
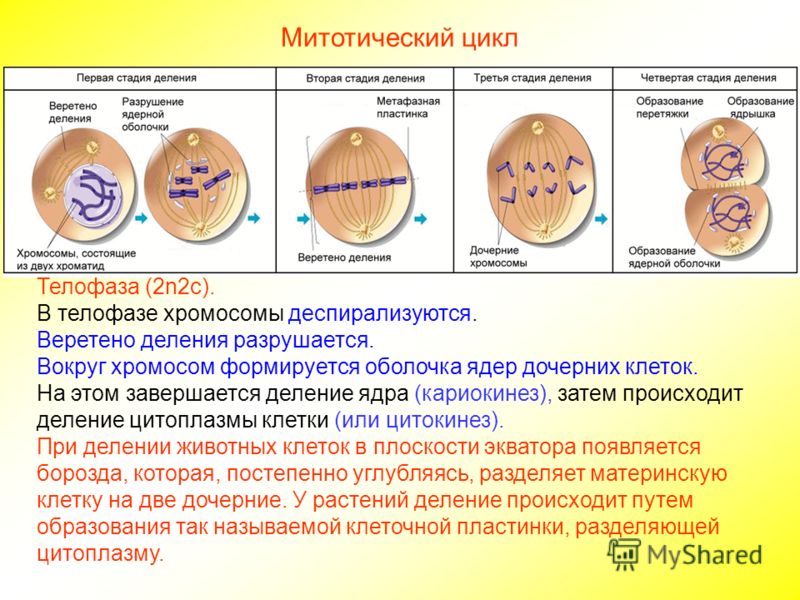
Стадии митоза
☰
Профаза — спирализация хромосом- Спирализация хромосом.
- Исчезновение ядрышек.
- Распад ядерной оболочки.
- Начало образования веретена деления.
- Прикрепление хромосом к нитям веретена и их движение к экваториальной плоскости клетки.
- Каждая хромосома стабилизируется в экваториальной плоскости за счет двух нитей, идущих с разных полюсов.
- Разрыв центромер хромосом.
- Каждая хроматида становится самостоятельной хромосомой.
- Сестринские хроматиды двигаются к разным полюсам клетки.


- Деспирализация хромосом и возобновление синтетических процессов в клетке.
- Образование ядрышек и ядерной оболочки.
- Цитокинез — деление тела клетки надвое.
Митоз — это деление клетки, при котором дочерние клетки генетически идентичны материнской и между собой. То есть при митозе хромосомы удваиваются и распределяются между дочерними клетками так, чтобы каждая получила по одной хроматиде каждой хромосомы.
В митозе выделяют несколько стадий (фаз). Однако самому митозу предшествует длительная интерфаза. Митоз и интерфаза вместе составляют клеточный цикл. В процессе интерфазы клетка растет, в ней образуются органоиды, активно идут процессы синтеза. В синтетическом периоде интерфазы редуплицируется, т.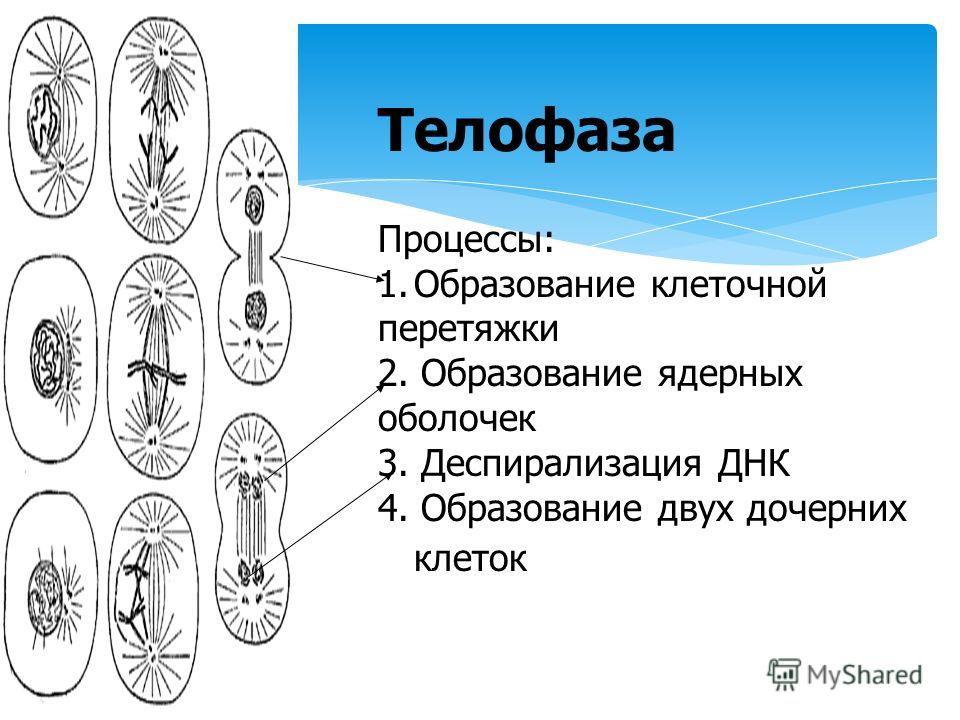
После удвоения хроматид они остаются соединенными в области центромеры, т. е. хромосома состоит из двух хроматид.
В самом митозе обычно выделяют четыре основные стадии (иногда больше).
Первая стадия митоза — профаза. В эту фазу хромосомы спирализуются и приобретают компактную скрученную форму. Из-за этого становятся невозможны процессы синтеза РНК. Исчезают ядрышки, а значит, рибосомы также не образуются, т. е. синтетические процессы в клетке приостанавливаются. Центриоли расходятся к полюсам (в разные концы) клетки, начинает образовываться веретено деления. В конце профазы распадается ядерная оболочка.
Прометафаза — это стадия, которую не всегда выделяют отдельно. Процессы, происходящие в ней, могут относить к поздней профазе или ранней метафазе. В прометафазе хромосомы оказываются в цитоплазме, беспорядочно перемещаются по клетке пока в районе центромеры не соединятся с нитью веретена деления.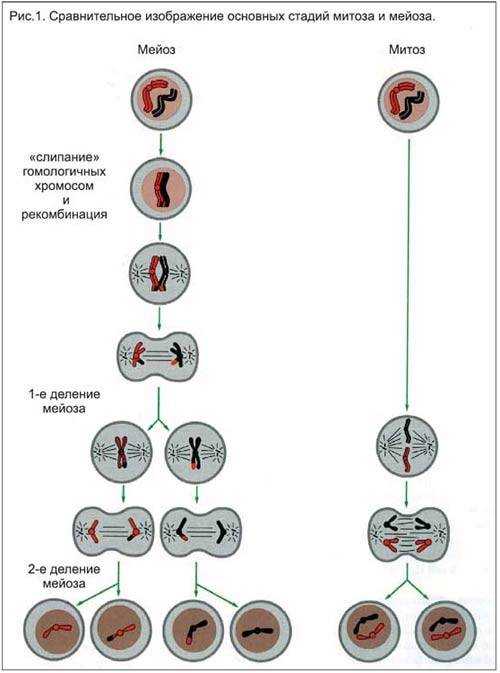
Нить представляет собой микротрубочку, построенную из белка тубулина. Она нарастает присоединяя новые тубулиновые субъединицы. При этом хромосома движется от полюса. Со стороны другого полюса к ней также присоединяется нить веретена и также толкает ее от полюса.
Вторая стадия митоза — метафаза. Все хромосомы располагаются в экваториальной области клетки рядом. К их центромерам прикреплено по две нити веретена деления. В митозе метафаза самая длительная стадия.
Третья стадия митоза — анафаза. В этой фазе хроматиды каждой хромосомы отделяются друг от друга и за счет тянущих их нитей веретена деления отходят к разным полюсам. Микротрубочки теперь не нарастают, а разбираются. Анафаза достаточно быстрая фаза митоза. При расхождении хромосом органоиды клетки примерно в равных количествах также расходятся ближе к полюсам.
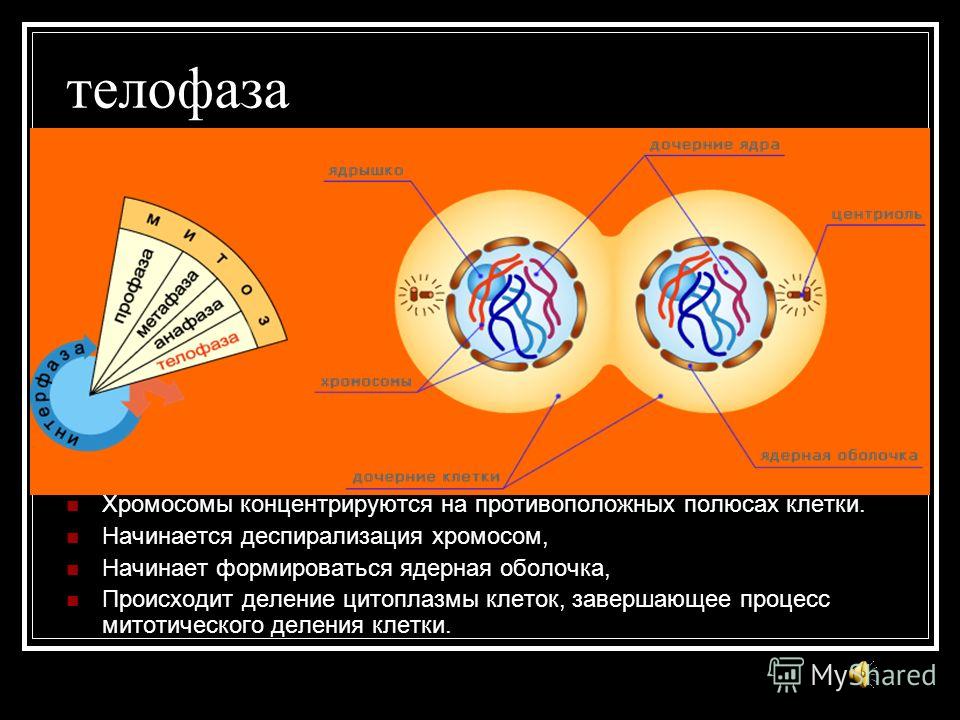
Четвертая стадия митоза — телофаза — во многом обратна профазе. Хроматиды собираются у полюсов клетки и раскручиваются, т.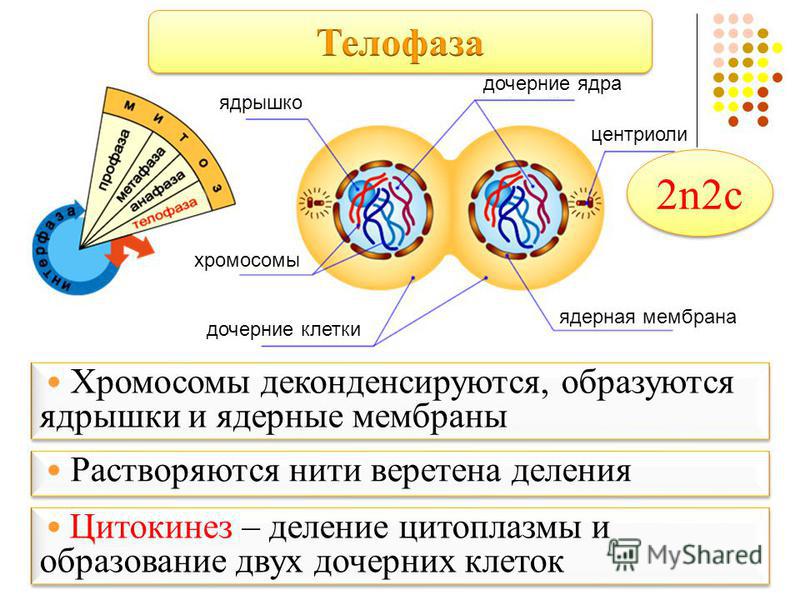 е. деспирализуются. Вокруг них формируются ядерные оболочки. Образуются ядрышки, начинается синтез РНК. Веретено деления начинает разрушаться. Далее происходит деление цитоплазмы — цитокинез. В клетках животных это происходит за счет впячивания мембраны внутрь и образования перетяжки. В клетках растений мембрана начинает формироваться внутри в экваториальной плоскости и идет к периферии.
е. деспирализуются. Вокруг них формируются ядерные оболочки. Образуются ядрышки, начинается синтез РНК. Веретено деления начинает разрушаться. Далее происходит деление цитоплазмы — цитокинез. В клетках животных это происходит за счет впячивания мембраны внутрь и образования перетяжки. В клетках растений мембрана начинает формироваться внутри в экваториальной плоскости и идет к периферии.
“Фазы митоза” таблица – интерфаза, профаза и анафаза, кратко
4.4
Средняя оценка: 4.4
Всего получено оценок: 984.
4.4
Средняя оценка: 4.4
Всего получено оценок: 984.
Рост и развитие живых организмов невозможен без процессов деления клеток. Одним из них является митоз – процесс деления эукариотических клеток, при котором образуются генетически идентичные клетки. Митоз можно назвать клонированием на клеточном уровне. В этой статье Вы подробнее узнаете об особенностях клеточного цикла, познакомитесь с характеристикой фаз митоза.
Материал подготовлен совместно с учителем высшей категории Макшаковой Натальей Алексеевной.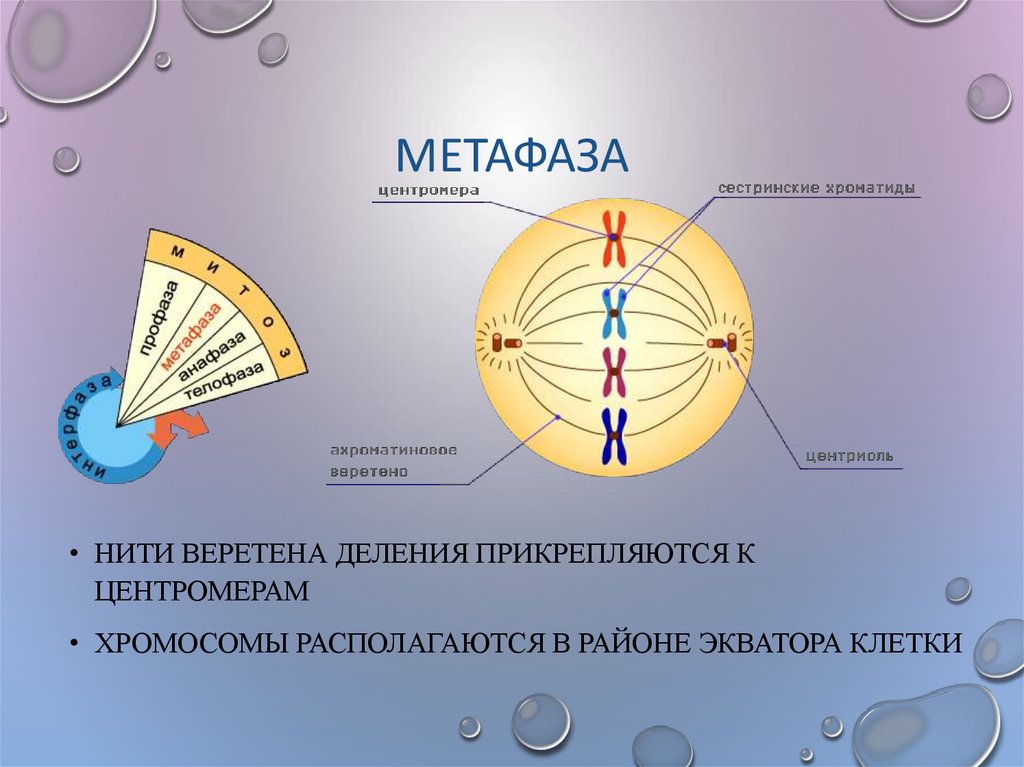
Опыт работы учителем биологии — 23 лет.
Понятие «клеточный цикл»
период существования клетки от момента её образования до следующего деления или гибели. Клеточный цикл включает в себя два периода: подготовка к делению (интерфаза) и собственно деление. Частью жизненного цикла также является состояние покоя клетки и период выполнения своих прямых функций.
Каким же образом при митозе из одной клетки образуются две точные ее генетические копии?
Это возможно благодаря двум важным процессам:- Самоудвоение или редупликация ДНК приводит к полному копированию всей наследственной информации, находящейся в клетке. Данный процесс происходит в период подготовки клетки к делению – интерфазе.
- Равномерное распределение генетического материала между дочерними клетками
Интерфаза митоза включает в себя три периода: пресинтетический, синтетический и постсинтетический. Именно в синтетический период происходит редупликация ДНК. В остальное время клетка растёт, идет синтез белка, накопление энергии в АТФ, удвоение органоидов.
В остальное время клетка растёт, идет синтез белка, накопление энергии в АТФ, удвоение органоидов.
Биологическое значение митоза состоит в строго одинаковом распределении молекул ДНК между дочерними клетками, что обеспечивает сохранение наследственной информации в ряду клеточных поколений. Митотическое деление клеток лежит в основе роста и развития многоклеточных форм жизни, регенерации утраченных клеток, бесполого размножения организмов.
Процесс непосредственного деления клетки, митоз, происходит в четыре фазы, соответствуя такой последовательности:
- профаза;
- метафаза;
- анафаза;
- телофаза.
Познакомиться с кратким описанием каждой фазы можно в таблице «Фазы митоза», которая представлена далее.
Таблица «Фазы митоза»
№ п/п | Фаза | Характеристика |
1. | Профаза | В профазе митоза происходит растворение ядерной оболочки и ядрышка, центриоли клеточного центра расходятся к разным полюсам, начинается формирование микротрубочек, так называемых нитей веретена деления. Происходит спирализация хромосом. Хромосомы, состоящие из двух хроматид (молекул ДНК) становятся видимыми в микроскоп. |
2. | Метафаза | На этом этапе хромосомы перемещаются и выстраиваются в экваториальной части веретена, образуя метафазную пластинку. Нити веретена деления прикрепляются к центромерам хромосом. |
3. | Анафаза | Является самой кратковременной фазой, во время которой происходит расхождение хроматид после распада центромер хромосом. Сестринские хромосомы (хроматиды) перемещаются к разным полюсам за счет сокращения нитей веретена деления. |
4. | Телофаза | Является заключительным этапом митоза, при котором новообразованные хромосомы располагаются на полюсах клетки и деспирализуются, обретая обычные размеры. |

 Вокруг них образуется новая ядерная оболочка с ядрышком внутри. Нити веретена распадаются и исчезают, начинается процесс деления цитоплазмы и её органоидов (цитотомия).
Вокруг них образуется новая ядерная оболочка с ядрышком внутри. Нити веретена распадаются и исчезают, начинается процесс деления цитоплазмы и её органоидов (цитотомия).Процесс цитотомии в животной клетке происходит при помощи борозды деления, а в растительной клетке – с помощью клеточной пластинки.
Нетипичные формы митоза
В природе иногда встречаются и нетипичные формы деления наследственного материала:
- Амитоз – способ прямого деления ядра, при котором сохраняется строение ядра, не происходит самоудвоения ДНК, хромосомы распределяются между ядрами случайным образом. В результате образуется двухъядерная клетка.
- Политения – кратно увеличивается число молекул ДНК, но без увеличения содержания хромосом.
- Эндомитоз – в ходе процесса после репликации ДНК нет разделения хромосом на дочерние, не следует процесс деления ядра и самой клетки.
Что мы узнали?
Процесс непрямого деления клеток эукариот проходит в несколько этапов, каждый из которых имеет свои особенности. Клеточный цикл состоит из интерфазы и непосредственного клеточного деления, состоящего из четырёх фаз: профазы, метафазы, анафазы и телофазы. Митоз обеспечивает точную передачу наследственной информации в ряду клеточных поколений. Иногда в природе встречаются нетипичные способы деления, к ним относятся амитоз, политения и эндомитоз.
Тест по теме
Доска почёта
Чтобы попасть сюда — пройдите тест.
Оценка доклада
4.4
Средняя оценка: 4.4
Всего получено оценок: 984.
А какая ваша оценка?
События М-фазы — Клетка
М-фаза — наиболее драматический период клеточного цикла, включающий серьезную реорганизацию практически всех клеточных компонентов.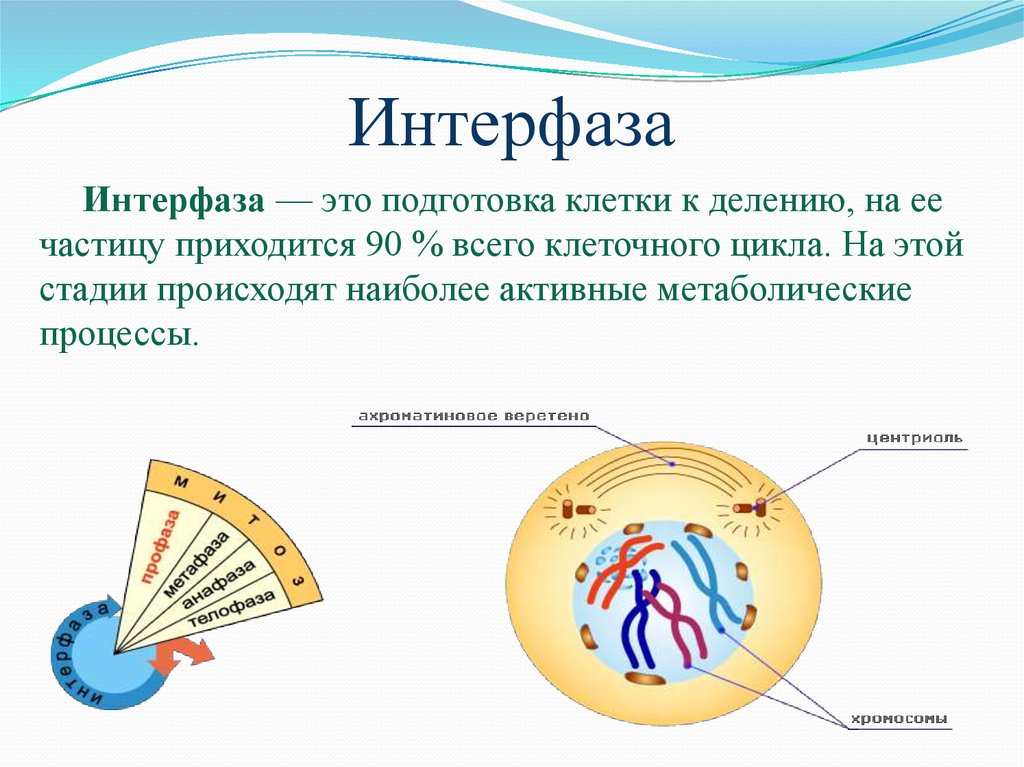 Во время митоза (деления ядра) хромосомы конденсируются, ядерная оболочка большинства клеток разрушается, цитоскелет реорганизуется, образуя митотическое веретено, и хромосомы расходятся к противоположным полюсам. За сегрегацией хромосом обычно следует деление клеток (цитокинез). Хотя многие из этих событий обсуждались в предыдущих главах в отношении структуры и функции ядра и цитоскелета, здесь они рассматриваются в контексте согласованного взгляда на М-фазу и действие MPF.
Во время митоза (деления ядра) хромосомы конденсируются, ядерная оболочка большинства клеток разрушается, цитоскелет реорганизуется, образуя митотическое веретено, и хромосомы расходятся к противоположным полюсам. За сегрегацией хромосом обычно следует деление клеток (цитокинез). Хотя многие из этих событий обсуждались в предыдущих главах в отношении структуры и функции ядра и цитоскелета, здесь они рассматриваются в контексте согласованного взгляда на М-фазу и действие MPF.
Стадии митоза
Хотя многие детали митоза различаются у разных организмов, фундаментальные процессы, обеспечивающие правильное расхождение сестринских хроматид, сохраняются у всех эукариот. Эти основные события митоза включают конденсацию хромосом, образование митотического веретена и прикрепление хромосом к микротрубочкам веретена. Затем сестринские хроматиды отделяются друг от друга и перемещаются к противоположным полюсам веретена деления с последующим образованием дочерних ядер.
Митоз условно делят на четыре стадии — профазу, метафазу, анафазу и телофазу, — которые проиллюстрированы для клетки животного в и . Начало профазы знаменуется появлением конденсированных хромосом, каждая из которых состоит из двух сестринских хроматид (дочерних молекул ДНК, образующихся в S-фазе). Эти вновь реплицированные молекулы ДНК остаются переплетенными на всем протяжении S и G 2 , распутываясь в процессе конденсации хроматина. Затем сконденсированные сестринские хроматиды удерживаются вместе на центромере, которая (как обсуждалось в главе 4) представляет собой последовательность ДНК, с которой связываются белки, образуя кинетохору — место возможного прикрепления микротрубочек веретена. В дополнение к конденсации хромосом в профазе инициируются цитоплазматические изменения, приводящие к развитию митотического веретена. Центросомы (удвоившиеся во время интерфазы) расходятся и расходятся к противоположным сторонам ядра. Там они служат двумя полюсами митотического веретена, которое начинает формироваться во время поздней профазы.
Начало профазы знаменуется появлением конденсированных хромосом, каждая из которых состоит из двух сестринских хроматид (дочерних молекул ДНК, образующихся в S-фазе). Эти вновь реплицированные молекулы ДНК остаются переплетенными на всем протяжении S и G 2 , распутываясь в процессе конденсации хроматина. Затем сконденсированные сестринские хроматиды удерживаются вместе на центромере, которая (как обсуждалось в главе 4) представляет собой последовательность ДНК, с которой связываются белки, образуя кинетохору — место возможного прикрепления микротрубочек веретена. В дополнение к конденсации хромосом в профазе инициируются цитоплазматические изменения, приводящие к развитию митотического веретена. Центросомы (удвоившиеся во время интерфазы) расходятся и расходятся к противоположным сторонам ядра. Там они служат двумя полюсами митотического веретена, которое начинает формироваться во время поздней профазы.
Рисунок 14.23
Стадии митоза в животной клетке. Во время профазы хромосомы конденсируются, и центросомы перемещаются к противоположным сторонам ядра, инициируя формирование митотического веретена. Затем разрушение ядерной оболочки позволяет прикрепить микротрубочки веретена (подробнее…)
Затем разрушение ядерной оболочки позволяет прикрепить микротрубочки веретена (подробнее…)
Рисунок 14.24
Флуоресцентные микрофотографии хроматина, кератина и микротрубочек во время митоза клеток легких тритона. Хроматин окрашивается в синий цвет, кератин — в красный, а микротрубочки — в зеленый. (Конли Л. Ридер/ Biological Photo Service.)
У высших эукариот окончание профазы соответствует распаду ядерной оболочки. Однако, как обсуждалось в главе 8, разрушение ядерной оболочки не является универсальным признаком митоза. В частности, дрожжи и многие другие одноклеточные эукариоты подвергаются «закрытому митозу», при котором ядерная оболочка остается неповрежденной (см. ). В этих клетках тела полюсов веретена погружены в ядерную оболочку, и ядро делится надвое после миграции дочерних хромосом к противоположным полюсам веретена.
После завершения профазы клетка вступает в прометафазу — переходный период между профазой и метафазой. Во время прометафазы микротрубочки митотического веретена прикрепляются к кинетохорам конденсированных хромосом. Кинетохоры сестринских хроматид ориентированы по разные стороны хромосомы, поэтому они прикрепляются к микротрубочкам, отходящим от противоположных полюсов веретена. Хромосомы перетасовываются взад и вперед, пока в конце концов не выровняются на метафазной пластинке в центре веретена. На этой стадии клетка достигла метафазы.
Кинетохоры сестринских хроматид ориентированы по разные стороны хромосомы, поэтому они прикрепляются к микротрубочкам, отходящим от противоположных полюсов веретена. Хромосомы перетасовываются взад и вперед, пока в конце концов не выровняются на метафазной пластинке в центре веретена. На этой стадии клетка достигла метафазы.
Большинство клеток лишь ненадолго остаются в метафазе, прежде чем перейти в анафазу. Переход от метафазы к анафазе запускается разрывом связи между сестринскими хроматидами, которые затем расходятся и расходятся к противоположным полюсам веретена. Митоз заканчивается телофазой, во время которой происходит переформирование ядер и деконденсация хромосом. Цитокинез обычно начинается во время поздней анафазы и почти завершается к концу телофазы, что приводит к образованию двух интерфазных дочерних клеток.
MPF и переход к метафазе
Митоз включает в себя драматические изменения во многих клеточных компонентах, ведущие к серьезной реорганизации всей структуры клетки.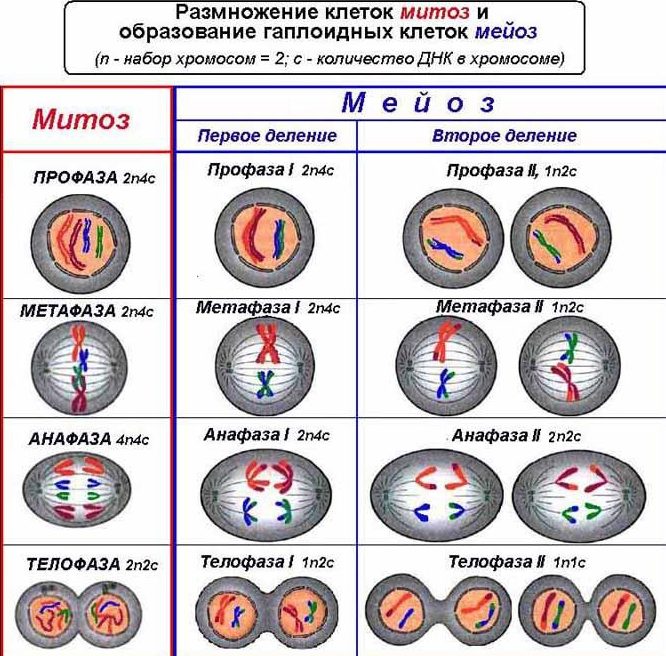 Как обсуждалось ранее в этой главе, эти события инициируются активацией протеинкиназы MPF (Cdc2/циклин B). По-видимому, MPF действует не только как мастер-регулятор М-фазного перехода, фосфорилируя и активируя другие нижележащие протеинкиназы, но также действует непосредственно, фосфорилируя некоторые структурные белки, участвующие в этой клеточной реорганизации (14).
Как обсуждалось ранее в этой главе, эти события инициируются активацией протеинкиназы MPF (Cdc2/циклин B). По-видимому, MPF действует не только как мастер-регулятор М-фазного перехода, фосфорилируя и активируя другие нижележащие протеинкиназы, но также действует непосредственно, фосфорилируя некоторые структурные белки, участвующие в этой клеточной реорганизации (14).
Рисунок 14.25
Цели MPF. MPF индуцирует множественные ядерные и цитоплазматические изменения в начале М-фазы как за счет активации других протеинкиназ, так и за счет фосфорилирования белков, таких как конденсины и ядерные ламины.
Конденсация интерфазного хроматина с образованием компактных хромосом митотических клеток является ключевым событием в митозе, критически важным для того, чтобы позволить хромосомам двигаться вдоль митотического веретена, не разрываясь и не спутываясь друг с другом. Как обсуждалось в главе 4, хроматин в интерфазных ядрах конденсируется почти в тысячу раз во время образования метафазных хромосом.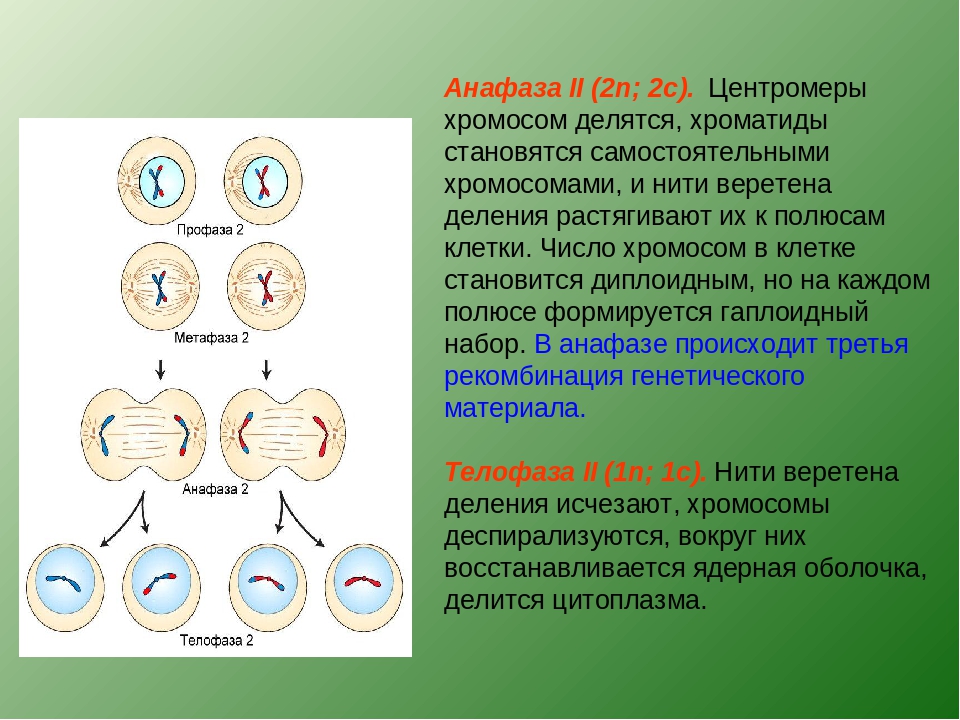 Такой высококонденсированный хроматин не может быть транскрибирован, поэтому транскрипция прекращается, когда происходит конденсация хроматина. Несмотря на фундаментальную важность этого события, мы не до конца понимаем ни строение метафазных хромосом, ни молекулярный механизм конденсации хроматина. Однако недавно было обнаружено, что белковые комплексы, называемые конденсинами, вызывают конденсацию хромосом, оборачивая ДНК вокруг себя, уплотняя хромосомы в конденсированную митотическую структуру. Конденсины фосфорилируются непосредственно протеинкиназой Cdc2, которая управляет конденсацией хроматина, активируя конденсины, когда клетки вступают в митоз. Одним молекулярным изменением, которое обычно сопровождает конденсацию хромосом, является фосфорилирование гистона h2, поэтому следует отметить, что гистон h2 также является субстратом для Cdc2. Однако фосфорилирование гистона h2 не требуется для митотической конденсации хромосом, поэтому значение фосфорилирования h2 с помощью Cdc2 неясно.
Такой высококонденсированный хроматин не может быть транскрибирован, поэтому транскрипция прекращается, когда происходит конденсация хроматина. Несмотря на фундаментальную важность этого события, мы не до конца понимаем ни строение метафазных хромосом, ни молекулярный механизм конденсации хроматина. Однако недавно было обнаружено, что белковые комплексы, называемые конденсинами, вызывают конденсацию хромосом, оборачивая ДНК вокруг себя, уплотняя хромосомы в конденсированную митотическую структуру. Конденсины фосфорилируются непосредственно протеинкиназой Cdc2, которая управляет конденсацией хроматина, активируя конденсины, когда клетки вступают в митоз. Одним молекулярным изменением, которое обычно сопровождает конденсацию хромосом, является фосфорилирование гистона h2, поэтому следует отметить, что гистон h2 также является субстратом для Cdc2. Однако фосфорилирование гистона h2 не требуется для митотической конденсации хромосом, поэтому значение фосфорилирования h2 с помощью Cdc2 неясно.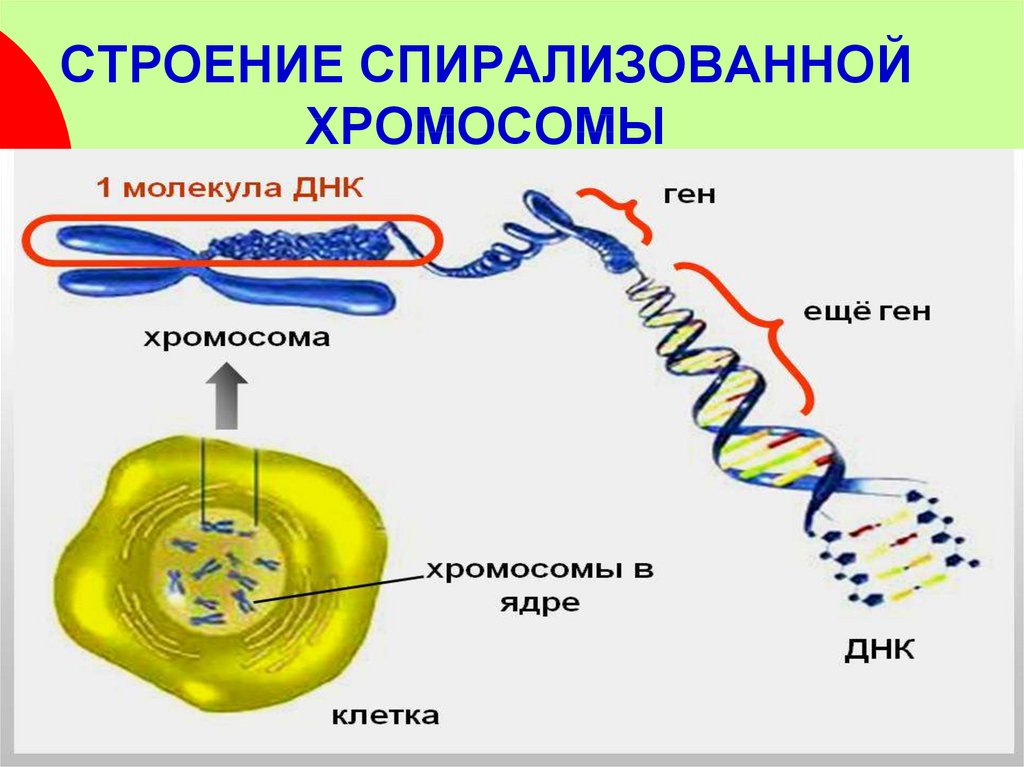 Напротив, было показано, что конденсация хромосом требует фосфорилирования гистона h4. Возможно неожиданно, однако, что гистон h4 не фосфорилируется с помощью Cdc2, и киназу, ответственную за фосфорилирование h4 в митотических клетках, еще предстоит идентифицировать.
Напротив, было показано, что конденсация хромосом требует фосфорилирования гистона h4. Возможно неожиданно, однако, что гистон h4 не фосфорилируется с помощью Cdc2, и киназу, ответственную за фосфорилирование h4 в митотических клетках, еще предстоит идентифицировать.
Разрушение ядерной оболочки, которое является одним из наиболее драматических событий митоза, представляет собой наиболее четко определенную цель для действия MPF. Как обсуждалось в главе 8, Cdc2 фосфорилирует ламины, что приводит непосредственно к деполимеризации ядерной ламины (see). Затем следует фрагментация ядерной мембраны на мелкие везикулы, которые в конце концов сливаются с образованием новых дочерних ядер в телофазе. Эндоплазматический ретикулум и аппарат Гольджи сходным образом фрагментируются на маленькие везикулы, которые затем могут распределяться по дочерним клеткам при цитокинезе. Разрушение этих мембран также индуцируется MPF и может быть частично опосредовано фосфорилированием Cdc2 матриксного белка Гольджи GM130, которое необходимо для прикрепления везикул, покрытых COPI, к мембране Гольджи.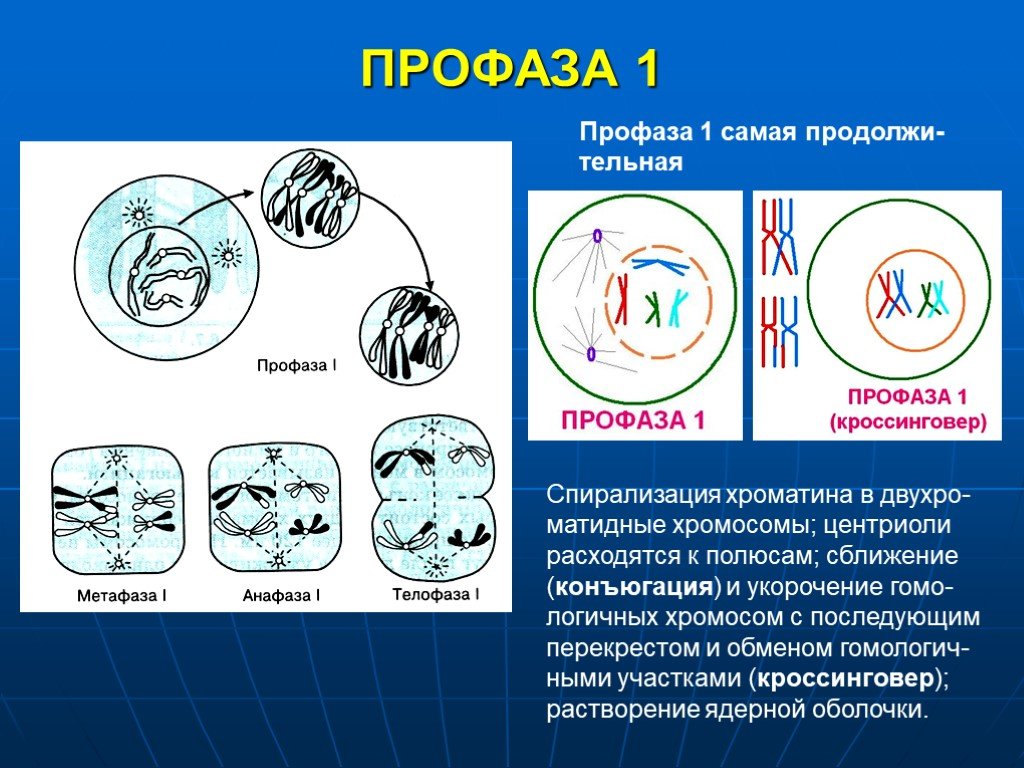 Фосфорилирование и инактивация GM130 с помощью Cdc2 ингибирует стыковку и слияние пузырьков, что приводит к фрагментации аппарата Гольджи. Однако могут быть задействованы и дополнительные мишени Cdc2, и механизмы, с помощью которых MPF приводит к фрагментации мембраны, еще предстоит полностью выяснить.
Фосфорилирование и инактивация GM130 с помощью Cdc2 ингибирует стыковку и слияние пузырьков, что приводит к фрагментации аппарата Гольджи. Однако могут быть задействованы и дополнительные мишени Cdc2, и механизмы, с помощью которых MPF приводит к фрагментации мембраны, еще предстоит полностью выяснить.
Реорганизация цитоскелета, завершающаяся образованием митотического веретена, является результатом динамической нестабильности микротрубочек (см. главу 11). В начале профазы центросомы расходятся к противоположным сторонам ядра. Затем повышение активности MPF вызывает резкое изменение динамического поведения микротрубочек. Во-первых, увеличивается скорость разборки микротрубочек, что приводит к деполимеризации и усадке интерфазных микротрубочек. Считается, что эта разборка является результатом фосфорилирования белков, ассоциированных с микротрубочками, либо самой MPF, либо другими MPF-активируемыми протеинкиназами. Кроме того, количество микротрубочек, отходящих от центросом, увеличивается, поэтому интерфазные микротрубочки замещаются большим количеством коротких микротрубочек, отходящих от центросом.
Затем разрушение ядерной оболочки позволяет некоторым микротрубочкам веретена прикрепляться к хромосомам в их кинетохорах (), инициируя процесс движения хромосом, который характеризует прометафазу. Белки, собранные на кинетохоре, включают моторы микротрубочек, которые направляют движение хромосом к минус-концам микротрубочек веретена, которые закреплены в центросоме. Действиям этих белков, которые притягивают хромосомы к центросоме, противостоит рост микротрубочек веретена, который отталкивает хромосомы от полюсов веретена. Следовательно, хромосомы в прометафазе перемещаются туда и обратно между центросомами и центром веретена.
Рисунок 14.26
Электронная микрофотография микротрубочек, прикрепленных к кинетохоре хромосомы. (Conly L. Rieder/ Biological Photo Service.)
Микротрубочки с противоположных полюсов веретена в конечном итоге прикрепляются к двум кинетохорам сестринских хроматид (которые расположены на противоположных сторонах хромосомы), и баланс сил, действующих на хромосомы приводит к их выравниванию на метафазной пластинке в центре веретена (). Как обсуждалось в главе 11, веретено состоит как из кинетохорных микротрубочек, прикрепленных к хромосомам, так и из полярных микротрубочек, перекрывающихся друг с другом в центре клетки. Кроме того, короткие астральные микротрубочки расходятся наружу от центросом к периферии клетки.
Как обсуждалось в главе 11, веретено состоит как из кинетохорных микротрубочек, прикрепленных к хромосомам, так и из полярных микротрубочек, перекрывающихся друг с другом в центре клетки. Кроме того, короткие астральные микротрубочки расходятся наружу от центросом к периферии клетки.
Рисунок 14.27
Веретено метафазы. (A) Веретено состоит из трех видов микротрубочек. Кинетохорные микротрубочки прикреплены к хромосомам, полярные микротрубочки перекрываются в центре клетки, а астральные микротрубочки расходятся от центросомы к клетке (подробнее…)
Протеолиз и инактивация MPF: анафаза и телофаза
Как обсуждалось ранее в этой главе важная контрольная точка клеточного цикла отслеживает выравнивание хромосом на метафазном веретене. После этого клетка переходит в анафазу и завершает митоз. Переход от метафазы к анафазе является результатом убиквитин-опосредованного протеолиза ключевых регуляторных белков, запускаемого активацией убиквитинлигазы (см. ), называемой комплексом, способствующим анафазе.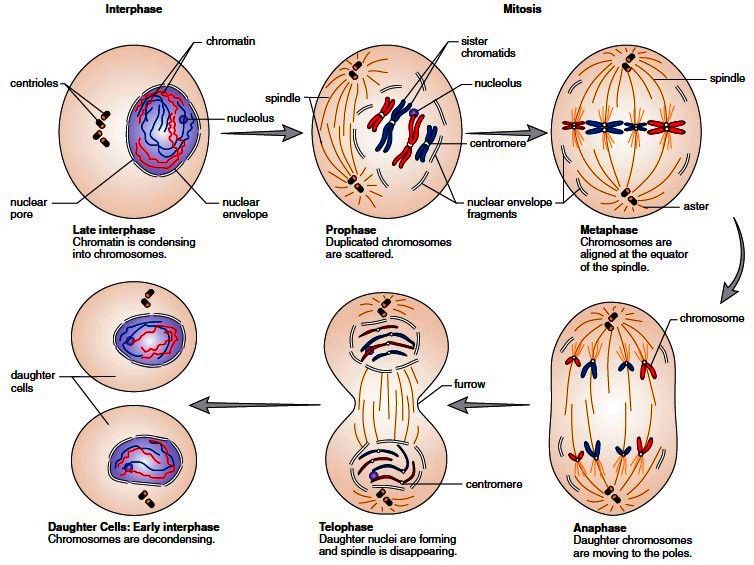 Активация комплекса, способствующего анафазе, индуцируется MPF в начале митоза, поэтому MPF в конечном итоге запускает собственное разрушение. Однако комплекс, стимулирующий анафазу, остается ингибированным до тех пор, пока клетка не пройдет контрольную точку метафазы, после чего активация системы деградации убиквитина приводит к переходу от метафазы к анафазе и прохождению через оставшуюся часть митоза.
Активация комплекса, способствующего анафазе, индуцируется MPF в начале митоза, поэтому MPF в конечном итоге запускает собственное разрушение. Однако комплекс, стимулирующий анафазу, остается ингибированным до тех пор, пока клетка не пройдет контрольную точку метафазы, после чего активация системы деградации убиквитина приводит к переходу от метафазы к анафазе и прохождению через оставшуюся часть митоза.
Активация комплекса, стимулирующего анафазу, приводит к деградации как минимум двух ключевых регуляторных белков (). Начало анафазы является результатом протеолитической деградации белка, называемого Scc1, компонента комплекса белков, называемых когезинами, которые поддерживают связь между сестринскими хроматидами, пока они выровнены на метафазной пластинке. Деградация Scc1 не катализируется непосредственно комплексом, способствующим анафазе, который вместо этого расщепляет регуляторный белок, называемый Pds1. Деградация Pds1, в свою очередь, активирует другой белок, называемый Esp1, что приводит к протеолизу cohesin Scc1.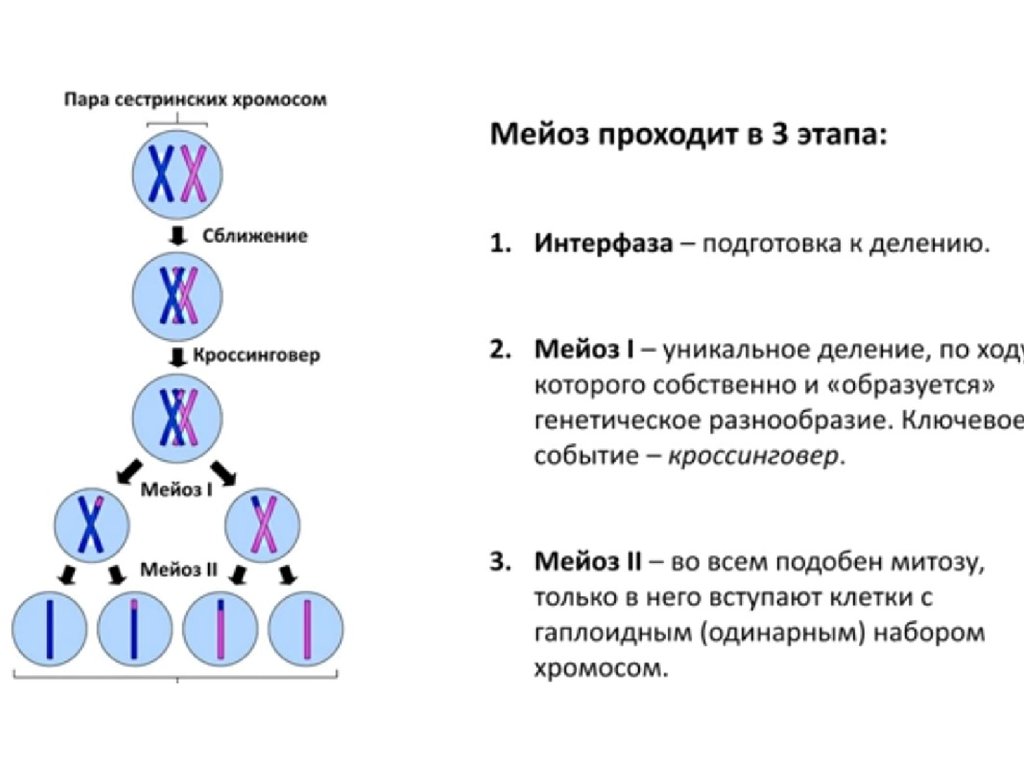 Расщепление Scc1 разрывает связь между сестринскими хроматидами, позволяя им расщепляться, перемещаясь к противоположным полюсам веретена (10). Затем разделение хромосом в анафазе происходит в результате действия нескольких типов моторных белков, связанных с микротрубочками веретена (см. и ).
Расщепление Scc1 разрывает связь между сестринскими хроматидами, позволяя им расщепляться, перемещаясь к противоположным полюсам веретена (10). Затем разделение хромосом в анафазе происходит в результате действия нескольких типов моторных белков, связанных с микротрубочками веретена (см. и ).
Рис. 14.28
Мишени системы протеолиза циклина В. Комплекс, стимулирующий анафазу, представляет собой убиквитинлигазу, которая активируется после прохождения контрольной точки метафазы. Его активация вызывает переход от метафазы к анафазе, приводя к (подробнее…)
Рисунок 14.29
Клетка сига в анафазе. (Michael Abbey/Photo Researchers, Inc.)
Другим ключевым регуляторным белком, предназначенным для убиквитинирования и деградации комплексом, стимулирующим анафазу, является циклин B. Деградация циклина B приводит к инактивации MPF, который необходим для выхода клетки из организма. митоз и возврат в интерфазу. Многие из клеточных изменений, связанных с этими переходами, являются просто обращением событий, вызванных MPF во время вступления в митоз.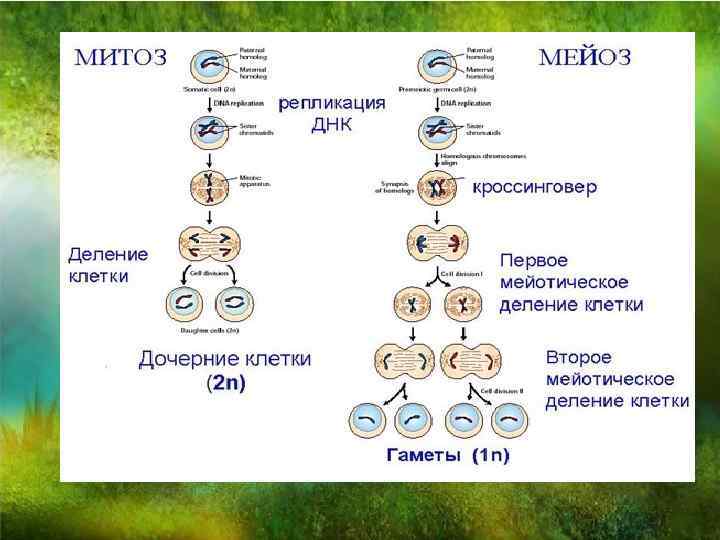 Напр., повторная сборка ядерной оболочки, деконденсация хроматина и возвращение микротрубочек в интерфазное состояние, вероятно, являются прямым следствием потери активности MPF и дефосфорилирования белков, которые были фосфорилированы MPF в начале митоза. Как обсуждается далее, инактивация MPF также запускает цитокинез.
Напр., повторная сборка ядерной оболочки, деконденсация хроматина и возвращение микротрубочек в интерфазное состояние, вероятно, являются прямым следствием потери активности MPF и дефосфорилирования белков, которые были фосфорилированы MPF в начале митоза. Как обсуждается далее, инактивация MPF также запускает цитокинез.
Цитокинез
Завершение митоза обычно сопровождается цитокинезом, в результате которого образуются две дочерние клетки. Цитокинез обычно инициируется в поздней анафазе и запускается инактивацией MPF, тем самым координируя ядерное и цитоплазматическое деление клетки. Как обсуждалось в главе 11, цитокинез животных клеток опосредован контрактильным кольцом актиновых и миозиновых филаментов II, которые формируются под плазматической мембраной (4). Расположение этого кольца определяется положением митотического веретена, поэтому клетка в конечном итоге расщепляется в плоскости, проходящей через метафазную пластинку перпендикулярно веретену. Расщепление происходит по мере того, как сокращение актин-миозиновых филаментов тянет плазматическую мембрану внутрь, в конечном итоге сжимая клетку пополам.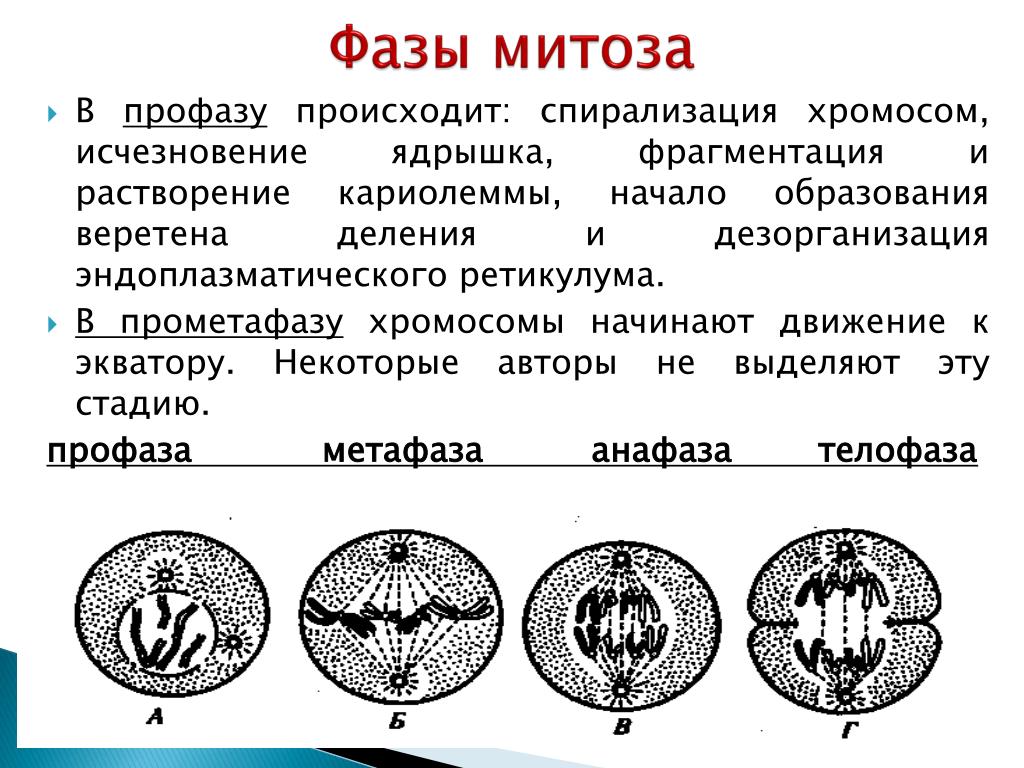
Рисунок 14.30
Цитокинез животных клеток. (A) Цитокинез возникает в результате сокращения кольца актиновых и миозиновых филаментов, которое сжимает клетку надвое. (B) Сканирующая электронная микрофотография яйца лягушки, подвергающегося цитокинезу. (Б, Дэвид М. Филлипс/Visuals Unlimited). (подробнее…)
Механизм цитокинеза иной для клеток высших растений, окруженных жесткими клеточными стенками. Эти клетки не сжимаются пополам сократительным кольцом, а делятся, образуя новые клеточные стенки и плазматические мембраны внутри клетки (4). В ранней телофазе везикулы, несущие предшественники клеточных стенок из аппарата Гольджи, связываются с микротрубочками веретена и накапливаются на прежнем месте метафазной пластинки. Затем эти везикулы сливаются, образуя большую, окруженную мембраной дискообразную структуру, а их полисахаридное содержимое собирается, образуя матрикс новой клеточной стенки (называемой ячеистая пластина ). Клеточная пластинка расширяется наружу, перпендикулярно веретену, пока не достигнет плазматической мембраны. Затем мембрана, окружающая клеточную пластинку, сливается с родительской плазматической мембраной, разделяя клетку на две части.
Затем мембрана, окружающая клеточную пластинку, сливается с родительской плазматической мембраной, разделяя клетку на две части.
Рисунок 14.31
Цитокинез высших растений. Везикулы Гольджи, несущие предшественники клеточной стенки, связываются с полярными микротрубочками на месте бывшей метафазной пластинки. Слияние этих везикул дает дискообразную структуру, окруженную мембраной (ранняя клеточная пластинка), которая (далее…)
Функция и стадии митоза
Определение, функция и время митоза
Митоз — тип деления, при котором образуются дочерние клетки с целью роста тканей, регенерации или бесполого (вегетативного) размножения. После митоза и цитокинеза дочерние клетки содержат ту же информацию о свойствах наследственности, что и материнская клетка: материнская и дочерняя клетки генетически идентичны .
Митоз следует за G2-фазой интерфазы в клеточном цикле. К тому времени ДНК уже продублирована и контролируется.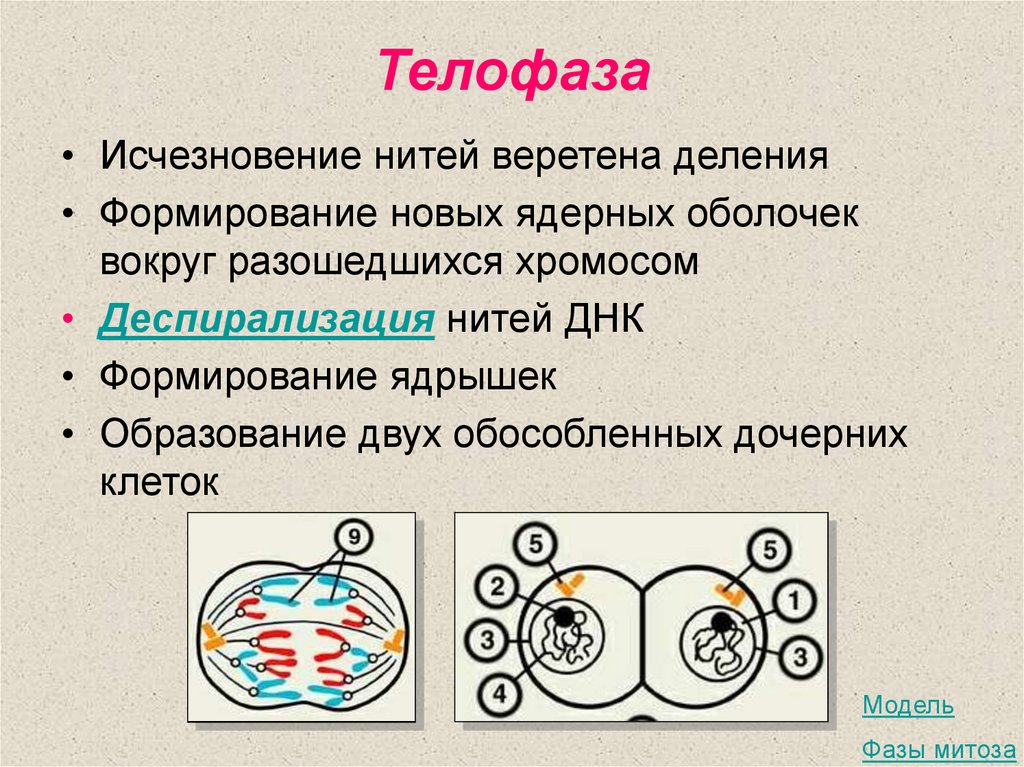 В начале митоза ядерная оболочка исчезает, и хромосомы сильно конденсируются, скручиваясь по спирали вокруг белковых молекул. Будучи компактными, хромосомы могут быть легче разделены по дочерним ядрам, чем если бы они были развернуты. Конденсированные одиночные хромосомы хорошо визуализируются под световым микроскопом.
В начале митоза ядерная оболочка исчезает, и хромосомы сильно конденсируются, скручиваясь по спирали вокруг белковых молекул. Будучи компактными, хромосомы могут быть легче разделены по дочерним ядрам, чем если бы они были развернуты. Конденсированные одиночные хромосомы хорошо визуализируются под световым микроскопом.
Схематично: для диплоидных материнских клеток (например, клеток листа)
или если материнская клетка была гаплоидной (например, слоевище у печеночников)
Интерфазные и митотические стадии
 Это первая стадия митоза. Хромосомы начинают спирализоваться, а ядерная оболочка и ядерные тельца (ядрышки = места в ядре, активные в синтезе рибосом) исчезают. Центросомы (состоящие каждая из пары центриолей — только в клетках животных), которые были продублированы во время G2-фазы, теперь разделяются. Все хромосомы становятся видимыми и теперь состоят из двух хроматид, соединенных центромерой. Фотографии ранней, средней и поздней стадии профазы.
Это первая стадия митоза. Хромосомы начинают спирализоваться, а ядерная оболочка и ядерные тельца (ядрышки = места в ядре, активные в синтезе рибосом) исчезают. Центросомы (состоящие каждая из пары центриолей — только в клетках животных), которые были продублированы во время G2-фазы, теперь разделяются. Все хромосомы становятся видимыми и теперь состоят из двух хроматид, соединенных центромерой. Фотографии ранней, средней и поздней стадии профазы.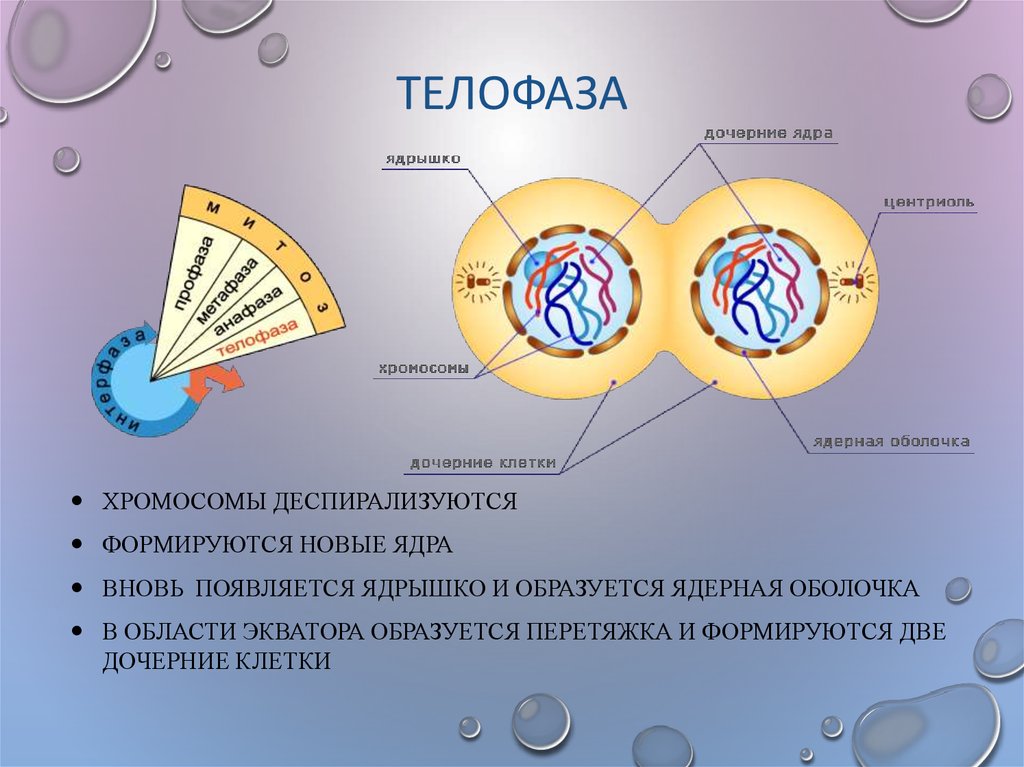 Две хроматиды каждой хромосомы раздвигаются микротрубочками, прикрепленными к кинетохоре (= специализированному участку центромеры) в направлении противоположных полюсов. С этого момента каждую отдельную хроматиду можно рассматривать как новую хромосому. (вид ранней анафазы и средней анафазы в Беллевалии)
Две хроматиды каждой хромосомы раздвигаются микротрубочками, прикрепленными к кинетохоре (= специализированному участку центромеры) в направлении противоположных полюсов. С этого момента каждую отдельную хроматиду можно рассматривать как новую хромосому. (вид ранней анафазы и средней анафазы в Беллевалии) У растений этот процесс характеризуется образованием и ростом клеточной пластинки (пример на Solanum sp.), который расширяется из пространства между двумя дочерними ядрами к периферии клетки. Иногда в прикреплении этой новой стенки участвуют остатки веретена (фрагмопласт).
У растений этот процесс характеризуется образованием и ростом клеточной пластинки (пример на Solanum sp.), который расширяется из пространства между двумя дочерними ядрами к периферии клетки. Иногда в прикреплении этой новой стенки участвуют остатки веретена (фрагмопласт).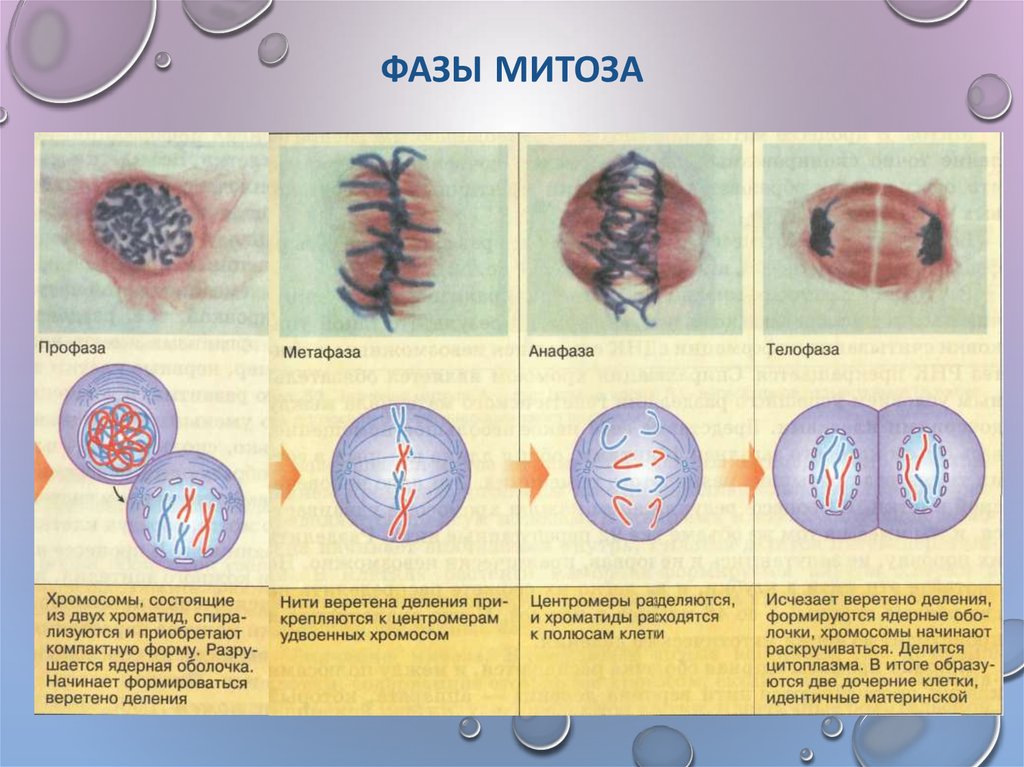
Leave A Comment