
Современные высшие растения возникли в результате сдвига экспрессии генов
Для жизненного цикла всех высших растений характерно чередование гаплоидных и диплоидных фаз, имеющих, соответственно, одну или две копии генома в каждой клетке. Гаплоидная фаза жизненного цикла растений называется гаметофитом, диплоидная — спорофитом. Судя по всему, на первых порах в жизненном цикле высших растений преобладал гаметофит, а спорофит был лишь зависимой «надстройкой» (такая ситуация сохраняется сейчас у мхов). Но у большинства современных высших растений спорофит живет самостоятельно и является главной жизненной формой: все папоротники, деревья, травы — спорофиты. Предложена гипотеза, согласно которой переход от крайне простого и зависимого спорофита к сложному и автономному произошел в результате переноса на спорофит экспрессии нескольких групп регуляторных генов, которые вначале обеспечивали развитие гаметофита. Это — генетический механизм важнейшего эволюционного события, изменившего облик Земли.
Жизненные циклы и эволюция
Любой живой организм, от вируса до слона, человека или дуба, обязательно проходит жизненный цикл — устойчивую последовательность фаз (или стадий), часть из которых в норме сопровождается размножением. Чередование жизненных циклов равносильно смене поколений. По ходу этого чередования жизненные циклы «редактируются», и вносимые в них небольшие изменения слагаются в историческое развитие жизни — эволюцию (рис. 1). Попросту говоря, биологическая эволюция — это сумма изменений жизненных циклов. Сопряжение жизненного цикла и эволюции — важнейшая особенность, свойственная всем живым системам и только им (А. С. Раутиан, 1993. О природе генотипа и наследственности). Одна из самых интересных проблем, с которыми имеют дело биологи в этой области — происхождение и эволюция жизненного цикла наземных растений.
Жизненный цикл, состоящий из повторяющихся стадий, есть абсолютно у любого живого существа. Но на этом однообразие кончается: например, жизненные циклы вируса и многоклеточного животного не имеют между собой почти ничего общего («почти» — потому что молекулярные механизмы копирования и считывания наследственной информации у них все же едины).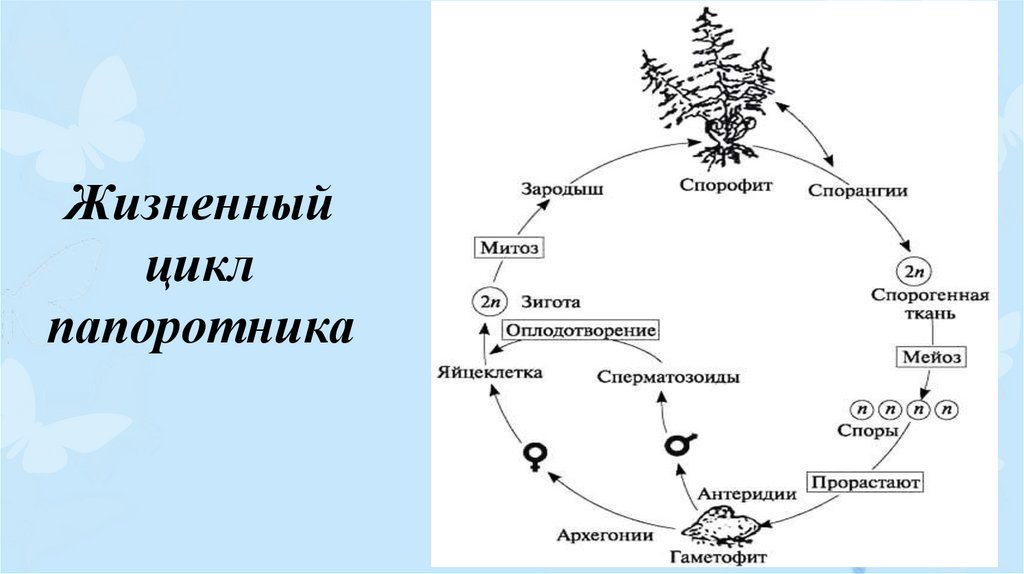
Есть три типа жизненных циклов эукариот, отличающихся способом чередования гаплоидной и диплоидной фаз.
1) Жизненный цикл, где «главная», питающаяся и растущая, стадия, — гаплоидная. В этом случае гаметы образуются митозом — обычным «повседневным» делением, в результате которого каждая клетка дает две себе подобных. Гаметы сливаются в зиготу, которая сразу же делится мейозом, вновь давая гаплоидные клетки. Такой жизненный цикл называется жизненным циклом с зиготической редукцией. Никаких диплоидных стадий, кроме зиготы, в нем нет.
Примеры обладателей жизненного цикла с зиготической редукцией — одноклеточная зеленая водоросль хламидомонада, колониальная зеленая водоросль вольвокс, нитчатая зеленая водоросль улотрикс.
2) Жизненный цикл, где питающаяся и растущая стадия — диплоидная. В этом случае зигота делится митозом, давая диплоидные клетки, а мейоз происходит только при образовании гамет. Такой жизненный цикл называется жизненным циклом с гаметической редукцией. Все стадии, кроме гамет, в нем диплоидные.
Все стадии, кроме гамет, в нем диплоидные.
Примеры обладателей жизненного цикла с гаметической редукцией — бурая водоросль фукус и все многоклеточные животные, включая, разумеется, и человека.
3) Жизненный цикл, в котором есть две питающихся и растущих стадии — одна гаплоидная, а другая диплоидная. В этом случае некоторые клетки диплоидного организма делятся мейозом, но дают не гаметы, а споры — гаплоидные клетки, любая из которых может дать новый организм без оплодотворения, то есть ни с кем не сливаясь. Из споры вырастает гаплоидный организм, который образует гаметы путем обычного деления — митоза. Из слившихся гамет образуется зигота, дающая диплоидный организм, после чего цикл повторяется. Такой жизненный цикл называется
У высших растений и водорослей, имеющих жизненный цикл со спорической редукцией, диплоидную стадию (производящую споры) принято называть спорофитом, а гаплоидную стадию (производящую гаметы) гаметофитом.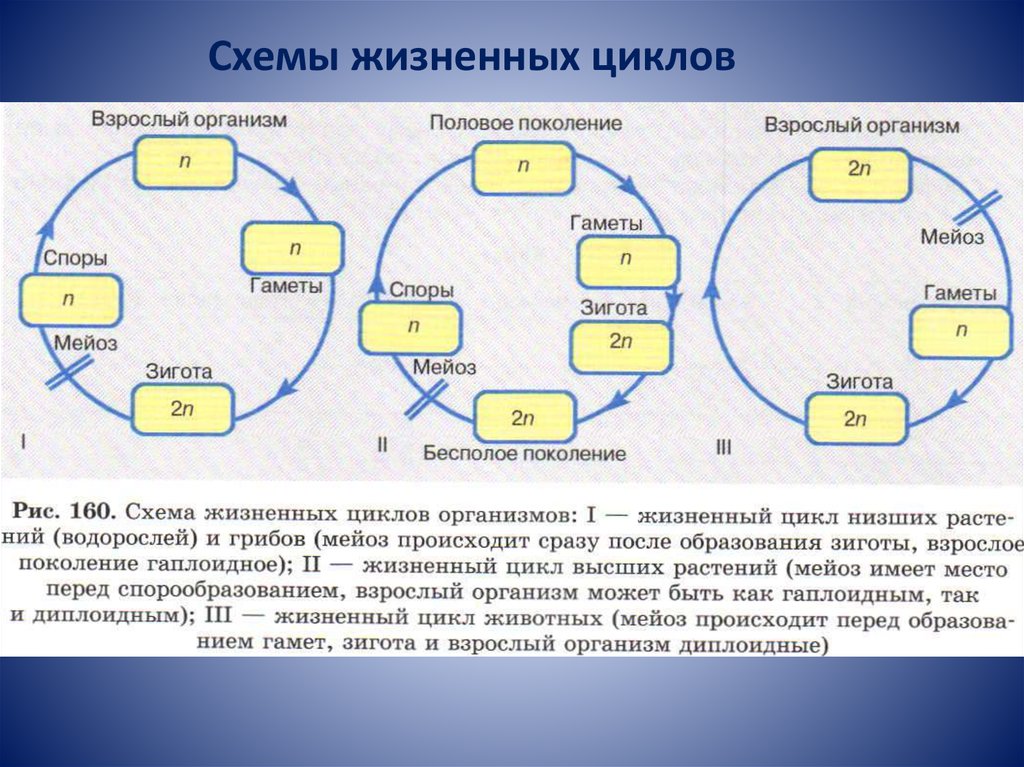
Например, у папоротников (вроде часто встречающегося в наших лесах орляка) спорофит — это хорошо всем знакомое растение с подземным стеблем, мощными корнями и крупными сложными перистыми листьями, а гаметофит — маленькая зеленая пластинка, растущая прямо на почве и называемая заростком. У хвощей ситуация аналогичная: спорофит — это сам хвощ, а гаметофит — маленький пластинчатый заросток, найти который неспециалисту очень трудно. У плаунов гаметофит зачастую подземный, бесцветный и неспособный фотосинтезировать — на пушистый ветвящийся зеленый спорофит, который мы встречаем в лесу и называем плауном в обычной жизни, он ничуть не похож.
Наконец, у голосеменных и цветковых растений женский гаметофит скрыт внутри семени, которое само является ничем иным, как органом спорофита, и из которого сразу вырастает спорофит следующего поколения.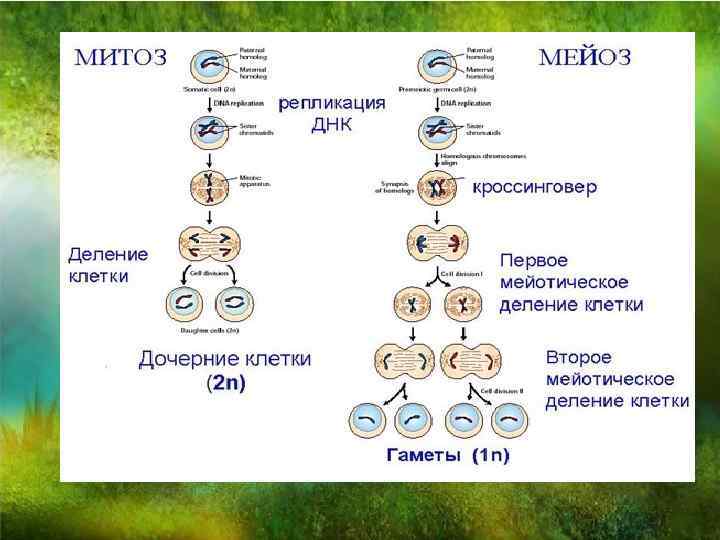
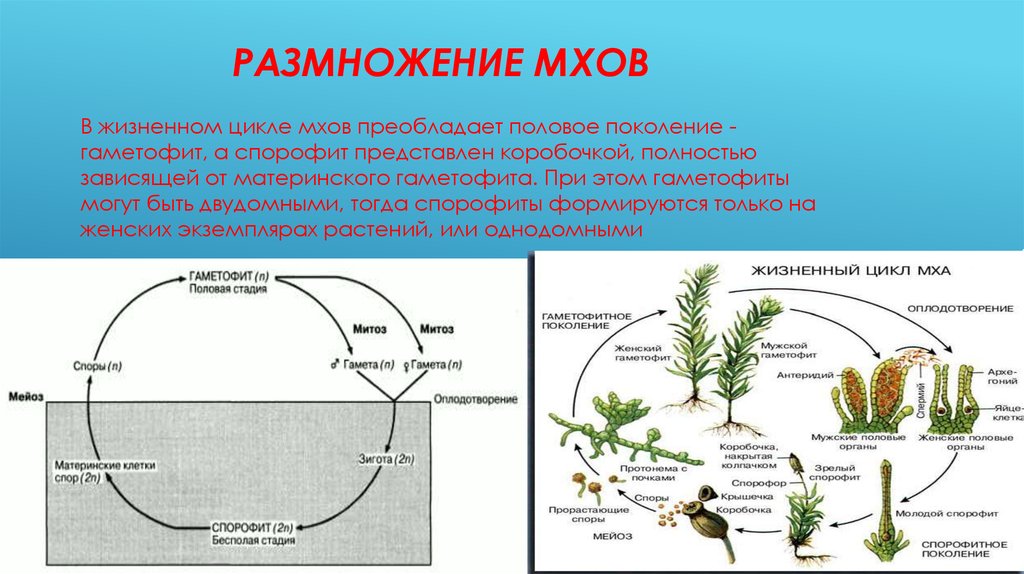
Единственная современная группа высших растений, где гаметофит доминирует над спорофитом, — мхи. Основная зеленая часть растения мха — это гаплоидный гаметофит. Что касается диплоидного спорофита, то он у мхов представляет собой стволик с коробочкой, который растет прямо на гаметофите и питается за его счет (фотосинтезировать он, как правило, неспособен).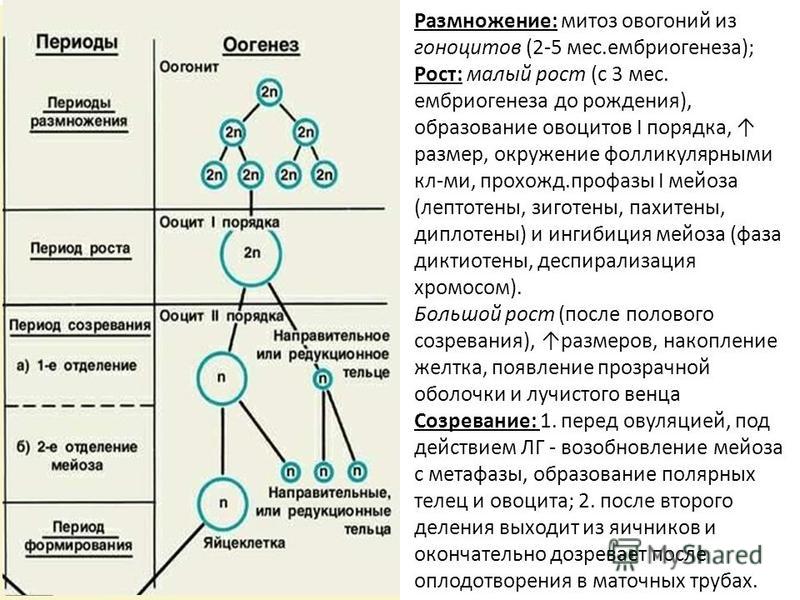
Мы видим, что жизненный цикл высших растений гораздо сложнее привычного нам жизненного цикла животных. Чередование гаплоидной и диплоидной многоклеточных фаз, еще и сильно отличающихся друг от друга, — чисто растительное «изобретение», животным оно неведомо. А поскольку земные биологи сами являются животными, то неудивительно, что разобраться в этом им удалось далеко не сразу — путаница продолжалась до середины XIX века, когда немецкий ботаник Вильгельм Гофмейстер (Friedrich Wilhelm Benedikt Hofmeister) сумел правильно «расшифровать» жизненный цикл цветковых.
А теперь зададимся вопросом о том, как этот жизненный цикл возник.
Сценарий происхождения высших растений
Начнем с самого начала. Высшие растения, несомненно, произошли от водорослей. Какой же жизненный цикл был у этих водорослей-предков?
Ответ известен.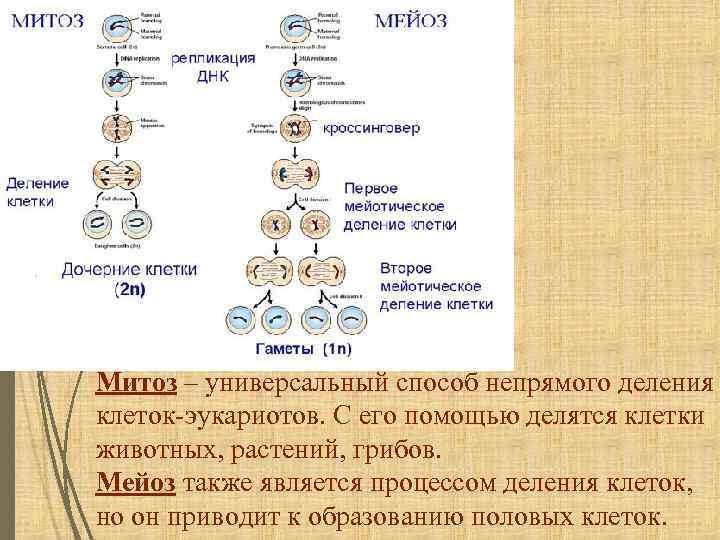 Скорее всего, у них был жизненный цикл с зиготической редукцией, то есть с единственной гаплоидной многоклеточной стадией (без спорофита). Из молекулярной систематики это следует вполне однозначно. Зеленые водоросли со сменой гаметофита и спорофита существуют, но — вот ирония судьбы! — все они настолько далеки от высших растений, что никак не могут быть их предками. А вот те водоросли, которые к наземным растениям близки — харовые, колеохетовые, сцеплянки — все без исключения имеют жизненный цикл с зиготической редукцией, в котором единственной диплоидной стадией является зигота. Это означает, что высшие растения не унаследовали многоклеточный спорофит от предков, а «изобрели» его совершенно самостоятельно. Стадия спорофита стала своего рода «вставкой» в древний жизненный цикл, который исходно был целиком гаплоидным (кроме зиготы).
Скорее всего, у них был жизненный цикл с зиготической редукцией, то есть с единственной гаплоидной многоклеточной стадией (без спорофита). Из молекулярной систематики это следует вполне однозначно. Зеленые водоросли со сменой гаметофита и спорофита существуют, но — вот ирония судьбы! — все они настолько далеки от высших растений, что никак не могут быть их предками. А вот те водоросли, которые к наземным растениям близки — харовые, колеохетовые, сцеплянки — все без исключения имеют жизненный цикл с зиготической редукцией, в котором единственной диплоидной стадией является зигота. Это означает, что высшие растения не унаследовали многоклеточный спорофит от предков, а «изобрели» его совершенно самостоятельно. Стадия спорофита стала своего рода «вставкой» в древний жизненный цикл, который исходно был целиком гаплоидным (кроме зиготы).
Как именно это произошло? По последним молекулярным данным, самыми близкими «внешними» родственниками высших растений являются водоросли-сцеплянки (B. R. Ruhfel et al., 2014. From algae to angiosperms–inferring the phylogeny of green plants (Viridiplantae) from 360 plastid genomes). К этой группе относится, например, широко распространенная пресноводная нитчатая водоросль спирогира. Такие данные вполне согласуются с предположением, что предком наземных растений была нитчатая (или, возможно, пластинчатая) водоросль, жившая на мелководье либо на влажном берегу. «Тело» этой водоросли, как и у современных сцеплянок, было гаплоидным, а ее зигота сразу делилась мейозом на четыре неподвижные споры (апланоспоры), которые разносились течением или ветром, давая новые колонии.
R. Ruhfel et al., 2014. From algae to angiosperms–inferring the phylogeny of green plants (Viridiplantae) from 360 plastid genomes). К этой группе относится, например, широко распространенная пресноводная нитчатая водоросль спирогира. Такие данные вполне согласуются с предположением, что предком наземных растений была нитчатая (или, возможно, пластинчатая) водоросль, жившая на мелководье либо на влажном берегу. «Тело» этой водоросли, как и у современных сцеплянок, было гаплоидным, а ее зигота сразу делилась мейозом на четыре неподвижные споры (апланоспоры), которые разносились течением или ветром, давая новые колонии.
Согласно общим закономерностям экологии, в нестабильных условиях пересыхающей прибрежной зоны конкурентное преимущество должны были при прочих равных условиях получить r-стратеги — виды с короткими жизненными циклами, быстрым размножением и большим количеством потомства (G. L. Stebbins, G. J. C. Hill, 1980. Did multicellular plants invade the land?). В нашем случае выиграли те водоросли, у которых зигота до начала мейоза стала несколько раз делиться обычным способом — митозом, превращаясь в многоклеточную структуру.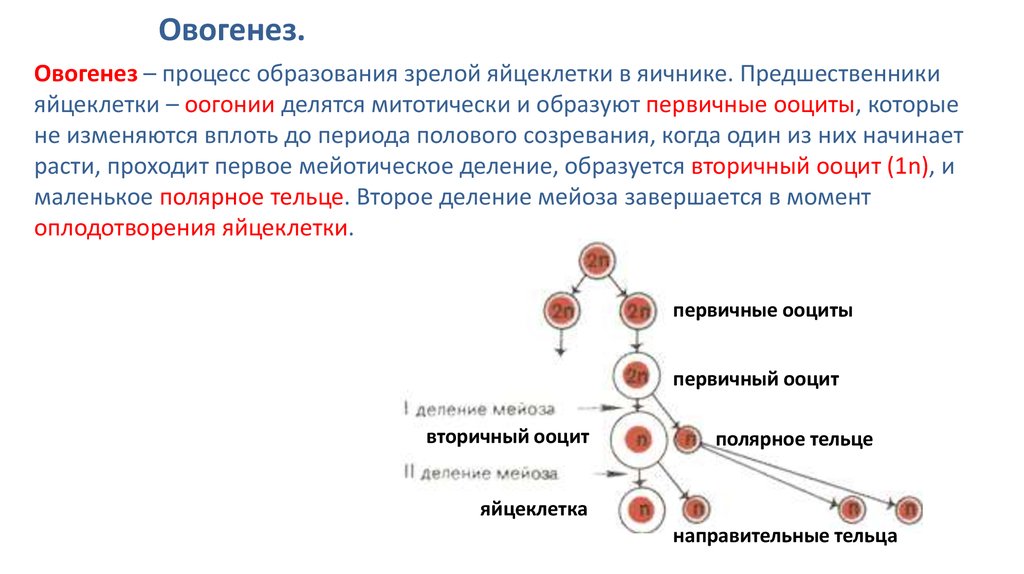 В результате из одной зиготы получалось не четыре споры, а много. Таким образом, одноклеточная зигота породила многоклеточный диплоидный спорофит (напомним, что при митозе плоидность клеток не меняется).
В результате из одной зиготы получалось не четыре споры, а много. Таким образом, одноклеточная зигота породила многоклеточный диплоидный спорофит (напомним, что при митозе плоидность клеток не меняется).
До появления сплошного растительного покрова реки на Земле не имели постоянных берегов, и почти вся суша представляла собой, по сути, сильно пересеченное мелководье (А. Г. Пономаренко, 1993. Основные события в эволюции биосферы). В этих условиях распространение спор ветром было гораздо эффективнее, чем их перенос водными течениями. Поэтому диплоидный спорофит вскоре принял форму стволика с приподнятой коробочкой, а споры стали воздушными — рассчитанными на дальний перенос ветром и покрытыми оболочкой, защищающей живые клетки от высыхания (L. E. Graham et al., 2000. The origin of plants: body plan changes contributing to a major evolutionary radiation). Однако из этого же эволюционного сценария следует, что диплоидный спорофит на первых порах был матротрофным: он рос прямо на гаметофите, питался за его счет и фактически составлял с ним единый организм (P.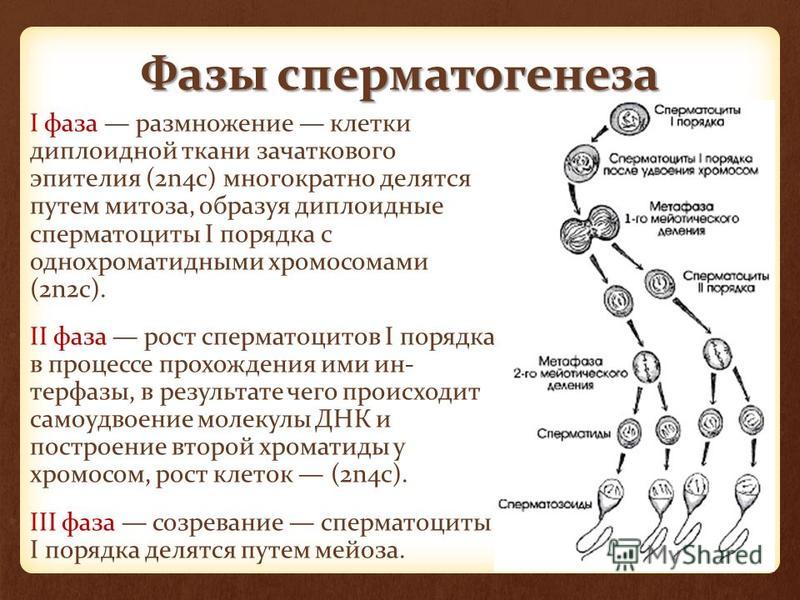 Kenrick, 1994. Alternation of generations in land plants: new phylogenetic and palaeobotanical evidence).
Kenrick, 1994. Alternation of generations in land plants: new phylogenetic and palaeobotanical evidence).
Именно такую картину мы видим у мхов — современных обладателей матротрофного спорофита в виде стволика с коробочкой, разбрасывающей воздушные споры. Есть ли основания считать, что мхи сохранили примитивное состояние, которое было когда-то свойственно всем высшим растениям? Да, молекулярная систематика прекрасно это подтверждает. Тут надо сказать, что все высшие растения делятся на две группы: мхи (в широком смысле слова) и сосудистые растения, к которым относятся папоротники, псилоты, хвощи, плауны, голосеменные и цветковые. Молекулярная генетика показывает, что мхи являются как минимум сестринской группой по отношению к сосудистым растениям (C. J. Cox et al., 2014. Conflicting phylogenies for early land plants are caused by composition biases among synonymous substitutions). Но многие авторы делают на основе молекулярных данных и более сильный вывод: мхи — это парафилетическая группа, то есть фактически целый эволюционный уровень, охватывающий в том числе и предков сосудистых растений (N. J. Wickett et al., 2014. Phylotranscriptomic analysis of the origin and early diversification of land plants). Если говорить совсем попросту, это означает, что сосудистые растения произошли от мхов.
J. Wickett et al., 2014. Phylotranscriptomic analysis of the origin and early diversification of land plants). Если говорить совсем попросту, это означает, что сосудистые растения произошли от мхов.
Следует заметить, что предок сосудистых растений, даже если он относился к мхам в широком смысле, совершенно не обязательно должен был выглядеть как современные листостебельные мхи (наподобие всем известного кукушкина льна). Гораздо вероятнее, что он напоминал антоцеротовые мхи, гаметофит которых — зеленая пластинка с лопастями, не разделенная на листья и стебель. Антоцеротовые мхи тоже дожили до наших дней, один их род — антоцерос — широко распространен в Европе (см. Из жизни антоцероса).
Итак, мы пришли к выводу, что у всех древнейших наземных растений спорофит был «надстройкой» на гаметофите, как у современных мхов (в широком смысле этого слова). Но как же, в таком случае, произошел переход от доминирования гаметофита к доминированию спорофита, которое у современных сосудистых растений стало просто подавляющим?
Чтобы в этом разобраться, надо взглянуть на то, как описанные события отображаются в палеонтологической летописи.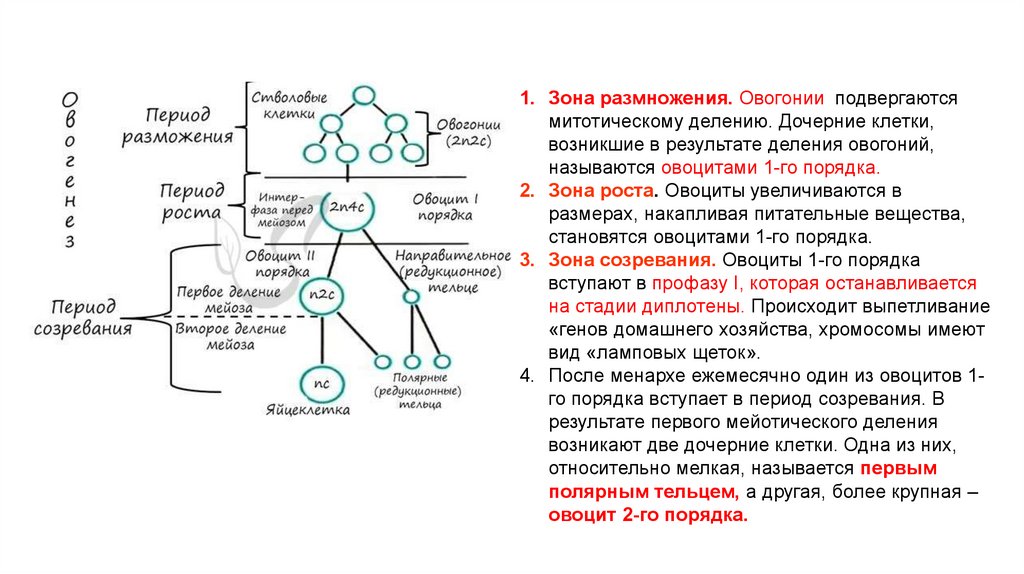
«Жизненные циклы, сохраненные в камне»
Новый обзор палеонтологического аспекта этой проблемы недавно опубликовал Пол Кенрик (Paul Kenrick) — известный британский палеоботаник, работающий в лондонском Музее естественной истории (Natural History Museum) и уже много лет занимающийся вопросами ранней эволюции наземных растений. «Жизненные циклы, сохраненные в камне» — название одной из главок его статьи.
Первые совершенно бесспорные остатки высших растений — это воздушные споры с плотной оболочкой и характерной трехлучевой щелью, которая образуется в процессе расхождения спор при мейозе (см. С. В. Мейен, 1981. Следы трав индейских). В наше время такие споры характерны для сосудистых растений и для некоторых мхов, в том числе и для антоцеротовых. В палеонтологической летописи самые древние споры с трехлучевой щелью появляются в конце ордовикского периода, около 450 миллионов лет назад (см. Палеонтологи уточнили время и место появления сосудистых растений, «Элементы», 18.
Парадокс в том, что сколько-нибудь достоверно определимые остатки «тел» высших растений появляются в летописи гораздо позже, чем их споры. Объясняется это просто: споры, с их прочными оболочками, изначально приспособлены к перенесению неблагоприятных условий, и поэтому сохраняются они гораздо лучше, чем зеленые гаметофиты или спорофиты (тем более что до появления древесины и других твердых растительных тканей тогда еще было далеко).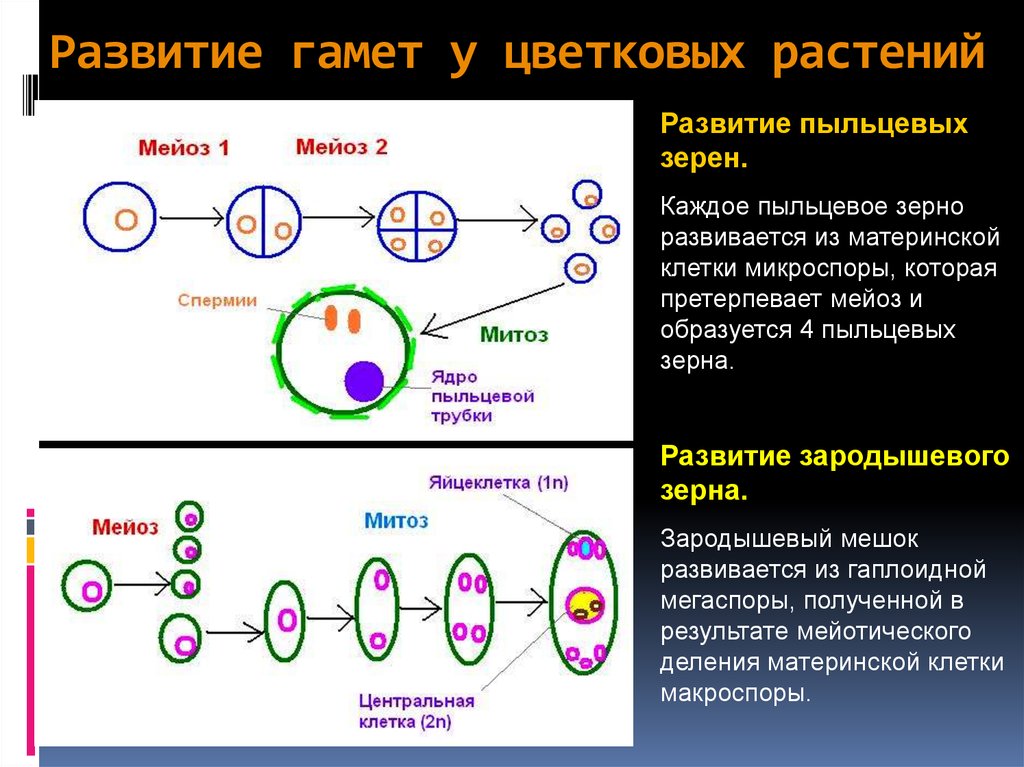 Так или иначе, это означает, что в начале эволюции высших растений была скрытая от нас фаза продолжительностью примерно 35 миллионов лет (C. H. Wellman, 2014. The nature and evolutionary relationships of the earliest land plants). На протяжении этой скрытой фазы весь жизненный цикл высших растений, кроме спор, остается палеонтологически «невидимым».
Так или иначе, это означает, что в начале эволюции высших растений была скрытая от нас фаза продолжительностью примерно 35 миллионов лет (C. H. Wellman, 2014. The nature and evolutionary relationships of the earliest land plants). На протяжении этой скрытой фазы весь жизненный цикл высших растений, кроме спор, остается палеонтологически «невидимым».
Достаточно целые остатки высших растений, по которым уже можно попытаться реконструировать их полный жизненный цикл, появляются в палеонтологической летописи только в начале девонского периода, примерно 407 миллионов лет назад. Именно такой возраст имеет флора знаменитого шотландского местонахождения Райни (Rhynie chert). Типичные обитатели этого местонахождения — риниофиты, стелющиеся наземные растения с ветвящимися осями, но без листьев и корней. До недавнего времени все их найденные более-менее полные экземпляры, как правило, считались спорофитами. Предполагалось, что гаметофиты риниофитов были маленькими нежными заростками, практически не имевшими шансов захорониться целыми. Иначе говоря, среди ботаников господствовало убеждение, что в жизненном цикле риниофитов уже доминировал спорофит — так же, как и у современных сосудистых растений.
Иначе говоря, среди ботаников господствовало убеждение, что в жизненном цикле риниофитов уже доминировал спорофит — так же, как и у современных сосудистых растений.
Эта точка зрения была обоснованно поставлена под сомнение в конце XX века, когда палеонтологи нашли на некоторых экземплярах девонских растений остатки многоклеточных женских и мужских половых органов — уж они-то по определению могут быть только на гаметофите (W. Remy et al., 1993. The gametophyte generation of some early devonian land plants). Действительно, сейчас палеонтологам известны гаметофиты не менее чем шести родов раннедевонских растений, причем не только из Райни, но и из других местонахождений. И, по мнению Пола Кенрика, разнообразие ископаемых гаметофитов еще недооценено. Надо оговорить, что сопоставление ископаемых гаметофитов с ископаемыми спорофитами тех же видов всегда остается более или менее предположительным, особенно если они захоронены отдельно друг от друга: на ископаемых экземплярах ведь не написано, относятся ли они к разным видам или к разным стадиям жизненного цикла одного и того же вида. Но, несмотря на это, кое-какие выводы насчет раннедевонских спорофитов и гаметофитов сделать уже можно (рис. 3).
Но, несмотря на это, кое-какие выводы насчет раннедевонских спорофитов и гаметофитов сделать уже можно (рис. 3).
Во-первых, раннедевонские гаметофиты по своему строению достаточно сильно похожи на спорофиты. Сходство так велико, что жизненный цикл этих видов вполне можно назвать изоморфным, как у упомянутой выше зеленой водоросли ульвы. Безошибочно различить фазы можно только благодаря тому, что гаметофит несет половые органы, а спорофит — спорангии, то есть коробочки, где формируются споры. Это в высшей степени интересное наблюдение, потому что ни у каких современных высших растений изоморфный жизненный цикл не встречается. Тут мы столкнулись с «вымершей» жизненной стратегией.
Во-вторых, по крайней мере у некоторых раннедевонских видов гаметофит, при общем сходстве со спорофитом, оказался существенно меньше него. Исходя из общих принципов биологии так и должно быть: ядра клеток гаметофита содержат ровно вдвое меньше ДНК, чем ядра клеток спорофита, а чем меньше в ядре ДНК, тем — при прочих равных условиях — будет меньше общий размер как каждой клетки, так и состоящего из этих клеток организма (см. Геномы хвостатых амфибий с самого начала были большими, «Элементы», 24.06.2015). Вот если бы соотношение размеров было обратным, это указывало бы или на ошибку исследователей, или на какую-то неизвестную закономерность. А так — все вполне ожидаемо.
Геномы хвостатых амфибий с самого начала были большими, «Элементы», 24.06.2015). Вот если бы соотношение размеров было обратным, это указывало бы или на ошибку исследователей, или на какую-то неизвестную закономерность. А так — все вполне ожидаемо.
Теперь сопоставим то, что мы знаем о жизненных циклах самых первых высших растений, предположительно возникших в ордовике, с тем, что мы знаем о жизненных циклах уже относительно продвинутых растений-риниофитов, которые появляются в палеонтологической летописи в раннем девоне.
Мы уже говорили, что самые первые высшие растения, скорее всего, были мхами. Это означает, что спорофит у них был матротрофным — привязанным к гаметофиту и зависимым от него. То, что известно о вероятных предках высших растений, полностью подтверждает эту гипотезу.
С другой стороны, у раннедевонских сосудистых растений спорофит и гаметофит, судя по всему, были автономны друг от друга и почти что изоморфны. Более того, спорофит у них крупнее гаметофита. Вероятно, это первый шаг к абсолютному преобладанию спорофита над гаметофитом, которое мы видим у сосудистых растений сейчас. Ведь, собственно говоря, все колоссальное многообразие современных сосудистых растений, покрывающих Землю, — это многообразие разросшихся спорофитов.
Вероятно, это первый шаг к абсолютному преобладанию спорофита над гаметофитом, которое мы видим у сосудистых растений сейчас. Ведь, собственно говоря, все колоссальное многообразие современных сосудистых растений, покрывающих Землю, — это многообразие разросшихся спорофитов.
Получается, что ключевым событием, сделавшим возможным само возникновение сосудистых растений, был переход от матротрофного спорофита мохообразных к автономному спорофиту, который в конце концов стал доминировать в жизненном цикле. Как же этот переход произошел? Каким образом спорофит обрел независимость?
«Перенос экспрессии» как эволюционный механизм
По мнению Пола Кенрика, ответ на этот вопрос может дать современная биология развития. Тесное взаимодействие биологии развития и палеонтологии, два-три десятилетия назад только намечавшееся, сейчас стало обычным делом в работе исследователей, стремящихся разобраться в механизмах крупных эволюционных событий.
Любая стадия развития любого многоклеточного организма характеризуется активностью (экспрессией) некоторых характерных для нее генов. Чаще всего исследователей интересуют регуляторные гены, которые, экспрессируясь на определенных стадиях в определенных частях тела, управляют через свои белковые продукты развитием этих частей. Непосредственный эффект такого регуляторного гена чаще всего (хотя и не всегда) состоит во «включении» или «выключении» каких-нибудь других генов посредством белка — фактора транскрипции. Очень часто подобные гены, «включающие» и «выключающие» друг друга, складываются в целые генно-регуляторные сети (gene regulatory networks, сокращенно GRN; см., например: У губок обнаружена генная сеть, которая могла бы управлять развитием глаз, «Элементы», 25.06.2013). Вся эта система генетической регуляции развития отлично представлена и у животных, и у растений. Но у растений (по крайней мере у высших) многоклеточных стадий, как мы теперь знаем, две — гаметофит и спорофит. Вполне можно было бы допустить, что их развитие управляется двумя отдельными группами генных сетей, не зависящих друг от друга.
Чаще всего исследователей интересуют регуляторные гены, которые, экспрессируясь на определенных стадиях в определенных частях тела, управляют через свои белковые продукты развитием этих частей. Непосредственный эффект такого регуляторного гена чаще всего (хотя и не всегда) состоит во «включении» или «выключении» каких-нибудь других генов посредством белка — фактора транскрипции. Очень часто подобные гены, «включающие» и «выключающие» друг друга, складываются в целые генно-регуляторные сети (gene regulatory networks, сокращенно GRN; см., например: У губок обнаружена генная сеть, которая могла бы управлять развитием глаз, «Элементы», 25.06.2013). Вся эта система генетической регуляции развития отлично представлена и у животных, и у растений. Но у растений (по крайней мере у высших) многоклеточных стадий, как мы теперь знаем, две — гаметофит и спорофит. Вполне можно было бы допустить, что их развитие управляется двумя отдельными группами генных сетей, не зависящих друг от друга.
Однако генетика развития растений показывает, что это не так. Например, существуют специфические гены, которые экспрессируются в ризоидах мхов и в корневых волосках цветковых растений (B. Menand et al., 2007. An ancient mechanism controls the development of cells with a rooting function in land plants). И ризоиды, и корневые волоски — это тончайшие выросты, обеспечивающие контакт подземной части растения с почвой: они «заякориваются» там, одновременно создавая поверхность для обмена водой и минеральными веществами. Нет ничего удивительного в том, что их развитие управляется одними и теми же регуляторными генами. Только ризоиды мхов находятся на гаметофите, а корневые волоски цветковых растений — разумеется, на спорофите. Маловероятно, чтобы гены, управляющие развитием тех и других, с самого начала имели двойную функцию. Исследователи уже давно предположили, что первоначально эти гены участвовали именно в развитии гаметофита, и только потом их экспрессия распространилась на спорофит, дав ему принципиально новые возможности.
Например, существуют специфические гены, которые экспрессируются в ризоидах мхов и в корневых волосках цветковых растений (B. Menand et al., 2007. An ancient mechanism controls the development of cells with a rooting function in land plants). И ризоиды, и корневые волоски — это тончайшие выросты, обеспечивающие контакт подземной части растения с почвой: они «заякориваются» там, одновременно создавая поверхность для обмена водой и минеральными веществами. Нет ничего удивительного в том, что их развитие управляется одними и теми же регуляторными генами. Только ризоиды мхов находятся на гаметофите, а корневые волоски цветковых растений — разумеется, на спорофите. Маловероятно, чтобы гены, управляющие развитием тех и других, с самого начала имели двойную функцию. Исследователи уже давно предположили, что первоначально эти гены участвовали именно в развитии гаметофита, и только потом их экспрессия распространилась на спорофит, дав ему принципиально новые возможности.
Группа генов с такой судьбой наверняка не одна. Аналогичные эволюционные сценарии предложены для регуляторных генов, контролирующих у высших растений развитие проводящих тканей и системы транспорта ауксина — важнейшего растительного гормона, управляющего ростом. Все эти гены исключительно важны для развития спорофита, но они экспрессируются и в гаметофите мхов, причем их функции там — приблизительно те же. Пол Кенрик делает твердый вывод: в самом начале эволюции сосудистых растений произошел «перенос» экспрессии этих генов с гаметофита на спорофит. Именно это позволило спорофиту приобрести сложную форму и стать совершенно автономным — с корнями, проводящими тканями и прочим (рис. 4, А).
Аналогичные эволюционные сценарии предложены для регуляторных генов, контролирующих у высших растений развитие проводящих тканей и системы транспорта ауксина — важнейшего растительного гормона, управляющего ростом. Все эти гены исключительно важны для развития спорофита, но они экспрессируются и в гаметофите мхов, причем их функции там — приблизительно те же. Пол Кенрик делает твердый вывод: в самом начале эволюции сосудистых растений произошел «перенос» экспрессии этих генов с гаметофита на спорофит. Именно это позволило спорофиту приобрести сложную форму и стать совершенно автономным — с корнями, проводящими тканями и прочим (рис. 4, А).
Может ли такое быть? Да, и не только у растений. Примеры аналогичного «переноса экспрессии» есть и у животных, хотя там они касаются не столько разных стадий (жизненный цикл животных для этого слишком беден), сколько разных органов одного и того же тела. Например, у современной кистеперой рыбы латимерии задний спинной и анальный плавники являются «мясистыми» — они имеют внутренний скелет, неожиданно напоминающий сложный внутренний скелет парных плавников (тех самых, которые соответствуют лапам наземных позвоночных).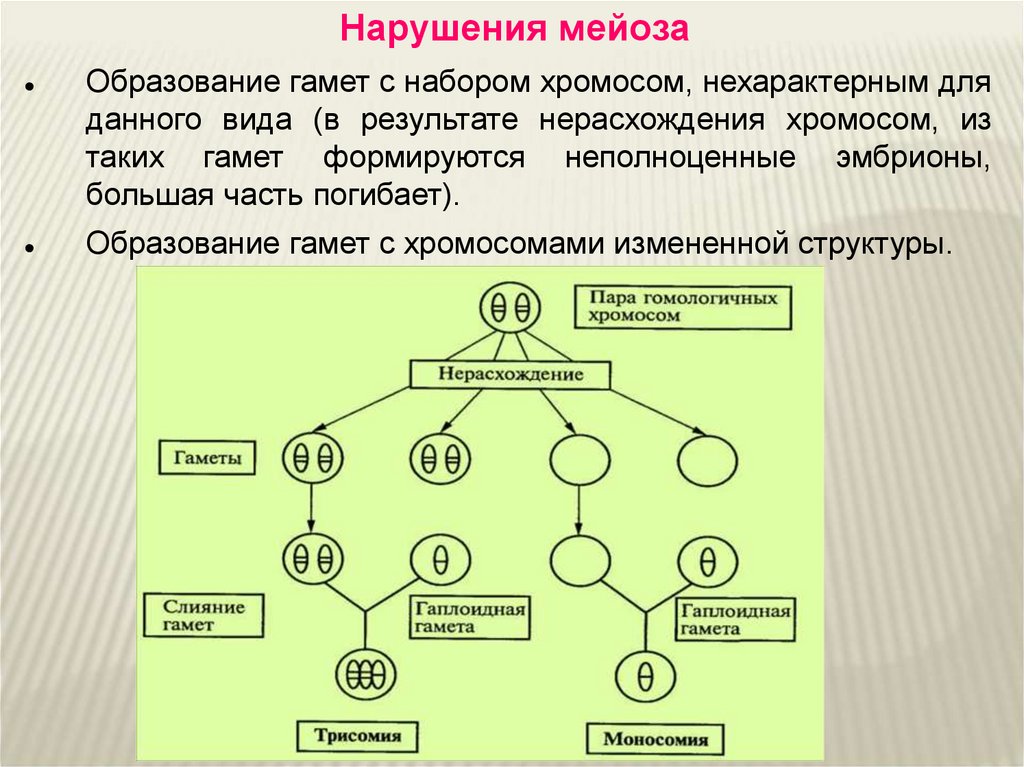 Для рыб в целом это необычный признак — как правило, непарные плавники у них имеют гораздо более простой скелет, чем парные (в переднем спинном плавнике у латимерии до сих пор так и есть). Предполагается, что у далеких предков латимерии в области заднего спинного и анального плавников почему-то включилась экспрессия определенной группы регуляторных генов, которые обычно управляют развитием только парных плавников (C. Tabin, E. Laufer, 1993. Hox genes and serial homology). Именно в результате «переноса экспрессии» у них и возникло анатомическое новшество (рис. 4, Б).
Для рыб в целом это необычный признак — как правило, непарные плавники у них имеют гораздо более простой скелет, чем парные (в переднем спинном плавнике у латимерии до сих пор так и есть). Предполагается, что у далеких предков латимерии в области заднего спинного и анального плавников почему-то включилась экспрессия определенной группы регуляторных генов, которые обычно управляют развитием только парных плавников (C. Tabin, E. Laufer, 1993. Hox genes and serial homology). Именно в результате «переноса экспрессии» у них и возникло анатомическое новшество (рис. 4, Б).
Если прав Пол Кенрик, то у предков сосудистых растений произошло нечто подобное — только в гораздо большем масштабе. «Перенос экспрессии» охватил там целую фазу жизненного цикла. В результате в спорофите запустилась программа развития, которая раньше была свойственна только гаметофиту, и жизненный цикл стал изоморфным — как у раннедевонских растений. В поддержке гаметофита «новая модель» спорофита уже не нуждалась, а потому она перешла к автономному существованию.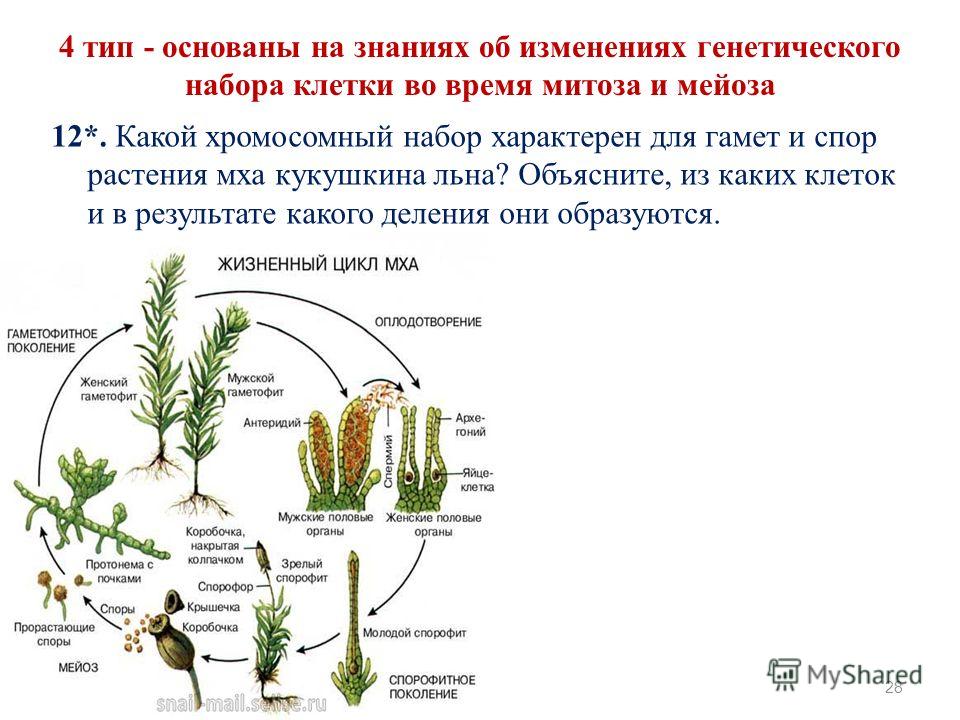 Еще позже стадия спорофита стала у сосудистых растений безусловно преобладающей. Растения пришли к доминированию в жизненном цикле диплоидной фазы, подобно животным, но абсолютно другим путем.
Еще позже стадия спорофита стала у сосудистых растений безусловно преобладающей. Растения пришли к доминированию в жизненном цикле диплоидной фазы, подобно животным, но абсолютно другим путем.
Интересно, что предполагаемая этим сценарием перестройка жизненного цикла растений могла произойти очень быстро. Она ведь на первых порах заключалась в изменении области действия уже существующей программы развития, а не в создании новой. Смещение областей экспрессии регуляторных генов может произойти даже за одно поколение — иной вопрос, будет ли оно поддержано естественным отбором (или хотя бы допущено им к существованию). На этом начальном этапе эволюция вполне может быть скачкообразной. Правда, чтобы результаты такого «скачка» охватили крупные группы организмов, они должны пройти долгую, длящуюся десятками миллионов лет, «подгонку и дошлифовку». Судя по всему, в эволюции сосудистых растений это и произошло.
Знаменитый генетик Рональд Фишер (Ronald Aylmer Fisher) не зря сравнивал эволюцию с настройкой оптического прибора.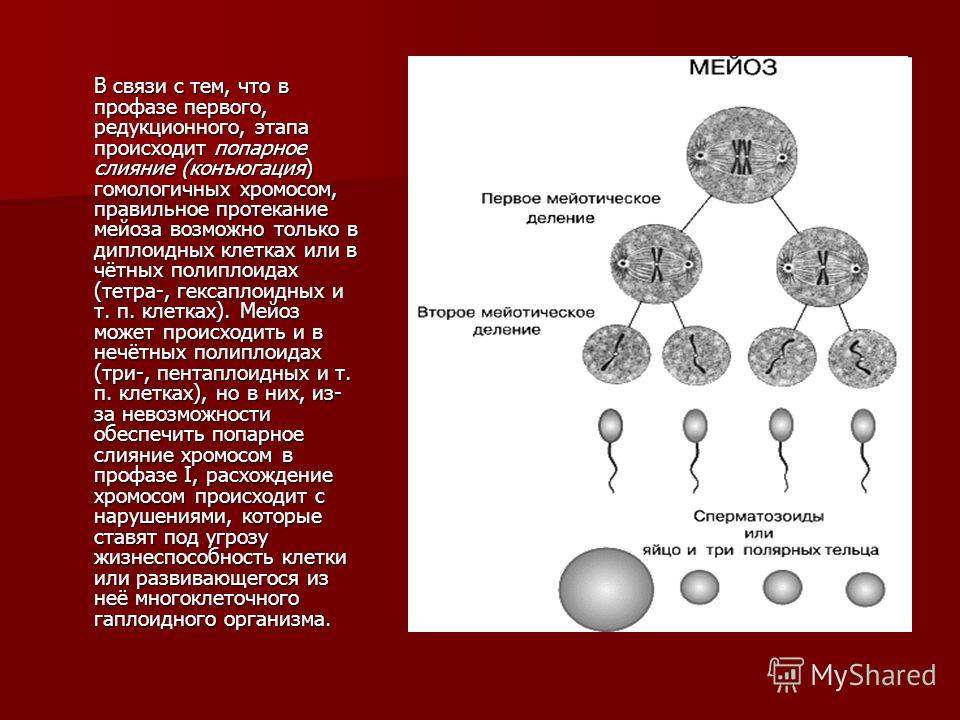 Ничто не мешает человеку, сидящему за микроскопом, резко изменить расположение линз, повернув макровинт. Но если он хочет что-то увидеть, то ему неизбежно придется после этого постепенно адаптировать прибор к своему зрению, последовательно совершая мелкие, требующие обратной связи пошаговые движения сначала макро-, а потом и микровинтом. Этот процесс занимает время, но без него хорошего результата не достичь. Вот и эволюция устроена похоже.
Ничто не мешает человеку, сидящему за микроскопом, резко изменить расположение линз, повернув макровинт. Но если он хочет что-то увидеть, то ему неизбежно придется после этого постепенно адаптировать прибор к своему зрению, последовательно совершая мелкие, требующие обратной связи пошаговые движения сначала макро-, а потом и микровинтом. Этот процесс занимает время, но без него хорошего результата не достичь. Вот и эволюция устроена похоже.
Источник: P. Kenrick. Changing expressions: a hypothesis for the origin of the vascular plant life cycle // Philosophical Transactions of Royal Society B. 2018. V. 373. № 1739. P. 20170149. DOI: 10.1098/rstb.2017.0149.
Сергей Ястребов
Тест по биологии по теме «Деление клетки»
Инструкция к тесту по теме «Деление клетки: митоз, мейоз»
Тест рекомендуется проводить при изучении итогового текста для закрепления полученных знаний
Тест имеет следующую структуру;
— Часть А содержит 15 заданий
-Часть В содержит 5 заданий
— Часть С содержит 10 заданий.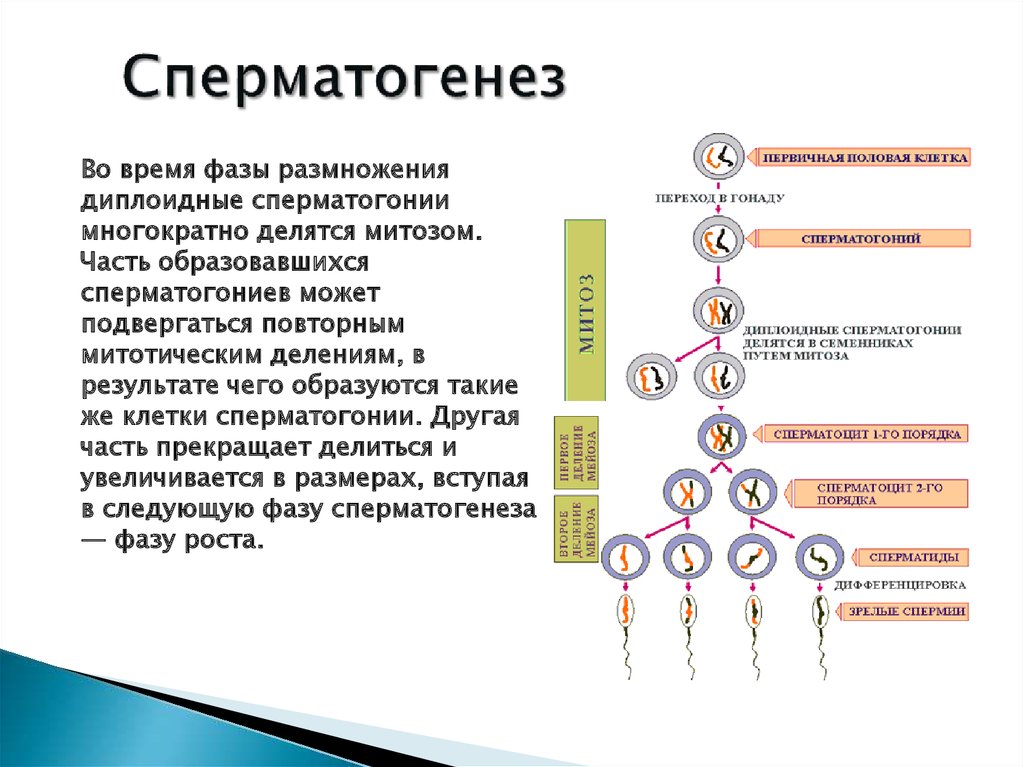
На выполнение теста отводится 40 мин. (1 урок). За каждое верно выполненное задание с выбором ответа (часть А) выставляется по 1 баллу; задание с выбором нескольких ответов(часть В) выставляется 2 балла, если допущена 1 ошибка – 1 балл, если более ошибок, то 0 баллов. За каждое верно выполненное задание с ответом «Да, нет» — 1 балл, всего – 10 баллов. За весь тест – 35 баллов
Успешность выполнения работы определяется в соответствии с ниже проведённой шкалой:
Удовлетворительно -27-29 баллов;
Хорошо – 32-30 баллов;
Удовлетворительно – 29-27 баллов.
Тест по теме «Деление клетки :митоз и мейоз»
Часть А. Выбрать один правильный вариант.
В какой фазе происходит расхождение хроматид к полюсам клетки?
В профазе 2) в метафазе 3) в анафазе
4) в телофазе
В чём заключается биологический смысл удвоения хромосом в интерфазе митоза ?
В процессе удвоения изменяется наследственная информация
Удвоенные хромосомы лучше видны
В результате удвоения хромосом наследственная информация новых половых клеток остаётся неизменной
В результате удвоения хромосом новые клетки содержат вдвое больше информации.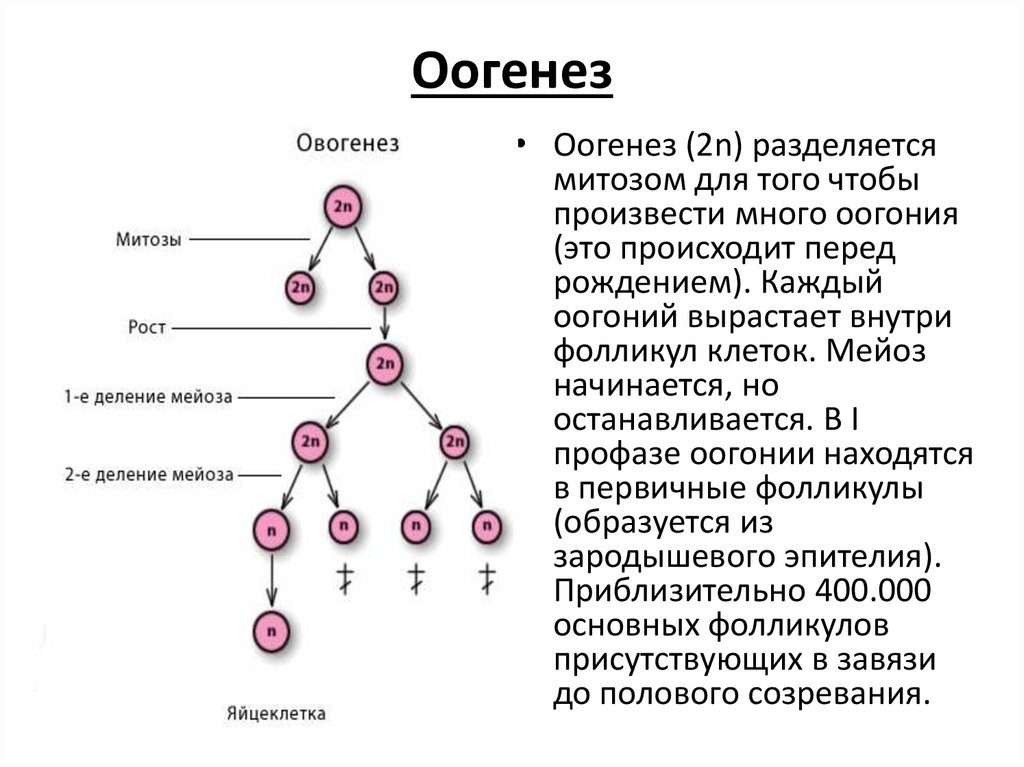
Укажите процессы, происходящие в интерфазе
Расхождение хромосом к полюсам клетки
Синтез белков, репликация ДНК, рост клетки
Формирование новых ядер, органоидов клетки
Митоз от греческого переводится как
верёвка 2)нить 3)появление 4) удвоение
В телофазе митоза
Увеличивается объём
) Хромосомы разделяются
3)Формируются ядра
4) Формируется веретено
Мейоз 2 заканчивается
Метафазой 2 3 ) Профазой 2
Телофазой 2 4) Анафазой 2
Мейоз 1 начинается
Интерфазой
Метафазой 1
Профазой 1
Анафазой 1
Мейоз от греческого переводится как
1)увеличение 2)уменьшение
3)деление
4) удвоение
9. В первой интерфазе происходит
1) уменьшение размеров клетки 2) удвоение ДНК
3) обмен участками
4) образование дочерних клеток
10. Что происходит в телофазе:
Что происходит в телофазе:
1) формирование веретена деления
2) формирование новых ядер и цитокинез
3) разделение хромосом
4) перемещение хромосом в центр клетки.
11. В анафазе митоза происходит:
1) расхождение двухроматидных хромосом к полюсам клетки
2) Спирализация хромосом
3) расхождение хроматид к полюсам клетки 4)деспирализация хромосом
12. Последовательность стадий митоза следующая:
1) профаза.анафаза,телофаза,метафаза
2) профаза,телофаза,метафаза.анафаза
3) профаза,метафаза.анафаза.телофаза 4)профаза,метафаза,телофаза.анафаза
13. При мейозе дочерние клетки имеют набор хромосом:
1)n 2) 2n 3)3n 4)4n
14. В результате митоза возникает
1)генетическое разнообразие видов
2) образование гамет
3)перекрёст хромосом
4) прорастание спор мха.
15. Что происходит в метафазе:
1) формирование веретена деления
2) формирование новых ядер и цитокинез
3) разделение хромосом
4)перемещение хромосом в центр клетки.
Часть В. Выбрать несколько правильных ответов.
В1 Во время метафазы 1 происходит
Спирализация и обмен участками гомологичных хромосом
Прикрепление к центромерам хромосом нитей веретена деления
Окончание формирования митотического аппарата
Коньюгация гомологичных хромосом
Выстраивание бивалентов хромосом на экваторе клетки с образованием метафазной пластинки
Деление хроматид и их расхождение к полюсам клетки
Расхождение гомологичных хромосом к полюсам клетки.
В 2. Выберите признаки мейоза :
В результате деления количество хромосом в клетке остаётся прежним
Процесс завершается в результате одного деления.
В результате одного деления образуется четыре ядра
Процесс проходит два этапа деления
Процесс обеспечивает рост и развитие организма, его бесполое размножение
Процесс обеспечивает образование гамет и половое размножение животных
В 3 Укажите процессы, в основе которых лежит митоз
1)мутации
2)рост
3)дробление зиготы
4) образование спермиев
5)регенерация тканей
6) оплодотворение
В4 Выберите процессы, происходящие в интерфазе митоза
Синтез белков
Уменьшение количества ДНК
Рост клетки
Удвоение хромосом
Расхождение хромосом
Деление ядра.
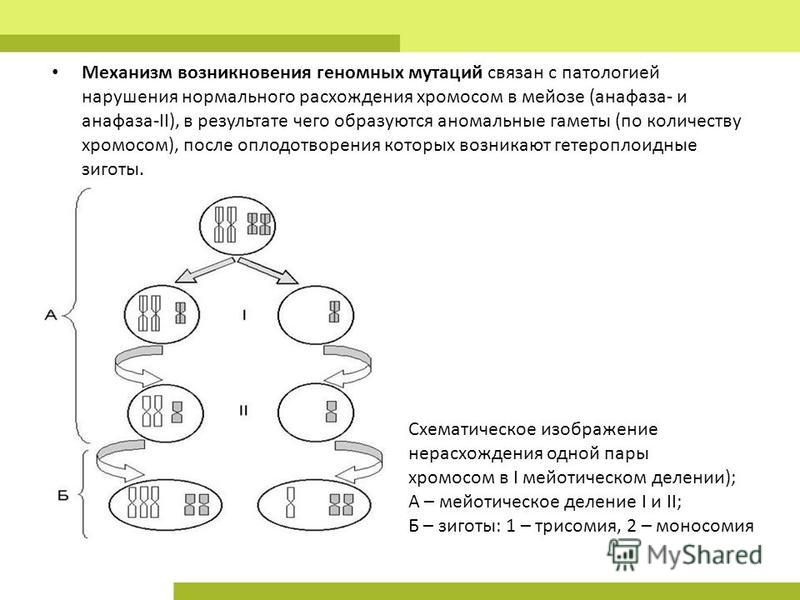
В 5 Биологическое значение мейоза заключается в (три правильных ответа)
1)поддержание постоянства видового числа хромосом
2) Создание условий для комбинативной изменчивости
3) произвольном расхождении родителских хромосом по гаметам
4)сохранение родительской наследственной информации без изменений
5) увеличении числа хромосом в клетке
6) сохранении полезных признаков организма при размножении
Часть С Верно ли суждение (если верно –отметить это суждение знаком «+»,если нет, то знаком «-»)
Интерфаза –это период клеточного цикла между делениями клетки
Кроссинговер – это перекрёст между гомологичными хромосомами, сопровождающийся обменом, соответствующими участками.
В результате мейоза образуются клетки точные копии материнской клетки.
Разделение цитоплазмы происходит во время анафазы митоза.
Бесполое размножение осуществляется при помощи гамет
Гаметы образуются в результате мейоза
В анафазе митоза хромосомы располагаются в экваториальной плоскости тела
Клетки делятся мейозом
В интерфазе происходит подготовка к делению и удвоение генетического материала.
В профазе происходит увеличение объёма ядра.
Ответы:
Часть А
1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 |
3 | 3 | 2 | 2 | 3 | 2 | 3 | 2 | 2 | 2 | 1 | 3 | 1 | 4 | 4 |
Часть В
В1 | В2 | В3 | В4 | В5 |
1,2,5 | 3,5,6 | 2,3,5 | 1,3,4 | 1,2,3 |
Часть С
1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 |
да | да | нет | нет | нет | да | нет | нет | да | да |
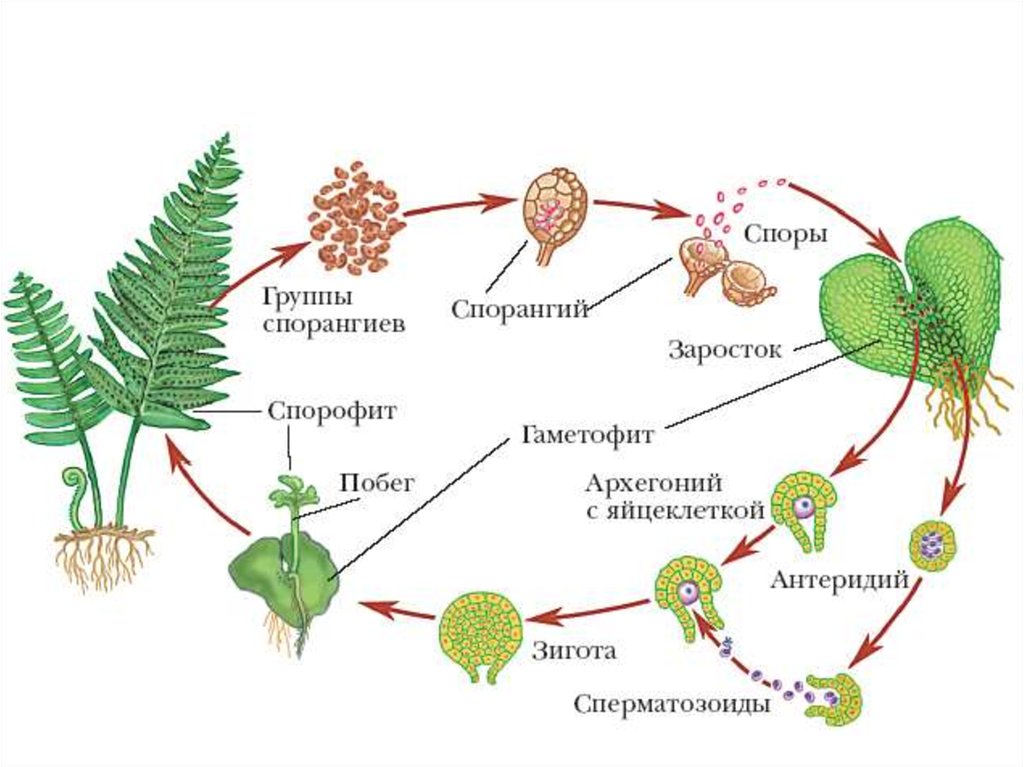
Жизненный цикл мха
В жизненном цикле мхов чередуются диплоидные и гаплоидные поколения, что является уникальным среди цветковых растений. Где происходит оплодотворение в жизненном цикле мха? Споры гаплоидны или диплоидны? Прокрутите до ключевых выводов, чтобы получить ответы, или начните сверху, чтобы узнать о жизненном цикле мха.
РЕПРОДУКЦИЯ МОХА
Как размножается мох?
Мхи имеют две формы размножения: половое размножение и бесполое / вегетативное размножение . Это верно для всех мохообразных .
Практически все цветковые растения диплоидны, но у мхов дело обстоит иначе. У мхов чередуются диплоидные поколения (как спорофиты) и гаплоидные поколения (как гаметофиты).
ПОЛОВОЕ РАЗМНОЖЕНИЕ
Вообще говоря, половое размножение — это процесс, при котором гены двух разных родителей смешиваются, чтобы произвести потомство с генетическим строением, сходным с генетическим составом каждого из родителей, но отличающимся от него.
Половое размножение мха (мохообразного) жизненного цикла чередуется между фазами диплоидного спорофита и гаплоидного гаметофита. Короче говоря, гаплоидные гаметофиты производят гаплоидные гаметы, которые могут быть сперматозоидами или яйцеклетками. Когда яйцеклетка и сперматозоид сливаются, они образуют диплоидную зиготу, которая вырастает в диплоидный спорофит. Спорофиты производят гаплоидные споры, содержащие генетическую информацию от обоих родителей-гаплоидных гаметофитов. Спора дает начало гаплоидному гаметофиту, завершающему цикл.
Гаметофит
Одно растение гаметофит мох может производить как сперму, так и яйцеклетки. Это может происходить на разных частях одного и того же растения: одна часть производит сперму, а другая часть — яйцеклетки. Однако растение обычно производит либо все органы, производящие сперму, либо все органы, производящие яйцеклетки, в любой момент времени. Таким образом, он не размножается сам с собой, способствуя генетической изменчивости. Женская структура для производства яйцеклеток известна как архегоний , а мужская структура для производства сперматозоидов известна как 9.
 0011 антеридий . Антеридии представляют собой крошечные, обычно на ножках, булавовидные или сферические образования. Архегонии представляют собой сосуды, похожие на бутылки, их стенки толщиной всего в одну клетку. Архегонии обычно формируются группами. Архегонии и антеридии обычно собраны в розетки листьев, похожие на цветы, называемые перихетиями . Удлиненные булавовидные клеточные нити, называемые Paraphyse , иногда встречаются на гаметофите, сохраняя воду и защищая антеридии песка архегоний от высыхания.
0011 антеридий . Антеридии представляют собой крошечные, обычно на ножках, булавовидные или сферические образования. Архегонии представляют собой сосуды, похожие на бутылки, их стенки толщиной всего в одну клетку. Архегонии обычно формируются группами. Архегонии и антеридии обычно собраны в розетки листьев, похожие на цветы, называемые перихетиями . Удлиненные булавовидные клеточные нити, называемые Paraphyse , иногда встречаются на гаметофите, сохраняя воду и защищая антеридии песка архегоний от высыхания.Когда антеридии созревают и цветок намокает от дождя, выделяются многочисленные антерозоиды (сперматозоиды / сперматозоиды). Антерозоиды способны передвигаться только под водой. Они плавают с помощью двух нитевидных хвостов. Некоторые успешно попадают на растения женского гаметофитного мха и химически привлекаются архегонием. Каждый архегоний содержит одно яйцо в вздутой части, называемой вентеральной частью .
Сперматозоиды попадают в архегоний через узкий канал в его горлышко . Оплодотворение происходит в архегонии с образованием диплоидной зиготы . Как только один архегоний в группе был оплодотворен, во многих случаях другие теряют способность к оплодотворению. Это вызвано ингибирующим гормоном, высвобождаемым из оплодотворенного архегония.Формирование зиготы начинается во второй фазе жизненного цикла мха, где зигота развивается в диплоидный спорофит (споровое растение).
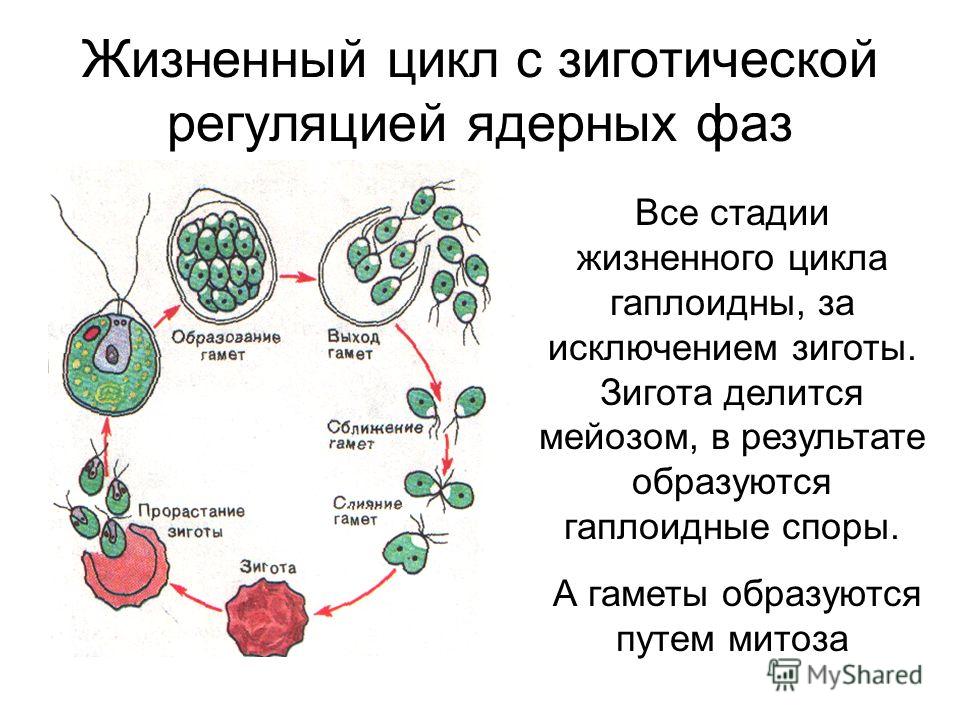 Сперматозоиды попадают в архегоний через узкий канал в его горлышко . Оплодотворение происходит в архегонии с образованием диплоидной зиготы . Как только один архегоний в группе был оплодотворен, во многих случаях другие теряют способность к оплодотворению. Это вызвано ингибирующим гормоном, высвобождаемым из оплодотворенного архегония.
Сперматозоиды попадают в архегоний через узкий канал в его горлышко . Оплодотворение происходит в архегонии с образованием диплоидной зиготы . Как только один архегоний в группе был оплодотворен, во многих случаях другие теряют способность к оплодотворению. Это вызвано ингибирующим гормоном, высвобождаемым из оплодотворенного архегония.Спорофит
Перевернутое фото мха после дождя. Спорофиты подняли руки в воздух, как будто им все равно. https://t.co/jDpYaHweHn pic.twitter.com/tFeU2ahzgO
— Меган Линч (@may_gun) 18 марта 2020 г.
После оплодотворения архегоний на гаметофитном растении превращается в защитную оболочку вокруг молодого спорофита. Спорофит начинает расти путем митоза (диплоидного деления клеток) из верхушки архегония.
Она удлиняется и после нескольких клеточных делений начинается дифференцировка. В этот момент спорофит практически является паразитом на гаметофитном растении, хотя на ранних стадиях роста он может производить некоторую собственную пищу посредством фотосинтеза.Зародышевый спорофит состоит из трех структур: ножки, щетинки и капсулы. Нога в нижней части прикрепляет спорофит к гаметофиту путем проникновения и помогает переносить воду и питательные вещества из гаметофита. Щетинка представляет собой длинный прямостоячий опорный стебель. На конце спорофита находится стручковидная капсула , в которой образуются спор . Щетинка встречается только у видов, у которых зрелая капсула имеет ножку.
Переносные клетки развиваются на границе спорофит-гаметофит у большинства мохообразных, но не у всех. Эти специализированные клетки обеспечивают эффективную передачу питательных веществ от гаметофита к спорофиту.
Они могут образовываться на гаметофите, спорофите или на обоих. Соединение гаметофит-спорофит часто бывает извилистым и лабиринтным. Это увеличивает площадь поверхности, позволяя переносить больше клеток, чем простая граница, тем самым увеличивая скорость, с которой питательные вещества могут поступать к спорофиту.Капсула может содержать от четырех до более миллиона спор, в зависимости от вида. Он также может быть на стебле или без стебля в зависимости от вида. У большинства мхов устье капсулы покрыто крышечкой operculum , которая отпадает, когда споры созревают. Перепончатый капюшон, калиптра , который также выбрасывается по мере созревания, дополнительно защищает жаберную крышку.
Во влажных условиях споры не могут распространяться далеко. Крошечная зубчатая структура вокруг устья капсулы контролирует высвобождение спор. Эти структуры, называемые перистом , состоят из одного или двух рядов зубов.
Они предотвращают высвобождение спор во влажных условиях, оставаясь закрытыми. В сухих условиях они раскрываются, высвобождая споры.Каждая спора содержит смесь генов двух родителей. Если спора упадет на влажный участок земли, она может прорасти в ветвящуюся нитевидную нитевидную протонему . Бугорки отпочковываются от протонемы, затем перерастают в лиственные мужские или женские гаметофиты, завершая жизненный цикл.
 Она удлиняется и после нескольких клеточных делений начинается дифференцировка. В этот момент спорофит практически является паразитом на гаметофитном растении, хотя на ранних стадиях роста он может производить некоторую собственную пищу посредством фотосинтеза.
Она удлиняется и после нескольких клеточных делений начинается дифференцировка. В этот момент спорофит практически является паразитом на гаметофитном растении, хотя на ранних стадиях роста он может производить некоторую собственную пищу посредством фотосинтеза.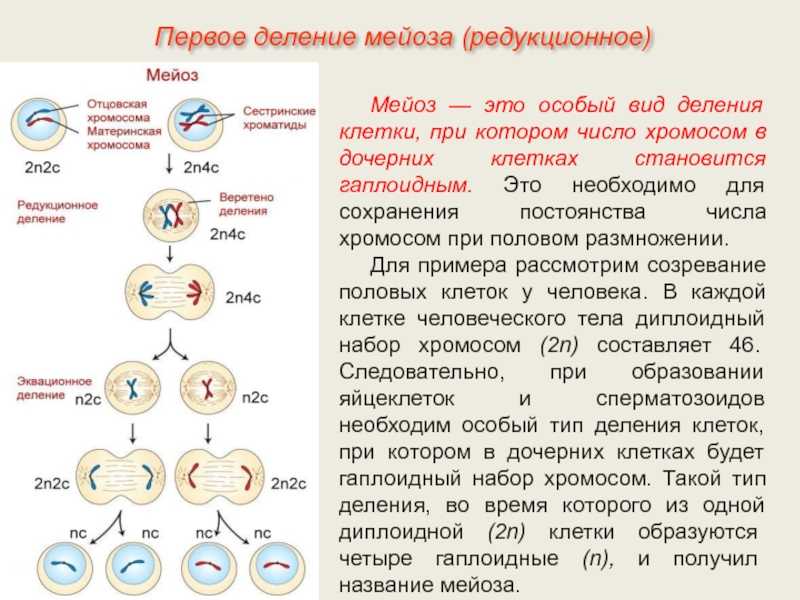 Они могут образовываться на гаметофите, спорофите или на обоих. Соединение гаметофит-спорофит часто бывает извилистым и лабиринтным. Это увеличивает площадь поверхности, позволяя переносить больше клеток, чем простая граница, тем самым увеличивая скорость, с которой питательные вещества могут поступать к спорофиту.
Они могут образовываться на гаметофите, спорофите или на обоих. Соединение гаметофит-спорофит часто бывает извилистым и лабиринтным. Это увеличивает площадь поверхности, позволяя переносить больше клеток, чем простая граница, тем самым увеличивая скорость, с которой питательные вещества могут поступать к спорофиту.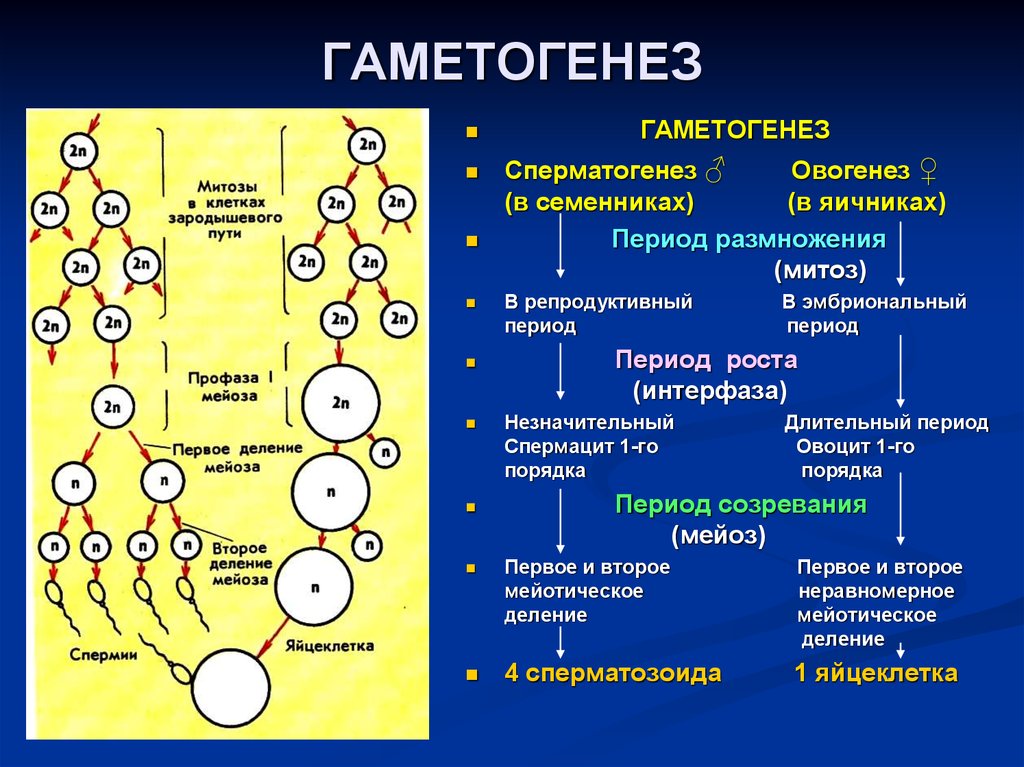 Они предотвращают высвобождение спор во влажных условиях, оставаясь закрытыми. В сухих условиях они раскрываются, высвобождая споры.
Они предотвращают высвобождение спор во влажных условиях, оставаясь закрытыми. В сухих условиях они раскрываются, высвобождая споры.БЕСПОЛОЕ РЕПРОДУКЦИЯ
Помимо полового размножения мхи могут размножаться бесполым (вегетативно). Метод, который они используют для достижения этой цели, зависит от ситуации, в которой они находятся.
Когда стебель большого скопления мха отмирает, скопление без стебля становится отдельными растениями.
Когда кусочки стебля или даже один лист мха отламываются, эти кусочки могут затем регенерировать, образуя новое растение.
ОСНОВНЫЕ ВЫВОДЫ
- В жизненном цикле мха оплодотворение происходит в
Этот пост предназначен только для подписчиков
Подпишитесь сейчас
Уже есть аккаунт? Войти
Для остальной части этой статьи требуется бесплатное членство
Присоединяйся сейчасУже есть учетная запись? Войти
наукациклыжизненные циклыЛоуренс Веру
Ларри является спикером TEDx, лауреатом стипендии декана Гарвардской медицинской школы, Университетом штата Флорида «Известный Ноул», а также выступал в качестве приглашенного докладчика в Гарварде, странах бывшего СССР и USF. Вклад Ларри представлен Fast Company и Gizmodo Japan, а также процитирован в книгах Routledge и No Starch Press. Его истории и мнения публикуются, среди прочего, в Slate, Vox, Toronto Star, Orlando Sentinel и Vancouver Sun.
Он иллюстрирует науку для более справедливого и устойчивого мира.
Вклад Ларри представлен Fast Company и Gizmodo Japan, а также процитирован в книгах Routledge и No Starch Press. Его истории и мнения публикуются, среди прочего, в Slate, Vox, Toronto Star, Orlando Sentinel и Vancouver Sun.
Он иллюстрирует науку для более справедливого и устойчивого мира.
Лекция 25: Биология 102
Протистины, растения Обновлено: |
Темы на 6 марта
Эволюционный тенденции в растениях
Гаметофит и тренды спорофита.
Независимость от жидкости водный тренд.
Мхи и прочие мохообразные
Самый гаметофит заметный
Восковидная кутикула, защищенная развитие гамет
Папоротники и прочие бессемянные сосудистые растения
Спорофит наиболее заметный
Проводящие ткани сосудов вода
Голосеменные: Хвойные и их родственники.
Структура и функция колбочек: репродуктивные структуры.

Эволюционный
тенденции наземных растений.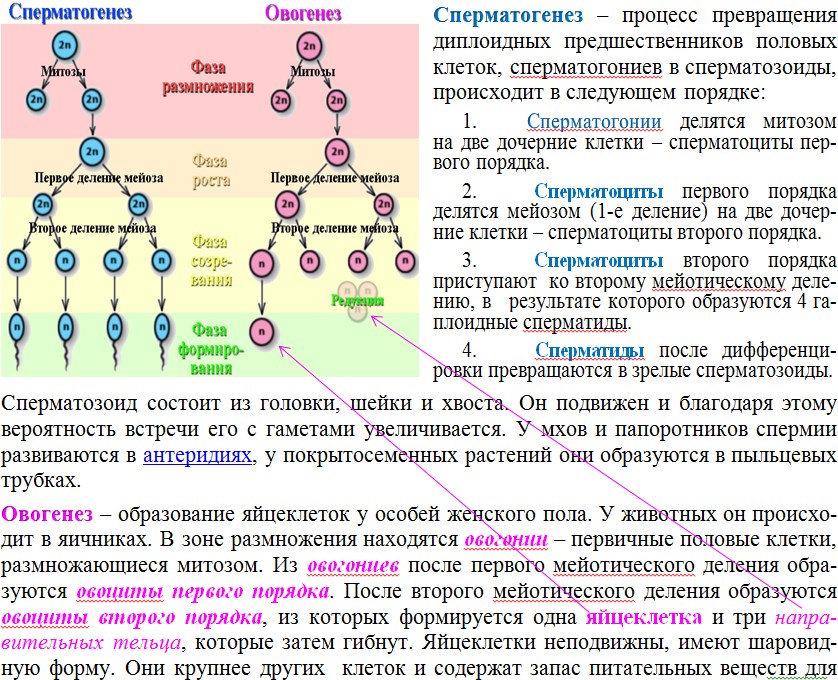
1. Смена из гаметофита в спорофит : В самом простом наземные растения, стадия гаметофит является наиболее заметный. У высших растений имеется гаметофит. только в составе репродуктивных структур.
2. Увеличение независимость от воды: Наземные растения стали все более независимым от жидкой воды различными приспособления для предотвращения потери воды и др.
Зеленый водоросли: возможные предки высших растений.
Зеленый водоросли имеют ряд общих биохимических признаков с высшие растения, включая химическую форму хлорофилл.
Как и многие другие протисты, зеленые водоросли способны к половому воспроизведение.
 Пример: хламидомонада. Обратите внимание, что
доминирующей фазой жизни является гаплоидная фаза. Большинство из
время гаплоидные клетки размножаются бесполым путем.
Пример: хламидомонада. Обратите внимание, что
доминирующей фазой жизни является гаплоидная фаза. Большинство из
время гаплоидные клетки размножаются бесполым путем.Жизнь цикл растений.
Растения чередуются гаплоидные фазы (гаметофит) и диплоидная фаза (спорофит).
гаметофит доминирует в более простых несосудистых растения.
спорофит доминирует у высших растений. (см. рисунок 19.2).
Эволюционный тенденции в растениях.
1. У более простых растений преобладает гаплоидная фаза; в диплоидная фаза доминирует у высших растений.
2. Простые растения очень зависят от жидкой воды; высшие растения менее зависимы от жидкой воды.
3. Высшие растения производят семена (стадия жизни, приспособленная к расселение).
мохообразные: простые наземные растения.
Примеры Мохообразные: мхи, печеночники, роголистники.
Мхи частично независимы от жидкой воды. Они могут растут во влажной земной среде.
А критическая стадия жизни зависит от жидкой воды: сперматозоиды должны доплыть до структур, производящих яйцеклетки.
Жизнь цикл мха. (фигура 19.4).
наиболее заметной стадией мха является гаметофит : Гаплоидная стадия.
Мужской гаметофиты производят сперму, которая должна доплыть до женский гаметофит.
Женский гаметофиты образуют яйца, которые сохраняются в специальные структуры на женском гаметофите.
оплодотворенная яйцеклетка — зигота, зависящая от самки гаметофит.

Жизнь цикл мха (продолж.)
зигота развивается как многоклеточный спорофит, но остается зависимым от женского гаметофита.
Мейоз происходит в специализированной структуре на спорофит (спорангий), образующий споры (не гаметы!).
споры делятся и производят мужские или женские гаметофиты.
спорофит мхов: зависимая стадия.
спорофитная фаза мхов остается прикрепленной к женский гаметофит.
В В отличие от высших растений спорофит мхов имеет краткое существование (как часть сексуального репродукция).
самая заметная фаза мхов — гаметофит фаза.
Адаптация к жизни на суше.
Мхи способны расти на суше, а не в воде (водоросли растут только в воде).
Адаптации на землю включают:
1. Восковидная кутикула, уменьшающая испарение.
2. Клеточная оболочка вокруг клеток, производящих сперму и яйцо (защищая их).
3. Стадия крупного гаметофита (гаплоидная), поддерживающая стадии спорофита.
Что удерживает мхи привязанными к влажным местам обитания?
Мхи производят сперму, зависящую от жидкой воды.
Из-за этой особенностью жизненного цикла мхи связаны с среды обитания, в которых есть хотя бы немного жидкой воды часть времени. (Капли дождя или водяная пленка достаточно.)
 Без наличия жидкой воды сперматозоиды
не может добраться до яйца.
Без наличия жидкой воды сперматозоиды
не может добраться до яйца.Без косточек сосудистые растения.
Бессемянный сосудистые растения (папоротники и их родственники) имеют самостоятельных долгоживущий спорофит на стадии . гаметофит стадия меньше, но независима.
спорофит обладает комплексом сосудистой ткани: сосудистая ткань допускает больший размер, потому что вода может переносится из почвы в растение.
Сосудистый салфетка.
Высшее растения имеют два типа сосудистой ткани: ксилему и флоэма.
 (см. стр. 380 и далее)
(см. стр. 380 и далее)Ксилем : Клетки ксилемы представляют собой трубки, по которым вода поступает из почвы к другим частям растения. Большинство клеток ксилемы на самом деле мертвы, но их структура продолжает функция.
Флоэма : Клетки флоэмы транспортируют сахара и другие пищевые вещества. по всему заводу. Клетки флоэмы остаются живыми.
Без косточек сосудистые растения: разнообразные виды растений.
Папоротники: Наиболее известны бессемянные сосудистые растения.
Ликофиты: Живые виды малы и незаметны, но их предки были доминирующими растениями Каменноугольный период.
Хвощи: Только несколько выживших видов, но, как и Lycophytes, когда-то они были доминирующими наземными растениями.
Жизнь цикл папоротника (см. рис. 19.8)
жизненный цикл папоротника включает свободноживущую стадия гаметофит. Он маленький и незаметный, а отсутствует сосудистая ткань.
Зигота начинает жизнь прикрепленным к гаметофиту, но вскоре развивается в крупный и самостоятельный спорофит. спорофит имеет сосудистую ткань и может достигать очень большой размер.
гаметофит папоротников.
гаметофит гаплоидный.
Оба сперма и яйцеклетки образуются на одном и том же растении (к митоз !)
гаметофит начинает жизнь с прорастания гаплоидная спора. Споры являются эффективным средством рассеивания фаза папоротников.
спорофит папоротников.
крупная и привычная фаза папоротников — спорофит. Это диплоидная фаза.
спорофит имеет сосудистую ткань и может проводить воду из почвы в другие части растения.
спорофит образует гаплоидные споры путем мейоза. ( Споры представляют собой пыльно-коричневый материал на нижней стороне «листа».)
Митоз
и мейоз в жизненном цикле папоротника.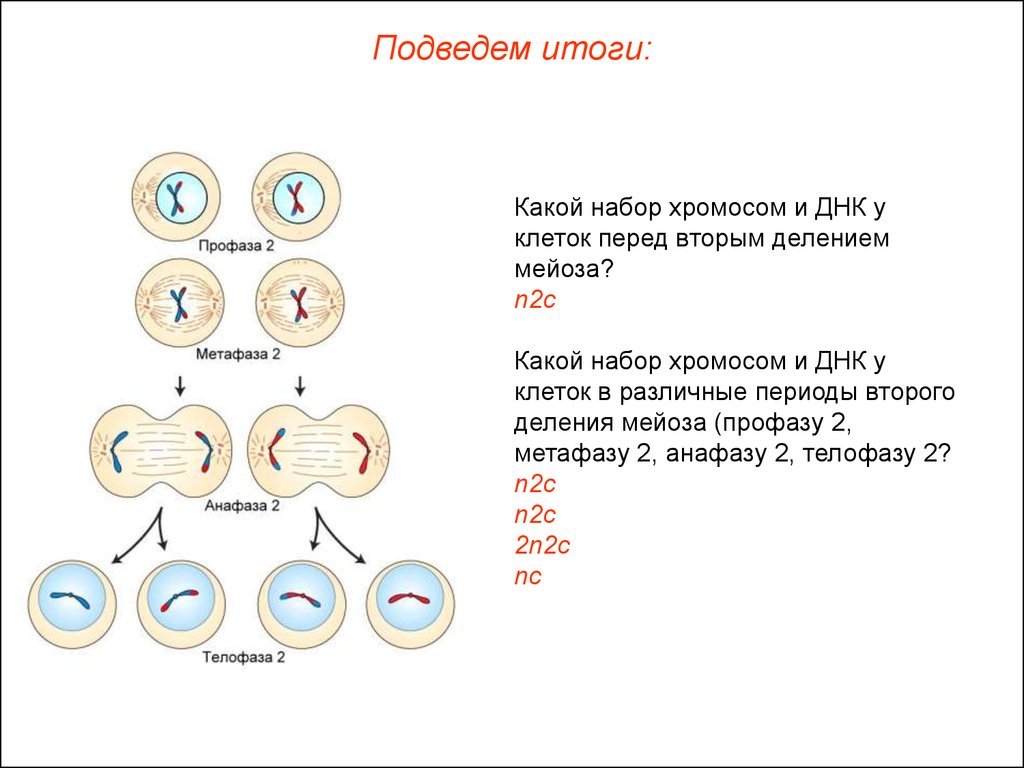
А гаплоидная спора прорастает и начинает делиться митоз с образованием небольшого многоклеточного гаметофита этап.
На стадии гаметофит образуются гаметы (путем митоза), которые сливаются, образуя зиготу.
зигота делится митозом, образуя большую стадия многоклеточного спорофита.
Митоз и мейоз у папоротников (продолжение).
спорофит образует гаплоидные споры путем мейоза. споры являются эффективной фазой распространения в жизни цикл папоротника.
Обратите внимание, что мейоз производит споры, а не гаметы. споры прорастает и превращается в самостоятельный гаметофит этап.
 (Мейоз не производит гамет в
эти растения!)
(Мейоз не производит гамет в
эти растения!)Голосеменные: Хвойные и их родственники.
Хвойные деревья древесные деревья и кустарники с игольчатыми листьями.
Хвойные деревья имеют шишки (отсюда и их название).
Конусы являются репродуктивными структурами хвойные: Шишки представляют собой диплоидную ткань, вырабатываемую доминирующая стадия спорофита.
развивается гаплоидный гаметофит и образуется гаметы внутри конуса .
Семена: важное эволюционное достижение хвойных.
Конусы производить семена. Семена развиваются на «обнаженные» части спорофита, следовательно, название «голосеменные» или «голые семена».
 Семена являются эффективными пропагулами для рассеивания
Население.
Семена являются эффективными пропагулами для рассеивания
Население.Семена являются очень устойчивыми стадиями и могут сохраняться в течение многих лет, поддержание популяции.
Пыльца: Важный эволюционный шаг. (рисунок 19.9)
Голосеменные (а также цветковые растения) производят пыльцу в качестве пакет для рассеивания спермы.
Пыльца зерна мужские гаметофиты . Они перевозят сперматозоиды (внутри пыльцевого зерна) ветром или насекомые: жидкая вода не требуется.
Конусы: мужские и женские репродуктивные структуры.
Женский колбочки диплоидной ткани вырабатываются доминирующая стадия спорофита.
Мейозы происходит внутри охватывающего конуса для получения мегаспоры.
Мегаспоры развиваться, еще будучи прикрепленным к конусу, в женские гаметофиты.
Внутри женский гаметофит , внутри образуются яйца особая структура — яйцеклетка.

Конусы: мужчина и женщина
Мужской колбочки представляют собой диплоидную ткань, прикрепленную к доминантному спорофит.
Внутри мужские шишки, происходит мейоз с образованием микроспоры.
Микроспоры развиваются в мужских гаметофитов: пыльцевые зерна .
пыльцевые зерна содержат мужские гаметы: сперматозоиды.
Leave A Comment