
Оплодотворение: основные этапы процесса. Этапы развития эмбриона
Оплодотворение: основные этапы процесса. Этапы развития эмбриона | Клиника «Центр ЭКО» в КурскеСкачать базы доноров:
Доноры ооцитов (Excel)
Оплодотворение – это процесс слияния женской (яйцеклетка) и мужской (сперматозоид) половых клеток, в результате которого зарождается новая жизнь. С этого момента начинает развиваться ребенок в теле материи, соответственно, начинается беременность.
Во время полового акта (семяизвержения) сперма партнера попадает во влагалище, среда которого имеет повышенную кислотность, что приводит к гибели значительной части сперматозоидов. Те же половые клетки, которые выжили, продвигаются к шейке матки и далее в саму полость матки. Из неё сперматозоиды попадают по маточным трубам к яичникам. Процесс оплодотворения происходит в самой широкой дистальной части маточной трубы – ампулярной.
Этапы оплодотворения яйцеклетки
Оплодотворение представляет собой сложный процесс, состоящий из ряда этапов. На каждом из них могут возникать определенные проблемы, что в свою очередь, может приводить к бесплодию.
- Пенетрация. В том случае, если произошла встреча яйцеклетки и мужских половых клеток, один из сперматозоидов должен преодолеть некоторые барьеры – лучистый венец и оболочку яйцеклетки, которая называется блестящей. С целью разрушения лучистого венца сперматозоид использует гиалуронидазу (специальный фермент), который расположен на поверхностной части его головки. После того, как разрушен первый барьер, происходит соприкосновение сперматозоида с блестящей оболочкой. Следует обратить внимание, что для успешного зачатия ребенка недостаточно только одного сперматозоида, ведь он не сможет преодолеть защитные оболочки самостоятельно. Доказано, что лишь при контакте определенного значительного количества сперматозоидов с яйцеклеткой становится возможным разрушение лучистого венца.

Следующий этап – преодоление блестящей оболочки яйцеклетки. Для этого у сперматозоида предусмотрена такая часть, как акросома. Она представляет собой пузырек с ферментами, располагающийся на переднем конце головки самого сперматозоида. После этого лиганды, имеющиеся на блестящей оболочке яйцеклетки, взаимодействуют с рецепторами головки сперматозоида. Далее ферменты акросомы мужской половой клетки разрушают блестящую оболочку женской половой клетки в месте соприкосновения. Сперматозоид проникает под преодоленную оболочку.
- Кортикальная реакция. После того, как ядро сперматозоида оказалось внутри яйцеклетки, в последней происходит кортикальная реакция – высвобождение из специальных гранул их ферментов в окружающую среду. Эти ферменты делают блестящую оболочку яйцеклетки непроницаемой для остальных сперматозоидов.
- Образование зиготы. После того, как произошло успешное слияние мужской и женской половых клеток (оплодотворение), образуется зигота – одноклеточная форма эмбриона. Эта стадия длится примерно 26 – 30 часов.


Оплодотворение: этапы развития эмбриона
- Дробление. Зигота продвигается по маточным трубам к полости матки и в ней начинается процесс дробления – митотического деления. На этом этапе размер эмбриона не увеличивается, а только количество его клеток. Длится процесс деления около 3 суток.
- Дифференциация. На 4-е сутки клетки эмбриона разделяются на два слоя – эмбриобласт и трофобласт.
- Стадия бластоцисты. На 5-е сутки после оплодотворения яйцеклетки сперматозоидом эмбрион формирует бластоцисту, которая состоит из порядка 30 клеток, а далее, к концу своего развития — из 200 клеток.
- Имплантация. На 7 – 10 сутки после того, как произошло оплодотворение, бластоциста прикрепляется к эндометрию (внутренний слой, выстилающий полость матки). На этапе имплантации происходит тесное физическое и химическое взаимодействие эндометрия и бластоцисты. В итоге эндометрий окружает её со всех сторон.

На всех из вышеперечисленных этапов возможен сбой вследствие ряда провоцирующих факторов внешней и внутренней среды женского организма и, даже если произошло оплодотворение, беременность не развивается. Если это повторяется систематически – возникает бесплодие.
Тщательное обследование и качественное лечение всех видов бесплодия проводится в клинике «Центр ЭКО» Курск.
← Назад
Отзывы пациентов:
(Все отзывы)
Популярные программы ЭКО:
«Стоимость программ ЭКО»
Экстракорпоральное оплодотворение (ЭКО) – современный и успешный метод лечения бесплодия у женщин и мужчин.
Комплексные программы
Оплодотворение яйцеклетки
Оплодотворение представляет собой процесс слияния двух половых клеток – мужской (сперматозоида) и женской (яйцеклетки), в результате которого образуется зигота. С этого момента берёт начало эмбриональный период развития организма. Он происходит одинаково и после спонтанного зачатия, в результате вспомогательных репродуктивных технологий.
Механизм оплодотворения
При нормальном цикле у женщины репродуктивного возраста ежемесячно созревает доминантный фолликул, он лопается и готовая к оплодотворению яйцеклетка выходит в брюшную полость. Там в случае хорошего функционирования фибрин она захватывается воронкой фаллопиевой трубы. В трубе она дожидается сперматозоидов.
Во время полового акта в момент эякуляции семенная жидкость, которая содержит около сорока миллионов сперматозоидов, изливается во влагалище женщины. Они движутся через шейку матки в её полость.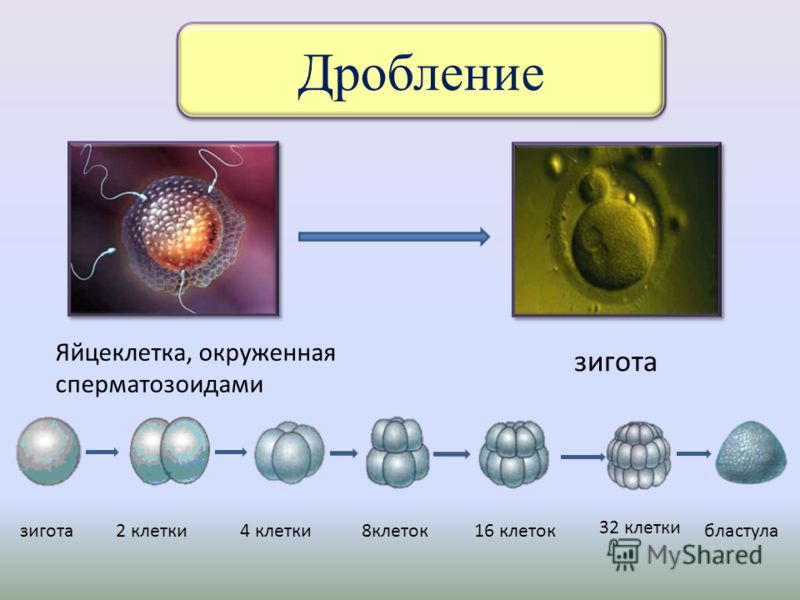 Этому способствует цервикальная слизь. Далее спермии направляются в маточную трубу, где на противоположном конце встречаются с яйцеклеткой. В трубе кислотность среды ниже, чем во влагалище. В ней содержатся питательные вещества. Благодаря этому сперматозоиды в маточной трубе сохраняют жизнеспособность более двух дней. Они ожидают, пока яйцеклетка созреет к оплодотворению.
Этому способствует цервикальная слизь. Далее спермии направляются в маточную трубу, где на противоположном конце встречаются с яйцеклеткой. В трубе кислотность среды ниже, чем во влагалище. В ней содержатся питательные вещества. Благодаря этому сперматозоиды в маточной трубе сохраняют жизнеспособность более двух дней. Они ожидают, пока яйцеклетка созреет к оплодотворению.
Сперматозоиды обладают высокой подвижностью. Они, благодаря поступательным движениям, продвигаются к цели, не останавливаются ни на мгновение и, встречая достаточно много преград, отсеивают нежизнеспособных и слабых. До конца трубы добираются наиболее сильные и выносливые спермии. У них на кончике головки находится так называемая акросома. Она содержит литические ферменты, которые растворяют блестящую оболочку яйцеклетки. Для того чтобы произошло оплодотворение, требуется достаточно большое количество сперматозоидов. Они совместными усилиями растворяют оболочку яйцеклетки. Этот процесс называется «акросомальная реакция».
Как только растворится оболочка яйцеклетки, наиболее шустрый сперматозоид проникает сквозь её клеточную мембрану. В этот же момент заряд клеточной мембраны меняется на противоположный. Остальные спермии отталкиваются от неё. По этой причине оплодотворение яйцеклетки возможно только одним сперматозоидом. Так случается в девяноста восьми процентах случаев оплодотворения.
Генетическая информация отца заложена в головке сперматозоида. Как только она проникает вовнутрь женской половой клетки, хвостик, который обеспечивает подвижность сперматозоида, тут же отпадает. В этот момент и происходит оплодотворение.
Изменения в организме женщины после оплодотворения
Организм женщины с момента оплодотворения начинает перестраиваться, чтобы сохранить беременность. На месте лопнувшего фолликула образуется жёлтое тело, которое синтезирует специальный гормон, поддерживающий необходимые физиологические процессы. Снижается уровень напряжённости иммунитета, чтобы не наступило отторжения плода.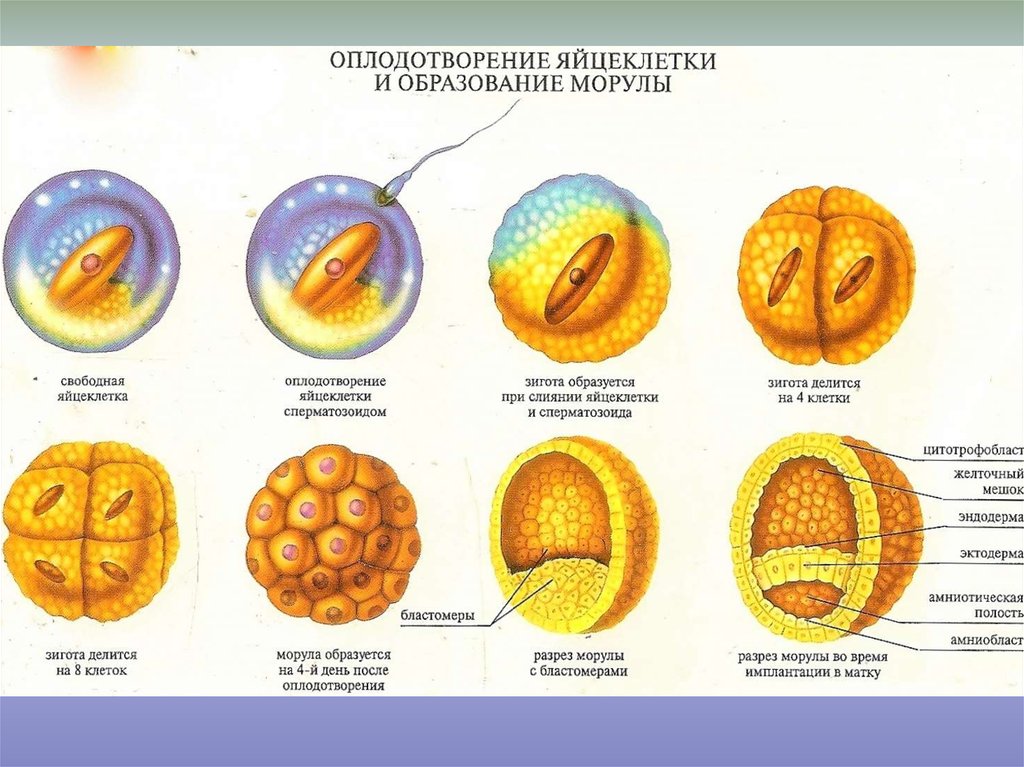
Начинает утолщаться кора надпочечников, которая вырабатывает большее количество глюкокортикоидов. Это гормоны, способствующие повышению устойчивости к стрессу, каковым для женщины является беременность. Они также обладают иммуносупрессорным действием, а плод ведь является наполовину чужеродным для женского организма.
Меняется уровень гонадотропинов и тиреотропных гормонов в крови беременной женщины. Плацента начинает синтезировать огромное количество хорионического гормона человека, являющегося гормоном беременности. Прогрессирующее повышение его уровня в крови указывает на то, что не только оплодотворение произошло успешно, но и эмбрион имплантировался в матку, а значит, беременность развивается нормально.
Физиологические процессы после оплодотворения
В процессе оплодотворения из двух клеток родителей, содержащих половинный набор хромосом, образуется одна. Она называется зигота. Она содержит генетический код, определяющий пол ребёнка, характер, разрез и цвет глаз, а также все фенотипические признаки.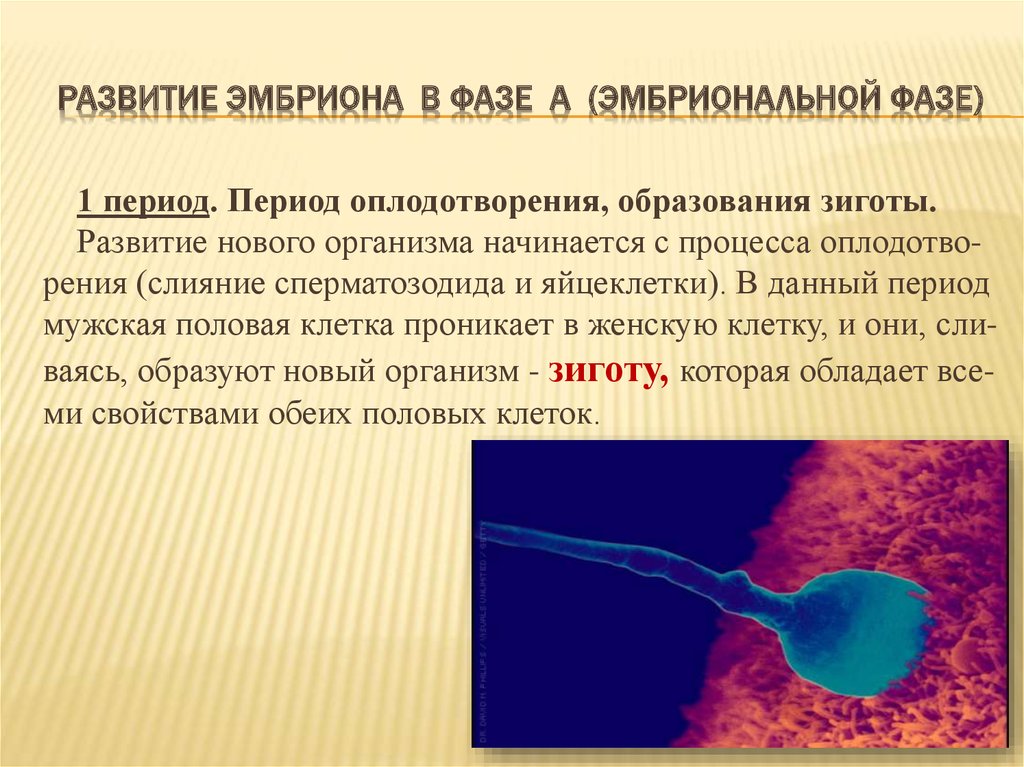
Зигота попадает в маточную трубу из брюшной полости. В течение трёх или четырёх суток она продвигается по направлению к матке. Этот процесс происходит благодаря перистальтическим движениям маточной трубы и колебаниям ресничек её эпителия. При нарушении их функции даже в том случае, когда оплодотворение произошло успешно, женщина может не забеременеть. Зародыш в маточной трубе дробится на дочерние клетки (бластомеры). Он попадает в матку на стадии морулы и состоит из множества клеток. Он покрыт защитной оболочкой. Но на пятый или шестой день происходит хетчинг – разрушение блестящей оболочки, и эмбрион выходит наружу. В это время он имплантируется в стенку матки.
Перед хетчингом эмбрион называется «бластоциста». Процесс развития эмбриона, получившегося в результате оплодотворения, происходит дальше таким образом:
- из тёмных бластомеров образуется эмбриобласт, который является основой для развития тела зародыша, а также некоторые внезародышевые образования;
- мелкие светлые трофобласты окружают со всех сторон эмбриобласт и дают начало трофобласту, являющемуся специфической рано дифференцирующейся тканью, которая дальнейшем обеспечивает имплантацию и питание зародыша;
- между зачатками эмбриобласта и трофобласта образуется небольшая полость, которая заполнена жидкостью.
В конце первой или начале второй недели после того, как произошло оплодотворение, происходит имплантация – плодное яйцо внедряется в слизистую оболочку матки. Далее начинает вырабатываться гормон беременности ХГЧ, наличие которого определяют в моче при помощи специального теста.
Тест на беременность проводят следующим образом: полоску необходимо опустить в утреннюю мочу на несколько секунд. Если оплодотворение и имплантация произошли, то в моче будет гормон беременности, и на тест-полоске появятся две чёрточки. Если же этого не произошло то может появиться всего лишь одна полоска. Домашние тесты на беременность, которые можно приобрести в аптеке, эффективны с четырнадцатого дня после оплодотворения или с первого дня задержки менструации. Для того чтобы убедиться в достоверности результата, исследование можно повторить.
Течение беременности после оплодотворения и имплантации эмбриона
Гинекологи условно выделяют три этапа, или триместра беременности. На первом из них происходит построение тела плода, а также органов, которые поддерживают его жизнеобеспечение. На втором этапе осуществляется закладка и настройка систем организма малыша. В третьем триместре плод готовится появиться на свет.
На первом из них происходит построение тела плода, а также органов, которые поддерживают его жизнеобеспечение. На втором этапе осуществляется закладка и настройка систем организма малыша. В третьем триместре плод готовится появиться на свет.
Для того чтобы плод развивался нормально, образуется амнион (околоплодный пузырь). Он представляет собой мешочек, который наполнен околоплодными водами, которые способствуют обмену веществ, защищают младенца от неравномерного давления стенок матки, ударов и шума, а также перепадов температуры. Неповторимым уникальным образованием является детское место, или плацента. Это источник жизни для маленького человечка.
По сосудам плаценты обогащённая кислородом кровь от матери поступает в пуповинную вену. Плацента — это одновременно почки, лёгкие и органы пищеварения ребёнка на этапе внутриутробного развития. Она создаёт заслон для токсинов и бактерий, служит в качестве фильтра, защищающего его от воздействия неблагоприятных факторов.
Кроме этого, в плаценте происходит синтез гормонов и биологически активных элементов, играющих важную роль в процессе внутриутробного развития плода. Пупочный канатик, или пуповина соединяет тело плода с плацентой. По её сосудам осуществляется кровоток от матери к плоду и обратно.
Пупочный канатик, или пуповина соединяет тело плода с плацентой. По её сосудам осуществляется кровоток от матери к плоду и обратно.
Если у вас не происходит оплодотворение, обращайтесь в «Центр ЭКО» Архангельск. Специалисты клиники выяснят, какие факторы являются этому причиной и определятся с методами лечения. При необходимости мы используем репродуктивные технологии.
Зигота | Определение, разработка, пример и факты
- Развлечения и поп-культура
- География и путешествия
- Здоровье и медицина
- Образ жизни и социальные вопросы
- Литература
- Философия и религия
- Политика, право и правительство
- Наука
- Спорт и отдых
- Технология
- Изобразительное искусство
- Всемирная история
- Этот день в истории
- Викторины
- Подкасты
- Словарь
- Биографии
- Резюме
- Популярные вопросы
- Инфографика
- Демистификация
- Списки
- #WTFact
- Товарищи
- Галереи изображений
- Прожектор
- Форум
- Один хороший факт
- Развлечения и поп-культура
- География и путешествия
- Здоровье и медицина
- Образ жизни и социальные вопросы
- Литература
- Философия и религия
- Политика, право и правительство
- Наука
- Спорт и отдых
- Технология
- Изобразительное искусство
- Всемирная история
- Britannica объясняет
В этих видеороликах Britannica объясняет различные темы и отвечает на часто задаваемые вопросы. - Britannica Classics
Посмотрите эти ретро-видео из архивов Encyclopedia Britannica. - Demystified Videos
В Demystified у Britannica есть все ответы на ваши животрепещущие вопросы. - #WTFact Видео
В #WTFact Britannica делится некоторыми из самых странных фактов, которые мы можем найти. - На этот раз в истории
В этих видеороликах узнайте, что произошло в этом месяце (или любом другом месяце!) в истории.

- Студенческий портал
Britannica — это главный ресурс для учащихся по ключевым школьным предметам, таким как история, государственное управление, литература и т. д. - Портал COVID-19
Хотя этот глобальный кризис в области здравоохранения продолжает развиваться, может быть полезно обратиться к прошлым пандемиям, чтобы лучше понять, как реагировать сегодня. - 100 женщин
Britannica празднует столетие Девятнадцатой поправки, выделяя суфражисток и политиков, творящих историю. - Спасение Земли
Британника представляет список дел Земли на 21 век. Узнайте об основных экологических проблемах, стоящих перед нашей планетой, и о том, что с ними можно сделать! - SpaceNext50
Britannica представляет SpaceNext50. От полета на Луну до управления космосом — мы изучаем широкий спектр тем, которые питают наше любопытство к космосу!
Содержание
- Введение
Краткие факты
- Связанный контент
Викторины
- Факты, которые вы должны знать: викторина о человеческом теле
Мейоз и оплодотворение — Клетка
Соматические клеточные циклы, обсуждавшиеся до сих пор в этой главе, приводят к образованию диплоидных дочерних клеток с идентичным генетическим дополнением. Мейоз, напротив, представляет собой особый вид клеточного цикла, при котором число хромосом уменьшается наполовину, что приводит к образованию гаплоидных дочерних клеток. Одноклеточные эукариоты, такие как дрожжи, могут подвергаться мейозу, а также размножаться путем митоза. Диплоид Saccharomyces cerevisiae , например, подвергаются мейозу и образуют споры при столкновении с неблагоприятными условиями окружающей среды. Однако у многоклеточных растений и животных мейоз ограничен зародышевыми клетками, где он играет ключевую роль в половом размножении. В то время как соматические клетки подвергаются митозу для пролиферации, зародышевые клетки подвергаются мейозу с образованием гаплоидных гамет (сперматозоидов и яйцеклеток). Затем в результате слияния этих гамет при оплодотворении инициируется развитие нового организма-потомка.
Мейоз, напротив, представляет собой особый вид клеточного цикла, при котором число хромосом уменьшается наполовину, что приводит к образованию гаплоидных дочерних клеток. Одноклеточные эукариоты, такие как дрожжи, могут подвергаться мейозу, а также размножаться путем митоза. Диплоид Saccharomyces cerevisiae , например, подвергаются мейозу и образуют споры при столкновении с неблагоприятными условиями окружающей среды. Однако у многоклеточных растений и животных мейоз ограничен зародышевыми клетками, где он играет ключевую роль в половом размножении. В то время как соматические клетки подвергаются митозу для пролиферации, зародышевые клетки подвергаются мейозу с образованием гаплоидных гамет (сперматозоидов и яйцеклеток). Затем в результате слияния этих гамет при оплодотворении инициируется развитие нового организма-потомка.
Процесс мейоза
В отличие от митоза, мейоз приводит к делению диплоидной родительской клетки на гаплоидное потомство, каждое из которых содержит только один член пары гомологичных хромосом, которые присутствовали в диплоидной родительской клетке (). Это уменьшение числа хромосом достигается за счет двух последовательных раундов деления ядер и клеток (называемых мейозом I и мейозом II), которые следуют за одним раундом репликации ДНК. Подобно митозу, мейоз I инициируется после завершения S-фазы и репликации родительских хромосом с образованием идентичных сестринских хроматид. Однако характер расхождения хромосом в мейозе I резко отличается от такового при митозе. Во время мейоза I гомологичные хромосомы сначала спариваются друг с другом, а затем расходятся в разные дочерние клетки. Сестринские хроматиды остаются вместе, поэтому завершение мейоза I приводит к образованию дочерних клеток, содержащих по одному члену каждой пары хромосом (состоящих из двух сестринских хроматид). За мейозом I следует мейоз II, который напоминает митоз тем, что сестринские хроматиды разделяются и расходятся в разные дочерние клетки. Таким образом, завершение мейоза II приводит к образованию четырех гаплоидных дочерних клеток, каждая из которых содержит только одну копию каждой хромосомы.
Это уменьшение числа хромосом достигается за счет двух последовательных раундов деления ядер и клеток (называемых мейозом I и мейозом II), которые следуют за одним раундом репликации ДНК. Подобно митозу, мейоз I инициируется после завершения S-фазы и репликации родительских хромосом с образованием идентичных сестринских хроматид. Однако характер расхождения хромосом в мейозе I резко отличается от такового при митозе. Во время мейоза I гомологичные хромосомы сначала спариваются друг с другом, а затем расходятся в разные дочерние клетки. Сестринские хроматиды остаются вместе, поэтому завершение мейоза I приводит к образованию дочерних клеток, содержащих по одному члену каждой пары хромосом (состоящих из двух сестринских хроматид). За мейозом I следует мейоз II, который напоминает митоз тем, что сестринские хроматиды разделяются и расходятся в разные дочерние клетки. Таким образом, завершение мейоза II приводит к образованию четырех гаплоидных дочерних клеток, каждая из которых содержит только одну копию каждой хромосомы.
Рисунок 14.32
Сравнение мейоза и митоза. И мейоз, и митоз инициируются после репликации ДНК, поэтому каждая хромосома состоит из двух сестринских хроматид. В мейозе I гомологичные хромосомы спариваются, а затем расходятся в разные клетки. Затем сестринские хроматиды разделяются (подробнее…)
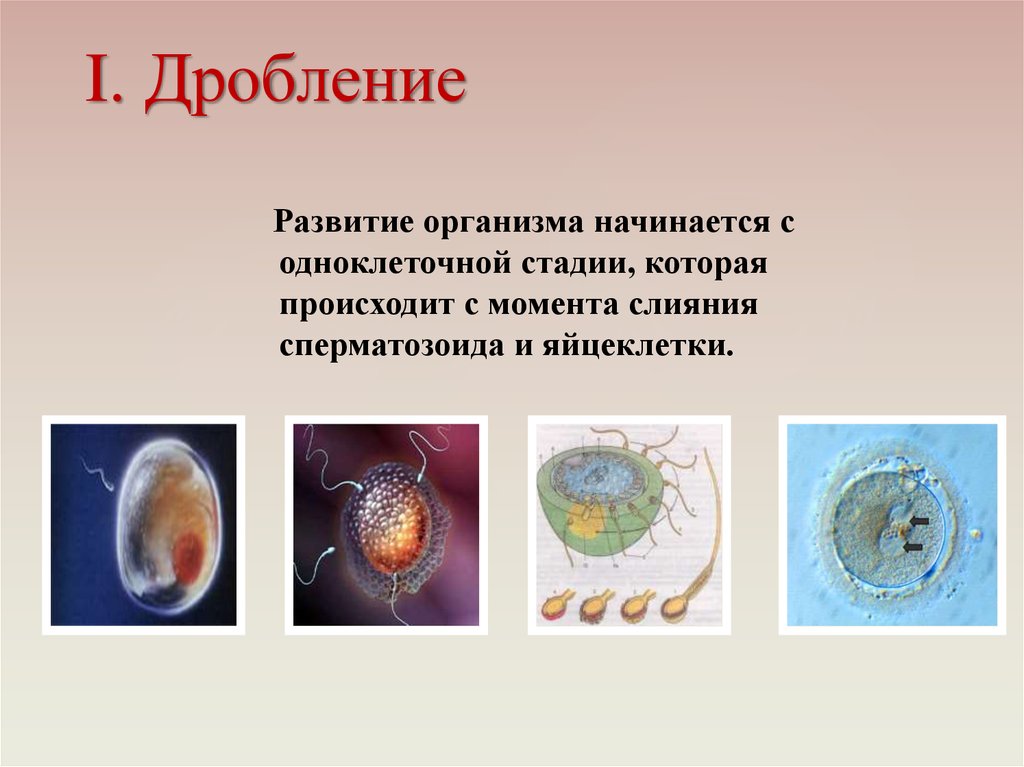
Спаривание гомологичных хромосом после репликации ДНК является не только ключевым событием, лежащим в основе мейотической сегрегации хромосом, но также делает возможной рекомбинацию между хромосомами отцовского и материнского происхождения. Это критическое спаривание гомологичных хромосом происходит во время расширенной профазы мейоза I, который подразделяется на пять стадий (лептотена, зиготена, пахитена, диплотена и диакинез) на основе морфологии хромосом (4). Считается, что первоначальная ассоциация гомологичных хромосом опосредована спариванием оснований между комплементарными цепями ДНК на стадии лептотены, прежде чем хроматин станет сильно конденсированным. Тесная ассоциация гомологичных хромосом (синапсис) начинается на стадии зиготены. Во время этой стадии по длине парных хромосом формируется подобная застежке-молнии белковая структура, называемая синаптонемным комплексом. Этот комплекс удерживает гомологичные хромосомы тесно связанными и выровненными друг с другом на стадии пахитены, которая может сохраняться в течение нескольких дней. Рекомбинация между гомологичными хромосомами завершается при их ассоциации на стадии пахитены, оставляя хромосомы сцепленными в местах кроссинговера (хиазмы; единичные числа, хиазмы). Синаптонемный комплекс исчезает на стадии диплотены и гомологичные хромосомы расходятся по своей длине. Важно, однако, что они остаются связанными в хиазмах, что имеет решающее значение для их правильного выравнивания в метафазе. На этой стадии каждая пара хромосом (называемая бивалентной) состоит из четырех хроматид с четко выраженными хиазмами (). Диакинез, завершающая стадия профазы I, представляет собой переход к метафазе, во время которой происходит полная конденсация хромосом.
Во время этой стадии по длине парных хромосом формируется подобная застежке-молнии белковая структура, называемая синаптонемным комплексом. Этот комплекс удерживает гомологичные хромосомы тесно связанными и выровненными друг с другом на стадии пахитены, которая может сохраняться в течение нескольких дней. Рекомбинация между гомологичными хромосомами завершается при их ассоциации на стадии пахитены, оставляя хромосомы сцепленными в местах кроссинговера (хиазмы; единичные числа, хиазмы). Синаптонемный комплекс исчезает на стадии диплотены и гомологичные хромосомы расходятся по своей длине. Важно, однако, что они остаются связанными в хиазмах, что имеет решающее значение для их правильного выравнивания в метафазе. На этой стадии каждая пара хромосом (называемая бивалентной) состоит из четырех хроматид с четко выраженными хиазмами (). Диакинез, завершающая стадия профазы I, представляет собой переход к метафазе, во время которой происходит полная конденсация хромосом.
Рисунок 14.
 33
33Стадии профазы мейоза I. Микрофотографии, иллюстрирующие морфологию хромосом лилии. (C. Hasenkampf/Biological Photo Service.)
Рис. 14.34
Синаптонемный комплекс. К боковым элементам прикреплены петли хроматина, которые соединены друг с другом центральным элементом в виде молнии.
Рис. 14.35
Двухвалентная хромосома на стадии диплотены. Бивалентная хромосома состоит из парных гомологичных хромосом. Сестринские хроматиды каждой хромосомы соединены центромерой. Хроматиды гомологичных хромосом соединяются в хиазмах, которых (далее…)
В метафазе I двухвалентные хромосомы выстраиваются на веретене. В отличие от митоза (см.), кинетохоры сестринских хроматид прилегают друг к другу и ориентированы в одном направлении, тогда как кинетохоры гомологичных хромосом направлены к противоположным полюсам веретена (см.). Следовательно, микротрубочки с одного полюса веретена прикрепляются к сестринским хроматидам, а микротрубочки с противоположных полюсов прикрепляются к гомологичным хромосомам. Анафаза I начинается с разрыва хиазм, в которых соединяются гомологичные хромосомы. Затем гомологичные хромосомы расходятся, а сестринские хроматиды остаются связанными в своих центромерах. Следовательно, по завершении мейоза I каждая дочерняя клетка приобретает по одному члену каждой гомологичной пары, состоящей из двух сестринских хроматид.
Анафаза I начинается с разрыва хиазм, в которых соединяются гомологичные хромосомы. Затем гомологичные хромосомы расходятся, а сестринские хроматиды остаются связанными в своих центромерах. Следовательно, по завершении мейоза I каждая дочерняя клетка приобретает по одному члену каждой гомологичной пары, состоящей из двух сестринских хроматид.
Рис. 14.36
Расхождение хромосом в мейозе I. В метафазе I кинетохоры сестринских хроматид сливаются или примыкают друг к другу. Таким образом, микротрубочки с одного полюса веретена прикрепляются к кинетохорам сестринских хроматид, тогда как микротрубочки (подробнее…)
Мейоз II инициируется сразу после цитокинеза, обычно до полной деконденсации хромосом. В отличие от мейоза I мейоз II напоминает нормальный митоз. В метафазе II хромосомы выстраиваются на веретене с микротрубочками от противоположных полюсов веретена, прикрепленными к кинетохорам сестринских хроматид. Связь между центромерами сестринских хроматид разрывается в анафазе II, и сестринские хроматиды расходятся к противоположным полюсам. Затем следует цитокинез, дающий начало гаплоидным дочерним клеткам.
Затем следует цитокинез, дающий начало гаплоидным дочерним клеткам.
Регуляция мейоза ооцитов
Ооциты позвоночных (развивающиеся яйца) были особенно полезными моделями для исследования клеточного цикла, отчасти из-за их большого размера и простоты манипуляций в лаборатории. Примечательным примером, обсуждавшимся ранее в этой главе, является открытие и последующая очистка MPF из ооцитов лягушки. Мейоз этих ооцитов, как и у других видов, регулируется в двух уникальных точках клеточного цикла, и исследования мейоза ооцитов выявили новые механизмы контроля клеточного цикла.
Первая регуляторная точка мейоза ооцитов приходится на стадию диплотены первого мейотического деления (). Ооциты могут оставаться задержанными на этой стадии в течение длительного периода времени — до 40–50 лет у человека. Во время этой остановки диплотены хромосомы ооцитов деконденсируются и активно транскрибируются. Эта транскрипционная активность отражается в огромном росте ооцитов в этот период. Ооциты человека, например, имеют диаметр около 100 мкм (более чем в сто раз больше объема типичной соматической клетки). Ооциты лягушки еще крупнее, их диаметр составляет примерно 1 мм. В этот период клеточного роста ооциты накапливают запасы материалов, включая РНК и белки, которые необходимы для поддержки раннего развития эмбриона. Как отмечалось ранее в этой главе, ранние эмбриональные клеточные циклы затем происходят в отсутствие клеточного роста, быстро деля оплодотворенную яйцеклетку на более мелкие клетки (см. Ресурсы).
Ооциты человека, например, имеют диаметр около 100 мкм (более чем в сто раз больше объема типичной соматической клетки). Ооциты лягушки еще крупнее, их диаметр составляет примерно 1 мм. В этот период клеточного роста ооциты накапливают запасы материалов, включая РНК и белки, которые необходимы для поддержки раннего развития эмбриона. Как отмечалось ранее в этой главе, ранние эмбриональные клеточные циклы затем происходят в отсутствие клеточного роста, быстро деля оплодотворенную яйцеклетку на более мелкие клетки (см. Ресурсы).
Рисунок 14.37
Мейоз ооцитов позвоночных. Мейоз останавливается на стадии диплотены, во время которой ооциты вырастают до больших размеров. Затем ооциты возобновляют мейоз в ответ на гормональную стимуляцию и завершают первое мейотическое деление с асимметричным цитокинезом (подробнее…)
Ооциты разных видов различаются в зависимости от того, когда возобновляется мейоз и происходит оплодотворение. У некоторых животных ооциты остаются задержанными на стадии диплотены до тех пор, пока они не будут оплодотворены, и только затем переходят к полному мейозу. Однако ооциты большинства позвоночных (включая лягушек, мышей и человека) возобновляют мейоз в ответ на гормональную стимуляцию и проходят через мейоз I до оплодотворения. Деление клеток после мейоза I асимметрично, что приводит к образованию небольшого полярное тельце и ооцит, сохранивший свой крупный размер. Затем ооцит вступает в мейоз II, не переформировав ядро и не деконденсировав свои хромосомы. Затем большинство ооцитов позвоночных снова задерживаются в метафазе II, где они остаются до оплодотворения.
Однако ооциты большинства позвоночных (включая лягушек, мышей и человека) возобновляют мейоз в ответ на гормональную стимуляцию и проходят через мейоз I до оплодотворения. Деление клеток после мейоза I асимметрично, что приводит к образованию небольшого полярное тельце и ооцит, сохранивший свой крупный размер. Затем ооцит вступает в мейоз II, не переформировав ядро и не деконденсировав свои хромосомы. Затем большинство ооцитов позвоночных снова задерживаются в метафазе II, где они остаются до оплодотворения.
Подобно М-фазе соматических клеток, мейоз ооцитов контролируется MPF. Тем не менее, регуляция MPF во время мейоза ооцитов обнаруживает уникальные особенности, ответственные за остановку метафазы II (4). Гормональная стимуляция ооцитов с задержкой диплотена первоначально запускает возобновление мейоза путем активации MPF, как в G 2 к М переход соматических клеток. Как и при митозе, MPF вызывает конденсацию хромосом, разрушение ядерной оболочки и образование веретена деления. Затем активация комплекса В, стимулирующего анафазу, приводит к переходу мейоза I из метафазы в анафазу, сопровождающемуся снижением активности MPF. Однако после цитокинеза активность MPF снова повышается и остается высокой, пока яйцеклетка останавливается в метафазе II. Регуляторный механизм, уникальный для ооцитов, таким образом, поддерживает активность MPF во время остановки метафазы II, предотвращая переход от метафазы к анафазе мейоза II и инактивацию MPF, которая является результатом протеолиза циклина B во время нормальной M-фазы.
Затем активация комплекса В, стимулирующего анафазу, приводит к переходу мейоза I из метафазы в анафазу, сопровождающемуся снижением активности MPF. Однако после цитокинеза активность MPF снова повышается и остается высокой, пока яйцеклетка останавливается в метафазе II. Регуляторный механизм, уникальный для ооцитов, таким образом, поддерживает активность MPF во время остановки метафазы II, предотвращая переход от метафазы к анафазе мейоза II и инактивацию MPF, которая является результатом протеолиза циклина B во время нормальной M-фазы.
Рисунок 14.38
Активность MPF во время мейоза ооцитов. Гормональная стимуляция диплотеновых ооцитов активирует MPF, что приводит к переходу в метафазу I. Затем активность MPF падает при переходе от метафазы I к анафазе I. После завершения мейоза I активность MPF (подробнее…)
Фактор, ответственный за метафазу Арест II был впервые обнаружен Йошио Масуи и Клементом Маркертом в 1971 году в той же серии экспериментов, которая привела к открытию MPF. В этом случае, однако, цитоплазму из яйцеклетки, задержанной в метафазе II, инъецировали в клетку раннего эмбриона, которая подвергалась митотическим клеточным циклам (14). Эта инъекция цитоплазмы яйца вызывала остановку эмбриональной клетки в метафазе, указывая на то, что остановка метафазы была вызвана цитоплазматическим фактором, присутствующим в яйцеклетке. Поскольку этот фактор останавливал митоз, его назвали цитостатический фактор ( CSF ).
В этом случае, однако, цитоплазму из яйцеклетки, задержанной в метафазе II, инъецировали в клетку раннего эмбриона, которая подвергалась митотическим клеточным циклам (14). Эта инъекция цитоплазмы яйца вызывала остановку эмбриональной клетки в метафазе, указывая на то, что остановка метафазы была вызвана цитоплазматическим фактором, присутствующим в яйцеклетке. Поскольку этот фактор останавливал митоз, его назвали цитостатический фактор ( CSF ).
Рисунок 14.39
Идентификация цитостатического фактора. Цитоплазму из яйца метафазы II микроинъецируют в одну клетку двухклеточного эмбриона. Инъецированная эмбриональная клетка останавливается в метафазе, в то время как неинъецированная клетка продолжает делиться. Фактор в цитоплазме яйца метафазы II (подробнее…)
Более поздние эксперименты выявили протеин-серин/треонинкиназу, известную как Mos , в качестве основного компонента ЦСЖ. Mos специфически синтезируется в ооцитах во время завершения мейоза I и затем необходим как для увеличения активности MPF во время мейоза II, так и для поддержания активности MPF во время остановки метафазы II. Действие Mos является результатом активации киназы ERK MAP, которая играет центральную роль в клеточных сигнальных путях, обсуждавшихся в предыдущей главе. Однако в ооцитах ERK играет другую роль; он активирует другую протеинкиназу, называемую Rsk, которая ингибирует действие комплекса, стимулирующего анафазу, и останавливает мейоз в метафазе II (4). Ооциты могут оставаться задержанными на этом этапе мейотического клеточного цикла в течение нескольких дней, ожидая оплодотворения.
Действие Mos является результатом активации киназы ERK MAP, которая играет центральную роль в клеточных сигнальных путях, обсуждавшихся в предыдущей главе. Однако в ооцитах ERK играет другую роль; он активирует другую протеинкиназу, называемую Rsk, которая ингибирует действие комплекса, стимулирующего анафазу, и останавливает мейоз в метафазе II (4). Ооциты могут оставаться задержанными на этом этапе мейотического клеточного цикла в течение нескольких дней, ожидая оплодотворения.
Рисунок 14.40
Поддержание остановки метафазы II с помощью протеинкиназы Mos. Протеинкиназа Mos поддерживает остановку метафазы II, ингибируя комплекс, способствующий анафазе. Действие Mos опосредуется протеинкиназами MEK, ERK и Rsk.
Оплодотворение
При оплодотворении сперматозоиды связываются с рецептором на поверхности яйцеклетки и сливаются с плазматической мембраной яйцеклетки, инициируя развитие нового диплоидного организма, содержащего генетическую информацию, полученную от обоих родителей (). Оплодотворение не только приводит к смешению отцовских и материнских хромосом, но и вызывает ряд изменений в цитоплазме яйцеклетки, имеющих решающее значение для дальнейшего развития. Эти изменения активируют яйцеклетку, что приводит к завершению мейоза ооцитов и инициации митотических клеточных циклов раннего эмбриона.
Оплодотворение не только приводит к смешению отцовских и материнских хромосом, но и вызывает ряд изменений в цитоплазме яйцеклетки, имеющих решающее значение для дальнейшего развития. Эти изменения активируют яйцеклетку, что приводит к завершению мейоза ооцитов и инициации митотических клеточных циклов раннего эмбриона.
Рисунок 14.41
Внесение удобрений. Сканирующая электронная микрофотография человеческого сперматозоида, оплодотворяющего яйцеклетку. (David M. Philips/Visuals Unlimited.)
Ключевым сигналом, возникающим в результате связывания сперматозоида с его рецептором на плазматической мембране яйцеклетки, является повышение уровня Ca 2+ в цитоплазме яйцеклетки, вероятно, как следствие стимуляции гидролиза фосфатидилинозитол 4,5-бисфосфата (PIP 2 ) (см. ). Один из эффектов этого повышения внутриклеточного Ca 2+ – индукция поверхностных изменений, препятствующих проникновению дополнительных сперматозоидов в яйцеклетку.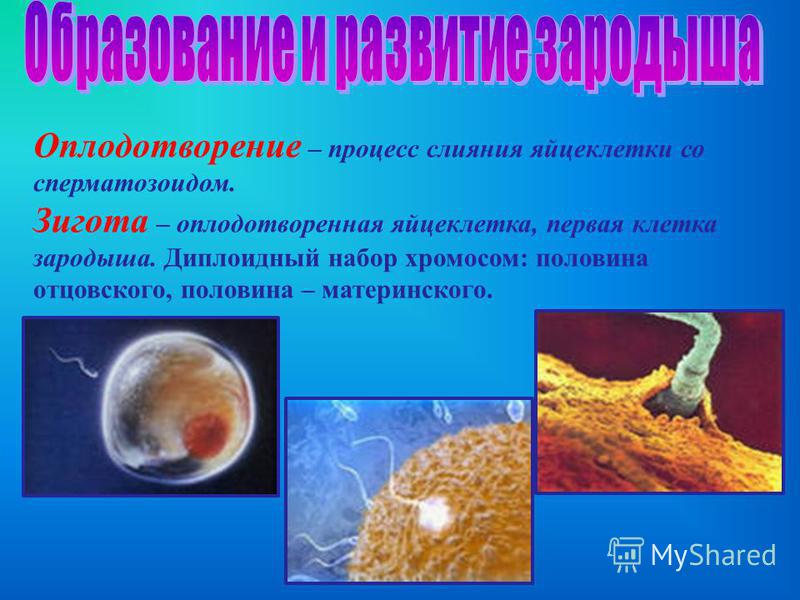 Поскольку яйцеклетки обычно одновременно подвергаются воздействию большого количества сперматозоидов, это критическое событие для обеспечения формирования нормального диплоидного эмбриона. Эти поверхностные изменения, как полагают, являются результатом, по крайней мере частично, Ca 2+ -индуцированного экзоцитоза секреторных везикул, которые присутствуют в большом количестве под плазматической мембраной яйцеклетки. Высвобождение содержимого этих везикул изменяет внеклеточную оболочку яйцеклетки, блокируя проникновение дополнительных сперматозоидов.
Поскольку яйцеклетки обычно одновременно подвергаются воздействию большого количества сперматозоидов, это критическое событие для обеспечения формирования нормального диплоидного эмбриона. Эти поверхностные изменения, как полагают, являются результатом, по крайней мере частично, Ca 2+ -индуцированного экзоцитоза секреторных везикул, которые присутствуют в большом количестве под плазматической мембраной яйцеклетки. Высвобождение содержимого этих везикул изменяет внеклеточную оболочку яйцеклетки, блокируя проникновение дополнительных сперматозоидов.
Увеличение цитозольного Ca 2+ после оплодотворения также свидетельствует о завершении мейоза (). В яйцах, остановленных в метафазе II, переход от метафазы к анафазе запускается Ca 2+ -зависимой активацией комплекса, способствующего анафазе. Результирующая инактивация MPF приводит к завершению второго мейотического деления с асимметричным цитокинезом (как в мейозе I), дающим начало второму маленькому полярному тельцу.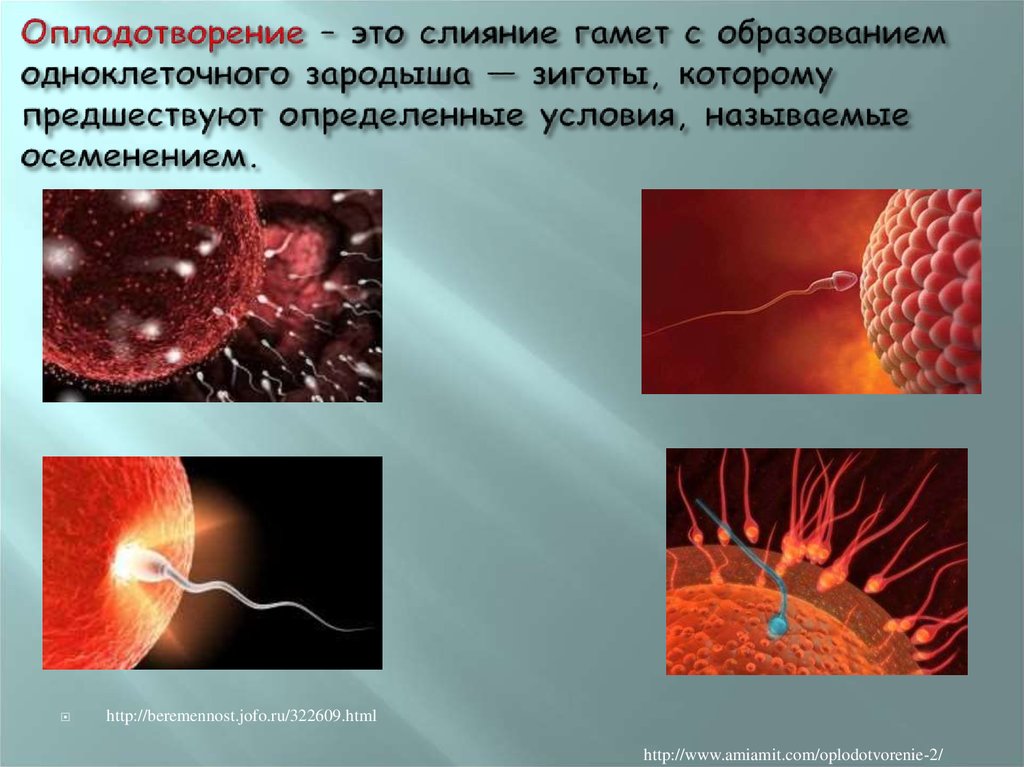
Рисунок 14.42
Оплодотворение и завершение мейоза. (A) Оплодотворение вызывает переход от метафазы II к анафазе II, что приводит к завершению мейоза ооцитов и выбросу второго полярного тельца (которое обычно дегенерирует). Ядро спермия деконденсируется, (далее…)
После завершения мейоза ооцита оплодотворенная яйцеклетка (теперь называемая зиготой) содержит два гаплоидных ядра (называемых пронуклеусами), по одному производному от каждого родителя. У млекопитающих два пронуклеуса затем вступают в S-фазу и реплицируют свою ДНК по мере миграции навстречу друг другу. Когда они встречаются, зигота вступает в М-фазу своего первого митотического деления. Две ядерные оболочки разрушаются, и конденсированные хромосомы как отцовского, так и материнского происхождения выстраиваются на общем веретене. После завершения митоза образуются две эмбриональные клетки, каждая из которых содержит новый диплоидный геном. Затем эти клетки начинают серию эмбриональных клеточных делений, которые в конечном итоге приводят к развитию нового организма.
Leave A Comment