность ДНК в геноме человека. Вскоре после того, как генетический код был расшифрован (в начале 60-х гг.), ученые пришли к выводу об избыточности ДНК в эукариотических клетках. По данным разных авторов, содержание ДНК в диплоидной клетке человека составляет примерно 7,3-10 г (размах от 6,6 до 8,0). Зная мол. массу оснований, можно подсчитать, что нуклеотидная пара А-Т (аденин—тимин) имеет массу 1,025 10 г, а нуклеотидная пара О—С (гуанин—цитозин)-1,027 10 г. Следовательно, весь диплоидный набор содержит приблизительно 7,1 10 нуклеотидных пар [c.114]
В нуклеиновых кислотах основными хромофорами являются пуриновые (аденин и гуанин) и пиримидиновые (цитозин и тимин у ДПК, цитозин и урацил у РПК) азотистые основания нуклеотидов. Наряду с к —основная полоса при 260 нм) вклад в обш ее поглош ение дают и п —> т1 -переходы ( плечи в области 280 — 320 нм) с участием неподеленной пары электронов гетероатомов азота и кислорода. Электронную структуру нуклеотидных оснований исследовали с помош ью метода молекулярных орбиталей, в том числе с учетом взаимодействия л-электронов. В результате удалось получить значения плотностей зарядов, локализованных у отдельных атомов. На основании этих данных можно судить о связи реакционной способности с отдельными участками молекулы (А. Пюльман, Б.Пюльман). Так, оказалось, что электрон-акцепторные свойства аденина обусловлены в основном атомом С в положении 6, а электрон-донорные свойства — атомом С в положении 8. Эта закономерность также подтверждается и на примере пиримидиновых [c.363]
Чаргафф [369] заметил, что молярное содержание гуанина и цитозина, с одной стороны, и аденина и тимина — с другой в любом данном препарате природной ДНК одинаковы. Это и понятно, и даже необходимо, в свете предложенной позже, в 1953 г., двойной спирали Крика и Уотсона. Аденин спаривается с тимином, а гуанин — с цитозином. Поэтому нуклеотидный состав ДНК любого вида можно выразить одним показателем. Этот показатель был определен для многих групп организмов у бактерий он варьирует между 23 и 74% ГЦ (см., например, [469, 470]). Разумеется, что в таком случае показатель для АТ-пар будет составлять 77— 26%. Предполагается, что количества ГЦ у родственных групп сравнительно близки. [c.30]
Репликация последовательности при ферментативных (Е. соН) синтезах ДНК была изучена с использованием меченных Р субстратов и ДНК затравки, изолированной из вирусных, бактериальных и животных источников. Гидролиз полученных полидезоксинуклеотидов диэстеразой до дезоксинуклеозид-З -фосфатов с последующим определением распределения радиоактивности показал, что в каждом случае присутствуют все 16 возможных динуклеотидных (ближайший сосед) последовательностей, что это распределение является уникальным, не случайным, репродуцируемым и не предопределяется нуклеотидным составом затравочной ДНК и что эта ферментативная репликация включает образование пар аденин — тимин и гуанин — цитозин в двух цепях с противоположной последовательностью оснований (т. е. с противоположной полярностью ), как в модели Уотсона и Крика. [c.322]
Уотсон и Крик выдвинули идею о специфическом спаривании на основании имевшихся в их распоряжении данных о нуклеотидном составе различных ДНК. Из этих даных следовало, что отношения аденин тимин и гуанин цитозин близки к единице. Уотсон и Крик дали наиболее вероятную схему образования пар. [c.217]
В 50-х годах нашего века впервые было высказано предположение, что нуклеиновые кислоты живой клетки, в частности ДНК, являются кодирующими системами по отношению к основным химическим компонентам протоплазмы — белкам. Отправным пунктом в анализе этого процесса было положение, что порядок аминокислот в молекуле белка (точнее — в его полипептидной цепи, т. е. первичных структурах) определяется порядком расположения мономеров ДНК — нуклеотидных пар в молекуле ДНК. Стало очевидным, что каждый из многих образующихся в клетке белков не может кодироваться ( шифроваться ) на нити ДНК всякий раз новым, качественно особым инфортиационным элементом. Тогда естественно было предположить, что в нити ДНК линейно расположены немногочисленные элементы (единицы информации), которые подобно буквам, складывающимся в слова, способны обеспечить специфические инструкции о том, какой белок необходимо синтезировать. В молекуле ДНК такими единицами являются азотистые основания, а именно аденин (А), гуанин (Г), цитозин (Ц) и тимин (Т), в нитях РНК вместо тимина — урацил (У). [c.57]
Метод седиментации в градиенте плотности СзС1 применяют, в частности, для разделения Н — и М -ДНК. В результате центрифугирования получают два отдельных пика, соответствующих разности в плотностях, равной всего 0,014 г/см . Относительные количества ДНК каждого типа определяют посредством фотографирования в ультрафиолете (интенсивность поглощения рассчитывают по степени почернения фотопластинки). Метод седиментации в градиенте плотности позволяет также изучать нуклеотидный состав ДНК из разных источников, поскольку различия в нуклеотидном составе приводят к различиям в плотности. (Препараты ДНК, содержащие большое количество пар гуанин — цитозин, обладают более высокой плотностью, чем препараты с высоким содержанием аденина и тимина.) Эти работы будут рассмотрены более подробно в разд. 3 и 4 гл. ХУП1. Применение описанного метода к белкам встречает ряд затруднений, связанных с тем, что молекулярный вес белков составляет всего 1% от молекулярного веса ДНК и они образуют в растворах с таким же градиентом плотности очень широкие полосы (в 10 раз шире, чем ДНК). Такая полоса может занять всю ячейку. Осаждение белков с помощью сульфата аммония, добавляемого в небольших количествах, позволяет заметно сузить полосы. [c.196]
www.chem21.info
Тимин цитозин урацил
Аденин гуанин
Пурин пиримидин
Пуриновых азотистых оснований два — аденин и гуанин (А и Г).
Пиримидиновых азотистых оснований три- тимин, цитозин и урацил (Т, Ц и У).
Название нуклеотида определяется его азотистым основанием и часто сокращается до одной буквы.
В молекуле ДНК 4 вида нуклеотидов: А, Г, Т, Ц.
Аденин (аденин + фосфорная кислота + рибоза или АФД) = А. Гуанин (ГФД)- Г. Тимин (ТФД) =У.Цитозин (ЦФД) =Ц.
В молекуле РНК тоже 4 вида нуклеотидов. А, Г, У, Ц.
Аденин (аденин + фосфорная кислота + рибоза или АФР) = А. Гуанин (ГФР)- Г. Урацил (ТФР) =У.Цитозин (ЦФР) =Ц.
Пример нуклеотидаРНК —АФР.Пример нуклеотида ДНК — АФД
Таким образом, на этом уровне ДНК и РНК отличаются по виду остатка сахара (рибоза и дезоксирибоза) и по одному виду азотистого основания. В ДНК это тимин, в РНК урацил.
Молекулы нуклеиновых кислот представляют собой цепочки, соединенные посредством прочных ковалентных связей между фосфорной кислотой одного нуклеотида и пентозой следующего нуклеотида.
Пример соединения аденина и цитозина в одной цепочке молекулы ДНК:
Вторичная структура ДНК имеет форму скрученной лестницы или двойной спирали. Две отдельные цепочки молекулы ДНК соединяются друг с другом посредством относительно слабых водородных связей между азотистыми основаниями нуклеотидов. Такие связи лежат в основе принципа комплементарного спаривания нуклеотидов. При комплементарном спаривании нуклеотидов соблюдаются строгие правила.
Большие (пуриновые) азотистые основания спариваются только с маленькими (пиримидиновыми).
Гуанин в любой молекуле (ДНК и РНК) может спариваться только с цитозином, образуя три водородные связи.
Аденин в молекуле может спариваться с тимином или с урацилом, образуя по две водородные связи. Если аденин спаривается с тимином, то синтезируется или работает молекула ДНК. Если аденин спаривается с урацилом, то синтезируется или работает молекула РНК.
Другие варианты спаривания нуклеотидов невозможны. Кратко принцип комплементарного спаривания азотистых оснований нуклеотидов записывают так:
Пример соединения двух пар нуклеотидов в молекуле ДНК
Этот принцип является не только способом связи параллельных цепочек ДНК. Он основа для репликации ДНК и реализации генетической информации (транскрипция при биосинтезе белка).
У некоторых вирусов двухцепочечной может быть РНК.
Связи между параллельными цепочками слабые (водородные), но многочисленные, что делает систему достаточно надежной.
Конкретная последовательность нуклеотидов в цепочках нуклеиновых кислот не случайная. Она передается из поколения в поколение по наследству и определяет генетический код.
Функции ДНК — хранение и реализация генетической информации.
РНК делят на 3 группы.
1. Матричная РНК (м-РНК) и близкая к ней информационная (и-РНК)- это комплементарная копия участка одной из цепочек ДНК. Она получена по принципу комплементарного спаривания азотистых оснований и представляет собой одну полинуклеотидную цепочку из разного количества нуклеотидов (от нескольких десятков до нескольких тысяч).
Функция и-РНК — перенос информации от хранилища (ДНК) к месту реализации, например, на рибосомы для сборки белка. Функционально и-РНК можно разделить на триплетные кодоны (тройки нуклеотидов, кодирующих аминокислоты).
2. Рибосомальные р-РНК, образуют большую и малую субъединицу рибосом. Их функция связана с ролью рибосом, как мест сборки белка.
3. Транспортные т-РНК, имеют сложную пространственную конфигурацию, но на плоскости изображаются в форме клеверного листа. По их главной части — триплетному антикодону — можно различить более 60 разновидностей т-РНК.
Функция т-РНК: перенос аминокислот к рибосомам на встречу с и-РНК для сборки белка.
Липиды— разнородные по строению вещества, как правило, не растворимые в воде и растворимые в органических растворителях.
Наиболее важными для нашего курса биологии являются 3 группы.
1. Нейтральные жиры- это сложные эфиры глицерина и жирных кислот.
Глицерин
3-net.ru
Азотистые основания пуриновые (аденин, гуанин) и пиримидиновые (тимин, урацил, цитозин). Строение, лактим-лактамная таутомерия.
Азо́тистые основа́ния— гетероциклические органические соединения, производные пиримидина и пурина, входящие в состав нуклеиновых кислот. Для сокращенного обозначения пользуются большими латинскими буквами. К азотистым основаниям относят аденин (A), гуанин (G), цитозин (C), которые входят в состав как ДНК, так и РНК. Тимин (T) входит в состав только ДНК, а урацил (U) встречается только в РНК.
Аденин и гуанин являются производными пурина, а цитозин, урацил и тимин — производными пиримидина.
Тимин, который присутствует только в ДНК, и урацил, который встречается только в РНК, обладают сходной химической структурой. Урацил отличается от тимина отсутствием метильной группы у 5-го атома углерода.
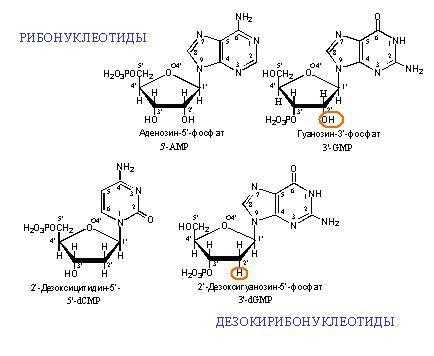
Азотистые основания, соединяясь ковалентной связью с 1′ атомом рибозы или дезоксирибозы, образуют N-гликозиды, которые называют нуклеозиды. Нуклеозиды, в которых к 5′-гидроксильной группе сахара присоединены одна или несколько фосфатных групп, называются нуклеотидами. Эти соединения являются строительными блоками молекул нуклеиновых кислот — ДНК и РНК.
После образования молекулы нуклеиновой кислоты входящие в её состав азотистые основания могут вступать в различные химические реакции под действием ферментов а также факторов внешней среды.
Азотистые основания одной из цепей соединены с азотистыми основаниями другой цепи водородными связями согласно принципу комплементарности.
Антипараллельностьцепей ДНК: противоположная направленность двух нитей двойной спирали ДНК; одна нить имеет направление от 5′ к 3′, другая — от 3′ к 5′.
ЛАКТИМ-ЛАКТАМНАЯ ТАУТОМЕРИЯ
Этот вид таутомерии характерен для азотсодержащих гетероциклов с фрагментом N=C—ОН.
Взаимопревращение таутомерных форм связано с переносом протона от гидроксильной группы, напоминающей фенольную ОН-группу, к основному центру — пиридиновому атому азота и наоборот. Обычно лактамная форма в равновесии преобладает.
Нуклеозиды: строение, номенклатура , гидролиз, характер связей
Нуклеиновые основания образуют связь за счет одного из атомов азота с аномерным центром пентозы (D-рибозы или 2-дезокси-D-рибозы). Этот тип связи аналогичен обычной гликозидной связи и известен как N-гликозидная связь, а сами гликозиды — как N-гликозиды. В химии нуклеиновых кислот их называют нуклеозидами.
В состав природных нуклеозидов пентозы входят в фуранозной форме (атомы углерода в них нумеруют цифрой со штрихом). Гликозидная связь осуществляется с атомом азота N-1 пиримидинового и N-9 пуринового оснований.
Природные нуклеозиды всегда являются β-аномерами.
В зависимости от природы углеводного остатка различают рибонуклеозиды и дезоксирибонуклеозиды. Для нуклеозидов употребительны названия, производимые от тривиального названия соответствующего нуклеинового основания с суффиксами -идин у пиримидиновых и -озин у пуриновых нуклеозидов.
infopedia.su
Leave A Comment