
Прокариотические водоросли — Справочник химика 21
Прокариотические сине-зеленые водоросли (цианобактерии) являются простейшими фотосинтезирующими организмами, выделяющими кислород. Они не содержат хлоропластов, но имеют более или менее развитые внутренние фотосинтетические мембраны, включающие хлорофилл а, каротиноиды и фикобилины. Опишите все изменения в морфологии, ультраструктуре и составе пигментов, которые можно ожидать при переносе цианобактерий из 1) условий низкой освещенности в условия высокой освещенности и высокого содержания кислорода, 2) условий высокой освещенности в условия низкой освещенности, 3) условий освещения белым светом в условия освещения зеленым или красным светом. [c.402]РЕСТРИКТАЗЫ (эндодезоксирибонуклеазы рестрикции), Арменты класса гидролаз, катализирующие гидролиз фосфодиэфирных связей чужеродных ДНК в большинстве прокариотических (бактерии и синезеленые водоросли) и иек-рых др.

Существует два больших класса клеток, отличающихся по строению и функциям. Наиболее древними и простыми по строению являются прокариотические клетки. Основные свойства, характерные для прокариот, можно рассмотреть на примере бактерий. Это одни из наиболее простых по строению клеток, отличающиеся малыми размерами и примитивным строением. Они не имеют ядра, и их генетический материал не защищен дополнительной внутриклеточной мембраной. Как правило, бактерии получают необходимую энергию из окружающей среды, причем глюкоза является основным ее источником. Разновидностью бактерий являются синезеленые водоросли, или цианобактерии, имеющие фотосистему, подобную растительным клеткам. Цианобактерии способны фиксировать азот, углекислый газ и выделять кислород. Таким образом, их нормальная жизнедеятельность может протекать при наличии только во- Электронная
До сих пор мы рассматривали нефтяные нентациклические углеводороды ряда гопана. Безусловно, эта структура является главной для тритерпанов любых нефтей. В геохимическом аспекте весьма симптоматично, что именно гопаны, скелет которых создается простейшей прокариотической клеткой бактерий или сине-зеленых водорослей, занимают такое ведуш ее положение в нефтях [48, 54]. Следует предположить, что углеводороды ряда гопана представляют собой результат деятельности древних микроорганизмов и среди прочих соединений входили в состав липидов их клеточных мембран, т. е. образование гопанов происходило на стадии раннего диагенеза органического вещества осадков.
Безусловно, эта структура является главной для тритерпанов любых нефтей. В геохимическом аспекте весьма симптоматично, что именно гопаны, скелет которых создается простейшей прокариотической клеткой бактерий или сине-зеленых водорослей, занимают такое ведуш ее положение в нефтях [48, 54]. Следует предположить, что углеводороды ряда гопана представляют собой результат деятельности древних микроорганизмов и среди прочих соединений входили в состав липидов их клеточных мембран, т. е. образование гопанов происходило на стадии раннего диагенеза органического вещества осадков.
Сине-зеленые водоросли 12, Е) были первыми организмами, начавшими выделять кислород в атмосферу, которая до того была в основном бескислородной. Понятно, что эти прокариотические водоросли были также первыми организмами, сумевшими выработать систему защиты от такого агрессивного элемента, каким является кислород, и стать толерантными к нему. Если первобытный океан содержал много ионов двухвалентного железа 24, А), то эти ионы могли быстро соединяться с высвобожденным кислородом, предоставляя, таким образом, водорослям длительное время для того, чтобы они привыкли к этому яду.
Если принять, что скользящие бактерии происходят от сине-зеленых водорослей, то, следовательно, они должны быть далеки от настоящих бактерий. Нитчатые серные бактерии тогда должны быть ближе к прокариотическим водорослям, чем к бактериям. Развилась ли их способность к использованию восстановленных соединений серы для энергетического метаболизма независимо от настоящих серных бактерий, или такая способность появилась заново, после того как она была в основном (но не совсем см. 12, Ж) латентной у сине-зеленых водорослей Сравнительное изучение деталей соответствующих механизмов позволит ответить на этот вопрос.
Ли [1107], однако, ставит под сомнение существование в прошлом прокариотических водорослей с другими пигмента- [c.196]
Всем прокариотическим организмам, включая грамположительные и грамотрицательные эубактерии, актиномицеты и синезеленые водоросли, а также архебактерии (метабактерии), свойственны 70S рибосомы.
Репликация, транскрипция и трансляция геномов органелл. В хлоропластах и митохондриях ДНК представлена небольшими двухцепочечными молекулами, обычно кольцевыми, и не связана с гистонами. Таким образом, генетическая информация органелл содержится в структурах, весьма сходных с хромосомами прокариот, хотя и значительно меньших по размерам. В каждой органелле имеется множество копий ДНК (до 40—50 в некоторых хлоропластах). Кроме того, хлоропласты и митохондрии содержат аппарат транскрипции и трансляции, включая специфические для органелл рибосомы, которые меньше цитоплазматических 808-рибосом и близки по величине к 708-рибосо-мам прокариот. Синтез белка в органеллах ингибируется хлорам нико-лом и некоторыми другими антибиотиками, подавляющими этот процесс и у прокариот, но не влияющими на синтез белка в цитоплазме эукариотической клетки.
В современном мире существуют два существенно различных по внутренней архитектуре типа клеток. Более сложная эукариотическая клетка является структурной единицей у растений, многоклеточных животных, простейших, грибов и всех групп, которые обычно относили к водорослям, кроме одной. Несмотря на крайнее разнообразие эукариотических клеток, обусловленное их специализацией в ходе эволюции этих групп, а также модификациями, которые они претерпевают во время дифференцировки у растений и животных, в основной архитектуре таких клеток всегда имеется много общих черт.
Все клетки имеют ограничивающую их плазматическую мембрану, цитоплазму, рибосомы и ядерную зону или ядро. Размеры и форма клеток определяются скоростями физической диффузии молекул питательных веществ и кислорода, а также соотношением между площадью поверхности и объемом клетки. Существуют два больших класса клеток прокариотические и эукариотические. Прокариоты, к которым относятся бактерии и сине-зеленые водоросли,-это простые клетки малых размеров, характеризующиеся тем, что содержащийся в них генетический материал не окружен мембраной.
В более примитивных прокариотических клетках ДНК не выделяется специальной дополнительной мембраной. Обычно эти клетки содержат одну гигантскую молекулу двуспиральной ДНК, состоящую из нескольких миллионов нуклеотидов. Иногда, по аналогии с эукариотической клеткой, ее называют хромосомной ДНК. В некоторых случаях в прокариотических клетках, в дополнение к этой ДНК, присутствуют еще и относительно маленькие молекулы ДНК (длиной в несколько тысяч- нуклеотидов), несущие дополнительную информацию их называют плазмидами. В большинстве случаев плазмиды копируются независимо от хромосомной ДНК и клетки могут содержать ряд подобных молекул. Несмотря на маленькие размеры, они придают клетке ряд особенностей, чрезвычайно важных для их выживания, например устойчивость к определенным антибиотикам. Прокариотические клетки обладают относительно маленькими размерами. Их линейные размеры имеют порядок 1 мкм, а самые маленькие из известных прокариотических клеток — микоплазмы — имеют размер около 0,3 мкм.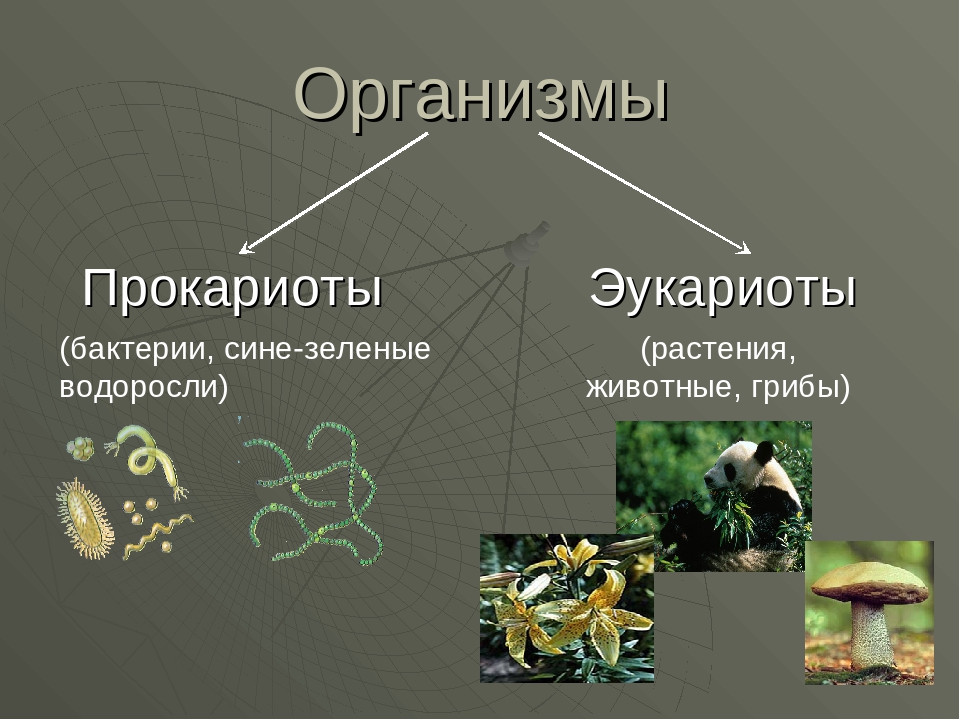
В гл. 1 мы уже говорили о том, что есть два больших класса клеток-прокариотические и эукариотические. Представители этих двух классов сильно различаются по размерам, по своей внутренней структуре, а также по генетической и метаболической организации. Прокариотические клетки, к которым принадлежат бактерии и сине-зеленые водоросли (цианобактерии),-это очень мелкие клетки, сравнительно простого строения, с одной-единственной мембранной системой, а именно мембраной, окружающей клетку.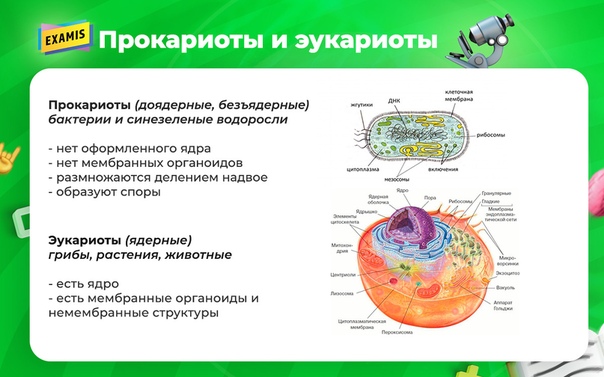 [c.396]
[c.396]
Микробиология — наука о живых организмах, не видимых невооруженным человеческим глазом, размерами менее 1 мм. Объектами микробиологии являются прокариотические организмы — бактерии и археи, а также эукариоты — простейшие, микроскопические водоросли, низшие грибы. [c.6]
Некоторые виды зеленых и других водорослей могут после соответствующего периода адаптации начать фотовосстановление СОг с помощью газообразного водорода как фотосинтезирующие бактерии, иными словами, у них появляется латентная в других условиях гидрогеназа [214, 633, 652, 654, 656, 983, 1707, 1771, 1813, 1814]. У таких растений протекают фотофосфорилирование и цикл Кальвина [705, 1582], но у них нет явления усиления интенсивности фотосинтеза [704] по-видимому, водород непосредственно попадает в фотосистему I. В отсутствие СОг некоторые водоросли могут выделять Нг на свету [76, 654, 661, 686]. Прокариотические и эукариотические растения могут до некоторой степени использовать на свету в качестве донора электронов HsS, хотя фотосистема II им, по-видимому, все-таки нужна [1018, 1801]. Длительное освещение хлоропластов красным светом, вероятно, подавляет эффект усиления у растений, например у элодеи [1489, 1490]. [c.133]
Длительное освещение хлоропластов красным светом, вероятно, подавляет эффект усиления у растений, например у элодеи [1489, 1490]. [c.133]
Однако хлоропласты и митохондрии эукариотических клеток содержат рибосомы, отличные от 80S типа. Рибосомы хлоропластов высших растений принадлежат к истинному 70S типу и практически не отличимы от рибосом эубактерий и синезеленых водорослей по вышеприведенным показателям и по более детальным молекулярным характеристикам. Митохондриальные рибосомы более разнообразны в зависимости от принадлежности организма к тому или иному царству эукариот. Наиболее изучены рибосомы митохондрий грибов и млекопитающих. Митохондриальные рибосомы грибов (Sa haromy es, Neurospora) похожи на прокариотические 70S рибосомы, но, может быть, лишь слегка крупнее (около 75S) и содержат относительно больше белка абсолютное содержание рибосомной РНК в них, повидимому, почти такое же, как в типичных 70S рибосомах. Митохондриальные рибосомы млекопитающих, однако, существенно мельче типичных 70S рибосом, имея также и существенно меньшее абсолютное количество рибосомной РНК на частицу их иногда называют мини-рибосомами . Действительно, коэффициент седиментации рибосом из митохондрий млекопитающих составляет всего около 55S, а тотальная масса рибосомной РНК на частицу более чем на 1/3 меньше, чем в типичных 70S рибосомах. В то же время, митохондриальные рибосомы млекопитающих содержат довольно много белка, так что общие размеры их как будто бы не сильно отличаются от таковых прокариотических рибосом. В целом, несмотря на ряд необычных черт, по ряду своих признаков, и в том числе по функциональному поведению, митохондриальные рибосомы млекопитающих все же близки к прокариотическим 70S рибосомам. [c.54]
Действительно, коэффициент седиментации рибосом из митохондрий млекопитающих составляет всего около 55S, а тотальная масса рибосомной РНК на частицу более чем на 1/3 меньше, чем в типичных 70S рибосомах. В то же время, митохондриальные рибосомы млекопитающих содержат довольно много белка, так что общие размеры их как будто бы не сильно отличаются от таковых прокариотических рибосом. В целом, несмотря на ряд необычных черт, по ряду своих признаков, и в том числе по функциональному поведению, митохондриальные рибосомы млекопитающих все же близки к прокариотическим 70S рибосомам. [c.54]
Попытки установить аминокислотную последовательность gg, (малый вариант) по последовательностям больших вариантов оказались неудачными. Только с помощью рентгеноструктурного анализа ggx удалось выявить родство обоих белков [571]. Благодаря этому рентгеноструктурному исследованию были ликвидированы неясности, связанные не только с установлением аминокислотной последовательности и не только у этого белка, но и у других цитохромов с известными последовательностями, например фотосинте-тических цитохромов с (называемых также /) прокариотических и эукариотических водорослей. [c.225]
[c.225]
Свободного хлорофилла в животных тканях обычно нет его можно обнаружить лишь в кишечнике травоядных. Некоторые виды животных, в частности водные беспозвоночные, содержат симбиотические одноклеточные водоросли. Интересным примером представляется одна из тропических асцидий, которая, как было обнаружено, содержит неизвестный ранее тип прокариотических зеленых водорослей РгосН1огоп, в настоящее время рассматриваемый многими исследователями как недостающее звено в эволюционной цепи высших растений и [c.166]
Однако следует подчеркнуть, что в вышеприведенной характеристике класса лучистых грибов опуш,ены представления об их ядер-ном аппарате. Между тем хорошо известно деление царства простейших организмов на высшие и низшие протисты соответственно организации ядра. У первых ядра оформленные — настоящие, у вторых — неоформленные, нуклеоиды. Ко вторым относятся бактерии и сине-зеленые водоросли. Несомненно, к ним относятся и прокариотические актиномицеты [22]. [c.150]
[c.150]
Особенности водных микроорганизмов. Водные микроорганизмы осуществляют в водоемах замкнутые циклы основных элементов, поскольку в микробных сообществах представлены и первичные продуценты органического вещества (эу- и прокариотические фотоавтотрофы, прокариоты-хемоавтотрофы), и консументы (про-стейщие), и деструкторы (большинство гетеротрофных прокариот и фибов). Водные микроорганизмы присутствуют в планктоне и бентосе, прикрепляясь к плавающим частицам и обитая в донных осадках. Важнейшие фуппы прокариот, обитающих в водных экосистемах, даны в табл. 30. Простейшие представлены фораминифе-рами и радиоляриями. Среди водорослей преобладают диатомеи. [c.285]
Как уже говорилось в гл. 1, прокариотический характер генетической системы органелл, особенно ярко выраженный у хлоропластов, позволяет предполагать, что митохондрии и хлоропласты произошли от бактерий, некогда поглощенных путем эндоцитоза Согласно этой эндосимбиотической гипотезе, клетки эукариот в начале своего эволюционного пути были анаэробными организмами без митохондрий и хлоропластов, а затем вступили в прочный симбиоз с бактериями и приспособили их систему окислительного фосфорилирования для своих нужд (рис.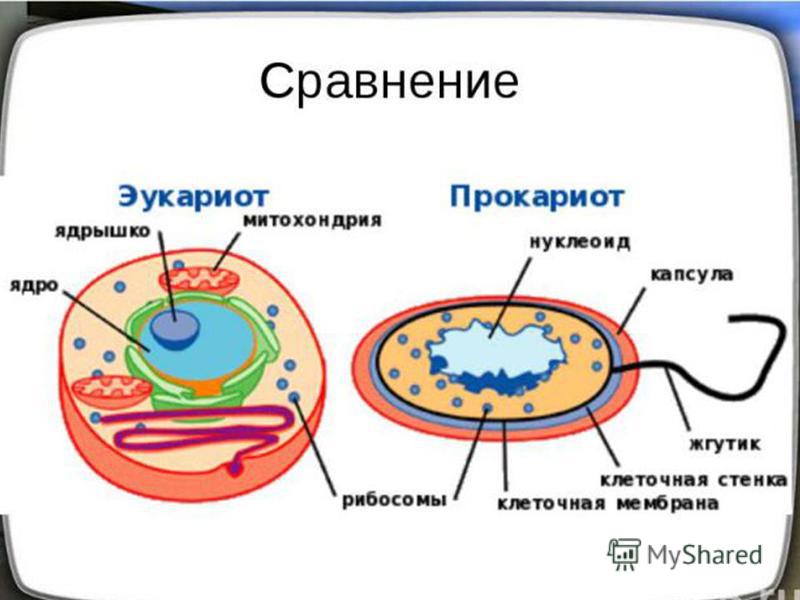 7-74) Полагают, что событие, приведшее к появлению митохондрий, произошло 1,5 млрд. лет назад, когда в атмосферу поступило значительное количество кислорода, еще до разделения линий животных и растений (см. рис. 7-61). Вероятно, хлоропласты растений и водорослей появились позднее в результате другого эндосимбиоза, когда клеткой были захвачены фотосинтезирующие бактерии, выделяющие молекулярный кислород. Обычно предполагают, что произошло по меньшей мере три независимых события этого рода, так как тогда можно было бы объяснить различие пигментов и других особенностей у современных высших растений и у зеленых, бурых и красных водорослей (см. рис. 7-62). [c.499]
7-74) Полагают, что событие, приведшее к появлению митохондрий, произошло 1,5 млрд. лет назад, когда в атмосферу поступило значительное количество кислорода, еще до разделения линий животных и растений (см. рис. 7-61). Вероятно, хлоропласты растений и водорослей появились позднее в результате другого эндосимбиоза, когда клеткой были захвачены фотосинтезирующие бактерии, выделяющие молекулярный кислород. Обычно предполагают, что произошло по меньшей мере три независимых события этого рода, так как тогда можно было бы объяснить различие пигментов и других особенностей у современных высших растений и у зеленых, бурых и красных водорослей (см. рис. 7-62). [c.499]
Хлоробиум-хлорофилл у зеленых фотосинтезирующих бактерий и хлорофилл Ь у большинства эукариотических растений, включая все высшие растения, характеризуются фотосенсибилизирующей активностью. Они переносят поглощенную световую энергию всех частей спектра, на которых бактериохлорофилл а и хлорофилл а поглощают плохо, к этим соединениям. Подобную же роль у прокариотических растений (сине-зеленых водорослей) и некоторых из эукариотических водорослей (красных водорослей) играют фико-билины — тетрапиррольные соединения с открытыми концами. [c.91]
Подобную же роль у прокариотических растений (сине-зеленых водорослей) и некоторых из эукариотических водорослей (красных водорослей) играют фико-билины — тетрапиррольные соединения с открытыми концами. [c.91]
Можно думать, что ранние протоводоросли [469, 470] еще не были способны к дыханию, а могли только переносить присутствие кислорода. По-видимому, такие организмы вымерли. В настоящее время не известны такие сине-зеленые водоросли (или другие растения), которые не были бы способны получать энергию путем дыхания, хотя, насколько известно, цикл лимонной кислоты у сине-зеленых водорослей неполон [1737]. Более того, редко наблюдался настоящий рост растений без кислорода. Даже зеленые водоросли [1339], адаптированные к водороду и не высвобождающие кислород 12, Ж), не могут расти без кислорода. Но быть может, кислород нужен не для дыхания, а для биосинтеза. Некоторое растения предпочитают пониженные давления кислорода [1801]. Но, как бы там ни было, прокариотические и эукариотические растения могут утрачивать фотосинтез и хлорофилл и жить только на дыхании 12, Е, 14, Е) [33, 1477—1479]. [c.141]
[c.141]
Стэниер [1777] же считает, что митоз возник еще до симбиоза в результате увеличения размеров и расширения функций прокариотического организма. Более того, по его мнению, длительность предполагаемого промежутка времени между появлением сине-зеленых водорослей и появлением дышащих организмов (для которых требуется хотя бы минимальное содержание кислорода в атмосфере) заставляет сомневаться в том, что нефотосинтезирующие эукариоты возникли первыми. Таким образом, предложенный им порядок должен быть следующим [c.195]
Действительно, большинство бактерий, как и грибы, являются гетеротрофами и осмотрофами. Но совершенно очевидно, что грибы, будучи эукариотами, лишь очень отдаленно связаны с прокариотическими бактериями. Однако тогда можно спросить почему лишь на основе их одинакового энергетического метаболизма сияе-зеленые водоросли группируют с эукариотическими, например зелеными или красными водорослями И все же эта терминология пока сохраняется. [c.204]
[c.204]
Предложение обратить этот порядок, внесенное Маргулис, стало возможным только благодаря концептуальному отделению прокариотических растений (сине-зеленых водорослей) от других растений, поскольку первые могли создать кислородную атмосферу, необходимую для простейших. Впрочем, большое разнообразие темновых биоэнергетических процессов, 01бнаруженных у простейших, само собой свидетельствует об их более раннем происхождении. [c.205]
Лишь часть протистов, а именно бактерии и сине-зеленые водоросли, имеют прокариотические клетки (табл. 12). Их относят к низшим протистам, а большинство водорослей, все грибы и простейшие имеют эукариотические клетки и относятся к высшим протистам. Таким образом, прокариотические низшие протисты существенно отличаются от эукариотических высших протистов, а также растений и животных (о предлагаемом делении всех организмов на четыре или пять царств см. [15]). Их надо считать самыми примитивными из живущих сейчас организмов, и этот факт лишь затемняется объединением высших и низших протистов в одно царствс. А ведь многие ботаники даже относят сине-зеленые к царству растений. [c.152]
А ведь многие ботаники даже относят сине-зеленые к царству растений. [c.152]
Эту главу, в которой собраны довольно разрозненные сведения о различных процессах, шедших на заре развития жизни, можно заключить короткой заметкой о сине-зеленых водорослях. Это довольно разнородная группа прокариотических организмов. Отдельные виды сине-зеленых водорослей сильно различаются по способам метаболизма. Но для нас важнее всего, что они обладают хлорофиллом, и, значит, способны к фотосинтезу [4, 3]. Из прокариотических фотосинтезирующих организмов наших дней сине-зеленые водоросли — самые важные. К тому же они фотолитотрофы. Хотя, как мы узнали из разд. 7 этой главы, ранняя жизнь, видимо, была фото органо трофной, производство кислорода должно было усилиться с появлением фотолитотрофов. Сейчас общепринято мнение, что сине-зеленые водоросли или другае, более примитивные организмы, родственные или не родственные им, но имеющие сходный обмен, сыграли ваншую роль в создании кислорода нашей атмосферы. Следующая стадия началась лишь с появлением эукариотической клетки и наших современных эукариотических зеленых растений. Как мы увидим, в гл. XIV, разд. 12, внешнее сходст- [c.157]
Следующая стадия началась лишь с появлением эукариотической клетки и наших современных эукариотических зеленых растений. Как мы увидим, в гл. XIV, разд. 12, внешнее сходст- [c.157]
Бактерии сине-зеленые
Сине-зеленые не имеют оформленных хроматофоров, а пигменты равномерно распределены в протоплазме (кроме центральной части). Ядерное вещество, так же как у бактерий, находится в диффузном состоянии. Оболочка часто покрыта слоем слизи. В результате слияния слизи отдельных клеток образуются колонии.[ …]
Бактерии, впервые увиденные в XVII в. изобретателем микроскопа голландцем Антони ван Левенгуком, представляют собой одноклеточные прокариотические организмы размером от 0,5 до 10—13 мкм. Подавляющее большинство бактерий — гетеротрофы, но среди них имеются и автотрофы —цианобактерии, обладающие фотосинтезирующей системой и содержащие хлорофилл, который придает им зеленую или сине-зеленую окраску. Собственно это объясняет, что часто цианобактерии именуют просто «синезеленые», а за внешнее сходство называют водорослями.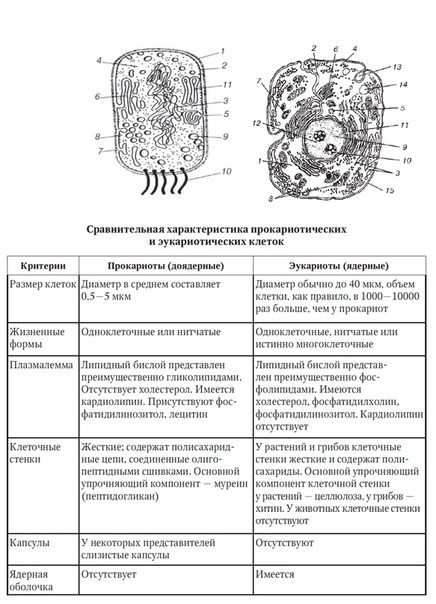 [ …]
[ …]
Бактерии являются самыми древними обитателями земли. Ископаемые бактерии», т. е. их отпечатки, найдены вместе с сине-зелеными водорослями в кембрийских .и докембрийских отложениях. От этих примитивных существ произошли все остальные организмы. Усложнение организмов привело к появлению нитчатых ветвящихся форм, от которых в дальнейшем произошли грибы.[ …]
Бактерии и сине-зеленые водоросли (циа-неи) — две филогенетически родственные группы — резко отличаются от всех остальных живых существ (в том числе от грибов) отсутствием истинного ядра и тем, что ДНК лежит в их клетке свободно, погруженная в так называемую нуклеоплазму, которая не отделена от цитоплазмы ядерной мембраной. У них нет также митохондрий и сложных жгутиков. Жгутики у них (когда они имеются) устроены проще и имеют принципиально иное строение, чем у остальных организмов; их клеточная стенка состоит из гетерополимерного вещества му-реина, которое не было обнаружено ни у одной другой группы организмов. Эти организмы называют прокариотами (Ргосагуо-1а — доядерные). У всех остальных организмов, как одноклеточных, так и многоклеточных, имеется настоящее ядро, окруженное ядерной мембраной и тем самым резко отграниченное от цитоплазмы. Такие организмы называют эукариотами (Еисагуо1а — ядерные). Кроме ясно дифференцированного ядра и цитоплазмы, у них имеются также митохондрии, а у многих также пластиды и сложные жгутики. Постепенно стало выясняться, что различия между прокариотами и эукариотами гораздо более глубокие и фундаментальные, чем, например, различия между высшими животными и высшими растениями (те и другие—эукариоты).[ …]
У всех остальных организмов, как одноклеточных, так и многоклеточных, имеется настоящее ядро, окруженное ядерной мембраной и тем самым резко отграниченное от цитоплазмы. Такие организмы называют эукариотами (Еисагуо1а — ядерные). Кроме ясно дифференцированного ядра и цитоплазмы, у них имеются также митохондрии, а у многих также пластиды и сложные жгутики. Постепенно стало выясняться, что различия между прокариотами и эукариотами гораздо более глубокие и фундаментальные, чем, например, различия между высшими животными и высшими растениями (те и другие—эукариоты).[ …]
У сине-зеленых водорослей (цианобактерий) фиксация азота может происходить как у свободноживущих форм, так и в симбиозах с грибами (в составе некоторых лишайников), или со мхами, папоротниками, а в одном известном случае — с семенным растением. На вайях мелкого плавающего водного папоротника Azolla имеются микроскопические поры, наполненные симбиотическими сине-зелеными водорослями АпаЪаепа, активно фиксирующими азот (Moore, 1969). Многие века этот папоротник играл важную роль на заливаемых рисовых полях Востока. До высаживания рассады риса залитые поля зарастают папоротником, который фиксирует достаточно азота для снабжения риса в период его созревания. Этот способ, а также стимуляция свободноживущих сине-зеленых водорослей позволяют выращивать рис сезон за сезоном на одном и том же поле, не внося удобрений. Как и в случае бактерий из клубеньков бобовых, симбиотические сине-зеленые водоросли более эффективны, чем свободноживущие [обзор фиксации азота сине-зелеными водорослями дал Питерс (Peters, 1978)].[ …]
Многие века этот папоротник играл важную роль на заливаемых рисовых полях Востока. До высаживания рассады риса залитые поля зарастают папоротником, который фиксирует достаточно азота для снабжения риса в период его созревания. Этот способ, а также стимуляция свободноживущих сине-зеленых водорослей позволяют выращивать рис сезон за сезоном на одном и том же поле, не внося удобрений. Как и в случае бактерий из клубеньков бобовых, симбиотические сине-зеленые водоросли более эффективны, чем свободноживущие [обзор фиксации азота сине-зелеными водорослями дал Питерс (Peters, 1978)].[ …]
Отдел сине-зеленых водорослей считают древнейшей группой автотрофных растений на Земле. Примитивное строение клетки, отсутствие полового размножения и жгутиковых стадий— все это серьезные доказательства их древности, lio цитологии сине-зеленые сходны с бактериями, а некоторые их пигменты (билипро-теины) встречаются и у красных водорослей. Однако, учитывая весь комплекс характерных для отдела признаков, можно предполагать, что сине-зеленые водоросли являются самостоятельной ветвью эволюции. Свыше трех миллиардов лет назад они отошли от основного ствола растительной эволюции и образовали тупиковую ветвь.[ …]
Свыше трех миллиардов лет назад они отошли от основного ствола растительной эволюции и образовали тупиковую ветвь.[ …]
Способность зеленых растений осуществлять фотосинтез обусловлена наличием у них пигментов. Максимальное поглощение света осуществляется хлорофиллом. Другие пигменты поглощают оставшуюся часть, преобразуя ее в различные виды энергии. В цветке покрытосеменных благодаря пигментации избирательно улавливается солнечный спектр с определенной длиной волны. Идея двух плазм в органическом мире предопределила симбиотрофное начало растений. Выделенные из всех частей растений симбиотические эндофиты класса Fungi imperfect синтезируют пигменты всех цветов, гормоны, ферменты, витамины, аминокислоты, липиды и поставляют их растению взамен полученных углеводов. Наследственная передача эндофитов гарантирует целостность системы. Некоторые виды растений имеют два вида экто-эндофитных микоризных грибов или грибов и бактерий, сочетание которых обеспечивает окраску цветков, рост и развитие растений (Гельцер, 1990). [ …]
[ …]
От пигментных бактерий могли взять начало сине-зеленые водоросли.[ …]
Эти данные указывают на возможность использования антагонистических бактерий, как и вирусов, для биологического контроля сине-зеленых водорослей. Возможно, что наиболее целесообразным подходом к такому контролю явится временное изменение окружающей среды, направленное на стимулирование природных антагонистов.[ …]
Действительно, сине-зеленые водоросли резко отличаются от других водорослей простотой внутренней организации клеток. Клетки их лишены оформленного ядра, что сближает их с бактериями. Вместе с бактериями сине-зеленые водоросли составляют раздел организмов, обозначаемый как прокариоты (Prokaryota), т. е. «доядерные», в отличие от всех остальных растений и животных, обладающих оформленным клеточным ядром и обозначаемых как эукариоты (Eukaryota), т. е. «истинно ядерные». Некоторые авторы придают этому признаку основное таксономическое значение и объединяют сине-зеленые водоросли с бактериями, т. е. вообще исключают их из числа собственно водорослей.[ …]
е. вообще исключают их из числа собственно водорослей.[ …]
Продукты распада сине-зеленых водорослей. Сине-зеленые водоросли относятся к группе низших, наиболее примитивных растений. В большинстве случаев это одноклеточные организмы, обычно соединяющиеся в колонии. У некоторых клетки при помощи слизи и выростов соединены в ценобии в виде нитей, давая внешнюю картину многоклеточности (рис. 9.1). Размножаются они преимущественно путем деления клеток. Живут сине-зеленые водоросли не только в воде, но и на суше (на берегах водоемов, в почвах и на их поверхности). Это самые распространенные растения земного шара. Они первыми заселяют бесструктурные почвы и совместно с бактериями подготавливают их для освоения другими растениями. Эти водоросли вообще аэробные организмы. Они способны к синтезу углеводов, но используют и распадающиеся органические вещества.[ …]
Эрой водорослей и бактерий называют протерозой, однако появились они еще раньше. Уже давно высказывались предположения, что архейские графиты и мраморы — результат жизнедеятельности каких-то древних организмов, а в последнее десятилетие удалось найти остатки живых существ, населявших древнейшие моря нашей планеты: в Южной Африке в архейских породах были обнаружены самые древние из известных в настоящее время организмов, жившие на Земле 3 млрд. 200 млн. лет назад. Они представляют собой шаровидные микроскопические тельца (диаметром от 5 до 30 мкм), по своему строению очень похожие на сине-зеленые водоросли.[ …]
200 млн. лет назад. Они представляют собой шаровидные микроскопические тельца (диаметром от 5 до 30 мкм), по своему строению очень похожие на сине-зеленые водоросли.[ …]
Существует несколько видов бактерий и сине-зеленых водорослей (к счастью, весьма многочисленных), которые способны фиксировать азот атмосферы. В результате их деятельности а также благодаря разложению органических остатков в почве растения-автотрофы получают возможность усваивать необходимый азот.[ …]
К этому виду близко примыкают бактерии Рэ. 8с1шПкППеп818, которые синтезируют сине-зеленый флуоресцирующий пигмент, диффундирующий в субстрат; температурный оптимум развития около 37 °С. Желатин разжижают медленно.[ …]
К ним относятся бактерии, сходные с простейшими и имеющие признаки сине-зеленых водорослей.[ …]
Значение фиксации азота некоторыми видами сине-зеленых водорослей в поставке связанного азота в озере показано в ряде исследований [17—19]. Способность фиксировать азот объясняет тот факт, что хотя сине-зе-леные водоросли часто встречаются в водах, содержащих мало связанного азота, они тем не менее имеют высокое содержание этого элемента на единицу массы. Бактерии также в некоторых случаях фиксируют азот в озерах [20].[ …]
Бактерии также в некоторых случаях фиксируют азот в озерах [20].[ …]
Помимо нескольких вирусов, которые заражают сине-зеленые водоросли, до сих пор пока неизвестны вирусы, которые поражали бы представителей этой группы растений. Однако в настоящее время па основании биохимических и улътраструктурных исследований сине-зеленых водорослей стало очевидно, что они в этом отношении имеют большее сходство с бактериями, чем с другими группами водорослей. Количество полученной в их опытах инфекционной РНК было невелико и нарастало медленно, в связи с чем данный вопрос требует дальнейшего изучения.[ …]
К прокариотическим организмам относятся (см. стр. 56) бактерии и так называемые сине-зеленые водоросли. Они выделяются отсутствием в их клетках ядер, что, вероятно, свидетельствует о сохранении ими древней организации клетки.[ …]
На градирнях биологические обрастания состоят 1[ …]
По-видимому, во всех водоемах-охладителях появляется больше бактерий и сине-зеленых, чем их было раньше. Этот факт зарегистрирован для Копорской губы у Ленинградской АЭС (Сазыкина, 1991), для озер Удомля и Песьво — водоемов-охладителей Калининской АЭС (Волкова, 1997).[ …]
Этот факт зарегистрирован для Копорской губы у Ленинградской АЭС (Сазыкина, 1991), для озер Удомля и Песьво — водоемов-охладителей Калининской АЭС (Волкова, 1997).[ …]
Некоторые виды микробов образуют различные пигменты: красные, синие, зеленые, оранжевые, бурые, черные и смешанных цветов. Большинство микробов не образуют пигментов, их колонии бесцветны. Пигменты — признак стабильный, который присущ определенным видам микробов. Этот признак может быть использован и в систематике бактерий. Биологическое значение пигментов пока малоизвестно.[ …]
Настоящий том посвящен прокариотным организмам (за исключением сине-зеленых водорослей, которые будут описаны в 3-м томе). Хотя раньше бактерии относили к растительным организмам, в настоящее время показано, что бактерии как прокариоты составляют особое царство живых существ, отличное от царств растений и животных. Следуя традиции, редакция начинает издание «Жизнь растений» с описания бактерий как низших организмов — протоцитов. В первой части раздела по бактериям (введении) кратко излагаются основные принципы классификации, строение клеток и обмен веществ у бактерий. Во второй части приводятся сведения об основных систематических группах бактерий. Заключают раздел главы, описывающие экологию, физиологические группы и биохимические особенности бактерий.[ …]
Во второй части приводятся сведения об основных систематических группах бактерий. Заключают раздел главы, описывающие экологию, физиологические группы и биохимические особенности бактерий.[ …]
В водной среде также существуют различные виды нитрофи-цирующих бактерий, но главная роль в фиксации атмосферного азота здесь принадлежит многочисленным видам способных к фотосинтезу сине-зеленых водорослей из родов Anabaena, Nostoc, Frichodesmium и др.[ …]
Особенно большую опасность представляет «цветение», вызываемое сине-зелеными и другими токсичными видами водорослей. Сине-зеленые водоросли играют особую роль в экосистемах современных водоемов. Они занимают промежуточное положение между бактериями и растениями, так что их часто называют цианобактериями. Сине-зеленые водоросли появились на Земле более 3 млрд. лет назад, были первыми фотосинтезирующими организмами, образовавшими аэробную систему Земли. Сине-зеленые водоросли обладают колоссальным потенциалом размножения: за 70 дней вегетационного периода одна клетка может дать Ю20 потомков.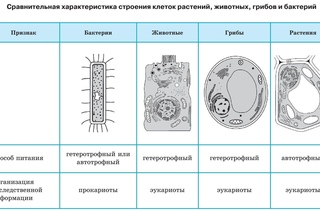 К благоприятным условиям для размножения сине-зеленых водорослей относятся низкое содержание кислорода, т.е. более восстановительная среда. Сине-зеленые водоросли — единственные обитатели Земли, которые способны усваивать четыре вида газов: С02 (фотосинтез, как у зеленых растений), 02 (дыхание), Ы2 (азотфиксация), Н23 (как бактерии в процессах хемосинтеза).[ …]
К благоприятным условиям для размножения сине-зеленых водорослей относятся низкое содержание кислорода, т.е. более восстановительная среда. Сине-зеленые водоросли — единственные обитатели Земли, которые способны усваивать четыре вида газов: С02 (фотосинтез, как у зеленых растений), 02 (дыхание), Ы2 (азотфиксация), Н23 (как бактерии в процессах хемосинтеза).[ …]
Все они синтезируют водорастворимые флуоресцирующие пигменты от желто-зеленого до оранжевого оттенка. Некоторые из них образуют синий пигмент, нерастворимый в воде. Бактерии этой группы активно разлагают углеводы. При росте на средах с гексозами образуют кислоты; на средах с лактозой и крахмалом не развиваются. Потребляют органические кислоты, растут на средах с ароматическими соединениями; аэробы. Восстанавливают нитраты до нитритов. Сероводород и индол не образуют. Распространены в воде, почве, сточных водах.[ …]
Низшие организмы. Концентрация хинона менее 1 мг/л селективно ядовита для сине-зеленых водорослей; но хинон не ядовит в этой концентрации для большинства зеленых водорослей и высших растений [121]. Для бактерий Escherichia coli предельно вредная концентрация хинона составляет 55 мг /л, для водорослей Scenedesmus она составляет 6 мг /л и для рачков Daphnia magna — 0,4 мг/л [139].[ …]
Для бактерий Escherichia coli предельно вредная концентрация хинона составляет 55 мг /л, для водорослей Scenedesmus она составляет 6 мг /л и для рачков Daphnia magna — 0,4 мг/л [139].[ …]
Солнечную энергию способны непосредственно использовать только клетки зеленых растений, одноклеточных водорослей, зеленых и пурпурных бактерий. За счет этой энергии они синтезируют органические соединения: углеводы, жиры, белки, нуклеиновые кислоты и др. Такой биосинтез, который происходит благодаря энергии света, и называют фотосинтезом. Отметим, что зеленый цвет фотосинтезирующих клеток зависит от наличия в них хлорофилла, поглощающего свет в красной и синей частях спектра и пропускающего лучи, которые дают при их смешении зеленый цвет. Некоторые водоросли и бактерии имеют и иные светопоглощающие пигменты, что придает им бурый, красный или пурпурный цвет.[ …]
Поступление органических загрязнений в водоем часто способствует бурному развитию сине-зеленых водорослей, что приводит к так называемому «цветению» воды и обрастанию подводных предметов, т. е. к развитию на их поверхности некоторых бактерий, грибов, водорослей, простейших. В зависимости от природы и количества развивающихся водорослей цветение воды может играть или положительную роль, ускоряя самоочищение воды, или отрицательную, ухудшая физико-химические ее свойства. Во время массового цветения вода становится мутной, зеленой, в ней появляются неприятные привкусы и запахи, и она делается непригодной для водоснабжения населения. При массовом отмирании водорослей образуются различные продукты их распада, поглощающие кислород из воды и токсические вещества. Все это вызывает вторичное загрязнение водоема.[ …]
е. к развитию на их поверхности некоторых бактерий, грибов, водорослей, простейших. В зависимости от природы и количества развивающихся водорослей цветение воды может играть или положительную роль, ускоряя самоочищение воды, или отрицательную, ухудшая физико-химические ее свойства. Во время массового цветения вода становится мутной, зеленой, в ней появляются неприятные привкусы и запахи, и она делается непригодной для водоснабжения населения. При массовом отмирании водорослей образуются различные продукты их распада, поглощающие кислород из воды и токсические вещества. Все это вызывает вторичное загрязнение водоема.[ …]
Фиксация — восстановительный процесс, осуществляемый такими микроорганизмами, как бактерии и сине-зеленые водоросли. Он требует аденозинтрифосфата (АТФ), который образуется при фотосинтезе, и, следовательно, данный процесс неэффективен в ночное время. В то же время процесс может подавляться при большом поступлений радиации, так что максимальные скорости фиксации часто отмечаются на некотором расстоянии от поверхности воды. Процесс идет интенсивнее и в том случае, если концентрация фосфора в водной среде относительно высока.[ …]
Процесс идет интенсивнее и в том случае, если концентрация фосфора в водной среде относительно высока.[ …]
Строение цианобактерий (по старой ботанической терминологии — сине-зеленых водорослей) до некоторой степени сходно со строением бактерий (рис. 2). Известно около 2500 видов. В основном они являются одноклеточными организмами разной формы (округлой, цилиндрической), но могут образовывать длинные многоклеточные нити или даже объединяться в колонии. Однако они отличаются от настоящих бактерий тем, что их клеточные стенки содержат некоторое количество целлюлозы и что они способны к фотосинтезу, т. к. в цитоплазме содержат хлорофилл (в гранулах, но не в хлоропластах) и другие пигменты (каротин, ксантофил и фикобилины), создающие их окраску. Некоторые клетки в многоклеточных цианобактериях обладают способностью фиксировать азот атмосферы.[ …]
Прокариоты — это простейшие организмы, клетки которых не имеют истинного ядра. К ним относятся бактерии и сине-зеленые водоросли.[ . ..]
..]
Для этого водоема характерно наличие трех максимумов развития фитопланктона: весной доминировали диатомовые и сине-зеленые водоросли, летом и осенью — диатомовые и криптомонады. По уровню развития водорослей озеро относилось к эвтрофному типу (Ми-неева, 1994; Корнева, 1994). В нем складывались относительно благоприятные условия для развития зооперифитона — наличие субстратов в виде макрофитов, затопленных пней и коряг, обилие пищи в виде бактерий и водорослей, удовлетворительный гидрохимический режим. Стрессовую ситуацию для гидробионтов могло создавать эпизодическое снижение pH воды до 5,5 во время половодья, но в этот период беспозвоночные находятся еще в малоактивном состоянии.[ …]
Автотрофные организмы используют неорганические источники для своего существования, тем самым создавая органическую материю из неорганической. К ним относятся фотосинтезирующие зеленые растения суши и водной среды, сине-зеленые водоросли, некоторые хемосинтезирующие бактерии и др.[ …]
Основная часть азота находится в воздухе (78%).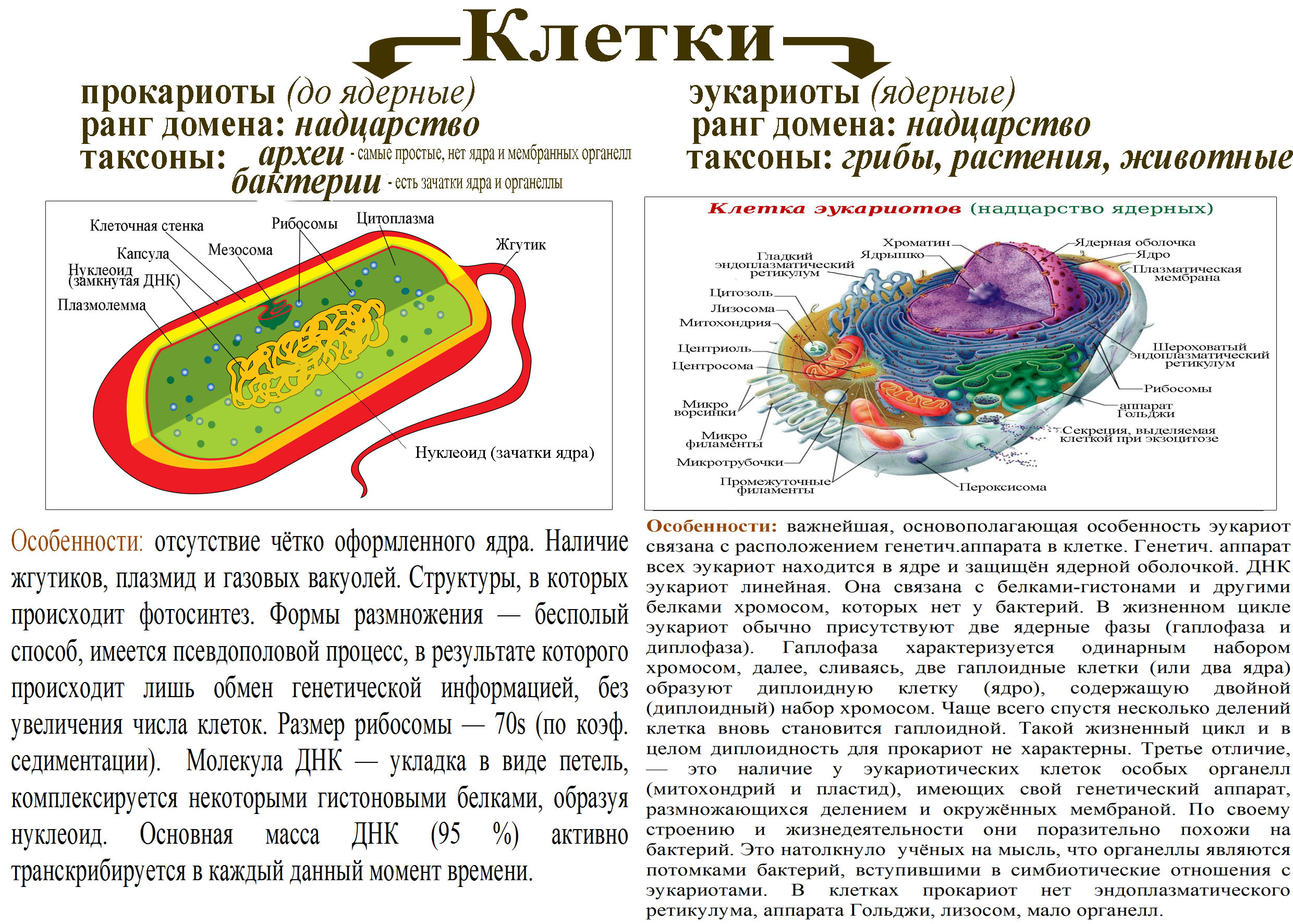 Однако растения не могут усваивать азот непосредственно, а только в виде ионов 1ЧНЦ+ и N03“. Фиксацию газообразного состояния осуществляют бактерии и сине-зеленые водоросли (цианеи). Основное значение в данной группе имеют бактерии, живущие на клубеньках бобовых растений. Растения обеспечивают бактерии местообитанием и пищей (сахарами), получая от них взамен доступную форму азота. По пищевым цепям органический азот передается от бобовых к другим организмам экосистемы. Органические соединения азота после гибели организмов при помощи бактерий разлагаются до аммиака и нитратов (N63). Нитраты частично вновь поглощаются растениями, частично восстанавливаются до N2, вновь поступающего в атмосферу.[ …]
Однако растения не могут усваивать азот непосредственно, а только в виде ионов 1ЧНЦ+ и N03“. Фиксацию газообразного состояния осуществляют бактерии и сине-зеленые водоросли (цианеи). Основное значение в данной группе имеют бактерии, живущие на клубеньках бобовых растений. Растения обеспечивают бактерии местообитанием и пищей (сахарами), получая от них взамен доступную форму азота. По пищевым цепям органический азот передается от бобовых к другим организмам экосистемы. Органические соединения азота после гибели организмов при помощи бактерий разлагаются до аммиака и нитратов (N63). Нитраты частично вновь поглощаются растениями, частично восстанавливаются до N2, вновь поступающего в атмосферу.[ …]
Цвет воды в озере испытывает сезонные колебания и неоднороден в различных частях озера, так же как и прозрачность. Так, в открытой части оз. Байкал при большой прозрачности вода имеет темно-синий цвет, в районе Селенгинского мелководья — серовато-зеленый, а вблизи р. Селенги — даже бурый. В Телецком озере в открытой части цвет воды зеленый, а у берегов желто-зеленый. Массовое развитие планктона уменьшает не только прозрачность, но и изменяет цвет озера, придавая ему цвет находящихся в воде организмов. Во время цветения зеленые водоросли окрашивают озеро в зеленый цвет, сине-зеленые придают ему бирюзовую окраску, диатомовые— желтую, а некоторые бактерии окрашивают озеро в малиновый и красный цвета.[ …]
В Телецком озере в открытой части цвет воды зеленый, а у берегов желто-зеленый. Массовое развитие планктона уменьшает не только прозрачность, но и изменяет цвет озера, придавая ему цвет находящихся в воде организмов. Во время цветения зеленые водоросли окрашивают озеро в зеленый цвет, сине-зеленые придают ему бирюзовую окраску, диатомовые— желтую, а некоторые бактерии окрашивают озеро в малиновый и красный цвета.[ …]
У одноклеточных организмов вегетативное размножение представлено такими формами, как деление, множественное деление и почкование. Деление путем простой перетяжки с образованием при этом из одного родительского организма двух дочерних присуще бактериям и сине-зеленым водорослям (цианобактериям). Напротив, размножение делением бурых и зеленых водорослей, а также одноклеточных животных (саркодовых, жгутиковых и инфузорий) происходит путем митотического деления ядра с последующей перетяжкой цитоплазмы.[ …]
Все процессы жизнедеятельности протекают при определенной температуре в основном от 10 до +40 °С. Лишь немногие организмы приспособлены к жизни при более высоких температурах. Например, некоторые моллюски живут в термальных источниках при температуре до +53°, сине-зеленые (цианобактерии) и бактерии могут обитать при +70° — +85 °С. Оптимальная температура для жизни большинства организмов колеблется в узких пределах от 10 до +30 °С. Однако диапазон колебания температур на суше значительно шире (от —50 до +40 °С), чем в воде (от 0 до —1-40 °С), поэтому предел устойчивости к температуре у водных организмов уже, чем у наземных.[ …]
Лишь немногие организмы приспособлены к жизни при более высоких температурах. Например, некоторые моллюски живут в термальных источниках при температуре до +53°, сине-зеленые (цианобактерии) и бактерии могут обитать при +70° — +85 °С. Оптимальная температура для жизни большинства организмов колеблется в узких пределах от 10 до +30 °С. Однако диапазон колебания температур на суше значительно шире (от —50 до +40 °С), чем в воде (от 0 до —1-40 °С), поэтому предел устойчивости к температуре у водных организмов уже, чем у наземных.[ …]
Слоевцовые эукариотические растения бывают и автотрофными, тогда их называют водорослями, и гетеротрофными; объединяющего общепринятого термина для обозначения последних нет. В эту категорию входят грибы и мик сомице ты (слизевики). Нередко эту категорию гетеротрофных низших растений понимают в широком смысле, присоединяя к ним бактерии из числа прокариотических организмов. Аналогично к числу водорослей относят прокариотические цианеи, называя их сине-зелеными водорослями.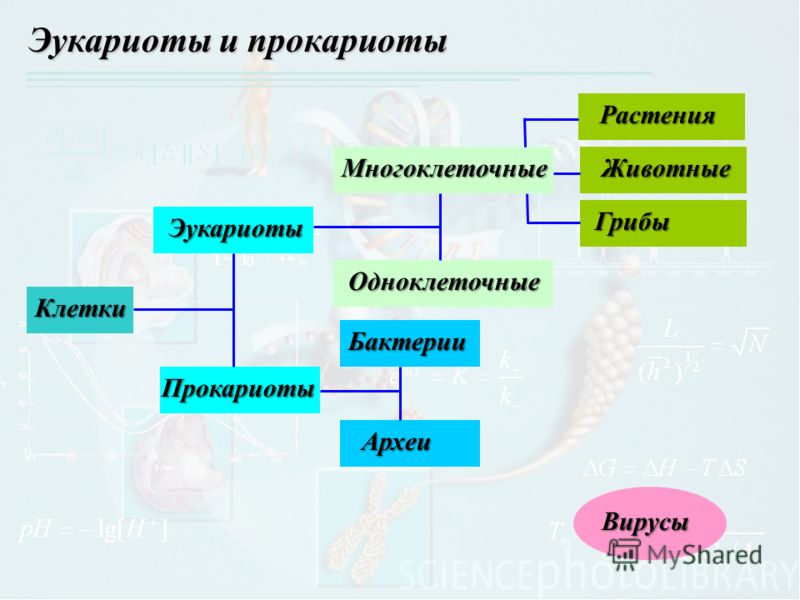 [ …]
[ …]
Большая часть микроорганизмов бесцветна. Колонии дрожжей обычно окрашены в слегка беловатые, кремоватые или сероватые тона. Дикие дрожжи иногда бывают окрашены в красные или розоватые цвета и редко в черные. Многие актиномицеты образуют различные пигменты и бывают окрашены в красные, розовые, зеленоватые и черные тона. У грибов окрашены споры, конидии и поверхностный слой гиф в черные, зеленые, желтые цвета. Окраска у микроорганизмов связана с наличием пигментов, которые являются отбросными продуктами обмена вещест» клетки. Только пигменты некоторых бактерий участвуют в процессах фотосинтеза. Они бывают окрашены в желтые, красные, сине-зеленые цвета.[ …]
Ядро бактериальной клетки. Примерно 1—2% веса сухой массы микроорганизмов приходится на ДНК, в которой заложена генетическая информация организма. У большинства микроорганизмов имеются области (или несколько областей), в которой сконцентрировано основное количество ДНК, имеющие определенную структуру (или органеллу) и называющиеся ядром. Ядро (или ядерное вещество) связано с цитоплазматической мембраной, независимо от того, окружено оно элементарными мембранами (как у амебы) или не имеет их (как у бактерий и сине-зеленых водорослей). Ядерное вещество активизируется в период размножения и при наступлении возрастных изменений, связанных со старением клетки.[ …]
Ядро (или ядерное вещество) связано с цитоплазматической мембраной, независимо от того, окружено оно элементарными мембранами (как у амебы) или не имеет их (как у бактерий и сине-зеленых водорослей). Ядерное вещество активизируется в период размножения и при наступлении возрастных изменений, связанных со старением клетки.[ …]
Проростки, выросшие в отсутствии света, называют этиолированными. Такие проростки, как правило, характеризуются измененпой формой (вытяпутые стебли, перазяившиеся листья) и слабой желтой окраской (хлорофилла в них пет). Вместе с тем, еще со времен Сакса (1864) известно, что в некоторых случвях хлорофилл образуется п в отсутствии света. Способность образовывать хлорофилл в темноте характерна для растений, стоящих иа нижней ступени эволюционного процесса. Так, при благоприятных условиях питания пепоторые бактерии могут синтезировать в темноте желто-зеленый пигмент — бак-териохлорофилл. Сине-зеленые водоросли при достаточном снабжении органическим веществом растут и образуют пигменты в темноте. [ …]
[ …]
Примечательной особенностью всех саговниковых являются растущие вверх над землей и дихотомически ветвящиеся коралловидные корни — кораллоиды. Они возникают как разветвления боковых корней эндогенно из многорядного перицикла напротив лучей первичной ксилемы. Благодаря интенсивному дихотомическому ветвлению коротких и тонких боковых корней образуются целые грозди клубеньков, окружающие ствол у его основания и напоминающие внешне кораллы. Сначала считали, что это происходит под влиянием бактерий, проникающих в клетки коры корней. В последнее время высказывается предположение, что бактерии, как и сине-зеленые водоросли, являются вторичными поселенцами в сформировавшихся уже клубеньках, а сам клубенек представляет разрастание несущего его корня, вызванное эндофитным грибом, мицелий которого обильно заполняет межклетники в коровой паренхиме этого корня.[ …]
На этой точке зрения особенно настаивает Маргулис (Margulis, 1975). Позднее поглощение клеток сине-зеленых водорослей могло бы привести к появлению автотрофного «организма», предка всего царства растений.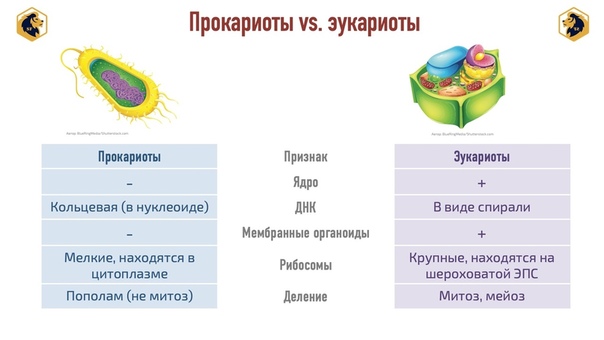 Эта последовательность событий является чисто гипотетической и вызывает много скептических замечаний, но, по крайней мере отчасти, она может соответствовать действительности. До сих пор существуют близкие к предполагаемым симбионтам формы, включая бактерию Paracoccus denitrificans, рядом признаков напоминающую гипотетического свободножи-вущего предшественника митохондрии. Если теория Маргулис верна, то основная часть этой главы была посвящена второй стадии развития мутуализма, когда все более интегрируются друг с другом пары видов, каждый из которых по происхождению является симбиотической ассоциацией.[ …]
Эта последовательность событий является чисто гипотетической и вызывает много скептических замечаний, но, по крайней мере отчасти, она может соответствовать действительности. До сих пор существуют близкие к предполагаемым симбионтам формы, включая бактерию Paracoccus denitrificans, рядом признаков напоминающую гипотетического свободножи-вущего предшественника митохондрии. Если теория Маргулис верна, то основная часть этой главы была посвящена второй стадии развития мутуализма, когда все более интегрируются друг с другом пары видов, каждый из которых по происхождению является симбиотической ассоциацией.[ …]
Изменения состава атмосферы оказали огромное влияние на жизнедеятельность обитателей нашей планеты. Первоначально в восстановительной среде простейшие одноклеточные организмы участвовали только в энергетически невыгодных реакциях гликолиза; лишь с появлением б атмосфере свободного кислорода стало возможным окисление и разрушение энергетически богатых субстратов. Продуктами распада органических соединений являются СОг и Н20. Этот процесс осуществляется с появлением таких организмов, как эукариоты , что привело к процессам дыхания, дало толчок бурному развитию новых форм и видов живых организмов; если в период, отстоящий от нашего времени на 3,5—1,5 млрд. лет, существовали только бактерии и сине-зеленые водоросли, то за последние 1,5 млрд. лет возникли все виды животных, а также водоросли, грибы и растения. За эти последние 1,5 млрд. лет установилось и содержание кислорода в атмосфере, составлявшее сначала около одного процента и достигшее в настоящее время 21%.[ …]
Этот процесс осуществляется с появлением таких организмов, как эукариоты , что привело к процессам дыхания, дало толчок бурному развитию новых форм и видов живых организмов; если в период, отстоящий от нашего времени на 3,5—1,5 млрд. лет, существовали только бактерии и сине-зеленые водоросли, то за последние 1,5 млрд. лет возникли все виды животных, а также водоросли, грибы и растения. За эти последние 1,5 млрд. лет установилось и содержание кислорода в атмосфере, составлявшее сначала около одного процента и достигшее в настоящее время 21%.[ …]
Происхождение эукариотических клеток (стр. 3 из 4)
Фотосинтезирующие эукариоты (водоросли и зеленые растения ) произошли от фотосинтезирующих прокариот ( сине-зеленых водорослей)
Растения и их фотосинтезируюшшие системы в докембрии эволюционировали монофилетически .
Животные и грибы произошли от автотрофов , утративщих пластиды .
Митохондрии дифференцировались внутриклеточно у фотосинтезирующих праводорослей .
У праводорослей внутриклеточно дифференцировались ундолиподии , митотический аппарат и все прочие эукариотические органеллы .
Все организмы произошли от предков – прокариот путем накопления одиночных мутаций .
СЛЕДСТВИЕ : согласно теории , должны существовать праводоросли ; трудно связать эту теорию с ископаемой летописью и построить соответствующую филогению .
Теория последовательных эндосимбиозов :
Главная дихотомия : прокариоты – эукариоты .
Фотосинтезирующие эукариоты (ядросодержащие водоросли и растения ) и нефотосинтезирующие эукариоты ( животные , грибы , простейшие ) произошли от общих гетеротрофных предков ( амебофлагеллят ).
Эволюция фотосинтеза в архее была монофилетической у бактерий ( включая циано- и хлорооксибактерий ), фотосин-тезирующие протисты возникли внезапно , когда их гетеро-трофные протисты – предки вступили в симбиоз с фото-синтезирующими прокариотами (протопластидами ).
Животные и большинство водорослей и грибов произошли непосредственно от гетеротрофных протистов .
Митохондрии уже были у гетеротрофных протистов ,когда последние в результате симбиоза приобрели пластиды . Промитохондрии были ранее приобретены симбиотическим путем .
Митоз выработался у гетеротрофных эукариотических протистов путем дифференциации ундолиподиальной системы .
Прокариоты произошли от ранних прокариотических предков путем ряда одиночных мутационных событий , эукариоты тоже произошли от эукариотических предков благодаря одиночным мутационным событиям , эукариотические же предки произошли от прокариот в результате серии симбиозов .
Следствия : теория объясняет биохимические пути и их избыточность у органелл . Она позволяет построить последо-вательную филогению , объясняющую “биологический разрыв “ между временами протерозоя и фанерозоя . Теория предска-зывает , что у органелл должны сохраняться остатки систем репликации .
Ботанический миф
Со времен Жюсье низшие растения называют таллофитами .а высшие — бриофитами и трахеофитами . Согласно ботаническому мифу ,общие предки фотосинтезирующих бактерий и сине-зеленых водорослей путем прямой филиации дали начало эукариотическим водорослям в том числе морским . Группы организмов , филогению которых в наибольшшей мере затра-гивает концепция прямой филиации , оказались в области ботаники — это бактерии , водоросли , грибы и растения .
Группы организмов , филогению которых в наибольшшей мере затра-гивает концепция прямой филиации , оказались в области ботаники — это бактерии , водоросли , грибы и растения .
Гипотетические организмы , которые могли бы быть связующим звеном между цианобактериями и зелеными водорослями , были названы родоначальными фито-монадами ( ancestral phytomonads ) или праводорослями . Некоторые авторы утверждали , что подходящими кандитами на роль праводорослей могут быть эукариоты Cyanidium и Cyanophora, сходные по своим пигментным системам с цианобактериями . У таллофитов не было обнаружено промежуточных форм между немитотическим состоянием цианобактерий , и вполне развитым типичным митозом зеленых водорослей и растений . Хотя грибы и цветковые растения полностью лишены ундулиподий , им свойственны митоз и мейоз , для которых необходимы веретена , состоящие из микротрубочек . Имеется общее согласие в том ,что все многоклеточные организмы ,обладающие микротрубочками ,происходят от эукариотических микроорганизмов с ундолиподиями , но никто не знает , от какой именно группы протистов . Подкреплением ботанического мифа служили древность и сложность фотосинтеза , а токже тот факт , что у многих организмов ( у бактерий , евгленовых , пара-зититических растений ) как в природе , так и в лаборатории , наблюдалась утрата фотосинтеза в результате мутаций . Поскольку фотосинтез — анаэробный процесс ,обеспечивающий в конечном итоге существование всех организмов , то он должен был развиться очень рано в истории жизни на нашей планете , еще до того как возник митоз , для которого небходим кис-лород . в фотосинтезе участвует множество пигментов , липидов и ферментов ,упорядоченно расположенных в очень сложных мембранах . При этом детали фотосинтетических механизмов у водорослей и растений , с одной стороны , и у цианобактерий , с другой — поразительно сходны . Все эти организмы освобождают газообразный кислород из воды , которая служит источником атомов водорода , используемых для восстановления СО2 до органических веществ клетки . Многие водоросли и простейщие не имеют ни митоза , ни генетической системы , сходной с таковой животных и растений .
Подкреплением ботанического мифа служили древность и сложность фотосинтеза , а токже тот факт , что у многих организмов ( у бактерий , евгленовых , пара-зититических растений ) как в природе , так и в лаборатории , наблюдалась утрата фотосинтеза в результате мутаций . Поскольку фотосинтез — анаэробный процесс ,обеспечивающий в конечном итоге существование всех организмов , то он должен был развиться очень рано в истории жизни на нашей планете , еще до того как возник митоз , для которого небходим кис-лород . в фотосинтезе участвует множество пигментов , липидов и ферментов ,упорядоченно расположенных в очень сложных мембранах . При этом детали фотосинтетических механизмов у водорослей и растений , с одной стороны , и у цианобактерий , с другой — поразительно сходны . Все эти организмы освобождают газообразный кислород из воды , которая служит источником атомов водорода , используемых для восстановления СО2 до органических веществ клетки . Многие водоросли и простейщие не имеют ни митоза , ни генетической системы , сходной с таковой животных и растений .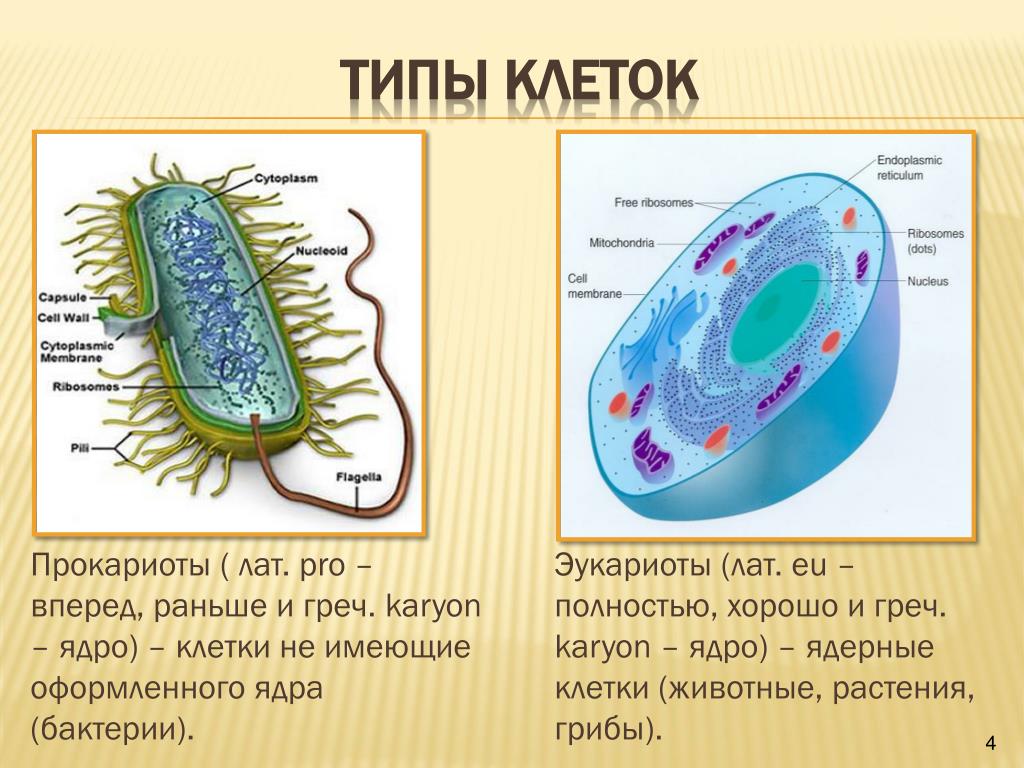 Инфузории имеют особую гене-тическую систему с ядрами 2 типов , причем клетки нередко многоядерные . Один тип яядер – макронуклеусы , а другой – намного меньшие микронуклеусы . Микронуклеусы физиологи-чески несущественны . однако они хранят копии генов , а при половом процессе — претерпевают мейоз . Физиологически необходимые макронуклеусы служат местом транскрипции РНК, и , таким образом , управляют физиологией клетки . Одна из трудностей при выяснении отношений между цианобактериями и эукариотическими водорослями связана с отсутствием данных о происхождении ундолиподий : не было найдено никаких промежуточных звеньев между цианобактериями , у которых ни-когда не бывает ундолиподий , и множеством водорослей , которые их имеют . Другая проблема – полное отсутствие полового процесса у цианобактерий : между бесполыми цианобактериями и водорослями , большинству которых свойственен половой процесс , не оказалось никаких промежуточных форм . Обнаружилось , что большинство водо-рослей имеют ундолиподии на какой – либо стадии жизненного цикла .
Инфузории имеют особую гене-тическую систему с ядрами 2 типов , причем клетки нередко многоядерные . Один тип яядер – макронуклеусы , а другой – намного меньшие микронуклеусы . Микронуклеусы физиологи-чески несущественны . однако они хранят копии генов , а при половом процессе — претерпевают мейоз . Физиологически необходимые макронуклеусы служат местом транскрипции РНК, и , таким образом , управляют физиологией клетки . Одна из трудностей при выяснении отношений между цианобактериями и эукариотическими водорослями связана с отсутствием данных о происхождении ундолиподий : не было найдено никаких промежуточных звеньев между цианобактериями , у которых ни-когда не бывает ундолиподий , и множеством водорослей , которые их имеют . Другая проблема – полное отсутствие полового процесса у цианобактерий : между бесполыми цианобактериями и водорослями , большинству которых свойственен половой процесс , не оказалось никаких промежуточных форм . Обнаружилось , что большинство водо-рослей имеют ундолиподии на какой – либо стадии жизненного цикла .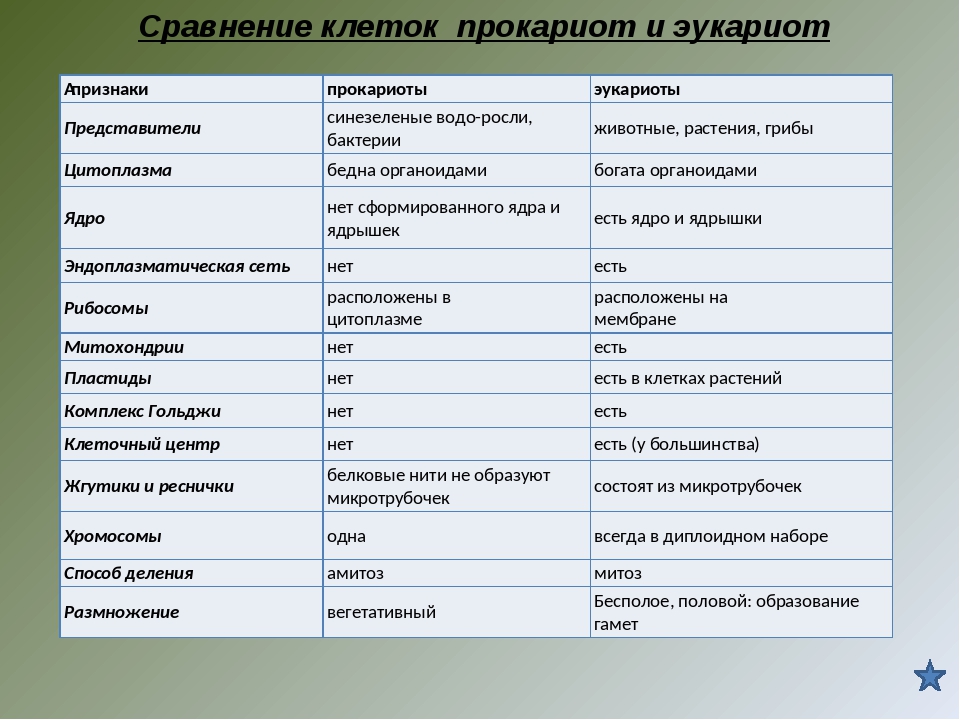 Подвижные структуры у водорослей , простейших , жи-вотных и растений оказались одинаковыми : все они состоят из микротрубочек диаметром 0,024 мкм . Выяснилось ,что мито-тическое веретено даже у грибов , у которых нет ундолипдий ,состоит из белковых микротрубочек того же диаметра , что и в ундолиподиях . На протяжении полувека казалось чрезвычайно правдоподобным , что эукариотичкские водоросли произошли от цианобактерий . Этот ботанический миф привел к предположе-ниям , которые многие годы служили как стимулом для исследования , так и системой координат ,в которой про-изводилась оценка получаемых результатов . Утверждение , что связующие звенья между цианобактериями и эукариотическими водорослями вымерли , не оставив ископаемых следов и живых реликтов , становилось все менее убедительным по мере поступ-ления новых данных о живых микробах и об окаменелостях . Главные промежуточные звенья между фотосинтезирующими прокариотами и эукариотами не вымерли бесследно , как того требует ботанический миф ,- они никогда не существовали .
Подвижные структуры у водорослей , простейших , жи-вотных и растений оказались одинаковыми : все они состоят из микротрубочек диаметром 0,024 мкм . Выяснилось ,что мито-тическое веретено даже у грибов , у которых нет ундолипдий ,состоит из белковых микротрубочек того же диаметра , что и в ундолиподиях . На протяжении полувека казалось чрезвычайно правдоподобным , что эукариотичкские водоросли произошли от цианобактерий . Этот ботанический миф привел к предположе-ниям , которые многие годы служили как стимулом для исследования , так и системой координат ,в которой про-изводилась оценка получаемых результатов . Утверждение , что связующие звенья между цианобактериями и эукариотическими водорослями вымерли , не оставив ископаемых следов и живых реликтов , становилось все менее убедительным по мере поступ-ления новых данных о живых микробах и об окаменелостях . Главные промежуточные звенья между фотосинтезирующими прокариотами и эукариотами не вымерли бесследно , как того требует ботанический миф ,- они никогда не существовали . Согласно теории последовательных эндосимбиозов , циано-бактерии – действительно родоначальники ,но только пластид , а не остальных частей эукариотической клетки , и уж во всяком случае не ее ядра . И все же в вопросе о происхождении ядра гипотеза прямой филиации и теория симбиоза совместимы . Мыслимые модификации крайнего варианта теории симбиоза включают , например ,идею о симбиотическом происхождении пластид , но не митохондрий , или о симбиотическом про-исхождении пластид и митохондрий , но не ундолиподий . Сторонники прямой филиации выдвинули также предположение о происхождении ядра путем симбиоза .
Согласно теории последовательных эндосимбиозов , циано-бактерии – действительно родоначальники ,но только пластид , а не остальных частей эукариотической клетки , и уж во всяком случае не ее ядра . И все же в вопросе о происхождении ядра гипотеза прямой филиации и теория симбиоза совместимы . Мыслимые модификации крайнего варианта теории симбиоза включают , например ,идею о симбиотическом происхождении пластид , но не митохондрий , или о симбиотическом про-исхождении пластид и митохондрий , но не ундолиподий . Сторонники прямой филиации выдвинули также предположение о происхождении ядра путем симбиоза .
Решающие доказательства эндосимбиотического происхождения хлоропластов и митохондрий .
Наиболее существенные доказательства происхождения важней-ших органелл эукариотических клеток — хлоропластов и мито-хондрий из прокариот получены в последние годы при изуче-нии структуры рибосомальных РНК . Молекудярный биолог
К. Воз показал уже в 80-х годах , что структура РНК из рибосом хлоропластов растений почти идентична структуре РНК из некоторых цианобактерий . В то же время группа исследователей установила , что РНК рибосом , полученных из митохондрий разных эукариотов схожа с РНК некоторых бактерий , в частности бактерий рода Paracoccus . Набор дыхательных ферментов указанной бактерии очень похож на
В то же время группа исследователей установила , что РНК рибосом , полученных из митохондрий разных эукариотов схожа с РНК некоторых бактерий , в частности бактерий рода Paracoccus . Набор дыхательных ферментов указанной бактерии очень похож на
“ дыхательный ансамбль “ митохондрий животных .
Эти данные достаточно ясно свидетельствуют в пользу большей вероятности эндосимбиотического происхождения эукариотических клеток . Явление эндосимбиоза и по сей день чрезвычайно широко распространено в природе . Известно , что на теле некоторых морских рыб часто поселяются светящиеся бактерии , которые не только сами получают большие преимущества для своей жизни , но и очень полезны для организмов хозяев . Все сказанное убедительно свидетельствует в пользу эндосимбиотического происхождения эукариотических клеток из гораздо меньщих по объему клеток прокариот . Однако главный вопрос заключается в установлении природы хозяйской клетки , ведь клетки эукариот обладают хорощо сформированным ядром , имеющим оболочку , а прокариоты такого ядра не имеют . Л. Маргелис , например , считает ,что роль хозяйских клеток , возможно , сыграли прокариоты – предшественники современных бактерий , относящиеся к микоплазмам — очень примитивно построенным прокариотам , практически лишенным клеточных стенок . В последнее время появились и широко обсуждаются работы японского ученого
Л. Маргелис , например , считает ,что роль хозяйских клеток , возможно , сыграли прокариоты – предшественники современных бактерий , относящиеся к микоплазмам — очень примитивно построенным прокариотам , практически лишенным клеточных стенок . В последнее время появились и широко обсуждаются работы японского ученого
Т. Ошимы . На основании тщательного изучения структуры рибосомных РНК многих про- и эукариот , в том числе и многих представителей до сих пор малоизученного надцарства архей , этот автор приходит к выводу , что хозяйской клеткой являлся один из предков современных архей . Следует сказать , что у представителей именно этого надцарства , в отличие от бактерий , имеется очень много общих с эукариотами биохи-мических и молекулярно – биологических особенностей . В по-следних работах Т. Ошима приводятся доводы в пользу того , что хозяйской клеткой при возникновении эукариот являлся предок архей , подобный одному из современных представителей этих прокариот — термоплазме . Каково будет окончательное решение проблемы установления природы хозяйской клетки , давшей начало эукариотической клетке в процессе множествен-ного эндосимбиоза , покажут дальнейшие исследования .
Уроки биологии
Уроки биологии
Многообразие органического мира
Классификация организмов
На Земле около 1,5 млн видов животных, более 0,5 млн видов растений, сотни тысяч видов грибов и множество микроорганизмов. Их классификацией — объединением в группы по сходству строения и жизнедеятельности — занимается биологическая дисциплина, зародившаяся в XVIII в. и называемая систематикой. Для ученых XVIII—XIX веков поиски системы в природе являлись прежде всего попыткой увидеть закономерности в плане Творца.
В настоящее время ученые выделяют две формы жизни.
Неклеточные формы жизни — вирусы.
Клеточные формы жизни включают:
Надцарство (или Империя) Прокариоты:
Царство Архебактерии (Архей),
Царство Истинные бактерии (Эубактерии),
Надцарство (или Империя) Эукариоты:
Царство Протисты,
Царство Животные,
Царство Растения,
Царство Грибы.
Надцарства Прокариоты и Эукариоты. Важнейший признак клетки — наличие или отсутствие отделенного от цито-плазмы ядра. Среди клеточных существ различают безъядерные (прокариоты) и ядерные (эукариоты). К прокариотам относят Истинные бактерии и Архебактерии, существенно различающиеся строением мембран и оболочки.
Цианобактерии (греч. kyanos синий) относят к истинным бактериям. Они обладают признаками бактерий (прочные клеточные стенки, отсутствие отделенного от цитоплазмы ядра) и растений (осуществляют фотосинтез, выделяя кислород), поэтому цианобактерии иногда называют синезелеными водорослями. Они живут в виде отдельных клеток или объединяются в колонии с образованием нитей. Наличие различных пигментов придает им различную окраску от сине-зеленой, фиолетовой и красной до почти черной. Некоторые цианобактерии (например, спирулину) люди употребляют в пищу.
Разделение эукариот на царства представляет для современных ученых определенные трудности, в особенности при классификации низших (чаще одноклеточных) эукариот. Современная систематика все чаще ориентируется не только на морфофизиологические, цитологические, биохимические признаки организмов, но и на строение ДНК, РНК, отдельных генов.
Современная систематика все чаще ориентируется не только на морфофизиологические, цитологические, биохимические признаки организмов, но и на строение ДНК, РНК, отдельных генов.
Царство Протисты объединяет простейших (одноклеточных), одноклеточные водоросли и низшие грибы (стенки их клеток, как и у растений, построены из целлюлозы). Эти организмы, согласно гипотезе эволюции, — предки остальных трех царств эукариот.
В ряде классификаций все водоросли относят к царству Растения; в других классификациях царство Протисты вовсе отсутствует: низшие грибы относят к царству Грибы, одноклеточные водоросли — к царству Растения, а простейших — к царству Животные. В некоторых классификациях надцарство Эукариоты подразделяют более чем на 26 царств. Систематика является сегодня одним из самых спорных разделов биологии. Среди главных причин разногласий ученые называют невыясненность основных механизмов эволюции, положенных в основу классификации.
Водоросли, по современным представлениям, являются сборной группой фототрофных организмов, ведущих преимущественно водный образ жизни. Приспособленностью к свету объясняется различная окраска водорослей. Спектральные компоненты солнечного света пронизывают воду на разную глубину. Красные лучи проникают лишь в верхние слои, а синие — значительно глубже. Для функционирования хлорофилла необходим красный свет, поэтому зеленые водоросли встречаются обычно на глубинах в несколько метров. Наличие пигмента, осуществляющего фотосинтез при желто-зеленом свете, позволяет бурым водорослям жить на глубинах до 200 метров. Пигмент красных водорослей использует зеленый и синий свет, поэтому красные водоросли населяют глубины до 268 метров. Из красных водорослей добывают полисахарид агар, используемый для изготовления мармелада, зефира и пастилы.
Приспособленностью к свету объясняется различная окраска водорослей. Спектральные компоненты солнечного света пронизывают воду на разную глубину. Красные лучи проникают лишь в верхние слои, а синие — значительно глубже. Для функционирования хлорофилла необходим красный свет, поэтому зеленые водоросли встречаются обычно на глубинах в несколько метров. Наличие пигмента, осуществляющего фотосинтез при желто-зеленом свете, позволяет бурым водорослям жить на глубинах до 200 метров. Пигмент красных водорослей использует зеленый и синий свет, поэтому красные водоросли населяют глубины до 268 метров. Из красных водорослей добывают полисахарид агар, используемый для изготовления мармелада, зефира и пастилы.
Царство Грибы объединяет гетеротрофные организмы, тело которых состоит из ветвящихся нитей (гиф), в совокупности образующих мицелий (грибницу). Грибы всасывают необходимые питательные вещества из окружающей среды (осмотрофное питание). Выделяя высокоактивные ферменты, грибы расщепляют (деполимеризуют) белки, нуклеиновые кислоты, целлюлозу, а затем всасывают образовавшиеся мономеры (аминокислоты, нуклеотиды, моносахариды). Царство грибов очень разнообразно, в него входят плесневые грибы и дрожжи, шляпочные грибы и трутовики.
Царство грибов очень разнообразно, в него входят плесневые грибы и дрожжи, шляпочные грибы и трутовики.
Грибы обладают признаками животных и растений. Они не содержат хлорофилла и не способны к фотосинтезу, а подобно животным питаются готовыми органическими веществами. Конечным продуктом метаболизма азота, как и у животных, является мочевина. Запасное питательное вещество, как и у животных, — полисахарид гликоген (в растениях запасающую функцию выполняет обычно крахмал). Прочность клеточным стенкам большинства грибов, так же как покрову жуков и панцирю крабов, придает хитин (клеточные стенки низших грибов, относимых к протистам, состоят из целлюлозы). Прикрепленный образ жизни, неограниченный рост и способы размножения (в основном, спорами и грибницей, но для ряда грибов возможно половое размножение) сближают грибы с растениями. Любопытный симбиоз представляют собой лишайники. Их тело состоит из грибницы, в которой живут цианобактерии и водоросли.
Царство Растения объединяет фотосинтезирующие организмы, выделяющие кислород и запасающие крахмал, имеющие плотные клеточные стенки (обычно из целлюлозы).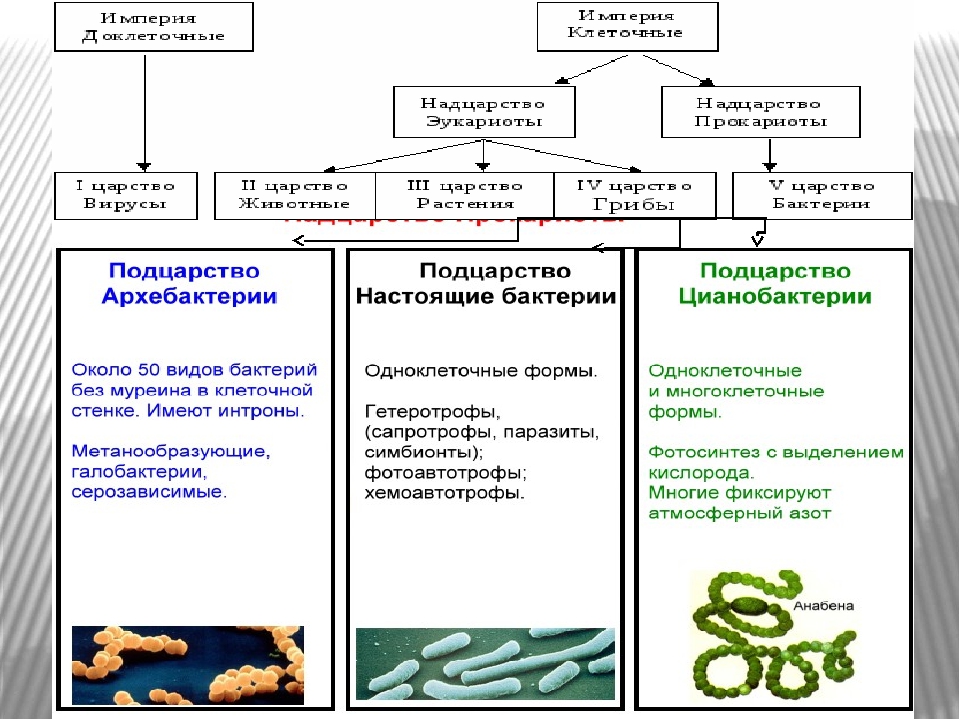 В царство растений входят 7 отделов: многоклеточные водоросли, мхи, плауны, хвощи, папоротники, голосеменные и покрытосеменные растения. Классификация проведена по ряду сходных признаков. Например, отдел покрытосеменных выделен по признаку наличия цветка и защищенного плодом семени. Высшие растения характеризуются разделением тела на органы (корень, стебель, лист). Тело низших растений не имеет разделения на органы, к ним относят, например, некоторые мхи.
В царство растений входят 7 отделов: многоклеточные водоросли, мхи, плауны, хвощи, папоротники, голосеменные и покрытосеменные растения. Классификация проведена по ряду сходных признаков. Например, отдел покрытосеменных выделен по признаку наличия цветка и защищенного плодом семени. Высшие растения характеризуются разделением тела на органы (корень, стебель, лист). Тело низших растений не имеет разделения на органы, к ним относят, например, некоторые мхи.
Царство Животные
Существенным их отличием является подвижный образ жизни. Но этот критерий не абсолютный. Так, коралловые полипы — неподвижные животные, а эвглена и вольвокс — подвижные водоросли. В связи с необходимостью движения большинство клеток животных не имеет плотной наружной оболочки, их основное запасающее вещество — легкорастворимый гликоген, а не крахмал.
Царство животных объединяет беспозвоночных и хордовых. Подцарство многоклеточных беспозвоночных включает 6 типов: кишечнополостные; плоские, круглые и кольчатые черви; моллюски и членистоногие. Тип членистоногих объединяет 3 класса: ракообразные, паукообразные и насекомые — эти существа имеют сегментарные конечности. Тип хордовых состоит из 3 подтипов: оболочники, бесчерепные и черепные (позвоночные). В подтип позвоночных входит 7 классов: круглоротые, надкласс рыб (классы хрящевых и костных), земноводные, пресмыкающиеся, птицы и млекопитающие. Классы последовательно подразделяются на отряды, семейства, роды и виды.
Тип членистоногих объединяет 3 класса: ракообразные, паукообразные и насекомые — эти существа имеют сегментарные конечности. Тип хордовых состоит из 3 подтипов: оболочники, бесчерепные и черепные (позвоночные). В подтип позвоночных входит 7 классов: круглоротые, надкласс рыб (классы хрящевых и костных), земноводные, пресмыкающиеся, птицы и млекопитающие. Классы последовательно подразделяются на отряды, семейства, роды и виды.
Основы современной классификации заложил выдающийся шведский натуралист Карл Линней. В своей работе «Система природы» (1775 г.) он описал около 8 000 видов растений, 4 000 видов животных. Линней определял вид как совокупность особей, сходных между собой по строению и дающих при скрещивании плодовитое потомство. Подчеркивая неизменность исходных сотворенных видов, Линней указывал: «Видов столько, сколько различных форм создал в начале мира Всемогущий». Группы похожих видов, по Линнею, входят в один предковый род: «И сказал Бог: да произведет земля душу живую по роду ее, скотов, и гадов, и зверей земных по роду их» (Быт. 1,24). Классификационное понятие рода при этом совпадает с библейским.
1,24). Классификационное понятие рода при этом совпадает с библейским.
В основу систематики ученый положил принцип иерархичности таксонов (единиц классификации организмов): сходные виды он объединил в роды, роды — в отряды, а отряды — в классы. В наименовании организмов Линней установил бинарную (двойную) номенклатуру: название каждого существа стало состоять из двух латинских слов, первое писалось с большой буквы и указывало родовую принадлежность (существительное), второе — видовую (прилагательное). Например, латинское название степного сурка байбака — Marmota bybak. В разных местностях этот вид сурков именуют по-разному: свистун, сугур и т. д. Единая номенклатура существенно облегчила взаимопонимание ученых разных стран, заменив прежние многословные описания видов. В дальнейшем система была дополнена категориями семейства, подкласса, подтипа, типа. Так, кошка домашняя (Felis domestica) входит в род мелких кошек семейства кошачьих отряда хищных класса млекопитающих подтипа позвоночных типа хордовых.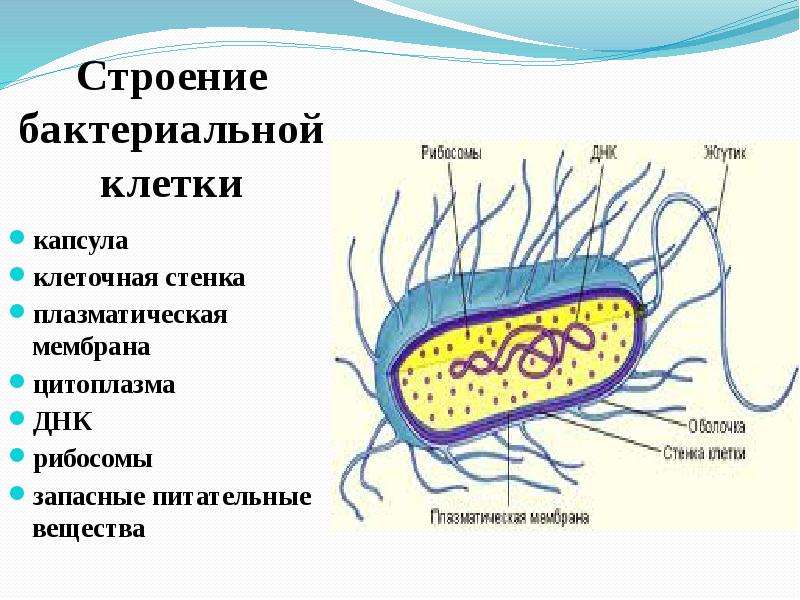 Кроме домашней кошки род мелких кошек включает амурского лесного кота, камышового кота, рысь. Подход Линнея революционизировал биологию, открыв новые возможности систематизации растительного и животного мира.
Кроме домашней кошки род мелких кошек включает амурского лесного кота, камышового кота, рысь. Подход Линнея революционизировал биологию, открыв новые возможности систематизации растительного и животного мира.
В системе Линнея виды образуют восходящую последовательность организмов от низших к высшим, но они не связаны эволюционным родством. Возможность систематизации организмов рассматривалась Карлом Линнеем как следствие существования единого плана сотворения мира. По убеждению Линнея, наличие анатомических сходств у разных видов свидетельствует о том, что Конструктор использовал типовые конструкции.
Вопросы учителя
1. Назовите формы жизни и надцарства организмов.
2. Охарактеризуйте четыре царства эукариот.
3. Какой принцип лежит в основе классификации организмов?
4. Чем объяснял Линней возможность классификации организмов? С какой классификационной единицей Линней отождествил библейское понятие рода?
© Все права защищены http://www.
 portal-slovo.ru
portal-slovo.ruГеномы одноклеточных водорослей проливают свет на эволюцию фотосинтезирующих эукариот
Отсеквенированы ядерные геномы представителей двух групп одноклеточных водорослей, появившихся в результате вторичного симбиоза. Фотосинтезирующие симбионты этих организмов — одноклеточные красные и зеленые водоросли — сохранили не только свои пластиды, служащие для фотосинтеза, но и ядро с рудиментарным геномом («нуклеоморф»), собственный аппарат синтеза белка и остатки цитоплазмы. Выявлена сложная система взаимодействий между четырьмя геномами, сосуществующими в одной клетке, — ядерным, митохондриальным, пластидным и нуклеоморфным. Сохранение нуклеоморфа у двух изученных видов, возможно, связано с тем, что фотосинтезирующий симбионт у них присутствует в клетке в единственном экземпляре, что затрудняет перенос генов из нуклеоморфа в ядро.
Первые эукариоты были гетеротрофными организмами: они питались готовой органикой. В дальнейшем представители разных эволюционных линий эукариот много раз независимо приобретали способность к фотосинтезу.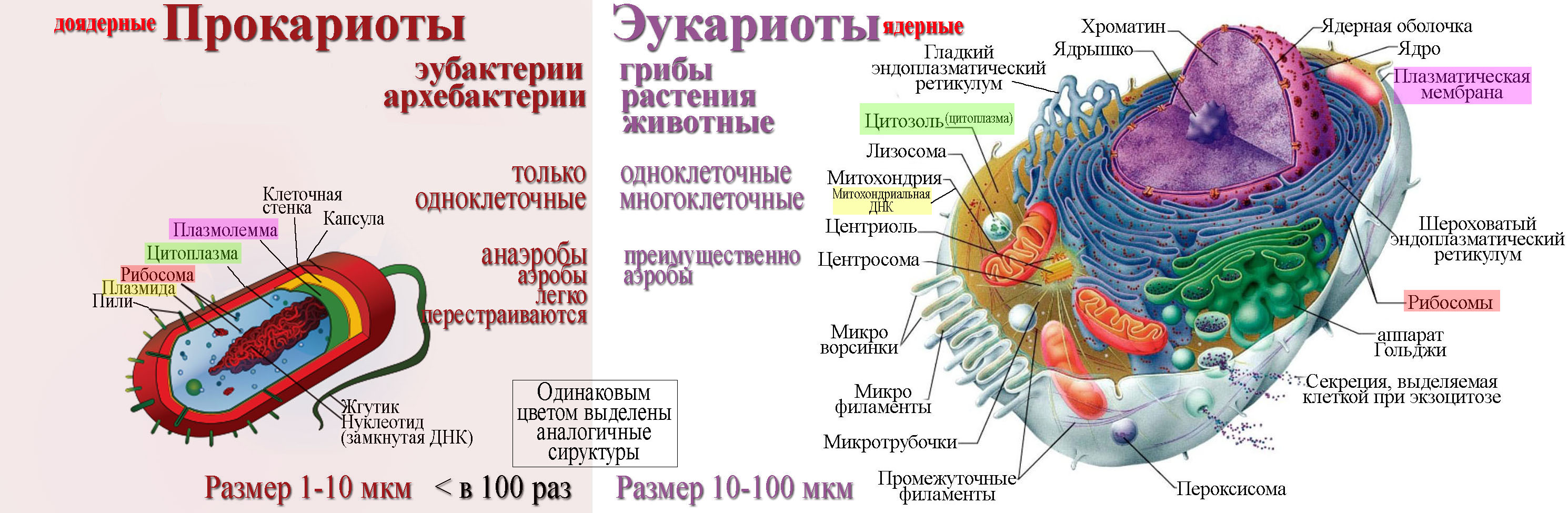 Происходило это всегда одним и тем же способом: путем эндосимбиоза, то есть приобретения фотосинтезирующих внутриклеточных симбионтов.
Происходило это всегда одним и тем же способом: путем эндосимбиоза, то есть приобретения фотосинтезирующих внутриклеточных симбионтов.
Первые фотосинтезирующие эукариоты (одноклеточные водоросли) появились свыше 1,2 миллиарда лет назад в результате «первичного» эндосимбиоза с цианобактерией, которая дала начало первичным пластидам, окруженным двойной мембраной. Потомки этих эукариот дали начало глаукофитовым, красным и зеленым водорослям, а от последних произошли наземные растения. Помимо этой «главной» эволюционной линии фотосинтезирующих эукариот — обладателей первичных пластид, существует много групп водорослей со вторичными пластидами, окруженными четырьмя мембранами. Они произошли от разных гетеротрофных эукариот, вступивших во вторичный симбиоз с одноклеточными красными или зелеными водорослями.
Вторичные пластиды, происходящие от симбиотических красных водорослей, характерны для гетероконт, к которым относятся бурые, диатомовые, золотистые и желто-зеленые водоросли, а также для динофлагеллят, гаптофитов и криптофитов (см.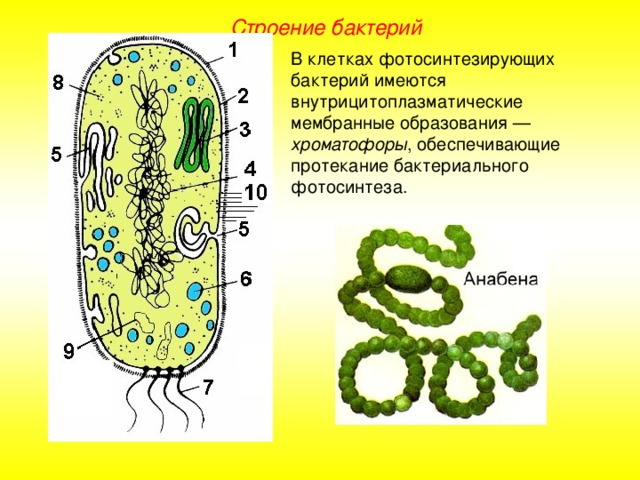 также: Чужие гены помогли диатомовым водорослям добиться успеха, «Элементы», 17.10.2008). Вторичные пластиды эвгленовых и хлорарахниофитовых (см. Chlorarachniophyte) водорослей происходят от симбиотических зеленых водорослей.
также: Чужие гены помогли диатомовым водорослям добиться успеха, «Элементы», 17.10.2008). Вторичные пластиды эвгленовых и хлорарахниофитовых (см. Chlorarachniophyte) водорослей происходят от симбиотических зеленых водорослей.
У большинства перечисленных групп от вторичного симбионта ничего не осталось, кроме хлоропласта, окруженного четырьмя мембранами (две внутренние унаследованы от первичного хлоропласта, две внешние — это бывшая оболочка симбионта и мембрана вакуоли, в которую он был заключен хозяйской клеткой). Цитоплазма симбионта, его рибосомы, ядро и всё прочее благополучно редуцировалось, а жизненно необходимые гены «переселились» в ядерный геном клетки-хозяина.
Особый интерес представляют те немногочисленные фотосинтезирующие эукариоты, у которых вторичный симбионт сохранил, помимо хлоропласта, также остатки цитоплазмы и маленькое ядро («нуклеоморф») с собственным геномом. У таких организмов в каждой клетке целых четыре генома: ядерный, митохондриальный, нуклеоморфный и пластидный. У их предков, очевидно, был еще и пятый, ныне утраченный — геном митохондрий симбионта. Кроме того, у них четыре разных аппарата синтеза белка, в том числе рибосомы четырех сортов: бактериальные в пластидах и митохондриях, эукариотические — в цитоплазме хозяина и в остаточной цитоплазме симбионта.
У их предков, очевидно, был еще и пятый, ныне утраченный — геном митохондрий симбионта. Кроме того, у них четыре разных аппарата синтеза белка, в том числе рибосомы четырех сортов: бактериальные в пластидах и митохондриях, эукариотические — в цитоплазме хозяина и в остаточной цитоплазме симбионта.
Существование таких промежуточных форм, у которых редукция симбионта не дошла до своего логического конца, — наглядное подтверждение теории вторичного эндосимбиоза. Ученые надеются, что их изучение прольет свет на формирование эндосимбиотических систем. Кроме того, чрезвычайно интересно узнать, как клетка координирует работу четырех геномов.
Большая международная команда исследователей отсеквенировала ядерные геномы двух таких водорослей: Guillardia theta (из группы криптофитов, вторичный симбионт — красная водоросль) и Bigelowiella natans (из группы хлорарахниофитов, вторичный симбионт — зеленая водоросль). Остальные три генома обоих видов тоже известны: некоторые были отсеквенированы ранее, а с недостающими авторы разобрались в рамках данного исследования.
По сравнению с геномами других эукариот с вторичными пластидами (диатомовых и бурых водорослей) геномы G. theta и B. natans оказались большими (87 и 95 млн пар нуклеотидов) и содержащими много белок-кодирующих генов (24 840 и 21 708 генов было предсказано на основе анализа геномной ДНК). Для 85% предсказанных генов анализ РНК (см. RNA-seq) подтвердил наличие транскрипции, то есть это действительно гены, и они работают. Многие белки (51% и 47%) оказались уникальными (не имеющими очевидных гомологов среди белков других организмов), что свидетельствует о быстрой молекулярной эволюции.
У B. natans выявлен рекордный для одноклеточных (и даже для всех изученных организмов, кроме животных) уровень альтернативного сплайсинга (см.: Почти все человеческие гены кодируют более одного белка, «Элементы», 08.11.2008). По-видимому, в большинстве случаев это не регулируемый альтернативный сплайсинг, а «случайный шум», порождаемый неточной работой сплайсосом. Однако для 246 генов можно предположить, что альтернативные изоформы белка транспортируются в разные отделы (компартменты) клетки, потому что в зависимости от попадания в зрелую мРНК тех или иных экзонов в состав белковой молекулы входят (или не входят) сигнальные пептиды, от которых зависит транспорт белков в органеллы.
Однако для 246 генов можно предположить, что альтернативные изоформы белка транспортируются в разные отделы (компартменты) клетки, потому что в зависимости от попадания в зрелую мРНК тех или иных экзонов в состав белковой молекулы входят (или не входят) сигнальные пептиды, от которых зависит транспорт белков в органеллы.
Геномы нуклеоморфов у G. theta и B. natans невелики. Они содержат небольшой набор «генов домашнего хозяйства», около 30 генов белков, предназначенных для транспортировки в пластиды, а также ряд генов неопознаваемых белков (ORFans), непохожих по своим аминокислотным последовательностям ни на какие известные белки. Геномы нуклеоморфов подверглись слишком сильной редукции, чтобы обеспечивать существование фотосинтезирующего симбионта. Поэтому жизнь нуклеоморфа, пластиды и остатков цитоплазмы симбионта («перипластидиального компартмента») поддерживается за счет белков, закодированных в ядерном геноме.
Авторы установили, что в ядерном геноме G. theta закодировано 2400 белков, предназначенных для транспортировки в перипластидиальный компартмент или нуклеоморф; у B. natans — 1002. В основном это белки, функции которых связаны с посттрансляционными модификациями других белков, с передачей сигналов, с метаболизмом и транспортом углеводов. По-видимому, репликация ДНК и транскрипция генов нуклеоморфа контролируются в основном белками, закодированными в ядерном геноме. Синтез белков (трансляция) в остаточной цитоплазме симбионта, наоборот, осуществляется «своими силами», то есть при помощи белков, закодированных в геноме нуклеоморфа.
theta закодировано 2400 белков, предназначенных для транспортировки в перипластидиальный компартмент или нуклеоморф; у B. natans — 1002. В основном это белки, функции которых связаны с посттрансляционными модификациями других белков, с передачей сигналов, с метаболизмом и транспортом углеводов. По-видимому, репликация ДНК и транскрипция генов нуклеоморфа контролируются в основном белками, закодированными в ядерном геноме. Синтез белков (трансляция) в остаточной цитоплазме симбионта, наоборот, осуществляется «своими силами», то есть при помощи белков, закодированных в геноме нуклеоморфа.
В системе взаимодействий между частями сложной клетки у двух изученных видов, по-видимому, больше сходств, чем различий. Это свидетельствует о параллельной эволюции и в какой-то мере — о закономерном характере эволюционных изменений, следующих за приобретением вторичного симбионта. К числу существенных различий относится наличие у B. natans большого количества (852) интронов в генах нуклеоморфа. Для вырезания этих интронов в нуклеоморфе присутствуют сплайсосомы, сделанные из белков, закодированных в ядерном геноме. У G. theta, по-видимому, в нуклеоморфе нет полноценных сплайсосом, а интронов в геноме нуклеоморфа всего 17. Зато у G. theta в цитоплазме симбионта есть свои протеасомы (см.: Белки попадают в протеасому через «преддверие» уже развернутыми, «Элементы», 05.11.2010), опять-таки собирающиеся из белков, закодированных в ядерном геноме. У симбионта B. natans протеасом, судя по всему, нет.
Для вырезания этих интронов в нуклеоморфе присутствуют сплайсосомы, сделанные из белков, закодированных в ядерном геноме. У G. theta, по-видимому, в нуклеоморфе нет полноценных сплайсосом, а интронов в геноме нуклеоморфа всего 17. Зато у G. theta в цитоплазме симбионта есть свои протеасомы (см.: Белки попадают в протеасому через «преддверие» уже развернутыми, «Элементы», 05.11.2010), опять-таки собирающиеся из белков, закодированных в ядерном геноме. У симбионта B. natans протеасом, судя по всему, нет.
Исследование подтвердило, что на ранних этапах становления симбиоза значительная часть генов из ядра симбионта была перенесена в ядро хозяина. Об этом свидетельствует тот факт, что у обоих видов в ядерном геноме обнаружены сотни генов явно «водорослевого» происхождения, причем у G. theta преобладают гены, характерные для красных водорослей, у B. natans — для зеленых.
Большинство генов симбионта, переселившихся в ядерный геном, обслуживают не симбионта, а хозяина, и лишь небольшая их часть кодирует белки, предназначенные для транспортировки в цитоплазму симбионта или нуклеоморф (см.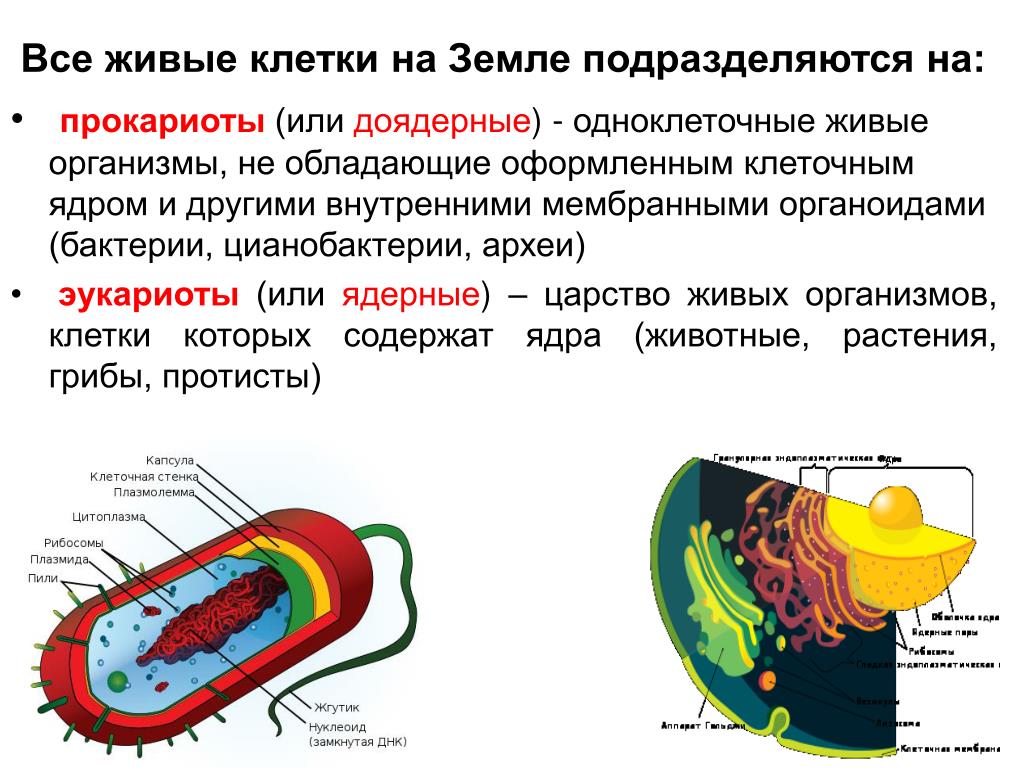 рисунок 3). Остальные ядерные гены, обслуживающие симбионта, — это «хозяйские» по своему происхождению гены, а не водорослевые. Многие гены хозяина подверглись дупликации, после чего одна из копий гена сохранила старую функцию, а другая взяла на себя заботу о симбионте.
рисунок 3). Остальные ядерные гены, обслуживающие симбионта, — это «хозяйские» по своему происхождению гены, а не водорослевые. Многие гены хозяина подверглись дупликации, после чего одна из копий гена сохранила старую функцию, а другая взяла на себя заботу о симбионте.
Почему же у криптофитов и хлорарахниофитов нуклеоморф не редуцировался полностью? Почему от симбионта не осталась только окруженная четырьмя мембранами пластида, как у других вторично-симбиотических водорослей? Чтобы получить ответ на этот вопрос, авторы просканировали ядерные геномы обоих исследованных видов в поисках следов недавнего переноса генетического материала из органелл в ядро.
В обоих геномах обнаружились короткие фрагменты митохондриальной ДНК, недавно инкорпорированные в ядерный геном. Эти фрагменты не содержат целых генов и, по-видимому, не выполняют никаких функций, но они свидетельствуют о том, что эпизодический перенос ДНК из митохондриального генома в ядерный продолжается и поныне. Однако ни одного узнаваемого фрагмента нуклеоморфной или пластидной ДНК ни в одном из двух ядерных геномов обнаружено не было. Это значит, что перенос ДНК из симбионта в ядро в настоящее время уже не происходит (хотя в далеком прошлом он, несомненно, происходил, как видно из наличия множества «водорослевых» генов в ядерном геноме).
Однако ни одного узнаваемого фрагмента нуклеоморфной или пластидной ДНК ни в одном из двух ядерных геномов обнаружено не было. Это значит, что перенос ДНК из симбионта в ядро в настоящее время уже не происходит (хотя в далеком прошлом он, несомненно, происходил, как видно из наличия множества «водорослевых» генов в ядерном геноме).
Что же мешает ДНК симбионта переноситься в ядерный геном? Возможно, одна из причин в том, что фотосинтезирующий симбионт у обоих изученных видов присутствует в клетке в единственном экземпляре. Чтобы ДНК симбионта попала в цитоплазму хозяина и затем в ядро, симбионт, очевидно, должен погибнуть (лизироваться). Но поскольку он всего один, его гибель будет фатальна для симбиотического организма. Митохондрий, напротив, в каждой клетке много, поэтому гибель некоторых из них не смертельна для клетки. Возможно, именно поэтому митохондриальная ДНК часто попадает в ядро, а ДНК симбионта — никогда или очень редко.
Затрудненный перенос ДНК симбионта в ядро в принципе может быть причиной сохранения нуклеоморфов у G. theta и B. natans. Ведь для того, чтобы какой-то жизненно важный ген нуклеоморфа был потерян, в ядерном геноме сначала должен появиться аналогичный ген, белковый продукт которого будет транспортироваться в симбионта (для этого он должен приобрести соответствующий сигнальный пептид). Такой ген может появиться в ядерном геноме двумя способами: в результате переноса ДНК симбионта в ядро и в результате приобретения каким-нибудь исходно ядерным геном соответствующей функции. Первый путь — более легкий (вероятный, быстрый). Если он оказался «запрещен» из-за того, что симбионт в каждой клетке остался только один, это могло замедлить редукцию нуклеоморфа (см. также: Половое размножение препятствует крупномасштабным изменениям генома, «Элементы», 27.03.2007).
theta и B. natans. Ведь для того, чтобы какой-то жизненно важный ген нуклеоморфа был потерян, в ядерном геноме сначала должен появиться аналогичный ген, белковый продукт которого будет транспортироваться в симбионта (для этого он должен приобрести соответствующий сигнальный пептид). Такой ген может появиться в ядерном геноме двумя способами: в результате переноса ДНК симбионта в ядро и в результате приобретения каким-нибудь исходно ядерным геном соответствующей функции. Первый путь — более легкий (вероятный, быстрый). Если он оказался «запрещен» из-за того, что симбионт в каждой клетке остался только один, это могло замедлить редукцию нуклеоморфа (см. также: Половое размножение препятствует крупномасштабным изменениям генома, «Элементы», 27.03.2007).
Впрочем, эти рассуждения выглядят не слишком убедительными, потому что у некоторых других криптофитов и хлорарахниофитов в каждой клетке не по одному, а по несколько фотосинтезирующих симбионтов с нуклеоморфами. По-видимому, придется отсеквенировать и их геномы тоже, чтобы окончательно разобраться в этой проблеме.
Источник: Bruce A. Curtis, Goro Tanifuji, Fabien Burki et al. Algal genomes reveal evolutionary mosaicism and the fate of nucleomorphs // Nature. V. 492. P. 59–65. Статья находится в открытом доступе.
См. также:
Чужие гены помогли диатомовым водорослям добиться успеха, «Элементы», 17.10.2008.
Александр Марков
Тема Возникновение жизни на Земле Задание 1 «Происхождение жизни на Земле»
Тема: Возникновение жизни на Земле
Задание 1. «Происхождение жизни на Земле»
Запишите номера предложений и допишите их:
1. Согласно теории креационизма жизнь на Земле появилась (_).
2. Сущность метафизического мировоззрения в том, что (_).
3. Сторонники гипотезы абиогенеза утверждали: (_).
4. Сторонники гипотезы биогенеза считали, что (_).
5. Рыбы могут зарождаться из ила, черви из почвы, мыши из грязи, мухи из мяса утверждали сторонники теории (_).
6. Франческо Реди нанес первый удар по этой теории, доказав (_).
7. Луи Пастер сумел доказать (_).
8. Теория панспермии утверждает, что живые организмы появились на Земле (_).
9. Теория биохимической эволюции А.И.Опарина предполагает (_).
Задание 2. «Колбы Спалланцани и Л.Пастера»
Какой опыт провел Спалланцани для доказательства невозможности самозарождения микроорганизмов? С чем не согласились виталисты?
Чем опыт Пастера отличался от опыта Спалланцани?
Задание 3. «Аппарат С. Миллера»
Какие газы входили в состав первичной атмосферы Земли?
Что произошло на первом этапе возникновения жизни на Земле по А.И.Опарину?
Какие органические мономеры были получены в опытах С.Миллера?
Какие биополимеры смогли получить в экспериментах?
Можно ли считать коацерваты первыми живыми существами?
Задание 4.
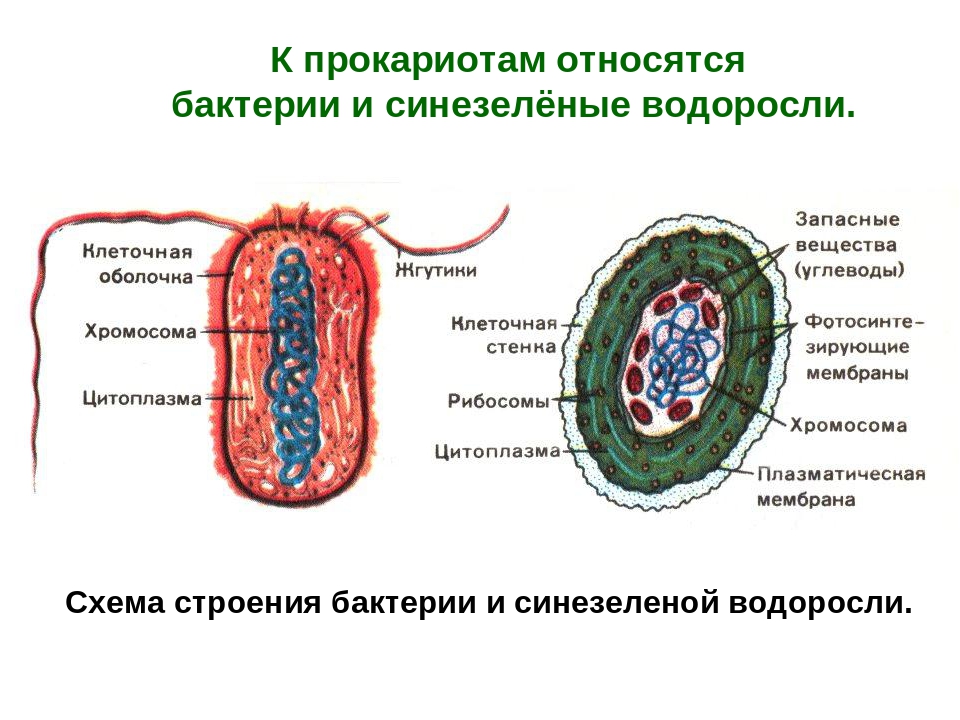 «Основные этапы биохимической эволюции»
«Основные этапы биохимической эволюции»Запишите номера предложений и допишите их:
Возраст Земли составляет (_).
Во время планетарной стадии первичная атмосфера Земли состояла из газов (_).
А.И.Опарин в 1924 году предположил, что на первом этапе зарождения жизни на Земле за счет энергии (_) произошел (_).
На втором этапе произошло образование (_), которые сливались в коллоидные капли – (_).
С.Миллер в 1953 году доказал (_).
На третьем этапе зарождения жизни появляется – (_).
Доказана (_) способность молекул РНК, обеспечивающая устойчивость коацерватов.
Эволюция РНК сопровождается взаимодействием с (_), появился белковый синтез, контролируемый РНК.
В дальнейшем преимущества получают ДНК, так как (_).

Первые организмы по типу питания были (_).
Важнейшим ароморфозом было появление двух типов автотрофного питания – (_).
Фотоавтотрофный тип питания бактерий с первой фотосистемой не сопровождался (_).
Кислород стал накапливаться в атмосфере благодаря (_).
Симбиоз архебактерий с бактериями окислителями привел к (_).
Симбиоз архебактерий с синезелеными привел к (_).
Обособление генетической информации привело к (_).
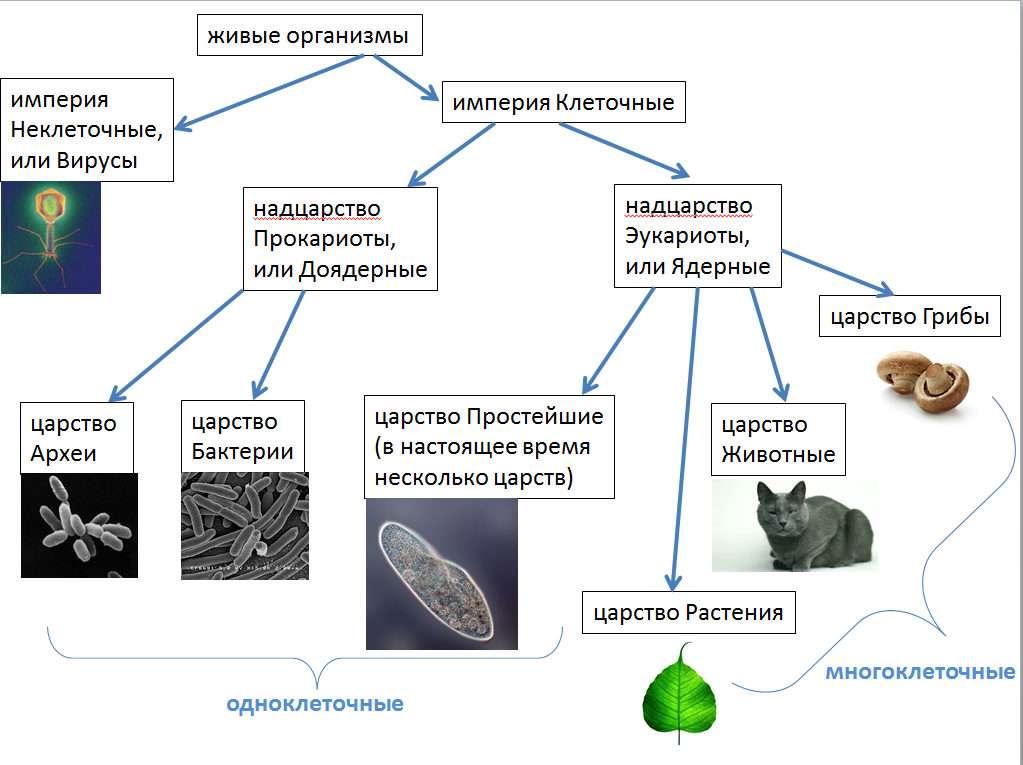
Задание 5. «Возникновение жизни на Земле»
Запишите номера тестов, против каждого – правильные варианты ответа
Тест 1. Доказал невозможность самозарождения личинок мух в сосудах с мясом, рыбой, угрями и змеей:
Л.Пастер.
Ф.Реди.
В.
Гельмонт.Спалланцани.
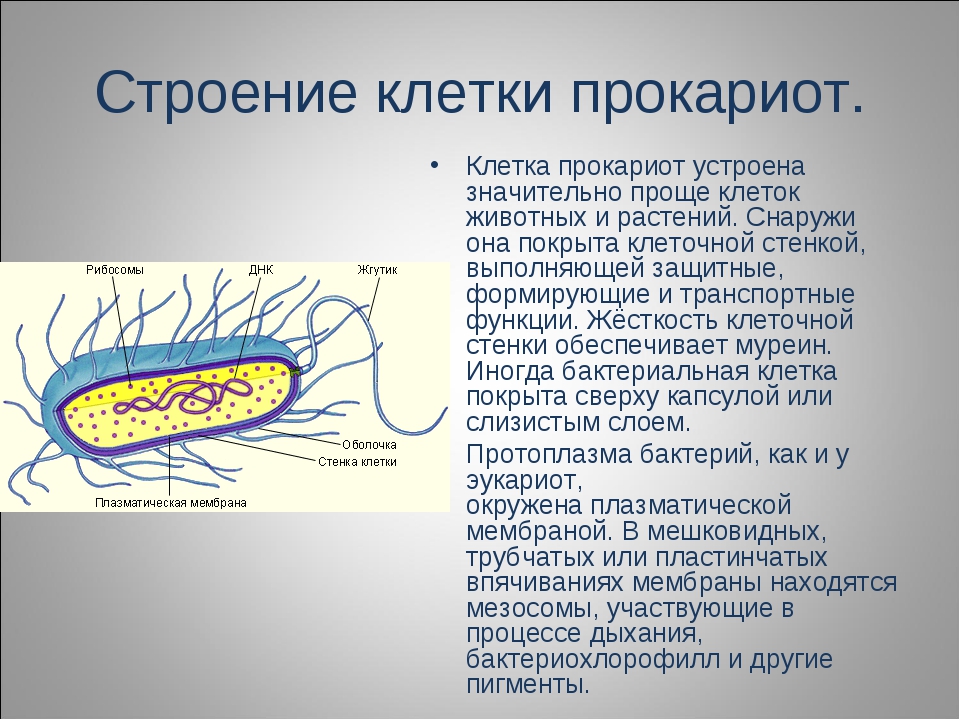 Гельмонт.
Гельмонт.Тест 2. Доказал невозможность самозарождения микроорганизмов в колбе с питательным бульоном при доступе воздуха:
Л.Пастер.
Ф.Реди.
В.Гельмонт.
Спалланцани.
Тест 3. Появление жизни на Земле с помощью «Высшей силы» объясняет:
Теория панспермии.
Теории биогенеза.
Теории абиогенеза.
Теория креационизма.
Тест 4. Утверждает, что жизнь на Землю занесена из космоса:
Теория панспермии.
Теория биогенеза.
Теория абиогенеза.
Теория креационизма.
Тест 5.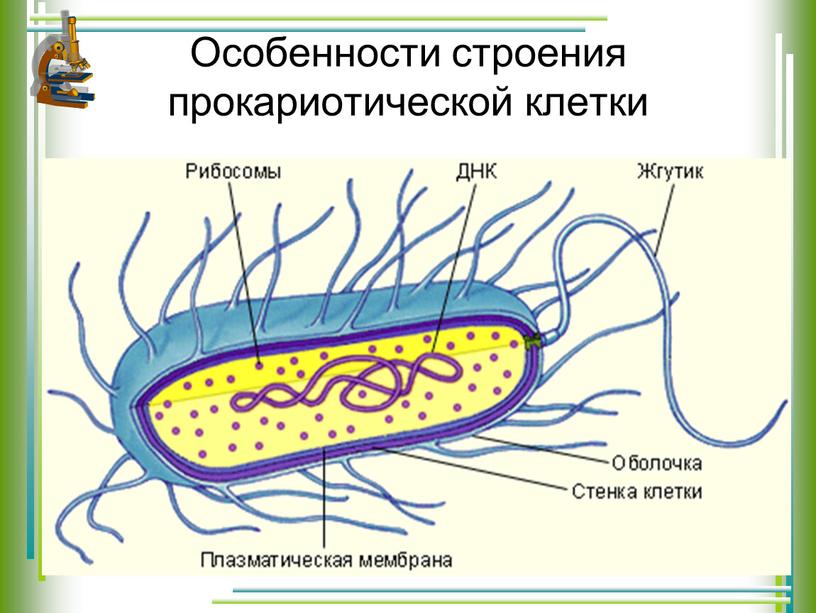 В 1924 году предположение, что в первичной атмосфере могли возникнуть простейшие органические соединения под влиянием электрических разрядов, было высказано:
В 1924 году предположение, что в первичной атмосфере могли возникнуть простейшие органические соединения под влиянием электрических разрядов, было высказано:
С.Миллером.
А.Г.Пасынским.
Т.Е.Павловской.
А.И.Опариным.
Л.Пастером.
Тест 6. Согласно гипотезе А.С.Опарина, на первом этапе зарождения жизни на Земле:
Появились коацерваты.
Происходил абиогенный синтез органических веществ.
Образовались биополимеры.
Появился матричный синтез.
Тест 7. На втором этапе зарождения жизни на Земле:
Появились прокариоты.
Появились эукариоты.
Образовались биополимеры.
Появился матричный синтез.

Тест 8. На третьем этапе зарождения жизни на Земле:
Появились прокариоты.
Появились эукариоты.
Образовались биополимеры.
Появился матричный синтез.
Тест 9. Первые живые организмы по типу обмена веществ были:
Аэробными автотрофами.
Анаэробными автотрофами.
Аэробными гетеротрофами.
Анаэробными гетеротрофами.
**Тест 10. В состав первичной атмосферы входили:
Кислород. 5. Метан.
Азот. 6. Аммиак.
Водород. 7. Пары воды.
Углекислый газ. 8. Угарный газ.

Задание 6. «Теория симбиогенеза»
Кем были первые пробионты по типу питания?
Какие организмы появились первыми: синезеленые или бактерии-окислители?
У каких организмов впервые появилась фотосистема-2?
Какие органеллы считаются эндосимбионтами эукариотической клетки?
Возможно ли возникновение жизни за счет биохимической эволюции на Земле сейчас? Ответ поясните.
Задание 7. Важнейшие термины и понятия: «Происхождение жизни»
Дайте определение терминам или раскройте понятия (одним предложением, подчеркнув важнейшие особенности):
1. Теории абиогенеза. 2. Теории биогенеза. 3. Теории самозарождения. 4. Теория панспермии. 5. Теория биохимической эволюции. 6. Коацерваты. 7. Теория симбиогенеза.
Задание 8. «Архей и протерозой»
Запишите номера вопросов и дайте ответ одним предложением:
Укажите временные рамки архейской эры.
Какие организмы жили в архейскую эру?
Какими по типу обмена веществ были первые организмы архея?
Какие организмы при фотосинтезе впервые стали выделять кислород в атмосферу?
Перечислите основные ароморфозы архейской эры.
Укажите временные рамки протерозойской эры.
Когда появились первые эукариоты?
Когда появились первые многоклеточные животные?
Перечислите основные ароморфозы протерозойской эры.
Чем был представлен растительный мир протерозоя?
Чем был представлен животный мир протерозоя?
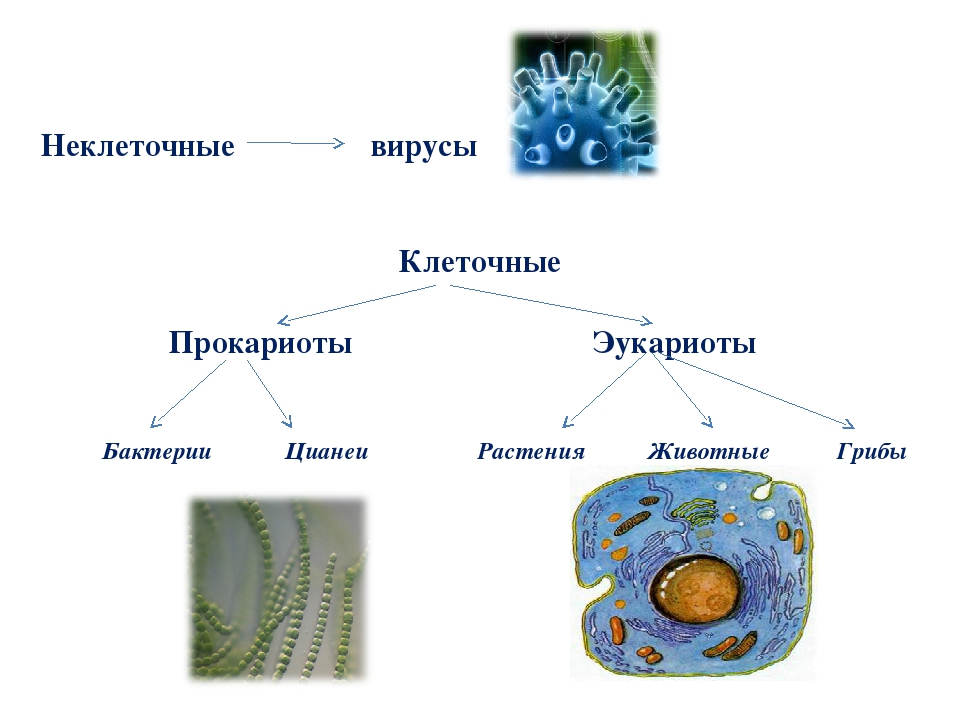
Задание 9. «Архейская эра»
Кем были первичные пробионты по типу питания?
Каким образом первичные пробионты получали энергию?
Какие организмы изменили атмосферу Земли, первыми начав выделение кислорода?
Какие организмы господствовали в архее?
Задание 10.
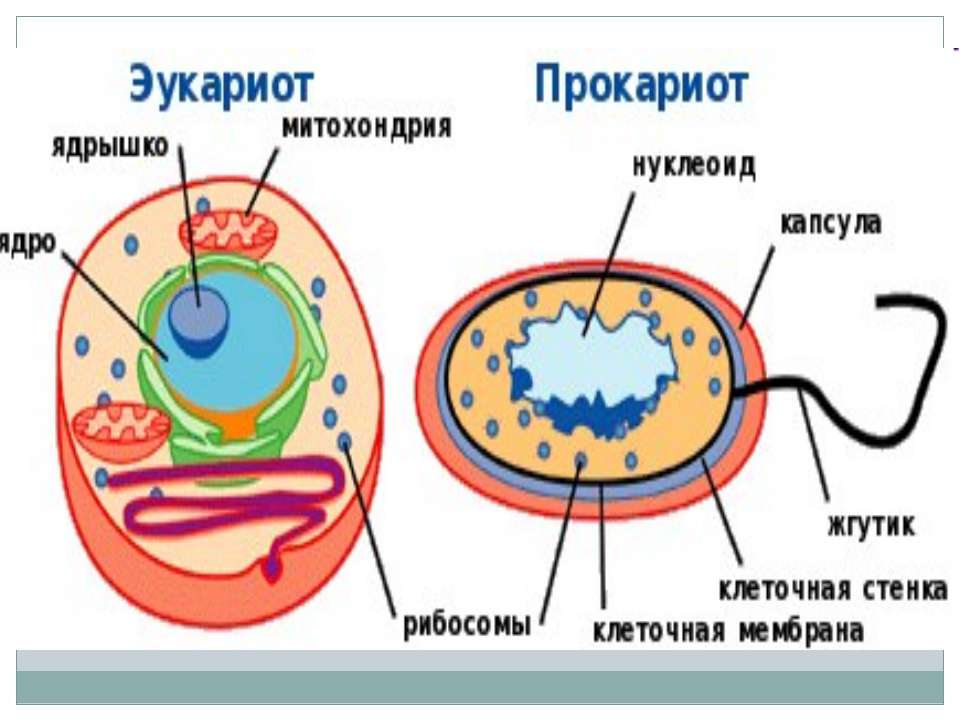 «Протерозойская эра»
«Протерозойская эра»Каковы временные рамки протерозойской эры?
Какие растения появились в протерозое?
Основные ароморфозы животных в протерозое.
Какие типы животных появились в протерозое?
Почему протерозойскую эры называют «;веком медуз»;?
Задание 11. «Эволюция растений в палеозое»
Заполните таблицу:
Периоды | Основные этапы эволюции растений |
Кембрий Ордовик Силурий Девон Карбон Пермь |
Задание 12. «Эволюция животных в палеозое»
Заполните таблицу:
Периоды | Основные этапы эволюции животных |
Кембрий Ордовик Силурий Девон Карбон Пермь |
Задание 13.
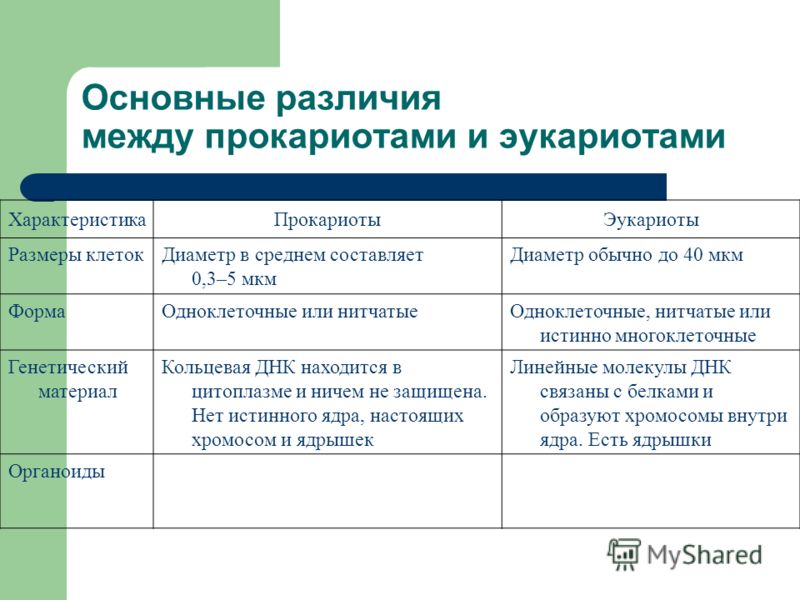 «Палеозойская эра»
«Палеозойская эра»Каковы временные рамки палеозойской эры?
На какие периоды делится палеозойская эра?
Какие ароморфозы привели к появлению псилофитов? В какой период?
Какие ароморфозы привели к появлению папоротникообразных? В какой период?
Какие ароморфозы привели к появлению семенных папоротников? В какой период?
Какие ароморфозы привели к появлению стегоцефалов? В какой период?
Какие ароморфозы привели к появлению пресмыкающихся? В какой период?
Задание 14. «Эволюция растений в мезозое»
Заполните таблицу:
Периоды | Основные этапы эволюции растений |
Триас Юра Мел |
Задание 15.
 «Эволюция животных в мезозое»
«Эволюция животных в мезозое»Заполните таблицу:
Периоды | Основные этапы эволюции животных |
Триас Юра Мел |
Задание 16. «Мезозойская эра»
Каковы временные рамки мезозойской эры?
На какие периоды делится мезозойская эра?
Какие растения и животные господствуют на Земле в мезозойскую эру?
Какие ароморфозы привели к появлению млекопитающих? В какие периоды они появились?
Какие ароморфозы привели к появлению птиц? В какой период появились птицы?
Когда появились первые цветковые растения? Благодаря каким ароморфозам?
Задание 17. «Эволюция растений и животных в кайнозое»
Заполните таблицу:
Периоды | Климат, растительность | Основные этапы эволюции животных |
Третичный — палеоген (40 млн. — неоген (25 млн. лет) Четвертичный (2 млн. лет) (антропоген) |
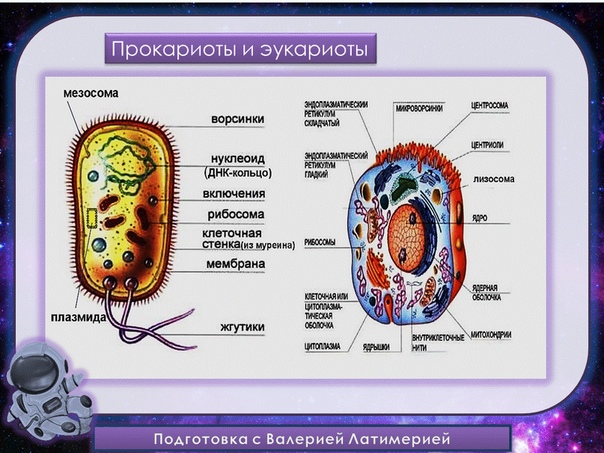 лет)
лет)Задание 18. «Эволюция в кайнозое»
Запишите номера тестов, против каждого – правильные варианты ответа
Тест 1. Временные рамки кайнозойской эры:
2500 — 570 млн лет назад.
570 — 230 млн лет назад.
230 — 67 млн лет назад.
67 млн лет — до наших дней.
Тест 2. Приматы появились от животных отряда:
Хищные.
Насекомоядные.
Грызуны.
Зверозубые пресмыкающиеся.
Тест 3. Первые приматы появились:
В конце мела мезозойской эры, 70 млн лет назад.
В конце палеогена.
В неогене.
В антропогене.
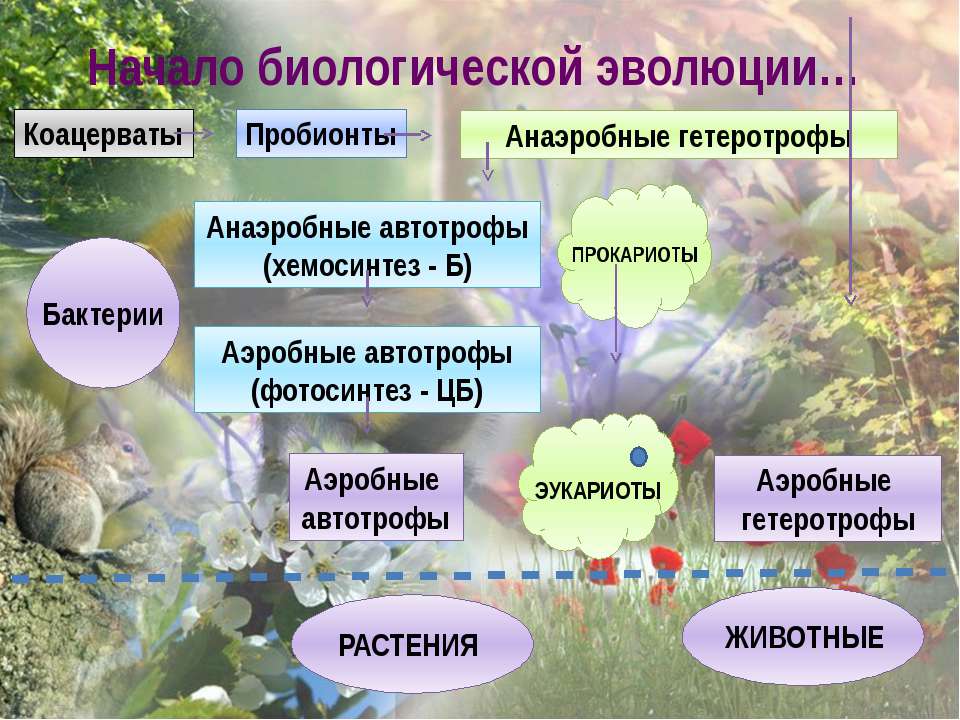
Тест 4. Климат в палеогене, (первой половине третичного периода):
Влажный, тропический.
Сухой, холодный.
Произошли четыре гигантских оледенения.
Тест 5. Климат в неогене (второй половине третичного периода):
Влажный, тропический.
Сухой, холодный.
Произошли четыре гигантских оледенения.
Тест 6. Климат в антропогене (четвертичном периоде):
Влажный, тропический.
Сухой, холодный.
Произошли четыре гигантских оледенения.
Тест 7. Гоминиды (прямоходящие приматы) появились:
В палеогене.
В неогене.
В антропогене.

Тест 8. Первые люди появились:
В начале палеогена.
В конце палеогена.
В начале неогена.
В антропогене.
Тест 9. Накопление гигантских запасов льда и снега на суше в антропогене кайнозойской эры привело к снижению уровня мирового океана:
На 20—40 метров.
На 40—60 метров.
На 60—90 метров.
На 90—120 метров.
**Тест 10. Сухопутные мосты в антропогене кайнозойской эры существовали:
Между Австралией и Азией.
Между Европой и Британскими островами.
Между Азией и Северной Америкой.
Между полуостровом Индокитай и островами Зондского архипелага.

Задание 19. «Эволюция органического мира»
В
Эры и периоды | Произошедшие события |
Архейская | |
Протерозойская | |
Палеозойская | |
Кембрий | |
Ордовик | |
Силурий | |
Девон | |
Карбон | |
Пермь | |
Мезозойская | |
Триас | |
Юра | |
Мел | |
Кайнозойская | |
Третичный | |
Четвертичный |
в конкретную эру и период:
1.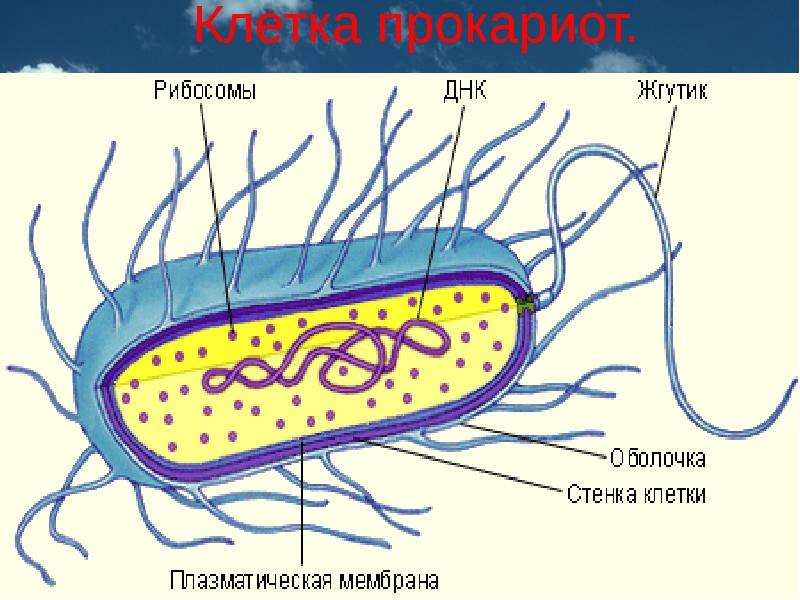 Вымирают мамонты, саблезубые тигры, шерстистые носороги; на 60-90 метров понизился уровень мирового океана. 2. Появились первые многоклеточные животные. 3. В атмосфере появился свободный кислород. 4. Появляются первые хордовые животные. 5. В этом периоде появляются панцирные «;рыбы»;, относящиеся к бесчелюстным позвоночным. 6. Период, в котором распространены леса тропического и субтропического типа, от насекомоядных млекопитающих обособляется отряд приматов. 7. По берегам пресных водоемов впервые появляются псилофиты. 8. От кистеперых рыб произошли древние земноводные — ихтиостеги. 9. Появились первые отряды крылатых насекомых — тараканы и стрекозы. 10. Появляются первые пресмыкающиеся. 11. Появляются первые голосеменные растения — семенные папоротники. 12. Появились первые
Вымирают мамонты, саблезубые тигры, шерстистые носороги; на 60-90 метров понизился уровень мирового океана. 2. Появились первые многоклеточные животные. 3. В атмосфере появился свободный кислород. 4. Появляются первые хордовые животные. 5. В этом периоде появляются панцирные «;рыбы»;, относящиеся к бесчелюстным позвоночным. 6. Период, в котором распространены леса тропического и субтропического типа, от насекомоядных млекопитающих обособляется отряд приматов. 7. По берегам пресных водоемов впервые появляются псилофиты. 8. От кистеперых рыб произошли древние земноводные — ихтиостеги. 9. Появились первые отряды крылатых насекомых — тараканы и стрекозы. 10. Появляются первые пресмыкающиеся. 11. Появляются первые голосеменные растения — семенные папоротники. 12. Появились первые
млекопитающие — яйцекладущие. 13. Появление бактерий и синезеленых. 14. Конец этой эры образно называют «;веком медуз»;. 15. Господство прокариот сменяется расцветом эукариот. 16. От пресмыкающихся, ведущих древесный образ жизни произошли птицы.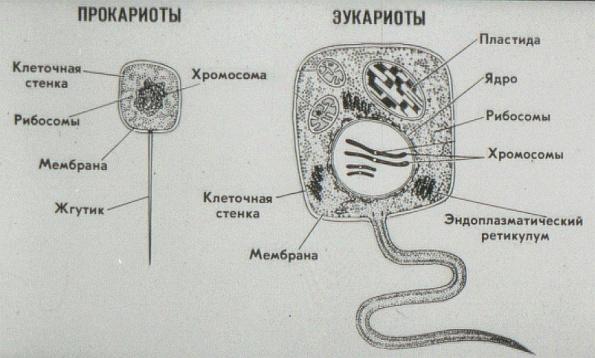 17. Эта эра получила название — эра пресмыкающихся и голосеменных. 18. Появляется большое количество животных с органическим или минеральным скелетом. 19. Появились первые сумчатые и плацентарные млекопитающие. 20. Появляются и чрезвычайно быстро распространяются покрытосеменные растения.
17. Эта эра получила название — эра пресмыкающихся и голосеменных. 18. Появляется большое количество животных с органическим или минеральным скелетом. 19. Появились первые сумчатые и плацентарные млекопитающие. 20. Появляются и чрезвычайно быстро распространяются покрытосеменные растения.
Задание 20. «Классификация живых организмов»
Заполните таблицу:
Систематические категории | Характерные особенности |
Империя Клеточные Надцарство Прокариоты Царство Дробянки Подцарство Архебактерии Подцарство Эубактерии Подцарство Оксифотобактерии (Синезеленые) Надцарство Эукариоты Царство Растения Подцарство Настоящие водоросли Подцарство Багрянковые Подцарство Высшие растения Царство Животные Подцарство Одноклеточные Подцарство Многоклеточные Царство Грибы | |
Империя Неклеточные Царство Вирусы |
Задание 21.
 «Эволюция органического мира»
«Эволюция органического мира»Запишите номера вопросов и дайте ответ одним предложением:
Каков возраст Земли?
Какие организмы появились в архейскую эру?
Какие организмы при фотосинтезе впервые стали выделять кислород в атмосферу?
Важнейшие ароморфозы архейской эры?
Растительный мир протерозоя?
Животный мир протерозоя?
Временные границы палеозойской эры?
Периоды палеозойской эры?
Временные границы мезозойской эры?
Периоды мезозойской эры?
Временные границы кайнозойской эры?
Периоды кайнозойской эры?
В какую эру и период появились псилофиты?
От какой группы водорослей произошли псилофиты?
Какие ароморфозы привели к появлению псилофитов?
В какую эру и период появились семенные папоротники?
Какие ароморфозы привели к появлению семенных папоротников?
В какую эру и период появились цветковые?
Какие ароморфозы привели к появлению цветковых?
В какую эру и период появились первые насекомые?
В какую эру и период появились крылатые насекомые?
В какую эру и период появились бесчелюстные «;рыбы»;?
В какую эру и период появились настоящие рыбы?
В какую эру и период появились стегоцефалы?
В какую эру и период появились первые пресмыкающиеся?
В какую эру и период появились яйцекладущие млекопитающие?
В какую эру и период появились сумчатые и плацентарные млекопитающие?
В какую эру и период появились первые птицы?
Какую эру можно назвать эрой млекопитающих и покрытосеменных?
В какую эру и период появился человек?
Какую эру можно назвать эрой медуз?
Какую эру можно назвать эрой папоротникообразных и земноводных?
Какую эру можно назвать эрой пресмыкающихся?
Какую эру можно назвать эрой цветковых и млекопитающих?
Какой климат в начале и в конце третичного периода?
В каком периоде было четыре великих оледенения?
Какие организмы относятся к империи Неклеточные?
Какие организмы относятся к надцарству Прокариот?
Какие организмы относятся к надцарству Эукариот?
Какие организмы способны фиксировать атмосферный азот?
Ответы:
Задание 1.
1. В результате ее сотворения высшим разумом. 2. Природа неизменна и изначально целесообразна, создана Богом-творцом, не развивается и не изменяется. 3. Что живые организмы возникают из неживого в результате самозарождения. 4. Живые организмы могут появиться только от живых организмов. 5. Самозарождения. 6. Невозможность самозарождения мух. 7. Невозможность самозарождения микроорганизмов. 8. Из космоса. 9. Образование органического вещества из неорганического на первом этапе, на втором – образование биополимеров и коацерватов. На третьем этапе в результате эволюции нуклеиновых кислот появились пробионты.
Задание 2.
1. В 1765 г. Ладзаро Спалланцани прокипятил мясные и овощные отвары в течение нескольких часов, затем он их запечатал. Исследовав их через несколько дней, Спалланцани не обнаружил в них никаких микроорганизмов. Он сделал вывод, что высокая температура убила все формы живых существ, и без них ничто живое уже не могло возникнуть. Но виталисты утверждали, что в запаянную колбу просто не могла попасть жизненная сила, которая находится в воздухе. 2. В 1860 г. Луи Пастер повторил опыты Спалланцани, но колбу не запаял, а длинную трубку горлышка изогнул S-образно. Воздух в колбу проходил свободно, а микробы оседали в горлышке и в бульон не попадали. Проходили месяцы, а содержимое колбы оставалось стерильным.
Но виталисты утверждали, что в запаянную колбу просто не могла попасть жизненная сила, которая находится в воздухе. 2. В 1860 г. Луи Пастер повторил опыты Спалланцани, но колбу не запаял, а длинную трубку горлышка изогнул S-образно. Воздух в колбу проходил свободно, а микробы оседали в горлышке и в бульон не попадали. Проходили месяцы, а содержимое колбы оставалось стерильным.
Задание 3.
1. Атмосфера была восстановительной и содержала СО2, СО, CH4, NH3, H2O, H2. 2. Абиогенный синтез органических молекул-мономеров из неорганических. 3. Некоторые аминокислоты, азотистые основания, простые сахара. 4. Белки, нуклеотиды и нуклеиновые кислоты, полисахариды. 5. Нет, в них отсутствует размножение на основе матричного синтеза.
Задание 4.
1. Более 6 млрд. лет. 2. СО2, СО, CH4, NH3, H2O, H2, была восстановительной.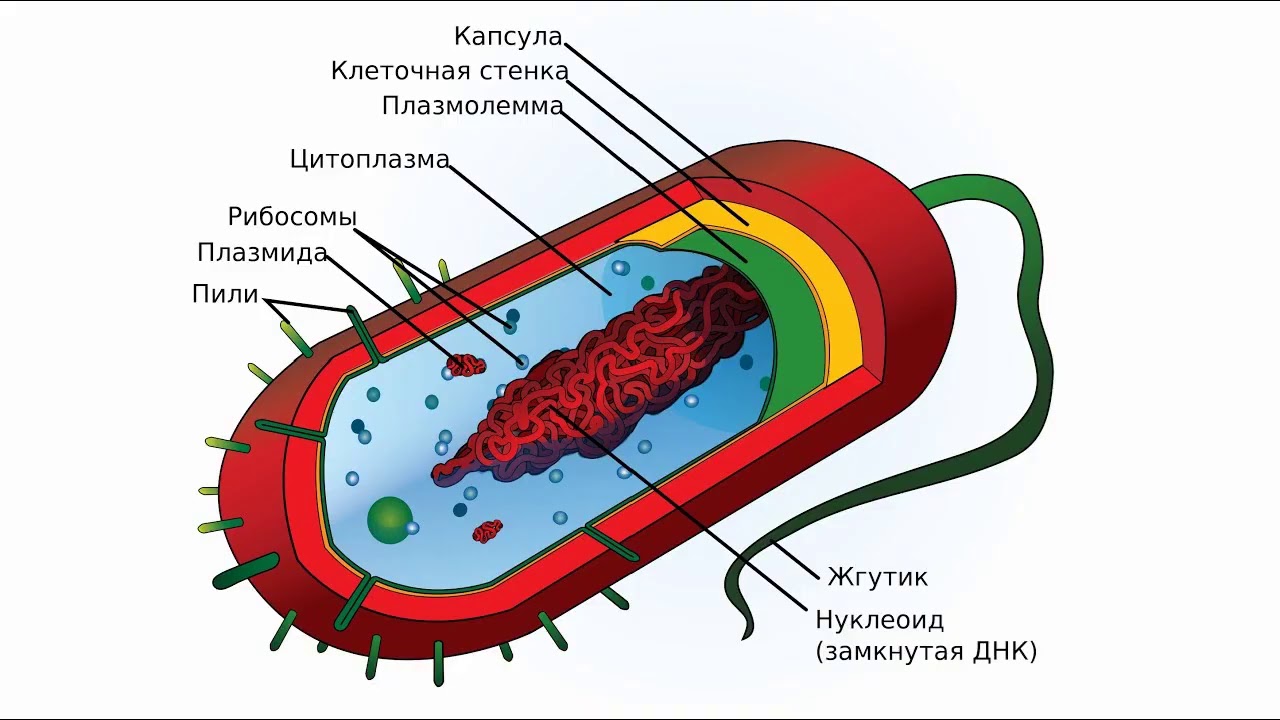 3. Грозовых разрядов, радиоактивного и ультрафиолетового излучения; абиогенный синтез органических мономеров из неорганических веществ. 4. Биополимеров – белков, жиров, углеводов, нуклеиновых кислот; коацерваты. 5. Возможность абиогенного синтеза некоторых аминокислот, азотистых оснований, рибозы, мочевины. 6. Доказана возможность саморепликации молекул РНК. 7. Каталитическая. 8. Аминокислотами и пептидами. 9. Их двуцепочечное строение обеспечивает точную репликацию. 10. Анаэробными гетеротрофами. 11. Хемоавтотрофного и фотоавтотрофного. 12. Выделением кислорода в атмосферу. 13. Появлению второй фотосистемы у цианобактерий. 14. К появлению митохондрий. 15. Появлению хлоропластов. 16. К появлению ядра.
3. Грозовых разрядов, радиоактивного и ультрафиолетового излучения; абиогенный синтез органических мономеров из неорганических веществ. 4. Биополимеров – белков, жиров, углеводов, нуклеиновых кислот; коацерваты. 5. Возможность абиогенного синтеза некоторых аминокислот, азотистых оснований, рибозы, мочевины. 6. Доказана возможность саморепликации молекул РНК. 7. Каталитическая. 8. Аминокислотами и пептидами. 9. Их двуцепочечное строение обеспечивает точную репликацию. 10. Анаэробными гетеротрофами. 11. Хемоавтотрофного и фотоавтотрофного. 12. Выделением кислорода в атмосферу. 13. Появлению второй фотосистемы у цианобактерий. 14. К появлению митохондрий. 15. Появлению хлоропластов. 16. К появлению ядра.
Задание 5.
Тест 1: 2. Тест 2: 1. Тест 3: 4. Тест 4: 1. Тест 5: 4. Тест 6: 2. Тест 7: 3. Тест 8: 4. Тест 9: 4. **Тест 10: 3, 4, 5, 6, 7, 8.
Задание 6.
1. Анаэробными гетеротрофами. 2. Сине-зеленые, для окислителей нужен кислород. 3. У цианобактерий. 4. Хлоропласты, митохондрии, жгутики. 5. Нет, сейчас атмосфера не восстановительная, а окислительная — произойдет окисление образовавшейся органики. Кроме того, образовавшиеся органические вещества будут поглощены гетеротрофными организмами.
Задание 7.
1. Теории, согласно которым живые организмы образовывались из неживой материи путем самозарождения. 2. Теории, утверждавшие, что живые организмы могут появиться только от живых, самозарождение невозможно. 3. Теории, господствовавшие до середины 19 века и утверждавшие в возможность зарождения живых организмов из неживой природы. 4. Теория, согласно которой жизнь на Землю попала из космоса. 5. Теория, объясняющая возможность зарождения жизни на Земле в результате биохимической эволюции. 6. Коллоидные капли, состоящие из органических макромолекул. 7. Теория, объясняющая происхождение митохондрий, пластид, возможно и жгутиков эукариот симбиозом архебактерий с различными видами прокариотических организмов.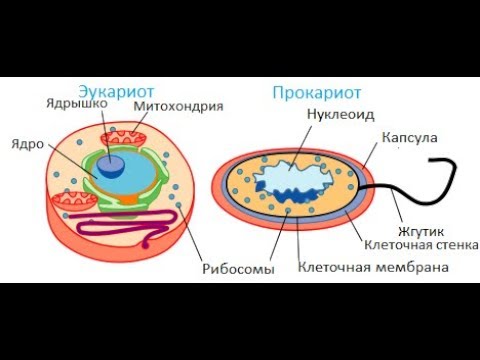
Задание 8.
1. 3,5 – 2,6 млрд.лет. 2. Прокариотические. 3. Анаэробными гетеротрофами. 4. Цианобактерии. 5. Появление автотрофного типа питания – хемотрофного и фототрофного. Причем фотосинтез зеленых и пурпурных бактерий с фотосистемой 1 не сопровождался выделением кислорода, и только появление цианобактерий с фотосистемой 2 привело к фотолизу воды и выделению кислорода. Важным ароморфозом можно считать появление дыхания – появление бактерий, способных использовать кислород в качестве окислителя. 6. 2,6 млрд. лет – 570 млн. лет. 7. Около 1,5 млрд. лет назад. 8. Около 1 млрд. лет назад. 9. Появление эукариот, многоклеточности, полового размножения, двусторонней симметрии. 10. Представителями всех отделов водорослей. 11. Появились все типы животных, кроме вторичноротых – иглокожих и хордовых.
Задание 9.
1. Анаэробными гетеротрофами. 2. За счет окисления готовых органических веществ. 3. Синезеленые. 4. Прокариоты.
4. Прокариоты.
Задание 10.
1. 2600—570 млн. лет. 2. Все отделы водорослей. 3. Многоклеточность, пищеварительный тракт, половое размножение, двусторонняя симметрия. 4. Все типы животных кроме иглокожих и хордовых. 5. Животные, имеющие органический или минеральный скелет, появились только в конце протерозойской эры.
Задание 11.
Периоды | Основные этапы эволюции растений |
Кембрий Ордовик Силурий Девон Карбон Пермь | В палеозое существовал один сверхматерик — Пангея. В морях и океанах распространяются водоросли. В конце силура, начале девона от зеленых водорослей прибрежной зоны появляются первые наземные растения — псилофиты (440 млн. лет). Ароморфозы: появление покровных, механических и проводящих тканей, возникает сосудистая система. В конце девона от псилофитов выделились высшие споровые растения: плауны, хвощи, папоротниковидные. Ароморфозы: появление корней, листьев. В карбоне появляются первые семенные растения — семенные папоротники. Ароморфозы: опыление ветром и слияние гамет без участия воды; образование семян, содержащих большой запас питательных веществ. Гаметофиты развиваются на спорофите и за счет питательных веществ спорофита, после оплодотворения формируется семя с зародышем и запасом питательных веществ. В связи с засушливым климатом начинается вытеснение споровых растений голосеменными. |

Задание 12.
Периоды | Основные этапы эволюции животных |
Кембрий Ордовик Силурий Девон Карбон Пермь | На суше только бактерии и синезеленые. Появляются первые хордовые животные. Появляются первые позвоночные — «;панцирные рыбы»;, которые не имели челюстей и парных конечностей и относились к бесчелюстным, или круглоротым. На суше появляются пауки. Потомки панцирных рыб дали хрящевых и костных рыб. Среди костных рыб выделилась группа кистеперых рыб, девон стал временем их расцвета. Парные плавники с мощным внутренним скелетом позволяли им передвигаться по мелководью, а глоточные карманы превратились в легкие, обеспечивающие дополнительное дыхание. В конце девона от пресноводных кистеперых появляются первые земноводные (ихтиостеги, от них произошли стегоцефалы). Появляются многоножки и бескрылые насекомые. Появляются крылатые насекомые. В конце карбона появляются первые пресмыкающиеся (котилозавры). Ароморфозы: внутреннее оплодотворение, хорошо развитые легкие, сухая кожа, откладывание амниотических яиц на суше. Распространяются пресмыкающиеся. |
 В морях — расцвет скелетных беспозвоночных. Характерны трилобиты. Появились иглокожие.
В морях — расцвет скелетных беспозвоночных. Характерны трилобиты. Появились иглокожие.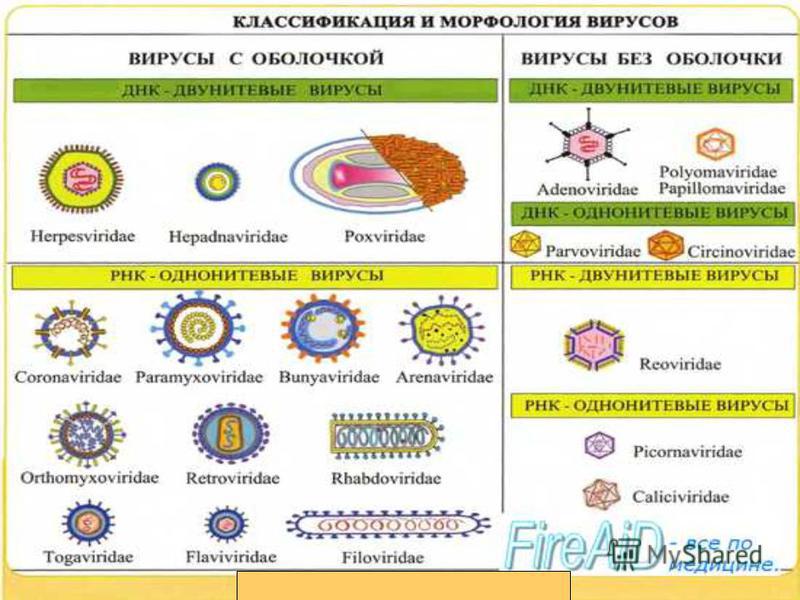
Задание 13.
1. 570 — 230 млн. лет. 2. Кембрий, ордовик, силурий, девон, карбон, пермь.3. Появление механических, покровных и проводящих тканей. В конце силурия. 4. Появление корней и листьев. В девоне. 5. Ветроопыление, образование семян. Гаметофиты развиваются на спорофите за счет питательных веществ спорофита, после оплодотворения формируется семя с зародышем и запасом питательных веществ. В карбоне. 6. Конечности рычажного типа, состоящие из трех отделов и легочное дыхание. В девоне. 7. Внутреннее оплодотворение, сухая кожа, усложнение легких, откладывание амниотических яиц. В карбоне.
Задание 14.
Периоды | Основные этапы эволюции растений |
Триас Юра Мел | Климат засушливый. В начале мелового периода появились первые цветковые растения. Цветок обеспечивал опыление, околоплодник защищал семена и способствовал их распространению, разнообразные жизненные формы позволили заселить различные среды обитания. |
 Почти полностью вымирают древовидные папоротники, хвощи, плауны. Происходит дальнейшее распространение голосеменных растений.
Почти полностью вымирают древовидные папоротники, хвощи, плауны. Происходит дальнейшее распространение голосеменных растений.Задание 15.
Периоды | Основные этапы эволюции животных |
Триас Юра Мел | Вымирает большинство земноводных. Происходит дальнейшее распространение пресмыкающихся. Появляются динозавры, размеры которых достигают 5—6 м. Возникли первые яйцекладущие млекопитающие и в конце периода появились первые птицы. Период расцвета пресмыкающихся, которые занимали господствующее положение на суше, в воздухе, многие вторично вернулись в водную среду. Птицы еще сохранили зубы. В конце мела происходит похолодание, вымирают аммониты и белемниты, растительноядные и хищные динозавры. Начинается расцвет теплокровных животных — птиц и млекопитающих. |
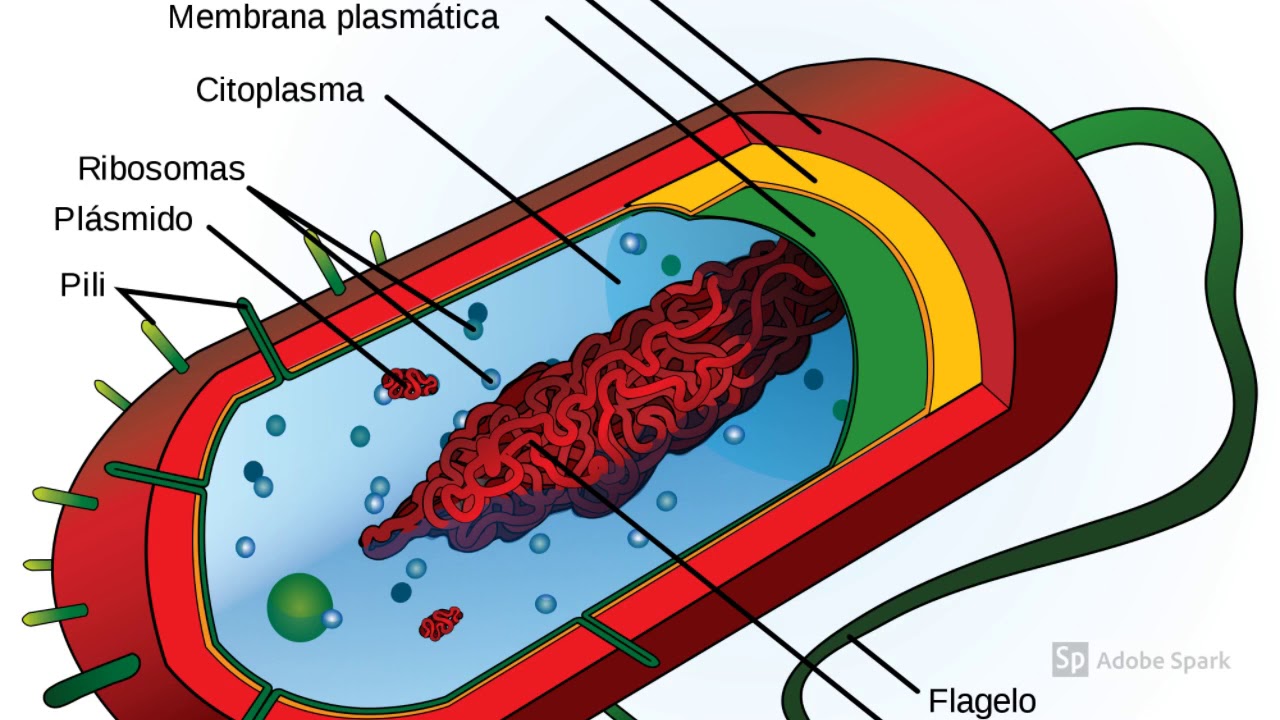 Появляются птицы. В морях многочисленны головоногие моллюски — аммониты и белемниты. Появились сумчатые и плацентарные млекопитающие.
Появляются птицы. В морях многочисленны головоногие моллюски — аммониты и белемниты. Появились сумчатые и плацентарные млекопитающие.Задание 16.
1. 230 — 67 млн. лет. 2. Триас, юра, мел. 3. Голосеменные и пресмыкающиеся. 4. Четырехкамерное сердце, волосяной покров, живорождение и выкармливание молоком, усложнение нервной системы. В триасе – яйцекладущие, юре – сумчатые и плацентарные млекопитающие. 5. Четырехкамерное сердце, оперение, крылья. В конце триаса. 6. В меловом периоде. Важнейшие ароморфозы — появление цветка и плода.
Задание 17.
Периоды | Климат, растительность | Основные этапы эволюции животных |
Третичный Четвертичный | В первой половине периода преобладает тропическая и субтропическая растительность, во второй происходит остепнение, однодольные вытесняют древесную растительность. Евразия и Северная Америка четырежды подвергались оледенениям. Накопление гигантских запасов льда привело к снижению уровня мирового океана на 60—90 м. Появились сухопутные мосты между Европой и Британскими островами, Азией и Северной Америкой. | В первой половине периода возникли все современные отряды млекопитающих. От насекомоядных млекопитающих обособляется отряд приматов. В середине периода появились предки понгид (человекообразных обезьян) и гоминид (прямоходящих приматов). В конце периода от гоминид произошли люди. Вымиранию мамонтов, шерстистых носорогов способствовали древние охотники. Сухопутные мосты способствовали обмену фауной и флорой между материками, расселению людей. Отсутствие моста сохранило жизнь примитивным млекопитающим Австралии. |
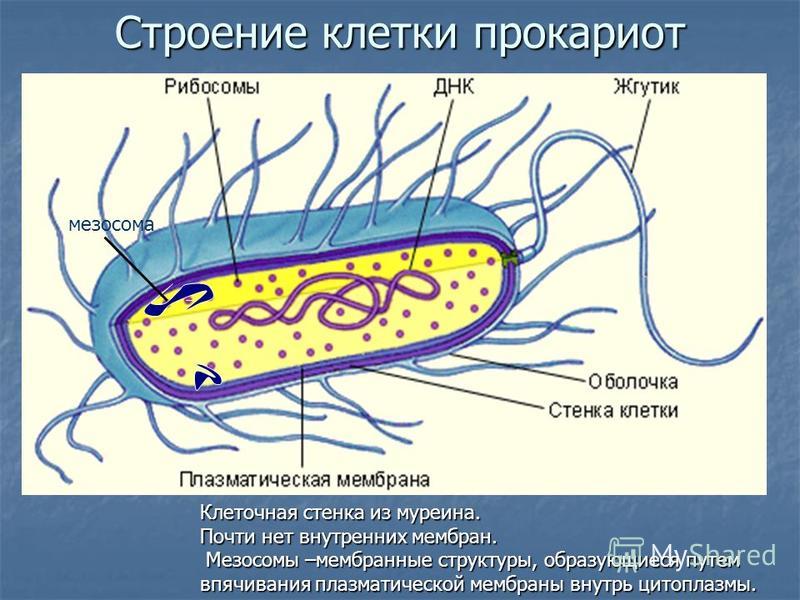
Задание 18.
Тест 1: 4. Тест 2: 2. Тест 3: 1. Тест 4: 1. Тест 5: 2. Тест 6: 3. Тест 7: 2. Тест 8: 4. Тест 9: 3. **Тест 10: 2, 3, 4.
Тест 6: 3. Тест 7: 2. Тест 8: 4. Тест 9: 3. **Тест 10: 2, 3, 4.
Задание 19.
Архейская эра (3, 13). Протерозойская эра (2, 14, 15). Палеозойская эра: кембрий (18), ордовик (4), силурий (5, 7), девон (8), карбон (9, 10 11), пермь. Мезозойская эра (17): триас (12,16), юра (19), мел (20). Кайнозойская эра: третичный (6), четвертичный (1).
Задание 20.
Систематические категории | Характерные особенности |
Империя Клеточные Надцарство Прокариоты Царство Дробянки Подцарство Архебактерии Подцарство Эубактерии Подцарство Оксифотобактерии Надцарство Эукариоты Царство Растения Подцарство Настоящие водоросли Подцарство Багрянковые Подцарство Высшие растения Царство Грибы Царство Животные Подцарство Одноклеточные Подцарство Многоклеточные | Организмы, имеющие клеточное строение. Доядерные организмы. Около 50 видов наиболее древних метанообразующих, галообразующих и серозависимых аэробных и анаэробных бактерий. Настоящие бактерии — одноклеточные организмы, без ядра, хлоропластов, митохондрий, центриолей, ЭПС, комплекса Гольджи. Аэробы и анаэробы. Нет ядра, вакуолей, нет полового размножения. Фотосинтез сопровождается фотолизом воды и выделением кислорода. Ядерные организмы. Фотосинтезирующие организмы. Низшие растения, не имеющие тканей и органов. Одноклеточные и многоклеточные организмы. Отличаются наличием пигментов красного и синего цвета и отсутствием подвижных стадий в циклах развития. Растения, имеющие ткани и органы (корень, листья, стебель). Делятся на споровые (мхи, плауны, хвощи, папоротники) и семенные (голосеменные и покрытосеменные). Гетеротрофные эукариоты, не способные к движению и совмещающие признаки как растений, так и животных. Гетеротрофные организмы, как правило, активно передвигаются, запасное вещество гликоген, у многоклеточных имеется нервная система. Тело простейших состоит из одной единственной клетки, которая является целым организмом. У многоклеточных происходит специализация клеток, образуются ткани и органы. |
Империя Неклеточные Царство Вирусы | Включает организмы, не имеющие клеточного строения. Объединяет организмы, являющиеся паразитами эукариотических и бактериальных клеток (бактериофаги), содержащие только один тип нуклеиновых кислот (ДНК или РНК) и использующие обмен веществ и энергию клетки—хозяина. |
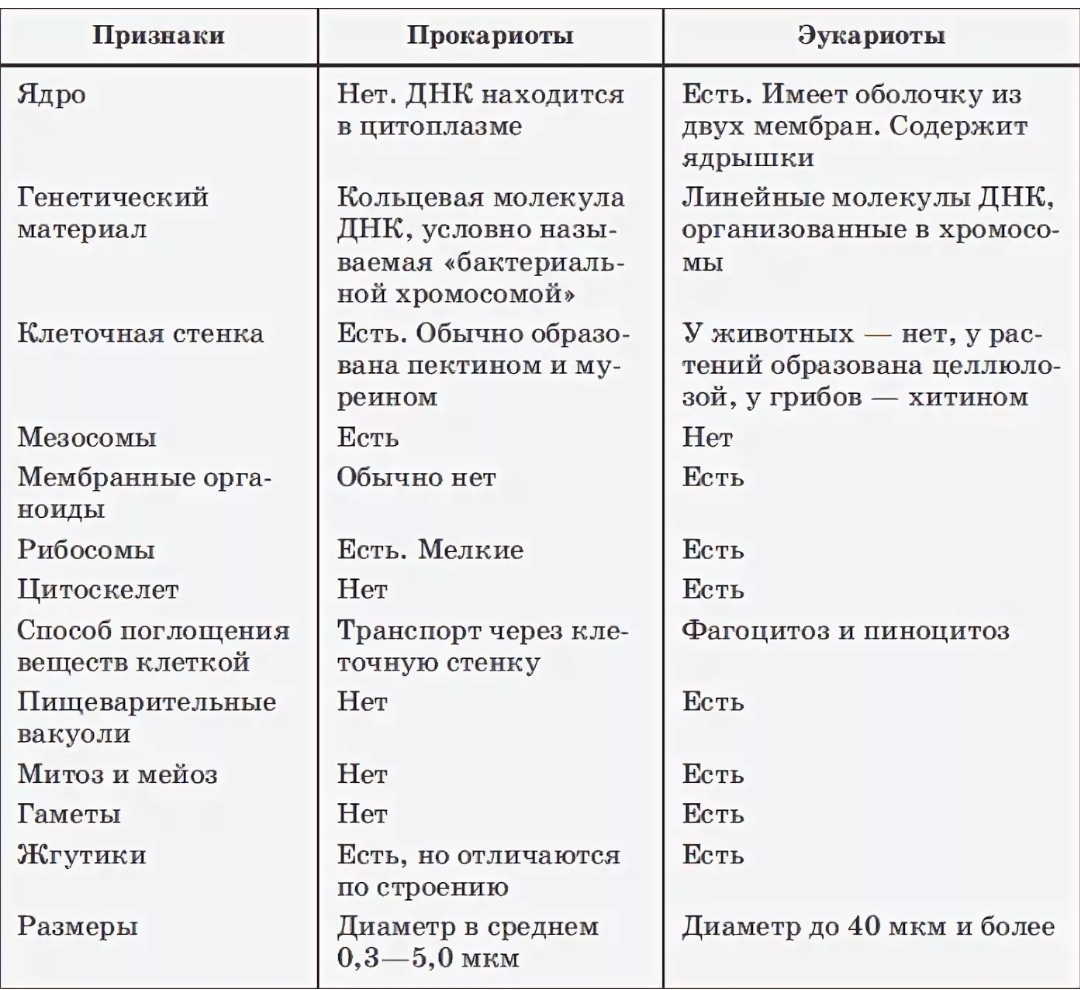
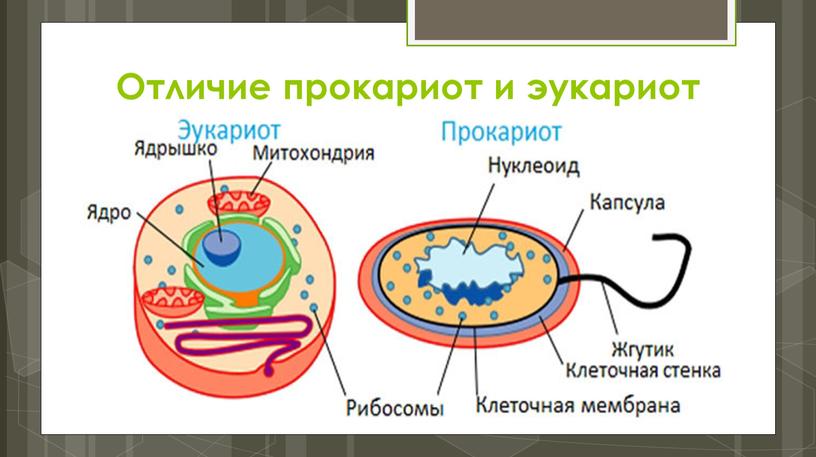
Задание 21.
1. Около 6—7 млрд. лет. 2. Прокариоты: архебактерии, эубактерии и цианобактерии. 3. Цианобактерии. 4. Появление фото- и хемоавтотрофных бактерий, появление дыхания. 5. Появились все отделы водорослей. 6. Появились все типы животных, кроме иглокожих и хордовых. 7. 570-230 млн. лет. 8. Кембрий, ордовик, силурий, девон, карбон, пермь. 9. 230-67 млн. лет. 10. Триас, юра, мел. 11. 67 млн. лет – настоящее время. 12. Третичный (палеоген и неоген) и четвертичный (антропоген). 13. Палеозойская эра, конец силурия. 14. От зеленых водорослей. 15. Механические, покровные и проводящие ткани. 16. Палеозойская эра, карбон. 17. Ветроопыление, развитие женского гаметофита на спорофите, образование семян. 18. Мезозойская эра, мел. 19. Появление цветка и плода. 20. Палеозойская эра, девон. 21. Палеозойская эра, карбон. 22. Палеозойская эра, силурий. 23. Палеозойская эра, девон. 24. Палеозойская эра, девон. 25. Палеозойская эра, карбон. 26. Мезозойская эра, триас. 27. Мезозойская эра, юра. 28. Мезозойская эра, триас. 29. Кайнозойскую. 30. В кайнозойскую эру, четвертичный период. 31. Протерозойскую. 32. Палеозойскую. 33. Мезозойскую. 34. Кайнозойскую. 35. В начале — тропический, в конце — континентальный.
Появились все отделы водорослей. 6. Появились все типы животных, кроме иглокожих и хордовых. 7. 570-230 млн. лет. 8. Кембрий, ордовик, силурий, девон, карбон, пермь. 9. 230-67 млн. лет. 10. Триас, юра, мел. 11. 67 млн. лет – настоящее время. 12. Третичный (палеоген и неоген) и четвертичный (антропоген). 13. Палеозойская эра, конец силурия. 14. От зеленых водорослей. 15. Механические, покровные и проводящие ткани. 16. Палеозойская эра, карбон. 17. Ветроопыление, развитие женского гаметофита на спорофите, образование семян. 18. Мезозойская эра, мел. 19. Появление цветка и плода. 20. Палеозойская эра, девон. 21. Палеозойская эра, карбон. 22. Палеозойская эра, силурий. 23. Палеозойская эра, девон. 24. Палеозойская эра, девон. 25. Палеозойская эра, карбон. 26. Мезозойская эра, триас. 27. Мезозойская эра, юра. 28. Мезозойская эра, триас. 29. Кайнозойскую. 30. В кайнозойскую эру, четвертичный период. 31. Протерозойскую. 32. Палеозойскую. 33. Мезозойскую. 34. Кайнозойскую. 35. В начале — тропический, в конце — континентальный. 36. В четвертичном кайнозойской эры. 37. Царство Вирусы. 38. Царство Дробянки. 39. Царства Растения, Животные, Грибы. 40. Некоторые прокариоты: клубеньковые бактерии, цианобактерии, некоторые свободноживущие эубактерии.
36. В четвертичном кайнозойской эры. 37. Царство Вирусы. 38. Царство Дробянки. 39. Царства Растения, Животные, Грибы. 40. Некоторые прокариоты: клубеньковые бактерии, цианобактерии, некоторые свободноживущие эубактерии.
Эукариоты, микробиология: бактерий и архей
Эукариоты, микробиология / Eukaryotes, Microbiology
Эукариоты ядерные (Nuclear eukaryotes) домен (надцарство) живых организмов, клетки которых содержат ядро. Эукариот — организм со сложными клетками , или одна клетка с сложными структурами. В этих клетках генетический материал организован в хромосомы в ядре клетки . Животные , растения , водоросли и грибы являются эукариотами. Есть также эукариоты среди одноклеточных протистов . Напротив, более простые организмы, такие как бактерии и археи , не имеют ядер и других сложных клеточных структур. Такие организмы называются прокариотами .
Все организмы, кроме прокариот (бактерий (bacteria) и архей (archaea)), являются ядерными.
Вирусы и вироиды также не являются ни прокариотами, ни эукариотами; более того, сам вопрос, считать ли их живыми организмами, является дискуссионным.
 Классификации активно обсуждается научным сообществом, и несколько таксономии были предложены. Все современные версии имеют пять королевств , но не согласны с тем, какие группы входят в каждое королевство.
Классификации активно обсуждается научным сообществом, и несколько таксономии были предложены. Все современные версии имеют пять королевств , но не согласны с тем, какие группы входят в каждое королевство.Описание Эукариоты / Description of Eukaryotes
Эукариотические организмы
Животные (Animals), растения (plants), грибы (mushrooms), а также группы организмов под общим названием протисты — все являются эукариотическими организмами. Они могут быть одноклеточными и многоклеточными, но все имеют общий план строения клеток.
Считается, что все эти столь несхожие организмы имеют общее происхождение, поэтому группа ядерных рассматривается как монофилетический таксон наивысшего ранга. Согласно наиболее распространённым гипотезам, эукариоты появились 1,6—2,1 млрд лет назад. Важную роль в эволюции эукариот сыграл симбиогенез — симбиоз между эукариотической клеткой, видимо, уже имевшей ядро и способной к фагоцитозу, и поглощёнными этой клеткой бактериями — предшественниками митохондрий и пластид.
Структура Строение эукариотической клетки / Structure Structure of a eukaryotic cell
Эукариотические клетки в среднем намного крупнее прокариотических, разница в объёме достигает тысяч раз. Клетки эукариот включают около десятка видов различных структур, известных как органеллы (другое название, реже употребляемое в научной литературе[комм. 1], — органоиды), из которых многие отделены от цитоплазмы одной или несколькими мембранами (в прокариотических клетках внутренние органоиды, окруженные мембраной, встречаются редко). Ядро — часть клетки, окружённая у эукариот двойной мембраной (двумя элементарными мембранами) и содержащая генетический материал: молекулы ДНК, «упакованные» в хромосомы. Ядро обычно одно, но бывают и многоядерные клетки.
Деление на царства / Division into kingdoms
Существует несколько вариантов деления надцарства эукариот на царства.
- Первыми были выделены царства растений и животных.
-
Затем было выделено царство грибов, которые, по мнению большинства биологов, не могут быть причислены ни к одному из этих царств из-за биохимических особенностей.
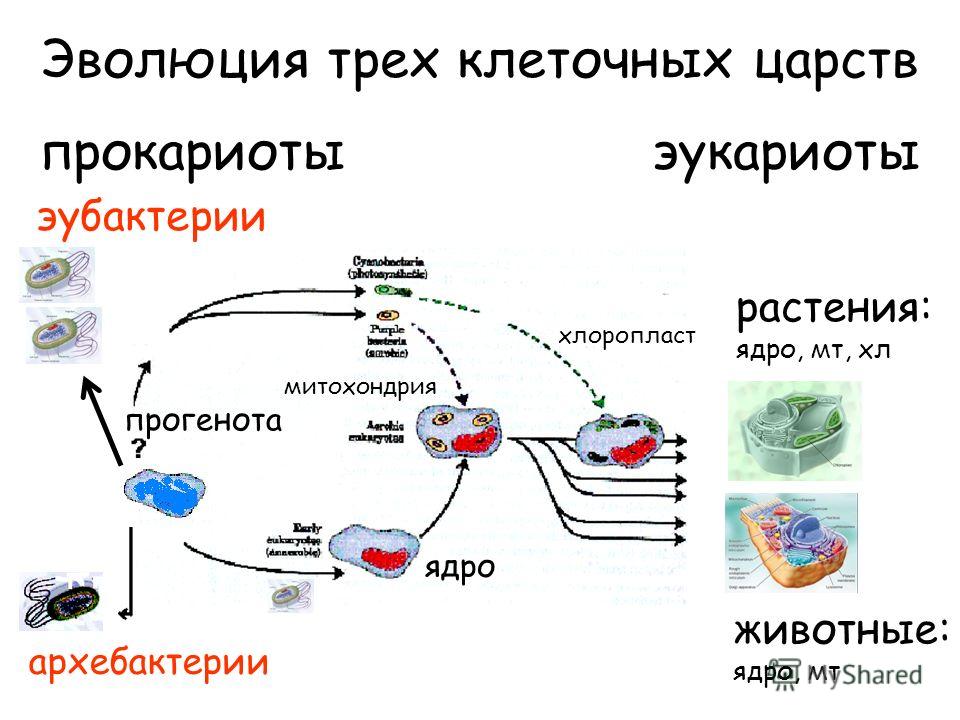
Также некоторые авторы выделяют царства протистов или простейших и хромистов. Некоторые системы насчитывают до 20 царств.
По системе Томаса Кавалье-Смита, все эукариоты подразделяются на два монофилетических таксона (поддомена) — униконтов (Unikonta) и биконтов (Bikonta). Положение таких эукариот, как Collodictyon и Diphylleia rotans, на данный момент не определено.
Отличия эукариот от прокариот / Differences of eukaryotes from prokaryotes
Важнейшая, основополагающая особенность эукариот связана с расположением генетического аппарата в клетке. Генетический аппарат всех эукариот находится в ядре и защищён ядерной оболочкой. ДНК эукариот линейная (у прокариот ДНК кольцевая и находится в особой области клетки — нуклеоиде, который не отделён мембраной от остальной цитоплазмы). Она связана с белками-гистонами и другими белками хромосом, которых нет у бактерий.
В жизненном цикле эукариот обычно присутствуют две ядерные фазы (гаплофаза и диплофаза). Первая фаза характеризуется гаплоидным (одинарным) набором хромосом, далее, сливаясь, две гаплоидные клетки (или два ядра) образуют диплоидную клетку (ядро), содержащую двойной (диплоидный) набор хромосом. Иногда при следующем делении, а чаще спустя несколько делений клетка вновь становится гаплоидной.
Первая фаза характеризуется гаплоидным (одинарным) набором хромосом, далее, сливаясь, две гаплоидные клетки (или два ядра) образуют диплоидную клетку (ядро), содержащую двойной (диплоидный) набор хромосом. Иногда при следующем делении, а чаще спустя несколько делений клетка вновь становится гаплоидной.
Такой жизненный цикл и в целом диплоидность (diploid) для прокариот не характерны.
Третье, пожалуй, самое интересное отличие, — это наличие у эукариотических клеток особых органелл, имеющих свой генетический аппарат, размножающихся делением и окружённых мембраной. Эти органеллы — митохондрии и пластиды. По своему строению и жизнедеятельности они поразительно похожи на бактерий. Это обстоятельство натолкнуло современных учёных на мысль, что подобные организмы являются потомками бактерий, вступившими в симбиотические отношения с эукариотами. Прокариоты характеризуются малым количеством органелл, и ни одна из них не окружена двойной мембраной.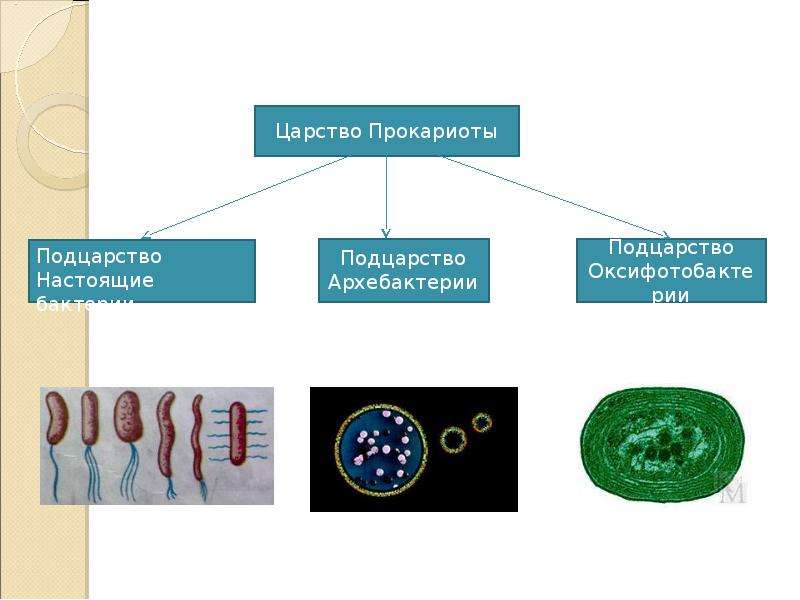 В клетках прокариот нет эндоплазматического ретикулума (no endoplasmic reticulum), аппарата Гольджи, лизосом.
В клетках прокариот нет эндоплазматического ретикулума (no endoplasmic reticulum), аппарата Гольджи, лизосом.
Эндоцитоз и фагоцитоз — захватт, заключение в мембранный пузырёк, переваривать твёрдых частиц
Ещё одно важное различие между прокариотами и эукариотами — наличие у эукариот эндоцитоза, в том числе у многих групп — фагоцитоза. Фагоцитозом (дословно «поедание клеткой») называют способность эукариотических клеток захватывать, заключая в мембранный пузырёк, и переваривать самые разные твёрдые частицы. Этот процесс обеспечивает в организме важную защитную функцию. Впервые он был открыт И. И. Мечниковым у морских звёзд. Появление фагоцитоза (phagocytosis) у эукариот скорее всего связано со средними размерами (далее о размерных различиях написано подробнее). Размеры прокариотических клеток несоизмеримо меньше, и поэтому в процессе эволюционного развития эукариот у них возникла проблема снабжения организма большим количеством пищи. Как следствие, среди эукариот появляются первые настоящие, подвижные хищники.
Большинство бактерий имеет клеточную стенку, отличную от эукариотической (далеко не все эукариоты имеют её). У прокариот это прочная структура, состоящая главным образом из муреина (у архей из псевдомуреина). Строение муреина таково, что каждая клетка окружена особым сетчатым мешком, являющимся одной огромной молекулой. Среди эукариот клеточную стенку имеют многие протисты, грибы и растения. У грибов она состоит из хитина и глюканов, у низших растений — из целлюлозы и гликопротеинов, диатомовые водоросли синтезируют клеточную стенку из кремниевых кислот, у высших растений она состоит из целлюлозы, гемицеллюлозы и пектина. Видимо, для более крупных эукариотических клеток стало невозможно создавать достаточно прочную клеточную стенку из одной молекулы. Это обстоятельство могло заставить их использовать для клеточной стенки иной материал.
Другое объяснение состоит в том, что общий предок эукариот в связи с переходом к хищничеству утратил клеточную стенку, а затем были утрачены и гены, отвечающие за синтез муреина.
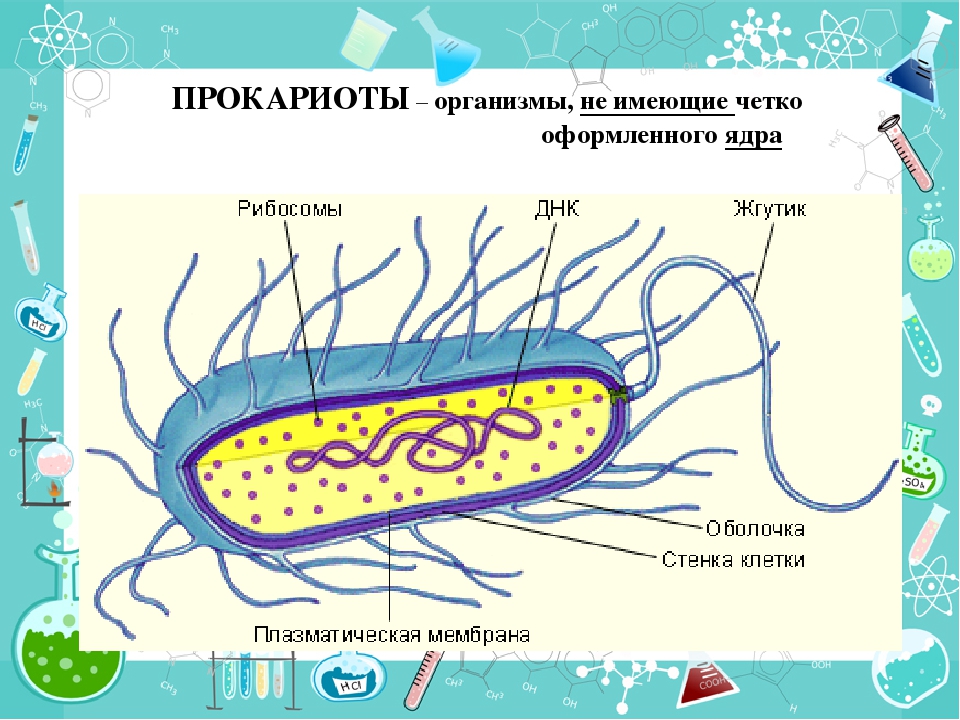 При возврате части эукариот к осмотрофному питанию клеточная стенка появилась вновь, но уже на другой биохимической основе.
При возврате части эукариот к осмотрофному питанию клеточная стенка появилась вновь, но уже на другой биохимической основе.Обмен веществ у бактерий\ Metabolism in bacteria
Разнообразен и обмен веществ у бактерий.
Четыре типа питания среди фототрофных и хемотрофных бактерий /Four types of nutrition among phototrophic and chemotrophic bacteria
Выделяют четыре типа питания, и среди фототрофных и хемотрофных бактерий встречаются все / There are four types of food, and among bacteria there are all.
| фотоавтотрофные | photoautotrophic | фототрофные используют энергию солнечного света | phototrophic use of sunlight energy |
| фотогетеротрофные | photoheterotrophic | ||
| хемоавтотрофные | chemoautotrophic | хемотрофные используют химическую энергию | chemotrophic use chemical energy |
| хемогетеротрофные | chemoheterotrophic |
Эукариоты же либо сами синтезируют энергию из солнечного света, либо используют готовую энергию такого происхождения.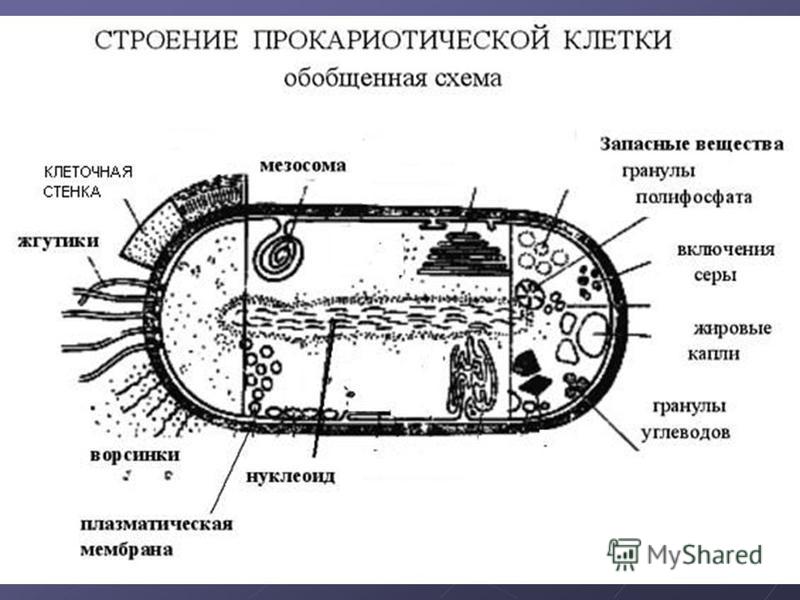 Это может быть связано с появлением среди эукариотов хищников, необходимость синтезировать энергию для которых отпала.
Это может быть связано с появлением среди эукариотов хищников, необходимость синтезировать энергию для которых отпала.
Ещё одно отличие — строение жгутиков. У бактерий жгутиками являются полые нити диаметром 15—20 нм из белка флагеллина. Строение жгутиков эукариот гораздо сложнее. Они представляют собой вырост клетки, окруженный мембраной, и содержат цитоскелет (аксонему) из девяти пар периферических микротрубочек и двух микротрубочек в центре. В отличие от вращающихся прокариотических жгутиков жгутики эукариот изгибаются или извиваются.
Две группы рассматриваемых нами организмов, как уже было сказано, сильно отличаются и по своим средним размерам. Диаметр прокариотической клетки составляет обычно 0,5—10 мкм, когда тот же показатель у эукариот составляет 10—100 мкм. Объём такой клетки в 1000—10 000 раз больше, чем прокариотической.
Рибосомы прокариот мелкие (70S-типа). Клетки эукариот содержат как более крупные рибосомы 80S-типа, находящиеся в цитоплазме, так и 70s-рибосомы прокариотного типа, расположенные в митохондриях и пластидах.
Различается время возникновения групп прокариот и эукариотических организмов / The time of occurrence of groups of prokaryotes and eukaryotic organisms differs
| около 3,5 млрд лет назад | в процессе эволюции возникли первые прокариоты | during the evolution of the first prokaryotes |
| около 1,2 млрд лет назад | от прокариот произошли эукариотические организмы | eukaryotic organisms originated from prokaryotes |
10.2 Что такое водоросли? | EGEE 439: Альтернативные виды топлива из источников биомассы
10.2 Что такое водоросли?
Водоросли — это эукариотические организмы, представляющие собой организмы, клетки которых содержат ядро и другие структуры (органеллы), заключенные в мембраны. Они живут во влажной среде, в основном водной, и содержат хлорофилл.
Водоросли — это не наземные растения, у которых есть 1) настоящие корни, стебли и листья, 2) сосудистые (проводящие) ткани, такие как ксилема и флоэма, и 3) отсутствие непродуктивных клеток в репродуктивных структурах.Водоросли — это не цианобактерии. Цианобактерии — это прокариоты, у которых отсутствуют мембраносвязанные органеллы и есть одна кольцевая хромосома. Рисунок 10.1a показывает клеточный состав синих водорослей, а 1b показывает микрофотографию клеток. Стенка клетки имеет студенистую оболочку. Прямо под клеточной стенкой находится плазматическая мембрана. Внутри клетки есть слои фикобилисом, фотосинтетические ламели, рибосомы, белковые гранулы и кольцевая ДНК, известная как нуклеоиды. Это типичные компоненты выращиваемых растений, однако нас интересуют липидные капли — масла, которые можно извлечь из водорослей.
Рисунок 10.1a: Клеточная структура синих водорослей.Щелкните, чтобы увидеть текстовое описание рисунка 10.1a.
Клеточная структура синих водорослей, включая жидкую каплю, нуклеоид (кольцевую ДНК), белковые гранулы, рибосому, фотосинтетические ламеллы, фикобилисомы (цианосомы), плазматическую мембрану, клеточную стенку, студенистую оболочку
Рисунок 10. 1b: Микрофотография синих водорослей.
1b: Микрофотография синих водорослей.
Водоросли состоят из ~ 50% углерода, 10% азота и 2% фосфора. В таблице 10.3 показан состав различных водорослей с учетом процентного содержания белка, углеводов, липидов и нуклеиновой кислоты.
| Виды | Белок | Углеводы | Липиды | Нуклеиновая кислота |
|---|---|---|---|---|
| Scenedesmus obliquus (зеленая водоросль) | 50-56 | 10-17 | 12-14 | 3-6 |
| Scenedesmus quadricauda | 47 | – | 1.9 | – |
| Scenedesmus dimorphus | 8-18 | 21-52 | 16-40 | – |
| Chlamydomonas rheinhardii (зеленая водоросль) | 48 | 17 | 21 | – |
| Chlorella vulgaris (зеленая водоросль) | 51-58 | 12-17 | 14-22 | 4-5 |
| Chlorella pyrenoidosa | 57 | 26 | 2 | – |
Spirogyra sp. | 6-20 | 33-64 | 11–21 | – |
| Dunaliella bioculata | 49 | 4 | 8 | – |
| Dunaliella salina | 57 | 32 | 6 | – |
| Euglena gracilis | 39-61 | 14-18 | 14-20 | – |
| Prymnesium parvum | 28-45 | 25-33 | 22-38 | 1-2 |
| Tetraselmis maculata | 52 | 15 | 3 | – |
| Porphyridium cruentum (красная водоросль) | 28-39 | 40-57 | 9-14 < | – |
Итак, каковы характеристики водорослей?
1.Эукариотические организмы:
Как упоминалось выше, водоросли — это эукариотические организмы. Структура эукариота (типичная растительная клетка) показана на рисунке 10.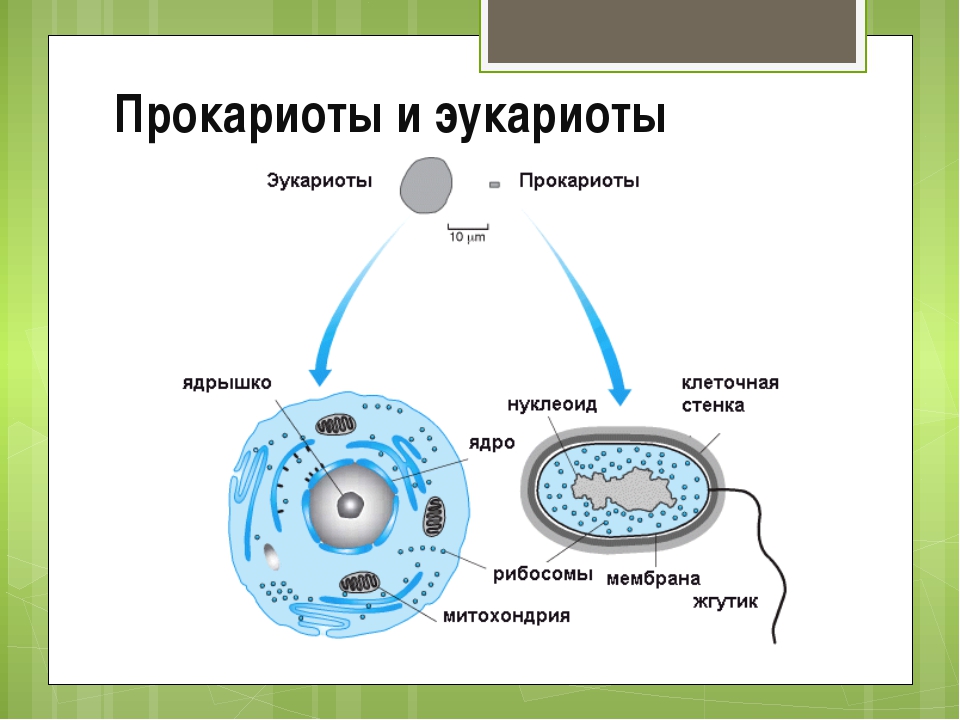 2а. Рисунок 10.2b показывает клеточную структуру прокариота, бактерии, одной из двух групп прокариотической жизни. Некоторые не считают прокариот настоящими водорослями, потому что они имеют другую структуру, но большинство включает их в семейство водорослей. Есть ярлыки для различных частей организмов, но я не буду требовать, чтобы вы знали эту информацию в деталях — они есть, так что, если у вас есть желание найти дополнительную информацию, вы можете.Таблица 10.4 показывает сравнение обоих этих типов клеток.
2а. Рисунок 10.2b показывает клеточную структуру прокариота, бактерии, одной из двух групп прокариотической жизни. Некоторые не считают прокариот настоящими водорослями, потому что они имеют другую структуру, но большинство включает их в семейство водорослей. Есть ярлыки для различных частей организмов, но я не буду требовать, чтобы вы знали эту информацию в деталях — они есть, так что, если у вас есть желание найти дополнительную информацию, вы можете.Таблица 10.4 показывает сравнение обоих этих типов клеток.
Щелкните, чтобы увидеть текстовое описание рисунка 10.2a.
Эукариотическая клетка включает в себя: ядро (ядерную пору, ядерную оболочку, ядрышко), грубую эндоплазматическую сеть, рибосомы, гладкую эндоплазматическую сеть, мелкие мембранозные пузырьки, нитчатый цитоскелет, плазмодесматы, плазматическую мембрану, клеточную стенку, хлоропластную (тилакоидную) мембрану зерна, крахмальную мембрану. , вакуоль (вакуоль, тонопласт), митохондрия (митохондрии), пероксисома, цитоплазма, везикулы Гольджи, тело Гольджи (аппарат Гольджи)
Рисунок 10. 2b: схематическая структура прокариот.
2b: схематическая структура прокариот.Щелкните, чтобы увидеть текстовое описание рисунка 10.2b.
Клетка прокариот содержит: нуклеоид (кольцевую ДНК), рибосомы, плазмиду, цитоплазму, плазматическую мембрану, клеточную стенку, капсулу, пили, бактериальный жгутик
| – | Эукариотические клетки | Прокариотические клетки |
|---|---|---|
| Размер | Довольно большой размер | Очень минутный размер |
| Ядерный регион | Ядерные материалы в мембране | Ядерная область (нуклеоид), не окруженная ядерной мембраной |
| Хромосома | Присутствует более одной хромосомы | Присутствует одна хромосома |
| Мембрана | Наличие мембраносвязанных клеточных органелл | Органеллы мембраносвязанных клеток отсутствуют |
2.
 Жить во влажной среде
Жить во влажной средеУ этих организмов отсутствует восковая кутикула (воск у наземных растений предотвращает потерю воды). Существует множество сред для выращивания водорослей. Типичные условия для водорослей — влажные тропические регионы, они могут расти как в морской, так и в пресной воде. Пресноводные водоросли растут в животных, водных растениях, фермерских дамбах, сточных водах, озерах, реках, лагунах, снеге, грязи / песке и почве.
3. Содержат хлорофилл
Водоросли, как и растения, в основном фотосинтезируют.У них есть пять видов фотосинтетических пигментов (хлорофилл a, b, c, d и f) и много дополнительных пигментов: синий, красный, коричневый и золотой. Хлорофилл — это зеленый пигмент, который содержится почти во всех растительных водорослях и цианобактериях. Он поглощает свет и передает световую энергию АТФ (аденозинтрифосфат).
Так как же классифицируются водоросли?
Водоросли принадлежат королевству протистов. На рис. 10.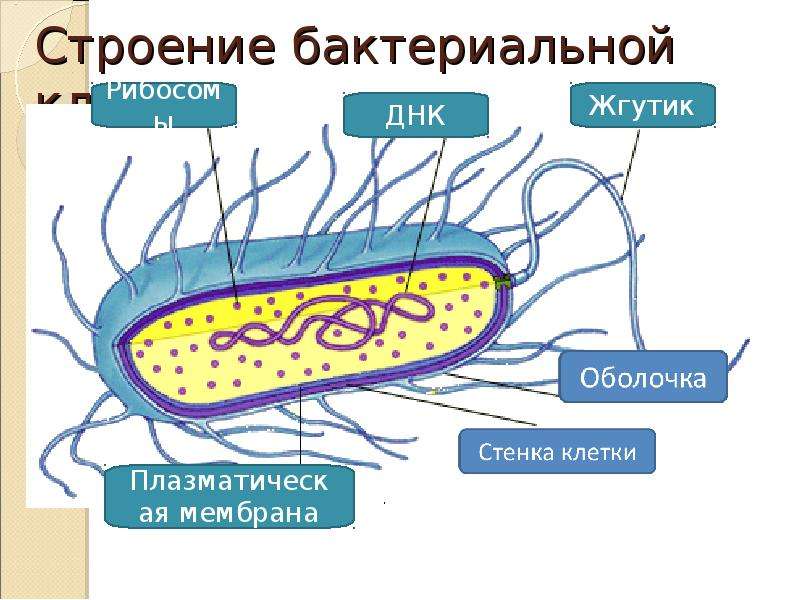 3 схематически показано, как Protista соответствует другим классификациям Plantae, Animalia, Fungi, Eubacteria и Archaebacteria.
3 схематически показано, как Protista соответствует другим классификациям Plantae, Animalia, Fungi, Eubacteria и Archaebacteria.
Водоросли также можно классифицировать по содержанию хлорофилла. Первый тип — хромиста. Эти типы водорослей содержат хлорофиллы а и с, и примеры водорослей включают бурые водоросли (золотисто-коричневые водоросли), ламинарии и диатомеи. Эти материалы являются подразделением Phaeophyta . Эти виды обитают на скалистых побережьях в зонах умеренного климата или в открытом море (холодные воды). Строения многоклеточные и могут вырастать до 50 м в длину.
Рисунок 10.3: Различные царства жизни.Щелкните, чтобы увидеть текстовое описание рисунка 10.3.
Эубактерии (одноклеточные, прокариотические)
Архебактерии (одноклеточные, прокариотические)
Протиста (эукариотический, одноклеточный и многоклеточный)
Plantae (многоклеточные, эукариотические)
Animalia (многоклеточные, эукариотические)
Грибы (многоклеточные, эукариотические)
Кредит: Халл (Google) [общественное достояние], через Wikimedia Commons
Рисунок 10.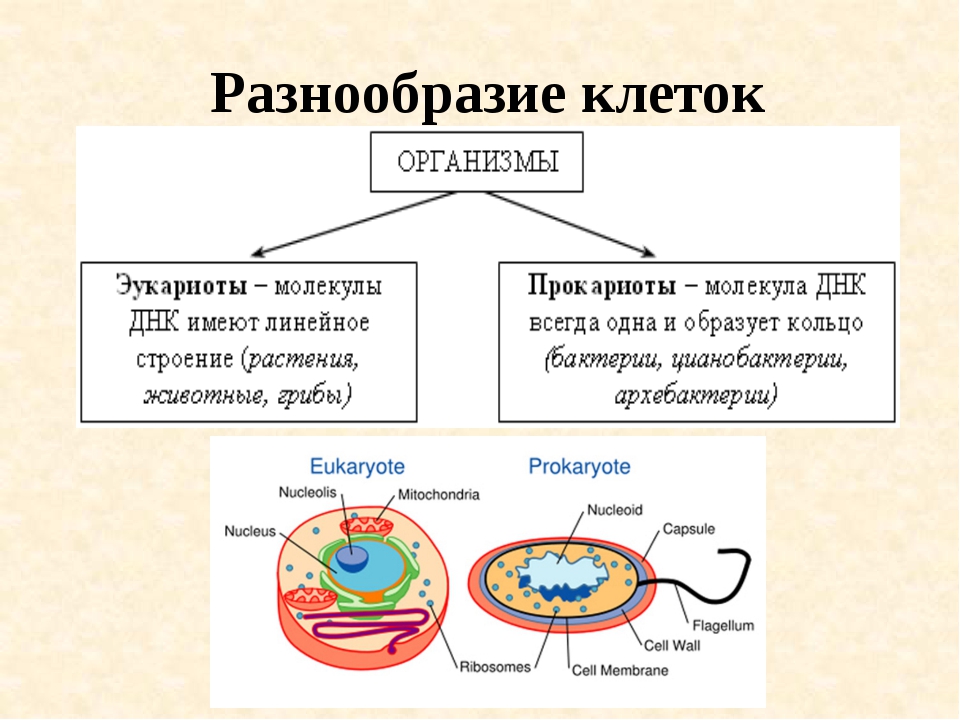 4: Филогенетическое дерево.
4: Филогенетическое дерево.
Красные водоросли относятся к другому типу и содержат хлорофилл а, например морские водоросли (морские водоросли). Эти организмы входят в подразделение Rhodophyta , насчитывающее более 4000 видов. Это одни из самых старых эукариотических организмов на Земле (есть окаменелости возрастом 2 миллиарда лет). Их много в тропических теплых водах. Они служат пищей и средой обитания для многих морских видов. Структура варьируется от тонких пленок до сложных нитевидных мембран. Эти водоросли имеют дополнительные пигменты, а фикобилины (красные) маскируют хлорофилл а.На рисунке 10.5b показаны различные красные водоросли. Динофлагелляты — одноклеточные протисты, которые связаны с красным приливом и биолюминесценцией.
Рисунок 10.5a: Келп
Кредит: Модуль A3 BEEMS
Зеленые водоросли содержат хлорофиллы a и b. Они находятся в отделе Chlorophyta. Это самая большая и разнообразная группа водорослей. Он встречается в основном в пресных водах, а также на суше (камни, деревья и почва).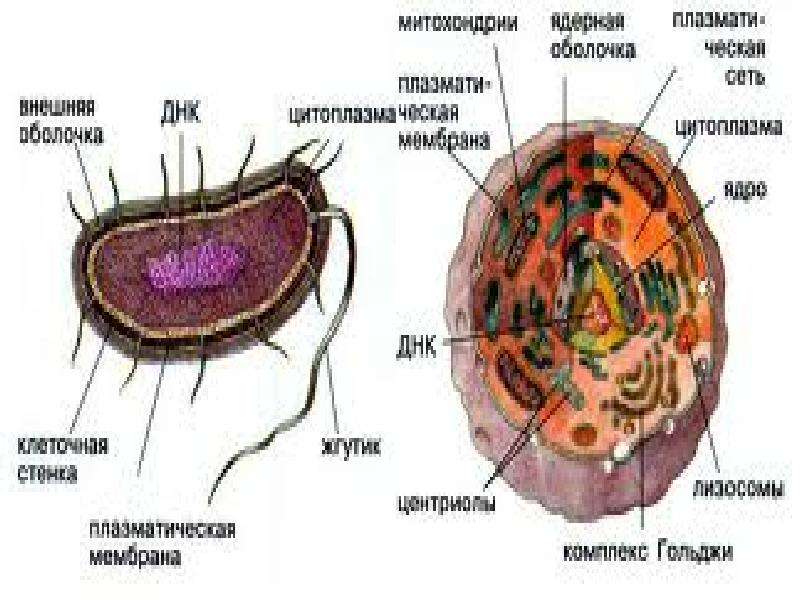 Структуры представляют собой одиночные клетки ( Micrasterias ), нитчатые водоросли, колонии ( Volvox ) и листообразную форму ( Thalli ).Наземные растения произошли от предка зеленых водорослей. Оба имеют одинаковые фотосинтетические пигменты (хлорофилл a и b). Некоторые зеленые водоросли имеют клеточную стенку из целлюлозы, похожую на наземные растения. На рисунке 5c показаны примеры зеленых водорослей.
Структуры представляют собой одиночные клетки ( Micrasterias ), нитчатые водоросли, колонии ( Volvox ) и листообразную форму ( Thalli ).Наземные растения произошли от предка зеленых водорослей. Оба имеют одинаковые фотосинтетические пигменты (хлорофилл a и b). Некоторые зеленые водоросли имеют клеточную стенку из целлюлозы, похожую на наземные растения. На рисунке 5c показаны примеры зеленых водорослей.
Рис. 10.5b: Фото и микрофотографии красных водорослей.
Кредит: Модуль A3 BEEMS
Рисунок 10.5c: Примеры зеленых водорослей.
Кредит: Модуль A3 BEEMS
Краткие сведения о прокариотических и эукариотических водорослях
Краткие сведения о прокариотических и эукариотических водорослях!
Есть два основных образца клеточной организации водорослей.Они бывают прокариотическими и эукариотическими.
1. Прокариотические водоросли: Сине-зеленые водоросли (Cyanophyceae или Cyanophycophyta) являются прокариотическими водорослями.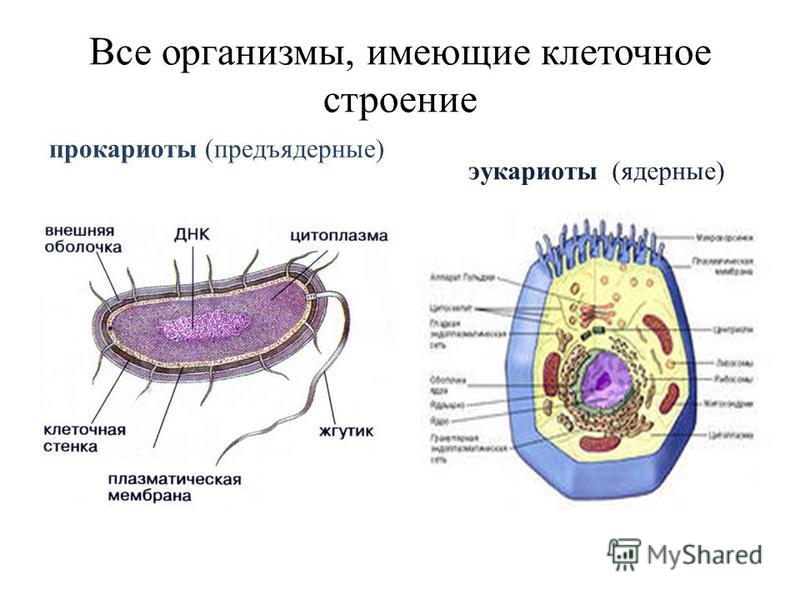 В этих водорослях их ядерный материал, дезоксирибонуклеиновая кислота (ДНК), не отделен от остальной части протоплазмы ядерной мембраной, а, скорее, в некоторой степени рассредоточен по всей клетке. Мембранные пластиды отсутствуют; фотосинтетические ламели свободно располагаются в цитоплазме.
В этих водорослях их ядерный материал, дезоксирибонуклеиновая кислота (ДНК), не отделен от остальной части протоплазмы ядерной мембраной, а, скорее, в некоторой степени рассредоточен по всей клетке. Мембранные пластиды отсутствуют; фотосинтетические ламели свободно располагаются в цитоплазме.
Изображение предоставлено: planetforward.org / sites / default / files / pond-algae4.jpg
Эндоплазматический ретикулум, митохондрии и аппарат Гольджи также отсутствуют. Большие водные вакуоли, подобные тем, которые встречаются у многих зеленых водорослей (Chlorophyceae или Chlorophycophyta), отсутствуют в клетках сине-зеленых водорослей. Кроме того, клеточные стенки сине-зеленых водорослей демонстрируют некоторое химическое сходство со стенками бактерий. Некоторые сине-зеленые водоросли могут быть инфицированы вирусами, которые напоминают бактериофаги, выступающие за дальнейшее сходство сине-зеленых водорослей и бактерий, поэтому их называют цианобактериями.
2.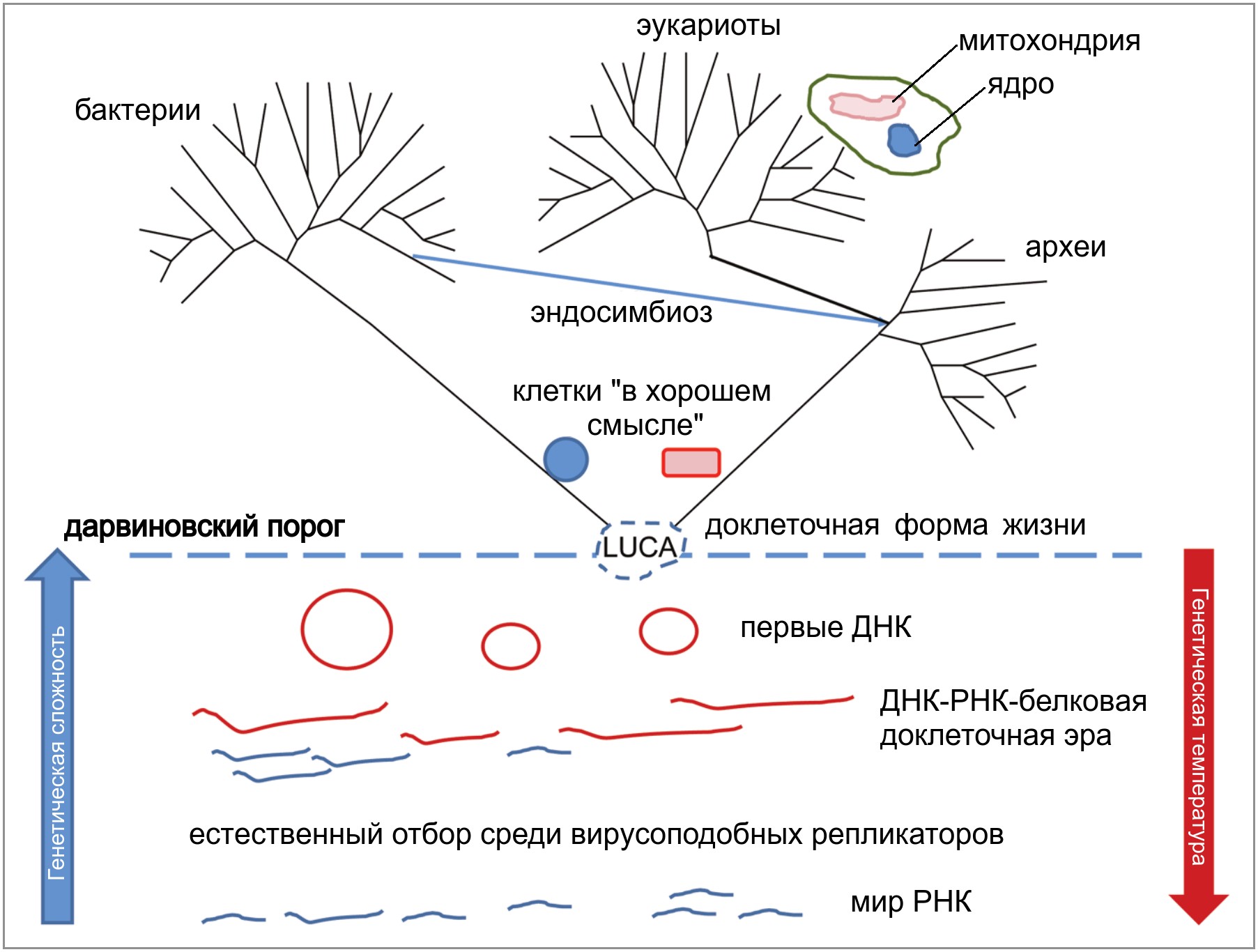 Эукариотические водоросли:
Эукариотические водоросли: С другой стороны, клетки эукариотических водорослей имеют ДНК, локализованную в тонко перфорированной ядерной мембране, причем их ядра по существу аналогичны ядрам высших растений. Эти фотосинтетические ламеллы заключены в мембраны в виде четко определенных хлоропластов. Хлоропласты могут быть массивными, париетальными или звездообразными структурами, встречающимися по отдельности внутри каждой клетки, или они могут быть ленточными, стержневидными, сетчатыми или дискоидными структурами, как у зеленых растений, не относящихся к водорослям.
У многих зеленых водорослей (Chlorophyceae), золотых водорослей (Chrysophyceae), бурых водорослей (Phaeophyceae) и красных водорослей (Rhodophyceae) в пластидах присутствуют специализированные области, пиреноиды. В зеленых водорослях пиреноиды обычно окружены крахмальными зернами. Считается, что пиреноиды являются центрами ферментативной конденсации глюкозы в крахмал.
Эндоплазматический ретикулум, митохондрии и тельца Гольджи присутствуют в клетках всех изученных эукариотических водорослей.
Подвижные клетки водорослей могут быть жгутиковидными. С подвижностью связаны такие структуры, как сократительные вакуоли, жгутики и рыльца.
клеток прокариотических водорослей: функция, определение и особенности — класс биологии
Что такое сине-зеленые водоросли?
Сине-зеленые водоросли — это общее название, используемое для типа (категории) бактерий, известных как Cyanobacteria . Чтобы запомнить научное название, просто напомните себе, что голубой — это сине-зеленый оттенок, используемый для описания цвета бактерий.Часть общего названия водоросли может вводить в заблуждение, поскольку большинство водорослей считаются растениями, которые классифицируются как эукариотические организмы. Но, как было сказано ранее, цианобактерии являются прокариотами, поэтому их структура сильно отличается от структуры растений.
Цианобактерии — удивительные существа (да, они живые; помните, это бактерии, то есть живой организм). Они встречаются почти во всех возможных средах или средах обитания, о которых мы знаем — от самых жарких пустынь до самых глубоких океанов и самых высоких гор.Цианобактерии чрезвычайно разнообразны и приспособились жить практически где угодно.
Они похожи на растения в том, что они проходят фотосинтез, хотя и совсем другим образом. У цианобактерий отсутствуют хлоропласты — мембранные органеллы, содержащие хлорофилл, ответственный за фотосинтез растений. Однако цианобактерии используют другие пигменты, такие как фикоцианин , , для процесса фотосинтеза. Видите ли, цианобактерии на самом деле берут электрон из воды или сероводорода, чтобы использовать фикоцианин для захвата солнечного света.
Затем они используют углекислый газ, электроны из воды (или сероводорода) и фикоцианин для производства сахаров для себя. У растений этот процесс происходит в хлоропластах, но у цианобактерий нет хлоропластов, поэтому вместо этого процесс происходит в некоторых складках бактерий. Если мы посмотрим на фотосинтез в растениях, мы обнаружим в хлоропластах другую органеллу, называемую тилакоидом, которая содержит хлорофилл. Здесь же, в тилакоидной мембране, находятся фотоситемы I и II.Здесь происходят реакции фотосинтеза.
Возвращаясь к цианобактериям, мы обнаруживаем, что организмы объединили фотосистемы I и II, чтобы работать через фотосинтез. Многие из этих цианобактерий все еще имеют тилакоиды, поэтому здесь происходит фотосинтетический процесс. Процесс фотосинтеза все еще в чем-то похож на таковой у растений, за исключением того, что все он происходит в одной фотосистеме, а не в двух.
Азотфиксация
При фотосинтезе организм поглощает и использует углекислый газ в качестве одного из своих компонентов.Цианобактерии обладают способностью впитывать азот, а также углекислый газ для использования в обработке и приготовлении пищи.
Для «фиксации азота» (полной азотфиксации) у цианобактерий есть специализированные клетки, называемые гетероцистами . Гетероцисты специально предназначены для фиксации азота.Именно в этих структурах содержится нитрогеназа. Помните, что все, что имеет суффикс «аза», обычно считается способным расщепить первую часть слова, поэтому задача нитрогеназы — расщеплять азот (отсюда азот + азе = нитрогеназа). Цианобактерии способны преобразовывать газообразный азот в аммиак, нитраты и нитриты, которые жизненно важны для роста растений.
Именно для этой азотфиксации мы чаще всего используем цианобактерии. Например, мы используем их в сельском хозяйстве, чтобы помочь вернуть азот в почву, тем самым позволяя нашим культурам расти лучше.
Структура клеток цианобактерий
Цианобактерии напоминают нечто среднее между клетками животных и клетками растений. У них есть клеточная стенка, но нет мембран. У них есть белковые оболочки, которые покрывают любые внутренние структуры, а также защищают саму клетку. Внутри у них есть ДНК, но ее нет ни в каком ядре.
Цианобактерии технически не могут двигаться, но могут распространяться с помощью гормогонии , которые представляют собой пучки нитей, которые отростки или ответвляются от основного тела и перемещаются, создавая новые клетки в другом месте.Это создает своего рода сеть клеток цианобактерий.
Краткое содержание урока
Цианобактерии — это прокариотические клетки, в которых отсутствуют мембраносвязанные органеллы и ядра. Их общее название — сине-зеленые водоросли из-за их сине-зеленого цвета, вызванного их пигментом фикоцианином. Большинство водорослей считаются растениями, но сине-зеленые водоросли — бактериями. Они могут фотосинтезировать, как растения, но используют воду и сероводород для захвата электронов. Они также могут «фиксировать» азот, то есть поглощать азот и преобразовывать его в полезную энергию.Мы часто используем сине-зеленые водоросли из-за их способности фиксировать азот в сельском хозяйстве и других практиках.
Истоки жизни на Земле
Все, что мы знаем о жизни, исходит из выборки размером один: жизнь здесь, на Земле. И дело в том, что мы действительно не знаем точно, как жизнь возникла из неорганической материи все эти миллиарды лет назад… и кто может сказать, что на разных планетах во Вселенной не могло быть разных процессов? Возможно, есть еще вопросы, которые нужно задать, больше возможностей для рассмотрения.Но давайте начнем с того, что мы знаем о некоторых из самых первых живых существ на Земле.
Цианобактерии, также известные как сине-зеленые водоросли, появились на Земле довольно давно. Возможные примеры окаменелостей были найдены в породах, возраст которых составляет около 3500 миллионов лет, в Западной Австралии.
Хотя цианобактерии обычно называют сине-зелеными водорослями, на самом деле это не водоросли. Цианобактерии и бактерии в целом являются прокариотическими формами жизни. По сути, это означает, что их клетки не имеют органелл (крошечных структур внутри клеток, которые выполняют определенные функции) и не имеют отдельных ядер — их генетический материал смешивается с остальной частью клетки.Эта характеристика характерна для бактерий и архей; все другие формы жизни на Земле, включая настоящие водоросли, состоят из эукариотических клеток с органеллами и генетическим материалом, содержащимся в одном месте (ядре).
Полосатый рисунок масс цианобактерий (сине-зеленых водорослей). Изображение адаптировано из: Bobby McKay; CC BY ND 2.0Бактерии (и археи) — выносливые существа. Они процветают в горячей, холодной, соленой, кислой и щелочной среде, в которой погибнет большинство эукариот.Несмотря на это, у них плохой имидж: ведь бактерии вызывают у человека множество болезней.
Но без них нас, возможно, вообще не было бы. Прокариоты были самыми ранними формами жизни, простыми существами, питавшимися соединениями углерода, которые накапливались в ранних океанах Земли. Постепенно появились другие организмы, которые использовали энергию Солнца вместе с такими соединениями, как сульфиды, для выработки собственной энергии. Затем цианобактерии пошли еще дальше: они начали использовать воду во время фотосинтеза, выделяя кислород в качестве побочного продукта.Со временем в атмосфере Земли накопилось достаточно кислорода, чтобы обеспечить эволюцию организмов, метаболизирующих кислород.
Когда мы выходим во Вселенную в поисках жизни за пределами нашей родной планеты, мы думаем, что, скорее всего, найдем ее где-нибудь, где есть вода …Но мы можем быть обязаны бактериям больше, чем воздухом, которым мы дышим. Вероятно, что эукариотические клетки, из которых состоит человек, произошли от бактерий около двух миллиардов лет назад. Одна из теорий состоит в том, что эукариотические клетки эволюционировали посредством симбиотических отношений между двумя независимыми прокариотическими бактериями.Одна бактерия была поглощена другой, а меньшая клетка продолжала существовать внутри другой, что было выгодно обеим. Они эволюционировали, чтобы стать более развитой эукариотической клеткой с ее заключенным в мембрану ядром.
Независимо от того, как это произошло, эволюция эукариотических клеток стала важной вехой в истории жизни на Земле. По мере того, как условия становились более благоприятными, начали развиваться более сложные организмы.
Одно из самых старых свидетельств существования жизни на Земле — 3 года.Окаменелые остатки микробных матов возрастом 49 миллиардов лет, которые выглядят как следы морщин на скалах, найдены в регионе Пилбара в Западной Австралии. Также в районе Пилбара обнаружены окаменелые остатки строматолитов. Это также похожие на циновки структуры микробов, которые живут в мелководной морской среде и все еще существуют сегодня. Песок скапливается поверх микробных матов, и микробы поднимаются к поверхности, чтобы снова попасть на свет, образуя характерные слои выпуклой формы, которые в конечном итоге затвердевают в камни.
Строматолиты в бассейне Хамелин, залив Шарк, Западная Австралия, Изображение адаптировано из: Пол Моррис; CC BY SA 2.0Когда мы ищем жизнь за пределами нашей родной планеты в другом месте Вселенной, мы думаем, что, скорее всего, найдем ее где-то там, где есть вода и где есть источник тепла, согревающий воду. Хотя мы знаем, что некоторые живые существа процветают в более экстремальных условиях, сочетание тепла и воды, по-видимому, является наиболее вероятным требованием для создания среды, способной поддерживать какую-то жизнь — по крайней мере, формы жизни, подобные тем, что мы делаем. найти на Земле.Но кто знает, какие еще виды живых существ могут существовать?
Эта статья была адаптирована из материалов веб-сайта Академии и проверена следующим экспертом: Профессор Малькольм Уолтер AM FAA Австралийский центр астробиологии, Школа биологических наук о Земле и окружающей среде, UNSW Сидней
Водоросли | Структура клетки | Прокариотические и эукариотические | Ядро | Defferences
В водорослях есть два основных типа клеток: прокариотических и эукариотических .В прокариотических клетках отсутствуют связанные с мембраной органеллы (пластиды, митохондрии, ядра, тельца Гольджи и флаги), и они встречаются у цианобактерий. Остальные водоросли являются эукариотическими и имеют органеллы.
Структура клеток цианобактерий
Бактериальная клетка: прокариотическая
Эукариотическая клетка часто окружена клеточной стенкой , состоящей из полисахаридов, которые частично продуцируются и секретируются тельцами Гольджи. Плазматическая мембрана (плазмалемма , ) окружает оставшуюся часть клетки; эта мембрана представляет собой живую структуру, отвечающую за контроль притока и оттока веществ в протоплазме . Локомоторных органы, жгутиков, передвижения клетка через среду по их избиению. Жгутики заключены в плазматическую мембрану и имеют определенное количество и ориентацию микротрубочек, . Ядро , , которое содержит генетический материал клетки, окружено двойной мембраной с порами в ней. Содержимое ядра — ядрышко , хромосомы и фоновый материал или кариолимфа. Хлоропласты имеют мембранные мешочки, называемые тилакоидами , которые осуществляют световые реакции фотосинтеза.Тилакоиды встраиваются в строму, где происходят темные реакции фиксации углерода. В строме есть небольшие рибосомы 70S, ДНК и в некоторых случаях продукт хранения. Хлоропласты окружены двумя мембранами оболочки хлоропласта. Иногда хлоропласты имеют плотную белковую зону, пиреноид , , которая связана с образованием продуктов хранения.
Митохондрии с двойной мембраной содержат 70S рибосомы и ДНК и содержат дыхательный аппарат.Тело Гольджи состоит из ряда мембранных мешочков, называемых цистернами , уложенных друг на друга. Тело Гольджи участвует в производстве и секреции полисахаридов. Цитоплазма также содержит большие 80S рибосомы и липидные тельца.
Структурные характеристики сине-зеленых водорослей
Сине-зеленые водоросли, самые примитивные организмы в мире растений, на самом деле не являются «настоящими» водорослями. По своей структуре они больше похожи на бактерии, и на самом деле они классифицируются как цианобактерии, большая группа в основном фототрофных бактерий.Клетки цианобактерий одноклеточные и поэтому имеют более простую структуру, чем многоклеточные эукариотические клетки растений и животных.
Цианобактерии Определение
Цианобактерии — это прокариотические оксигенные фототрофы, которые содержат зеленый пигмент, называемый хлорофиллом, и синий фотосинтетический пигмент, называемый фикобилинами. Прокариотика означает, что у них нет мембраносвязанного ядра, митохондрий или других мембраносвязанных органелл (как у настоящих водорослей). Фототроф — это организм, который использует энергию солнца для синтеза органических соединений для еды.
Структура цианобактерий
Клетки цианобактерий, размер которых обычно составляет от одной десятой до одной двадцатой размера эукариотических клеток, имеют округлую форму.
Типичная клетка цианобактерии состоит из внешнего клеточного покрытия, цитоплазмы и нуклеинового материала. Наружное клеточное покрытие состоит из слизистого слоя, который защищает клетку от факторов окружающей среды, сложной многослойной клеточной стенки, состоящей из полисахаридов и мукопептидов, и внутренней живой плазматической мембраны.Это основы строения цианобактерий.
Цитоплазма имеет пигментированные пластинки (мембранные складки) по периферии, происходящие из плазматической мембраны. Пигменты включают хлорофиллы, каротины, ксантофиллы, ц-фикоэритрин и ц-фикоцианин. C-фикоэритрин и c-фикоцианин уникальны для сине-зеленых водорослей.
Нуклеоплазма, в которой расположена ДНК, состоит из множества нитевидных волокон или нитей и находится в центре клетки. Нет ядерной границы или ядрышка.Нуклеоплазматический материал, разбросанный по клетке, в процессе деления клетки разделяется на две части.
В то время как клетки цианобактерий не имеют органелл, таких как митохондрии, хлоропласты, эндоплазматический ретикулум или аппарат Гольджи, которые все присутствуют в эукариотических клетках, они обе имеют рибосомы. Рибосомы содержат РНК (рибонуклеиновую кислоту) и отвечают за синтез белка. Рибосомы в клетках цианобактерий примерно на треть меньше, чем рибосомы в эукариотических клетках, но они выполняют аналогичные функции.
Характеристики цианобактерий
Определяющими характеристиками цианобактерий являются устойчивость к экстремальным условиям и способность существовать без витаминов. В качестве источника азота они используют фосфор, железо и другие питательные микроэлементы, а также аммиак или нитрат. Некоторые виды цианобактерий нитчатые и не нуждаются в солнечном свете. Вместо этого они растут в темноте, полагаясь на сахар из глюкозы или сахарозы в качестве источника углерода и энергии.
Цианобактерии не размножаются путем митоза, в отличие от эукариотических клеток.Клетка цианобактерий удлиняется, и ДНК реплицируется. Хромосома разделяется, и одна клетка разделяется на две клетки в процессе, называемом бинарным делением.
Эукариотическое происхождение | Безграничная биология
Ранние эукариоты
Протисты — это эукариоты, впервые появившиеся примерно 2 миллиарда лет назад с повышением уровня кислорода в атмосфере.
Цели обучения
Обсудить происхождение эукариот с точки зрения геологической временной шкалы
Ключевые выводы
Ключевые моменты
- На геологической временной шкале простейшие являются одними из первых организмов, которые эволюционировали после прокариот.
- Современные эукариоты произошли от общего предка со следующими особенностями: ядро, разделенное митозом, ДНК, связанная с гистонами, цитоскелет и эндомембранная система, способность создавать реснички / жгутики.
- Протисты сильно различаются по размеру: от отдельных клеток размером примерно 10 мкм до многоклеточных водорослей, которые видны невооруженным глазом.
Ключевые термины
- цианобактерии : фотосинтезирующие прокариотические микроорганизмы типа Cyanobacteria, когда-то известные как сине-зеленые водоросли
- аэробный : живет или встречается только в присутствии кислорода
- эндомембрана : все мембранные компоненты внутри эукариотической клетки, включая ядерную оболочку, эндопластический ретикулум и аппарат Гольджи
Происхождение эукариот
Люди были знакомы с макроскопическими организмами (организмами, достаточно большими, чтобы их можно было видеть невооруженным глазом) еще до того, как появилась письменная история.Вероятно, что большинство культур различали животных и наземные растения, но наиболее вероятно, что макроскопические грибы были включены в состав растений. Таким образом, с появлением микроскопов несколько столетий назад стало интересно иметь дело с миром микроорганизмов. За последние пару столетий использовалось множество различных схем именования, но наиболее распространенной практикой стало называть эукариот, не являющихся наземными растениями, животными или грибами, протистами.
Большинство протистов — микроскопические одноклеточные организмы, которые в изобилии встречаются в почве, пресной воде, солоноватой и морской среде.Они также распространены в пищеварительном тракте животных и в тканях сосудов растений. Другие вторгаются в клетки других простейших, животных и растений. Не все протисты микроскопические. Некоторые из них имеют огромные макроскопические клетки, такие как плазмодии (гигантские амебы) миксомицетной слизистой плесени или морская зеленая водоросль Caulerpa , которые могут иметь отдельные клетки, которые могут достигать размеров нескольких метров. Некоторые простейшие являются многоклеточными, например, красные, зеленые и коричневые водоросли. Именно среди протистов можно найти множество способов, которыми могут развиваться организмы.Они являются одними из первых организмов, эволюционировавших с появлением эукариот.
Эукариоты в геологической временной шкале
Самым древним ископаемым свидетельствам эукариот с клетками размером 10 мкм и более около 2 миллиардов лет. Все окаменелости старше этого, по-видимому, являются прокариотами. Вероятно, что современные эукариоты произошли от предка, имевшего прокариотическую клеточную организацию. Последний общий предок (LCA) сегодняшней Eukarya имел несколько характеристик, которые включали: клетки с ядрами, которые делятся митотически и содержат линейные хромосомы, в которых ДНК связана с гистонами; цитоскелет и эндомембранная система; и способность создавать реснички / жгутики, по крайней мере, в течение части своего жизненного цикла.LCA была аэробной, потому что в ней были митохондрии, которые были результатом действия аэробных альфа-протеобактерий, которые жили внутри клетки-хозяина. Было ли у этого хозяина ядро во время начального симбиоза, остается неизвестным. У LCA могла быть клеточная стенка, по крайней мере, в течение части своего жизненного цикла, но для подтверждения этой гипотезы необходимы дополнительные данные. Современные эукариоты очень разнообразны по форме, организации, жизненным циклам и количеству клеток на человека.
В то время как сегодняшняя атмосфера на одну пятую состоит из молекулярного кислорода (O 2 ), геологические данные показывают, что изначально в нем не было O 2 .Без кислорода нельзя было бы ожидать аэробного дыхания; вместо этого живые существа полагались бы на ферментацию. В какой-то момент, примерно 3,5 миллиарда лет назад, некоторые прокариоты развили способность к фотосинтезу. Цианобактерии использовали воду в качестве источника водорода и выпустили O 2 в качестве побочного продукта. Первоначально богатая кислородом среда, вероятно, была локализована вокруг мест, где были активны цианобактерии, но примерно 2 миллиарда лет назад геологические данные показывают, что концентрация кислорода в атмосфере увеличивалась до более высоких.Уровни кислорода, аналогичные сегодняшним, возникли только за последние 700 миллионов лет. Напомним, что первые окаменелости, которые, как мы полагаем, являются эукариотами, датируются примерно 2 миллиардами лет, поэтому они появились при повышении уровня кислорода.
Разновидности протистов. на сотни футов в подводных «лесах».«Характеристики эукариотической ДНК
Эукариоты, вероятно, произошедшие от прокариот, имеют более сложные черты как в организации клеток, так и в организации ДНК.
Цели обучения
Сравнить и сопоставить прокариотическую ДНК с эукариотической ДНК
Ключевые выводы
Ключевые моменты
- Прокариотическая геномная ДНК прикрепляется к плазматической мембране в виде нуклеоида, в отличие от эукариотической ДНК, которая расположена в ядре.
- Эукариотическая ДНК линейна, уплотнена в хромосомы гистонами и имеет теломеры на каждом конце для защиты от порчи.
- Прокариоты содержат кольцевую ДНК в дополнение к более мелким, трансферабельным ДНК-плазмидам.
- Эукариотические клетки содержат митохондриальную ДНК в дополнение к ядерной ДНК.
- Эукариоты разделяют реплицированные хромосомы митозом, используя цитоскелетные белки, тогда как прокариоты делятся проще посредством бинарного деления.
Ключевые термины
- теломеры : любая из повторяющихся нуклеотидных последовательностей на каждом конце эукариотической хромосомы, которые защищают хромосому от деградации
- плазмида : круг из двухцепочечной ДНК, отделенный от хромосом, который встречается у бактерий и простейших
Характеристики ДНК эукариот по сравнению с ДНК прокариот
Известно, что прокариотические клетки намного менее сложны, чем эукариотические клетки, поскольку считается, что эукариотические клетки присутствуют на более позднем этапе эволюции.Вероятно, эукариотические клетки произошли от прокариотических клеток. Различия в сложности можно увидеть на клеточном уровне.
Единственная характеристика, которая необходима и достаточна для определения организма как эукариота, — это ядро, окруженное ядерной оболочкой с ядерными порами. У всех существующих эукариот есть клетки с ядрами; большая часть генетического материала эукариотической клетки содержится в ядре. Напротив, прокариотическая ДНК не содержится в ядре, а скорее прикреплена к плазматической мембране и содержится в форме нуклеоида, области неправильной формы, которая не окружена ядерной мембраной.
Клеточное расположение эукариотической и прокариотической ДНК : Эукариотическая ДНК хранится в ядре, тогда как прокариотическая ДНК находится в цитоплазме в форме нуклеоида.
Эукариотическая ДНК упакована в пучки хромосом, каждая из которых состоит из линейной молекулы ДНК, намотанной вокруг основных (щелочных) белков, называемых гистонами, которые скручивают ДНК в более компактную форму. Прокариотическая ДНК находится в кольцевой, нехромосомной форме. Кроме того, у прокариот есть плазмиды, которые представляют собой более мелкие фрагменты кольцевой ДНК, которые могут реплицироваться отдельно от геномной ДНК прокариот.Из-за линейной природы эукариотической ДНК повторяющиеся некодирующие последовательности ДНК, называемые теломерами, присутствуют на обоих концах хромосом в качестве защиты от разрушения.
Митоз, процесс деления ядра, при котором реплицированные хромосомы делятся и разделяются с использованием элементов цитоскелета, повсеместно присутствует у эукариот. Цитоскелет содержит структурные и подвижные компоненты, называемые актиновыми микрофиламентами и микротрубочками. Все современные эукариоты имеют эти цитоскелетные элементы.Прокариоты, с другой стороны, подвергаются бинарному делению в процессе репликации ДНК, затем разделения на два полюса клетки, и, наконец, клетка полностью делится.
Основное различие ДНК между эукариотами и прокариотами — это присутствие митохондриальной ДНК (мтДНК) у эукариот. Поскольку у эукариот есть митохондрии, а у прокариот их нет, эукариотические клетки содержат митохондриальную ДНК в дополнение к ДНК, содержащейся в ядре и рибосомах. МтДНК состоит из значительно меньшего количества пар оснований, чем ядерная ДНК, и кодирует всего несколько десятков генов, в зависимости от организма.
Эндосимбиоз и эволюция эукариот
Эукариоты могли быть продуктом одной клетки, поглощающей другую, и эволюционировали со временем, пока отдельные клетки не стали единым организмом.
Цели обучения
Опишите общую концепцию эндосимбиоза и эволюцию эукариот
Ключевые выводы
Ключевые моменты
- Эндосимбиоз — это концепция, когда одна клетка поглощает другую, и обе клетки получают выгоду от взаимосвязи.
- Эндосимбиоз был первоначально рассмотрен после наблюдения сходства между хлоропластами растений и свободноживущими цианобактериями.
- Пероксисомы, возможно, были первыми эндосимбионтами, вызванными увеличением количества атмосферного кислорода в тот момент геологического времени.
- Со временем эндосимбионты могли перенести часть своей ДНК в ядро хозяина, таким образом, становясь зависимыми от хозяина для выживания и завершения полной интеграции в единый организм.
Ключевые термины
- цианобактерии : фотосинтезирующие прокариотические микроорганизмы типа Cyanobacteria, когда-то известные как сине-зеленые водоросли
- пероксисома : эукариотическая органелла, являющаяся источником ферментов, которые катализируют производство и распад перекиси водорода и отвечают за окисление длинноцепочечных жирных кислот
- эндосимбионт : организм, который живет в теле или клетках другого организма
Эндосимбиоз и эволюция эукариот
Чтобы полностью понять эукариотические организмы, необходимо понимать, что все существующие эукариоты являются потомками химерного организма, который был составной частью клетки-хозяина и клетки (ов) альфа-протеобактерии, которая «поселилась» внутри хозяина. .Эта главная тема происхождения эукариот известна как эндосимбиоз, когда одна клетка поглощает другую, так что поглощенная клетка выживает, и обе клетки получают выгоду. На протяжении многих поколений симбиотические отношения могут привести к появлению двух организмов, которые настолько зависят друг от друга, что ни один из них не сможет выжить самостоятельно. Эндосимбиотические события, вероятно, способствовали возникновению последнего общего предка (LCA) современных эукариот.
Эндосимбиоз : Современные эукариотические клетки произошли от более примитивных клеток, которые поглотили бактерии с такими полезными свойствами, как производство энергии.Вместе некогда независимые организмы процветали и превратились в единый организм.
Теория эндосимбиотиков
Эндосимбиотическая теория была впервые сформулирована русским ботаником Константином Мерещковским в 1905 году. Мерешковский был знаком с работой ботаника Андреаса Шимпера, который в 1883 году заметил, что деление хлоропластов в зеленых растениях очень похоже на деление свободноживущих цианобактерий. Шимпер предварительно предположил, что зеленые растения возникли в результате симбиотического союза двух организмов.Иван Валлин распространил идею эндосимбиотического происхождения на митохондрии в 1920-х годах. Эти теории изначально отвергались или игнорировались. Более подробное электронно-микроскопическое сравнение цианобактерий и хлоропластов в сочетании с открытием того, что пластиды (органеллы, связанные с фотосинтезом) и митохондрии содержат свою собственную ДНК, привело к возрождению этой идеи в 1960-х годах. Теория эндосимбиотиков была выдвинута и подтверждена микробиологическими данными Линн Маргулис в 1967 году.
Хлоропласты в растениях : Эукариот с митохондриями поглотил цианобактерии в случае серийного первичного эндосимбиоза, создав линию клеток с обеими органеллами. Эти цианобактерии превратились в хлоропласты в современных клетках растений. У цианобактериального эндосимбионта уже была двойная мембрана.
В 1981 году она утверждала, что эукариотические клетки возникли как сообщества взаимодействующих сущностей, включая эндосимбиотические спирохеты, которые развились в эукариотические жгутики и реснички.Эта последняя идея не получила широкого признания, потому что жгутикам не хватает ДНК и они не проявляют ультраструктурного сходства с бактериями или археями. По словам Маргулиса и Дориона Сагана, «жизнь захватила земной шар не в битвах, а в сетях» (, то есть , благодаря сотрудничеству). Также рассматривалась возможность того, что органеллы пероксисом могут иметь эндосимбиотическое происхождение, хотя в них отсутствует ДНК. Кристиан де Дюв предположил, что они, возможно, были первыми эндосимбионтами, позволяющими клеткам выдерживать растущее количество свободного молекулярного кислорода в атмосфере Земли.Однако теперь выясняется, что они могут быть образованы de novo , что противоречит идее о симбиотическом происхождении.
Считается, что на протяжении тысячелетий эти эндосимбионты передали часть своей собственной ДНК ядру клетки-хозяина во время эволюционного перехода от симбиотического сообщества к установленной эукариотической клетке (так называемый «серийный эндосимбиоз»). Эта гипотеза считается возможной, потому что сегодня из научных наблюдений известно, что передача ДНК происходит между видами бактерий, даже если они не являются тесно связанными.Бактерии могут брать ДНК из своего окружения и иметь ограниченную способность включать ее в свой собственный геном.
Эволюция митохондрий
Митохондрии — это органеллы, производящие энергию, которые, как считается, когда-то были типом свободноживущих альфа-протеобактерий.
Цели обучения
Объясните связь между эндосимбиозом и митохондриями и эволюцией эукариот
Ключевые выводы
Ключевые моменты
- Эукариотические клетки содержат различное количество митохондрий, в зависимости от энергетических потребностей клеток.
- Митохондрии обладают многими особенностями, которые позволяют предположить, что они ранее были независимыми организмами, включая их собственную ДНК, клеточно-независимое деление и физические характеристики, аналогичные альфа-протеобактериям.
- Некоторые митохондриальные гены со временем перешли в ядерный геном, но митохондрии сохранили некоторый генетический материал по причинам, до конца не выясненным.
- Предполагаемый перенос генов из митохондрий в ядро клетки-хозяина, вероятно, объясняет, почему митохондрии не могут выжить вне клетки-хозяина.
Ключевые термины
- crista : cristae (единственная криста) — это внутренние компартменты, образованные внутренней мембраной митохондрии
- вакуоль : большой, связанный с мембраной, заполненный жидкостью отсек в цитоплазме клетки
- эндосимбиоз : когда один симбиотический вид попадает в цитоплазму другого симбиотического вида, и оба становятся эндосимбиотическими
Связь между эндосимбиозом и митохондриями
Одним из основных отличительных признаков прокариот от эукариот является наличие митохондрий.Эукариотические клетки содержат от одной до нескольких тысяч митохондрий, в зависимости от уровня потребления энергии клеткой. Каждая митохондрия имеет длину от 1 до 10 мкм и существует в клетке в виде органеллы, которая может быть яйцевидной, червеобразной или сложно разветвленной. Митохондрии возникают в результате разделения существующих митохондрий. Они могут сливаться вместе. Они перемещаются внутри клетки за счет взаимодействия с цитоскелетом. Однако митохондрии не могут выжить вне клетки. Поскольку количество кислорода в атмосфере увеличивалось миллиарды лет назад и по мере развития успешных аэробных прокариот, данные свидетельствуют о том, что предковая клетка с некоторой компартментализацией мембран поглотила свободноживущие аэробные прокариоты, в частности альфа-протеобактерии, тем самым дав клетке-хозяину способность использовать кислород для высвобождения энергии, хранящейся в питательных веществах.Альфа-протеобактерии — это большая группа бактерий, в которую входят виды, симбиотические с растениями, болезнетворные организмы, которые могут заражать людей через клещей, и многие свободноживущие виды, которые используют свет для получения энергии. Несколько линий доказательств подтверждают происхождение митохондрий от этого эндосимбиотического события. Большинство митохондрий имеют форму альфа-протеобактерий и окружены двумя мембранами, в результате чего один мембраносвязанный организм поглощает другой в вакуоль. Внутренняя мембрана митохондрий включает значительные складки, называемые кристами, которые напоминают текстурированную внешнюю поверхность альфа-протеобактерий.Матрикс и внутренняя мембрана богаты ферментами, необходимыми для аэробного дыхания.
Микрофотография митохондрий млекопитающих : На этой просвечивающей электронной микрофотографии митохондрий в клетке легких млекопитающих в поперечном сечении видны кристы, складки внутренней митохондриальной мембраны.
Митохондрии делятся независимо друг от друга в результате процесса, который напоминает деление на бинарное у прокариот. В частности, митохондрии не образуются de novo эукариотической клеткой; они воспроизводятся внутри клетки и распределяются между двумя клетками при делении клеток.Следовательно, хотя эти органеллы сильно интегрированы в эукариотическую клетку, они все равно воспроизводятся, как если бы они были независимыми организмами внутри клетки. Однако их размножение синхронизировано с активностью и делением клетки. Митохондрии имеют собственную кольцевую хромосому ДНК, которая стабилизируется за счет прикрепления к внутренней мембране и несет гены, аналогичные генам, экспрессируемым альфа-протеобактериями. Митохондрии также имеют особые рибосомы и передающие РНК, которые напоминают эти компоненты у прокариот.Все эти особенности подтверждают, что митохондрии когда-то были свободноживущими прокариотами.
Митохондриальные гены
Митохондрии, осуществляющие аэробное дыхание, имеют свои собственные геномы с генами, аналогичными генам альфа-протеобактерий. Однако многие гены респираторных белков расположены в ядре. Когда эти гены сравниваются с генами других организмов, оказывается, что они имеют альфа-протеобактериальное происхождение. Кроме того, в некоторых группах эукариот такие гены находятся в митохондриях, тогда как в других группах они находятся в ядре.Это было интерпретировано как доказательство того, что гены были перенесены из хромосомы эндосимбионта в геном хозяина. Эта потеря генов эндосимбионтом, вероятно, является одним из объяснений того, почему митохондрии не могут жить без хозяина.
Несмотря на перенос генов между митохондриями и ядром, митохондрии сохраняют большую часть своего собственного независимого генетического материала. Одно из возможных объяснений того, что митохондрии сохраняют контроль над некоторыми генами, заключается в том, что может быть трудно транспортировать гидрофобные белки через митохондриальную мембрану, а также обеспечить их доставку в правильное место, что предполагает, что эти белки должны производиться в митохондриях.Другое возможное объяснение состоит в том, что существуют различия в использовании кодонов между ядром и митохондриями, что затрудняет полную передачу генов. Третье возможное объяснение состоит в том, что митохондриям необходимо производить свой собственный генетический материал, чтобы обеспечить метаболический контроль в эукариотических клетках, что указывает на то, что мтДНК напрямую влияет на дыхательную цепь и процессы восстановления / окисления митохондрий.
Эволюция пластидов
Пластиды могут происходить из цианобактерий, поглощенных посредством эндосимбиоза ранними эукариотами, что дает клеткам способность проводить фотосинтез.
Цели обучения
Объясните связь между эндосимбиозом и пластидами и эволюцией эукариот
Ключевые выводы
Ключевые моменты
- Каждый из хлоропластов, хромопластов и лейкопластов представляет собой тип пластид.
- Пластиды у эукариот происходят от первичного эндосимбиоза с участием древних цианобактерий.
- Chlorarachniophytes — это тип водорослей, возникших в результате вторичного эндосимбиоза, когда эукариот поглотил зеленую водоросль (которая сама была продуктом первичного эндосимбиоза с цианобактериями).
- Пластиды имеют несколько общих черт с митохондриями, в том числе наличие собственной ДНК и способность к репликации путем бинарного деления.
Ключевые термины
- хлоропласт : органелла, обнаруженная в клетках зеленых растений и фотосинтезирующих водорослей, где происходит фотосинтез
- тилакоид : складчатая мембрана в хлоропластах растений, из которой образуются граны, используется в фотосинтезе
- пластида : любая из различных органелл, обнаруженных в клетках растений и водорослей, часто связанных с фотосинтезом
Пластиды
Некоторые группы эукариот фотосинтезируют: их клетки содержат, помимо стандартных эукариотических органелл, органеллы другого типа, называемые пластидами.Есть три типа пластид: хлоропласты, хромопласты и лейкопласты. Хлоропласты — это пластиды, которые проводят фотосинтез. Хромопласты — это пластиды, которые синтезируют и хранят пигменты. Лейкопласты представляют собой пластиды, расположенные в несинтетических тканях растения (, например, , корни) и обычно хранящие непигментные молекулы.
Подобно митохондриям, пластиды, по-видимому, имеют первичное эндосимбиотическое происхождение, но отличаются тем, что происходят от цианобактерий, а не от альфа-протеобактерий.Цианобактерии — это группа фотосинтезирующих бактерий со всеми обычными структурами прокариот. Однако, в отличие от большинства прокариот, у них есть обширные внутренние мембранные компартменты, называемые тилакоидами, которые содержат хлорофилл и являются местом светозависимых реакций фотосинтеза. Помимо тилакоидов, хлоропласты, обнаруженные у эукариот, имеют кольцевую хромосому ДНК и рибосомы, аналогичные таковым у цианобактерий. Каждый хлоропласт окружен двумя мембранами, что указывает на первичный эндосимбиоз.Считается, что внешняя мембрана, окружающая пластиду, произошла от вакуоли хозяина, а внутренняя мембрана, как полагают, произошла от плазматической мембраны эндосимбионта.
Хлоропласт : (a) Это поперечное сечение хлоропласта иллюстрирует его сложную внутреннюю мембранную организацию. Стеки тилакоидных мембран разделяют фотосинтетические ферменты и обеспечивают основу для ДНК хлоропластов. (б) Хлоропласты можно увидеть как маленькие зеленые сферы.
Также, как и в случае с митохондриями, есть убедительные доказательства того, что многие гены эндосимбионта передаются в ядро.Пластиды, как и митохондрии, не могут жить независимо вне хозяина. Кроме того, как и митохондрии, пластиды образуются в результате бинарного деления других пластид. Исследователи предположили, что эндосимбиотическое событие, которое привело к появлению Archaeplastida (наземные растения, красные и зеленые водоросли), произошло от 1 до 1,5 миллиарда лет назад, по крайней мере, через 500 миллионов лет после того, как летопись окаменелостей предполагает присутствие эукариот.
Вторичный эндосимбиоз у Chlorarachniophytes
Эндосимбиоз заключается в том, что одна клетка поглощает другую, чтобы со временем произвести коэволюционирующие отношения, в которых ни одна клетка не может выжить в одиночку.Например, хлоропласты красных и зеленых водорослей образуются в результате поглощения фотосинтетической цианобактерии ранним прокариотом. Это приводит к вопросу о возможности клетки, содержащей эндосимбионт, поглотить себя, что приведет к вторичному эндосимбиозу. Не все пластиды эукариот происходят непосредственно от первичного эндосимбиоза. Некоторые из основных групп водорослей стали фотосинтезировать в результате вторичного эндосимбиоза; то есть, поглощая либо зеленые водоросли, либо красные водоросли в качестве эндосимбионтов.Этот вывод подтверждают многочисленные микроскопические и генетические исследования; вторичные пластиды окружены тремя и более мембранами; некоторые вторичные пластиды даже имеют явные остатки ядра эндосимбиотических водорослей.
Первичный и вторичный эндосимбиоз : Показан гипотетический процесс эндосимбиотических событий, ведущих к эволюции хлорахниофитов. При первичном эндосимбиотическом событии гетеротрофный эукариот потреблял цианобактерию. При вторичном эндосимбиозе клетка, возникшая в результате первичного эндосимбиоза, была поглощена второй клеткой.Образовавшаяся органелла стала пластидой у современных хлорахниофитов.
Красные и зеленые водоросли : (а) красные водоросли и (б) зеленые водоросли (визуализированные с помощью световой микроскопии) имеют сходные последовательности ДНК с фотосинтезирующими цианобактериями. Ученые предполагают, что в процессе, называемом эндосимбиозом, предковые прокариоты поглотили фотосинтезирующие цианобактерии, которые превратились в современные хлоропласты.
Молекулярные и морфологические данные свидетельствуют о том, что простейшие хлорарахниофиты являются производными вторичного эндосимбиотического события.Хлорарахниофиты — это редкие водоросли, обитающие в тропических морях и песках. Считается, что эти протисты возникли, когда эукариот поглотил зеленую водоросль, последняя из которых уже установила эндосимбиотические отношения с фотосинтетической цианобактерией. Несколько линий доказательств подтверждают, что хлорарахниофиты произошли от вторичного эндосимбиоза. Хлоропласты, содержащиеся в эндосимбионтах зеленых водорослей, способны к фотосинтезу, заставляя хлорарахниофиты фотосинтезировать.Эндосимбионт зеленых водорослей также имеет низкорослое рудиментарное ядро. Фактически, кажется, что хлорахниофиты являются продуктом недавнего (в масштабе эволюции) вторичного эндосимбиотического события. Пластиды хлорахниофитов окружены четырьмя мембранами: первые две соответствуют внутренней и внешней мембранам фотосинтетической цианобактерии, третья соответствует зеленой водоросли, а четвертая соответствует вакуоли, окружавшей зеленую водоросль, когда она была поглощена предок хлорарахниофита.
Процесс вторичного эндосимбиоза характерен не только для хлорарахниофитов.
Leave A Comment