
Экологическое и географическое образование новых видов в природе
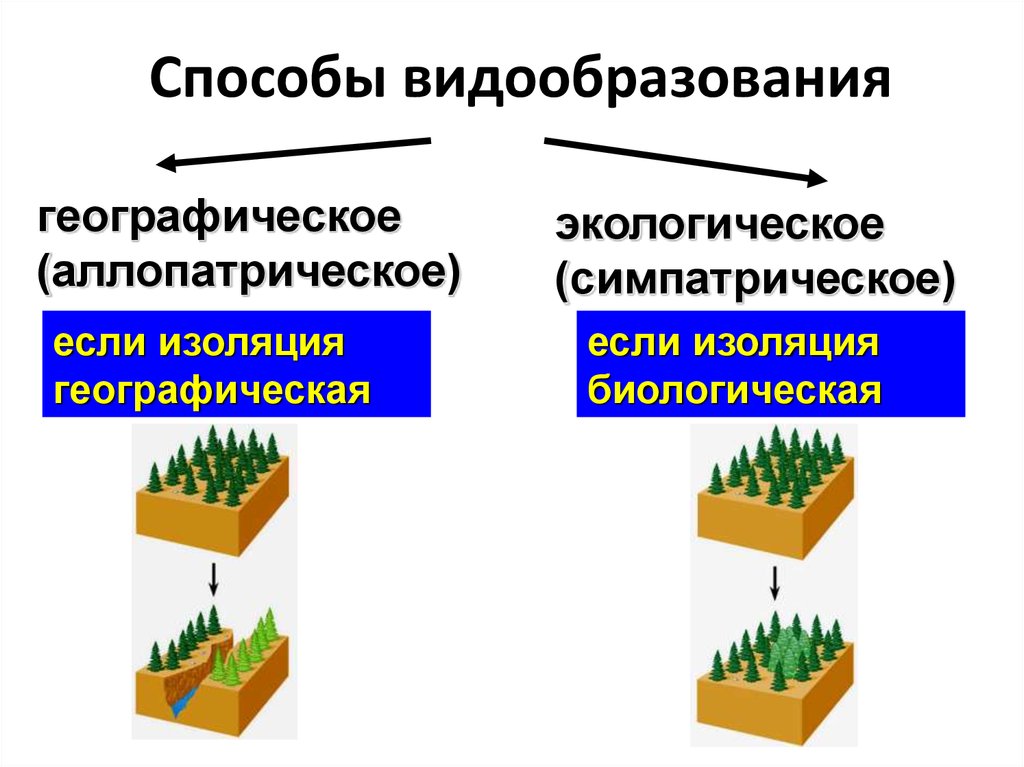
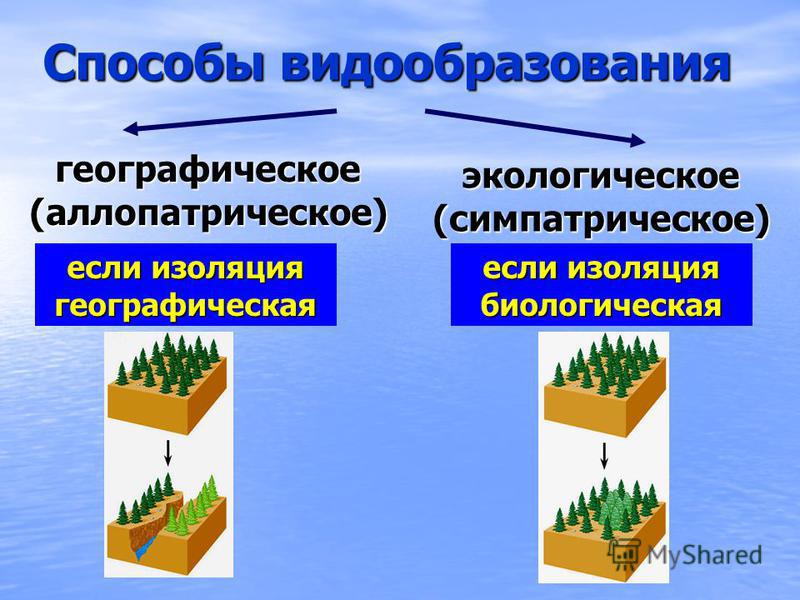
Основные способы видообразования
Что такое видообразование?
Определение 1Видообразование — это процесс эволюционного развития видов в природе.
Размножающиеся половым путем живые организмы представляют собой виды, под которыми понимают совокупность связанных между собой популяций. Вид представляет собой относительно целостную систему, а особи различных популяций могут скрещиваться между собой и обмениваться генетическим материалом. В случаях, если обмен генов по какой-либо из причин затруднен, происходит видовое расчленение. Так как поток генов останавливается, то генофонд становится более-менее самостоятельным. Со временем в каждой популяции происходит накопление оригинальных мутаций — возникают конкретные различия.
Есть несколько форм видообразования, среди которых выделяются географическое и экологическое. Проще всего разобраться в способах видообразования в таблице. В таблице также представлены географическое и экологическое видообразование.
Таблица вариантов видообразования:
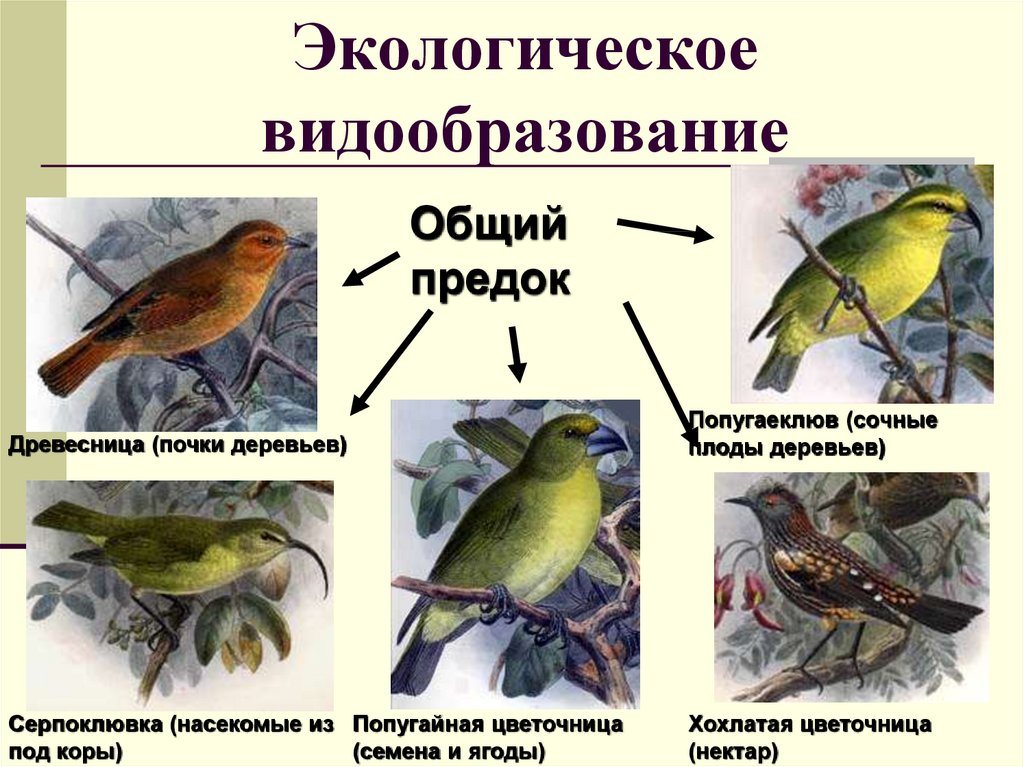
Особенности экологического видообразования
Определение 2Под экологическим видообразованием понимают путь возникновения нового вида в природе, в случае которого вид не выходит за рамки сложившегося ареала.
Также этот процесс образования видов получил название симпатрического — он протекает внутри одной популяции.
Среди признаков такого типа видообразования можно выделить:
- наличие конкретных факторов, в частности, динамики специфических условий обитания того или иного исходного вида. Обычно среди причин видового расхождения — сезонность воспроизведения потомства, различия в способах питания;
- еще один мощный фактор — генетическая изоляция. В общем, можно сказать, что она образованна на мутациях. Благодаря ей потом возникает репродуктивная изоляция;
- расселение особи в пределах одного ареала. Однако экологические условия в этом случае будут различаться;
- образование мутаций, с помощью которых занимаемые экологические ниши расширяются;
- действие естественного отбора;
- закрепление и передача признаков по наследству;
- появление репродуктивной изоляции.


Как происходит экологическое видообразование в природе?
Пример экологического видообразования — становление нескольких видов люцерны. Места произрастания серповидной люцерны — подножия гор. Клейкая разновидность растения встречается в горах. Этот пример экологического видообразования позволяет ученым полагать, что оба вида вышли из одной популяции и заняли экологические ниши без смены непосредственно ареала. Есть множество других примеров экологического видообразования.
Географический способ видообразования
Такой способ подразумевает, что образование новых видов в природе происходит в результате расширения исходного ареала или его разделения на части, в случае возникновения физических препятствий.
При этом важно, чтобы возникающие препятствия являлись непреодолимыми для всех особей популяции. Такой вид видообразования также называется аллопатрическим. Географическое видообразования можно считать наиболее изученным, но, тем не менее, оно не является самым важным.
Как осуществляется видообразование географическим способом? В его основе лежит пространственное разобщение популяции. Его создают географические барьеры: моря, реки, хребты, а также различия в местах обитания. Когда ареал вида расширяется, то популяции сталкиваются с другими почвенно-климатическими условиями и сообществами животных, растений и микроорганизмов. Внутри популяций постоянно осуществляются наследственные изменения, а также борьба за существование и естественный отбор. Перечисленные факторы дают возможность эволюционному процессу протекать относительно стабильно, а системе образования новых видов в природе оставаться непрерывной.
Имеющиеся географические барьеры не позволяют возникать непрерывному генетическому потоку — у гамет утрачивается возможность встречаться. В результате длительного разобщения происходит образование нового вида и генетической изоляции.
В качестве примера можно привести сибирскую лиственницу, для популяций которой характерен довольно широкий ареал обитания. За счет мутаций и систематического скрещивания новые гены получают возможность соединяться в оригинальные комбинации, и популяция становится генетически неоднородной.
Борьба за существование и естественный отбор обуславливали выживание тех особей, у которых отмечались полезные изменения, свойственные данным условиям. Со временем у популяций появлялись более резкие различия. Это привело к возникновению биологической изоляции, невозможности скрещивания особей различных популяций одного вида.
Связи между экологическим видообразованием и разобщением популяций в период возникновения генетической изоляции. Оно происходит только при генетической изоляции, и только в том случае, когда виды находятся в пределах одного и того же ареала обитания. Генный состав популяций довольно сильно меняется под действием конкретных эволюционных процессов. Изменения могут доходить до момента возникновения биологической изоляции, характерной для различных видов живых организмов и приводящей к накоплению мутаций.
К примеру, сегодня существует пять видов синиц, образованных, предположительно, в процессе пищевой специализации. Сюда относят:
- синицу большую;
- синицу хохлатую.
Основная пища первой группы — крупные насекомые. Хохлатая же синица питается в основном семенами различных хвойных деревьев. Есть и другие виды синиц: все это свидетельствует о правдивости теории различных типов видообразования.
Как видно, в ходе микроэволюции один способ видообразования сменяет другой, либо они действуют одновременно. Поэтому так трудно понять, где границы действия всех типов видообразования и каждого способа в отдельности.
Изучение путей видообразования ведется давно и не дается так просто. Причина кроется в длительности процесса видообразования. В чем различие географической и экологической изоляции и образования видов — можно говорить много и долго: их довольно много. Но, тем не менее, оба этих способа представляют ценность в природе и жизни человека.
Особенности географического видообразования и наиболее яркие примеры
Сущность географического видообразования
Определение 1Географическое видообразование представляет собой путь формирования видов в ходе эволюции, при котором первостепенную роль играет пространственная или территориальная изоляция.
Названый путь видообразования также называют аллопатрическим. При географическом видообразовании формирование нового вида происходит в результате изоляции. Одно из важнейших условий образования новых видов — изоляция популяции в конкретных условиях.
Генофонд популяции заметно меняется в результате воздействия окружающей среды. Кроме того, происходит формирование почвы, способствующей расширению спектра мутаций и образованию новых продуктивных генных сочетаний.
Географическая изоляция становится основой для возникновения новой формы этого явления — репродуктивной изоляции. Именно она становится решающим моментом процесса возникновения в природе новых видов.
Репродуктивная изоляция — это невозможность скрещивания между особями разных популяций.
Несмотря на то, что на данный момент известно о множестве теорий видообразования, ни одну из них нельзя назвать целиком доказанной. Это объясняется тем, что любую из них достаточно сложно обосновать эмпирическим путем.
Однако теория географического образования прочно вошла в науку и успешно составляет конкуренцию другим представлениям о системе эволюционного процесса.
Механизм географического видообразования
Появление новых видов — важнейший этап эволюции. Образование новых природных видов формируется под воздействием движущих сил эволюции — это доказал Ч. Дарвин.
Если условия существования внутри вида изменяются, происходит процесс расхождения признаков — дивергенция. Этот процесс приводит к появлению новых группировок особей внутри вида. Поэтому один вид может стать источником целого спектра форм, каждая из которых способна к воспроизведению плодовитого потомства и выживанию в неблагоприятной среде.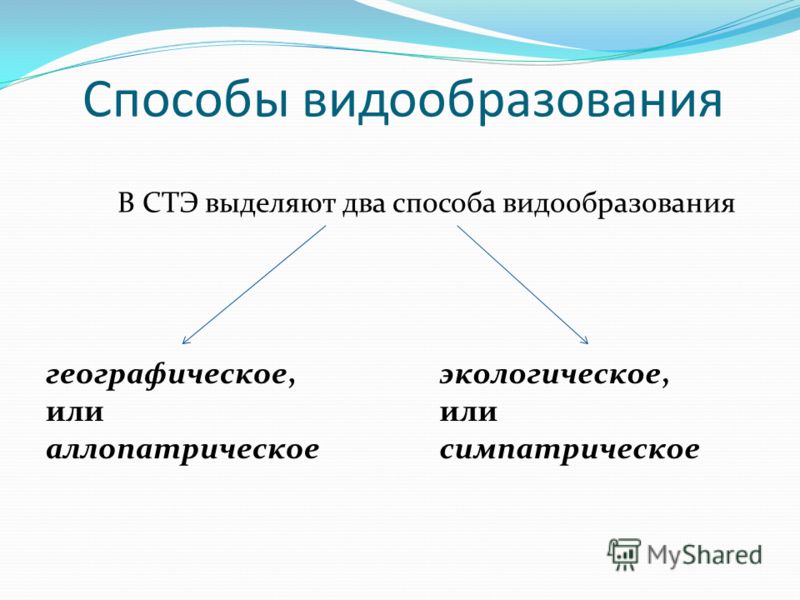
В сравнении с экологическим, географическое видообразование может быть основано только лишь на смене организмом своего ареала обитания. Обычно такое происходит, когда популяция расселяется. Факторам географической изоляции в таких случаях выступают:
- реки;
- озера;
- моря;
- горы;
- леса и др.
Однако нужно понимать, что препятствие само по себе еще не является фактором географической изоляции. К примеру, для одной популяции отдельное препятствие может стать фактором географической изоляции, а для другой популяции это препятствие вообще не будет считаться препятствием.
Когда популяция остается на новой территории, то приобретает адаптации, меняющие ее генотип. Это, в свою очередь, сказывается на фенотипе.
Замечание 2Аллопатрический тип видообразования дал жизнь уникальным видам, проживающим на территориях Австралии, Антарктиды, пустынь и северных широт.
В основе географического видообразования лежат определенные механизмы:
- наличие пространственной изоляции популяций внутри одного вида дает начало новым видам. Пространственная изоляция в этом случае — разделение географических ареалов;
- возникновение генетической изоляции в результате продолжительного разобщения популяций. Генетическая изоляция сохраняется даже тогда, когда географическая изоляция перестает действовать;
- появившиеся в результате географического видообразования новые виды называются географическими расами.
 Пространственная изоляция в этом случае — разделение географических ареалов;
Пространственная изоляция в этом случае — разделение географических ареалов;Причины формирования географического разнообразия
Основные причины формирования географического видообразования:
- миграция особей;
- расширение ареала обитания популяции;
- естественный отбор;
- репродуктивная изоляция;
- накопление мутаций, которые влияют на образование фенотипических и генотипических различий.
Все эти причины проявляются по-разному и зависят от изначального местонахождения популяции и ее генофонда.
Возникновение географической изоляции связано с динамикой развития климата и рельефа. Приспособившиеся к новым условиям обитания особи передают приобретенные навыки адаптации своим потомкам. Поэтому эти потомки способны выжить только в этих условиях.
Поэтому эти потомки способны выжить только в этих условиях.
Аллопатрия — процесс длительный. Он включает 6 этапов:
- Изменение среды обитания видов, расширение ареала (в результате климатических катаклизмов и изменений ландшафта).
- Изменение хода естественного отбора (в результате мутагенеза и адаптаций).
- Сохранение приобретенных полезных адаптаций с последующим закреплением их в генотипе.
- Сохранение географической изоляции.
- Появление репродуктивной изоляции.
- Возникновение новых видов.
Более четкие различия между приспособленными и неадаптированными видами обусловлены меньшим обменом генами между ними. Так образуется репродуктивная изоляция.
Между географическим типом видообразования и изменениями рельефа наблюдается заметная связь. К таким изменениям рельефа относят горообразование, формирование рек и озер, расхождение континентов и др. Все эти процессы длятся по несколько сотен тысяч лет.
Примеры географического видообразования
Примеры географического видообразования помогают понять сам процесс.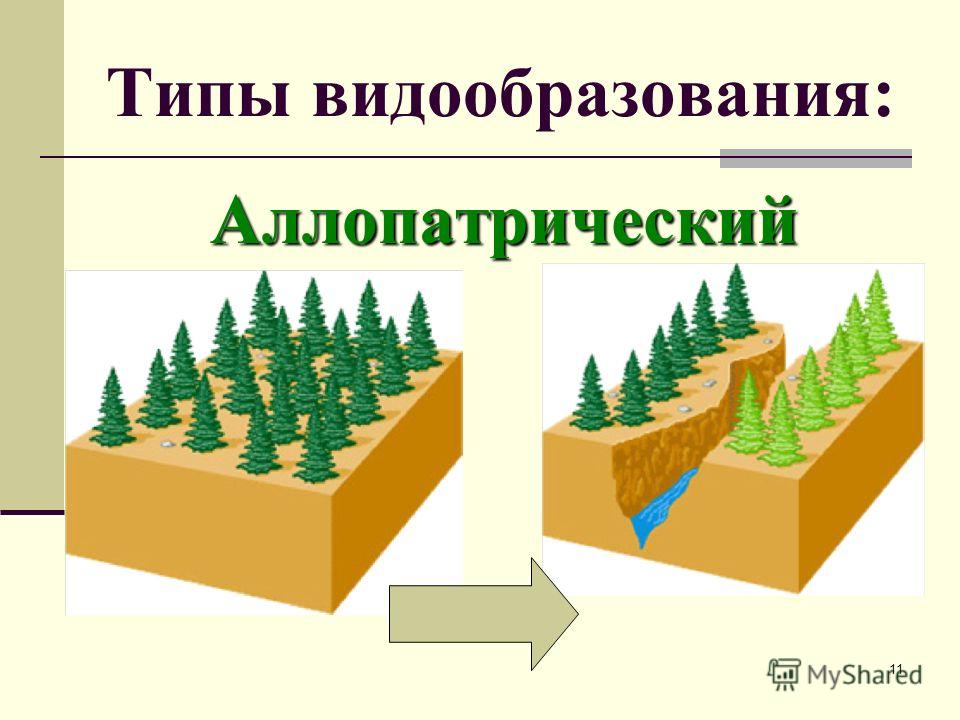
Видовое разнообразие Байкала, которое без сомнения считается уникальным (а это белоголовые чайки, различные виды хлопчатника и др) обусловлено горным хребтом, изолировавшем эту территории от других водных бассейнов.
Схожие примеры видообразования (географического) объясняются парой причин:
- непреодолимыми препятствиями, возникшими на территории вида;
- расширением ареала популяции, в результате которого возможность скрещивания свелась к минимуму или вообще исчезла.
Как следует из текста выше, географическое видообразование — это длительный эволюционный процесс. В его основе лежат климатические изменения и естественные преграды.
Чтобы такое видообразование прошло успешно, важна территориальная изоляция особей внутри одной популяции. Когда возникает полная репродуктивная изоляции, можно говорить об окончании формирования видов.
География видообразования: тематические исследования птиц | Evolution: Education and Outreach
Volume 5 Supplement 4
Biogeography: The Geography of Evolution
- Geography and Evolution
- Открытый доступ
- Опубликовано:
- Роберт М. Цинк 1
Эволюция: образование и просветительская деятельность том 5 , страницы 541–546 (2012 г.)Процитировать эту статью
6724 Доступ
4 Цитаты
1 Альтметрика
Сведения о показателях
Abstract
Наше понимание происхождения видов или видообразования иногда считают неполным, «загадкой тайн».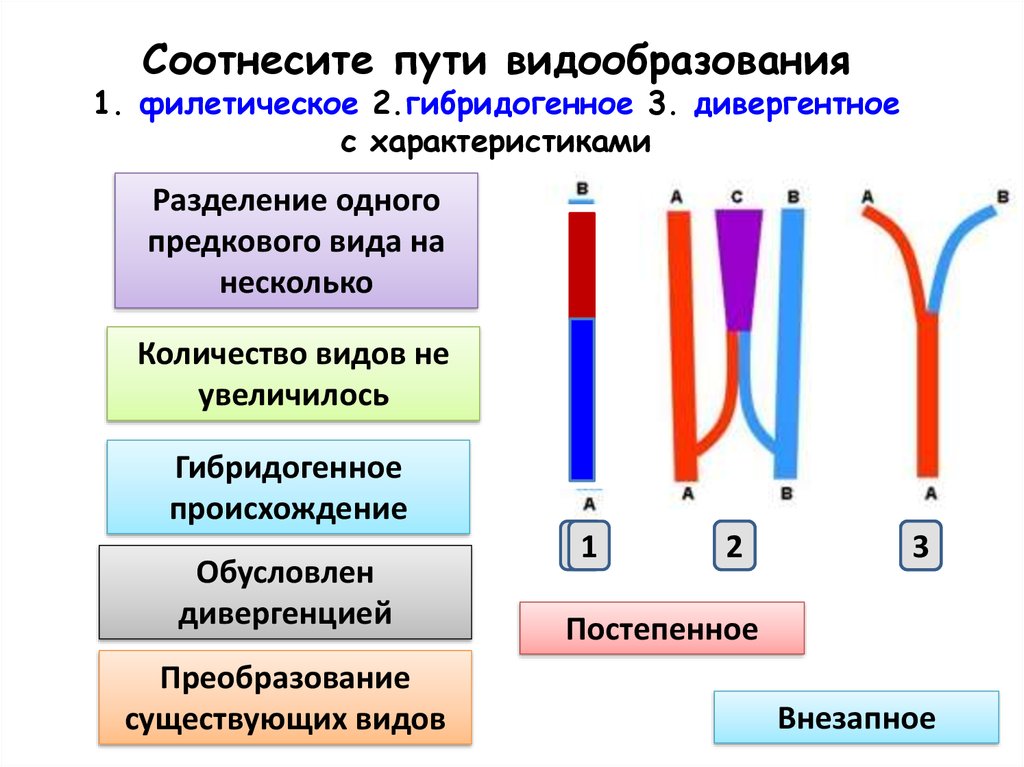 На самом деле мы многое знаем о видообразовании, особенно если рассматривать два его основных компонента — географию видообразования и биологию видообразования (изменения фенотипа и генотипа, происходящие в процессе). Наше понимание географии видообразования достаточно ясно. Этот процесс включает в себя разделение когда-то непрерывного ареала на две или более географически изолированные или аллопатрические части, в которых со временем накапливаются генетические изменения, приводящие к появлению новых дочерних видов. Текущие распределения показывают, что недавно эволюционировавшие виды в настоящее время являются аллопатричными, что подтверждает роль аллопатрии в видообразовании. Однако многие виды возникли в период от раннего до середины плейстоцена, а это означает, что они сохранились в результате экологических возмущений многочисленных ледниковых циклов. Предполагалось, что виды сохраняли аллопатрическое распространение во время этих эпизодов наступления и отступления ледников. Я использовал нишевые модели Гриннеля для оценки распределения видов во время последнего ледникового максимума и последнего межледниковья.
На самом деле мы многое знаем о видообразовании, особенно если рассматривать два его основных компонента — географию видообразования и биологию видообразования (изменения фенотипа и генотипа, происходящие в процессе). Наше понимание географии видообразования достаточно ясно. Этот процесс включает в себя разделение когда-то непрерывного ареала на две или более географически изолированные или аллопатрические части, в которых со временем накапливаются генетические изменения, приводящие к появлению новых дочерних видов. Текущие распределения показывают, что недавно эволюционировавшие виды в настоящее время являются аллопатричными, что подтверждает роль аллопатрии в видообразовании. Однако многие виды возникли в период от раннего до середины плейстоцена, а это означает, что они сохранились в результате экологических возмущений многочисленных ледниковых циклов. Предполагалось, что виды сохраняли аллопатрическое распространение во время этих эпизодов наступления и отступления ледников. Я использовал нишевые модели Гриннеля для оценки распределения видов во время последнего ледникового максимума и последнего межледниковья. У пары недавно выделенных видов камышевок и пары относительно старых сестринских видов мухоловок аллопатрии наблюдались во все периоды времени. Таким образом, в географии видообразования нет никакой тайны: по крайней мере, у птиц преобладает аллопатрия. Мнение о том, что видообразование является таинственным, исходит, как я утверждаю, из концепции биологических видов, которая требует, чтобы популяции были репродуктивно изолированы, прежде чем признавать их видами. Репродуктивная изоляция — сложный процесс, который редко повторяется дважды, и я утверждаю, что это отсутствие общности было неправильно истолковано как загадка.
У пары недавно выделенных видов камышевок и пары относительно старых сестринских видов мухоловок аллопатрии наблюдались во все периоды времени. Таким образом, в географии видообразования нет никакой тайны: по крайней мере, у птиц преобладает аллопатрия. Мнение о том, что видообразование является таинственным, исходит, как я утверждаю, из концепции биологических видов, которая требует, чтобы популяции были репродуктивно изолированы, прежде чем признавать их видами. Репродуктивная изоляция — сложный процесс, который редко повторяется дважды, и я утверждаю, что это отсутствие общности было неправильно истолковано как загадка.
Видообразование, или происхождение видов, часто называют дарвиновской «тайной тайн». На самом деле, Виа (2009:9939) заметил: «Сейчас происхождение видов лишь немногим менее загадочно, чем 150 лет назад, когда Дарвин опубликовал свою знаменитую книгу». Отчасти эта характеристика обескураживает, потому что на самом деле мы очень много знаем о том, как формируются новые виды. Я считаю, что представление о том, что видообразование является загадкой, проистекает из определения самих видов. Различные концепции видов различаются по своей сложности и типам биологических изменений, которые считаются необходимыми и достаточными, чтобы судить о том, что видообразование произошло. Чем сложнее концепция вида, тем загадочнее становится видообразование. Чтобы разрушить представление о таинственности видообразования, полезно разделить происхождение видов на два компонента, которые мы могли бы для простоты назвать географией и биологией видообразования (Буш 19).75). В этом эссе я сосредоточусь на нашем понимании географии видообразования у птиц, возможно, одной из самых известных групп позвоночных. Я показываю, что география видообразования проста при всех концепциях видов и что на самом деле видообразование загадочно только при определенных определениях видов.
Я считаю, что представление о том, что видообразование является загадкой, проистекает из определения самих видов. Различные концепции видов различаются по своей сложности и типам биологических изменений, которые считаются необходимыми и достаточными, чтобы судить о том, что видообразование произошло. Чем сложнее концепция вида, тем загадочнее становится видообразование. Чтобы разрушить представление о таинственности видообразования, полезно разделить происхождение видов на два компонента, которые мы могли бы для простоты назвать географией и биологией видообразования (Буш 19).75). В этом эссе я сосредоточусь на нашем понимании географии видообразования у птиц, возможно, одной из самых известных групп позвоночных. Я показываю, что география видообразования проста при всех концепциях видов и что на самом деле видообразование загадочно только при определенных определениях видов.
Представления о видах и видообразование
Чтобы проиллюстрировать взаимосвязь между представлениями о видах и видообразованием, я кратко рассмотрю три общих представления о видах. Это важно, потому что нельзя обсуждать видообразование без предварительного определения критериев распознавания видов (Wiens 2004). Концепция биологических видов (BSC) была в значительной степени сформулирована генетиком Феодосием Добжанским и популяризирована многими последующими биологами-эволюционистами, особенно Эрнстом Майром. Одна из формулировок BSC гласит: «Виды — это группы фактически или потенциально скрещивающихся естественных популяций, которые репродуктивно изолированы от других таких групп» (19 мая 2009 г.).42). Таким образом, видообразование требует эволюции характеристик, обеспечивающих репродуктивную изоляцию популяциям, происходящим от общего предка. Эти характеристики могут помешать особям выбирать себе пару (репродуктивная изоляция до спаривания) или привести к бесплодному или нежизнеспособному потомству в случае спаривания (репродуктивная изоляция после спаривания). Между таксономическими группами мало единообразия в отношении того, какие конкретные генетические, поведенческие или экологические атрибуты способствуют репродуктивной изоляции, что, по сути, является эпифеноменом дивергенции признаков.
Это важно, потому что нельзя обсуждать видообразование без предварительного определения критериев распознавания видов (Wiens 2004). Концепция биологических видов (BSC) была в значительной степени сформулирована генетиком Феодосием Добжанским и популяризирована многими последующими биологами-эволюционистами, особенно Эрнстом Майром. Одна из формулировок BSC гласит: «Виды — это группы фактически или потенциально скрещивающихся естественных популяций, которые репродуктивно изолированы от других таких групп» (19 мая 2009 г.).42). Таким образом, видообразование требует эволюции характеристик, обеспечивающих репродуктивную изоляцию популяциям, происходящим от общего предка. Эти характеристики могут помешать особям выбирать себе пару (репродуктивная изоляция до спаривания) или привести к бесплодному или нежизнеспособному потомству в случае спаривания (репродуктивная изоляция после спаривания). Между таксономическими группами мало единообразия в отношении того, какие конкретные генетические, поведенческие или экологические атрибуты способствуют репродуктивной изоляции, что, по сути, является эпифеноменом дивергенции признаков.
Вторая концепция вида — это концепция филогенетического вида (PSC), которая была определена Кракрафтом (1983): «Вид — это наименьшая диагностируемая группа отдельных организмов, внутри которой существует родительский образец происхождения и происхождения». Здесь видообразование — это просто происхождение диагностических признаков, которые показывают, что популяция или группа популяций имели независимую эволюционную историю. Это может быть короткая или долгая история, а характеристики могут быть фенотипическими, экологическими, молекулярными, физиологическими или поведенческими — короче говоря, любой наследственный диагностический признак. Нет требования, чтобы виды были репродуктивно изолированы (Zink and McKitrick 19).95).
Например, третья (среди многих других) концепция вида — это концепция эволюционного вида, вариант которой гласит: «Эволюционный вид — это отдельная линия популяций предков-потомков, которая сохраняет свою идентичность от других таких линий и которая имеет свои эволюционные тенденции и историческую судьбу» (Wiley 1978). Здесь не нужны никакие доказательства характера, а только знание того, что популяция или группа популяций «отправились» на самостоятельную эволюционную траекторию. Например, популяция, изолированная на острове, теоретически является видом независимо от того, отличима ли она от своего материкового предка.
Здесь не нужны никакие доказательства характера, а только знание того, что популяция или группа популяций «отправились» на самостоятельную эволюционную траекторию. Например, популяция, изолированная на острове, теоретически является видом независимо от того, отличима ли она от своего материкового предка.
Основные отличительные характеристики этих представлений о видах приведены в Таблице 1. Каждое из представлений о видах определяет видообразование как начало аллопатрического разделения предкового вида. Ниже я сосредоточусь на географических аспектах видообразования, которое правильно рассматривать как длительный процесс во времени, а не как мгновенное событие.
Таблица 1 Некоторые характеристики, необходимые для того, чтобы популяции считались отдельными видами в соответствии с тремя различными концепциями видовПолноразмерная таблица
География видообразования
История Земли за последние два миллиона лет, примерно в плейстоцене, характеризовалась регулярными и массовыми изменениями среды обитания. Конечно, конечной причиной было глобальное изменение климата, которое привело к многочисленным эпизодам наступления и отступления ледников с более или менее регулярными интервалами. Последствия этих ледниковых циклов очевидны. Ареалы видов регулярно сдвигались, сжимались, дробились, элиминировались или сильно расширялись. Другими словами, широко распространенный вид, возможно, универсальная среда обитания, будет вытеснен на юг по мере ухудшения климата (похолодания). По мере продвижения ледникового цикла и оттеснения ледяных щитов на юг, что привело к изменению распределения растительных сообществ, от которых зависел вид, ареал вида мог не только сместиться на юг, но и раздробиться на географически изолированные компоненты.
Конечно, конечной причиной было глобальное изменение климата, которое привело к многочисленным эпизодам наступления и отступления ледников с более или менее регулярными интервалами. Последствия этих ледниковых циклов очевидны. Ареалы видов регулярно сдвигались, сжимались, дробились, элиминировались или сильно расширялись. Другими словами, широко распространенный вид, возможно, универсальная среда обитания, будет вытеснен на юг по мере ухудшения климата (похолодания). По мере продвижения ледникового цикла и оттеснения ледяных щитов на юг, что привело к изменению распределения растительных сообществ, от которых зависел вид, ареал вида мог не только сместиться на юг, но и раздробиться на географически изолированные компоненты.
Когда когда-то непрерывные популяции географически изолированы друг от друга, что называется аллопатрией (проживание в разных местах), они начинают накапливать мутации, уникальные для их географической группы, и начинается процесс генетического расхождения. Процесс может быть обусловлен естественным отбором, половым отбором или даже генетическим дрейфом. Если вновь изолированные популяции изолированы не полностью, т. е. некоторые особи могут перемещаться из одной вновь изолированной области в другую, процесс будет замедлен или, возможно, предотвращен. Аксиома популяционной генетики состоит в том, что небольшой поток генов имеет большое значение — поток генов через рассредоточенных особей является «генетическим клеем», который удерживает популяции от появления на их собственной отдельной эволюционной траектории. Но если популяции останутся изолированными, они накопит достаточное разнообразие в своей генетике, экологии или морфологии, чтобы стать разными видами (19 мая).42). Природа дивергенции будет определять, произошло ли видообразование в соответствии с концепциями разных видов. Если аллопатрические таксоны демонстрируют различия в диагностических признаках, они являются видами согласно PSC. Только в том случае, если эти диагностические различия также функционируют как механизмы репродуктивной изоляции, таксоны могут считаться биологическими видами.
Процесс может быть обусловлен естественным отбором, половым отбором или даже генетическим дрейфом. Если вновь изолированные популяции изолированы не полностью, т. е. некоторые особи могут перемещаться из одной вновь изолированной области в другую, процесс будет замедлен или, возможно, предотвращен. Аксиома популяционной генетики состоит в том, что небольшой поток генов имеет большое значение — поток генов через рассредоточенных особей является «генетическим клеем», который удерживает популяции от появления на их собственной отдельной эволюционной траектории. Но если популяции останутся изолированными, они накопит достаточное разнообразие в своей генетике, экологии или морфологии, чтобы стать разными видами (19 мая).42). Природа дивергенции будет определять, произошло ли видообразование в соответствии с концепциями разных видов. Если аллопатрические таксоны демонстрируют различия в диагностических признаках, они являются видами согласно PSC. Только в том случае, если эти диагностические различия также функционируют как механизмы репродуктивной изоляции, таксоны могут считаться биологическими видами.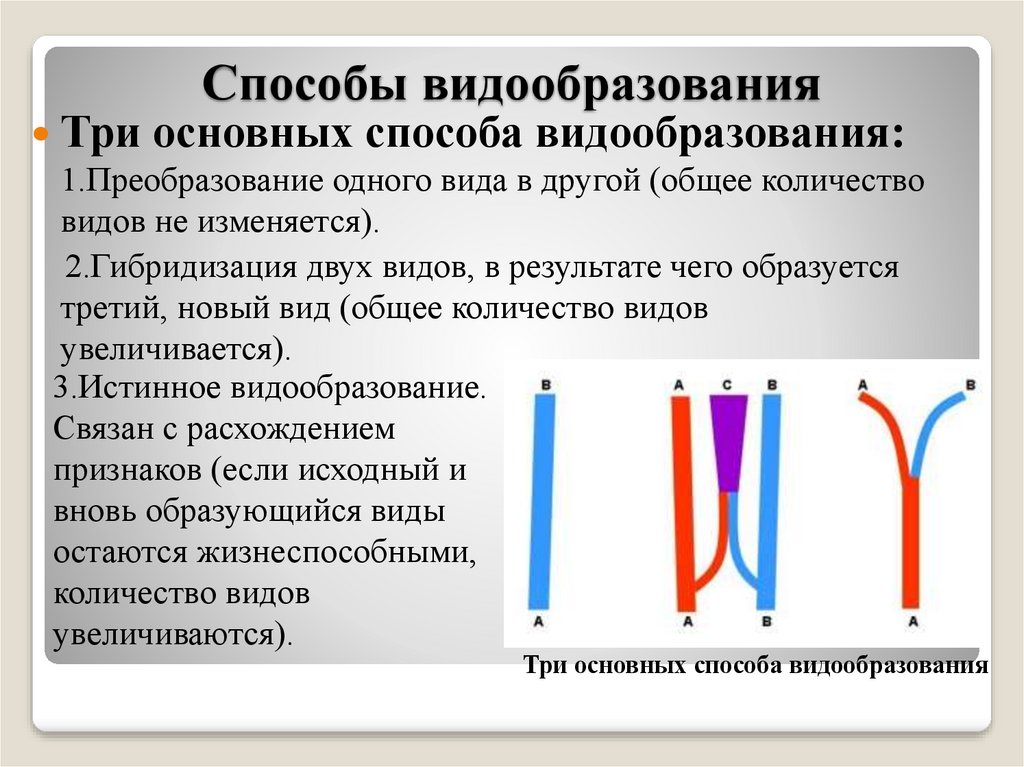 В любом случае представляется, что виды регулярно развивались на протяжении плейстоцена в северных районах с умеренным климатом в результате ледниковых циклов и вызываемых ими изменений среды обитания (Klicka and Zink 19).97, 1999).
В любом случае представляется, что виды регулярно развивались на протяжении плейстоцена в северных районах с умеренным климатом в результате ледниковых циклов и вызываемых ими изменений среды обитания (Klicka and Zink 19).97, 1999).
Изучение распространения видов птиц сыграло важную роль в определении важности географической изоляции в процессе видообразования. В частности, важную роль сыграл анализ ареалов вновь отделившихся или сестринских видов. Большинство существующих сестринских видов птиц аллопатричны или почти таковы; те, которые частично перекрываются, эволюционно «старее», что означает, что они развили достаточные экологические различия, чтобы обеспечить сосуществование в некоторых областях, обычно там, где встречаются ареалы (Chesser and Zink 19).94). Таким образом, пространственная изоляция сестринских видов является признаком аллопатрического видообразования.
Всегда существуют потенциальные исключения из аллопатрического видообразования, такие как видообразование, происходящее без географической изоляции, которое называется симпатрическим видообразованием. Однако доказательств существования этого способа видообразования у птиц мало, если они вообще есть.
Однако доказательств существования этого способа видообразования у птиц мало, если они вообще есть.
Два тематических исследования роли аллопатрии в видообразовании птиц, выявленные с помощью моделирования ниши
Тот факт, что сестринские виды сегодня имеют аллопатрическое распространение, является убедительным свидетельством важности географической изоляции в процессе видообразования. Однако до сих пор мы только предполагали, что аллопатрия сохранялась с точки первоначального расхождения. Например, хотя сестринские виды сегодня являются аллопатрическими (или «парапатрическими», что означает, что ареалы прилегают друг к другу), мы можем только предполагать, что в более ранние времена, в разные ледниковые режимы, таксоны также были аллопатрическими. Недавние разработки в области моделирования ниш (Петерсон и др., 2012) позволяют нам исследовать распространение современных видов в более ранние моменты времени, чтобы оценить постоянство аллопатрии на протяжении истории видов. Я представляю два тематических исследования, в которых участвуют две пары ныне аллопатрических видов, пара североамериканских видов камышевок и пара североамериканских видов мухоловок. Я реконструирую их историческое распределение, чтобы проверить, были ли они аллопатрическими на протяжении всего своего эволюционного расхождения.
Я представляю два тематических исследования, в которых участвуют две пары ныне аллопатрических видов, пара североамериканских видов камышевок и пара североамериканских видов мухоловок. Я реконструирую их историческое распределение, чтобы проверить, были ли они аллопатрическими на протяжении всего своего эволюционного расхождения.
Две камышевки по-разному рассматриваются как виды и подвиды: миртовая камышевка ( Dendroica coronata ) и камышевка Одюбона ( Dendroica auduboni ). Они очень похожи по размеру и форме, с основным отличием в том, что у первого горло белое, а у второго желтое. Они также очень похожи по своей последовательности ДНК, а это означает, что различия в их оперении возникли недавно (Milá et al. 2007, Brelsford and Irwin 2009). В основном они аллопатрические, но их ареалы перекрываются на небольшой территории в Британской Колумбии, где происходит некоторая гибридизация. Вот почему некоторые исследователи считают их одним и тем же видом (в соответствии с BSC), в то время как другие, такие как текущий автор, считают это неуместным и считают их двумя видами в соответствии с PSC.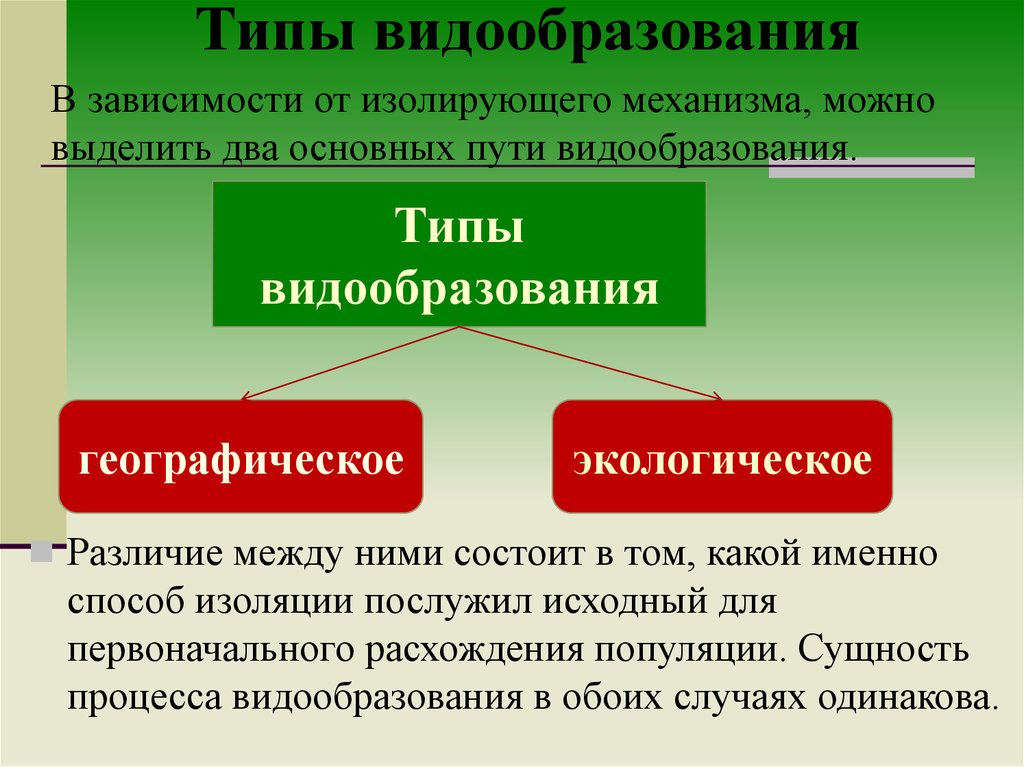
Были ли эти виды камышевок исторически аллопатричными или в основном таковыми были? Чтобы ответить на этот вопрос, можно использовать нишевые модели. Они включают в себя набор современных записей событий и набор различных климатических переменных, таких как температура и осадки, и построение модели, которая предсказывает их текущее распределение. Это называется моделью распространения вида или моделью ниши и в основном описывает условия окружающей среды, благоприятные для вида; ее иногда называют «гриннеллианской» нишей в честь известного орнитолога из Беркли Джозефа Гриннелла. Вид может иметь меньшее распространение, чем предполагалось, из-за присутствия конкурентов (иногда называемых элтоновской или реализованной нишей).
Если нишевая модель может предсказать, где вид встречается в настоящее время, мы затем берем набор климатических переменных из прошлого и используем модель, чтобы предсказать, где вид мог встречаться в то время (Петерсон, 2001 г., Петерсон и др., 2012 г. ). , Валтари и др., 2007). Эта процедура предполагает, что ниша вида не изменилась с течением времени. Кроме того, модели только предсказывают, где условия, в которых вид живет сегодня, имели место в какое-то предыдущее время, а не то, действительно ли этот вид существовал там (т. Е. Конкурент мог исключить вид из какой-то области). Однако это обычная процедура, и многие ученые пришли к выводу, что эти методы надежны. Я использовал моделирование ниш, чтобы определить, были ли две пары видов камышевок аллопатричными во время последнего ледникового максимума (LGM) примерно за 21 000 лет до настоящего времени (ybp) и во время последнего межледниковья (LIG), ок. 120 000 лн, время, для которого доступны климатические данные.
). , Валтари и др., 2007). Эта процедура предполагает, что ниша вида не изменилась с течением времени. Кроме того, модели только предсказывают, где условия, в которых вид живет сегодня, имели место в какое-то предыдущее время, а не то, действительно ли этот вид существовал там (т. Е. Конкурент мог исключить вид из какой-то области). Однако это обычная процедура, и многие ученые пришли к выводу, что эти методы надежны. Я использовал моделирование ниш, чтобы определить, были ли две пары видов камышевок аллопатричными во время последнего ледникового максимума (LGM) примерно за 21 000 лет до настоящего времени (ybp) и во время последнего межледниковья (LIG), ок. 120 000 лн, время, для которого доступны климатические данные.
Реконструированное распределение камышевок в настоящее время (рис. 1А) хорошо соответствует их известному распределению, что позволяет с уверенностью строить реконструкции ареалов предков. Предсказанные распределения камышевок на LGM (рис. 1B) и LIG (рис. 1C) предполагают долгую историю аллопатрии. Кроме того, предсказанные распределения в LIG (рис. 1C) очень похожи на текущие, что свидетельствует о непрерывности распределений в течение двух последних межледниковых периодов (LIG и настоящего). Таким образом, славки были аллопатричными на протяжении как минимум 120 000 лет, что усиливает роль аллопатрии в поддержании (и инициировании) их эволюционного расхождения. Кстати, этот результат также дает минимальную оценку времени, прошедшего с тех пор, как они стали отдельными видами. Модель ниши предполагает, что они были изолированы, по крайней мере, со времен LIG, но, вероятно, не намного дольше из-за небольшого количества генетических различий между ними (Milá et al. 2007; но см. Brelsford and Irwin 2009).).
Кроме того, предсказанные распределения в LIG (рис. 1C) очень похожи на текущие, что свидетельствует о непрерывности распределений в течение двух последних межледниковых периодов (LIG и настоящего). Таким образом, славки были аллопатричными на протяжении как минимум 120 000 лет, что усиливает роль аллопатрии в поддержании (и инициировании) их эволюционного расхождения. Кстати, этот результат также дает минимальную оценку времени, прошедшего с тех пор, как они стали отдельными видами. Модель ниши предполагает, что они были изолированы, по крайней мере, со времен LIG, но, вероятно, не намного дольше из-за небольшого количества генетических различий между ними (Milá et al. 2007; но см. Brelsford and Irwin 2009).).
Прогнозное распределение камышевки Одюбона и миртовой камышевки в настоящее время ( a ), максимум последнего ледникового периода ( b ) и последнее межледниковье ( c ). Нишевые модели были построены путем ввода записей о размножении из обследования гнездящихся птиц (доступ на www. pwrc.usgs.gov/bbs) в MAXENT версии 3.2.2. (Филлипс и др., 2006). Климатические данные (19 слоев) были получены из биоклиматической базы данных Worldclim (Hijmans et al. 2005), и Maxent использовал для обучения 30% значений. Карты распределения были закодированы так, чтобы показать наличие или отсутствие на основе минимального порога прогнозируемого появления
pwrc.usgs.gov/bbs) в MAXENT версии 3.2.2. (Филлипс и др., 2006). Климатические данные (19 слоев) были получены из биоклиматической базы данных Worldclim (Hijmans et al. 2005), и Maxent использовал для обучения 30% значений. Карты распределения были закодированы так, чтобы показать наличие или отсутствие на основе минимального порога прогнозируемого появления
Изображение в натуральную величину
Два других родственных вида, калифорнийская комахоловка ( Polioptila californica ) и чернохвостая комахоловка ( Polioptila melanura ) сегодня имеют аллопатрические ареалы. Мухоловки отличаются от камышевок, по крайней мере, в двух важных аспектах: они ведут оседлый образ жизни (камышевки мигрируют) и сильно отличаются генетически. В частности, расхождение нуклеотидов между видами составляет почти 5% в гене митохондриальной ДНК (Zink and Blackwell 19).98). Молекулярные эволюционисты считают, что замены ДНК происходят примерно как часы, и, учитывая некоторые стандартные калибровки скорости эволюции мтДНК у птиц, вполне вероятно, что два вида мокроловов в последний раз имели общего предка более миллиона лет назад. Это означает, что они были отдельными видами и оставались разными на протяжении многих ледниковых наступлений и отступлений, которые произошли за последний миллион лет. Таким образом, учитывая их своеобразие, мы можем предсказать, что мы должны, как и в случае с камышевками, наблюдать аллопатрию на LGM и LIG.
Это означает, что они были отдельными видами и оставались разными на протяжении многих ледниковых наступлений и отступлений, которые произошли за последний миллион лет. Таким образом, учитывая их своеобразие, мы можем предсказать, что мы должны, как и в случае с камышевками, наблюдать аллопатрию на LGM и LIG.
Предсказанные текущие распределения (рис. 2A) очень близки к известным распределениям двух мокроловов, что позволяет предположить, что модель отражает основные климатические особенности, определяющие распределение мокроловов. Реконструированные LGM (рис. 2B) и LIG (рис. 2C) распределения для двух видов мокроловов ясно показывают, что аллопатрии была доминирующей чертой их исторических ареалов.
Рис. 2 Прогноз распространения калифорнийской коммунарки и чернохвостой мухоловки в настоящее время ( a ), Последний ледниковый максимум ( b ) и Последнее межледниковье ( c ). Методы см. в легенде к рис. 1.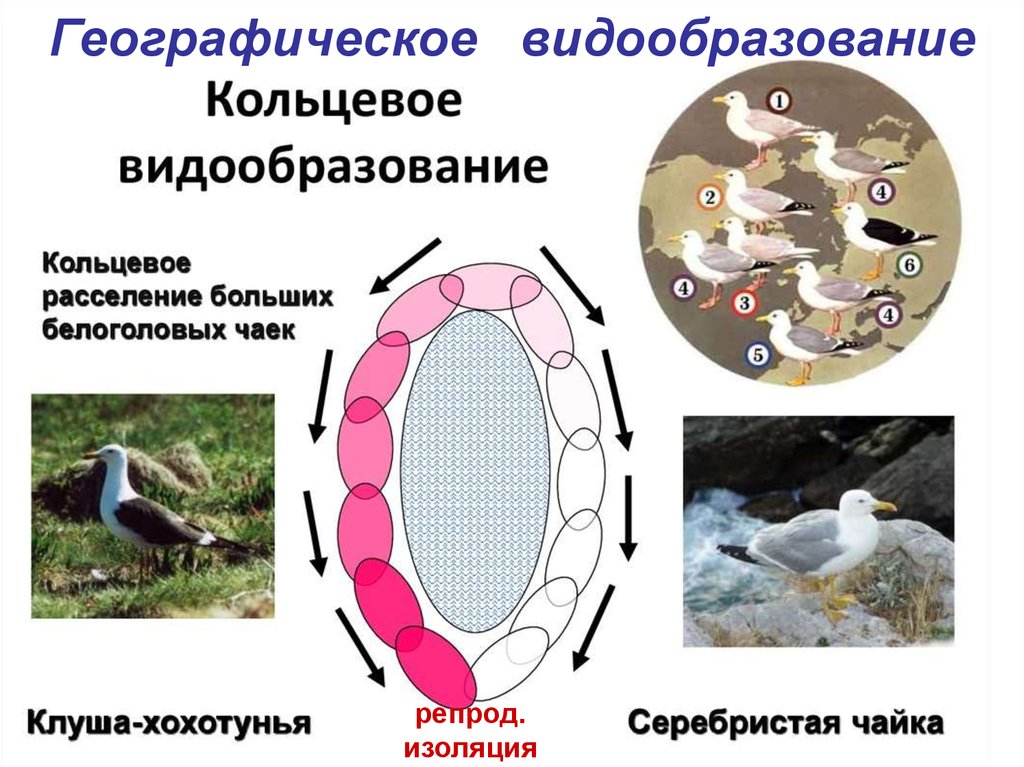 То есть можно было бы ожидать, что с момента их видообразования прошло достаточно времени, чтобы у них развились экологические различия, которые позволили бы им «вторгаться» в ареалы друг друга и сосуществовать на одной и той же территории. Тот факт, что они являются аллопатрическими, предполагает, что они слишком похожи экологически, чтобы сосуществовать (называемый консерватизмом ниши; см. McCormick et al. 2010), и, следовательно, они остаются аллопатрически распределенными. Однако для подтверждения этой гипотезы требуются более сложные нишевые тесты. В качестве альтернативы два вида могут быть поведенчески антагонистическими по отношению друг к другу, препятствуя сосуществованию.
То есть можно было бы ожидать, что с момента их видообразования прошло достаточно времени, чтобы у них развились экологические различия, которые позволили бы им «вторгаться» в ареалы друг друга и сосуществовать на одной и той же территории. Тот факт, что они являются аллопатрическими, предполагает, что они слишком похожи экологически, чтобы сосуществовать (называемый консерватизмом ниши; см. McCormick et al. 2010), и, следовательно, они остаются аллопатрически распределенными. Однако для подтверждения этой гипотезы требуются более сложные нишевые тесты. В качестве альтернативы два вида могут быть поведенчески антагонистическими по отношению друг к другу, препятствуя сосуществованию.
География видообразования — не тайна: аллопатрии преобладает
На протяжении десятилетий мы изучали современное распространение видов и наблюдали признаки аллопатрической дивергенции. Исследования птиц сыграли важную роль в предоставлении многочисленных примеров. Однако мы только недавно начали использовать моделирование ниш для изучения роли аллопатрии в истории видов. Я показал здесь на двух примерах, что аллопатрия действительно является отличительной чертой процесса видообразования. То есть география видообразования почти наверняка представляет собой процесс, связанный с аллопатрией. Таким образом, в этой части процесса видообразования нет ничего загадочного.
Я показал здесь на двух примерах, что аллопатрия действительно является отличительной чертой процесса видообразования. То есть география видообразования почти наверняка представляет собой процесс, связанный с аллопатрией. Таким образом, в этой части процесса видообразования нет ничего загадочного.
Где тайна видообразования?
Ответ на этот вопрос — тема для отдельного эссе. Однако он включает в себя вторую часть процесса видообразования, а именно то, как развиваются признаки в процессе видообразования, или, как я назвал это выше, «биологию» видообразования. Некоторые концепции видов, такие как BSC, требуют, чтобы произошел сложный набор событий, прежде чем две диагностически разные популяции будут признаны двумя видами и, следовательно, сделан вывод о том, что видообразование произошло (таблица 1). Существует множество способов, которыми две аллопатрические популяции могут приобрести различия в характерах, необходимые им для репродуктивной изоляции. В этом процессе для животных и растений мало общего, если оно вообще есть, и то, что существует, ограничивается конкретными группами. Я считаю, что из-за того, что существует так мало общего, видообразование считается таинственным. Короче говоря, если вы осознаете, что два вида могут стать репродуктивно изолированными с помощью уникальных средств, в этом нет никакой тайны, только отсутствие общности. Таким образом, биология видообразования также не является загадкой. Просто биология видообразования сложнее географии видообразования. Это, на мой взгляд, нельзя назвать биологической загадкой, это просто еще раз раскрывает сложную природу биологических вариаций, которая так интересует нас в понимании жизни в первую очередь.
Я считаю, что из-за того, что существует так мало общего, видообразование считается таинственным. Короче говоря, если вы осознаете, что два вида могут стать репродуктивно изолированными с помощью уникальных средств, в этом нет никакой тайны, только отсутствие общности. Таким образом, биология видообразования также не является загадкой. Просто биология видообразования сложнее географии видообразования. Это, на мой взгляд, нельзя назвать биологической загадкой, это просто еще раз раскрывает сложную природу биологических вариаций, которая так интересует нас в понимании жизни в первую очередь.
Ссылки
Брелсфорд А., Ирвин Д.Е. Начальное видообразование, несмотря на небольшое ассортативное спаривание: гибридная зона желтопоясничной камышевки. Эволюция. 2009;63:3050–60.
Артикул Google Scholar
Буш Г. Способы видообразования животных. Энн Рев Экол Сист. 1975; 6: 339–64.
Артикул Google Scholar
Чессер РТ, Цинк РМ. Способы видообразования у птиц: приложение Линча (19).89) метод. Эволюция. 1994;48:490–7.
Артикул Google Scholar
Кракрафт Дж. Концепции видов и анализ видообразования. Карр Орнито. 1983; 1: 159–87.
Артикул Google Scholar
Хиджманс Р., Камерон С., Парра Дж., Джонс П., Джарвис А. Интерполированные климатические поверхности с очень высоким разрешением для глобальных участков суши. Int J Климатол. 2005; 25:1965–78.
Артикул Google Scholar
Клика Дж., Цинк Р.М. Значение недавних ледниковых периодов в видообразовании: несостоявшаяся парадигма. Наука. 1997; 277:1666–9.
Артикул КАС Google Scholar
Кличка Дж.
, Цинк Р.М. Филогеографические эффекты плейстоцена на эволюцию птиц. Proc Royal Soc Lond B. 1999; 266: 695–700.Артикул Google Scholar
Майр Э. Систематика и происхождение видов. 1942. Колумбийский ун-т. Нажимать.
McCormack JE, Zellmer AJ, Knowles LL. Сопровождается ли дивергенция ниши аллопатрической дивергенцией у соек Aphelocoma , как это предсказывает экологическое видообразование? Выводы из тестов с нишевыми моделями. Эволюция. 2010;64–5:1231–44.
Google Scholar
Мила Б., Смит Т.Б., Уэйн Р.К. Видообразование и быстрая фенотипическая дифференциация желтопоясничной камышевки Dendroica coronata 9Комплекс 0024. Мол Экол. 2007; 16: 159–73.
Артикул Google Scholar
Петерсон А. Прогнозирование географического распространения видов на основе моделирования экологических ниш.
Кондор. 2001; 103: 599–605.Артикул Google Scholar
Петерсон А.Т., Соберон Дж., Андерсон Р.П., Пирсон Р.Г., Мартинес-Мейер Э., Накамура М., Араужо М.Б. Экологические ниши и географическое распространение: перспектива моделирования. 2012. Издательство Принстонского университета, Принстон.
Филлипс С., Андерсон Р., Шапир Р. Моделирование максимальной энтропии географического распространения видов. Эколь Модель. 2006; 190: 231–59.
Артикул Google Scholar
Виа С. Естественный отбор в действии во время видообразования. Proc Nat Acad Sci USA. 2009; 106: 9939–46.
Артикул КАС Google Scholar
Уолтари Э., Перкинс С., Хийманс Р., Петерсон А.Т., Ньяри А., Гуральник Р. Обнаружение убежищ плейстоцена: сравнение предсказаний филогеографической и экологической модели ниши. ПЛОС Один.
2007;2:e563.Артикул Google Scholar
Вена Дж.Дж. Что такое видообразование и показать, должны ли мы его изучать? Я натуралист. 2004; 164:914–23.
Артикул Google Scholar
Wiley EO. Пересмотр концепции эволюционного вида. Сист Зоол. 1978; 27:17–26.
Артикул Google Scholar
Zink RM, Blackwell RC. Молекулярная систематика и биогеография аридных мухоловок (род Polioptila ), а также доказательства, подтверждающие статус вида калифорнийского комарика ( P. californica ). Крот Фило Эвол. 1998; 9: 26–32.
Артикул КАС Google Scholar
Цинк Р.М., МакКитрик М.С. Дебаты о концепциях видов и их значении для орнитологии. Аук. 1995; 112: 701–19.
Google Scholar
 , Цинк Р.М. Филогеографические эффекты плейстоцена на эволюцию птиц. Proc Royal Soc Lond B. 1999; 266: 695–700.
, Цинк Р.М. Филогеографические эффекты плейстоцена на эволюцию птиц. Proc Royal Soc Lond B. 1999; 266: 695–700. Кондор. 2001; 103: 599–605.
Кондор. 2001; 103: 599–605. 2007;2:e563.
2007;2:e563.Скачать ссылки
Благодарности
Я думаю, К. -М. Hung, H. Vazquez-Miranda, B. McKay и двум анонимным рецензентам за полезные комментарии к рукописи.
-М. Hung, H. Vazquez-Miranda, B. McKay и двум анонимным рецензентам за полезные комментарии к рукописи.
Информация о авторе
Авторы и принадлежности
Музей и факультет экологии, эволюции и поведения, Университет Миннесоты, Сент
- Robert M. Zink
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
Автор, ответственный за корреспонденцию
Роберт М. Зинк.
Права и разрешения
Открытый доступ Эта статья распространяется на условиях международной лицензии Creative Commons Attribution 2.0 ( https://creativecommons.org/licenses/by/2.0 ), который разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии надлежащего цитирования оригинальной работы.
Перепечатки и разрешения
Об этой статье
География видообразования — Эволюционная биология
Логин
Войти
Имя пользователя
Пожалуйста, введите ваше имя пользователя
Пароль
Пожалуйста, введите ваш пароль
Забыли пароль?
Нет учетной записи?
Введение
Видообразование – это процесс разделения одной линии на две или более дочерних линии.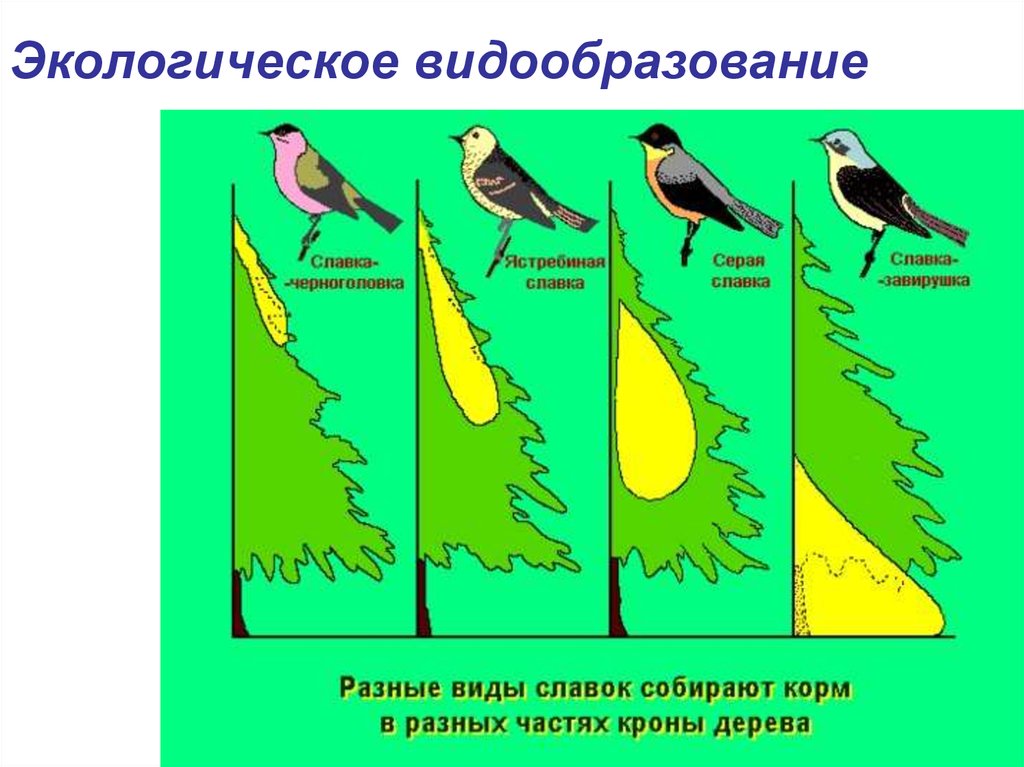 География играет ключевую роль в этом процессе. Когда популяции пространственно изолированы, они больше не обмениваются генами и могут дивергировать. Побочным продуктом этого расхождения является эволюция внутренних репродуктивных барьеров — признаков, которые препятствуют скрещиванию видов и сигнализируют о завершении видообразования. Аллопатрическое видообразование – это возникновение новых видов в пространственно изолированных популяциях. Аллопатрическое видообразование является самой простой и легкой моделью видообразования, потому что между популяциями нет потоков генов, которые могли бы подорвать дивергенцию. При наличии достаточного количества времени ожидается аллопатрическое видообразование, и широко признано, что это наиболее распространенный способ видообразования. Действительно, аллопатрическое видообразование было описано Койном и Орром в Вид (2004 г.) как «настолько правдоподобный, что вряд ли стоит его документировать». Видообразование, конечно, может происходить и в других географических контекстах, с разными уровнями потока генов.
География играет ключевую роль в этом процессе. Когда популяции пространственно изолированы, они больше не обмениваются генами и могут дивергировать. Побочным продуктом этого расхождения является эволюция внутренних репродуктивных барьеров — признаков, которые препятствуют скрещиванию видов и сигнализируют о завершении видообразования. Аллопатрическое видообразование – это возникновение новых видов в пространственно изолированных популяциях. Аллопатрическое видообразование является самой простой и легкой моделью видообразования, потому что между популяциями нет потоков генов, которые могли бы подорвать дивергенцию. При наличии достаточного количества времени ожидается аллопатрическое видообразование, и широко признано, что это наиболее распространенный способ видообразования. Действительно, аллопатрическое видообразование было описано Койном и Орром в Вид (2004 г.) как «настолько правдоподобный, что вряд ли стоит его документировать». Видообразование, конечно, может происходить и в других географических контекстах, с разными уровнями потока генов. Одна панмиктическая популяция может быть разделена на две изолированные популяции посредством разрушительного отбора, известного как симпатрическое видообразование, или новые виды могут возникнуть из популяций, которые не перекрываются напрямую, но находятся достаточно близко к обмену генами, известному как парапатрическое видообразование. Эти три географических режима представляют собой разные точки на континууме пространственной изоляции, определяющей связность или объем потока генов между дивергирующими популяциями. Однако вряд ли это дискретные категории, и термин «видообразование с потоком генов» (также «дивергенция с потоком генов») часто используется для обозначения любой географической модели видообразования, отличной от строгой аллопатрии. В этой статье будет представлен обзор различных географических контекстов видообразования с акцентом на случаи, когда пространственная изоляция варьируется от полной (аллопатрическое видообразование) до некоторой формы контакта между дивергирующими популяциями.
Одна панмиктическая популяция может быть разделена на две изолированные популяции посредством разрушительного отбора, известного как симпатрическое видообразование, или новые виды могут возникнуть из популяций, которые не перекрываются напрямую, но находятся достаточно близко к обмену генами, известному как парапатрическое видообразование. Эти три географических режима представляют собой разные точки на континууме пространственной изоляции, определяющей связность или объем потока генов между дивергирующими популяциями. Однако вряд ли это дискретные категории, и термин «видообразование с потоком генов» (также «дивергенция с потоком генов») часто используется для обозначения любой географической модели видообразования, отличной от строгой аллопатрии. В этой статье будет представлен обзор различных географических контекстов видообразования с акцентом на случаи, когда пространственная изоляция варьируется от полной (аллопатрическое видообразование) до некоторой формы контакта между дивергирующими популяциями.
Общие сведения
География видообразования находится в центре внимания исследований видообразования. В результате большинство основных текстов по видообразованию посвящают этой теме несколько разделов или глав. На сегодняшний день наиболее полным обзором является Coyne and Orr 2004. С тех пор в области видообразования было сделано много достижений, но он остается отличным источником для понимания истории этой области. Добжанский 1937 является одним из самых ранних обсуждений географии видообразования и подчеркивает семантические трудности определения видообразования только с точки зрения географии. 19 мая42 и Mayr 1963 несколько догматически канонизируют преобладающую роль аллопатрии в видообразовании, представляя обширные доказательства ее важности для многих организмов. Грант 1981 расширяет эти синтезы, предоставляя углубленный обзор видообразования у растений. После этих основополагающих книг вышли антологии Otte and Endler 1989 и Rice, et al. 1998 г., в которых представлены обновления по основным темам, включая географию видообразования. Avise 2000 обрисовал в общих чертах изучение генетической изменчивости в пространственном и филогенетическом контексте и обеспечил методологическую основу для многих современных исследователей географии видообразования. Две недавние книги, Price 2008 и Nosil 2012, имеют более узкую направленность, но представляют собой самые свежие обзоры в этой области.
Avise 2000 обрисовал в общих чертах изучение генетической изменчивости в пространственном и филогенетическом контексте и обеспечил методологическую основу для многих современных исследователей географии видообразования. Две недавние книги, Price 2008 и Nosil 2012, имеют более узкую направленность, но представляют собой самые свежие обзоры в этой области.
Avise, JC 2000. Филогеография: история и формирование видов . Кембридж, Массачусетс: Гарвардский ун-т. Нажимать.
DOI: 10.2307/j.ctv1nzfgj7
Хотя эта книга не посвящена видообразованию как таковому , она явилась крупным шагом вперед в изучении генетической изменчивости в космосе. Эвис задокументировал соответствие между прошлыми и настоящими географическими барьерами и генетическими разрывами внутри и между видами. Во многих примерах генетические разрывы совпадают с географическими особенностями у многих различных видов, что указывает на общие биогеографические способы аллопатрического видообразования.
Койн, Дж. А. и Х. А. Орр. 2004. Вид . Сандерленд, Массачусетс: Sinauer Associates.
Всесторонний обзор литературы по видообразованию. Обсуждение географии видообразования тщательное, с главой, посвященной аллопатрическому и парапатрическому видообразованию, и отдельной главой, посвященной симпатрическому видообразованию.
Добжанский Т. 1937. Генетика и происхождение видов . Нью-Йорк: Колумбийский ун-т. Нажимать.
Содержит обзор изучения географии видообразования. Добжанский подчеркнул семантические трудности определения видообразования с точки зрения географии и предположил, что видообразование может быть лучше определено балансом отбора и потока генов. Впоследствии эта книга была дополнена вторым изданием в 1982 г.
Грант, В., 1981 г. Нью-Йорк: Колумбийский ун-т. Нажимать.
DOI: 10.7312/gran92318
Подробный обзор видообразования у растений, включая обсуждение аллопатрического видообразования.
Mayr, E. 1942. Систематика и происхождение видов . Нью-Йорк: Колумбийский ун-т. Нажимать.
В этой книге изложены обширные аргументы в пользу того, почему аллопатрическое видообразование важно для многих различных организмов, и подчеркнута главная роль географической изоляции в видообразовании.
Mayr, E. 1963. Виды животных и эволюция . Кембридж, Массачусетс: Гарвардский ун-т. Нажимать.
DOI: 10.4159/harvard.9780674865327
Обновленная версия Mayr 1942, в которой автор расширяет свое обсуждение географии видообразования и приводит больше примеров и непреклонных аргументов в пользу преобладания аллопатрического видообразования.
Носиль, П. 2012. Экологическое видообразование . Нью-Йорк: Оксфордский ун-т. Нажимать.
DOI: 10.1093/acprof:osobl/9780199587100.001.0001
Самая последняя книга о видообразовании, но особое внимание уделено роли экологии в видообразовании.
Хороший ресурс для относительно недавнего взгляда на поле.Отте Д. и Дж. А. Эндлер, ред. 1989. Видообразование и его последствия . Сандерленд, Массачусетс: Sinauer Associates.
Обзор области видообразования. Многие главы этой книги представляют собой классические работы в области видообразования и являются хорошим ресурсом для изучения истории этой области.
Цена, т. 2008 г. Видообразование у птиц . Гринвуд-Виллидж, Колорадо: Робертс.
Хотя эта книга полностью посвящена птицам, она охватывает многие основные темы видообразования. Особое значение для географии видообразования имеют главы, посвященные географической изменчивости, географической изоляции и островному эндемизму.
Rice, W.R., D.J. Howard, and S.H. Berlocher, eds. 1998. Бесконечные формы: виды и видообразование . Нью-Йорк: Оксфордский ун-т. Нажимать.
Обновленный обзор области видообразования после Otte and Endler 1989.
Эта коллекция посвящена Гаю Бушу, чей вклад в изучение симпатрического видообразования яблоневой мухи Rhagoletis pomonella поставил под сомнение мнение Майра о том, что аллопатрическое видообразование единственный важный способ видообразования. Есть много важных глав, посвященных географии видообразования.


 Хороший ресурс для относительно недавнего взгляда на поле.
Хороший ресурс для относительно недавнего взгляда на поле.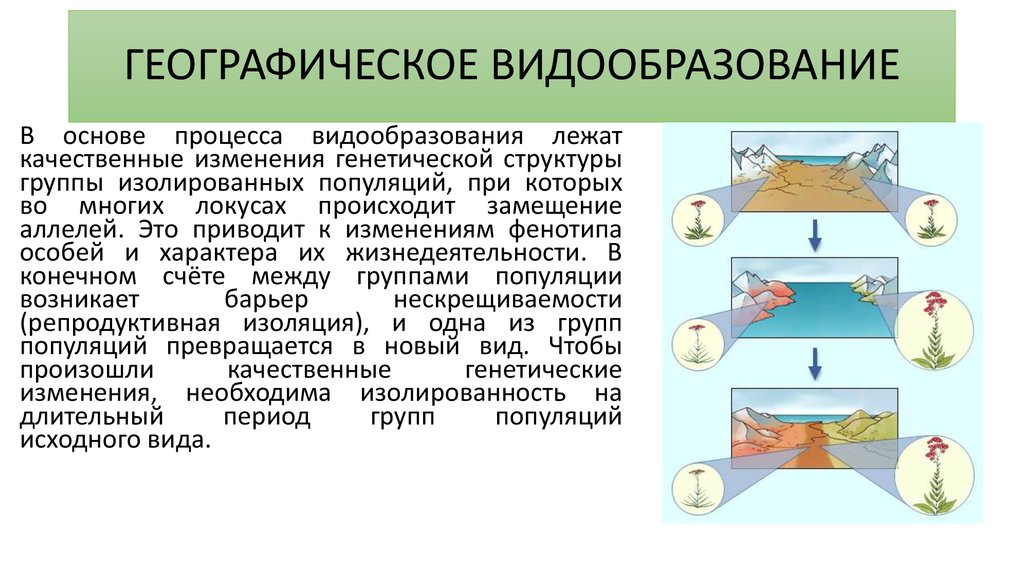 Эта коллекция посвящена Гаю Бушу, чей вклад в изучение симпатрического видообразования яблоневой мухи Rhagoletis pomonella поставил под сомнение мнение Майра о том, что аллопатрическое видообразование единственный важный способ видообразования. Есть много важных глав, посвященных географии видообразования.
Эта коллекция посвящена Гаю Бушу, чей вклад в изучение симпатрического видообразования яблоневой мухи Rhagoletis pomonella поставил под сомнение мнение Майра о том, что аллопатрическое видообразование единственный важный способ видообразования. Есть много важных глав, посвященных географии видообразования.наверх
Пользователи без подписки не могут видеть весь контент на эта страница. Пожалуйста, подпишитесь или войдите.
Как подписаться
Oxford Bibliographies Online доступен по подписке и с бессрочным доступом к учреждениям. Чтобы получить дополнительную информацию или связаться с торговым представителем Oxford, нажмите здесь.
Эволюционная биология
- Об эволюционной биологии »
- Познакомьтесь с редакционной коллегией »
Перейти к другим статьям:
Артикул
Вверх
- Приспособление
- Адаптивное излучение
- Альтруизм
- Древняя ДНК
- Поведенческая экология
- Канализация и надежность
- Рак, Эволюционные процессы в
- Смещение персонажа
- Коэволюция
- Познание, Эволюция
- Ограничения, Эволюция
- Современная эволюция
- Конвергентная эволюция
- Сотрудничество и конфликт: от микробов к людям
- Совместное разведение насекомых и позвоночных
- креационизм
- Загадочный женский выбор
- Дарвин, Чарльз
- дарвинизм
- Болезнь Вирулентность, Эволюция
- Диверсификация, зависящая от разнообразия
- Экологическое видообразование
- Эндосимбиоз
- Эпигенетика и поведение
- Эпистаз и эволюция
- Эусоциальные насекомые как модель для понимания альтруизма, Ко. ..
- Эусоциальность
- Доказательства эволюции,
- Эволюция
- Эволюция и развитие: гены и мутации, лежащие в основе…
- Эволюция и развитие индивидуальных поведенческих вариаций…
- Эволюция, Культура
- Эволюция систем спаривания животных
- Эволюция устойчивости к антибиотикам
- Эволюция новых генов
- Эволюция систем спаривания растений
- Эволюция специализации
- Эволюционная биология старения
- Эволюционная биомеханика
- Эволюционные вычисления
- Эволюционная биология развития
- Эволюционная экология сообществ
- Экспериментальная эволюция
- Вымирание
- Полевые исследования естественного отбора
- Окаменелости
- Эффект основателя
- Выбор в зависимости от частоты
- грибы, эволюция
- Дублирование генов
- Экспрессия генов, эволюция
- Генный поток
- Генетика, Экология
- Эволюция генома
- Географическая вариация
- Постепенность
- Выбор группы
- гетерохрония
- гетерозиготность
- История эволюционной мысли, 1860–1925 гг.
- История эволюционной мысли до Дарвина
- История эволюционной мысли с 1930 года
- Экология поведения человека
- Эволюция человека
- Гибридное видообразование
- Гибридные зоны
- Выявление геномной основы, лежащей в основе фенотипических вариаций…
- Инбридинг и инбредная депрессия
- Инклюзивный фитнес
- Инновации, Эволюция
- Острова как эволюционные лаборатории
- Выбор родственников
- Наземные растения, эволюция
- Ландшафтная генетика
- Пейзажи, Адаптивные
- Язык, Эволюция
- Градиент широтного разнообразия, The
- Макроэволюция
- Макроэволюционные темпы
- Мужской-Мужской конкурс
- Массовое вымирание
- Выбор помощника
- Материнские эффекты
- Медицина, Эволюция
- Мейотический драйв
- Мимикрия
- Современный синтез, The
- Молекулярные часы
- Молекулярная филогенетика
- Скорость мутации и спектр
- Мутуализм, Эволюция
- Естественный отбор в человеческих популяциях
- Естественный отбор в геноме, обнаружение
- Нейтральная теория
- Новая Зеландия, Эволюционная биогеография
- Строительство ниши
- Эволюция ниши
- Нечеловеческие животные, культурная эволюция в
- Происхождение и ранняя эволюция животных
- Происхождение эукариот
- Происхождение жизни,
- Парадокс секса
- Родительская забота, эволюция
- Партеногенез
- Различия личности, эволюция
- Фенотипическая пластичность
- Филогенетические сравнительные методы и тесты макроэволюци.
 ..
..
Leave A Comment