
§17. Митохондрии. Пластиды. Клеточный центр. Биология 9 класс Пасечник
1. Каково строение и функции АТФ?
Аденозинтрифосфат (АТФ) — нуклеотид, состоящий из азотистого основания аденина, углевода рибозы и трёх остатков фосфорной кислоты.
АТФ — универсальный источник энергии для всех реакций, протекающих в клетке.
2. Какие виды пластид вам известны?
В зависимости от окраски пластиды делят на лейкопласты, хлоропласты и хромопласты.
3. Какие способы движения клеток вам известны?
1. Амебоидное движение.
2. Движения при помощи жгутиков и ресничек.
3. Движение с помощью мышц.
4. В каком виде клетка хранит питательные вещества?
В виде липидов и гликогена.
Вопросы
1. Какова функция митохондрий?
Функция митохондрий – синтез АТФ.
2. Какие виды пластид вы знаете?
В зависимости от окраски пластиды делят на лейкопласты, хлоропласты и хромопласты.
3. Чем отличается каждый вид пластид от другого?
Лейкопласты — неокрашенные пластиды, как правило выполняют запасающую функцию. В лейкопластах клубней картофеля накапливается крахмал. Лейкопласты высших растений могут превращаться в хлоропласты или хромопласты.
Хромопласты — пластиды, окрашенные в жёлтый, красный или оранжевый цвет. Окраска хромопластов связана с накоплением в них каротиноидов. Хромопласты определяют окраску осенних листьев, лепестков цветов, корнеплодов, созревших плодов.
Хлоропласты — пластиды, несущие фотосинтезирующие пигменты — хлорофиллы. Имеют зелёную окраску у высших растений, харовых и зелёных водорослей.
4. Почему граны в хлоропласте расположены в шахматном порядке?
Граны расположены в шахматном порядке, чтобы до каждой из них мог доходить свет солнца.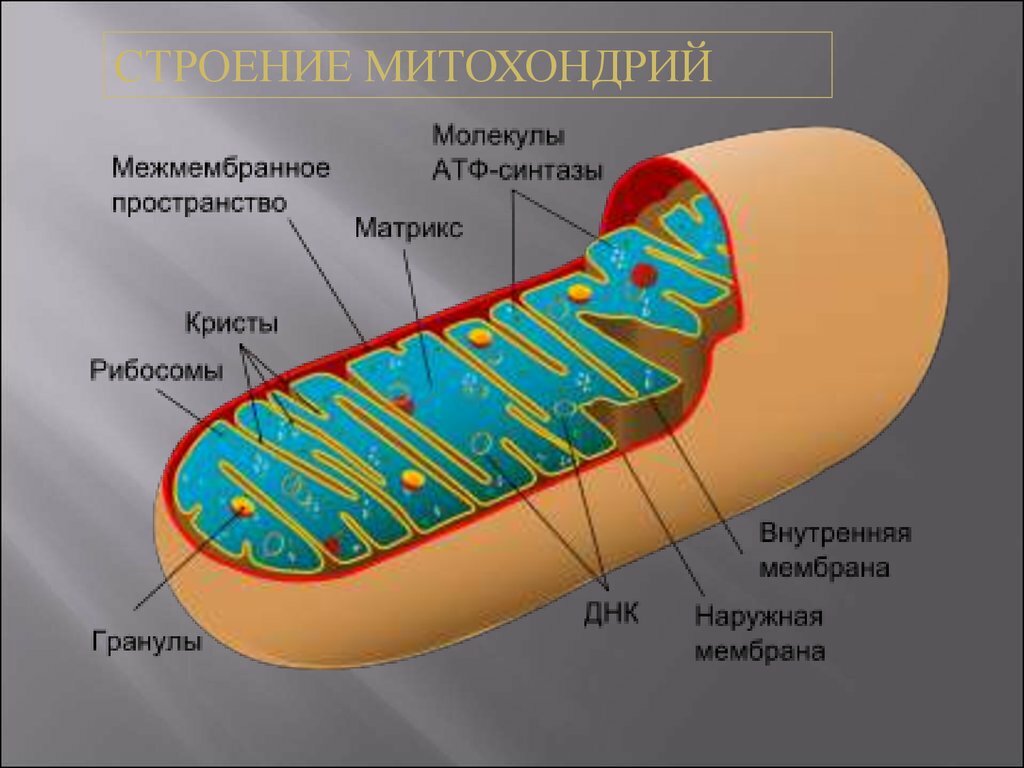
5. В чём сходство митохондрий и пластид?
Как и митохондрии, пластиды содержат собственные молекулы ДНК. Поэтому они также способны самостоятельно размножаться, независимо отделения клетки.
Клеточный центр играет важнейшую роль в формировании внутреннего скелета клетки — цитоскелета. Из области клеточного центра расходятся многочисленные микротрубочки, поддерживающие форму клетки и играющие роль своеобразных рельсов для движения органоидов по цитоплазме.
Велика роль клеточного центра при делении клеток, когда центриоли расходятся к полюсам делящейся клетки и образуют веретено деления.
7. Приведите примеры клеточных включений.
Это могут быть мелкие капли жира, гранулы крахмала или гликогена, реже — гранулы белка, кристаллы солей.
Задания
Сравнив строение и функции митохондрий и пластид.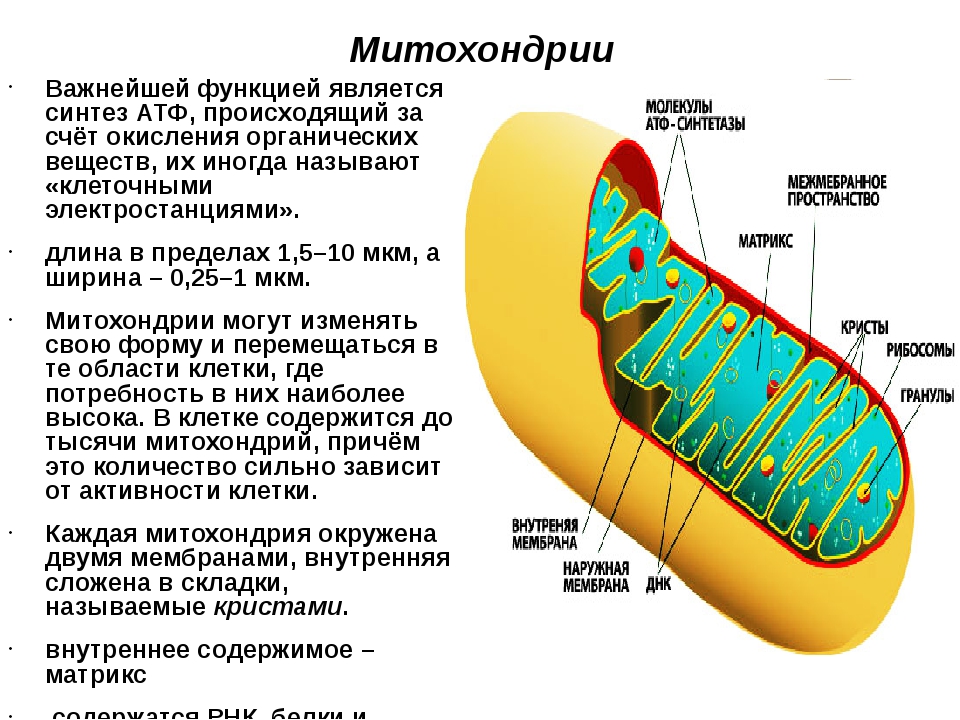 В чём их сходство и различия?
В чём их сходство и различия?
Сходство:
● Двумембранные органоиды. Наружная мембрана ровная, а внутренняя образует многочисленные впячивания, служащие для увеличения площади поверхности. Между мембранами имеется межмембранное пространство.
● Имеют собственные кольцевые молекулы ДНК, все типы РНК и рибосомы.
● Способны к росту и размножению путём деления.
● В них осуществляется синтез АТФ.
Различия:
● Впячивания внутренней мембраны митохондрий (кристы) имеют вид складок или гребней, а впячивания внутренней мембраны хлоропластов образуют замкнутые дисковидные структуры (тилакоиды), собранные в стопки (граны).
● Митохондрии содержат ферменты, участвующие в процессе клеточного дыхания. Внутренняя мембрана хлоропластов содержит фотосинтетические пигменты и ферменты, участвующие в преобразовании энергии света.
● Основная функция митохондрий – синтез АТФ. Основная функция хлоропластов – осуществление фотосинтеза.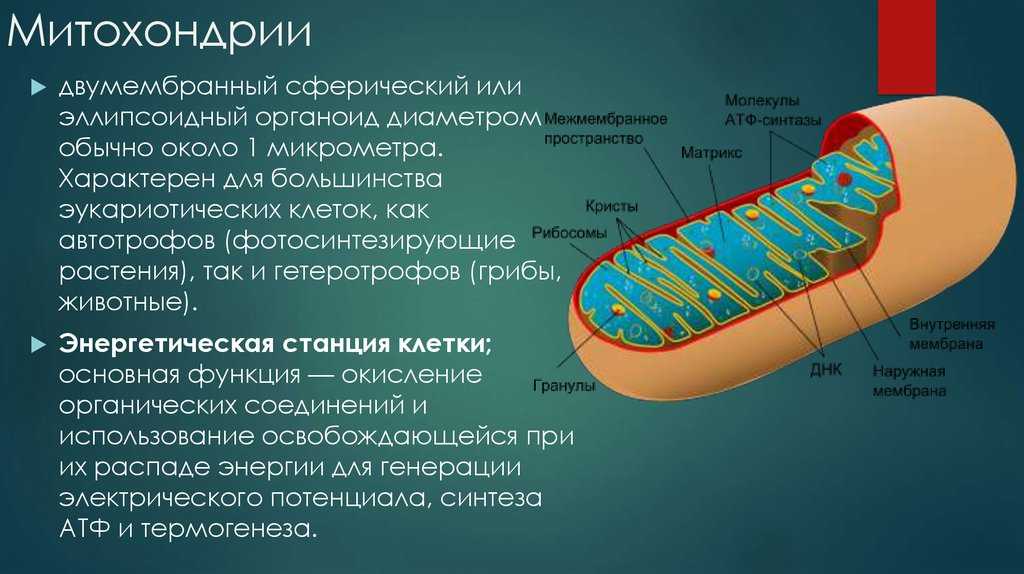
Строение и функции митохондрий и пластид
Митохондрии есть у всех типов эукариотных клеток (рис. 1). Они имеют вид либо округлых телец, либо палочек, реже — нитей. Их размеры колеблются от 1 до 7 мкм. Число митохондрий в клетке составляет от нескольких сотен до десятков тысяч (у крупных простейших).
Рис. 1. Митохондрии. Вверху — митохондрии (?) в мочевых канальцах, видимые в световом микроскопе. Внизу — трехмерная модель организации митохондрии: 1 — кристы; 2 — внешняя мембрана; 3 — внутренняя мембрана; 4 — матрикс
Митохондрия образована двумя мембранами — внешней и внутренней, между которыми расположено межмембранное пространство. Внутренняя мембрана образует множество впячиваний — крист, представляющих собой либо пластины, либо трубочки. Такая ее организация обеспечивает огромную площадь внутренней мембраны. На ней располагаются ферменты, обеспечивающие преобразование энергии, заключенной в органических веществах (углеводах, липидах), в энергию АТФ, необходимую для жизнедеятельности клетки.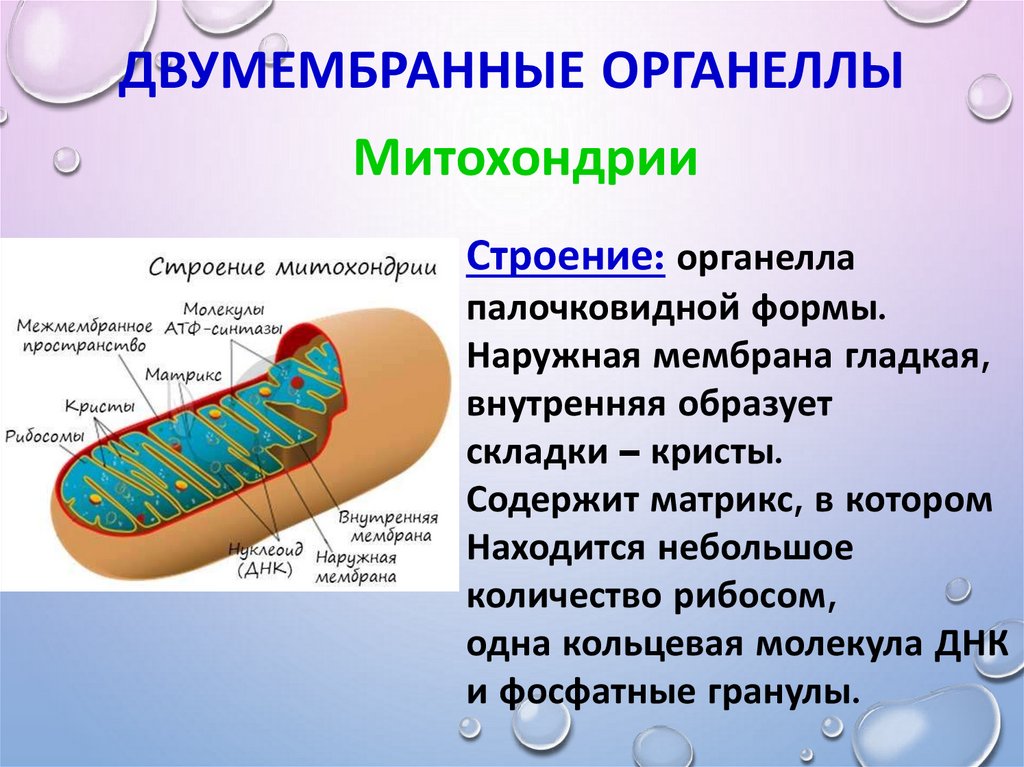 Следовательно, функция митохондрий — участие в энергетических клеточных процессах. Именно поэтому большое количество митохондрий присуще, например, мышечным клеткам, выполняющим большую работу.
Следовательно, функция митохондрий — участие в энергетических клеточных процессах. Именно поэтому большое количество митохондрий присуще, например, мышечным клеткам, выполняющим большую работу.
Пластиды. В растительных клетках обнаруживаются особые органоиды — пластиды, имеющие чаще веретеновидную или округлую форму, иногда более сложную. Различают три вида пластид — хлоропласты (рис. 2), хромопласты и лейкопласты.
Хлоропласты отличаются зеленым цветом, который обусловлен пигментом — хлорофиллом, обеспечивающим процесс фотосинтеза, т. е. синтеза органических веществ из воды (Н2О) и углекислого газа (СО2) с использованием энергии солнечного света. Хлоропласты содержатся преимущественно в клетках листьев (у высших растений). Они сформированы двумя параллельно расположенными друг другу мембранами, окружающими содержимое хлоропластов — строму. Внутренняя мембрана образует многочисленные уплощенные мешочки — тилакоиды, которые сложены в стопки (наподобие стопки монет) — граны — и лежат в строме.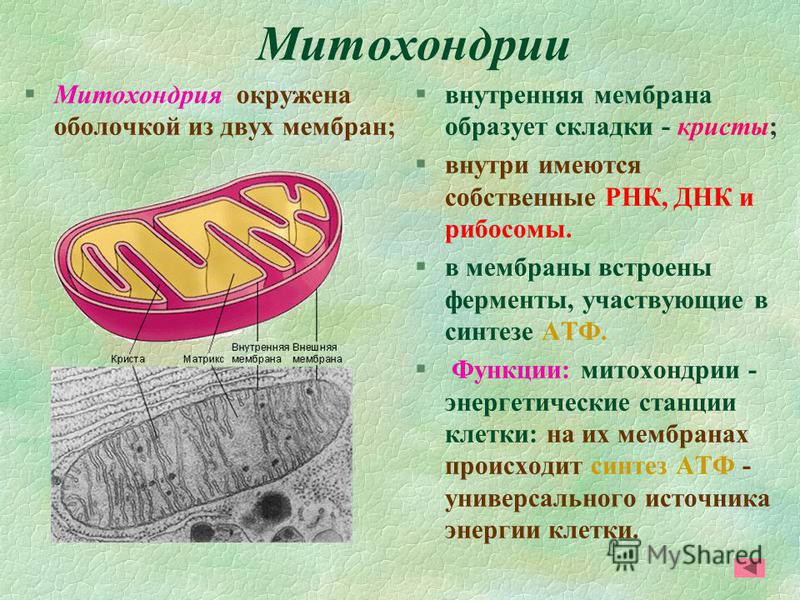 Именно в тила-коидах и содержится хлорофилл.
Именно в тила-коидах и содержится хлорофилл.
Хромопласты определяют желтый, оранжевый и красный цвет многих цветков и плодов, в клетках которых присутствуют в большом количестве. Основными пигментами в их составе являются каротины. Функциональное назначение хромопластов состоит в цветовом привлечении животных, обеспечивающих опыление цветков и распространение семян.
Рис. 2. Пластиды: а — хлоропласты в клетках листа элодеи, видимые в световом микроскопе; б — схема внутреннего строения хлоропласта с гранами, представляющими собой стопки плоских мешочков, расположенных перпендикулярно поверхности хлоропласта; в — более подробная схема, на которой видны анастомозирующие трубочки, соединяющие отдельные камеры гран
Лейкопласты — это бесцветные пластиды, содержащиеся в клетках подземных частей растений (например, в клубнях картофеля), семян и сердцевины стеблей. В лейкопластах, главным образом, происходит образование из глюкозы крахмала и накапливание его в запасающих органах растений.
Пластиды одного вида могут превращаться в другой. Например, при осеннем изменении цвета листьев хлоропласты превращаются в хромопласты.
Источник: Краснодембский Е. Г.»Общая биология: Пособие для старшеклассников и поступающих в вузы»
Митохондрии – Структура – Функция
звезда звезда звезда звезда
на основе 36 оценок (ы): Аби Бадрик
Последнее обновление: 10 апреля 2023 г.
Редакции: 15
format_list_bulleted Содержание добавить удалить
- 1 Структура
- 2 Функция
- 3 ДНК и наследование
- 4 Клиническая значимость – Наследственная оптическая нейропатия Лебера
Митохондрии 056 ) представляют собой клеточные органеллы с двойными мембранами типичного размера 0,75-3 мкм². Они обнаружены в большинстве клеток млекопитающих, за заметными исключениями, включая зрелые эритроциты
Они обнаружены в большинстве клеток млекопитающих, за заметными исключениями, включая зрелые эритроциты
В этой статье мы рассмотрим структуру и функцию митохондрий и рассмотрим их клиническое значение.
СтроениеМитохондрии имеют внутреннюю и внешнюю мембраны, между которыми межмембранное пространство . внешняя мембрана содержит белки, известные как порины, которые обеспечивают перемещение ионов в митохондрию и из нее. Ферменты, участвующие в удлинении жирных кислот и окислении адреналина, также можно найти на внешней мембране.
Пространство во внутренней мембране митохондрии известно как матрикс , который содержит ферменты цикла Кребса (TCA) и цикла жирных кислот, а также ДНК, РНК, рибосомы и гранулы кальция.
Внутренняя мембрана содержит различные ферменты. Он содержит АТФ-синтазу, которая генерирует АТФ в матрице, и транспортные белки, которые регулируют перемещение метаболитов в матрицу и из нее.
Внутренняя мембрана состоит из cristae для увеличения площади поверхности, доступной для производства энергии посредством окислительного фосфорилирования .
Rice University [CC BY 4.0], via openstax CNX
Рис. 1. Электронная микрофотография митохондрии
ФункцияТаким образом, количество митохондрий, обнаруженных в клетке, является хорошим показателем скорости метаболической активности клетки; клетки, которые очень метаболически активны, такие как гепатоциты, будут иметь много митохондрий.
Митохондрии также играют роль в поддержании внутриклеточной среды. Они:
- Хранят каспаз , ответственных за запуск апоптоза.
- Способны кратковременно хранить кальций , способствуя гомеостазу кальция.

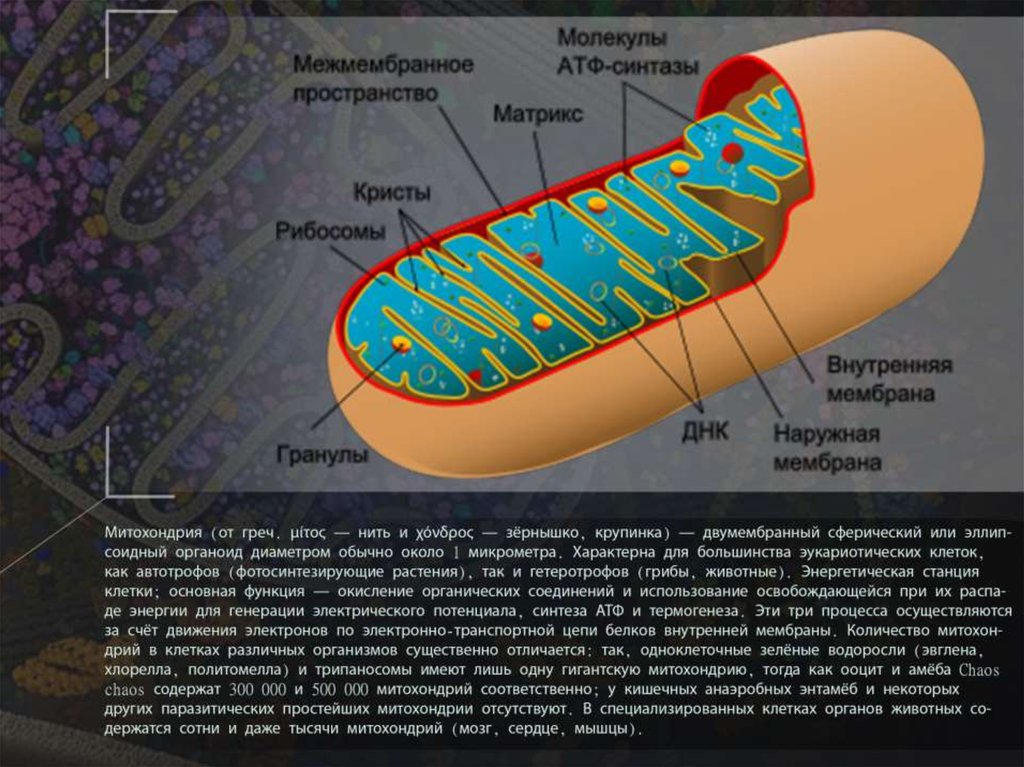
В бурой жировой ткани митохондрии выполняют альтернативную функцию по производству тепла с использованием цепи переноса электронов.
Мариана Руис Вильярреал LadyofHats [общественное достояние], через Wikimedia Commons
Рис. 2. Митохондрия с ее основными характеристиками
ДНК и НаследованиеМитохондрии реплицируют свою ДНК с помощью процесса, называемого бинарным делением, для образования одной митохондрии и могут использовать ее.
Их ДНК имеет материнскую линию , что означает, что их ДНК передается от матери к ребенку с небольшими изменениями.
Клиническая значимость — наследственная нейропатия зрительного нерва Лебера
В результате того, насколько важны митохондрии для выживания клетки,
Некоторые общие черты митохондриальных заболеваний включают непереносимость физической нагрузки, миопатию и мышечную слабость.
Наиболее частым митохондриальным заболеванием является наследственная оптическая нейропатия Лебера (LHON), которая поражает зрительный нерв , вызывая нечеткость центрального зрения и потерю цветового зрения и сопряжена с риском развития слепоты.
Абу-Амеро, KK. [CC BY 2.0], via OPENi
Рис. 3 – Митохондрии зрительного нерва
print Print this Article
Mitochondria — The Cell — NCBI Bookshelf
Митохондрии играют решающую роль в выработке метаболической энергии в эукариотических клетках. Как уже говорилось в главе 2, они ответственны за большую часть полезной энергии, получаемой при расщеплении углеводов и жирных кислот, которая в процессе окислительного фосфорилирования превращается в АТФ. Большинство митохондриальных белков транслируются на свободных цитозольных рибосомах и импортируются в органеллы с помощью специфических сигналов-мишеней. Кроме того, митохондрии уникальны среди уже рассмотренных цитоплазматических органелл тем, что содержат собственную ДНК, которая кодирует тРНК, рРНК и некоторые митохондриальные белки.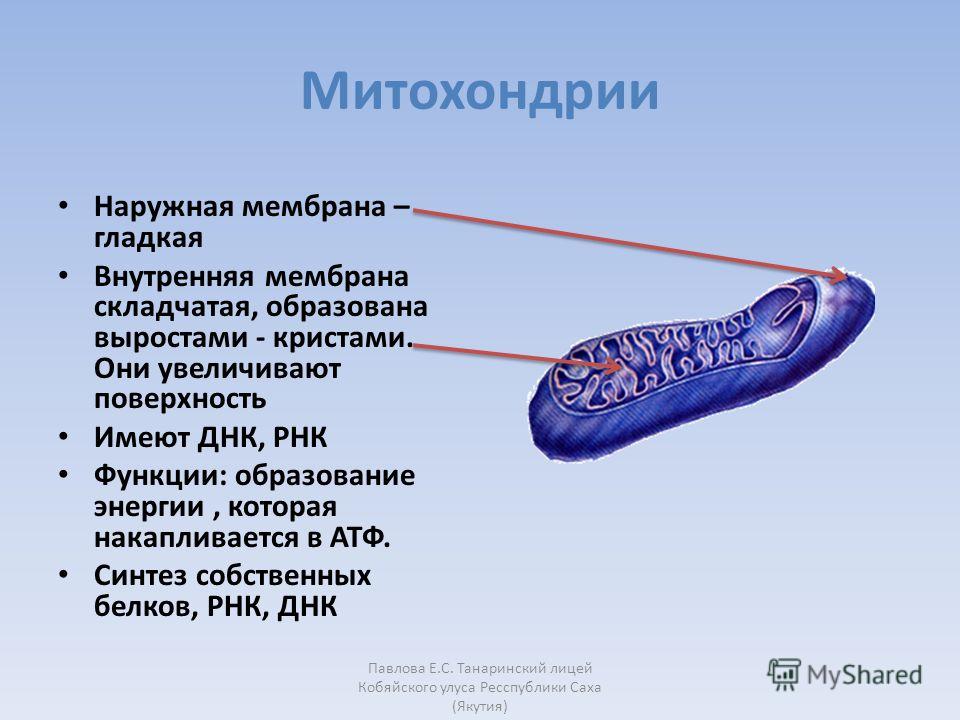 Таким образом, в сборке митохондрий участвуют белки, кодируемые их собственными геномами и транслируемые внутри органеллы, а также белки, кодируемые ядерным геномом и импортируемые из цитозоля.
Таким образом, в сборке митохондрий участвуют белки, кодируемые их собственными геномами и транслируемые внутри органеллы, а также белки, кодируемые ядерным геномом и импортируемые из цитозоля.
Организация и функция митохондрий
Митохондрии окружены двойной мембранной системой, состоящей из внутренней и внешней митохондриальных мембран, разделенных межмембранным пространством (). Внутренняя мембрана образует многочисленные складки ( cristae ), которые простираются внутрь (или в матрикс) органеллы. Каждый из этих компонентов играет различные функциональные роли, при этом матрикс и внутренняя мембрана представляют собой основные рабочие отсеки митохондрий.
Рисунок 10.1
Структура митохондрии. Митохондрии ограничены двойной мембранной системой, состоящей из внутренней и внешней мембран. Складки внутренней мембраны (кристы) вдаются в матрикс. (Микрофотография K. R. Porter/Photo Researchers, Inc.)
Матрица содержит митохондриальную генетическую систему, а также ферменты, ответственные за центральные реакции окислительного метаболизма (). Как обсуждалось в главе 2, окислительный распад глюкозы и жирных кислот является основным источником метаболической энергии в клетках животных. Начальные стадии метаболизма глюкозы (гликолиз) происходят в цитозоле, где глюкоза превращается в пируват (см.). Затем пируват транспортируется в митохондрии, где происходит его полное окисление до CO 2 дает большую часть полезной энергии (АТФ), полученной в результате метаболизма глюкозы. Это включает первоначальное окисление пирувата до ацетил-КоА, который затем расщепляется до CO
Как обсуждалось в главе 2, окислительный распад глюкозы и жирных кислот является основным источником метаболической энергии в клетках животных. Начальные стадии метаболизма глюкозы (гликолиз) происходят в цитозоле, где глюкоза превращается в пируват (см.). Затем пируват транспортируется в митохондрии, где происходит его полное окисление до CO 2 дает большую часть полезной энергии (АТФ), полученной в результате метаболизма глюкозы. Это включает первоначальное окисление пирувата до ацетил-КоА, который затем расщепляется до CO
Рисунок 10.2
Метаболизм в матриксе митохондрий. Пируват и жирные кислоты импортируются из цитозоля и превращаются в ацетил-КоА в митохондриальном матриксе.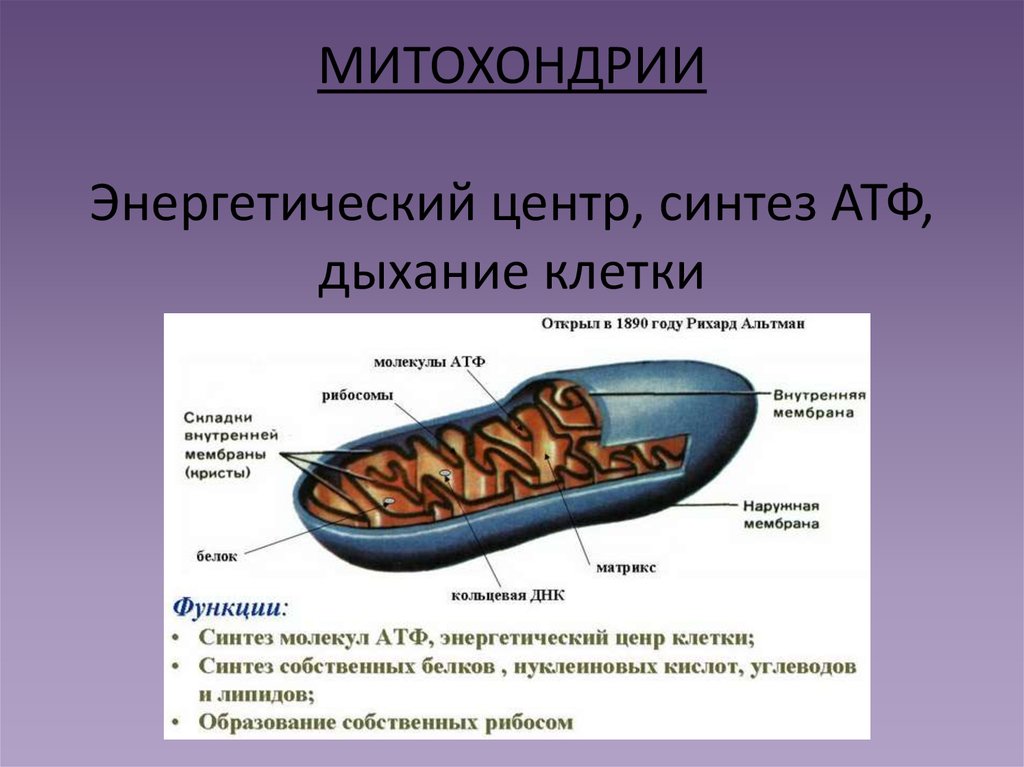 Затем ацетил-КоА окисляется до CO 2 через цикл лимонной кислоты, центральный путь окислительного метаболизма. (подробнее…)
Затем ацетил-КоА окисляется до CO 2 через цикл лимонной кислоты, центральный путь окислительного метаболизма. (подробнее…)
Окисление ацетил-КоА до СО 2 сопряжено с восстановлением НАД + и ФАД до НАДН и ФАДН 2 соответственно. Большая часть энергии, полученной в результате окислительного метаболизма, затем производится в процессе окислительного фосфорилирования (подробно обсуждаемого в следующем разделе), которое происходит во внутренней митохондриальной мембране. Высокоэнергетические электроны от NADH и FADH 2 переносятся через ряд носителей в мембране на молекулярный кислород. Энергия, полученная в результате этих реакций переноса электронов, преобразуется в потенциальную энергию, хранящуюся в градиенте протонов через мембрану, которая затем используется для запуска синтеза АТФ. Таким образом, внутренняя митохондриальная мембрана представляет собой основное место образования АТФ, и эта важная роль отражена в ее структуре.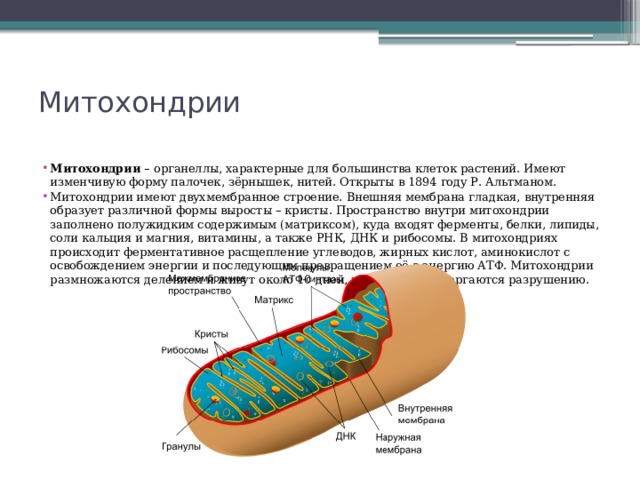 Во-первых, площадь его поверхности существенно увеличивается за счет складывания в кристы. Кроме того, внутренняя митохондриальная мембрана содержит необычно высокий процент (более 70%) белков, которые участвуют в окислительном фосфорилировании, а также в транспорте метаболитов (например, пирувата и жирных кислот) между цитозолем и митохондриями. В противном случае внутренняя мембрана непроницаема для большинства ионов и малых молекул — свойство, имеющее решающее значение для поддержания протонного градиента, который запускает окислительное фосфорилирование.
Во-первых, площадь его поверхности существенно увеличивается за счет складывания в кристы. Кроме того, внутренняя митохондриальная мембрана содержит необычно высокий процент (более 70%) белков, которые участвуют в окислительном фосфорилировании, а также в транспорте метаболитов (например, пирувата и жирных кислот) между цитозолем и митохондриями. В противном случае внутренняя мембрана непроницаема для большинства ионов и малых молекул — свойство, имеющее решающее значение для поддержания протонного градиента, который запускает окислительное фосфорилирование.
В отличие от внутренней мембраны, внешняя митохондриальная мембрана свободно проницаема для малых молекул. Это связано с тем, что он содержит белки, называемые поринами , которые образуют каналы, обеспечивающие свободную диффузию молекул размером менее 6000 дальтон. Таким образом, состав межмембранного пространства подобен цитозолю в отношении ионов и малых молекул. Следовательно, внутренняя митохондриальная мембрана является функциональным барьером для прохождения малых молекул между цитозолем и матриксом и поддерживает протонный градиент, который запускает окислительное фосфорилирование.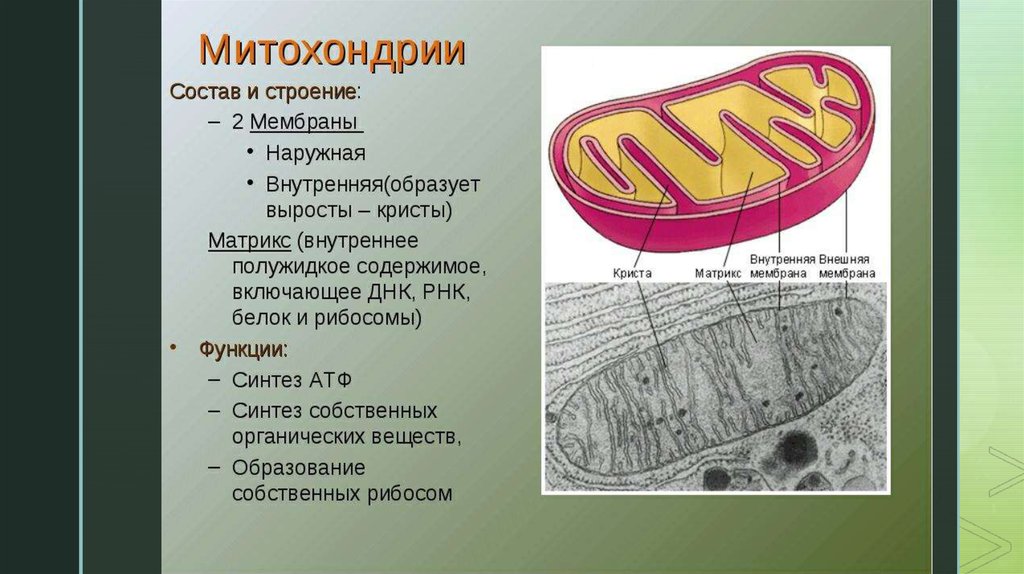
Генетическая система митохондрий
Митохондрии содержат свою собственную генетическую систему, которая отделена и отличается от ядерного генома клетки. Как было рассмотрено в главе 1, считается, что митохондрии произошли от бактерий, которые развили симбиотические отношения, в которых они жили внутри более крупных клеток (эндосимбиоз). Эта гипотеза недавно была подтверждена результатами анализа последовательности ДНК, выявившего поразительное сходство между геномами митохондрий и бактерии 9.0003 Риккетсия провазеки
Митохондриальные геномы обычно представляют собой кольцевые молекулы ДНК, как и у бактерий, которые присутствуют в нескольких копиях на органеллу.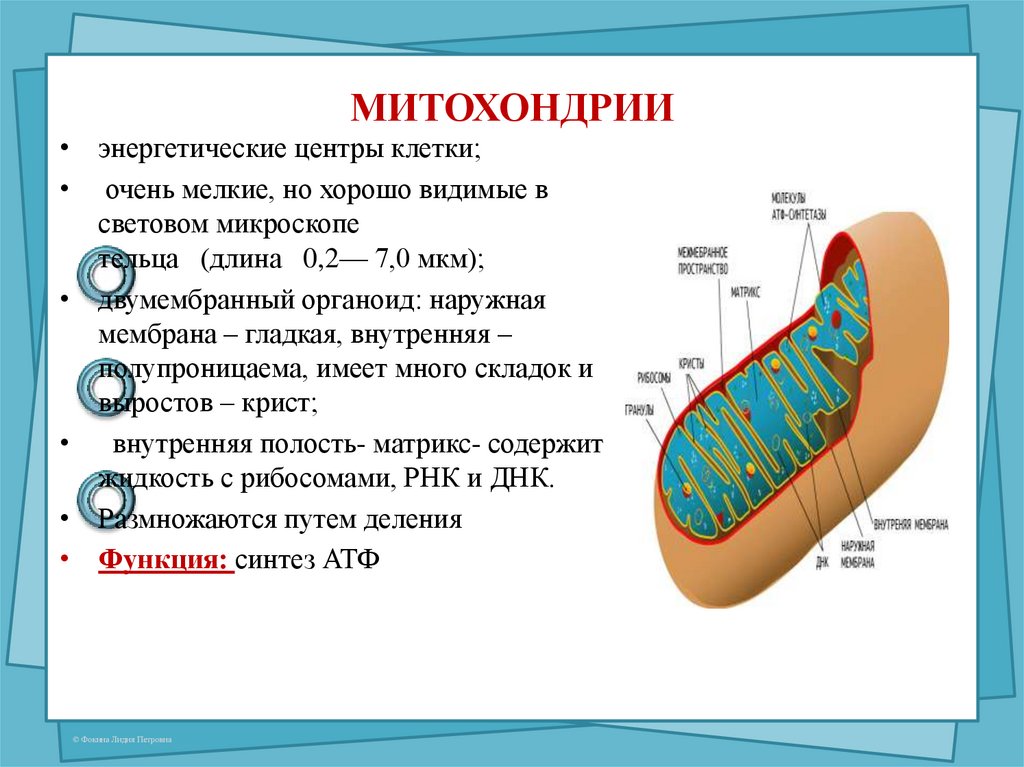 Они значительно различаются по размеру у разных видов. Геномы митохондрий человека и большинства других животных составляют всего около 16 т.п.н., но значительно большие митохондриальные геномы обнаружены у дрожжей (примерно 80 т.п.н.) и растений (более 200 т.п.н.). Однако эти более крупные митохондриальные геномы состоят преимущественно из некодирующих последовательностей и, по-видимому, не содержат значительно большего количества генетической информации. Например, самый большой секвенированный митохондриальный геном принадлежит 9 растениям.0003 Arabidopsis thaliana . Хотя митохондриальная ДНК Arabidopsis составляет приблизительно 367 т.п.н., она кодирует только 32 белка: чуть более чем в два раза больше, чем митохондриальная ДНК человека. Наибольшее количество митохондриальных генов обнаружено в митохондриальной ДНК простейшего Reclinomonas americana , которая составляет 69 т.п.н. и содержит 97 генов. Митохондриальный геном Reclinomonas , по-видимому, больше напоминает бактериальный геном, из которого произошли митохондрии, чем большинство современных митохондриальных геномов, которые кодируют лишь небольшое количество белков, являющихся важными компонентами системы окислительного фосфорилирования.
Они значительно различаются по размеру у разных видов. Геномы митохондрий человека и большинства других животных составляют всего около 16 т.п.н., но значительно большие митохондриальные геномы обнаружены у дрожжей (примерно 80 т.п.н.) и растений (более 200 т.п.н.). Однако эти более крупные митохондриальные геномы состоят преимущественно из некодирующих последовательностей и, по-видимому, не содержат значительно большего количества генетической информации. Например, самый большой секвенированный митохондриальный геном принадлежит 9 растениям.0003 Arabidopsis thaliana . Хотя митохондриальная ДНК Arabidopsis составляет приблизительно 367 т.п.н., она кодирует только 32 белка: чуть более чем в два раза больше, чем митохондриальная ДНК человека. Наибольшее количество митохондриальных генов обнаружено в митохондриальной ДНК простейшего Reclinomonas americana , которая составляет 69 т.п.н. и содержит 97 генов. Митохондриальный геном Reclinomonas , по-видимому, больше напоминает бактериальный геном, из которого произошли митохондрии, чем большинство современных митохондриальных геномов, которые кодируют лишь небольшое количество белков, являющихся важными компонентами системы окислительного фосфорилирования.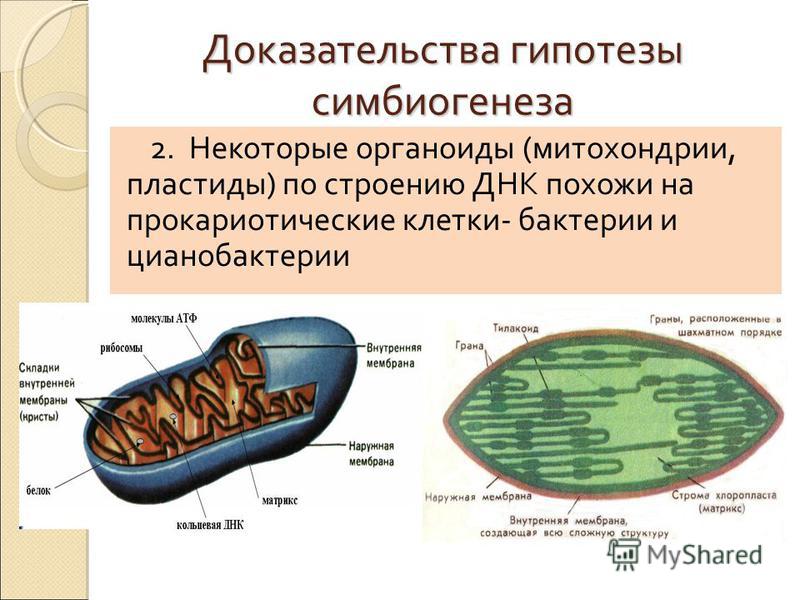 Кроме того, митохондриальные геномы кодируют все рибосомные РНК и большинство транспортных РНК, необходимых для трансляции этих кодирующих белок последовательностей в митохондриях. Другие митохондриальные белки кодируются ядерными генами, которые, как полагают, были перенесены в ядро из предкового митохондриального генома.
Кроме того, митохондриальные геномы кодируют все рибосомные РНК и большинство транспортных РНК, необходимых для трансляции этих кодирующих белок последовательностей в митохондриях. Другие митохондриальные белки кодируются ядерными генами, которые, как полагают, были перенесены в ядро из предкового митохондриального генома.
Митохондриальный геном человека кодирует 13 белков, участвующих в транспорте электронов и окислительном фосфорилировании (). Кроме того, митохондриальная ДНК человека кодирует 16S и 12S рРНК и 22 тРНК, которые необходимы для трансляции белков, кодируемых геномом органеллы. Две рРНК являются единственными РНК-компонентами митохондриальных рибосом животных и дрожжей, в отличие от трех рРНК бактериальных рибосом (23S, 16S и 5S). Однако митохондриальные ДНК растений также кодируют третью рРНК 5S. Митохондрии растений и простейших также различаются по импорту и использованию тРНК, кодируемых как ядерным, так и митохондриальным геномом, тогда как в митохондриях животных все тРНК кодируются органеллой.
Рисунок 10.3
Митохондриальный геном человека. Геном содержит 13 белоккодирующих последовательностей, которые обозначены как компоненты дыхательных комплексов I, III, IV или V. Кроме того, геном содержит гены 12S и 16S рРНК и 22 тРНК, которые обозначены (подробнее…)
Небольшое количество тРНК, кодируемых митохондриальным геномом, подчеркивает важную особенность митохондриальной генетической системы — использование несколько иного генетического кода, отличного от «универсального» генетического кода, используемого как прокариотическими, так и эукариотическими клетками () . Как обсуждалось в главе 3, существует 64 возможных триплетных кодона, 61 из которых кодирует 20 различных аминокислот, входящих в состав белков (см. табл. 3.1). Многие тРНК как в прокариотических, так и в эукариотических клетках способны распознавать более одного кодона в мРНК из-за «колебания», которое допускает некоторое неправильное спаривание между антикодоном тРНК и третьим положением некоторых комплементарных кодонов (см. Однако для трансляции универсального кода в соответствии с правилами колебания требуется не менее 30 различных тРНК. Тем не менее, митохондриальная ДНК человека кодирует только 22 вида тРНК, и это единственные тРНК, используемые для трансляции митохондриальных мРНК. Это достигается крайней формой колебания, при которой U в антикодоне тРНК может спариваться с любым из четырех оснований в третьем положении кодона мРНК, позволяя одной тРНК распознавать четыре кодона. Кроме того, некоторые кодоны определяют в митохондриях другие аминокислоты, чем в универсальном коде.
Однако для трансляции универсального кода в соответствии с правилами колебания требуется не менее 30 различных тРНК. Тем не менее, митохондриальная ДНК человека кодирует только 22 вида тРНК, и это единственные тРНК, используемые для трансляции митохондриальных мРНК. Это достигается крайней формой колебания, при которой U в антикодоне тРНК может спариваться с любым из четырех оснований в третьем положении кодона мРНК, позволяя одной тРНК распознавать четыре кодона. Кроме того, некоторые кодоны определяют в митохондриях другие аминокислоты, чем в универсальном коде.
Таблица 10.1
Различия между универсальным и митохондриальным генетическими кодами.
Как и ДНК ядерных геномов, митохондриальная ДНК может быть изменена мутациями, которые часто вредны для органеллы. Поскольку почти все митохондрии оплодотворенных яйцеклеток формируются ооцитом, а не спермой, мутации зародышевой линии в митохондриальной ДНК передаются следующему поколению от матери. Такие мутации связаны с рядом заболеваний.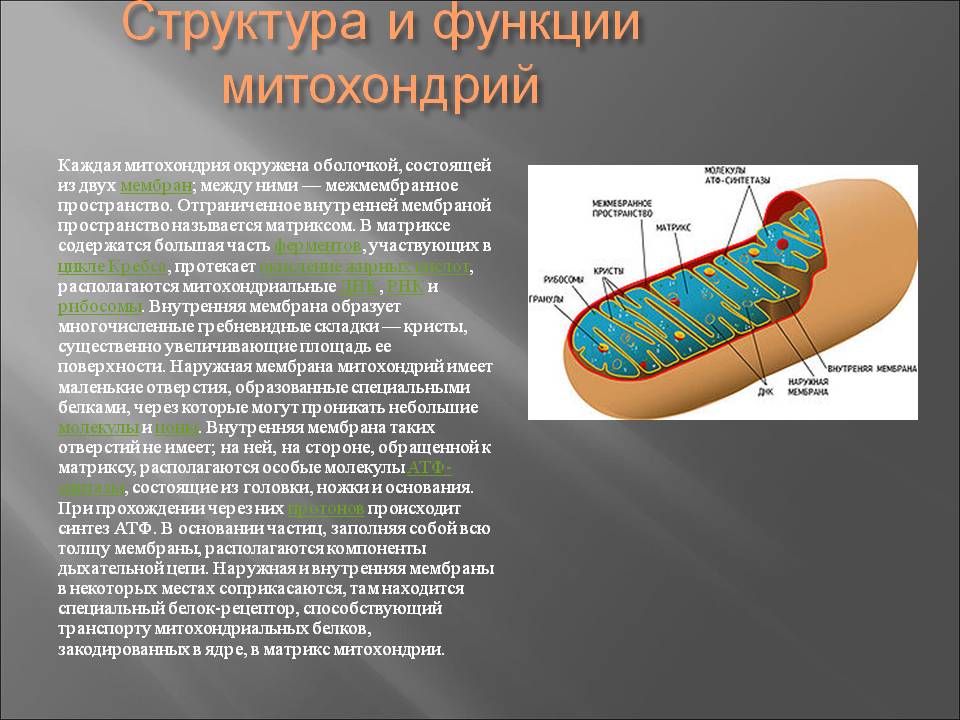 Например, наследственная оптическая нейропатия Лебера, заболевание, приводящее к слепоте, может быть вызвано мутациями в митохондриальных генах, кодирующих компоненты цепи переноса электронов. Кроме того, было высказано предположение, что прогрессирующее накопление мутаций в митохондриальной ДНК в течение жизни людей способствует процессу старения.
Например, наследственная оптическая нейропатия Лебера, заболевание, приводящее к слепоте, может быть вызвано мутациями в митохондриальных генах, кодирующих компоненты цепи переноса электронов. Кроме того, было высказано предположение, что прогрессирующее накопление мутаций в митохондриальной ДНК в течение жизни людей способствует процессу старения.
Импорт белков и митохондриальная сборка
В отличие от РНК-компонентов митохондриального аппарата трансляции (рРНК и тРНК), большинство митохондриальных геномов не кодируют белки, необходимые для репликации, транскрипции или трансляции ДНК. Вместо этого гены, кодирующие белки, необходимые для репликации и экспрессии митохондриальной ДНК, содержатся в ядре. Кроме того, ядро содержит гены, кодирующие большинство митохондриальных белков, необходимых для окислительного фосфорилирования, и все ферменты, участвующие в митохондриальном метаболизме (например, ферменты цикла лимонной кислоты). Белки, кодируемые этими генами (более 95% митохондриальных белков) синтезируются на свободных цитозольных рибосомах и импортируются в митохондрии в виде законченных полипептидных цепей. Из-за двухмембранной структуры митохондрий импорт белков значительно сложнее, чем перенос полипептида через одиночный фосфолипидный бислой. Белки, нацеленные на матрикс, должны пересекать как внутреннюю, так и внешнюю митохондриальные мембраны, в то время как другие белки должны быть отсортированы по отдельным компартментам внутри органеллы (например, межмембранное пространство).
Из-за двухмембранной структуры митохондрий импорт белков значительно сложнее, чем перенос полипептида через одиночный фосфолипидный бислой. Белки, нацеленные на матрикс, должны пересекать как внутреннюю, так и внешнюю митохондриальные мембраны, в то время как другие белки должны быть отсортированы по отдельным компартментам внутри органеллы (например, межмембранное пространство).
Импорт белков в матрикс является наиболее изученным аспектом сортировки митохондриальных белков (). Большинство белков нацелены на митохондрии с помощью амино-концевых последовательностей из 20-35 аминокислот (называемых препоследовательностями ), которые удаляются протеолитическим расщеплением после их импорта в органеллы. Препоследовательности митохондриальных белков, впервые описанные Готфридом Шацем, содержат множество положительно заряженных аминокислотных остатков, обычно в амфипатической α-спирали. Первым шагом в импорте белков является связывание этих препоследовательностей с рецепторами на поверхности митохондрий. Затем полипептидные цепи встраиваются в белковый комплекс, который направляет транслокацию через внешнюю мембрану (транслоказа внешней мембраны или комплекс Тома). Затем белки переносятся во второй белковый комплекс внутренней мембраны (транслоказу внутренней мембраны или комплекс Тима). Для непрерывной транслокации белка требуется электрохимический потенциал, устанавливаемый на внутренней митохондриальной мембране во время переноса электронов. Как обсуждалось в следующем разделе этой главы, перенос высокоэнергетических электронов от НАДН и ФАДН 2 к молекулярному кислороду сопряжен с переносом протонов из митохондриального матрикса в межмембранное пространство. Поскольку протоны являются заряженными частицами, этот перенос создает электрический потенциал на внутренней мембране, а матрица является отрицательной. Во время импорта белка этот электрический потенциал управляет перемещением положительно заряженной препоследовательности.
Затем полипептидные цепи встраиваются в белковый комплекс, который направляет транслокацию через внешнюю мембрану (транслоказа внешней мембраны или комплекс Тома). Затем белки переносятся во второй белковый комплекс внутренней мембраны (транслоказу внутренней мембраны или комплекс Тима). Для непрерывной транслокации белка требуется электрохимический потенциал, устанавливаемый на внутренней митохондриальной мембране во время переноса электронов. Как обсуждалось в следующем разделе этой главы, перенос высокоэнергетических электронов от НАДН и ФАДН 2 к молекулярному кислороду сопряжен с переносом протонов из митохондриального матрикса в межмембранное пространство. Поскольку протоны являются заряженными частицами, этот перенос создает электрический потенциал на внутренней мембране, а матрица является отрицательной. Во время импорта белка этот электрический потенциал управляет перемещением положительно заряженной препоследовательности.
Рисунок 10.4
Импорт белков в митохондрии. Белки направляются в митохондрии с помощью амино-концевой последовательности, содержащей положительно заряженные аминокислоты. Белки поддерживаются в частично развернутом состоянии за счет ассоциации с цитозольным Hsp70 и являются (далее…)
Белки направляются в митохондрии с помощью амино-концевой последовательности, содержащей положительно заряженные аминокислоты. Белки поддерживаются в частично развернутом состоянии за счет ассоциации с цитозольным Hsp70 и являются (далее…)
Для перемещения через митохондриальную мембрану белки должны быть хотя бы частично развернуты. Следовательно, импорт белка в митохондрии требует молекулярных шаперонов в дополнение к мембранным белкам, участвующим в транслокации (see). На цитозольной стороне члены семейства шаперонов Hsp70 поддерживают белки в частично развернутом состоянии, так что они могут быть встроены в митохондриальную мембрану. Когда они пересекают внутреннюю мембрану, развернутые полипептидные цепи связываются с другим членом семейства Hsp70, который связан с комплексом Tim и действует как двигатель, который управляет импортом белка. Затем полипептид переносится на шаперон семейства Hsp60 (шаперонин), внутри которого происходит сворачивание белка. Поскольку эти взаимодействия полипептидных цепей с молекулярными шаперонами зависят от АТФ, для импорта белка требуется АТФ как снаружи, так и внутри митохондрий, в дополнение к электрическому потенциалу на внутренней мембране.
Как отмечалось выше, некоторые митохондриальные белки нацелены на внешнюю мембрану, внутреннюю мембрану или межмембранное пространство, а не на матрикс, поэтому необходимы дополнительные механизмы для направления этих белков в правильный субмитохондриальный компартмент. Эти белки направляются к месту назначения с помощью второго сортировочного сигнала, следующего за положительно заряженной предварительной последовательностью, которая направляет митохондриальный импорт. Нацеливание белков на митохондриальные мембраны, по-видимому, опосредовано гидрофобными последовательностями остановки-переноса, которые останавливают транслокацию полипептидных цепей через комплексы Тима или Тома, что приводит к их встраиванию во внутреннюю или внешнюю митохондриальные мембраны, соответственно. Белки могут направляться в межмембранное пространство с помощью нескольких разных механизмов (14). Некоторые белки переносятся через внешнюю мембрану через комплекс Тома, но затем высвобождаются в межмембранном пространстве вместо переноса в комплекс Тима.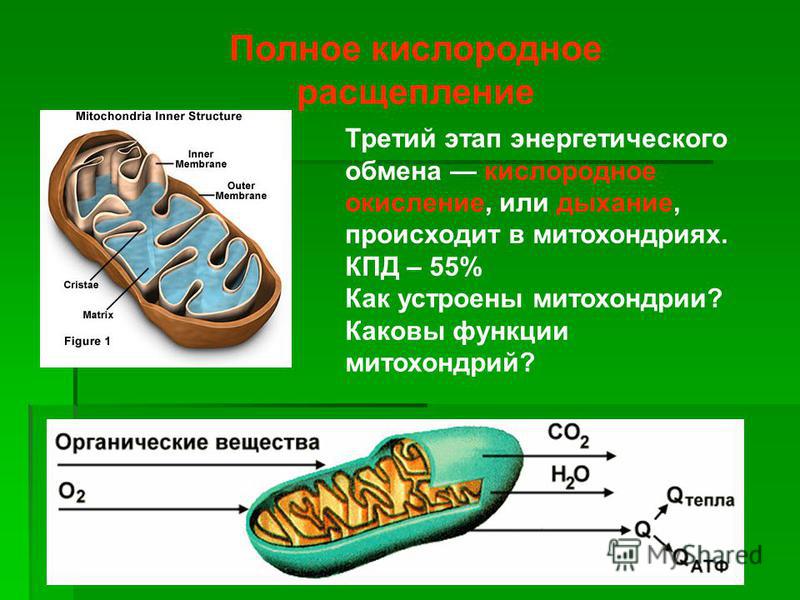 Другие белки переносятся в комплекс Tim, но затем высвобождаются в межмембранное пространство в результате расщепления гидрофобных стоп-трансферных последовательностей. Другие белки могут быть полностью импортированы в митохондриальный матрикс, а затем экспортированы обратно через внутреннюю мембрану в межмембранное пространство.
Другие белки переносятся в комплекс Tim, но затем высвобождаются в межмембранное пространство в результате расщепления гидрофобных стоп-трансферных последовательностей. Другие белки могут быть полностью импортированы в митохондриальный матрикс, а затем экспортированы обратно через внутреннюю мембрану в межмембранное пространство.
Рисунок 10.5
Вставка белков митохондриальной мембраны. Белки, предназначенные для митохондриальных мембран, содержат гидрофобные последовательности остановки-переноса, которые останавливают их транслокацию через комплексы Тома или Тима и приводят к их включению во внешний или (подробнее…)
Рисунок 10.6
Сортировка белков в межмембранное пространство. Белки могут направляться в межмембранное пространство с помощью нескольких механизмов. Некоторые белки (I) транслоцируются через комплекс Тома и высвобождаются в межмембранное пространство. Переносятся другие белки (II) (далее…)
Из цитозоля импортируются не только белки, но и фосфолипиды митохондриальных мембран.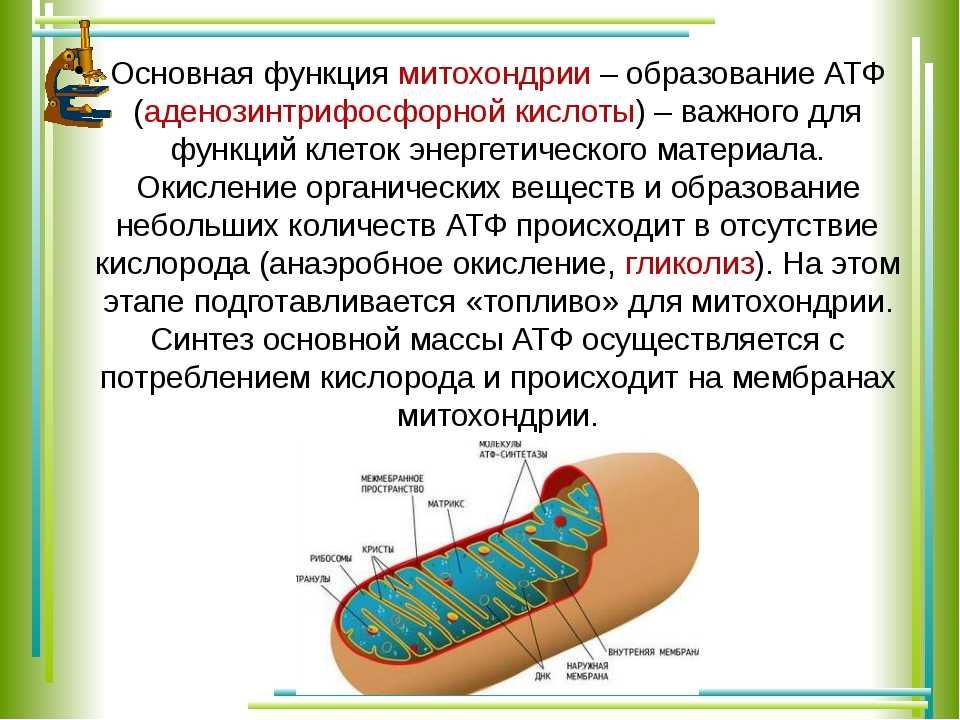
Leave A Comment