
Открыт хищный родственник красных водорослей • Сергей Ястребов • Новости науки на «Элементах» • Микробиология, Альгология, Систематика, Эволюция
Не дело философа определять, как далеко простирается царство растений и где начинается класс животных или где начинается жизнь. Это проблемы, относящиеся к отдельным наукам. Скорее всего, до самого конца мира мы так и не узнаем, где в точности проходят эти границы.
Стефан Свежавски, «Святой Фома, прочитанный заново»
Описан новый род одноклеточных организмов, которые являются ближайшими родственниками красных водорослей, но совершенно не похожи на них по образу жизни: это хищные жгутиконосцы, успевшие вторично утратить способность к фотосинтезу. По всей вероятности, это означает, что общие предки красных водорослей и зеленых растений были одноклеточными хищниками, которые даже после приобретения хлоропластов долго сохраняли смешанный тип питания (хищничество + фотосинтез). Жгутиконосец Rhodelphis — это дожившее до наших дней одноклеточное хищное растение.
30 мая 2019 года безвременно, в возрасте всего лишь 66 лет, умер известный российский биолог Александр Петрович Мыльников. Доктор наук, сотрудник Института биологии внутренних вод имени И. Д. Папанина (ИБВВ РАН), он был одним из крупнейших современных протистологов. Мыльников прославился как мастер электронной микроскопии и сделал много открытий, серьезно прояснивших наши представления об эволюции одноклеточных организмов. Установленные им факты уже давно вошли в университетские курсы. Мыльников был очень активным исследователем, признанным во всем мире (см. его страницу на

Клетки в клетках
В последние два с лишним миллиарда лет облик жизни на Земле определяют в основном эукариоты, обладатели сложных клеток с ядрами. К ним относятся животные, растения, грибы и многочисленные одноклеточные существа, которых собирательно называют протистами. Отличие сложной эукариотной клетки от простой прокариотной огромно: не вдаваясь сейчас в детали, вполне можно сказать, что это принципиально разные уровни организации живой материи. Понятно, что проблема становления эукариот очень важна для эволюционной биологии.
Основой для ответа на вопрос «откуда взялись эукариоты?» является симбиогенетическая теория, согласно которой эукариотная клетка возникла в результате взаимовыгодного объединения (симбиоза) двух или нескольких прокариот. Принципиально важно, что при этом происходило вселение одних клеток в другие (эндосимбиоз). Эту теорию нельзя назвать новой: она сформировалась на рубеже XIX–XX веков, после долгого периода забвения возродилась в 1960-х благодаря Линн Маргулис (Lynn Margulis), а на русском языке её современную версию впервые изложил еще в 1973 году Армен Леонович Тахтаджян (А. Л. Тахтаджян, 1973. Четыре царства органического мира (DjVu, 6 Мб)). Молекулярная биология подтверждает симбиогенетическую теорию настолько однозначно, что споры о её верности или неверности давно прекратились. Иное дело — как конкретно всё это происходило. Поток данных, добываемых современными биологами, порождает множество вопросов на эту тему, и исследователи планомерно ищут ответы на них.
Л. Тахтаджян, 1973. Четыре царства органического мира (DjVu, 6 Мб)). Молекулярная биология подтверждает симбиогенетическую теорию настолько однозначно, что споры о её верности или неверности давно прекратились. Иное дело — как конкретно всё это происходило. Поток данных, добываемых современными биологами, порождает множество вопросов на эту тему, и исследователи планомерно ищут ответы на них.
У эукариот есть два типа внутриклеточных структур (органелл), которые точно имеют симбиотическое происхождение, то есть являются бывшими бактериями. Это митохондрии, обеспечивающие дыхание, и хлоропласты, обеспечивающие фотосинтез. В наши дни никто из серьезных биологов не сомневается, что эти органеллы возникли эндосимбиотически. Симбиоз с предками митохондрий начался очень давно, на заре становления эукариот. Есть все основания считать, что у общего предка всех современных эукариот митохондрии уже были (см. Теория симбиогенеза 50 лет спустя: параллельной эукариотизации, скорее всего, не было, «Элементы», 22.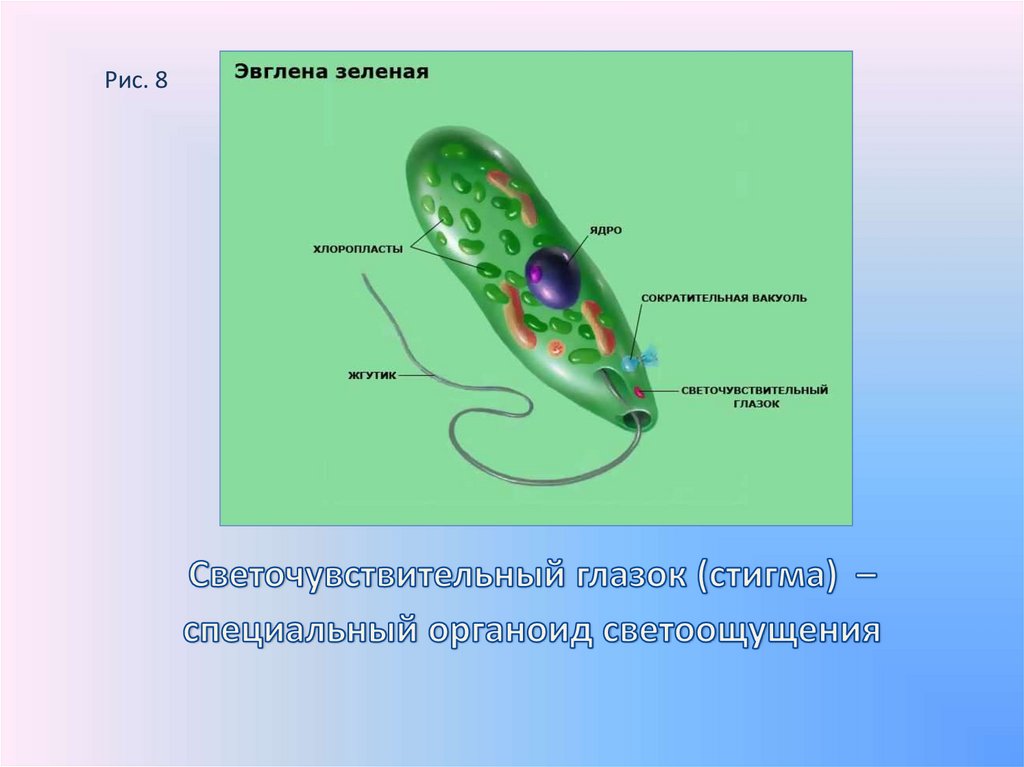 11.2017). Сейчас на наших глазах идут оживленные дискуссии о том, в какой именно момент и по какому сценарию предок эукариот, который сам был не бактерией, а археей, вступил в симбиоз с предком митохондрий (см. Генеалогия белков свидетельствует о позднем приобретении митохондрий предками эукариот, «Элементы», 08.02.2016). Но в любом случае это было до того, как сложились «нормальные» эукариотные клетки, знакомые нам из современной живой природы. В общем, союз с митохондриями по большому счету относится скорее к предыстории эукариот.
11.2017). Сейчас на наших глазах идут оживленные дискуссии о том, в какой именно момент и по какому сценарию предок эукариот, который сам был не бактерией, а археей, вступил в симбиоз с предком митохондрий (см. Генеалогия белков свидетельствует о позднем приобретении митохондрий предками эукариот, «Элементы», 08.02.2016). Но в любом случае это было до того, как сложились «нормальные» эукариотные клетки, знакомые нам из современной живой природы. В общем, союз с митохондриями по большому счету относится скорее к предыстории эукариот.
С хлоропластами дело обстоит совсем иначе. Во-первых, они есть далеко не у всех эукариот: многие современные группы (в том числе и та, к которой относится человек) не имели их никогда. Во-вторых, в отличие от приобретения митохондрий, которое, по-видимому, было уникальным и неповторимым событием, приобретение хлоропластов совершенно точно происходило много раз. Хлоропласты имеют богатейшую историю, насыщенную неожиданными явлениями (см., например: P. J.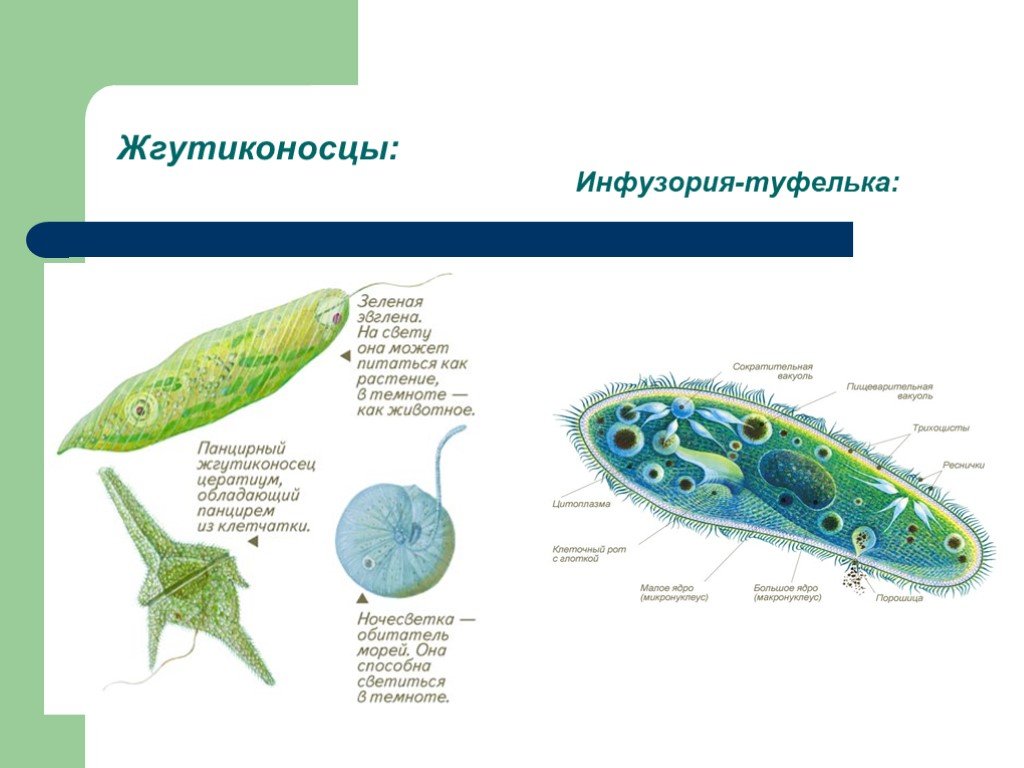 Keeling, 2013. The number, speed, and impact of plastid endosymbioses in eukaryotic evolution; рис. 2). Тому, кто захочет разобраться в ней детально, можно посоветовать сначала взглянуть на современную версию эволюционного древа эукариот (её беглое описание можно найти, например, в статье: Случайно открытый жгутиконосец обновляет систему эукариот, «Элементы», 06.02.2019). Здесь же достаточно сказать, что это древо включает в себя от 6 до 10 (в зависимости от способа подсчета) крупных ветвей, ни одна из которых — увы — не соответствует традиционным царствам животных, растений или грибов. Хорошо это или плохо, но система живой природы, знакомая нам из учебников XX века, ушла в прошлое безвозвратно. Крупные эволюционные ветви эукариот, приблизительно соответствующие по значению классическим царствам, сейчас принято называть супергруппами (supergroup).
Keeling, 2013. The number, speed, and impact of plastid endosymbioses in eukaryotic evolution; рис. 2). Тому, кто захочет разобраться в ней детально, можно посоветовать сначала взглянуть на современную версию эволюционного древа эукариот (её беглое описание можно найти, например, в статье: Случайно открытый жгутиконосец обновляет систему эукариот, «Элементы», 06.02.2019). Здесь же достаточно сказать, что это древо включает в себя от 6 до 10 (в зависимости от способа подсчета) крупных ветвей, ни одна из которых — увы — не соответствует традиционным царствам животных, растений или грибов. Хорошо это или плохо, но система живой природы, знакомая нам из учебников XX века, ушла в прошлое безвозвратно. Крупные эволюционные ветви эукариот, приблизительно соответствующие по значению классическим царствам, сейчас принято называть супергруппами (supergroup).
Краткая история хлоропластов
Итак, предками всех хлоропластов в конечном счете являются цианобактерии, прокариотные организмы, более известные как синезеленые водоросли.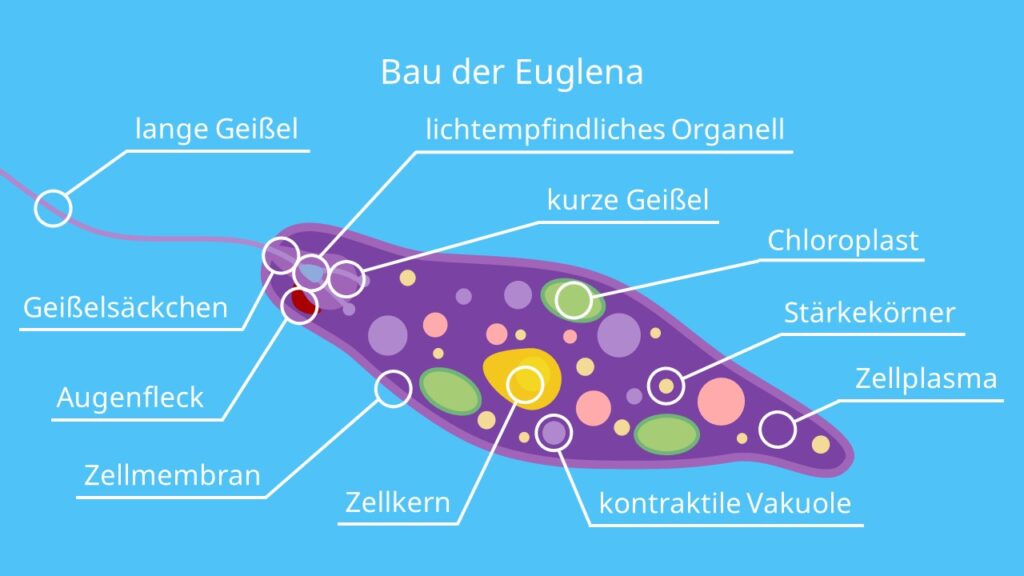 Наша история началась с того, что некий жгутиконосец поглотил цианобактерию, которая превратилась в хлоропласт. Потомки этого жгутиконосца разделились на три эволюционные ветви: микроскопические глаукофитовые водоросли, красные водоросли и зеленые растения (см. рис. 3). Все вместе они образуют супергруппу Archaeplastida. Именно с возникновения эволюционного ствола архепластид началась история фотосинтезирующих эукариот. Другое название этой супергруппы — Plantae, что значит попросту «растения». Здесь и далее мы будем использовать названия «архепластиды» и «растения» как синонимы.
Наша история началась с того, что некий жгутиконосец поглотил цианобактерию, которая превратилась в хлоропласт. Потомки этого жгутиконосца разделились на три эволюционные ветви: микроскопические глаукофитовые водоросли, красные водоросли и зеленые растения (см. рис. 3). Все вместе они образуют супергруппу Archaeplastida. Именно с возникновения эволюционного ствола архепластид началась история фотосинтезирующих эукариот. Другое название этой супергруппы — Plantae, что значит попросту «растения». Здесь и далее мы будем использовать названия «архепластиды» и «растения» как синонимы.
Цианобактерии относятся к грамотрицательным бактериям, главная особенность которых состоит в уникальном строении клеточной оболочки. Любая грамотрицательная бактерия имеет две настоящие клеточные мембраны — внутреннюю и наружную. Между ними находится тонкая клеточная стенка, состоящая, как и у всех бактерий, из пептидогликана. Получается эдакий сэндвич. Когда цианобактерия становится хлоропластом, обе ее мембраны остаются на своем месте.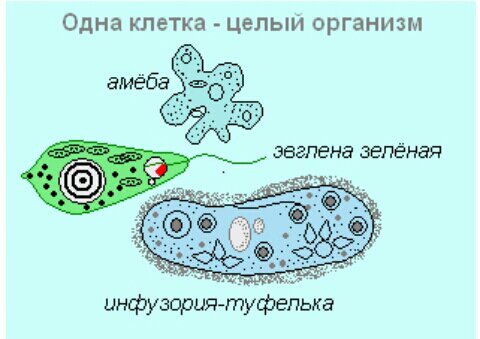
Но это — только часть разнообразия хлоропластов. Другая часть, причем с некоторой точки зрения гораздо более значительная, образуется вот как: хищный эукариот поглощает другого эукариота, внутри которого уже есть первичный хлоропласт, и заставляет этого эукариота в свою очередь работать хлоропластом. Такие хлоропласты называются вторичными. В одних случаях они возникли из зеленых водорослей, в других — из красных. Типичный вторичный хлоропласт имеет четыре мембраны: к двум мембранам цианобактерии добавляется внешняя мембрана первого хозяина (зеленой или красной водоросли) и мембрана пищеварительной вакуоли второго хозяина (хищника, захватившего эту водоросль).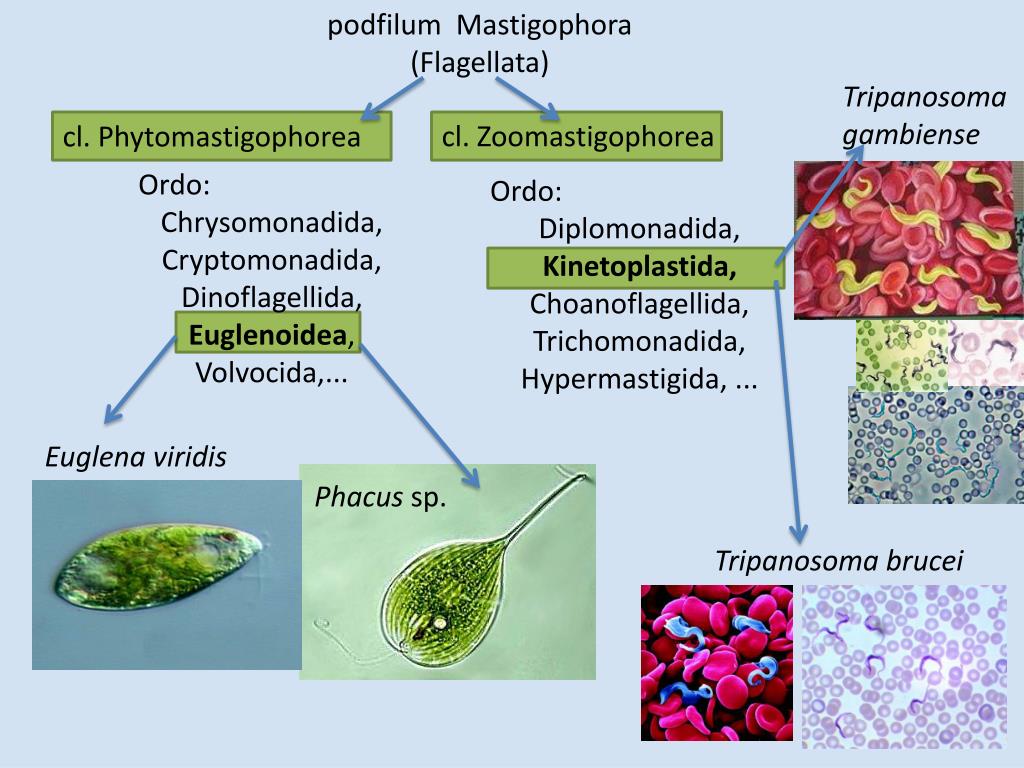
Зеленые водоросли стали вторичными хлоропластами у эвгленовых жгутиконосцев (супергруппа Discoba) и у микроскопических морских хлорарахниофитовых водорослей, которые часто имеют своеобразный облик фотосинтезирующих амеб (супергруппа Rhizaria). Во времена, когда это не было известно, такой характер эволюции изрядно мешал исследователям разобраться в родственных связях эукариот. Например, ботаников постоянно сбивал с толку тот факт, что эвглена зелёная по структуре хлоропластов очень близка к зеленым водорослям, но при этом ни по каким другим признакам ничего общего с зелеными водорослями у нее нет! Разгадка состоит в том, что зеленые водоросли родственны не эвглене — она относится к совершенно другой супергруппе, — а ее хлоропластам.
Судьба тех вторичных хлоропластов, которые образовались из красных водорослей, была еще сложнее. В числе их обладателей — бурые, золотистые, желтозеленые и диатомовые водоросли, входящие в супергруппу Stramenopiles, а также еще три группы преимущественно одноклеточных водорослей: динофлагелляты (супергруппа Alveolata), криптофиты (супергруппа Cryptista) и гаптофиты (супергруппа Haptista; тут надо отметить, что родственные связи криптофит и гаптофит до сих пор неясны, и их отнесение к самостоятельным супергруппам может оказаться временным). Ситуация дополнительно усложняется тем, что многие обладатели вторичных хлоропластов потом их потеряли. Например, в супергруппе Alveolata есть две большие группы, потеря хлоропластов у которых подтверждена электронной микроскопией и молекулярной генетикой, причем обе они фигурируют в школьных учебниках зоологии. Это паразитические споровики, к которым относится малярийный плазмодий, и инфузории, к которым относится всем известная туфелька. Строго говоря, как минимум у споровиков вторичный хлоропласт, происходящий из красной водоросли, даже и не исчез: он сохранился в виде бесцветной структуры, называемой
В числе их обладателей — бурые, золотистые, желтозеленые и диатомовые водоросли, входящие в супергруппу Stramenopiles, а также еще три группы преимущественно одноклеточных водорослей: динофлагелляты (супергруппа Alveolata), криптофиты (супергруппа Cryptista) и гаптофиты (супергруппа Haptista; тут надо отметить, что родственные связи криптофит и гаптофит до сих пор неясны, и их отнесение к самостоятельным супергруппам может оказаться временным). Ситуация дополнительно усложняется тем, что многие обладатели вторичных хлоропластов потом их потеряли. Например, в супергруппе Alveolata есть две большие группы, потеря хлоропластов у которых подтверждена электронной микроскопией и молекулярной генетикой, причем обе они фигурируют в школьных учебниках зоологии. Это паразитические споровики, к которым относится малярийный плазмодий, и инфузории, к которым относится всем известная туфелька. Строго говоря, как минимум у споровиков вторичный хлоропласт, происходящий из красной водоросли, даже и не исчез: он сохранился в виде бесцветной структуры, называемой

В ходе эволюции устройство вторичных хлоропластов может упрощаться. Например, у эвглен и у панцирных водорослей динофлагеллят они имеют уже не четыре мембраны, а три (исчезает, скорее всего, та мембрана, которая соответствует внешней мембране зеленой или красной водоросли — она наименее физиологически необходима). У криптомонад и у хлорарахниофит между второй и третьей мембранами хлоропласта сохраняется нуклеоморф — маленькое, но функционирующее ядро красной (у криптомонад) или зеленой (у хлорарахниофит) поглощенной водоросли. В других группах оно успело исчезнуть.
У некоторых эукариот приобретение хлоропластов происходило неоднократно: они теряли их, а потом опять приобретали. Особенно это свойственно почему-то динофлагеллятам. Такого бешеного разнообразия хлоропластов, как у динофлагеллят, нет ни в одной другой группе эукариот. Представим себе одноклеточную динофлагелляту, которая потеряла хлоропласт, вернулась к хищничеству, а потом «приняла решение» завести хлоропласт заново.
Как мы видим, большинство случаев приобретения эукариотами хлоропластов связано с поглощением не цианобактерий (как можно было бы ожидать), а других эукариот, внутри которых поглощенные цианобактерии уже есть. Таким образом, в конечном счете все эукариоты обязаны своими хлоропластами общему предку супергруппы Archaeplastida, который некогда «приручил» захваченную цианобактерию и тем самым сделал возможными все описанные эволюционные авантюры. Известно одно-единственное исключение: пресноводная раковинная амеба
За этим исключением, вся живая природа Земли обязана своими хлоропластами архепластидам, то есть растениям в эволюционном, а не экологическом смысле этого слова. Вот почему так важно понять, как именно они их приобрели.
Родельфис и эволюция
О том, как появились самые первые хлоропласты, существуют разные мнения. Например, знаменитый биоинформатик Евгений Викторович Кунин с коллегами в свое время отважились допустить, что в эволюционной линии растений вообще никогда не существовало фагоцитоза, то есть свойственного хищным эукариотам механизма активного поглощения пищевых объектов (N. Yutin et al., 2009. The origins of phagocytosis and eukaryogenesis (PDF, 1 Мб)). Тогда способ приобретения первичных хлоропластов архепластид должен быть принципиально иным, чем способ приобретения всех вторичных хлоропластов (и первичных хлоропластов паулинеллы тоже), поскольку уж в этих-то случаях фагоцитоз определенно был задействован. Действительно, предполагать тут можно всякое: ранние этапы эволюции растений, прямо скажем, очень темны.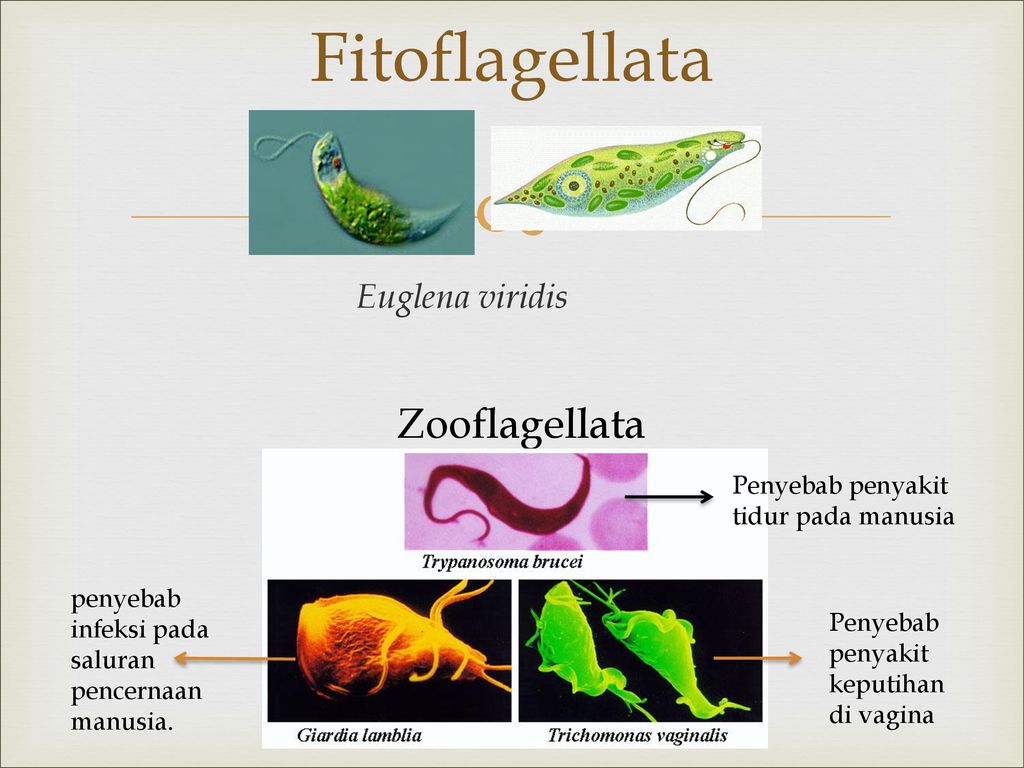
Новое исследование российских, канадских и немецких протистологов бросает луч света на эти таинственные события. Героем статьи стал хищный жгутиконосец Rhodelphis, один вид которого открыли в небольшом пресном озере в бассейне реки Десна, а другой — в морском песке у берега Южно-Китайского моря. По внешнему облику родельфисы — довольно заурядные бесцветные жгутиконосцы с двумя жгутиками. Однако анализ транскриптомов показал, что на эволюционном древе эукариот родельфисы занимают довольно неожиданное положение, а именно — являются сестринской группой по отношению ко всем красным водорослям. Это означает, что они входят в супергруппу Archaeplastida, и не просто входят, а располагаются на древе этой группы довольно близко к ее общему предку, хотя и не у самого корня.
Между тем родельфис вовсе не похож ни на какое растение. Он — настоящий хищник. С помощью ложноножек, образующихся в задней части клетки, он запросто поглощает не только бактерий, но и других эукариот. Надо сказать, что для примитивных эукариот такое хищничество в целом довольно обычно — ни по общему облику, ни по образу жизни, ни по способу питания родельфис среди них особо не выделяется (см. , например: Выделена новая супергруппа эукариот, «Элементы», 18.04.2018). Нет оснований сомневаться, что хищничество родельфиса — примитивная черта. А это, в свою очередь, означает, что предки растений все-таки были хищниками, владевшими фагоцитозом. И более того, даже внутри эволюционного ствола Archaeplastida хищничество продержалось еще довольно долго.
, например: Выделена новая супергруппа эукариот, «Элементы», 18.04.2018). Нет оснований сомневаться, что хищничество родельфиса — примитивная черта. А это, в свою очередь, означает, что предки растений все-таки были хищниками, владевшими фагоцитозом. И более того, даже внутри эволюционного ствола Archaeplastida хищничество продержалось еще довольно долго.
Интересной особенностью родельфисов, которую отметили исследователи, оказались трубчатые кристы митохондрий. Что это значит? Митохондрии образовались из альфа-протеобактерий, которые, как и цианобактерии, являются грамотрицательными. Поэтому митохондрия, точно так же как и первичный хлоропласт, имеет две мембраны — наружную и внутреннюю. Кристами называются складки внутренней мембраны, предназначенные для увеличения ее поверхности (этого требует физиологическая функция митохондрии). Самые распространенные варианты формы крист — пластинчатые и трубчатые, причем еще в 1970-х было замечено, что в крупных эволюционных ветвях этот признак очень стабилен.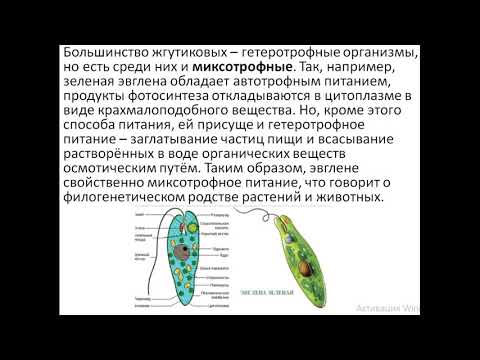 Предлагалось даже разделить большинство эукариот на группы Lamellicristata (с пластинчатыми кристами) и Tubulicristata (с трубчатыми кристами; см. Я. И. Старобогатов, 1986. К вопросу о числе царств эукариотных организмов (DjVu, 1 Мб)). И хотя эта идея столкновения с фактами не выдержала, форма крист митохондрий осталась признаком, важным для крупномасштабной системы. Интересно то, что — как опять же давно известно — у представителей супергруппы Archaeplastida, то есть у растений, кристы, как правило, пластинчатые. Трубчатые кристы родельфиса — редкий для этой супергруппы признак, который может оказаться очень древним.
Предлагалось даже разделить большинство эукариот на группы Lamellicristata (с пластинчатыми кристами) и Tubulicristata (с трубчатыми кристами; см. Я. И. Старобогатов, 1986. К вопросу о числе царств эукариотных организмов (DjVu, 1 Мб)). И хотя эта идея столкновения с фактами не выдержала, форма крист митохондрий осталась признаком, важным для крупномасштабной системы. Интересно то, что — как опять же давно известно — у представителей супергруппы Archaeplastida, то есть у растений, кристы, как правило, пластинчатые. Трубчатые кристы родельфиса — редкий для этой супергруппы признак, который может оказаться очень древним.
Ну а как же с хлоропластами? Они есть у глаукофитовых водорослей, которые, по общепринятой версии, являются древнейшей эволюционной ветвью растений (более древней, чем ветвь красных водорослей и родельфиса). У красных водорослей они тоже есть. Все это означает, что хлоропласты наверняка уже были у ближайшего общего предка всех современных архепластид. Но тогда родельфис тоже должен был бы их получить в наследство. Ну, и где они у него?
Ну, и где они у него?
Электронная микроскопия не обнаружила у родельфиса хлоропластов (во всяком случае, пока). Но вот молекулярная биология, как оказалось, может к этому кое-что добавить. Хорошо известно, что белки, необходимые для работы хлоропластов, но кодируемые ядерными генами, всегда синтезируются за пределами хлоропласта и только потом транспортируются в него. При этом они обязательно распознаются специальными белковыми комплексами, которые встроены во внешнюю и внутреннюю мембраны хлоропласта (см. TIC/TOC complex). Так вот у родельфиса удалось обнаружить не менее четырех белков, входящих в эти комплексы. Нашлись у него и некоторые другие белки, связанные с хлоропластами — в их числе ферредоксин, обычно участвующий в фотосинтезе. В то же время большинство других белков фотосинтеза у родельфиса не найдено, и клетки его, как уже говорилось бесцветные, а не окрашенные, как у фотосинтезирующих растений. Вывод: у родельфиса, вероятно, всё-таки есть первичный хлоропласт, но только маленький, рудиментарный и бесцветный. От фотосинтеза эта эволюционная линия успела отказаться.
От фотосинтеза эта эволюционная линия успела отказаться.
Подводя итог, родельфис вполне можно описать как одноклеточное хищное растение. Причем, что самое главное, его хищность — первичная, унаследованная прямо от общего предка всех современных эукариот, а не вторичная, как у растущей в наших болотах росянки или у триффидов из великого романа Уиндема.
Что касается общих предков родельфисов и красных водорослей, то они скорее всего были миксотрофами, то есть организмами со смешанным типом питания. Они совмещали фотосинтез и хищничество. Затем красные водоросли (как и зеленые растения) отказались от хищничества, а родельфис — от фотосинтеза. Ну а общий предок всех архепластид, вероятно, был обыкновенным хищным жгутиконосцем.
Таким образом, последовательность древнейшей эволюции хлоропластов теперь проясняется. Очевидно, все началось с хищного жгутиконосца, который просто захватывал цианобактерий как пищевые объекты. Потом цианобактерии были «приручены» и наступил этап миксотрофного питания, который, судя по положению родельфисов на эволюционном древе, был достаточно долгим: следуя этой логике, приходится сделать вывод, что даже общий предок красных и зеленых водорослей еще мог быть только миксотрофом.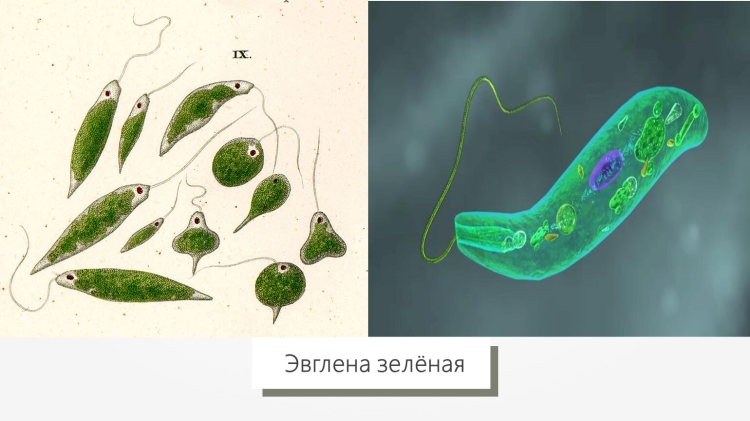 И наконец, в нескольких эволюционных ветвях миксотрофия исчезла. Так возникли растения в наиболее классическом смысле слова — организмы, относящиеся к эволюционной ветви Archaeplastida (= Plantae) и питающиеся только фотосинтезом.
И наконец, в нескольких эволюционных ветвях миксотрофия исчезла. Так возникли растения в наиболее классическом смысле слова — организмы, относящиеся к эволюционной ветви Archaeplastida (= Plantae) и питающиеся только фотосинтезом.
Загадка красных водорослей
Уже упоминалось, что ближайшими современными родственниками родельфисов оказались красные водоросли. Эти организмы давно и хорошо знакомы человеку; многие из них удостоились народных названий, например ирландский мох (Irish moss, Chondrus crispus), в изобилии заселяющий литораль Атлантического океана. Красные водоросли живут почти исключительно в морях, хотя надо отметить, что пресноводные формы среди них все же попадаются: например, известный научно-популярный журнал «Батрахоспермум» получил свое название именно в честь пресноводной красной водоросли. Иногда красные водоросли бывают одноклеточными, но чаще они многоклеточные — нитчатые, пластинчатые или ветвящиеся. Благодаря особым фотосинтетическим пигментам они могут жить глубоко в море, там, где никаким другим водорослям света для фотосинтеза уже не хватает, уходя таким образом от конкуренции.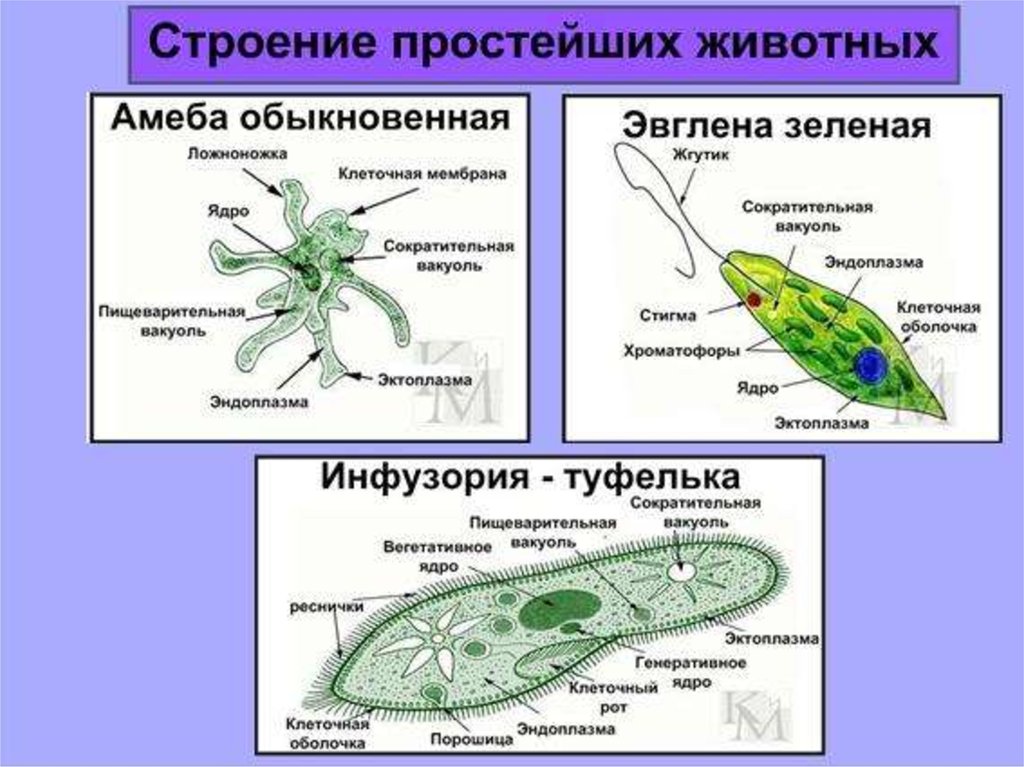 Группа эта очень древняя: самые ранние красные водоросли, обнаруженные палеонтологами, имеют возраст 1,2 миллиарда лет (см. N. J. Butterfield, 2015. Proterozoic photosynthesis — a critical review). Причем они относятся к современному классу бангиевых водорослей, которые, таким образом, являют собой поразительный по глубине пример живых ископаемых (для сравнения — ближайшие ископаемые родичи прославленной латимерии имеют возраст всего-то 70 миллионов лет). По-видимому, это самые древние живые ископаемые среди всех многоклеточных организмов — во всяком случае, эукариотных (вездесущие колониальные цианобактерии не в счет).
Группа эта очень древняя: самые ранние красные водоросли, обнаруженные палеонтологами, имеют возраст 1,2 миллиарда лет (см. N. J. Butterfield, 2015. Proterozoic photosynthesis — a critical review). Причем они относятся к современному классу бангиевых водорослей, которые, таким образом, являют собой поразительный по глубине пример живых ископаемых (для сравнения — ближайшие ископаемые родичи прославленной латимерии имеют возраст всего-то 70 миллионов лет). По-видимому, это самые древние живые ископаемые среди всех многоклеточных организмов — во всяком случае, эукариотных (вездесущие колониальные цианобактерии не в счет).
У красных водорослей есть одна удивительная особенность, давно ставящая биологов в тупик. Это — полное отсутствие жгутиков. В этой статье уже не раз встречалось слово «жгутиконосцы», которое, собственно, и обозначает эукариот, имеющих жгутики. Когда-то такими были все эукариоты без исключения. Жгутики — это универсальные двигательные структуры, унаследованные современными эукариотами от их одноклеточного общего предка.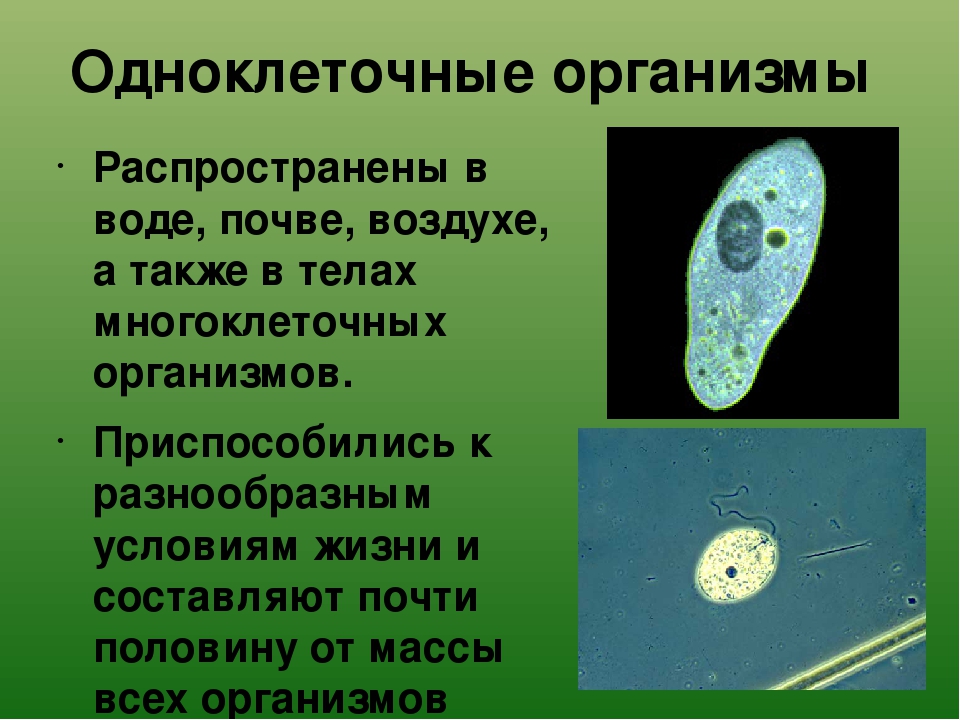 Они часто сохраняются и после перехода к многоклеточности: например, у большинства многоклеточных животных, включая человека, с помощью жгутиков движутся сперматозоиды. Случаи отказа от жгутиков не очень многочисленны и связаны, как правило, с наземным образом жизни. Например, жгутики совершенно утрачены у высших грибов и у цветковых растений. Этим организмам они просто не нужны: жизненные циклы высших грибов и цветковых растений рассчитаны на полностью сухопутное существование, поэтому размножение у них давным-давно идет без участия активно плавающих половых клеток. Но вот многоклеточные животные такой высокой степени адаптации к наземности не достигли. А красные водоросли — это вообще чисто водные существа. Никаких видимых причин для отказа от жгутиков у них нет. Однако ни у одной известной красной водоросли жгутики не появляются ни на какой стадии жизненного цикла. Это очень странно, тем более что красные водоросли испытывают по этой причине явные трудности: например, из-за того, что мужские половые клетки у них неподвижны, им приходится «изобретать» специальные сложные структуры для оплодотворения, хотя со жгутиковыми половыми клетками всё было бы намного проще.
Они часто сохраняются и после перехода к многоклеточности: например, у большинства многоклеточных животных, включая человека, с помощью жгутиков движутся сперматозоиды. Случаи отказа от жгутиков не очень многочисленны и связаны, как правило, с наземным образом жизни. Например, жгутики совершенно утрачены у высших грибов и у цветковых растений. Этим организмам они просто не нужны: жизненные циклы высших грибов и цветковых растений рассчитаны на полностью сухопутное существование, поэтому размножение у них давным-давно идет без участия активно плавающих половых клеток. Но вот многоклеточные животные такой высокой степени адаптации к наземности не достигли. А красные водоросли — это вообще чисто водные существа. Никаких видимых причин для отказа от жгутиков у них нет. Однако ни у одной известной красной водоросли жгутики не появляются ни на какой стадии жизненного цикла. Это очень странно, тем более что красные водоросли испытывают по этой причине явные трудности: например, из-за того, что мужские половые клетки у них неподвижны, им приходится «изобретать» специальные сложные структуры для оплодотворения, хотя со жгутиковыми половыми клетками всё было бы намного проще.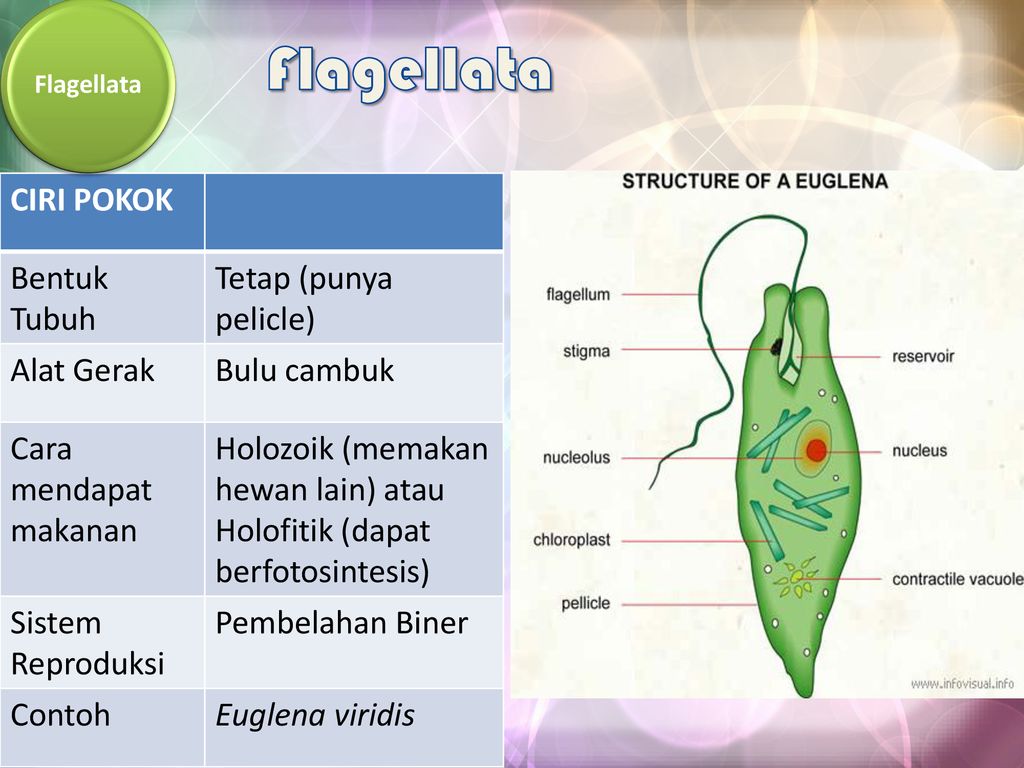
Лет тридцать назад было популярно мнение, что красные водоросли — это чрезвычайно примитивные эукариоты, у которых жгутики еще просто не успели появиться. Увы, современная филогенетика отвергает эту гипотезу напрочь. Здесь надо искать какое-то другое объяснение.
В 2015 году вышла очень интересная работа, указывающая путь к решению этой проблемы (H. Qiu et al., 2015. Evidence of ancient genome reduction in red algae (Rhodophyta)). Она посвящена геномам красных водорослей. Прежде всего, оказалось, что эти геномы по меркам эукариот маленькие — всего 5–10 тысяч генов. Причем это относится не только к одноклеточным, но и к достаточно крупным и сложным многоклеточным красным водорослям, включая тот же ирландский мох. Детальный анализ показал, что многие гены, свойственные почти всем эукариотам, у красных водорослей почему-то потеряны. В частности, у них отсутствует целый набор генов, обеспечивающих сборку жгутиков и работу центриолей (опорных структур, которые всегда располагаются в основаниях жгутиков).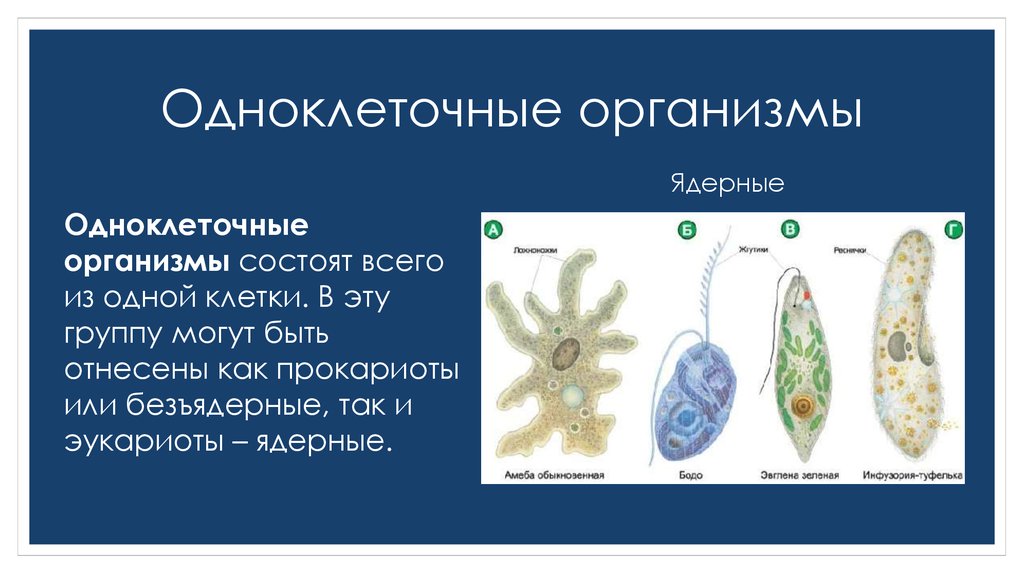 Красные водоросли не образуют жгутиков не потому, что им это не пригодилось бы, а потому, что они на это физически неспособны: у них нет генов, кодирующих необходимые белки. Ни жгутик, ни центриоль они сформировать не могут. Кроме того, они потеряли ряд генов, никак не связанных со жгутиками, но имеющих отношение к разным другим клеточным структурам и механизмам. Например, у красных водорослей не оказалось генов, обычно контролирующих синтез некоторых компонентов клеточных мембран (см. гликозилфосфатидилинозитол). Причем известно, что для многих самых разных эукариот — грибы, трипаносома, мышь — потеря этих генов вообще смертельна. За счет утраты множества генов у красных водорослей заметно обеднен метаболизм, то есть обмен веществ; некоторые биохимические пути у них целиком исчезли.
Красные водоросли не образуют жгутиков не потому, что им это не пригодилось бы, а потому, что они на это физически неспособны: у них нет генов, кодирующих необходимые белки. Ни жгутик, ни центриоль они сформировать не могут. Кроме того, они потеряли ряд генов, никак не связанных со жгутиками, но имеющих отношение к разным другим клеточным структурам и механизмам. Например, у красных водорослей не оказалось генов, обычно контролирующих синтез некоторых компонентов клеточных мембран (см. гликозилфосфатидилинозитол). Причем известно, что для многих самых разных эукариот — грибы, трипаносома, мышь — потеря этих генов вообще смертельна. За счет утраты множества генов у красных водорослей заметно обеднен метаболизм, то есть обмен веществ; некоторые биохимические пути у них целиком исчезли.
Авторы делают твердый вывод: все современные красные водоросли произошли от «генетически обнищавшего предка» (gene depauperate ancestor). И это «обнищание» в самом деле составляет для них проблему: показано, что для восполнения недостающих элементов метаболизма красные водоросли склонны «рекрутировать» бактериальные гены, проникающие в их клетки путем постоянно идущего в природе горизонтального переноса генов (ГПГ).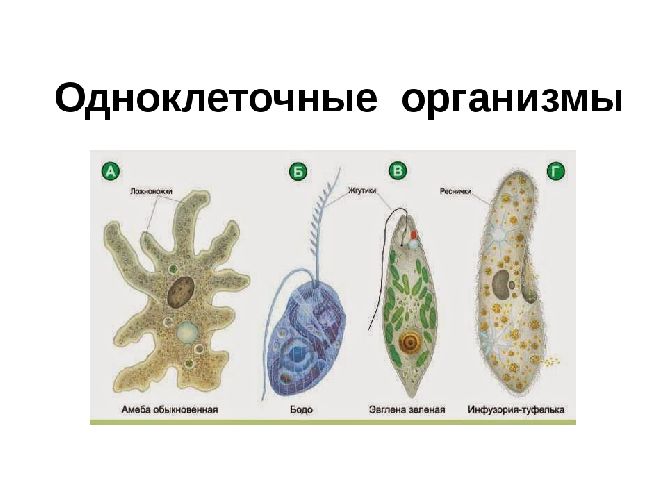 А вот жгутики восстановить так легко не получается.
А вот жгутики восстановить так легко не получается.
Возможно, что именно из-за сужения адаптивных возможностей, вызванного массовой потерей генов, красные водоросли не смогли завоевать сушу (J. Collen, 2015. Win some, lose some: genome evolution in red algae (PDF)). В конце концов, почему бы земным лесам и травам не быть красными? Однако зеленым растениям освоить сушу удалось, а красным водорослям нет. Зато красные водоросли внесли в эволюцию жизни на Земле другой важный вклад: как мы уже знаем, они «подарили» хлоропласты многим другим группам эукариот — бурым водорослям и иже с ними.
У современных морских или пресноводных красных водорослей нет ни малейших причин отказываться ни от жгутиков, ни от многих других полезных вещей. Это означает, что их эволюционный путь был на редкость непрямым. Судя по всему, в начале эволюционной ветви красных водорослей произошла какая-то генетическая катастрофа: за короткое (конечно, по эволюционным меркам) время они, по грубой оценке, потеряли около четверти генов, свойственных зеленым растениям и большинству других эукариот.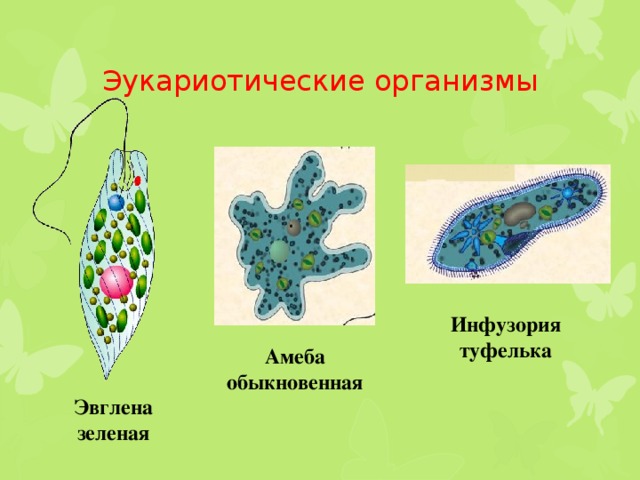 Почему это произошло — пока никто не знает. Может быть, предки красных водорослей на каком-то эволюционном этапе были паразитами — ведь известно, что паразитизм часто приводит к редукции генома (см. Бубонная чума была уже 3800 лет назад, «Элементы», 15.08.2018). Но тогда совершенно непонятно, почему они сохранили фотосинтез. А может быть, потеря генов была связана с жизнью в каком-то экстремальном местообитания, но в каком — пока трудно даже представить. Интересующая нас «генная катастрофа» должна была произойти не менее полутора миллиардов лет назад (позже разные ветви красных водорослей уже разделились), так что реконструировать ее условия — дело не самое простое.
Почему это произошло — пока никто не знает. Может быть, предки красных водорослей на каком-то эволюционном этапе были паразитами — ведь известно, что паразитизм часто приводит к редукции генома (см. Бубонная чума была уже 3800 лет назад, «Элементы», 15.08.2018). Но тогда совершенно непонятно, почему они сохранили фотосинтез. А может быть, потеря генов была связана с жизнью в каком-то экстремальном местообитания, но в каком — пока трудно даже представить. Интересующая нас «генная катастрофа» должна была произойти не менее полутора миллиардов лет назад (позже разные ветви красных водорослей уже разделились), так что реконструировать ее условия — дело не самое простое.
Открытие родельфиса добавляет к складывающемуся «пазлу» очень важный фрагмент. У родельфиса нет ни малейших признаков той массивной потери генов, которая произошла у красных водорослей. Все те гены, отсутствие которых специфично для красных водорослей, у него оказались на месте. Общий предок красных водорослей и родельфисов был в этом отношении совершенно «нормален». Особенно интересно, что геном родельфиса богат внутригенными некодирующими последовальностями — интронами, в то время как у красных водорослей число интронов резко уменьшено. Дело в том, что интроны сами по себе нефункциональны. Их массовая потеря обычно бывает связана просто с уменьшением общего размера генома, которое, в свою очередь, связано с уменьшением размера клеток (см. Геномы хвостатых амфибий с самого начала были большими, «Элементы», 24.06.2015). Итак, возможно, что одним из факторов постигшей красные водоросли «генной катастрофы» было предельное уменьшение абсолютного размера клеток, случившееся на стадии, когда эти водоросли еще были одноклеточными. Такие примеры в других эволюционных ветвях известны (H. Qiu, 2016. Travel light: reductive genome evolution in free-living eukaryotes (PDF, 1 Мб)). Полной разгадки тайны красных водорослей эти соображения не дают, но ключ к ней — быть может.
Особенно интересно, что геном родельфиса богат внутригенными некодирующими последовальностями — интронами, в то время как у красных водорослей число интронов резко уменьшено. Дело в том, что интроны сами по себе нефункциональны. Их массовая потеря обычно бывает связана просто с уменьшением общего размера генома, которое, в свою очередь, связано с уменьшением размера клеток (см. Геномы хвостатых амфибий с самого начала были большими, «Элементы», 24.06.2015). Итак, возможно, что одним из факторов постигшей красные водоросли «генной катастрофы» было предельное уменьшение абсолютного размера клеток, случившееся на стадии, когда эти водоросли еще были одноклеточными. Такие примеры в других эволюционных ветвях известны (H. Qiu, 2016. Travel light: reductive genome evolution in free-living eukaryotes (PDF, 1 Мб)). Полной разгадки тайны красных водорослей эти соображения не дают, но ключ к ней — быть может.
Источник: Ryan M. R. Gawryluk, Denis V. Tikhonenkov, Elisabeth Hehenberger, Filip Husnik, Alexander P. Mylnikov and Patrick J. Keeling. Non-photosynthetic predators are sister to red algae // Nature. 2019. Published 17 July 2019. DOI: 10.1038/s41586-019-1398-6
Mylnikov and Patrick J. Keeling. Non-photosynthetic predators are sister to red algae // Nature. 2019. Published 17 July 2019. DOI: 10.1038/s41586-019-1398-6
Сергей Ястребов
Саркомастигофоры | Саркодовые | Амеба протей (обыкновенная), амеба дизентерийная, радиолярия | |
Жгутиковые | Эвглена зеленая, вольвокс, трипаносома африканская, лейшмания, трихомонада, лямблия печеночная | ||
Споровики | Кокцидиеобразные | Малярийный плазмодий | |
Инфузории | Ресничные | Инфузоря-балантидий, инфузория-туфелька, инфузория-трубач | |
Трихофриоз |
Признаки простейших одноклеточных | Амеба обыкновенная (Класс Корненожки) | Эвглена зеленая (класс Жгутиконосцы) | Инфузория ту-фелька (класс Ин-фузории) |
Строение | Состоит из цитоплазмы, ядра, сократительной вакуоли, ложноножки, пищеварительной вакуоли (см. рис) | Состоит из оболочки, ядра, жгутика, глазок, сократительной вакуоли, питательных веществ, хлоропласты (см. рис.) | Состоит из мембраны, малого и большого ядра, сократительной и пищеварительной вакуоли, рот, порошица, реснички (см. |
Движение | «Перетекание» с по-мощью ложноножек | Передвижение с помощью жгутика | Передвижение с помощью рес-ничек |
Кормом могут быть бактерии, микроскопические водоросли. Амёба захватывает пищу, вытягивая ложноножки на любом участке тела. Они обволакивают добычу и вместе с небольшим количеством воды погружают её в цитоплазму. Так образуется пищеварительная вакуоль — фагоцитоз, за-хват капель жидкости — пиноцитоз. Из пищеварительной вакуоли растворимые продукты пищеварения поступают в цитоплазму, а непереваренные остатки выводятся из организма в любой части клетки. | Автотрофное (фото-синтез) или гетеро-трофное (фагоцитоз и пиноцитоз) | Питаются различными микроорганизмами, преимущественно бактериями. Движением ресничек, расположенных вдоль ротового углубления, загоняют в него добычу. Вместе с водой она попадает в клеточный рот, затем в глотку. | |
Размноже-ние | Амеба размножается делением. При этом ядро делится надвое. Образовавшиеся новые ядра расходятся в стороны, и между ними появляется поперечная перетяжка, разделяющая амёбу на две дочерние клетки, которые живут самостоятельно. Через некоторое время молодые амёбы также начинают делиться. Для размножения благоприятна температура воды около +20 °С. | Размножение организмов данного вида эвглен бесполое — делением клетки пополам, в отличие от инфузории-туфельки, для которой характерен еще и половой процесс. | Инфузории размножаются бесполым путём — поперечным делением, как амёбы. Первым делится надвое малое ядро, затем большое. Одновременно появляется поперечная перетяжка. Она со временем разделяет инфузорию на две молодые (дочерние) клетки. Они растут и при хорошем питании и оптимальной температуре уже на следующие сутки становятся взрослыми и снова могут делиться. Для инфузорий характерен и половой процесс в форме конъюгации (слия-ние двух клеток и обмен генети-ческой информа-цией) |
_______________
Источник информации: Биология в таблицах и схемах./ Издание 2е, — СПб.: 2004.
Класс Жгутиковые
Строение . У жгутиковых имеются жгутики, служащие органоидами движения и способствующие захвату пищи. Их может быть один, два или множество. Движением жгутика в окружающей воде вызывается водоворот, благодаря которому мелкие взвешенные в воде частички увлекаются к основанию жгутика, где имеется небольшое отверстие — клеточный рот, ведущий в глубокий канал-глотку.
Почти все жгутиковые покрыты плотной эластичной оболочкой, которая наряду с развитыми элементами цитоскелета определяет постоянную форму тела.
Генетический аппарат у большинства жгутиковых представлен одним ядром, но существуют также двуядерные (например, лямблии) и многоядерные (например, опалина) виды.
Цитоплазма четко делится на тонкий наружный слой — прозрачную эктоплазму и глубже лежащую эндоплазму.
Способ питания. По способу питания жгутиковые делятся на три группы. Автотрофные организмы как исключение в царстве животных синтезируют органические вещества (углеводы) из углекислого газа и воды при помощи хлорофилла и энергии солнечного излучения. Хлорофилл находится в хроматофорах, сходных по организации с пластидами растений. У многих жгутиконосцев с растительным типом питания имеются особые аппараты, воспринимающие световые раздражения, — стигмы.
Гетеротрофные организмы (трипаносома — возбудитель сонной болезни) не имеют хлорофилла и поэтому не могут синтезировать углеводы из неорганических веществ. Миксотрофные организмы способны к фотосинтезу, но питаются также минеральными и органическими веществами, созданными другими организмами (эвглена зеленая).
Осморегуляторная и отчасти выделительная функции выполняются у жгутиковых, как у саркодовых, сократительными вакуолями, которые имеются у свободноживущих пресноводных форм.
Размножение. У жгутиковых отмечается половое и бесполое размножение. Обычная форма бесполого размножения — продольное деление.
Среда обитания. Жгутиковые широко распространены в пресных водоемах, особенно небольших и загрязненных органическими остатками, а также в морях. Многие виды паразитируют у различных животных и человека и тем самым приносят большой вред (трипоносомы, паразиты кишечника и др.).
Общими чертами организации простейших являются следующие:
Большинство простейших-одноклеточные, реже колониальные организмы. Их одноклеточное тело обладает функциями целостного организма, которые выполняются органеллами общего назначения (ядро, эндоплазматическая сеть, комплекс Гольджи, лизосомы, митохондрии, рибосомы и др.) и специального (пищеварительные и сократительные вакуоли, жгутики, реснички и др.). Согласованно функционируя, они обеспечивают отдельной клетке возможность существования в качестве самостоятельного организма.
Покровы
простейших представлены либо
только плазматической
мембраной, либо
еще и плотной, довольно гибкой и
эластичной оболочкой — пелликулой, придающей
им относительное постоянство формы
тела. В цитоплазме четко различаются
два слоя: поверхностный, более плотный
—эктоплазма, и
внутренний, более жидкий и зернистый
— эндоплазма, в
которой располагаются органеллы
простейшего. Благодаря коллоидным
свойствам цитоплазмы эти два слоя могут
взаимно переходить друг в друга.
Органоиды движения большинства видов — ложноножки, жгутики или многочисленные короткие реснички.
У пресноводных одноклеточных имеется 1 -2 сократительные вакуоли, основная функция которых состоит в поддержании постоянства осмотического давления, осуществляемого за
Раздражимость у простейших проявляется в форме таксисов.
Большинство
простейших обладает способностью
переносить неблагоприятные условия в
состоянии покоящейся стадии —цисты. При
этом клетка округляется, втягивает или
отбрасывает органоиды движения и
покрывается плотной защитной оболочкой.
Стадия цисты дает возможность простейшему
не только переживать в неактивном
состоянии неблагоприятные условия, но
и расселяться. Попав в благоприятные
условия, простейшее покидает оболочку
цисты и начинает питаться и размножаться.
Простейшие подразделяются на классы: Корненожки, Жгутиковые, Инфузории, Споровики.
Эволюция одноклеточных, растительного и животного мира
Эволюция одноклеточных организмов
До
1950-х годов не удавалось обнаружить следы
докембрийской жизни на уровне одноклеточных
организмов, поскольку микроскопические
останки этих существ невозможно выявить
обычными методами палеонтологии. Важную
роль в их обнаружении сыграло открытие,
сделанное в начале XX в. Ч. Уолкотом. В
докембрийских отложениях на западе
Северной Америки он нашел слоистые
известняковые образования в виде
столбов, названные позднее строматолитами.
В 1954 г. было установлено, что строматолиты
формации Ганфлинт (Канада) образованы
останками бактерий и сине-зеленых
водорослей. У берегов Австралии обнаружены
и живые строматолиты, состоящие из этих
же организмов и очень сходные с ископаемыми
докембрийскими строматолитами. К
настоящему времени остатки микроорганизмов
найдены в десятках строматолитов, а
также в глинистых сланцах морских
побережий.
Самые ранние из бактерий (прокариоты) существовали уже около 3,5 млрд. лет назад. К настоящему времени сохранились два семейства бактерий: древние, или археобактерии (галофильные, метановые, термофильные), и эубактерии (все остальные). Таким образом, единственными живыми существами на Земле в течение 3 млрд. лет были примитивные микроорганизмы. Возможно, они представляли собой одноклеточные существа, сходные с современными бактериями, например клостридиями, живущими на основе брожения и использования, богатых энергией органических соединений, возникающих абиогенно под действием электрических разрядов и ультрафиолетовых лучей. Следовательно, в эту эпоху живые существа были потребителями органических веществ, а не их производителями.
Гигантский
шаг на пути эволюции жизни был связан
с возникновением основных биохимических
процессов обмена — фотосинтеза и дыхания
и с образованием клеточной организации,
содержащей ядерный аппарат (эукариоты). Эти «изобретения», сделанные еще на
ранних стадиях биологической эволюции,
в основных чертах сохранились у
современных организмов. Методами
молекулярной биологии установлено
поразительное единообразие биохимических
основ жизни при огромном различии
организмов по другим признакам. Белки
почти всех живых существ состоят из 20
аминокислот. Нуклеиновые кислоты,
кодирующие белки, монтируются из четырех
нуклеотидов. Биосинтез белка осуществляется
по единообразной схеме, местом их синтеза
являются рибосомы, в нем участвуют и-РНК
и т-РНК. Подавляющая часть организмов
использует энергию окисления, дыхания
и гликолиза, которая запасается в АТФ.
Рассмотрим
подробнее особенности эволюции на
клеточном уровне организации жизни.
Наибольшее различие существует не между
растениями, грибами и животными, а между
организмами, обладающими ядром (эукариоты)
и не имеющими его (прокариоты). Последние
представлены низшими организмами —
бактериями и сине-зелеными водорослями
(цианобактерии, или цианеи), все остальные
организмы — эукариоты, которые сходны
между собой по внутриклеточной
организации, генетике, биохимии и
метаболизму.
Различие между прокариотами и эукариотами заключается еще и в том, что первые могут жить как в бескислородной (облигатные анаэробы), так и в среде с разным содержанием кислорода (факультативные анаэробы и аэробы), в то время как для эукариотов, за немногим исключением, обязателен кислород. Все эти различия имели существенное значение для понимания ранних стадий биологической эволюции.
Сравнение прокариот и эукариот по потребности в кислороде приводит к заключению, что прокариоты возникли в период, когда содержание кислорода в среде изменилось. Ко времени же появления эукариот концентрация кислорода была высокой и относительно постоянной.
Первые
фотосинтезирующие организмы появились
около 3 млрд. лет назад. Это были анаэробные
бактерии, предшественники современных
фотосинтезирующих бактерий. Предполагается,
что именно они образовали самые древние
среди известных строматолитов. Обеднение
среды азотистыми органическими
соединениями вызывало появление живых
существ, способных использовать
атмосферный азот. Такими организмами,
способными существовать в среде,
полностью лишенной органических
углеродистых и азотистых соединений,
являются фотосинтезирующие азотфиксирующие
сине-зеленые водоросли. Эти организмы
осуществляли аэробный фотосинтез. Они
устойчивы к продуцируемому ими кислороду
и могут использовать его для собственного
метаболизма. Поскольку сине-зеленые
водоросли возникли в период, когда
концентрация кислорода в атмосфере
колебалась, вполне допустимо, что они
— промежуточные организмы между
анаэробами и аэробами.
С уверенностью предполагается, что фотосинтез, в котором источником атомов водорода для восстановления углекислого газа является сероводород (такой фотосинтез осуществляют современные зеленые и пурпурные серные бактерии), предшествовал более сложному двустадийному фотосинтезу, при котором атомы водорода извлекаются из молекул воды. Второй тип фотосинтеза характерен для цианей и зеленых растений.
Фотосинтезирующая
деятельность первичных одноклеточных
имела три последствия, оказавшие решающее
влияние на всю дальнейшую эволюцию
живого. Во-первых, фотосинтез освободил
организмы от конкуренции за природные
запасы абиогенных органических
соединений, количество которых в среде
значительно сократилось. Развившееся
посредством фотосинтеза автотрофное
питание и запасание питательных готовых
веществ в растительных тканях создали
затем условия для появления громадного
разнообразия автотрофных и гетеротрофных
организмов. Во-вторых, фотосинтез
обеспечивал насыщение атмосферы
достаточным количеством кислорода для
возникновения и развития организмов,
энергетический обмен которых основан
на процессах дыхания. В-третьих, в
результате фотосинтеза в верхней части
атмосферы образовался озоновый экран,
защищающий земную жизнь от губительного
ультрафиолетового излучения космоса,
Еще
одно существенное отличие прокариот и
эукариот заключается в том, что у вторых
центральным механизмом обмена является
дыхание, у большинства же прокариот
энергетический обмен осуществляется
в процессах брожения. Сравнение
метаболизма прокариот и эукариот
приводит к выводу об эволюционной связи
между ними. Вероятно, анаэробное брожение
возникло на более ранних стадиях
эволюции. После появления в атмосфере
достаточного количества свободного
кислорода аэробный метаболизм оказался
намного выгоднее, так как при окислении
углеводов в 18 раз увеличивается выход
биологически полезной энергии в сравнении
с брожением. Таким образом, к анаэробному
метаболизму присоединился аэробный
способ извлечения энергии одноклеточными
организмами.
Когда же появились эукариотические клетки? На этот вопрос нет точного ответа, но значительное количество данных об ископаемых эукариотах позволяет сказать, что их возраст составляет около 1,5 млрд. лет. Относительно того, каким образом возникли эукариоты, существуют две гипотезы.
Одна
из них (аутогенная гипотеза) предполагает,
что эукариотическая клетка возникла
путем дифференциации исходной
прокариотической клетки. Вначале
развился мембранный комплекс: образовалась
наружная клеточная мембрана с впячиваниями
внутрь клетки, из которой сформировались
отдельные структуры, давшие начало
клеточным органоидам. От какой именно
группы прокариот возникли эукариоты,
сказать невозможно.
Другую
гипотезу (симбиотическую) предложила
недавно американский ученый Маргулис.
В ее обоснование она положила новые
открытия, в частности обнаружение у
пластид и митохондрий внеядерной ДНК
и способности этих органелл к
самостоятельному делению. Л. Маргулис
предполагает, что эукарио-тическая
клетка возникла вследствие нескольких
актов симбиогенеза. Вначале произошло
объединение крупной амебовидной
прокариотной клетки с мелкими аэробными
бактериями, которые превратились в
митохондрии. Затем эта симбиотическая
прокариотная клетка включила в себя
спирохетоподобные бактерии, из которых
сформировались кинетосомы, центросомы
и жгутики. После обособления ядра в
цитоплазме (признак эукариот) клетка с
этим набором органелл оказалась исходной
для образования царств грибов и животных.
Объединение прокариотной клетки с
цианеями привело к образованию пластидной
клетки, что дало начало формированию
царства растений. Гипотеза Маргулис
разделяется не всеми и подвергается
критике. Большинство авторов придерживается
аутогенной гипотезы, более соответствующей
дарвиновским принципам монофилии,
дифференциации и усложнения организации
в ходе прогрессивной эволюции.
В эволюции одноклеточной организации выделяются промежуточные ступени, связанные с усложнением строения организма, совершенствованием генетического аппарата и способов размножения.
Самая примитивная стадия — агамная прокариотная — представлена цианеями и бактериями. Морфология этих организмов наиболее проста в сравнении с другими одноклеточными (простейшими). Однако уже на этой стадии появляется дифференциация на цитоплазму, ядерные элементы, базальные зерна, цитоплазматическую мембрану. У бактерий известен обмен генетическим материалом посредством конъюгации. Большое разнообразие видов бактерий, способность существовать в самых разных условиях среды свидетельствуют о высокой адаптивности их организации.
Следующая
стадия — агамная эукариотная —
характеризуется дальнейшей дифференциацией
внутреннего строения с формированием
высокоспециализированных органоидов
(мембраны, ядро, цитоплазма, рибосомы,
митохондрии и др. ). Особо существенной
здесь была эволюция ядерного аппарата
— образование настоящих хромосом в
сравнении с прокариотами, у которых
наследственное вещество диффузно
распределено по всей клетке. Эта стадия
характерна для простейших, прогрессивная
эволюция которых шла по пути увеличения
числа одинаковых органоидов (полимеризация),
увеличения числа хромосом в ядре
(полиплоидизация), появления генеративных
и вегетативных ядер — макронуклеуса и
микронуклеуса (ядерный дуализм). Среди
одноклеточных эукариотных организмов
имеется много видов с агамным размножением
(голые амебы, раковинные корненожки,
жгутиконосцы).
Прогрессивным
явлением в филогенезе простейших было
возникновение у них полового размножения
(гамогонии), которое отличается от
обычной конъюгации. У простейших имеется
мейоз с двумя делениями и кроссинговером
на уровне хроматид, и образуются гаметы
с гаплоидным набором хромосом. У некоторых
жгутиковых гаметы почти неотличимы от
бесполых особей и нет еще разделения
на мужские и женские гаметы, т. е.
наблюдается изогамия. Постепенно в ходе
прогрессивной эволюции происходит
переход от изогамии к анизогамии, или
разделению генеративных клеток на
женские и мужские, и к анизогамной
копуляции. При слиянии гамет образуется
диплоидная зигота. Следовательно, у
простейших наметился переход от агамной
эукариотной стадии к зиготной — начальной
стадии ксеногамии (размножение путем
перекрестного оплодотворения). Последующее
развитие уже многоклеточных организмов
шло по пути совершенствования способов
ксеногамного размножения.
Животные, состоящие из единственной клетки, располагающей ядром, называются одноклеточными организмами.
В них сочетаются характерные особенности клетки и независимого организма.
Одноклеточные животные
Животные подцарства Одноклеточных или Простейших обитают в жидких средах. Внешние формы их разнообразны — от аморфных особей, не имеющих определенных очертаний, до представителей со сложными геометрическими формами.
Насчитывается около 40 тысяч видов одноклеточных животных. К наиболее известным относятся:
- амеба;
- зеленая эвглена;
- инфузория-туфелька.
Амеба
Принадлежит классу корненожки и отличается непостоянной формой.
Она состоит из оболочки, цитоплазмы, сократительной вакуоли и ядра.
Усвоение питательных веществ осуществляется с помощью пищеварительной вакуоли, а кормом служат другие простейшие, такие как водоросли и . Для респирации амебе необходим кислород, растворенный в воде и проникающий через поверхность тела.
Зеленая эвглена
Обладает вытянутой веерообразной формой. Питается за счет превращения углекислого газа и воды в кислород и продукты питания благодаря световой энергии, а также готовыми органическими веществами при отсутствии света.
Относится к классу жгутиковые.
Инфузория-туфелька
Класс инфузории, своими очертаниями напоминает туфельку.
Пищей служат бактерии.
Одноклеточные грибы
Грибы отнесены к низшим бесхлорофилльным эукариотам. Они отличаются наружным пищеварением и содержанием хитина в клеточной стенке. Тело образует грибницу, состоящую из гифов.
Одноклеточные грибы систематизированы в 4 основных классах:
- дейтеромицеты;
- хитридиомицеты;
- зигомицеты;
- аскомицеты.
Ярким примером аскомицетов служат дрожжи, широко распространенные в природе. Скорость их роста и размножения велика благодаря особенному строению. Дрожжи состоят из одиночной клетки округлой формы, размножающейся почкованием.
Одноклеточные растения
Типичным представителем низших одноклеточных растений, часто встречающихся в природе, являются водоросли:
- хламидомонада;
- хлорелла;
- спирогира;
- хлорококк;
- вольвокс.
Хламидомонада отличается от всех водорослей подвижностью и наличием светочувствительного глазка, определяющего места наибольшего скопления солнечной энергии для фотосинтеза .
Многочисленные хлоропласты заменены одним большим хроматофором. Роль насосов, откачивающих излишки жидкости, выполняют сократительные вакуоли. Передвижение осуществляется при помощи двух жгутиков.
Зеленые водоросли хлореллы, в отличие от хламидомонады, обладают типичными растительными клетками. Плотная оболочка защищает мембрану, а в цитоплазме расположено ядро и хроматофор. Функции хроматофора сходны с ролью хлоропласт наземных растений.
С хлореллой схожа водоросль шарообразной формы хлорококк. Местом ее обитания служит не только вода, но и суша, стволы деревьев, растущих во влажной среде.
Кто открыл одноклеточные организмы
Честь открытия микроорганизмов принадлежит голландскому ученому А. Левенгуку.
В 1675 году он разглядел их в микроскоп собственного изготовления. За мельчайшими существами закрепилось название инфузория, а с 1820 года их стали называть простейшими животными.
Зоологами Келлекером и Зибольдом в 1845 году одноклеточные были отнесены к особому типу животного царства и разделены на две группы:
- корненожки;
- инфузории.

Как выглядит клетка одноклеточного животного
Строение одноклеточных организмов возможно изучить лишь с помощью микроскопа. Тело простейших существ состоит из единственной клетки, выполняющей роль независимого организма.
В состав клетки входят:
- цитоплазма;
- органоиды;
- ядро.
Со временем, в результате приспособления к окружающей среде, у отдельных видов одноклеточных появились специальные органоиды движения, выделения и питания.
Кто такие простейшие
Современная биология относит простейших к парафилетической группе животноподобных протистов. Наличие в клетке ядра, в отличие от бактерий, включает их в список эукариотов.
Клеточные структуры разнятся с клетками многоклеточных. В живой системе простейших присутствуют пищеварительные и сократительные вакуоли, у некоторых наблюдаются схожие с ротовой полостью и анальным отверстием органеллы.
Классы простейших
В современной классификации по признакам отсутствует отдельный ранг и значение одноклеточных.
Лабиринтула
Их принято подразделять на следующие типы:
- саркомастигофоры;
- апикомплексы;
- миксоспоридии;
- инфузории;
- лабиринтулы;
- асцестоспородии.
Устаревшей классификацией считается деление простейших на жгутиковых, саркодовых, ресничных и споровиков.
В каких средах обитают одноклеточные
Средой обитания простейших одноклеточных служит любая влажная среда. Амеба обыкновенная, эвглена зеленая и инфузория-туфелька являются типичными обитателями загрязненных пресных водных источников.
Наука долгое время относила опалин к инфузориям, благодаря внешнему сходству жгутиков с ресничками и наличию двух ядер. В результате тщательных исследований родство было опровергнуто. Половое размножение опалин происходит в результате копуляции, ядра одинаковые, а ресничный аппарат отсутствует.
Заключение
Биологическую систему невозможно представить без одноклеточных организмов, являющихся источником питания других животных.
Простейшие организмы способствуют образованию горных пород, служат показателями загрязненности водоемов, участвуют в круговороте углерода . Широкое применение микроорганизмы нашли в биотехнологиях.
Эвглена | Определение, схема, движение, классификация и факты
- Развлечения и поп-культура
- География и путешествия
- Здоровье и медицина
- Образ жизни и социальные вопросы
- Литература
- Философия и религия
- Политика, право и правительство
- Наука
- Спорт и отдых
- Технология
- Изобразительное искусство
- Всемирная история
- Этот день в истории
- Викторины
- Подкасты
- Словарь
- Биографии
- Резюме
- Популярные вопросы
- Обзор недели
- Инфографика
- Демистификация
- Списки
- #WTFact
- Товарищи
- Галереи изображений
- Прожектор
- Форум
- Один хороший факт
- Развлечения и поп-культура
- География и путешествия
- Здоровье и медицина
- Образ жизни и социальные вопросы
- Литература
- Философия и религия
- Политика, право и правительство
- Наука
- Спорт и отдых
- Технология
- Изобразительное искусство
- Всемирная история
- Britannica объясняет
В этих видеороликах Britannica объясняет различные темы и отвечает на часто задаваемые вопросы. - Britannica Classics
Посмотрите эти ретро-видео из архивов Encyclopedia Britannica. - Demystified Videos
В Demystified у Britannica есть все ответы на ваши животрепещущие вопросы. - #WTFact Видео
В #WTFact Britannica делится некоторыми из самых странных фактов, которые мы можем найти. - На этот раз в истории
В этих видеороликах узнайте, что произошло в этом месяце (или любом другом месяце!) в истории.
- Студенческий портал
Britannica — это главный ресурс для учащихся по ключевым школьным предметам, таким как история, государственное управление, литература и т. д. - Портал COVID-19
Хотя этот глобальный кризис в области здравоохранения продолжает развиваться, может быть полезно обратиться к прошлым пандемиям, чтобы лучше понять, как реагировать сегодня. - 100 женщин
Britannica празднует столетие Девятнадцатой поправки, выделяя суфражисток и политиков, творящих историю. - Спасение Земли
Британника представляет список дел Земли на 21 век. Узнайте об основных экологических проблемах, стоящих перед нашей планетой, и о том, что с ними можно сделать! - SpaceNext50
Britannica представляет SpaceNext50. От полёта на Луну до управления космосом — мы исследуем широкий спектр тем, которые подпитывают наше любопытство к космосу!
Содержание
- Введение
Краткие факты
- Связанный контент
Читать Далее
- Познакомьтесь с микробами: 5 крошечных простейших и водорослей
одноклеточные водоросли – неживая жизнь
Эвглена — это род одноклеточных пресноводных организмов, очень часто встречающихся в прудах и небольших водоемах, особенно если они богаты питательными веществами и, следовательно, содержат большое количество водорослей (также известных как «прудовая пена»). Как отмечено ниже, Euglena сама по себе иногда фотосинтезирует и является компонентом зеленого ила в таких прудах. Но в других случаях он не является фотосинтезирующим и является компонентом разнообразной группы организмов, которые поедают зеленый ил или, возможно, едят другие существа, которые едят зеленый ил.
Euglena относятся к небольшой группе (менее 1000 видов), которая в прошлом была востребована как зоологами (поскольку они подвижны, а некоторые из них гетеротрофны), так и ботаниками (поскольку некоторые представители фотосинтез). Соответственно, зоологи иногда называли эту группу «Euglenozoa» («зоа» относится к животным), а ботаники называли ее «Euglenophyta» («фита» относится к растениям). В прошлом группа была помещена в Королевство протистов. Недавние филогенетические исследования показали, что они очень рано отделились от других эукариот и, следовательно, поместили их в очень небольшую группу, состоящую из очень незнакомых одноклеточных организмов. Некоторые близкие родственники Euglena включает возбудитель сонной болезни и болезни Шагаса. Их таксономию усложняет тот факт, что некоторые в группе явно являются составными организмами, являясь продуктом вторичного эндосимбиоза, когда зеленая водоросль потреблялась, но не переваривалась жгутиконосцем.
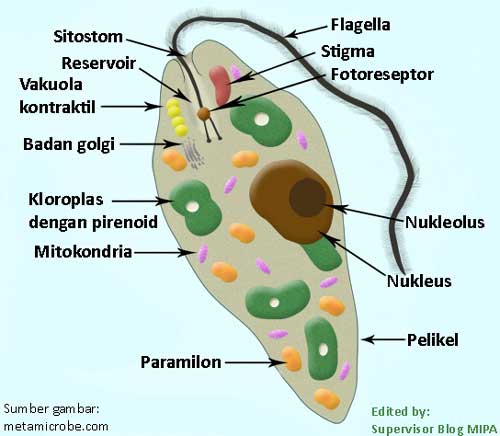
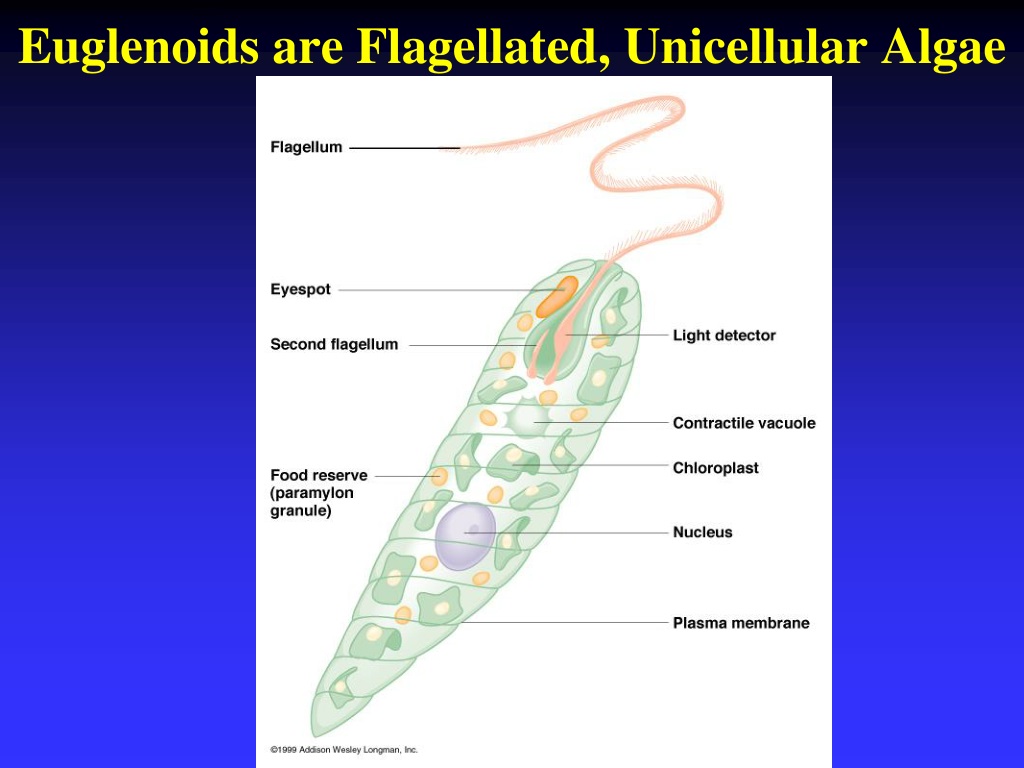
Эвглена — это одноклеточный организм со сложной внутренней структурой, который включает сократительную вакуоль, способную выталкивать воду, и красное «глазное пятно». Фотосинтезирующие формы содержат хлоропласт. У них есть два жгутика, один длинный, другой короткий, которые позволяют организмам двигаться. Euglena также может двигаться, изменяя свою форму (см. ссылки на видео). Снаружи клеточной мембраны находится гибкая белковая структура, называемая пелликулой. Хотя обычно она не считается клеточной стенкой, она выполняет аналогичные функции, обеспечивая некоторую жесткость и прочность, которые не может обеспечить мембрана. Однако пленка намного более гибкая, чем большинство клеточных стенок, и допускает изменение формы, которое часто наблюдается при движении Euglena .
Эвглена размножается бесполым путем, когда клетки делятся. Полового размножения внутри группы не обнаружено.
Исчерченность, видимая на правой стороне, представляет собой пелликулу, белковое покрытие, обнаруженное у эвгленоидов. Материя и энергия Иногда Euglena являются типичными фотоавтотрофами, использующими энергию солнечного света для синтеза углеводов из углекислого газа, а затем использующими углеводы в качестве источника энергии в клеточном дыхании и в качестве строительных материалов для синтеза разнообразных биомолекул. Эвгленоиды хранят углеводы в другом полимере глюкозы, чем обычный крахмал — звенья глюкозы объединены в 1,3-связь, а не 1,4-связь, как в обычном крахмале. Эвгленоиды также могут вести себя как гетеротрофы и приобретать материал путем проглатывания (фагоцитоз) или путем поглощения растворенных веществ из водной среды. Некоторые формы Euglena лишены хлоропластов и исключительно гетеротрофны.
Эвглена может быть важным компонентом определенных водных сред и играть роль как первичного продуцента, поедаемого другими организмами, так и разлагателя (гетеротрофа), потребляющего другие организмы и разрушающего их, или потребляет мертвый органический материал и разрушает его.
Некоторые виды Euglena (например, Euglena sanguinea) может окрашивать пруд в красный цвет, а также выделять токсины, убивающие рыбу.
- «Введение в эвглениды (эвгленоиды), некоторые из которых проявляют свойства как растений, так и животных», доктор Роберт Бердан.
- https://www.canadiannaturephotographer.com/euglenoids.html
- Hall Group в Институте Эрлхэма. Эвглена изящная.
- https://www.earlham.ac.uk/research-group/hall-group
Leave A Comment