
Способы движения рыб
Разнообразие условий обитания рыб определяет и способы их движения. У рыб известны три способа передвижения – плавание, ползание и полет.
Плавание– основной тип движения, который осуществляется в основном за счет боковых изгибов тела и хвоста.
Различают два типа плаванияпри помощи боковых изгибов тела:
Скумбриевидный – у рыб при плавании большое значение имеет хвост, при помощи которого рыба отталкивается от воды и продвигается вперед, на долю которого приходится около 40% всей движущей силы (скумбрия, лосось).
Угревидный (змеевидный) – у рыб при движении волнообразно изгибается все тело. Это наиболее экономичный тип движения, скорость плавания при этом невелика (минога, угорь, вьюн).
Рыбы плавают с различной скоростью. Наиболее быстрой является меч-рыба, способная развивать скорость до 33 м/с (118,8 км/час), тунец плавает со скоростью до 20 м/с (72 км/час), лосось – 5 м/с (18 км/час).
Скорость движения рыб находится в определенной зависимости и от длины тела. В соответствии с этим определяется коэффициент скорости — отношение абсолютной скорости к корню квадратному от ее длины:
По скорости движения выделяют следующие группы рыб:
1)очень быстрые (меч-рыба, тунцы) – коэффициент скорости около 70;
2) быстрые (лососи, скумбрии) – 30–60;
3) умеренно быстрые (кефали, треска, сельди) – 20–30;
4) небыстрые (сазан, лещ) – 10–20;
5) медленные (бычки) – 5–10;
6) очень медленные (колюшка, луна-рыба) – 5.
Рыбы одного вида могут плавать с различной скоростью. Различают:
1. Бросковая скорость (коэффициент скорости 30–70), которая
развивается в течение очень короткого времени (при испуге, броске на добычу).
2. Крейсерская скорость (коэффициент скорости 1–4), с которой рыбы плавают в течение длительного времени.
Ползание по грунту является одним из способов передвижения рыб, которое осуществляется в основном при помощи грудных плавников и хвоста (ползун, морской черт, многопер, прыгун, морской петух). Так, прыгун живет в мангровых зарослях и значительную часть времени проводит на берегу. По суше двигается прыжками, которые совершает при помощи хвоста и грудных плавников, питается наземными беспозвоночными.
Полет (воздушное парение) свойствен немногим летучим рыбам, обитающим в пелагиали тропических и субтропических вод Мирового океана. У этих рыб длинные и широкие грудные плавники выполняют роль крыльев. Хвост с сильно развитой нижней лопастью является двигателем, дающим начальную скорость. Выскочив на поверхность воды, летучая рыба первое время скользит по водной поверхности, с нарастанием скорости движения отрывается от воды, пролетая при этом расстояние до 200 и даже 400 м.
Ход работы
1. Ознакомится с содержанием теоретического материала изложенного в методическом указании.
2. Рассмотреть формы, типы, расположение и строение плавников рыб приготовленных для лабораторной работы. Схематично изобразить лосося и выделить на схеме парные и непарные плавники. Назовите функции различных плавников.
3. Перечислите различные положения брюшных плавников и приведите примеры.
4. Перечислите и зарисуйте типы хвостовых плавников по строению и по форме и расположению относительно конца позвоночника.
5. Рассмотрите строение спинных плавников окуня, выделите не ветвистые (колючие) и ветвистые (членистые) лучи. Запишите формулу спинного плавника окуня и спинного и анального плавников серебряного карася или другой рыбы на выбор.
6. Приведите примеры рыб с различными видами плавания.
7. Используя компьютерный калькулятор, определите коэффициент скорости — отношение абсолютной скорости к корню квадратному от ее длины. При необходимости переведите скорость в км/час.
для меч-рыбы,(V=33 м/с, L= 170 см),
тунца (V=20 м/с, L= 120 см 20 м/с),
лосося – (V=33 м/с, L= 70 см).
8. Составите список рекомендуемой литературы.
Похожие статьи:
poznayka.org
Движения и перемещения у рыб

Основная форма движения у рыб — плавание. У всех рыб, имеющих веретенообразное, обычно в большей или меньшей степени сжатое с боков тело, поступательное движение совершается волнообразным изгибанием всего тела в горизонтальной плоскости.
Хвостовой плавник главным образом парализует тормозящее движение конца тела и ослабляет обратные токи. Другие непарные плавники (спинной и анальный) в большинстве случаев играют роль киля, придавая телу устойчивость, тогда как грудные и брюшные плавники служат в основном рулями глубины; с их помощью осуществляются повороты рыбы в горизонтальной плоскости. Лучшие пловцы относятся к рыбам, имеющим этот тип тела. Так, лосось плавает со скоростью 5 м/сек, тунец —6, а голубая акула — 10 м/сек. Но в отдельных случаях некоторые рыбы могут развивать еще большую скорость. Летучая рыба, например, перед тем как выскочить из воды, плывет со скоростью 18 м/сек, а меч-рыба при нанесении удара длинной верхней челюстью развивает скорость даже до 25 м/сек.
Илистый прыгун Periophthalmus

Изменение окраски камбалы в зависимости от окраски субстрата
Но у разных рыб, в связи с формой их тела и степенью развития тех или иных плавников, плавание осуществляется весьма различно. Так, у кузовков с неподвижным панцирем главную роль в движении играет хвостовой плавник, у электрического угря — волнообразные колебания анального плавника в горизонтальной плоскости, у камбал — волнообразные движения всего тела, спинного и брюшного плавников в вертикальной плоскости, у скатов — такие же колебания грудных плавников, у морского конька, плавающего в противоположность предыдущим рыбам в вертикальном положении, — усиленной работой спинного и грудных плавников и волнообразными движениями конца тела, лишенного хвостового плавника, и т. д.

Тряпичник Phyllopteryx eques (сем. морские коньки)
В течение всей жизни из всех рыб пассивно плавает, по-видимому, только луна-рыба. Но временно так же может плавать иглобрюх, когда, набрав воздух в кишечник и раздувшись подобно шару, он носится по воле волн. Рыба-прилипало, присосавшись головной присоской к какой-нибудь большой рыбе (акуле) или дну корабля, переносится ими порой на значительное расстояние. Наконец, парусник (Istiophorus greyi), выставив из воды свой очень длинный и высокий спинной плавник, при соответствующем ветре может пользоваться им как парусом.
Еще интересные статьи по теме:
www.zoofirma.ru
Способы передвижения рыб в воде
Введение
Угорь скользит меж камней и, ускоряясь, одним рывком скрывается в узкой расщелине. В центральной колонне застыла зебрасома, в то время как нахальный губанчик делает резкие выпады, хватая и поглощая небольшими порциями закуску из паразитов. Поодаль, вокруг своего домика из актиний, безучастно взирая на остальных, неспешно плавает рыба-клоун.
Как правило, мы употребляем слово «плавать» для объяснения перемещения рыб под водой, но на самом деле их движения это нечто большее. Да, рыбы плавают, но сказать, что они всего лишь плавают – чрезмерное упрощение. Они ведь так по-разному это делают. Только представьте змееобразных извивающихся рыб, бороздящих океаны тунцов и хрупких морских коньков. Все они являются подтверждением эволюционной уникальности, каждая из которых адаптирована к своему образу жизни. Превосходная обтекаемость форм позволяет рыбам рассекать плотную среду обитания, прорезать это густое вязкое вещество коим является вода.
Вероятно, было бы более справедливым называть это не просто плаванием, а классифицировать различные его формы. Во многих случаях большую часть массы тела рыбы составляют плавательные мышцы, в то время как внутренние органы, такие как сердце и печень, едва ли не собраны где-то в стороне, как нечто второстепенное.
Когда мы представляем себе плывущую рыбу, многие из нас видят величавые, стремительные движение из стороны в сторону. Их обычно совершают акулы и другие быстрые морские обитатели: сельдь, скумбрия, марлин. И в этом нет ничего предосудительного, потому что этих рыб можно считать эталоном.
Суб-карангиформный и карангиформный тип локомоции
Такая классическая техника плавания носит одно из двух названий, зависящих от того, насколько задействовано тело. Если для продвижения используется большая часть тела, где-то между половиной и двумя третями её длины, тогда рыба имеет суб-карангиформный тип локомоции (subcarangiform). Если мышцы, приводящие тело в движение, перенести на последнюю треть длины тела (в основном хвост), то получится карангиформный тип (carangiform).
Форель и лосось характеризуются суб-карангиформным типом локомоции. Особенности их внешнего строения позволяют длительное время плыть прямиком через озёра или вверх по рекам. Они могут совершать быстрые маневры в случае нападения хищников, во время охоты за добычей или же борьбы с постоянно меняющимися течениями.

Радужная форель
Виды, обладающие карангиформной манерой передвижения, лучше подготовлены к длительному быстрому плаванию. Океанические обитатели, такие как барракуда, сталкиваются с различными течениями, с которыми сталкивается и речная рыба. Часто эти рыбы объединяются в огромные косяки, что исключает необходимость совершать резкие выверенные движения в скоростном потоке.

Косяк барракуд (илл. uwphotographyguide.com)
 Многие из них, если не все, очень быстры и при необходимости удивительно проворны. Вряд ли читатель наблюдал скумбрию в действии, гораздо более вероятно, он мог ощутить силу и проворство сельди среднего размера, которая попала к нему на рыболовный крючок.
Многие из них, если не все, очень быстры и при необходимости удивительно проворны. Вряд ли читатель наблюдал скумбрию в действии, гораздо более вероятно, он мог ощутить силу и проворство сельди среднего размера, которая попала к нему на рыболовный крючок.
Амииформный тип локомоции
Следующая техника, которую демонстрирует скромный морской конёк, встречается достаточно редко. Её представители демонстрируют амииформную манеру перемещения (amiiform). При этом движения ограничены невероятно быстрыми колебаниями спинного плавника. Для большинства рыб, спинной плавник является единственным плавником в верхней части тела.
У морских коньков, учитывая их необычное вертикальное положение тела, спинной плавник расположен по центру вдоль спины и ориентирован в противоположном от головы направлении. Как таковой, он используется в качестве основного управляющего механизма.
Тем не менее, даже с учетом высоких скоростей, которые морскому коньку дают плавники с их удобным расположением, он представляется весьма жалким пловцом. Его постоянно сносит в сторону малейшим изменением течения, создаваемым более быстрыми собратьями по водоёму.
Тунниформный тип локомоции
Наиболее распространенными мореплавателями считаются, как правило, обладатели тунниформного типа локомоции (thunniforms). Классическим примером является, конечно же, тунец, в честь которого и назван данный тип передвижения. Тунец чемпион среди скоростных пловцов на длинные дистанции. Используя еле заметные движения туловищем, он управляет при этом крупным серповидным хвостом. Моторика хвоста обусловлена поразительно развитой мускулатурой и похожими на провода сухожилиями.
Но дело не только в хвосте. Совершаемые хвостом гребки определяют до 85% движения передней части тела при субкарангиформном, карангиформном и тунниформном типах локомоции.

Полосатый тунец (Katsuwonus pelamis) (илл. portal.nfrdi.re.kr).
Тем не менее, другие плавники также выполняют ряд важных функций. В частности, спинной плавник служит для совершения резких поворотов и торможения. Рыбы, располагающие вторым спинным плавником, используют его также, чтобы совершать рывок вперед. В некоторых случаях он задействуется на 15%.
Анальные плавники выступают в роли тормоза, брюшные обеспечивают и торможение и всплытие, а грудные помогают совершать быстрые повороты. У каждого вида плавников есть своё назначение и зачастую не одно.
Лабриформный тип локомоции
Губаны (Labridae, семейство окунеобразных) демонстрируют технику плавания окунеобразных (лабриформный тип или labriform). Они вращают своими грудными плавниками, как олимпийские спортсмены, плывущие в стиле «баттерфляй». Помимо губанов, многие аквариумные виды рыб могут также её демонстрировать. Важно не путать этот стиль с тем, что используют скаты.
Рейформный тип локомоции
Скаты и, в меньшей степени, спинопёры имеют собственную технику, обозначаемую как скатообразный или рейформный (rajiformes) тип локомоции. Преимущество скатов перед многими другими рыбами состоит в том, что они более гибкие, благодаря хрящевому скелету, который в полной мере способствует подобному передвижению.
В отличие от жёстких плавниковых лучей губанов, скат может изгибать свои «крылья» и совершать замысловатые колебательные движения. Это позволяет оптимизировать процесс отталкивания плавниками от очень большой площади поверхности воды.

Манта или гигантский морской дьявол (Manta birostris).
Острацииформный тип локомоции
В условиях аквариума наблюдатель сможет разглядеть, главным образом, движения грудных плавников, если только аквариумист не содержит ёмкость полную угрей! Два грудных плавника, как правило, наиболее активно используемых, предназначены для совершения координированных движений. Акулам они необходимы в качестве рулей, в особенности, для перемещения вверх или вниз, тогда как значительная часть других рыб всё равно используют грудные плавники для совершения толчков и поступательного движения вперед (острацииформный тип локомоции или ostraciiforms).
Классическим примером последних являются представители надсемейства Кузовкоподобных (Ostracioidea). Семейство Кузовковые (Ostraciidae), например, рыба-собака, типичная таксономическая единица данного надсемейства. Их ключевая особенность заключается в переложении функции руля на хвостовой плавник, в то время как грудные плавники проталкивают тело через толщу воды.
Балистиформный тип локомоции
Парные плавники не обязательно должны располагаться по бокам рыбы. Иглобрюхи и кузовки, любопытные любимчики туристов, используют спинные и анальные, а не грудные плавники, чтобы продвигать тело вперёд.
Эта техника, известная как балистиформное передвижение (balistiform), встречается редко. Тем не менее, её можно наблюдать у спинорога Пикассо колючего (Rhinecanthus aculeatus), либо у широко известной, возможно не только рыбозаводчикам, огромной выразительной Луны-рыбы (Mola mola). Она неспешно передвигается, главным образом, используя спинной и анальный плавники, потому что в процессе эволюционного развития хвостовой плавник был утрачен.
Передвижение спинорога Пикассо колючего (Rhinecanthus aculeatus) по аквариуму. Можно отметить активную работу спинного и анального плавников.
Передвижения рыбы луны, у которой отсутствует хвостовой плавник.
Ангуиллиформный тип локомоции
Наконец, любителя природы ни могут не завораживать гипнотические движения угрей, демонстрирующих ангуиллиформный тип локомоции (anguilliform). Это слово происходит от «Anguis» или «Anguilla», означающие «змея» и «угорь», соответственно. Как змеи, так и угри решили сказать решительное «нет» необходимости иметь конечности для передвижения, по крайней мере, там, где речь идёт о плавании.
Наблюдая за волнообразными движениями их длинного тела, нельзя не отметить чрезвычайно развитую мускулатуру угрей, достойную самых заядлых бодибилдеров. Хотя в некоторых случаях грудные плавники все ещё могут сохраняться, например, у представителей семейства Колючих угрей (Mastacemblidae), они играют незначительную роль в перемещении.
Хвост угрей может быть таким же цепким как хвост обезьяны. Стоит привести в пример мурену, которая способна молниеносно выбрасываться из своего логова и настолько же быстро забираться обратно, прихватив с собой добычу. При этом тело используется для сцепления с неподвижным предметом в норе, что повышает тяговую силу.
Просто удивительно, насколько обладатели ангуиллиформного типа локомоции гидродинамически идеально подходят как для движения вперед, так и назад. Как правило, их тело не покрыто чешуей, либо имеет очень мелкие чешуйки. Это в некоторой степени способствует беспрепятственному передвижению назад, и немного объясняет, почему угри оставляют после себя скользкий след.
Тема передвижения рыб очень сложная и практически неисчерпаемая. Представленная характеристика даёт общее понимание о техниках плавания и того, насколько легко не различать, а принимать все эти типы передвижений как должное.
Помимо описанных в данной статье рыб, также существуют виды с «ногами» и, использующие для передвижения выталкивание струй воды.
——
www.рrаcticаlfishkееping.cо.uk=5241
en.wikipedia.org/wiki/Fish_locomotion
aquavitro.org
Способ скоростного передвижения для меч-рыбы
Но коль рыба плавает, значит, в науке что-то обстоит не правильно. Рыбе никак нельзя приписать использование внеземных технологий. Причину незнания следует икать в науке и… в неизвестных по сей день свойствах воды. Вода является средой обитания животных, и выявление ее новых свойств объясняет секрет поведения рыб, млекопитающих и т. д.
Не отрицая достижений современной науки и техники, хочу заметить: если на протяжении нескольких столетий ученый мир не может объяснить полет майского жука и скоростное передвижение рыбы, значит надо перестать бояться их ужасных формул и принять на вооружение, прежде всего, человеческую логику. А она, как известно, создана по образу и подобию ТОГО, кто создал и рыбу и жука и воду. А математика придет потом. Архимед, Ньютон и другие были в науке прежде всего философами – они умели думать.
Общеизвестно, что вода практически не сжимается и плотность ее увеличивается очень незначительно даже на глубине Марианской впадины. А может ли вода расширяться, уменьшая свою плотность (становиться легче)? Вообще-то, каждая хозяйка ежедневно на кухне превращает воду в пар, только делает это очень энергозатратным способом. И техника использует этот способ в паровых машинах, используя принципы науки, называемой термодинамика. Сформулируем задачу по другому: а можно ли этот процесс осуществить более экономным способом? И сделать это можно тем способом, который используют меч-рыба, тунец, дельфин… С помощью хвоста и головы они делают с водой нечто такое, чего до сих пор не научился делать человек. То есть надо, используя механическое движение и познания в физике, снизить плотность воды перед лобовой частью подводного транспортного средства. Тогда подводный аппарат будет передвигаться со скоростью самолета (пар легче воздуха), а самолет — с космическими скоростями.
Перейдем к описанию маленького эксперимента, который позволяет сделать большие выводы
Автор изготовил подводную модель (см. фото 1 и фото 2) со сменной носовой частью (самая обыкновенная механика). Модель оснащена маломощным двигателем и передвигалась под водой на колесиках по металлической дорожке. При этом замерялось время прохождения одинакового расстояния.
Фото 1. На модели установлен обтекатель традиционной формы.
Фото 2. Экспериментальная модель с вогнутым экраном, по форме напоминающим голову меч-рыбы.
Как видите, аппарат не соответствует современным понятиям об обтекаемой форме лобовой части. Тем не менее, экспериментальная модель передвигалась на 11-15 % быстрее, чем традиционная. Механизм прост, но теория оригинальна. Не может существовать теоретически простая мысль там, где споткнулись современная наука и техника. Дело здесь не в сложных формулах, а в логике физического процесса. Логика присуща каждому человеку. Поэтому студент 3-4 курса политехнического института при желании вполне справится с освоением теоретических основ способа. Опуская физическую аналитику и небольшие математические расчеты, сделаю вывод. Описанный принцип увеличения скорости передвижения под водой основан на уменьшении плотности жидкости перед движущимся телом. Вода имеет удельный вес 1 кг/дм3, но перед меч-рыбой (как показывает расчет) плотность воды уменьшается до 0,13 кг/дм3. В этом и заключается главная часть секрета.
Для тех, кого интересует теория, привожу ссылку в Интернете, где размещена более полная статья.
На этой основе разъясним способ передвижения меч-рыбы.
Рыба обладает рядом физиологических особенностей. Ее «конструктивное устройство» полностью соответствует принципам теории и подтверждаются приведенными фотографиями и комментариями к ним.
Передвигаясь в пространстве, рыба совершает два движения. Первое – волнообразное колебательное движение всем телом и хвостом в горизонтальной плоскости, знакомое каждому человеку, наблюдавшему за плывущей рыбой. И второе – синхронно с первым движением рыба качает головой вверх и вниз. Причем, кончик меча остается неподвижным относительно оси передвижения. Меч рыбы, по сути, представляет собой лопасть высокоэффективного вентилятора, рассасывающего воду от оси передвижения, чем обеспечивает передвижение рыбы через высокоразреженное пространство. Важной особенностью является вогнутая форма меча и головы, которые обеспечивают снижение лобового сопротивления.
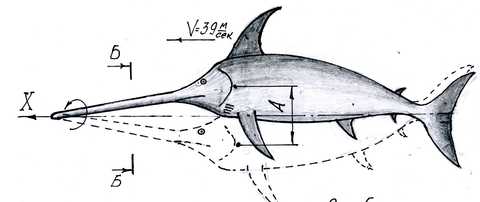
Рисунок 1. Наглядно иллюстрирует движения меч-рыбы.
Рисунок 2.
Это не только красиво, но имеет практическое значение. Жаберная часть головы имеет вогнутую форму. Как ни парадоксально, на лобовой части головы достигается наименьшая плотность воды. Движущееся тело изогнуто в вертикальной плоскости. И это при вертикальном-то расположении хвоста!!! Это объясняет причину, по которой рыба плывет в три раза быстрее дельфина. Рыба вращает своим мечом вокруг оси движения, а дельфин только качает своей узкой удлиненной мордой (подобной короткому мечу) вверх — вниз. Рыба рассасывает воду от оси передвижения в направлении двух осей координат, а дельфин — в направлении одной. Это значит, что меч-рыба плывет через более разреженное пространство, чем дельфин.
Плавным продолжением меча является вогнутая лобовая поверхность головы.

Фото 3. Меч совсем не острый: выпуклый закругленный кончик по мере приближения к голове постепенно превращается в прямолинейную, а у головы — в вогнутую коническую поверхность.

Фото 4. Пора сбрасывать «прилипший» разреженный поток — органы управления должны находиться в более плотной среде. Поэтому перед верхним килем вогнутая поверхность приобретает выпуклую форму. Нижние боковые плавники обеспечивают грубую корректировку положения рыбы в движении. Для них важна повышенная мощность поперечного движения. Гребенчатая выпуклая поверхность перед ними расслаивает и закручивает часть цельного вихря в шесть малых вихревых потоков. От них и отталкиваются плавники. Жабры еще вогнуты: в этом направлении спешить незачем — разреженная оболочка над извивающимся телом уменьшит силу трения.
 Фото 5. Рыба-парусник опустила нос вниз в предчувствии плохого исхода. Эта и следующая фотографии свидетельствуют о том, что рыба может кивать головой в вертикальном направлении. Ось кивания расположена сразу за жабрами под верхним «килем»
Фото 5. Рыба-парусник опустила нос вниз в предчувствии плохого исхода. Эта и следующая фотографии свидетельствуют о том, что рыба может кивать головой в вертикальном направлении. Ось кивания расположена сразу за жабрами под верхним «килем»
Фото 6. При жизни рыба-меч явно стремилась задирать свой нос вверх.
Рисунок 3. Высокие плавники и хвост выходят за габариты разреженной зоны вокруг тела рыбы, обеспечивая регулируемое передвижение в пространстве. Места изменения кривизны и наклона плавников указывают на места, где плотность воды возвращается в нормальное состояние. Судя по размерам и по углу наклона, малые хвостовые плавники расположены в зоне, где вода возвращается к исходной плотности. Назначение этих плавников – обеспечить неизменное положение центра колебательных движений рыбы относительно траектории ее передвижения. Рыба должна обладать динамометрической системой слежения за отклонением кончика меча от траектории движения. Это обеспечивает наименьшее лобовое сопротивление. Хвост оттолкнется уже от воды, вернувшейся к исходной плотности, обеспечивая скоростное передвижение через высокоразреженное пространство.
В полноформатной статье рассматривается физическая сущность таких физических явлений, как турбулентность и кавитация. Рассматриваются условия их возникновения. И то, каким образом рыбы в отдельных случаях используют разреженное состояние воды в турбулентном вихре.
Дополнительно скажу, что все сказанное распространяется на полет в воздухе. В принципе теория способна объяснить секрет полета майского жука, что является загадкой уже для авиации.
factopedia.ru
Органы движения — плавники / Зоология для учителя
Присмотритесь к движениям рыбы в воде, и вы увидите, какая часть тела принимает в этом главное участие (рис. 8). Рыба несётся вперёд, быстро двигая вправо и влево хвостом, который оканчивается широким хвостовым плавником. В этом движении принимает участие и туловище рыбы, но в основном оно осуществляется хвостовым отделом тела.
Поэтому хвост у рыбы очень мускулист и массивен, почти незаметно сливается с туловищем (сравните в этом отношении с наземными млекопитающими вроде кошки или собаки), например, у окуня туловище, внутри которого заключаются все внутренности, оканчивается лишь немного дальше половины общей длины его тела, а все остальное есть уже его хвост.
Кроме хвостового плавника у рыбы имеется ещё два непарных плавника — сверху спинной (у окуня, судака и некоторых других рыб он состоит из двух отдельных выступов, расположенных один, за другим) и снизу подхвостовой, или анальный, который называется так потому, что сидит на нижней стороне хвоста, как раз позади заднепроходного отверстия.
Эти плавники препятствуют вращению тела вокруг продольной оси (рис. 9) и, подобно килю на корабле, помогают рыбе сохранять в воде нормальное положение; у некоторых рыб спинной плавник служит и надёжным орудием защиты. Такое значение он может иметь в том случае если, поддерживающие его плавниковые лучи представляют собой твёрдые колючие иглы, препятствующие более крупному хищнику проглотить рыбу (ёрш, окунь).
Затем мы видим у рыбы ещё парные плавники — пару грудных и пару брюшных.
Грудные плавники сидят выше, почти по бокам тела, а брюшные более сближены между собой и расположены на брюшной стороне.
Расположение плавников у различных рыб неодинаково. Обыкновенно брюшные плавники находятся позади грудных, как мы видим это, например, у щуки (брюхопёрые рыбы; см. рис. 52), у других рыб брюшные плавники переместились на переднюю часть тела и находятся между двумя грудными (грудопёрые рыбы, рис. 10), и, наконец, у налима и некоторых морских рыб, например трески, пикши (рис. 80, 81) и наваги, брюшные плавники сидят впереди грудных, как бы на горле рыбы (горлопёрые рыбы).
Парные плавники не имеют сильной мускулатуры (проверьте это на вяленой вобле). Поэтому они не могут влиять на скорость движения, и рыба гребёт ими только при самом медленном перемещении в спокойной стоячей воде (карп, карась, золотая рыбка).
Главное же их назначение — поддерживать равновесие тела. Мёртвая или ослабевшая рыба опрокидывается брюхом кверху, так как спина у рыбы оказывается тяжелее, чем её брюшная сторона (почему — это мы увидим при вскрытии). Значит, живой рыбе приходится всё время делать некоторое усилие, чтобы не опрокинуться на спину или не свалиться набок; это и достигается работой парных плавников.
Убедиться в этом можно путём несложного опыта, лишив рыбу возможности пользоваться своими парными плавниками и привязав их к телу шерстяными нитками.
У рыбы с подвязанными грудными плавниками более тяжёлый головной конец перетягивает и опускается вниз; рыбы, у которых отрезаны или привязаны грудной или брюшной плавники с одной стороны, ложатся на бок, а рыба, у которой обвязаны нитками все парные плавники, точно мёртвая опрокидывается вверх брюхом.
(Здесь, впрочем, имеются исключения: у тех видов рыб, у которых плавательный пузырь расположен ближе к спинной стороне, брюхо может оказаться тяжелее спины, и рыба не перевернётся.)
Кроме того, парные плавники помогают рыбе делать повороты: желая повернуть направо, рыба загребает левым плавником, а правый прижимает к телу, и наоборот.
Возвратимся ещё раз к уточнению роли спинного и подхвостового плавников. Иногда не только в ответах учащихся, но и в объяснениях учителя дело представляется так, будто именно они придают телу нормальное положение — спиной вверх.
В действительности, как мы видели, такую роль выполняют парные плавники, тогда как спинной и подхвостовой при движении рыбы препятствуют её веретеновидному телу крутиться вокруг продольной оси и тем самым поддерживают нормальное положение, которое придали телу парные плавники (у ослабевшей рыбы, плывущей на боку или вверх брюхом, те же непарные плавники поддерживают уже принятое телом ненормальное положение).
www.5zaklepok.ru
Приспособления, позволяющие рыбам расходовать меньше энергии при движении в воде
Природа поступила очень разумно, когда наделила рыб особыми приспособлениями, помогающими обитателям водоемов, преодолевать сопротивление воды во время движения в ее толщах. Разумеется, есть они и у декоративных аквариумных рыбок.
За сотни миллионов лет, которые существуют рыбы на нашей планете, можно с уверенностью утверждать, что они, как никто другой, в результате своей длительной эволюции обзавелись множеством уловок и «секретов», позволяющих этим удивительным представителям подводной фауны с огромной выгодой для себя использовать свою природную среду обитания. Одним из таких секретов как раз и является способность передвигаться в воде долго и с большой скоростью, не чувствуя при этом усталости и не тратя попусту накопленную энергию.
Способы передвижения рыб, скорость и выносливость

Для рыб характерны такие способы передвижения как плавание, ползание, полет. Нас интересует первый способ, потому что он присущ подавляющему виду рыб. Однако и эти виды тоже могут летать (например, если в панике выскакивают из аквариума от преследующего их хищника) и ползать, когда, плюхнувшись на пол, продолжают движение по суше.
Нас все-таки интересует скорость плавания рыб и их выносливость. Для плавающих рыб, независимо от места их обитания (озеро Титикака или домашний аквариум), характерны четыре вида скорости – бросковая, максимальная, крейсерская, промежуточная.
- с бросковой скоростью рыбы кидаются на добычу и спасаются от обнаруженной явной близкой угрозы. Ихтиологи установили, что долго с такой скоростью рыбы плыть не могут – лишь какие-то доли секунды. После броска они снижают скорость движения до максимальной,
- на максимальной скорости рыбы «убегают» от пасти хищника. Сохранять ее они способны в зависимости от своего размера и строения от 20 секунд (декоративные аквариумные рыбки) до нескольких минут (более крупные по размеру и развитию мускулатуры обитатели природных водоемов: рек, морей, океанов),
- устав, рыбы переходят на крейсерскую скорость, которую способны поддерживать довольно долго,
- с промежуточной скоростью (или обычной) рыба не может плавать постоянно, потому что ее поведение в воде постоянно меняется: то она увидела кусочек корма и сделала за ним бросок, то она увидела опасность со стороны крупного соседа по аквариуму, то еще что-то заставило рыбку «переключиться» на другую скорость.
Рыбам, как и нам, людям, присуща одна закономерность: чем быстрее мы двигаемся, тем скорее устаем. Как установили ихтиологи, мелкие рыбки – гораздо более выносливы и их мышцы лучше приспособлены к частому «переключению» скоростей.
Что способствует тому, чтобы рыбы двигались быстро и не утомлялись:

- обтекаемая форма тела помогает рыбам набирать большую скорость. Если обитатели аквариума правильно пообобраны и миниатюрным рыбкам ничто не грозит, необходимость прибегать к броскам и максимальной скорости у них если и возникает, то крайне редко, так как владелец аквариум следит, чтобы корма для всех хватало. Так или иначе, в погоне за едой опережают соперников те рыбки, тельце которых имеет вытянутую обтекаемую форму, лучше противостоящую сопротивлению воды. В этой связи нет ничего удивительного в том, что в природе рекордсменом среди рыб по скорости признан парусник, обитающий в западных водах Тихого и тропических водах Индийского океана. Своим названием рыба обязана спинному плавнику в виде паруса. Ученые установили, что эта рыба способна за короткое время развить скорость до 110 км/ч. Из сухопутных животных такая скорость не по зубам даже быстроногому гепарду. Такой прыткостью рыба-парус обязана, в первую очередь, строению своего тела;
- выделяемое кожей рыб слизистое вещество снижает силу сопротивления воды. Выделение такой слизистой смазки характерно для большинства видов рыб (морских, пресноводных, промысловых, декоративных). Чем больше слизи выделяется у рыбы, тем быстрее она плавает. Такая естественная смазка сглаживает все неровности на теле рыбы и кратно уменьшает трение тела рыбы о воду;
- плавники и хвост способствуют сохранению сил у рыб и экономии энергии,
- благодаря жабрам усиливается приток кислорода к сосудам. Они в свою очередь снабжают жабры рыб кровью, что усиливает снабжение кислородом клеток тканей мышц подводных обитателей.
Все эти анатомические, биологические и физиологические особенности рыб необходимы им для экономии энергии при неустанном постоянном передвижении в воде.
Поделиться в соц. сетях:
kwitri.ru
Скорость плавания движения рыб под водой
Скорость плавания рыбы в воде пропорциональна частоте и амплитуде колебаний тела и хвоста и укладывается в рамки определенных математических уравнений. Например, для карася, ельца и форели предельная скорость плавания v=(1/4)[L(3f-4)], где L - длина тела; f- частота колебаний тела (хвоста).
Несмотря
на то что вода является более плотной средой по сравнению с воздухом, рыбам
удается достичь очень высоких скоростей (км/ч):
Рыба-меч 130
Голубой марлин 90
Голубой тунец 80
Терпуг 70
Голубая акула 60
Атлантический лосось 40
Естественно, что для обеспечения таких скоростей природа наделила рыб рядом морфологических, гидродинамических и метаболических приспособлений. Поскольку предельные абсолютные скорости рыб зависят от их чиненных размеров, специалисты предлагают использовать для сопоставления скоростных возможностей рыб относительный показатель — коэффициент скорости: Этот коэффициент характеризует скорость рыбы, равную числу ее корпусов (длин) в секунду. Тогда все виды рыб можно классифицировать, как минимум, шестью категориями (табл. 5.4). Понятно, что рыбы с одинаковыми максимальными скоростями движения, но с различной длиной тела согласно этой классификации могут относиться к разным категориям.
|
5.4. Коэффициент скорости разных видов рыб
|
Рыбы, у которых изгибы тела затруднены, используют для передвижения плавники (см. рис. 5.13). Плавники совершают либо волнообразные (ундулярные), либо гребковые движения. При этом скат и морской конек используют грудные, угорь — анальный, а кузовок — хвостовой плавники.
Следует принимать во внимание и то, что многие виды рыб при относительно небольших крейсерских скоростях движения могут развивать высокую бросковую скорость (например, щука, акулы). При небольшой крейсерской скорости (1-4 L/C) во время скоротечного броска рыбы доводят скорость своего перемещения до 30-70 L/C.
Для уменьшения гидродинамического сопротивления рыбы применяют две тактические уловки. Во-первых, они сохраняют ламинарность обтекающего потока по всей длине тела от головы до хвостового плавника. Достигается это сглаживанием неровностей тела. У активных пловцов даже глаза могут быть закрыты жировыми веками, создающими своеобразные обтекатели. Все плавники, за исключением хвостового, прижимаются к телу, а у тунцов даже убираются в специальные желоба и впадины тела.
Многие рыбы при движении с большими скоростями переходят на так называемое пассивное дыхание. При этом вода как бы самотеком проходит через ротовую полость и жабры. На выходе из жаберного аппарата вода не создает турбулентных завихрений как у малоподвижных видов при активном прокачивании воды через жабры, а ламинируется.
Снижение сопротивления достигается и за счет снижения трения тела о водную массу. Этому способствуют эластические свойства кожи, чешуя и кожная слизь. В опытах со щукой искусственное удаление слизи с ее тела повышало гидродинамическое сопротивление на 50%. Слизь выступает главным фактором ламинирования обтекающего тело водяного потока у таких рыб, как угри и сомы. Эти рыбы не отличаются высокой скоростью плавания, но способны на короткие броски с высокой стартовой скоростью, что требует ламинирования потока. Однако у быстрых рыб — тунцов и акул — слизи на коже очень мало и кожа имеет шершавую, а не гладкую поверхность.
Подмечено, что размер и распределение чешуи по телу также связаны с гидродинамическими характеристиками рыбы. Наличие чешуи на туловище препятствует образованию складок кожи при мышечных сокращениях, т. е. сохраняет обтекаемость тела рыбы. Крупная чешуя обычна для малоподвижных рыб с коротким, но высоким телом. Мелкая чешуя характерна для рыб с вытянутым телом, совершающих угревидные движения. Лучшие пловцы среди рыб имеют среднюю и мелкую чешую, причем у последних чешуя может вообще отсутствовать в наиболее гибкой части хвостового стебля.
Следует признать, что способность рыб к снижению гидродинамического сопротивления до сих пор остается во многом загадочным явлением природы. Физические методы измерения сопротивления тел живых и мертвых рыб показали, что у живых рыб гидродинамическое сопротивление значительно ниже. Для поддержания скорости движения 4 L/C живая рыба затрачивает энергии в 2 раза меньше ожидаемой расчетной величины. Предполагают, что решающая роль в преодолении гидродинамического сопротивления принадлежит особым свойствам живого тела — мышечным сокращениям и взаимодействиям кожных покровов с водяными потоками.
Кроме того, живые рыбы умело используют энергию вихревых потоков воды, возникающих при каждом резком изгибе тела. В интересных опытах М. V. Rosen рыб помещали в сосуд, заполненный водой и слоями молока. Оказалось, что при каждом движении тела или хвоста рыбы в области жабр возникают местные водовороты. Рыба движется вдоль дорожки («дорожка Кармана»), состоящей из водяных завихрений. Тело рыбы получает дополнительную энергию ускорения при контакте с каждым завихрением, так как завихрение слегка подталкивает тело рыбы по оси движения. Это явление описано не только у рыб, но и у всех подвижных водных животных от насекомых до дельфинов. Особенно эффективно используются водяные вихри животными при движении в плотной стае. Частота изгибов тела рыб, или ундуляция, довольно высока (табл. 5.5). Она в основном и определяет относительную скорость рыб. Чем чаще рыба совершает изгибы тела, тем большую скорость она развивает.
|
5.5. Максимальная ундуляция некоторых видов рыб
|
|
Карп |
25 |
10 |
|
Вьюн |
20 |
10 |
|
Бычок |
15 |
20 |
|
Гольян |
7 |
15 |
|
Колюшка |
7 |
15 |
|
Хамса |
12 |
30 |
|
Тиляпия |
21 |
12 |
Физики добавляют к уже отмеченной способности рыб снижать гидродинамическое сопротивление еще и способность сохранять энергию при импульсном характере движения.
В соответствии с этим законом при волнообразном и импульсном движении тела в воде импульсный характер движущей силы распространяет в воде свое действие на движущееся тело и при расслаблении скелетной мускулатуры рыбы. Этот эффект дополняется вихревым потоком, возникающим позади рыбы.
Исследования показали некорректность сопоставления движения рыбы и неживых физических тел в воде. В физиологии известен так называемый «парадокс Грея». Еще в 30-е годы американский исследователь Дж. Грей при сопоставлении расчетных энергетических затрат, необходимых для движения в воде дельфина, с реальными затратами энергии животного обнаружил парадоксальное явление. Для движения со скоростью 40 км/ч дельфину длиной 180 см необходимо развить мощность не менее 2,5л. с. Реально дельфин не способен развить мощность даже в 0,5 л. с.
Подобный парадокс обнаружен и у рыб. Например, лососи не обладают необходимыми энергетическими и мощностными ресурсами для нерестовых миграций. В действительности лосось при миграциях затрачивает энергии в несколько раз меньше по сравнению с идеальной физической (неживой) моделью. Коэффициент полезного действия энергетических затрат, расходуемых на двигательные усилия у рыб, в разных экспериментах оценивается 60-90%. Максимальный КПД зарегистрирован при скоростях движения от 1 до 8 L/C. Удельная мощность, развивается при этом рыбой, составляет около 8 Вт/(кг o с). Интересно, что при быстром плавании или резком броске обмен энергии возрастает в 2-7 раз по сравнению со стандартным обменом на фоне большой кислородной задолженности. Величина кислородного долга оценивается в 45 % величины стандартного обмена и ликвидируется в течение нескольких часов относительного покоя.
Неожиданно высокая мощность скелетной мускулатуры рыб отчасти объясняется повышением температуры тела во время движения. Так, у тунцов разница между температурой воды и температурой тела составляет 5-13 °C. Однако, оставаясь пойкилотермными животными, рыбы проявляют большую зависимость от температуры окружающей среды. Показано, что максимальную скорость движения каждый вид рыбы проявляет в определенных температурных диапазонах. Для нерки температурным оптимумом является температура воды 15 °C. Только при этой температуре она развивает крейсерскую скорость 5 L/C в течение 1 ч движения. Как понижение температуры воды до 10 °C, так и ее повышение до 20 °C снижает скоростные возможности рыбы.
Подобная закономерность обнаружена и у других видов рыб — карася, пикши, нототении, но в другом температурном диапазоне, зачастую очень узком. Например, нототения наиболее активна при минус 1,8 °C и уже при 2 °C прекращает движение.
Влияние температуры окружающей среды на крейсерскую скорость рыб осуществляется через обмен веществ и изменение вязкости воды в зоне контакта с кожными покровами рыбы.
У рыб с хорошо выраженным бросковым характером двигательной активности несколько другие свойства. Бросковые скорости остаются высокими в широком диапазоне температур, что увязывают с некоторым прогревом мышц при броске. Двигательная активность рыбы зависит и от некоторых дополнительных факторов. Так, предельные скорости движения у большинства рыб с хорошим зрением достигаются лишь при достаточном уровне освещенности. Более того, у рыб при этом возникает потребность в зрительных ориентирах. Например, у верховки увеличение зрительных ориентиров с 0 до 50 на 1 м пути сопровождалось ростом линейной скорости движения с 32 до 47 см/с.
Влияет на скорость движения и гидрохимический состав воды. Так, снижение концентрации кислорода в воде с 2 до 1 мг/л сопровождалось снижением скорости движения с 3 до 1 L/C, т. е. в 3 раза. Еще большей чувствительностью к содержанию кислорода в воде отличается форель. Уменьшение концентрации кислорода в воде с 2,5 мг/л всего на 0,5 мг/л сопровождается четырехкратным падением крейсерской скорости рыбы.
Скорость и характер движения рыбы меняются при изменении солености, осмотического давления, содержания диоксида углерода в водной среде. Скорость движения зависит и от физиологического состояния рыбы. Так, критические скорости движения леща после нереста уменьшаются в 3-5 раз. Лососи с незрелыми половыми продуктами (1-3-я стадия зрелости) развивают большую скорость и проявляют большую выносливость, чем рыбы перед нерестом. Отмечены половые различия в скоростных возможностях рыб одного вида. Самцы развивают более высокую скорость по сравнению с самками. У стайных рыб изоляция отдельной особи меняет характер двигательной активности. При этом крейсерская скорость и выносливость уменьшаются, а бросковые скорости могут увеличиваться.
Известно и влияние сытости (голода), наличия (отсутствия) течений на двигательную активность рыб, причем эти влияния различны. Голодные рыбы более активны по сравнению с сытыми, однако предельные крейсерские скорости выше у сытых рыб.
biofile.ru
Leave A Comment