
Взаимоотношения гриба и водоросли в теле лишайника
Вопрос взаимоотношения гриба и водоросли в слоевище лишайника занимал умы ученых еще в конце прошлого столетия, да и в наше время продолжает волновать лихенологов. Со дня открытия С. Швенденера прошло более 100 лет. За этот период появилось не менее десятка теорий, пытающихся объяснить отношения между грибом и водорослью, однако среди них нет ни одной общепризнанной и окончательно доказапной. С. Швенденер, обнаружив, что лишайник состоит из гриба и водоросли, предположил, что гриб в слоевище паразитирует па водоросли. Однако он ошибочно отвел грибу роль хозяина, а водоросли — раба.[ …]
Наибольшее распространение среди ученых того времени получила теория мутуалистиче-ского симбиоза. Сторонники этой теории считали, что в слоевище лишайника гриб и водоросль находятся во взаимовыгодном симбиозе: водоросль «снабжает» гриб органическими веществами, а гриб «защищает» водоросль от чрезмерного нагревания и освещения и «обеспечивает» ее водой и неорганическими солями. Однако в 1873 г. этой идеалистической теории был нанесен удар. Известный французский исследователь Е. Б о р н е, изучая анатомическое строение слоевища лишайников, обнаружил внутри водорослевых клеток грибные отростки — гаустории, всасывающие органы гриба. Это позволяло думать, что гриб использует содержимое клеток водорослей, т. е. ведет себя как паразит.[ …]
За прошедшие со времен Борне 100 лет в слоевище лишайников было открыто и описано много различных фо£м абсорбционных, или всасывающих, гиф гриба. Эти гифы плотно прижимаются к клетке водоросли или проникают в нее и служат, как предполагают, для передачи веществ, которые образуют водоросли в результате своей жизнедеятельности, грибному компоненту.[ …]
Гриб должен использовать лишь часть водорослей, оставляя резерв — здоровые и нормальные водоросли, содержимым которых он мог бы питаться.[ …]
Учеными были замечены любопытные защитные реакции со стороны лишайниковых водорослей.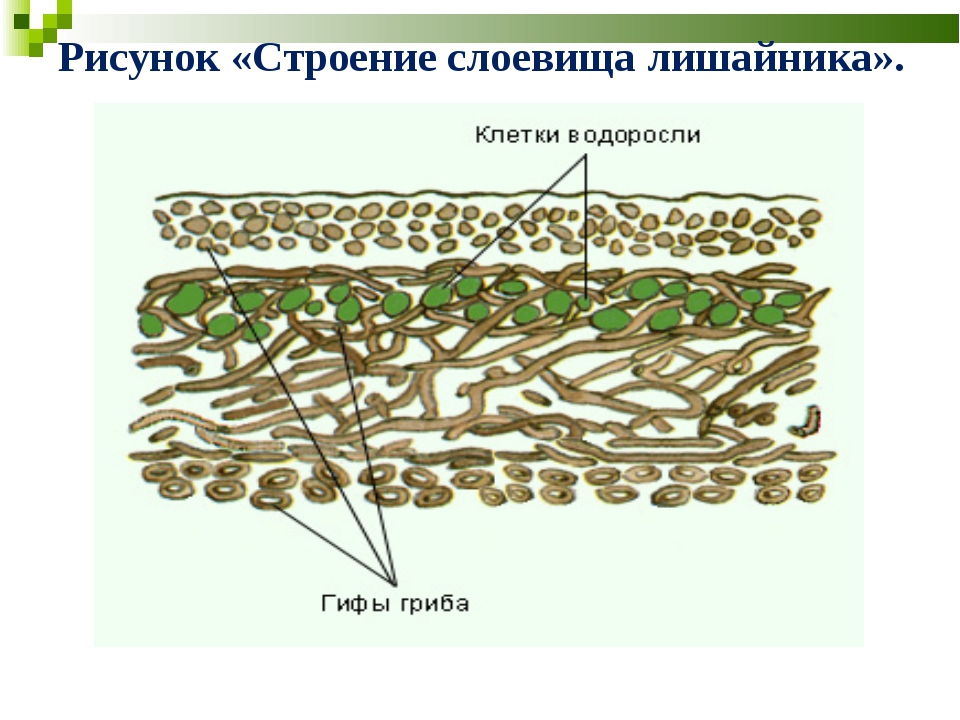 Например, одновременно с проникновением гаустория в клетку водоросли эта клетка делилась. При этом плоскость деления, как правило, проходила как раз через участок, занятый гаусторием, а образовавшиеся в результате этого процесса дочерние клетки были свободны от гаусториев. Было замечено также, что обычно гриб поражает водоросли, уже достигшие определенной стадии зрелости. В молодых растущих водорослях происходит энергичное отложение веществ в оболочке клетки и быстрое ее утолщение. Эта толстая оболочка клетки фикобионта препятствует проникновению абсорбционных органов гриба. Однако большей частью защитная реакция водорослей против активности грибного компонента очень слаба.[ …]
Например, одновременно с проникновением гаустория в клетку водоросли эта клетка делилась. При этом плоскость деления, как правило, проходила как раз через участок, занятый гаусторием, а образовавшиеся в результате этого процесса дочерние клетки были свободны от гаусториев. Было замечено также, что обычно гриб поражает водоросли, уже достигшие определенной стадии зрелости. В молодых растущих водорослях происходит энергичное отложение веществ в оболочке клетки и быстрое ее утолщение. Эта толстая оболочка клетки фикобионта препятствует проникновению абсорбционных органов гриба. Однако большей частью защитная реакция водорослей против активности грибного компонента очень слаба.[ …]
Способность водорослей нормально развиваться и даже размножаться в слоевище лишайника сохраняется скорее благодаря умеренности паразитизма самого гриба.[ …]
Ученые отмечают, что степень паразитизма гриба на водоросли различна не только у разных видов лишайников, но даже в одном и том же слоевище. Резкий паразитизм обнаружен лишь у примитивных лишайников. Гаустории, проникающие глубоко внутрь протопласта водоросли, пока что были найдены лишь у наиболее просто организованных форм, в слоевище которых еще нельзя различить оформленных дифференцированных слоев. В слоевищах более высокоорганизованных лишайников часть клеток водорослей поражена грибными гифами, а остальные продолжают нормально жить и развиваться. Обычно у высокоорганизованных форм лишайников паразитизм гриба на водоросли носит весьма умеренный характер: прежде чем гриб убьет пораженные им клетки, успевает вырасти одно или несколько поколений водорослей.[ …]
Интересную мысль о взаимоотношении компонентов в слоевище лишайника высказал в 60-х годах нашего столетия крупнейший советский лихенолог А. Н. О к с н е р. Он пришел к выводу, что водоросль в слоевище лишайника, полностью изолированная от внешней среды грибной тканью, обязательно должна забирать у грибного компонента все необходимые для своего существования вещества, за исключением тех органических соединений, которые она сама вырабатывает на свету в процессе ассимиляции углекислоты.
Таким образом, ученые считают, что водорослевый и грибной компоненты лишайника находятся в очень сложных взаимоотношениях. Микобионт ведет себя как паразит и сапрофит на теле водоросли, а фикобионт, в свою очередь, паразитирует на лишайниковом грибе. При этом паразитизм фикобионта всегда носит более умеренный характер, чем паразитизм гриба.[ …]
Одной из причин неудач подобных попыток можно считать чрезвычайно медленный рост лишайников. Лишайники — многолетние растения. Обычно возраст взрослых слоевищ, которые можно увидеть где-нибудь в лесу на стволе деревьев или на почве, составляет не менее 20—50 лет. В северных тундрах возраст некоторых кустистых лишайников рода кладония достигает 300 лет. Слоевище лишайников, имеющих вид корочки, в год дает прирост всего 0,2—0,3 мм.[ …]
Рисунки к данной главе:
Вернуться к оглавлениюЛишайники — урок. Биология, Бактерии. Грибы. Растения (5–6 класс).
Лишайники — это группа симбиотических организмов, насчитывающая свыше \(20\) тыс. видов.
Тело лишайника — слоевище — состоит из гриба и водоросли, живущих в симбиозе как один организм. Оно образовано переплетающимися нитями грибницы, между которыми расположены одноклеточные зелёные водоросли. Нити гриба лишайника поглощают воду и растворённые в ней минеральные вещества, а в клетках зелёных водорослей образуются органические вещества
.
Водоросль, входящая в организм лишайника, отделённая от гриба, как правило, может существовать самостоятельно. Гриб отдельно от водоросли жить не может.
Гриб отдельно от водоросли жить не может.
Размножаются лишайники главным образом кусочками слоевища, а также особыми группами клеток гриба и водоросли, во множестве образующимися внутри его тела. Под их давлением слоевище лишайника разрывается, группы клеток подхватывает ветер или смывают струи дождя.
Многообразие лишайников
Окраска лишайников разная: белая, жёлтая, серая, коричневая и т. д. По внешнему виду слоевище может быть кустистым, листоватым и накипным.
Кустистые лишайники выглядят как небольшие кустики или деревца. Они поднимаются вверх, если растут на почве, или свисают сверху вниз, если прикреплены к стволам деревьев. Известный под названием «олений мох» кустистый лишайник ягель широко распространён в тундре.
На камнях поселяются накипные лишайники, похожие на застывшую коричнево-серую пену.
Широко распространены листоватые лишайники в виде пластинок разной окраски. Они разрастаются на коре деревьев или на камнях. На коре осин часто можно увидеть лишайник золотисто-жёлтого цвета. Это ксантория настенная.
Значение лишайников
Лишайники могут расти в самых бесплодных местах — в пустынях, на камнях, на крышах, на скалах, даже на бетоне, стекле и железе. Всей поверхностью тела они впитывают влагу дождей, росы и туманов.
Лишайники первыми заселяют безжизненные участки и постепенно превращают их в пригодные для заселения другими организмами. Их таллом выделяет особые лишайниковые кислоты, которые медленно разрушают горные породы.
Лишайники — долгожители. Они могут жить до \(50\)–\(100\) лет. Растут лишайники очень медленно: от нескольких миллиметров до нескольких сантиметров в год.
При недостатке влаги лишайники высыхают и кажутся безжизненными. Но при увлажнении они быстро оживают.
Лишайники служат кормом для животных. Их с удовольствием поедают олени, маралы, косули, лоси, а также некоторые улитки.
Лишайники издавна используются человеком. Из них получают лекарства, лакмус, красители. В некоторых странах лишайники используют в пищу.
Строение и размножение лишайников реферат 2010 по биологии
Лишайники представляют собой своеобразную группу комплексных организмов, тело которых состоит из двух компонентов — гриба и водоросли. Как организмы, лишайники были известны задолго до открытия их сущности, Еще великий Теофраст, «отец ботаники» (IV-III вв. до н. э.), дал описание двух лишайников — успей и рочеллы — которые уже тогда использовали для получения ароматических и красящих веществ. Правда, в те времена их нередко называли то мхами, то водорослями, то даже «хаосом природы» и «убогой нищетой растительности», Сейчас известно около 20 000 видов лишайников. Наука о лишайниках называется лихенологией. Специфический признак лишайников — симбиоз двух разных организмов: гетеротрофного гриба (микобионт) и автотрофной водоросли (фикобионт), В лишайнике оба эти компонента вступают в тесные взаимоотношения: гриб окружает водоросли и даже может проникать в их клетки. Лишайники образуют особые морфологические типы — жизненные формы, которые не встречаются у отдельных слагающих их организме». Метаболизм лишайников имеет специфический характер: только в них образуются лишайниковые кислоты, не встречающиеся у других организмов. Специфичны также и способы размножения лишайников как целостных организмов. Отношения гриба и водоросли в лишайнике очень сложны. С одной стороны, они взаимополезны: водоросль снабжает гриб синтезируемыми ею органическими веществами, а гриб обеспечивает доставку воды и минеральных солей, а также защищает от воздействия неблагоприятных факторов среды, таких, как перегрев, высыхание, излишняя инсоляция. С другой же, гриб ведет себя в слоевище лишайника как паразитический организм. Для сохранения как самого себя, так и лишайника в целом необходимо, чтобы водоросль, окруженная со всех сторон грибными гифами, все-таки могла жить. Если гриб начнет вести себя слишком активно, использовать не только продуцируемые водорослью вещества, но и поражать фикобионт, это может привести к гибели всего водорослевого компонента, а следом погибнет и сам гриб, и лишайник перестанет существовать».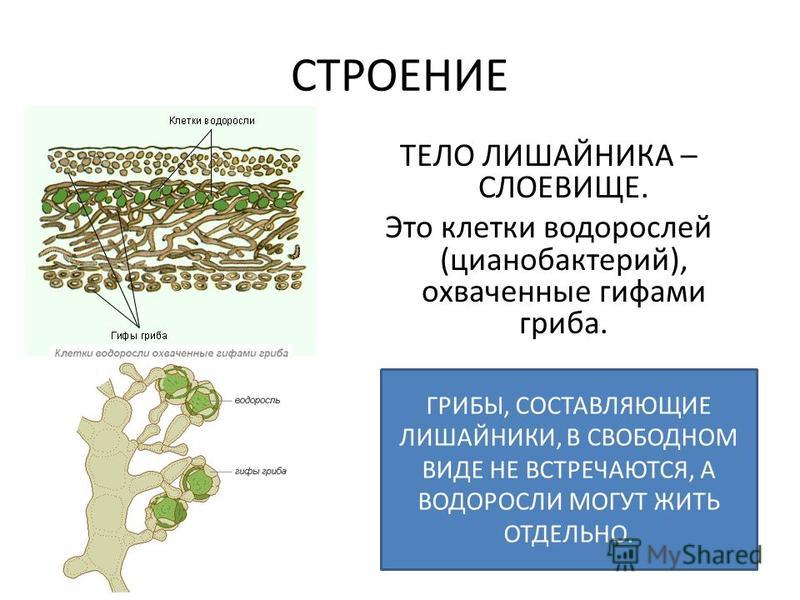 Исходя из этого можно назвать взаимоотношения гриба и водоросли в лишайнике умеренным паразитизмом. Проникая в клетку водоросли, гифы гриба образуют гаустории, то есть расширения на концах гиф для более активного всасывания питательных веществ, что характерно для паразитических грибов. Слоевище (так называется тело лишайника) разнообразно по форме, размерам, окраске и строению. Окраска лишайников варьирует: они бывают белые, серые, желтые, оранжевые, зеленые, черные; это определяется характером пигментов, содержащихся в оболочке гиф. Пигментация способствует защите водорослевого компонента от чрезмерного освещения. Иногда бывает и наоборот: лишайники Антарктиды окрашены в черный цвет, который поглощает тепловые лучи. По форме слоевища лишайники делятся на накипные, листоватые и кустистые. Слоевище накипных лишайников имеет вид корочки, плотно сросшейся с субстратом сердцевинными гифами. Иногда оно представлено порошковидным налетом. Листоватые лишайники имеют вид пластинки, горизонтально расположенной на субстрате, прикрепляясь к нему выростами гиф — ризинами. Слоевище может быть цельным или рассеченным прижатым к субстрату или приподнимающимся над ним. Слоевище остистых лишайником имеет вид разветвленного стоячего или повислого кустика либо неразветвленных стоячих столбиков. К субстрату они прикрепляются короткой ножкой, расширенной на конце пяточкой. По анатомическому строению лишайники бывают: 1) го-меомерными, когда водоросли разбросаны по всему телу лишайника; 2)гетеромерными, когда водоросли образуют в слоевище обособленный слой. Сверху слоевище покрыто коревым слоем, состоящим из срастающихся своими стенками http://www.ebio.ru/images/03020401.jpg Строение лишайника Постоянный компонент лишайников – водоpосли (сине-зелёные, жёлто- зелёные или зелёные). Обычно каждому виду лишайника соответствует свой вид водоpосли. По строению тела (слоевища) различают накипные (коpковые), листоватые и кустистые лишайники. Вегетативное тело накипных лишайников наиболее пpимитивно, оно бывает зеpнистым, пористым или в виде коpочек.
Исходя из этого можно назвать взаимоотношения гриба и водоросли в лишайнике умеренным паразитизмом. Проникая в клетку водоросли, гифы гриба образуют гаустории, то есть расширения на концах гиф для более активного всасывания питательных веществ, что характерно для паразитических грибов. Слоевище (так называется тело лишайника) разнообразно по форме, размерам, окраске и строению. Окраска лишайников варьирует: они бывают белые, серые, желтые, оранжевые, зеленые, черные; это определяется характером пигментов, содержащихся в оболочке гиф. Пигментация способствует защите водорослевого компонента от чрезмерного освещения. Иногда бывает и наоборот: лишайники Антарктиды окрашены в черный цвет, который поглощает тепловые лучи. По форме слоевища лишайники делятся на накипные, листоватые и кустистые. Слоевище накипных лишайников имеет вид корочки, плотно сросшейся с субстратом сердцевинными гифами. Иногда оно представлено порошковидным налетом. Листоватые лишайники имеют вид пластинки, горизонтально расположенной на субстрате, прикрепляясь к нему выростами гиф — ризинами. Слоевище может быть цельным или рассеченным прижатым к субстрату или приподнимающимся над ним. Слоевище остистых лишайником имеет вид разветвленного стоячего или повислого кустика либо неразветвленных стоячих столбиков. К субстрату они прикрепляются короткой ножкой, расширенной на конце пяточкой. По анатомическому строению лишайники бывают: 1) го-меомерными, когда водоросли разбросаны по всему телу лишайника; 2)гетеромерными, когда водоросли образуют в слоевище обособленный слой. Сверху слоевище покрыто коревым слоем, состоящим из срастающихся своими стенками http://www.ebio.ru/images/03020401.jpg Строение лишайника Постоянный компонент лишайников – водоpосли (сине-зелёные, жёлто- зелёные или зелёные). Обычно каждому виду лишайника соответствует свой вид водоpосли. По строению тела (слоевища) различают накипные (коpковые), листоватые и кустистые лишайники. Вегетативное тело накипных лишайников наиболее пpимитивно, оно бывает зеpнистым, пористым или в виде коpочек. Более pазвиты листоватые лишайники, имеющие вид более или менее pассечённых пластинок. Высоко оpганизованы кустистые лишайники, имеющие вид кустиков, свисающих нитей или пpямостоящих выpостов. http://www.ebio.ru/images/03020402.jpg Корковые (слева), листоватые (в центре) и кустистые (справа) лишайники Большинство лишайников состоят из образованной гифами плотной коры, в которой имеются необходимые для дыхания поры. Кора позволяет всасывать влагу из воздуха и защищает лишайник от переохлаждения или перегрева. Под корой гифы более рыхлые, между ними располагаются клетки водоросли. Обычно клетки фотобионта сконцентрированы по периферии – ближе к свету – образуя фотосинтезирующий слой. По анатомическому стpоению pазличают лишайники гомеомеpные (в котоpых водоpосли pаспpеделены более или менее pавномеpно по всему телу) и гетеpомеpные (водоpосли находятся только под слоем коры). Некоторые лишайники симбиотируют также с бактериями, за счёт чего получают азот прямо из воздуха, или паразитируют на мхах и других лишайниках. Pазмножение лишайников осуществляется половым и бесполым (вегетативным) способами. В pезультате полового пpоцесса обpазуются споpы гpиба лишайника, котоpые pазвиваются в закpытых плодовых телах – пеpитециях, имеющих узкое выводное отвеpстие ввеpху, или в апотециях, шиpоко откpытых к низу. Пpоpосшие споpы, встpетив соответствующую своему виду водоpосль, обpазуют с ней новое слоевище. Вегетативное pазмножение заключается в pегенеpации слоевища из небольших его участков (обломков, веточек). У многих лишайников есть специальные выросты – изидии, которые легко отламываются и дают начало новому слоевищу. В других лишайниках образуются крошечные гранулы (соредии), в которых клетки водоросли окружены плотным скоплением гиф; эти гранулы легко разносятся ветром. Лишайники растут на почве (эпигейные), камнях (эпилитные) или древесных стволах (эпифитные), получая необходимую для жизни влагу из атмосферы. Некоторые виды обитают на морской литорали. Впеpвые поселяясь на бесплодных местах, лишайники обpазуют пpи отмиpании пеpегной, на котоpом потом могут поселиться дpугие pастения.
Более pазвиты листоватые лишайники, имеющие вид более или менее pассечённых пластинок. Высоко оpганизованы кустистые лишайники, имеющие вид кустиков, свисающих нитей или пpямостоящих выpостов. http://www.ebio.ru/images/03020402.jpg Корковые (слева), листоватые (в центре) и кустистые (справа) лишайники Большинство лишайников состоят из образованной гифами плотной коры, в которой имеются необходимые для дыхания поры. Кора позволяет всасывать влагу из воздуха и защищает лишайник от переохлаждения или перегрева. Под корой гифы более рыхлые, между ними располагаются клетки водоросли. Обычно клетки фотобионта сконцентрированы по периферии – ближе к свету – образуя фотосинтезирующий слой. По анатомическому стpоению pазличают лишайники гомеомеpные (в котоpых водоpосли pаспpеделены более или менее pавномеpно по всему телу) и гетеpомеpные (водоpосли находятся только под слоем коры). Некоторые лишайники симбиотируют также с бактериями, за счёт чего получают азот прямо из воздуха, или паразитируют на мхах и других лишайниках. Pазмножение лишайников осуществляется половым и бесполым (вегетативным) способами. В pезультате полового пpоцесса обpазуются споpы гpиба лишайника, котоpые pазвиваются в закpытых плодовых телах – пеpитециях, имеющих узкое выводное отвеpстие ввеpху, или в апотециях, шиpоко откpытых к низу. Пpоpосшие споpы, встpетив соответствующую своему виду водоpосль, обpазуют с ней новое слоевище. Вегетативное pазмножение заключается в pегенеpации слоевища из небольших его участков (обломков, веточек). У многих лишайников есть специальные выросты – изидии, которые легко отламываются и дают начало новому слоевищу. В других лишайниках образуются крошечные гранулы (соредии), в которых клетки водоросли окружены плотным скоплением гиф; эти гранулы легко разносятся ветром. Лишайники растут на почве (эпигейные), камнях (эпилитные) или древесных стволах (эпифитные), получая необходимую для жизни влагу из атмосферы. Некоторые виды обитают на морской литорали. Впеpвые поселяясь на бесплодных местах, лишайники обpазуют пpи отмиpании пеpегной, на котоpом потом могут поселиться дpугие pастения. Лишайники обнаружены даже в бесплодных арктических пустынях и внутри антарктических горных пород. Лишайники распространены по всему миру, но особенно разнообразны в тропиках, высокогорьях и в тундре. А вот в лабораториях лишайники достаточно быстро погибают. И только в 1980 году Более примитивным считается слоевище гомеомерного строения. Если рассмотреть поперечный срез такого слоевища под микроскопом, то хорошо видно, что оно образовано беспорядочными переплетениями гиф гриба, среди которых разбросаны отдельные клетки или нити водорослей. Такое строение наиболее характерно для тех лишайников, фикобионтом которых являются сине-зеленые водоросли — носток, глеокапса и некоторые другие. Эти лишайники образуют группу, известную под названием слизистых лишайников. У лишайников, которые имеют фикобионтом зеленые водоросли, слоевище гомеомерного строения встречается очень редко, лишь у наиболее примитивных накипных форм. Слоевища слизистых лишайников в сухом состоянии имеют вид темных или даже черных твердых и хрупких корок или пленок. Но они обладают одной интересной особенностью — способностью впитывать огромное количество воды, в 20 — 30 раз превышающее их собственную массу. При увлажнении они сильно разбухают, ослизняются, приобретают характер студня или желе и становятся оливково-зелеными или грязно-зелеными. Во внешнем облике этих лишайников имеется много общего с некоторыми свободноживущими водорослями. Действительно, на формирование внешнего облика этих слоевищ оказывают большое влияние морфологические особенности их фикобионта — сине-зеленых водорослей. Для клеток этих водорослей чрезвычайно характерны слизистые желатинные образования, которые формируются чаще всего в результате ослизнения толстых оболочек клеток или как продукт выделения их содержимого. До последнего времени даже считали, что сине-зеленые водоросли полностью определяют форму слоевища у большинства слизистых лишайников. Однако поляризационно- оптические исследования, проведенные в последние годы, показали, что слизистая масса этих слоевищ образована не только водорослью, но и грибом.
Лишайники обнаружены даже в бесплодных арктических пустынях и внутри антарктических горных пород. Лишайники распространены по всему миру, но особенно разнообразны в тропиках, высокогорьях и в тундре. А вот в лабораториях лишайники достаточно быстро погибают. И только в 1980 году Более примитивным считается слоевище гомеомерного строения. Если рассмотреть поперечный срез такого слоевища под микроскопом, то хорошо видно, что оно образовано беспорядочными переплетениями гиф гриба, среди которых разбросаны отдельные клетки или нити водорослей. Такое строение наиболее характерно для тех лишайников, фикобионтом которых являются сине-зеленые водоросли — носток, глеокапса и некоторые другие. Эти лишайники образуют группу, известную под названием слизистых лишайников. У лишайников, которые имеют фикобионтом зеленые водоросли, слоевище гомеомерного строения встречается очень редко, лишь у наиболее примитивных накипных форм. Слоевища слизистых лишайников в сухом состоянии имеют вид темных или даже черных твердых и хрупких корок или пленок. Но они обладают одной интересной особенностью — способностью впитывать огромное количество воды, в 20 — 30 раз превышающее их собственную массу. При увлажнении они сильно разбухают, ослизняются, приобретают характер студня или желе и становятся оливково-зелеными или грязно-зелеными. Во внешнем облике этих лишайников имеется много общего с некоторыми свободноживущими водорослями. Действительно, на формирование внешнего облика этих слоевищ оказывают большое влияние морфологические особенности их фикобионта — сине-зеленых водорослей. Для клеток этих водорослей чрезвычайно характерны слизистые желатинные образования, которые формируются чаще всего в результате ослизнения толстых оболочек клеток или как продукт выделения их содержимого. До последнего времени даже считали, что сине-зеленые водоросли полностью определяют форму слоевища у большинства слизистых лишайников. Однако поляризационно- оптические исследования, проведенные в последние годы, показали, что слизистая масса этих слоевищ образована не только водорослью, но и грибом.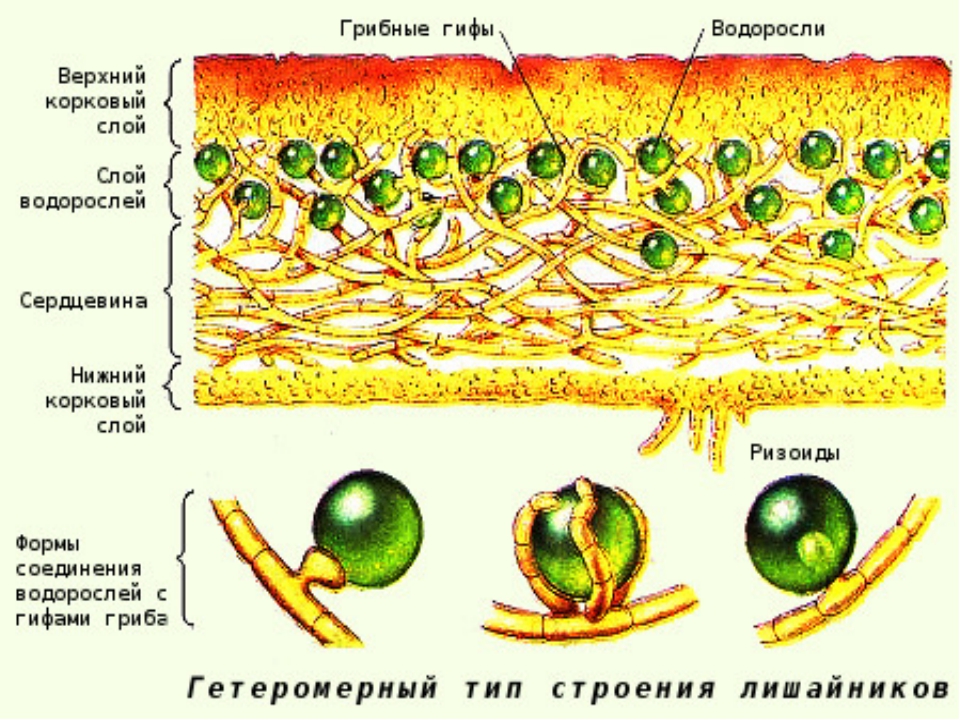 Значительная часть желатина этих лишайников продуцируется грибным компонентом. Гриб определяет размеры слоевища, формирует лопасти и основные детали общего вида. Среди слизистых лишайников встречаются как накипные, так и листоватые и кустистые формы. Но, в отличие от других лишайников, у слизистых усложнение морфологического строения почти не приводит к анатомическим преобразованиям в слоевище. Как у накипных, так и у более высокоразвитых листоватых и кустистых форм этих лишайников структура слоевища гомеомерная, т. е. водоросли всегда разбросаны по всей толще слоевища. Слизистые лишайники насчитывают небольшое число видов, всего 750, что составляет толь ко 3% от общего числа всех известных видов лишайников. Сравнительно небольшое количество этих лишайников, по- видимому, можно объяснить более поздним вовлечением сине-зеленых водорослей в процесс образования слоевища лишайников. Но также возможно, что захват грибом сине-зеленых водорослей был менее успешным по сравнению с зелеными водорослями. В силу особенностей строения сине- зеленых водорослей, нередко собранных в колонии и обладающих плотными, богатыми слизью оболочками, эти водоросли оказались более трудным объектом для гриба при установлении с ними контакта и при построении слоевища. Безусловно, при захвате этих водорослей и их передвижении гриб испытывает гораздо больше трудностей, чем с одноклеточными и другими формами зеленых водорослей. Может быть, именно этим можно объяснить и примитивное анатомическое строение слизистых лишайников. Для остального огромного большинства лишайников характерна гетеромерная структура, при которой в слоевищах можно различить дифференцированные слои. При этом чем сложнее морфологическое строение слоевища лишайника, тем сложнее и его анатомическая структура. У этих растений в ходе эволюции усложнение морфологического строения слоевища шло параллельно усложнению его анатомической структуры. Так, в слоевище накипных лишайников, наиболее примитивных форм, можно различить только три анатомических слоя: коровой слой, слой водорослей и сердцевину.
Значительная часть желатина этих лишайников продуцируется грибным компонентом. Гриб определяет размеры слоевища, формирует лопасти и основные детали общего вида. Среди слизистых лишайников встречаются как накипные, так и листоватые и кустистые формы. Но, в отличие от других лишайников, у слизистых усложнение морфологического строения почти не приводит к анатомическим преобразованиям в слоевище. Как у накипных, так и у более высокоразвитых листоватых и кустистых форм этих лишайников структура слоевища гомеомерная, т. е. водоросли всегда разбросаны по всей толще слоевища. Слизистые лишайники насчитывают небольшое число видов, всего 750, что составляет толь ко 3% от общего числа всех известных видов лишайников. Сравнительно небольшое количество этих лишайников, по- видимому, можно объяснить более поздним вовлечением сине-зеленых водорослей в процесс образования слоевища лишайников. Но также возможно, что захват грибом сине-зеленых водорослей был менее успешным по сравнению с зелеными водорослями. В силу особенностей строения сине- зеленых водорослей, нередко собранных в колонии и обладающих плотными, богатыми слизью оболочками, эти водоросли оказались более трудным объектом для гриба при установлении с ними контакта и при построении слоевища. Безусловно, при захвате этих водорослей и их передвижении гриб испытывает гораздо больше трудностей, чем с одноклеточными и другими формами зеленых водорослей. Может быть, именно этим можно объяснить и примитивное анатомическое строение слизистых лишайников. Для остального огромного большинства лишайников характерна гетеромерная структура, при которой в слоевищах можно различить дифференцированные слои. При этом чем сложнее морфологическое строение слоевища лишайника, тем сложнее и его анатомическая структура. У этих растений в ходе эволюции усложнение морфологического строения слоевища шло параллельно усложнению его анатомической структуры. Так, в слоевище накипных лишайников, наиболее примитивных форм, можно различить только три анатомических слоя: коровой слой, слой водорослей и сердцевину. Такое же анатомическое строение имеют и некоторые листоватые лишайники, например представители рода пельтигера. Но у большинства листоватых лишайников, более эволюционно продвинутых форм, в связи с отделением от поверхности субстрата происходит образование еще одного корового слоя — с нижней стороны слоевища. В таких слоевищах можно различить уже четыре слоя: верхний коровой слой, слой водорослей, сердцевину и нижний коровой слой. У кустистых лишайников с плоскими лентовидными лопастями образуется еще один анатомический слой — слой водорослей с нижней стороны слоевища. У этих лишайников в слоевище можно различить уже пять анатомических слоев: два коровых слоя на верхней и нижней поверхности слоевища, два слоя водорослей, тоже с верхней и нижней стороны лопастей, и сердцевину.И наконец, кустистые лишайники с округлыми лопастями, обладающие наиболее высокоорганизованными слоевищами, имеют радиальную структуру: снаружи лопасти этих лишайников покрыты коровым слоем, под ним расположен слой водорослей, а центральная часть слоевища заполнена сердцевиной. Каждый из перечисленных анатомических слоев слоевища выполняет в жизни лишайника ту или иную функцию и в зависимости от этого имеет совершенно определенное строение. Коровой слой играет в жизни лишайника очень важную роль. Он выполняет сразу две функции: защитную и укрепляющую. Он защищает внутренние слои слоевища от воздействия внешней среды, прежде всего водоросли от чрезмерного освещения. Поэтому коровой слой лишайников обычно бывает плотного строения и окрашен в сероватый, коричневый, оливковый, желтый, оранжевый или красноватый цвет. Коровой слой служит и для укрепления слоевища. Чем выше слоевище поднимается над субстратом, тем более оно нуждается в укреплении. Укрепляющие механические функции в таких случаях нередко выполняет толстый коровой слой. Гифы корового слоа срастаются своими стенками и могут образовывать очень плотные и сложные плектенхимы — клеточные ткани различного строения. Реже гифы корового слоя лишайников не срастаются, а лежат свободно параллельно друг другу.
Такое же анатомическое строение имеют и некоторые листоватые лишайники, например представители рода пельтигера. Но у большинства листоватых лишайников, более эволюционно продвинутых форм, в связи с отделением от поверхности субстрата происходит образование еще одного корового слоя — с нижней стороны слоевища. В таких слоевищах можно различить уже четыре слоя: верхний коровой слой, слой водорослей, сердцевину и нижний коровой слой. У кустистых лишайников с плоскими лентовидными лопастями образуется еще один анатомический слой — слой водорослей с нижней стороны слоевища. У этих лишайников в слоевище можно различить уже пять анатомических слоев: два коровых слоя на верхней и нижней поверхности слоевища, два слоя водорослей, тоже с верхней и нижней стороны лопастей, и сердцевину.И наконец, кустистые лишайники с округлыми лопастями, обладающие наиболее высокоорганизованными слоевищами, имеют радиальную структуру: снаружи лопасти этих лишайников покрыты коровым слоем, под ним расположен слой водорослей, а центральная часть слоевища заполнена сердцевиной. Каждый из перечисленных анатомических слоев слоевища выполняет в жизни лишайника ту или иную функцию и в зависимости от этого имеет совершенно определенное строение. Коровой слой играет в жизни лишайника очень важную роль. Он выполняет сразу две функции: защитную и укрепляющую. Он защищает внутренние слои слоевища от воздействия внешней среды, прежде всего водоросли от чрезмерного освещения. Поэтому коровой слой лишайников обычно бывает плотного строения и окрашен в сероватый, коричневый, оливковый, желтый, оранжевый или красноватый цвет. Коровой слой служит и для укрепления слоевища. Чем выше слоевище поднимается над субстратом, тем более оно нуждается в укреплении. Укрепляющие механические функции в таких случаях нередко выполняет толстый коровой слой. Гифы корового слоа срастаются своими стенками и могут образовывать очень плотные и сложные плектенхимы — клеточные ткани различного строения. Реже гифы корового слоя лишайников не срастаются, а лежат свободно параллельно друг другу. При этом они могут образовывать два различных типа корового слоя. Если гифы расположены параллельными рядами перпендикулярно поверхности слоевища, то образуется палисадный коровой слой. В слоевищах лишайников верхний и нижний коровые слои могут быть одинаковыми по окраске и окраски сердцевины. Если кристаллы лишайниковых веществ окрашены в тот или иной цвет, то в зависимости от этого и сердцевинный слой приобретает золотисто-желтую, кроваво-красную, серую или другую окраску. Лишайниковые вещества обладают одной важной особенностью: они не растворимы или очень слабо растворимы в холодной воде. Благодаря этому свойству кристаллы, покрывая поверхность сердцевинных гиф, препятствуют их смачиванию. Поэтому даже во влажном слоевище лишайника его сердцевинный слой может частично оставаться сухим и содержать воздух, необходимый для клеток водорослей. У некоторых кустистых лишайников, слоевище которых далеко отстает от субстрата, сердцевинный слой, помимо проводящей функции, выполняет еще одну — укрепляющую. Как отмечалось, у большинства лишайников укрепление слоевища происходит с помощью корового слоя, который у высокоразвитых кустистых и листоватых форм постепенно становится все более толстым и плотным. Однако дальнейшее развитие в этом направлении могло бы привести к ухудшению нормальных ассимиляционных процессов: очень толстый и плотный коровой слой препятствовал бы проникновению в слоевище воздуха и света. Поэтому в процессе развития кустистых форм происходит постепенное снижение роли корового слоя в укреплении слоевища и образование механической ткани под зоной водорослей, т. е. в сердцевинном слое. В сердцевинном слое высокоразвитых кустистых лишайников возникают тяжи, которые расположены вдоль ветвей слоевища и состоят из толстостенных, плотно сросшихся друг с другом гиф. Как же воздух проникает в слоевище лишайника? У некоторых форм на поверхности слоевища вовсе не образуется корового слоя. Поверхность их слоевищ образована рыхлыми сердцевинными гифами. В таком случае воздух беспрепятственно по промежуткам между рыхло расположенными гифами проникает к водорослям, которые разбросаны отдельными группами.
При этом они могут образовывать два различных типа корового слоя. Если гифы расположены параллельными рядами перпендикулярно поверхности слоевища, то образуется палисадный коровой слой. В слоевищах лишайников верхний и нижний коровые слои могут быть одинаковыми по окраске и окраски сердцевины. Если кристаллы лишайниковых веществ окрашены в тот или иной цвет, то в зависимости от этого и сердцевинный слой приобретает золотисто-желтую, кроваво-красную, серую или другую окраску. Лишайниковые вещества обладают одной важной особенностью: они не растворимы или очень слабо растворимы в холодной воде. Благодаря этому свойству кристаллы, покрывая поверхность сердцевинных гиф, препятствуют их смачиванию. Поэтому даже во влажном слоевище лишайника его сердцевинный слой может частично оставаться сухим и содержать воздух, необходимый для клеток водорослей. У некоторых кустистых лишайников, слоевище которых далеко отстает от субстрата, сердцевинный слой, помимо проводящей функции, выполняет еще одну — укрепляющую. Как отмечалось, у большинства лишайников укрепление слоевища происходит с помощью корового слоя, который у высокоразвитых кустистых и листоватых форм постепенно становится все более толстым и плотным. Однако дальнейшее развитие в этом направлении могло бы привести к ухудшению нормальных ассимиляционных процессов: очень толстый и плотный коровой слой препятствовал бы проникновению в слоевище воздуха и света. Поэтому в процессе развития кустистых форм происходит постепенное снижение роли корового слоя в укреплении слоевища и образование механической ткани под зоной водорослей, т. е. в сердцевинном слое. В сердцевинном слое высокоразвитых кустистых лишайников возникают тяжи, которые расположены вдоль ветвей слоевища и состоят из толстостенных, плотно сросшихся друг с другом гиф. Как же воздух проникает в слоевище лишайника? У некоторых форм на поверхности слоевища вовсе не образуется корового слоя. Поверхность их слоевищ образована рыхлыми сердцевинными гифами. В таком случае воздух беспрепятственно по промежуткам между рыхло расположенными гифами проникает к водорослям, которые разбросаны отдельными группами. У крупнолистоватых лишайников на нижней стороне слоевищ также не развивается коровой слой, ее образуют рыхло расположенные сердцевинные гифы. И у этих лишайников воздух свободно проникает в сердцевину и далее по промежуткам между гифами к клеткам водорослей. Но такие случаи являются исключением. У большинства лишайников на поверхности слоевища образуется плотный коровой слой и доступ воздуха во внутренние слои слоевища сильно затруднен. Для осуществления нормального газообмена на поверхности слоевища лишайников образуются особые отверстия — специальные органы, которые служат для аэрации внутренних частей растения. Это разрывы коры, через которые воздух, как через маленькие форточки, проникает внутрь слоевища. Они являются органами газообмена лишайников и носят на звание псевдоцифел. Помимо псевдоцифелл, у лишайников образуются и другие органы газообмена. Так, на нижней поверхности листоватых лишайников можно заметить круглые правильной формы белые углубления. Эти разрывы нижней коры лишайника, по форме напоминающие маленькие чашечки, называют цифеллами. Дно их выстлано рыхлорасположенными, шарообразными клетками сердцевины, через промежутки между которыми воздух легко проникает в сердцевину. У некоторых лишайников органы аэрации слоевища имеют вид маленьких точковидных пор, расположенных в нижней коре лишайников. Иногда такие поры возникают на вершине бородавочек, разбросанных на верхней поверхности лишайников. В том случае, когда на поверхности слоевища лишайников не образуются специальные органы газообмена, на помощь приходят различные трещинки и разрывы в коровом слое. Эти трещинки могут возникнуть как в результате особенностей роста слоевища, так и в результате повреждения слоевища ветром, животными, человеком.
У крупнолистоватых лишайников на нижней стороне слоевищ также не развивается коровой слой, ее образуют рыхло расположенные сердцевинные гифы. И у этих лишайников воздух свободно проникает в сердцевину и далее по промежуткам между гифами к клеткам водорослей. Но такие случаи являются исключением. У большинства лишайников на поверхности слоевища образуется плотный коровой слой и доступ воздуха во внутренние слои слоевища сильно затруднен. Для осуществления нормального газообмена на поверхности слоевища лишайников образуются особые отверстия — специальные органы, которые служат для аэрации внутренних частей растения. Это разрывы коры, через которые воздух, как через маленькие форточки, проникает внутрь слоевища. Они являются органами газообмена лишайников и носят на звание псевдоцифел. Помимо псевдоцифелл, у лишайников образуются и другие органы газообмена. Так, на нижней поверхности листоватых лишайников можно заметить круглые правильной формы белые углубления. Эти разрывы нижней коры лишайника, по форме напоминающие маленькие чашечки, называют цифеллами. Дно их выстлано рыхлорасположенными, шарообразными клетками сердцевины, через промежутки между которыми воздух легко проникает в сердцевину. У некоторых лишайников органы аэрации слоевища имеют вид маленьких точковидных пор, расположенных в нижней коре лишайников. Иногда такие поры возникают на вершине бородавочек, разбросанных на верхней поверхности лишайников. В том случае, когда на поверхности слоевища лишайников не образуются специальные органы газообмена, на помощь приходят различные трещинки и разрывы в коровом слое. Эти трещинки могут возникнуть как в результате особенностей роста слоевища, так и в результате повреждения слоевища ветром, животными, человеком.
Лишайники
Строение тела
Лишайники — группа симбиотических организмов, морфологическую основу тела которых образует гриб. Под симбиозом понимают взаимополезное сожительство организмов, принадлежащих к разным видам. В теле лишайников сочетаются два компонента: автотрофный —
В теле лишайников сочетаются два компонента: автотрофный —
По форме и величине лишайники разнообразны, их размеры — от нескольких до десятков сантиметров. Вегетативное тело представлено слоевищем, или талломом. В зависимости от строения слоевища выделяют накипные, имеющие вид корочки, листовамые и кустистые.
Лишайники представляют собой своеобразную группу комплексных организмов, тело которых состоит из двух компонентов — гриба и водоросли. Как организмы, лишайники были известны задолго до открытия их сущности, Еще великий Теофраст, «отец ботаники» (IV-III вв. до н. э.), дал описание двух лишайников — успей и рочеллы — которые уже тогда использовали для получения ароматических и красящих веществ. Правда, в те времена их нередко называли то мхами, то водорослями, то даже «хаосом природы» и «убогой нищетой растительности»,
Сейчас известно около 20 000 видов лишайников. Наука о лишайниках называется лихенологией. Специфический признак лишайников — симбиоз двух разных организмов: гетеротрофного гриба (микобионт) и автотрофной водоросли (фикобионт), В лишайнике оба эти компонента вступают в тесные взаимоотношения: гриб окружает водоросли и даже может проникать в их клетки. Лишайники образуют особые морфологические типы — жизненные формы, которые не встречаются у отдельных слагающих их организме». Метаболизм лишайников имеет специфический характер: только в них образуются лишайниковые кислоты, не встречающиеся у других организмов. Специфичны также и способы размножения лишайников как целостных организмов.
Отношения гриба и водоросли в лишайнике очень сложны. С одной стороны, они взаимополезны: водоросль снабжает гриб синтезируемыми ею органическими веществами, а гриб обеспечивает доставку воды и минеральных солей, а также защищает от воздействия неблагоприятных факторов среды, таких, как перегрев, высыхание, излишняя инсоляция.
Слоевище (так называется тело лишайника) разнообразно по форме, размерам, окраске и строению. Окраска лишайников варьирует: они бывают белые, серые, желтые, оранжевые, зеленые, черные; это определяется характером пигментов, содержащихся в оболочке гиф. Пигментация способствует защите водорослевого компонента от чрезмерного освещения. Иногда бывает и наоборот: лишайники Антарктиды окрашены в черный цвет, который поглощает тепловые лучи.
По форме слоевища лишайники делятся на накипные, листоватые и кустистые.
Слоевище накипных лишайников имеет вид корочки, плотно сросшейся с субстратом сердцевинными гифами. Иногда оно представлено порошковидным налетом.
Листоватые лишайники имеют вид пластинки, горизонтально расположенной на субстрате, прикрепляясь к нему выростами гиф — ризинами. Слоевище может быть цельным или рассеченным прижатым к субстрату или приподнимающимся над ним.
Слоевище остистых лишайником имеет вид разветвленного стоячего или повислого кустика либо неразветвленных стоячих столбиков. К субстрату они прикрепляются короткой ножкой, расширенной на конце пяточкой.
По анатомическому строению лишайники бывают: 1) го-меомерными, когда водоросли разбросаны по всему телу лишайника; 2)гетеромерными, когда водоросли образуют в слоевище обособленный слой. Сверху слоевище покрыто коревым слоем, состоящим из срастающихся своими стенками клеток и имеющим вид клеточной ткани — плектенхимы, Кора играет защитную функцию, а также укрепляет слоевище. Органы прикрепления листоватых лишайников ризоиды и ризины; первые состоят из одного ряда клеток, а вторые — из соединенных в тяжи ризоидов.
Сверху слоевище покрыто коревым слоем, состоящим из срастающихся своими стенками клеток и имеющим вид клеточной ткани — плектенхимы, Кора играет защитную функцию, а также укрепляет слоевище. Органы прикрепления листоватых лишайников ризоиды и ризины; первые состоят из одного ряда клеток, а вторые — из соединенных в тяжи ризоидов.
Лишайники размножаются либо спорами, которые образует гриб, либо фрагментами слоевища, то есть вегетативно,
Половое размножение лишайников обеспечивают апотеции находящиеся на верхней стороне слоевища и имеющие блюдцевидную форму. Там формируются споры в результате слияния половых клеток. Споры распространяются ветром и, попав в благоприятные условия, прорастают в гифу, но новый лишайник сформируется только в том случае, если гифа встретит подходящую водоросль.
Вегетативно лишайники размножаются изидиями и соредиями — выростами на слоевище, содержащими оба компонента лишайника.
Широкое распространение лишайников на земном шаре свидетельствует об их огромном значении. Особенно велика их роль в тундре и лесотундре, где они составляют заметную часть растительного покрова и где с ними связана жизнь большой группы животных: они являются убежищем для беспозвоночных и мелких позвоночных животных, пищей для них и для крупных позвоночных, таких как северный олень. Лишайник исландский мох в северных странах используется в качестве дополнения к корму домашних животных и добавки при выпечке хлеба,
Во всех биогеоценозах лишайники выполняют фотосинтетическую, почвообразовательную функции. Особенно при заселении свежеобнажённых субстратов, каменистых, скальных, бедных органикой.
В хозяйственной деятельности-человека лишайники могут использоваться как продуценты лишайниковых кислот — соединений, обладающих антибиотическими свойствами. Широкое применение лишайников в медицине основано на их тонизирующих и антисептических свойствах. Вырабатываемые ими лишайниковые кислоты обладают антимикробной активностью в отношении стафилококков, стрептококков, туберкулезной палочки, а также успешно применяются при лечении дерматитов.
С древних времен известно использование лишайников в парфюмерии, основанное на высоком содержании в их слоевищах ароматических веществ и эфирных масел. В частности, дубовый мох используется при изготовлении духов.
В качестве красителей эта группа растений известна также очень давно, а шотландский твид до сих пор окрашивается экстрактами лишайников. Широко используемый в химии индикатор лакмус также является производным лишайников.
Лишайники чувствительны к наличию в воздухе вредных примесей, особенно содержащих тяжелые металлы, В последнее время они широко применяются при оценке загрязнения воздуха и для контроля радиационной обстановки.
Также использованы материалы личной страницы Иванова Андрея http://www.chat.ru/~dronisimo/homepage1/
Лишайники
Лишайники ─ это группа симбиотических организмов. Что это значит?
В переводе с греческого (симбиос) συμ-βίωσις — «совместная жизнь». А необычное в этом растительном организме то, что тело лишайника ― слоевище ─ состоит из грибов и микроскопических водорослей. Их совместная жизнь тесно переплелась и стала единым целым.
Слоевище состоит из множества переплетенных грибных нитей (гиф). Между ними группами или одиночно расположены одноклеточные зелёные водоросли. Гифы гриба плотно оплетают водоросли. А в некоторых местах гифы прорастают внутрь клетки водоросли.
Лишайники способны поглощать воду как из субстрата (то есть из своего места обитания), так и из воздуха всем телом, светолюбивы, нетребовательны к субстрату, чувствительны к загрязнению воздуха.
И гриб, и водоросль получают от совместного существования пользу.
Гриб
от водоросли получает питательные вещества, в основном глюкозу, производимые водорослью
в результате фотосинтеза, а также азотсодержащие вещества. А водоросль получает
от гриба воду и минеральные вещества. Также гриб создаёт водоросли
благоприятный микроклимат. Он защищает её от высыхания, закрывая от
ультрафиолетового излучения солнца. Обеспечивает жизнь на кислых субстратах,
смягчает действие ряда других неблагоприятных факторов.
А водоросль получает
от гриба воду и минеральные вещества. Также гриб создаёт водоросли
благоприятный микроклимат. Он защищает её от высыхания, закрывая от
ультрафиолетового излучения солнца. Обеспечивает жизнь на кислых субстратах,
смягчает действие ряда других неблагоприятных факторов.
Если отделить водоросль от гриба, то она спокойно может существовать и без гриба, а вот гриб без водоросли существовать не может.
Лишайники довольно разнообразны. Их тела окрашены в различные цвета в зависимости от того, какие пигменты в них находятся: от белого до ярко-жёлтого, коричневого, оранжевого, зелёного, синего, серого, чёрного.
По внешнему виду различают лишайники накипные, листоватые и кустистые.
Тело накипных, или корковых, лишайников представляет собой корочку («накипь»), его нижняя поверхность плотно срастается с субстратом и не отделяется без значительных повреждений.
К накипным относится лишайник ― бацидия. Тело его обычно красновато-коричневого цвета. Он встречается в лесной зоне, на юге в горах, нередко в степях и полупустынях, но заходит также и в Арктику.
Накипные лишайники могут жить на камнях, крутых склонах гор, деревьях и даже на бетонных стенах.
Листоватые лишайники имеют вид пластин разной формы и окраски, они более или менее плотно прикрепляются к субстрату при помощи выростов нижнего коркового слоя.
Листоватые лишайники разрастаются на камнях и на коре деревьев. Из таких лишайников особенно часто встречается золотисто-жёлтая ксантория настенная.
Листоватый лишайник пармелия обладает лекарственными свойствами. Этот лишайник широко используют в народной медицине степные жители.
Отвар
из пармелии обладает антисептическим, бактерицидным, ранозаживляющим и
кровоостанавливающим эффектом.
Тела кустистых лишайников образуют множество округлых или плоских веточек. Слоевище этой формы характеризуются верхушечным ростом. В них водоросли могут максимально использовать свет. Такие лишайники могут расти как на земле, так и свисать с деревьев, скал.
Интересным представителем кустистых лишайников является лишайник-бородач. Его тело, как борода свисает со старых ветвей до самой земли, обычно в тёмном еловом лесу.
К кустистым лишайникам относят и ягель (олений мох), которым питается северный олень. Он широко распространен в тундре. Как и другие лишайники, ягель растёт очень медленно ─ 3–5 мм в год. Для восстановления пастбища после выпаса оленей может потребоваться несколько десятилетий.
Продолжительность жизни всех лишайников от 50 до 100 лет. А возраст некоторых, например ризокарпона географического, может достигать более 4000 лет.
Лишайники достаточно неприхотливые организмы. Это позволяет им поселятся на голых, бесплодных скалах и камнях, на крышах домов и других местах где, казалось бы, жизнь невозможна.
Не имея корней, они довольно крепко прикрепляются к субстрату особыми выростами, похожими на корни, расположенными на нижней стороне слоевища. Однако в отличие от корней высших растений они служат лишь для прикрепления.
В пустынях и полупустынях встречаются лишайники, имеющие форму шариков (напр., аспицилия), они легко переносятся ветром на большие расстояния, отчего их часто называют кочующими.
Существует легенда о том, как в бесплодной пустыне люди, истощенные голодом и трудным переходом, встретили на земле массу мелких сухих крупинок, похожих на манную крупу. Насытившись этими крупинками, они обрели силы, которые помогли закончить трудный путь.
Предполагают,
что крупинки, о которых рассказано в легенде, ― это аспицилия
съедобная, или манна лишайниковая.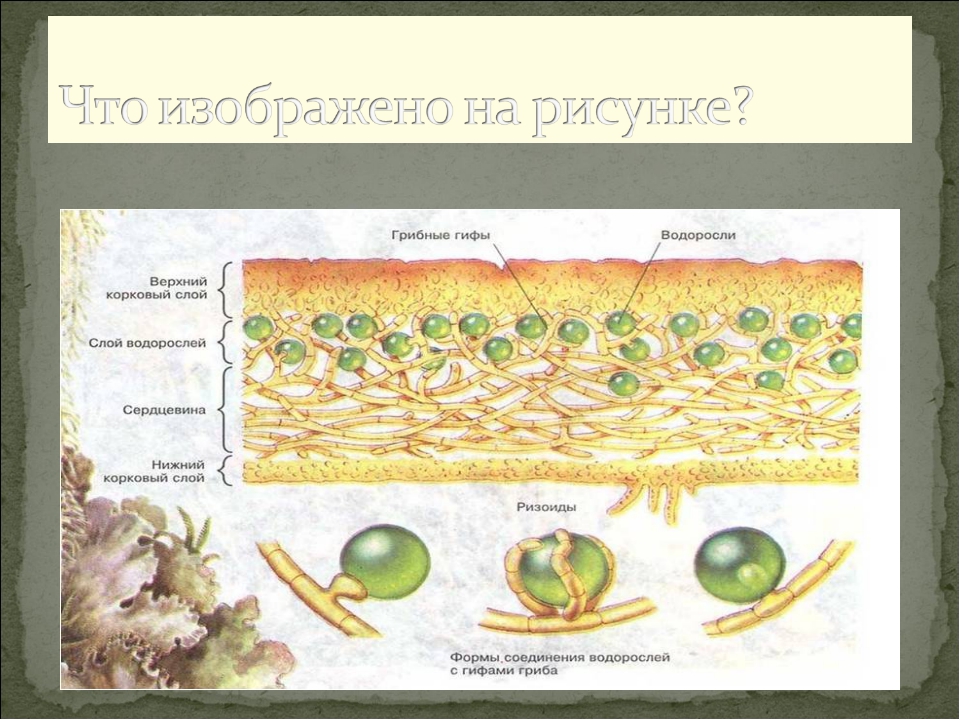
Отсюда пошло выражение «манна небесная».
Лишайники очень чувствительны к загрязнению воздуха. В присутствии дымов и ядовитых газов они быстро гибнут. По состоянию лишайников (особенно в городах) определяют чистоту воздуха.
При загрязнении атмосферы сначала вымирают кустистые, потом листоватые, а затем и накипные лишайники. Изучите лишайники, которые произрастают в вашей местности и сделайте вывод о чистоте воздуха.
Однако лишайники спокойно переживают неблагоприятные климатические условия. Они настолько выносливы, что растут даже там, где отсутствует другая растительность, например в Арктике и Антарктике. Живым организмам здесь приходится жить при очень низких температурах. Осадки выпадают всегда в виде снега, а лишайник не может поглощать воду в такой форме.
Но его выручает чёрная окраска слоевища. Тёмная поверхность тела лишайника, благодаря высокой солнечной радиации, быстро нагревается даже при низких температурах. Снег, попав на нагретое слоевище, тает. Лишайник сразу впитывает появившуюся влагу, обеспечивая себя водой, необходимой ему для дыхания и фотосинтеза.
В жару лишайники настолько высыхают, что кажутся совершенно безжизненными. Но после дождя быстро оживают вновь. Всей поверхностью своего тела они впитывают влагу дождей и туманов.
Лишайники, как и любые другие живые организмы, размножаются. Размножение их происходит, главным образом, кусочками слоевища.
А также особыми группами клеток гриба и водоросли. Группы клеток образуются внутри тела лишайника, которое затем под давлением разрывается, и клетки разносятся ветром и дождевыми потоками.
В природе лишайники играют важную роль, первыми поселяясь в самых бесплодных местах. Выделяя особые кислоты, они медленно разрушают горные породы.
Скопившаяся
на них пыль и песчинки создают тонкий слой почвы, в котором могут закрепиться
мхи и другие наземные растения. Мхи и травы, разрастаясь, затеняют наземные
лишайники, засыпают их отмершими частями своих тел, и лишайники исчезают со
временем с этого места.
Мхи и травы, разрастаясь, затеняют наземные
лишайники, засыпают их отмершими частями своих тел, и лишайники исчезают со
временем с этого места.
Среди лишайников почти нет ядовитых видов. Поэтому практическое значение их довольно велико. Многие виды лишайников используют в медицине благодаря высокому содержанию усниновой кислоты, которая способна убивать вирусы и бактерии. Благодаря этим свойствам лишайники используются в косметике, стоматологии и других областях медицины.
Препараты исландского лишайника обладают антибактериальными свойствами. Их используют при лечении инфицированных ран и ожогов. Из этого лишайника готовят отвары для лечения простуды и диареи.
Также лишайники являются пищей и убежищем для многих мелких организмов.
III Международный конкурс научно-исследовательских и творческих работ учащихся Старт в науке
БИОИНДИКАЦИОННЫЕ ИССЛЕДОВАНИЯ ЧИСТОТЫ ВОЗДУХА П. Г. Т. АЛЕКСЕЕВКА Г. О. КИНЕЛЬ САМАРСКОЙ ОБЛАСТИ
Волощук Е.А. 11ГБОУ СОШ №4 п.г.т.Алексеевка
Петина О.В. 1Петина О.В. 11ГБОУ СОШ №4 п.г.т.Алексеевка г.о.Кинель
Текст работы размещён без изображений и формул.
Полная версия работы доступна во вкладке «Файлы работы» в формате PDF
ВВЕДЕНИЕ
Глобальный экологический кризис — печальная реальность наших дней. Если с 1850 по 1950 годы каждые десять лет биосфера теряла один вид растений, то, начиная с 2000 года, один вид уничтожается каждый час, а 23 гектара леса — каждую минуту! Под угрозой уничтожения находятся 30 тысяч видов растений, повсеместно деградирует почвенный покров планеты, сокращаются запасы пресной воды, в южных районах плодородные земли превращаются в пустыни, в тундрах при нефтяных разработках полностью уничтожается растительный покров (Петров, Абрамов, 1994).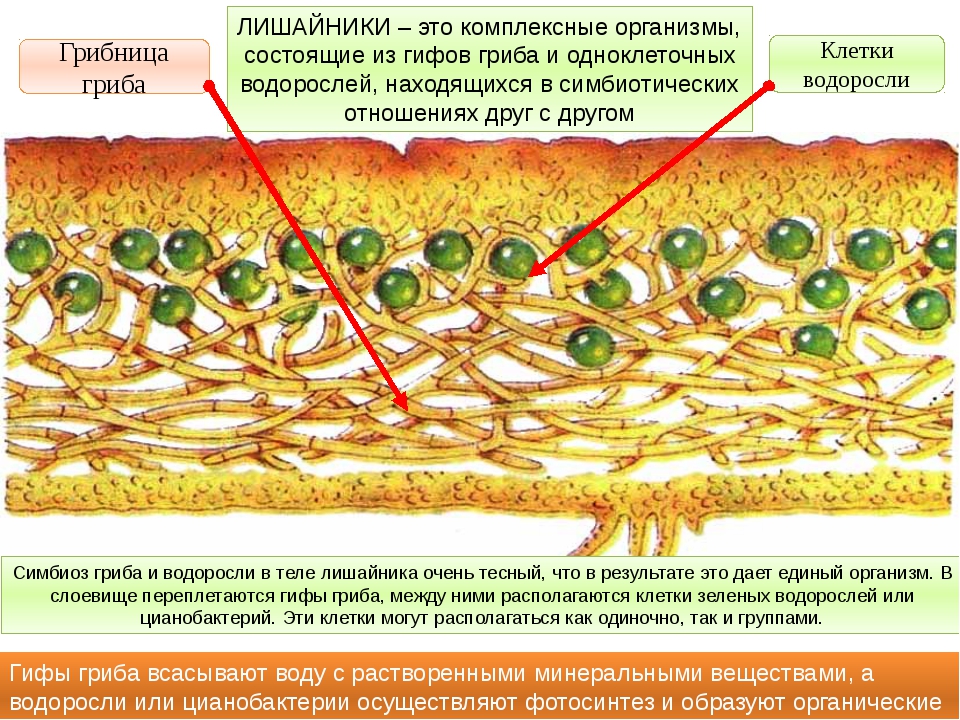
В этих условиях ученым-биологам все труднее становится работать, ведь для изучения требуется изъятие из природы некоторых видов, а это в последнее время становится невозможным.
1. ОБЩАЯ ХАРАКТЕРИСТИКА ЛИШАЙНИКОВ
Лишайники представляют своеобразную группу комплексных организмов, тело которых всегда состоит из двух компонентов — гриба и водоросли. Сейчас каждый школьник знает, что в основе биологии лишайников лежит явление симбиоза — сожительства различных организмов. Но еще немногим более ста лет назад лишайники были для ученых великой загадкой, и открытие Симоном Швенденером в 1867 г. их сущности оценивалось как одно из наиболее удивительных открытий того времени.
Однако как организмы лишайники были известны ученым и в народе задолго до открытия их сущности. Еще великий Теофраст (371—286 до н.э.), «отец ботаники», дал описание двух лишайников — уснеи (Usnea) и ро-челлы (Roccella). Последнюю уже тогда использовали для получения красящих веществ. Постепенно количество известных видов лишайников возрастало. Правда, в те времена их называли нередко то мхами, то водорослями, то даже «хаосом природы» и «убогой нищетой растительности».
Сейчас известно более 20 000 лишайников. И каждый год ученые обнаруживают и описывают десятки и сотни новых неизвестных видов.
В настоящее время лихенология (лат. lichen — лишайник)— наука о лишайниках — изучает сложный комплекс проблем, связанных с возникновением, филогенией, строением, систематикой, биохимией, физиологией, распространением и экологией лишайников (Жизнь растений, 1977).
1.1. Строение лишайников
Чем же лишайники отличаются от других растений?
Во-первых, специфичный признак лишайников—симбиотическое сожительство двух разных организмов — гетеротрофного гриба (микобионт) и автотрофной водоросли (фикобионт).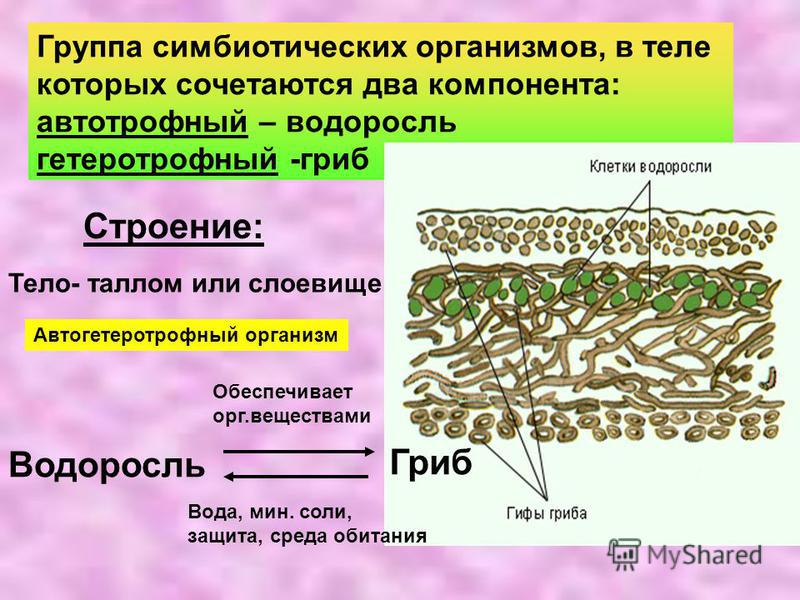 Не всякое сожительство гриба и водоросли образует лишайник. Лишайниковое сожительство должно быть постоянным и исторически выработавшимся, а не случайным, кратковременным. В природе бывают случаи, когда гриб и водоросль образуют временное смешанное скопление, но это еще не лишайник. В настоящем лишайнике гриб и водоросль вступают в тесные взаимоотношения, грибной компонент окружает водоросли и может даже проникать в их клетки.
Не всякое сожительство гриба и водоросли образует лишайник. Лишайниковое сожительство должно быть постоянным и исторически выработавшимся, а не случайным, кратковременным. В природе бывают случаи, когда гриб и водоросль образуют временное смешанное скопление, но это еще не лишайник. В настоящем лишайнике гриб и водоросль вступают в тесные взаимоотношения, грибной компонент окружает водоросли и может даже проникать в их клетки.
Во-вторых, лишайники образуют особые морфологические типы, жизненные формы, не встречающиеся отдельно у слагающих лишайниковое слоевище грибов и водорослей, т.е. лишайники прошли исторический, длительный формообразующий процесс на основе симбиоза, приведший к формированию специфичных морфологических форм внешнего и внутреннего строения.
В-третьих, для лишайников в целом и каждого из их компонентов в отдельности характерен особый тип метаболизма. Физиология гриба и водоросли в слоевище лишайника во многом отличается от физиологии свободно-живущих грибов и водорослей.
В-четвертых, весьма специфична биохимия лишайников, образование в них вторичных продуктов обмена — лишайниковых веществ, не встречающихся в других группах организмов.
Наконец, лишайникисущественно отличаются от других групп организмов, в том числе и от свободноживущих грибов и водорослей, особой биологией: способами размножения, медленным ростом, отношением к экологическим условиям и др. На основе этих специфических свойств лишайникам можно дать следующее определение: лишайниками являются организмы, тело (слоевище) которых постоянно состоит из двух компонентов — автотрофного фикобионта и гетеротрофного микобионта, образующих единое симбиотическое сожительство, отличающееся особыми морфологическими типами и особыми физиолого-биохимическими процессами (Жизнь растений, 1977).
1.1.1. Грибной компонент (микобионт)
Представители трех классов грибов — аскомицетов, базидиомицетов и фикомицетов, вступив в контакт с водорослями, дали начало образованию лишайников. О близости с грибами говорит строение вегетативного тела лишайников и их плодовых тел. Вегетативное тело лишайников, называемое, как у всех низших растений, талломом или слоевищем, целиком состоит из переплетения грибных гиф. Водоросли же либо разбросаны в беспорядке среди грибных гиф по всей толще слоевища, либо расположены отдельным дифференцированным слоем несколько ниже его верхней поверхности.
Как известно, вегетативное тело обычных грибов, состоящее из грибных гиф и называемое грибницей или мицелием, в большинстве случаев развивается целиком внутри субстрата, а на поверхности мы видим лишь плодовые тела гриба. В отличие от этого вегетативное тело лишайников, также образованное грибными гифами, целиком живет в воздушной среде (исключение составляет лишь небольшая группа лишайников, слоевище которых развивается внутри камня или древесного субстрата). Лишайники иногда даже называют воздушными грибами.
Гифы, образующие слоевище лишайника, представляют собой простые или разветвленные тонкие (шириной 3—10 мкм) нити, растущие вершиной. Гифа покрыта двухслойной оболочкой, под которой находится протоплазма. Обычно гифы бывают поделены поперечными перегородками на клетки. У большинства лишайников в центральной части поперечных перегородок имеется одно маленькое отверстие, называемое перфорацией. Через перфорацию протягивается тончайший цитоплазматический тяж, который носит название плазмодесмы. Плазмодесма соединяет протопласты соседних клеток. У некоторых лишайников поперечная перегородка имеет не одну, а три перфорации, расположенные на равных расстояниях друг от друга. Кроме того, перфорации образуются не только на поперечных перегородках гиф, но и на их продольных стенках, в результате плазмодесмами соединяются протопласты двух параллельно расположенных соседних гиф.
Протоплазма лишайников слегка зерниста. В вегетативных клетках почти всегда имеется одно ядро, но у некоторых видов бывает по два или по многу ядер в одной клетке.
У целого ряда лишайников оболочки гиф могут сильно разбухать и ослизняться. Это происходит за счет содержания во внешних слоях оболочек пектиновых веществ, легко растворимых в воде углеводов. Особенно сильно ослизняются и набухают при увлажнении гифы так называемых слизистых лишайников, в слоевище которых водорослевым партнером обычно бывает сине-зеленая водоросль. Гифы обычных грибов, как правило, не обладают этой способностью, исключение составляют лишь дрожалковые грибы.
Интересные, специфичные для лишайниковых гиф образования, которые у свободноживущих грибов в природных условиях не встречаются,—так называемые жировые клетки или жировые гифы. Они развиваются обычно в нижней части слоевища, в местах прикрепления к субстрату. Жировые клетки имеют зеленоватую окраску, обусловленную содержанием жира. Они, как правило, крупнее обычных клеток и имеют вздутую мешковидную, иногда почти шаровидную форму. Было замечено, что чаще всего жировые клетки встречаются у лишайников, растущих на известковом субстрате, особенно в слоевищах, глубоко погруженных в известняк. В слоевищах лишайников, которые произрастают на гранитных породах и на древесном субстрате, жировые клетки образуются очень редко. Более того, если лишайник растет на горной породе смешанного состава, то жировые клетки можно заметить только в гифах, проходящих через участки карбонатсодержащих пород, а в гифах того же слоевища на бескарбонатных участках жировые клетки отсутствуют. Это позволило сделать вывод, что образование жира обусловливает углекислота, освобождающаяся при разложении карбонатов гифами лишайников. Однако это пока лишь одна из гипотез, пытающихся объяснить возникновение этих специфичных для лишайников образований.
Благодаря воздушному образу жизни и симбиозу с водорослями у гриба появилась тенденция к развитию сложной вегетативной структуры. Как правило, микобионт лишайников образует сложно устроенные талломы, с хорошо дифференцированными анатомическими слоями, с особыми органами прикрепления, которые встречаются лишь у лишайников (Жизнь растений, 1977).
1.1.2. Водорослевый компонент (фикобионт)
Водоросли, встречающиеся в слоевище лишайников, называют фикобионтом лишайников. По своему систематическому положению они относятся к различным отделам: к сине-зеленым (Cyanophyta), зеленым (Chlorophyta), желто-зеленым (Xanthophyta) и бурым (Phaeophyta) водорослям. Долгое время считали, что каждому виду лишайника соответствует определенный вид водоросли. Однако, как показали дальнейшие исследования, сравнительно небольшое количество водорослей оказались способными существовать в симбиозе с грибом. Только самые неприхотливые из них, обладающие высокой устойчивостью по отношению к факторам внешней среды, приспособились к жизни в окружении грибных гиф. Экспериментальные исследования показали, что лишайниковый гриб при образовании слоевища, как правило, не проявляет избирательной способности по отношению к водорослям. Если взять чистую культуру гриба, выделенного из слоевища лишайника, и поместить в нее какую-либо свободноживущую водоросль (несвойственную данному лишайнику, а другого типа), то грибные гифы начнут разрастаться, давая сложные переплетения, как в начале образования слоевища. Однако слоевище в таком случае все-таки не развивается. Большинство водорослей, введенных в культуру лишайникового гриба, рано или поздно погибает: многие не могут перенести воздействия гиф гриба, причиной гибели других является недостаток света. Водоросли, окруженные со всех сторон грибной тканью, получают гораздо меньше света, чем это необходимо для их нормальной жизнедеятельности. И если водоросль не может приспособиться к осуществлению процесса фотосинтеза при минимальной световой интенсивности, она неизбежно погибает. Значит, при образовании слоевища лишайников не гриб выбирает себе подходящего партнера из числа водорослей. Все зависит от того, сможет ли водоросль существовать в тесном контакте с грибом и при этом нормально жить и развиваться.
Как показало изучение лишайниковых фикобионтов, проведенное в последние годы, в слоевище лишайников встречаются представители 28 родов водорослей. Из сине-зеленых водорослей наиболее обычным фикобионтом в слоевище лишайников являются водоросли рода носток и глеокапса. Встречаются и другие сине-зеленые: хроококкус (Chroococcus) и гиелла (Hyella), калотрикс (Calothrix), дихотрикс (Dichothrix) и ривулярия (Rivularia), спитонема (Scytonema) и стигонема (Stigonema). Из зеленых водорослей в слоевище лишайников встречаются представители хлорококковых и улотриксовых.
Существует взаимосвязь между географическим распространением лишайников и преобладанием в их слоевищах определенных водорослей. Так, в умеренной зоне земного шара около 8% лишайников имеют фикобионтом сине-зеленые водоросли, 9% лишайников — нитчатые или пластинчатые зеленые водоросли типа трентеполии и 83% лишайников — хлорококковые зеленые водоросли. В тропических и субтропических странах увеличивается количество лишайников, в слоевище которых встречается водоросль трентеполия. Так, в тропиках от 5 до 10% лишайников содержат в слоевищах сине-зеленые водоросли. 45—48% лишайников — нитчатые зеленые водоросли типа трентеполии и такое же количество лишайников имеют фикобионтом остальные зеленые водоросли.
Водоросли в слоевище лишайника очень сильно изменяют свой внешний облик. Особенно это касается нитчатых водорослей, которые под влиянием гриба распадаются на отдельные клетки и часто деформируются до неузнаваемости. Размножаются водоросли внутри слоевища обычно делением и апланоспорами. Подвижных клеток размножения, характерных для свободноживущих водорослей, они не образуют. Но в лаборатории, в условиях культуры, фикобионты лишайников, как правило, принимают свой обычный, присущий им в свободноживущем состоянии облик и образуют подвижные зооспоры и гаметы.
Лишайниковые водоросли очень устойчивы к воздействию высокой температуры, способны переносить длительное высушивание. Опыты показали, например, что водоросль коккомикса в слоевище одного из лишайников выдерживала высушивание в течение 23 недель, а после нормализации условий восстанавливала все процессы своей жизнедеятельности. В культуре же этот фикобионт погибал без воды уже через 5 недель.
Все эти данные показывают,что в связи сособенностями жизни в контакте с грибом лишайниковые водоросли претерпевают не только морфологические,но и физиологические изменения (Жизнь растений, 1977).
1.1.3. Взаимоотношения гриба и водоросли в теле лишайника
Вопрос взаимоотношения гриба и водоросли в слоевище лишайника занимал умы ученых еще в конце прошлого столетия, да и в наше время продолжает волновать лихенологов. Со дня открытия С. Швенденера прошло более 100 лет. За этот период появилось не менее десятка теорий, пытающихся объяснить отношения между грибом и водорослью, однако среди них нет ни одной общепризнанной и окончательно доказанной. С. Швенденер, обнаружив, что лишайник состоит из гриба и водоросли, предположил, что гриб в слоевище паразитирует, на водоросли. Однако он ошибочно отвел грибу роль хозяина, а водоросли — раба.
Но уже в те времена некоторые ученые выдвинули мысль о двустороннем паразитизме компонентов лишайника — гриба на водоросли и водоросли на грибе. При этом было высказано предположение, что гриб и водоросль в слоевище лишайника находятся в полном морфо-физиологическом единстве и связаны между собой так же, как корни и листья цветковых растений. Такое сравнение, безусловно, было совсем необоснованным.
Наибольшее распространение среди ученых того времени получила теория мутуалистического симбиоза. Сторонники этой теории считали, что в слоевище лишайника гриб и водоросль находятся во взаимовыгодном симбиозе: водоросль «снабжает» гриб органическими веществами, а гриб «защищает» водоросль от чрезмерного нагревания и освещения и «обеспечивает» ее водой и неорганическими солями.
О том, что в слоевище лишайника происходит обмен веществами между грибом и водорослью, ученые стали говорить сразу после открытия двойственной природы лишайников. Однако некоторые экспериментальные подтверждения этим предположениям были получены лишь за последние три десятилетия. Применение новейших методов физиологических исследований с использованием меченых атомов углерода и азота, особых красящих веществ и некоторых других позволило установить, что гриб получает вещества, ассимилируемые водорослью, и ведет себя в слоевище лишайника как паразитический организм. Однако для существования как самого гриба, так и лишайника в целом необходимо, чтобы водоросль, окруженная со всех сторон грибными гифами, все-таки могла жить и более или менее нормально развиваться. Если гриб начнет проявлять себя слишком активно, поражать все без исключения водоросли и, использовав их содержимое, уничтожать их, это в конце концов может привести к гибели всех водорослей слоевища. Но тогда, уничтожив весь свой запас питания, погибнет и сам гриб, а значит, перестанет существовать и лишайник.
Гриб должен использовать лишь часть водорослей, оставляя резерв — здоровые и нормальные водоросли, содержимым которых он мог бы питаться.
Ученые отмечают, что степень паразитизма гриба на водоросли различна не только у разных видов лишайников, но даже в одном и том же слоевище. Резкий паразитизм обнаружен лишь у примитивных лишайников. Гаустории, проникающие глубоко внутрь протопласта водоросли, пока что были найдены лишь у наиболее просто организованных форм, в слоевище которых еще нельзя различить оформленных дифференцированных слоев. В слоевищах более высокоорганизованных лишайников часть клеток водорослей поражена грибными гифами, а остальные продолжают нормально жить и развиваться. Обычно у высокоорганизованных форм лишайников паразитизм гриба на водоросли носит весьма умеренный характер: прежде чем гриб убьет пораженные им клетки, успевает вырасти одно или несколько поколений водорослей.
Отношения между мико- и фикобионтом в слоевище лишайника не сводятся только к паразитизму гриба на водоросли. Ученые предполагают, что эти отношения гораздо сложнее. Еще в начале нашего века крупнейший русский лихенолог А.А. Еленкин, изучая анатомическое строение лишайников, обнаружил в их слоевище некральные зоны водорослей — скопления отмерших, потерявших зеленую окраску клеток, расположенные несколько ниже зоны живых водорослей. К этим бесцветным мертвым клеткам водорослей тоже тянулись грибные гифы. Это привело А. А. Еленкина к мысли, что гриб в слоевище лишайника вначале проявляет себя как паразитический организм, поражая живые клетки водоросли и используя их содержимое. Затем, убив водоросль, гриб переходит к сапрофитному способу питания, поглощая и ее мертвые остатки. Таким образом, гриб в слоевище лишайника ведет себя и как паразит, и как сапрофит. И отношения между грибом и водорослью в слоевище лишайника А.А. Еленкип назвал эндопаразитосапрофи-тизмом.
Таким образом, ученые считают, что водорослевый и грибной компоненты лишайника находятся в очень сложных взаимоотношениях. Микобионт ведет себя как паразит и сапрофит на теле водоросли, а фикобионт, в свою очередь, паразитирует на лишайниковом грибе. При этом паразитизм фикобионта всегда носит более умеренный характер, чем паразитизм гриба.
У ряда лишайников, в слоевище которых встречаются нитчатые улотриксовые водоросли, можно наблюдать еще один тип контакта. Как правило, в таком случае нити водорослей бывают целиком покрыты грибными гифами. Причем лишь иногда гифы образуют на поверхности водорослевой нити рыхлую сетку. Чаще же они располагаются очень густо и, срастаясь своими стенками, образуют сплошной чехол. Отдельная лопасть такого лишайника имеет вид тончайшего волоса. Под микроскопом она напоминает полую трубку, стенки которой образованы сросшимися грибными гифами; внутри трубки тянется нить водоросли (Жизнь растений, 1977).
1.1.4. Морфология слоевища лишайников
Слоевище лишайников очень разнообразно по окраске, размерам, форме и строению.
Лишайники окрашены в самые различные цвета: белый, розовый, ярко-желтый, оранжевый, оранжево-красный, серый, голубовато-серый, серовато-зеленый, желтовато-зеленый, оливково-коричневый, коричневый, черный и некоторые другие. Окраска слоевища лишайников зависит от наличия пигментов, которые откладываются в оболочках гиф, реже в протоплазме. Наиболее богаты пигментами гифы корового слоя лишайников и различные части их плодовых тел. У лишайников различают пять групп пигментов: зеленые, синие, фиолетовые, красные, коричневые. Механизм образования их до сих пор не выяснен, но совершенно очевидно, что важнейшим фактором, влияющим на этот процесс, является свет.
Иногда цвет слоевища зависит от окраски лишайниковых кислот, которые откладываются в виде кристаллов или зернышек на поверхности гиф. Большинство лишайниковых кислот бесцветны, но некоторые из них окрашены, и иногда очень ярко — в желтый, оранжевый, красный и другие цвета. Окраска кристаллов этих веществ определяет и окраску всего слоевища. И здесь важнейшим фактором, способствующим образованию лишайниковых веществ, является свет. Чем ярче освещение в месте произрастания лишайника, тем ярче он окрашен. Примером может служить широко распространенный, часто встречающийся на стволах осин лишайник ксантория (Xanthoria parietina). Слоевище у него яркое, красновато-оранжевое. Этот цвет придает ему особое лишайниковое вещество — париетин, которое в виде оранжевых кристаллов покрывает гифы корового слоя. Если ксантория растет на солнце, ее слоевище имеет яркую красновато-оранжевую окраску. В условиях слабого освещения этот лишайник полностью теряет оранжевый цвет и становится серовато-зеленоватым. Как правило, очень ярко окрашены лишайники высокогорий и полярных районов Арктики и Антарктики. Это тоже связано с условиями освещения. Для высокогорных и полярных районов земного шара характерны большая прозрачность атмосферы и высокая интенсивность прямой солнечной радиации, обеспечивающие здесь значительную яркость освещения. В таких условиях в наружных слоях слоевищ концентрируется большое количество пигментов и лишайниковых кислот, обусловливая яркую окраску лишайников. Предполагают, что окрашенные наружные слои защищают нижележащие клетки водорослей от чрезмерной интенсивности освещения.
Интересно, что слоевища многих антарктических лишайников окрашены в черный или другой темный цвет. Это тоже объясняется высокой концентрацией зеленых, синих и фиолетовых пигментов в коровом слое и плодовых телах этих лишайников. Хорошо пигментированные наружные слои антарктических лишайников не только защищают клетки водорослей от чрезмерно интенсивного освещения, в данном случае темная окраска слоевища является приспособительной и служит для притягивания тепловых лучей. Как известно, условия в Антарктике крайне суровы; здесь растениям приходится переносить постоянное воздействие очень низкой температуры и жить фактически без воды. Среднегодовая температура воздуха ледяного континента —16°С. Зимой она падает до —45 °С, а летом, которое наступает в январе — феврале, только днем поднимается выше нуля, ночью же снова опускается до —10 °С.
Из-за низкой температуры осадки выпадают в Антарктике только в виде снега. В такой форме они не могут быть использованы растениями. Вот здесь-то темная окраска лишайников и приходит им на помощь. Темноокрашенные слоевища антарктических лишайников за счет высокой солнечной радиации быстро нагреваются до положительной температуры даже при отрицательной температуре воздуха. Снег, падающий на эти нагретые слоевища, тает, превращаясь в воду, которую лишайник сразу же впитывает. Таким образом он обеспечивает себя водой, необходимой для осуществления процессов дыхания и фотосинтеза (Жизнь растений, 1977).
1.1.5. Анатомия слоевища лишайников
У лишайников в зависимостиот анатомического строения различают два типа слоевищ: 1) гомеомерное слоевище, когда водоросли разбросаны по всей толще слоевища; 2) гетеромерное слоевище, когда водоросли образуют в слоевище обособленный слой.
Более примитивным считается слоевище гомеомерного строения. Если рассмотреть поперечный срез такого слоевища под микроскопом, то хорошо видно, что оно образовано беспорядочными переплетениями гиф гриба, среди которых разбросаны отдельные клетки или нити водорослей. Такое строение наиболее характерно для тех лишайников, фикобионтом которых являются сине-зеленые водоросли — носток, глеокапса и некоторые другие. Эти лишайники образуют группу, известную под названием слизистых лишайников. У лишайников, которые имеют фикобионтом зеленые водоросли, слоевище гомеомерного строения встречается очень редко, лишь у наиболее примитивных накипных форм.
Слоевища слизистых лишайников в сухом состоянии имеют вид темных или даже черных твердых и хрупких корок или пленок. Но они обладают одной интересной особенностью — способностью впитывать огромное количество воды, в 20 — 30 раз превышающее их собственную массу. При увлажнении они сильно разбухают, ослизняются, приобретают характер студня или желе и становятся оливково-зелеными или грязно-зелеными. Примером может служить влажное слоевище слизистого лишайника лептогиума. Во внешнем облике этих лишайников имеется много общего с некоторыми свободноживущими водорослями.
Слизистые лишайники насчитывают небольшое число видов, всего 750, что составляет только 3% от общего числа всех известных видов лишайников. Сравнительно небольшое количество этих лишайников, по-видимому, можно объяснить более поздним вовлечением сине-зеленых водорослей в процесс образования слоевища лишайников. Но также возможно, что захват грибом сине-зеленых водорослей был менее успешным по сравнению с зелеными водорослями. В силу особенностей строения сине-зеленых водорослей, нередко собранных в колонии и обладающих плотными, богатыми слизью оболочками, эти водоросли оказались более трудным объектом для гриба при установлении с ними контакта и при построении слоевища. Безусловно, при захвате этих водорослей и их передвижении гриб испытывает гораздо больше трудностей, чем с одноклеточными и другими формами зеленых водорослей. Может быть, именно этим можно объяснить и примитивное анатомическое строение слизистых лишайников.
Для остального огромного большинства лишайников характерна гетеромерная структура, при которой в слоевищах можно различить дифференцированные слои. При этом, чем сложнее морфологическое строение слоевища лишайника, том сложнее и его анатомическая структура. У этих растений в ходе эволюции усложнение морфологического строения слоевища шло параллельно усложнению его анатомической структуры. Так, в слоевище накипных лишайников, наиболее примитивных форм, можно различить только три анатомических слоя: коровой слой, слой водорослей и сердцевину. Такое же анатомическое строение имеют и некоторые листоватые лишайники, например представители рода пельтигера. Но у большинства листоватых лишайников, более эволюционно продвинутых форм, в связи с отделением от поверхности субстрата происходит образование еще одного корового слоя — с нижней стороны слоевища. В таких слоевищах можно различить уже четыре слоя: верхний коровой слой, слой водорослей, сердцевину и нижний коровой слой. У кустистых лишайников с плоскими лентовидными лопастями образуется еще один анатомический слой — слой водорослей с нижней стороны слоевища. У этих лишайников в слоевище можно различить уже пять анатомических слоев: два коровых слоя на верхней и нижней поверхности слоевища, два слоя водорослей, тоже с верхней и нижней стороны лопастей, и сердцевину. И наконец, кустистые лишайники с округлыми лопастями, обладающие наиболее высокоорганизованными слоевищами, имеют радиальную структуру: снаружи лопасти этих лишайников покрыты коровым слоем, под ним расположен слой водорослей, а центральная часть слоевища заполнена сердцевиной.
Каждый из перечисленных анатомических слоев слоевища выполняет в жизни лишайника ту или иную функцию и в зависимости от этого имеет совершенно определенное строение.
Коровой слой играет в жизни лишайника очень важную роль. Он выполняет сразу две функции: защитную и укрепляющую.Он защищает внутренние слои слоевища от воздействия внешней среды, прежде всего водоросли от чрезмерного освещения. Поэтому коровой слой лишайников обычно бывает плотного строения и окрашен в сероватый, коричневый, оливковый, желтый, оранжевый или красноватый цвет.
Как уже отмечалось, у некоторых листоватых лишайников, имеющих вид более или менее округлой листовидной пластинки, слоевище прикрепляется лишь в своей центральной части с помощью короткой ножки — гомфа. Гомф — это также вырост нижней поверхности слоевища, и в его образовании принимают участие нижний коровой слой и сердцевина лишайника. Снаружи эта толстая ножка покрыта параплектенхимным коровым слоем, а внутри ее проходят вытянутые сердцевинные гифы. На конце гомфа, на месте прикрепления к субстрату, эти гифы имеют очень толстые темные оболочки и образуют зубцы, с помощью которых слоевище очень плотно прикрепляется к поверхности скал. Некоторые кустистые лишайники, виды родов уснея, нейропогон и др., прикрепляются к скалам с помощью псевдогомфа, также имеющего вид короткой расширенной на конце ножки. В отличие от гомфа в образовании псевдогомфа принимает участие только сердцевина лишайников. Псевдогомф образован вытянутыми, склеенными сердцевинными гифами, также образующими на конце темные зубцы, с помощью которых лишайник плотно прикрепляется к субстрату.
В зоне водорослей осуществляются процессы ассимиляции углекислоты и накопление органических веществ. Как известно, для осуществления процессов фотосинтеза водорослям необходим солнечный свет. Поэтому слой водорослей обычно размещается вблизи верхней поверхности слоевища, непосредственно под верхним коровым слоем, а у вертикально стоящих кустистых лишайников еще и над нижним коровым слоем. Слой водорослей чаще всего бывает небольшой толщины, и водоросли размещаются в нем так, что находятся почти в одинаковых условиях освещения. Водоросли в слое-вище лишайника могут образовывать непрерывный слой, но иногда гпфы микобионта делят его на отдельные участки. Для осуществления процессов ассимиляции углекислоты и дыхания водорослям необходим также нормальный газообмен. Поэтому грибные гифы в зоне водорослей не образуют плотных сплетений, а расположены рыхло на некотором расстоянии друг от друга.
Под слоем водорослей расположен сердцевинный слой. Обычно сердцевина по толщине значительно превышает коровой слой и зону водорослей. Особенно мощно она развита по сравнению с другими слоями у накипных лишайников. От степени развития сердцевины зависит толщина самого слоевища. Основная функция сердцевинногослоя — проведение воздуха к клеткам водорослей, содержащим хлорофилл. Поэтому для большинства лишайников характерно рыхлое расположение гиф в сердцевине. Воздух, попадающий в слоевище, легко проникает к водорослям по промежуткам между гифами.
У некоторых кустистых лишайников, слоевище которых далеко отстает от субстрата, сердцевинный слой, помимо проводящей функции, выполняет еще одну — укрепляющую. Как отмечалось, у большинства лишайников укрепление слоевища происходит с помощью корового слоя, который у высокоразвитых кустистых и листоватых форм постепенно становится все более толстым и плотным. Однако дальнейшее развитие в этом направлении могло бы привести к ухудшению нормальных ассимиляционных процессов: очень толстый и плотный коровой слой препятствовал бы проникновению в слоевище воздуха и света. Поэтому в процессе развития кустистых форм происходит постепенное снижение роли коревого слоя в укреплении слоевища и образование механической ткани под зоной водорослей, т. е. в сердцевинном слое (Жизнь растений, 1977).
1.2. Биоразнообразие лишайников (экологические группы)
Медленный рост слоевища не дает возможности лишайникам в более или менее благоприятных местообитаниях конкурировать с быстрорастущими цветковыми растениями или мхами. Поэтому обычно лишайники заселяют такие экологические ниши, где условия существования слишком суровы для других растений. Так, они обладают удивительной способностью расти на самых разнообразных субстратах: каменистых породах (известняках, гранитах, гнейсах, кварцах и др.), почве, коре деревьев, на хвое, листьях вечнозеленых растений, на мхах, гниющей древесине, на гниющих растительных остатках. Лишайники поселяются также на стекле, костях, коже, железе, тряпках и других предметах, при этом важно лишь одно — чтобы эти предметы пролежали неподвижно довольно длительное время.
Хотя среди лишайников и встречаются такие, которые способны расти на самых разных субстратах, все же большинство видов обладают избирательной способностью и поселяются на немногих или даже только на одном определенном субстрате.
По отношению к субстрату и другим условиям местообитания среди лишайников различают несколько основных крупных экологических групп: эпилитные лишайники — живущие на поверхности горных пород, эпифитные — растущие на коре деревьев и кустарников, эпиксильные — обитающие на гниющей древесине, эпигейные — растущие на поверхности почвы, эпифилльные — развивающиеся на хвое и листьях вечнозеленых растений, эпибриофитные — обитающие на дерновинках мхов, и другие.
В различных местообитаниях на скалах, почве, стволах деревьев и т.д. лишайники образуют растительные группировки — синузии, которые характеризуются определенным видовым составом и определенными морфологическими типами. На распределение лишайников влияют как физические, так и химические свойства субстрата. Так, среди эпилитных лишайников можно различить группу кальцефилов, поселяющихся исключительно на известняках, доломитах и других горных породах, содержащих известь, и группу кальцефобов, растущих только на поверхностях неизвестковых пород: гранитах, гнейсах, кварцах и др. Обе эти группы отличаются друг от друга видовым составом. Например, на известняках встречается довольно большое количество лишайников с эндолитными слоевищами, целиком или частично погруженными в субстрат. Это связано с хорошей растворимостью известняков: гифы лишайников, выделяя лишайниковые кислоты, легко растворяют известковые породы и проникают на довольно большую глубину — до 10 мм, а иногда даже более 30 мм. Среди кальцефобов очень мало эндолитных лишайников и почти нет видов со слоевищем, целиком погруженным в субстрат. В то же время для них характерно преобладание видов со слоевищем, имеющим форму ареолированной корочки.
Обычно на поверхности скал очень обильно развиваются накипные лишайники (ризокарпоны, лецидеи, калоплаки, леканоры, гематоммьг и др.), слоевища которых образуют на скалах пестрые яркие пятна. Но нередко вместе с ними на скалах растут и некоторые листоватые лишайники — пармелии, умбиликарии, ксантории, лептогиумы и др. В горных и арктических районах одним из очень характерных эпилитных лишайников является, например, листоватый красно-оранжевый лишайник ксантория элегантная (Xanthoria elegans). В некоторых горных районах Центральной Азии этот вид достигает такого колоссального развития, что отдельные горные массивы принимают сплошной оранжевый оттенок. Среди кустистых лишайников типичных эпилптов сравнительно немного (лишайники рода Neuropogon, некоторые виды Alectoria, Ramalina и др.).
Эпигейные лишайники чаще всего поселяются на песчаных, торфянистых и сильно щебнистых почвах, бедных питательными веществами и непригодных для произрастания других растений. Многие из этих лишайников являются ацидофильными, т.е. живущими на почве с кислой реакцией. Например, кладонии и цетрарии обычно лучше растут на кислых почвах при концентрации водородных ионов (рН) в пределах 3—7,4. Среди эпигейных лишайников много кустистых видов — кладонии, цетрарии, алектории, стереокаулона и др.. Из листоватых эпигейных лишайников наиболее широко распространены виды пельтигеры, а из накипных на почвах чаще всего встречаются представители семейства лецидеевых (Lecidea, Biatora, Bacidia, Топила и др.). В степных и пустынных областях земного шара среди напочвенных лишайников преобладают виды с чешуйчатым слоевищем (роды Psora Dermatocarpon, Endocarpon и др.).
На группировки эпигейных лишайников субстрат оказывает существенное влияние. Так, по видовому составу лишайниковые синузии песчаных почв отличаются от синузии торфянистых почв или почв, богатых известью; в значительной степени это обусловлено реакцией субстрата (рН). Например, на бедных песчаных почвах широко встречается синузия накипных лишайников Lecidea uliginosa и Baeomyces roseus; для торфянистых почв характерна другая синузия, образованная накипными лишайниками Icmadophila ericetorum и Lecidea granulosa, а для почв, богатых известью, — синузия Toninia coeruleonigricans и Fulgensia bracteata.
Но особенно сильно физические и химические свойства субстрата влияют на эпифитные лишайники. Для них большое значение имеют структура коры, ее расчленение, жесткость, частота отслаивания и другие особенности. Эпифитная лишайниковая растительность разных древесных пород, как правило, различается по составу. Например, на стволах сосен обычна синузия, образованная гипогим-нией и псевдэвернией (Hypogymnia physodes и Pseudevernia furfuracea). На дубах, липах и других лиственных породах эти лишайники обычно не встречаются, зато здесь обильно развиваются виды листоватой пармелии (особенно Parmelia sulcata, P. caperata и некоторые другие) и кустистые рамалины и эвернии (Evernia prunastri). Для стволов осин характерны группировки, образованные видами фисции, ксантории, калоплаки и др.
Видовой состав лишайниковых группировок зависит не только от вида дерева, но и от его возраста. В данном случае особенно сильно проявляется влияние физических свойств коры. Так, например, на молодых деревьях лиственных пород, имеющих гладкую тонкую кору, обычно развиваются накипные лишайники с эндофлеодным слоевищем (виды опеграфы, артонии, графиса и др.). С возрастом свойства коры меняются: она становится грубее, на ней появляются трещины и шероховатости. Меняется и состав живущих на ней лишайников. На такой коре поселяются уже листоватые и кустистые лишайники (виды пармелии, эвернии, рамалины), а из накипных — многочисленные виды леканор, лецидеи и др.
Кроме того, состав эпифитных синузий лишайников на одном и том же стволе дерева различен в зависимости от высоты над землей; в этом случае на распределение лишайников оказывают влияние не только физические особенности коры, но и экологические факторы — освещенность, влажность и некоторые другие.
Однако, несмотря на тесную зависимость, существующую между лишайниками и субстратом, на котором они поселяются, до сих пор с достоверностью еще неизвестно, используют лишайники субстрат только как место прикрепления или они извлекают из него некоторые питательные вещества, необходимые для их жизнедеятельности. С одной стороны, способность лишайников расти на субстратах, бедных питательными веществами, дает основание считать, что они используют субстрат лишь как место прикрепления. Однако, с другой стороны, избирательная способность, проявляемая лишайниками при расселении, строгая приуроченность большинства из них к определенному субстрату, зависимость видового состава лишайниковой растительности не только от физических, но и от химических свойств субстрата невольно наводят на мысль, что лишайники используют субстрат и как дополнительный источник питания. Это подтверждается и биохимическими исследованиями, проведенными в последние годы. Например, выяснилось, что у одного и того же вида лишайника, растущего на разных древесных породах, состав лишайниковых веществ может быть неодинаковым. Еще более очевидным доказательством служит открытие у лишайников внеклеточных ферментов, которые выделяются во внешнюю среду. Внеклеточные ферменты, такие, например, как инвертаза, амилаза, целлюлаза и многие другие, представлены в лишайниках довольно широко и обладают достаточно высокой активностью. Причем, как оказалось, они наиболее активны в нижней части слоевища, которой лишайник прикреплен к субстрату. Это указывает на возможность активного воздействия слоевища лишайников на субстрат с целью извлечения из него дополнительных питательных веществ.
Помимо свойств субстрата, на расселение лишайников большое влияние оказывают также климатические факторы — свет, влажность, температура.
Насколько разнообразны слоевища лишайников по окраске, настолько же разнообразны они и по форме. Слоевище может иметь вид корочки, листовидной пластинки или кустика. В зависимости от внешнего облика различают три основных морфологических типа: накипные, листоватые и кустистые лишайники (Шапиро, 1991).
1.3. Роль микроорганизмов в лишайниковом симбиозе
Рассматривая микроорганизмы в качестве сожителей лишайников более или менее приспособленных к узко специфическим условиям существования, Красильников отмечает две стороны их деятельности. С одной стороны, несомненна их роль в почвообразовании и подготовке субстрата для развития данной группы низших растений, а с другой — участие в различных процессах, протекающих в лишайниках как в самостоятельной экосистеме, синтезе и деструкции веществ.
Размеры бактерий позволяют им проникать в мельчайшие поры бесплодных каменистых субстратов. В результате своей жизнедеятельности они способствуют разрушению и выветриванию таких горных пород, как гранит, базальт, туф и др. Приспособленные к существованию на бедных субстратах, в которых отсутствуют сложные органические вещества и ограничено количество минерального азота, бактерии, по мнению Красильникова, являются первыми поселенцами скальных пород. Они, как и свободноживущие водоросли, подготавливают почву для последующего развития низших растений, в частности лишайников. Наличие среди бактерий большого числа азотфиксаторов (преимущественно олигонитрофилов) играет существенную роль в обогащении природных субстратов азотом. Учитывая, что в лишайниках обитают различные группы микроорганизмов, можно предположить, что и функции, выполняемые бактериями в лишайниках, также разнообразны. Известно, например, что развитие водорослей и грибов влечет за собой накопление в среде органических веществ,что является как результатом прижизненных выделений клеток, так и следствием их автолиза. Сопутствующие бактерии, несомненно, используют органические вещества, выделяемые грибами и водорослями. Можно предположить, что выделения водорослей и грибов изменяются в зависимости не только от вида лишайника, но и от различных экологических условий. Эти различия в свою очередь вероятно, приводят к изменениям количественного и качественного составов сопутствующих микроорганизмов. Однако основные принципы участия всех организмов в жизнедеятельности данной системы, по-видимому, остаются общими.
Среди микроорганизмов, обитающих в слоевище лишайников, особый интерес представляют олигонитрофильные бактерии, которые связывают атмосферный азот и тем самым регулируют азотное питание лишайников. Кроме того, как показано работами Н.И. Мальцевой и Н.А. Карпушина (1969) и Мальцевой и др. (1977), олигонитрофильные микроорганизмы продуцируют внеклеточные слизистые вещества полисахаридной природы. Слизи микробного происхождения, по некоторым литературным данным, могут использоваться другими микроорганизмами, в частности актиномицетами, нокардиями и плесневыми грибами в реакциях энергетического и конструктивного обмена. В то же время известно, что олигонитрофилы, входящие в состав микробных ассоциаций, снабжают сопутствующую микрофлору биологически активными веществами, органическими кислотами и азотом, фиксированным из воздуха. Считают, что одной из причин возникновения тесных взаимоотношений, складывающихся в почве между олигонитрофилами и другими микроорганизмами, и образования прочных микробных ассоциаций являются внеклеточные слизи олигонитрофилов, которые используются как самими продуцентами, так и сопутствующими микроорганизмами в качестве источника энергии. Возможно, этот фактор объясняет отсутствие в чистых культурах роста некоторых микроорганизмов из группы «карликовых», которые могут развиваться только при смешанных посевах (Добровольская, Соколов, 1962).
Таким образом, внеклеточные слизистые вещества, выделяемые олигонитрофилами, вероятно, можно рассматривать как важный экологический фактор формирования микробных ассоциаций и жизнедеятельности отдельных микроорганизмов в лишайниках. Продуцируя биотические вещества, некоторые микроорганизмы стимулируют рост ряда гетероауксотрофных бактерий, грибов и дрожжей (Красильников, 1949).
Пигментные формы бактерий и грибов выполняют определенную защитную функцию в лишайниках. Вырабатывая каротиноидные пигменты, они защищают лишайник от действия ультрафиолетовых лучей. Красильников отмечает, что пигментные формы микроорганизмов хорошо приспособлены к условиям очень низкой влажности, поэтому устойчивы к высушиванию.
Целлюлозоразрушающие микроорганизмы, разрушают оболочки отмерших водорослей, чем обеспечивают круговорот углерода в лишайниках как своеобразной экосистеме. Образующиеся при расщеплении целлюлозы углеводы могут быть использованы микобионтом и другими микроорганизмами. Хитин — основной компонент клеточной стенки микобионта лишайника — может служить хорошим субстратом для развития актин омицетов (Шлегель, 1972).
Накопленные к настоящему времени литературные данные свидетельствуют о наличии многочисленных и разнообразных групп микроорганизмов, связанных сложными метаболическими взаимоотношениями как с симбионтами лишайников, так и между собой. Однако имеющихся данных еще недостаточно для определения взаимосвязи и взаимовлияния симбионтов лишайников и обитающих в них бактерий. Для установления и определения этих связей большое значение может иметь ресинтез лишайников в стерильных условиях. По-видимому, в естественно складывающемся лишайниковом сообществе устанавливаются определенные типы взаимоотношений между бактериями и симбионтами лишайника. Очевидно, какие-то физиологические процессы жизнедеятельности лишайников влияют на выживаемость определенных видов бактерий и способствуют развитию форм, приспособленных к совместному существованию.
Таким образом, более глубокое изучение микроорганизмов, обитающих в лишайниках и продуктов метаболизма как симбионтов лишайников, так и сопутствующих им микроорганизмов позволит понять и изучить характер взаимосвязей и взаимозависимостей, обеспечивающих формирование и существование такого уникального биоценоза как лишайник (Красильников, 1949).
1.4. Хозяйственное значение лишайников
Будучи одним из основных напочвенных растений тундровой зоны, некоторые лишайники (кладонии и цетрарии) являются главным кормовым растением для северных оленей. Олени обладают способностью чувствовать запах лишайников и сквозь снеговой покров, из-под которого они их достают, разгребая снег копытами. При кормежке олени скусывают лишь верхушки кустиков кладоний и цетрарии, и лишайники продолжают свой рост. Однако растут они медленно, и для восстановления пастбищ требуется от 10 до 30 лет. Поэтому в оленеводческих хозяйствах Севера ведется плановое использование лишайниковых пастбищ с учетом их возобновления. Лишайники используют в пищу и другие животные; например, свиньи и овцы охотно поедают кладонию листоватую, кладонию оленевидную и др.
В Японии употребляется в пищу и даже служит предметом экспорта в страны Юго-Восточной Азии растущий на скалах листоватый лишайник гирофора съедобная (Gyrophora esculenta). Съедобен и лишайник аспицилия съедобная (Aspicilia esculenta), растущий в степной и полупустынной зонах. Он содержит до 55-65% щавелевокислого кальция. Однако усвояемость питательных веществ лишайников очень незначительна.
В результате взаимодействия гриба и водоросли, составляющих организм лишайника, образуются специфические вещества, которые в природе нигде больше не встречаются. Это так называемые лишайниковые вещества, или лишайниковые кислоты.
Некоторые из этих кислот обладают антибиотическим действием, например усниновая кислота, образуемая 70 видами лишайников. Это сильный антибиотик, который под названием «бинан» введен в медицинскую практику для лечения некоторых заболеваний и рекомендован к применению в ветеринарии.
Некоторые лишайниковые вещества действуют как стимуляторы, поднимающие тонус организма. На этом основано использование в народной медицине отваров цетрарии исландской («исландский мох»). В ее состав входит паралихестериновая кислота, обладающая тонизирующим действием.Из широко распространенного лишайника эверния сливовая («дубовый мох») извлечено вещество резиноид, обладающее ароматическими свойствами и являющееся хорошим закрепителем аромата. Этот препарат заменяет соответствующее импортное сырье для парфюмерной промышленности. К духам, в состав которых входит резиноид, относятся «Шипр», «Кристалл», «Кремль», «Кармен», «Маска», «Свежее сено» и др. Этот же лишайник и реже другой вид — эверния шелушащаяся — применялись в странах Северной Африки для ароматизации хлеба.
Некоторые лишайники из рода рочелла, растущие на морских побережьях, а также охролехия виннокаменная, растущая на скалах и на почве в северных районах России, применяются местным населением как красители.
В наши дни, когда актуальнейшей проблемой стала борьба с загрязнением окружающей среды, лишайники могут сослужить человеку еще одну службу. Многие виды лишайников — хорошие индикаторы степени загрязненности воздуха. Вблизи больших промышленных городов они растут плохо и постепенно вымирают. Так, очень чувствительны к загрязнению воздуха накипные лишайники охролехия двуполая и леканора выпуклоплодная, растущие обычно на коре деревьев и обнаженной древесине. Разработаны шкалы и простые математические формулы для определения степени загрязненности воздуха на основе наличия или отсутствия определенных лишайниковых группировок. Поэтому вместе с врачами-гигиенистами и химиками, занимающимися оценкой чистоты воздуха в городах и промышленных районах, теперь часто работают и лихенологи.
В перспективе при широком и углубленном изучении лишайники могут стать источниками ценных биологически активных веществ (медицинских препаратов и т. д.). Однако использование лишайников должно вестись планомерно, на заранее разработанной научной основе, чтобы не нанести непоправимый ущерб природе, в которой они играют свою определенную важную роль (Шапиро, 1991).
2. ЛИШАЙНИКИ КАК ИНДИКАТОРЫ ЗАГРЯЗНЕНИЯ ОКРУЖАЮЩЕЙ СРЕДЫ
Проблема загрязнения природной среды — одна из глобальных проблем современного мира. В связи с интенсивным развитием промышленности и транспорта в атмосферу, гидросферу, литосферу поступает все большее количество вредных выбросов. На земном шаре практически невозможно найти место, где бы ни присутствовали, в той или иной концентрации, загрязняющие вещества (поллютанты).
Среди веществ, загрязняющих воздух, наибольшее значение имеет сернистый газ, галогены и их соединения, оксид углерода, сероводород, аммиак, этилен. А также копоть, пепел, твердые частицы пыли (цемента, извести, кремния, каменного угля, металлов и их соединений).
Благодаря уникальным свойствам лишайников их стали использовать для общей оценки степени загрязненности атмосферного воздуха. На этой основе стало развиваться особое направление индикационной экологии — лихеноиндикация (Шкараба, Селиванов, 2001).
Как и большинство биологических методов оценки состояния окружающей среды, метод лихеноиндикации не позволяет различить конкретные вредные вещества, загрязняющие атмосферный воздух, но зато позволяет выделить территории, подверженные воздействию загрязненного воздуха. Для этого иногда бывает достаточно даже неполного описания разнообразия и обилия лишайников на единице площади в данном массиве (http://www.eco.nw.ru).
Общие изменения структуры лишайниковых сообществ под воздействием загрязнения проявляются в уменьшении числа видов и обилия чувствительных видов, смене субстратов и увеличении обилия устойчивых к загрязнению видов, изменение спектра жизненных форм (уменьшение доли кустистых и, в меньшей степени, листоватых лишайников). В основе этих изменений лежит дифференциальная чувствительность различных видов к воздействию поллютантов (Голубкова, Малышева, 1978).
Чувствительность лишайников к загрязнению обусловлена несколькими причинами:
-
лишайники представляют собой симбиоз гриба и водорослей и любое, даже не значительное, влияние может изменять баланс взаимодействия между симбионтами, что сказывается на их жизнеспособности;
-
лишайники поглощают аэрозоли и газы всей поверхностью талломов, а также периодически подвергаются дегидратации талломов (обезвоживанию), что приводит к росту концентрации загрязняющих веществ в талломах до высоких уровней;
-
водоросль требуксия (Trebuxia), входящая в состав 80% видов лишайников, обладает высокой чувствительностью к повышенным концентрациям сернистого газа в атмосфере;
-
четкая зависимость лишайников от величины кислотности субстрата (рН среды), поллютанты могут изменять значения рН в ту или иную строну, и эти значения могут выходить за пределы выносливости одних видов и поселению на данном субстрате других (Инсарова, Инсаров, 1989).
Установлено, что наиболее удобными для изучения являются эпифитные лишайники (обитающие на стволах и ветвях деревьев). Это связано с тем, что стволы деревьев подвергаются более сильной циркуляции воздуха в течении всего года, чем напочвенная растительность. К тому же все необходимые вещества эпифиты получают только из атмосферы, а субстрат служит им только местом для прикрепления. Эпифиты удобны для изучения еще и потому, что существуют в более-менее однородных условиях местообитания, тогда как напочвенные и эпилитные лишайники могут обитать на целой мозаике из различных микроусловий и их распространение может в большей степени зависеть от случайных факторов, а не от загрязнения (http://fadr.msu.ru/ecocoop/monitor).
Также установлено, что при повышении степени загрязнения воздуха первыми исчезают кустистые, затем листоватые и последними накипные (корковые) формы лишайников (Шкараба, Селиванов, 2001).
3. МЕСТО И МЕТОДИКА ИССЛЕДОВАНИЯ
Исследования были проведены на территории пгт Алексеевка. Были выбраны два района исследований, которые на наш взгляд коардинально отличаются по степени загрязнения атмосферного воздуха:
1. Деревья, растущие вдоль проезжей части по улицы в районе учебного корпуса ГБОУ СОШ №4.
2. Участок древостоя в парке (ближе к центру парка) на расстоянии приблизительно 40-50м пгт Алексеевка
В основу методики оценки относительной численности эпифитных лишайников был положен метод линейных пересечений, который заключается в наложении гибкой ленты с миллиметровыми делениями на поверхность ствола дерева с фиксированием всех пересечений её со слоевищами лишайников. В качестве ленты использовался «портняжный метр» с миллиметровыми делениями.
После выбора дерева определили на стволе точку, находящуюся на высоте 1,5 метра от основания ствола с северной стороны. Затем на ствол наложили мерную ленту с делениями таким образом, чтобы ноль шкалы ленты совпадал с выбранной точкой, а возрастание чисел на шкале соответствовало движению по часовой стрелке (с севера на восток). После полного оборота ствола лента закрепляется на стволе булавкой в нулевой точке. Совмещая последнее деление и ноль ленты, определили длину окружности ствола.
При измерении отмечали начало и конец каждого пересечения ленты с талломами лишайников. Измерения проводили с точностью до 1 мм.
По завершении измерений провели расчёт проективного покрытия лишайников на основе линейных пересечений, который определяет отношение «заросшей» лишайниками части ствола к общей поверхности. Зная общую длину окружности ствола и принимая её за 100%, рассчитали проективное покрытие лишайников (Малышева, 1996).
Проективное покрытие определяется для всех видов лишайников в сумме.
Видовую принадлежность лишайников определили в лабораторных условиях по определителю.
По чувствительности к атмосферным загрязнителям лишайники делят на среднечувствительные и высокочувствительные. К первой группе относят, например, некоторые виды пармелий (бороздчатую, скальную) и кладоний (порошистую, бахромчатую). Высокой чувствительностью отличаются уснеи (хохлатая, пышная), цетрария сизая,кладония неприглаженная, ксантория настенная (золотянка).
При больших концентрациях часто встречающегося загрязнителя – двуокиси серы – лишайники преждевременно стареют. Так называют состояние организма, когда слоевище нарастает по краям, а сердцевинные его части отстают от субстрата и выпадают. И тогда колонии листоватых или кустистых лишайников приобретают очертания полумесяца.
Есть и другой, не менее заметный показатель качества окружающей среды. По мере приближения к источнику загрязнения слоевища становятся более толстыми, компактными.
4. РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЙ
Первой была обследована территория в районе учебного корпуса МОУ СОШ №4:
-
Длина окружности первого ствола из обследуемых (ясень) – 28 см.
Пересечение ленты с талломами наблюдались на отметках: 2,0 – 4,3; 5,5 – 6,6; 14,0 – 14,7.
-
Длина окружности второго ствола (клен американский) – 29 см.
Пересечение ленты с талломами наблюдались на отметках: 2,4 – 9,7; 17,7 – 18,2.
-
Длина окружности третьего ствола (береза повислая) – 22 см.
Пересечение ленты с талломами наблюдались на отметках: 3,0 – 9,0; 10,1 – 11,3; 11,7 – 12,7; 13,0 – 15,5; 15,6 – 18,0; 19,3 – 22,0.
-
Длина окружности четвертого ствола (вяз мелколистный) – 31 см.
Пересечение ленты с талломами наблюдались на отметках: 2,6 – 4,1; 5,3 – 6,1; 12,2 – 14,9.
-
Длина окружности пятого ствола (вяз мелколистный) – 29 см.
Пересечение ленты с талломами наблюдались на отметках: 2,2 – 4,0; 6,3 – 8,1; 11,9 – 15,3.
Общая сумма «протяжённости» лишайников на каждом дереве составила:
1 дерево: 4,1 см (1,1 + 2,3 + 0,7)
2 дерево: 7,8 см (7,3 + 0,5)
3 дерево: 2,2 см (6,0 + 1,2 + 1,0 + 2,5 + 2,4 + 2,7)
4 дерево: 5,0 см (1,5 + 0,8 + 2,7)
5 дерево: 7,0 см (1,8 + 1,8 + 3,4)
Находим величину проективного покрытия лишайников
-
-
4,1: 28 х 100 = 14,64%
-
7,8 : 29 х 100 = 26,9%
-
2,2 : 22 х 100 = 10%
-
5,0 : 31 х 100 = 16,1%
-
7,0 : 29 х 100 = 24,1%
-
Большого видового разнообразия лишайников на данной территории не обнаружено, нами определены на обследуемых стволах следующие виды: Ксантория настенная (Xanthoria parietina) и Пармелия бороздчатая (Parmelia sulcfta).
В парковой зоне для достоверности сравнения результатов нами были выбраны те же древесные породы и с приблизительно такой же окружностью ствола.
Величина проективного покрытия лишайников в данном случае была следующей
-
-
7,1: 28,5 х 100 = 24,9%
-
11,3 : 28,5 х 100 = 39,6%
-
8,6 : 23 х 100 = 37,4%
-
8,1 : 32 х 100 = 25,3%
-
10,4 : 28 х 100 = 37,1%
-
И видовое разнообразие лишайников в парке несколько больше: кроме указанных для первого участка видов нами так же были зарегистрированы еще 2 вида: Ксантория многоплодная (Xanthoria polycarpa) и Псора ступенчатая (Psora scalaris).
Таким образом, сравнивая площади проективного покрытия лишайников на деревьях одних и тех же пород и с приблизительно равными окружностями ствола на двух обследуемых территориях, мы можем сделать вывод, что состояние атмосферного воздуха в парковой зоне более благополучно сточки зрения чистоты, так как и количество, и видовое разнообразие лишайников, в этой зоне больше.
Так же хочется отметить, что данная методика довольно проста в использовании и может с успехом применяться для исследовательских работ школьников.
ВЫВОДЫ:
Чем больше индустриализирован будет поселок, чем сильнее загрязнен воздух, тем меньше встречается в нем видов лишайников и ниже их жизнеспособность; при повышении степени загрязненности воздуха первыми исчезают кустистые лишайники, за ними — листоватые и последними — накипные.
При изучении лишайников в пгт Алексеевка было определено: кустистых лишайников на исследуемых участках не было обнаружено, значит загрязнение уже есть и достаточно высокой степени.
Интересным фактом является тот, что накипных лишайников очень мало, особенно на улице вблизи проезжей части. Листоватые лишайники встречаются. Было обнаружено 4 вида этих лишайников. Больше их в количественном отношении нами зарегистрировано в парковой зоне и меньше на улице с большим движением транспорта. Значит, интенсивность движения сказывается на численности лишайников.
Выбранный нами район пгт Алексеевка в экологических сводках характеризуется средней загрязненностью. Для лишайников это «зона угнетения». Чем сильнее загрязнен воздух, тем меньшую площадь покрывают лишайники на стволах деревьев.
СПИСОК ЛИТЕРАТУРЫ
-
Бирюкова А. Определение чистоты воздуха по лишайникам // http://fadr.msu.ru/ecocoop/monitor
-
Верясова Ю, Пареева М. Биоиндикация загрязненности воздуха // http://www.eco.nw.ru
-
Голубкова Н.С., Малышева Н.В. Влияние роста города на лишайники и лихеноиндикация атмосферных загрязнений г. Казани // Ботан. журн., 1978. Т. 63, № 8. C. 1145-1152.
-
Добровольская Т.Г., Соколов А.А. Азотфиксирующие бактерии в лишайниках. // // Бюлл. МОИП. Отдел. микробиологии. Серия 81(1). М.: АН РСФСР, 1962. С. 124-125.
-
Жизнь растений в 6. т. Т. 3. Водоросли. Лишайники. М.: Просвещение, 1977. С. 379-381, 382-385, 490-419, 420-423, 426-431.
-
Инсарова И.Д., Инсаров Г.Э. Сравнительные оценки чувствительности эпифитных лишайников различных видов к загрязнению воздуха // Проблемы экологического мониторинга и моделирования экосистем. Л.: Гидрометеоиздат, 1989. Т. 12. C. 113-175.
-
Красильников Н.А. Микрофлора лишайников. // Бюлл. МОИП. Отдел. микробиологии. Серия 81(1). М.: АН РСФСР, 1949. С. 84-86.
-
Лишайники как индикаторы чистоты воздуха // http://miksike.net/docs
-
Малышева Н.В. Биоразнообразие лишайников и оценка экологического состояния парковых ландшафтов с помощью лишайников (на примере парков окрестностей Санкт-Петербурга) // Новости систематики низших растений. СПб., 1996. Т. 31. С. 135-137.
-
Петров В.В., Абрамова Л.И. Общая ботаника с основами геоботаники. М.: Высшая школа, 1994. С. 142-147.
-
Перцева Е.В., Биоиндикация экосистем: методические указания для проведения лабораторных занятий.-Кинель, РИЦ СГСХА.с.21-22.
-
Шапиро И.А. Загадки растения сфинкса. Лишайники и экологический мониторинг. Л.: Гидрометеоиздат, 1991. 80 с.
-
Шкараба Е.М., Селиванов А.Е. Использование лишайников в качестве индикаторов загрязнения окружающей среды: Учебное пособие. — Пермь. Изд. ПГПУ, 2001
-
Шлегель Г.М. Общая микробиология. М.: Мир, 1972. С. 211-218.
Просмотров работы: 360
Цетрария и Кладония применение в фитодизайне
Все мы любим бывать в лесу, гулять, собирать грибы или ягоды, вдыхать чистый и вкусный, пьянящий аромат, слушать пение птиц, наслаждаться видами и затейливыми узорами ковров из мхов и лишайников.
Вот о двух повсеместно распространённых лишайниках и пойдёт речь. Это Кладония (ягель) и Цетрария («Исландский мох»).
Несомненно, вы все видели их в лесу, преимущественно в сухих сосновых борах. Многие думают, что это мхи, но это не так, это лишайники — удивительные живые организмы, представляющие из себя симбиоз представителей двух разных царств живого мира – грибов и одноклеточных синезелёных водорослей.
Тело (слоевище) лишайника состоит из грибных гиф, а водоросли синтезируют в процессе фотосинтеза органические вещества, обеспечивая лишайник питательными веществами.
Лишайники очень устойчивы во внешней среде. Для жизни кладонии и цетрарии не важны ни качество почвы, ни особо благоприятные погодные условия, они терпимы к экстремальным условиям (засухе, высоким и низким температурам, ультрафиолетовому излучению). Почва служит для них только в качестве места прикрепления (субстрата). Именно поэтому кладонии заселяют неплодородные почвы сухих сосновых боров. Едва попадется чуть более плодородный участок, места лишайникам уже не остается. Здесь уже растут мхи, плауны, травы.
Не только почва служит субстратом для лишайников. Некоторые виды охотно заселяют нижнюю часть ствола деревьев, селятся на пнях, на валежнике. Они могут жить на открытых незащищённых поверхностях, таких как камни, кора деревьев, бетон и даже ржавеющий металл. Небольшое количество минеральных веществ, необходимое для жизнедеятельности, лишайники получают большей частью из пыли в воздухе или с дождевой водой.
Растут лишайники очень медленно, всего несколько миллиметров в год.
Поверхность лишайника может удерживать воду на короткое время, в сухих условиях вода быстро теряется и лишайник переходит в фотосинтетически неактивное состояние, при котором вода может составлять не более 10% массы. Время, которое лишайник может провести высушенным, зависит от вида, известны случаи «воскрешения» после 40 лет в сухом состоянии. Когда поступает пресная вода в форме дождя, росы или влажности, лишайники быстро переходят в активное состояние, возобновляя метаболизм.
Эти лишайники прекрасно живут в условиях квартир при регулярном опрыскивании. Они абсолютно безвредны для человека и домашних животных. Более того даже полезны, т.к. с давних пор используются как сырьё и лечебное средство в фармацевтике и народной медицине. А также являются незаменимым кормом для северных оленей.
Идеи для применения.
Кладонии и цетрарии относятся к кустистым лишайникам, они красивы и грациозны.
Именно поэтому, лишайники этих двух родов – это прекрасный материал для различного фитодизайна. Их можно использовать в сухих открытых флорариумах, они прекрасно сочетаются с суккулентами. Можно делать различные природные композиции, сочетая с корягами и камнями, фитопанно. Детям можно использовать для поделок, создания игровых ландшафтов и диарам.
Несколько слов о сборе, обработке и уходе в домашних условиях.
Отправляться за Цетрарией и Кладонией нужно в сухой, преимущественно сосновый лес.
Лучше взять несколько, необходимых вам кустиков из места с богатым лишайниковым покровом. Отделить от субстрата, потрясти от мусора. Дома погрузить в воду на несколько минут. Это избавит от мелких насекомых, позволит напитаться слоевищу влагой и легче перенести адаптацию. При этом лишайник становится мягким, цетрария становится зелёной и «мясистой». Начинает источать приятный свежий грибной аромат.
Затем можно разместить на подходящем субстрате, например, какой-нибудь живописной коряге или коре. Далее обильно опрыскивать водой 2-3 раза в неделю, но периодичность опрыскивания может быть и более редкой.
Эти лишайники – это прекрасный вариант соприкоснуться с кусочком дикой природы для городских людей, у которых не хватает времени и желания ухаживать за требующими более сложного ухода домашними растениями.
Купить Цетрарию и Кладонию можно в нашем интернет-магазине Gu-Ga.ru
Лишайников | Микробиология
Цели обучения
- Объясните, почему лишайники включены в исследование микробиологии
- Опишите уникальные характеристики лишайника и роль каждого партнера в симбиотических отношениях лишайника
- Опишите, чем лишайники полезны для окружающей среды
Никто не должен беспокоиться о том, чтобы заболеть лишайниковой инфекцией, но лишайники интересны с микробиологической точки зрения и являются важным компонентом большинства наземных экосистем.Лишайники предоставляют возможности для изучения тесных взаимоотношений между неродственными микроорганизмами. Лишайники способствуют производству почвы, разрушая горные породы, и они являются первыми колонизаторами в беспочвенных средах, таких как потоки лавы. Цианобактерии в некоторых лишайниках могут связывать азот и действовать как источник азота в некоторых средах. Лишайники также являются важными стабилизаторами почвы в некоторых пустынных условиях и являются важным источником зимней пищи для карибу и северных оленей. Наконец, лишайники производят соединения, обладающие антибактериальным действием, и дальнейшие исследования могут обнаружить соединения, которые с медицинской точки зрения полезны для человека.
Характеристики
A лишайник представляет собой комбинацию двух организмов, зеленой водоросли или цианобактерии и грибка аскомицета, живущих в симбиотических отношениях. В то время как водоросли обычно растут только в водной или чрезвычайно влажной среде, лишайники потенциально могут быть найдены практически на любой поверхности (особенно на скалах) или в виде эпифитов (что означает, что они растут на других растениях).
Рис. 1. На этом поперечном срезе слоевища лишайника показаны его различные компоненты.Верхняя кора грибковых гиф обеспечивает защиту. Фотосинтез происходит в зоне водорослей. Мозговое вещество состоит из гиф гриба. Нижняя кора также обеспечивает защиту. Корневища прикрепляют слоевище к субстрату.
В некотором смысле симбиотические отношения между лишайниками и водорослями кажутся мутуализмом (отношения, в которых выигрывают оба организма). Гриб может получать фотосинтаты из водорослей или цианобактерий, а водоросли или цианобактерии могут расти в более сухой среде, чем они могли бы в противном случае.Однако большинство ученых считают эти симбиотические отношения контролируемым паразитизмом (отношения, при которых один организм получает выгоду, а другой страдает), потому что фотосинтезирующий организм растет хуже, чем без гриба. Важно отметить, что такие симбиотические взаимодействия являются континуумом между конфликтом и сотрудничеством.
Лишайники медленно растут и могут жить веками. Их использовали в пищевых продуктах и для извлечения химических веществ, таких как красители или противомикробные вещества.Некоторые из них очень чувствительны к загрязнению и используются в качестве индикаторов окружающей среды.
Лишайникиимеют тело, называемое слоевищем, внешний плотно упакованный грибковый слой, называемый кортикального мозга , и внутренний, слабо упакованный грибковый слой, называемый мозговым веществом (рис. 1). Лишайники используют пучки гиф, называемые ризинами , , для прикрепления к субстрату.
Разнообразие лишайников
Лишайники классифицируются как грибы, а их партнеры по грибам принадлежат к Ascomycota и Basidiomycota.Лишайники также могут быть сгруппированы по типам в зависимости от их морфологии. Есть три основных типа лишайников, хотя существуют и другие типы. Лишайники, которые плотно прикреплены к субстрату и придают им твердый вид, называются корковыми лишайниками . Листообразные лепестки имеют листовые лишайники ; они могут быть прикреплены только в одной точке формы роста, и они также имеют вторую кору ниже мозгового вещества. Наконец, кустистые лишайники имеют округлую структуру и в целом разветвленный вид.На рисунке 2 показан пример каждой из форм лишайников.
Рис. 2. Здесь показаны примеры трех типов лишайников. (а) Это корковый лишайник, встречающийся в основном на морских породах, Caloplaca marina . (б) Это листовой лишайник, Flavoparmelia caperata . (c) Это кустистый лишай, Letharia vulpina, который настолько ядовит, что когда-то из него делали наконечники стрел. (кредит b, c: модификация работы Джейсона Холлингера)
Подумай об этом
- Какие типы организмов встречаются в лишайниках?
- Какие три формы роста лишайников?
Клиническая направленность: Энтони, Разрешение
Этот пример продолжает историю Энтони, которая началась с одноклеточных эукариотических паразитов, паразитических гельминтов и грибов.
Мать Энтони спрашивает врача, что ей делать, если прописанный Энтони крем от стригущего лишая не работает. Врач объясняет, что стригущий лишай — это общий термин для обозначения состояния, вызванного несколькими видами. Первым делом нужно сдать соскоб для исследования под микроскопом, что врач уже сделал. Он объясняет, что определил инфекцию как грибок и что противогрибковый крем действует против наиболее распространенных грибков, связанных со стригущим лишаем.Однако крем может не действовать против некоторых видов грибка. Если через пару недель крем не подействует, Энтони должен прийти еще раз, и врач примет меры, чтобы определить вид грибка.
Положительная идентификация дерматофитов требует культивирования. Для этой цели можно использовать агар Сабуро . В случае инфекции Энтони, которая исчезла в течение 2 недель лечения, культура имела бы зернистую текстуру и выглядела бы бледно-розовой сверху и красной снизу.Эти особенности предполагают, что гриб Trichophyton rubrum , частая причина стригущего лишая.
Ключевые концепции и резюме
- Лишайники представляют собой симбиотическую ассоциацию между грибами и водорослями или цианобактериями
- Симбиотическая ассоциация, обнаруженная в лишайниках, в настоящее время считается контролируемым паразитизмом , при котором гриб приносит пользу, а водоросли или цианобактерии причиняют вред
- Лишайники медленно растут и могут веками жить в самых разных средах обитания
- Лишайники важны для окружающей среды, они помогают создавать почву, дают пищу и служат индикаторами загрязнения воздуха
Множественный выбор
Вы встретили лишайник с листвой.Каким термином описывается этот лишай?
- крастозе
- лист
- фрутикоза
- агароза
Ответ б. Этот лишай — листовой.
Что из нижеперечисленного обозначает внешний слой лишайника?
- кора
- мозгового вещества
- слоевище
- theca
Ответ а. Кора — это внешний слой лишайника.
Что из перечисленного является грибком в лишайнике?
- базидиомицет
- аскомицет
- зигомицет
- апикомплекс
Ответ б.Грибок при лишайнике — аскомицет.
Подумай об этом
Какими тремя способами лишайники имеют экологическую ценность?
Характеристики грибов | Безграничная биология
Характеристики грибов
Грибы, латинское слово «гриб», — это эукариоты, которые отвечают за разложение и круговорот питательных веществ в окружающей среде.
Цели обучения
Опишите роль грибов в экосистеме
Ключевые выводы
Ключевые моменты
- Грибы более тесно связаны с животными, чем с растениями.
- Грибы гетеротрофны: они используют сложные органические соединения в качестве источников энергии и углерода, а не фотосинтеза.
- Грибки размножаются бесполым, половым или обоими способами.
- Большинство грибов продуцируют споры, которые определяются как гаплоидные клетки, которые могут подвергаться митозу с образованием многоклеточных гаплоидных особей.
- Грибы взаимодействуют с другими организмами, образуя полезные или мутуалистические ассоциации (микоризы и лишайники) или вызывая серьезные инфекции.
Ключевые термины
- микориза : симбиотическая ассоциация между грибком и корнями сосудистого растения
- спора : репродуктивная частица, обычно одна клетка, выделяемая грибком, водорослью или растением, которая может прорасти в другую
- лишайник : любой из многих симбиотических организмов, являющихся ассоциациями грибов и водорослей; часто встречается в виде белых или желтых пятен на старых стенах и т. д.
- Ascomycota : таксономическое подразделение в королевстве Fungi; те грибы, которые производят споры в микроскопическом спорангии, называемом аск
- гетеротрофные : организмы, использующие сложные органические соединения в качестве источников энергии и углерода
Знакомство с грибами
Слово «гриб» происходит от латинского слова «грибы».Действительно, знакомый гриб — это репродуктивная структура, используемая многими видами грибов. Однако есть также много видов грибов, которые вообще не производят грибов. Являясь эукариотами, типичная грибковая клетка содержит истинное ядро и множество мембраносвязанных органелл. Королевство грибов включает в себя огромное разнообразие живых организмов, вместе называемых Ascomycota, или настоящие грибы. Хотя ученые идентифицировали около 100 000 видов грибов, это лишь часть из 1,5 миллиона видов грибов, которые, вероятно, присутствуют на Земле.Съедобные грибы, дрожжи, черная плесень и производитель антибиотика пенициллина Penicillium notatum — все они являются членами королевства Fungi, которое принадлежит домену Eukarya.
Примеры грибов : Многие виды грибов производят знакомый гриб (а), который является репродуктивной структурой. Этот (б) коралловый гриб демонстрирует ярко окрашенные плодовые тела. Эта электронная микрофотография показывает (c) споровые структуры Aspergillus, типа токсичных грибов, встречающихся в основном в почве и растениях.
Грибы, которые когда-то считались растительными организмами, более тесно связаны с животными, чем с растениями. Грибы не способны к фотосинтезу: они гетеротрофны, потому что используют сложные органические соединения в качестве источников энергии и углерода. Некоторые грибковые организмы размножаются только бесполым путем, тогда как другие подвергаются как бесполому, так и половому размножению со сменой поколений. Большинство грибов производят большое количество спор, которые представляют собой гаплоидные клетки, которые могут подвергаться митозу с образованием многоклеточных гаплоидных особей.Как и бактерии, грибы играют важную роль в экосистемах, потому что они являются разложителями и участвуют в круговороте питательных веществ, расщепляя органические и неорганические материалы на простые молекулы.
Грибы часто взаимодействуют с другими организмами, образуя полезные или мутуалистические ассоциации. Например, большинство наземных растений вступают в симбиотические отношения с грибами. Корни растения соединяются с подземными частями гриба, образуя микоризы. Через микоризу гриб и растение обмениваются питательными веществами и водой, в значительной степени способствуя выживанию обоих видов. Альтернативно, лишайники являются ассоциацией между грибком и его фотосинтетическим партнером (обычно водорослями).Грибы также вызывают серьезные инфекции у растений и животных. Например, болезнь голландского вяза, которая вызывается грибком Ophiostoma ulmi , представляет собой особенно разрушительный тип грибкового заражения, который уничтожает многие местные виды вяза ( Ulmus sp.), Заражая сосудистую систему дерева. Вяз-короед действует как переносчик, передавая болезнь с дерева на дерево. Случайно завезенный в 1900-х годах, гриб уничтожил ильмы по всему континенту. Многие европейские и азиатские ильмы менее восприимчивы к голландскому вязу, чем американские.
Считается, что у людей грибковые инфекции трудно поддаются лечению. В отличие от бактерий, грибы не реагируют на традиционную терапию антибиотиками, потому что они эукариоты. Грибковые инфекции могут оказаться смертельными для людей с ослабленной иммунной системой.
Грибы находят множество коммерческих применений. В пищевой промышленности дрожжи используются в выпечке, пивоварении, производстве сыра и вина. Многие промышленные соединения являются побочными продуктами грибковой ферментации. Грибы являются источником многих коммерческих ферментов и антибиотиков.
Строение и функции клеток грибов
Грибы — это одноклеточные или многоклеточные гетеротрофные разлагатели с толстыми клетками, которые поедают разлагающееся вещество и образуют клубки нитей.
Цели обучения
Опишите физические структуры, связанные с грибами
Ключевые выводы
Ключевые моменты
- Стенки клеток грибов жесткие и содержат сложные полисахариды, называемые хитином (добавляет структурной прочности) и глюканами.
- Эргостерол — это стероидная молекула в клеточных мембранах, которая заменяет холестерин, обнаруженный в мембранах клеток животных.
- Грибы могут быть одноклеточными, многоклеточными или диморфными, то есть тогда гриб может быть одноклеточным или многоклеточным в зависимости от условий окружающей среды.
- Грибы на морфологической вегетативной стадии состоят из клубка тонких нитевидных гиф, тогда как репродуктивная стадия обычно более очевидна.
- Грибы любят находиться во влажной и слабокислой среде; они могут расти как на свете, так и без кислорода.
- Грибы являются сапрофитными гетеротрофами в том смысле, что они используют мертвое или разлагающееся органическое вещество в качестве источника углерода.
Ключевые термины
- глюкан : любой полисахарид, являющийся полимером глюкозы
- эргостерин : функциональный эквивалент холестерина, обнаруженный в клеточных мембранах грибов и некоторых простейших, а также стероидный предшественник витамина D2
- мицелий : вегетативная часть любого гриба, состоящая из массы ветвящихся нитевидных гиф, часто подземных
- гифа : длинная ветвистая нитчатая структура гриба, являющаяся основным способом вегетативного роста
- перегородка : деление клеточной стенки между гифами гриба
- слоевище : вегетативное тело гриба
- сапрофит : любой организм, который живет за счет мертвого органического вещества, например, некоторые грибы и бактерии
- хитин : сложный полисахарид, полимер N-ацетилглюкозамина, обнаруженный в экзоскелетах членистоногих и в клеточных стенках грибов; считается ответственным за некоторые формы астмы у людей
Структура и функции ячеек
Грибы — эукариоты со сложной клеточной организацией.Как и эукариоты, клетки грибов содержат связанное с мембраной ядро, в котором ДНК обернута вокруг гистоновых белков. Некоторые типы грибов имеют структуру, сравнимую с бактериальными плазмидами (петли ДНК). Клетки грибов также содержат митохондрии и сложную систему внутренних мембран, включая эндоплазматический ретикулум и аппарат Гольджи.
В отличие от клеток растений, клетки грибов не содержат хлоропластов или хлорофилла. Многие грибы проявляют яркие цвета, обусловленные другими клеточными пигментами, от красного до зеленого и черного.Ядовитый мухомор Amanita muscaria (мухомор) узнаваем по ярко-красной шляпке с белыми пятнами. Пигменты у грибов связаны с клеточной стенкой. Они играют защитную роль от ультрафиолетового излучения и могут быть токсичными.
Ядовитый Amanita muscaria : Ядовитый Amanita muscaria произрастает в умеренных и северных регионах Северной Америки.
Жесткие слои клеточных стенок грибов содержат сложные полисахариды, называемые хитином и глюканами.Хитин, также содержащийся в экзоскелете насекомых, придает структурную прочность клеточным стенкам грибов. Стенка защищает клетку от высыхания и хищников. У грибов есть плазматические мембраны, подобные другим эукариотам, за исключением того, что структура стабилизируется эргостеролом: молекулой стероида, которая заменяет холестерин, обнаруженный в мембранах клеток животных. Большинство членов королевства Грибки неподвижны.
Рост
Вегетативное тело гриба представляет собой одноклеточное или многоклеточное слоевище.Диморфные грибы могут переходить из одноклеточного в многоклеточное состояние в зависимости от условий окружающей среды. Одноклеточные грибы обычно называют дрожжами. Saccharomyces cerevisiae (пекарские дрожжи) и Candida видов (возбудители молочницы, распространенной грибковой инфекции) являются примерами одноклеточных грибов.
Пример одноклеточного гриба : Candida albicans является дрожжевой клеткой и возбудителем кандидоза и молочницы. Этот организм имеет морфологию, сходную с кокковыми бактериями; однако дрожжи — это эукариотический организм (обратите внимание на ядро).
Большинство грибов — многоклеточные организмы. У них есть две отчетливые морфологические стадии: вегетативная и репродуктивная. Вегетативная стадия состоит из клубка тонких нитевидных структур, называемых гифами (единичные, гифы), тогда как репродуктивная стадия может быть более заметной. Масса гиф — мицелий. Он может расти на поверхности, в почве или разлагающемся материале, в жидкости или даже на живой ткани. Хотя отдельные гифы необходимо наблюдать под микроскопом, мицелий гриба может быть очень большим, а некоторые виды действительно являются «огромным грибом».«Гигантский Armillaria solidipes (опята) считается самым большим организмом на Земле, распространяющимся на более чем 2 000 акров подземной почвы в восточном Орегоне; по оценкам, ему не менее 2400 лет.
Пример мицелия гриба : Мицелий гриба Neotestudina rosati может быть патогенным для человека. Грибок проникает через порез или царапину и развивает мицетому, хроническую подкожную инфекцию.
Большинство гиф грибов разделены на отдельные клетки торцевыми стенками, называемыми перегородками (единичные, перегородки) (а, в).У большинства типов грибов крошечные отверстия в перегородках обеспечивают быстрый поток питательных веществ и небольших молекул от клетки к клетке вдоль гифы. Их описывают как перфорированные перегородки. Гифы в формах для хлеба (принадлежащих к Phylum Zygomycota) не разделены перегородками. Вместо этого они образованы большими клетками, содержащими множество ядер, расположение которых описывается как ценоцитарные гифы (b). Грибы процветают во влажных и слабокислых средах; они могут расти как со светом, так и без него.
Разделение гиф на отдельные клетки : Гифы грибов могут быть (а) септированными или (б) ценоцитарными (цено- = «общий»; -цитарный = «клеточный») с множеством ядер, присутствующих в одной гифе.На микрофотографии в светлом поле (c) Phialophora richardsiae видны перегородки, разделяющие гифы.
Питание
Как и животные, грибы являются гетеротрофами: они используют сложные органические соединения в качестве источника углерода, а не фиксируют углекислый газ из атмосферы, как некоторые бактерии и большинство растений. Кроме того, грибы не фиксируют азот из атмосферы. Как и животные, они должны получать его из своего рациона. Однако, в отличие от большинства животных, которые проглатывают пищу, а затем переваривают ее в специализированных органах, грибы выполняют эти шаги в обратном порядке: пищеварение предшествует приему пищи.Во-первых, экзоферменты выводятся из гиф, где они перерабатывают питательные вещества в окружающей среде. Затем более мелкие молекулы, образующиеся в результате внешнего пищеварения, абсорбируются через большую площадь поверхности мицелия. Как и в случае с клетками животных, запасающий полисахарид — это гликоген, а не крахмал, содержащийся в растениях.
Грибы — это в основном сапробионты (сапрофит — эквивалентный термин): организмы, которые получают питательные вещества из разлагающихся органических веществ. Они получают свои питательные вещества из мертвых или разлагающихся органических веществ, в основном из растительного материала.Экзоферменты грибов способны расщеплять нерастворимые полисахариды, такие как целлюлоза и лигнин мертвой древесины, на легко усваиваемые молекулы глюкозы. Таким образом, углерод, азот и другие элементы выбрасываются в окружающую среду. Из-за разнообразия метаболических путей грибы выполняют важную экологическую роль и исследуются как потенциальные инструменты для биоремедиации.
Некоторые грибы паразитируют, поражая растения или животных. Головня и голландский вяз поражают растения, тогда как микоз и кандидоз (молочница) являются важными с медицинской точки зрения грибковыми инфекциями у людей.
Размножение грибов
Грибы могут размножаться бесполым путем путем фрагментации, образования почки или образования спор, либо половым путем с помощью гомоталлического или гетероталлического мицелия.
Цели обучения
Описать механизмы полового и бесполого размножения грибов
Ключевые выводы
Ключевые моменты
- Новые колонии грибов могут расти в результате фрагментации гиф.
- Во время бутонизации сбоку клетки образуется выпуклость; почка в конечном итоге отделяется после митотического деления ядра.
- Бесполые споры генетически идентичны родительским и могут выделяться либо вне, либо внутри особого репродуктивного мешка, называемого спорангием.
- Неблагоприятные условия окружающей среды часто вызывают половое размножение грибов.
- Мицелий может быть гомоталлическим или гетероталлическим при половом размножении.
- Половое размножение грибов включает следующие три стадии: плазмогамия, кариогамия и гаметангия.
Ключевые термины
- гомоталлический : мужские и женские репродуктивные структуры присутствуют в одном мицелии растений или грибов
- гаметангий : орган или клетка, в которых вырабатываются гаметы, которые обнаруживаются во многих многоклеточных простейших, водорослях, грибах и гаметофитах растений
- спора : репродуктивная частица, обычно одна клетка, выделяемая грибком, водорослью или растением, которая может прорасти в другую
- спорангий : корпус, капсула или контейнер, в котором споры производятся организмом
- кариогамия : слияние двух ядер внутри клетки
- плазмогамия : стадия полового размножения, слияние цитоплазмы двух родительских мицелиев без слияния ядер
Репродукция
Грибки размножаются половым и / или бесполым путем.Совершенные грибы размножаются как половым, так и бесполым способом, тогда как несовершенные грибы размножаются только бесполым (митозом).
Как при половом, так и при бесполом размножении грибы производят споры, которые распространяются от родительского организма, либо плывя по ветру, либо запрягая животное. Споры грибов меньше и легче семян растений. Гигантский гриб-клубень лопается и высвобождает триллионы спор. Огромное количество выпущенных спор увеличивает вероятность попадания в среду, которая будет поддерживать рост.
Выделение спор грибов : (а) гигантский снежно-шариковый гриб выпускает (б) облако спор, когда достигает зрелости.
Бесполое размножение
Грибы размножаются бесполым путем путем фрагментации, образования почки или образования спор. Из фрагментов гиф могут расти новые колонии. Фрагментация мицелия происходит, когда мицелий гриба разделяется на части, при этом каждый компонент превращается в отдельный мицелий. Соматические клетки дрожжей образуют бутоны. Во время почкования (тип цитокинеза) на стороне клетки образуется выпуклость, ядро делится митотически, и почка в конечном итоге отделяется от материнской клетки.
Наиболее распространенным способом бесполого размножения является образование бесполых спор, которые производятся только одним родителем (посредством митоза) и генетически идентичны этому родителю. Споры позволяют грибам расширять свое распространение и заселять новые среды обитания. Они могут выделяться из родительского слоевища как снаружи, так и внутри особого репродуктивного мешка, называемого спорангием.
Типы размножения грибов : Грибы могут иметь как бесполую, так и половую стадии размножения; Половое размножение часто происходит в ответ на неблагоприятные условия окружающей среды.
Есть много видов бесполых спор. Конидиоспоры представляют собой одноклеточные или многоклеточные споры, которые выделяются непосредственно с кончика или стороны гифы. Другие бесполые споры возникают в результате фрагментации гифы с образованием отдельных клеток, которые высвобождаются в виде спор; некоторые из них имеют толстую стенку, окружающую фрагмент. Третьи отпочковываются от вегетативной родительской клетки. Спорангиоспоры образуются в спорангии.
Высвобождение спор из спорангия : На этой микрофотографии в светлом поле показано высвобождение спор из спорангия на конце гифы, называемого спорангиофором.Изображенный организм — это Mucor sp. грибок: плесень, часто встречающаяся в помещении.
Половое размножение
Половое размножение вносит генетические изменения в популяцию грибов. У грибов половое размножение часто происходит в ответ на неблагоприятные условия окружающей среды. Производятся два типа вязки. Когда оба типа спаривания присутствуют в одном и том же мицелии, он называется гомоталлическим или самооплодотворяющимся. Гетероталлический мицелий требует двух разных, но совместимых мицелий для размножения половым путем.
Хотя существует множество вариаций полового размножения грибов, все они включают следующие три стадии. Во-первых, во время плазмогамии (буквально «брак или объединение цитоплазмы») две гаплоидные клетки сливаются, что приводит к дикариотической стадии, когда два гаплоидных ядра сосуществуют в одной клетке. Во время кариогамии («ядерного брака») гаплоидные ядра сливаются, образуя диплоидное ядро зиготы. Наконец, мейоз происходит в органах гаметангий (единичных, гаметангиев), в которых образуются гаметы разных типов спаривания.На этом этапе споры распространяются в окружающую среду.
Взаимодействие грибов и водорослей
Термин «водоросли» (единственное число: водоросль) используется для обозначения огромного множества организмов, похожих на растения. Современные генетические исследования убедительно показали, что организмы, называемые водорослями, принадлежат к нескольким различным царствам и в большинстве случаев вообще не являются растениями. Вероятно, нам следует полностью отказаться от этого термина, но он имеет долгую историю и на самом деле не совсем бесполезен.
Организмы, которые мы называем водорослями, представляют собой разнообразную группу, часто очень отдаленно родственных видов.У всех из них отсутствуют корни и настоящие листья, хотя у некоторых из более крупных есть структуры, похожие на корни, называемые фиксаторами и плоскими листовыми лезвиями. Все они содержат хлорофилл и фотосинтезируют, но во многих группах зеленый цвет маскируется множеством «дополнительных пигментов». Самые маленькие водоросли одноклеточные и всего несколько микрометров в диаметре. Nanochlorum eukaryoticum может быть всего 1 мкм в диаметре. На другом конце шкалы, Nereocystis lutkeana , гигантские водоросли, распространенные на нашем западном побережье, могут достигать 30 метров в длину.На фотографии справа вы можете увидеть длинные листовые пластинки на одном конце слоевища, которые затем змеятся вправо от изображения.
ЛИШАНИИ
Лишайники — это мутуализм, образованный грибами и различными группами водорослей. Невозможно переоценить важность этого взаимопонимания с точки зрения экологии, биоразнообразия и глобального благополучия. Наиболее характерной особенностью лишайников является то, что сочетание двух мутуалистов, водоросли (называемой фотобионтом или фитобионтом ) и гриба (называемым микобионтом ), приводит к форме, называемой слоевищем (множественное число: талломы ). ), который полностью не похож ни на одного из двух участников.Слоевище лишайника может быть довольно большим и бросающимся в глаза, больше напоминающим мох или небольшое растение, чем гриб или водоросль.
Таллос
Талломы лишайников обычно подразделяются на три типа: корковые, листовые или кустистые. Четвертый тип слоевища, называемый чешуйчатым, также часто распознается лихенологами. Распознать эти основные типы несложно.
Слоевища ракообразных плотно прикреплены к субстрату, поэтому имеют хорошо развитую верхнюю поверхность, но просто сливаются с субстратом с нижней стороны.Обычно вы не можете отделить корковый лишай от субстрата, и при этом вы не увидите, как его края загибаются назад, открывая компактную нижнюю поверхность. На иллюстрации слева корковый лишайник Dimelaena oreina растет на поверхности гранитной скалы, к которой он прочно прикреплен. Места, где он, по-видимому, взлетел, вероятно, были вызваны сильными силами, такими как зимний лед или поедание животного.
Листовые лишайники похожи на листья (отсюда и их название) и имеют различную верхнюю и нижнюю поверхность.На снимке слева вид листовых лишайников из рода Peltigera растет на мшистом месте на лесной подстилке. Восходящие доли делают его похожим на растение с листьями. В некоторых местах можно увидеть почти чисто-белую нижнюю часть лепестков
Раковидные лишайники похожи на небольшие деревья или кустарники, у них нет верхней и нижней поверхности; вместо этого их ветви имеют радиальную симметрию, как у растения. Если вы отрежете одну из их ветвей, вы увидите, что у них может быть внутренняя и внешняя поверхность, но никогда не будет ничего, что вы бы пометили сверху и снизу.На фотографии слева представлены два вида: Cladina alpestris , густо-кустистый, и Cladonia rangiferina , менее разветвленный. Cladonia rangiferina , обычно называемая оленьим лишайником, является основным источником корма для карибу и северных оленей в зимний период.
Чешуйчатые лишайники очень похожи на листовые лишайники тем, что имеют двусторонний слоевище с четко выраженными верхом и низом. Их главное отличие — небольшой размер; редко превышает 4 мм.Обычно они образуют плотно упакованные слоевища вместе, образуя колонию, похожую на гальку. Вид Cladonia (слева) типичен для своего рода тем, что дает не только чешуйчатое слоевище, но также вертикальные структуры, называемые podetia , которые в конечном итоге поддерживают апотеции (см. Дальнейшее обсуждение этих структур ниже). Чашевидная структура на фотографии — развивающийся подеций.
Половое размножение
Большинство лишайников являются членами Ascomycota и размножаются с помощью асков и аскоспор.Дополнительные сведения об этих структурах у грибов см. В разделе «Размножение в дикарии».
На изображении справа можно увидеть лишайник Lecanora grantii , распространенный супралиторальный вид в нашей области, с несколькими красновато-коричневыми дискообразными апотециями . Апотеции — это структуры, которые производят частокол асков, которые открыто подвергаются воздействию окружающей среды. Лишайники, продуцирующие апотеции, рассматриваются далее в разделе, посвященном дискомицетам, образующим лишайники.В случае L. grantii слой асков и промежуточных тканей красновато-коричневый и окружен белесым краем апотеция. Несмотря на то, что край выглядит беловатым, он содержит зеленые водоросли, фотобионт, и при продольном разрезе он будет выглядеть зеленым.
Некоторые лишайники образуют аски в перитециях, как показано на фотографии Hydropunctaria maura справа. Перитеции — это структуры в форме колб, которые открываются наружу через пору, называемую устьем .Лишайники, продуцирующие апотеции, рассматриваются далее в разделе, посвященном лихенообразующим пиреномицетам. Устье обоих перитеций справа можно увидеть как небольшие круглые области на вершине.
Есть и другие способы производства аски лишайниками, но наиболее распространены перитеции и апотеции. На фотографиях выше показаны два других типа, с которыми вы можете столкнуться. Первые — это структуры под названием лиреллы . Они похожи на апотеции, но сильно удлинены.В случае Graphis scripta , показанном на фотографии, лиреллы сильно разветвлены и могут напоминать какое-то загадочное письмо. Вторая фотография, Calicium trabinellum , , иллюстрирует mazaedium , разновидность апотеция со стеблями, в котором аски растворяются и оставляют аскоспоры накапливаться в виде порошкообразной массы.
Некоторые базидиомицеты также способны образовывать лишайники. Обычно они не считаются высокоразвитыми взаимоотношениями, но, несомненно, функционируют как лишайники.Первая из двух фотографий выше показывает Multiclavula mucida . У этого вида базидии и базидиоспоры выстилают поверхность вертикальных «пальцев» и в прохладных влажных условиях выпускают споры, которые разносятся ветром. Фотобионт, зеленая водоросль, образует толстую корку субстрата, в данном случае гнилую древесину. Водоросли окружены гифами микобионта. На втором рисунке микобионт — это гриб Lichenomphalia umbellifera . Фотобионт и его связь с фитобионтом такие же, как у M.муцида
Бесполое размножение
До сих пор мы обсуждали только структуры, участвующие в половом размножении лишайников. Поскольку эти половые структуры воспроизводят только гриб, полученные споры должны быть достаточно удачливыми, чтобы приземлиться на подходящую водоросль или погибнуть. Однако есть другой способ. Если лишайник может распространять отростки, содержащие как мико-, так и фотобионты, он сможет развиваться в любой подходящей среде обитания. Однако этот тип воспроизводства является строго клональным и не допускает генетической рекомбинации, которая происходит во время полового размножения.
Клональное размножение лишайников может происходить несколькими способами. Самый простой из них — просто отделить кусок слоевища, содержащий водоросли и грибок, и отправить его ветром или водой для развития на новом месте. Этот вид размножения распространен среди лишайников и в целом эффективен. Есть более развитые формы клонального размножения, две из которых представлены на фотографиях выше. В первом из лишайников образовалось соредий .Соредии — это небольшие пучки водорослей, которые удерживаются гифами грибов. Они достаточно малы, чтобы их переносил ветер, но при этом гарантируют присутствие обоих партнеров. На иллюстрации вверху слева показан молодой слоевище листового лишайника Peltigera didactyla . У этого вида верхняя поверхность усеяна соралиями, — специальными структурами для производства соредий. На фотографии соралии высвободили зернистые массы соредий. Другая фотография выше представляет собой сильно увеличенное изображение isidia , маленьких коралловидных ветвей, содержащих обоих мутуалистов, которые могут отломиться и переместиться в новую среду обитания.На фотографии изображен лишайник Xanthoparmelia conspersa , обычный лишайник на обнаженной скале в Нью-Брансуике.
Места обитания лишайников
Один из захватывающих аспектов биологии лишайников — способность этих организмов занимать места обитания, которые были бы совершенно негостеприимны для других организмов. Таким образом, мы можем найти их растущими на земле в пустынях, по сторонам сухих скал, свисающими с ветвей деревьев и даже растущими на спинах черепах.Их почти так же легко найти и изучить среди зимы, как и в теплые месяцы.
Первая из трех фотографий выше была сделана в Саскачеване, в открытой прерии. Скала на переднем плане — самая высокая точка в непосредственной близости; сидящие там животные получают панорамный вид на луга и все, что там происходит. Это излюбленное место птиц, особенно хищных птиц, которые поджидают мышь или полевку, которые могут передвигаться по траве.Оранжевый лишайник — это разновидность Xanthoria , которая процветает на богатом азотом птичьем помете, оставленном на камнях.
Подобные виды Xanthoria , а также представители родственного рода Caloplaca можно найти на нашем побережье на скалах, часто посещаемых чайками и бакланами. На второй из двух фотографий выше изображен остров Уайт-Хорс, небольшой остров в заливе Фанди, на котором обитают большие колонии гнездящихся птиц. Белый цвет скалы обусловлен толстым слоем птичьего помета; оранжевый материал является разновидностью Caloplaca .
Третий снимок был сделан на Старом кладбище в Киттери-Пойнт, штат Мэн. Надгробие слева отмечает место упокоения Роланда Такстера (1858-1932), профессора Гарвардского университета и блестящего миколога, известного, в частности, своими монументальными исследованиями Лабульбениалес. Рядом с могилой Роланда находится его брат Карл. Оба надгробия заселены лишайниками, и теперь их трудно прочитать. Нажмите на фотографию, чтобы увидеть надгробие Роланда
в увеличенном масштабе.Еще одна интересная особенность наших прибрежных лишайников заключается в том, что некоторые из них очень толерантны к соли — веществу, токсичному для большинства грибов, в том числе лишайниковых.На картинке справа изображены прибрежные скалы в заливе Фанди возле Сент-Джона. Внизу изображения — пучки бурых водорослей, в основном Fucus vesiculosus и Ascophyllum nodosum , обычно называемые каменистыми водорослями. Эти водоросли растут на участках вдоль берега, где они будут погружены в морскую воду, по крайней мере, во время прилива. На самой вершине скалы находится оранжевый участок, вероятно, Xanthoria parietina . Между ними находится черная зона, состоящая из лишайника обыкновенного Hydropunctaria maura . Hydropunctaria maura может расти там, где ее периодически погружают в морскую воду, но также может расти и в области чуть выше той, где она получает только брызги от волн. Эта «черная зона» занимает область, которая часто бывает в течение нескольких дней или даже недель без погружения в морскую воду, но в конечном итоге на нее может попасть брызг. Это тяжелое место для жизни: слишком соленое для наземных организмов и слишком высоко над морем для морских. Как раз место для лишайника!
На рисунке справа изображен еще один вид Verrucaria mucosa , близкий родственник H.Маура . В отличие от H. maura , V. mucosa растет там, где она ежедневно затопляется морской водой. Фактически, он высвобождает свои аскоспоры, когда находится над водой и, следовательно, зависит от контакта с воздухом. Однако он не растет в верхних частях прилива, как H. maura , и может наблюдаться только во время отлива. На снимке V. mucosa видны круглые черные пятна, напоминающие деготь. На высохших частях камня их труднее увидеть, но вы можете заметить, что они слегка зеленые, что указывает на присутствие фотобионта.Красные пятна — это водоросль Hildenbrandia polytypa , аналогичная по размеру и характеру роста водоросле V. mucosa , но не лишайниковая и совершенно не связанная с ней.
На последнем снимке снова показаны Verrucaria mucosa , на этот раз растущие под водой во время прилива. Обратите внимание, что даже у этого лишайника есть свои пределы; на большинстве скал на снимке лишайников нет. Это может быть связано с тем, что камни слишком малы и могут перемещаться течениями во время приливов и отливов, или может быть, что их поверхность непригодна для лишайников.Еще одна проблема, с которой сталкиваются лишайники, — это поедание животных. Многие из них содержат кислоты и другие соединения, которые делают их неприятными для животных, но V. mucosa не является полностью иммунным. Обратите внимание на большой камень над камнем с лишайниками. На его поверхности изображена небольшая улитка, называемая барвинком. Некоторые виды барвинков, особенно барвинок грубый, поедают V. mucosa и могут удалить его из среды обитания. Здесь этого еще не произошло, но на самом деле есть несколько барвинков, а также белые ракушки и мидия.Сколько здесь барвинков? На первый взгляд, немного, но вы можете быть удивлены. Нажмите на картинку, чтобы увеличить ее и посмотреть, сколько барвинков вы можете сосчитать.
ВЗАИМОДЕЙСТВИЕ МЕЖДУ ASCOPHYLLUM NODOSUM И MYCOPHYCIAS ASCOPHYLLI :
Один из наиболее интригующих мутуализмов, обнаруженных в нашем регионе, связан с бурой водорослью Ascophyllum nodosum и грибком Mycophycias ascophylli . Ascophyllum nodosum , обычно называемый каменистым водорослем, встречается в приливной зоне, где он остается открытым для воздуха после отлива. Mycophycias ascophylli , член группы грибов Verrucariales, образующих лишайник, растет в теле (слоевище) A. nodosum , где, как считается, помогает водорослям сопротивляться высыханию при воздействии на них. В свою очередь, гриб имеет доступ к углеводам и другим питательным веществам в своей защитной среде. ДокторД.Дж. Гарбэри и его коллеги из Университета Святого Франциска Ксавьера в Новой Шотландии подробно изучили этот мутуализм и показали, что гриб не только формирует отношения с морскими водорослями, но и, по-видимому, формирует мутуализм с Polysiphonia lanosa , обычным эпифитом, прикрепленным к A. nodosum .
На приведенных выше рисунках показаны некоторые аспекты взаимосвязи Ascophyllum / Mycophycias . Слева слева на скале разложено слоевище A. nodosum .Возможно, вы захотите вернуться на эту страницу и увидеть фотографию среды обитания A. nodosum . На следующем рисунке показана деталь из первой панели. Маленькие миндалевидные образования вдоль стебля — это сосуды. Каждая емкость несет на себе несколько концептуальных элементов, структур, которые каждую весну выпускают в океан сперматозоиды и яйцеклетки. На второй фотографии они видны как неровные участки, но на третьей, более увеличенной панели, они видны легче и показывают поры, через которые выходят сперматозоиды и яйцеклетки.Следующая панель еще больше увеличена, а концептуальные решения еще более ясны. На этой панели также можно увидеть крошечные черные точки, напоминающие крупинки перца; это перитеции (плодовые тела) Mycophycias ascophylli . Синяя рамка, нарисованная вокруг одного из них, ведет к следующей фотографии, сделанной с помощью сложного микроскопа, демонстрирующей подробный вид одного перитеция, частично погруженного в сосуд. Перитеции содержат аски и аскоспоры. На последней панели показан один аск, содержащий восемь двухклеточных аскоспор.Аскоспоры на этой картинке не очень четкие, но, тем не менее, почти созрели.
В нашем регионе Ascophyllum nodosum выпускает свои сперматозоиды и яйцеклетки в конце мая. В теплый день во время отлива эти крошечные клетки сочатся из концептакулов, как зубная паста из тюбика. Когда наступает прилив, их выпускают в воду. Сперма, выпущенная в количестве, достаточном для того, чтобы окрасить воду в оранжевый цвет, активно плавает в поисках яиц и прикрепляется к ним, когда они их находят.Можно наблюдать, как яйца дико крутятся, питаясь сотнями сперматозоидов, прикрепленных к их поверхности. Наконец, один сперматозоид оплодотворяет яйцеклетку, которая затем опускается на дно, чтобы вырасти в новое растение. Одновременно с выделением спермы и яйцеклетки аски Mycophycias ascophylli выпускают свои споры в воздух. Вполне вероятно, что эти споры способны колонизировать только что развивающиеся растения, но детали этой стадии развития все еще не очень хорошо изучены. Оранжевый цвет воды, вызванный огромным количеством сперматозоидов и яйцеклеток, можно принять за токсичный «красный прилив», но это совершенно не связанное с этим явление.
Были некоторые споры по поводу отношения Ascophyllum / Mycophycias . Большинство биологов соглашаются с утверждением доктора Гарбэри о мутуалистическом подходе, но не всегда соглашаются с его значением. Некоторые предполагают, что это тип лишайника, утверждая, что мутуализм между водорослями и грибами по определению является лишайником. Однако другие утверждали, что в настоящем лишайнике гриб является экзобионтом, то есть он окружает водоросль и образует заметную часть слоевища.В мутуализме Ascophyllum / Mycophycias гриб явно является эндобионтом и мало влияет на внешний вид водорослей. Недавние генетические доказательства того, что Mycophycias ascophylli связаны с лишайообразующими Verrucariales, поддерживают сторонников лишайников.
Главная >> Где растут грибы >> Мутуалисты >> Взаимоотношения с водорослями
Экология грибов — Биология 2e
Цели обучения
К концу этого раздела вы сможете делать следующее:
- Описание роли грибов в различных экосистемах
- Описать мутуалистические отношения грибов с корнями растений и фотосинтезирующими организмами
- Опишите полезные отношения между некоторыми грибами и насекомыми
Грибы играют решающую роль в постоянно меняющемся «балансе» экосистем.Они колонизируют большинство мест обитания на Земле, предпочитая темные влажные условия. Они могут процветать в кажущейся враждебной среде, такой как тундра, благодаря наиболее успешному симбиозу с фотосинтезирующими организмами, такими как водоросли, для производства лишайников. В их сообществах грибки не так очевидны, как крупные животные или высокие сокровища. Подобно бактериям, они негласно действуют как основные разложители. Благодаря своему универсальному метаболизму грибы разрушают органические вещества, которые в противном случае не были бы переработаны.
Среда обитания
Хотя грибы в первую очередь связаны с влажной и прохладной средой, обеспечивающей поставку органических веществ, они заселяют удивительное разнообразие сред обитания, от морской воды до кожи и слизистых оболочек человека.Читриды обитают в основном в водной среде. Другие грибы, такие как Coccidioides immitis , вызывающие пневмонию при вдыхании спор, процветают в сухой и песчаной почве на юго-западе США. Грибы, паразитирующие на коралловых рифах, обитают в океане. Однако большинство членов Королевства грибов растут на лесной подстилке, где темная и влажная среда богата разлагающимися остатками растений и животных. В этих средах грибы играют важную роль в качестве разлагателей и переработчиков, позволяя членам других царств получать питательные вещества и жить.
Декомпозиторы и рециклеры
Пищевая сеть была бы неполной без организмов, разлагающих органические вещества ((Рисунок)). Некоторые элементы, такие как азот и фосфор, требуются биологическим системам в больших количествах, но их не так много в окружающей среде. Действие грибов высвобождает эти элементы из разлагающегося вещества, делая их доступными для других живых организмов. Микроэлементы, присутствующие в небольших количествах во многих местах обитания, необходимы для роста и будут оставаться связанными с гниющими органическими веществами, если грибы и бактерии не вернут их в окружающую среду благодаря своей метаболической активности.
Скоба грибов. Грибы — важная часть круговорота питательных веществ экосистемы. Эти кустовые грибы, растущие на стороне дерева, являются плодовыми структурами базидиомицета. Питательные вещества они получают через гифы, которые проникают в ствол дерева и разрушают его. (Кредит: Кори Занкер)
Способность грибов разрушать многие крупные и нерастворимые молекулы обусловлена их режимом питания. Как было замечено ранее, пищеварение предшествует приему пищи. Грибы производят множество экзоферментов для переваривания питательных веществ.Ферменты либо высвобождаются в субстрат, либо остаются связанными с внешней стороной клеточной стенки грибка. Большие молекулы распадаются на маленькие, которые транспортируются в клетку системой белков-носителей, встроенных в клеточную мембрану. Поскольку движение небольших молекул и ферментов зависит от присутствия воды, активный рост зависит от относительно высокого процента влаги в окружающей среде.
Как сапробионты, грибы помогают поддерживать устойчивую экосистему для животных и растений, обитающих в одной среде обитания.Помимо пополнения окружающей среды питательными веществами, грибы напрямую взаимодействуют с другими организмами полезными, а иногда и вредными способами ((Рисунок)).
Шельфовые грибы. Полочные грибы, названные так потому, что они растут на деревьях в штабеле, атакуют и переваривают ствол или ветви дерева. В то время как некоторые шельфовые грибы встречаются только на мертвых деревьях, другие могут паразитировать на живых деревьях и вызывать в конечном итоге смерть, поэтому они считаются серьезными патогенами деревьев. (Кредит: Кори Занкер)
Взаимоотношения
Симбиоз — это экологическое взаимодействие двух организмов, живущих вместе.Это определение не описывает тип или качество взаимодействия. Когда выигрывают оба члена ассоциации, симбиотические отношения называются мутуалистическими. Грибы образуют мутуалистические ассоциации со многими типами организмов, включая цианобактерии, водоросли, растения и животных.
Грибок / мутуализм растений
Одной из самых замечательных ассоциаций между грибами и растениями является установление микоризы . Микориза, происходящая от греческих слов myco , означающих гриб, и rhizo , означающих корень, относится к грибовому партнеру мутуалистической ассоциации между корнями сосудистых растений и их симбиотическими грибами.Почти 90 процентов всех видов сосудистых растений имеют микоризных партнеров. В микоризной ассоциации грибной мицелий использует свою обширную сеть гиф и большую площадь поверхности в контакте с почвой, чтобы направлять воду и минералы из почвы в растения. Взамен растение поставляет продукты фотосинтеза, которые подпитывают метаболизм гриба.
Существует несколько основных типов микоризы. Эктомикориза («внешняя» микориза) зависит от грибов, окружающих корни оболочкой (так называемой мантией).Гифы прорастают из мантии в корень и охватывают внешние слои корневых клеток сетью гиф, называемой сетью Хартига ((рисунок)). Грибковый партнер может принадлежать к Ascomycota, Basidiomycota или Zygomycota. Эндомикориза («внутренняя» микориза), также называемая арбускулярной микоризой , образуется, когда грибы растут внутри корня в разветвленной структуре, называемой арбускулой (от латинского «маленькие деревья»). Все грибковые партнеры эндомикоризных ассоциатов принадлежат к Glomeromycota.Грибковые арбускулы проникают в клетки корня между клеточной стенкой и плазматической мембраной и являются местом метаболического обмена между грибком и растением-хозяином ((Рисунок) b и (Рисунок) b ). Орхидеи полагаются на третий тип микоризы. Орхидеи — это эпифиты, которые обычно производят очень маленькие семена, переносимые воздухом, без достаточного хранения для поддержания прорастания и роста. Их семена не прорастут без микоризного партнера (обычно базидиомицета). После того, как питательные вещества в семенах истощаются, грибковые симбионты поддерживают рост орхидеи, обеспечивая необходимые углеводы и минералы.Некоторые орхидеи продолжают оставаться микоризными на протяжении всего жизненного цикла.
Визуальное соединение
Два типа микоризы. (а) Эктомикориза и (б) арбускулярная или эндомикориза имеют разные механизмы взаимодействия с корнями растений. (кредит b: М.С. Турмель, Университет Манитобы, факультет растениеводства)
Если бы симбиотические грибы отсутствовали в почве, как вы думаете, какое влияние это оказало бы на рост растений?
Без микоризы растения не могут усваивать достаточное количество питательных веществ, что замедляет их рост.Добавление спор грибов в стерильную почву может облегчить эту проблему.
Микориза. (А) Заражение корней Pinus radiata (сосна монтерейская) гифами Amanita muscaria (мухомор мухомор) приводит к образованию на сосне множества мелких разветвленных корешков. Гифы Amanita покрывают эти маленькие корни белой мантией. (b) Споры (круглые тела) и гифы (нитевидные структуры) видны на этой светлой микрофотографии арбускулярной микоризы, вызванной грибком на корне кукурузы.(кредит а: модификация работы Рэнди Молина, USDA; кредит b: модификация работы Сары Райт, USDA-ARS; данные шкалы от Мэтта Рассела)
Другие примеры мутуализма между грибами и растениями включают эндофиты: грибы, которые живут внутри тканей, не повреждая растение-хозяин. Эндофиты выделяют токсины, которые отпугивают травоядных животных или придают устойчивость к факторам стресса окружающей среды, таким как заражение микроорганизмами, засуха или тяжелые металлы в почве.
Evolution Connection
Коэволюция наземных растений и микоризы Как мы видели, микоризы являются грибковыми партнерами взаимовыгодной симбиотической ассоциации, которая одновременно развивалась между корнями сосудистых растений и грибов.Хорошо обоснованная теория предполагает, что грибы сыграли важную роль в эволюции корневой системы растений и способствовали успеху покрытосеменных растений. У мохообразных (мхи и печеночники), которые считаются самыми древними растениями и первыми выжили и адаптировались на суше, есть простые подземные ризоиды, а не настоящая корневая система, и поэтому они не могут выжить в засушливых районах. Однако некоторые мохообразные имеют арбускулярную микоризу, а некоторые нет.
Истинные корни впервые появились у предковых сосудистых растений: сосудистые растения, которые развили систему тонких отростков от своих корней, имели бы избирательное преимущество перед несосудистыми растениями, потому что они имели большую площадь поверхности контакта с грибковыми партнерами, чем ризоиды мхи и печеночники.Первые настоящие корни позволили бы сосудистым растениям получать больше воды и питательных веществ из почвы.
летописи окаменелостей показывают, что грибы на самом деле предшествовали вторжению пресноводных пресноводных растений на сушу. Первая ассоциация между грибами и фотосинтезирующими организмами на суше была связана с мохоподобными растениями и эндофитами. Эти ранние ассоциации возникли до того, как у растений появились корни. Постепенно преимущества взаимодействия эндофитов и ризоидов для обоих партнеров привели к современной микоризе: около 90 процентов современных сосудистых растений имеют ассоциации с грибами в своей ризосфере.
Грибы, участвующие в микоризах, обладают многими характеристиками предковых грибов; они производят простые споры, мало разнообразны, не имеют полового репродуктивного цикла и не могут жить вне микоризной ассоциации. Растения выиграли от ассоциации, потому что микоризы позволили им переехать в новые среды обитания и увеличили потребление питательных веществ, что дало им огромное селективное преимущество перед растениями, которые не установили симбиотических отношений.
Лишайники
Лишайники имеют разные цвета и текстуры ((Рисунок)) и могут выжить в самых необычных и враждебных средах обитания. Они покрывают камни, надгробия, кору деревьев и землю в тундре, куда не могут проникнуть корни растений. Лишайники могут пережить длительные периоды засухи, когда они полностью высыхают, а затем быстро становятся активными, когда вода снова становится доступной.
Ссылка на обучение
Исследуйте мир лишайников с помощью сайта Университета штата Орегон.
Лишайники. У лишайников много форм. Они могут быть (а) корковидными, (б) волосковыми или (в) листовыми. (кредит а: модификация работы Джо Нейлор; кредит б: модификация работы «djpmapleferryman» / Flickr; кредит c: модификация работы Кори Занкер)
Важно отметить, что лишайники — это не один организм, а, скорее, еще один замечательный пример мутуализма, в котором гриб (обычно член Ascomycota или Basidiomycota) живет в физических и физиологических отношениях с фотосинтетическим организмом. (эукариотическая водоросль или прокариотическая цианобактерия) ((Рисунок)).Как правило, ни грибок, ни фотосинтезирующий организм не могут выжить в одиночку вне симбиотических отношений. Тело лишайника, называемое слоевищем, состоит из гиф, обернутых вокруг фотосинтетического партнера. Фотосинтезирующий организм обеспечивает углерод и энергию в виде углеводов. Некоторые цианобактерии дополнительно фиксируют азот из атмосферы, внося в ассоциацию азотистые соединения. В свою очередь, гриб поставляет минералы и защищает от сухости и чрезмерного света, заключая водоросли в свой мицелий.Грибок также прикрепляет лишайник к своему субстрату.
Состав лишайника. На этом поперечном срезе слоевища лишайника показана (а) верхняя кора грибковых гиф, обеспечивающая защиту; (b) зона водорослей, где происходит фотосинтез, (c) мозговой слой гиф грибов и (d) нижняя кора головного мозга, которая также обеспечивает защиту и может иметь (e) ризина для прикрепления слоевища к субстрату.
Слоевище лишайников растет очень медленно, увеличивая свой диаметр на несколько миллиметров в год.И грибок, и водоросль участвуют в образовании единиц распространения, называемых соредиями — скоплений клеток водорослей, окруженных мицелием. Соредии разносятся ветром и водой и образуют новые лишайники.
Лишайники чрезвычайно чувствительны к загрязнению воздуха, особенно к аномальным уровням азотистых и сернистых соединений. Лесная служба США и Служба национальных парков могут отслеживать качество воздуха, измеряя относительную численность и состояние популяции лишайников в определенной области. Лишайники выполняют множество экологических функций.Карибу и северный олень поедают лишайники и служат укрытием для мелких беспозвоночных, которые прячутся в мицелии. При производстве тканей ткачи использовали лишайники для окрашивания шерсти на протяжении многих веков, вплоть до появления синтетических красителей. Пигменты, используемые в лакмусовой бумаге, также извлекаются из лишайников.
Ссылка на обучение
Лишайники используются для контроля качества воздуха. Подробнее читайте на этом сайте Лесной службы США.
Грибок / мутуализм животных
Грибы развили взаимные отношения с многочисленными насекомыми типа членистоногих: суставноногие беспозвоночные с хитиновым экзоскелетом.Членистоногие зависят от грибка для защиты от хищников и патогенов, в то время как гриб получает питательные вещества и способ распространения спор в новую среду. Одним из примеров является связь между видами Basidiomycota и щитовками. Грибной мицелий покрывает и защищает колонии насекомых. Щитовки способствуют поступлению питательных веществ от зараженного растения к грибку.
Во втором примере муравьи-листорезы Центральной и Южной Америки буквально выращивают грибы.Они срезают с растений диски из листьев и складывают их в подземных садах ((Рисунок)). В этих дисковых садах выращивают грибы, которые переваривают целлюлозу в листьях, которую муравьи не могут разрушить. Когда более мелкие молекулы сахара производятся и потребляются грибами, они, в свою очередь, становятся пищей для муравьев. Насекомые также патрулируют свой сад, охотясь на конкурирующие грибы. От этой мутуалистической ассоциации выигрывают и муравьи, и грибы. Гриб получает постоянный запас листьев и свободу от конкуренции, в то время как муравьи питаются грибами, которые они культивируют.
Листорез муравейник. Муравей-листорез переносит лист, которым питается выращиваемый на ферме гриб. (кредит: Скотт Бауэр, USDA-ARS)
Грибоядные
Расселение животных важно для некоторых грибов, поскольку животные могут переносить споры грибов на значительные расстояния от источника. Споры грибов редко полностью разлагаются в желудочно-кишечном тракте животного, и многие из них способны прорастать, попадая с фекалиями. Некоторым «навозным грибам» действительно требуется пройти через пищеварительную систему травоядных, чтобы завершить свой жизненный цикл.Черный трюфель — ценный деликатес для гурманов — плодовое тело подземного аскомицета. Почти все трюфели эктомикоризны и обычно находятся в тесной связи с деревьями. Животные поедают трюфели и разносят споры. В Италии и Франции охотники за трюфелями используют самок свиней для обнюхивания трюфелей (самок свиней привлекают трюфели, потому что гриб выделяет летучие соединения, тесно связанные с феромоном, вырабатываемым самцами свиней).
Сводка раздела
Грибы колонизировали почти все среды на Земле, но часто встречаются в прохладных, темных, влажных местах с запасом разлагающегося материала.Грибы — это сапробионты, разлагающие органические вещества. Многие успешные взаимоотношения связаны с грибком и другим организмом. Многие грибы устанавливают сложные микоризные ассоциации с корнями растений. Некоторые муравьи выращивают грибы в качестве пищи. Лишайники — это симбиотические отношения между грибком и фотосинтезирующим организмом, обычно водорослями или цианобактериями. Фотосинтезирующий организм обеспечивает энергию из хранимых углеводов, в то время как гриб обеспечивает минералами и защитой. Некоторые животные, потребляющие грибы, способствуют распространению спор на большие расстояния.
Вопросы о визуальном подключении
(Рисунок) Если симбиотические грибы отсутствуют в почве, как вы думаете, какое влияние это окажет на рост растений?
(Рисунок) Без микоризы растения не могут усваивать достаточное количество питательных веществ, что замедляет их рост. Добавление спор грибов в стерильную почву может облегчить эту проблему.
Обзорные вопросы
Какой термин описывает тесную связь грибка с корнем дерева?
- ризоид
- лишайник
- микориза
- эндофит
Почему грибы являются важными деструкторами?
- Они производят много спор.
- Они могут расти в самых разных средах.
- Они производят мицелий.
- Они перерабатывают углерод и неорганические минералы в процессе разложения.
Рассмотрим экосистему, в которой уничтожены все грибы, не участвующие в микоризах. Как это повлияет на потребление азота растениями?
- Потребление азота увеличится.
- Потребление азота не изменится.
- Потребление азота уменьшится.
- Потребление азота прекратится.
Вопросы о критическом мышлении
Почему защита от света приносит пользу партнеру по фотосинтезу лишайников?
Защита от избыточного света, который может обесцветить фотосинтетические пигменты, позволяет партнеру по фотосинтезу выживать в окружающей среде, неблагоприятной для растений.
Жуки-короеды Ambrosiella переносят споры грибка Ambrosiella к деревьям, затем сверлят норы и откладывают яйца с грибком. Когда вылупляются новые личинки, они поедают проросший в лунках грибок.Опишите, как эти отношения можно классифицировать как мутуалистические.
Жуки-короеды и гриб имеют мутуалистические отношения, поскольку каждый партнер получает выгоду от взаимодействия друг с другом. Жук может дать пищу своему потомству, а грибок может распространяться на новые деревья.
Экологи часто пытаются внедрить новые растения для восстановления деградированных земель. В засушливом климате ученые рекомендуют заносить растения с арбускулярной микоризой. Как микориза может увеличить выживаемость растений по сравнению с растениями без микоризы?
Растения с арбускулярной микоризой заселяются грибами, которые проникают в клетки корня и обмениваются с растением метаболитами.Сеть гифов грибов простирается от корневых клеток в окружающую среду, охватывая большую площадь, чем только корневая система растения. Это позволяет растению набирать воду с большей площади, повышая вероятность того, что оно сможет удовлетворить свои повседневные потребности.
Глоссарий
- арбускулярная микориза
- микоризная ассоциация, при которой гифы гриба проникают в клетки корня и образуют обширные сети
- эктомикориза
- микоризные грибы, которые окружают корни мантией и имеют сеть Хартига, которая проникает в корни между клетками
- лишайник
- тесная связь грибка с фотосинтезирующей водорослью или бактерией, которая приносит пользу обоим партнерам
- микориза
- мутуалистическая ассоциация грибов и корней сосудистых растений
- soredia
- кластеров клеток водорослей и мицелия, которые позволяют лишайникам размножаться
Симбиоз водорослей и грибов приводит к фотосинтетическому мицелию
Прежде всего, мы хотим поблагодарить вас за конструктивное рассмотрение нашей рукописи и сказать, что мы нашли ваши комментарии ценными.Следуя вашему совету, мы провели множество дополнительных экспериментов, которые были предложены, в том числе серию новых экспериментов с растровой электронной микроскопией, чтобы детализировать взаимодействие клеточной стенки между двумя типами клеток, контроль ПЦР для выделения водорослей из грибов, анализы фотосинтеза / хлорофилла совместно выращиваемых растений. водоросли, анализы роста / биомассы совместно культивируемых водорослей и грибов, световая и конфокальная микроскопия и анализы жизнеспособности как водорослей, так и грибов в зеленых гифах. Мы подготовили новые рисунки и пересмотрели все рисунки для ясности, включая новые микрофотографии, полученные с помощью просвечивающего электронного микроскопа, на которых видны эндомикроводоросли, окруженные грибковыми мембранами и пузырьками.Мы чувствуем ваши предложения, и эти дополнения значительно улучшили нашу новую заявку, и надеемся, что вы и рецензенты согласны.
Рецензент # 1:
Авторы делают интересное открытие: при совместном выращивании водоросль Nannochloropsis oceanica и гриб Mortierella elongata образуют консорциум. Авторы предоставляют некоторые свидетельства перемещения углерода и азота между симбионтами, некоторые свидетельства специфичности взаимодействия в том смысле, что это происходит не только с какими-либо грибами, а также свидетельства роста N.oceanica внутри гиф M. elongate, хотя не так ясно, живы ли гифы. Данные интригуют, и система может дать представление о возникновении новых симбиозов и о том, как развиваются мутуализмы, а также о возможных приложениях. Однако в настоящее время я считаю, что данных недостаточно, чтобы поддержать широкие утверждения, которые делаются, а доказательства эндосимбиоза особенно слабы. В некоторых случаях отсутствуют важные элементы управления, и делаются убедительные заявления без достаточной экспериментальной поддержки.
Мы провели дополнительные эксперименты, чтобы подтвердить выводы, и смягчили грубые выражения.
1) Взаимодействие Noc с различными грибами оценивали на основе внешнего вида агрегатов, и поскольку агрегаты наблюдались только для этой пары организмов, эти данные, безусловно, свидетельствуют в пользу чего-то уникального, происходящего во время совместного культивирования этих двух видов. Однако количественные данные, показывающие рост этих двух организмов с течением времени, отсутствуют, хотя утверждается, что их можно совместно культивировать вместе в течение длительных периодов времени и что совместное культивирование им выгодно.Есть некоторые свидетельства того, что N. oceanica демонстрирует небольшое улучшение роста при совместном культивировании в среде -C или -N, но не -P. Однако исходный процент жизнеспособности в этих экспериментах различается (очень высокий в выражении -P), поэтому неясно, действительно ли отсутствие разницы в -P. Кроме того, количество подсчитанных клеток было в 5 раз меньше в эксперименте P. Мне не удалось найти никаких данных, подтверждающих утверждение, что Mortierella «растет в PBS за счет включения метаболитов, полученных из водорослей» (заявлено в подразделе «Дефицит питательных веществ и преимущества совместного культивирования для N.oceanica и M. elonga »). Фактически, данные о росте Mortierella, похоже, полностью отсутствуют.
Наблюдения интересны, но данные о росте важны (и желательно при различных условиях питания) для подтверждения утверждений о взаимном обмене питательными веществами, приводящем к взаимному взаимопониманию.
Поскольку не было показано, что P играет важную роль во взаимодействии и не является основной частью этого исследования, мы удалили результат из рукописи и подготовили новый рисунок 3 и новый рисунок 3 — приложение к рисунку 1.
Количественное определение грибковой биомассы технически сложно. Мы провели покадровую микроскопию и обнаружили, что гифы грибов продолжали расти в буфере PBS в присутствии партнера водорослей, тогда как сами грибы в PBS не демонстрировали никакого очевидного нового роста. Мы предоставляем параллельное видео, чтобы показать разницу (Видео 1).
В этом новом материале мы приложили большие усилия для количественной оценки роста водорослей и грибов во время совместного культивирования. Мы обнаружили, что Mortierella elongata содержит основные уникальные жирные кислоты, такие как α-линоленовая кислота (C18: 3), которые являются лишь второстепенным компонентом в Nannochloropsis .Следовательно, он может служить биомаркером для оценки численности грибов. Мы обнаружили, что соотношение C18: 3 к биомассе у гриба Mortierella очень стабильно после инкубации в среде f / 2-N (Рисунок 3 — приложение к рисунку 2A). Таким образом, мы использовали C18: 3 в качестве маркера для количественной оценки биомассы грибов в агрегатах водоросли-грибы. Мы измерили содержание C18: 3 и общую биомассу агрегатов водорослей и грибов. Мы действительно наблюдали увеличение биомассы как водорослей, так и грибов после совместного культивирования в среде f / 2-N (среда, лишенная питательных веществ как для грибов, так и для водорослей), чего не наблюдалось, когда партнеров инкубировали по отдельности в одной и той же среде (рис. 3 — приложение к рисунку 2Б).
2) Интересна идея, что Noc может расти внутри гиф. Изображения, полученные с помощью светового микроскопа с малым увеличением, безусловно, демонстрируют наличие клеток водорослей внутри гиф. Однако данные не убеждают меня в том, что эти гифы живые. Главным образом потому, что на ЭМ изображениях гифы, которые явно содержат Noc, не содержат органелл. Кроме того, данные, подтверждающие утверждение, что Noc окружен мембраной клетки-хозяина (рис. 4), очень слабы. Подходы химической фиксации не обеспечивают уровень разрешения, необходимый для подтверждения заявлений о новых мембранах.На этих изображениях очень сложно визуализировать мембраны, поэтому следует использовать замораживание под высоким давлением. К сожалению, видео не дает четкого изображения Noc внутри живых гиф. Если они действительно живут вместе, с Noc внутри гиф Mortierella, должно быть возможно получить изображения гиф, на которых есть четкие доказательства цитоплазматического потока вместе с живыми клетками водорослей.
Интернализация интересна, но требует дополнительных данных, чтобы подтвердить утверждения о том, что оба организма действительно живы и растут.Если, как отмечают авторы, эти культуры можно фрагментировать и поддерживать в течение многих месяцев, тогда не составит труда продемонстрировать, что мицелий жив и что организмы сосуществуют.
Мы понимаем озабоченность рецензентов. Чтобы проверить, живут ли клетки водорослей внутри гиф грибов и являются ли сами гифы грибами (имеют ли они клеточное содержимое), мы выполнили дополнительную микроскопию, включая световую ДИК, конфокальную и электронную микроскопию.Наши новые данные демонстрируют, что клетки водорослей живы в живых клетках грибов.
В этой редакции мы заменили изображения ПЭМ на рисунке 4 изображением, показывающим, что интактные клетки водорослей находятся внутри гифы гриба и окружены цитоплазмой грибов, включая органеллы (AG77 и Noc, рис. 4F и G; NVP64 и Noc, Рисунок 4 — приложение к рисунку 2С).
Конфокальные изображения SYTOX, полученные зеленой микроскопией на рис. 4 — дополнение к рисунку 5 A-C, демонстрируют, что как интернализованные водоросли, так и клетки-хозяева живы.Зонд SYTOX проникает в мертвые клетки, выделяя их, и сигнал становится очевидным, когда мертвые клетки используются в качестве контроля (Рисунок 4 — приложение к рисунку с 5D по H). В агрегатах наблюдали несколько мертвых клеток водорослей, но внутри гифов грибов не наблюдалось. Затем мы использовали световой микроскоп DIC, чтобы проверить, были ли гифы грибов полыми или были ли в них цитоплазма и органеллы. Мы наблюдали, что гифы грибов содержат цитоплазму, а клетки водорослей в гифах грибов окружены грибковыми пузырьками, включая липидные капли, которые присутствуют в живых клетках (Рисунок 4 — рисунок в приложении 6).Мы записали видео в реальном времени, чтобы показать трехмерное содержание зеленых гиф, и видео 5 является репрезентативным. Мы также отмечаем, что цитоплазматический поток является обычным для молодых активно растущих клеток в среде с низким содержанием углерода f / 2, но через 2-3 недели цитоплазматический поток не проявляется в более старых частях мицелиальной сети. Следовательно, неудивительно, что поток цитоплазмы не наблюдался в зрелых гифах грибов в течение одного месяца совместного культивирования.
Существенные изменения:
1) В качестве общего комментария некоторые эксперименты четко не описаны.Например, на рисунке 1 наблюдалась агрегация и совместное присутствие клеток водорослей и грибов. Утверждается, что интерфейс напоминает лишайники, но не описаны особенности, на которых основан этот вывод. Агрегируются ли когда-либо клетки Nostoc при выращивании в одиночку в среде f / 2? Присутствуют ли поверхностные структуры на клетках Nostoc только при совместном культивировании с Mortierella или это общая особенность клеток Nostoc. Как минимум должно быть включено изображение клеток Nostoc, выращенных в отсутствие Mortierella.Какой состав у f / 2?
Благодарим рецензента за эти комментарии. Мы соответствующим образом отредактировали рукопись, чтобы предоставить более подробные описания. Чтобы уточнить, мы работаем с Nannochloropsis (Noc), одноклеточной микроводорослью, а не с цианобактерией Nostoc . Мы демонстрируем с помощью дополнительных контролируемых экспериментов, что морфология и агрегация клеток Nannochloropsis (Noc) значительно различаются между монокультурами и совместными культурами с живыми Mortierella. После продолжительной инкубации после стационарной фазы в среде f / 2 Клетки Nannochloropsis (Noc) могут агрегировать при выращивании по отдельности, но в сравнительно небольших кластерах (до десятков клеток) (Рисунок 1 — приложение к рисунку 1A). С помощью сканирующей электронной микроскопии (SEM) мы демонстрируем, что клетки Noc, инкубированные в виде монокультуры, имеют гладкую поверхность наружной клеточной стенки (рис. 1D; рис. 1-S1A), тогда как клетки Noc, культивируемые совместно с живыми клетками Mortierella , демонстрируют волокнистое расширение от их клеточные стенки (рис. 1C, E и F; рис. 1 — приложение к рисунку 1D).В частности, тонкий внешний слой отделяется от клеток Noc при инкубации с Mortierella , и находящиеся под ним волокнистые отростки становятся обнаженными (рис. 1 — рис. Большие части сломанной мембраны видны в клетках Noc (желтые стрелки, Рисунок 1 — рисунок в приложении 1D), тогда как другие имеют меньшие остаточные фрагменты (Рисунок 1E и F). Мы также исследовали свободные клетки Noc в супернатанте совместной культуры Noc- Mortierella и обнаружили, что клетки имеют частично поврежденную покровную мембрану (не такую гладкую, как в контроле), но волокнистые отростки не обнажены (рис. — приложение к рисунку 1С).
Как и в контрольных монокультурах водорослей, тонкая внешняя мембрана, покрывающая клетки Noc, также очевидна при совместном культивировании с не взаимодействующими грибами ( Clavulina sp. PMI390 и Morchella americana GB760 — Рисунок 3 — приложение к рисунку 3 и Рисунок 3 — приложение к рисунку 4) или с мертвыми клетками Mortierella (рисунок 1 — приложение к рисунку 1B). Волокнистые отростки, по-видимому, важны для физического взаимодействия клеток Noc и AG77. Мы включили эти новые находки в рукопись.Мы также включили состав среды f / 2 в раздел «Метод», который является распространенной и широко используемой универсальной обогащенной средой для выращивания морских водорослей.
2) Рисунок 2 — дополнение к рисунку 1. ПЦР-тесты на загрязнение — очень хорошая идея, но положительные контроли отсутствуют. Показывают ли 3 образца гиф положительную реакцию ПЦР с геном гриба и то же самое для Noc?
Мы провели ПЦР-тесты с положительными и отрицательными ПЦР-контролями.Три образца гиф показали положительную реакцию ПЦР на маркер грибкового гена EF1a , а два образца водорослей показали положительную амплификацию гена водорослей AURE04 . Не было ПЦР-амплификации ни для одной из реакций отрицательного контроля (Рисунок 2 — рисунок в приложении 1D и E).
3) В экспериментах по маркировке нормализована ли радиоактивность по отношению к сухой массе организма или по отношению к сухой массе измеряемой молекулы (например, белков, липидов или углеводов).Это важно, потому что пропорции этих макромолекул, вероятно, различаются у двух организмов.
Для экспериментов по маркировке использовали общий сухой вес клеток. Теперь у нас есть четкое описание этого в разделе «Метод» и пояснение в легенде рисунка.
4) Есть ли различия в степени агрегации в условиях дефицита питательных веществ?
Мы не наблюдали очевидной разницы в агрегатах Noc-AG77 в условиях дефицита питательных веществ.
5) Утверждается, что они могут быть совместными культивациями долгое время. Сколько?
Мы определяем долгосрочную совместную культуру продолжительностью от одного до трех месяцев. И Noc, и AG77 — быстрые производители. Noc-клетки могут удвоиться в течение 48 часов, а AG77 может заполнять чашку или распространяться по колбе в течение 2-3 дней после инокуляции.
Рецензент № 2:
Рукопись Чжи-Янь Ду и его коллег описывает новое взаимодействие, установленное между водорослями, производящими биотопливо (Nannochloropsis oceanica), и грибком Mortierella elongata.Авторы предлагают подробное описание ассоциации (начиная с агрегации водорослей, в конечном итоге приводящей к интернализации водорослей), а также некоторые функциональные характеристики (включая обмен питательными веществами между партнерами). Авторы предполагают, что их открытие может иметь соответствующие биотехнологические приложения, поскольку оба микробы важны для производства липидов и топлива.
1) Рукопись интересна и оригинальна. Приятно видеть, что базальные грибы, такие как Mucoromycota, могут взаимодействовать не только с большинством линий растений, но и с водорослями.Однако есть много слабых мест в экспериментальных подходах, в качестве снимков, получаемых с помощью электронного микроскопа, а также в использовании терминологии, связанной с симбиозом.
Мы улучшили наши электронные микрофотографии и пересмотрели терминологию после комментариев рецензента.
2) В качестве общего комментария (и авторы могут или не могут согласиться) сравнение с лишайниками иногда вводит в заблуждение: организация лишайников очень разнообразна, с небольшими гифами, которые окружают клетки водорослей, и очень часто эти гифы проникают внутрь клеток водорослей. продуцирующие колышки или внутриклеточные гаустории (см., например, Honegger, 1986).Описанная здесь система полностью разнообразна (Noc очень мал в диаметре по сравнению с Mortierella hyphae) и напоминает другие взаимодействия, при которых водоросли, например, эукариотические эндосимбионты, размножаются внутри гетеротрофных протистов, таких как Paramecium или Hydra. Результатом этих взаимодействий является фотосинтетическая ассоциация, в которой не образуются псевдоткани (в отличие от лишайников). Авторы могут взглянуть на работу Анджелы Дуглас (2009) или на обширный обзор Новака и Мелконяна (2010), где проиллюстрированы эти симбиозы.Главный вывод состоит в том, что первая функция, которую выполняют эукариотические эндосимбионты, когда они участвуют в стабильных взаимодействиях с живыми протистами, — это фотосинтез. Эта общая концепция также может помочь авторам лучше охарактеризовать функциональность описываемой ассоциации (зеленый фотосинтетический мицелий).
Благодарим рецензента за эти предложения и соглашаемся. Мы отредактировали и включили их в раздел «Обсуждение».
Основные комментарии:
3) Качество изображений, показывающих взаимодействие двух партнеров, не полностью удовлетворяет.На рис. 1 панели C, E, D очень плохие. И гифы, и клетки водорослей, кажется, разрушены материалом, присутствующим на поверхности, что очень трудно интерпретировать (см., Например, поверхность Noc на панели D). Это результат артефакта подготовки? А как насчет контрольной водорослевой клетки, которая сохраняется в отсутствие грибка? его поверхность гладкая? Возникают ли бородавки / выступы при наличии грибка? На мой взгляд, контрольный эксперимент отсутствует. N oceanica обычно описывается с гладкой поверхностью.Также необходимо тщательно проверить легенду. Если авторы говорят, что клетки Noc захватываются, это подразумевает активный механизм действия гриба (как и в случае грибов, которые улавливают нематод). Напротив, эксперименты предполагают агрегацию (см. Ниже).
Мы благодарим рецензента за эти предложения.
Следуя этим предложениям, мы провели контролируемые эксперименты для анализа клеток водорослей, выращенных в монокультуре в среде f / 2, а также с неживыми Mortierella и несовместимыми грибами.Как упоминалось выше, мы обнаружили, что эти клетки Noc сами по себе имеют гладкую поверхность (рисунок 1D; рисунок 1 — рисунок в приложении 1A) при выращивании в монокультуре или с неживыми Mortierella или несовместимыми грибами (например, Morchella, Clavulina spp.). Волокнистые удлинения наблюдались только в клетках Noc, совместно культивируемых с живыми клетками Mortierella (Рисунок 1C, E и F; Рисунок 1 — рисунок в приложении 1D). Волокнистые отростки покрыты тонким слоем относительно гладкой мембраны.При инкубации с Mortierella этот слой распадается, обнажая фиброзные отростки ниже (Рисунок 1 — приложение к рисунку 1D). Некоторые клетки Noc все еще имеют большие части разорванной мембраны (желтые стрелки, рис. 1 — рисунок в приложении 1D), тогда как другие имеют небольшие остатки (рис. 1E и F).
Scholz et al., 2014 выполнили крио-ЭМ сверхвысокого разрешения и предположили, что волокнистые отростки находились на внешнем слое Nannochloropsis gaditana , родственного вида водорослей.Однако, по нашим данным, после разрыва покровной мембраны фиброзные расширения обнажаются. Фактически, покровная мембрана также видна на изображениях Cryo-EM, сделанных Scholz et al., 2014. Авторы, возможно, пропустили мембрану как часть клеточной стенки Noc из-за того, как они обрабатывали и готовили свои образцы (рис. 2B). и C в Scholz et al., 2014). Дальнейшие исследования N. gaditana также показали, что клетки имеют гладкую поверхность с помощью SEM (Jazzar et al., 2015).
Мы согласны с комментариями рецензента по легенде и также изменили терминологию как «агрегирование», а не захваченные ячейки.
4) Агрегация клеток водорослей. Имеются сообщения, демонстрирующие, что N.oceanica может легко агрегировать в присутствии бактериальных штаммов или биофлокулянтов (Wang et al., 2012). В некоторых случаях активные молекулы, которые действуют как биофлокулянты, были идентифицированы (Wan et al., 2012) как протеогликаны.
Поскольку данные по геномике и метаболомике доступны для Mortierella elongata, могут ли Авторы предоставить некоторую экспериментальную поддержку агрегации, которую они описывают? Могут ли они провести эксперимент с течением времени? Автор может лечить Nocs экссудатами Mortierella, просто чтобы посмотреть, происходит ли агрегация.
Мы благодарим рецензента за эти предложения. Мы думаем, что важные открытия в этой рукописи — это детали физического взаимодействия клеточной стенки и взаимопонимания между водорослями и грибами, а также долгосрочное совместное культивирование, которое привело к образованию зеленых гиф. Что касается флокуляции водорослей грибами, мы недавно опубликовали способность Mortierella к биофлокуляции Nannochloropsis как эффективный, экономичный и экологически безопасный подход к сбору микроводорослей по сравнению с традиционными методами сбора, такими как центрифугирование и химическая флокуляция. .Мы предоставляем данные, включая результаты краткосрочной флокуляции и профилирования липидов / жирных кислот, а также другие данные о продуктивности нефти для биотехнологических целей (Du et al., 2018). В документе Biotechnology for Biofuels не обсуждались никакие аспекты обмена питательными веществами или зеленые гифы, наблюдаемые после длительной инкубации, поскольку эти данные являются сутью этого представления. Определение состава внешней стенки водорослей и средств, с помощью которых Mortierella влияет на это, является частью более крупного текущего проекта, поддерживаемого JGI, и выходит за рамки настоящего исследования.
5) Эксперименты по переносу питательных веществ хорошо разработаны и точно описаны. В дополнительном материале содержится много интересных деталей. Однако некоторые аспекты до конца не ясны. Эксперименты с питательными веществами: Рисунок 2 непросто интерпретировать. Если я хорошо понимаю рис. 2А (слева), только уменьшенное количество меченого углерода перемещается к грибку (менее 1 dpm / мг), в то время как в тексте это описывается как релевантное количество. Какой термин сравнения используется для определения «релевантного» значения радиоактивности? Является ли разница между меченой глюкозой, обнаруженной в прикрепленных и свободных клетках Noc, значительно? На панели C тот же эксперимент повторяется, но значение радиоактивности у гриба намного выше (12,7%).Кроме того, почему N экспериментов представлены по-разному?
Прежде всего, благодарим вас за комментарий и постарались более четко описать представленные данные и цифры. Одним из ключевых моментов является то, что большая часть перенесенного 14 C-углерода оказалась в липидной фракции грибковых клеток. Более поздние относительные значения на фиг.2, панели C и D, представляют собой общее перенесенное 14 C-углерода в различных типах образцов, таких как немеченые грибы, при физическом контакте с мечеными водорослями (фиг.2, панель C # 1, 12.7%) и с физическим контактом с водорослями, убитыми нагреванием (Рисунок 2, панель C # 3, 1,3%) по сравнению с общей радиоактивностью предварительно меченных образцов, таких как 14 C-меченых водорослей (Рисунок 2, панель C, 100 %). Эти анализы были нацелены на то, чтобы проверить, важны ли физический контакт и живые клетки для переноса углерода 14 . Мы изменили текст и рисунок, чтобы облегчить понимание. Мы добавляем «100%» после 14 C-Noc и 14 C-AG77 на рис. 2, панели C и D.
По сравнению с экспериментами по обмену углерода, в которых используется 14 C-углерод, который можно измерить сцинтилляционным счетчиком и легко представить, эксперимент по обмену азота проводился с использованием 15 N, стабильного нерадиоактивного изотопа азота в сочетании масс-спектрометрия изотопного отношения для расчета [ 15 N / ( 15 N + 14 N), моль / моль]. По общему признанию, последние эксперименты более сложны, чем результаты углеродной маркировки (Рисунок 2 — приложение к рисунку 3J и K).Чтобы сделать эти результаты более понятными для широкой аудитории, мы суммируем результаты в виде простого рисунка (рис. 2, панель E) и предоставляем более подробную информацию в дополнительных файлах.
6) Механизмы переноса углерода. Результаты ясно показывают, что углерод перемещается от водорослей к грибам. Дает ли геном Mortierella какое-то предположение о лежащем в основе механизме? Наличие переносчика глюкозы? С другой стороны, результаты, показывающие перемещение углерода от гриба к водорослям даже при отсутствии физического контакта, очень трудно понять.Noc является фотосинтетическим: влияют ли условия окружающей среды на его фотосинтетическую активность? проверили ли Авторы некоторые параметры фотосинтеза в этих условиях?
Мы согласны с тем, что это интересные и очевидные оставшиеся вопросы, но решили, что рассмотрение механизма транспорта углерода должно быть следующим шагом, требующим дополнительных метаболических, транскриптомных экспериментов и системы трансформации гриба. Мы считаем, что эти эксперименты выходят за рамки текущей рукописи, которая уже содержит большое количество исходных данных.
Что касается переноса углерода от грибов к водорослям, мы сообщаем, что грибов Mortierella выделяют органический углерод и азот в окружающую среду / среду, когда они инкубируются отдельно в среде f / 2 (рис. 3D и E). Это позволит непривязанным водорослям получать углерод и азот от совместно культивируемых грибов (рис. 2).
Условия окружающей среды обязательно влияют на фотосинтез. Мы проанализировали содержание хлорофилла, которое можно экстрагировать с помощью растворителей ацетон: ДМСО, поскольку это может быть использовано в качестве показателя фотосинтетического потенциала.Содержание хлорофилла снижается, когда клетки Noc подвергаются стрессу. Мы действительно наблюдали значительно более высокое содержание хлорофилла в клетках Noc, совместно культивируемых с грибами AG77, чем в одних только клетках Noc, служащих контролем, после 10-дневной продолжительной инкубации в среде f / 2, что указывает на то, что клетки Noc с грибами Mortierella обладают более высоким фотосинтетическим потенциалом (Рисунок 3 — рисунок в приложении 1A).
7) Условия питания: Noc реагирует на краткосрочное и долгосрочное голодание по азоту, активируя специфические молекулярные ответы (Dong et al., 2012). Проверяли ли авторы эти механизмы восстановления, когда клетки Noc поддерживаются в присутствии Mortierella?
Мы наблюдали повышенную жизнеспособность клеток Noc после длительного (10 дней) голодания по N в присутствии грибов Mortierella (рис. 3A – C), которые выделяют органический углерод и азот в среду (рис. 3D и E). Клетки Noc без Mortierella были значительно более подвержены стрессу, о чем свидетельствует деградация хлорофилла после продолжительной инкубации (Рисунок 3 — рисунок в приложении 1A).
8) Эксперимент со специфичностью. Этот эксперимент очень интересен, учитывая также отчеты, которые показывают, как бактерии могут агрегировать маленькие клетки Noc (см. Предыдущий комментарий). На мой взгляд, некоторая информация отсутствует: каково поведение исходного штамма Mortierella, штамма, содержащего Mycoavidus? проверили ли Авторы лихенизированные грибы? Было бы интересно посмотреть на поведение Rhizopus, родственного Mucoromycota, который является хозяином B. rhizoxinica.Эта бактерия проникает внутрь гиф Rhizopus в соответствии с модальностями, которые напоминают описанные для Mortierella-Noc, используя хитиназу для разрушения грибковой стенки на кончике (Moebius et al., 2014).
Кроме того, я бы прокомментировал отрицательные результаты по Saccharomyces. Хом и Мюррей, 2014 ясно демонстрируют, что взаимодействие между дрожжами и водорослями зависит от условий окружающей среды! окружающая среда (например, недостаток питательных веществ) является движущей силой мутуалистической ассоциации.В этом контексте авторам следует также рассмотреть очень интересные результаты, проиллюстрированные в Li Chien et al., 2017, где была разработана синтетическая платформа путем связывания различных дрожжей с фотосинтетическими цианобактериями.
Мы благодарим рецензента за эти предложения. Мы протестировали Mortierella elongata AG77 дикого типа, который несет эндобактерию Mycoavidus cysteinexigens . Мы наблюдали значительно сниженную жизнеспособность клеток Noc с WT AG77 по сравнению с вылеченным AG77, а также серьезную деградацию хлорофилла и накопление ROS в клетках Noc при совместном культивировании с AG77 WT.Однако мы повторили эксперименты несколько раз, и результаты не были согласованными. Это могло произойти из-за колебаний активности бактерий. Интересно, что присутствие эндобактерий усложняет симбиоз и не является предметом настоящего исследования. Таким образом, мы удалили данные WT AG77 и пересмотрели раздел «Обсуждение». Хотя мы не проводили специального скрининга на Rhizopus или лишайниковые грибы, мы провели скрининг разнообразных по филогенезу грибов, включая родственников этих организмов.Мы обнаружили, что указанный здесь фенотип уникален для Mortierella .
9) Подраздел «Длительное совместное культивирование приводит к интернализации N. oceanica внутри M. elongata hyphae»: WGA — это лектин, который, как ожидается, будет связываться с N-ацетилглюкозамином, мономером хитина. Содержит ли Noc этот грибковый компонент стенок?
Да. Сообщалось, что Noc содержит N-ацетилглюкозамин в клеточной стенке (Scholz et al., 2014), и мы действительно знаем, что Mortierella эффективно разрушает этот субстрат.
10) Глядя на конфокальные изображения, (Рисунок 4 — Приложение 1 к рисунку) нет сомнений в том, что WGA окрашивает стенки грибов, но стенки водорослей не помечены (Рисунок 4 — Приложение 1 к рисунку, панель B), только разделение перегородка между двумя делящимися клетками показывает флуоресценцию. Напротив, красная флуоресценция хлорофилла четко позволяет идентифицировать как изолированные клетки водорослей, так и клетки гиф. На последней панели (справа) хлоропласт четко виден внутри гифы, что свидетельствует об интернализации водорослей.Предлагаю переписать описание.
Мы благодарим рецензента за эти предложения. Рисунок 4 — дополнение к рисунку 1B — это не просто обычное конфокальное изображение, сфокусированное на одной панели. Это совмещенное конфокальное изображение, которое показывает трехмерное изображение сигналов WGA и хлорофилла. Вот почему сигнал WGA распространяется по всем ячейкам, а не по кругу. Разделительная перегородка показала более сильный сигнал. Мы снизили усиление WGA, чтобы показать внутренний сигнал хлорофилла.В противном случае это будут ярко-зеленые клетки. У нас есть видео вращающейся 3D-модели, чтобы показать, как совмещены конфокальные изображения, а также для рисунка 4 — приложение к рисунку 1С (видео 2). Мы переписали описание по запросу.
11) Рисунок 4, вероятно, необходимо реорганизовать, чтобы облегчить чтение: сначала изображения ДИК, четко показывающие кластер водорослей внутри на кончике гифы, а затем несколько изображений ПЭМ, выбирая лучшее: я бы предложил Рисунок 4 — дополнение к рисунку 2, первые два изображения слева, так как гриб кажется живым..На всех остальных изображениях гифы пусты: нет органелл, нет мембран, что говорит о том, что водоросли входят в пустую нишу … В отличие от этого рисунка 4 — рисунка, добавленного к рисунку 2, кажется, что некоторые грибковые мембраны присутствуют. А затем увеличенный рисунок 4 панели C, чтобы показать ультраструктуру Noc. Однако снимки не решают вопрос, окружены ли клетки Noc грибковой мембраной. Опять же, авторы могут проверить свои изображения с водорослями, живущими внутри одноклеточных протистов.Например, красивые картинки из Song et al., (2017).
Благодарим рецензента за предложения. Сначала мы заменили изображения ПЭМ на рис. 4 на изображения, показывающие, что интактные клетки водорослей находятся внутри гифа гриба и окружены грибковыми мембранами и органеллами (AG77 и Noc, рис. 4F и G; NVP64 и Noc, рис. 4 — дополнение к рисунку). 2С). Предыдущие ПЭМ-изображения, показывающие клетки водорослей внутри гифы гриба с четкой ультраструктурой хлоропласта и некоторых других органелл водорослей, были перенесены на Рисунок 4 — приложение к рисунку 2B.Клетки водорослей имеют уникальные и различимые хлоропластные и тилакоидные мембраны. Для грибковых органелл мы включаем ТЕА-контроль грибов, инкубированных отдельно в среде f / 2, для сравнения органелл водорослей и грибов (Рисунок 4 — рисунок в приложении 2A). Некоторые грибковые мицелии становятся полыми после жестких этапов подготовки образца ТЕА, как и другие изображения совместно культивируемых грибковых тканей.
К сожалению, с Cyro-EM нам не удалось получить изображения с более высоким разрешением, чтобы определить, окружает ли грибковая мембрана клетки водорослей.Однако мы использовали световую ДИК-микроскопию и конфокальную микроскопию с SYTOX green и продемонстрировали, что клетки водорослей живут внутри живых клеток грибов. Рис. 4 — дополнения к рисункам 5A – C — типичный пример того, что и водоросли, и гифы живы. Зонд SYTOX проникает только в мертвые клетки, и сигнал очевиден в контроле мертвых клеток (Рисунок 4 — приложение к рисунку с 5D по H).
Затем мы использовали световую микроскопию DIC и наблюдали, что клетки водорослей внутри гифов грибов окружены грибковыми пузырьками и липидными капельками, которые обычно присутствуют в живых клетках (Рисунок 4 — приложение к рисунку 6).Учитывая, что образцы обычно не находятся на одной и той же фокальной панели при большом увеличении, мы предоставляем видео, чтобы показать трехмерное содержание зеленых гиф, и видео 5 является репрезентативным.
Рецензент № 3:
Эта рукопись документирует очень захватывающее открытие: эндосимбиоз клеток водорослей Nannochloropsis грибком Mortierella в Mucoromycota, типе, который подвергается более тщательному изучению в связи с эволюцией ассоциаций растений и грибов. Текст хорошо написан, а описанные эксперименты убедительны и демонстрируют внутриклеточную ассоциацию водорослей (с помощью световой и электронной микроскопии) и обмен углерода и азота между водорослями и грибами.Хотя механизмы, лежащие в основе (вертикальной?) Передачи и поддержания этого нового «индуцированного» симбиоза совместной культуры, не исследуются, эта работа создает основу для дальнейших исследований.
Существенные изменения:
1) В аннотации написано: «Этот симбиоз начинается с хемотаксического влечения…». Мне не ясно, какие доказательства есть в пользу этого утверждения. Основано ли это утверждение на агрегации клеток водорослей на концах гиф грибов? Можно ли с уверенностью заключить, что на этом основании было хемотаксическое влечение?
Мы перефразировали это.Взаимодействие Nannochloropsis-Mortierella начинается с флокуляции клеток водорослей мицелием и потери покрытия внешней клеточной стенки фотобионтом водорослей.
2) Можно ли предоставить серию изображений или видеоролик, показывающий, как плотная кластеризация клеток водорослей на концах гиф изменяется или прогрессирует с течением времени (раздел для обсуждения)? Возможно, от более низкой плотности к более высокой плотности?
Это отличный момент. Мы приложили огромные усилия, чтобы попытаться решить эту проблему, но это серьезная проблема.Мы можем наблюдать зеленые гифы при длительном совместном культивировании, а зеленые гифы различимы под микроскопом DIC (рис. 4B – E, рис. 4 — приложение к рисунку 4). Однако зафиксировать прогресс через покадровую съемку крайне сложно. Это связано с тем, что процесс длится несколько недель, а фокальная плоскость сильно ограничена при световой микроскопии. Учитывая, что это живая растущая система (3D), мы не можем предсказать, где в колонии будут развиваться эти гифы и на какой точке трехмерной колонии сосредоточиться в течение нескольких дней.Например, рисунки 4-S3C-F были сняты как покадровые изображения за 3 дня в случайно выбранной области. Чтобы это увидеть, нам потребовались тысячи часов микроскопического времени. Еще больше усложняет ситуацию то, что водоросли и грибы образуют агрегаты после совместного культивирования (Рисунок 4 — рисунок в приложениях 1D и E), и если количество клеток слишком велико, видимость затрудняется. Тем не менее, нам очень повезло записать серию изображений, показывающих группу клеток водорослей, прикрепленных к кончику гифы, растущую внутри гиф (Рисунок 4 — приложение к рисунку 3A и B).
3) Ссылка на видео 4 (подраздел «Долгосрочное совместное культивирование приводит к интернализации N. oceanica в гифах M. elongata») как свидетельство того, что клетки водорослей и грибов могут проходить через фрагментацию и оставаться жизнеспособными. Это видео показывает очаговую гифу с эндосимбиотическими водорослями (в контексте более крупной «зеленой ткани») за 6 дней (не месяцы совместного культивирования), но не показывает какой-либо фрагментации и / или пассирования. Был загружен не тот файл? Есть ли другое видео или набор рисунков / изображений, которые действительно можно использовать для подтверждения заявленного утверждения?
Рецензент прав.Видео 4 показывает рост и деление клеток водорослей в гифах грибов, а не их фрагментацию. Однако совместное культивирование на видео было фрагментировано (вегетативное размножение) из более старой материнской колонии, как это обычно делается с грибковым мицелием в лаборатории. Мы изменили рукопись, чтобы она гласила: «Хотя нет никаких указаний на то, что водоросли передаются вертикально через репродуктивные структуры грибов, клетки водорослей остаются жизнеспособными (растут и делятся) в течение нескольких месяцев совместного культивирования (видео 4)».
[Примечание редакции: ответы на повторную проверку приведены ниже.]
Существенные изменения:
Рисунок 4G. Авторы умерили свои комментарии о грибковых оболочках, и мы это ценим. Однако изображения ПЭМ (рис. 4) не решают полностью вопрос: окружены ли клетки водорослей мембраной, полученной из грибов? Мы считаем, что этого нельзя сделать на основании представленных данных. Во многих случаях очевидно, что гифы Mortierella пусты, и тот факт, что грибок растет, когда помещается в новую чашку, просто означает, что некоторые гифы живы и способны возобновить свой рост.Но это не обязательно гифы с Noc. Итак, если у вас нет данных, подтверждающих это, мы просим вас удалить предложение, в котором говорится, что «предполагаемые мембраны грибов окружают интернализованные клетки водорослей».
Мы удалили предложение.
Мы очень ценим новые изображения сканирования, показывающие структуру клеточной стенки Nannochloropsis. И действительно, они показывают, что одна только водоросль имеет гладкую поверхность, как сообщается в литературе. Однако, чтобы сделать вывод о наличии выступов под гладким внешним покрытием, необходима просвечивающая электронная микроскопия поперечных сечений.Возможно, что расширения были вызваны контактом с грибком. Следовательно, мы просим изменить выводы и обсудить этот вопрос (если вы не можете включить поперечные сечения, на которых видны выступы под слоем покрытия).
Мы добавили пояснение к разделу «Результаты» следующим образом: «Хотя возможно, что волокнистые расширения были вызваны контактом с грибком, в наших наблюдениях очевидны остатки внешнего покрытия, покрывающие нижележащие расширения, как показано на Рисунки 1E и F и рисунок 1 — дополнение к рисунку 1D.Поэтому кажется вероятным, что эти расширения присутствуют под внешним гладким слоем ».
Abstract: Первое предложение не имеет смысла (… совместная эволюция наземных растений и лишайников). Реферат был исправлен после предыдущего предложения в некоторых документах, но не в PDF «объединенной новой версии». Кроме того, я считаю, что термин «коэволюция» требует двух членов. (например, совместная эволюция растений и грибов, грибов и насекомых). В заключение: предложение нужно переписать.
Мы согласны с рецензентами и заменили «коэволюцию» на «эволюцию».
Раздел для обсуждения: кончики гиф относятся к наименее развитым тканям. Обратите внимание, что ткань означает: группы клеток, которые имеют схожую структуру и происхождение и действуют вместе для выполнения определенной функции. Следовательно, термин «ткань» нельзя использовать для обозначения части клетки (в данном случае гифы). В этом контексте я бы написал: кончики гиф являются наименее дифференцированными частями мицелиальной сети
Мы согласны, и предложение было переписано, как было предложено.
Подраздел «Дефицит питательных веществ и преимущества совместного культивирования N. oceanica и M. elongate»: мы думаем, что где-то здесь уместно признать, что маркер линоленовой кислоты для биомассы был стандартизован в полных условиях монокультур. Авторы не исключают возможность того, что соответствие маркера биомассе может нарушиться в условиях совместного культивирования, тем более что ремоделирование липидного состава, безусловно, возможно и не является беспрецедентным в симбиозах (например.g., между растениями и арбускулярными микоризными грибами). Мы считаем правильным добавить простое и честное предложение, в котором говорится, что незначительные изменения C18: 3 по сравнению с биомассой предполагаются для любых физиологических изменений, которые могут возникнуть у M. elongata в сокультуре. (Я думаю, что это, вероятно, несущественно, но следует быть точным, а не строить предположения.) Это относится и к рисунку 3 — дополнению к рисунку 2.
Мы согласны и добавили обсуждение, как было предложено. Подраздел «Дефицит питательных веществ и преимущества совместного культивирования для N.oceanica и M. elongata ».
Эксперименты по обмену питательных веществ красиво выполнены и представлены, но обсуждение их биологического значения отсутствует. M.elongata — сильный сапротроф, которому не нужен углерод, поступающий из водорослей. С другой стороны, он также выделяет N в водоросли. Итак, в чем польза грибка? Единственное преимущество, по-видимому, заключается в увеличении биомассы грибов; однако это зависит от указанного выше момента. Количественное определение C18: 3 может отражать другой метаболизм (запас липидов) в большей степени, чем активный рост.Пожалуйста, добавьте точку обсуждения, включая условия C и N среды, в которой проводились эти эксперименты.
Совместное культивирование экспериментов по обмену питательных веществ проводили в среде f / 2, содержащей 14 N (2,5 мМ NaNO 3 ) и 12 C (20 мМ NaHCO 3 и окружающий CO 2 ). Таким образом, водоросли должны быть единственным источником органического углерода для гриба. Поскольку M. elongata является сильным сапротрофом, мы проверили, погибла ли водоросль (а также грибок) в совместной культуре (Рисунок 2 — рисунок в приложении 3), и мы также кормили гриб убитым нагреванием 14 C-меченой водоросли и скармливали водоросли убитым нагреванием 14 C-меченым грибком.Эти результаты суммированы на Рисунке 2C и обсуждаются в подразделе «Перенос углерода и азота между N. oceanica и M. elongata ».
Подраздел «Долгосрочное совместное культивирование приводит к интернализации N. oceanica в M. elongata hyphae»: что касается водорослей, находящихся внутри живых грибов, как вы можете быть уверены, что они действительно живы, основываясь только на этих данных? Свободные / внеклеточные водоросли могут быть доступны для SYTOX Green, но водоросли в живых грибах могут никогда не подвергаться воздействию SYTOX Green, да? (Живые грибы исключили бы это, поэтому невозможно определить внутриклеточную жизнеспособность водорослей.Мы не считаем это важным моментом, потому что данные ЭМ показывают, что водоросли живы, но может оказаться невозможным определить истинную жизнеспособность внутриклеточных водорослей с помощью исключающих красителей, и это следует признать.
Отличная точка зрения, и мы признаем недостатки этого эксперимента. Наше предположение о том, что водоросли внутри гриба живы, в основном основано на том факте, что они делятся, и в некоторой степени на том, что они представляют собой флуоресценцию хлорофилла, что в конечном итоге указывает на собранную фотосинтетическую мембрану.
Реферат — Долгосрочное совместное культивирование — это субъективный термин. Здесь следует указать продолжительность наблюдаемого времени.
Добавлено время: «больше месяца».
Рисунок 3 — дополнение к рисунку 2 — Опять же, следует добавить подробности использованных статистических тестов.
Добавлены детали статистических тестов.
Рисунок 4 G -Что такое Mo? Его нет в легенде.
Mo означает « органелл Mortierella, ».Это было добавлено в условных обозначениях рисунка 4G и рисунка 4 — дополнение к рисунку 2.
Подраздел «Долгосрочное совместное культивирование приводит к интернализации N. oceanica в M. elongata hyphae»: мы предлагаем изменить формулировку, чтобы быть более консервативной: «Результаты согласуются с представлением о том, что и гриб-хозяин, и водоросли внутри живы. (Рисунок 4 — рисунок в приложении 5), хотя ДИК-микроскопия… окружена грибковыми органеллами и чем-то вроде липидных капель… »
Изменено «соответствует» для «показано», добавлено «хотя» перед «микроскопией ДИК», удалено «» перед «грибковыми органеллами» и изменено «и то, что кажется» вместо «например».Капельки липидов не являются грибковыми органеллами…. В качестве примечания: как узнать, что это липидные капли? Они окрашивают Nile Red? Это заявлено, но не обосновано.
Мы внесли поправки в текст. Мы окрашивали и визуализировали липидные капли с помощью BODIPY с помощью конфокальной микроскопии (рисунок 4 — приложение к рисунку 6). Капли липидов различимы по цвету (сине-зеленый) и размеру под световым микроскопом.
Раздел обсуждения: Мы предлагаем изменить формулировку, чтобы быть более консервативной: «зеленые гифы кажутся живыми через 2 месяца…».Заменено «оставаться» на «осталось».
С внесенными в него поправками.
Пожалуйста, укажите характер использованных статистических тестов в условных обозначениях и / или в разделе «Материалы и методы».
Статистические тесты были добавлены в условные обозначения, как было предложено.
https://doi.org/10.7554/eLife.47815.031грибов | Определение, характеристики, типы и факты
Грибок , множественное число грибов , любой из примерно 144 000 известных видов организмов царства грибов, включая дрожжи, ржавчину, головню, плесень, плесень и грибы.Есть также много грибоподобных организмов, включая слизевики и оомицеты (водяные формы), которые не принадлежат к царству грибов, но часто называются грибами. Многие из этих грибовидных организмов входят в королевство Хромиста. Грибы являются одними из наиболее широко распространенных организмов на Земле и имеют большое экологическое и медицинское значение. Многие грибы свободно живут в почве или воде; другие образуют паразитические или симбиотические отношения с растениями или животными.
Популярные вопросы
Как грибы получают питание?
- Сапротрофные грибы получают пищу из мертвого органического материала и являются экологически полезными деструкторами.
- Паразитические грибы питаются живыми организмами (обычно растениями), вызывая болезни.
- Для питания оба типа грибов выделяют пищеварительные ферменты на питательную поверхность, на которой они растут. Ферменты расщепляют углеводы и белки, которые затем всасываются через стенки гиф.
- Некоторые паразитические грибы также производят специальные поглощающие органы, называемые гаусториями, для более глубокого проникновения в живые ткани хозяина.
Что такое спора гриба?
Почти все грибы образуют и выделяют огромное количество спор в рамках своего жизненного цикла.Споры являются основными репродуктивными единицами грибов и обычно представляют собой одиночные клетки. Они могут быть произведены либо напрямую бесполым путем, либо косвенно половым размножением. Споры обычно образуются в результате фрагментации мицелия или внутри специализированных структур (спорангии, гаметангии, спорофоры и т. Д.). Некоторые споры, особенно у примитивных грибов, имеют жгутики и могут плавать, хотя большинство из них неподвижны. Когда спора попадает в подходящее место, она прорастает и растет, образуя новую грибковую особь.
Где растут грибы?
Грибы растут в самых разных средах по всему миру. Большинство грибов являются наземными и встречаются во всех регионах с умеренным и тропическим климатом. Некоторые виды обитают в арктических и антарктических регионах, обычно в составе лишайников. Почва, богатая органическими веществами, является идеальной средой обитания для многих видов, и лишь небольшое количество грибов встречается в более засушливых районах или в местах обитания с небольшим содержанием органических веществ или без них. Некоторые грибы паразитируют на растениях или животных и живут на своих хозяевах или внутри них, по крайней мере, часть своего жизненного цикла.Водные грибы обычно обитают в чистой прохладной пресной воде, хотя некоторые виды обитают в слегка солоноватой воде, а некоторые процветают в сильно загрязненных ручьях.
Грибы — эукариотические организмы; т.е. их клетки содержат связанные с мембраной органеллы и четко определенные ядра. Исторически грибы были включены в царство растений; однако, поскольку в грибах отсутствует хлорофилл и они отличаются уникальными структурными и физиологическими особенностями (т.е. компонентами клеточной стенки и клеточной мембраны), они были отделены от растений.Кроме того, грибы четко отличаются от всех других живых организмов, включая животных, по их основным способам вегетативного роста и потребления питательных веществ. Грибы растут из кончиков нитей (гиф), составляющих тела организмов (мицелий), и они переваривают органическое вещество извне, прежде чем впитать его в свой мицелий.
Хотя грибы и поганки (ядовитые грибы) ни в коем случае не являются самыми многочисленными или экономически значимыми грибами, их легче всего распознать.Латинское слово, обозначающее гриб, гриб (множественное число гриб ) стало обозначать всю группу. Точно так же изучение грибов известно как микология — широкое применение греческого слова, обозначающего гриб, mykēs . Грибы, отличные от грибов, иногда собирательно называют плесенью, хотя этот термин лучше ограничивать грибами, представляющими собой плесень для хлеба. (Для получения информации о слизистых плесени, которые проявляют черты как животного, так и грибного мира, см. protist.)
Белые грибыСъедобные белые грибы ( Boletus edulis ). Белые грибы широко распространены в Северном полушарии и образуют симбиотические ассоциации с рядом древесных пород.
© Henk Bentlage / Fotolia24.2B: Взаимоотношения с грибами и грибными животными
Члены Королевства грибов формируют экологически выгодные взаимные отношения с цианобатериями, растениями и животными.
Задачи обучения
- Описать мутуалистические отношения с грибами
Ключевые моменты
- Взаимоотношения — это отношения, в которых выигрывают оба члена ассоциации; Грибы образуют такие отношения с различными другими Царствами жизни.
- Микориза, образованная в результате ассоциации корней растений и примитивных грибов, способствует увеличению усвоения растениями питательных веществ; в свою очередь, растение снабжает грибы продуктами фотосинтеза для их метаболического использования.
- В лишайниках грибы живут в непосредственной близости с фотосинтетическими цианобатериями; водоросли снабжают грибы углеродом и энергией, а грибы снабжают водоросли минералами и защитой.
- Взаимоотношения между грибами и животными связаны с многочисленными насекомыми; Членистоногие зависят от грибов для защиты, в то время как грибы получают взамен питательные вещества и обеспечивают способ распространения спор в новую среду.
Ключевые термины
- микориза : симбиотическая ассоциация между грибком и корнями сосудистого растения
- лишайник : любой из многих симбиотических организмов, являющихся ассоциациями грибов и водорослей; часто встречается в виде белых или желтых пятен на старых стенах и т. д.
- слоевище : вегетативное тело гриба
Взаимоотношения
Симбиоз — это экологическое взаимодействие двух организмов, живущих вместе.Однако определение не описывает качество взаимодействия. Когда выигрывают оба члена ассоциации, симбиотические отношения называются мутуалистическими. Грибы образуют мутуалистические ассоциации со многими типами организмов, включая цианобактерии, растения и животных.
Мутуализм грибов и растений
Mycorrhiza, происходящее от греческих слов «мико», что означает гриб, и «ризо», что означает корень, относится к ассоциации между корнями сосудистых растений и их симбиотическими грибами.Около 90 процентов всех видов растений имеют микоризных партнеров. В микоризной ассоциации грибной мицелий использует свою обширную сеть гиф и большую площадь поверхности в контакте с почвой для направления воды и минералов из почвы в растения, тем самым увеличивая поглощение растениями питательных веществ. Взамен растение поставляет продукты фотосинтеза, которые подпитывают метаболизм гриба.
Микоризы обладают многими характеристиками примитивных грибов: они образуют простые споры, мало разнообразны, не имеют полового репродуктивного цикла и не могут жить вне микоризной ассоциации.Существует ряд видов микоризы. Эктомикориза («внешняя» микориза) зависит от грибов, окружающих корни оболочкой (называемой мантией), и сети гиф Хартига, которая простирается в корни между клетками. Грибковый партнер может принадлежать к Ascomycota, Basidiomycota или Zygomycota. Во втором типе грибы Glomeromycete образуют везикулярно-арбускулярные взаимодействия с арбускулярной микоризой (иногда называемой эндомикоризой). В этой микоризе грибы образуют арбускулы, которые проникают в клетки корня и являются местом метаболического обмена между грибом и растением-хозяином.Арбускулы (от латинского «маленькие деревья») имеют вид кустарника. Орхидеи полагаются на третий тип микоризы. Орхидеи — это эпифиты, которые образуют маленькие семена без особого хранения для поддержания прорастания и роста. Их семена не прорастут без микоризного партнера (обычно базидиомицета). После того, как питательные вещества в семенах истощаются, грибковые симбионты поддерживают рост орхидеи, обеспечивая необходимые углеводы и минералы. Некоторые орхидеи продолжают оставаться микоризными на протяжении всего своего жизненного цикла.
Рисунок \ (\ PageIndex {1} \): Микоризные грибы : (а) эктомикориза и (б) арбускулярная микориза имеют разные механизмы взаимодействия с корнями растений.Лишайники
Лишайники имеют разные цвета и текстуры. Они могут выжить в самых необычных и враждебных местах обитания. Они покрывают камни, надгробия, кору деревьев и землю в тундре, куда не могут проникнуть корни растений. Лишайники могут пережить длительные периоды засухи: они полностью высыхают, а затем быстро становятся активными, когда вода снова становится доступной.Лишайники выполняют множество экологических функций, в том числе выступают в качестве видов-индикаторов, которые позволяют ученым отслеживать состояние среды обитания из-за их чувствительности к загрязнению воздуха.
Рисунок \ (\ PageIndex {1} \): Лишайники: грибы и цианобатерии. : Лишайники имеют множество форм. Они могут быть (а) корковидными, (б) волосковыми или (в) листовыми.Лишайники — это не единый организм, а, скорее, пример мутуализма, в котором гриб (обычно член типов Ascomycota или Basidiomycota) живет в тесном контакте с фотосинтезирующим организмом (эукариотическая водоросль или прокариотическая цианобактерия).Как правило, ни грибок, ни фотосинтезирующий организм не могут выжить в одиночку вне симбиотических отношений. Тело лишайника, называемое слоевищем, состоит из гиф, обернутых вокруг фотосинтетического партнера. Фотосинтезирующий организм обеспечивает углерод и энергию в виде углеводов. Некоторые цианобактерии фиксируют азот из атмосферы, внося в ассоциацию азотистые соединения. В свою очередь, гриб поставляет минералы и защищает от сухости и чрезмерного света, заключая водоросли в свой мицелий.Грибок также прикрепляет симбиотический организм к субстрату.
Рисунок \ (\ PageIndex {1} \): Слоевище лишайника : Это поперечное сечение слоевища лишайника показывает (а) верхнюю часть коры гиф грибка, которая обеспечивает защиту; (б) зона водорослей, где происходит фотосинтез, (в) мозговое вещество гиф грибов и (г) нижняя кора головного мозга, которая также обеспечивает защиту и может иметь (д) ризины для прикрепления слоевища к субстрату.Слоевище лишайников растет очень медленно, увеличивая свой диаметр на несколько миллиметров в год.И гриб, и водоросль участвуют в формировании единиц распространения для размножения. Лишайники производят соредии, скопления клеток водорослей, окруженные мицелием. Соредии разносятся ветром и водой и образуют новые лишайники.
Грибки и мутуализм животных
Грибы развили взаимопонимание с многочисленными насекомыми. Членистоногие (членистоногие беспозвоночные, такие как насекомые) зависят от грибка для защиты от хищников и патогенов, в то время как гриб получает питательные вещества и способ распространения спор в новую среду.Одним из примеров является связь между видами Basidiomycota и щитовками. Грибной мицелий покрывает и защищает колонии насекомых. Щитовки способствуют поступлению питательных веществ от зараженного растения к грибку. Во втором примере муравьи-листорезы Центральной и Южной Америки буквально выращивают грибы. Они срезают с растений диски из листьев и складывают их в садах. В этих дисковых садах выращивают грибы, которые переваривают целлюлозу в листьях, которую муравьи не могут разрушить.Когда более мелкие молекулы сахара производятся и потребляются грибами, они, в свою очередь, становятся пищей для муравьев. Насекомые также патрулируют свой сад, охотясь на конкурирующие грибы. От ассоциации выигрывают и муравьи, и грибы. Гриб получает постоянный запас листьев и свободу от конкуренции, в то время как муравьи питаются грибами, которые они культивируют.
ЛИЦЕНЗИИ И АТРИБУТЫ
CC ЛИЦЕНЗИОННЫЙ КОНТЕНТ, ПРЕДЫДУЩИЙ РАЗДЕЛ
- Курирование и проверка. Предоставлено : Без границ.com. Лицензия : CC BY-SA: Attribution-ShareAlike
CC ЛИЦЕНЗИОННОЕ СОДЕРЖАНИЕ, СПЕЦИАЛЬНАЯ АТРИБУЦИЯ
- Колледж OpenStax, Биология. 22 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44632/latest…ol11448/latest . Лицензия : CC BY: Attribution
- Колледж OpenStax, Биология.12 ноября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44632/latest…ol11448/latest . Лицензия : CC BY: Attribution
- экзофермент. Предоставлено : Викисловарь. Расположен по адресу : en.wiktionary.org/wiki/exoenzyme . Лицензия : CC BY-SA: Attribution-ShareAlike
- декомпозитор. Предоставлено : Викисловарь. Расположен по адресу : en.wiktionary.org/wiki/decomposer . Лицензия : CC BY-SA: Attribution-ShareAlike
- сапроб. Предоставлено : Викисловарь. Расположен по адресу : en.wiktionary.org/wiki/saprobe . Лицензия : CC BY-SA: Attribution-ShareAlike
- Колледж OpenStax, Биология.12 ноября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44632/latest…ol11448/latest . Лицензия : CC BY: Attribution
- Колледж OpenStax, Биология. 12 ноября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44632/latest…ol11448/latest . Лицензия : CC BY: Attribution
- Безграничный. Предоставлено : Безграничное обучение. Расположен по адресу : www.boundless.com//biology/definition/thallus . Лицензия : CC BY-SA: Attribution-ShareAlike
- Колледж OpenStax, Биология. 17 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44632/latest…ol11448/latest . Лицензия : CC BY: Attribution
- Колледж OpenStax, Биология.22 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44632/latest…ol11448/latest . Лицензия : CC BY: Attribution
- микориза. Предоставлено : Википедия. Расположен по адресу : en.Wikipedia.org/wiki/mycorrhiza . Лицензия : CC BY-SA: Attribution-ShareAlike
- лишайник. Предоставлено : Викисловарь. Расположен по адресу : en.wiktionary.org/wiki/lichen . Лицензия : CC BY-SA: Attribution-ShareAlike
- Колледж OpenStax, Биология. 12 ноября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44632/latest…ol11448/latest . Лицензия : CC BY: Attribution
- Колледж OpenStax, Биология.12 ноября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44632/latest…ol11448/latest . Лицензия : CC BY: Attribution
- Колледж OpenStax, Экология грибов.
Leave A Comment