
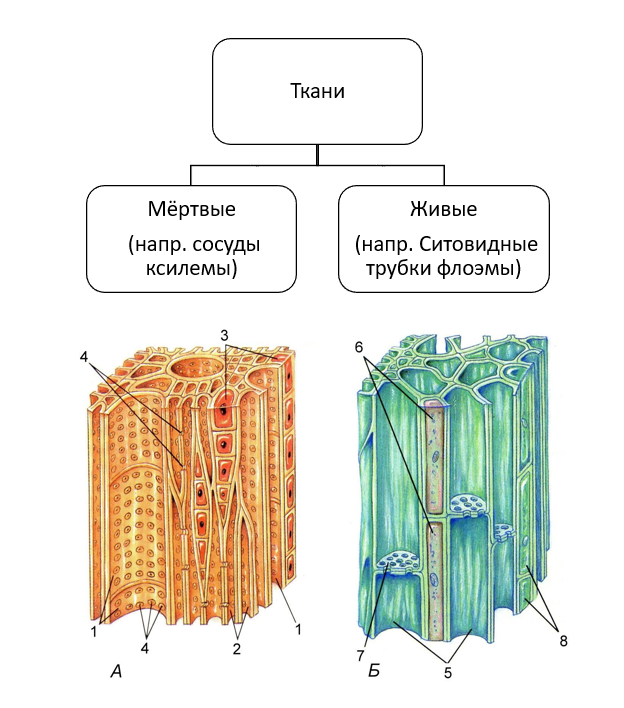
Проводящие ткани. Флоэма
Высшее растение представляет собой сложный организм с четкой дифференциацией тканей и специализацией органов, выполняющих различные жизненно важные функции.
При этом специализированные органы часто удалены друг от друга на значительное. расстояние. Например, фотосинтез происходит главным образом в листьях, поглощение воды и минеральных веществ — в корнях, отложение запасных питательных веществ — в особых запасающих тканях.
Основным условием нормальной жизнедеятельности растения является существование специального аппарата передвижения продуктов метаболизма от одного органа к другому. Передача веществ на большие расстояния осуществляется в растении достаточно экономично и с большой скоростью по специализированным проводящим тканям — флоэме и ксилеме.
Флоэма — ткань, главная функция которой состоит в проведении пластических веществ (нисходящий ток).
Ксилема — ткань, проводящая воду и растворенные в ней вещества (восходящий ток). Обычно обе проводящие ткани объединяются во флоэмно-ксилемные пучки, совокупность которых составляет проводящую систему растения.
Обычно обе проводящие ткани объединяются во флоэмно-ксилемные пучки, совокупность которых составляет проводящую систему растения.
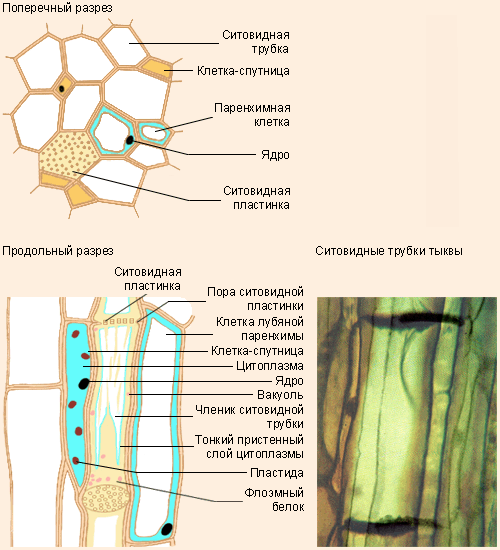
Флоэма — сложная ткань, включающая различные по структуре и функциональному значению анатомические элементы. Основным элементом флоэмы являются ситовидные трубки.
Каждая ситовидная трубка состоит из ряда отдельных клеток, соединенных между собой поперечными стенками. Такие трубки обычно тянутся вдоль продольной оси органа, но есть и поперечно идущие ситовидные трубки, входящие в состав анастомозов, тянущихся от одного продольно расположенного сосудисто-волокнистого пучка к другому. Оболочки ситовидных трубок целлюлозные. Лишь к концу вегетации растения некоторые ситовидные трубки одревесневают. В полостях ситовидных трубок весьма долго сохраняется живой протопласт в виде пристенного слоя. Ядро в зрелых ситовидных трубках отсутствует.
Протопласты ситовидных трубок содержат ряд включений. В некоторых ситовидных трубках находили пластиды и митохондрии.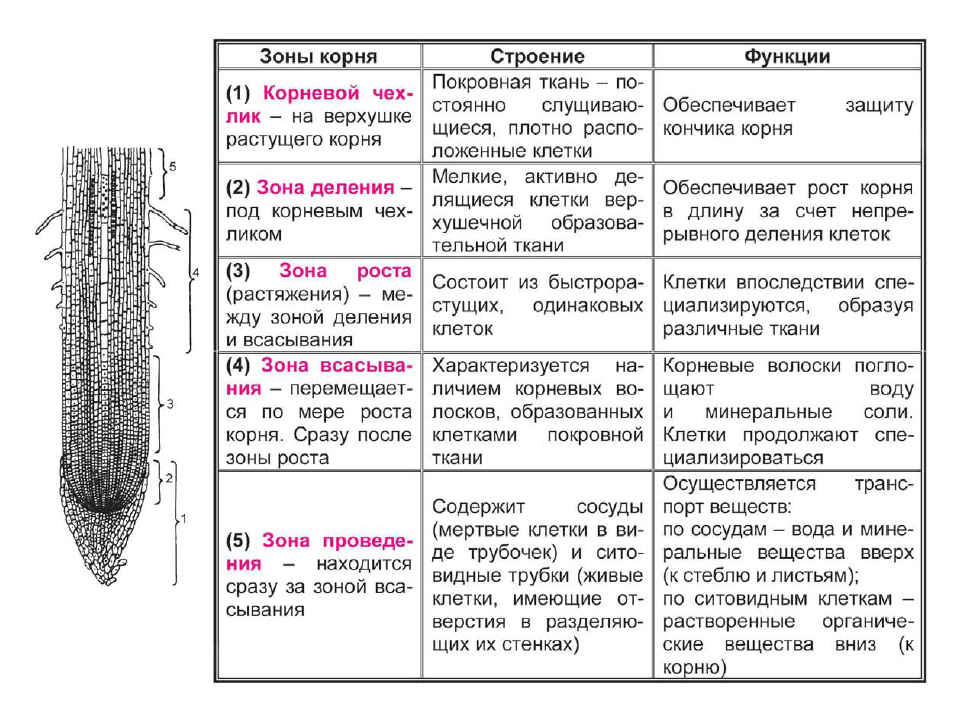 Ситовидные трубки предназначены преимущественно для проведения пластических веществ. Особенно важна их роль в проведения азотсодержащих веществ, служащих для построения белков.
Ситовидные трубки предназначены преимущественно для проведения пластических веществ. Особенно важна их роль в проведения азотсодержащих веществ, служащих для построения белков.
Клетки-членики ситовидных трубок живут сравнительно недолго. Как показали электронно-микроскопические исследования, в их протопласте в процессе дифференциации наблюдаются постепенные структурные изменения. В прокамбиальной или камбиальной (меристематической) стадии протопласт молодого ситовидного элемента обладает тонкой структурой, типичной для нормальной клетки. Однако уже на довольно ранней стадии дифференциации в нем происходит заметное разрыхление (разжижение) цитоплазмы. Затем ядро и тонопласт разрушаются, а вакуоля наполняется тонкофибриллярными структурами. Несмотря на отсутствие тонопласта, отделяющего цитоплазму от клеточного сока, митохондрии и пластиды остаются в постенном слое и обычно сохраняются во взрослых ситовидных трубках. Эндоплазматическая сеть и диктиосомы в дифференцированных ситовидных элементах покрытосеменных распадаются на многочисленные пузырьки и теряют свою структуру.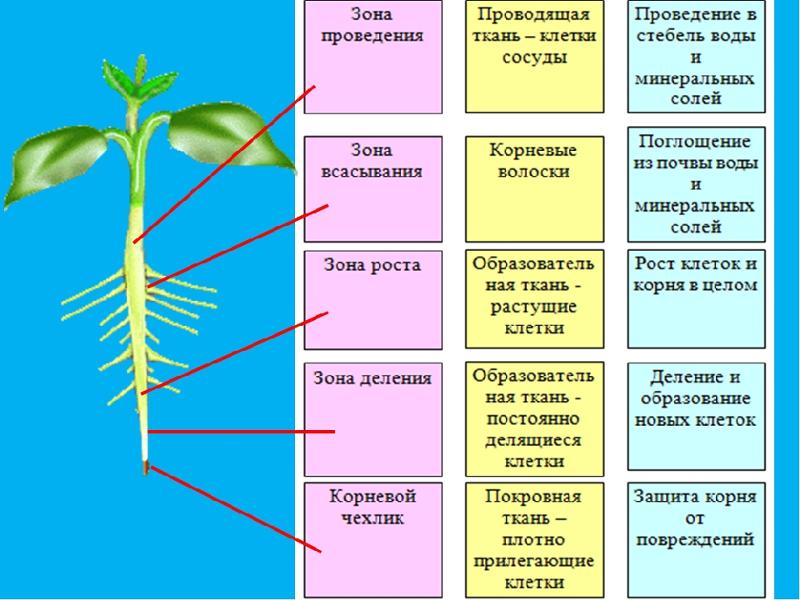 У голосеменных эндоплазматическая сеть может некоторое время сохраняться в полостях дифференцированных ситовидных клеток, но в конце концов также разрушается.
У голосеменных эндоплазматическая сеть может некоторое время сохраняться в полостях дифференцированных ситовидных клеток, но в конце концов также разрушается.
Наиболее своеобразной особенностью ситовидных трубок является строение их поперечных стенок, испещренных многочисленными мелкими перфорациями наподобие сита, откуда и сами клетки получили название ситовидных, а поперечные стенки с ситами — ситовидных пластинок. Перфорации обеспечивают непрерывность протопластов элементов ситовидных трубок. Эта непрерывность была показана с помощью электронного микроскопа. Осенью ситовидные пластинки в большинстве случаев затягиваются особым веществом, называемым каллозой. В некоторых ситовидных трубках каллоза закупоривает сита окончательно, а в большинстве трубок она к весне растворяется, открывая сообщение между отдельными члениками.
Ситовидные участки имеются и на продольных стенках. Строение и функция сит на продольных стенках такие же, как и на поперечных.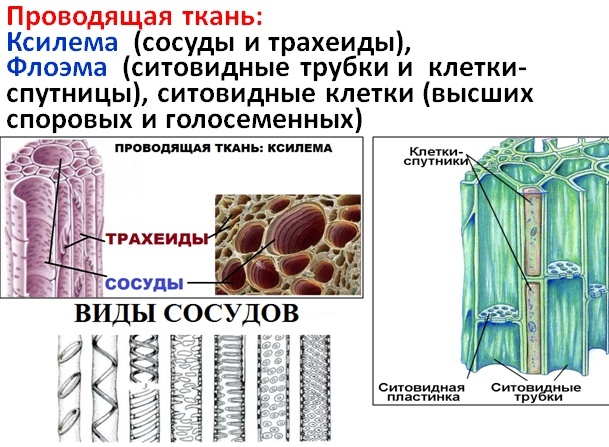 Так как продольные стенки оболочек ситовидных трубок имеют более обширную площадь, чем поперечные, то сита на продольных стенках не занимают всей их поверхности, а собраны в группы, называемые ситовидными полями.
Так как продольные стенки оболочек ситовидных трубок имеют более обширную площадь, чем поперечные, то сита на продольных стенках не занимают всей их поверхности, а собраны в группы, называемые ситовидными полями.
Ситовидные трубки функционально связаны с другими специализированными элементами флоэмы — клетками-спутниками. Ситовидная трубка происходит из той же инициальной клетки, что и сопровождающая ее клетка-спутник.
Инициальная клетка делится продольной перегородкой на две клетки неодинакового диаметра. Более крупная из дочерних клеток дифференцируется как ситовидная трубка, а более мелкая несколько раз делится в поперечном направлении и образует цепочку клеток-спутников. В этих клетках полностью сохраняется живой протопласт с ядрами. Оболочки этих клеток, примыкающие к ситовидным трубкам, тонкие, целлюлозные и имеют простые поры. Связь ситовидных трубок со спутниками настолько прочная, что они не отделяются друг от друга даже при мацерации.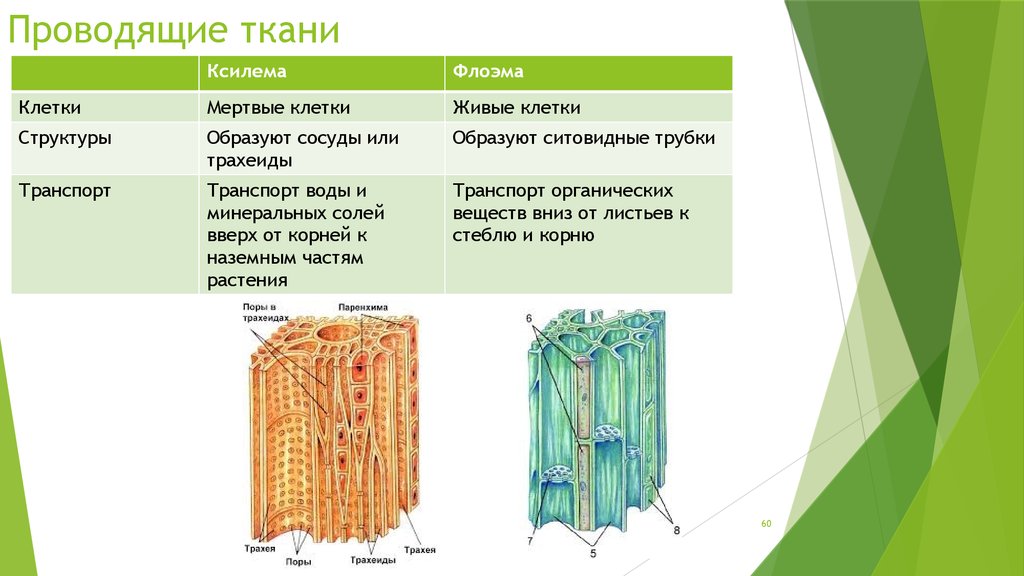
Присутствие в клетках-спутниках ядер и цитоплазмы, а также тесная связь этих клеток с ситовидными трубками, в значительной степени утративших эти атрибуты самостоятельной живой системы, указывают на активную роль спутников в метаболизме флоэмы. Предполагают, что в спутниках с особой интенсивностью вырабатываются различные ферменты, которые передаются в ситовидные трубки.
Ситовидные трубки и спутники соприкасаются не только между собой, но и с клетками лубяной паренхимы. Связь с этими клетками также обеспечивается посредством простых пор. Простые поры, соединяющие продольные стенки ситовидных трубок с паренхимой, собраны группами и со стороны ситовидных трубок вполне напоминают ситовидные пластинки. Клетки паренхимы, соприкасающиеся с ситовидными трубками, более или менее удлинены. Они располагаются среди ситовидных элементов без какого-либо определенного порядка. Эта паренхима называется лубяной. Оболочки таких клеток целлюлозные, тонкие, протопласт содержит ряд пластических веществ, периодически накапливающихся или переходящих в растворенное состояние, как во всякой живой и вполне жизнедеятельной клетке.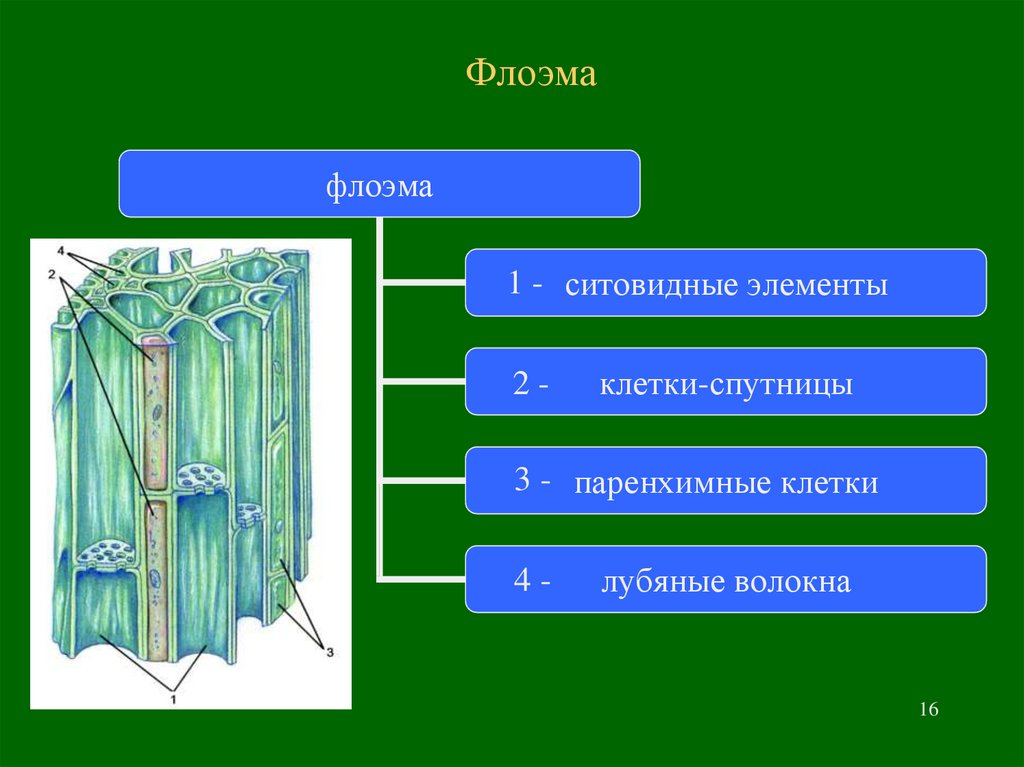
У некоторых растений группы ситовидных трубок с клетками-спутниками и лубяной паренхимой перемежаются с группами лубяных волокон. Такая структура особенно характерна для древесных растений (виноградная лоза, липа и др.). Весь комплекс анатомических элементов, состоящий из ситовидных трубок и примыкающих к ним клеток, называется мягким лубом, а пучки лубяных волокон — твердым лубом. Лубяные волокна, как уже говорилось, часто одревесневают и притом весьма рано, элементы же мягкого луба или совсем не одревесневают, или же одревесневают лишь старые элементы (у растения, кончающего свою вегетацию).
Ситовидные трубки не у всех растений хорошо развиты. Особенно широкими ситовидными трубками с ясно выраженной перфорацией отличаются лианы и вообще растения с вьющимися и цепляющимися побегами (тыква, виноградная лоза, глициния) и водные растения (водяной орех, водяная лилия и др.). У многих растений ситовидные трубки очень узкие, перфорации выражены слабо (картофель, лен и др. ).
).
Продолжительность существования ситовидных трубок у различных растений различна и колеблется от одного вегетационного периода до нескольких лет. В общем же ситовидные трубки, лишенные ядер, недолговечны. Срок существования каждой клетки (членика) ситовидной трубки тесно связан с сохранностью ее живого содержимого — протопласта. С разрушением протопласта оболочка каждой клетки ситовидной трубки может одревесневать и сохраняться или же сдавливаться соседними живыми паренхимными клетками. В последнем случае происходит облитерация ситовидной трубки, и она становится трудно различимой.
В редких случаях паренхимные клетки образуют сосочковидные выросты в полость ситовидной трубки. Эти выросты, называемые тиллами, закупоривают ситовидную трубку. Образование тилл в ситовидных трубках можно наблюдать у виноградной лозы в месте срастания привоя и подвоя, причем тиллы в данных случаях имеют неодревесневшие оболочки. Хорошо и часто тиллы развиваются в сосудах.
В общих чертах строение ситовидных трубок у всех растений одинаково, но в деталях имеются различия.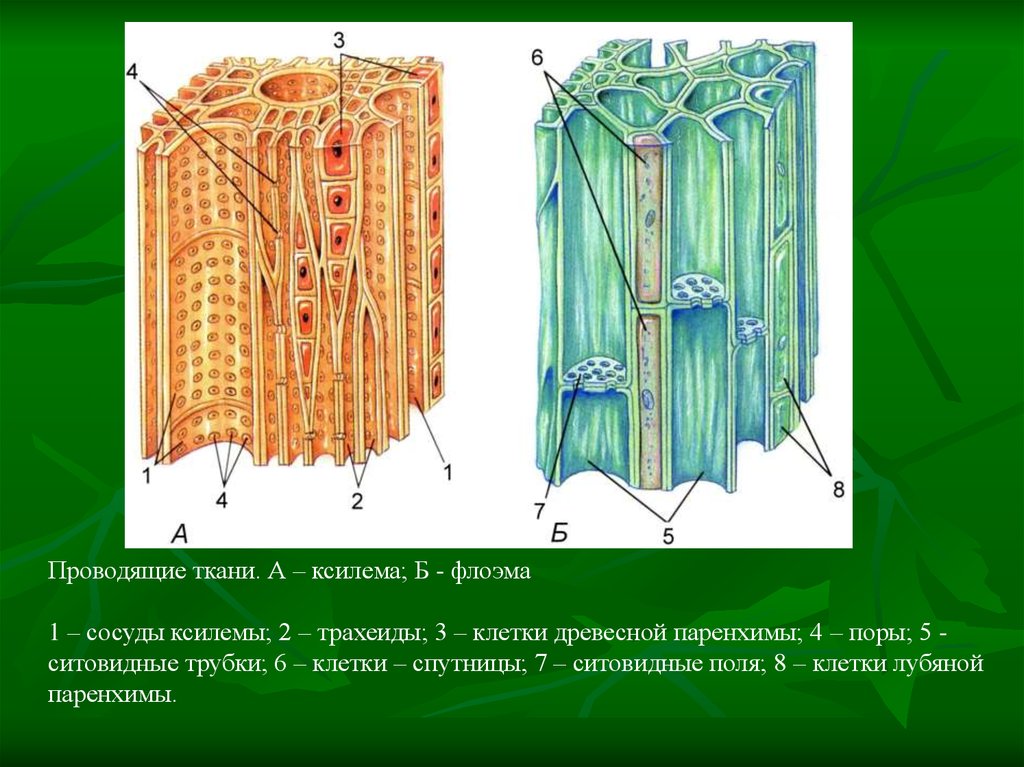 Прежде всего, у разных растений различен просвет ситовидных трубок, размеры перфораций и ситовидных полей, составленных из них, очертания ситовидных полей как на поперечных, так и на продольных стенках, и само распределение полей, неодинаковы также толщина оболочек, степень развития каллозы. У голосеменных и папоротникообразных флоэмные элементы имеют ситовидные пластинки только на продольных стенках. Они называются ситовидными клетками.
Прежде всего, у разных растений различен просвет ситовидных трубок, размеры перфораций и ситовидных полей, составленных из них, очертания ситовидных полей как на поперечных, так и на продольных стенках, и само распределение полей, неодинаковы также толщина оболочек, степень развития каллозы. У голосеменных и папоротникообразных флоэмные элементы имеют ситовидные пластинки только на продольных стенках. Они называются ситовидными клетками.
Даже в одном и том же растении, например, в стеблях виноградной лозы, не все ситовидные трубки построены одинаково. Часть из них не имеет клеток-спутников. Ситовидные трубки, возникшие в начале формирования побега, т. е. первичного происхождения, имеют ситовидные участки только на поперечных стенках, а у ситовидных трубок, возникших позднее (вторичного происхождения), они возникают и на продольных стенках. Тиллы образуются лишь в полостях ситовидных трубок вторичного происхождения. Ситовидные трубки первичного происхождения относительно скоро облитерируются и в дальнейшем, если участок коры, содержащий эти трубки, сохраняется на растении живым, окончательно исчезают, растворяясь соответствующими ферментами.
Транспорт сахара в растениях: флоэма
Цели обучения
- Различать источники и поглотители сахара в тканях растений
- Объясните модель потока под давлением для транслокации сахара в ткани флоэмы
- Описать роль протонных насосов, котранспортеров и облегченной диффузии в модели потока под давлением
- Знать, как разные концентрации сахара в источниках и разные типы поглотителей влияют на транспортный путь, используемый для загрузки или выгрузки сахаров
- Сравните и сопоставьте механизмы транспорта жидкости в ксилеме и флоэме
Приведенная ниже информация была адаптирована из OpenStax Biology 30.5
Для роста растениям нужен источник энергии. У растущих растений фотосинтаты (сахара, образующиеся в результате фотосинтеза) вырабатываются в листьях в результате фотосинтеза, а затем транспортируются в места активного роста, где сахара необходимы для поддержки роста новых тканей. В течение вегетационного периода зрелые листья и стебли производят излишки сахаров , которые переносятся в места хранения, включая наземную ткань в корнях или луковицах (тип модифицированного стебля).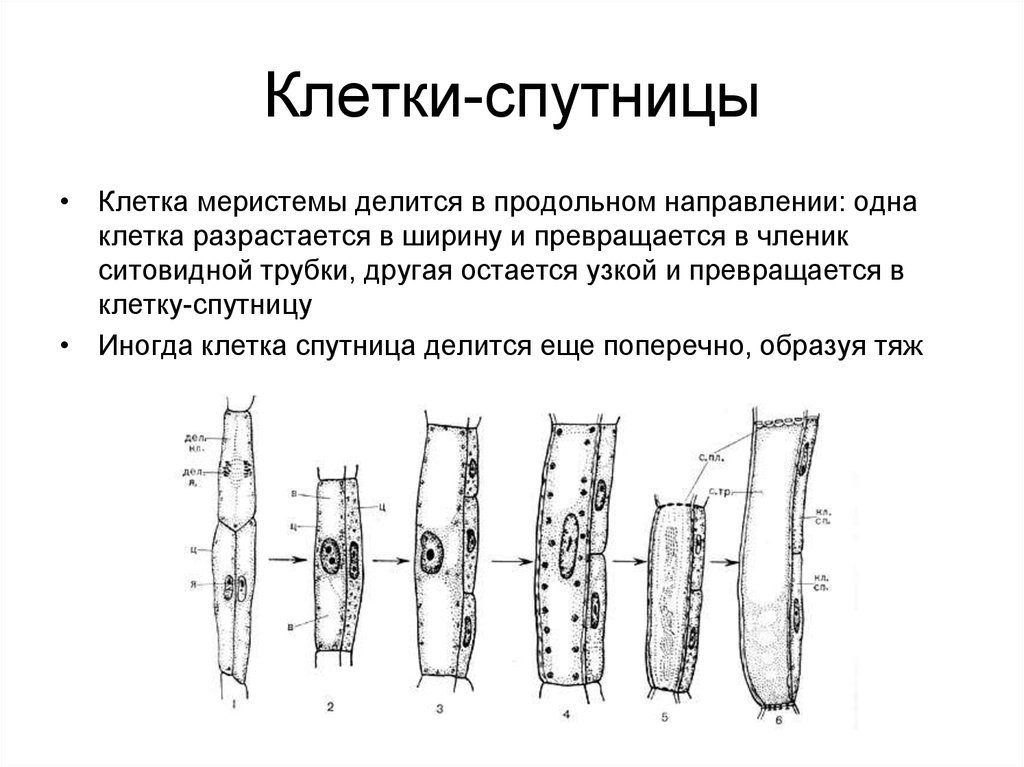
Места, которые производят или выделяют сахар для выращивания растений, называются источниками . Сахара, образующиеся в таких источниках, как листья, необходимо доставлять к растущим частям растения через флоэму в процессе, называемом транслокацией или перемещением сахара. Места поступления сахара, такие как корни, молодые побеги и развивающиеся семена, называются
Фотосинтеты из источника обычно перемещаются в ближайший сток через элементы ситовидных трубок флоэмы. Например, самые высокие листья направляют сахар вверх к верхушке растущего побега, тогда как нижние листья направляют сахар вниз к корням. Промежуточные листья будут посылать продукты в обоих направлениях, в отличие от потока в ксилеме, который всегда однонаправлен (почва-лист-атмосфера). Обратите внимание, что жидкость в одном элементе ситовой трубки может течь только в одном направлении за раз, но жидкость в соседних элементах ситовой трубки может двигаться в разных направлениях. Направление потока также меняется по мере роста и развития растения:
Например, самые высокие листья направляют сахар вверх к верхушке растущего побега, тогда как нижние листья направляют сахар вниз к корням. Промежуточные листья будут посылать продукты в обоих направлениях, в отличие от потока в ксилеме, который всегда однонаправлен (почва-лист-атмосфера). Обратите внимание, что жидкость в одном элементе ситовой трубки может течь только в одном направлении за раз, но жидкость в соседних элементах ситовой трубки может двигаться в разных направлениях. Направление потока также меняется по мере роста и развития растения:
- В середине вегетационного периода активно фотосинтезирующие зрелые листья и стебли служат источниками, производящими излишки сахаров, которые транспортируются в стоки, где потребление сахара велико. Поглотители в течение вегетационного периода включают области активного роста меристем, новых листьев и репродуктивных структур. Раковины также включают места хранения сахара, такие как корни, клубни или луковицы. В конце вегетационного периода растение сбрасывает листья и больше не имеет активно фотосинтезирующих тканей.

- Рано в начале следующего вегетационного периода растение должно возобновить рост после периода покоя (зимнего или засушливого сезона). Поскольку у растения нет существующих листьев, его единственным источником сахара для роста является сахар, хранящийся в корнях, клубнях или луковицах с последнего вегетационного периода. Эти хранилища теперь служат источниками, а активно развивающиеся листья — стоками. Как только листья созреют, они станут источником сахара в течение вегетационного периода.

Обзор транслокации: транспортировка от источника к приемнику
Сахара перемещаются (перемещаются) из источника в сток, но как? Наиболее общепринятой гипотезой для объяснения движения сахаров во флоэме является модель напорного потока для транспорта флоэмы. Эта гипотеза объясняет несколько наблюдений:
- Флоэма находится под давлением
- Транслокация прекращается, если ткань флоэмы погибает
- Транслокация ингибируется соединениями, которые останавливают выработку АТФ в источнике сахара
В общих чертах модель напорного потока работает следующим образом: высокая концентрация сахара в источнике создает низкий потенциал растворенного вещества (Ψs), который втягивает воду во флоэму из соседней ксилемы.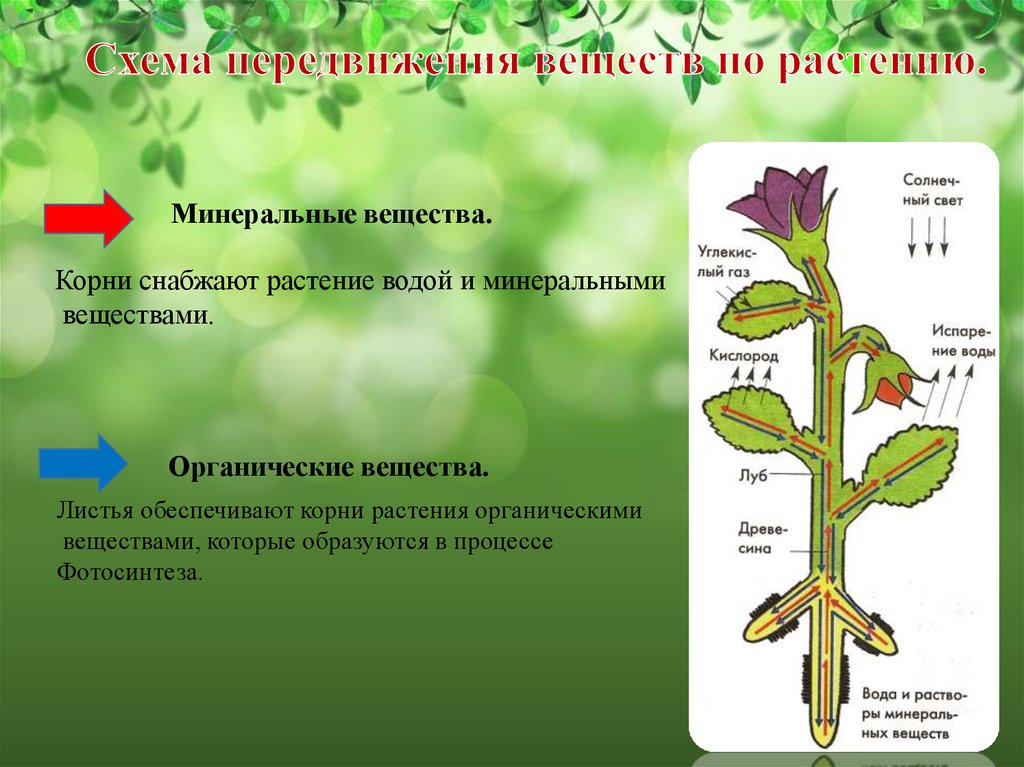 Это создает потенциал высокого давления (Ψp) или высокое тургорное давление во флоэме. Высокое тургорное давление вызывает движение сока флоэмы «объемным потоком» от источника к стоку, где сахара быстро удаляются из флоэмы в стоке. Удаление сахара увеличивает Ψs, что приводит к тому, что вода покидает флоэму и возвращается в ксилему, уменьшая Ψp.
Это создает потенциал высокого давления (Ψp) или высокое тургорное давление во флоэме. Высокое тургорное давление вызывает движение сока флоэмы «объемным потоком» от источника к стоку, где сахара быстро удаляются из флоэмы в стоке. Удаление сахара увеличивает Ψs, что приводит к тому, что вода покидает флоэму и возвращается в ксилему, уменьшая Ψp.
В этом видео представлен краткий обзор источников сахара, поглотителей и гипотезы напорного потока:
Транспортные пути при транслокации сахара
Прежде чем мы углубимся в детали того, как работает модель напорного потока, давайте сначала вернемся к некоторым транспортным путям, которые мы обсуждали ранее:
- Диффузия происходит, когда молекулы перемещаются из области с высокой концентрацией в область с низкой концентрацией. Диффузия не требует энергии, потому что молекулы движутся по градиенту их концентрации (из областей с высокой концентрацией в области с низкой концентрацией).
- Протонные насосы используют энергию АТФ для создания электрохимических градиентов с высокой концентрацией протонов на одной стороне плазматической мембраны. Затем этот электрохимический градиент можно использовать в качестве источника энергии для перемещения других молекул против градиента их концентрации с помощью котранспортеров.
- Котранспортеры — это каналы, выполняющие тип вторичного активного (энергоемкого) транспорта. Котранспортеры перемещают две молекулы одновременно: одна молекула транспортируется по градиенту ее концентрации («вниз»), что высвобождает энергию, которая используется для транспортировки другой молекулы против градиента ее концентрации.
- Симпортеры — это тип котранспортера, который переносит две молекулы в одном направлении; оба в клетку, или оба из клетки.
- Антипортеры — тип котранспортера, который переносит две молекулы в противоположных направлениях; один в клетку, а другой из клетки.
- Симпортеры — это тип котранспортера, который переносит две молекулы в одном направлении; оба в клетку, или оба из клетки.
 Диффузия не требует энергии, потому что молекулы движутся по градиенту их концентрации (из областей с высокой концентрацией в области с низкой концентрацией).
Диффузия не требует энергии, потому что молекулы движутся по градиенту их концентрации (из областей с высокой концентрацией в области с низкой концентрацией).
Симпортеры перемещают две молекулы в одном направлении; Антипортеры перемещают две молекулы в противоположных направлениях. Изображение предоставлено: Академия Хана, https://www.khanacademy.org/science/biology/membranes-and-transport/active-transport/a/active-transportИзображение изменено из биологии OpenStax. Исходное изображение предоставлено Lupask/Wikimedia Commons.
Каждый из этих транспортных путей играет роль в модели напорного потока для транспорта по флоэме.
Детали модели напорного потока для транспорта флоэмы
Фотосинтаты, такие как сахароза, вырабатываются в клетках мезофилла (тип паренхимных клеток) фотосинтезирующих листьев. Сахара активно транспортируются из клеток-источников в клетки-компаньоны ситовидной трубки, которые связаны с элементами ситовидной трубки в сосудистых пучках. Этот активный транспорт сахара в клетки-компаньоны происходит через симпортер протон-сахароза ; клетки-компаньоны используют протонный насос, работающий от АТФ, для создания электрохимического градиента вне клетки. Котранспорт протона с сахарозой позволяет перемещать сахарозу против градиента ее концентрации в клетки-компаньоны. имеет место.
Этот активный транспорт сахара в клетки-компаньоны происходит через симпортер протон-сахароза ; клетки-компаньоны используют протонный насос, работающий от АТФ, для создания электрохимического градиента вне клетки. Котранспорт протона с сахарозой позволяет перемещать сахарозу против градиента ее концентрации в клетки-компаньоны. имеет место.
Из клеток-компаньонов сахар диффундирует в элементы ситовидной трубки флоэмы через плазмодесмы, которые связывают клетку-компаньон с элементами ситовидной трубки. Элементы ситовидных трубок флоэмы имеют уменьшенное цитоплазматическое содержимое и соединены ситовидной пластинкой с порами, которые обеспечивают объемный поток или перемещение флоэмного сока под давлением.
Флоэма состоит из клеток, называемых элементами ситовидных трубок. Сок флоэмы проходит через отверстия, называемые пластинками ситовидных трубок. Соседние клетки-спутницы выполняют метаболические функции для элементов ситовидной трубки и обеспечивают их энергией.
Наличие высоких концентраций сахара в элементах ситовидных трубок резко снижает Ψs, что приводит к перемещению воды путем осмоса из ксилемы в клетки флоэмы. Это перемещение воды в клетки ситовидных трубок приводит к увеличению Ψp, увеличивая как тургорное давление во флоэме, так и общий водный потенциал во флоэме у источника. Это увеличение водного потенциала приводит к увеличению потока флоэмы от источника к стоку.
Разгрузка на стоковом конце флоэмной трубки может происходить либо путем диффузии , если концентрация сахарозы на стоке ниже, чем во флоэме, либо путем активного транспорта , если концентрация сахарозы выше на стоке чем во флоэме. Если сток представляет собой область активного роста, такую как новый лист или репродуктивная структура, то концентрация сахарозы в стоковых клетках обычно ниже, чем в элементах ситовидных трубок флоэмы, потому что стоковая сахароза быстро метаболизируется для роста.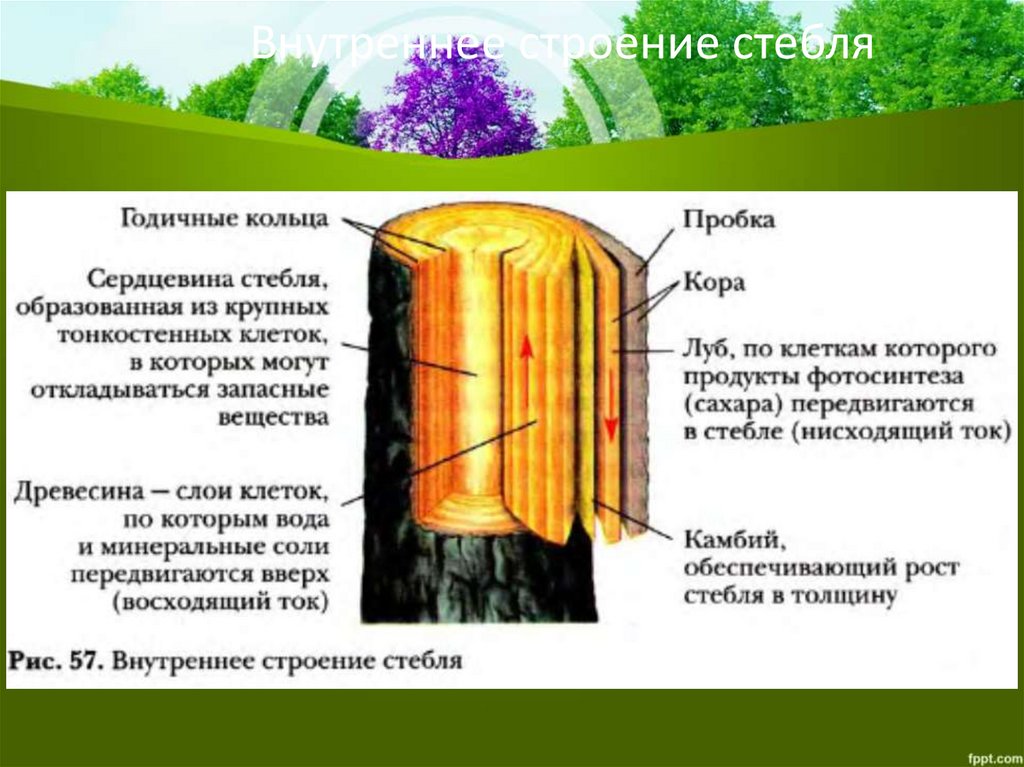 Если раковина представляет собой место хранения, где сахар превращается в крахмал, например, в корнях или луковицах, то концентрация сахара в раковине обычно ниже, чем в ситовидных элементах флоэмы, потому что сахароза раковины быстро превращается в крахмал для хранилище. Но если раковина представляет собой место хранения, где сахар хранится в виде сахарозы, например, сахарная свекла или сахарный тростник, то в раковине может быть более высокая концентрация сахара, чем в клетках ситовидных трубок флоэмы. В этой ситуации активный транспорт протонно-сахарозного антипортера используется для транспортировки сахара из клеток-спутников в запасающие вакуоли в запасных клетках.
Если раковина представляет собой место хранения, где сахар превращается в крахмал, например, в корнях или луковицах, то концентрация сахара в раковине обычно ниже, чем в ситовидных элементах флоэмы, потому что сахароза раковины быстро превращается в крахмал для хранилище. Но если раковина представляет собой место хранения, где сахар хранится в виде сахарозы, например, сахарная свекла или сахарный тростник, то в раковине может быть более высокая концентрация сахара, чем в клетках ситовидных трубок флоэмы. В этой ситуации активный транспорт протонно-сахарозного антипортера используется для транспортировки сахара из клеток-спутников в запасающие вакуоли в запасных клетках.
Как только сахар выгружается из стоковых клеток, Ψs увеличивается, вызывая диффузию воды за счет осмоса из флоэмы обратно в ксилему. Это движение воды из флоэмы приводит к уменьшению Ψp, снижению тургорного давления во флоэме в стоке и сохранению направления основного потока от источника к стоку.
Сахароза активно транспортируется из клеток-источников в клетки-спутницы, а затем в элементы ситовидных трубок.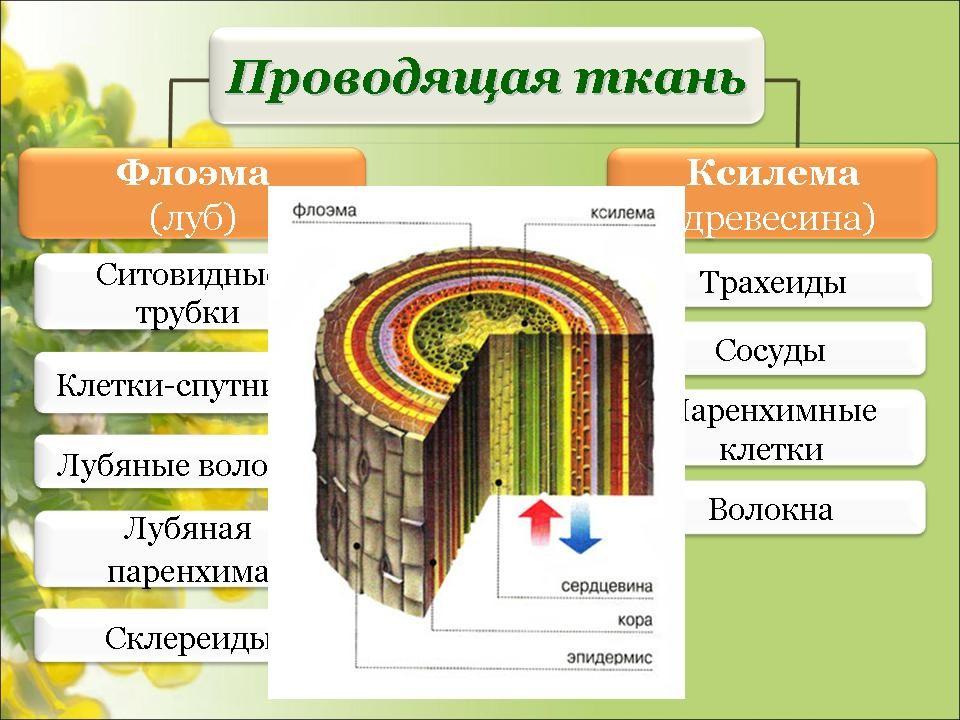 Это снижает водный потенциал, из-за чего вода поступает во флоэму из ксилемы. Возникающее в результате положительное давление выталкивает смесь сахарозы и воды вниз к корням, где сахароза выгружается. Благодаря транспирации вода возвращается к листьям по сосудам ксилемы. Изображение предоставлено: OpenStax Biology
Это снижает водный потенциал, из-за чего вода поступает во флоэму из ксилемы. Возникающее в результате положительное давление выталкивает смесь сахарозы и воды вниз к корням, где сахароза выгружается. Благодаря транспирации вода возвращается к листьям по сосудам ксилемы. Изображение предоставлено: OpenStax Biology
В этом видео (начало 5:03) более подробно обсуждается гипотеза напорного потока:
Движение жидкости в ксилеме по сравнению с флоэмой
Должно быть ясно, что движение сахаров во флоэме зависит от движения воды во флоэме. Но есть некоторые важные различия в механизмах движения жидкости в этих двух разных сосудистых тканях:
- Движущая сила движения жидкости:
- Ксилема: транспирация (испарение) из листьев в сочетании со сцеплением и натяжением воды в элементах сосудов и трахеидах (пассивная; не требует энергии)
- Флоэма: активный транспорт сахарозы из исходных клеток в элементы ситовидной трубки флоэмы (требуется энергия)
- Клетки, обеспечивающие движение жидкости:
- Ксилема: неживые элементы сосудов и трахеиды
- Флоэма: элементы живой ситовидной трубки (поддерживаемые клетками-спутницами)
- Ксилема: отрицательный результат из-за натяжения сверху (транспирация, напряжение)
- Флоэма: положительная из-за толчка от источника (Ψp увеличивается из-за притока воды, который увеличивает тургорное давление в источнике)
Флоэма: схема, структура, функции, адаптации
Флоэма представляет собой специализированную живую ткань, которая переносит аминокислоты и сахара из листьев (источник) в растущие части растения (приемник) в процессе, называемом транслокацией . Этот процесс является двунаправленным.
Этот процесс является двунаправленным.
Источник — это область растения, производящая органические соединения, такие как аминокислоты и сахара. Примерами источников являются зеленые листья и клубни.
Раковина — активно растущая область растения. Примеры включают корни и меристемы.
Структура флоэмы
Флоэма содержит четыре специализированных типа клеток, выполняющих свои функции. К ним относятся:
- Элементы ситовидной трубки — ситовидная трубка представляет собой непрерывный ряд клеток, играющих ключевую роль в поддержании клеток и транспортировке аминокислот и сахаров (ассимилятов). Они тесно взаимодействуют с клетками-спутницами.
- Клетки-компаньоны — клетки, ответственные за транспортировку ассимилятов в ситовидные трубки и из них.
- Волокна флоэмы представляют собой клетки склеренхимы, представляющие собой неживые клетки флоэмы, обеспечивающие структурную поддержку растения.
- Клетки паренхимы представляют собой постоянную основную ткань, которая образует основную массу растения.

Ассимиляты растений относятся к аминокислотам и сахарам (сахароза).
Рис. 1 — Показано строение флоэмы
Адаптации флоэмы
Клетки, составляющие флоэму, адаптированы к своим функциям: ситовидные трубки , которые специализируются на транспорте и лишены ядер, и клетки-компаньоны s, которые являются необходимыми компонентами в перемещении ассимилирует. Ситовидные трубки имеют перфорированные концы, поэтому их цитоплазма соединяет одну клетку с другой. Ситовидные трубки транспортируют сахара и аминокислоты в своей цитоплазме.
И ситовидные трубки, и клетки-компаньоны встречаются исключительно у покрытосеменных растений (растений, которые цветут и дают семена, заключенные в плодолистика).
Приспособления ячеек ситовидных трубок
- Ситовидные пластинки соединяют их (концевые пластинки ячеек) поперечно (проходя в поперечном направлении), позволяя ассимилятам течь между ячейками ситовидных элементов.
- Они не имеют ядра и имеют уменьшенное количество органелл, чтобы максимизировать пространство для ассимилятов.
- У них толстые и жесткие клеточные стенки, способные противостоять высокому гидростатическому давлению, создаваемому перемещением.

Адаптация клеток-компаньонов
- Их плазматическая мембрана загибается внутрь, чтобы увеличить площадь поверхности для поглощения материала (дополнительную информацию см. в нашей статье «Отношение площади поверхности к объему»).
- Они содержат много митохондрий для производства АТФ для активного транспорта ассимилятов между источниками и приемниками.
- Они содержат много рибосом для синтеза белка.
Таблица 1. Различия между ситовидными трубками и клетками-спутницами.
Ситчатые трубки | Сопутствующие ячейки |
Относительно большие ячейки | Относительно маленькие ячейки |
Содержит ядро | |
Поры в поперечных стенках | Поры отсутствуют |
Относительно низкая метаболическая активность | Относительно высокая метаболическая активность |
Рибосомы отсутствуют | Много рибосом |
Присутствуют лишь несколько митохондрий | Большое количество митохондрий |
Функция флоэмы
Ассимиляты, такие как амино кислоты и сахара (сахароза) транспортируются во флоэме путем транслокации от источников к стокам.
Ознакомьтесь с нашей статьей «Перенос масс в растениях», чтобы узнать больше о гипотезе массового расхода.
Загрузка флоэмы
Сахароза может поступать в элементы ситовидной трубки двумя путями:
- Путь апопластный
- Путь симпластический
Плазмодесмы представляют собой межклеточные каналы вдоль клеточной стенки растений, которые облегчают обмен сигнальными молекулами и сахарозой между клетками. Они действуют как цитоплазматические соединения и играют ключевую роль в клеточной коммуникации (за счет транспорта сигнальных молекул).
Цитоплазматические соединения относятся к межклеточным или межклеточным соединениям внеклеточного матрикса через цитоплазму.
Рис. 2. Движение веществ через пути апопласта и симпласта
Массовый поток
Массовый поток относится к движению веществ по градиенту температуры или давления. Транслокация описывается как массовый поток и происходит во флоэме. В этом процессе участвуют элементы ситовидной трубки и клетки-спутницы. Он перемещает вещества из мест их производства (источников) туда, где они необходимы (стоки). Примером источника являются листья, а стоком — любые растущие или запасающие органы, такие как корни и побеги.
Гипотеза массового расхода часто используется для объяснения перемещения веществ, хотя она не полностью принята из-за отсутствия доказательств. Здесь мы подытожим процессы.
Сахароза поступает в ситовидные трубки из клеток-спутниц посредством активного транспорта (требуется энергия). Это приводит к уменьшению водного потенциала в ситовых трубках, и вода поступает внутрь за счет осмоса. В свою очередь гидростатическое (водяное) давление увеличивается. Это вновь созданное гидростатическое давление вблизи источников и более низкое давление в поглотителях позволит веществам течь по градиенту. Растворенные вещества (растворенные органические вещества) перемещаются в раковины. Когда раковины удаляют растворенные вещества, водный потенциал увеличивается, и вода покидает флоэму путем осмоса. При этом гидростатическое давление поддерживается.
Это вновь созданное гидростатическое давление вблизи источников и более низкое давление в поглотителях позволит веществам течь по градиенту. Растворенные вещества (растворенные органические вещества) перемещаются в раковины. Когда раковины удаляют растворенные вещества, водный потенциал увеличивается, и вода покидает флоэму путем осмоса. При этом гидростатическое давление поддерживается.
В чем разница между ксилемой и флоэмой?
Флоэма состоит из живых клеток, поддерживаемых клетками-спутницами, тогда как сосуды ксилемы состоят из неживой ткани.
Ксилема и флоэма представляют собой транспортные структуры, которые вместе образуют сосудистый пучок . Ксилема несет воду и растворенные минералы, начиная с корней (приемник) и заканчивая листьями растений (источник). Движение воды обусловлено транспирацией в однонаправленном потоке.
Транспирация описывает потерю водяного пара через устьица.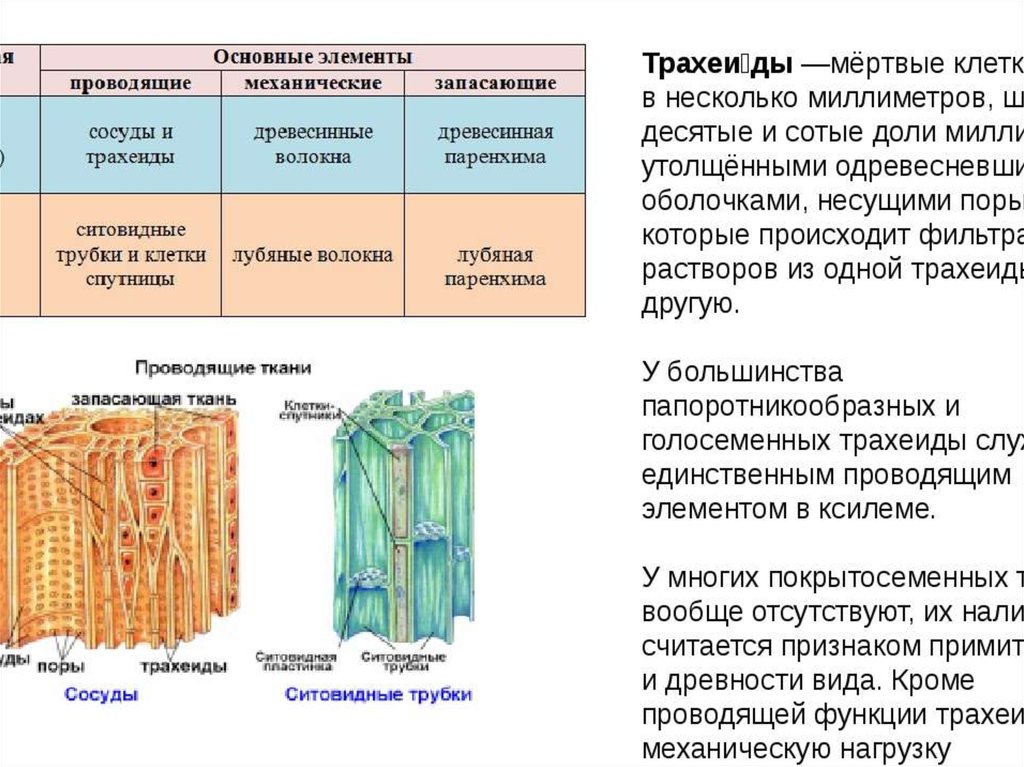
Флоэма транспортирует ассимилированные к запасающим органам путем транслокации. Примеры запасающих органов включают запасающие корни (модифицированный корень, например, морковь), луковицы (модифицированные основания листьев, например, лук) и клубни (подземные стебли, которые хранят сахара, например, картофель). Поток вещества внутри флоэмы двунаправленный.
Рис. 3 – Различия между тканями ксилемы и флоэмы
Таблица 2. Резюме сравнения ксилемы и флоэмы.
Ксилема | Флоэма |
В основном неживая ткань | В основном живая ткань |
Присутствует во внутренней части растения | Присутствует на наружная часть сосудистого пучка |
Движение материалов однонаправленное | Двунаправленное перемещение материалов |
Перевозит воду и минералы | Перевозит сахара и аминокислоты | Обеспечивает механическую структуру растения (содержит лигнин) | Содержит волокна, придающие прочность стеблю (но не по шкале лигнина в ксилеме) |
Нет торцевых стенок между клетками | Содержит ситовидные пластины |
Флоэма — Ключевые выносы
- Основная функция флоэмы заключается в транспортировке ассимилятов в стоки посредством транслокации.
Leave A Comment