
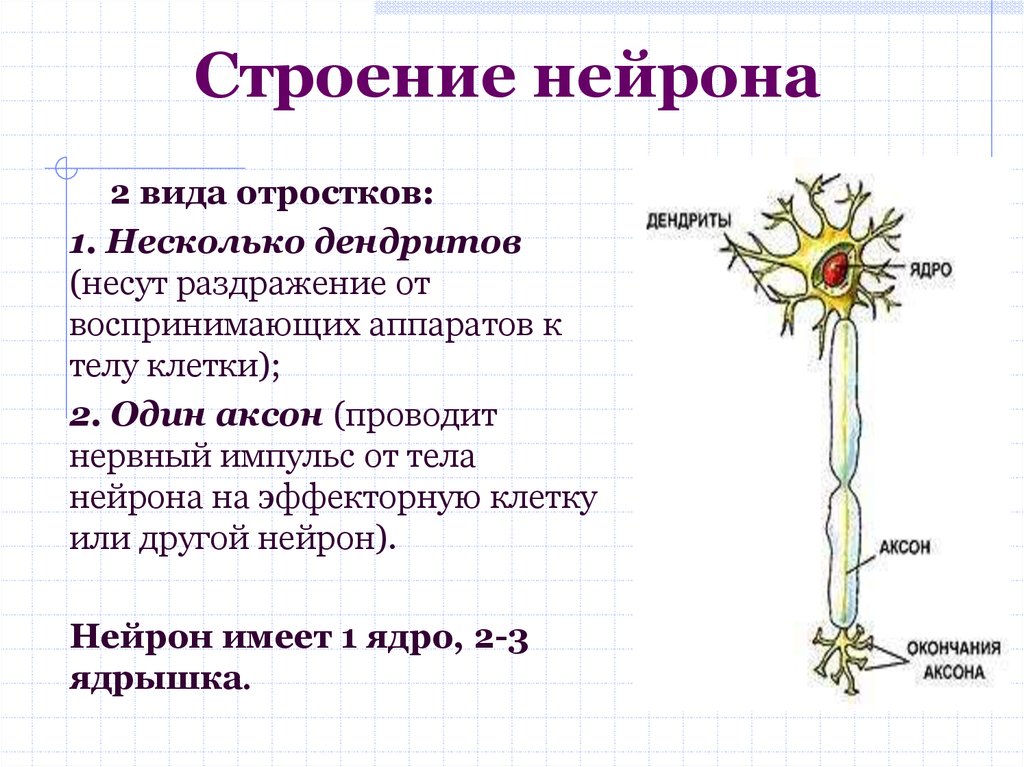
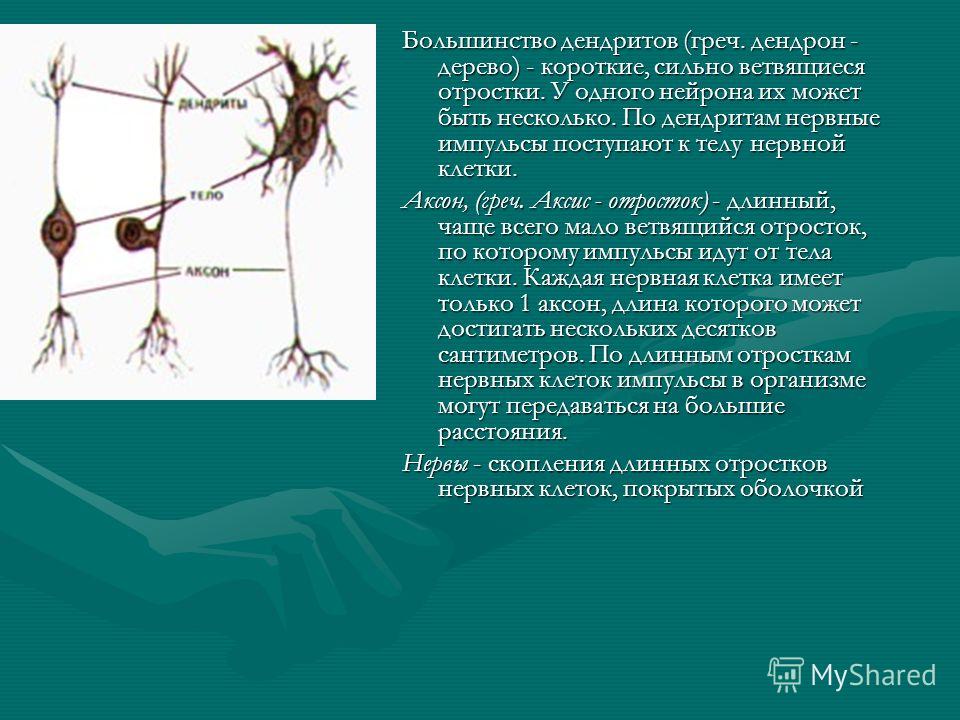
дендриты и аксоны. Отростки могут быть длинными и короткими.
Большинство дендритов — короткие, сильно ветвящиеся отростки. У одного нейрона их может быть несколько. По дендритам нервные импульсы поступают к телу нервной клетки.
Аксон — длинный, чаще всего мало ветвящийся отросток, по которому импульсы идут от тела клетки. Каждая нервная клетка имеет только 1 аксон, длина которого может достигать нескольких десятков сантиметров. По длинным отросткам нервных клеток импульсы в организме могут передаваться на большие расстояния.
Длинные отростки часто покрыты оболочкой из жироподобного вещества белого цвета. Их скопления в центральной нервной системе образуют белое вещество. Короткие отростки и тела нейронов не имеют такой оболочки. Их скопления образуют серое вещество.
Нейроны
различаются по форме и функциям. Одни
нейроны, чувствительные, передают
импульсы от органов чувств в спинной и
головной мозг. Тела чувствительных
нейронов лежат на пути к центральной
нервной системе в нервных узлах.
Нервы — скопления длинных отростков нервных клеток, покрытых оболочкой. Нервы, состоящие из аксонов двигательных нейронов, называются двигательными нервами. Чувствительные нервы состоят из дендритов чувствительных нейронов. Большинство нервов содержат и аксоны и дендриты. Такие нервы называются смешанными. По ним импульсы идут в двух направлениях — к центральной нервной системе и от нее к органам.
Отделы нервной
системы. Нервная система состоит из
центрального и периферического отделов.
Центральный отдел представлен головным
и спинным мозгом, защищенным оболочками
из соединительной ткани.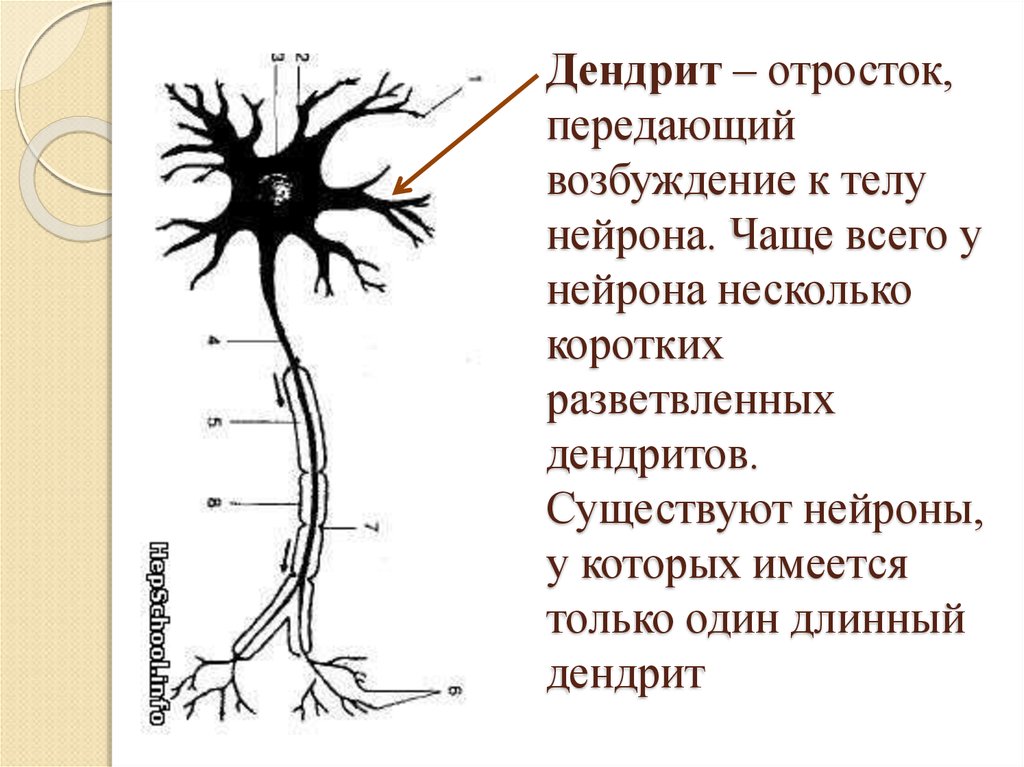 К периферическому
отделу относятся нервы и нервные узлы.
К периферическому
отделу относятся нервы и нервные узлы.
Часть нервной системы, которая регулирует работу скелетных мышц, называют соматической . Посредством соматической нервной системы человек может управлять движениями, произвольно вызывать или прекращать их. Часть нервной системы, регулирующую деятельность внутренних органов (сердца, желудка, желез и т.д.), называют автономной .Работа автономной нервной системы не подчиняется человека. Нельзя, например, по желанию остановить сердце, ускорить процесс пищеварения, задержать потоотделение.
В автономной нервной системе различают два отдела: симпатический и парасимпатический. Большинство внутренних органов снабжаются нервами этик двух отделов. Как правило, они оказывают противоположные влияния на органы. Например, симпатический нерв усиливает и ускоряет работу сердца, а парасимпатический — замедляет и ослабляет ее.
Рефлекс.
Рефлекторная дуга. Ответную реакцию
организма на раздражение, осуществляемую
и контролируемую центральной нервной
системой, называют рефлексом.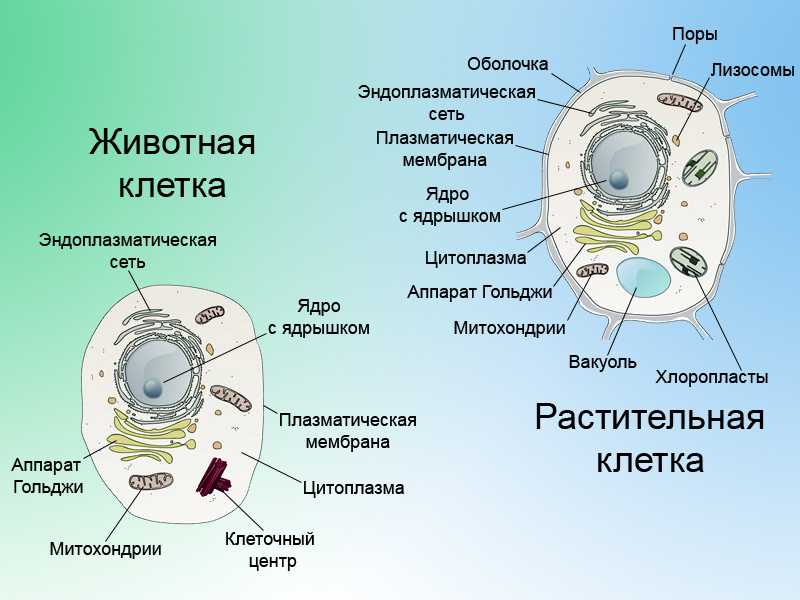 Путь, по
которому проводятся нервные импульсы
при осуществлении рефлекса, называют
рефлекторной дугой. Рефлекторная дуга
состоит из пяти частей: рецептора,
чувствительного пути, участка центральной
нервной системы, двигательного пути и
рабочего органа.
Путь, по
которому проводятся нервные импульсы
при осуществлении рефлекса, называют
рефлекторной дугой. Рефлекторная дуга
состоит из пяти частей: рецептора,
чувствительного пути, участка центральной
нервной системы, двигательного пути и
рабочего органа.
Рефлекторная дуга начинается рецептором. Каждый рецептор воспринимает определенный раздражитель: свет, звук, прикосновение, запах, температуру и др. Рецепторы преобразуют чти раздражители в нервные импульсы-сигналы нервной системы. Нервные импульсы имеют электрическую природу, распространяются но мембранам длинных отростков нейронов и одинаковы у животных и человека.
От рецептора
нервные импульсы по чувствительному
пути передаются в центральную нервную
систему. Этот путь образован чувствительным
нейроном. От центральной нервной системы
импульсы по двигательному пути идут к
рабочему органу. В состав большинства
рефлекторных дуг входят и вставочные
нейроны, которые находятся как в спинном,
так и в головном мозге.
Рефлекс — ответная реакция организма на раздражение, осуществляемая при участии нервной системы.
Рефлекторная дуга — путь, по которому проходят нервные импульсы (рис.5).
Рефлексы человека разнообразны. Некоторые из них очень просты. Например, одергивание руки в ответ на укол или ожог кожи, обильное выделение слез под действием веществ, раздражающих глаза, чихание при попадании посторонних частиц в носовую полость. Во время рефлекторной реакции рецепторы рабочих органов передают сигналы в центральную нервную систему, которая контролирует, насколько реакция эффективна.
Таким образом, основной принцип работы нервной системы рефлекторный.
Рефлекторное кольцо — совокупность структур нервной системы, участвующих в осуществлении рефлекса и передаче информации о характере и силе рефлекторного действия в центральной нервной системе.
СИ́НАПС ,зона
контакта между нейронами и другими
образованиями (нервными, мышечными или
железистыми клетками), служащая для
передачи информации от клетки, генерирующей
нервный импульс к другим клеткам.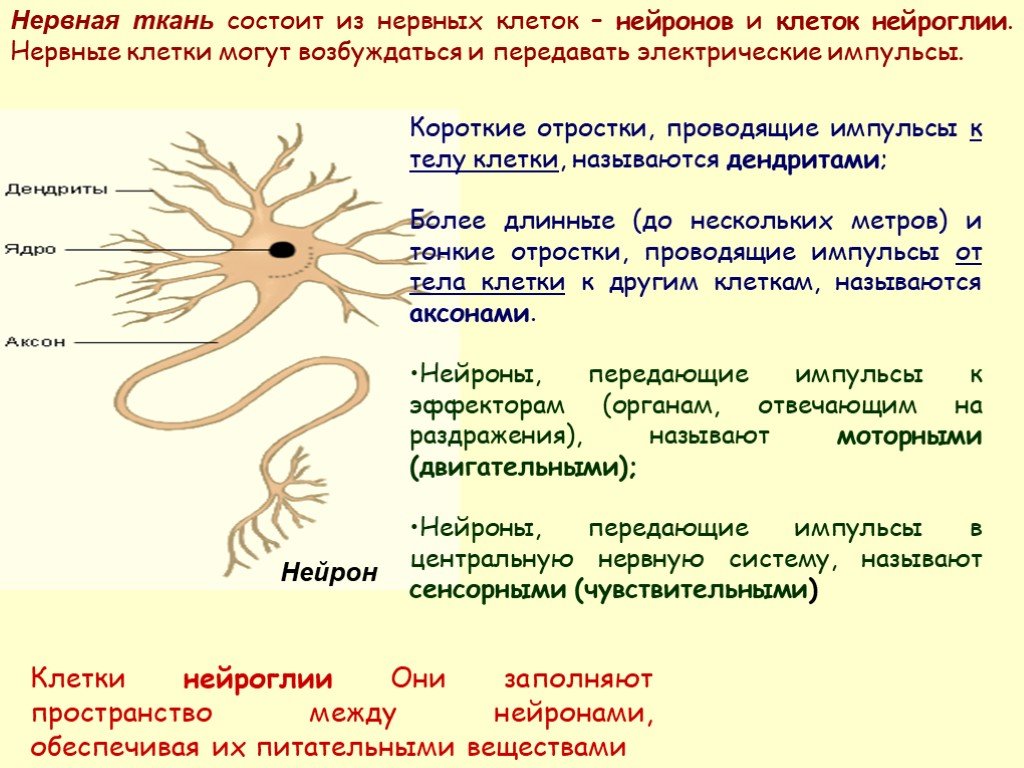 Термин
ввел Ч. Шеррингтон в 1897.
Термин
ввел Ч. Шеррингтон в 1897.
Синапс состоит из трех отделов: пресинаптического (нейрон, посылающий сигналы), постсинаптического (клетка, принимающая сигналы) и соединяющей их структуры (синаптическая щель). В тех случаях, когда речь идет о контактах между нервными клетками, синапсы могут образовываться между аксонамии сомой, аксонами и дендритами, аксонами и аксонами, дендритами и дендритами, а также между сомой и дендритами нейронов. В зависимости от способа передачи возбуждения выделяют химические (наиболее распространенные) и электрические синапсы. Существуют также смешанные синапсы, сочетающие оба механизма передачи.
Рецепторы ,специальные чувствительные образования, воспринимающие и преобразующие раздражения из внешней или внутренней среды организма и передающие информацию о действующем агенте в нервную систему.
Маленькие митохондрии усиливают взаимодействие между нейронами
Нейроны — это клетки с очень необычной формой: от компактного «тела» отходят множественные тонкие отростки — дендриты и аксоны.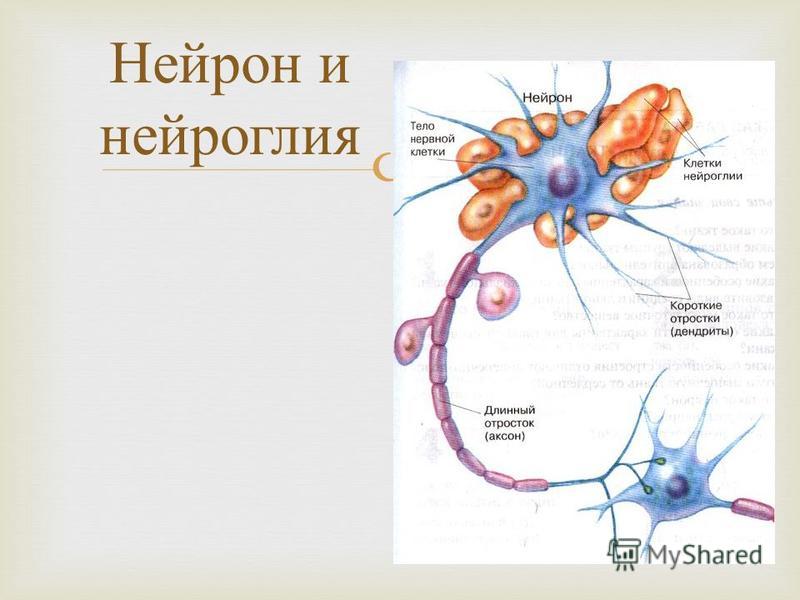
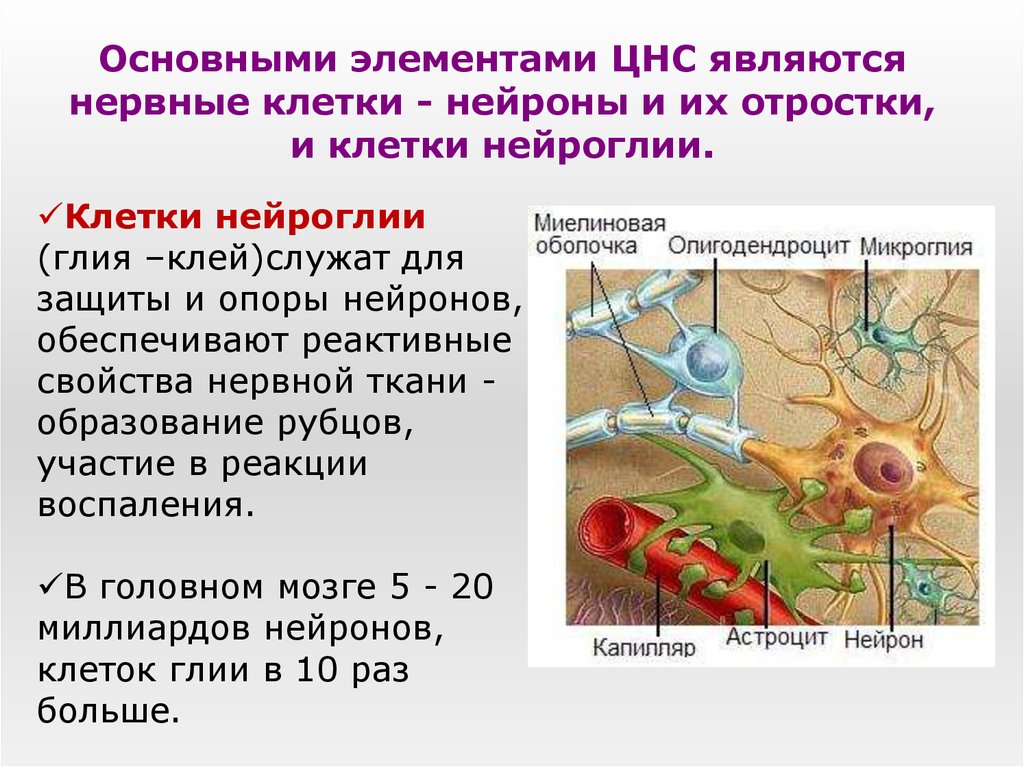
Рис. 1. Пирамидальный нейрон (a) и митохондрии, подсвеченные зеленым в его дендритах (b) и аксонах (с). На графиках показаны линейные размеры митохондрий в аксонах и дендритах (g) и доля длины аксонов и дендритов, которая занята митохондриями (h). In vivo — это данные, полученные на зафиксированных срезах коры головного мозга мышей, в которые предварительно были введены плазмиды белков, избирательно окрашивающих матрикс и митохондрии. In vitro — данные, полученные аналогичным образом в культуре нейронов. Фотография отображает инвертированные цвета, поэтому зеленый флуоресцирующий белок (GFP) здесь выглядит пурпурным, а красный флуоресцирующий белок, окрашивающий митохондрии (mt-DsRED) — зеленым. Рисунок из обсуждаемой статьи в Nature Communications
Нейроны — это клетки, составляющие основные функциональные элементы нервной системы. Структурными элементами нейрона являются тело клетки, в котором располагается ядро с генетическим материалом, и два типа отростков: дендриты(обычно их много, они тонкие и обильно ветвящиеся) и аксон (более толстый отросток, который, однако, обычно тоже ветвится на некотором расстоянии от тела клетки; он всегда один). Оба типа отростков могут формировать множественные контакты с другими клетками, однако функция у них разная: дендриты принимают возбуждающие или тормозящие сигналы от других нейронов и передают их к телу нейрона, а аксон обеспечивает передачу сигналов к следующим нейронам (либо к клеткам мышц).
Оба типа отростков могут формировать множественные контакты с другими клетками, однако функция у них разная: дендриты принимают возбуждающие или тормозящие сигналы от других нейронов и передают их к телу нейрона, а аксон обеспечивает передачу сигналов к следующим нейронам (либо к клеткам мышц).
Передача сигналов от одного нейрона к другому осуществляется через особые образования — синапсы, в которых мембраны передающей и принимающей клеток формируют выпячивания и тесно сближаются друг с другом. Нейроны млекопитающих (таких, как мышь или человек) общаются при помощи химических синапсов. Это значит, что передающий нейрон выделяет через пресинаптическую мембрану сигнальные молекулы — нейромедиаторы, а принимающая клетка улавливает эти молекулы при помощи специальных рецепторов, встроенных в постсинаптическую мембрану. Связывание рецептора с нейромедиатором и вызывает в принимающей клетке те или иные реакции.
Однако в контексте обсуждаемого ниже исследования нам особенно важно рассмотреть то, что происходит в пресинапсической части.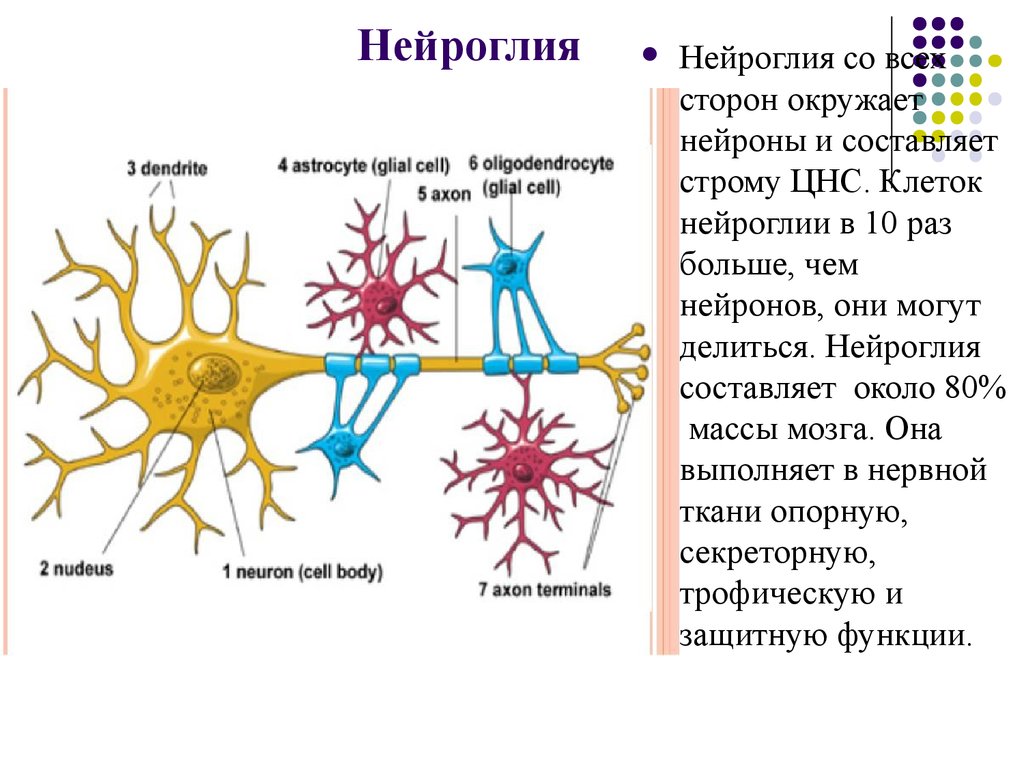 Потенциал действия, генерируемый нейроном в ответ на входящие сигналы, распространяется по аксону (в форме токов ионов натрия и калия), достигает терминалей с синапсами и там вызывает открывание кальциевых каналов. Кальций поступает внутрь из межклеточного пространства и активирует белки, встроенные в мембраны синаптических везикул. Благодаря этой активации мембраны везикул сливаются с пресинаптической мембраной, и их содержимое — нейромедиаторы — выбрасывается в синаптическую щель. Таким образом, критическим моментом для передачи сигнала является повышение концентрации кальция в терминали аксона. Весь этот процесс замечательно описан в комиксе «Как происходит выделение нейромедиатора».
Потенциал действия, генерируемый нейроном в ответ на входящие сигналы, распространяется по аксону (в форме токов ионов натрия и калия), достигает терминалей с синапсами и там вызывает открывание кальциевых каналов. Кальций поступает внутрь из межклеточного пространства и активирует белки, встроенные в мембраны синаптических везикул. Благодаря этой активации мембраны везикул сливаются с пресинаптической мембраной, и их содержимое — нейромедиаторы — выбрасывается в синаптическую щель. Таким образом, критическим моментом для передачи сигнала является повышение концентрации кальция в терминали аксона. Весь этот процесс замечательно описан в комиксе «Как происходит выделение нейромедиатора».
Но затем избыток кальция откачивается из цитоплазмы. И тогда активаторный сигнал прекращается до индукции нового потенциала действия.
Работа нейронов очень энергозатратна. Неудивительно поэтому, что митохондрии чрезвычайно важны для их функционирования. Пирамидальные нейроны коры головного мозга содержат огромное количество митохондрий, причем форма этих митохондрий различается в разных частях клетки.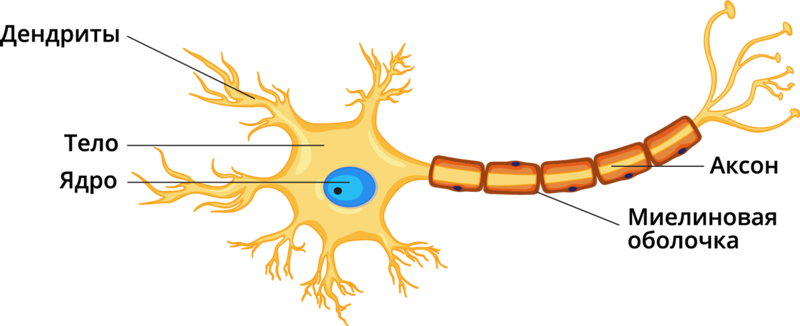 В дендритах митохондрии имеют сильно удлиненную форму, напоминая под микроскопом макаронины. В теле клетки такие же удлиненные митохондрии сливаются между собой, формируя густую сеть. Но в аксонах митохондрии выглядят иначе: они гораздо более короткие, многие — практически шарообразной формы. Эти митохондрии без конца снуют туда и обратно, используя в качестве рельсов цитоскелетные элементы — микротрубочки, протянутые вдоль аксона. Эта динамика хорошо видна на видео, снятом в ходе прошлогодней работы польской группы исследователей (T. M. Stępkowski et al., 2017. mitoLUHMES: An Engineered Neuronal Cell Line for the Analysis of the Motility of Mitochondria)
В дендритах митохондрии имеют сильно удлиненную форму, напоминая под микроскопом макаронины. В теле клетки такие же удлиненные митохондрии сливаются между собой, формируя густую сеть. Но в аксонах митохондрии выглядят иначе: они гораздо более короткие, многие — практически шарообразной формы. Эти митохондрии без конца снуют туда и обратно, используя в качестве рельсов цитоскелетные элементы — микротрубочки, протянутые вдоль аксона. Эта динамика хорошо видна на видео, снятом в ходе прошлогодней работы польской группы исследователей (T. M. Stępkowski et al., 2017. mitoLUHMES: An Engineered Neuronal Cell Line for the Analysis of the Motility of Mitochondria)
Группа ученых из США, решила продвинуться дальше и разобраться в механизме формирования особенных аксональных митохондрий и в том, есть ли какой-то специфический функциональный смысл в этой их особенности. Их статья на прошлой неделе была опубликована в журнале Nature Communications.
Ученые предположили, что размеры митохондрий могут зависеть от контроля их деления.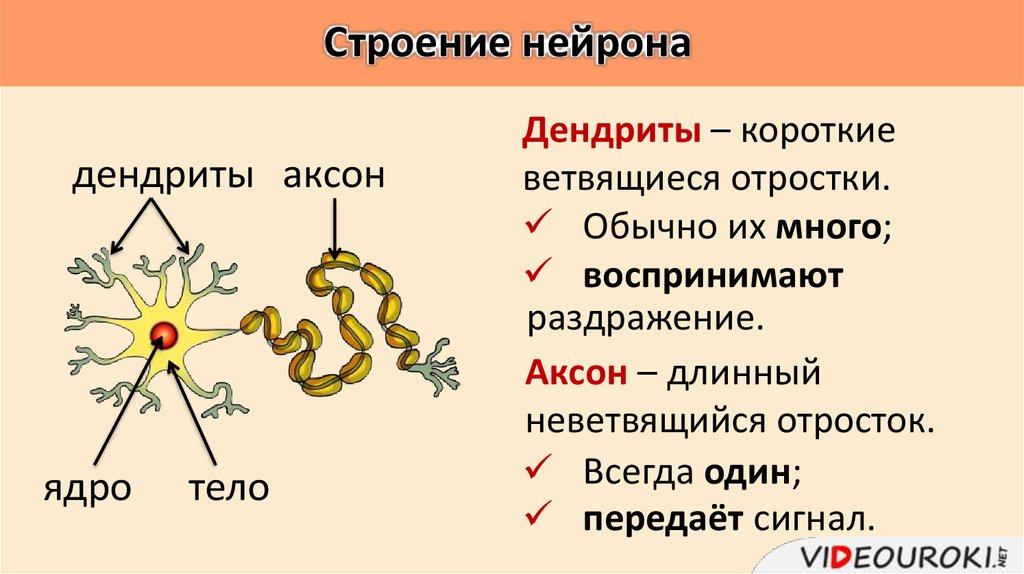 На сегодняшний день главные молекулярные игроки, задействованные в этом процессе известны (K. Basu et al., 2017. Molecular mechanism of DRP1 assembly studied in vitro by cryo-electron microscopy). Один из этих игроков — белок Mff. Этот белок встраивается в наружную мембрану митохондрий и служит рецептором для моторного белка Drp1. Drp1 формирует кольцо, перетягивающее митохондрию пополам (будто ремнем) и таким образом разделяет ее на две отдельные митохондрии меньшего размера. На рис. 2 показаны и другие молекулы, участвующие в этом процессе. С другой стороны, митохондрии могут и увеличиваться в размере за счет слияния друг с другом, зависящего от другого комплекта белков-регуляторов. В итоге размер и морфология митохондрии определяется, по-видимому, соотношением частоты слияний и разделений.
На сегодняшний день главные молекулярные игроки, задействованные в этом процессе известны (K. Basu et al., 2017. Molecular mechanism of DRP1 assembly studied in vitro by cryo-electron microscopy). Один из этих игроков — белок Mff. Этот белок встраивается в наружную мембрану митохондрий и служит рецептором для моторного белка Drp1. Drp1 формирует кольцо, перетягивающее митохондрию пополам (будто ремнем) и таким образом разделяет ее на две отдельные митохондрии меньшего размера. На рис. 2 показаны и другие молекулы, участвующие в этом процессе. С другой стороны, митохондрии могут и увеличиваться в размере за счет слияния друг с другом, зависящего от другого комплекта белков-регуляторов. В итоге размер и морфология митохондрии определяется, по-видимому, соотношением частоты слияний и разделений.
Рис. 2. Механизм разделения митохондрий при участии Drp1 и MFF рецептора. Рисунок из статьи K. Basu et al., 2017. Molecular mechanism of DRP1 assembly studied in vitro by cryo-electron microscopy
Главным инструментом в руках исследователей стала РНК-интерференция(см.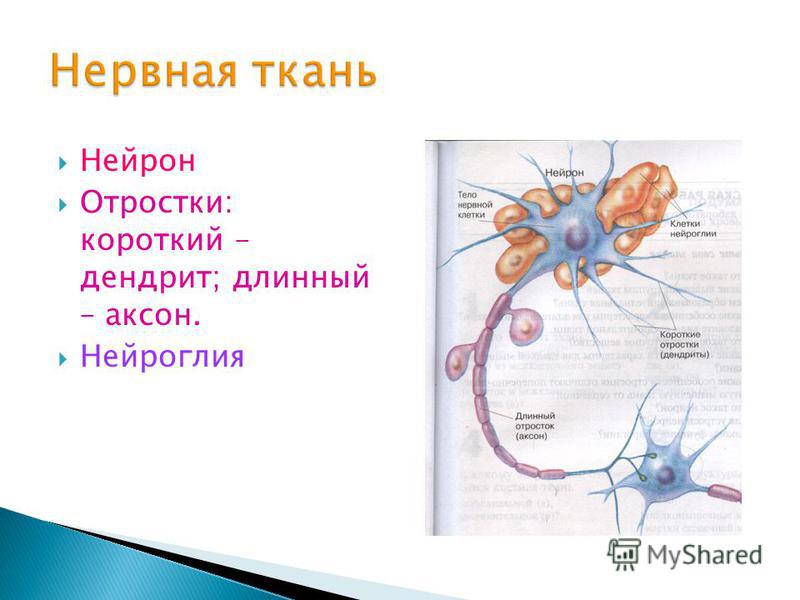 новость У низших животных обнаружены системы управления генами и транспозонами при помощи малых РНК, «Элементы», 03.10.2008). В мышиных нейронах при помощи РНК-интерференции подавляли работу гена Mff, вводя в клетки короткие шпилечные РНК. Контрольные образцы также обрабатывали короткими шпилечными РНК, которые, однако, не должны были влиять на работу каких-либо генов. В итоге было установлено, что при подавлении гена Mff в аксонах действительно стали появляться удлиненные митохондрии. Причем в сумме эти митохондрии стали занимать значительно большую часть внутреннего пространства аксона, чем в контрольных клетках (рис. 3).
новость У низших животных обнаружены системы управления генами и транспозонами при помощи малых РНК, «Элементы», 03.10.2008). В мышиных нейронах при помощи РНК-интерференции подавляли работу гена Mff, вводя в клетки короткие шпилечные РНК. Контрольные образцы также обрабатывали короткими шпилечными РНК, которые, однако, не должны были влиять на работу каких-либо генов. В итоге было установлено, что при подавлении гена Mff в аксонах действительно стали появляться удлиненные митохондрии. Причем в сумме эти митохондрии стали занимать значительно большую часть внутреннего пространства аксона, чем в контрольных клетках (рис. 3).
Рис. 3. Слияние и деление митохондрий внутри аксона. Здесь показана серия фотографий, зафиксировавших динамику митохондрий на протяжении нескольких минут (время от начала съемки в минутах показано в правом верхнем углу каждого фотоснимка). В клетки ввели плазмиду с геном белка mt-mEos2, который метит митохондрии, встраиваясь в их мембраны. Облучая небольшую зону аксона светом определенной волны, получали фотоконверсию, из-за которой меняется спектр флуоресценции репортерного белка. На данных фото митохондрии без фотоконверсии выглядят зелеными, а с фотоконверсией — пурпурными. Спустя время некоторые митохондрии сливаются и в результате смешения цветов выглядят белесоватыми. Верхняя серия фотографий демонстрирует нормальный процесс слияния-деления митохондрий в аксоне контрольных клеток. Несмотря на наличие обоих процессов, митохондрии остаются короткими. Нижняя серия показывает результаты аналогичного исследования для нейронов с подавленным Mff (при помощи коротких шпилечных РНК — Mff shRNA). Здесь преобладает слияние, и митохондрии со временем удлиняются. Рисунок из обсуждаемой статьи в Nature Communications
На данных фото митохондрии без фотоконверсии выглядят зелеными, а с фотоконверсией — пурпурными. Спустя время некоторые митохондрии сливаются и в результате смешения цветов выглядят белесоватыми. Верхняя серия фотографий демонстрирует нормальный процесс слияния-деления митохондрий в аксоне контрольных клеток. Несмотря на наличие обоих процессов, митохондрии остаются короткими. Нижняя серия показывает результаты аналогичного исследования для нейронов с подавленным Mff (при помощи коротких шпилечных РНК — Mff shRNA). Здесь преобладает слияние, и митохондрии со временем удлиняются. Рисунок из обсуждаемой статьи в Nature Communications
Сделав видеозаписи изменения митохондрий, помеченных флуоресцентными красителями, в клетках с интерференцией и без интерференции гена Mff (видео 1, видео 2, видео 3), исследователи убедились, что причиной наблюдаемых изменений стало значительное уменьшение частоты делений митохондрий и некоторое повышение частоты их слияний. При подавленном гене Mff уже на входе в аксон оказывались более длинные митохондрии, чем при нормально работающем Mff. Соотношение же частоты разделений и слияний митохондрий внутри самого аксона при подавлении Mff снизилось более чем в 3 раза. Фото, сделанные во время одного из подобных экспериментов, приведены на рис. 3. Чтобы застраховаться от возможных ошибок в интерпретации результатов, проводили также эксперименты, в которых наряду со шпилечными РНК против Mff вводилась «спасительная» копия самого Mff гена. Фенотип аксональных митохондрий в этом случае восстанавливался — и это подтверждает, что наблюдаемые эффекты не связаны с какими-то неспецифическими влияниями производимых над клетками манипуляций.
Соотношение же частоты разделений и слияний митохондрий внутри самого аксона при подавлении Mff снизилось более чем в 3 раза. Фото, сделанные во время одного из подобных экспериментов, приведены на рис. 3. Чтобы застраховаться от возможных ошибок в интерпретации результатов, проводили также эксперименты, в которых наряду со шпилечными РНК против Mff вводилась «спасительная» копия самого Mff гена. Фенотип аксональных митохондрий в этом случае восстанавливался — и это подтверждает, что наблюдаемые эффекты не связаны с какими-то неспецифическими влияниями производимых над клетками манипуляций.
Функциональное значение слияния и деления митохондрий обычно объясняют с двух точек зрения. Во-первых, энергетическая эффективность маленьких митохондрий оказывается несколько ниже, чем у больших — просто вследствие большей утечки протонов из-за более высокой суммарной площади поверхности по отношению к объему. Соответственно, митохондрии более склонны сливаться при голодании, а при избытке питательных веществ дробятся на более мелкие. Во-вторых, деление митохондрий позволяет сегрегировать «высококачественные» митохондрии, выполняющие свои функции как следует, от дефектных, которые функционируют хуже вследствие поломок в геноме или по иным причинам — такие митохондрии подвергаются уничтожению при помощи системы аутофагии. Но поскольку в данном случае морфология митохондрий оказывается специфичной для клеточных компартментов, ни одно из изложенных объяснений не кажется удовлетворительным.
Во-вторых, деление митохондрий позволяет сегрегировать «высококачественные» митохондрии, выполняющие свои функции как следует, от дефектных, которые функционируют хуже вследствие поломок в геноме или по иным причинам — такие митохондрии подвергаются уничтожению при помощи системы аутофагии. Но поскольку в данном случае морфология митохондрий оказывается специфичной для клеточных компартментов, ни одно из изложенных объяснений не кажется удовлетворительным.
Быть может размер митохондрий отрицательно сказывается на их мобильности внутри аксона или на возможности захвата митохондрий в пресинаптическую область? Ученые измерили соответствующие показатели в клетках с нормальным и подавленным Mff, сравнили результаты — это предположение не подтвердилось.
Далее измерили электрохимический потенциал на мембране митохондрий, окислительно-восстановительный потенциал матрикса, скорость синтеза АТФ длинными и короткими митохондриями в аксонах экспериментальных клеток (кому интересно как именно — рекомендую обратиться к оригинальной статье, но все использованные подходы так или иначе опираются на явление флуоресценции), сравнили, и снова не выявили значимых различий.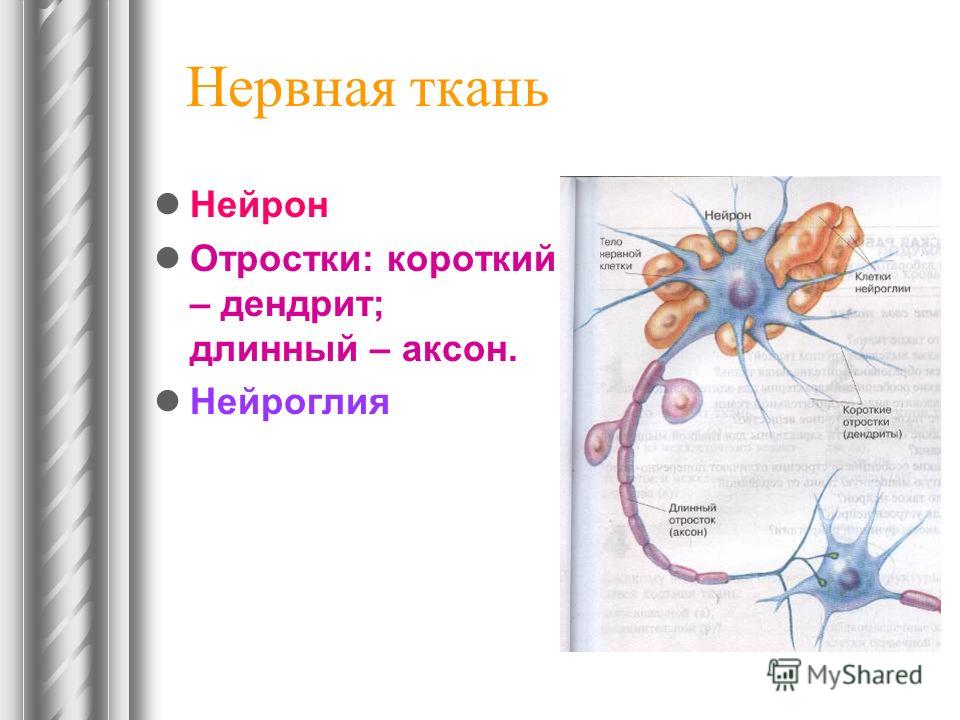
Наконец, решили проверить, не влияют ли митохондрии каким-то образом на динамику ионов кальция. Напомню, что именно поступление избыточного кальция в терминали аксона заставляет нейрон высвободить нейромедиатор, отправляя сигналы к принимающей клетке. Куда этот избыточный кальций уходит потом? Частично снова выводится наружу, часть захватывается трубочками эндоплазматической сети, и еще какая-то часть поглощается митохондриями (это было установлено авторами в работе, опубликованной ранее). Может быть именно здесь и кроется ключ к ответу? Эту гипотезу проверяли, замеряя уровни кальция в аксонах пирамидальных нейронов коры головного мозга мышей до, во время и после индукции потенциалов действия. Выяснили следующее:
1) Суммарное количество ионов кальция, удерживаемого внутри длинных митохондрий в состоянии покоя выше, чем в коротких митохондриях. Это и логично, учитывая больший объем матрикса в первых по сравнению со вторыми.
2) Немедленно после индукции потенциала действия кальций, поступивший внутрь терминали аксона, быстрее откачивается близлежащими крупными митохондриями, чем мелкими.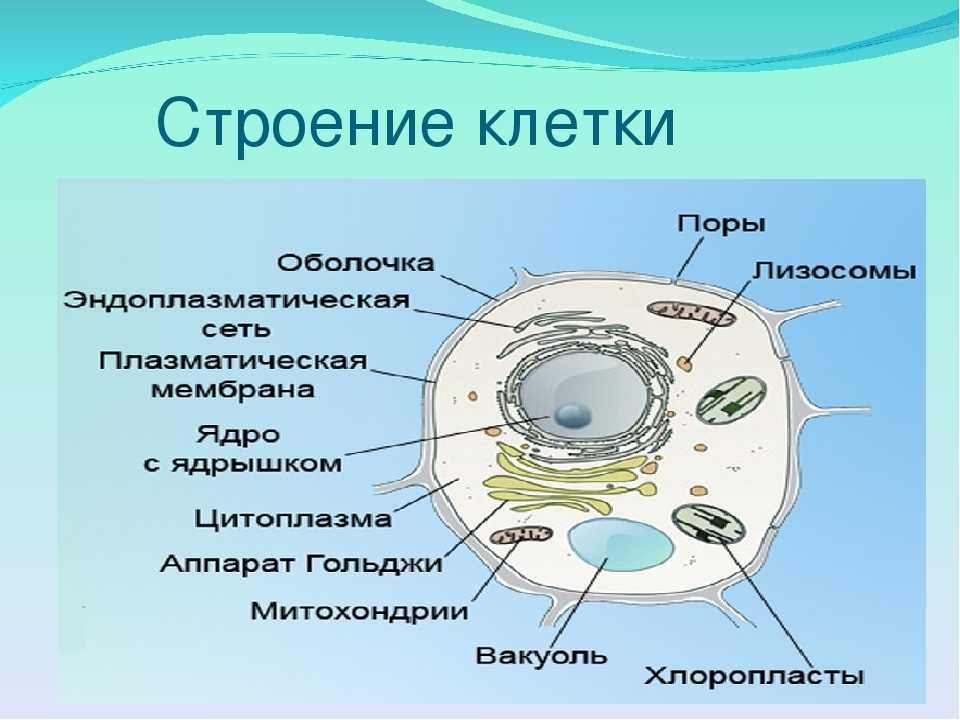 В итоге концентрация ионов кальция в цитоплазме внутри терминали оказывается в первом случае более низкой, чем в норме при мелких митохондриях.
В итоге концентрация ионов кальция в цитоплазме внутри терминали оказывается в первом случае более низкой, чем в норме при мелких митохондриях.
3) В присутствии длинных митохондрий в синаптическую щель выделяется меньше нейромедиатора, следовательно, к принимающей клетке приходит ослабленный сигнал по сравнению с тем, что происходит в случае контрольных нейронов с мелкими аксональными митохондриями. Это, собственно, вполне ожидаемое следствие более низкого уровня ионов кальция в цитоплазме.
Рис. 4. Активность Mff существенна для формирования ветвлений аксонов пирамидальных нейронов в ходе развития коры головного мозга. В левой частирисунка — полные срезы мозга мыши. Зеленым светятся зоны расположения нейронов и их отростков, в которые на эмбриональной стадии развития при помощи электропорации вводили указанные по левому краю конструкции. Самый верхний срез (a) — контрольный образец, ниже (d) — образец с подавленным геном Mff, в самом низу (g) — образец, где проводилось введение шпилечной РНК против гена Mff одновременно со «спасительной» копией того же гена. Во всех случаях конструкции вводились в левую половину мозга (здесь мы видим ярко-зеленое свечение), но отростки нейронов, расположенных в этой области, тянутся к противоположному полушарию, где их отростки формируют светло-зеленые «облачка». Зоны с этими «облачками» выделены прямоугольниками и показаны правее в большем увеличении. Можно заметить, что в мозге с подавленным Mff эти «облачка» заметно слабее, особенно верхнее, заходящее в слой, обозначенный как L2/3 — это указывает на уменьшение ветвления терминалей аксонов. Рисунок из обсуждаемой статьи в Nature Communications
Во всех случаях конструкции вводились в левую половину мозга (здесь мы видим ярко-зеленое свечение), но отростки нейронов, расположенных в этой области, тянутся к противоположному полушарию, где их отростки формируют светло-зеленые «облачка». Зоны с этими «облачками» выделены прямоугольниками и показаны правее в большем увеличении. Можно заметить, что в мозге с подавленным Mff эти «облачка» заметно слабее, особенно верхнее, заходящее в слой, обозначенный как L2/3 — это указывает на уменьшение ветвления терминалей аксонов. Рисунок из обсуждаемой статьи в Nature Communications
Кроме того исследователи проверили и влияние размера митохондрий на развитие коры головного мозга. Для этого в определенную область коры головного мозга мышиных эмбрионов (на 15-й день жизни) вводили шпилечные РНК (контрольные или интерферирующие ген Mff). Одновременно вводили ген флуоресцирующего белка для последующей визуализации. Образцы срезов мозга получали через три недели после рождения мышат. В контрольных образцах аксоны от пирамидальных нейронов соответствующей области направлялись в противоположное полушарие и там формировали две зоны обильного ветвления: в слое 2/3 и в слое 5 коры головного мозга.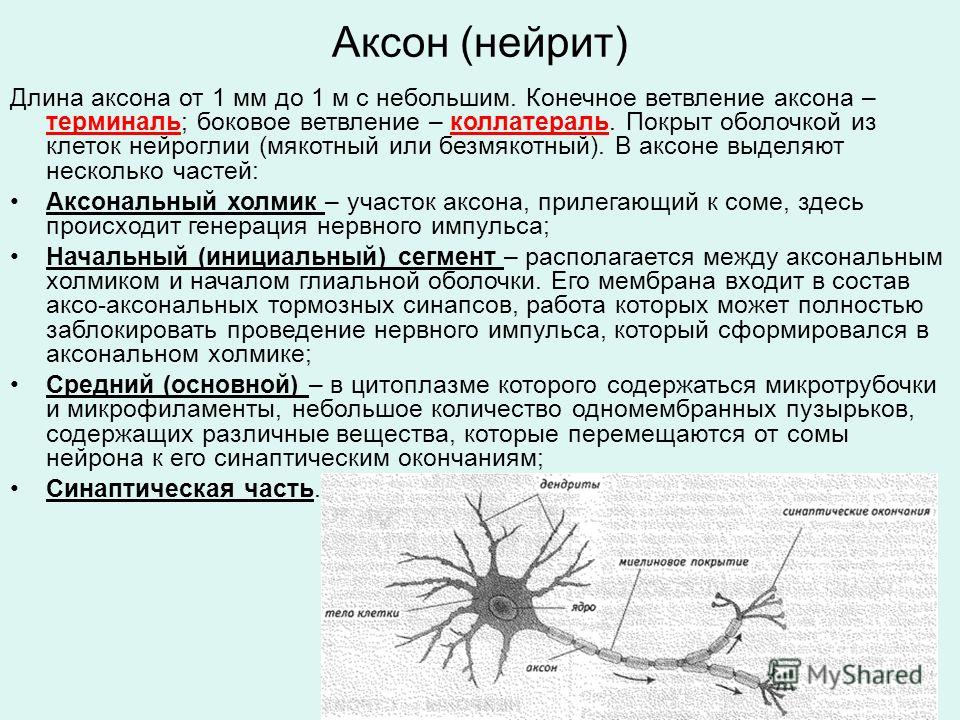 Но это ветвление было значительно слабее (особенно в слое 2/3) в случае, когда Mff был подавлен. Между тем «спасительная» копия гена Mff частично восстанавливала ветвление, так что картина получалась более близка к норме (рис. 4).
Но это ветвление было значительно слабее (особенно в слое 2/3) в случае, когда Mff был подавлен. Между тем «спасительная» копия гена Mff частично восстанавливала ветвление, так что картина получалась более близка к норме (рис. 4).
Общая картина, вырисовавшаяся по итогам этого исследования, отражена на рис. 5.
Рис. 5. Общая схема влияния размера митохондрий, контролируемого Mff, на функции и морфологию нейрона. В контрольных нейронах в аксон входят небольшие митохондрии и их размер поддерживается при перемещении вдоль аксона благодаря балансу между процессами слияния и деления митохондрий (a). Потеря активности Mff приводит к увеличению размера митохондрий, входящих в аксон, а также к сокращению частоты делений по сравнению с частотой слияний митохондрий (b). В итоге размеры митохондрий увеличиваются, а ветвление аксона уменьшается. В контрольных нейронах около 50% пресинаптических расширений («бутонов») содержит вблизи небольшую митохондрию (c). При активации нейрона, митохондрия буферизует некоторое количество поступившего Ca2+, вследствие чего здесь высвобождается меньше нейромедиаторов, чем в «бутонах» без митохондрии. При подавлении Mff, увеличенные митохондрии захватывают больше кальция из цитоплазмы, что значительно уменьшает выброс нейромедиатора в синаптическую щель (d). Вероятнее всего, именно это и приводит в конечном итоге к уменьшению ветвления аксона, поскольку активация синапсов служит поддерживающим стимулом для инициации ветвления по механизму обратной связи. Рисунок из обсуждаемой статьи в Nature Communications
При подавлении Mff, увеличенные митохондрии захватывают больше кальция из цитоплазмы, что значительно уменьшает выброс нейромедиатора в синаптическую щель (d). Вероятнее всего, именно это и приводит в конечном итоге к уменьшению ветвления аксона, поскольку активация синапсов служит поддерживающим стимулом для инициации ветвления по механизму обратной связи. Рисунок из обсуждаемой статьи в Nature Communications
Итак, обсуждаемая работа выявила еще одну важную функцию митохондрий в качестве регулятора уровня кальция в нейронах. Вместе с тем картина не ясна в полной мере. К примеру, остается вопрос, каким образом обеспечивается формирование морфологически разных митохондрий в дендритах и в аксонах одних и тех же нейронов (между прочим, напоминают авторы, движение митохондрий, располагающихся внутри аксонов и внутри дендритов обеспечивается двумя разными моторными белками: TRAK1 и TRAK2, соответственно — может ответ где-то тут)? И как их функция в качестве депо кальция задействована (если задействована) в дендритах? Что ж, будем ждать продолжения.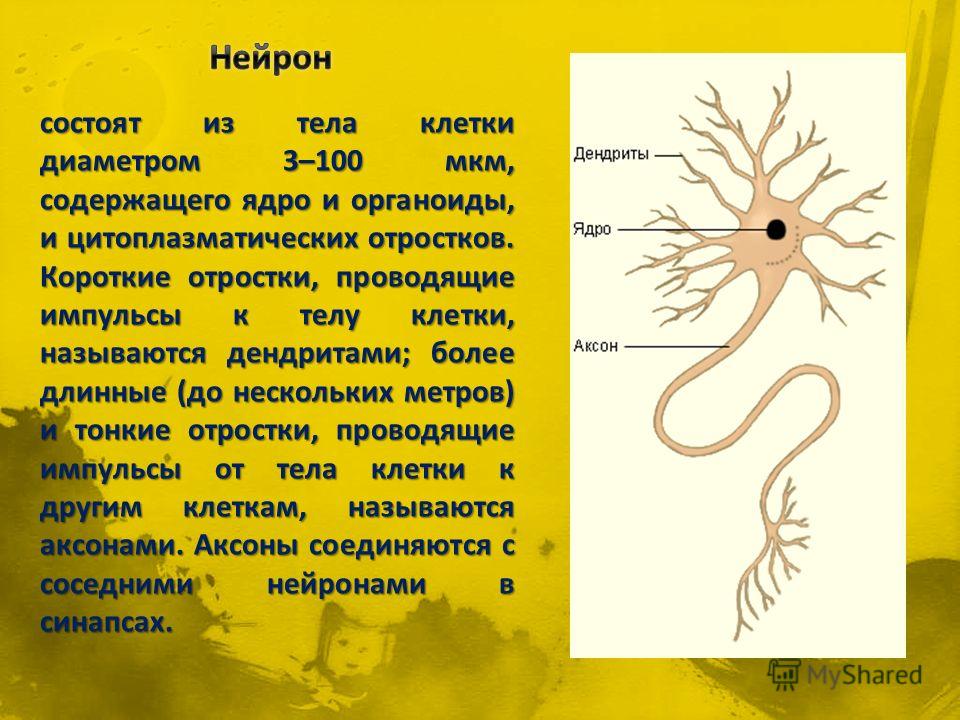 Авторы представленного исследования наверняка не остановятся на достигнутом.
Авторы представленного исследования наверняка не остановятся на достигнутом.
Татьяна Романовская
Источник: Элементы
Нервные клетки — Неврология — Книжная полка NCBI
Несмотря на специфические молекулярные, морфологические и функциональные особенности любого
определенного типа нервных клеток, основная структура нейронов напоминает структуру других
клетки. Таким образом, каждая нервная клетка имеет клеточное тело, содержащее ядро, эндоплазматический
ретикулум, рибосомы, аппарат Гольджи, митохондрии и другие органеллы,
необходимы для функционирования всех клеток (). Эти особенности лучше всего распознаются при большом увеличении и
разрешение, которое дает электронный микроскоп. Отличительной чертой
нервные клетки – это их специализация для межклеточной коммуникации. Этот атрибут
проявляется в их общей морфологии, в специализации их мембран
для электрической сигнализации, а также в структурных и функциональных тонкостях
синаптические контакты между ними.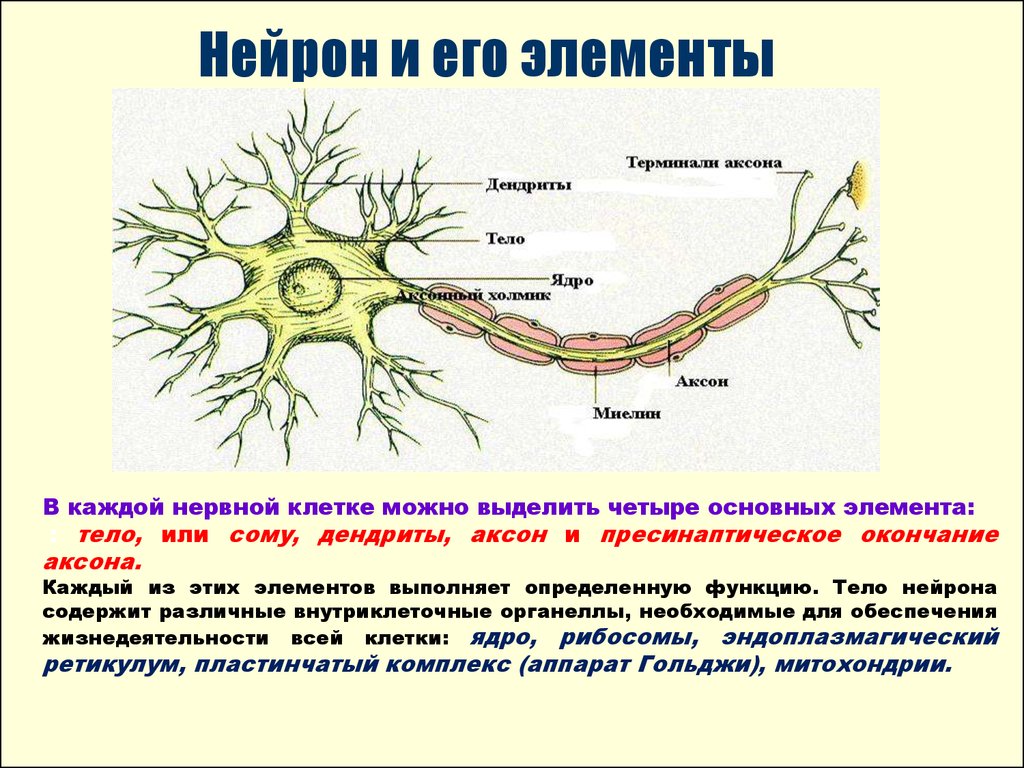
Рисунок 1.3
(A) Схема нервных клеток и их составных частей. (B) Исходный аксон сегмент (синий), входящий в миелиновую оболочку (золотой). (C) Терминальные бутоны (синий) загружен синаптическими пузырьками (наконечниками стрел), образующими синапсы (стрелки) с дендритом (фиолетовый). (D) Поперечный (подробнее…)
Особенно заметной морфологической особенностью большинства нервных клеток является сложная
разветвления дендритов (также называемых дендритными ветвями или дендритными
процессы), которые возникают из тела нейрона. Спектр нейронов
Геометрия варьируется от небольшого меньшинства клеток, полностью лишенных дендритов, до
нейроны с дендритными разветвлениями, которые соперничают по сложности со зрелым деревом (см.
). Количество входов, которые
конкретный нейрон получает зависит от сложности его дендритной ветви: Нерв
клетки, у которых отсутствуют дендриты, иннервируются только одной или несколькими другими нервными клетками,
тогда как те, у которых все более сложные дендриты, иннервируются
соизмеримо большее количество других нейронов.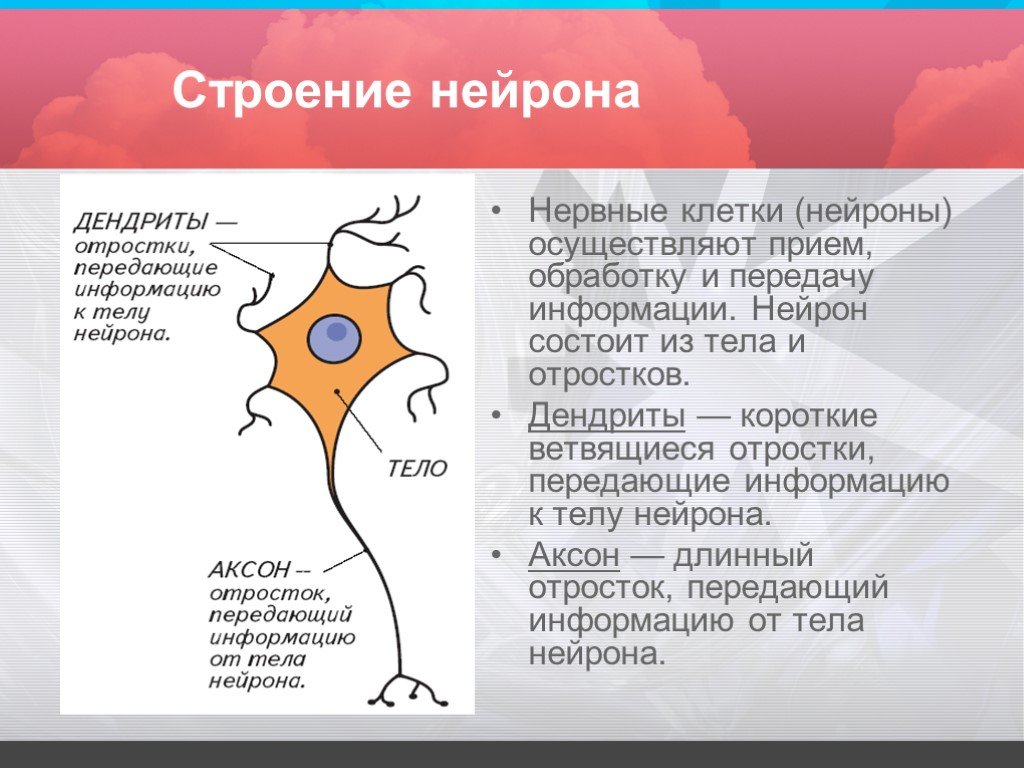
Дендриты (вместе с телом клетки) обеспечивают основное место для синаптических
терминалы образованы аксональными окончаниями других нервных клеток. синаптический
Сам контакт представляет собой особую разработку секреторного аппарата, обнаруживаемую у большинства
поляризованные эпителиальные клетки. Как правило, пресинаптический терминал непосредственно примыкает к постсинаптической специализации контактируемого
клетка. Для подавляющего большинства синапсов нет физической непрерывности между
эти пре- и постсинаптические элементы. Вместо этого пре- и постсинаптические компоненты
общаются посредством секреции молекул из пресинаптического окончания, которые связываются с
рецепторы постсинаптической специализации. Эти молекулы должны пройти
внеклеточное пространство между пре- и постсинаптическими элементами; это прерывание
называется синаптической щелью. Количество синаптических входов, полученных каждой нервной клеткой
в нервной системе человека колеблется от 1 до примерно 100 000. Этот диапазон входов
отражает основное предназначение нервных клеток, а именно интегрировать информацию от
другие нейроны. Таким образом, количество входов в любую конкретную ячейку равно
особенно важный детерминант функции нейронов.
Этот диапазон входов
отражает основное предназначение нервных клеток, а именно интегрировать информацию от
другие нейроны. Таким образом, количество входов в любую конкретную ячейку равно
особенно важный детерминант функции нейронов.
Информация от входов, которые воздействуют на дендриты нейронов, интегрируется
и «считывать» в начале аксона часть
нервная клетка, специализирующаяся на проведении сигнала к следующему участку синаптического
взаимодействие (см. и ). Аксон является уникальным продолжением
тело нейронной клетки, которое может перемещаться на несколько сотен микрометров или намного дальше,
в зависимости от типа нейрона и размера вида. Многие нервные клетки в
человеческий мозг имеет аксоны длиной не более нескольких миллиметров, а некоторые не имеют аксонов
вообще (см., например, амакриновую клетку сетчатки в ; на самом деле, амакрин средство
«отсутствие длительного процесса»). Эти короткие аксоны являются определяющими
особенность нейронов локальной цепи или интернейронов по всему мозгу.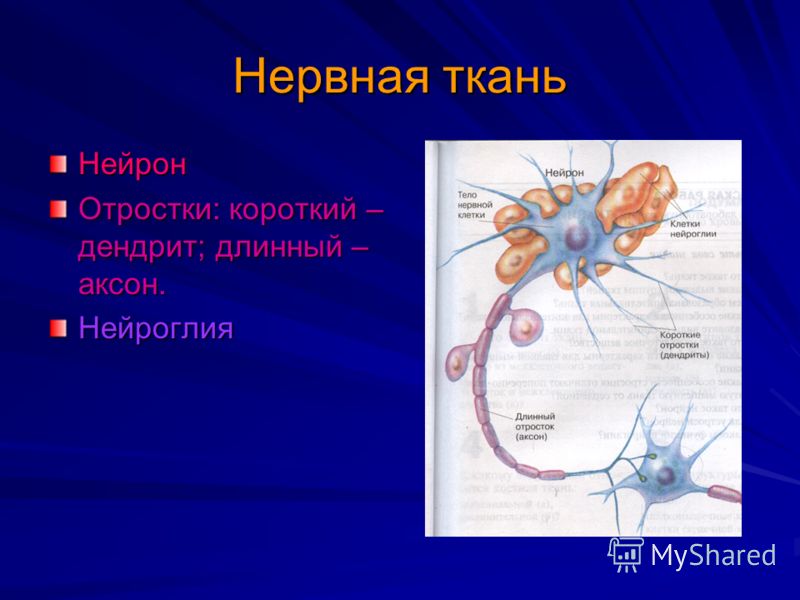 Многие аксоны,
однако распространяется на более удаленные цели. Например, аксоны, отходящие от
спинного мозга человека до стопы имеют длину около метра. Аксональный механизм, который
передает сигналы на такие расстояния, называется потенциалом действия.
самовоспроизводящаяся волна электрической активности, которая распространяется от своей точки
от тела клетки (называемого аксонным холмиком) до окончания аксона.
На конце аксона другой набор синаптических контактов создается на других клетках.
клетки-мишени нейронов включают другие нервные клетки в головном, спинном мозге и
вегетативные ганглии, а также клетки мышц и желез по всему телу.
Многие аксоны,
однако распространяется на более удаленные цели. Например, аксоны, отходящие от
спинного мозга человека до стопы имеют длину около метра. Аксональный механизм, который
передает сигналы на такие расстояния, называется потенциалом действия.
самовоспроизводящаяся волна электрической активности, которая распространяется от своей точки
от тела клетки (называемого аксонным холмиком) до окончания аксона.
На конце аксона другой набор синаптических контактов создается на других клетках.
клетки-мишени нейронов включают другие нервные клетки в головном, спинном мозге и
вегетативные ганглии, а также клетки мышц и желез по всему телу.
Процесс, посредством которого информация, закодированная потенциалами действия, передается в
синаптические контакты со следующей клеткой на пути называются синаптических
передача . Пресинаптические окончания (также называемые синаптическими окончаниями, аксоны
терминали, или терминальные бутоны) и их постсинаптическая специализация обычно
химические синапсы, наиболее распространенный тип синапсов в нервной системе (еще один
тип, называемый электрическим синапсом, описан в главе 5).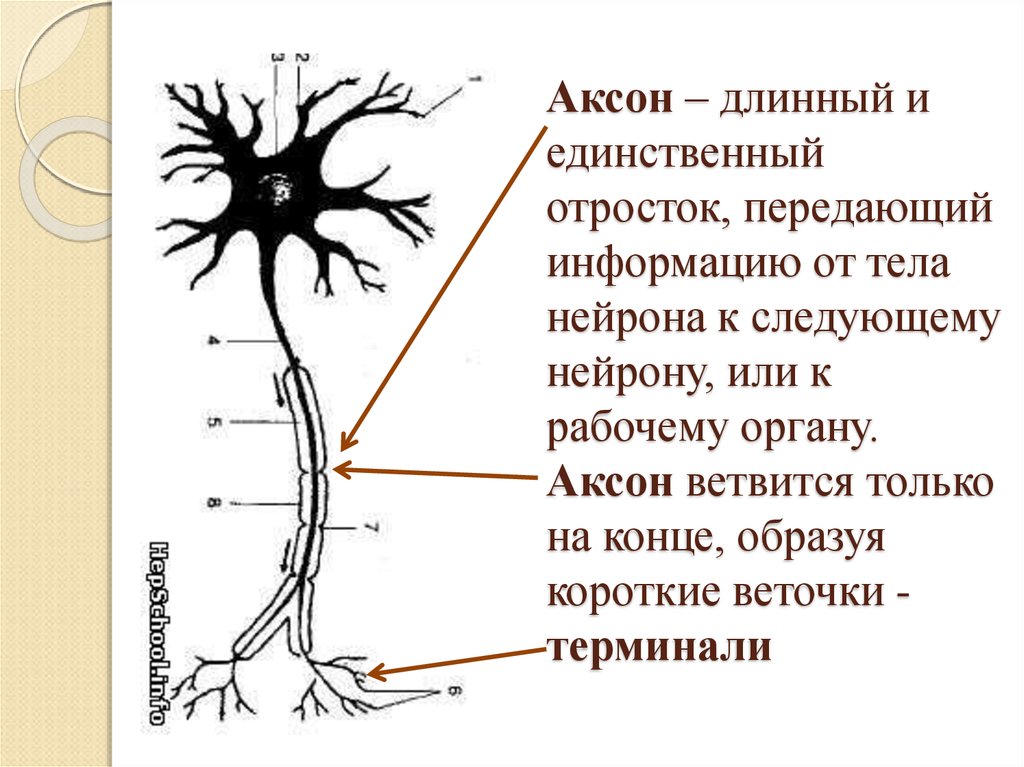 Секреторные органеллы в пресинаптических
окончания химических синапсов называются синаптические пузырьки, которые заполнены
молекулы нейротрансмиттеров. Нейротрансмиттеры высвобождаются из синаптических пузырьков.
изменять электрические свойства клетки-мишени путем связывания с нейротрансмиттерных рецепторов , которые локализованы преимущественно в
постсинаптическая специализация. Нейротрансмиттеры, рецепторы и все, что с ними связано.
молекулы трансдукции — это механизм, который позволяет нервным клеткам взаимодействовать с
друг с другом, а также с эффекторными клетками в мышцах и железах.
Секреторные органеллы в пресинаптических
окончания химических синапсов называются синаптические пузырьки, которые заполнены
молекулы нейротрансмиттеров. Нейротрансмиттеры высвобождаются из синаптических пузырьков.
изменять электрические свойства клетки-мишени путем связывания с нейротрансмиттерных рецепторов , которые локализованы преимущественно в
постсинаптическая специализация. Нейротрансмиттеры, рецепторы и все, что с ними связано.
молекулы трансдукции — это механизм, который позволяет нервным клеткам взаимодействовать с
друг с другом, а также с эффекторными клетками в мышцах и железах.
Неврология для детей — клетки нервной системы
Типы нейронов (нервных клеток)
Тело человека состоит из триллионов клеток. Клетки нервной
система, называемая нервными клетками или нейронов , специализируются на
нести «сообщения» посредством электрохимического процесса. Человеческий мозг имеет
примерно 86 миллиардов нейронов. Чтобы узнать, как нейроны передают сообщения,
читайте про потенциал действия.
Нейроны бывают разных форм и размеров. Одни из самых маленьких нейроны имеют клеточные тела шириной всего 4 микрона. Некоторые из самые большие нейроны имеют клеточные тела шириной 100 микрон. (Помнить что 1 микрон равен одной тысячной миллиметра!).
Нейроны похожи на другие клетки организма, потому что:
- Нейроны окружены клеточной мембраной.
- Нейроны имеют ядро, содержащее гены.
- Нейроны содержат цитоплазму, митохондрии и другие органеллы.
- Нейроны осуществляют основные клеточные процессы, такие как белок синтез и производство энергии.
Однако нейроны отличаются от других клеток организма, потому что:
- Нейроны имеют специализированные клеточные части, называемые дендриты и аксонов . Дендриты передают электрические сигналы тело клетки и аксоны забирают информацию от тела клетки.
- Нейроны общаются друг с другом посредством электрохимического
процесс.

- Нейроны содержат некоторые специализированные структуры (например, синапсы) и химические вещества (например, нейротрансмиттеры).
Нейрон
Один из способов классификации нейронов — по количеству расширений, которые отходят от тела нейрона (сомы).
Биполярные нейроны имеют два отростка
от тела клетки (примеры: клетки сетчатки,
клетки обонятельного эпителия).
Псевдоуниполярные клетки (пример: дорсальные
клетки корневых ганглиев). На самом деле, эти клетки имеют 2 аксона, а не один.
аксон и дендрит. Один аксон идет центрально к спинному мозгу,
другой аксон простирается к коже или мышце.
Мультиполярные нейроны имеют множество отростков, отходящих от
тело клетки. Однако каждый нейрон имеет только один аксон (примеры: спинномозговой
двигательные нейроны, пирамидные нейроны, клетки Пуркинье).
Нейроны также можно классифицировать по направлению, в котором они посылают
информация.
- Сенсорные (или афферентные) нейроны: отправить информация от сенсорных рецепторов (например, кожи, глаз, носа, языка, уши) К центральной нервной системе.
- Двигательные (или эфферентные) нейроны: отправка информации ОТСУТСТВУЕТ из центральной нервной системы в мышцы или железы.
- Интернейроны: посылают информацию между сенсорными нейроны и двигательные нейроны. Большинство вставочных нейронов расположены в центральной нервная система.
Посетите галерею нейронов, чтобы увидеть некоторые фотографии реальных нейронов или «Тротуар Клетки», чтобы увидеть фотографии нейронов на улице. «Аксон» | «Дендрит» |
Между аксонами и дендриты:
| Аксоны | Дендриты |
|
|
Что находится внутри нейрона? Нейрон имеет много одинаковых органеллы , такие как митохондрии, цитоплазма и
ядро, как и другие клетки организма.
- Ядро — содержит генетический материал (хромосомы), включая информацию для развития клеток и синтеза белков, необходимых для поддержания и выживания клеток. Покрытый мембрана.
- Ядрышко — производит необходимые рибосомы для трансляции генетической информации в белки
- Тельца Ниссля — используемые группы рибосом для синтеза белка.
- Эндоплазматический ретикулум (ER) — система трубы для транспорта веществ внутри цитоплазмы. Может иметь рибосомы (грубый ЭР) или без рибосом (гладкий ЭР). С рибосомами ER важен для синтез белка.
- Аппарат Гольджи — мембранный структура, важная для упаковки пептидов и белков (включая нейротрансмиттеры) в везикулы.
- микрофиламенты/нейротубулы — система транспорт материалов внутри нейрона и может использоваться для структурных поддерживать.
- Митохондрии —
производят энергию для подпитки клеточной активности.
Leave A Comment