
Решение заданий части С. Цитологические задачи (С5)
Похожие презентации:
Эндокринная система
Анатомо — физиологические особенности сердечно — сосудистой системы детей
Хронический панкреатит
Топографическая анатомия верхних конечностей
Анатомия и физиология сердца
Мышцы головы и шеи
Эхинококкоз человека
Черепно-мозговые нервы
Анатомия и физиология печени
Топографическая анатомия и оперативная хирургия таза и промежности
1. Решение заданий части С
РЕШЕНИЕ ЗАДАНИЙЧАСТИ С
ЦИТОЛОГИЧЕСКИЕ ЗАДАЧИ (С5)
2. Задача 1
ЗАДАЧА 1В процессе транскрипции участвовало 120
нуклеотидов. Определите число аминокислот,
которые кодируются этими нуклеотидами, а
также число т – РНК, которые будут участвовать
в трансляции, число триплетов в молекуле ДНК,
которые кодируют этот белок.
3. теория
ТЕОРИЯ• 1. транскрипция – это биосинтез молекул и – РНК на
основе молекулы ДНК (происходит в ядре)
• 2.
 трансляция – биосинтез белка на рибосоме
трансляция – биосинтез белка на рибосоме• 3. триплет – последовательность из трех нуклеотидов
• 4. одна молекула т – РНК переносит одну аминокислоту на
рибосому
• 5. один триплет кодирует одну аминокислоту
4. Решение задачи
РЕШЕНИЕ ЗАДАЧИ• 1. одну аминокислоту кодирует три
нуклеотида, следовательно,
число аминокислот = 120 : 3= 40
• 2. число т – РНК = числу аминокислот, т. к.
каждая т – РНК транспортирует одну
аминокислоту
число т – РНК = 40
• 3. три нуклеотида = 1 триплет
число триплетов = 120 : 3 = 40
5. Задача 2
ЗАДАЧА 2В процессе трансляции участвовало 30 молекул т –
РНК. Определите число аминокислот, входящих в
состав синтезируемого белка, а также число
триплетов и нуклеотидов в гене, который кодирует
этот белок.
6. Решение задачи
РЕШЕНИЕ ЗАДАЧИ1. одна молекула т – РНК транспортирует
одну аминокислоту
число аминокислот = число т – РНК = 30
2. одну аминокислоту кодирует один триплет
число триплетов = число аминокислот = 30
3.
 Триплет – это последовательность из трех
Триплет – это последовательность из трехнуклеотидов
Число нуклеотидов = число триплетов × 3
Число нуклеотидов = 30 × 3 = 90
7. Задача 3
ЗАДАЧА 3При исследовании клеток различных органов
млекопитающих было обнаружено, что % — ное
содержание митохондрий в клетках сердечной
мышцы в 2 раза выше, чем в клетках печени, и в
5 раз выше, чем в клетках поджелудочной
железы. Как можно объяснить полученные
результаты?
8. теория
ТЕОРИЯМитохондрии «энергетические
станции клетки»
Основная функция –
синтез АТФ
(Аденозинтрифосфо
рная кислота –
универсальный
источник энергии)
9. Решение задачи
РЕШЕНИЕ ЗАДАЧИ• Митохондрии – органоиды клетки, в которых
происходит энергетический обмен, синтез и
накопление АТФ
• Для работы сердца требуется много
энергии (АТФ), поэтому в клетках сердечной
мышцы наибольшее содержание
митохондрий.
• Обмен веществ в клетках печени выше, чем
в клетках поджелудочной железы, поэтому
клетки этого органа содержат больше
митохондрий
10.
 Задача 4ЗАДАЧА 4
Задача 4ЗАДАЧА 4Фрагмент цепи ДНК имеет последовательность
нуклеотидов: ГТГТАТГГААГТ.
Определите последовательность нуклеотидов на и
– РНК, антикодоны соответствующих т – РНК и
последовательность аминокислот в фрагменте
молекулы белка, пользуясь таблицей
генетического кода.
11. теория
ТЕОРИЯ1.
Принцип комплементарности – избирательное
соединение нуклеотидов. В основе этого принципа лежит
Образование и – РНК на одной из цепочек
ДНК – матрицы.
ДНК
и — РНК
Г (гуанин) – Ц (цитозин)
Ц (цитозин) – Г (гуанин)
А (аденин) – У (урацил)
Т (тимин) – А (аденин)
и – РНК
т – РНК
Г (гуанин) – Ц (цитозин)
Ц (цитозин) – Г (гуанин)
А (аденин) – У (урацил)
У (урацил) – А (аденин)
12. Таблица генетического кода
ТАБЛИЦА ГЕНЕТИЧЕСКОГО КОДА13. Решение задачи
РЕШЕНИЕ ЗАДАЧИ1. Последовательность нуклеотидов на
и – РНК:
ЦАЦАУАЦЦУУЦА
2. антикодоны молекул т – РНК:
ГУГ, УАУ, ГГА, АГУ
3.
 последовательность аминокислот в молекуле
последовательность аминокислот в молекулебелка:
гис-иле-про-сер
14. Задача 5
ЗАДАЧА 5В результате гликолиза образовалось 56 молекул
пировиноградной кислоты (ПВК). Определите,
какое количество молекул глюкозы подверглось
расщеплению и сколько молекул АТФ
образовалось при гидролизе и при полном
окислении. Ответ поясните.
15. теория
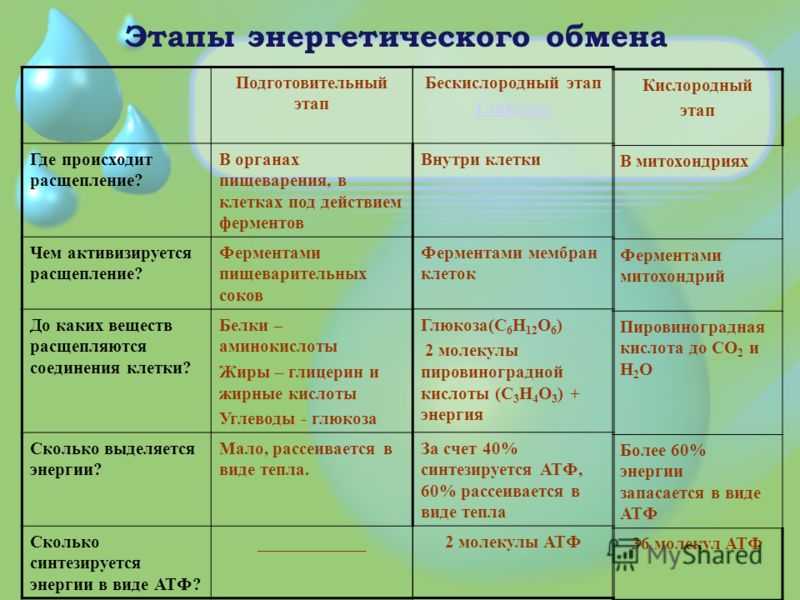
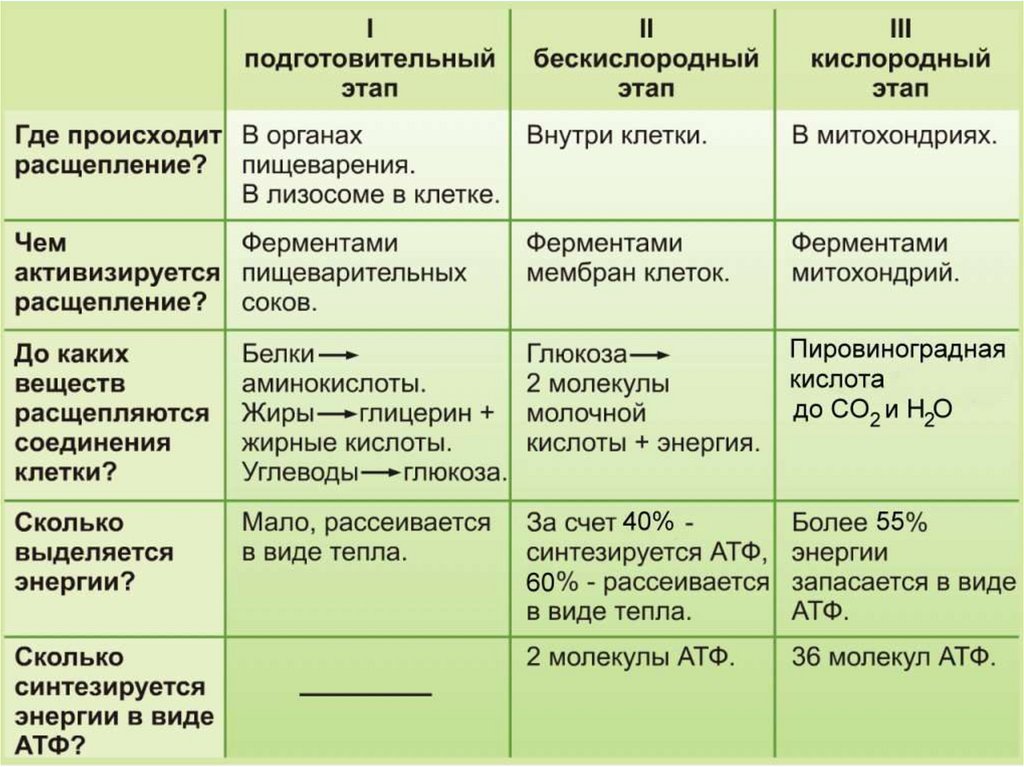
ТЕОРИЯЭнергетический обмен
1 этап: подготовительный
Сложные органические вещества расщепляются на более простые,
энергия рассеивается в виде тепла
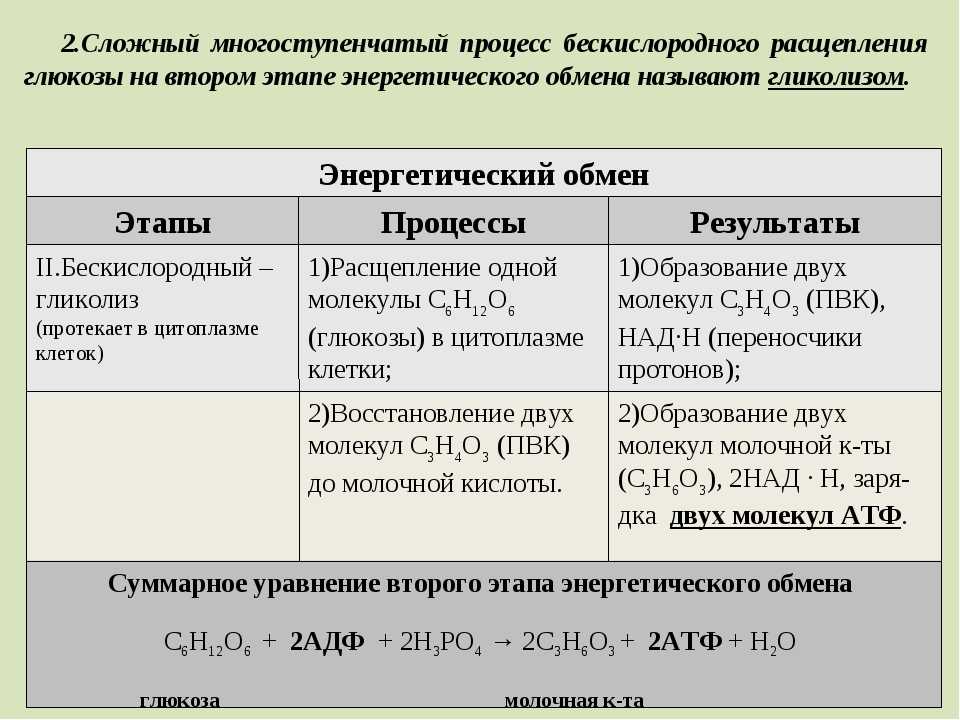
2 этап: гликолиз (бескислородный)
Осуществляется в цитоплазме,
Образуется 2 молекулы ПВК, 2 молекулы АТФ
3 этап – кислородный (гидролиз)
Протекает в митохондриях
Образуется 36 молекул АТФ, углекислый газ, вода
16. Решение задачи
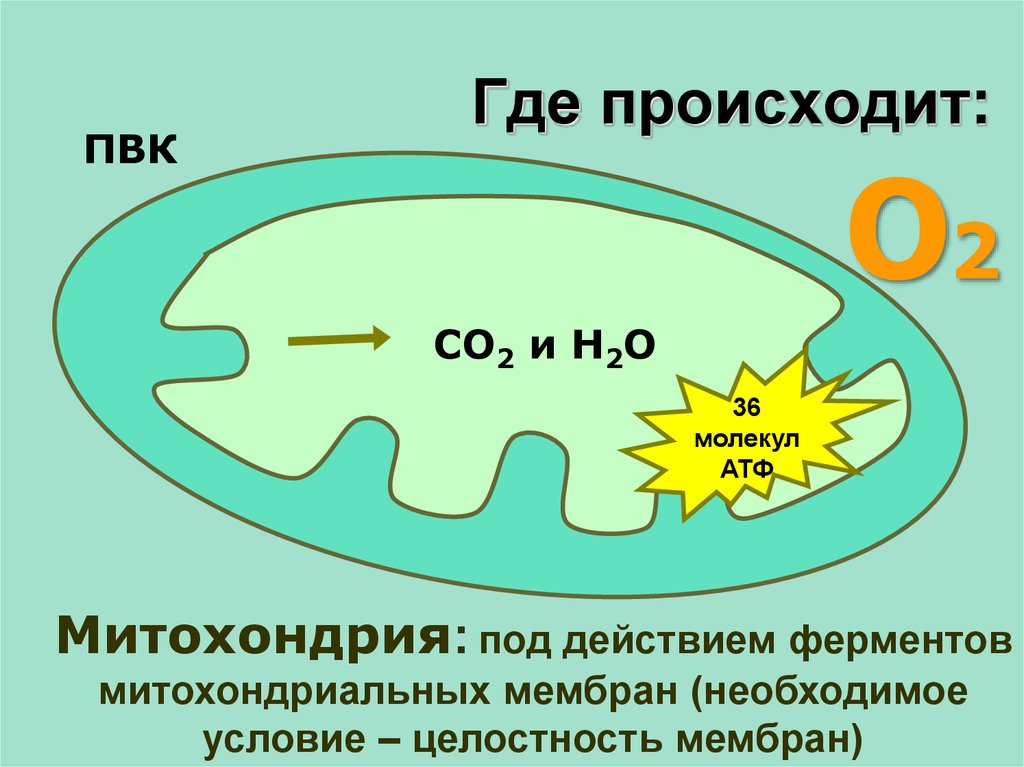
РЕШЕНИЕ ЗАДАЧИ1.
при гликолизе одна молекула глюкозы
расщепляется до 2 молекул ПВК и 2 молекул АТФ
Число молекул глюкозы = 56 : 2 = 28
2. При гидролизе образуется 36 молекул АТФ из одной
молекулы глюкозы
Число АТФ(гидролиз) = 28 * 36 = 1008
3.
 При полном окислении из одной молекулы глюкозы
При полном окислении из одной молекулы глюкозыобразуется 38 молекул АТФ
Число АТФ (полное окисление) = 28 * 38 = 1064
17. Задача 6
ЗАДАЧА 6Сколько молекул АТФ будет синтезировано в
клетках молочнокислых бактерий и клетках
мышечной ткани при окислении 30 молекул
глюкозы?
18. Решение задачи
РЕШЕНИЕ ЗАДАЧИ1.
2.
3.
В клетках молочнокислых бактерий происходит
только гликолиз, а в клетках мышечной ткани и
гликолиз и гидролиз
Из одной молекулы глюкозы при гликолизе
синтезируется 2 молекулы АТФ, значит в клетках
молочнокислых бактерий образуется
30 × 2 = 60 молекул АТФ
При полном окислении одной молекулы глюкозы
синтезируется 38 молекул АТФ, поэтому в клетках
мышечной ткани образуется
30 × 38 = 1140 молекул АТФ
19. Самостоятельно решите
САМОСТОЯТЕЛЬНО РЕШИТЕ• В процессе полного расщепления глюкозы
выход АТФ составил 760 молекул. Сколько
молекул глюкозы подверглись расщеплению, и
каков выход молекул АТФ в процессе гликолиза?
Объясните полученные результаты.

20. Решение задачи
РЕШЕНИЕ ЗАДАЧИ• в результате полного расщепления 1 молекулы глюкозы
выход АТФ составит 38 молекул, из которых 2 молекулы в
процессе гликолиза и 36 молекул в процессе дыхания;
• гликолизу подверглось: 760:38=20 молекул глюкозы;
• при гликолизе 20 молекул глюкозы выход АТФ: 20×2=40
молекул АТФ.
21. Повторение (все варианты С5 заданий)
ПОВТОРЕНИЕ (ВСЕ ВАРИАНТЫ С5ЗАДАНИЙ)
• В молекуле ДНК содержится 19% цитозина.
Определите, сколько (в %) в этой молекуле
содержится других нуклеотидов.
• Г=19%. А=Т=31%.
• В трансляции участвовало 110 молекул т-РНК.
Определите количество аминокислот, входящих
в состав образующегося белка, а также число
триплетов и нуклеотидов в гене, который
кодирует этот белок.
• 110 аминокислот, 110 триплетов, 330 нуклеотидов.
• Фрагмент ДНК состоит из 102 нуклеотидов.
иРНК, а также количество аминокислот,
входящих в состав образующегося белка.

• 34 триплета, 34 аминокислоты, 34 молекулы т-РНК.
• Фрагмент одной из цепей ДНК имеет
следующее строение: ЦТАТЦЦГЦТГТЦ. Постройте
на ней и-РНК и определите последовательность
аминокислот во фрагменте молекулы белка
(для этого используйте таблицу генетического
кода).
• и-РНК: ГАУ-АГГ-ЦГА-ЦАГ. Аминокислотная
последовательность: асп-арг-арг-глн.
• Фрагмент и-РНК имеет следующее строение:
ГАЦАГАЦУЦААГУЦУ. Определите антикодоны тРНК и последовательность аминокислот,
закодированную в этом фрагменте. Также
была синтезирована эта и-РНК (для этого
используйте таблицу генетического кода).
• Фрагмент ДНК: ЦТГТЦТГАГТТЦАГА. Антикодоны тРНК: ЦУГ, УЦУ, ГАГ, УУЦ, АГА. Аминокислотная
последовательность: асп-арг-лей-лиз-сер.
• Фрагмент ДНК имеет следующую
последовательность нуклеотидов ТГТЦЦАТЦАААЦ.
Установите нуклеотидную последовательность тРНК, которая синтезируется на данном фрагменте,
и аминокислоту, которую будет переносить эта тРНК, если третий триплет соответствует
антикодону т-РНК.
используйте таблицу генетического кода.
• т-РНК: АЦА-ГГУ-АГУ-УУГ. Антикодон АГУ, кодон иРНК – УЦА, переносимая аминокислота – сер.
• В клетке животного диплоидный набор
хромосом равен 30. Определите количество
молекул ДНК перед митозом, после митоза,
после первого и второго деления мейоза.
• 2n=30. Генетический набор:
a. перед митозом 60 молекул ДНК;
b. после митоза 30 молекулы ДНК;
c. после первого деления мейоза 30 молекул
ДНК;
d. после второго деления мейоза 15 молекул ДНК.
• В клетке животного диплоидный набор
хромосом равен 56. Определите количество
молекул ДНК и хромосом перед мейозом, после
• В диссимиляцию вступило 23 молекулы глюкозы.
Определите количество АТФ после гликолиза,
после энергетического этапа и суммарный
эффект диссимиляции.
• Поскольку из одной молекулы глюкозы образуется 2
молекулы ПВК и 2АТФ, следовательно, синтезируется
46 АТФ.
• После энергетического этапа диссимиляции
образуется 36 молекул АТФ (при распаде 1 молекулы
глюкозы), следовательно, синтезируется 828 АТФ.
• Суммарный эффект диссимиляции равен
828+46=874 АТФ.
• В цикл Кребса вступило 10 молекул ПВК.
энергетического этапа, суммарный эффект
диссимиляции и количество молекул глюкозы,
вступившей в диссимиляцию.
• В цикл Кребса вступило 10 молекул ПВК,
следовательно, распалось 5 молекул глюкозы.
Количество АТФ после гликолиза – 10 молекул, после
энергетического этапа – 180 молекул, суммарный
эффект диссимиляции 190 молекул АТФ.
• Известно, что длина 1 нуклеотида равна 0,34нм,
его масса 345, а масса аминокислоты 100.
Определить длину гена, кодирующего белок с
массой 82000, его массу и число аминокислот в
белке. Что тяжелее – белок или ген?
English Русский Правила
Подборка заданий по цитологии — материалы для подготовки к ЕГЭ по Биологии
- Примеры задач второго типа
- Примеры задач третьего типа
- Примеры задач четвертого типа
- Примеры задач шестого типа
- Примеры задач седьмого типа
- Ответы
 org/ListItem»>
Примеры задач первого типа
org/ListItem»>
Примеры задач первого типа org/ListItem»>
Приложение I Генетический код (и-РНК)
org/ListItem»>
Приложение I Генетический код (и-РНК)Эта подборка задач содержит все основные типы заданий по цитологии, встречающиеся в ЕГЭ, и предназначена, прежде всего, для самостоятельной подготовки абитуриента к решению задания С5 на экзамене. Для удобства задачи сгруппированы по основным разделам и темам, включенным в программу по биологии (раздел «Цитология»). В конце приведены ответы для самопроверки.
Примеры задач первого типа
- В молекуле ДНК содержится 26% тимина. Определите, сколько (в %) в этой молекуле содержится других нуклеотидов.
- В молекуле ДНК содержится 11% тимина. Определите, сколько (в %) в этой молекуле содержится других нуклеотидов.
- В молекуле ДНК содержится 7% гуанина. Определите, сколько (в %) в этой молекуле содержится других нуклеотидов.
- В молекуле ДНК содержится 23% гуанина.
 Определите, сколько (в %) в этой молекуле содержится других нуклеотидов.
Определите, сколько (в %) в этой молекуле содержится других нуклеотидов. - В молекуле ДНК содержится 19% цитозина. Определите, сколько (в %) в этой молекуле содержится других нуклеотидов.
- В молекуле ДНК содержится 40% цитозина. Определите, сколько (в %) в этой молекуле содержится других нуклеотидов.
 Определите, сколько (в %) в этой молекуле содержится других нуклеотидов.
Определите, сколько (в %) в этой молекуле содержится других нуклеотидов.Примеры задач второго типа
- В трансляции участвовало 80 молекул т-РНК. Определите количество аминокислот, входящих в состав образующегося белка, а также число триплетов и нуклеотидов в гене, который кодирует этот белок.
- В трансляции участвовало 75 молекул т-РНК. Определите количество аминокислот, входящих в состав образующегося белка, а также число триплетов и нуклеотидов в гене, который кодирует этот белок.
- В трансляции участвовало 110 молекул т-РНК. Определите количество аминокислот, входящих в состав образующегося белка, а также число триплетов и нуклеотидов в гене, который кодирует этот белок.
- Фрагмент ДНК состоит из 72 нуклеотидов. Определите число триплетов и нуклеотидов в иРНК, а также количество аминокислот, входящих в состав образующегося белка.
- Фрагмент ДНК состоит из 51 нуклеотида. Определите число триплетов и нуклеотидов в иРНК, а также количество аминокислот, входящих в состав образующегося белка.
- Фрагмент ДНК состоит из 93 нуклеотидов. Определите число триплетов и нуклеотидов в иРНК, а также количество аминокислот, входящих в состав образующегося белка.
- Фрагмент ДНК состоит из 102 нуклеотидов. Определите число триплетов и нуклеотидов в иРНК, а также количество аминокислот, входящих в состав образующегося белка.
- Фрагмент ДНК состоит из 114 нуклеотидов. Определите число триплетов и нуклеотидов в иРНК, а также количество аминокислот, входящих в состав образующегося белка.

к оглавлению ▴
Примеры задач третьего типа
- Фрагмент одной из цепей ДНК имеет следующее строение: ААГЦГТГЦТЦАГ. Постройте на ней и-РНК и определите последовательность аминокислот во фрагменте молекулы белка (для этого используйте таблицу генетического кода).
- Фрагмент одной из цепей ДНК имеет следующее строение: ЦЦАТАТЦЦГГАТ. Постройте на ней и-РНК и определите последовательность аминокислот во фрагменте молекулы белка (для этого используйте таблицу генетического кода).
- Фрагмент одной из цепей ДНК имеет следующее строение: АГТТТЦТГГЦАА. Постройте на ней и-РНК и определите последовательность аминокислот во фрагменте молекулы белка (для этого используйте таблицу генетического кода).
- Фрагмент одной из цепей ДНК имеет следующее строение: ГАТТАЦЦТАГТТ. Постройте на ней и-РНК и определите последовательность аминокислот во фрагменте молекулы белка (для этого используйте таблицу генетического кода).
- Фрагмент одной из цепей ДНК имеет следующее строение: ЦТАТЦЦГЦТГТЦ. Постройте на ней и-РНК и определите последовательность аминокислот во фрагменте молекулы белка (для этого используйте таблицу генетического кода).
- Фрагмент одной из цепей ДНК имеет следующее строение: ААГЦТАЦАГАЦЦ. Постройте на ней и-РНК и определите последовательность аминокислот во фрагменте молекулы белка (для этого используйте таблицу генетического кода).
- Фрагмент одной из цепей ДНК имеет следующее строение: ГГТГЦЦГГАААГ. Постройте на ней и-РНК и определите последовательность аминокислот во фрагменте молекулы белка (для этого используйте таблицу генетического кода).
- Фрагмент одной из цепей ДНК имеет следующее строение: ЦЦЦГТАААТТЦГ. Постройте на ней и-РНК и определите последовательность аминокислот во фрагменте молекулы белка (для этого используйте таблицу генетического кода).
 Постройте на ней и-РНК и определите последовательность аминокислот во фрагменте молекулы белка (для этого используйте таблицу генетического кода).
Постройте на ней и-РНК и определите последовательность аминокислот во фрагменте молекулы белка (для этого используйте таблицу генетического кода).
к оглавлению ▴
Примеры задач четвертого типа
- Фрагмент и-РНК имеет следующее строение: ГАУГАГУАЦУУЦААА. Определите антикодоны т-РНК и последовательность аминокислот, закодированную в этом фрагменте. Также напишите фрагмент молекулы ДНК, на котором была синтезирована эта и-РНК (для этого используйте таблицу генетического кода).
- Фрагмент и-РНК имеет следующее строение: ЦГАГГУАУУЦЦЦУГГ. Определите антикодоны т-РНК и последовательность аминокислот, закодированную в этом фрагменте. Также напишите фрагмент молекулы ДНК, на котором была синтезирована эта и-РНК (для этого используйте таблицу генетического кода).
- Фрагмент и-РНК имеет следующее строение: УГУУЦААУАГГААГГ. Определите антикодоны т-РНК и последовательность аминокислот, закодированную в этом фрагменте. Также напишите фрагмент молекулы ДНК, на котором была синтезирована эта и-РНК (для этого используйте таблицу генетического кода).
- Фрагмент и-РНК имеет следующее строение: ЦЦГЦААЦАЦГЦГАГЦ. Определите антикодоны т-РНК и последовательность аминокислот, закодированную в этом фрагменте. Также напишите фрагмент молекулы ДНК, на котором была синтезирована эта и-РНК (для этого используйте таблицу генетического кода).
- Фрагмент и-РНК имеет следующее строение: АЦАГУГГЦЦААЦЦЦУ. Определите антикодоны т-РНК и последовательность аминокислот, закодированную в этом фрагменте. Также напишите фрагмент молекулы ДНК, на котором была синтезирована эта и-РНК (для этого используйте таблицу генетического кода).
- Фрагмент и-РНК имеет следующее строение: ГАЦАГАЦУЦААГУЦУ. Определите антикодоны т-РНК и последовательность аминокислот, закодированную в этом фрагменте. Также напишите фрагмент молекулы ДНК, на котором была синтезирована эта и-РНК (для этого используйте таблицу генетического кода).
- Фрагмент и-РНК имеет следующее строение: УГЦАЦУГААЦГЦГУА. Определите антикодоны т-РНК и последовательность аминокислот, закодированную в этом фрагменте. Также напишите фрагмент молекулы ДНК, на котором была синтезирована эта и-РНК (для этого используйте таблицу генетического кода).
- Фрагмент и-РНК имеет следующее строение: ГЦАГГЦЦАГУУАУАУ. Определите антикодоны т-РНК и последовательность аминокислот, закодированную в этом фрагменте. Также напишите фрагмент молекулы ДНК, на котором была синтезирована эта и-РНК (для этого используйте таблицу генетического кода).
- Фрагмент и-РНК имеет следующее строение: ГЦУААУГУУЦУУУАЦ. Определите антикодоны т-РНК и последовательность аминокислот, закодированную в этом фрагменте. Также напишите фрагмент молекулы ДНК, на котором была синтезирована эта и-РНК (для этого используйте таблицу генетического кода).

 Также напишите фрагмент молекулы ДНК, на котором была синтезирована эта и-РНК (для этого используйте таблицу генетического кода).
Также напишите фрагмент молекулы ДНК, на котором была синтезирована эта и-РНК (для этого используйте таблицу генетического кода).
к оглавлению ▴
Примеры задач пятого типа
- Фрагмент ДНК имеет следующую последовательность нуклеотидов ТАТГГГЦТАТТГ. Установите нуклеотидную последовательность т-РНК, которая синтезируется на данном фрагменте, и аминокислоту, которую будет переносить эта т-РНК, если третий триплет соответствует антикодону т-РНК. Для решения задания используйте таблицу генетического кода.
- Фрагмент ДНК имеет следующую последовательность нуклеотидов ЦААГАТТТТГТТ. Установите нуклеотидную последовательность т-РНК, которая синтезируется на данном фрагменте, и аминокислоту, которую будет переносить эта т-РНК, если третий триплет соответствует антикодону т-РНК. Для решения задания используйте таблицу генетического кода.
- Фрагмент ДНК имеет следующую последовательность нуклеотидов ГЦЦАААТЦЦТГА. Установите нуклеотидную последовательность т-РНК, которая синтезируется на данном фрагменте, и аминокислоту, которую будет переносить эта т-РНК, если третий триплет соответствует антикодону т-РНК. Для решения задания используйте таблицу генетического кода.
- Фрагмент ДНК имеет следующую последовательность нуклеотидов ТГТЦЦАТЦАААЦ. Установите нуклеотидную последовательность т-РНК, которая синтезируется на данном фрагменте, и аминокислоту, которую будет переносить эта т-РНК, если третий триплет соответствует антикодону т-РНК. Для решения задания используйте таблицу генетического кода.
- Фрагмент ДНК имеет следующую последовательность нуклеотидов ЦАТГААААТГАТ. Установите нуклеотидную последовательность т-РНК, которая синтезируется на данном фрагменте, и аминокислоту, которую будет переносить эта т-РНК, если третий триплет соответствует антикодону т-РНК. Для решения задания используйте таблицу генетического кода.
 Для решения задания используйте таблицу генетического кода.
Для решения задания используйте таблицу генетического кода. Для решения задания используйте таблицу генетического кода.
Для решения задания используйте таблицу генетического кода.к оглавлению ▴
Примеры задач шестого типа
- В клетке животного диплоидный набор хромосом равен 8. Определите количество молекул ДНК перед митозом, после митоза, после первого и второго деления мейоза.
- В клетке животного диплоидный набор хромосом равен 42. Определите количество молекул ДНК перед митозом, после митоза, после первого и второго деления мейоза.
- В клетке животного диплоидный набор хромосом равен 16. Определите количество молекул ДНК перед митозом, после митоза, после первого и второго деления мейоза.
- В клетке животного диплоидный набор хромосом равен 48. Определите количество молекул ДНК перед митозом, после митоза, после первого и второго деления мейоза.
- В клетке животного диплоидный набор хромосом равен 12. Определите количество молекул ДНК перед митозом, после митоза, после первого и второго деления мейоза.
- В клетке животного диплоидный набор хромосом равен 30. Определите количество молекул ДНК перед митозом, после митоза, после первого и второго деления мейоза.
- В клетке животного диплоидный набор хромосом равен 4. Определите количество молекул ДНК перед митозом, после митоза, после первого и второго деления мейоза.
- В клетке животного диплоидный набор хромосом равен 24. Определите количество молекул ДНК перед митозом, после митоза, после первого и второго деления мейоза.
к оглавлению ▴
Примеры задач седьмого типа
- В диссимиляцию вступило 18 молекул глюкозы. Определите количество АТФ после гликолиза, после энергетического этапа и суммарный эффект диссимиляции.
- В диссимиляцию вступило 23 молекулы глюкозы. Определите количество АТФ после гликолиза, после энергетического этапа и суммарный эффект диссимиляции.
- В диссимиляцию вступило 27 молекул глюкозы. Определите количество АТФ после гликолиза, после энергетического этапа и суммарный эффект диссимиляции.
- В диссимиляцию вступило 32 молекулы глюкозы. Определите количество АТФ после гликолиза, после энергетического этапа и суммарный эффект диссимиляции.
- В цикл Кребса вступило 6 молекул ПВК. Определите количество АТФ после энергетического этапа, суммарный эффект диссимиляции и количество молекул глюкозы, вступившей в диссимиляцию.
- В цикл Кребса вступило 8 молекул ПВК. Определите количество АТФ после энергетического этапа, суммарный эффект диссимиляции и количество молекул глюкозы, вступившей в диссимиляцию.
- В цикл Кребса вступило 10 молекул ПВК. Определите количество АТФ после энергетического этапа, суммарный эффект диссимиляции и количество молекул глюкозы, вступившей в диссимиляцию.
- В цикл Кребса вступило 14 молекул ПВК. Определите количество АТФ после энергетического этапа, суммарный эффект диссимиляции и количество молекул глюкозы, вступившей в диссимиляцию.
- В цикл Кребса вступило 28 молекул ПВК. Определите количество АТФ после энергетического этапа, суммарный эффект диссимиляции и количество молекул глюкозы, вступившей в диссимиляцию.
- В цикл Кребса вступило 40 молекул ПВК. Определите количество АТФ после энергетического этапа, суммарный эффект диссимиляции и количество молекул глюкозы, вступившей в диссимиляцию.


к оглавлению ▴
Приложение I Генетический код (и-РНК)
| Первое основание | Второе основание | Третье основание | |||
| У | Ц | А | Г | ||
| У | Фен | Сер | Тир | Цис | У |
| Фен | Сер | Тир | Цис | Ц | |
| Лей | Сер | — | — | А | |
| Лей | Сер | — | Три | Г | |
| Ц | Лей | Про | Гис | Арг | У |
| Лей | Про | Гис | Арг | Ц | |
| Лей | Про | Глн | Арг | А | |
| Лей | Про | Глн | Арг | Г | |
| А | Иле | Тре | Асн | Сер | У |
| Иле | Тре | Асн | Сер | Ц | |
| Иле | Тре | Лиз | Арг | А | |
| Мет | Тре | Лиз | Арг | Г | |
| Г | Вал | Ала | Асп | Гли | У |
| Вал | Ала | Асп | Гли | Ц | |
| Вал | Ала | Глу | Гли | А | |
| Вал | Ала | Глу | Гли | Г |
к оглавлению ▴
Ответы
- А=26%. Г=Ц=24%.
- А=11%. Г=Ц=39%.
- Ц=7%. А=Т=43%.
- Ц=23%. А=Т=27%.
- Г=19%. А=Т=31%.
- Г=40%. А=Т=10%.
- 80 аминокислот, 80 триплетов, 240 нуклеотидов.
- 75 аминокислот, 75 триплетов, 225 нуклеотидов.
- 110 аминокислот, 110 триплетов, 330 нуклеотидов.
- 24 триплета, 24 аминокислоты, 24 молекулы т-РНК.
- 17 триплетов, 17 аминокислот, 17 молекул т-РНК.
- 31 триплет, 31 аминокислота, 31 молекула т-РНК.
- 34 триплета, 34/span> аминокислоты, 34 молекулы т-РНК.
- 38 триплетов, 38 аминокислот, 38 молекул т-РНК.
- и-РНК: УУЦ-ГЦА-ЦГА-ГУЦ. Аминокислотная последовательность: фен-ала-арг-вал.
- и-РНК: ГГУ-АУА-ГГЦ-ЦУА. Аминокислотная последовательность: гли-иле-гли-лей.
- и-РНК: УЦА-ААГ-ЦЦГ-ГУУ. Аминокислотная последовательность: сер-лиз-про-вал.
- и-РНК: ЦУА-АУГ-ГАУ-ЦАА. Аминокислотная последовательность: лей-мет-асп-глн.
- и-РНК: ГАУ-АГГ-ЦГА-ЦАГ. Аминокислотная последовательность: асп-арг-арг-глн.
- и-РНК: УУЦ-ГАУ-ГУЦ-УГГ. Аминокислотная последовательность: фен-асп-вал-три.
- и-РНК: ЦЦА-ЦГГ-ЦЦУ-УУЦ. Аминокислотная последовательность: про-арг-про-фен.
- и-РНК: ГГГ-ЦАУ-УУА-АГЦ. Аминокислотная последовательность: гли-гис-лей-сер.
- Фрагмент ДНК: ЦТАЦТЦАТГААГТТТ. Антикодоны т-РНК: ЦУА, ЦУЦ, АУГ, ААГ, УУУ. Аминокислотная последовательность: асп-глу-тир-фен-лиз.
- Фрагмент ДНК: ГЦТЦЦАТААГГГАЦЦ. Антикодоны т-РНК: ГЦУ, ЦЦА, УАА, ГГГ, АЦЦ. Аминокислотная последовательность: арг-гли-иле-про-три.
- Фрагмент ДНК: АЦААГТТАТЦЦТТЦЦ. Антикодоны т-РНК: АЦА, АГУ, УАУ, ЦЦУ, УЦЦ. Аминокислотная последовательность: цис-сер-иле-гли-арг.
- Фрагмент ДНК: ГГЦГТТГТГЦГЦТЦГ. Антикодоны т-РНК: ГГЦ, ГУУ, ГУГ, ЦГЦ, УЦГ. Аминокислотная последовательность: про-глн-гис-ала-сер.
- Фрагмент ДНК: ТГТЦАЦЦГГТТГГГА. Антикодоны т-РНК: УГУ, ЦАЦ, ЦГГ, УУГ, ГГА. Аминокислотная последовательность: тре-вал-ала-асн-про.
- Фрагмент ДНК: ЦТГТЦТГАГТТЦАГА. Антикодоны т-РНК: ЦУГ, УЦУ, ГАГ, УУЦ, АГА. Аминокислотная последовательность: асп-арг-лей-лиз-сер.
- Фрагмент ДНК: АЦГТГАЦТТГЦГЦАТ. Антикодоны т-РНК: АЦГ, УГА, ЦУУ, ГЦГ, ЦАУ. Аминокислотная последовательность: цис-тре-глу-арг-вал.
- Фрагмент ДНК: ЦГТЦЦГГТЦААТАТА. Антикодоны т-РНК: ЦГУ, ЦЦГ, ГУЦ, ААУ, АУА. Аминокислотная последовательность: ала-гли-глн-лей-тир.
- Фрагмент ДНК: ЦГАТТАЦААГАААТГ. Антикодоны т-РНК: ЦГА, УУА, ЦАА, ГАА, АУГ. Аминокислотная последовательность: ала-асн-вал-лей-тир.
- т-РНК: АУА-ЦЦЦ-ГАУ-ААЦ. Антикодон ГАУ, кодон и-РНК — ЦУА, переносимая аминокислота — лей.
- т-РНК: ГУУ-ЦУА-ААА-ЦАА. Антикодон ААА, кодон и-РНК — УУУ, переносимая аминокислота — фен.
- т-РНК: ЦГГ-УУУ-АГГ-АЦУ. Антикодон АГГ, кодон и-РНК — УЦЦ, переносимая аминокислота — сер.
- т-РНК: АЦА-ГГУ-АГУ-УУГ. Антикодон АГУ, кодон и-РНК — УЦА, переносимая аминокислота — сер.
- т-РНК: ГУА-ЦУУ-УУА-ЦУА. Антикодон УУА, кодон и-РНК — ААУ, переносимая аминокислота — асн.
- 2n=8. Генетический набор:
- перед митозом 16 молекул ДНК;
- после митоза 8 молекулы ДНК;
- после первого деления мейоза 8 молекул ДНК;
- после второго деления мейоза 4 молекул ДНК.
- 2n=42. Генетический набор:
- перед митозом 84 молекул ДНК;
- после митоза 42 молекулы ДНК;
- после первого деления мейоза 42 молекул ДНК;
- после второго деления мейоза 21 молекул ДНК.
- 2n=16. Генетический набор:
- перед митозом 32 молекул ДНК;
- после митоза 16 молекулы ДНК;
- после первого деления мейоза 16 молекул ДНК;
- после второго деления мейоза 8 молекул ДНК.
- 2n=48. Генетический набор:
- перед митозом 96 молекул ДНК;
- после митоза 48 молекулы ДНК;
- после первого деления мейоза 48 молекул ДНК;
- после второго деления мейоза 24 молекул ДНК.
- 2n=12. Генетический набор:
- перед митозом 24 молекул ДНК;
- после митоза 12 молекулы ДНК;
- после первого деления мейоза 12 молекул ДНК;
- после второго деления мейоза 6 молекул ДНК.
- 2n=30. Генетический набор:
- перед митозом 60 молекул ДНК;
- после митоза 30 молекулы ДНК;
- после первого деления мейоза 30 молекул ДНК;
- после второго деления мейоза 15 молекул ДНК.
- 2n=4. Генетический набор:
- перед митозом 8 молекул ДНК;
- после митоза 4 молекулы ДНК;
- после первого деления мейоза 4 молекул ДНК;
- после второго деления мейоза 2 молекул ДНК.
- 2n=24. Генетический набор:
- перед митозом 48 молекул ДНК;
- после митоза 24 молекулы ДНК;
- после первого деления мейоза 24 молекул ДНК;
- после второго деления мейоза 12 молекул ДНК.
- Поскольку из одной молекулы глюкозы образуется 2 молекулы ПВК и 2АТФ, следовательно, синтезируется 36 АТФ. После энергетического этапа диссимиляции образуется 36 молекул АТФ (при распаде 1 молекулы глюкозы), следовательно, синтезируется 648 АТФ. Суммарный эффект диссимиляции равен 648+36=684 АТФ.
- Поскольку из одной молекулы глюкозы образуется 2 молекулы ПВК и 2АТФ, следовательно, синтезируется 46 АТФ. После энергетического этапа диссимиляции образуется 36 молекул АТФ (при распаде 1 молекулы глюкозы), следовательно, синтезируется 828 АТФ. Суммарный эффект диссимиляции равен 828+46=874 АТФ.
- Поскольку из одной молекулы глюкозы образуется 2 молекулы ПВК и 2АТФ, следовательно, синтезируется 54 АТФ. После энергетического этапа диссимиляции образуется 36 молекул АТФ (при распаде 1 молекулы глюкозы), следовательно, синтезируется 972 АТФ. Суммарный эффект диссимиляции равен 972+54=1026 АТФ.
- Поскольку из одной молекулы глюкозы образуется 2 молекулы ПВК и 2АТФ, следовательно, синтезируется 64 АТФ. После энергетического этапа диссимиляции образуется 36 молекул АТФ (при распаде 1 молекулы глюкозы), следовательно, синтезируется 1152 АТФ. Суммарный эффект диссимиляции равен 1152+64=1216 АТФ.
- В цикл Кребса вступило 6 молекул ПВК, следовательно, распалось 3 молекулы глюкозы. Количество АТФ после гликолиза — 6 молекул, после энергетического этапа — 108 молекул, суммарный эффект диссимиляции 114 молекул АТФ.
- В цикл Кребса вступило 8 молекул ПВК, следовательно, распалось 4 молекулы глюкозы. Количество АТФ после гликолиза — 8 молекул, после энергетического этапа — 144 молекул, суммарный эффект диссимиляции 152 молекул АТФ.
- В цикл Кребса вступило 10 молекул ПВК, следовательно, распалось 5 молекул глюкозы. Количество АТФ после гликолиза — 10 молекул, после энергетического этапа — 180 молекул, суммарный эффект диссимиляции 190 молекул АТФ.
- В цикл Кребса вступило 14 молекул ПВК, следовательно, распалось 7 молекул глюкозы. Количество АТФ после гликолиза — 14 молекул, после энергетического этапа — 252 молекул, суммарный эффект диссимиляции 266 молекул АТФ.
- В цикл Кребса вступило 28 молекул ПВК, следовательно, распалось 14 молекул глюкозы. Количество АТФ после гликолиза — 28 молекул, после энергетического этапа — 504 молекул, суммарный эффект диссимиляции 532 молекул АТФ.
- В цикл Кребса вступило 40 молекул ПВК, следовательно, распалось 20 молекул глюкозы. Количество АТФ после гликолиза — 40 молекул, после энергетического этапа — 720 молекул, суммарный эффект диссимиляции 760 молекул АТФ.
 Г=Ц=24%.
Г=Ц=24%.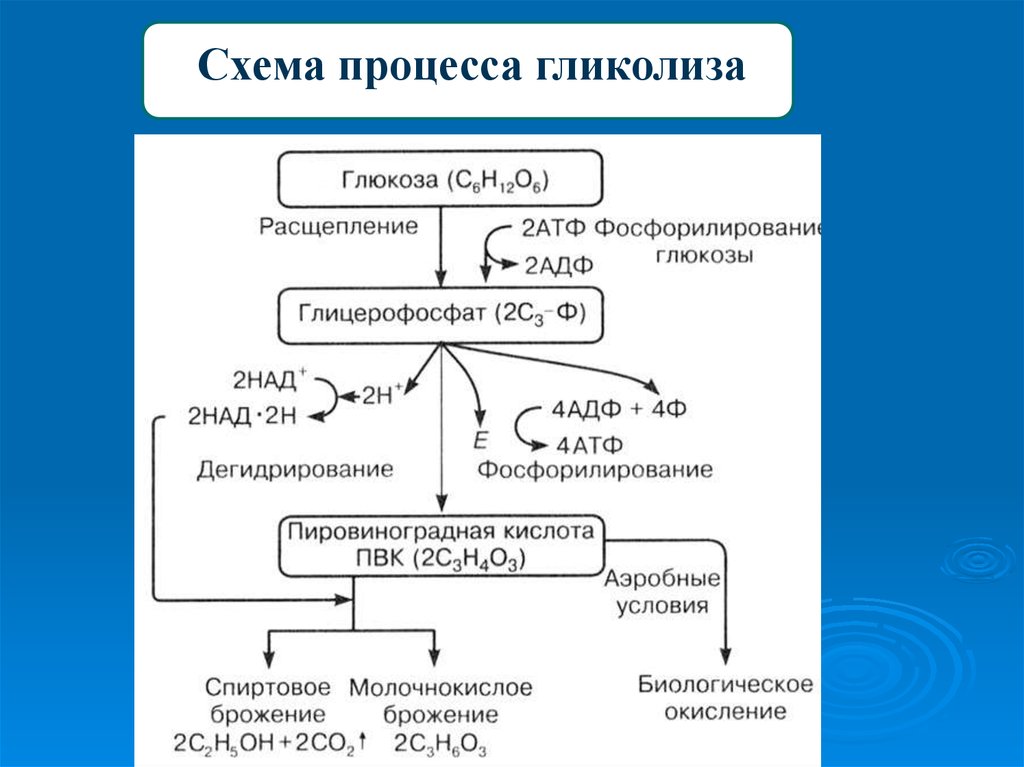
 Аминокислотная последовательность: асп-арг-лей-лиз-сер.
Аминокислотная последовательность: асп-арг-лей-лиз-сер.

 Суммарный эффект диссимиляции равен 828+46=874 АТФ.
Суммарный эффект диссимиляции равен 828+46=874 АТФ.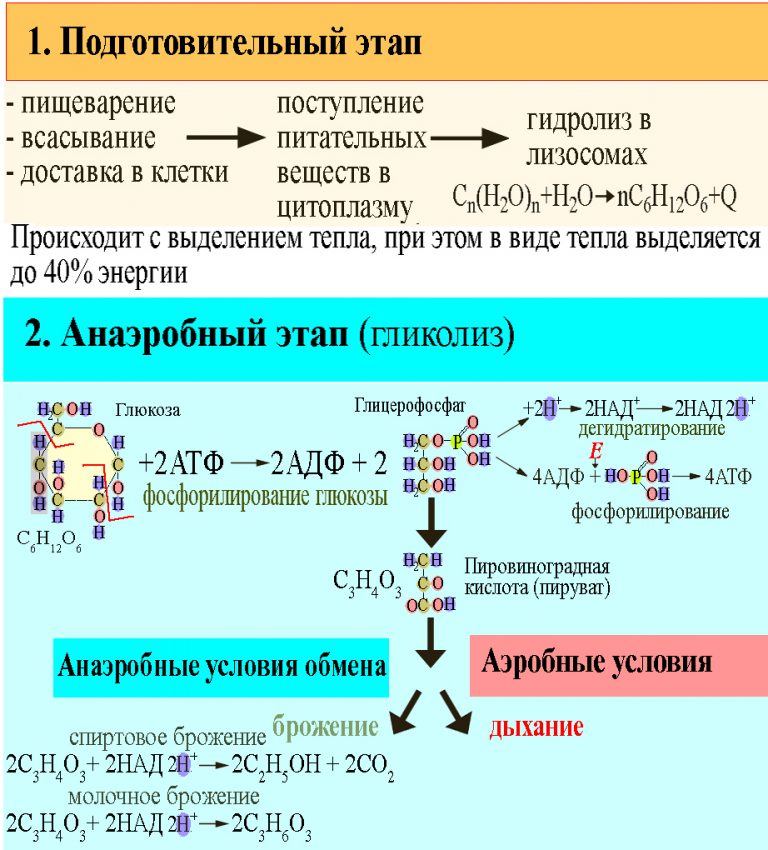
Спасибо за то, что пользуйтесь нашими статьями.
Информация на странице «Подборка заданий по цитологии» подготовлена нашими авторами специально, чтобы помочь вам в освоении предмета и подготовке к экзаменам. Чтобы успешно сдать нужные и поступить в высшее учебное заведение или техникум нужно использовать все инструменты: учеба, контрольные, олимпиады, онлайн-лекции, видеоуроки, сборники заданий.
Также вы можете воспользоваться другими материалами из данного раздела.
Чтобы успешно сдать нужные и поступить в высшее учебное заведение или техникум нужно использовать все инструменты: учеба, контрольные, олимпиады, онлайн-лекции, видеоуроки, сборники заданий.
Также вы можете воспользоваться другими материалами из данного раздела.
Публикация обновлена: 09.04.2023
Документ без названия
Документ без названияГомеостаз глюкозы
Большинство животных обязаны катаболизировать пищу и использовать высвобожденную энергию для запуска анаболического синтеза. Другими словами, мы потребляем сложные вещества, расщепляем их для высвобождения энергии и используем эту энергию для подпитки, создания и восстановления наших собственных клеточных компонентов.
От плесени до млекопитающих глюкоза в количественном отношении является наиболее важным источником топлива для жизни на Земле. Это основное топливо для нашей нервной системы и предпочтительный источник энергии во время начальной физической активности. Глюкоза также является важным строительным материалом для клеточных структур. Когда организму необходимо производить лактозу, гликопротеины и гликолипиды, все они синтезируются с использованием глюкозы.
Глюкоза также является важным строительным материалом для клеточных структур. Когда организму необходимо производить лактозу, гликопротеины и гликолипиды, все они синтезируются с использованием глюкозы.
У нас есть два источника глюкозы: 1) пища, 2) продукты метаболизма. Пища содержит углеводы, липиды, белки и т. д. Пищевые углеводы перевариваются с образованием простых молекул сахара в кишечнике. Простые сахара, такие как глюкоза, галактоза и фруктоза, попадают из просвета кишечника в печень через портальное кровообращение. Глюкоза составляет около 80% абсорбированных диетических сахаров. Галактоза и фруктоза составляют разницу.
Помимо пищевых углеводов мы можем синтезировать глюкозу из неуглеводных продуктов метаболизма (глюконеогенез). Глюконеогенез особенно важен во время поста и голодания, потому что яички, эритроциты, почки, хрусталик и роговица зависят от глюкозы как от единственного источника энергии. Глюкоза также является основным топливом для мозга, но если глюкозы мало, он может использовать кетоновые тела для восполнения около 20% потребности в глюкозе.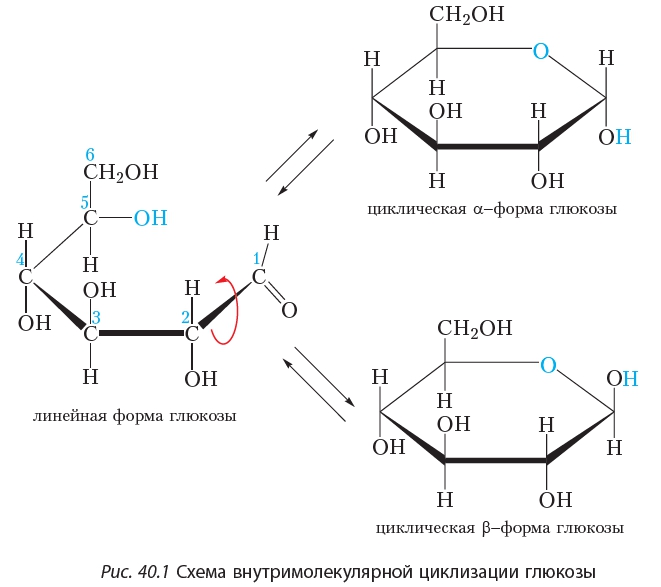 Глюконеогенез может обеспечить нервную систему стабильным поступлением глюкозы даже при длительном голодании.
Глюконеогенез может обеспечить нервную систему стабильным поступлением глюкозы даже при длительном голодании.
Повседневная деятельность требует от нас потребления большего количества питательных веществ при каждом приеме пищи, чем мы можем использовать сразу. Однако организм может хранить лишь небольшое количество глюкозы. Всего во внеклеточной жидкости взрослого человека растворено около 20 г глюкозы. Для удовлетворения потребности в энергии между приемами пищи и во время сна организм запасает глюкозу в виде гликогена или жирных кислот.
Жировая ткань, мышцы и клетки печени являются основными местами накопления энергии. Жировая ткань может использовать глюкозу для синтеза жирных кислот и триглицеридов. Жир служит практически неограниченным долговременным хранилищем липидов с высокой плотностью энергии. Мышцы хранят глюкозу в виде гликогена (≈400 г), который может быть быстро преобразован обратно в глюкозу для мышечной активности. Печень может хранить глюкозу в виде гликогена (≈75 г) и жирных кислот. Печень способна преобразовывать накопленный гликоген в глюкозу и выделять ее в кровь для использования остальными частями тела между приемами пищи, во время голодания и стресса. Печень может поставлять глюкозу примерно на 16 часов, после чего должны быть мобилизованы другие источники энергии.
Печень способна преобразовывать накопленный гликоген в глюкозу и выделять ее в кровь для использования остальными частями тела между приемами пищи, во время голодания и стресса. Печень может поставлять глюкозу примерно на 16 часов, после чего должны быть мобилизованы другие источники энергии.
У здоровых людей чувствительные к инсулину ткани, такие как жировая ткань, мышцы и печень, быстро поглощают глюкозу, чтобы поддерживать постпрандиальный уровень глюкозы в крови в довольно узком диапазоне. У людей с диабетом типа I (СД1) и типа II (СД2) инсулин либо отсутствует, либо недостаточен, либо неэффективен. Поэтому основные действия инсулина, включая всасывание глюкозы и подавление глюкагона, снижаются, что приводит к гипергликемии, характерной для сахарного диабета
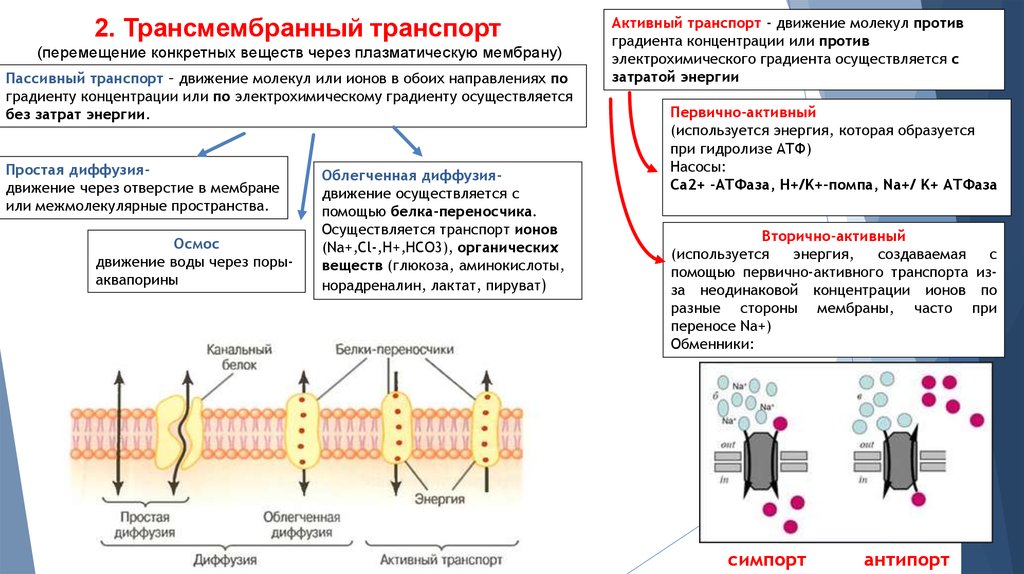
ГЛЮТ | ||
| ГЛЮТ1 | Все ячейки | Низкое сродство/высокая емкость |
| ГЛЮТ2 | Печень, В-клетки, кишечник | Низкое сродство/высокая емкость |
| ГЛЮТ3 | Центральные и периферические нейроны, плацента, яички, тромбоциты | Низкое сродство/высокая емкость |
| ГЛЮТ4 | Поперечно-полосатые и сердечные мышцы, жир | инсулин и активированная физическая нагрузка |
| ГЛЮТ5 | Слизистая оболочка кишечника, сперматозоиды | Носитель фруктозы |
Скорость диффузии глюкозы в клетку зависит от концентрации глюкозы, количества и сродства переносчиков в плазматической мембране. | ||
Гомеостаз глюкозы требует постоянного баланса между транспортом, хранением и метаболизмом глюкозы. Глюкоза не может попасть в клетку путем простой диффузии. Диффузия глюкозы через плазматическую мембрану должна быть облегчена молекулами транспорта глюкозы (GLUT). Чем больше молекул GLUT встроено в плазматическую мембрану, тем больше глюкозы может диффундировать в клетку. Чем больше глюкозы диффундирует в клетки, тем быстрее уровень глюкозы в крови возвращается к норме.
Все клетки экспрессируют GLUT 1 в своей плазматической мембране. Считается, что GLUT 1 отвечает за базальное поглощение глюкозы. Некоторые клетки, такие как мышечные и жировые, могут перемещать дополнительные молекулы GLUT 4 из внутриклеточного хранилища в плазматическую мембрану в ответ на инсулин. Другие клетки, такие как энтероциты в кишечнике, могут уменьшать количество портов GLUT 2 в ответ на инсулин. Это приводит к задержке или уменьшению всасывания глюкозы с пищей. Большинство клеток, особенно печени и поджелудочной железы, могут регулировать синтез генов и экспрессию молекул GLUT в ответ на повышенную концентрацию глюкозы в плазме.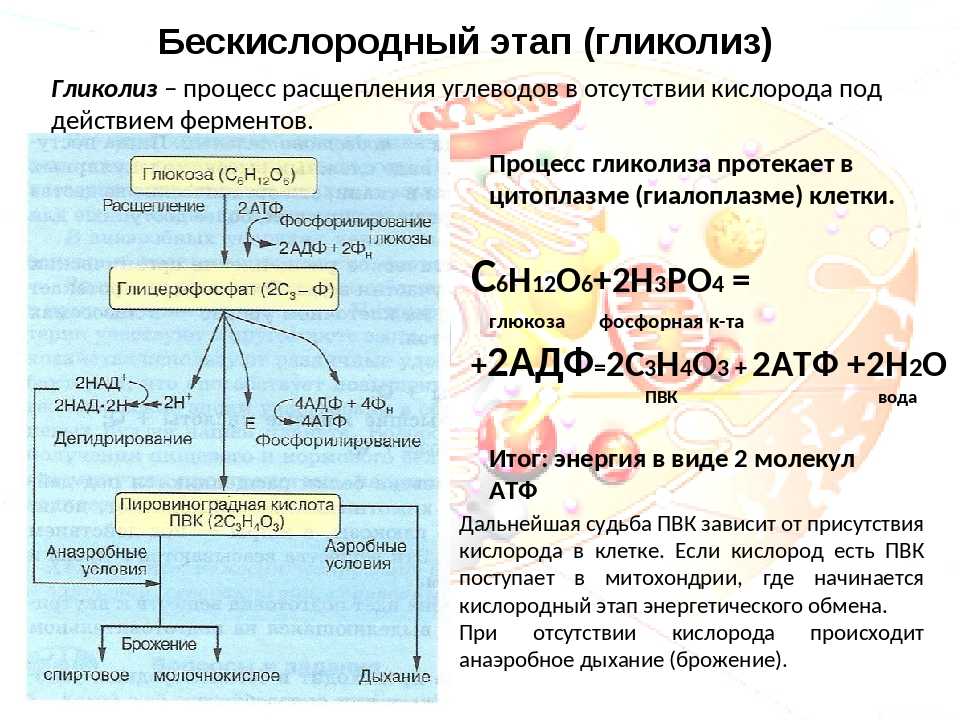
Необходимо контролировать внутриклеточную концентрацию глюкозы. Низкий уровень внутриклеточной глюкозы приводит к стрессу и голоданию. Было показано, что высокие внутриклеточные уровни глюкозы способствуют некротической гибели клеток посредством образования h3O2 (пероксида), который может участвовать в развитии диабетических васкулопатий и связанных с ними заболеваний.
Млекопитающие координируют четыре метаболических процесса для регулирования внутриклеточной глюкозы, независимо от активности или рациона питания. Этими процессами являются: гликолиз, гликогенез, гликогенолиз, гликонеогенез.
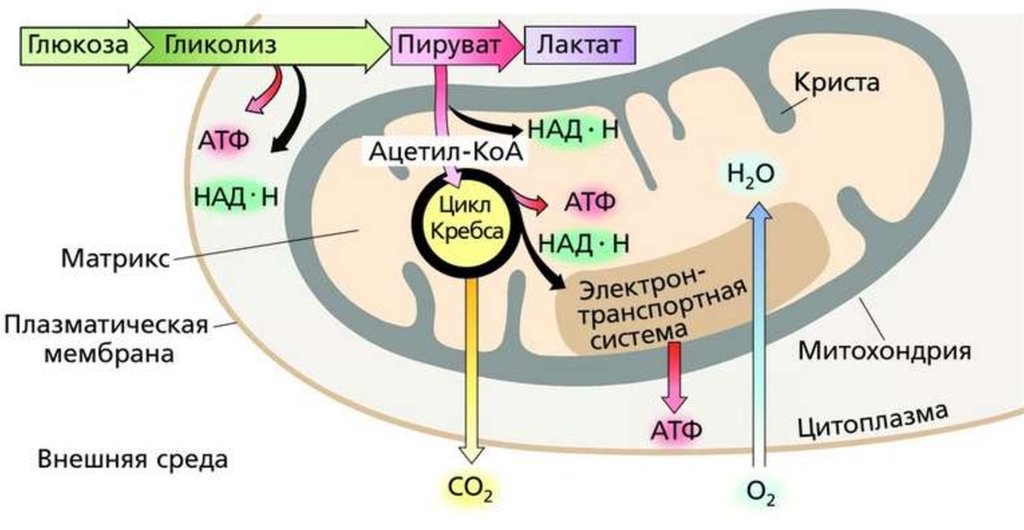
Гликолиз представляет собой ферментативный катаболизм (6 атомов углерода) сахара-глюкозы в две (3 атома углерода) молекулы пирувата + 2 АТФ. Гликолиз снижает внутриклеточную глюкозу, что позволяет безопасно поступать дополнительной глюкозе. Гликолиз представляет собой десятиэтапный процесс. Каждый шаг связан с определенным ферментным катализатором.
Гликогенез — это анаболический процесс, требующий энергии АТФ для сборки избыточных молекул глюкозы в более сложные гранулы гликогена. Одна гранула гликогена может содержать 30 000 единиц глюкозы. Гликоген синтезируется преимущественно гепатоцитами и мышцами. Постпрандиальный запас гликогена в гепатоцитах может составлять до 10% массы печени.
Гликогенез инициируется анаболическим гормоном инсулином. Переваривание пищи дает глюкозу, которая транспортируется из кишечника в циркулирующую кровь. Повышенный уровень глюкозы в плазме заставляет бета-клетки поджелудочной железы высвобождать инсулин. Повышенный уровень инсулина в плазме активирует анаболические пути в печени, мышцах, жире, почках и головном мозге. Эти органы реагируют: 1) облегчая синтез ГЛЮТ, экспрессию и диффузию глюкозы в клетку, 2) усиливая метаболизм глюкозы, 3) увеличивая накопление избыточной глюкозы, 4) ингибируя катаболическое высвобождение глюкозы путем подавления действия глюкагона, 5) усиление продукции липидов, 6) активация генных каскадов, которые регулируют рост и дифференцировку клеток.
Гликогенолиз — это катаболический процесс, при котором накопленный гликоген расщепляется до глюкозы. Гликогенолиз обеспечивает глюкозу во время коротких периодов голодания, между приемами пищи, во время сна и т. д. Глюкагон, продуцируемый альфа-клетками поджелудочной железы, и надпочечниковые катехоламины адреналин и норадреналин являются катаболическими гормонами, регулирующими гликогенолиз. Глюкагон и катехоламины активируют фермент гликогенфосфорилазу . Печень и мышцы являются основными мишенями гликогенолиза, индуцированного глюкагоном. Мышцы дополнительно подвергаются воздействию катехоламинов, которые высвобождаются во время стресса и напряженных упражнений.
Печень и мышцы способны поглощать большое количество глюкозы и сохранять ее в виде гликогена в ответ на инсулин. Между приемами пищи, когда уровень инсулина в плазме низкий, глюкагон увеличивает доступность глюкозы. Глюкагон активирует фермент гликогенфосфорилазу , который разрывает связи, связывающие отдельные молекулы глюкозы с макромолекулами гликогена, хранящимися в гепатоцитах и миоцитах.
Глюкоза, катаболизируемая из гликогена в миоците, используется только в этом миоците. Гликоген фосфорилаза отделяет глюкозу от гликогена в виде глюкозо-1-фосфата. Присутствие фосфатной группы в положении углерода-1 делает глюкозо-1-фосфат слишком большим, чтобы диффундировать из миоцита через портал GLUT 4. Глюкозо-1-фосфат должен быть преобразован в глюкозо-6-фосфат, чтобы подвергнуться гликолизу. Фермент фосфоглюкомутаза удаляет фосфатную группу из положения углерода-1 и добавляет фосфатную группу в положение углерода-6. Затем миоциты могут использовать доступный глюкозо-6-фосфат в гликолизе или возвращать его в хранилище в виде гликогена посредством гликогенеза.
Гепатоциты, в отличие от миоцитов, продуцируют глюкозо-6-фосфатазу . Фермент глюкозо-6-фосфатаза может гидролизовать глюкозо-6-фосфат на фосфатную группу и свободную глюкозу.
Катехоламины адреналин и норэпинефрин являются гормонами, секретируемыми мозговым веществом надпочечников. Эти гормоны подготавливают тело к стрессу, например, к борьбе или бегству. Эти гормоны также активируют фермент гликогенфосфорилазу . Гликогенфосфорилаза разрывает связи, удерживающие отдельные молекулы глюкозы в макромолекуле гликогена. Это обеспечивает мышцы топливом, необходимым для быстрых спринтов и подъемов тяжестей.
Недавнее исследование Dufour S. и Lebon V. et al. (2009 г.) продемонстрировали 2,5-кратное увеличение уровня глюкозы в плазме после инфузии эпинефрина.
Печень и почки экспрессируют фермент глюкозо-6-фосфатазу. Эти органы используют фермент глюкозо-6-фосфатазу для удаления фосфатной группы глюкозо-6-фосфата. В результате этой реакции образуется свободная молекула глюкозы и фосфатная группа. Затем свободная глюкоза может диффундировать из клетки в кровоток.
Глюконеогенез представляет собой синтез новой глюкозы из таких метаболитов, как лактат, пируват, глицерин и аланин. Например, лактат, образующийся в мышцах и эритроцитах как метаболический продукт гликолиза, переносится кровью в печень, где он превращается в глюкозу и высвобождается обратно в кровь.
 (Нажмите здесь, чтобы просмотреть инструкции)
(Нажмите здесь, чтобы просмотреть инструкции) 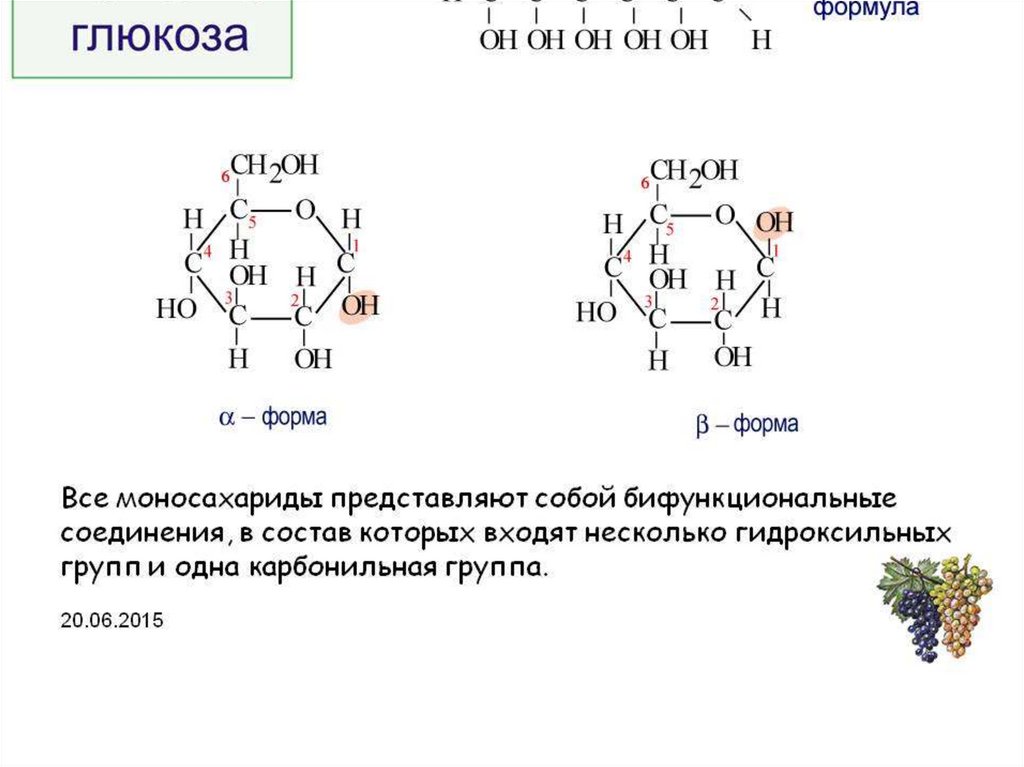 Все эти действия снижают избыток глюкозы в плазме.
Все эти действия снижают избыток глюкозы в плазме.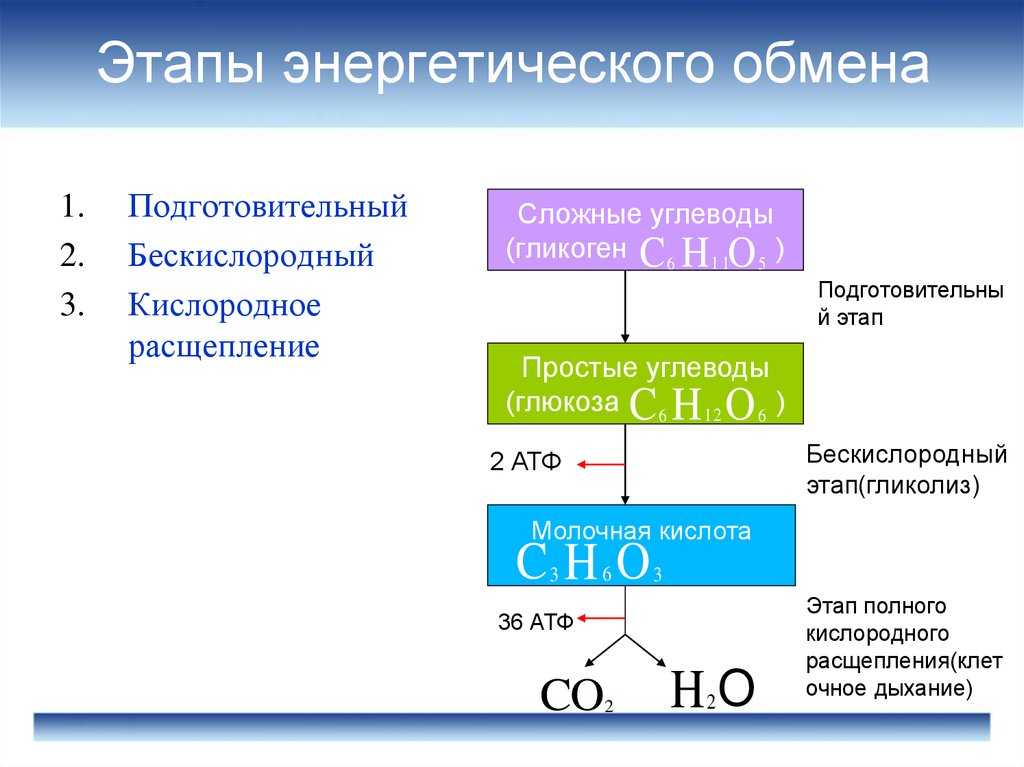 При СД1 и СД2 продукция глюкагона не подавляется инсулином. Беспрепятственное действие глюкагона у диабетиков, по крайней мере частично, ответственно за характерную гипергликемию.
При СД1 и СД2 продукция глюкагона не подавляется инсулином. Беспрепятственное действие глюкагона у диабетиков, по крайней мере частично, ответственно за характерную гипергликемию.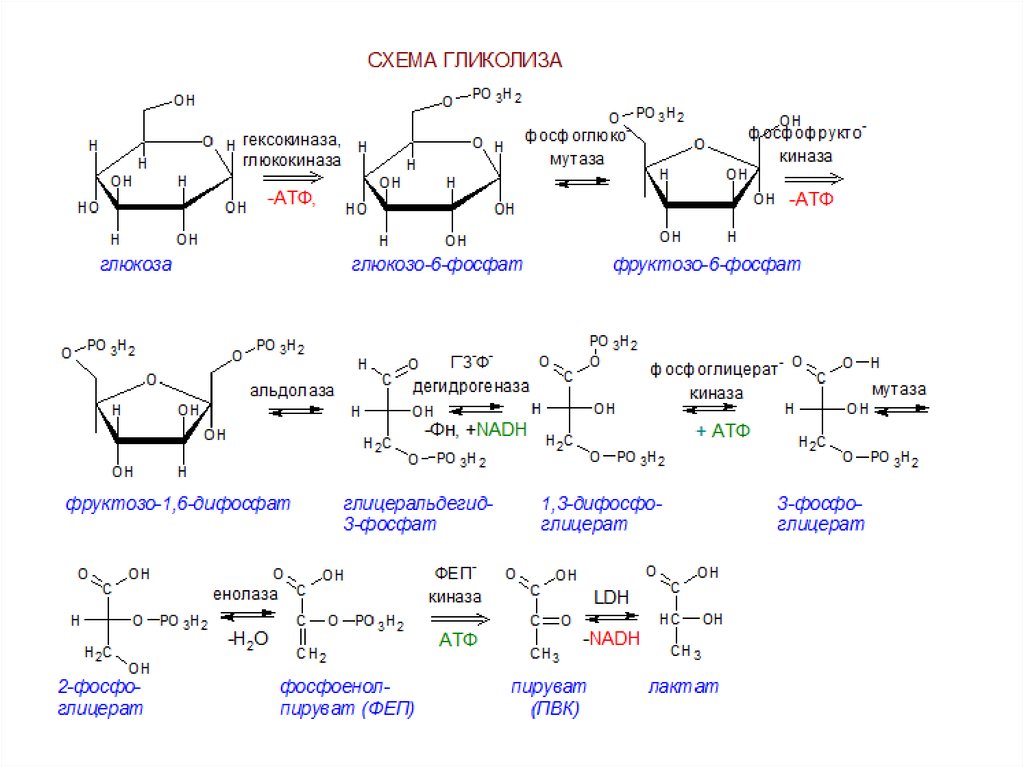 Свободная молекула глюкозы достаточно мала, чтобы диффундировать обратно в плазму через портал GLUT4. Наличие глюкозо-6-фосфатазы позволяет печени накапливать и высвобождать глюкозу для использования всеми клетками организма. Глюкоза, выделяемая печенью, является основным источником энергии между приемами пищи. Он также является источником гипергликемии у инсулинорезистентных диабетиков.
Свободная молекула глюкозы достаточно мала, чтобы диффундировать обратно в плазму через портал GLUT4. Наличие глюкозо-6-фосфатазы позволяет печени накапливать и высвобождать глюкозу для использования всеми клетками организма. Глюкоза, выделяемая печенью, является основным источником энергии между приемами пищи. Он также является источником гипергликемии у инсулинорезистентных диабетиков. Была продемонстрирована обратная связь между запасами гликогена в печени и уровнем глюкозы в плазме после инфузии адреналина. Был сделан вывод, что большая часть печеночного гликогена превращается в глюкозу в первые 60 минут после инфузии адреналина, а уровень глюкозы в плазме нормализуется в течение 90 минут из-за двукратного увеличения печеночного гликонеогенеза.
Была продемонстрирована обратная связь между запасами гликогена в печени и уровнем глюкозы в плазме после инфузии адреналина. Был сделан вывод, что большая часть печеночного гликогена превращается в глюкозу в первые 60 минут после инфузии адреналина, а уровень глюкозы в плазме нормализуется в течение 90 минут из-за двукратного увеличения печеночного гликонеогенеза.
- Сахарный диабет 2 типа увеличивает скорость глюконеогенеза. Магнуссон и его коллеги связывают увеличение выработки глюкозы натощак на ≈18% у диабетиков с СД2 глюконеогенезом.
- Сахарный диабет СД1 увеличивает выработку глюкозы печенью во время физической нагрузки и в состоянии покоя. Петерсон и его коллеги продемонстрировали, что гипергликемия при СД1 в покое и во время физической нагрузки может быть полностью объяснена повышенным глюконеогенезом. В то время как повышенная выработка глюкозы печенью во время физических упражнений у здоровых людей является результатом гликогенолиза.
- «Введение лактата Рингера пациенту с диабетом, перенесшему операцию, может заметно повысить уровень глюкозы. Лактат является предшественником глюконеогенеза, и в стрессовых ситуациях, таких как операция, скорость глюконеогенеза может повышаться». — Сахарный диабет Джослина
- Некоторые виды рака, особенно с быстрым ростом клеток, требуют больших затрат энергии. Производство и метаболизм глюкозы часто намного выше у этих больных раком, чем у здоровых людей. Дж. А. Тайек и Дж. Кац определили, что многие больные раком полагаются на глюконеогенез для большей части производства глюкозы, в то время как здоровые люди получают таким образом менее половины своей глюкозы.
- G. Bongaerts and H. van Halteren et al. выдвигают гипотезу: «В растущих опухолях концентрация O2 критически низка». «При отсутствии достаточного количества О2 им приходится переключаться на анаэробную диссимиляцию, при этом на 1 моль глюкозы приходится всего 2 моля АТФ и 2 моля молочной кислоты». «Поэтому для роста этих опухолевых клеток потребуется примерно в 40 раз больше глюкозы, чем необходимо при наличии достаточного количества O2». «Компенсаторная глюкоза обеспечивается печеночным глюконеогенезом из молочной кислоты. Однако для синтеза глюкозы печень должна затрачивать в 3 раза больше энергии, чем может быть извлечена опухолевыми клетками анаэробным путем. Печень извлекает необходимую энергию из аминокислот и особенно из жирных кислоты окислительным путем. Это может быть причиной потери веса, даже когда прием пищи кажется адекватным».
 Производство и метаболизм глюкозы часто намного выше у этих больных раком, чем у здоровых людей. Дж. А. Тайек и Дж. Кац определили, что многие больные раком полагаются на глюконеогенез для большей части производства глюкозы, в то время как здоровые люди получают таким образом менее половины своей глюкозы.
Производство и метаболизм глюкозы часто намного выше у этих больных раком, чем у здоровых людей. Дж. А. Тайек и Дж. Кац определили, что многие больные раком полагаются на глюконеогенез для большей части производства глюкозы, в то время как здоровые люди получают таким образом менее половины своей глюкозы.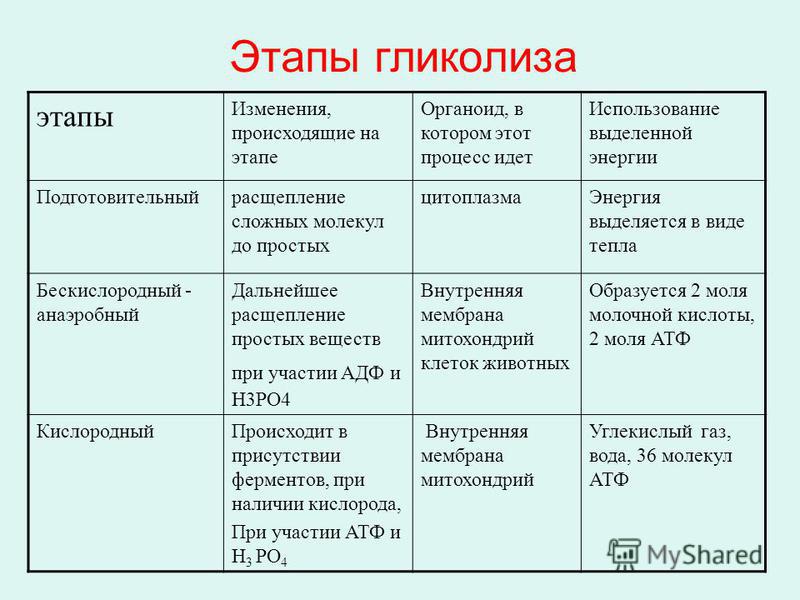 Это может быть причиной потери веса, даже когда прием пищи кажется адекватным».
Это может быть причиной потери веса, даже когда прием пищи кажется адекватным».Каталожные номера
Bongaerts GP, van Halteren HK, Verhagen CA, Wagener DJ. (2006). Раковая кахексия демонстрирует энергетическое влияние глюконеогенеза на метаболизм человека. Медицинские гипотезы . 67(5):1213-22. doi: 10.1016/j.mehy.2006.04.048. Epub 2006 23 июня.
Тайек Дж. А., Кац Дж. (1997) Производство, переработка глюкозы, цикл Кори и глюконеогенез у людей: связь с кортизолом в сыворотке. Am J Physiol . 272 (3 часть 1): E476-84. дои: 10.1152/ajpendo.1997.272.3.Е476. PMID: 9124555.
Пути диссимиляции глюкозы в организме
РЕКЛАМА:
Следующие пункты выделяют три основных пути диссимиляции глюкозы в организмах. Пути: 1. Путь Эмбдена-Мейергофа или гликолиз 2. Пентозофосфатный цикл (PPC) 3. Путь Энтнера-Дудорова (EDP).
Диссимиляция глюкозы: путь №
1.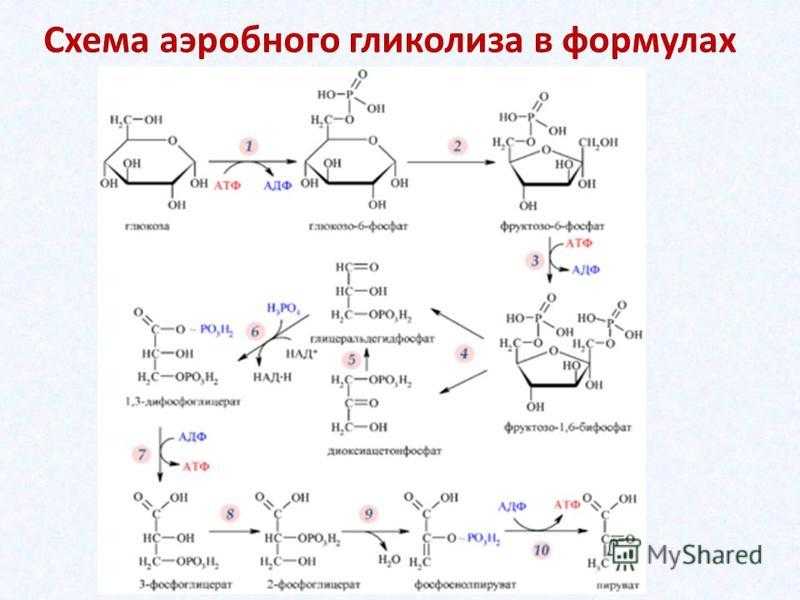 Путь Эмбдена-Мейергофа или гликолиз :
Путь Эмбдена-Мейергофа или гликолиз : Этот катаболический путь приводит к расщеплению глюкозы до двух молекул пировиноградной кислоты, т.е. шестиуглеродное соединение расщепляется до двух молекул трехуглеродных соединений. Это преобразование происходит в несколько этапов, и в процессе фосфорилирования на уровне субстрата образуются две молекулы АТФ. Всего задействовано 10 различных ферментов, которые присутствуют в растворимом состоянии в цитозоле в виде свободных единиц.
Стадии гликолиза и названия ферментов приведены ниже на рис. 8.43:
Обсуждаются детали отдельных стадий гликолитического пути:
На первом этапе нейтральная молекула глюкозы активируется в отрицательно заряженный глюкозо-6-фосфат ферментом гексокиназой с использованием АТФ в качестве донора фосфата и Mg ++ в качестве кофактора. Гексокиназная реакция сильно экзергонична и не является свободно обратимой. Обратная реакция регенерации глюкозы из глюкозо-6-фосфата катализируется другим ферментом глюкозо-6-фосфатазой, который высвобождает неорганический фосфат.
На третьем этапе происходит дальнейшее фосфорилирование фруктозо-6-фосфата до фруктозодифосфата (правильнее его следует называть фруктозодифосфатом). Реакция катализируется фруктозо-6-фосфаткиназой, также известной как фосфофруктокиназа. Он требует АТФ и Mg ++ и управляет реакцией в одном направлении. Обратная реакция (ФБП —> F 6-P) катализируется дифосфофруктозофосфатазой.
Четвертая стадия состоит из расщепления FBP на два триозофосфата, GAP и DHAP, катализируемого лиазой, называемой альдолазой FBP. Альдолазная реакция обратима, а обратная реакция представляет собой альдольную конденсацию. Следовательно, фермент известен как альдолаза. Реакция расщепления распределяет атомы углерода 1, 2 и 3 FBP на GAP и атомы углерода 4, 5 и 6 на DHAP.
Два триозофосфата свободно превращаются друг в друга под действием изомеразы, триозофосфатизомеразы, но равновесие явно в пользу DHAP (указано жирной стрелкой), так что в любой момент времени около 90% триозофосфаты находятся в форме DHAP. Дальнейшая разбивка происходит только от GAP. По мере того, как все больше и больше GAP потребляется на последующих этапах гликолиза, DHAP превращается в GAP для поддержания запаса GAP.
Дальнейшая разбивка происходит только от GAP. По мере того, как все больше и больше GAP потребляется на последующих этапах гликолиза, DHAP превращается в GAP для поддержания запаса GAP.
Пятая стадия включает сложную реакцию, в которой GAP окисляется до дифосфоглицериновой кислоты, а NAD действует как H-акцептор с образованием NADH 2 . Реакция катализируется ферментом глицеральдегид-3-фосфатдегидрогеназой, который имеет активную группу -SH. Группа фермента -SH связывается с альдегидной группой GAP, образуя тиоэфирную связь с последующим окислением альдегидной группы до кислотной группы, NAD действует как кофермент.
Богатая энергией тиоэфирная связь используется для добавления другой фосфатной группы, и фермент высвобождается в своей исходной форме SH-фермента, чтобы инициировать новый цикл катализа. Продукт представляет собой 1,3-дифосфоглицериновую кислоту (DPGA).
Шестой этап состоит из переноса богатой энергией фосфатной группы DPGA в DPGA в АДФ с образованием АТФ и 3-фосфоглицериновой кислоты (3-PGA).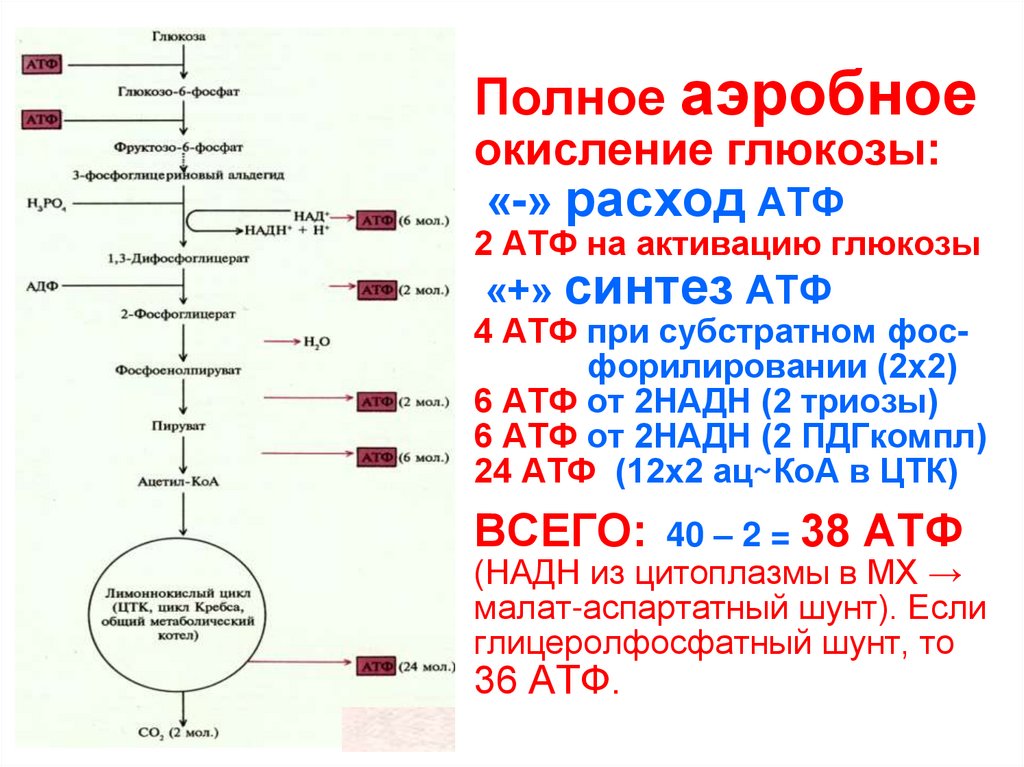 Реакция катализируется ферментом киназой фосфоглицериновой кислоты. Способ образования АТФ, включающий перенос богатой энергией фосфатной группы на АДФ, известен как фосфорилирование на уровне субстрата.
Реакция катализируется ферментом киназой фосфоглицериновой кислоты. Способ образования АТФ, включающий перенос богатой энергией фосфатной группы на АДФ, известен как фосфорилирование на уровне субстрата.
На седьмом этапе 3-ФГК превращают в 2-фосфоглицериновую кислоту (2-ФГК) путем переноса фосфатной группы на средний атом углерода. Реакцию катализирует фермент фосфоглицеромутаза. Реакция свободно обратима и требует Mg ++.
Восьмая стадия, катализируемая ферментом енолоазой, состоит из дегидратации 2-PGA с образованием фосфоенолпировиноградной кислоты (PEP). В результате удаления H 2 O из 2-PGA фосфатная связь становится более энергоемкой. Фермент требует – MG ++ или Mn ++ в качестве кофактора и сильно ингибируется фторидом, который связывает кофакторы и предотвращает функцию енолазы.
Девятый и последний этап гликолитического пути катализируется ферментом киназой пировиноградной кислоты, которая переносит богатую энергией фосфатную группу ФЕП на АДФ с образованием АТФ, тем самым вызывая фосфорилирование на втором уровне субстрата. Реакция сильно экзергонична и необратима.
Реакция сильно экзергонична и необратима.
ОБЪЯВЛЕНИЙ:
Можно отметить, что кроме трех реакций гликолитического пути, а именно. первая стадия катализируется гексокиназой, третья стадия катализируется фосфофруктокиназой и последняя стадия катализируется киназой пировиноградной кислоты, остальные реакции обратимы. Во всех этих трех необратимых стадиях АТФ является одним из реагентов.
Подводя итог, гликолитический путь приводит к расщеплению молекулы глюкозы сначала на две триозы, которые взаимопревращаются. Триоза окисляется до глицериновой кислоты, а затем превращается в пировиноградную кислоту. Таким образом, из каждой половины глюкозы образуется одна пировиноградная кислота, т.е. из одной молекулы глюкозы образуется две молекулы пировиноградной кислоты.
Существует только одна окислительно-восстановительная стадия, катализируемая глицеральдегидфосфатдегидрогеназой, на которой образуется NADH 2 , т. е. две NADH 2 образуются на молекулу глюкозы, по одной из каждой половины. Кроме того, два этапа потребляют АТФ, а именно. превращение глюкозы в глюкозо-6-фосфат и фруктозо-6-фосфат в фруктозобисфосфат, катализируемое соответствующими киназами; и есть две стадии производства АТФ для каждой половины глюкозы, а именно. превращение дифосфоглицериновой кислоты в фосфоглицериновую кислоту (этап 6) и превращение фосфоенолпировиноградной кислоты в пировиноградную кислоту (этап 9).).
Кроме того, два этапа потребляют АТФ, а именно. превращение глюкозы в глюкозо-6-фосфат и фруктозо-6-фосфат в фруктозобисфосфат, катализируемое соответствующими киназами; и есть две стадии производства АТФ для каждой половины глюкозы, а именно. превращение дифосфоглицериновой кислоты в фосфоглицериновую кислоту (этап 6) и превращение фосфоенолпировиноградной кислоты в пировиноградную кислоту (этап 9).).
При составлении баланса входов и выходов гликолитического пути наблюдается, что на каждую молекулу глюкозы, расщепленную до двух молекул пировиноградной кислоты, расходуется две молекулы АТФ и образуется четыре молекулы АТФ, а также две молекулы АТФ. молекулы НАД восстанавливаются до НАДН 2 . Таким образом, чистый выпуск равен
.ОБЪЯВЛЕНИЙ:
Глюкоза -> 2 Пировиноградная кислота + 2 АТФ + 2 НАДН 2
В аэробных организмах, осуществляющих кислородное дыхание, пировиноградная кислота далее разлагается в цикле трикарбоновых кислот (ЦТК или цикл Кребса), где атомы углерода пировиноградной кислоты окисляются до CO 2 и атом Н переносятся на определенные коферменты, такие как НАД и ФАД. Водород от этих коферментов подается в цепь переноса электронов для образования АТФ и, наконец, переносится в воду, производящую кислород. NADH 2 в процессе окисляется обратно до NAD и может использоваться на окислительно-восстановительной стадии гликолиза. Таким образом, обеспечивается непрерывное поступление НАД для запуска гликолиза.
Водород от этих коферментов подается в цепь переноса электронов для образования АТФ и, наконец, переносится в воду, производящую кислород. NADH 2 в процессе окисляется обратно до NAD и может использоваться на окислительно-восстановительной стадии гликолиза. Таким образом, обеспечивается непрерывное поступление НАД для запуска гликолиза.
Анаэробные организмы, у которых отсутствует цикл ТСА и дыхательная цепь, неспособны регенерировать НАД из НАДН 2 так, как это делают аэробные организмы. Они должны восстанавливать пировиноградную кислоту либо до молочной кислоты, либо до этанола с помощью NADH 2 , который при этом окисляется до NAD. Полученный таким образом НАД затем используется для запуска гликолиза. Более того, анаэробные организмы должны зависеть от фосфорилирования на уровне субстрата для образования АТФ.
Диссимиляция глюкозы: путь #
2. Пентозофосфатный цикл (PPC) : Этот путь также известен как гексозомонофосфатный шунт или фосфоглюконатный путь. PPC не является основным путем катаболизма глюкозы, но он действует во многих организмах бок о бок с EMP. Его основное значение заключается в производстве NADPH 2 , который требуется в качестве кофермента для многих ферментов биосинтеза.
PPC не является основным путем катаболизма глюкозы, но он действует во многих организмах бок о бок с EMP. Его основное значение заключается в производстве NADPH 2 , который требуется в качестве кофермента для многих ферментов биосинтеза.
ОБЪЯВЛЕНИЙ:
Еще одной важной функцией является производство пентозы, особенно рибозы, которая является неотъемлемой частью всех нуклеиновых кислот. PPC считается циклическим путем, в отличие от EMP, потому что два продукта, а именно. фруктозо-6-фосфат и глицеральдегидфосфат могут быть превращены обратно в исходное соединение, глюкозо-6-фосфат.
Схема КПП приведена на рис. 8.44:
PPC начинается с фосфорилирования глюкозы до глюкозо-6-фосфата, катализируемого гексокиназой и АТФ, как и в EMP.
На следующем этапе глюкозо-6-фосфат окисляется НАДФ-связанным ферментом, глюкозо-6-фосфатдегидрогеназой, с образованием 6-фосфоглюконовой кислоты и НАДФН 2 через нестабильное промежуточное соединение 6-фосфоглюконолактон.
6-фосфоглюконовая кислота затем превращается в процессе окисления и декарбоксилирования в рибулозо-5-фосфат с помощью фермента дегидрогеназы 6-фосфоглюконовой кислоты, в которой НАДФ снова действует как акцептор водорода. На этом этапе завершаются реакции окисления ППК.
Реакция остальной части цикла включает перенос двухуглеродных и трехуглеродных звеньев различных сахаров двумя классами ферментов – транскетолазой, переносящей гликоальдегидную группу (CH 2 OH-CHO), и трансальдолазой, переносящей дигидроксиацетон (CH 2 OH.CO.CH 2 OH) группа.
Показаны две реакции, катализируемые этими ферментами:
Баланс PPC показывает, что начиная с 6-молекул глюкозо-6-фосфата (без учета общего первого шага между ЭМИ и PPC) получаются следующие входы и выходы:
ОБЪЯВЛЕНИЙ:
6 Глюкоза-6-фосфат + 12 -> НАДФ 5 Глюкоза-6-фосфат + 6CO 2 + 12 НАДФН 2 + Pi.
Удалив 5 глюкозо-6-фосфатов с обеих сторон, получим
Глюкоза-6-фосфат + 12 НАДФ -> 12 НАДФН 2 + 6 СО 2 + Pi
Таким образом, видно, что при окислении каждой молекулы глюкозо-6-фосфата образуется 12 молекул НАДФ3. Основное значение PPC заключается в продукции восстановителя NADPh3 во внемитохондриальной цитоплазме клетки. Основное применение в многочисленных биосинтетических реакциях окисления — восстановления.
Следует отметить, что НАДФ3 не может окисляться напрямую через электрон-транспортную систему, как НАДФ3. Трансгидрирование НАД под действием НАДФ3 возможно, но происходит редко (НАДФН 2 —> НАДФ + НАДН 2 ).
Другим значением PPC является производство ряда полезных промежуточных продуктов, таких как эритроза, которая необходима для биосинтеза ароматических соединений, особенно ароматических аминокислот. Пентозы, образующиеся в ППК, используются для синтеза нуклеиновых кислот. Кроме того, ГАП, образующийся в ФПК, может быть направлен в ЭМИ для выработки энергии через ЦТК и систему переноса электронов.
Некоторые из окислительно-восстановительных реакций, требующих НАДФН 2 в качестве Н-донора:
1. Биосинтез аминокислот. Основная часть поступления неорганического Nh4 в органические соединения – восстановительное аминирование α-кетоглутаровой кислоты с образованием глутаминовой кислоты. Последний затем может переносить аминогруппу путем переаминирования с образованием других аминокислот.
Детали баланса КПП показаны на рис. 8.45. Используемые сокращения: GAP для глицеральдегидфосфата, G6P для глюкозо-6-фосфата, PGLA для фосфоглюконовой кислоты, Ru5P для рибулозо-5-фосфата, Xu5P для ксилулозо-5-фосфата, S7P для седогептулозо-7-фосфата, E4P для эритрозо-4-фосфата, F6P. для фруктозо-6-фосфата и FBP для фруктозодифосфата.
Диссимиляция глюкозы: Путь #
3. Путь Энтнера-Дудорова (EDP) : EDP обнаружен только у некоторых прокариотических организмов, особенно у грамотрицательных бактерий.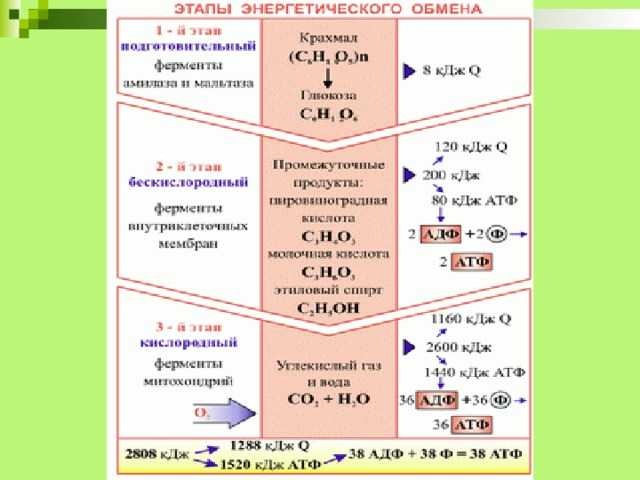 Некоторые из этих бактерий, такие как Pseudomonas saccharophila и Alcaligenes eutrophus, используют исключительно этот путь для диссимиляции глюкозы. У этих организмов явно отсутствуют некоторые ферменты ЭМИ. Некоторые другие организмы, такие как Pseudomonas aeruginosa, используют EDP для расщепления большей части глюкозы, в то время как остальная часть диссимилируется PPC.
Некоторые из этих бактерий, такие как Pseudomonas saccharophila и Alcaligenes eutrophus, используют исключительно этот путь для диссимиляции глюкозы. У этих организмов явно отсутствуют некоторые ферменты ЭМИ. Некоторые другие организмы, такие как Pseudomonas aeruginosa, используют EDP для расщепления большей части глюкозы, в то время как остальная часть диссимилируется PPC.
Характерным промежуточным продуктом EDP является 2-кето-3-дезокси-6-фосфоглюконовая кислота (KDPG). Следовательно, этот путь также известен как путь KDPG. Ключевым ферментом является КДФГ-альдолаза. Первые две реакции ЭДП такие же, как и в ФПК.
Глюкоза фосфорилируется гексокиназой и АТФ с образованием глюкозо-6-фосфата, который затем дегидрируется глюкозо-6-фосфатдегидрогеназой, при этом НАДФ действует как H-акцепторный кофермент с образованием 6-фосфоглюконовой кислоты.
Следующий шаг характерен для ЭОД. Он состоит из дегидратации и дегидрирования 6-фосфоглюконовой кислоты с образованием 2-кето-3-дезокси-6-фосфоглюконовой кислоты (КДФГ). Фермент, катализирующий эту стадию, представляет собой дегидрогеназу 6-фосфоглюконовой кислоты. Затем следует еще одна характерная реакция, катализируемая KDPG-альдолазой, в которой KDPG расщепляется на пировиноградную кислоту и глицеральдегидфосфат (GAP).
Фермент, катализирующий эту стадию, представляет собой дегидрогеназу 6-фосфоглюконовой кислоты. Затем следует еще одна характерная реакция, катализируемая KDPG-альдолазой, в которой KDPG расщепляется на пировиноградную кислоту и глицеральдегидфосфат (GAP).
Из двух продуктов альдолазной реакции КДФГ пировиноградная кислота далее расщепляется по циклу трикарбоновых кислот, а ГАП превращается в другую молекулу пировиноградной кислоты, как это делается в случае ЭМП через 1,3-дифосфоглицериновую кислоту, 3 -фосфоглицериновая кислота, 2-фосфоглицериновая кислота, фосфоенолпировиноградная кислота и, наконец, пировиноградная кислота. Таким образом, чистые продукты окисления одной молекулы глюкозы через ЭДФ такие же, как и в ЭМИ, а именно. две пировиноградные кислоты.
Бухгалтерский баланс ЭДП приведен:
Сравнение ЭДП с ЭМИ показывает, что в обоих продуктах распада глюкозы две молекулы пировиноградной кислоты на молекулу глюкозы, но в ЭМП две молекулы АТФ являются чистым приростом, в то время как в ЭДП образуется только одна АТФ.
Leave A Comment