
4. Строение и функции РНК. Виды РНК
Молекулы РНК находятся в ядре, цитоплазме, рибосомах, митохондриях и пластидах клетки.
Рибонуклеиновая кислота (РНК) — линейный полимер, состоящий из одной цепочки нуклеотидов. Мономеры (нуклеотиды) РНК состоят из пятиуглеродного сахара — рибозы, остатка фосфорной кислоты и азотистого основания.
Три азотистых основания в молекулах РНК такие же, как и у ДНК — аденин, гуанин, цитозин, а четвертым является урацил.

Образование полимера РНК происходит (также как и у ДНК) через ковалентные связи между рибозой и остатком фосфорной кислоты соседних нуклеотидов.
Информационные, или матричные, РНК (иРНК) составляют около \(5\) % всей клеточной РНК. Они синтезируются в ядре (на участке одной из цепей молекулы ДНК) при участии фермента РНК-полимеразы.
Функция иРНК — снятие информации с ДНК и передача её к месту синтеза белка — на рибосомы.

Рибосомные (рибосомальные) РНК (рРНК) — синтезируются в ядрышке, входят в состав рибосом. Они участвуют в формировании активного центра рибосомы, где происходит процесс биосинтеза белка. рРНК составляют примерно \(85\) % всех РНК клетки.
Транспортные РНК (тРНК) — образуются в ядре на ДНК, затем переходят в цитоплазму. Они составляют около \(10\) % клеточной РНК и являются самыми небольшими по размеру (состоят из \(70\)–\(90\) нуклеотидов).
тРНК транспортируют аминокислоты к месту синтеза белка на рибосоме. Для переноса каждого вида аминокислот к рибосоме нужен отдельный вид тРНК.
Строение всех тРНК сходно. Их молекулы образуют своеобразные структуры, напоминающие по форме лист клевера.
Виды тРНК различаются по триплету нуклеотидов, расположенному «на верхушке». Этот триплет (антикодон) по генетическому коду комплементарен кодону иРНК, кодирующему соответствующую аминокислоту.
Аминокислота прикрепляется специальным ферментом к «черешку листа» и транспортируется в активный центр рибосомы.
Таким образом, различные виды РНК представляют собой единую функциональную систему, направленную на реализацию наследственной информации через синтез белка.
Источники:
Каменский А. А., Криксунов Е. А., Пасечник В. В. Биология. 9 класс // ДРОФА.
Каменский А. А., Криксунов Е. А., Пасечник В. В. Биология. Общая биология (базовый уровень) 10–11 класс // ДРОФА.
Лернер Г. И. Биология: Полный справочник для подготовки к ЕГЭ: АСТ, Астрель.
http://distant-lessons.ru/molekula-rnk.html
http://900igr.net/kartinki/biologija/Sintez-belkov/015-Protsess-uznavanija.html
http://900igr.net/kartinki/khimija/Stroenie-DNK-i-RNK/065-Obrazovanie-vtorichnoj-struktury-RNK.html
РИБОНУКЛЕИ́НОВЫЕ КИСЛО́ТЫ (РНК), класс нуклеиновых кислот. Так же как и дезоксирибонуклеиновые кислоты (ДНК) – биополимеры, в нуклеотидной последовательности которых может быть записана генетич. информация. В то же время они выполняют в клетке многие другие, характерные для белков функции: ферментативные (см. Рибозимы), регуляторные, транспортные, защитные и структурообразующие. В клетке и вирусных частицах РНК всегда связаны с белками, т. е. функционируют в виде рибонуклеопротеиновых комплексов.
РНК открыты в 1889 нем. гистологом Р. Альтманом в дрожжах. В последующие 60 лет установлена химич. структура РНК и доказано, что они присутствуют в цитоплазме любой живой клетки, выполняя ключевую роль в биосинтезе белков; РНК были обнаружены также в вирусах растений и животных.
Строение РНК
В отличие от ДНК, все клеточные и большинство вирусных РНК представляют собой линейные однотяжевые полирибонуклеотиды. Они построены из четырёх видов нуклеозидных остатков (н. о.) – аденозина (А), гуанозина (G), цитидина (С) и уридина (U), связанных друг с другом 3’–5′-фосфодиэфирными связями. Углеводные остатки в РНК представлены D-рибозой. Некоторые виды РНК содержат небольшое количество т. н. минорных нуклеотидов, в осн. псевдоуридиловую кислоту (ψ), и нуклеотидные звенья, метилированные по остаткам рибозы или гетероциклич. оснований. Длина полинуклеотидных цепей РНК колеблется от нескольких десятков до нескольких тысяч нуклеотидных остатков. Накоплена значит. информация о нуклеотидных последовательностях (н. п.), или первичной структуре, РНК; её получают либо прямым секвенированием индивидуальных РНК, либо анализируя транскриптомы, в которых представлены все РНК, синтезирующиеся в данный момент в клетке, либо выводят теоретически из н. п. ДНК, в которых РНК кодированы. Анализ массивов данных о н. п. РНК осуществляется методами биоинформатики.
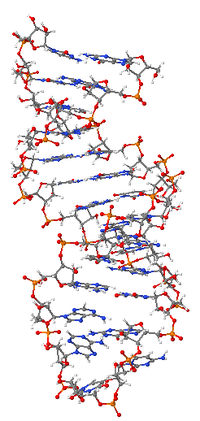
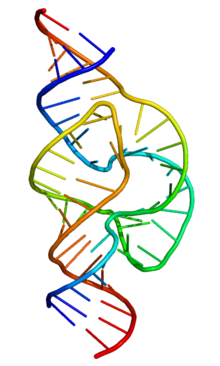
Принципы организации макромолекулярной структуры РНК сформулированы на рубеже 1950–60-х гг. Ж. Фреско и П. Доти (США) и А. С. Спириным. Было постулировано, что осн. элементом вторичной структуры РНК являются короткие, зачастую несовершенные (с «выпетливаниями») антипараллельные двойные спирали, образуемые за счёт комплементарных взаимодействий смежных участков полинуклеотидной цепи. Двуспиральные участки РНК формируются как за счёт стандартного уотсон-криковского спаривания оснований (G-C и A-U), так и за счёт образования сравнительно небольшого количества нестандартных пар (G-U, G-A, U-U, A-C). При компактном сворачивании цепи двуспиральные области в РНК могут быть образованы не только между соседними сегментами, но и между достаточно удалёнными областями полинуклеотидной цепи. С одного края двуспиральные участки замкнуты однотяжевыми сегментами разл. длины и т. о. имеют вид «шпильки»; соседние «шпильки» также связаны друг с другом однотяжевыми участками. Доказано, что такой способ организации вторичной структуры является универсальным для всех однотяжевых РНК.
Элементы вторичной структуры РНК взаимодействуют друг с другом, в результате чего формируется компактная уникальная третичная структура РНК. Третичные контакты в РНК создаются взаимодействием её однотяжевых сегментов друг с другом и с двуспиральными сегментами. Примером достаточно широко распространённого третичного контакта в РНК могут служить т. н. А-минорные взаимодействия, когда основания адениловых н. о. однотяжевых сегментов РНК встраиваются в желобки спиральных районов РНК, образуя там специфич. водородные связи. Третичная структура РНК стабилизируется белками и ионами магния. Ныне известны пространственные структуры разл. РНК как в свободном, так и в связанном с белками состоянии, полученные с атомным разрешением методами ядерного магнитного резонанса и рентгеноструктурного анализа.
Биосинтез РНК
Все клеточные РНК образуются в результате транскрипции генов, в которых они кодированы, с помощью ферментов РНК-полимераз. Процесс транскрипции протекает по принципу комплементарного копирования одной из цепей двуспиральной ДНК; т. о., РНК представляют собой полирибонуклеотидную копию одной из цепей ДНК и комплементарны другой её цепи. Процессы биосинтеза вирусных РНК весьма разнообразны, и для вируса иммунодефицита человека (ВИЧ), напр., включают стадию синтеза на РНК как на матрице однотяжевой ДНК (обратная транскрипция) с последующим превращением её в двуспиральную ДНК и интеграцией последней в геном клетки. В случае др. РНК-содержащих вирусов реализуется механизм РНК-репликации, когда на вирионной РНК синтезируется комплементарная цепь, а образующаяся двуспиральная РНК служит матрицей для последующего синтеза точной копии вирусной РНК.
Все клеточные РНК и РНК мн. вирусов синтезируются в виде предшественников, существенно превышающих по длине «зрелые» РНК. РНК-предшественник подвергается специфич. для каждого вида РНК и тонко регулируемому процессингу. Если гены, кодирующие данную РНК, содержат интроны, первичный транскрипт подвергается сплайсингу (в т. ч. альтернативному). Как правило, РНК-предшественник «разрезается» специфич. эндонуклеазами на фрагменты, которые укорачиваются соответствующими экзонуклеазами. Во многих случаях вслед за этим происходит модификация концов молекулы РНК, напр. кэпирование 5′-концов (присоединение остатка 7-метилгуанозина) эукариотич. матричных рибонуклеиновых кислот (мРНК) и полиаденилирование их 3′ -концов. Регуляция биосинтеза РНК происходит на всех стадиях транскрипции и процессинга с помощью спец. регуляторных белков. Кроме того, в состав самих РНК могут входить элементы, предназначенные для регуляции их синтеза. Нуклеотидная последовательность РНК может быть подвергнута редактированию. Так, в мРНК, синтезирующейся в митохондриях трипаносомы, происходит вставка большого числа уридиловых остатков, некодированных в ДНК, в результате чего её информац. содержание кардинально изменяется. Редактируются также транскрипты мн. генов человека и животных. В этом случае происходит замена (путём дезаминирования) определённых единичных остатков А в мРНК на остатки инозина (I), а также С на U.
Разнообразие РНК и их функций
РНК принято подразделять на кодирующие и некодирующие. Среди кодирующих РНК гл. место занимают мРНК. В соответствии с «центр. догмой» молекулярной биологии (ДНК – РНК – белок) они переносят генетич. информацию от ДНК к рибосомам, где она декодируется и реализуется в виде аминокислотных последовательностей белков. Кодирующей функцией обладает РНК-компонент фермента теломеразы (теломеразная РНК), определённый сегмент которой служит матрицей для синтеза теломерных ДНК-повторов на концах линейных хромосом. Со специфич. белками связана также короткая матричная РНК, называемая направляющей РНК (англ. guide RNA), участвующая в процессе редактирования мРНК. Она определяет, в какие участки мРНК будут введены дополнит. остатки U. У всех бактерий существует транспортно-матричная РНК (тмРНК), кодирующая короткий пептид, наращиваемый на С-концы дефектных белков, синтез которых по какой-то причине не смогли довести до конца рибосомы. Кодируемый тмРНК пептид служит сигналом для протеаз, уничтожающих дефектные белки. Т. о., в этом случае РНК осуществляет контроль качества белков, синтезируемых бактериальной клеткой. К кодирующим РНК относятся все вирусные РНК, которые служат матрицами для синтеза ДНК и РНК, а также вирусных белков.
К числу некодирующих РНК относится группа, формирующая белоксинтезирующий аппарат клетки. Более 80% РНК любой клетки представлено рибосомными рибонуклеиновыми кислотами (рРНК) – гл. структурными и функциональными компонентами рибосом.
Функцию декодирования генетич. информации на рибосомах под контролем рРНК осуществляют транспортные рибонуклеиновые кислоты (тРНК). Они переносят в рибосому аминокислотные остатки для синтеза белков. Когда рибосомы синтезируют секреторные или мембранные белки, с ними ассоциированы сигналузнающие РНК-белковые комплексы – т. н. SRP (от англ. signal recognition particles). В этих частицах РНК выполняют роль каркаса, на котором собираются белки, распознающие спец. сигнальные последовательности в синтезирующихся полипептидных цепях.
В регуляции процессов транскрипции и трансляции в клетке принимают участие многочисл. низкомолекулярные, или малые, и высокомолекулярные, или длинные, некодирующие РНК (мнкРНК и длнкРНК соответственно). Некоторые из них обладают рибозимной активностью и участвуют в процессинге др. РНК (напр., РНК-компонент бактериальной рибонуклеазы). В ядрах клеток эукариот присутствует семейство из 6–7 малых ядерных РНК (мяРНК), выполняющих гл. функцию на всех стадиях сплайсинга мРНК. Возможно, некоторые из них обладают рибозимной активностью. мнкРНК, локализующиеся в ядрышках эукариотич. клеток, отвечают за специфич. метилирование предшественников рРНК, за превращение в них определённых остатков уридина в псевдоуридин; они играют ключевую роль в РНК-интерференции. Многие из длнкРНК транскрибируются с цепи ДНК, противоположной кодирущей мРНК. Образуя комплементарные комплексы с мРНК вместе со специфич. белками, они полностью или частично подавляют экспрессию генов на уровне трансляции. Некодирующими РНК представлены транскрипты существенно большей части геномов всех организмов, чем кодирующими, т. е. разнообразие таких РНК в десятки раз превышает разнообразие индивидуальных белков в клетке.
Открытие у РНК столь высокой структурной и функциональной пластичности, в частности способности к самовоспроизведению и самопревращениям, позволяет предполагать, что при зарождении жизни на Земле эта нуклеиновая кислота была первым информац. биополимером, предшествовавшим появлению ДНК и белков.
Рибонуклеи́новая кисло́та (РНК) — одна из трёх основных макромолекул (две другие — ДНК и белки), которые содержатся в клетках всех живых организмов.
Так же, как ДНК (дезоксирибонуклеиновая кислота), РНК состоит из длинной цепи, в которой каждое звено называется нуклеотидом. Каждый нуклеотид состоит из азотистого основания, сахара рибозы и фосфатной группы. Последовательность нуклеотидов позволяет РНК кодировать генетическую информацию. Все клеточные организмы используют РНК (мРНК) для программирования синтеза белков.
Клеточные РНК образуются в ходе процесса, называемого транскрипцией, то есть синтеза РНК на матрице ДНК, осуществляемого специальными ферментами — РНК-полимеразами. Затем матричные РНК (мРНК) принимают участие в процессе, называемом трансляцией. Трансляция — это синтез белка на матрице мРНК при участии рибосом. Другие РНК после транскрипции подвергаются химическим модификациям, и после образования вторичной и третичной структур выполняют функции, зависящие от типа РНК.
Для одноцепочечных РНК характерны разнообразные пространственные структуры, в которых часть нуклеотидов одной и той же цепи спарены между собой. Некоторые высокоструктурированные РНК принимают участие в синтезе белка клетки, например, транспортные РНК служат для узнавания кодонов и доставки соответствующих аминокислот к месту синтеза белка, а рибосомные РНК служат структурной и каталитической основой рибосом.
Однако функции РНК в современных клетках не ограничиваются их ролью в трансляции. Так, малые ядерные РНК принимают участие в сплайсинге эукариотических матричных РНК и других процессах.
Помимо того, что молекулы РНК входят в состав некоторых ферментов (например, теломеразы), у отдельных РНК обнаружена собственная ферментативная активность: способность вносить разрывы в другие молекулы РНК или, наоборот, «склеивать» два РНК-фрагмента. Такие РНК называются рибозимами.
Геномы ряда вирусов состоят из РНК, то есть у них она играет роль, которую у высших организмов выполняет ДНК. На основании разнообразия функций РНК в клетке была выдвинута гипотеза, согласно которой РНК — первая молекула, которая была способна к самовоспроизведению в добиологических системах.
История изучения
Нуклеиновые кислоты были открыты в 1868 году швейцарским учёным Иоганном Фридрихом Мишером, который назвал эти вещества «нуклеин», поскольку они были обнаружены в ядре (лат. nucleus)[1]. Позже было обнаружено, что бактериальные клетки, в которых нет ядра, тоже содержат нуклеиновые кислоты. Значение РНК в синтезе белков было предположено в 1939 году в работе Торбьёрна Оскара Касперссона, Жана Брачета и Джека Шульца.[2] Джерард Маирбакс выделил первую матричную РНК, кодирующую гемоглобин кролика и показал, что при её введении в ооциты образуется тот же самый белок.[3] В 1956—1957 годах проводились работы (А. Белозёрский, А. Спирин, Э. Волкин, Л. Астрахан) по определению состава РНК клеток, которые привели к выводу, что основную массу РНК в клетке составляет рибосомальная РНК.[4] Северо Очоа получил Нобелевскую премию по медицине в 1959 году за открытие механизма синтеза РНК.[5] Последовательность 77 нуклеотидов одной из тРНК дрожжей S. cerevisiae была определена в 1965 году в лаборатории Роберта Холея, за что в 1968 году он получил Нобелевскую премию по медицине.[6] В 1967 Карл Вёзе предположил, что РНК обладают каталитическими свойствами. Он выдвинул так называемую Гипотезу РНК-мира, в котором РНК прото-организмов служила и в качестве молекулы хранения информации (сейчас эта роль выполняется в основном ДНК) и молекулы, которая катализировала метаболические реакции (сейчас это делают в основном ферменты).[7] В 1976 Уолтер Фаэрс и его группа в Гентском Университете (Голландия) определили первую последовательность генома РНК-содержащего вируса, бактериофага MS2.[8] В начале 1990-х было обнаружено, что введение чужеродных генов в геном растений приводит к подавлению выражения аналогичных генов растения.[9] Приблизительно в это же время было показано, что РНК длиной около 22 оснований, которые сейчас называются микроРНК, играют регуляторную роль в онтогенезе нематод C. elegans.[10]
Химический состав и модификации мономеров
Химическое строение полинуклеотида РНКНуклеотиды РНК состоят из сахара — рибозы, к которой в положении 1′ присоединено одно из оснований: аденин, гуанин, цитозин или урацил. Фосфатная группа соединяет рибозы в цепочку, образуя связи с 3′ атомом углерода одной рибозы и в 5′ положении другой. Фосфатные группы при физиологическом рН отрицательно заряжены, поэтому РНК — полианион. РНК транскрибируется как полимер четырёх оснований (аденина (A), гуанина (G), урацила (U) и цитозина (C), но в «зрелой» РНК есть много модифицированных оснований и сахаров[11]. Всего в РНК насчитывается около 100 разных видов модифицированных нуклеотидов, из которых 2′-О-метилрибоза наиболее частая модификация сахара, а псевдоуридин — наиболее часто встречающееся модифицированное основание[12].
У псевдоуридина (Ψ) связь между урацилом и рибозой не C — N, а C — C, этот нуклеотид встречается в разных положениях в молекулах РНК. В частности, псевдоуридин важен для функционирования тРНК[13]. Другое заслуживающее внимания модифицированное основание — гипоксантин, деаминированный гуанин, нуклеозид которого носит название инозина. Инозин играет важную роль в обеспечении вырожденности генетического кода.
Роль многих других модификаций не до конца изучена, но в рибосомальной РНК многие пост-транскрипционные модификации находятся в важных для функционирования рибосомы участках. Например, на одном из рибонуклеотидов, участвующем в образовании пептидной связи[14].
Структура
Азотистые основания в составе РНК могут образовывать водородные связи между цитозином и гуанином, аденином и урацилом, а также между гуанином и урацилом[15]. Однако возможны и другие взаимодействия, например, несколько аденинов могут образовывать петлю, или петля, состоящая из четырёх нуклеотидов, в которой есть пара оснований аденин — гуанин[16].
Разные формы нуклеиновых кислот. На рисунке (слева направо) представлены A (типична для РНК), B (ДНК) и Z (редкая форма ДНК)Важная структурная особенность РНК, отличающая её от ДНК — наличие гидроксильной группы в 2′ положении рибозы, которая позволяет молекуле РНК существовать в А, а не В-конформации, наиболее часто наблюдаемой у ДНК[17]. У А-формы глубокая и узкая большая бороздка и неглубокая и широкая малая бороздка[18]. Второе последствие наличия 2′ гидроксильной группы состоит в том, что конформационно пластичные, то есть не принимающие участие в образовании двойной спирали, участки молекулы РНК могут химически атаковать другие фосфатные связи и их расщеплять[19].
«Рабочая» форма одноцепочечной молекулы РНК, как и у белков, часто обладает третичной структурой. Третичная структура образуется на основе элементов вторичной структуры, образуемой с помощью водородных связей внутри одной молекулы. Различают несколько типов элементов вторичной структуры — стебель-петли, петли и псевдоузлы[20]. В силу большого числа возможных вариантов спаривания оснований предсказание вторичной структуры РНК — гораздо более сложная задача, чем предсказание вторичной структуры белков, но в настоящее время есть эффективные программы, например, mfold[21].
Примером зависимости функции молекул РНК от их вторичной структуры являются участки внутренней посадки рибосомы (IRES). IRES — структура на 5′ конце информационной РНК, которая обеспечивает присоединение рибосомы в обход обычного механизма инициации синтеза белка, требующего наличия особого модифицированного основания (кэпа) на 5′ конце и белковых факторов инициации. Первоначально IRES были обнаружены в вирусных РНК, но сейчас накапливается всё больше данных о том, что клеточные мРНК также используют IRES-зависимый механизм инициации в условиях стресса[22].
Многие типы РНК, например, рРНК и мяРНК в клетке функционируют в виде комплексов с белками, которые ассоциируют с молекулами РНК после их синтеза или (у эукариот) экспорта из ядра в цитоплазму. Такие РНК-белковые комплексы называются рибонуклеопротеиновыми комплексами или рибонуклеопротеидами.
Сравнение с ДНК
Между ДНК и РНК есть три основных отличия:
- ДНК содержит сахар дезоксирибозу, РНК — рибозу, у которой есть дополнительная, по сравнению с дезоксирибозой, гидроксильная группа. Эта группа увеличивает вероятность гидролиза молекулы, то есть уменьшает стабильность молекулы РНК.
- Нуклеотид, комплементарный аденину, в РНК не тимин, как в ДНК, а урацил — неметилированная форма тимина.
- ДНК существует в форме двойной спирали, состоящей из двух отдельных молекул. Молекулы РНК, в среднем, гораздо короче и преимущественно одноцепочечные.
Структурный анализ биологически активных молекул РНК, включая тРНК, рРНК, мяРНК и другие молекулы, которые не кодируют белков, показал, что они состоят не из одной длинной спирали, а из многочисленных коротких спиралей, расположенных близко друг к другу и образующих нечто, похожее на третичную структуру белка. В результате этого РНК может катализировать химические реакции, например, пептидил-трансферазный центр рибосомы, участвующий в образовании пептидной связи белков, полностью состоит из РНК[23][24].
Синтез
Синтез РНК в живой клетке проводится ферментом — РНК-полимеразой. У эукариот разные типы РНК синтезируются разными, специализированными РНК-полимеразами. В целом матрицей синтеза РНК может выступать как ДНК, так и другая молекула РНК. Например, полиовирусы используют РНК-зависимую РНК-полимеразу для репликации своего генетического материала, состоящего из РНК[25]. Но РНК-зависимый синтез РНК, который раньше считался характерным только для вирусов, происходит и в клеточных организмах, в процессе так называемой РНК-интерференции[26].
Как в случае ДНК-зависимой РНК-полимеразы, так и в случае РНК-зависимой РНК-полимеразы фермент присоединяется к промоторной последовательности. Вторичная структура молекулы матрицы расплетается с помощью хеликазной активности полимеразы, которая при движении субстрата в направлении от 3′ к 5′ концу молекулы синтезирует РНК в направлении 5′ → 3′. Терминатор транскрипции в исходной молекуле определяет окончание синтеза. Многие молекулы РНК синтезируются в качестве молекул-предшественников, которые подвергаются «редактированию» — удалению ненужных частей с помощью РНК-белковых комплексов[27].
Например, у кишечной палочки гены рРНК расположены в составе одного оперона (в rrnB порядок расположения такой: 16S — tRNAGlu 2 — 23S —5S) считываются в виде одной длинной молекулы, которая затем подвергается расщеплению в нескольких участках с образованием сначала пре-рРНК, а затем зрелых молекул рРНК[28]. Процесс изменения нуклеотидной последовательности РНК после синтеза носит название процессинга или редактирования РНК.
После завершения транскрипции РНК часто подвергается модификациям (см. выше), которые зависят от функции, выполняемой данной молекулой. У эукариот процесс «созревания» РНК, то есть её подготовки к синтезу белка, часто включает сплайсинг: удаление некодирующих белок последовательностей (интронов) с помощью рибонуклеопротеида сплайсосомы. Затем к 5′ концу молекулы пре-мРНК эукариот добавляется особый модифицированный нуклеотид (кэп), а к 3′ концу несколько аденинов, так назваемый «полиА-хвост»[27].
Типы РНК
 Структура молоточкового (hammerhead) рибозима, который расщепляет РНК
Структура молоточкового (hammerhead) рибозима, который расщепляет РНКМатричная (информационная) РНК — РНК, которая служит посредником при передаче информации, закодированной в ДНК к рибосомам, молекулярным машинам, синтезирующим белки живого организма. Кодирующая последовательность мРНК определяет последовательность аминокислот полипептидной цепи белка[29]. Однако подавляющее большинство РНК не кодируют белок. Эти некодирующие РНК могут транскрибироваться с отдельных генов (например, рибосомальные РНК) или быть производными интронов[30]. Классические, хорошо изученные типы некодирующих РНК — это транспортные РНК (тРНК) и рРНК, которые участвуют в процессе трансляции[31]. Существуют также классы РНК, ответственные за регуляцию генов, процессинг мРНК и другие роли. Кроме того, есть и молекулы некодирующих РНК, способные катализировать химические реакции, такие, как разрезание и лигирование молекул РНК[32]. По аналогии с белками, способными катализировать химические реакции — энзимами (ферментами), каталитические молекулы РНК называются рибозимами.
Участвующие в трансляции
Роль разных типов РНК в синтезе белка (по Уотсону)Информация о последовательности аминокислот белка содержится в мРНК. Три последовательных нуклеотида (кодон) соответствуют одной аминокислоте. В эукариотических клетках транскирибированный предшественник мРНК или пре-мРНК процессируется с образованием зрелой мРНК. Процессинг включает удаление некодирующих белок последовательностей (интронов). После этого мРНК экспортируется из ядра в цитоплазму, где к ней присоединяются рибосомы, транслирующие мРНК с помощью соединённых с аминокислотами тРНК.
В безъядерных клетках (бактерии и археи) рибосомы могут присоединяться к мРНК сразу после транскрипции участка РНК. И у эукариот, и у прокариот цикл жизни мРНК завершается её контролируемым разрушением ферментами рибонуклеазами[29].
Транспортные (тРНК) — малые, состоящие из приблизительно 80 нуклеотидов, молекулы с консервативной третичной структурой. Они переносят специфические аминокислоты в место синтеза пептидной связи в рибосоме. Каждая тРНК содержит участок для присоединения аминокислоты и антикодон для узнавания и присоединения к кодонам мРНК. Антикодон образует водородные связи с кодоном, что помещает тРНК в положение, способствующее образованию пептидной связи между последней аминокислотой образованного пептида и аминокислотой, присоединённой к тРНК[30].
Рибосомальные РНК (рРНК) — каталитическая составляющая рибосом. Эукариотические рибосомы содержат четыре типа молекул рРНК: 18S, 5.8S, 28S и 5S. Три из четырёх типов рРНК синтезируются в ядрышке. В цитоплазме рибосомальные РНК соединяются с рибосомальными белками и формируют нуклеопротеин, называемый рибосомой[29]. Рибосома присоединяется к мРНК и синтезирует белок. рРНК составляет до 80 % РНК, обнаруживаемой в цитоплазме эукариотической клетки[33].
Необычный тип РНК, который действует в качестве тРНК и мРНК (тмРНК) обнаружен во многих бактериях и пластидах. При остановке рибосомы на дефектных мРНК без стоп-кодонов тмРНК присоединяет небольшой пептид, направляющий белок на деградацию[34].
Участвующие в регуляции генов
В живых клетках обнаружено несколько типов РНК, которые могут уменьшать степень выражения гена при комплементарности мРНК или самому гену. Микро-РНК (21-22 нуклеотида в длину) найдены у эукариот и оказывают воздействие через механизм РНК-интерференции. При этом комплекс микро-РНК и ферментов может приводить к метилированию нуклеотидов в ДНК промотора гена, что служит сигналом для уменьшения активности гена. При использовании другого типа регуляции мРНК, комплементарная микро-РНК, деградируется[35]. Однако есть и миРНК, которые увеличивают, а не уменьшают экспрессию генов[36]. Малые интерферирующие РНК (миРНК, 20-25 нуклеотидов) часто образуются в результате расщепления вирусных РНК, но существуют и эндогенные клеточные миРНК[37]. Малые интерферирующие РНК также действуют через РНК-интерференцию по сходным с микро-РНК механизмам[38]. У животных найдены так называемыме РНК, взаимодействующие с Piwi (piRNA, 29-30 нуклеотидов), действующие в половых клетках против транспозиции и играющие роль в образовании гамет[39][40]. Кроме того, piRNA могут эпигенетически наследоваться по материнской линии, передавая потомству своё свойство ингибировать экспрессию транспозонов[41].
Антисмысловые РНК широко распространены у бактерий, многие из них подавляют выражение генов, но некоторые активируют экспрессию[42]. Действуют антисмысловые РНК, присоединяясь к мРНК, что приводит к образованию двуцепочечных молекул РНК, которые деградируются ферментами.[43]. У эукариот обнаружены высокомолекулярные, мРНК-подобные молекулы РНК. Эти молекулы также регулируют выражение генов[44]. В качестве примера можно привести Xist, присоединяющуюся и инактивирующую одну из двух Х-хромосом у самок млекопитающих[45].
Кроме роли отдельных молекул в регуляции генов, регуляторные элементы могут формироваться в 5′ и 3′ нетранслируемых участках мРНК. Эти элементы могут действовать самостоятельно, предотвращая инициацию трансляции, либо присоединять белки, например, ферритин или малые молекулы, например, биотин[46].
В процессинге РНК
Многие РНК принимают участие в модификации других РНК. Интроны вырезаются из пре-мРНК сплайсосомами, которые, кроме белков, содержат несколько малых ядерных РНК (мяРНК)[31]. Кроме того, интроны могут катализировать собственное вырезание[47]. Синтезированая в результате транскрипции РНК также может быть химически модифицирована. У эукариот химические модификации нуклеотидов РНК, например, их метилирование, выполняется малыми ядерными РНК (мяРНК, 60-300 нуклеотидов). Этот тип РНК локализуется в ядрышко и тельцах Кахаля[30]. После ассоциации мяРНК с ферментами, мяРНК связываются с РНК-мишенью путём образования пар между основаниями двух молекул, а ферменты модифицируют нуклеотиды РНК-мишени. Рибосомальные и транспортные РНК содержат много подобных модификаций, конкретное положение которых часто сохраняется в процессе эволюции. Также могут быть модифицированы мяРНК и сами мяРНК[48][49]. Гидовые РНК осуществляют процесс редактирования РНК в кинетопласте — особом участке митохондрии протистов-кинетопластид (например, трипаносом).
Геномы, состоящие из РНК
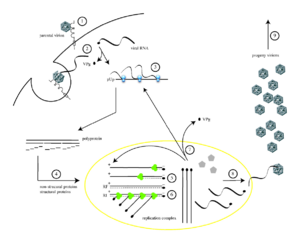 Жизненный цикл вируса с РНК геномом на примере полиовируса: 1 — присоединение исходного вириона к рецептору; 2 — вирион попадает в клетку; 3 — трансляция белков вируса с его РНК с образованием полипетида; 4 — полимеразы вируса размножают его РНК
Жизненный цикл вируса с РНК геномом на примере полиовируса: 1 — присоединение исходного вириона к рецептору; 2 — вирион попадает в клетку; 3 — трансляция белков вируса с его РНК с образованием полипетида; 4 — полимеразы вируса размножают его РНККак и ДНК, РНК может хранить информацию о биологических процессах. РНК может использоваться в качестве генома вирусов и вирусоподобных частиц. РНК-геномы можно разделить на те, которые не имеют промежуточной стадии ДНК и те, которые для размножения копируются в ДНК-копию и обратно в РНК (ретровирусы).
РНК-содержащие вирусы
Многие вирусы, например, вирус гриппа, на всех стадиях содержат геном, состоящий исключительно из РНК. РНК содержится внутри обычно белковой оболочки и реплицируется с помощью закодированных в ней РНК-зависимых РНК-полимераз. Вирусные геномы, состоящие из РНК разделяются на
- содержащие «плюс-цепь РНК», которая используется в качестве и мРНК, и генома;
- «минус-цепь РНК», которая служит только геномом, а в качестве мРНК используется комплементарная ей молекула;
- двухцепоченые вирусы.
Вироиды — другая группа патогенов, содержащих РНК-геном и не содержащих белок. Они реплицируются РНК-полимеразами организма хозяина[50].
Ретровирусы и ретротранспозоны
У других вирусов РНК-геном есть в течение только одной из фаз жизненного цикла. Вирионы так называемых ретровирусов содержат молекулы РНК, которые при попадании в клетки хозяина служат матрицей для синтеза ДНК-копии. В свою очередь, с матрицы ДНК считывается РНК-геном. Кроме вирусов обратную транскрипции применяют и класс мобильных элементов генома — ретротранспозоны[51].
Гипотеза РНК-мира
Способность молекул РНК одновременно служить как в качестве носителя информации, так и в качестве катализатора химических реакций, позволила выдвинуть гипотезу о том, что РНК была первым сложным полимером, появившимся в процессе добиологической эволюции. Эта гипотеза названа «гипотеза РНК-мира»[52][53]. Согласно ей, РНК на первых этапах эволюции автокатализировала синтез других молекул РНК, а затем и ДНК. На втором этапе эволюции синтезированные молекулы ДНК, как более стабильные, стали хранилищем генетической информации. Синтез белка на матрице РНК с помощью пра-рибосом, полностью состоящих из РНК, расширил свойства добиологических систем, постепенно белок заменил РНК в структурных аспектах. Из этой гипотезы делается вывод, что многие РНК, принимающие участие в синтезе белка в современных клетках, в особенности рРНК и тРНК — это реликты РНК-мира.
См. также
Примечания
- ↑ Dahm R (2005). «Friedrich Miescher and the discovery of DNA». Developmental Biology 278 (2): 274–88. PMID 15680349.
- ↑ Nierhaus KH, Wilson DN. Protein Synthesis and Ribosome Structure. — Wiley-VCH, 2004. — С. 3. — ISBN 3-527-30638-2
- ↑ Carlier M L’ADN, cette «simple» molécule. Esprit libre (июнь 2003). Архивировано из первоисточника 23 августа 2011. Проверено ???.
- ↑ А. С. Спирин. Биоорганическая химия. — М.: Высшая школа, 1986. — С. 10.
- ↑ Ochoa S. Enzymatic synthesis of ribonucleic acid. Nobel Lecture (1959). Архивировано из первоисточника 23 августа 2011. Проверено ???.
- ↑ Holley RW et al. Structure of a ribonucleic acid // Science. — 1965. — Vol. 147. — № 1664. — P. 1462–65. — DOI:10.1126/science.147.3664.1462
- ↑ Szathmáry E. The origin of the genetic code: amino acids as cofactors in an RNA world // Trends Genet.. — 1999. — Vol. 15. — № 6. — P. 223–9. — DOI:10.1016/S0168-9525(99)01730-8
- ↑ Fiers W et al. Complete nucleotide-sequence of bacteriophage MS2-RNA: primary and secondary structure of replicase gene // Nature. — 1976. — Vol. 260. — P. 500–7. — PMID 1264203.
- ↑ Napoli C, Lemieux C, Jorgensen R. Introduction of a chimeric chalcone synthase gene into petunia results in reversible co-suppression of homologous genes in trans. // Plant Cell. — 1990. — Vol. 2. — № 4. — P. 279–89. — PMID 12354959.
- ↑ Ruvkun G. Glimpses of a tiny RNA world // Science. — 2001. — Vol. 294. — № 5543. — P. 797–99. — DOI:10.1126/science.1066315
- ↑ Jankowski JAZ, Polak JM Clinical gene analysis and manipulation: tools, techniques and troubleshooting. — Cambridge University Press. — P. 14. — ISBN 0521478960
- ↑ Kiss T (2001). «Small nucleolar RNA-guided post-transcriptional modification of cellular RNAs». The EMBO Journal 20: 3617–22. DOI:10.1093/emboj/20.14.3617.
- ↑ Yu Q, Morrow CD (2001). «Identification of critical elements in the tRNA acceptor stem and TΨC loop necessary for human immunodeficiency virus type 1 infectivity». J Virol. 75 (10): 4902–6. DOI:10.1128/JVI.75.10.4902-4906.2001.
- ↑ King TH, Liu B, McCully RR, Fournier MJ (2002). «Ribosome structure and activity are altered in cells lacking snoRNPs that form pseudouridines in the peptidyl transferase center». Molecular Cell 11 (2): 425–35. DOI:10.1016/S1097-2765(03)00040-6.
- ↑ Barciszewski J, Frederic B, Clark C RNA biochemistry and biotechnology. — Springer. — P. 73–87. — ISBN 0792358627
- ↑ Lee JC, Gutell RR (2004). «Diversity of base-pair conformations and their occurrence in rRNA structure and RNA structural motifs». J. Mol. Biol. 344 (5): 1225–49. DOI:10.1016/j.jmb.2004.09.072. PMID 15561141.
- ↑ Salazar M, Fedoroff OY, Miller JM, Ribeiro NS, Reid BR (1992). «The DNA strand in DNAoRNA hybrid duplexes is neither B-form nor A-form in solution». Biochemistry 1993 (32): 4207–15. PMID 7682844.
- ↑ Hermann T, Patel DJ (2000). «RNA bulges as architectural and recognition motifs». Structure 8 (3): R47–R54. DOI:10.1016/S0969-2126(00)00110-6.
- ↑ Mikkola S, Nurmi K, Yousefi-Salakdeh E, Strömberg R, Lönnberg H (1999). «The mechanism of the metal ion promoted cleavage of RNA phosphodiester bonds involves a general acid catalysis by the metal aquo ion on the departure of the leaving group». Perkin transactions 2: 1619–26. DOI:10.1039/a903691a.
- ↑ Mathews DH, Disney MD, Childs JL, Schroeder SJ, Zuker M, Turner DH (2004). «Incorporating chemical modification constraints into a dynamic programming algorithm for prediction of RNA secondary structure». Proc. Natl. Acad. Sci. USA 101 (19): 7287–92. DOI:10.1073/pnas.0401799101.
- ↑ Redirect
- ↑ Spriggs KA, Stoneley M, Bushell M, Willis AE. (2008). «Re-programming of translation following cell stress allows IRES-mediated translation to predominate». Biol Cell. 100 (1): 27–38.
- ↑ Higgs PG (2000). «RNA secondary structure: physical and computational aspects». Quarterly Reviews of Biophysics 33: 199–253. DOI:10.1017/S0033583500003620.
- ↑ Nissen P, Hansen J, Ban N, Moore PB, Steitz TA (2000). «The structural basis of ribosome activity in peptide bond synthesis». Science 289 (5481): 920–30. DOI:10.1126/science.289.5481.920.
- ↑ Jeffrey L Hansen, Alexander M Long, Steve C Schultz (1997). «Structure of the RNA-dependent RNA polymerase of poliovirus». Structure 5 (8): 1109–22. DOI:10.1016/S0969-2126(97)00261-X.
- ↑ Ahlquist P (2002). «RNA-Dependent RNA Polymerases, Viruses, and RNA Silencing». Science 296 (5571): 1270–73. DOI:10.1126/science.1069132.
- ↑ 1 2 Alberts Bruce Molecular Biology of the Cell; Fourth Edition. — New York and London: Garland Science. — P. 302–303. — ISBN ISBN 0-8153-3218-1
- ↑ Wagner R., Theissen G., Zacharias Regulation of Ribosomal RNA synthesis and Control of ribosome Formation in E.coli. — 1993. — P. 119–129.
- ↑ 1 2 3 Cooper GC, Hausman RE The Cell: A Molecular Approach. — 3rd edition. — Sinauer. — P. 261–76, 297, 339–44. — ISBN 0-87893-214-3
- ↑ 1 2 3 Wirta W Mining the transcriptome – methods and applications. — ISBN 91-7178-436-5
- ↑ 1 2 Berg JM, Tymoczko JL, Stryer L Biochemistry. — 5th edition. — WH Freeman and Company, 2002. — P. 118–19, 781–808. — ISBN 0-7167-4684-0
- ↑ Rossi JJ (2004). «Ribozyme diagnostics comes of age». Chemistry & Biology 11 (7): 894–95. DOI:10.1016/j.chembiol.2004.07.002.
- ↑ Kampers T, Friedhoff P, Biernat J, Mandelkow E-M, Mandelkow E (1996). «RNA stimulates aggregation of microtubule-associated protein tau into Alzheimer-like paired helical filaments». FEBS Letters 399: 98–100, 344–49. PMID 8985176.
- ↑ Gueneau de Novoa P, Williams KP (2004). «The tmRNA website: reductive evolution of tmRNA in plastids and other endosymbionts». Nucleic Acids Res. 32 (Database issue): D104-8. DOI:10.1093/nar/gkh202. PMID 14681369.
- ↑ Matzke MA, Matzke AJM (2004). «Planting the seeds of a new paradigm». PLoS Biology 2 (5): e133. DOI:10.1371/journal.pbio.0020133. PMID 15138502.
- ↑ Check E (2007). «RNA interference: hitting the on switch». Nature 448 (7156): 855–58. DOI:10.1038/448855a. PMID 17713502.
- ↑ Vazquez F, Vaucheret H, Rajagopalan R, Lepers C, Gasciolli V, Mallory AC, Hilbert J, Bartel DP, Crété P (2004). «Endogenous trans-acting siRNAs regulate the accumulation of Arabidopsis mRNAs». Molecular Cell 16 (1): 69–79. DOI:10.1016/j.molcel.2004.09.028. PMID 15469823.
- ↑ Doran G (2007). «RNAi – Is one suffix sufficient?». Journal of RNAi and Gene Silencing 3 (1): 217–19.
- ↑ name=fruitfly_piRNA>Horwich MD, Li C Matranga C, Vagin V, Farley G, Wang P, Zamore PD (2007). «The Drosophila RNA methyltransferase, DmHen1, modifies germline piRNAs and single-stranded siRNAs in RISC». Current Biology 17: 1265–72. DOI:10.1016/j.cub.2007.06.030. PMID 17604629.
- ↑ Girard A, Sachidanandam R, Hannon GJ, Carmell MA (2006). «A germline-specific class of small RNAs binds mammalian Piwi proteins». Nature 442: 199–202. DOI:10.1038/nature04917. PMID 16751776.
- ↑ Brennecke J, Malone CD, Aravin AA, Sachidanandam R, Stark A, Hannon GJ (November 2008). «An epigenetic role for maternally inherited piRNAs in transposon silencing». Science (journal) 322 (5906): 1387–92. DOI:10.1126/science.1165171. PMID 19039138.
- ↑ Wagner EG, Altuvia S, Romby P (2002). «Antisense RNAs in bacteria and their genetic elements». Adv Genet. 46: 361–98. PMID 11931231.
- ↑ Gilbert SF Developmental Biology. — 7th ed. — Sinauer, 2003. — P. 101–3. — ISBN 0878932585
- ↑ Hüttenhofer A, Schattner P, Polacek N (2005). «Non-coding RNAs: hope or hype?». Trends Genet. 21 (5): 289–97. DOI:10.1016/j.tig.2005.03.007. PMID 15851066.
- ↑ Heard E, Mongelard F, Arnaud D, Chureau C, Vourc’h C, Avner P (1999). «Human XIST yeast artificial chromosome transgenes show partial X inactivation center function in mouse embryonic stem cells». Proc. Natl. Acad. Sci. USA 96 (12): 6841–46. DOI:10.1073/pnas.96.12.6841. PMID 10359800.
- ↑ Batey RT (2006). «Structures of regulatory elements in mRNAs». Curr. Opin. Struct. Biol. 16 (3): 299–306. DOI:10.1016/j.sbi.2006.05.001. PMID 16707260.
- ↑ Steitz TA, Steitz JA (1993). «A general two-metal-ion mechanism for catalytic RNA». Proc. Natl. Acad. Sci. U.S.A. 90 (14): 6498–502. DOI:10.1073/pnas.90.14.6498. PMID 8341661.
- ↑ Covello PS, Gray MW (1989). «RNA editing in plant mitochondria». Nature 341: 662–66. DOI:10.1038/341662a0. PMID 2552326.
- ↑ Omer AD, Ziesche S, Decatur WA, Fournier MJ, Dennis PP (2003). «RNA-modifying machines in archaea». Molecular Microbiology 48 (3): 617–29. DOI:10.1046/j.1365-2958.2003.03483.x. PMID 12694609.
- ↑ Daròs JA, Elena SF, Flores R (2006). «Viroids: an Ariadne’s thread into the RNA labyrinth». EMBO Rep. 7 (6): 593–8. DOI:10.1038/sj.embor.7400706. PMID 16741503.
- ↑ Kalendar R, Vicient CM, Peleg O, Anamthawat-Jonsson K, Bolshoy A, Schulman AH (2004). «Large retrotransposon derivatives: abundant, conserved but nonautonomous retroelements of barley and related genomes». Genetics 166 (3): D339. DOI:10.1534/genetics.166.3.1437. PMID 15082561.
- ↑ Gilbert, Walter (Feb 1986). «The RNA World». Nature 319: 618. DOI:10.1038/319618a0.
- ↑ Woese Carl The Genetic Code. — Harper & Row, 1968. — ISBN 978-0060471767
Литература
Ссылки
Рибонуклеи́новые кисло́ты (РНК) — нуклеиновые кислоты, полимеры нуклеотидов, в состав которых входят остаток ортофосфорной кислоты, рибоза (в отличие от ДНК, содержащей дезоксирибозу) и азотистые основания — аденин, цитозин, гуанин и урацил (в отличие от ДНК, содержащей вместо урацила тимин). Эти молекулы содержатся в клетках всех живых организмов, а также в некоторых вирусах.
Клеточные РНК образуются в ходе процесса, называемого транскрипцией, то есть синтеза РНК на матрице ДНК, осуществляемого специальными ферментами — РНК-полимеразами. Затем матричные РНК (мРНК) подвергаются сплайсингу и принимают участие в процессе, называемом трансляцией. Трансляция — это синтез белка на матрице мРНК при участии рибосом. Другие РНК после транскрипции подвергаются химическим модификациям, и после образования вторичной и третичной структур выполняют функции, зависящие от типа РНК.
Для одноцепочечных РНК характерны разнообразные пространственные структуры, в которых часть нуклеотидов одной и той же цепи спарены между собой. Некоторые высокоструктурированные РНК принимают участие в синтезе белка клетки, например, транспортные РНК служат для узнавания кодонов и доставки соответствующих аминокислот к месту синтеза белка, а рибосомные РНК служат структурной и каталитической основой рибосом.
Однако функции РНК в современных клетках не ограничиваются их ролью в трансляции. Так малые ядерные РНК принимают участие в сплайсинге эукариотических матричных РНК и других процессах.
Помимо того, что молекулы РНК входят в состав некоторых ферментов (например, теломеразы) у отдельных РНК обнаружена собственная энзиматическая активность, способность вносить разрывы в другие молекулы РНК или, наоборот, «склеивать» два РНК-фрагмента. Такие РНК называются рибозимами.
Геномы ряда вирусов состоят из РНК, то есть у них она играет роль, которую у высших организмов выполняет ДНК. На основании разнообразия функций РНК в клетке была выдвинута гипотеза, согласно которой РНК — первая молекула, которая была способна к самовоспроизведению в добиологических системах.
История изучения
Нуклеиновые кислоты были открыты в 1868 году швейцарским учёным Иоганном Фридрихом Мишером, который назвал эти вещества «нуклеин», поскольку они были обнаружены в ядре (лат. nucleus)[1]. Позже было обнаружено, что бактериальные клетки, в которых нет ядра, тоже содержат нуклеиновые кислоты. Значение РНК в синтезе белков было предположено в 1939 году в работе Торбьёрна Оскара Касперссона, Жана Брачета и Джека Шульца [2]. Джерард Маирбакс выделил первую матричную РНК, кодирующую гемоглобин кролика и показал, что при её введении в ооциты образуется тот же самый белок[3]. В Советском Союзе в 1956-57 годах проводились работы (А. Белозёрский, А. Спирин, Э. Волкин, Ф. Астрахан) по определению состава РНК клеток, которые привели к выводу, что основную массу РНК в клетке составляет рибосомальная РНК.[4] Северо Очоа получил Нобелевскую премию по медицине в 1959 году за открытие механизма синтеза РНК[5]. Последовательность 77 нуклеотидов одной из тРНК дрожжей S. cerevisiae была определена в 1965 году в лаборатории Роберта Холея, за что в 1968 году он получил Нобелевскую премию по медицине [6]. В 1967 Карл Вёзе предположил, что РНК обладают каталитическими свойствами. Он выдвинул так называемую Гипотезу РНК-мира, в котором РНК прото-организмов служила и в качестве молекулы хранения информации (сейчас эта роль выполняется в основном ДНК) и молекулы, которая катализировала метаболические реакции (сейчас это делают в основном ферменты)[7]. В 1976 Уолтер Фаэрс и его группа в Гентском Университете (Голландия) определили первую последовательность генома РНК-содержащего вируса, бактериофага MS2[8]. В начале 1990-х было обнаружено, что введение чужеродных генов в геном растений приводит к подавлению выражения аналогичных генов растения[9]. Приблизительно в это же время было показано, что РНК длиной около 22 оснований, которые сейчас называются микроРНК, играют регуляторную роль в онтогенезе нематод C.elegans [10].
Химический состав и модификации мономеров
Химическое строение полинуклеотида РНКНуклеотиды РНК состоят из сахара — рибозы, к которой в положении 1′ присоединено одно из оснований: аденин, гуанин, цитозин или урацил. Фосфатная группа соединяет рибозы в цепочку, образуя связи с 3′ атомом углерода одной рибозы и в 5′ положении другой. Фосфатные группы при физиологическом рН отрицательно заряжены, поэтому РНК — полианион. РНК транскрибируется как полимер четырёх оснований (аденина (A), гуанина (G), урацила (U) и цитозина (C), но в «зрелой» РНК есть много модифицированных оснований и сахаров[11]. Всего в РНК насчитывается около 100 разных видов модифицированных нуклеозидов, из которых 2′-О-метилрибоза наиболее частая модификация сахара, а псевдоуридин — наиболее часто встречающееся модифицированное основание [12]. У псевдоуридина (Ψ) связь между урацилом и рибозой не C — N, а C — C, этот нуклеотид встречается в разных положениях в молекулах РНК. В частности, псевдоуридин важен для функционирования тРНК[13]. Другое заслуживающее внимания модифицированное основание — гипоксантин, деаминированный гуанин, нуклеозид которого носит название инозина. Инозин играет важную роль в обеспечении вырожденности генетического кода. Роль многих других модификаций не до конца изучена, но в рибосомальной РНК многие пост-транскрипционные модификации находятся в важных для функционирования рибосомы участках. Например, на одном из рибонуклеотидов, участвующим в образовании пептидной связи[14].
Структура
Азотистые основания в составе РНК могут образовывать водородные связи между цитозином и гуанином, аденином и урацилом, а также между гуанином и урацилом [15]. Однако возможны и другие взаимодействия, например, несколько аденинов могут образовывать петлю, или петля, состоящая из четырёх нуклеотидов, в которой есть пара оснований аденин — гуанин[16].
Разные формы нуклеиновых кислот. На рисунке (слева направо) представлены A (типична для РНК), B (ДНК) и Z (редкая форма ДНК)Важная структурная особенность РНК, отличающая её от ДНК — наличие гидроксильной группы в 2′ положении рибозы, которая позволяет молекуле РНК существовать в А, а не В-конформации, наиболее часто наблюдаемой у ДНК[17]. У А-формы глубокая и узкая большая бороздка и неглубокая и широкая малая бороздка[18]. Второе последствие наличия 2′ гидроксильной группы состоит в том, что конформационно пластичные, то есть не принимающие участие в образовании двойной спирали, участки молекулы РНК могут химически атаковать другие фосфатные связи и их расщеплять [19].
«Рабочая» форма одноцепочечной молекулы РНК, как и у белков, часто обладает третичной структурой. Третичная структура образуется на основе элементов вторичной структуры, образуемой с помощью водородных связей внутри одной молекулы. Различают несколько типов элементов вторичной структуры — стебель-петли, петли и псевдоузлы[20]. В силу большого числа возможных вариантов спаривания оснований предсказание вторичной структуры РНК — гораздо более сложная задача, чем предсказание вторичной структуры белков, но в настоящее время есть эффективные программы, например, mfold [21].
Примером зависимости функции молекул РНК от их вторичной структуры являются участки внутренней посадки рибосомы (IRES). IRES — структура на 5′ конце информационной РНК, которая обеспечивает присоединение рибосомы в обход обычного механизма инициации синтеза белка, требующего наличия особого модифицированного основания (кэпа) на 5′ конце и белковых факторов инициации. Первоначально IRES были обнаружены в вирусных РНК, но сейчас накапливается всё больше данных о том, что клеточные мРНК также используют IRES-зависимый механизм инициации в условиях стресса[22].
Многие типы РНК, например, рРНК и мяРНК в клетке функционируют в виде комплексов с белками, которые ассоциируют с молекулами РНК после их синтеза или (у эукариот) экспорта из ядра в цитоплазму. Такие РНК-белковые комплексы называются рибонуклеопротеиновыми комплексами или рибонуклеопротеидами.
Сравнение с ДНК
Между ДНК и РНК есть три основных отличия:
- ДНК содержит сахар дезоксирибозу, РНК — рибозу, у которой есть дополнительная, по сравнению с дезоксирибозой, гидроксильная группа. Эта группа увеличивает вероятность гидролиза молекулы, то есть уменьшает стабильность молекулы РНК.
- Нуклеотид, комплементарный аденину, в РНК не тимин, как в ДНК, а урацил — неметилированная форма тимина.
- ДНК существует в форме двойной спирали, состоящей из двух отдельных молекул. Молекулы РНК, в среднем, гораздо короче и преимущественно одноцепочечные.
Структурный анализ биологически активных молекул РНК, включая тРНК, рРНК, мяРНК и другие молекулы, которые не кодируют белков, показал, что они состоят не из одной длинной спирали, а из многочисленных коротких спиралей, расположенных близко друг к другу и образующих нечто, похожее на третичную структуру белка. В результате этого РНК может катализировать химические реакции, например, пептидил-трансферазный центр рибосомы, участвующий в образовании пептидной связи белков, полностью состоит из РНК [23][24].
Синтез
Синтез РНК в живой клетке проводится ферментом — РНК-полимеразой. У эукариот разные типы РНК синтезируются разными, специализированными РНК-полимеразами. В целом матрицей синтеза РНК может выступать как ДНК, так и другая молекула РНК. Например, полиовирусы используют РНК-зависимую РНК-полимеразу для репликации своего генетического материала, состоящего из РНК[25]. Но РНК-зависимый синтез РНК, который раньше считался характерным только для вирусов, происходит и в клеточных организмах, в процессе так называемой РНК-интерференции[26].
Как в случае ДНК-зависимой РНК-полимеразы, так и в случае РНК-зависимой РНК-полимеразы фермент присоединяется к промоторной последовательности. Вторичная структура молекулы матрицы расплетается с помощью хеликазной активности полимеразы, которая при движении субстрата в направлении от 5′ к 3′ концу молекулы синтезирует РНК в 3′ —> 5′ направлении. Терминатор транскрипции в исходной молекуле определяет окончание синтеза. Многие молекулы РНК синтезируются в качестве молекул-предшественников, которые подвергаются «редактированию» — удалению ненужных частей с помощью РНК-белковых комплексов[27].
Например, у кишечной палочки гены рРНК расположены в составе одного оперона (в rrnB порядок расположения такой: 16S — tRNAGlu 2 — 23S —5S) считываются в виде одной длинной молекулы, которая затем подвергается расщеплению в нескольких участках с образованием сначала пре-рРНК, а затем зрелых молекул рРНК[28]. Процесс изменения нуклеотидной последовательности РНК после синтеза носит название процессинга или редактирования РНК.
После завершения транскрипции РНК часто подвергается модификациям (см. выше), которые зависят от функции, выполняемой данной молекулой. У эукариот процесс «созревания» РНК, то есть её подготовки к синтезу белка, часто включает сплайсинг: удаление некодирующих белок последовательностей (интронов) с помощью рибонуклеопротеида сплайсосомы. Затем к 5′ концу молекулы пре-мРНК эукариот добавляется особый модифицированный нуклеотид (кэп), а к 3′ концу несколько аденинов, так назваемый «полиА-хвост»[27].
Структура молоточкового (hammerhead) рибозима, который расщепляет РНК.
Типы РНК
Матричная (информационная) РНК — РНК, которая служит посредником при передаче информации, закодированной в ДНК к рибосомам, молекулярным машинам, синтезирующим белки живого организма. Кодирующая последовательность мРНК определяет последовательность аминокислот полипептидной цепи белка [29]. Однако подавляющее большинство РНК не кодируют белок. Эти некодирующие РНК могут транскрибироваться с отдельных генов (например, рибосомальные РНК) или быть производными интронов [30]. Классические, хорошо изученные типы некодирующих РНК — это транспортные РНК (тРНК) и рРНК, которые участвуют в процессе трансляции [31]. Существуют также классы РНК, ответственные за регуляцию генов, процессинг мРНК и другие роли. Кроме того, есть и молекулы некодирующих РНК, способные катализировать химические реакции, такие, как разрезание и лигирование молекул РНК[32]. По аналогии с белками, способными катализировать химические реакции — энзимами (ферментами), каталитические молекулы РНК называются рибозимами.
Участвующие в трансляции
Информация о последовательности аминокислот белка содержится в мРНК. Три последовательных нуклеотида (кодон) соответствуют одной аминокислоте. В эукариотических клетках транскирибированный предшественник мРНК или пре-мРНК процессируется с образованием зрелой мРНК. Процессинг включает удаление некодирующих белок последовательностей (интронов). После этого мРНК экспортируется из ядра в цитоплазму, где к ней присоединяются рибосомы, транслирующие мРНК с помощью соединённых с аминокислотами тРНК.
В безъядерных клетках (бактерии и археи) рибосомы могут присоединяться к мРНК сразу после транскрипции участка РНК. И у эукариот, и у прокариот цикл жизни мРНК завершается её контролируемым разрушением ферментами рибонуклеазами [29].
Транспортные (тРНК) — малые, состоящие из приблизительно 80 нуклеотидов, молекулы с консервативной третичной структурой. Они переносят специфические аминокислоты в место синтеза пептидной связи в рибосоме. Каждая тРНК содержит участок для присоединения аминокислоты и антикодон для узнавания и присоединения к кодонам мРНК. Антикодон образует водородные связи с кодоном, что помещает тРНК в положение, способствующее образованию пептидной связи между последней аминокислотой образованного пептида и аминокислотой, присоединённой к тРНК [30].
Рибосомальные РНК (рРНК) — каталитическая составляющая рибосом. Эукариотические рибосомы содержат четыре типа молекул рРНК: 18S, 5.8S, 28S и 5S. Три из четырёх типов рРНК синтезируются в ядрышке. В цитоплазме рибосомальные РНК соединяются с рибосомальными белками и формируют нуклеопротеин, называемый рибосомой[29]. Рибосома присоединяется к мРНК и синтезирует белок. рРНК составляет до 80 % РНК, обнаруживаемой в цитоплазме эукариотической клетки [33].
Необычный тип РНК, который действует в качестве тРНК и мРНК (тмРНК) обнаружен во многих бактериях и пластидах. При остановке рибосомы на дефектных мРНК без стоп-кодонов тмРНК присоединяет небольшой пептид, направляющий белок на деградацию [34].
Участвующие в регуляции генов
В живых клетках обнаружено несколько типов РНК, которые могут уменьшать степень выражения гена при комплементарности мРНК или самому гену. Микро-РНК (21-22 нуклеотида в длину) найдены у эукариот и оказывают воздействие через механизм РНК-интерференции. При этом комплекс микро-РНК и ферментов может приводить к метилированию нуклеотидов в ДНК промотора гена, что служит сигналом для уменьшения активности гена. При использовании другого типа регуляции мРНК, комплементарная микро-РНК, деградируется.[35]. Однако есть и миРНК, которые увеличивают, а не уменьшают экспрессию генов.[36]. Малые интерферирующие РНК (миРНК, 20-25 нуклеотидов) часто образуются в результате расщепления вирусных РНК, но существуют и эндогенные клеточные миРНК[37]. Малые интерферирующие РНК также действуют через РНК-интерференцию по сходным с микро-РНК механизмам[38]. У животных найдены так называемыме РНК, взаимодействующие с Piwi (piRNA, 29-30 нуклеотидов), действующие в половых клетках против транспозиции и играющие роль в образовании гамет[39][40]. Кроме того, piRNA могут эпигенетически наследоваться по материнской линии, передавая потомству своё свойство ингибировать экспрессию транспозонов[41].
Антисмысловые РНК широко распространены у бактерий, многие из них подавляют выражение генов, но некоторые активируют экспрессию[42]. Действуют антисмысловые РНК, присоединяясь к мРНК, что приводит к образованию двуцепочечных молекул РНК, которые деградируются ферментами.[43]. У эукариот обнаружены высокомолекулярные, мРНК-подобные молекулы РНК. Эти молекулы также регулируют выражение генов,[44]. В качестве примера можно привести Xist, присоединяющуюся и инактивирующую одну из двух Х-хромосом у самок млекопитающих.[45].
Кроме роли отдельных молекул в регуляции генов, регуляторные элементы могут формироваться в 5′ и 3′ нетранслируемых участках мРНК. Эти элементы могут действовать самостоятельно, предотвращая инициацию трансляции, либо присоединять белки, например, ферритин или малые молекулы, например, биотин.[46].
В процессинге РНК
Многие РНК принимают участие в модификации других РНК. Интроны вырезаются из пре-мРНК сплайсосомами, которые, кроме белков, содержат несколько малых ядерных РНК (мяРНК)[31]. Кроме того, интроны могут катализировать собственное вырезание[47]. Синтезированая в результате транскрипции РНК также может быть химически модифицирована. У эукариот химические модификации нуклеотидов РНК, например, их метилирование, выполняется малыми ядерными РНК (мяРНК, 60-300 нуклеотидов). Этот тип РНК локализуется в ядрышко и тельцах Кахаля [30]. После ассоциации мяРНК с ферментами, мяРНК связываются с РНК-мишенью путём образования пар между основаниями двух молекул, а ферменты модифицируют нуклеотиды РНК-мишени. Рибосомальные и транспортные РНК содержат много подобных модификаций, конкретное положение которых часто сохраняется в процессе эволюции. Также могут быть модифицированы мяРНК и сами мяРНК[48][49] Гидовые РНК осуществляют процесс редактирования РНК в кинетопласте — особом участке митохондрии протистов-кинетопластид (например, трипаносом).
Геномы, состоящие из РНК
Жизненный цикл вируса с РНК геномом на примере полиовируса: 1 — присоединение исходного вириона к рецептору; 2 — вирион попадает в клетку; 3 — трансляция белков вируса с его РНК с образованием полипетида; 4 — полимеразы вируса размножают его РНККак и ДНК, РНК может хранить информацию о биологических процессах. РНК может использоваться в качестве генома вирусов и вирусоподобных частиц. РНК-геномы можно разделить на те, которые не имеют промежуточной стадии ДНК и те, которые для размножения копируются в ДНК-копию и обратно в РНК (ретровирусы).
РНК-содержащие вирусы
Многие вирусы, например, вирус гриппа, на всех стадиях содержат геном, состоящий исключительно из РНК. РНК содержится внутри обычно белковой оболочки и реплицируется с помощью закодированных в ней РНК-зависимых РНК-полимераз. Вирусные геномы, состоящие из РНК разделяются на
- содержащие «плюс-цепь РНК», которая используется в качестве и мРНК, и генома;
- «минус-цепь РНК», которая служит только геномом, а в качестве мРНК используется комплементарная ей молекула;
- двухцепоченые вирусы.
Вироиды — другая группа патогенов, содержащих РНК-геном и не содержащих белок. Они реплицируются РНК-полимеразами организма хозяина[50].
Ретровирусы и ретротранспозоны
У других вирусов РНК-геном есть в течение только одной из фаз жизненного цикла. Вирионы так называемых ретровирусов содержат молекулы РНК, которые при попадании в клетки хозяина служат матрицей для синтеза ДНК-копии. В свою очередь, с матрицы ДНК считывается РНК-геном. Кроме вирусов обратную транскрипции применяют и класс мобильных элементов генома — ретротранспозоны [51]
Гипотеза РНК-мира
Способность молекул РНК одновременно служить как в качестве носителя информации, так и в качестве катализатора химических реакций, позволила выдвинуть гипотезу о том, что РНК была первым сложным полимером, появившимся в процессе добиологической эволюции. Эта гипотеза названа «гипотеза РНК-мира» [52][53]. Согласно ей, РНК на первых этапах эволюции автокатализировала синтез других молекул РНК, а затем и ДНК. На втором этапе эволюции синтезированные молекулы ДНК, как более стабильные, стали хранилищем генетической информации. Синтез белка на матрице РНК с помощью пра-рибосом, полностью состоящих из РНК, расширил свойства добиологических систем, постепенно белок заменил РНК в структурных аспектах. Из этой гипотезы делается вывод, что многие РНК, принимающие участие в синтезе белка в современных клетках, в особенности рРНК и тРНК — это реликты РНК-мира.
См. также
Примечания
Рекомендуемая литература
Ссылки
| Типы нуклеиновых кислот | |
|---|---|
| Азотистые основания | Пурины (Аденин, Гуанин) | Пиримидины (Урацил, Тимин, Цитозин) |
| Нуклеозиды | Аденозин | Гуанозин | Уридин | Тимидин | Цитидин |
| Нуклеотиды | монофосфаты (АМФ, ГМФ, UMP, ЦМФ) | дифосфаты (АДФ, ГДФ, УДФ, ЦДФ) | трифосфаты (АТФ, ГТФ, УТФ, ЦТФ) | циклические (цАМФ, цГМФ, cADPR) |
| Рибонуклеиновые кислоты | РНК | мРНК | тРНК | рРНК | антисмысловые РНК | gRNA | микроРНК | некодирующие РНК | piwi-interacting RNA | shRNA | малые интерферирующие РНК | малые ядерные РНК | малые ядрышковые РНК | тмРНК |
| Дезоксирибонуклеиновые кислоты | ДНК | кДНК | Геном | msDNA | Митохондриальная ДНК |
| Аналоги нуклеиновых кислот | en:glycerol nucleic acid | en:locked nucleic acid | ПНК | ТНК | Морфолино |
| Типы векторов | en:phagemid | Плазмиды | Фаг лямбда | en:cosmid | en:P1 phage | en:fosmid | en:Bacterial artificial chromosome | en:Yeast artificial chromosome | en:Human artificial chromosome |
РИБОНУКЛЕИНОВЫЕ КИСЛОТЫ (РНК) — фосфорсодержащие биополимеры, имеющие универсальное распространение в живой природе и являющиеся неотъемлемым компонентом всех микроорганизмов, растительных и животных клеток, а также многих вирусов. РНК представляют собой один из двух известных в живой природе типов нуклеиновых кислот (см.). Биол. функция РНК связана с их центральной ролью в реализации генетической информации — уникальной программы развития всех признаков и свойств каждого живого организма. Носителями наследственной информации в большинстве случаев служат молекулы ДНК (см. Дезоксирибонуклеиновые кислоты), однако РНК также может выполнять аналогичную роль (роль генетической нуклеиновой к-ты), напр, у вирусов, обеспечивая в этом случае не только реализацию, но и хранение и наследственную передачу генетической программы.
РНК представляют собой высокомолекулярные соединения с линейной структурой молекул. Мономерные звенья — рибонуклеотиды — связаны между собой с помощью эфирных связей между 5′-фосфатом одного нуклеотида и 3′-гидроксилом углеводного остатка следующего нуклеотида (5′,3′-фосфодиэфирная связь) и образуют длинную неразветвленную цепь полирибонуклеотида. Углеводный компонент РНК представлен пентозой — пятиуглеродным сахаром — D-рибозой (см. Рибоза), отсюда первоначальное название рибонуклеиновых к-т — пентозонуклеиновые к-ты. Азотистыми компонентами РНК являются гетероциклические основания, два из к-рых производные пурина (см. Пуриновые основания) — аденин (А) и гуанин (Г) и два производные пиримидина (см. Пиримидиновые основания) — цитозин (Ц) и урацил (У). Характерными структурными элементами нек-рых РНК являются так наз. минорные основания; соответствующие им нуклеотиды обычно входят в состав транспортных РНК (тРНК) и других РНК в очень небольших количествах. Азотистые основания связаны с углеводным остатком через его первый (гликозид-ный) углеродный атом. Пурины присоединяются к углеводу через атом азота в положении 9 пуринового кольца, пиримидины — через атом азота в положении 3. Конец цепи, несущий свободный или фосфорилированный 5′-гидроксил рибозы, называют 5′-концом, а конец цепи, содержащий свободный или фосфорилированный З’-гидроксил рибозы,-3′-концом молекулы.
 отрезок полирибонуклеотидной цепи молекулы РНК
отрезок полирибонуклеотидной цепи молекулы РНКПолинуклеотидные цепи РНК обладают гибкой структурой, их длина в зависимости от вида РНК может варьировать в очень широких пределах — от нескольких десятков до нескольких десятков тысяч нуклеотидных остатков. Мол. вес (масса) РНК в среднем 104—106. Последовательность нуклеотидных звеньев, соединенных фосфодиэфирной связью в непрерывную и неразветвлен-ную полинуклеотидную цепь, называют первичной структурой РНК; она строго специфична и уникальна для каждого вида природной РНК. Первичная структура РНК представляет собой форму записи биол. информации, многократно и точно воспроизводящуюся в процессах биосинтеза РНК (см. Генетический код). Это обусловливает большое разнообразие индивидуальных молекул РНК. Информация о структуре белка в виде уникальной последовательности азотистых оснований полирибонуклеотида переносится от ДНК (см. Транскрипция) к рибосомам (см.), чтобы транслироваться там в соответствующую последовательность аминокислот (см. Трансляция).
Вторичная и третичная структура РНК, определяемая как пространственная конфигурация полинукле-отидной цепи, формируется в основном за счет водородных связей и меж-плоскостных гидрофобных взаимодействий между азотистыми основаниями. Если для молекулы нативной ДНК характерно устойчивое спиральное строение, то макромолеку-лярная структура РНК гораздо более вариабельна и лабильна. В р-рах с низкой ионной силой молекулы РНК ведут себя как типичные сильно разбухшие цепи полиэлектролита, но при повышении ионной силы эти цепи сжимаются, их характеристическая вязкость уменьшается, а скорость седиментации увеличивается. Это объясняется образованием на отдельных участках гибкой цепи РНК, к-рая, перегибаясь, навивается сама на себя, и двуспиральных структур в результате так наз. комплементарного спаривания, аналогичного комплемен-тарности в двуспиральных молекулах ДНК. Стабилизация таких структур в РНК достигается за счет образования водородных связей между противолежащими азотистыми основаниями антипараллельных участков цепи; специфическими парами азотистых оснований между комплементарными участками цепи являются классические А—У, Г—Ц и, реже, Г—У.
Наличие в азотистых основаниях сопряженных двойных связей обусловливает интенсивное поглощение РНК в УФ-области спектра с максимумом при длине волны ок. 260 нм. Образование спиральной структуры сопровождается ослаблением поглощения при 260 нм (так наз. гипо-хромный эффект). Обратный процесс — разрушение двуспиральной структуры, происходящее при понижении ионной силы р-ра РНК или при его нагревании,— называют молекулярным плавлением. Оно объясняется конформационным переходом спираль —> беспорядочный клубок и связано с ослаблением стабилизирующих взаимодействий в молекуле РНК. В этом случае наблюдается гиперхромный эффект — увеличение поглощения при 260 нм.
Молекулы РНК, состоящие из двух комплементарных полинуклео-тидных цепей, обнаружены в нек-рых вирусах растений и животных. Кроме того, двуцепочечные молекулы РНК образуются как промежуточные продукты биосинтеза многих вирусных РНК в клетке; их называют репликативными формами РНК. По многим параметрам (по величине шага спирали, числу пар азотистых оснований на виток — 11—12 пар, углам их наклона к оси спирали, а также по конфигурации сахарофосфатного остова) двуспиральные молекулы или участки молекул РНК похожи на двуспиральные молекулы ДНК в A-форме. Нек-рые двутяжевые РНК подобно ДНК могут существовать в форме кольцевых молекул и в случае, если обе полинуклеотидные цепи ковалентно замкнуты, образовывать супер-спирализованные кольца. РНК способна к формированию двутяжевых комплексов, в к-рых один из тяжей представлен полирибонукле-отидной, а другой — полидезоксирибонуклеотидной цепью. Образование таких ДНК—РНК-гибридов происходит во время репликации ДНК с участием затравочных фрагментов РНК (см. Репликация), а также в процессе транскрипции РНК на матрице ДНК. Кроме того, ДНК— РНК-гибриды формируются после заражения клеток нек-рыми РНК-содержащими вирусами в результате синтеза на вирусной РНК комплементарной ей ДНК с помощью вирусоспецифического фермента — обратной транскриптазы (см. Ревертаза).
Подавляющее большинство природных РНК относится к однотяжевым полинуклеотидам. Однако в полинуклеотидных цепях РНК имеются участки различной длины, состоящие из комплементарных друг другу нуклеотидных последовательностей, включающих от десятков до тысяч нуклеотидных остатков и расположенных на небольшом удалении друг от друга. Благодаря этому в молекулах РНК возникают как короткие, так и весьма протяженные двутяжевые (биспиральные) участки, принадлежащие одной цепи, так наз. шпильки. Модель вторичной структуры РНК со шпилькообразными элементами была создана в конце 50-х —начале 60-х гг. 20 в. в лабораториях А. С. Спирина и Доти (P. Doty).
Первые подходы к определению нуклеотидной последовательности РНК были разработаны в середине 60-х гг. 20 в. в лабораториях Р. Холли, Цахау (H. Zachau) и А. А. Баева; это положило начало анализу структурно-функциональной организации индивидуальных РНК.
Содержание РНК в живых клетках (за исключением сперматозоидов) значительно выше, чем содержание ДНК, и распределение их внутри клетки сложнее. Основная масса РНК локализована в цитоплазме, они входят в состав собственно цитоплазматических рибосом (см.), а также рибосом митохондрий (см.) и присутствуют в свободном виде или в виде нерибосомных комплексов с белками. В ядре РНК являются составной частью хроматина (см.). Часть РНК хроматина является продуктом текущих процессов транскрипции генов, в т. ч. и регуляторных (см. Ген), однако имеются косвенные и прямые указания на существование специальных форм хроматиновой РНК, играющей регуляторную роль.
Большинство РНК животных клеток, бактерий и ДНК-содержащих вирусов синтезируется на матрице двуцепочечной ДНК в процессе транскрипции. Одноцепочечные РНК ряда вирусов образуются на матрице дву цепочечных РНК.
Матричный синтез одноцепочечных РНК существенно отличается от репликации ДНК и двуцепочечных РНК: он является консервативным, а не полуконсервативным, т. е. продукт синтеза не включает в себя каких-либо компонентов матрицы (см. Репликация). Консервативный характер синтеза и необходимость реплицировать не обе цепочки матрицы, а лишь одну, и не на всем протяжении матрицы, а только на определенных участках обусловливают существование специальных механизмов узнавания инициаторных и терминаторных последовательностей, определяющих начало и конец синтеза молекулы РНК.
В клетке биосинтез РНК на матрице ДНК осуществляют ферменты РНК-полимеразы (см. Полимеразы). В клетках эукариотов обнаружено по крайней мере три фермента, ответственных за синтез разных типов РНК. В отличие от изученных ДНК-полимераз РНК-полимеразы проявляют определенную специфичность по отношению к разным матрицам и даже участкам матриц. РНК-полимераза катализирует образование 3′,5′-фосфодиэфирных связей между мономерными рибонуклеози-дами, используя в качестве субстратов нуклеозидтрифосфаты. Синтез начинается с образования связи между двумя мононуклеотидами, при этом первый из них, как правило, является пуриновым нуклеотидом. Растущая полинуклеотидная цепь РНК удлиняется в направлении 5′->3′. Синтезированная РНК комплементарна матрице ДНК, и порядок включения нуклеотидов в цепь РНК определяется последовательностью нуклеотидов в матрице ДНК, поскольку в основе матричного синтеза лежат закономерности образования комплементарных цепей. Как правило, РНК синтезируются в виде молекул-предшественников (пре-РНК), имеющих больший молекулярный вес, чем функционально активные молекулы. Эти молекулы-предшественники проходят многостадийный процесс созревания — так наз. посттранскрипционный процессинг, к-рый сводится к вырезанию специализированными клеточными ферментами нек-рых последовательностей и модификации первичной структуры в результате ферментативного метилирования, дегидрирования и т. д. азотистых оснований, а также изомеризации нуклеотидов.
Функции РНК в клетке сложны и многообразны. В соответствии с функциональным назначением и структурными особенностями в любой клетке различают три основных типа РНК: рибосомные РНК (рРНК), транспортные РНК (тРНК) и информационные, или матричные, РНК (иРНК). Кроме указанных типов РНК, в клеточных ядрах и в цитоплазме в небольших количествах встречаются и другие разновидности молекул РНК. Установлено, что в нек-рых случаях они являются предшественниками РНК вышеперечисленных типов. В цитоплазме и в ядре имеется набор так наз. малых РНК, функции их пока неизвестны.
Рибосомная РНК имеет большой молекулярный вес и характеризуется метаболической стабильностью. Она составляет ок. 80% от всех клеточных РНК. Выделяют ее из очищенных рибосом или их субчастиц путем обработки водным р-ром фенола, к-рый денатурирует белки и делает их нерастворимыми. По весу рРНК составляет от 50 до 65% всего материала рибосом. Рибосомы всех организмов состоят из двух субъединиц: малой и большой. В состав большой субъединицы рибосомы клеток эукариотов входит РНК с мол. весом ок. 1,65•106 (26—28S-PHK), в состав малой субъединицы — РНК с мол. весом ок. 0,65•106 (18S-PHK). Большая и малая субъединицы рибосом клеток прокариотов содержат РНК с мол. весом соответственно ок. 1,1•106 (23S-PHK) и ок. 0,5•106 (16S-PHK).
В каждой субъединице молекула рРНК служит как бы каркасом, на к-ром собираются рибосомные белки; сформировавшийся рибонуклео-протеидный комплекс — так наз. рибонуклеопротеидный тяж (РНП-тяж) — организуется в сложную компактную частицу — собственно рибосомную субъединицу. Концепция РНП-тяжа как основы структурной организации рибосомы разработана в 60-х гг. 20 в. А. С. Спириным.
С рибосомами ассоциирована также РНК с относительно низким мол. весом: 5S-PHK, содержащая ок. 120 нуклеотидов и связанная с большой рибосомной субъединицей; такая РНК обнаружена в бактериальных и животных клетках. Кроме того, в рибосомах эукариотов, как правило, присутствует еще одна низкомолекулярная РНК, так наз. 5,8S-РНК, к-рая довольно прочно ассоциирована с помощью водородных связей с 28S-PHK большой рибосомной субъединицы. Эти РНК являются структурными компонентами рибосомы.
Роль рРНК в белоксинтезирующей системе клетки не исчерпывается их структурными функциями. У прокариотов на 3′-конце молекулы 16S-PHK имеется богатая пиримиди-нами последовательность, комплементарная небольшому участку иРНК, расположенному на 5′-конце ее молекулы. Комплементарное спаривание этих участков, по-видимому, способствует первоначальному связыванию иРНК с рибосомой. Не исключено, что нек-рые участки рРНК играют определенную роль в формировании пептидил-трансферазного центра рибосомы, ответственного за образование пептидных связей при синтезе белка.
Биосинтез рРНК в клетках эукариотов происходит в ядре и осуществляется при участии фермента РНК-полимеразы I. Геном эукариотов содержит много копий генов, кодирующих рРНК; рибосомные гены сгруппированы в виде тандемных повторов и локализованы в одной или нескольких хромосомах (см.). Эти участки генома являются составной частью нуклеопротеида (см.), образующего ядрышки (см.) — клеточные органеллы, в пределах к-рых и синтезируется рРНК. Образование этой РНК представляет собой сложный, многостадийный процесс. Молекулы рРНК синтезируются в виде гигантского предшественника (пре-рРНК) с мол. весом 4-106 (45S-PHK), к-рый в дальнейшем подвергается модификации (метилирование, изомеризация и т. д.) и специфическому расщеплению с образованием промежуточных форм зрелых рРНК. При этом почти половина исходной молекулы предшественника деградирует. Рибонуклеазы (см.), отделяющие от предшественников рРНК те участки их нуклеотидных цепей, к-рых уже нет в рибосоме, изучены еще мало.
Транспортная РНК составляет примерно 15% от общего количества клеточных РНК. Это относительно низкомолекулярные РНК: их нуклеотидные цепи содержат всего 75—90 нуклеотидов, а мол. вес находится в пределах 23 000—30 000. Ввиду небольшого размера тРНК легко отделяются от молекул других РНК, как правило, гораздо более крупных. Этот тип РНК — наиболее изученный в молекулярном плане элемент белоксинтезирующей системы. Для большинства тРНК определена полная последовательность нуклеотидов в молекуле.
Способ выделения тРНК состоит в обработке одноклеточных организмов или гомогенизированных тканей водным р-ром фенола, осаждении спиртом с последующим отделением рРНК, примесей ДНК и полисахаридов. В результате получают препарат суммарной тРНК. Фракционирование препаратов суммарной тРНК осуществляют с помощью ряда физических, химических или комбинированных методов.
Особенностью тРНК, отличающей ее от других РНК, является относительно высокое содержание минорных нуклеотидов. На основании данных о первичной структуре тРНК была предложена и экспериментально подтверждена модель вторичной структуры, плоское изображение к-рой напоминает клеверный лист. Сравнение структур различных тРНК, организованных в «клеверный лист», выявляет ряд общих черт. Во всех этих структурах имеется 4 двуцепочечных спиральных участка, 3 из к-рых являются «шпильками», несущими петли из неспаренных нуклеотидов; 3′- и 5′- концы полинуклеотидной цепи объединены в наиболее длинный спирализован-ный участок, содержащий 7 пар азотистых оснований, завершающийся неспаренным акцепторным тринук-леотидом ЦЦА, к к-рому присоединяется аминокислота. Противолежащая акцепторному концу петля содержит тринуклеотид антикодон, к-рый обеспечивает специфичность взаимодействия с комплементарным ему триплетом-кодоном в иРНК. Нуклеотиды, образующие антикодон, всегда расположены в середине петли. Боковые петли, видимо, играют важную роль в связывании тРНК с аминоацил-тРНК-синтетазой и с комплексом рибосома — и PH К.
Дальнейшее исследование структуры тРНК показало, что нативные молекулы имеют компактную форму: отдельные двуспиральные «шпильки» «клеверного листа» складываются в специфичную третичную структуру, к-рая является близкой у всех тРНК. Способность тРНК образовывать кристаллы позволила применить для изучения пространственной структуры ее молекулы метод рентгеноструктурного анализа. В 1973 —1975 гг. третичная структура одной из тРНК была расшифрована в лабораториях Рича (A. Rich) и Клуга (A. Klug). Согласно модели Рича — Клуга макромолекула тРНК имеет L-образную форму, причем основные функциональные центры молекулы — антикодоновая петля и акцепторный конец — находятся на ее концах. Расстояние между ними составляет 7,6 нм. За стабилизацию третичной структуры ответственны взаимодействия азотистых оснований, отличные от тех, к-рые обусловливают комплементарное спаривание по Уотсону—Крику.
Структура тРНК отличается большой консервативностью, что, по-видимому, связано с высокой степенью ее функциональной специализации. Этот класс молекул в ходе биосинтеза белков (см.) выполняет по отношению к аминокислотам функцию адапторов, к-рые с помощью высокоспецифичных ферментов — аминоацил-тРНК-синтетаз — присоединяют к себе ту или иную аминокислоту и переносят ее на рибосому. Связывание аминокислоты с тРНК происходит за счет образования ковалентной связи между СООН-группой аминокислоты и остатком рибозы З’-концевого аденозина тРНК. Фермент, осуществляющий этот процесс, — аминоацил-тРНК-синтетаза — способен «узнавать» как аминокислоту, так и соответствующую ей тРНК.
Существует целый набор различных тРНК, в к-ром каждая тРНК специфична по отношению к какой-либо одной аминокислоте. В клетке имеется примерно 20 различных аминокислот, однако установлено, что в ряде случаев для одной и той же аминокислоты имеется две или более — иногда пять или шесть — видов тРНК. Такие тРНК называют изоакцепторными тРНК.
В ходе синтеза полипептида на рибосоме тРНК «узнает» специфическую аминоацил-тРНК-синтетазу, от к-рой она принимает соответствующую активированную аминокислоту; затем присоединяется к кодону иРНК на рибосоме и тем самым обеспечивает строгую специфичность выбора и встраивания аминокислоты в аминокислотную последовательность растущего полипептида; после образования пептидной связи тРНК удерживает на рибосоме растущую полипептидную цепь.
В геноме E. coli насчитывается несколько десятков генов, контролирующих синтез тРНК. В клетках эукариотов их число много больше; по всей видимости, структурные гены тРНК распределены по разным хромосомам. Нуклеотидная цепь тРНК синтезируется в ядре при помощи фермента РНК-полимеразы III и появляется в цитоплазме в виде макромолекулярного предшественника, к-рый не содержит метилированных оснований. Предшественник длиннее функционально активной тРНК и имеет несколько менее компактную третичную структуру. Кроме того, в ряде случаев РНК-полимераза строит нуклеотидную цепь без трех последних нуклеотидов: 3′-конец каждой молекулы тРНК, заканчивающийся триплетом ЦЦА, образуется позже при участии особого фермента.
Как и рРНК, тРНК проходит стадию созревания, по завершении к-рой молекулы тРНК приобретают окончательную конформацию. Процесс превращения предшественника в тРНК сводится к вырезанию специальными клеточными ферментами дополнительных последовательностей и к модификации первичной структуры. Одним из многочисленных типов модификации является метилирование нуклеотидов с помощью тРНК-метилаз.
Информационная, или матричная, РНК составляет весьма незначительную часть общей массы клеточной РНК, всего 5—10% . В отличие от рРНК итРНК фракция иРНК характеризуется выраженной гетерогенностью по размеру молекул (мол. вес иРНК доходит до 2-106), т. к. она представляет собой совокупность молекул, программирующих синтез всех клеточных белков. В связи с этим относительное содержание индивидуальной иРНК в суммарном препарате РНК может составлять тысячные доли процента.
В то время как рРНК и тРНК метаболически устойчивы, иРНК в ряде случаев, особенно у прокариотических организмов, является относительно короткоживущей. Ее нуклеотидный состав близок к составу ДНК, выделенной из того же организма. В составе большинства иРНК обнаружены последовательности полиА, ковалентно связанные с З’-концом молекулы. Полиаденило-вый «хвост» иРНК содержит от нескольких десятков до сотен нуклеотидов и является характерной особенностью этого типа РНК.
Значительная часть цитоплазматической и PH К животной клетки локализуется в составе полисом (по-лирибосом), компонентами к-рых является транслируемая иРНК и связанная с ней группа рибосом, находящихся на разных стадиях трансляции и содержащих растущие по-липептидные цепи разной длины. Полисомы или их субфракции, полученные тем или иным способом, служат основным источником иРНК, выделение к-рой можно проводить путем непосредственной депроте-инизации полирибосом или после предварительного выделения полисомного иРНП (рибонуклеопро-теида). Наличие полиА в и РНК позволяет применять рациональные способы ее выделения из гетерогенных препаратов РНК, основанные на использовании специфических свойств полиА. В нек-рых случаях полиаденилированную иРНК выделяют непосредственно из нефракци-онированного цитоплазматического экстракта. Эти методы основаны на образовании водородных связей между полиА в и РНК и комплементарными олиго- или полинуклеотидами, иммобилизованными на инертных носителях. В качестве таких аффинных носителей (сорбентов) применяют полиУ-сефарозу и олиго (дезокситимидин)-целлюлозу. Иногда для выделения иРНК используют избирательную сорбцию полиА-содержащих иРНК на нитроцеллюлоз-ных фильтрах или колонках с химически модифицированными целлюлозами, обусловленную аномальными сорбционными свойствами аденилового гомополирибонуклеотида.
Из структурных признаков, характеризующих почти любую иРНК животного происхождения, следует отметить наличие блокированного метилированного 5′-конца, так наз. кэпа, содержащего 7-метилгуанозин, связанный с Б’-концевым нуклеотидом 5’—5′-трифосфатным мостиком. Кэп необходим для эффективной инициации синтеза белка на иРНК и, вероятно, для защиты Б’-конца молекулы иРНК от клеточных экзо-нуклеаз (см. Нуклеазы). Между кэпом и инициаторным кодоном АУГ, определяющим начало синтеза белковой цепи, существует 5′-конце-вая зона, не транслируемая в последовательность аминокислот. Далее следует транслируемая, т. е. кодирующая белок, область, длина к-рой варьирует в широких пределах в зависимости от молекулярных размеров кодируемого полипептида. В клетках прокариотических организмов транслируемая область иРНК обычно содержит информацию для синтеза нескольких полипептидных цепей, т. е. относится к так наз. поли-цистронному типу. Сегменты поли-цистронной иРНК, соответствующие каждому гену, транслируются раздельно благодаря наличию между ними внутримолекулярных сигналов инициации и терминации трансляции. иРНК животных клеток являются, как правило, моноцистронны-ми. Это справедливо и для таких иРНК, к-рые кодируют образование нескольких полипептидных цепей; образование зрелых полипептидов, закодированных в такой матрице, обеспечивается посттрансляционным протеолитическим расщеплением первичного продукта синтеза.
В З’-концевой зоне молекулы иРНК между терминаторным кодоном, ограничивающим транслируемую область, и полиА локализована нуклеотидная последовательность значительной длины, не несущая информации о структуре кодируемого белка. Функция ее пока не ясна.
В изучении первичной структуры иРНК достигнуты заметные успехи. Полностью установлена первичная структура иРНК а- и Р-цепей гемоглобина кролика и человека (см. Гемоглобин), проинсулина крысы (см. Проинсулин), куриного овальбумина и многих других. Определение нуклеотидных последовательностей индивидуальных иРНК представляет большой интерес. Это, в частности, путь идентификации мутантных генов и расшифровки молекулярных механизмов, лежащих в основе синтеза аномальных белков и генетических дефектов регуляции синтеза белков при различных формах наследственной патологии человека и животных. Круг этих патол. состояний достаточно широк. К ним относятся злокачественные новообразования, сопровождающиеся нарушением дифференцировки клеток и качественной перестройкой работы генома, различные эндокринопатии с нарушениями регуляции синтеза гормонов, заболевания печени с недостаточностью синтеза белков сыворотки крови, болезни почек и аномалии системы свертывания крови. Количественные дефекты синтеза индивидуальных белков (недостаточный или избыточный синтез) играют важную роль в патогенезе этих заболеваний. Наиболее детально изучена наследственная патология синтеза белка, к-рая представлена гетерогенной группой дефектов синтеза альфа- или бета-цепей глобина гемоглобина. Предполагают, что отсутствие активности p-иРНК при бета-талассемии (см. Талассемия) может быть связано с мутацией в 5′-концевой области, нарушающей связывание иРНК с рибосомами или вызывающей преждевременную терминацию белкового синтеза.
Аналогичные данные были получены и в случае болезни Вильсона— Коновалова (см. Гепато-церебральная дистрофия), клин, проявления к-рой связаны с интоксикацией организма человека медью. Накопление меди в токсической и неутилизиру-емой форме обусловлено генетически детерминированным нарушением синтеза церулоплазмина (см. Дыхательные пигменты) — основного медьсодержащего гликопротеида плазмы крови. Количественная недостаточность этого белка объясняется, вероятно, дефектом структурной организации Б’-конца церуло-плазминовой иРНК, к-рая определяет как связывание иРНК с рибосомой, так и синтез сигнальной пептидной последовательности.
Нек-рые формы наследственной патологии щитовидной железы человека, как полагают, обусловлены аномалией в структуре пре-иРНК, приводящей к нарушениям ее пост-транскрипционного процессинга и переноса в цитоплазму.
Различные иРНК имеют выраженную вторичную структуру; в состав двуцепочечных участков вовлечено до 75% всех нуклеотидных последовательностей иРНК. Значительная часть участков вторичной структуры в иРНК идентифицирована как «шпилечные» структуры. Внутримолекулярные участки вторичной структуры локализованы, по-видимому, как в транслируемой зоне иРНК, так и в З’-концевой и 5′-кон-цевой нетранслируемых частях ее молекулы. Роль участков вторичной структуры и реализации матричных функций иРНК пока не установлена. Возможно, они обеспечивают адекватное пространственное взаимодействие кэпа и инициирующего кодона, необходимое для их одновременного участия в связывании иРНК на рибосоме. Предполагают также, что «шпильки» играют роль специфических структур, обусловливающих узнавание определенных участков иРНК белковыми компонентами ядерных и цитоплазматических иРНП, так наз. информосом.
В ядре и в цитоплазме животных клеток иРНК в свободном состоянии не переносится, а всегда находится в комплексе со специализированными белками. Белковые компоненты различаются в ядерных и цитоплазматических РНП-частицах: часть ядерных белков-носителей оставляет иРНК в момент ее перехода в цитоплазму и заменяется на транспортирующие белки другого типа — образуются цитоплазматические инфор-мосомы. Функции информосомных белков не ограничиваются внутриклеточным транспортом иРНК и защитой ее от повреждений; предполагают, что они способствуют отделению новообразованной иРНК от ДНК матрицы, ее посттранскрипционному созреванию в ядре, а также играют регуляторную роль в процессах созревания и депонирования нек-рых и PH К в цитоплазме.
Цитоплазматические и ядерные иРНП-частицы были открыты и широко исследованы в нашей стране в начале 70-х гг. 20 в. в лабораториях А. С. Спирина и Г. П. Георгиева.
Как и все перечисленные типы клеточных РНК, иРНК представляют собой популяцию молекул, «скопированных» с соответствующих участков генетической нуклеиновой к-ты (чаще всего ДНК). Однако, если рРНК и тРНК относятся к обслуживающему аппарату белоксинтези-рующей системы клетки, то иРНК является прямым посредником между ДНК и белками и является матрицей для синтеза последних. Т. о. в форму иРНК переводится большая часть информации, содержащейся в ДНК.
В бактериальных клетках еще до завершения синтеза молекулы иРНК может начинаться ее трансляция: рибосомы связываются с отделившимся готовым участком иРНК и образуют полисому, синтезирующую белок. В животных клетках места синтеза белка и РНК пространственно разобщены: иРНК синтезируется в клеточном ядре в виде гигантского предшественника (пре-иРНК). Помимо последовательности иРНК, в молекулах пре-иРНК имеется большое количество участков, выполняющих регуляторные и различные вспомогательные функции. Молекулы пре-иРНК не имеют кэпа и полиаденилового «хвоста» — последний наращивается сразу после завершения транскрипции при помощи так наз. безматричного синтеза, катализируемого концевой поли-А-синтетазой. Из ядра пре-иРНК в составе РНП-частиц — информосом — транспортируется в цитоплазму. В процессе выхода пре-иРНК в цитоплазму основная часть вспомогательных нуклеотидных последовательностей в ней разрушается — молекула претерпевает ряд весьма существенных структурных превращений, включающих образование кэпа, специфическое фрагментирование, метилирование, изомеризацию и т. д., т. е. посттранскрипционный процессинг.
Характерной особенностью многих пре-иРНК эукариотов является наличие внутри информационной зоны молекулы некодирующих участков — интронов, размеры к-рых могут быть весьма значительными; количество таких вставок, не несущих информации для синтеза белка, в разных генах различно. Сложная внутренняя топография молекул пре-иРНК обусловливает сложную картину последовательных реакций посттранскрипционного созревания и формирования зрелой иРНК, кодирующая зона к-рой непрерывна и не содержит интронов. Высокоспецифичные процессы вырезания некодирующих вставок из молекулы пре-иРНК и их объединения (сшивки) получили название сплайсинг. Ферментная система сплайсинга, а также его биол. роль остаются пока совершенно неисследованными.
Гистохимические методы определения рибонуклеиновых кислот в тканях. В основе гистохим. методов выявления РНК в тканях и клетках лежат реакции на компоненты, образующиеся в результате гидролиза этих к-т. Для выявления РНК в ядре и цитоплазме обычно применяют метод Браше, сущность к-рого состоит в специфической деполимеризации РНК рибонуклеазой, ДНК при этом не затрагивается. Параллельно используются два среза ткани, из к-рых один предварительно подвергается действию рибонуклеазы. Затем оба среза окрашивают метиловым зеленым — пиронином, к-рый обладает избирательным сродством к РНК. Считают, что материал, окрашивающийся пиронином в красный цвет и исчезающий после обработки рибонуклеазой, представляет собой РНК. Срезы можно окрашивать также 1% водным р-ром толуидинового синего или 1% р-ром толуидинового синего в 95% этиловом спирте; РНК при этом окрашиваются в синий цвет. Для выявления не только РНК, но и их связи со структурами клетки после окрашивания толуидиновым синим срезы докрашивают 2% р-ром оранжевого G в 5% фосфорновольфрамовой к-те.
Кроме метода Браше, для выявления РНК пользуются красителем, носящим название галлоционин-хромовые квасцы, к-рый дает устойчивую окраску, не меняющуюся при обезвоживании в спирте и просветлении в ксилоле. Окрашивание можно проводить при любых значения pH в пределах от 0,8 до 4,3, но при низких значениях pH (1,5—1,75) специфическое окрашивание РНК максимально. Нек-рые исследователи считают метод выявления РНК с помощью красителя галлоционин-хромовые квасцы более надежным, чем метод Браше. Его используют и для количественного гистохим. определения нуклеиновых к-т.
Для изучения синтеза и обмена РНК применяют и гистоавторадиографию часто в сочетании с электронной авторадиографией (см.). В качестве радиоактивной метки используют 3H-урацил.
При гистохим. изучении локализации РНК в индивидуальных клетках или на тканевых срезах пуриновые и пиримидиновые основания выявляют по интенсивности поглощения в УФ-свете. Нуклеиновые к-ты благодаря наличию гетероциклических пуриновых и пиримидиновых оснований интенсивно поглощают УФ-свет с длиной волны 260 нм. При фотографировании клеток в этих лучах структуры, содержащие нуклеиновые к-ты, идентифицируются довольно легко. Однако с помощью этого метода нельзя проводить прямое дифференциальное определение ДНК и РНК.
Локализацию РНК в клетках и срезах можно определять по углеводному компоненту. После мягкого гидролиза соляной к-той, вызывающей отщепление пуриновых оснований и высвобождение реакционно-способных альдегидных групп остатка сахара, проводят реакцию с метилтриоксифлуореноном. При этом углеводный компонент РНК — рибоза — реагируя с красителем, образует соединение, окрашенное в желто-розовый цвет.
Гистохимически РНК идентифицируют также по выявлению фосфорной к-ты. Фосфатные группы нуклеиновых к-т можно избирательно определять с помощью окрашивания основными красителями. Напр., использование акролеина и толуидинового синего при наличии РНК приводит к появлению нежно-красного окрашивания (ДНК в этих условиях дает темно-синий цвет). Кре-зиловый фиолетовый реагируете РНК в стехиометрических количествах при pH 4,2 и поэтому может быть использован для цитофотометрического количественного определения РНК с применением УФ-микроспектрофотометрии (см. Спектрофотометрия, Цитофотометрия).
Библиография: Гайцхоки В. С. Информационные РНК клеток животных, М., 1980, библиогр.; Дэвидсон Д ж. Н. Биохимия нуклеиновых кислот, пер. с англ., М., 1976; Органическая химия нуклеиновых кислот, под ред. Н. К. Кочеткова и Э. И. Будовского, М., 1970; Спирин А. С. и Гаврилова JI. П. Рибосома, М., 1971; Уотсон Дж. Д. Молекулярная биология гена, пер. с англ., М., 1978; Шабарова 3. А. и Богданов А. А. Химия нуклеиновых кислот и их компонентов, М., 1978; Akzesso-rische Methoden in der Histochemie, hrsg. v. G. Geyer u. H. Luppa, Jena, 1975; Bra-ch et J. Embryologie chimique, P., 1944; Culling Ch. F. A. Handbook of his-topathological and histochemical techniques, L., 1974; Pearse A. G. Histochemistry, v. 1—2, L., 1968 —1972.
П. Л. Иванов; В. В. Серов (гист.).
Нуклеиновые кислоты (от лат. нуклеус — «ядро») впервые были обнаружены в \(1868\) г. в ядрах лейкоцитов швейцарским учёным Ф. Мишером. Позже было выяснено, что нуклеиновые кислоты содержатся во всех клетках (в цитоплазме, ядре и во всех органоидах клетки).
Первичная структура молекул нуклеиновых кислот
Нуклеиновые кислоты — самые крупные из молекул, образуемые живыми организмами. Они являются биополимерами, состоящими из мономеров — нуклеотидов.
Обрати внимание!
Каждый нуклеотид состоит из азотистого основания, пятиуглеродного сахара (пентозы) и фосфатной группы (остатка фосфорной кислоты).

В зависимости от вида пятиуглеродного сахара (пентозы), различают два типа нуклеиновых кислот:
- дезоксирибонуклеиновые кислоты (сокращённо ДНК) — молекула ДНК содержит пятиуглеродный сахар — дезоксирибозу.
- рибонуклеиновые кислоты (сокращённо РНК) — молекула РНК содержит пятиуглеродный сахар — рибозу.
Есть различия и в азотистых основаниях, входящих в состав нуклеотидов ДНК и РНК.
Нуклеотиды ДНК: А — аденин, Г — гуанин, Ц — цитозин, Т — тимин.
Нуклеотиды РНК: А — аденин, Г — гуанин, Ц — цитозин, У — урацил.

Вторичная структура молекул ДНК и РНК
Вторичная структура — это форма молекул нуклеиновых кислот.
Пространственная структура молекулы ДНК была смоделирована американскими учёными Джеймсом Уотсоном и Френсисом Криком в \(1953\) г.
Дезоксирибонуклеиновая кислота (ДНК) — состоит из двух спирально закрученных цепей, которые по всей длине соединены друг с другом водородными связями. Такую структуру (свойственную только молекулам ДНК), называют двойной спиралью.
Рибонуклеиновая кислота (РНК) — линейный полимер, состоящий из одной цепи нуклеотидов.

Исключение составляют вирусы, у которых встречаются одноцепочечные ДНК и двухцепочечные РНК.
Подробнее о ДНК и РНК будет рассказано в разделе «Хранение и передача генетической информации. Генетический код».
Источники:
Каменский А. А., Криксунов Е. А., Пасечник В. В. Биология. 9 класс // ДРОФА.
Каменский А. А., Криксунов Е. А., Пасечник В. В. Биология. Общая биология (базовый уровень) 10–11 класс // ДРОФА.
Лернер Г. И. Биология: Полный справочник для подготовки к ЕГЭ: АСТ, Астрель.
http://distant-lessons.ru/nukleinovye-kisloty.html
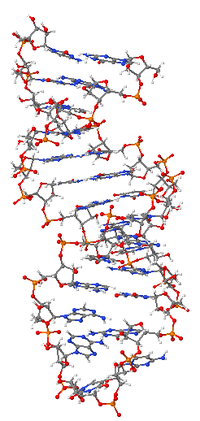
Рибонуклеи́новая кислота́ (РНК) — одна из трёх основных макромолекул (две другие — ДНК и белки), которые содержатся в клетках всех живых организмов и играют важную роль в кодировании, прочтении, регуляции и выражении генов.
Так же, как ДНК (дезоксирибонуклеиновая кислота), РНК состоит из длинной цепи, в которой каждое звено называется нуклеотидом. Каждый нуклеотид состоит из азотистого основания, сахара рибозы и фосфатной группы. Последовательность нуклеотидов позволяет РНК кодировать генетическую информацию. Все клеточные организмы используют РНК (мРНК) для программирования синтеза белков.
Клеточные РНК образуются в ходе процесса, называемого транскрипцией, то есть синтеза РНК на матрице ДНК, осуществляемого специальными ферментами — РНК-полимеразами. Затем матричные РНК (мРНК) принимают участие в процессе, называемом трансляцией. Трансляция — это синтез белка на матрице мРНК при участии рибосом. Другие РНК после транскрипции подвергаются химическим модификациям, и после образования вторичной и третичной структур выполняют функции, зависящие от типа РНК.
Для одноцепочечных РНК характерны разнообразные пространственные структуры, в которых часть нуклеотидов одной и той же цепи спарены между собой. Некоторые высокоструктурированные РНК принимают участие в синтезе белка клетки, например, транспортные РНК служат для узнавания кодонов и доставки соответствующих аминокислот к месту синтеза белка, а рибосомные РНК служат структурной и каталитической основой рибосом.
Однако функции РНК в современных клетках не ограничиваются их ролью в трансляции. Так, малые ядерные РНК принимают участие в сплайсинге эукариотических матричных РНК и других процессах.
Помимо того, что молекулы РНК входят в состав некоторых ферментов (например, теломеразы), у отдельных РНК обнаружена собственная ферментативная активность: способность вносить разрывы в другие молекулы РНК или, наоборот, «склеивать» два РНК-фрагмента. Такие РНК называются рибозимами.
Геномы ряда вирусов состоят из РНК, то есть у них она играет роль, которую у высших организмов выполняет ДНК. На основании разнообразия функций РНК в клетке была выдвинута гипотеза, согласно которой РНК — первая молекула, которая была способна к самовоспроизведению в добиологических системах.
История изучения
Нуклеиновые кислоты были открыты в 1868 году швейцарским учёным Иоганном Фридрихом Мишером, который назвал эти вещества «нуклеин», поскольку они были обнаружены в ядре (лат. nucleus)[1]. Позже было обнаружено, что бактериальные клетки, в которых нет ядра, тоже содержат нуклеиновые кислоты. Значение РНК в синтезе белков было предположено в 1939 году в работе Торбьёрна Оскара Касперссона, Жана Брачета и Джека Шульца[2]. Джерард Маирбакс выделил первую матричную РНК, кодирующую гемоглобин кролика и показал, что при её введении в ооциты образуется тот же самый белок[3]. В 1956—1957 годах А. Белозёрским, А. Спириным, Э. Волкиным, Л. Астраханом проводились работы по определению состава РНК клеток, которые привели к выводу, что основную массу РНК в клетке составляет рибосомальная РНК[4]. Северо Очоа получил Нобелевскую премию по медицине в 1959 году за открытие механизма синтеза РНК[5]. Последовательность 77 нуклеотидов одной из тРНК дрожжей S. cerevisiae была определена в 1965 году в лаборатории Роберта Холея, за что в 1968 году он получил Нобелевскую премию по медицине[6]. В 1967 Карл Вёзе предположил, что РНК обладают каталитическими свойствами. Он выдвинул так называемую гипотезу мира РНК, в котором РНК прото-организмов служила и в качестве молекулы хранения информации (сейчас эта роль выполняется в основном ДНК) и молекулы, которая катализировала метаболические реакции (сейчас это делают в основном ферменты)[7]. В 1976 Уолтер Фаэрс и его группа в Гентском Университете в Бельгии определили первую последовательность генома РНК-содержащего вируса, бактериофага MS2[8]. В начале 1990-х было обнаружено, что введение чужеродных генов в геном растений приводит к подавлению выражения аналогичных генов растения[9]. Приблизительно в это же время было показано, что РНК длиной около 22 оснований, которые сейчас называются микроРНК, играют регуляторную роль в онтогенезе нематод C. elegans[10].
Химический состав и модификации мономеров
Химическое строение полинуклеотида РНКНуклеотиды РНК состоят из сахара — рибозы, к которой в положении 1′ присоединено одно из оснований: аденин, гуанин, цитозин или урацил. Фосфатная группа соединяет рибозы в цепочку, образуя связи с 3′ атомом углерода одной рибозы и в 5′ положении другой. Фосфатные группы при физиологическом рН отрицательно заряжены, поэтому РНК — полианион. РНК транскрибируется как полимер четырёх оснований (аденина (A), гуанина (G), урацила (U) и цитозина (C), но в «зрелой» РНК есть много модифицированных оснований и сахаров[11]. Всего в РНК насчитывается около 100 разных видов модифицированных нуклеотидов, из которых 2′-О-метилрибоза наиболее частая модификация сахара, а псевдоуридин — наиболее часто встречающееся модифицированное основание[12].
У псевдоуридина (Ψ) связь между урацилом и рибозой не C — N, а C — C, этот нуклеотид встречается в разных положениях в молекулах РНК. В частности, псевдоуридин важен для функционирования тРНК[13]. Другое заслуживающее внимания модифицированное основание — гипоксантин, дезаминированный аденин, нуклеозид которого носит название инозина. Инозин играет важную роль в обеспечении вырожденности генетического кода.
Роль многих других модификаций не до конца изучена, но в рибосомальной РНК многие пост-транскрипционные модификации находятся в важных для функционирования рибосомы участках. Например, на одном из рибонуклеотидов, участвующем в образовании пептидной связи[14].
Структура
Азотистые основания в составе РНК могут образовывать водородные связи между цитозином и гуанином, аденином и урацилом, а также между гуанином и урацилом[15]. Однако возможны и другие взаимодействия, например, несколько аденинов могут образовывать петлю, или петля, состоящая из четырёх нуклеотидов, в которой есть пара оснований аденин — гуанин[16].
Разные формы нуклеиновых кислот. На рисунке (слева направо) представлены A (типична для РНК), B (ДНК) и Z (редкая форма ДНК)Важная структурная особенность РНК, отличающая её от ДНК — наличие гидроксильной группы в 2′ положении рибозы, которая позволяет молекуле РНК существовать в А, а не В-конформации, наиболее часто наблюдаемой у ДНК[17]. У А-формы глубокая и узкая большая бороздка и неглубокая и широкая малая бороздка[18]. Второе последствие наличия 2′ гидроксильной группы состоит в том, что конформационно пластичные, то есть не принимающие участие в образовании двойной спирали, участки молекулы РНК могут химически атаковать другие фосфатные связи и их расщеплять[19].
«Рабочая» форма одноцепочечной молекулы РНК, как и у белков, часто обладает третичной структурой. Третичная структура образуется на основе элементов вторичной структуры, образуемой с помощью водородных связей внутри одной молекулы. Различают несколько типов элементов вторичной структуры — стебель-петли, петли и псевдоузлы[20]. В силу большого числа возможных вариантов спаривания оснований предсказание вторичной структуры РНК — гораздо более сложная задача, чем предсказание вторичной структуры белков, но в настоящее время есть эффективные программы, например, mfold[21].
Примером зависимости функции молекул РНК от их вторичной структуры являются участки внутренней посадки рибосомы (IRES). IRES — структура на 5′ конце информационной РНК, которая обеспечивает присоединение рибосомы в обход обычного механизма инициации синтеза белка, требующего наличия особого модифицированного основания (кэпа) на 5′ конце и белковых факторов инициации. Первоначально IRES были обнаружены в вирусных РНК, но сейчас накапливается всё больше данных о том, что клеточные мРНК также используют IRES-зависимый механизм инициации в условиях стресса[22].
Многие типы РНК, например, рРНК и мяРНК в клетке функционируют в виде комплексов с белками, которые ассоциируют с молекулами РНК после их синтеза или (у эукариот) экспорта из ядра в цитоплазму. Такие РНК-белковые комплексы называются рибонуклеопротеиновыми комплексами или рибонуклеопротеидами.
Сравнение с ДНК
Между ДНК и РНК есть три основных отличия:
- ДНК содержит сахар дезоксирибозу, РНК — рибозу, у которой есть дополнительная, по сравнению с дезоксирибозой, гидроксильная группа. Эта группа увеличивает вероятность гидролиза молекулы, то есть уменьшает стабильность молекулы РНК.
- Нуклеотид, комплементарный аденину, в РНК не тимин, как в ДНК, а урацил — неметилированная форма тимина.
- ДНК существует в форме двойной спирали, состоящей из двух отдельных молекул. Молекулы РНК, в среднем, гораздо короче и преимущественно одноцепочечные.
Структурный анализ биологически активных молекул РНК, включая тРНК, рРНК, мяРНК и другие молекулы, которые не кодируют белков, показал, что они состоят не из одной длинной спирали, а из многочисленных коротких спиралей, расположенных близко друг к другу и образующих нечто, похожее на третичную структуру белка. В результате этого РНК может катализировать химические реакции, например, пептидил-трансферазный центр рибосомы, участвующий в образовании пептидной связи белков, полностью состоит из РНК[23][24].
Синтез
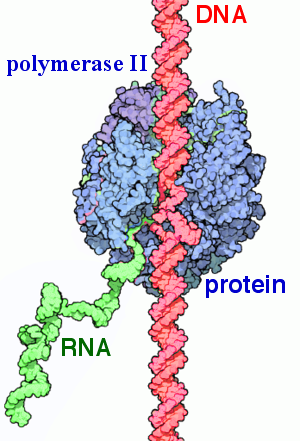
Синтез РНК в живой клетке проводится ферментом — РНК-полимеразой. У эукариот разные типы РНК синтезируются разными, специализированными РНК-полимеразами. В целом матрицей синтеза РНК может выступать как ДНК, так и другая молекула РНК. Например, полиовирусы используют РНК-зависимую РНК-полимеразу для репликации своего генетического материала, состоящего из РНК[25]. Но РНК-зависимый синтез РНК, который раньше считался характерным только для вирусов, происходит и в клеточных организмах, в процессе так называемой РНК-интерференции[26].
Как в случае ДНК-зависимой РНК-полимеразы, так и в случае РНК-зависимой РНК-полимеразы фермент присоединяется к промоторной последовательности. Вторичная структура молекулы матрицы расплетается с помощью хеликазной активности полимеразы, которая при движении субстрата в направлении от 3′ к 5′ концу молекулы синтезирует РНК в направлении 5′ → 3′. Терминатор транскрипции в исходной молекуле определяет окончание синтеза. Многие молекулы РНК синтезируются в качестве молекул-предшественников, которые подвергаются «редактированию» — удалению ненужных частей с помощью РНК-белковых комплексов[27].
Например, у кишечной палочки гены рРНК расположены в составе одного оперона (в rrnB порядок расположения такой: 16S — tRNAGlu 2 — 23S —5S) считываются в виде одной длинной молекулы, которая затем подвергается расщеплению в нескольких участках с образованием сначала пре-рРНК, а затем зрелых молекул рРНК[28]. Процесс изменения нуклеотидной последовательности РНК после синтеза носит название процессинга или редактирования РНК.
После завершения транскрипции РНК часто подвергается модификациям (см. выше), которые зависят от функции, выполняемой данной молекулой. У эукариот процесс «созревания» РНК, то есть её подготовки к синтезу белка, часто включает сплайсинг: удаление некодирующих белок последовательностей (интронов) с помощью рибонуклеопротеида сплайсосомы. Затем к 5′ концу молекулы пре-мРНК эукариот добавляется особый модифицированный нуклеотид (кэп), а к 3′ концу несколько аденинов, так называемый «полиА-хвост»[27].
Типы РНК
 Структура молоточкового (hammerhead) рибозима, который расщепляет РНК
Структура молоточкового (hammerhead) рибозима, который расщепляет РНКМатричная (информационная) РНК — РНК, которая служит посредником при передаче информации, закодированной в ДНК к рибосомам, молекулярным машинам, синтезирующим белки живого организма. Кодирующая последовательность мРНК определяет последовательность аминокислот полипептидной цепи белка[29]. Однако подавляющее большинство РНК не кодируют белок. Эти некодирующие РНК могут транскрибироваться с отдельных генов (например, рибосомальные РНК) или быть производными интронов[30]. Классические, хорошо изученные типы некодирующих РНК — это транспортные РНК (тРНК) и рРНК, которые участвуют в процессе трансляции[31]. Существуют также классы РНК, ответственные за регуляцию генов, процессинг мРНК и другие роли. Кроме того, есть и молекулы некодирующих РНК, способные катализировать химические реакции, такие, как разрезание и лигирование молекул РНК[32]. По аналогии с белками, способными катализировать химические реакции — энзимами (ферментами), каталитические молекулы РНК называются рибозимами.
Участвующие в трансляции
Роль разных типов РНК в синтезе белка (по Уотсону)Информация о последовательности аминокислот белка содержится в мРНК. Три последовательных нуклеотида (кодон) соответствуют одной аминокислоте. В эукариотических клетках транскрибированный предшественник мРНК или пре-мРНК процессируется с образованием зрелой мРНК. Процессинг включает удаление некодирующих белок последовательностей (интронов). После этого мРНК экспортируется из ядра в цитоплазму, где к ней присоединяются рибосомы, транслирующие мРНК с помощью соединённых с аминокислотами тРНК.
В безъядерных клетках (бактерии и археи) рибосомы могут присоединяться к мРНК сразу после транскрипции участка РНК. И у эукариот, и у прокариот цикл жизни мРНК завершается её контролируемым разрушением ферментами рибонуклеазами[29].
Транспортные (тРНК) — малые, состоящие из приблизительно 80 нуклеотидов, молекулы с консервативной третичной структурой. Они переносят специфические аминокислоты в место синтеза пептидной связи в рибосоме. Каждая тРНК содержит участок для присоединения аминокислоты и антикодон для узнавания и присоединения к кодонам мРНК. Антикодон образует водородные связи с кодоном, что помещает тРНК в положение, способствующее образованию пептидной связи между последней аминокислотой образованного пептида и аминокислотой, присоединённой к тРНК[30].
Рибосомальные РНК (рРНК) — каталитическая составляющая рибосом. Эукариотические рибосомы содержат четыре типа молекул рРНК: 18S, 5.8S, 28S и 5S. Три из четырёх типов рРНК синтезируются в ядрышке. В цитоплазме рибосомальные РНК соединяются с рибосомальными белками и формируют нуклеопротеин, называемый рибосомой[29]. Рибосома присоединяется к мРНК и синтезирует белок. рРНК составляет до 80 % РНК, обнаруживаемой в цитоплазме эукариотической клетки[33].
Необычный тип РНК, который действует в качестве тРНК и мРНК (тмРНК) обнаружен во многих бактериях и пластидах. При остановке рибосомы на дефектных мРНК без стоп-кодонов тмРНК присоединяет небольшой пептид, направляющий белок на деградацию[34].
Участвующие в регуляции генов
В живых клетках обнаружено несколько типов РНК, которые могут уменьшать степень выражения гена при комплементарности мРНК или самому гену. Микро-РНК (21-22 нуклеотида в длину) найдены у эукариот и оказывают воздействие через механизм РНК-интерференции. При этом комплекс микро-РНК и ферментов может приводить к метилированию нуклеотидов в ДНК промотора гена, что служит сигналом для уменьшения активности гена. При использовании другого типа регуляции мРНК, комплементарная микро-РНК, деградируется[35]. Однако есть и миРНК, которые увеличивают, а не уменьшают экспрессию генов[36]. Малые интерферирующие РНК (миРНК, 20-25 нуклеотидов) часто образуются в результате расщепления вирусных РНК, но существуют и эндогенные клеточные миРНК[37]. Малые интерферирующие РНК также действуют через РНК-интерференцию по сходным с микро-РНК механизмам[38]. У животных найдены так называемыме РНК, взаимодействующие с Piwi (piРНК, 29-30 нуклеотидов), противодействующие в половых клетках увеличению числа копий транспозонов и играющие роль в образовании гамет[39][40]. Кроме того, piRNA могут эпигенетически наследоваться по материнской линии, передавая потомству своё свойство ингибировать экспрессию транспозонов[41].
Антисмысловые РНК широко распространены у бактерий, многие из них подавляют выражение генов, но некоторые активируют экспрессию[42]. Действуют антисмысловые РНК, присоединяясь к мРНК, что приводит к образованию двуцепочечных молекул РНК, которые деградируются ферментами[43]. У эукариот обнаружены высокомолекулярные, мРНК-подобные молекулы РНК, не кодирующие белков. Эти молекулы также регулируют выражение генов[44]. В качестве примера можно привести Xist, присоединяющуюся и инактивирующую одну из двух X-хромосом у самок млекопитающих[45].
Кроме роли отдельных молекул в регуляции генов, регуляторные элементы могут формироваться в 5′ и 3′ нетранслируемых участках мРНК. Эти элементы могут действовать самостоятельно, предотвращая инициацию трансляции, либо присоединять белки, например, ферритин или малые молекулы, например, биотин[46].
В процессинге РНК
Многие РНК принимают участие в модификации других РНК. Интроны вырезаются из пре-мРНК сплайсосомами, которые, кроме белков, содержат несколько малых ядерных РНК (мяРНК)[31]. Кроме того, интроны могут катализировать собственное вырезание[47]. Синтезированная в результате транскрипции РНК также может быть химически модифицирована. У эукариот химические модификации нуклеотидов РНК, например, их метилирование, выполняется малыми ядерными РНК (мяРНК, 60-300 нуклеотидов). Этот тип РНК локализуется в ядрышке и тельцах Кахаля[30]. После ассоциации мяРНК с ферментами, мяРНК связываются с РНК-мишенью путём образования пар между основаниями двух молекул, а ферменты модифицируют нуклеотиды РНК-мишени. Рибосомальные и транспортные РНК содержат много подобных модификаций, конкретное положение которых часто сохраняется в процессе эволюции. Также могут быть модифицированы мяРНК и сами мяРНК[48][49]. Гидовые РНК осуществляют процесс редактирования РНК в кинетопласте — особом участке митохондрии протистов-кинетопластид (например, трипаносом).
Геномы, состоящие из РНК
 Жизненный цикл вируса с РНК геномом на примере полиовируса: 1 — присоединение исходного вириона к рецептору; 2 — вирион попадает в клетку; 3 — трансляция белков вируса с его РНК с образованием полипетида; 4 — полимеразы вируса размножают его РНК
Жизненный цикл вируса с РНК геномом на примере полиовируса: 1 — присоединение исходного вириона к рецептору; 2 — вирион попадает в клетку; 3 — трансляция белков вируса с его РНК с образованием полипетида; 4 — полимеразы вируса размножают его РНККак и ДНК, РНК может хранить информацию о биологических процессах. РНК может использоваться в качестве генома вирусов и вирусоподобных частиц. РНК-геномы можно разделить на те, которые не имеют промежуточной стадии ДНК и те, которые для размножения копируются в ДНК-копию и обратно в РНК (ретровирусы).
РНК-содержащие вирусы
Многие вирусы, например, вирус гриппа, на всех стадиях содержат геном, состоящий исключительно из РНК. РНК содержится внутри обычно белковой оболочки и реплицируется с помощью закодированных в ней РНК-зависимых РНК-полимераз. Вирусные геномы, состоящие из РНК разделяются на
- содержащие «плюс-цепь РНК», которая используется в качестве и мРНК, и генома;
- «минус-цепь РНК», которая служит только геномом, а в качестве мРНК используется комплементарная ей молекула;
- двухцепоченые вирусы.
Вироиды — другая группа патогенов, содержащих РНК-геном и не содержащих белок. Они реплицируются РНК-полимеразами организма хозяина[50].
Ретровирусы и ретротранспозоны
У других вирусов РНК-геном есть в течение только одной из фаз жизненного цикла. Вирионы так называемых ретровирусов содержат молекулы РНК, которые при попадании в клетки хозяина служат матрицей для синтеза ДНК-копии. В свою очередь, с матрицы ДНК считывается РНК-геном. Кроме вирусов обратную транскрипцию применяют и класс мобильных элементов генома — ретротранспозоны[51].
Гипотеза РНК-мира
Мир РНК — гипотетический этап эволюционной истории жизни на Земле, в котором самореплицирующиеся молекулы РНК размножались до эволюции ДНК и белков. Концепция мира РНК была предложена в 1962 году Александром Ричем<[52], термин был придуман Вальтером Гилбертом в 1986 году[53]. Кроме мира РНК, были предложены и другие химические пути возникновения жизни [54], и жизнь на основе РНК, возможно, не была первой[53][55]. Тем не менее, найдено достаточно доказательств возможности существования мира РНК, поэтому гипотеза получила широкое признание[52][56][57].
Подобно ДНК, РНК может хранить и реплицировать генетическую информацию, в виде ферментов — рибозимов она может катализировать (начинать или ускорять) химические реакции, которые имеют решающее значение для существования жизни [58]. Один из наиболее важных компонентов клетки, рибосомы, состоят, в основном, из РНК. Рибонуклеотидные фрагменты во многих коферментах, таких как ацетил-КоА, NADH, FADH и F420, уже давно считаются сохранившимися остатками ковалентно связанных коферментов в мире РНК [59]. Если РНК-мир существовал, то вероятно, за ним следовала стадия эволюции рибонуклеопротеинов (мир РНП) [60], которым, в свою очередь, наследовали ДНК и более длинные белки. Причина, по которой ДНК стала преобладающей молекулой хранения генетической информации, может быть связана с тем, что она более стабильна и долговечна, чем РНК [60]. Ферменты-белки, возможно, пришли на смену рибозимам на основе РНК в качестве биокатализаторов, поскольку разнообразие мономеров (аминокислот) делают их более универсальными. Поскольку некоторые кофакторы содержат как нуклеотидные, так и аминокислотные характеристики, может быть, аминокислоты, пептиды и, наконец, белки первоначально были сопутствующими факторами рибозимов[59].
См. также
Примечания
- ↑ Dahm R (2005). «Friedrich Miescher and the discovery of DNA». Developmental Biology 278 (2): 274–88. PMID 15680349.
- ↑ Nierhaus KH, Wilson DN. Protein Synthesis and Ribosome Structure. — Wiley-VCH, 2004. — С. 3. — ISBN 3-527-30638-2.
- ↑ Carlier M. L’ADN, cette «simple» molécule. Esprit libre (июнь 2003). Проверено ???. Архивировано 23 августа 2011 года.
- ↑ А. С. Спирин. Биоорганическая химия. — М.: Высшая школа, 1986. — С. 10.
- ↑ Ochoa S. Enzymatic synthesis of ribonucleic acid. Nobel Lecture (1959). Проверено ???. Архивировано 23 августа 2011 года.
- ↑ Holley RW et al. Structure of a ribonucleic acid // Science. — 1965. — Vol. 147, № 1664. — P. 1462–65. — DOI:10.1126/science.147.3664.1462.
- ↑ Szathmáry E. The origin of the genetic code: amino acids as cofactors in an RNA world // Trends Genet.. — 1999. — Vol. 15, № 6. — P. 223–9. — DOI:10.1016/S0168-9525(99)01730-8.
- ↑ Fiers W et al. Complete nucleotide-sequence of bacteriophage MS2-RNA: primary and secondary structure of replicase gene // Nature. — 1976. — Vol. 260. — P. 500–7. — PMID 1264203.
- ↑ Napoli C, Lemieux C, Jorgensen R. Introduction of a chimeric chalcone synthase gene into petunia results in reversible co-suppression of homologous genes in trans. // Plant Cell. — 1990. — Vol. 2, № 4. — P. 279–89. — PMID 12354959.
- ↑ Ruvkun G. Glimpses of a tiny RNA world // Science. — 2001. — Vol. 294, № 5543. — P. 797–99. — DOI:10.1126/science.1066315.
- ↑ Jankowski JAZ, Polak JM. Clinical gene analysis and manipulation: tools, techniques and troubleshooting. — Cambridge University Press, 1996. — P. 14. — ISBN 0521478960.
- ↑ Kiss T (2001). «Small nucleolar RNA-guided post-transcriptional modification of cellular RNAs». The EMBO Journal 20: 3617–22. DOI:10.1093/emboj/20.14.3617.
- ↑ Yu Q, Morrow CD (2001). «Identification of critical elements in the tRNA acceptor stem and TΨC loop necessary for human immunodeficiency virus type 1 infectivity». J Virol. 75 (10): 4902–6. DOI:10.1128/JVI.75.10.4902-4906.2001.
- ↑ King TH, Liu B, McCully RR, Fournier MJ (2002). «Ribosome structure and activity are altered in cells lacking snoRNPs that form pseudouridines in the peptidyl transferase center». Molecular Cell 11 (2): 425–35. DOI:10.1016/S1097-2765(03)00040-6.
- ↑ Barciszewski J, Frederic B, Clark C. RNA biochemistry and biotechnology. — Springer, 1999. — P. 73–87. — ISBN 0792358627.
- ↑ Lee JC, Gutell RR (2004). «Diversity of base-pair conformations and their occurrence in rRNA structure and RNA structural motifs». J. Mol. Biol. 344 (5): 1225–49. DOI:10.1016/j.jmb.2004.09.072. PMID 15561141.
- ↑ Salazar M, Fedoroff OY, Miller JM, Ribeiro NS, Reid BR (1992). «The DNA strand in DNAoRNA hybrid duplexes is neither B-form nor A-form in solution». Biochemistry 1993 (32): 4207–15. PMID 7682844.
- ↑ Hermann T, Patel DJ (2000). «RNA bulges as architectural and recognition motifs». Structure 8 (3): R47–R54. DOI:10.1016/S0969-2126(00)00110-6.
- ↑ Mikkola S, Nurmi K, Yousefi-Salakdeh E, Strömberg R, Lönnberg H (1999). «The mechanism of the metal ion promoted cleavage of RNA phosphodiester bonds involves a general acid catalysis by the metal aquo ion on the departure of the leaving group». Perkin transactions 2: 1619–26. DOI:10.1039/a903691a.
- ↑ Mathews DH, Disney MD, Childs JL, Schroeder SJ, Zuker M, Turner DH (2004). «Incorporating chemical modification constraints into a dynamic programming algorithm for prediction of RNA secondary structure». Proc. Natl. Acad. Sci. USA 101 (19): 7287–92. DOI:10.1073/pnas.0401799101.
- ↑ Redirect
- ↑ Spriggs KA, Stoneley M, Bushell M, Willis AE. (2008). «Re-programming of translation following cell stress allows IRES-mediated translation to predominate». Biol Cell. 100 (1): 27–38.
- ↑ Higgs PG (2000). «RNA secondary structure: physical and computational aspects». Quarterly Reviews of Biophysics 33: 199–253. DOI:10.1017/S0033583500003620.
- ↑ Nissen P, Hansen J, Ban N, Moore PB, Steitz TA (2000). «The structural basis of ribosome activity in peptide bond synthesis». Science 289 (5481): 920–30. DOI:10.1126/science.289.5481.920.
- ↑ Jeffrey L Hansen, Alexander M Long, Steve C Schultz (1997). «Structure of the RNA-dependent RNA polymerase of poliovirus». Structure 5 (8): 1109–22. DOI:10.1016/S0969-2126(97)00261-X.
- ↑ Ahlquist P (2002). «RNA-Dependent RNA Polymerases, Viruses, and RNA Silencing». Science 296 (5571): 1270–73. DOI:10.1126/science.1069132.
- ↑ 1 2 Alberts, Bruce. Molecular Biology of the Cell; Fourth Edition. — New York and London : Garland Science, 2002. — P. 302–303. — ISBN ISBN 0-8153-3218-1.
- ↑ Wagner R., Theissen G., Zacharias. Regulation of Ribosomal RNA synthesis and Control of ribosome Formation in E.coli. — 1993. — P. 119–129.
- ↑ 1 2 3 Cooper GC, Hausman RE. The Cell: A Molecular Approach. — 3rd edition. — Sinauer, 2004. — P. 261–76, 297, 339–44. — ISBN 0-87893-214-3.
- ↑ 1 2 3 Wirta W. Mining the transcriptome – methods and applications. — 2006. — ISBN 91-7178-436-5.
- ↑ 1 2 Berg JM, Tymoczko JL, Stryer L. Biochemistry. — 5th edition. — WH Freeman and Company, 2002. — P. 118–19, 781–808. — ISBN 0-7167-4684-0.
- ↑ Rossi JJ (2004). «Ribozyme diagnostics comes of age». Chemistry & Biology 11 (7): 894–95. DOI:10.1016/j.chembiol.2004.07.002.
- ↑ Kampers T, Friedhoff P, Biernat J, Mandelkow E-M, Mandelkow E (1996). «RNA stimulates aggregation of microtubule-associated protein tau into Alzheimer-like paired helical filaments». FEBS Letters 399: 98–100, 344–49. PMID 8985176.
- ↑ Gueneau de Novoa P, Williams KP (2004). «The tmRNA website: reductive evolution of tmRNA in plastids and other endosymbionts». Nucleic Acids Res. 32 (Database issue): D104-8. DOI:10.1093/nar/gkh202. PMID 14681369.
- ↑ Matzke MA, Matzke AJM (2004). «Planting the seeds of a new paradigm». PLoS Biology 2 (5): e133. DOI:10.1371/journal.pbio.0020133. PMID 15138502.
- ↑ Check E (2007). «RNA interference: hitting the on switch». Nature 448 (7156): 855–58. DOI:10.1038/448855a. PMID 17713502.
- ↑ Vazquez F, Vaucheret H, Rajagopalan R, Lepers C, Gasciolli V, Mallory AC, Hilbert J, Bartel DP, Crété P (2004). «Endogenous trans-acting siRNAs regulate the accumulation of Arabidopsis mRNAs». Molecular Cell 16 (1): 69–79. DOI:10.1016/j.molcel.2004.09.028. PMID 15469823.
- ↑ Doran G (2007). «RNAi – Is one suffix sufficient?». Journal of RNAi and Gene Silencing 3 (1): 217–19.
- ↑ Horwich MD, Li C Matranga C, Vagin V, Farley G, Wang P, Zamore PD (2007). «The Drosophila RNA methyltransferase, DmHen1, modifies germline piRNAs and single-stranded siRNAs in RISC». Current Biology 17: 1265–72. DOI:10.1016/j.cub.2007.06.030. PMID 17604629.
- ↑ Girard A, Sachidanandam R, Hannon GJ, Carmell MA (2006). «A germline-specific class of small RNAs binds mammalian Piwi proteins». Nature 442: 199–202. DOI:10.1038/nature04917. PMID 16751776.
- ↑ Brennecke J, Malone CD, Aravin AA, Sachidanandam R, Stark A, Hannon GJ (November 2008). «An epigenetic role for maternally inherited piRNAs in transposon silencing». Science (journal) 322 (5906): 1387–92. DOI:10.1126/science.1165171. PMID 19039138.
- ↑ Wagner EG, Altuvia S, Romby P (2002). «Antisense RNAs in bacteria and their genetic elements». Adv Genet. 46: 361–98. PMID 11931231.
- ↑ Gilbert SF. Developmental Biology. — 7th ed. — Sinauer, 2003. — P. 101–3. — ISBN 0878932585.
- ↑ Hüttenhofer A, Schattner P, Polacek N (2005). «Non-coding RNAs: hope or hype?». Trends Genet. 21 (5): 289–97. DOI:10.1016/j.tig.2005.03.007. PMID 15851066.
- ↑ Heard E, Mongelard F, Arnaud D, Chureau C, Vourc’h C, Avner P (1999). «Human XIST yeast artificial chromosome transgenes show partial X inactivation center function in mouse embryonic stem cells». Proc. Natl. Acad. Sci. USA 96 (12): 6841–46. DOI:10.1073/pnas.96.12.6841. PMID 10359800.
- ↑ Batey RT (2006). «Structures of regulatory elements in mRNAs». Curr. Opin. Struct. Biol. 16 (3): 299–306. DOI:10.1016/j.sbi.2006.05.001. PMID 16707260.
- ↑ Steitz TA, Steitz JA (1993). «A general two-metal-ion mechanism for catalytic RNA». Proc. Natl. Acad. Sci. U.S.A. 90 (14): 6498–502. DOI:10.1073/pnas.90.14.6498. PMID 8341661.
- ↑ Covello PS, Gray MW (1989). «RNA editing in plant mitochondria». Nature 341: 662–66. DOI:10.1038/341662a0. PMID 2552326.
- ↑ Omer AD, Ziesche S, Decatur WA, Fournier MJ, Dennis PP (2003). «RNA-modifying machines in archaea». Molecular Microbiology 48 (3): 617–29. DOI:10.1046/j.1365-2958.2003.03483.x. PMID 12694609.
- ↑ Daròs JA, Elena SF, Flores R (2006). «Viroids: an Ariadne’s thread into the RNA labyrinth». EMBO Rep. 7 (6): 593–8. DOI:10.1038/sj.embor.7400706. PMID 16741503.
- ↑ Kalendar R, Vicient CM, Peleg O, Anamthawat-Jonsson K, Bolshoy A, Schulman AH (2004). «Large retrotransposon derivatives: abundant, conserved but nonautonomous retroelements of barley and related genomes». Genetics 166 (3): D339. DOI:10.1534/genetics.166.3.1437. PMID 15082561.
- ↑ 1 2 (Apr 2013) «The «strong» RNA world hypothesis: fifty years old». Astrobiology 13 (4): 391–403. DOI:10.1089/ast.2012.0868. PMID 23551238. Bibcode: 2013AsBio..13..391N. “[The RNA world’s existence] has broad support within the community today.”
- ↑ 1 2 (Jul 2012) «The RNA worlds in context». Cold Spring Harbor Perspectives in Biology 4 (7): a006742. DOI:10.1101/cshperspect.a006742. PMID 21441585.
- ↑ (Apr 2015) «Common origins of RNA, protein and lipid precursors in a cyanosulfidic protometabolism». Nature Chemistry 7 (4): 301–7. DOI:10.1038/nchem.2202. PMID 25803468. Bibcode: 2015NatCh…7..301P.
- ↑ (May 2012) «The origins of the RNA world». Cold Spring Harbor Perspectives in Biology 4 (5): a003608. DOI:10.1101/cshperspect.a003608. PMID 20739415.
- ↑ Wade, Nicholas. Making Sense of the Chemistry That Led to Life on Earth, New York Times (May 4, 2015). Архивировано 9 июля 2017 года. Проверено 10 мая 2015.
- ↑ (Dec 2007) «The origin of the RNA world: co-evolution of genes and metabolism». Bioorganic Chemistry 35 (6): 430–43. DOI:10.1016/j.bioorg.2007.08.001. PMID 17897696. “The proposal that life on Earth arose from an RNA World is widely accepted.”
- ↑ Zimmer, Carl. A Tiny Emissary from the Ancient Past, New York Times (September 25, 2014). Архивировано 27 сентября 2014 года. Проверено 26 сентября 2014.
- ↑ 1 2 (2011) «Small Cofactors May Assist Protein Emergence from RNA World: Clues from RNA-Protein Complexes». PLoS One 6. DOI:10.1371/journal.pone.0022494. PMID 21789260.
- ↑ 1 2 Garwood, Russell J. (2012). «Patterns In Palaeontology: The first 3 billion years of evolution». Palaeontology Online 2 (11): 1–14. Проверено June 25, 2015.
Литература
Ссылки
(РНК), нуклеиновой кислоты, универсально распространенной в природе, которая содержит рибозу в качестве углеводного компонента и аденин и гуанин (пуриновые основания) и урацил и цитозин (пиримидиновые основания) в качестве азотистых оснований. Некоторые другие производные пурина и пиримидина также обнаруживаются в небольших количествах в РНК.
РНК являются линейными полинуклеотидами, которые имеют цепи, состоящие из нескольких десятков до десятков тысяч нуклеотидов и молекулярных масс, которые варьируются от 10-20 × 10 3 до 5-6 × 10 6 .Каждая отдельная РНК имеет определенную последовательность нуклеотидов. В организме РНК находится в основном в рибонуклеопротеинах, которые представляют собой комплексы РНК с белками.
РНК очень важна биологически, потому что она участвует в реализации генетической информации и в биосинтезе белков во всех живых организмах. Макромолекулярная структура РНК в основном состоит из одноцепочечных полинуклеотидных цепей, которые образуют двухспиральные сегменты в соответствии с принципом комплементарности оснований.Многие вирусы содержат РНК в качестве своего единственного нуклеинового компонента; РНК может служить матрицей для биосинтеза не только РНК, но и дезоксирибонуклеиновой кислоты (ДНК), процесс, известный как обратная транскрипция.
РНК биосинтезируется в основном в ядре клетки из рибонуклеозидтрифосфатов под действием полимераз либо на матрицу ДНК (ДНК-зависимые РНК-полимеразы), либо, в некоторых вирусах, на матрицу РНК (РНК-зависимые РНК-полимеразы).
В клетках бактерий, животных и растений различные типы РНК выполняют различные биологические функции.Они также различаются по структуре и метаболизму. Далее следует обсуждение наиболее важных типов РНК.
Рибосомная РНК (рРНК) обнаружена в рибосомах и составляет основную часть клеточной РНК; он представлен РНК с константами седиментации (S) 23S, 16S и 5S. В кишечной палочке первичная структура рРНК, то есть последовательность нуклеотидов, полностью установлена для 5S РНК, почти полностью установлена для 16S РНК и частично установлена для 23S РНК.У разных типов организмов размеры и структура рРНК различны. Биологическая роль рРНК до конца не выяснена. Целостность молекул рРНК необходима для биосинтеза белков в рибосомах.
Передающая РНК (тРНК) имеет константу седиментации около 4S и молекулярную массу около 25000. Он обладает относительно низким полимеризмом, имеет около 80 нуклеотидных остатков и содержит много метилированных и других минорных оснований. Биологическая роль тРНК заключается в связывании активированных аминокислотных остатков и переносе их на рибосомы, то есть на сайт синтеза полипептидных цепей.Каждая аминокислота имеет свою специфическую тРНК, обычно более одной. Трансферная РНК имеет сложную, частично двухспиральную макромолекулярную структуру, которая представлена в форме листа клевера. Этот тип РНК содержит сегменты, которые присоединены к рибосоме — триплету нуклеотидов или антикодону — который связан с кодоном мессенджерной РНК (мРНК), и концевой сегмент, который связывает аминокислотный остаток. Первичные структуры из более чем 60 тРНК были полностью созданы.
Messenger RNA — самая разнообразная группа РНК.Он выступает в качестве матрицы в биосинтезе белков в процессе трансляции, который представляет собой формирование определенной последовательности аминокислот в полипептидной цепи белка на основе нуклеотидного кода. РНК синтезируются в клетке на матрице ДНК и образуют последовательность рибонуклеотидов, комплементарных последовательности дезоксирибонуклеотидов в ДНК (процесс транскрипции). Гигантские молекулы, которые являются предшественниками мРНК, были обнаружены в ядре клетки. Большинство из этих гигантских молекул разлагаются внутри ядра, и только относительно небольшая часть каждой молекулы переносится в цитоплазму и образует фактическую мРНК.
РНК, которая быстро разлагается в ядре клетки, вероятно, участвует в процессах регуляции. Также были найдены некоторые другие типы РНК, например, относительно стабильная нуклеиновая РНК, которая проявляет низкий уровень полимеризации — ее функция до сих пор неясна.
СПИСОК ЛИТЕРАТУРЫ
Дэвидсон Дж. Биохимия нуклеиновых кислот. Москва, 1968. (пер. С англ.)Химия и биохимия нуклеиновых кислот. Под редакцией И. Б. Збарского и С. С. Дебова.Ленинград, 1968.
Спирин, А. С. и Л. П. Гаврилова. Рибосома , 2-е изд. Москва, 1971.
Дарнелл Дж. «Рибонуклеиновые кислоты из клеток животных». Бактериологические обзоры , 1968, вып. 32, нет 3, с. 262.
Аттарди Дж. И Ф. Амальди. «Структура и синтез рибосомальной РНК». Ежегодный обзор биохимии , 1970, том. 39, стр. 183.
Винберг Р. А. «Метаболизм ядерной РНК». Ежегодный обзор биохимии , 1973, том. 42, стр. 329..
Нуклеиновая кислота , химическое соединение природного происхождения, способное расщепляться с образованием фосфорной кислоты, сахаров и смеси органических оснований (пуринов и пиримидинов). Нуклеиновые кислоты являются основными переносящими информацию молекулами клетки, и, направляя процесс синтеза белка, они определяют наследственные характеристики каждого живого существа. Двумя основными классами нуклеиновых кислот являются дезоксирибонуклеиновая кислота (ДНК) и рибонуклеиновая кислота (РНК).ДНК является основным планом жизни и составляет генетический материал всех свободно живущих организмов и большинства вирусов. РНК является генетическим материалом некоторых вирусов, но она также обнаружена во всех живых клетках, где она играет важную роль в определенных процессах, таких как производство белков.
полинуклеотидной цепи дезоксирибонуклеиновой кислоты (ДНК) Часть полинуклеотидной цепи дезоксирибонуклеиновой кислоты (ДНК). На вставке показаны соответствующие пентозный сахар и пиримидиновое основание в рибонуклеиновой кислоте (РНК). Encyclopædia Britannica, Inc.Популярные вопросы
Что такое нуклеиновые кислоты?
Нуклеиновые кислоты — это природные химические соединения, которые служат первичными переносящими информацию молекулами в клетках. Они играют особенно важную роль в управлении синтезом белка. Двумя основными классами нуклеиновых кислот являются дезоксирибонуклеиновая кислота (ДНК) и рибонуклеиновая кислота (РНК).
Какова основная структура нуклеиновой кислоты?
Нуклеиновые кислоты — это длинные цепочечные молекулы, состоящие из ряда почти идентичных строительных блоков, называемых нуклеотидами.Каждый нуклеотид состоит из азотсодержащего ароматического основания, присоединенного к пентозному (пятиуглеродному) сахару, который, в свою очередь, присоединен к фосфатной группе.
Какие азотсодержащие основания встречаются в нуклеиновых кислотах?
Каждая нуклеиновая кислота содержит четыре из пяти возможных азотсодержащих оснований: аденин (A), гуанин (G), цитозин (C), тимин (T) и урацил (U). А и G подразделяются на пурины, а С, Т и U называются пиримидинами. Все нуклеиновые кислоты содержат основания A, C и G; T, однако, обнаруживается только в ДНК, а U — в РНК.
Когда были обнаружены нуклеиновые кислоты?
В этой статье рассматривается химия нуклеиновых кислот, описываются структуры и свойства, которые позволяют им служить в качестве передатчиков генетической информации. Для обсуждения генетического кода см. Наследственность , а для обсуждения роли, которую играют нуклеиновые кислоты в синтезе белка, см. Метаболизм .
Нуклеотиды: строительные блоки нуклеиновых кислот
Базовая структура
Нуклеиновые кислоты — это полинуклеотиды, то есть длинные цепочечные молекулы, состоящие из серии почти идентичных строительных блоков, называемых нуклеотидами.Каждый нуклеотид состоит из азотсодержащего ароматического основания, присоединенного к пентозному (пятиуглеродному) сахару, который, в свою очередь, присоединен к фосфатной группе. Каждая нуклеиновая кислота содержит четыре из пяти возможных азотсодержащих оснований: аденин (A), гуанин (G), цитозин (C), тимин (T) и урацил (U). A и G подразделяются на пурины, а C, T и U вместе называются пиримидинами. Все нуклеиновые кислоты содержат основания A, C и G; T, однако, обнаруживается только в ДНК, а U — в РНК. Пентозный сахар в ДНК (2′-дезоксирибоза) отличается от сахара в РНК (рибозе) отсутствием гидроксильной группы (―OH) на 2′-углероде сахарного кольца.Без присоединенной фосфатной группы сахар, присоединенный к одному из оснований, известен как нуклеозид. Фосфатная группа соединяет последовательные сахарные остатки, соединяя 5′-гидроксильную группу на одном сахаре с 3′-гидроксильной группой следующего сахара в цепи. Эти нуклеозидные связи называются фосфодиэфирными связями и одинаковы в РНК и ДНК.
Нуклеотиды синтезируются из легко доступных предшественников в клетке. Рибозофосфатная часть как пуриновых, так и пиримидиновых нуклеотидов синтезируется из глюкозы через пентозофосфатный путь.Шестиатомное пиримидиновое кольцо синтезируется первым, а затем присоединяется к рибозофосфату. Два кольца в пуринах синтезируются при присоединении к рибозофосфату во время сборки адениновых или гуаниновых нуклеозидов. В обоих случаях конечным продуктом является нуклеотид, несущий фосфат, присоединенный к 5′-углероду на сахаре. Наконец, специализированный фермент, называемый киназой, добавляет две фосфатные группы, используя аденозинтрифосфат (АТФ) в качестве донора фосфата, с образованием рибонуклеозидтрифосфата, непосредственного предшественника РНК.Что касается ДНК, 2′-гидроксильная группа удаляется из рибонуклеозиддифосфата с получением дезоксирибонуклеозиддифосфата. Дополнительная фосфатная группа из АТФ затем добавляется другой киназой с образованием дезоксирибонуклеозидтрифосфата, непосредственного предшественника ДНК.
Получите эксклюзивный доступ к контенту из нашего первого издания 1768 года с вашей подпиской. Подпишитесь сегодняВо время нормального клеточного метаболизма РНК постоянно производится и расщепляется. Остатки пурина и пиримидина повторно используются несколькими путями спасения, чтобы сделать больше генетического материала.Пурин спасен в виде соответствующего нуклеотида, тогда как пиримидин спасен в виде нуклеозида.
- Образование
- Наука
- Биология
- Какова роль нуклеиновых кислот в живых существах?
Нуклеиновые кислоты — это большие молекулы, несущие тонны мелких деталей: вся генетическая информация. Нуклеиновые кислоты находятся в каждом живом организме — растениях, животных, бактериях, вирусах, грибах — которые используют и преобразуют энергию. У каждого живого существа есть что-то общее.
Люди, животные, растения и многое другое связаны генетическим материалом. Все живые существа могут выглядеть по-разному и действовать по-разному, но в глубине — глубоко в ядре клеток — живые существа содержат одни и те же химические «ингредиенты», образующие очень похожий генетический материал.
Существует два типа нуклеиновых кислот: ДНК (что означает дезоксирибонуклеиновая кислота) и РНК (что означает рибонуклеиновая кислота). Нуклеиновые кислоты состоят из нитей нуклеотидов , которые состоят из основания, содержащего азот (называемый азотистым основанием ), сахара, содержащего молекулы с пятью атомами углерода, и фосфорной кислоты.
Весь ваш генетический состав, личность, возможно, даже интеллект зависят от молекул, содержащих соединение азота, немного сахара и кислоты. Азотистые основания представляют собой молекулы, называемые пуринами или пиримидинами.
пуринов включают
пиримидинов включают
Цитозин
Тимин (в РНК)
урацил (в ДНК)
Дезоксирибонуклеиновая кислота (ДНК)
ДНКсодержит две нити нуклеотидов, расположенных таким образом, что она выглядит как витая лестница (так называемая двойная спираль ).Азотистыми основаниями, на которых ДНК строит свою двойную спираль, являются аденин (A), гуанин (G), цитозин (C) и тимин (T). Сахар в составе ДНК — 2-дезоксирибоза.
Аденин всегда в паре с тимином (A-T), а гуанин всегда в паре с цитозином (G-C). Эти основания удерживаются вместе водородными связями, которые образуют «ступеньки» «витой лестницы». Стороны лестницы составлены из молекул сахара и фосфата.
Определенные участки азотистых оснований вдоль цепи ДНК образуют ген .Ген — это единица, которая содержит генетическую информацию или коды для конкретного продукта и передает наследственную информацию следующему поколению.
Но гены встречаются не только в репродуктивных клетках. Каждая клетка в организме содержит ДНК (и, следовательно, гены), потому что ДНК также кодирует белки, которые организм производит. И белки контролируют функцию клеток и обеспечивают структуру. Итак, основа жизни происходит в каждой клетке.
Всякий раз, когда в организме создается новая клетка, генетический материал воспроизводится и помещается в новую клетку.Новая клетка может затем создавать белки внутри себя, а также передавать генетическую информацию следующей новой клетке.
Порядок азотистых оснований на цепи ДНК (или в части ДНК, которая содержит ген) определяет, какая аминокислота производится. И порядок, в котором аминокислоты связаны вместе, определяет, какой белок производится. То, какой белок вырабатывается, определяет, какой структурный элемент вырабатывается в вашем теле (например, мышечная ткань, кожа или волосы) или какую функцию можно выполнять (например, вырабатывается ли гемоглобин для доставки кислорода ко всем клеткам).
рибонуклеиновая кислота (РНК)
Азотистыми основаниями, которые использует РНК, являются аденин, гуанин, цитозин и урацил (вместо тимина). И сахар в РНК представляет собой рибозу (вместо 2-дезоксирибозы). Это основные различия между ДНК и РНК.
У большинства животных РНК не является основным генетическим материалом. Многие вирусы, такие как вирус иммунодефицита человека (ВИЧ), вызывающий СПИД, содержат РНК в качестве своего генетического материала. Тем не менее, у животных, РНК работает вместе с ДНК, чтобы производить белки, необходимые по всему организму.
Например, РНК имеет три основных подтипа: мессенджер РНК (мРНК), трансфер РНК (тРНК) и рибосомальная РНК (рРНК). Все три из этих подтипов участвуют в синтезе белка.
,Нуклеотиды и нуклеиновые кислоты
 Нуклеотиды и нуклеиновые кислоты Краткая история 1 1869 — Мишер Выделенный нуклеин из загрязненных повязок 1902 — Гаррод Изучил редкое генетическое заболевание: алкаптонурия; пришел к выводу, что специфический ген связан с
Нуклеотиды и нуклеиновые кислоты Краткая история 1 1869 — Мишер Выделенный нуклеин из загрязненных повязок 1902 — Гаррод Изучил редкое генетическое заболевание: алкаптонурия; пришел к выводу, что специфический ген связан с
Белки и нуклеиновые кислоты
 Белки и нуклеиновые кислоты Глава 5 Макромолекулы: белки Белки Наиболее структурно и функционально разнообразная группа биомолекул.: o Участвует почти во всем o Ферменты o Структура (кератин,
Белки и нуклеиновые кислоты Глава 5 Макромолекулы: белки Белки Наиболее структурно и функционально разнообразная группа биомолекул.: o Участвует почти во всем o Ферменты o Структура (кератин,
СТРУКТУРЫ ЯДЕРНЫХ КИСЛОТ
 ГЛАВА 2 СТРУКТУРЫ НУКЛЕИНОВЫХ КИСЛОТ Какова химическая структура молекулы дезоксирибонуклеиновой кислоты (ДНК)? ДНК представляет собой полимер дезоксирибонуклеотидов. Все нуклеиновые кислоты состоят из нуклеотидов как строение
ГЛАВА 2 СТРУКТУРЫ НУКЛЕИНОВЫХ КИСЛОТ Какова химическая структура молекулы дезоксирибонуклеиновой кислоты (ДНК)? ДНК представляет собой полимер дезоксирибонуклеотидов. Все нуклеиновые кислоты состоят из нуклеотидов как строение
Структура и функция ДНК
 Структура и функции ДНК ДНК и РНК Структура ДНК и РНК являются нуклеиновыми кислотами.Они состоят из химических единиц, называемых нуклеотидами. Нуклеотиды соединены сахарофосфатным остовом. Четыре
Структура и функции ДНК ДНК и РНК Структура ДНК и РНК являются нуклеиновыми кислотами.Они состоят из химических единиц, называемых нуклеотидами. Нуклеотиды соединены сахарофосфатным остовом. Четыре
Рабочий лист ДНК BIOL 1107L DNA
 Рабочий лист BIOL 1107L Название День / Время См. Главу 5 и главу 16 (рис. 16.5, 16.7, 16.8 и рисунок, встроенный в текст на стр. 310) в учебнике по биологии, 9-е издание, для получения информации и его структуры
Рабочий лист BIOL 1107L Название День / Время См. Главу 5 и главу 16 (рис. 16.5, 16.7, 16.8 и рисунок, встроенный в текст на стр. 310) в учебнике по биологии, 9-е издание, для получения информации и его структуры
ПРАКТИКА ТЕСТОВЫЕ ВОПРОСЫ
 ЧАСТЬ A: ВОПРОСЫ ДЛЯ МНОЖЕСТВА ВЫБОРА ПРАКТИКА ВОПРОСЫ ДЛЯ СИНТЕЗА ДНК И СИНТЕЗА БЕЛКОВ B 1.Одна из функций ДНК состоит в том, чтобы А. секретировать вакуоли. Б. копировать себя. C. соединяют аминокислоты друг с другом.
ЧАСТЬ A: ВОПРОСЫ ДЛЯ МНОЖЕСТВА ВЫБОРА ПРАКТИКА ВОПРОСЫ ДЛЯ СИНТЕЗА ДНК И СИНТЕЗА БЕЛКОВ B 1.Одна из функций ДНК состоит в том, чтобы А. секретировать вакуоли. Б. копировать себя. C. соединяют аминокислоты друг с другом.
Молекулы клеток
 Молекулы клеток I. Введение A. Большинство населения мира не может переваривать продукты на основе молока. 1. Эти люди не переносят лактозу, потому что им не хватает фермента лактазы. 2. Это иллюстрирует
Молекулы клеток I. Введение A. Большинство населения мира не может переваривать продукты на основе молока. 1. Эти люди не переносят лактозу, потому что им не хватает фермента лактазы. 2. Это иллюстрирует
Глава 3 Молекулы клеток
 Bio 100 Молекулы клеток 1 Глава 3 Молекулы клеток Соединения, содержащие углерод, называются органическими соединениями Молекулы, такие как метан, состоящие только из углерода и водорода, называются углеводородами
Bio 100 Молекулы клеток 1 Глава 3 Молекулы клеток Соединения, содержащие углерод, называются органическими соединениями Молекулы, такие как метан, состоящие только из углерода и водорода, называются углеводородами
3120-1 — стр. 1.Название:
 Название: 1) Какие серии расположены в правильном порядке в соответствии с уменьшающимся размером структур? A) ДНК, ядро, хромосома, нуклеотид, азотистое основание B) хромосома, ядро, азотистое основание, нуклеотид,
Название: 1) Какие серии расположены в правильном порядке в соответствии с уменьшающимся размером структур? A) ДНК, ядро, хромосома, нуклеотид, азотистое основание B) хромосома, ядро, азотистое основание, нуклеотид,
Примеры вопросов к экзамену 3
 Примеры вопросов к экзамену 3 1. Все перечисленное происходит во время прометафазы митоза в клетках животных, за исключением a.центриоли движутся к противоположным полюсам. б. ядрышко больше не видно. с.
Примеры вопросов к экзамену 3 1. Все перечисленное происходит во время прометафазы митоза в клетках животных, за исключением a.центриоли движутся к противоположным полюсам. б. ядрышко больше не видно. с.
БИОЛОГИЧЕСКИЕ МОЛЕКУЛЫ ЖИЗНИ
 БИОЛОГИЧЕСКИЕ МОЛЕКУЛЫ ЖИЗНИ C A R B O H Y D R A T E S, L I P I D S, P R O T E I N S, A N D N U C L E I C A C I D S Центр академической поддержки @ Daytona State College (Science 115, стр. 1 из 29) Carbon
БИОЛОГИЧЕСКИЕ МОЛЕКУЛЫ ЖИЗНИ C A R B O H Y D R A T E S, L I P I D S, P R O T E I N S, A N D N U C L E I C A C I D S Центр академической поддержки @ Daytona State College (Science 115, стр. 1 из 29) Carbon
12.1 Роль ДНК в наследственности
 12.1 Роль ДНК в наследственности Только за последние 50 лет ученые поняли роль ДНК в наследственности. Это понимание началось с открытия структуры ДНК. В 1952 году Розалинда Франклин
12.1 Роль ДНК в наследственности Только за последние 50 лет ученые поняли роль ДНК в наследственности. Это понимание началось с открытия структуры ДНК. В 1952 году Розалинда Франклин
Биологические молекулы:
 Биологические молекулы: все они органические (на основе углерода). Мономеры противполимеры: мономеры относятся к субъединицам, которые при полимеризации образуют более крупный полимер. Мономеры могут функционировать самостоятельно в некоторых
Биологические молекулы: все они органические (на основе углерода). Мономеры противполимеры: мономеры относятся к субъединицам, которые при полимеризации образуют более крупный полимер. Мономеры могут функционировать самостоятельно в некоторых
Лаборатория № 12: ДНК и РНК
 115 116 Понятия, которые необходимо изучить: Структура ДНК Нуклеотиды Аминокислоты Белки Генетический код Мутация Транскрипция РНК в РНК Трансляция в белок Рисунок 12. 1: Двойная спираль ДНК Введение Длинный
115 116 Понятия, которые необходимо изучить: Структура ДНК Нуклеотиды Аминокислоты Белки Генетический код Мутация Транскрипция РНК в РНК Трансляция в белок Рисунок 12. 1: Двойная спираль ДНК Введение Длинный
Глава 2 Химические принципы
 Глава 2 Химические принципы I.Химия. [Студенты должны прочитать этот раздел самостоятельно]. а. Химия — это изучение взаимодействий между атомами и молекулами. б. Атом самая маленькая единица
Глава 2 Химические принципы I.Химия. [Студенты должны прочитать этот раздел самостоятельно]. а. Химия — это изучение взаимодействий между атомами и молекулами. б. Атом самая маленькая единица
Учебное пособие по репликации
 Учебное пособие по репликации Это учебное пособие является письменной версией материала, который вы видели в блоке репликации. Самовоспроизведение — это функция жизни, которую человеческие системы имеют
Учебное пособие по репликации Это учебное пособие является письменной версией материала, который вы видели в блоке репликации. Самовоспроизведение — это функция жизни, которую человеческие системы имеют
Биохимия клеток
 Биохимия клеток 1 Молекулы на основе углерода Хотя клетка состоит в основном из воды, остальная часть клетки состоит в основном из молекул на основе углерода Органическая химия — это исследование соединений углерода Углерод
Биохимия клеток 1 Молекулы на основе углерода Хотя клетка состоит в основном из воды, остальная часть клетки состоит в основном из молекул на основе углерода Органическая химия — это исследование соединений углерода Углерод
РНК и синтез белка
 РНК и синтез белка Гены посылают сообщения клеточному механизму. РНК играет главную роль в процессе. Процесс имеет три фазы (генетическая) транскрипционная (генетическая) трансляция синтез белка. РНК-синтез
РНК и синтез белка Гены посылают сообщения клеточному механизму. РНК играет главную роль в процессе. Процесс имеет три фазы (генетическая) транскрипционная (генетическая) трансляция синтез белка. РНК-синтез
Генетический модуль B, Якорь 3
 Генетический модуль B, Якорь 3 Основные понятия: — Характеристики человека определяются факторами, которые передаются от одного поколения родителей к следующему.- Во время образования гамет аллели на
Генетический модуль B, Якорь 3 Основные понятия: — Характеристики человека определяются факторами, которые передаются от одного поколения родителей к следующему.- Во время образования гамет аллели на
Биомолекул. отражать
 отражают, что строительные блоки ребенка являются относительно простыми структурами. Однако когда они собираются вместе, они могут образовывать великолепные структуры. Сложная городская сцена справа сделана из маленькой простой
отражают, что строительные блоки ребенка являются относительно простыми структурами. Однако когда они собираются вместе, они могут образовывать великолепные структуры. Сложная городская сцена справа сделана из маленькой простой
Руководство по переводу
 Учебное пособие по переводу Это учебное пособие является письменной версией материала, который вы видели в блоке тиражирования.В переводе клетка использует генетическую информацию, содержащуюся в мрне до
Учебное пособие по переводу Это учебное пособие является письменной версией материала, который вы видели в блоке тиражирования.В переводе клетка использует генетическую информацию, содержащуюся в мрне до
Краткая история ДНК
 Краткая история ДНК Содержание Введение … 1 Структура ДНК … 2 Ссылки … 6 Видео … 6 Введение 2 апреля 1953 г. Дж. Д. Уотсон и Ф. Х. Крик опубликовали свою теперь знаменитую статью о структуре
Краткая история ДНК Содержание Введение … 1 Структура ДНК … 2 Ссылки … 6 Видео … 6 Введение 2 апреля 1953 г. Дж. Д. Уотсон и Ф. Х. Крик опубликовали свою теперь знаменитую статью о структуре
13.2 рибосомы и синтез белка
 13.2. Рибосомы и синтез белка. Введение: * Определенная последовательность оснований в ДНК содержит указания по формированию полипептида, цепи аминокислот (существует 20 различных типов аминокислот).
13.2. Рибосомы и синтез белка. Введение: * Определенная последовательность оснований в ДНК содержит указания по формированию полипептида, цепи аминокислот (существует 20 различных типов аминокислот).
2007 7.013 Набор задач 1 КЛЮЧ
 2007 7.013 Задача: 1 КЛЮЧ, подлежащий исполнению до 17:00 в пятницу, 16 февраля 2007 г.Включите ответы в поле за пределами 68-120. ПОЖАЛУЙСТА, НАПИШИТЕ СВОИ ОТВЕТЫ НА ЭТОТ ПРИНТУТ. 1. Где в эукариотической клетке ты
2007 7.013 Задача: 1 КЛЮЧ, подлежащий исполнению до 17:00 в пятницу, 16 февраля 2007 г.Включите ответы в поле за пределами 68-120. ПОЖАЛУЙСТА, НАПИШИТЕ СВОИ ОТВЕТЫ НА ЭТОТ ПРИНТУТ. 1. Где в эукариотической клетке ты
Предварительная викторина MFM
 Предварительная MFM Викторина 1. Основным носителем химической энергии во всех клетках является: A) аденозинмонофосфат B) аденозиндифосфат C) аденозин трифосфат D) гуанозин трифосфат E) карбамоилфосфат
Предварительная MFM Викторина 1. Основным носителем химической энергии во всех клетках является: A) аденозинмонофосфат B) аденозиндифосфат C) аденозин трифосфат D) гуанозин трифосфат E) карбамоилфосфат
ДНК: структура и репликация
 7 ДНК: структура и репликация РАБОТА С ЦИФРАМИ 1.В таблице 7-1, почему нет записей для первых четырех источников ткани? Для последних трех записей, что является наиболее вероятным объяснением для
7 ДНК: структура и репликация РАБОТА С ЦИФРАМИ 1.В таблице 7-1, почему нет записей для первых четырех источников ткани? Для последних трех записей, что является наиболее вероятным объяснением для
Элементы в биологических молекулах
 Глава 3: Биологические молекулы 1. Углеводы 2. Липиды 3. Белки 4. Элементы нуклеиновых кислот в биологических молекулах Биологические макромолекулы почти полностью состоят всего из 6 элементов: Углерод (C)
Глава 3: Биологические молекулы 1. Углеводы 2. Липиды 3. Белки 4. Элементы нуклеиновых кислот в биологических молекулах Биологические макромолекулы почти полностью состоят всего из 6 элементов: Углерод (C)
Углеводы, белки и липиды
 Углеводы, белки и липиды Глава 3 МАКРОМОЛЕКУЛЫ Макромолекулы: полимеры с молекулярной массой> 1000 Функциональных групп ЧЕТЫРЕ МАКРОМОЛЕКУЛЫ В ЖИЗНИ Молекулы в живых организмах: белки,
Углеводы, белки и липиды Глава 3 МАКРОМОЛЕКУЛЫ Макромолекулы: полимеры с молекулярной массой> 1000 Функциональных групп ЧЕТЫРЕ МАКРОМОЛЕКУЛЫ В ЖИЗНИ Молекулы в живых организмах: белки,
Терминология судебной экспертизы ДНК
 Терминология криминалистического анализа ДНК Генетический анализатор ABI 310 — инструмент для капиллярного электрофореза, используемый лабораториями криминалистической ДНК для разделения коротких тандемных повторяющихся (STR) локусов на основе их размера.
Терминология криминалистического анализа ДНК Генетический анализатор ABI 310 — инструмент для капиллярного электрофореза, используемый лабораториями криминалистической ДНК для разделения коротких тандемных повторяющихся (STR) локусов на основе их размера.
Генетика Тест Биология I
 Генетический тест Биология I Множественный выбор Определите выбор, который наилучшим образом дополняет утверждение или отвечает на вопрос. 1. Эксперименты Эйвери показали, что бактерии трансформируются. РНК. с. белки.
Генетический тест Биология I Множественный выбор Определите выбор, который наилучшим образом дополняет утверждение или отвечает на вопрос. 1. Эксперименты Эйвери показали, что бактерии трансформируются. РНК. с. белки.
АТОМЫ И СВЯЗЫ. облигации
 АТОМЫ И СВЯЗЫ Атомы элементов — самые простые единицы организации в мире природы.Атомы состоят из протонов (положительный заряд), нейтронов (нейтральный заряд) и электронов (отрицательный заряд).
АТОМЫ И СВЯЗЫ Атомы элементов — самые простые единицы организации в мире природы.Атомы состоят из протонов (положительный заряд), нейтронов (нейтральный заряд) и электронов (отрицательный заряд).
 v vi vii viii ix 1 2 для старшеклассников. Для этого нужно было провести исследование, чтобы найти популярный и привлекательный стиль анимации для этой возрастной группы. Третий шаг был разработать анимацию так
v vi vii viii ix 1 2 для старшеклассников. Для этого нужно было провести исследование, чтобы найти популярный и привлекательный стиль анимации для этой возрастной группы. Третий шаг был разработать анимацию так
Репликация ДНК у прокариот
 Модуль OpenStax-CNX: m44488 1 Репликация ДНК в Prokaryotes OpenStax College Это произведение подготовлено OpenStax-CNX и лицензировано по лицензии Creative Commons Attribution License 3.0 К концу этого раздела
Модуль OpenStax-CNX: m44488 1 Репликация ДНК в Prokaryotes OpenStax College Это произведение подготовлено OpenStax-CNX и лицензировано по лицензии Creative Commons Attribution License 3.0 К концу этого раздела
Leave A Comment