
Типы клеток кишечнополостных (стрекательные, промежуточные, кожно-мускульные, нервные, пищеварительные, железистые) | Биология. Реферат, доклад, сообщение, краткое содержание, лекция, шпаргалка, конспект, ГДЗ, тест
Раздел: Типы Губки и Кишечнополостные
| Рис. 63. 2. Типы клеток |
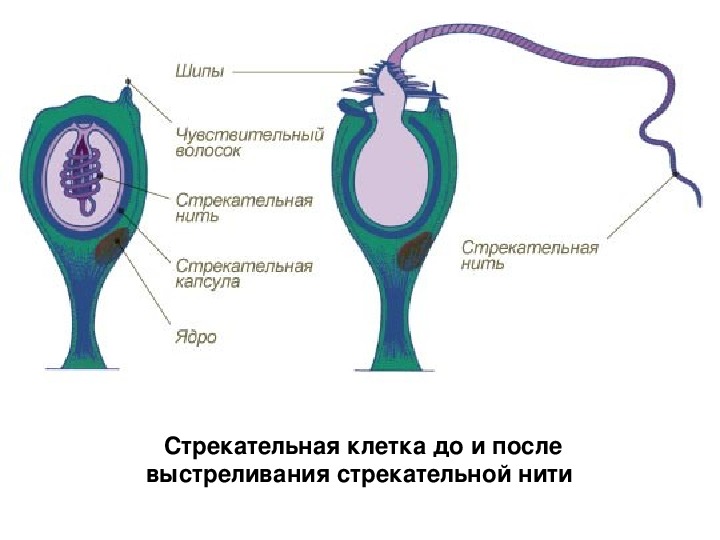
В наружном слое кишечнополостных есть стрекательные клетки (рис. 63. 2). Это их «визитная карточка», они больше ни у кого не встречаются. Стрекательные клетки служат для защиты, поражения добычи и ее удержания; наибольшее их количество расположено на щупальцах. Внутри стрекательной клетки находится капсула со спирально закрученной стрекательной нитью. Когда добыча, проплывая мимо гидры, задевает торчащий наружу чувствительный волосок, из стрекательной клетки выбрасывается стрекательная нить и впивается в тело жертвы или хищника.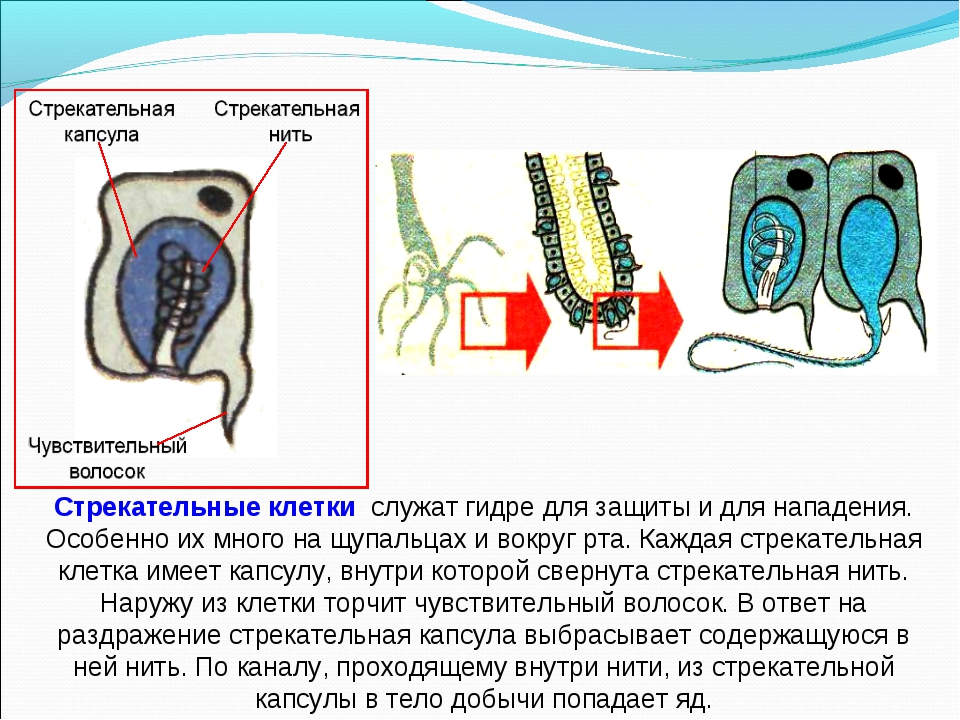 Вместе с ней вводится ядовитое вещество, парализующее добычу. После того как стрекательная нитка выходит из клетки, назад она уже не втягивается.
Вместе с ней вводится ядовитое вещество, парализующее добычу. После того как стрекательная нитка выходит из клетки, назад она уже не втягивается.
Новые стрекательные клетки, так же как и клетки других типов, образуются за счет промежуточных (рис. 63. 2). Это неспециализированные клетки, способные делиться и давать начало клеткам других типов. По своим функциям промежуточные клетки можно сравнить с клетками образовательной ткани растений. Благодаря этим клеткам происходят процессы регенерации.
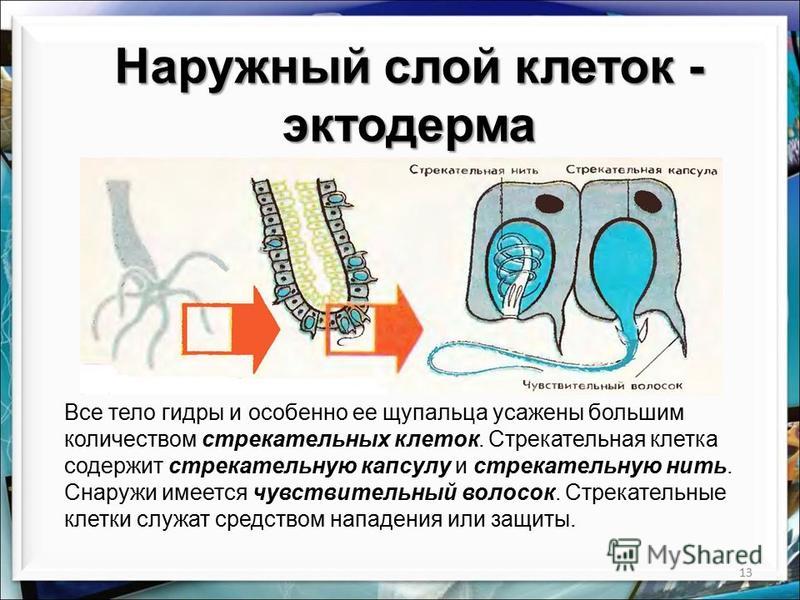
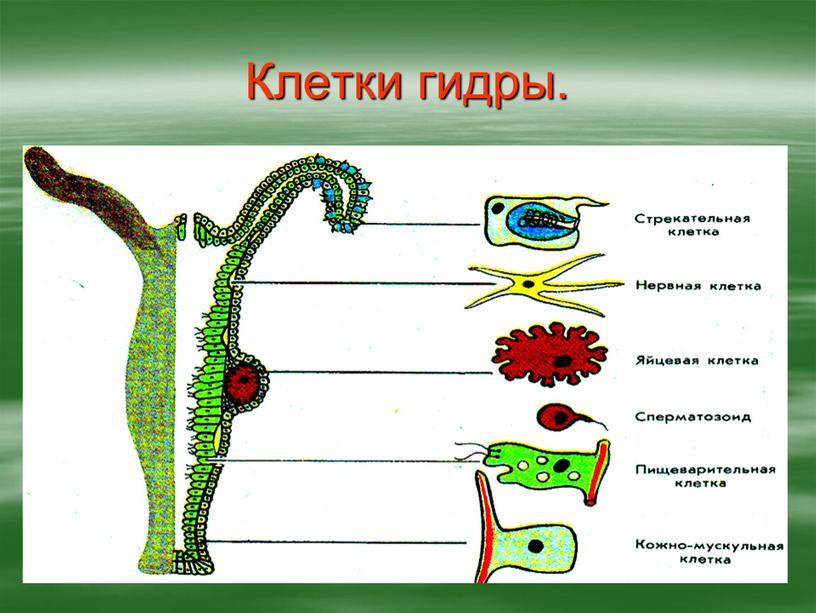
Покровы гидры преимущественно образованы кожно-мускульными клетками (рис. 63. 2). Благодаря их сокращениям тело гидры сжимается или наклоняется в определенную сторону. Между кожно-мускульных клеток разбросаны
Среди клеток внутреннего слоя преобладают пищеварительные клетки со жгутиками.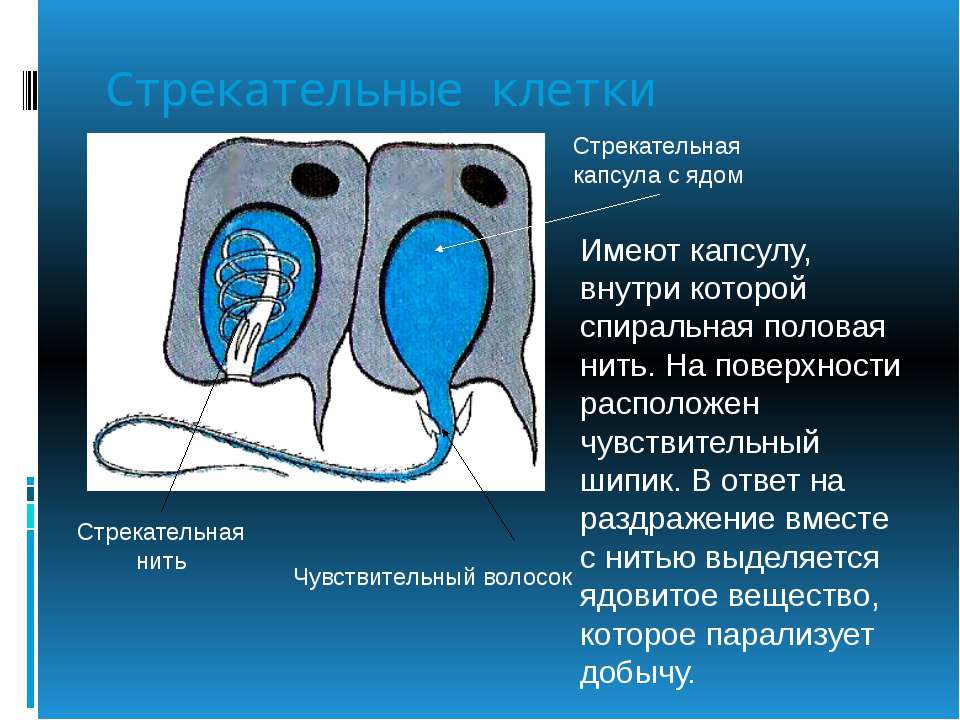 Они обеспечивают внутриклеточное переваривание пищи. Железистые клетки вырабатывают и выделяют в кишечную полость пищеварительные соки, под влиянием которых пища распадается на кусочки.
Они обеспечивают внутриклеточное переваривание пищи. Железистые клетки вырабатывают и выделяют в кишечную полость пищеварительные соки, под влиянием которых пища распадается на кусочки.
Стенки тела кишечнополостных состоят из двух слоев клеток — наружного и внутреннего, между которыми расположен слой межклеточного вещества.
У кишечнополостных формируются ткани и органы.
Реферат на тему стрекательные клетки кишечнополостных
Схема промежуточной коетки у кешечнополосчаиых
Функции кожно-мускульных клеток у губки
Нервные, стрекательные, промежуточные клетки находятся в слое
У кого стрекательные клетки
Какие типы клеток входят в состав тела кишечнополостных?
Каковы строение и функции стрекательных клеток?
(PDF) Стрекательные клетки Polypodium hydriforme (Cnidaria, Polypodiozoa)
ЦИТОЛОГИЯ том 61 № 3 2019
СТРЕКАТЕЛЬНЫЕ КЛЕТКИ POLYPODIUM HYDRIFORME 201
ные двуядерными клетками – инвазивной стадией
развития полиподия, заражающей рыбу (рис. 1г).
1г).
Стрекательные клетки служат для прикрепления га-
метофоров к хозяину (Смольянов, Райкова, 1961).
У P. h yd rif or me обнаружены три размерных катего-
рии стрекательных капсул (книдоцист или немато-
цист) с диаметром 12, 7 и 5 мкм (в среднем). Капсулы
размером 7 мкм сосредоточены в области рта и на
сенсорных щупальцах (рис. 1а, 1б), а капсулы разме-
ром 12 и 5 мкм в основном усеивают концы опорных
щупалец (рис. 1а, 1в). Поскольку свободноживущие
особи активно расходуют стрекательные клетки, об-
разование новых капсул (книдогенез) не прекраща-
ется, и развивающиеся книдобласты мигрируют к
местам их использования (Lipin, 1911). Развитие всех
стрекательных капсул сходно (Lipin, 1911; Райкова,
1978) и типично для книдарий. Будущая капсула раз-
вивается из последней большой цистерны аппарата
Гольджи. Стрекательная нить закладывается вне-
капсулярно и состоит из чередующихся светлых и
темных фрагментов одинакового размера.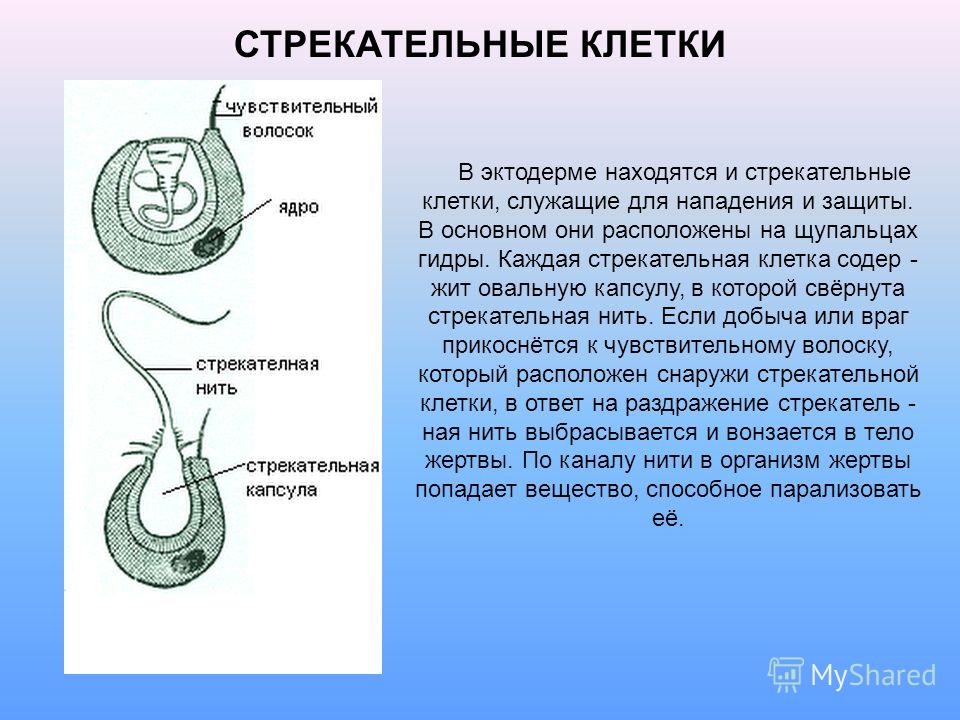 Затем
Затем
нить втягивается внутрь капсулы, в стенке которой к
этому времени различаются внутренний и внешний
слои, и капсула замыкается крышечкой, к которой
крепится стрекательная нить (Raikova, 1990).
ТИПЫ СТРЕКАТЕЛЬНЫХ
КАПСУЛ P. HY D R I F OR ME
Зрелые стрекательные капсулы P. hy dr if o r m e по
классификации Шульце (Schultze, 1917) относятся к
большим и малым глютинантам, а по классифика-
ции Вейла (Weill,1934) – к типу atrichous и ho-
lotrichous isorhiza.
Atrichous isorhiza (атрихи) – большие капсулы
диаметром около 12 мкм – устилают концы опор-
ных щупалец (рис. 1а, 1в, 2а). Свернутая стрекатель-
ная нить в них занимает все пространство капсулы
(рис. 1в). Нить уложена по принципу двойной
складки и на срезах демонстрирует сложные профили
(Raikova, 1990), но не “профиль пропеллера”, как в
капсулах большинства книдарий, где прослеживается
трехскладчатость (Mariscal, 1974). Все книдоцили та-
Все книдоцили та-
ких капсул прямые и одинаковой длины (рис. 1в, 2а).
Эти капсулы используются при передвижении поли-
подия по субстрату и при заякоривании на нем. При
этом выстреливающая стрекательная нить приклеи-
вается к песчинкам, а капсула может легко отделять-
ся от щупальца полиподия. Выстрелившая стрека-
тельная нить длинная и ровная, на ней часто заметен
обвивающий ее слизистый тяж (рис. 3в, 3г) (Ибраги-
мов, 1999, 2002; Ibragimov, Raikova, 2004).
Holotrichous isorhiza (холотрихи) – средние и мелкие
капсулы, диаметром соответственно 7 и 5 мкм, лока-
лизуются вокруг ротового отверстия и на сенсорных
щупальцах, а также единично встречаются на по-
верхности опорных щупалец. Капсулы диаметром
7 мкм окрашиваются гистологическими красителя-
ми. В капсулах вокруг рта (7 мкм) стрекательная
нить отходит от крышечки капсулы под прямым уг-
лом, делает 2–3 витка, а затем укладывается на дне
капсулы (рис. 1б). На электронограммах нить в кап-
1б). На электронограммах нить в кап-
суле демонcтрирует S-образные профили, т.е. двух-
складчатость (Ibragimov, Raikova, 2004). Капсулы же
размером 5 мкм при гистологической окраске обыч-
но остаются бесцветными, а нить в них, как и у ат-
рих, заполняет все пространство капсулы (рис. 1в).
Выстрелившие нити капсул холотрих отличаются
друг от друга длиной, они покрыты мелкими шипа-
ми, расположенными в два ряда (рис. 3д), что явля-
ется их основной характеристикой, тогда как у дру-
гих книдарий стрекательные нити имеют три ряда
шипов. Некоторые нити суживаются к концу, поэто-
му их предлагается именовать holotrichous anisorhiza.
Но внешне капсулы, ещe не выстрелившие, неразли-
чимы (Ибрагимов, 1999). Главное же отличие холо-
трих от атрих состоит в том, что холотрихи у полипо-
дия обладают уникальными пенетрантными свой-
ствами: их нить пронзает покровы тубифекса и
парамеции (Raikova, 1990; Ibragimov, Raikova, 2004).
Стрекательные клетки у P. hy dr if or me (и атрихи, и
холотрихи) относятся к самому примитивному типу
нематоцист (Werner, 1965; Боженова, 1988; Östman,
1999, 2000; David et al., 2008; Fautin, 2009). Эволюция
стрекательных клеток в настоящее время интенсив-
но разрабатывается, но книдоциты нашего объекта
изучаются очень слабо, хотя благодаря своим не-
обычным признакам интересны как определенная
ступень эволюции стрекательных клеток, как воз-
можное связующее звено между книдоцитами сво-
бодноживущих книдарий и полярными капсулами
миксоспоридий.
ТОНКОЕ СТРОЕНИЕ СТРЕКАТЕЛЬНОЙ
КЛЕТКИ И КНИДОЦИЛЬНОГО АППАРАТА
Каждая зрелая стрекательная клетка P. hy dr if or me
имеет книдоцильный аппарат (рис. 1б, 1в; 2а–2д; 3а),
состоящий из книдоциля – чувствительного механо-
сенсорного неподвижного жгутика, окруженного коль-
цом выростов апикальной поверхности стрекательной
клетки, называемых стереоцилиями (рис.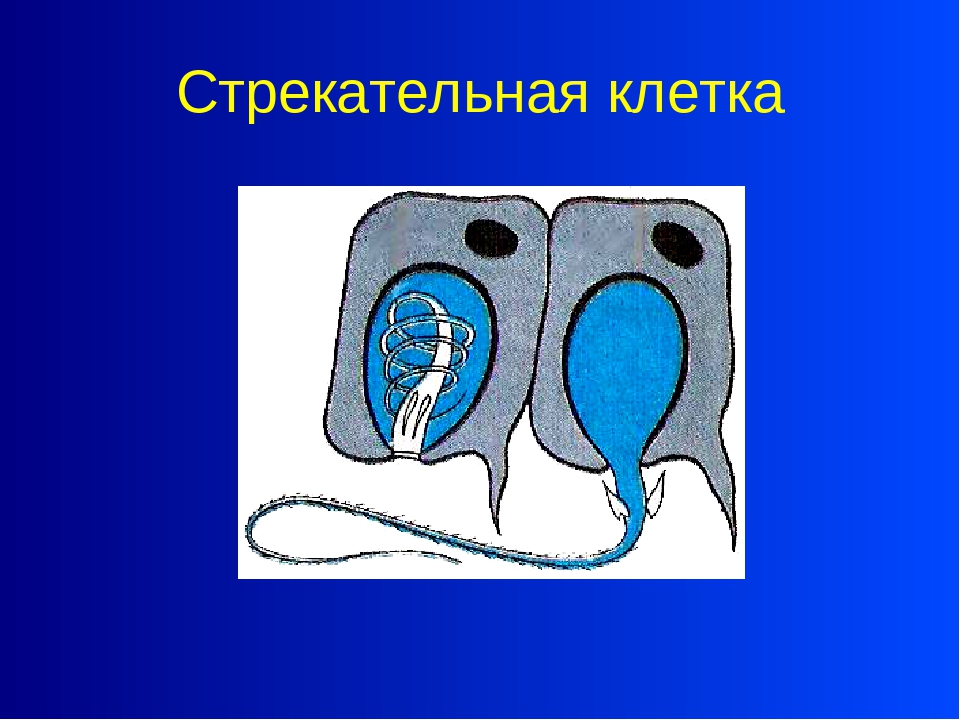 2д; 3а). Все
2д; 3а). Все
стрекательные капсулы у полиподия имеют округ-
лую форму, что не типично ни для глютинант, опи-
санных в литературе (обычно продолговатых капсул)
(Fautin, 2009), ни для других категорий книдоцитов
большинства книдарий. Вторая их особенность –
положение книдоциля непосредственно над кры-
шечкой капсулы (рис. 1б, 1в; 2а–2д; 3а). Эта особен-
Тест по биологии по теме «Беспозвоночные животные» с ответами
Тест по биологии по теме «Беспозвоночные животные» с ответами — Gee Test наверх Страница 1 из 14Страница 2 из 14Страница 3 из 14Страница 4 из 14Страница 5 из 14Страница 6 из 14Страница 7 из 14Страница 8 из 14Страница 9 из 14Страница 10 из 14Страница 11 из 14Страница 12 из 14Страница 13 из 14Страница 14 из 14- 1. выделительная
- 2. защитная
- 3. сократительная
- 4. пищеварительная
- 1. 1, 3, 5
- 2. 1, 2, 7
- 3.
 2, 6, 7
2, 6, 7 - 4. 2, 4, 5
 2, 6, 7
2, 6, 7- 1. 1, 5
- 2. 2, 4
- 3. 3, 5
- 4. 3, 4
- 1. нервное сплетение
- 2. спинной мозг
- 3. нервный столб
- 4. нервный узел
- 1. простейшие, кишечнополостные
- 2. моллюски, членистоногие
- 3. плоские черви, круглые черви
- 4. простейшие, моллюски
- 1. с железистыми
- 2. с пищеварительными
- 3. с кожно-мускульными
- 4. с половыми
- 1. В, С, D
- 2. коралловый полип
- 3. актиния
- 4. гидра
- 1. поражение добычи, защитная
- 2. двигательная
- 3. выделительная
- 4. пищеварительная
- 1. 1, 3, 4
- 2. 2, 6
- 3. 3, 5
- 4. 3, 4, 7
- 1. из кремнезема
- 2. из извести
- 3. из кальция
- 4. из калия
- 1. зигота
- 2. личинка
- 3. спорофилл
- 4. молодой организм, отличающийся от взрослых особей только размером
- 1. стрекательные
- 2. кожно-мускульные
- 3. промежуточные
- 4. покровные
 стрекательные
стрекательные- 1. оболочка, цитоплазма, ядро, стрекательная капсула, стрекательная нить, чувствительный волосок
- 2. оболочка, цитоплазма, пищеварительная вакуоль, реснички
- 3. оболочка, ядро, цитоплазма, сократительная вакуоль, жгутик
- 4. оболочка, цитоплазма, стрекательная капсула, стрекательная нить, мускульное волоконце
- 1. в митохондриях клеток
- 2. в кишечной полости
- 3. в энтодермальных клетках
- 4. в эктодермальных клетках
- 1. регенерация
- 2. пищеварение
- 3. защитная
- 4. размножение
- 1. амёбу
- 2. клетку из колонии вольвокса
- 3. эвглену зеленую
- 4. инфузорию туфельку
- 1. медуза аурелия, актинии
- 2. коралловые полипы, актинии
- 3. корнерот, коралловые полипы
- 4. гидра, медуза крестовичок
- 1. виноградная улитка, моллюски
- 2. дождевой червь, бычий цепень
- 3. гидра, медуза
- 4. белая планария, вольвокс
 дождевой червь, бычий цепень
дождевой червь, бычий цепень- 1. гребневиков, иглокожих
- 2. кишечнополостных, иглокожих
- 3. гребневиков, кишечнополостных
- 4. губок, споровиков
- 1. 1, 2
- 2. 2, 3
- 3. 1, 4
- 4. 5, 6
- 1. аурелия
- 2. большеротая медуза
- 3. крестовичок
- 4. корнерот
- 1. промежуточным, кожно-мышечным
- 2. стрекательным, нервным
- 3. половым, промежуточным
- 4. нервным, кожно-мышечным
- 1. гидрообразные, коралловые полипы, медузы
- 2. коралловые полипы, гидрообразные, медузы
- 3. медузы, коралловые полипы, гидрообразные
- 4. медузы, гидрообразные, коралловые полипы
- 1. 1, 4, 5
- 2. 2, 4, 6
- 3. 1, 3, 6
- 4. 2, 3, 5
- 1. половые клетки
- 2. пищеварительные клетки эктодермы
- 3. стрекательные клетки эктодермы
- 4.
- 1. 3
- 2. 2
- 3. пищеварительные клетки не имеют жгутиков
- 4. 1
- 1. из зиготы появляется свободноплавающая личинка
- 2. из зиготы появляется свободноплавающая личинка, которая превращается в сидячий полип
- 3. почкованием
- 4. происходит деление тела на две равные части
- 1. личинками
- 2. почкованием
- 3. делением тела на две равные части
- 4. зооспорами
- 1. прикрепленным образом жизни, расположением ротового отверстия на переднем конце тела, двухслойной стенкой тела с очень большим содержанием промежуточного вещества
- 2. свободноплавающим образом жизни, расположением ротового отверстия в центре нижней стороны тела, двухслойной стенкой тела с очень большим содержанием промежуточного вещества
- 3. свободноплавающим образом жизни, расположением ротового отверстия на переднем конце тела, однослойной стенкой тела
- 4. ничем
- 1. служат пищей для морских хищников
- 2. являются сырьем для получения шлифовального материала
- 3. являются сырьем для получения извести
- 4. В, С
 служат пищей для морских хищников
служат пищей для морских хищников- 1. наличие слепо замкнутой пищеварительной системы
- 2. способность к регенерации
- 3. двуслойное строение тела
- 4. стрекательные клетки
- 1. пищеварение
- 2. размножение
- 3. выведение продуктов обмена веществ
- 4. дыхание
- 1. всей поверхностью тела
- 2. с помощью ротового отверстия
- 3. гидра является свободноплавающим животным
- 4. с помощью подошвы
- 1. дыхальца
- 2. трахеи
- 3. жабры
- 4. специальные органы дыхания отсутствуют, дышит поверхностью тела
- 1. кожные покровы
- 2. порошица
- 3. разветвленные канальцы
- 4. выделительные железы
- 1. 1, 3, 4
- 2. 5
- 3. 1, 4
- 4. 1, 2, 3, 4
- 1. с помощью ротового отверстия
- 2.
- 3. с помощью глотки
- 4. всей поверхностью тела
- 1. способом питания
- 2. прикрепленным образом жизни
- 3. активным передвижением
- 4. паразитическим образом жизни
- 1. кольчатые черви
- 2. плоские черви
- 3. простейшие
- 4. кишечнополостные
- 1. покровная ткань, соединительная ткань, мышечная ткань
- 2. покровная ткань, слизистая оболочка, продольные мышцы
- 3. кутикула, эпителиальная ткань, продольные мышцы
- 4. хитин, покровная ткань, продольные мышцы
- 1. с особенностями строения нервной системы
- 2. с наличием только продольных мышц
- 3. с наличием кутикулы
- 4. с паразитическим способом питания
- 1. личинка с шести крючьями
- 2. членик червя с хорошо развитой половой системой
- 3. пузырек, заполненный жидкостью, внутри которого находится головка паразита с присосками
- 4. яйцо
- 1. I – 2, 6, 7, 8; II – б
- 2. I – 1, 2, 5, 6, 7; II – в
- 3. I – 2, 5, 6; II – а
- 4. I – 1, 3, 4, 5, 8; II – а
 I – 2, 6, 7, 8; II – б
I – 2, 6, 7, 8; II – б- 1. нервное сплетение
- 2. нервные стволы
- 3. диффузная
- 4. диффузно-узловая
- 1. наличие легких
- 2. наличие трахеид
- 3. наличие жабер и жаберных крышек
- 4. дыхательная система отсутствует
- 1. в водоёмах
- 2. в почве
- 3. во внутренних органах крупного рогатого скота
- 4. в тонком кишечнике человека
- 1. в шейной области
- 2. в задних члениках
- 3. во всех члениках одновременно
- 4. в головке с присосками
- 1. за счет увеличения размеров члеников тела
- 2. за счет роста головки с присосками
- 3. за счет появления новых члеников в шейной области тела
- 4. за счет деления каждого членика надвое
- 1. один яичник, одна матка, один семенник в каждом членике
- 2. пара яичников, одна матка, множество семенников на целый организм
- 3. множество яичников, одна матка, множество семенников в каждом членике
- 4. пара яичников, одна матка, множество семенников в каждом членике
 множество яичников, одна матка, множество семенников в каждом членике
множество яичников, одна матка, множество семенников в каждом членике- 1. выведение продуктов метаболизма
- 2. проведение пищи в пищевод
- 3. переваривание пищи
- 4. захват и проведение пищи в кишечник
Стрекательные клетки — Справочник химика 21
До недавнего времени ацетилхолин считался специфическим метаболитом мира животных. Однако на самом деле и холин, и его ацетильное производное 6.3 широко распространены в самых разнообразных растениях. В частности, ацетилхолин присутствует в стрекательных клетках крапивы. Именно он, совместно с гистамином и серотонином, вызывает болевую реакцию и ожог на коже при соприкосновении с растением. [c.429]
[c.429] Основа клеточной организации медузы — два слоя клеток (рис. 2.1Б), наружный слой — эктодерма и внутренний — энтодерма. Поэтому медузу (а также гидру) называют диплобласти-ческим организмом. Как показано на рис. 2.1В, некоторые клетки эктодермы специализируются для рецепции раздражителей, а некоторые для сокращения или для защиты (нематоцисты, или стрекательные клетки). Точно так же клетки энтодермы могут дифференцироваться на пищеварительные или железистые клетки. Эта дифференцировка, однако, неполная, поскольку обычно у таких клеток сохраняется несколько функций (например, отдельная клетка может быть одновременно и эпителиальной и сократительной). Кроме того, эти клетки не объединяются в органы. Можно поэтому сказать, что кишечнополостные в своей организации достигли тканевого уровня, но еще не органного. [c.40]
Подобно тому как отростки нейронов растут и соединяются со специфическими участками-мишенями, так и целые клетки нередко направленно перемещаются в процессе эмбрионального развития. Полагают, что в основе миграции клеток лежит хемотаксис (разд. Б. 7). Значительная часть работ по изучению миграции клеток выполнена на гидре (рис. 1-10), примитивном животном, содержащем клетки только 10 типов. Один из этих типов представлен эмбриональным резервом клеток мезодермы. Это клетки стволовой линии, образующие, помимо прочих клеток, нематоциты (стрекательные клетки), которые, сформировавшись, продвигаются вверх по телу гидры и располагаются в конце концов в щупальцах [159, 160]. [c.358]
Полагают, что в основе миграции клеток лежит хемотаксис (разд. Б. 7). Значительная часть работ по изучению миграции клеток выполнена на гидре (рис. 1-10), примитивном животном, содержащем клетки только 10 типов. Один из этих типов представлен эмбриональным резервом клеток мезодермы. Это клетки стволовой линии, образующие, помимо прочих клеток, нематоциты (стрекательные клетки), которые, сформировавшись, продвигаются вверх по телу гидры и располагаются в конце концов в щупальцах [159, 160]. [c.358]
Кишечнополостные (Сое1еп1ега1а). Кишечнополостные (медузы, кораллы, морские анемоны) — двухслойные многоклеточные, обладающие истинно тканевыми образованиями в виде экто- и эндодермы. Между этими слоями находится неструктурированная, желеобразная мезоглея. Среди клеток, входящих в тканевые слои, описаны такие формы как стрекательные клетки, реснитчатые клетки, гликоциты, интерстициальные клетки. В мезоглее пред- [c.395]
Токсины морской анемоны. Если ТТХ и STX действуют на максимальную натриевую проводимость gNa и, таким образом, блокируют открытие ворот, то стрекательные клетки морской анемоны содержат яд, который тормозит активацию натриевого канала и оставляет ворота открытыми. Из Anemonia sul ata было выделено три различных токсина, анемонотоксины I, II и III (АТХ I, АТХ II и АТХ III) [19]. Для этих токсинов были установлены структуры построены они из небольших полипептидных цепей, содержащих по 46, 47 и 27 аминокислотных остатков соответственно. АТХ I и АТХ II в высшей степени гомологичны, в то время как АТХ III резко от них отличается. Пространственная структура АТХ I и АТХ II стабилизируется тремя дисульфидными связями, нарушение которых приводит к потере токсичности. [c.148]
Если ТТХ и STX действуют на максимальную натриевую проводимость gNa и, таким образом, блокируют открытие ворот, то стрекательные клетки морской анемоны содержат яд, который тормозит активацию натриевого канала и оставляет ворота открытыми. Из Anemonia sul ata было выделено три различных токсина, анемонотоксины I, II и III (АТХ I, АТХ II и АТХ III) [19]. Для этих токсинов были установлены структуры построены они из небольших полипептидных цепей, содержащих по 46, 47 и 27 аминокислотных остатков соответственно. АТХ I и АТХ II в высшей степени гомологичны, в то время как АТХ III резко от них отличается. Пространственная структура АТХ I и АТХ II стабилизируется тремя дисульфидными связями, нарушение которых приводит к потере токсичности. [c.148]
Книга написана людьми, судя по всему, слабо разбирающимися в биологии у некоторых (одноклеточных) организмов весь организм заключен в эту одну клетку (с. 3) и называется этот код универсальный код жизни (с.
 4) генная инженерия — это наука из пробирки (с. 8) растения имеют стрекательные клетки (с. 3) нидерландские исследователи… обнаружили, что живые и целые гены устойчивости могут перепрыгивать из ГМ-продукта в кишечник человека и выживать там до нескольких минут (с. 20). Что такое живой ген и как он может прыгать Пытаясь объяснить простые биологические процессы, авторы строят фантастические конструкции. Например, они хотят доказать, что маки с красными листьями нельзя получить традиционными методами, а можно лишь генно-ин-женерными. (Это сомнительно. В листьях мака есть красные пигменты, и их количество можно увеличить путем селекции. В лепестки листья не превратятся, так как выполняют иные, несвойственные лепесткам, функции. Даже генная инженерия не избавит листья от необходимости осуществлять фотосинтез и содержать хлорофилл.) Существует барьер, предотвращающий покраснение листьев. Этот барьер может быть обусловлен двумя причинами красный ген во [c.150]
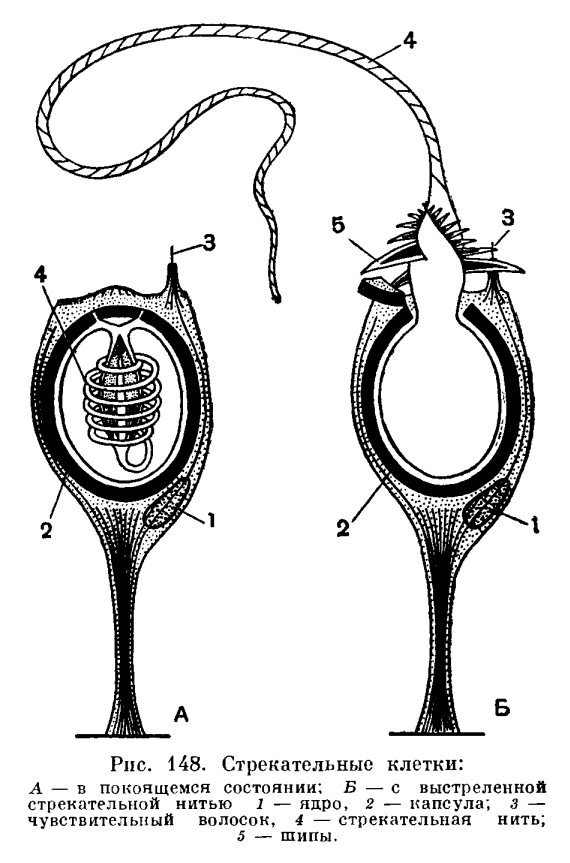
4) генная инженерия — это наука из пробирки (с. 8) растения имеют стрекательные клетки (с. 3) нидерландские исследователи… обнаружили, что живые и целые гены устойчивости могут перепрыгивать из ГМ-продукта в кишечник человека и выживать там до нескольких минут (с. 20). Что такое живой ген и как он может прыгать Пытаясь объяснить простые биологические процессы, авторы строят фантастические конструкции. Например, они хотят доказать, что маки с красными листьями нельзя получить традиционными методами, а можно лишь генно-ин-женерными. (Это сомнительно. В листьях мака есть красные пигменты, и их количество можно увеличить путем селекции. В лепестки листья не превратятся, так как выполняют иные, несвойственные лепесткам, функции. Даже генная инженерия не избавит листья от необходимости осуществлять фотосинтез и содержать хлорофилл.) Существует барьер, предотвращающий покраснение листьев. Этот барьер может быть обусловлен двумя причинами красный ген во [c.150] I. Расположение клеток. 1 — эпидермис, 2 — гастродерма, 3 — стрекательная клетка, 4 — интерстициальная клетка, S — мышечная клетка, 6 — нервная клетка, 7 — пищеварительная клетка, 8 — железистая клетка, 9 — клетка со жгутиками, 10 — сенсорная клетка. II. Мышечные клетки гидры. 1 — ядро, 2 — сократимый отросток. III. Нервные клетки морской лилии. [c.141]
1 — эпидермис, 2 — гастродерма, 3 — стрекательная клетка, 4 — интерстициальная клетка, S — мышечная клетка, 6 — нервная клетка, 7 — пищеварительная клетка, 8 — железистая клетка, 9 — клетка со жгутиками, 10 — сенсорная клетка. II. Мышечные клетки гидры. 1 — ядро, 2 — сократимый отросток. III. Нервные клетки морской лилии. [c.141]
В эктодерме, например, есть специальные клетки (стрекательные клетки), содержащие капсулу, заполненную ядовитой жидкостью. Плотная стенка этой капсулы в дистальной части образует спирально закрученную стрекательную нить, способную выбрасываться и парализовать тех мелких животных, которыми гидра питается. Большинство других экто- и энтодермальных клеток обладают мышечноподобными свойствами, что дает возможность гидре двигаться, как и положено хищнику. [c.45]
Преимущества, которые предоставляет членство в некоем сообществе клеток, не утрачиваются с увеличением размеров. Составляющие сообщество клетки могут специализироваться, в результате чего каждая из них начинает выполнять свою особую задачу более эффективно.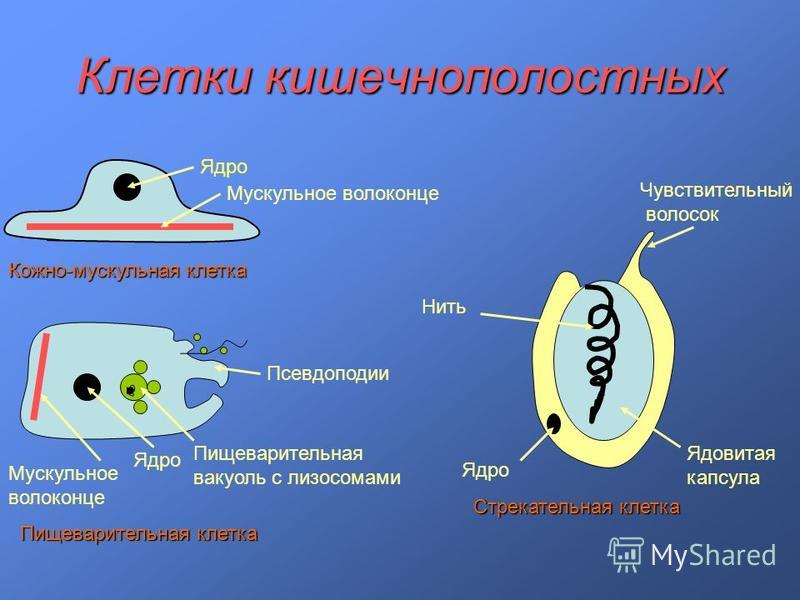 Специализированные клетки обслуживают другие клетки и в свою очередь пользуются услугами других клеток, специализированных в иных отношениях. Если клеток много, то одни становятся сенсорными клетками, обнаруживающими жертву, другие передают информацию, стрекательные клетки парализуют жертву, мышечные приводя в движение шупальцы, захватывающие жертву, секреторные клетки растворяют ее, а всасывающие абсорбируют соки. Не следует забывать, что все эти клетки, во всяком случае в таких телах, как наше, относятся к одному клону. Все они содержат одни и те же гены, хотя в клетках, специализированных к различным функциям, во включенном состоянии находятся различные гены. В клетках каждого типа гены непосредственно помогают тем собственным копиям, которые находятся в немногочисленных специализированных клетках, обеспечивающих размножение, — клетках бессмертной зародышевой линии. [c.199]
Специализированные клетки обслуживают другие клетки и в свою очередь пользуются услугами других клеток, специализированных в иных отношениях. Если клеток много, то одни становятся сенсорными клетками, обнаруживающими жертву, другие передают информацию, стрекательные клетки парализуют жертву, мышечные приводя в движение шупальцы, захватывающие жертву, секреторные клетки растворяют ее, а всасывающие абсорбируют соки. Не следует забывать, что все эти клетки, во всяком случае в таких телах, как наше, относятся к одному клону. Все они содержат одни и те же гены, хотя в клетках, специализированных к различным функциям, во включенном состоянии находятся различные гены. В клетках каждого типа гены непосредственно помогают тем собственным копиям, которые находятся в немногочисленных специализированных клетках, обеспечивающих размножение, — клетках бессмертной зародышевой линии. [c.199]
У обеих форм ротовое отверстие окружено щупальцами, которые содержат специальные стрекательные клетки, называемые книдобластами (отсюда и название типа nidaria). Под влиянием внешнего стимула они выбрасывают нити, выделяющие сильно-действующие яды, которые парализуют жертву (небольших ракообразных и личинок насекомых). Об этих ядах известно очень мало. Немногие изученные, по-видимому, представляют собой белки. Если токсические вещества стрекательных клеток ввести кроликам, наблюдается поражение нервной системы и образование антител, как и при инъекции змеиных ядов, столбнячного токсина и других ядовитых белков. [c.143]
Под влиянием внешнего стимула они выбрасывают нити, выделяющие сильно-действующие яды, которые парализуют жертву (небольших ракообразных и личинок насекомых). Об этих ядах известно очень мало. Немногие изученные, по-видимому, представляют собой белки. Если токсические вещества стрекательных клеток ввести кроликам, наблюдается поражение нервной системы и образование антител, как и при инъекции змеиных ядов, столбнячного токсина и других ядовитых белков. [c.143]
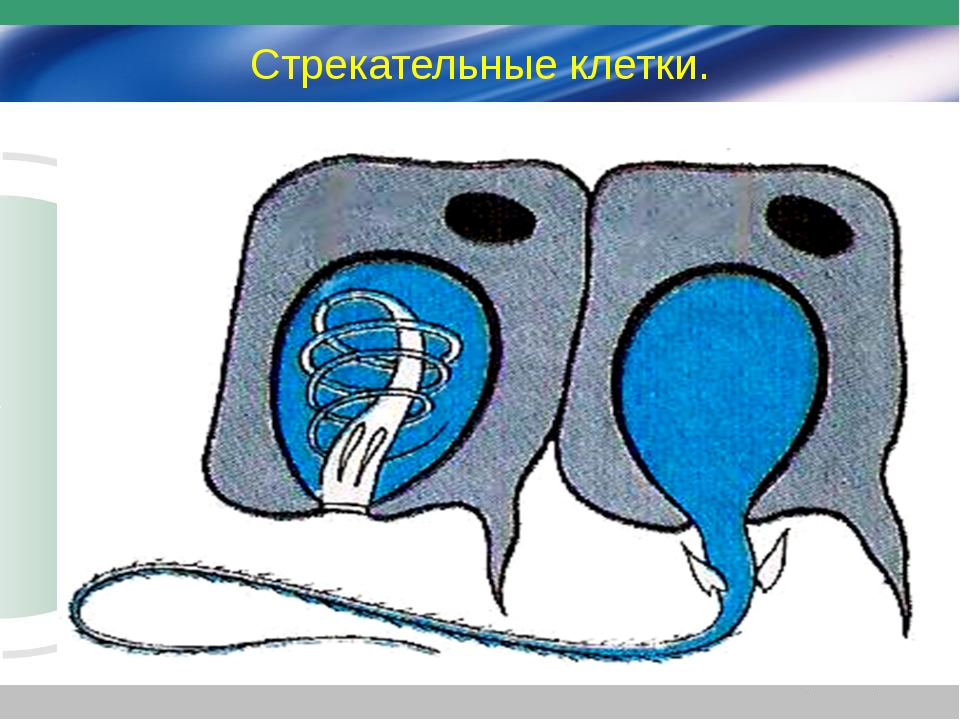
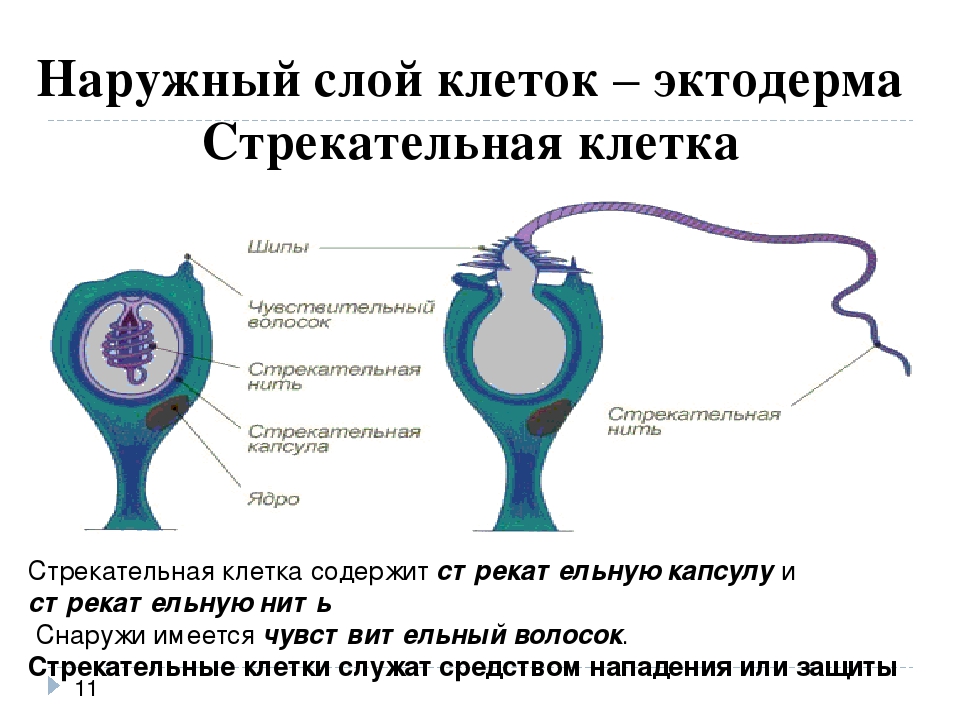
В наружном слое — эктодерме — прежде всего обращают на себя внимание так называемые стрекательные клетки, которые служат для защиты и нападения. Внутри такой клетки находится капсула, или пузырек, в котором спирально закручена нить. Снаружи этой клетки находится чувствительный шипик — книдоциль. При раздражении книдоциля нить с ядовитым содержимым энергично выбрасывается наружу. Во второй раз использовать стрекательные клетки гидра не может, так как они дегенерируют и вместо них развиваются новые. Благодаря присутствию стрекательных клеток рассматриваемый подтип кишечнополостных получал название стрекающих.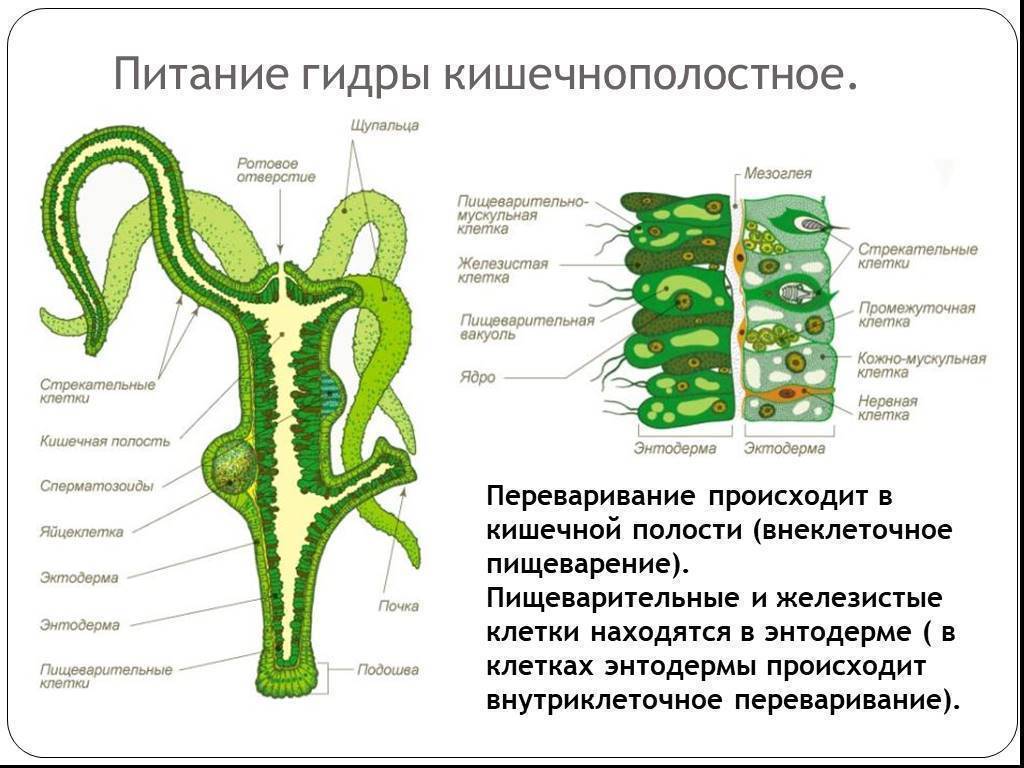 [c.324]
[c.324]
Морской червь Азр1до811оп, так же как и рак-отшельник, поселяется в пустых раковинах моллюсков. Затем иа таких раковинах находят пристанище актинии (рис. 186). Актииия использует своих симбионтов как средство передвижения. Защищаясь от врагов стрекательными клетками, оиа охраняет и своих партнеров. [c.459]
Стрекательные клетки кишечнополостных и действие их яда.
Стрекательное оружие медуз, коралов и полипов
Характернейшая особенность всех кишечнополостных, в том числе медуз, полипов, кораллов, — наличие стрекательных, или крапивных, клеток, которые развиваются из недифференцированных промежуточных клеток.
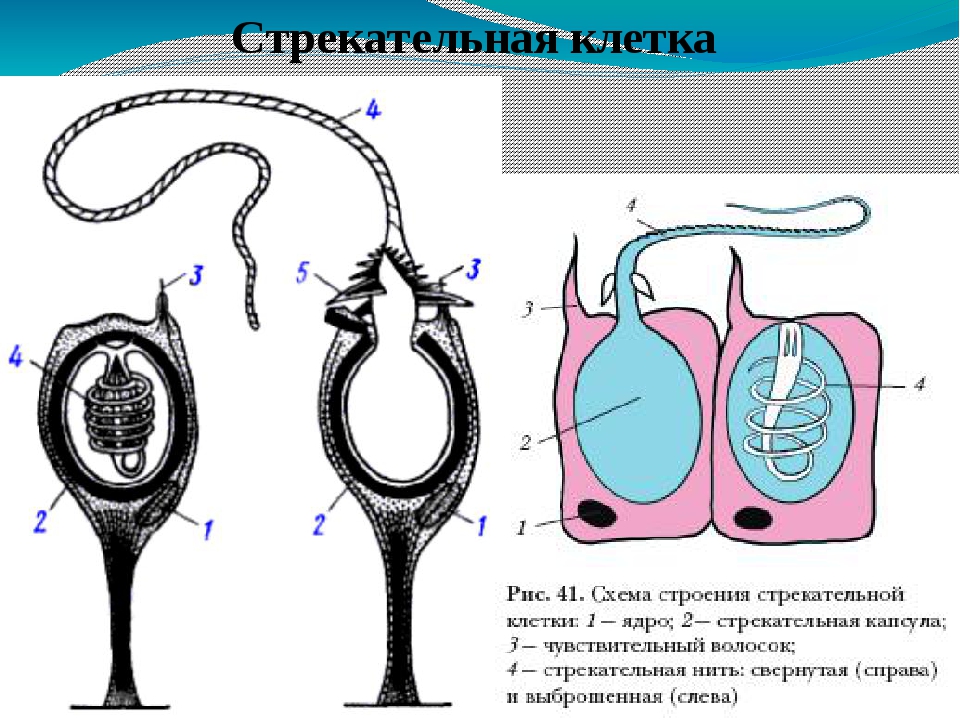
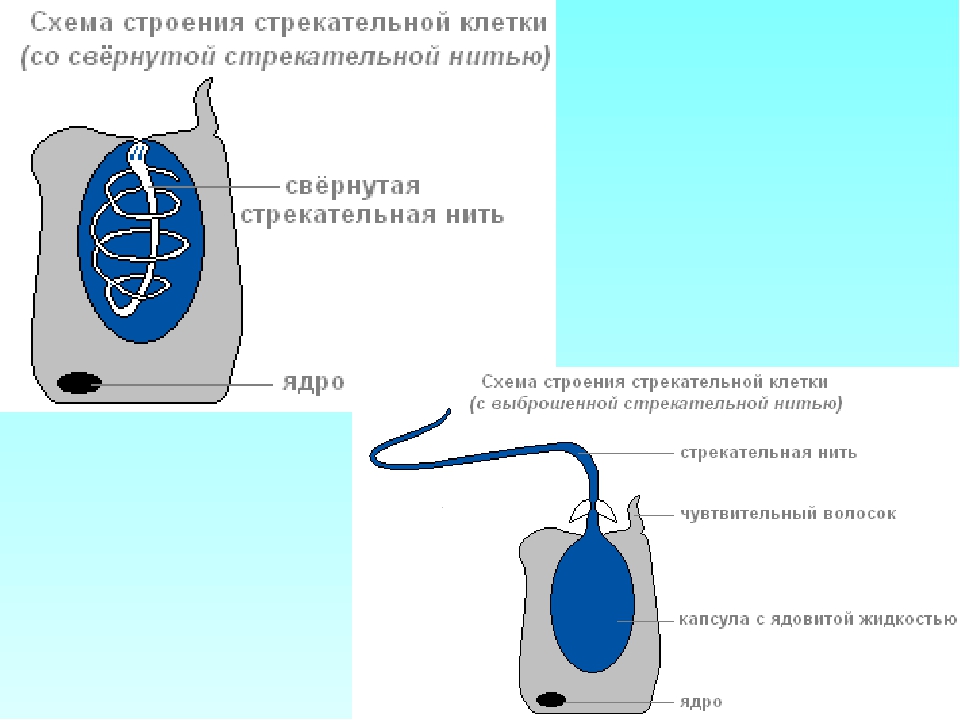
Каждая стрекательная клетка содержит хитиновую капсулу овальной или продолговатой формы. Стенки капсулы двухслойны. За счет первого (наружного) слоя на переднем конце капсулы формируется маленькая крышечка, второй же слой впячивается внутрь и дает начало тонкой спирально закрученной трубочке, называемой стрекательной нитью.
Полость капсулы заполнена ядовитой жидкостью.
На внешней поверхности стрекательной клетки имеется чувствительный волосок — книдоциль. Он представляет собой жгутик, сходный по устройству со жгутиком простейших, но, в отличие от них, неподвижный. Книдоциль окружен пальцевидными выростами цитоплазмы — микроворсинками, которые можно увидеть только с помощью микроскопа.
Малейшее прикосновение к книдоцилю вызывает возбуждение стрекательной клетки, проявляющееся в стремительном, подобном выстрелу, выворачивании стрекательной нити. В зависимости от деталей строения и способа воздействия на врага или жертву, различают множество типов строения стрекательных капсул.
Рассмотрим лишь главнейшие из них.
Одни имеют длинную, снабженную шипами нить. При «выстреле» такой капсулы нить вонзается в тело жертвы и изливает содержимое капсулы, вызывающее местное или общее отравление.
У других капсул нить короткая, лишенная шипов. Такие нити только опутывают жертву.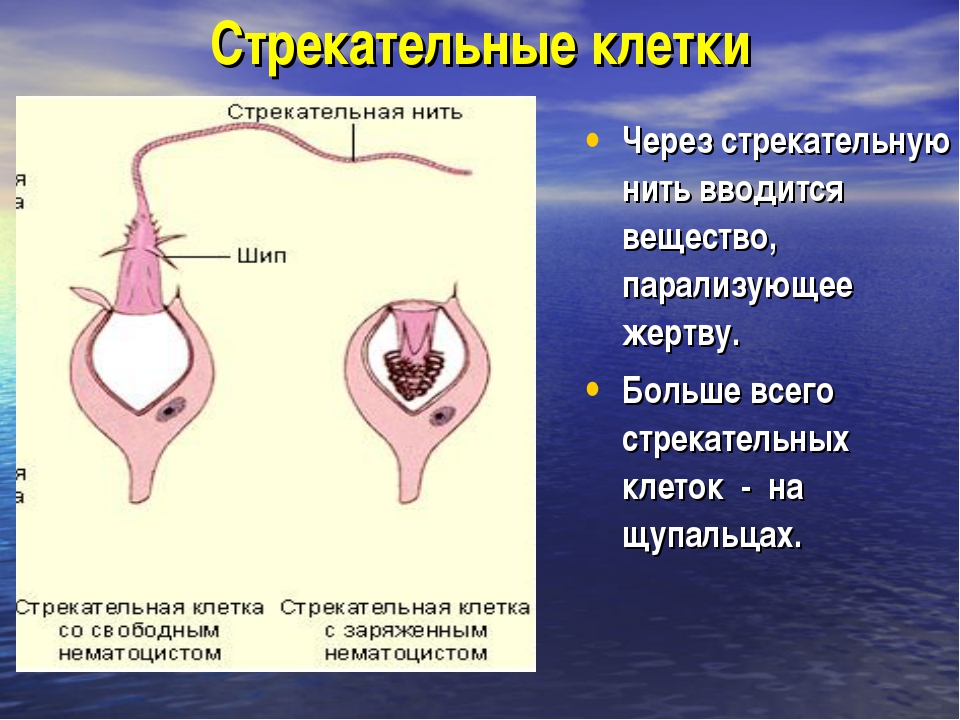
Наконец, имеются липкие нити, которые удерживают добычу приклеиванием. Они же могут служить и для временного приклеивания животного при передвижении.
Стрекательные клетки располагаются по всей поверхности тела кишечнополостных, но их наибольшие скопления наблюдаются на щупальцах и вокруг ротового отверстия, т.е. в местах, где они нужнее всего.
После «выстрела» стрекательная клетка погибает и на ее месте или поблизости от него развивается новая.
Интересно, что стрекательные капсулы могут действовать и после гибели животного. Так, прикосновение к мертвым, выброшенным на берег медузам может вызвать сильные покраснения кожи, сопровождающиеся жжением.
При поедании кишечнополостных некоторыми турбелляриями и заднежаберными моллюсками, часть стрекательных клеток переходит в тело хищника. Здесь стрекательные клетки располагаются в наружных покровах и могут нормально функционировать.
Стрекательные клетки, особенно содержащие ядовитую жидкость в капсулах, — грозное оружие, которое используется кишечнополостными (медузами, полипами, кораллами) и для защиты и для нападения.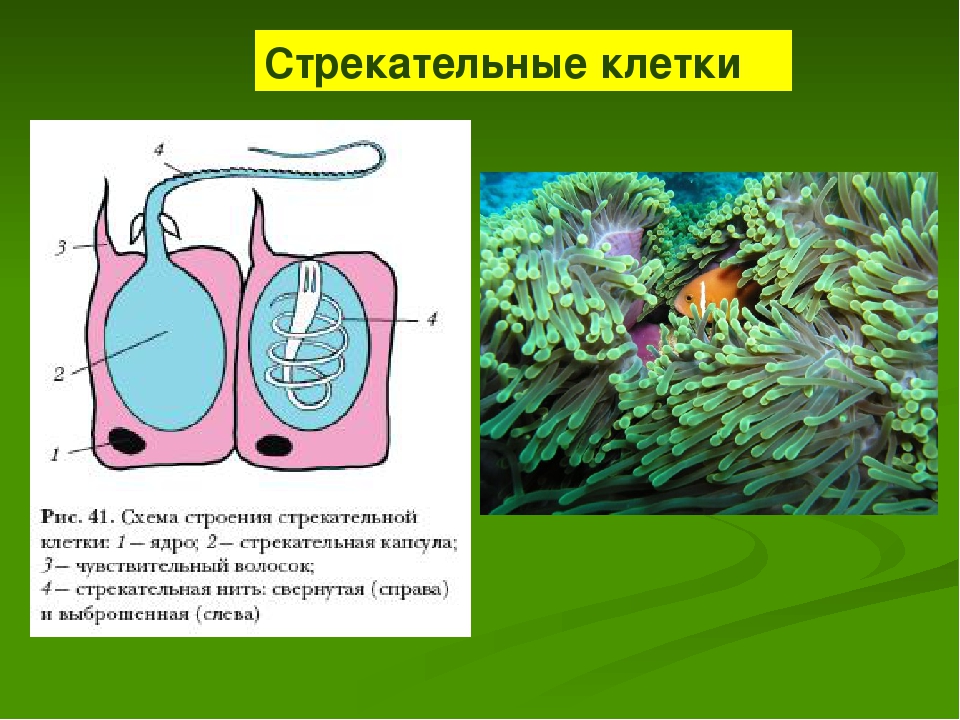 Мелкие животные, соприкасаясь с кишечнополостными, приклеиваются, опутываются и пронзаются стрекательными нитями, а в их тело попадает яд, вызывающий паралич или смерть.
Мелкие животные, соприкасаясь с кишечнополостными, приклеиваются, опутываются и пронзаются стрекательными нитями, а в их тело попадает яд, вызывающий паралич или смерть.
После этого жертва переносится щупальцами к ротовому отверстию и проглатывается. Даже довольно крупные животные часто оказываются пораженными ядом стрекательных капсул, вызывающим сильнейшую боль.
Сила воздействия яда различных видов кишечнополостных на человека неодинакова: некоторые из них совершенно безвредны, другие представляют собой серьезную опасность.
После прикосновения к нашим черноморским медузам корнеротам или аурелиям, вы почувствуете жжение, которое тем сильнее, чем нежнее кожа в месте контакта с медузой и похоже на ожог крапивой. Эти неприятные ощущения быстро проходят, не оставляя следа.
Однако в море имеется много гораздо более опасных кишечнополостных, прикосновение к которым может вызвать тяжелое заболевание и даже смерть человека. Так, многие актинии, обитающие, в том числе, и в Японском море, вызывают не только «ожоги», но и длительное опухание места контакта.
Тем не менее, наиболее неприятные последствия заключаются не в «ожогах» и опухолях, а в общем отравлении организма ядом стрекательных клеток.
У моряков, плавающих в тропических морях, уже издавна пользуется дурной славой красивая сифонофора физалия (Physalia physalis). Физалия снабжена большим, до 20 см длиной, плавательным пузырем, поднимающимся над поверхностью воды, от которого свешиваются вниз длиннейшие (до 30 м) ловчие щупальца, снабженные многочисленными стрекательными клетками.
Физалия очень ярко окрашена — пузырь отливает голубым, фиолетовым и пурпурным цветом, все свешивающиеся вниз придатки ультрамариновые. За яркую окраску физалия получила также название «португальского военного кораблика» — в средние века португальцы любили пестро окрашивать свои военные суда.
Плавающая на поверхности моря физалия видна издалека, и пловцы всегда остерегаются соприкосновения с ней, так как можно получить сильнейший «ожог», вызывающий жгучую боль. Пораженный физалией человек, даже если он отличный пловец, с трудом удерживается на воде. Вскоре после этого может наступить общее тяжелое заболевание с повышением температуры, длящееся несколько дней.
Вскоре после этого может наступить общее тяжелое заболевание с повышением температуры, длящееся несколько дней.
Физалия распространена исключительно в тропических морях и очень редко заносится течением и ветром в более теплые моря умеренных широт.
Нередко появляются сообщения о серьезных и даже смертельных отравлениях, вызываемых крупными сцифоидными медузами Chirodropus, Chirqsalmus, и, в особенности, Chironex из отряда кубомедуз, которые считаются одними из самых опасных существ, обитающих на нашей планете. Это тоже обитатели тропических морей. Их еще называют «морскими осами».
Высота колокола этих медуз достигает 10-15 см, по краю его сидят четыре разветвленных щупальца. Яд, содержащийся в их стрекательных клетках, попадая на кожу, вызывает омертвение эпидермиса и плохо заживающие язвы, после которых остаются глубокие рубцы.
Но самая большая опасность таится в воздействии яда на нервную систему, которое может привести к поражению дыхательного центра и вызвать смерть человека.
Впервые на ядовитость этих медуз обратили внимание в годы Второй Мировой войны в Австралии, куда было эвакуировано значительное число европейцев.
Стало известно несколько случаев загадочной гибели людей во время купания, причем на теле погибших не находили явных признаков повреждений. Долгое время тайна оставалась неразгаданной, но потом удалось установить, что причиной гибели является медуза хиродропус.
Получивший «ожог» человек захлебывался и тонул. Было отмечено, что среди пострадавших или погибших были, преимущественно, приезжие. Местные жители, особенно из числа коренного населения Австралии, могли купаться безбоязненно. По-видимому, у них выработался иммунитет к яду этой медузы.
В фауне наших дальневосточных морей также имеется один вид медуз, относящаяся к классу гидроидных, которая вызывает серьезные заболевания при соприкосновении с ней. Местные жители называют эту медузу «крестовичком» (научное название — Gonionemus vertens) за крестообразное расположение четырех темных радиальных каналов, вдоль которых тянутся четыре также темноокрашенные половые железы.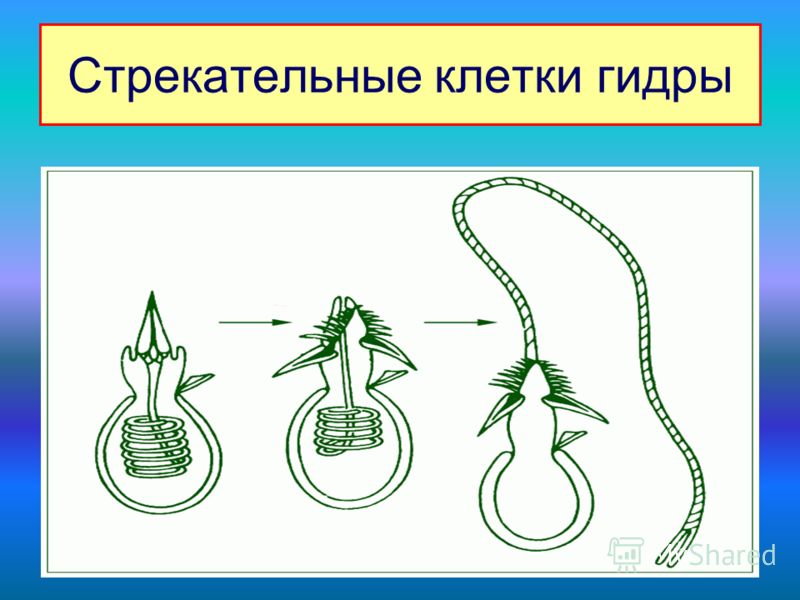
Зонтик медузы прозрачный, слабого желтовато-зеленого цвета, у отдельных экземпляров достигает 25 мм в диаметре, но обычно — 15-18 мм. На краю зонтика крестовичка имеется до 80 щупалец, способных сильно втягиваться и сокращаться. Щупальца этих медуз густо усажены стрекательными клетками, которые располагаются поясками. Посередине длины щупальца имеется присоска, при помощи которой медуза может прикрепляться к различным предметам под водой.
Крестовички обитают в Японском море и вблизи Курильских островов. Они обычно держатся на мелководье, в зарослях морской травы зостеры. В дождливые годы, когда вода у берегов сильно опресняется, медуз-крестовичков почти нет, они погибают. В засушливое лето они могут появляться массами.
Когда одно из щупалец крестовичка случайно коснется плавающего человека, медуза устремляется в этом направлении, пытаясь прикрепиться к нему присоской на щупальце. В этот момент купальщик получает сильнейший «ожог», через несколько минут кожа на теле «ожога» краснеет, покрывается волдырями. Уже через 10-30 минут наступает общая слабость, появляются боли в пояснице, затрудняется дыхание, немеют руки и ноги.
Уже через 10-30 минут наступает общая слабость, появляются боли в пояснице, затрудняется дыхание, немеют руки и ноги.
Очень сильный яд вырабатывают стрекательные клетки зоантарий — относительно мало изученных тропических шестилучевых кораллов. Отравление ядом (названным палитоксином) сказывается на деятельности сердца и всей системы кровообращения позвоночных, в том числе и человека. Яд зоантарий оказался в 100 раз сильнее яда кобры.
Статьи по теме: Проверочная работа по теме: ТИП КИШЕЧНОПОЛОСТНЫЕ
ТИП КИШЕЧНОПОЛОСТНЫЕ
Вставьте пропущенное слово.
1. Закончите предложения, вставив необходимые по смыслу слова.
A. Кишечнополостные – это … животные.
Б. Тело кишечнополостных имеет … форму.
B. Большинство кишечнополостных имеют … симметрию.
Г. Рот у кишечнополостных находится …
Д. Стенка тела кишечнополостных состоит из … клеток
Е. Отличительный признак кишечнополостных – наличие …
Ж. Тип «Кишечнополостные» делятся на классы: …, …, …, …
Тип «Кишечнополостные» делятся на классы: …, …, …, …
Найдите соответствие.
2. Напишите, какие клетки составляют: I – эктодерму, II – энтодерму и какую функцию они выполняют.
Клетки:
A. Стрекательная
Б. Нервная клетка
B. Кожно-мускульная
Г. Чувствительная
Д. Промежуточная
Е. Железистая
Ж. Эпителиально-мускульная
Функции:
1. Дают начало всем другим клеткам
2. Пищеварение
3. Движение
4. Чувствительная
5. Защита и нападение
6. Защитная
3. Выпишите признаки (обозначенные буквами), характерные для представителей кишечнополостных (обозначенные цифрами).
I. Гидра
II. Корнерот
III. Актиния
IV. Кораллы
A. Два слоя клеток
Б. Имеют стрекательные клетки
B. Радиальная симметрия
Г. Многоклеточный организм
Д. Прикрепляются к субстрату
Е. Обитают в пресных водоемах
Ж. Тело имеет вид зонтика или колокола
3. Прикрепляются к телу морских животных
И. Разрастаясь, образуют рифы
Разрастаясь, образуют рифы
К. Хорошо развит известковый скелет
Л. Одиночный полип
М. Исключительно колониальные формы
4. Из предложенных терминов составьте схему жизненного цикла пресноводной гидры:
A. Яйцеклетка
Б. Сперматозоид
B. Бугорок на эктодерме
Г. Яйцеклетка под оболочкой
Д. Оплодотворение
Е. Выход наружу
Ж. Дробление
3. Гаструляция
Выберите правильный ответ.
5. Эктодерма и энтодерма разделяются:
A. Мембраной
Б. Базальной мембраной
B. Оболочкой
Г. Мезоглеей
6. Стрекательные клетки характерны:
A. Для всех кишечнополостных
Б. Только для медуз
B. Только для гидры
Г. Только для актиний
7. Раздражимостью называют:
A. Любое движение животного
Б. Ответную реакцию на нападение
B. Защитную реакцию
Г. Способность организма отвечать на воздействие окружающей среды своей деятельностью
8. Гермафродиты – это организмы с признаками:
A.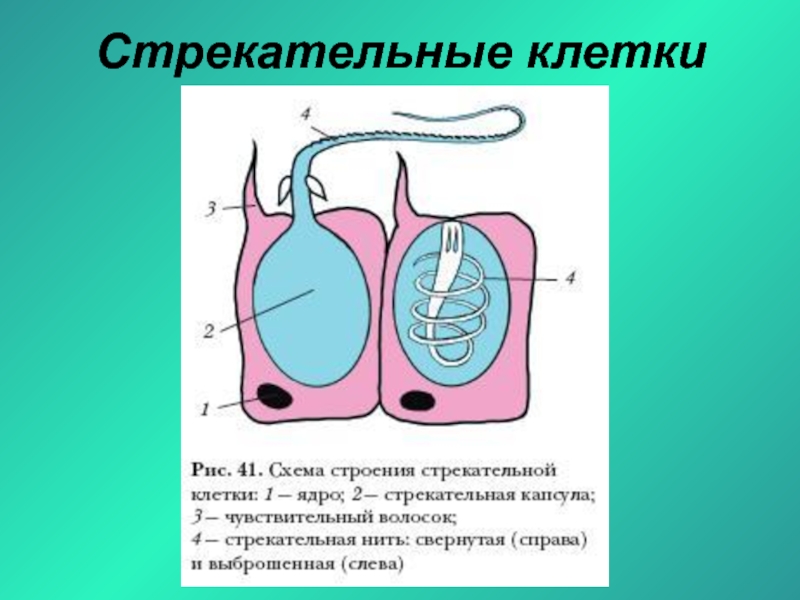 Мужского организма
Мужского организма
Б. Женского организма
B. Мужского и женского организма
Г. Однополые
9. Планула – это:
A. Личинка, покрытая ресничками
Б. Название медузы
B. Вид клетки
Г. Слой стенки
10. Гаструляция – это:
A. Образование двухслойного организма
Б. Вид размножения
B. Способ питания
Г. Восстановление утраченных частей тела
Выберите верное утверждение.
11. 1. В эктодерме у кишечнополостных присутствуют клетки: стрекательные, эпителиально-мускульные, промежуточные, нервные.
2. В энтодерме у кишечнополостных присутствуют железистые и эпителиально-мускульные клетки.
3. У кишечнополостных между эктодермой и энтодермой находится базальная мембрана.
4. У кишечнополостных чувствительные (нервные) клетки образуют диффузную нервную систему.
5. Железистые клетки эктодермы гидры выделяют клейкий секрет и распределены равномерно по всей эктодерме.
6. Стрекательные клетки – отличительный признак кишечнополостных, и находятся на переднем конце тела.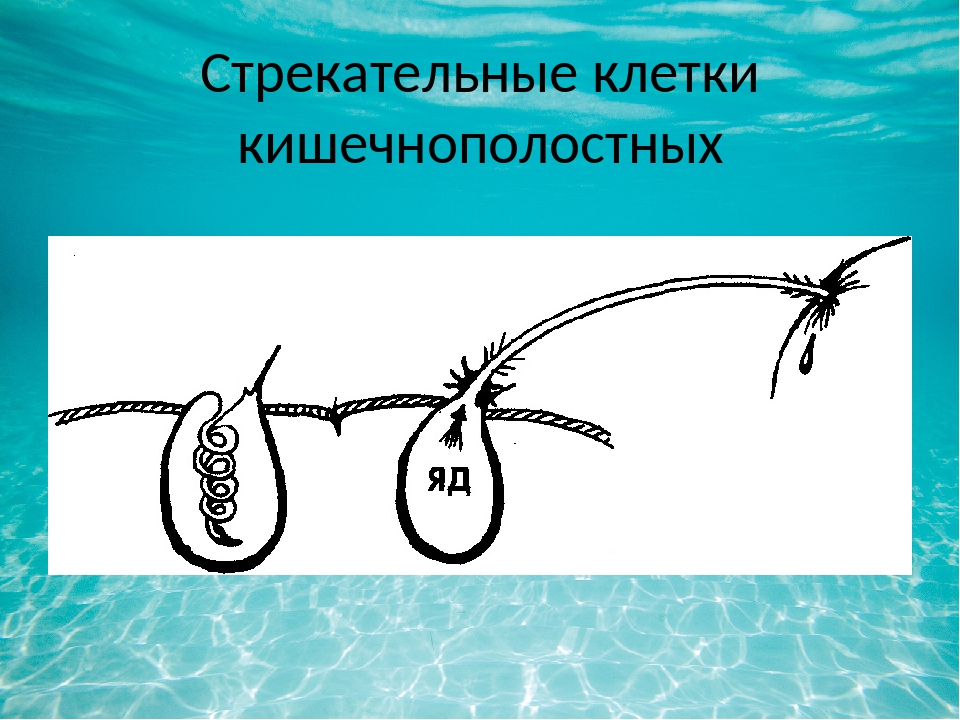
7. Половые клетки у гидры образуются из промежуточных клеток.
8. Гидры могут быть обоеполыми.
9. Оплодотворение у гидры происходит на ее теле.
10. Эмбриональный период у гидры включает два этапа: дробление и гаструляцию.
11. У некоторых кишечнополостных формируются специальные половые железы, где образуются половые клетки. Оплодотворение и развитие яйца происходит вне материнского организма.
12. У гидроидных из яйца выходит личинка – планула, которая проходит стадию паренхимулы.
Комментирование и размещение ссылок запрещено.
Гидра: стрекательные клетки
Гидра: стрекательные клетки
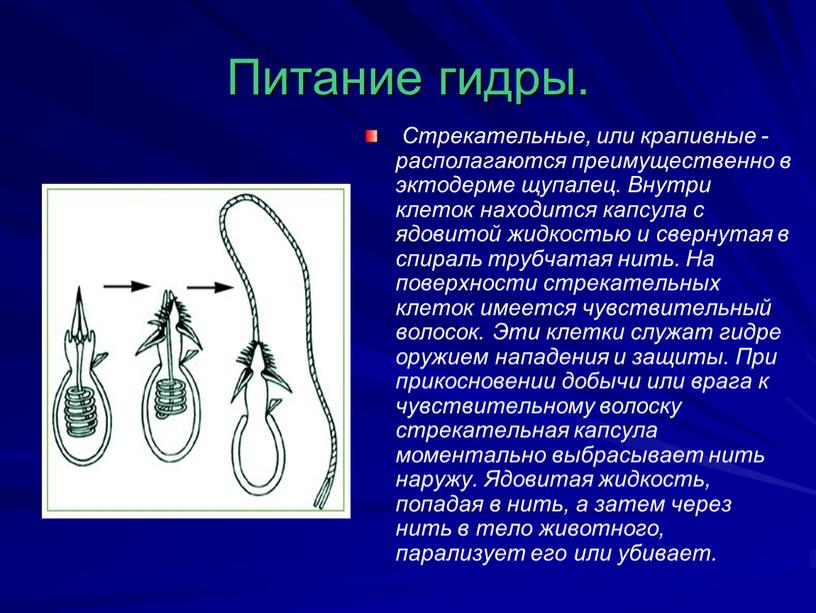
Характерная черта кишечнополостных — присутствие в покровах стрекательных
клеток (
рис. 93
). Они развиваются из промежуточных клеток и содержат особую овальную
стрекательную капсулу с плотными стенками. Капсула наполнена жидкостью, а
на одном конце капсулы стенка ее впячена внутрь в виде очень тонкого, но
полого отростка, который закручивается в капсуле в спирально завитую
стрекательную нить. Стрекательные клетки служат гидре орудием нападения и
защиты.
Стрекательные клетки служат гидре орудием нападения и
защиты.
На наружной поверхности клетки имеется тонкий чувствительный волосок —
книдоциль. Изучение стрекательных клеток с помощью электронного микроскопа
показало значительную сложность строения книдоциля (
рис. 93
). Он состоит из длинного жгутика, окруженного 18-22 тонкими пальцевидными
выростами цитоплазмы — микроворсинками. По строению жгутик книдоциля очень
сходен со жгутиками и ресничками простейших, но в отличие от них
неподвижен. При прикосновении добычи или врага к жгутику последний
отклоняется и задевает одну или несколько микроворсинок, что приводит к
возбуждению стрекательной клетки. При этом стрекательная капсула
выбрасывает выворачивающуюся из нее наружу упругую нить, которая
распрямляется, как стрела. Нить наподобие гарпуна усажена обращенными назад
шипиками, а в основании несет более крупные шипы. Уколы нити ядовиты и
могут парализовать мелких животных. После выбрасывания нити стрекательная
клетка погибает.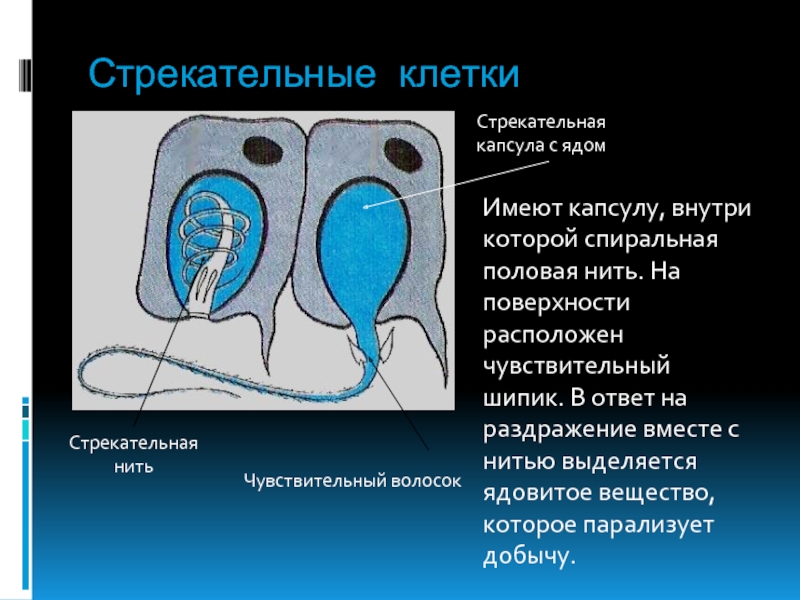
У гидры имеется несколько категорий капсул, функции которых различны. Рассмотренные крупные капсулы, служащие для пробивания покровов и поражения добычи, называются пенетрантами ( рис. 93 ). Значительно более мелкие — вольвенты — имеют короткие спирально закрученные нити, которые обвиваются вокруг различных выступов (щетинок, волосков и т.п.) на теле добычи и таким путем удерживают ее. Наконец, вытянутые стрекательные капсулы — глютинанты — приклеиваются к телу добычи длинными липкими нитями.
Ссылки:
© Jonathan Bird / ORG Тип Cnidaria (произносится как nid-AIR-ee-ah) содержит около 9000 живых видов во всем мире. Они относятся к числу самых простых из так называемых «высших» организмов, но также относятся к числу самых красивых. Существа этого типа радиально симметричны. Книдарийцы включают гидроидов, медуз, анемонов и кораллов. У всех книдарийцев есть щупальца с жалящими клетками на концах, которые используются для захвата и подчинения добычи. Фактически, название типа «Книдарианец» буквально означает «жалящее существо». Жалящие клетки называются книдоцитами и содержат структуру, называемую нематоцистой. Нематоциста представляет собой спиралевидное жало, похожее на нитку.Когда нематоциста запускается, нить разматывается и выпрямляется. Гарпуноподобная нить проникает сквозь стенку книдоцита в жертву. У большинства книдарийцев также есть токсин в жале, который помогает вывести добычу из строя. Нематоциста запускается либо прикосновением щупальца к чему-либо, либо, в некоторых случаях, нервным импульсом от животного, который приказывает ему стрелять. Большинство нематоцист Книдарианцев не опасны для человека, поскольку жало не может проникнуть в кожу человека в достаточной степени, чтобы нанести какой-либо вред.Однако есть некоторые медузы, которые могут доставить людям очень болезненные, а в некоторых случаях даже смертельные укусы. Книдариец может иметь один из двух основных типов телосложения: полиповидное или медузоидное. Полипоид (POL-ip-oyd) представляет собой конфигурацию кораллов и анемонов, щупальца и рот обычно обращены вверх, а другая сторона прикреплена к субстрату или соединена с колонией других существ того же вида. У Медузоида организм в основном перевернут, а рот и щупальца обычно направлены вниз.Эти типы книдарийцев обычно свободно плавают, как медузы. Вот почему анемон иногда называют перевернутой медузой; вот примерно то, что есть! Тип делится на 3 класса, называемых Hydrozoans (что означает «водные животные»), Anthozoans (что означает «животные-цветы») и Scyphozoans (что означает «животные-чаши»). HYDROZOANS СКИФОЗОНЫ Смотрите новый бесплатный интернет-сериал о подводном мире: СИНИЙ МИР ДЖОНАТАНА ПТИЦЫ | Две разные формы тела книдарийцев. Колония кораллов состоит из сотен или тысяч крошечных полипов. Каждый полип — это отдельное животное (в основном небольшой анемон), но они живут вместе, как группа. Медуза с львиной гривой — ядовитый скифоз, который своими длинными щупальцами может ужалить людей. Поскольку щупальца могут свисать так далеко вниз, желе может использовать эти щупальца, чтобы убивать рыбу, которая плывет через них, даже не видя само желе вверху! Это пример книдария с формой медузоида . |
 Это означает, что части тела выходят наружу из центра, как спицы велосипедного колеса.Распространенный пример радиальной симметрии — морская звезда (член филума иглокожих) или анемон, книдариец (см. Ниже).
Это означает, что части тела выходят наружу из центра, как спицы велосипедного колеса.Распространенный пример радиальной симметрии — морская звезда (член филума иглокожих) или анемон, книдариец (см. Ниже).


 Они выглядят как нежные растения, но это животные, которые кусают и захватывают пищу.
Они выглядят как нежные растения, но это животные, которые кусают и захватывают пищу. Определение профилей экспрессии по типу клеток раскрывает развитие и эволюцию стрекательных клеток морского анемона | BMC Biology
Дэвид К.Н., Озбек С., Адамчик П., Мейер С., Поли Б., Чепмен Дж., Хванг Дж. С., Годжобори Т., Голштинская обл. Эволюция сложных структур: миниколлагены формируют книдарийную нематоцисту. Тенденции Genet. 2008. 24 (9): 431–8.
CAS Статья Google ученый

Касс-Саймон Г., Скаппатиччи А.А. Поведенческая физиология и физиология развития нематоцист. Может J Zool. 2002. 80 (10): 1772–94.
Артикул Google ученый
Layden MJ, Rentzsch F, Rottinger E. Возникновение звездчатого морского анемона Nematostella vectensis в качестве модельной системы для исследования развития и регенерации. Wiley Interdiscip Rev Dev Biol. 2016; 5 (4): 408–28.
Артикул Google ученый
Meier S, Jensen PR, Adamczyk P, Bachinger HP, Holstein TW, Engel J, Ozbek S, Grzesiek S. Анализ последовательности-структуры и структурно-функции в богатых цистеином доменах, образующих ультрастабильную стенку нематоцисты.J Mol Biol. 2007. 368 (3): 718–28.
CAS Статья Google ученый
 org/ScholarlyArticle»> 5.
org/ScholarlyArticle»> 5.Adamczyk P, Meier S, Gross T, Hobmayer B, Grzesiek S, Bachinger HP, Holstein TW, Ozbek S. Миниколлаген-15, новый миниколлаген, выделенный из гидры, образует структуры канальцев в нематоцистах. J Mol Biol. 2008. 376 (4): 1008–20.
CAS Статья Google ученый
Hwang JS, Takaku Y, Momose T., Adamczyk P, Ozbek S, Ikeo K, Khalturin K, Hemmrich G, Bosch TC, Holstein TW, et al.Нематогалектин, белок нематоцисты с доменами GlyXY и галектина, демонстрирует специфический для нематоцитов альтернативный сплайсинг у гидры. Proc Natl Acad Sci U S. A. 2010; 107 (43): 18539–44.
CAS Статья Google ученый
Nuchter T, Benoit M, Engel U, Ozbek S, Holstein TW. Наносекундная кинетика разряда нематоцист. Curr Biol. 2006; 16 (9): R316–8.
Наносекундная кинетика разряда нематоцист. Curr Biol. 2006; 16 (9): R316–8.
Артикул Google ученый
Balasubramanian PG, Beckmann A, Warnken U, Schnoelzer M, Schueler A, Bornberg-Bauer E, Holstein TW, Ozbek S. Протеом нематоцисты гидры. J Biol Chem. 2012; 287 (13): 9672-81.
Hwang JS, Ohyanagi H, Hayakawa S, Osato N, Nishimiya-Fujisawa C, Ikeo K, David CN, Fujisawa T., Gojobori T. Эволюционное появление генов, специфичных для определенного типа клеток, на основе анализа экспрессии генов Гидры. Proc Natl Acad Sci U S. A. 2007; 104 (37): 14735–40.
CAS Статья Google ученый

Milde S, Hemmrich G, Anton-Erxleben F, Khalturin K, Wittlieb J, Bosch TC. Характеристика таксономически ограниченных генов в ограниченном по филуму типу клеток. Genome Biol. 2009; 10 (1): R8.
Артикул Google ученый
Technau U, Steele RE. Эволюционный перекресток в биологии развития: Cnidaria. Разработка. 2011. 138 (8): 1447–58.
CAS Статья Google ученый
Hsu PD, Lander ES, Zhang F. Разработка и применение CRISPR-Cas9 для геномной инженерии. Клетка. 2014. 157 (6): 1262–78.
CAS Статья Google ученый
Райт А. В., Нуньез Дж. К., Дудна Дж. А. Биология и приложения систем CRISPR: использование природных инструментов для геномной инженерии. Клетка. 2016; 164 (1–2): 29–44.
В., Нуньез Дж. К., Дудна Дж. А. Биология и приложения систем CRISPR: использование природных инструментов для геномной инженерии. Клетка. 2016; 164 (1–2): 29–44.
CAS Статья Google ученый
Зенкерт К., Такахаши Т., Диснер М.О., Озбек С. Морфологический и молекулярный анализ cnidom Nematostella vectensis. PLoS One. 2011; 6 (7): e22725.
CAS Статья Google ученый
Shaner NC, Campbell RE, Steinbach PA, Giepmans BN, Palmer AE, Tsien RY. Улучшенные мономерные красные, оранжевые и желтые флуоресцентные белки, полученные из Discosoma sp. красный флуоресцентный белок. Nat Biotechnol. 2004. 22 (12): 1567–72.
CAS Статья Google ученый
 org/ScholarlyArticle»> 16.
org/ScholarlyArticle»> 16.Ikmi A, McKinney SA, Delventhal KM, Gibson MC. TALEN и CRISPR / Cas9-опосредованное редактирование генома в раннем ветвлении многоклеточного животного Nematostella vectensis. Nat Commun. 2014; 5: 5486.
CAS Статья Google ученый
Renfer E, Amon-Hassenzahl A, Steinmetz PR, Technau U. Мышечно-специфическая трансгенная репортерная линия морского анемона Nematostella vectensis. Proc Natl Acad Sci U S. A. 2010; 107 (1): 104–8.
CAS Статья Google ученый
Колумб-Шенкар Ю.Ю., Сачкова М.Ю., Макрандер Дж., Фридрих А., Модепалли В., Рейцель А.М., Сунагар К., Моран Ю. Динамика состава яда на протяжении сложного жизненного цикла. Элиф. 2018; 7: e35014.
Моран Ю., Прахер Д., Шлезингер А., Аялон А., Тал И., Технау Ю. Анализ содержания растворимого белка в нематоцистах модельной морской анемона проливает свет на эволюцию яда. Mar Biotechnol (Нью-Йорк). 2013. 15 (3): 329–39.
CAS Статья Google ученый
Marlow HQ, Srivastava M, Matus DQ, Rokhsar D, Martindale MQ. Анатомия и развитие нервной системы Nematostella vectensis, антозойных книдарий. Dev Neurobiol. 2009. 69 (4): 235–54.
CAS Статья Google ученый
Nakanishi N, Renfer E, Technau U, Rentzsch F. Нервные системы морского анемона Nematostella vectensis генерируются эктодермой и энтодермой и формируются с помощью различных механизмов.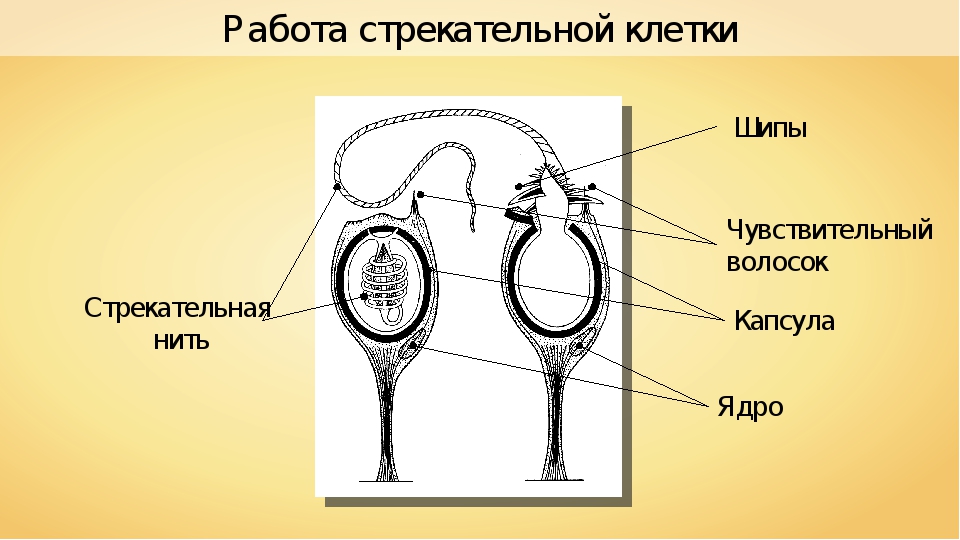 Разработка.2012. 139 (2): 347–57.
Разработка.2012. 139 (2): 347–57.
CAS Статья Google ученый
Воленски Ф.С., Лейден М.Дж., Мартиндейл М.К., Гилмор Т.Д., Финнерти-младший. Характеристика пространственно-временной экспрессии РНК и белков в морской ветренице, Nematostella vectensis. Nat Protoc. 2013. 8 (5): 900–15.
Артикул Google ученый
Babonis LS, Martindale MQ. PaxA, но не PaxC, необходим для развития книдоцитов у морского анемона Nematostella vectensis.Еводево. 2017; 8: 14.
Артикул Google ученый
Озбек С. Книдарная нематоциста: миниатюрный внеклеточный матрикс внутри секреторного пузырька. Протоплазма. 2011; 248 (4): 635–40.
Протоплазма. 2011; 248 (4): 635–40.
Артикул Google ученый
Бушар С., Прайс РБ, Манипенни К.Г., Томпсон Л.Ф., Зиллхардт М., Сталхейм Л., Андерсон, Пенсильвания. Клонирование и функциональная экспрессия субъединиц потенциал-управляемых ионных каналов из книдоцитов португальского человека О’Вара Physalia Physalis.J Exp Biol. 2006. 209 (Pt 15): 2979–89.
CAS Статья Google ученый
Li X, Martinson AS, Layden MJ, Diatta FH, Sberna AP, Simmons DK, Martindale MQ, Jegla TJ. Управляемые напряжением K + -каналы семейства Ether-a-go-go развились у предка многоклеточного животного и функционально разнообразны у предка книдарий-билатерий. J Exp Biol. 2015; 218 (Pt 4): 526–36.
Артикул Google ученый
 org/ScholarlyArticle»> 27.
org/ScholarlyArticle»> 27.Moran Y, Zakon HH. Эволюция четырех субъединиц потенциалзависимых кальциевых каналов: древние корни, возрастающая сложность и множественные потери. Genome Biol Evol. 2014; 6 (9): 2210–7.
Артикул Google ученый
Эльран Р., Раам М., Краус Р., Брехман В., Шер Н., Плашкес И., Халифа-Каспи В., Лотан Т. Ранняя и поздняя реакция транскриптома Nematostella vectensis на тяжелые металлы. Mol Ecol. 2014. 23 (19): 4722–36.
CAS Статья Google ученый
Hess J, Angel P, Schorpp-Kistner M. Подразделения AP-1: ссоры и согласие между братьями и сестрами. J Cell Sci. 2004; 117 (Pt 25): 5965–73.
CAS Статья Google ученый
 org/ScholarlyArticle»> 30.
org/ScholarlyArticle»> 30.Мэн Кью, Ся Й. ц-Цзюнь, на перекрестке сигнальной сети. Белковая клетка. 2011; 2 (11): 889–98.
CAS Статья Google ученый
Лейден М.Дж., Роттингер Э., Воленски Ф.С., Гилмор Т.Д., Мартиндейл М.К.Микроинъекция мРНК или морфолино для обратного генетического анализа в морской ветреницы, Nematostella vectensis. Nat Protoc. 2013; 8 (5): 924–34.
Артикул Google ученый
Magie CR, Daly M, Martindale MQ. Гаструляция у Nematostella vectensis книдарии происходит через инвагинацию, а не проникновение. Dev Biol. 2007. 305 (2): 483–97.
CAS Статья Google ученый
 org/ScholarlyArticle»> 33.
org/ScholarlyArticle»> 33.Рентч Ф., Фритценванкер Дж. Х., Шольц С. Б., Технау У. Передача сигналов FGF контролирует формирование апикального сенсорного органа у книдарий Nematostella vectensis. Разработка. 2008. 135 (10): 1761–9.
CAS Статья Google ученый
Shaner NC, Lin MZ, McKeown MR, Steinbach PA, Hazelwood KL, Davidson MW, Tsien RY. Повышение фотостабильности ярких мономерных оранжевых и красных флуоресцентных белков. Нат методы. 2008. 5 (6): 545–51.
CAS Статья Google ученый
Генихович Г., Технау Ю. Сложные функции вариантов сплайсинга Mef2 в дифференцировке энтодермы и нейронального типа клеток у морского анемона. Разработка. 2011. 138 (22): 4911–9.
CAS Статья Google ученый
 org/ScholarlyArticle»> 36.
org/ScholarlyArticle»> 36.Mouhat S, Jouirou B, Mosbah A, De Waard M, Sabatier JM. Разнообразие складок в токсинах животных, действующих на ионные каналы.Biochem J. 2004; 378 (Pt 3): 717–26.
CAS Статья Google ученый
Engel U, Ozbek S, Streitwolf-Engel R, Petri B, Lottspeich F, Holstein TW. Nowa, новый белок с миниколлагеновыми Cys-богатыми доменами, участвует в образовании нематоцист у гидры. J Cell Sci. 2002; 115 (Pt 20): 3923–34.
CAS Статья Google ученый
Friedberg F, Rhoads AR.Эволюционные аспекты кальмодулина. IUBMB Life. 2001. 51 (4): 215–21.
CAS Статья Google ученый
 org/ScholarlyArticle»> 39.
org/ScholarlyArticle»> 39.Лаббок Р., Гупта Б.Л., Холл ТА. Новая роль кальция в экзоцитозе: механизм выделения нематоцист, показанный с помощью рентгеновского микроанализа. Proc Natl Acad Sci U S. A. 1981; 78 (6): 3624–8.
CAS Статья Google ученый
Агрон М., Брехман В., Моргенштерн Д., Лотан Т.Регулирование AP-1 с помощью передачи сигналов MAPK у морского анемона, подвергшегося стрессу металлом. Cell Physiol Biochem. 2017; 42 (3): 952–64.
CAS Статья Google ученый
Синдзато К., Сёгучи Е., Кавасима Т., Хамада М., Хисата К., Танака М., Фуджи М., Фудзивара М., Коянаги Р., Икута Т. и др. Использование генома Acropora digitifera для понимания реакции кораллов на изменение окружающей среды.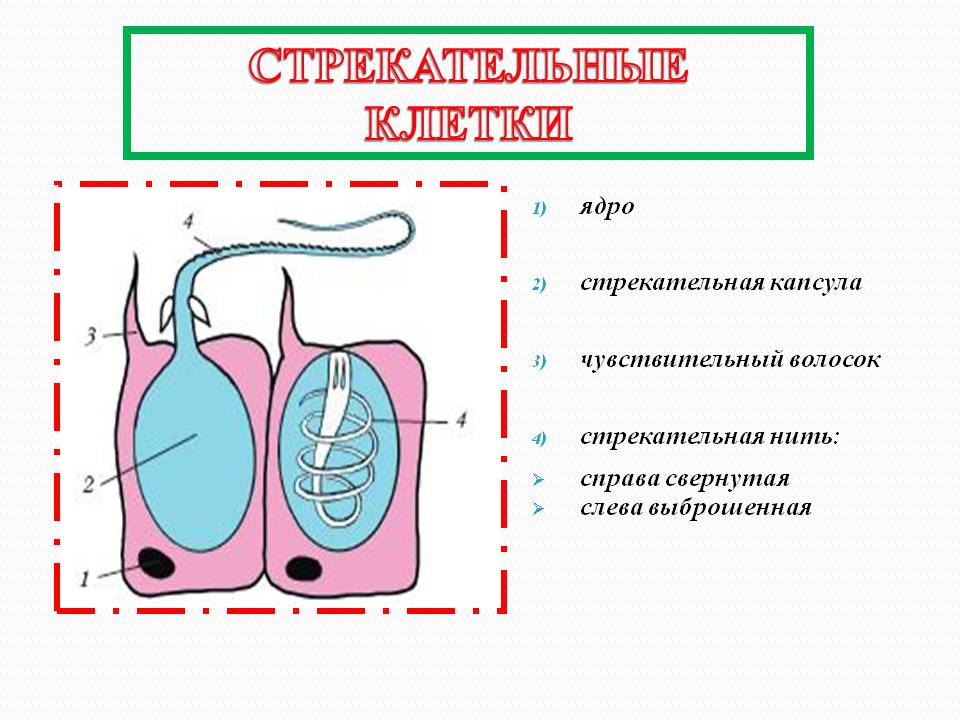 Природа. 2011. 476 (7360): 320–3.
Природа. 2011. 476 (7360): 320–3.
CAS Статья Google ученый
Рахамим Т., Моргенштерн Д., Ахаронович Д., Брехман В., Лотан Т., Шер Д. Динамически развивающееся содержание нематоцист антозоя, сцифоза и гидрозоя. Mol Biol Evol. 2015; 32 (3): 740–53.
CAS Статья Google ученый
Grunder S, Assmann M. Пептидно-управляемые ионные каналы и простая нервная система гидры. J Exp Biol. 2015; 218 (Pt 4): 551–61.
Артикул Google ученый
Гур Барзилай М., Рейцель А.М., Краус Дж. Э., Гордон Д., Технау У., Гуревиц М., Моран Ю.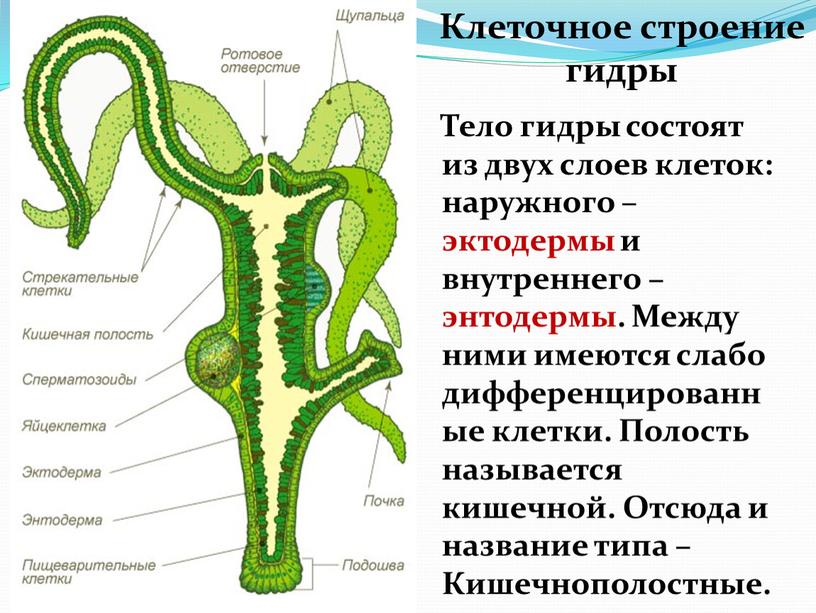 Конвергентная эволюция селективности по ионам натрия в передаче сигналов нейронов у многоклеточных животных. Cell Rep. 2012; 2 (2): 242–8.
Конвергентная эволюция селективности по ионам натрия в передаче сигналов нейронов у многоклеточных животных. Cell Rep. 2012; 2 (2): 242–8.
Артикул Google ученый
Jegla T, Marlow HQ, Chen B, Simmons DK, Jacobo SM, Martindale MQ. Расширенное функциональное разнообразие шейкерных K (+) каналов у книдарий обусловлено экспансией генов. PLoS One. 2012; 7 (12): e51366.
CAS Статья Google ученый
Генихович Г., Технау У. Индукция нереста морской ветреницы Nematostella vectensis, экстракорпоральное оплодотворение гамет и удаление желе из зигот. Cold Spring Harb Protoc. 2009; 2009 (9): pdb.prot5281.
PubMed Google ученый
 org/ScholarlyArticle»> 47.
org/ScholarlyArticle»> 47.Bolger AM, Lohse M, Usadel B. Trimmomatic: гибкий триммер для данных последовательности Illumina. Биоинформатика. 2014; 30 (15): 2114–20.
CAS Статья Google ученый
Andrews S. FastQC: инструмент контроля качества для данных последовательности с высокой пропускной способностью. 2010. Доступно в Интернете по адресу: http://www.bioinformatics.babraham.ac.uk/projects/fastqc. По состоянию на 23 сентября 2018 г.
Putnam NH, Srivastava M, Hellsten U, Dirks B, Chapman J, Salamov A, Terry A, Shapiro H, Lindquist E, Kapitonov VV, et al. Геном морского анемона раскрывает репертуар и геномную организацию предкового гена эуметазоана. Наука. 2007. 317 (5834): 86–94.
CAS Статья Google ученый
 org/ScholarlyArticle»> 50.
org/ScholarlyArticle»> 50.Добин А., Дэвис К.А., Шлезингер Ф., Дренков Дж., Залески С., Джа С., Батут П., Шассон М., Джингерас Т.Р. STAR: сверхбыстрый универсальный выравниватель RNA-seq. Биоинформатика. 2013. 29 (1): 15–21.
CAS Статья Google ученый
Андерс С., Пил П. Т., Хубер В. HTSeq — среда Python для работы с данными высокопроизводительного секвенирования. Биоинформатика. 2015; 31 (2): 166–9.
CAS Статья Google ученый
Moran Y, Fredman D, Praher D, Li XZ, Wee LM, Rentzsch F, Zamore PD, Technau U, Seitz H. Книдариевые микроРНК часто регулируют мишени путем расщепления. Genome Res. 2014; 24 (4): 651–63.
CAS Статья Google ученый
 org/ScholarlyArticle»> 53.
org/ScholarlyArticle»> 53.Schwaiger M, Schonauer A, Rendeiro AF, Pribitzer C, Schauer A, Gilles AF, Schinko JB, Renfer E, Fredman D, Technau U. Эволюционное сохранение регуляторного ландшафта гена эвметазоа. Genome Res.2014; 24 (4): 639–50.
CAS Статья Google ученый
Лав М.И., Хубер В., Андерс С. Умеренная оценка кратного изменения и дисперсии данных РНК-секвенирования с помощью DESeq2. Genome Biol. 2014; 15 (12): 550.
Артикул Google ученый
Робинсон, доктор медицины, Маккарти, ди-джей, Смит, Г.К. edgeR: пакет Bioconductor для анализа дифференциальной экспрессии цифровых данных экспрессии генов.Биоинформатика. 2010. 26 (1): 139–40.
CAS Статья Google ученый
 org/ScholarlyArticle»> 56.
org/ScholarlyArticle»> 56.Gotz S, Garcia-Gomez JM, Terol J, Williams TD, Nagaraj SH, Nueda MJ, Robles M, Talon M, Dopazo J, Conesa A. Высокопроизводительная функциональная аннотация и интеллектуальный анализ данных с помощью пакета Blast2GO. . Nucleic Acids Res. 2008. 36 (10): 3420–35.
CAS Статья Google ученый
Jungo F, Bougueleret L, Xenarios I, Poux S.Программа UniProtKB / Swiss-Prot Tox-Prot: центральный узел интегрированных данных о белках яда. Токсикон. 2012; 60 (4): 551–7.
CAS Статья Google ученый
Генихович Г., Технау Ю. Гибридизация зародышей, личинок и полипов звездчатого анемона ( Nematostella vectensis ) in situ. Cold Spring Harb Protoc. 2009; 2009: pdb. prot5282.
prot5282.
PubMed Google ученый
Моран Ю., Генихович Г., Гордон Д., Винкооп С., Зенкерт С., Озбек С., Технау Ю., Гуревиц М. Локализация нейротоксина в клетках эктодермальных желез открывает альтернативный механизм доставки яда у морских актиний. Proc Biol Sci. 2012. 279 (1732): 1351–8.
CAS Статья Google ученый
Кафри П., Хасенсон С.Е., Кантер И., Шейнбергер Дж., Кинор Н., Юнгер С., Шав-Таль Ю. Количественная оценка субклеточной динамики бета-катенина и транскрипции мРНК циклина D1 во время передачи сигналов Wnt в отдельных живых клетках.Элиф. 2016; 5: e16748.
Альтшул С.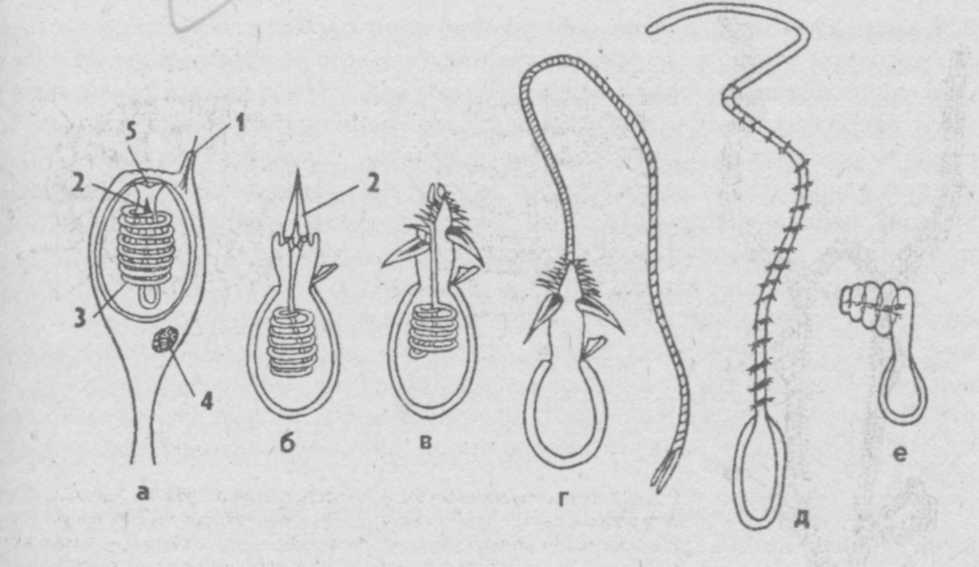 Ф., Гиш В., Миллер В., Майерс Е.В., Липман Д. Базовый инструмент поиска локального выравнивания. J Mol Biol. 1990. 215 (3): 403–10.
Ф., Гиш В., Миллер В., Майерс Е.В., Липман Д. Базовый инструмент поиска локального выравнивания. J Mol Biol. 1990. 215 (3): 403–10.
CAS Статья Google ученый
Стефаник Д.Д., Любински Т.Дж., Грейнджер Б.Р., Берд А.Л., Рейцель А.М., ДеФилиппо Л., Лоренк А., Финнерти-младший. Создание эталонного транскриптома и базы данных транскриптомов (EdwardsiellaBase) для морского анемона Edwardsiella lineata, паразитического книдария.BMC Genomics. 2014; 15:71.
Артикул Google ученый
Kitchen SA, Crowder CM, Poole AZ, Weis VM, Meyer E. De novo сборка и характеристика четырех транскриптомов антозойных (тип Cnidaria). G3 (Bethesda). 2015; 5 (11): 2441–52.
CAS Статья Google ученый
 org/ScholarlyArticle»> 64.
org/ScholarlyArticle»> 64.Guindon S, Dufayard JF, Lefort V, Anisimova M, Hordijk W., Gascuel O.Новые алгоритмы и методы для оценки филогении максимального правдоподобия: оценка производительности PhyML 3.0. Syst Biol. 2010. 59 (3): 307–21.
CAS Статья Google ученый
Жалящие клетки с лета! — Желатиновый жало
Я очень занят в Смитсоновском университете, но мне хотелось поделиться фотографиями одной из моих любимых сложных особенностей животного: жалящей клетки! Жалящие клетки содержат нематоцисты, органеллы жалящих клеток, характерные для всех книдарий (отсюда латинский перевод — крапивница).Нематоцисты представляют собой нитевидные капсулы, выделяемые жалящими клетками, которые механически или химически разряжаются через механизм высокого давления, выделяя яд. Жалящие клетки бывают более тридцати различных морфологических типов, но номенклатура меняется, и все время появляются новые определения. Но вместо того, чтобы пытаться описать чрезвычайно сложную терминологию, морфологию, физиологию и эволюционную историю жалящих клеток прямо сейчас (я мог бы потратить целую докторскую диссертацию на любую из них), я хотел показать, насколько чертовски круты эти структуры!
Эти безумные структуры представляют собой проникающие жалящие клетки, большие гетеронемы, называемые бирхропалоидами. Хотя это немного трудно увидеть, на стержне есть два отчетливых расширения или выступа. Древко выглядит как наконечник стрелы, выходящий из капсулы. Вдоль древка вы можете увидеть шипы, которые используются для зацепления цели, а чрезвычайно длинная нить доставляет яд. Я имею в виду, посмотрите на это! Я не думаю, что у меня даже половина нити на этом снимке! Когда эти жалящие клетки не разряжаются, вы можете увидеть нематоцисты, свернувшиеся внутри. В сочетании с проникающими жалящими клетками я также нахожу более мелкие и округлые клетки.Их называют изоризами, гаплонемными жалящими клетками, у которых нет стержня. Но это не значит, что эти морфологически более простые жалящие клетки не так безумно круто выглядят при выстреле! Одной из главных целей для меня этим летом было описать книдому Кассиопеи xamachana, Флоридская перевернутая медуза. Это включает изучение количества, типов и размеров жалящих клеток на разных стадиях развития (медуза, полип и т. Д.). Поскольку эти клетки имеют размер 20-200 мкм, я использую микроскоп, чтобы делать фотографии для измерений.
Хотя это немного трудно увидеть, на стержне есть два отчетливых расширения или выступа. Древко выглядит как наконечник стрелы, выходящий из капсулы. Вдоль древка вы можете увидеть шипы, которые используются для зацепления цели, а чрезвычайно длинная нить доставляет яд. Я имею в виду, посмотрите на это! Я не думаю, что у меня даже половина нити на этом снимке! Когда эти жалящие клетки не разряжаются, вы можете увидеть нематоцисты, свернувшиеся внутри. В сочетании с проникающими жалящими клетками я также нахожу более мелкие и округлые клетки.Их называют изоризами, гаплонемными жалящими клетками, у которых нет стержня. Но это не значит, что эти морфологически более простые жалящие клетки не так безумно круто выглядят при выстреле! Одной из главных целей для меня этим летом было описать книдому Кассиопеи xamachana, Флоридская перевернутая медуза. Это включает изучение количества, типов и размеров жалящих клеток на разных стадиях развития (медуза, полип и т. Д.). Поскольку эти клетки имеют размер 20-200 мкм, я использую микроскоп, чтобы делать фотографии для измерений.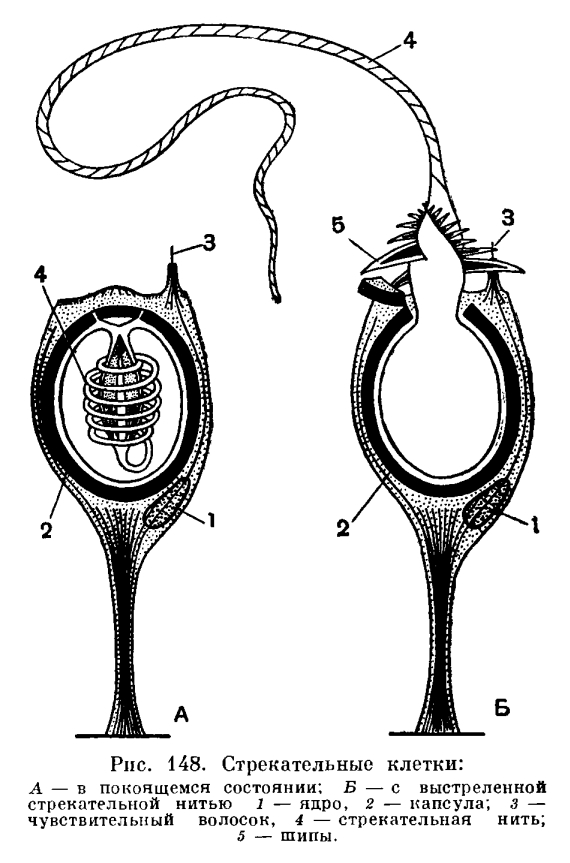
Кстати, очень мало исследований о том, как эти разные морфологические типы коррелируют с составом яда. Мы знаем, что определенные типы нематоцист (органелл жалящих клеток) и нематоцитов (жалящие клетки) важны с таксономической и функциональной точек зрения. То есть они могут рассказать нам о том, кто с кем связан эволюционно, и как определенные типы стрекательных клеток используются для определенной цели (захват добычи, защита, конкуренция, переваривание и т. Д.). Но коррелирует ли морфология с определенными разновидностями токсинов, выделяемых этими структурами? Содержат ли высокопенетрантные клетки с подобными вышеупомянутыми бихопалоидами и эврителами больше порообразующих токсинов или протеаз? Изоризы содержат больше нейротоксинов? Имеют ли стрекательные клетки, используемые для соревнования, более сильное укус, чем те, которые используются для общей защиты? Есть ли какие-нибудь стрекательные клетки, которые практически не содержат яда?
Даже когда я считаю и измеряю несколько тысяч из них, не так уж сложно понять, насколько удивительны эти структуры и животные, которые их создают.
Все изображения сделаны AK. Фотография была сделана в сотрудничестве с доктором Алленом Коллинзом (Лаборатория систематики NOAA) с использованием животных из аквариума, расположенного в отделе зоологии беспозвоночных Смитсоновского института NMNH. Этот проект был поддержан грантом Лернера Грея для морских наук, предоставленным ANMH, и финансированием аспирантуры KU.
Определения, выделенные жирным шрифтом :
Нематоцит: Жалящая клетка, содержит нематоцист
Нематоциста: Органелла жалящей клетки, общая со всеми книдариями, секретируется из аппарата Гольджи нематоцита
Книда (множественное число cnidae): Капсулы (клетки), содержащие межклеточные секреторные продукты книдаирны, общий термин, включающий книдоцисты, птихоцисты и спироцисты
Книдобласты: Клетки, из которых состоят книдобласты, общий термин, включающий книдобласты, птихобласты и спиробласты
Книдоциты: Зрелые книдобласты, общий термин, который включает нематоциты, птихоциты и спироциты; взаимозаменяемый с нематоцистой
Морфология:
Колючки: Зубцы вдоль поверхности выделившихся нематоцист
Стилетов: Большие шипы, используемые для первоначального проникновения в цель
Трубочка: Нитевидная структура, выходящая из капсулы
Вал: Увеличенная часть на конце канальца рядом с капсулой (стрекательная клетка)
Гаплонемы: Трубочка без стержня
Гетеронемы: Трубочка с четко выраженным стержнем
Упомянутые выше типы:
Birhopaloid: Два отчетливых расширения (выпуклости) вдоль вала
Eurytele: Одинарное расширение проксимального отдела вдоль стержня
Isorhiza: Трубочки округлые, одинаковой толщины; без дилатаций
Артикул:
- Бекманн, А. , & Озбек, С. (2012). Нематоциста: молекулярная карта стрекательной органеллы книдарий. Международный журнал биологии развития , 56 (6-7-8), 577-582.
- Корралес-Угальде, М., Колин, С. П., и Сазерленд, К. Р. (2017). Распределение нематоцист соответствует месту поимки добычи у гидромедуз с разными режимами хищничества. Marine Ecology Progress Series , 568 , 101-110.
- SmartEverDay. (2014). Жалящая медуза в МИКРОСКОПИЧЕСКОМ ЗАМЕДЛЕННОМ ДВИЖЕНИИ — Умнее с каждым днем 120.Получено с https://www.youtube.com/watch?v=7WJCnC5ebf4
- David, CN, Özbek, S., Adamczyk, P., Meier, S., Pauly, B., Chapman, J.,… & Гольштейн, TW (2008). Эволюция сложных структур: миниколлагены формируют книдарийную нематоцисту. Тенденции в генетике , 24 (9), 431-438.
- Фаутин, Д. Г. (2009). Структурное разнообразие, систематика и эволюция книд. Toxicon , 54 (8), 1054-1064.
- Heins, A., Glatzel, T. , & Холст, С. (2015). Пересмотренные описания нематоцист и способов бесполого размножения сцифозной медузы Cassiopea andromeda (Forskål, 1775). Зооморфология , 134 (3), 351-366.
- Касс-Саймон, Г.С.А.А., и Скаппатиччи, младший, А.А. (2002). Поведенческая физиология и физиология развития нематоцист. Канадский зоологический журнал , 80 (10), 1772-1794.
- Остман, К. (2000). Руководство по номенклатуре и классификации нематоцист, а также некоторые примечания по систематическому значению нематоцист. Scientia Marina , 64 (S1), 31-46.
 , & Озбек, С. (2012). Нематоциста: молекулярная карта стрекательной органеллы книдарий. Международный журнал биологии развития , 56 (6-7-8), 577-582.
, & Озбек, С. (2012). Нематоциста: молекулярная карта стрекательной органеллы книдарий. Международный журнал биологии развития , 56 (6-7-8), 577-582.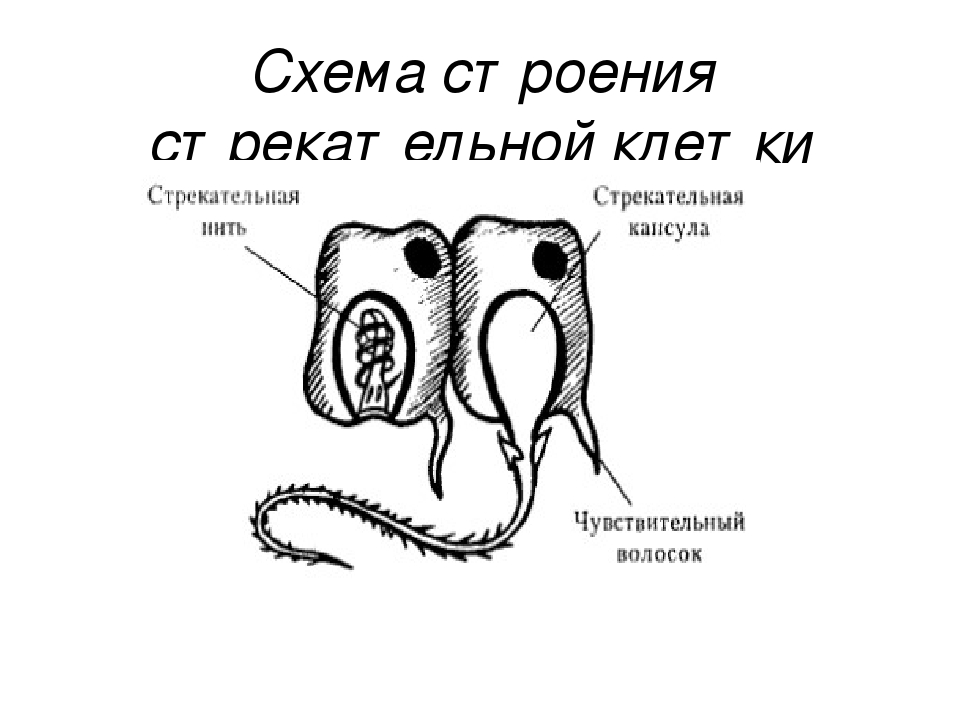 , & Холст, С. (2015). Пересмотренные описания нематоцист и способов бесполого размножения сцифозной медузы Cassiopea andromeda (Forskål, 1775). Зооморфология , 134 (3), 351-366.
, & Холст, С. (2015). Пересмотренные описания нематоцист и способов бесполого размножения сцифозной медузы Cassiopea andromeda (Forskål, 1775). Зооморфология , 134 (3), 351-366.Нравится:
Нравится Загрузка …
СвязанныеТип Cnidaria — Биология 2e
Цели обучения
К концу этого раздела вы сможете делать следующее:
- Сравните структурные и организационные характеристики Porifera и Cnidaria
- Опишите прогрессивное развитие тканей и их отношение к сложности животных
- Определите две основные формы тела, обнаруженные в Книдарии
- Опишите отличительные признаки основных классов книдарий
Тип Cnidaria включает животных, которые проявляют радиальную или бирадиальную симметрию и являются диплобластами, что означает, что они развиваются из двух эмбриональных слоев, эктодермы и энтодермы.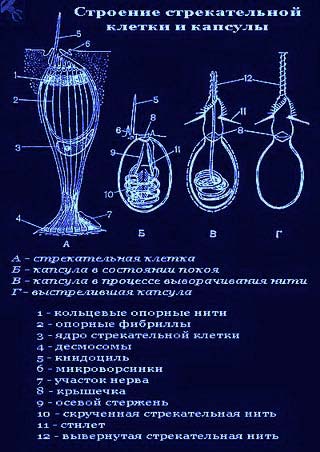 Почти все (около 99 процентов) книдарийцы — морские виды.
Почти все (около 99 процентов) книдарийцы — морские виды.
В то время как определяющим типом клеток для губок является хоаноцит, определяющим типом клеток для книдарий является книдоцит или стрекательная клетка. Эти клетки расположены вокруг рта и на щупальцах и служат для захвата добычи или отпугивания хищников. У книдоцитов есть большие жалящие органеллы, называемые нематоцистами, которые обычно содержат зазубрины в основании длинной спиральной нити. На внешней стенке клетки есть волосовидный выступ под названием книдоцил , чувствительный к тактильной стимуляции.Если прикоснуться к книдоцилам, полые нити разворачиваются с огромным ускорением, в 40 000 раз превышающим силу тяжести. Затем микроскопические нити либо запутывают добычу, либо мгновенно проникают в плоть жертвы или хищника, высвобождая токсины (включая нейротоксины и порообразующие токсины, которые могут привести к лизису клеток) в цель, тем самым обездвиживая или парализуя ее (см. )).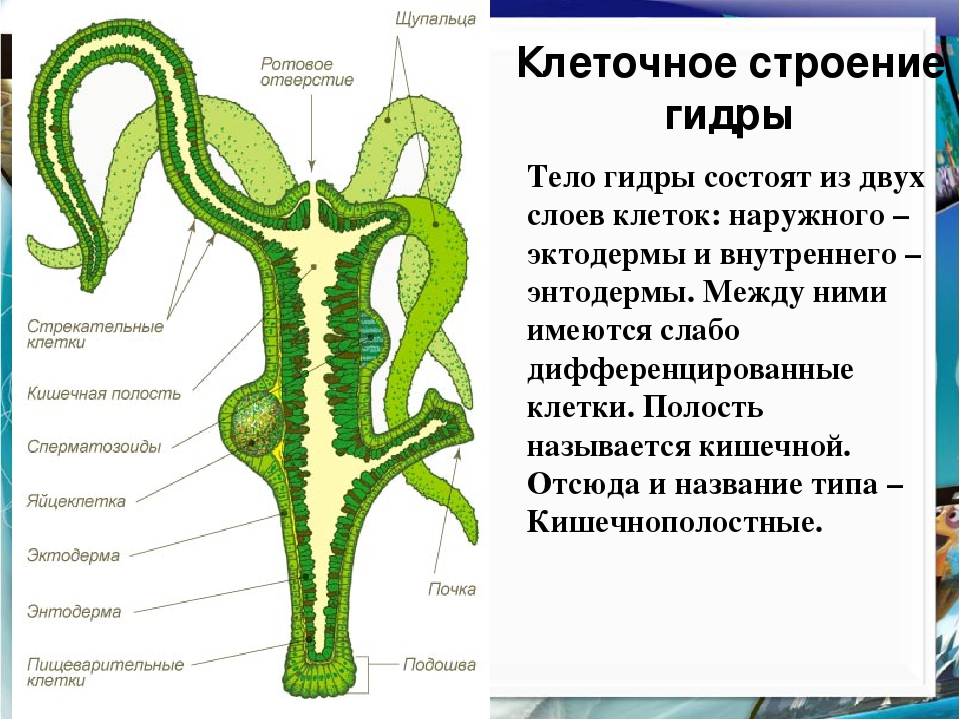
Книдоцитов. У животных из филума Cnidaria есть стрекательные клетки, называемые книдоцитами.Книдоциты содержат большие органеллы, называемые (а) нематоцистами, которые хранят спиральную нить и зазубрины, нематоцисты. При прикосновении даже легкого прикосновения к волосовидному книдоцилу на поверхности клетки (б) из органеллы выделяются нить, зазубрины и токсин.
Ссылка на обучение
Просмотрите этот анимационный видеоролик, в котором показаны два анемона, участвующие в битве.
У книдарийцев обнаружены два отчетливых плана тела: форма полипа или тюльпаноподобного «стебля» и форма медузы или «колокольчика».((Фигура)). Пример формы полипа обнаруживается в роду Hydra , тогда как наиболее типичная форма медузы встречается в группе, называемой «морские желе» (медузы). Формы полипов сидячие у взрослых особей, с единственным отверстием (ртом / анусом) в пищеварительную полость, обращенным вверх, с окружающими его щупальцами. Формы медузы подвижны, с ртом и щупальцами, свисающими с колокольчика в форме зонтика.
Формы медузы подвижны, с ртом и щупальцами, свисающими с колокольчика в форме зонтика.
Формы тела книдарий. Книдарийцы имеют два различных плана тела: медузу (а) и полип (б).У всех книдарий есть два мембранных слоя, между которыми находится желеобразная мезоглея.
Некоторые книдарии диморфны, то есть у них в течение своего жизненного цикла проявляются оба плана тела. У этих видов полип служит бесполой фазой, а медуза — половым этапом и производит гаметы. Однако обе формы тела диплоидны.
Пример диморфизма книдарий можно увидеть у колониального гидроида Obelia . Сидячая бесполая колония имеет два типа полипов, показанных на (Рисунок).Первый — гастрозооид , приспособленный для поимки добычи и кормления. В Obelia все полипы связаны через общую пищеварительную полость, называемую ценосарком . Другой тип полипа — гонозооид , адаптированный для бесполого почкования и образования половых медуз. Репродуктивные почки гонозооида отламываются и созревают в свободно плавающие медузы, которые бывают мужскими или женскими (раздельнополые). У каждой медузы есть несколько семенников или несколько яичников, в которых происходит мейоз с образованием сперматозоидов или яйцеклеток.Интересно, что продуцирующие гаметы клетки не возникают внутри самой гонады, а мигрируют в нее из тканей гонозооида. Это отдельное происхождение гонад и гамет характерно для всей эуметазоа. Гаметы попадают в окружающую воду, и после оплодотворения зигота превращается в бластулу, которая вскоре превращается в ресничную, двусторонне симметричную личинку планулы. Планула какое-то время свободно плавает, но в конце концов прикрепляется к субстрату и становится единым полипом, из которого образуется новая колония полипов путем отпочкования.
Репродуктивные почки гонозооида отламываются и созревают в свободно плавающие медузы, которые бывают мужскими или женскими (раздельнополые). У каждой медузы есть несколько семенников или несколько яичников, в которых происходит мейоз с образованием сперматозоидов или яйцеклеток.Интересно, что продуцирующие гаметы клетки не возникают внутри самой гонады, а мигрируют в нее из тканей гонозооида. Это отдельное происхождение гонад и гамет характерно для всей эуметазоа. Гаметы попадают в окружающую воду, и после оплодотворения зигота превращается в бластулу, которая вскоре превращается в ресничную, двусторонне симметричную личинку планулы. Планула какое-то время свободно плавает, но в конце концов прикрепляется к субстрату и становится единым полипом, из которого образуется новая колония полипов путем отпочкования.
Обелия . Колониальная сидячая форма Obelia geniculata имеет два типа полипов: гастрозооиды, которые приспособлены для поимки добычи, и гонозооиды, которые бесполым образом зачатки образуют медузы.
Ссылка на обучение
Щелкните здесь, чтобы следить за анимацией жизненного цикла Obelia и викториной.
Все книдарии являются диплобластами и, следовательно, имеют два «эпителиальных» слоя в теле, которые происходят из энтодермы и эктодермы эмбриона.Внешний слой (от эктодермы) называется эпидермисом и выстилает внешнюю часть животного, тогда как внутренний слой (от энтодермы) называется gastrodermis и выстилает пищеварительную полость. У личинки планулы слой эктодермы окружает твердую массу энтодермы, но по мере развития полипа пищеварительная или гастроваскулярная полость открывается внутри энтодермы. Между этими двумя слоями эпителия лежит неживая желеобразная мезоглея. Что касается клеточной сложности, книдарии показывают присутствие дифференцированных типов клеток в каждом слое ткани, таких как нервные клетки, сократительные эпителиальные клетки, клетки, секретирующие ферменты, и клетки, поглощающие питательные вещества, а также наличие межклеточных связей. Однако, за некоторыми примечательными исключениями, такими как статоцисты и rhopalia (см. Ниже), развитие органов или систем органов в этом типе не развито.
Однако, за некоторыми примечательными исключениями, такими как статоцисты и rhopalia (см. Ниже), развитие органов или систем органов в этом типе не развито.
Нервная система рудиментарна, нервные клетки организованы в сеть, разбросанную по всему телу. Эта нервная сеть может показывать присутствие групп клеток, которые образуют нервные сплетения (единственное число: сплетение) или нервные связки. Организация нервной системы подвижной медузы более сложна, чем у полипа на сидячей кости, с нервным кольцом по краю колокольчика медузы, которое контролирует работу щупалец.Книдариевые нервные клетки демонстрируют смешанные характеристики моторных и сенсорных нейронов. Преобладающими сигнальными молекулами в этих примитивных нервных системах являются пептиды, которые выполняют как возбуждающую, так и тормозную функции. Несмотря на простоту нервной системы, примечательно то, что она координирует сложные движения щупалец, притягивание захваченной добычи ко рту, переваривание пищи и выброс отходов.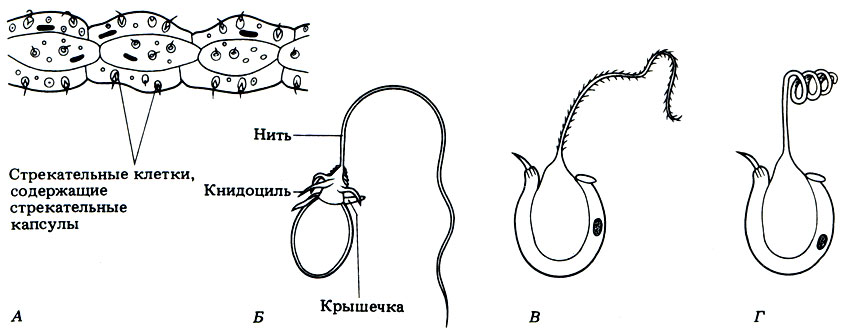
Гастроваскулярная полость имеет только одно отверстие, которое служит одновременно ртом и анусом; такое устройство называется неполноценной пищеварительной системой.В желудочно-сосудистой полости происходит внеклеточное пищеварение, когда пища поступает в желудочно-сосудистую полость, ферменты выделяются в полость, а клетки, выстилающие полость, поглощают питательные вещества. Однако также происходит некоторое внутриклеточное пищеварение. Гастроваскулярная полость распределяет питательные вещества по всему телу животного, при этом питательные вещества проходят из пищеварительной полости через мезоглею к эпидермальным клеткам. Таким образом, эта полость выполняет функции пищеварения и кровообращения.
Книдариевые клетки обменивают кислород и углекислый газ путем диффузии между клетками эпидермиса и водой в окружающей среде, а также между клетками гастродермы и водой в желудочно-сосудистой полости.Отсутствие системы кровообращения для перемещения растворенных газов ограничивает толщину стенки тела и требует наличия неживой мезоглеи между слоями. У книдарий с более толстой мезоглеей ряд каналов помогает распределять как питательные вещества, так и газы. Нет ни выделительной системы, ни органов, а азотистые отходы просто диффундируют из клеток в воду вне животного или в желудочно-сосудистую полость.
У книдарий с более толстой мезоглеей ряд каналов помогает распределять как питательные вещества, так и газы. Нет ни выделительной системы, ни органов, а азотистые отходы просто диффундируют из клеток в воду вне животного или в желудочно-сосудистую полость.
Тип Cnidaria содержит около 10 000 описанных видов, разделенных на две монофилетические клады: Anthozoa и Medusozoa.К Anthozoa относятся кораллы, морские веера, морские кнуты и морские анемоны. Medusozoa включает несколько классов Cnidaria в двух кладах: Hydrozoa включают сидячие формы, некоторые медузоидные формы и плавающие колониальные формы, такие как португальский военный корабль. Другая клада содержит различные типы студней, включая как Scyphozoa, так и Cubozoa. Anthozoa содержат только сидячие формы полипов, в то время как Medusozoa включают виды, имеющие в своем жизненном цикле как полипы, так и формы медузы.
Класс Anthozoa
Класс Anthozoa («цветочные животные») включает морских анемонов ((Рисунок)), морских загонов и кораллов, количество которых оценивается в 6 100 описанных видов.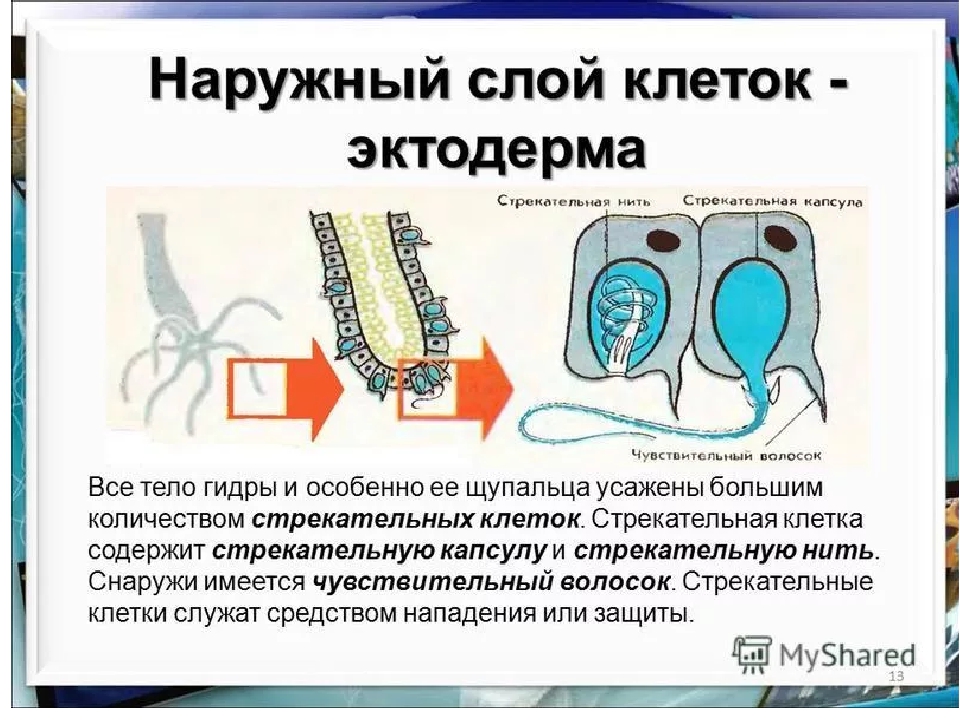 Морские анемоны обычно ярко окрашены и могут достигать размера от 1,8 до 10 см в диаметре. Отдельные животные имеют цилиндрическую форму и прикрепляются непосредственно к субстрату.
Морские анемоны обычно ярко окрашены и могут достигать размера от 1,8 до 10 см в диаметре. Отдельные животные имеют цилиндрическую форму и прикрепляются непосредственно к субстрату.
Морской анемон. Морской анемон показан (а) на фотографии и (б) на диаграмме, иллюстрирующей его морфологию. (кредит А: модификация работы «Dancing With Ghosts» / Flickr; кредит b: модификация работы NOAA)
Пасть морского анемона окружена щупальцами, несущими книдоциты.Щелевидное ротовое отверстие и уплощенный зев выстлан эктодермой. Такое строение глотки делает анемоны билатерально симметричными. Ресничная бороздка, называемая сифоноглифом, находится на двух противоположных сторонах глотки и направляет в нее воду. Глотка — это мышечная часть пищеварительной системы, которая служит как для приема пищи, так и для ее приема, и может занимать до двух третей длины тела, прежде чем открыться в желудочно-сосудистую полость. Эта полость разделена на несколько камер продольными перегородками, называемыми брыжейками .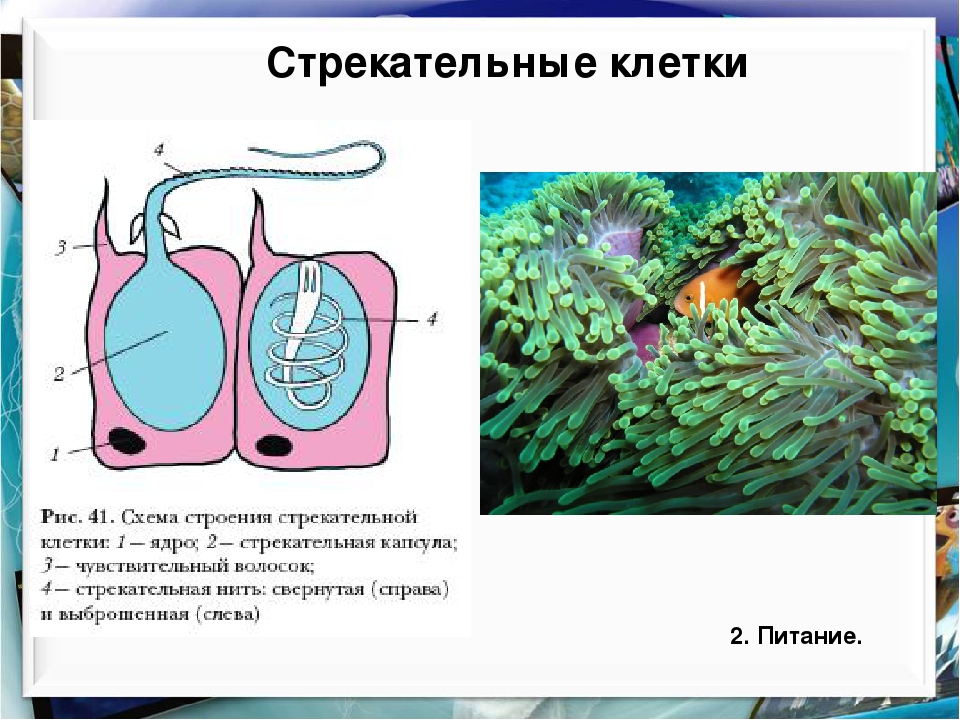 Каждая брыжейка состоит из складки гастродермальной ткани со слоем мезоглеи между листками гастродермы. Брыжейки не полностью разделяют желудочно-сосудистую полость, и более мелкие полости сливаются у глоточного отверстия. Адаптивным преимуществом брыжейки является увеличение площади поверхности для поглощения питательных веществ и газообмена, а также дополнительная механическая поддержка тела анемона.
Каждая брыжейка состоит из складки гастродермальной ткани со слоем мезоглеи между листками гастродермы. Брыжейки не полностью разделяют желудочно-сосудистую полость, и более мелкие полости сливаются у глоточного отверстия. Адаптивным преимуществом брыжейки является увеличение площади поверхности для поглощения питательных веществ и газообмена, а также дополнительная механическая поддержка тела анемона.
Морские анемоны питаются мелкой рыбой и креветками, обычно обездвиживая свою добычу нематоцистами.Некоторые морские анемоны устанавливают мутуалистические отношения с крабами-отшельниками, когда краб хватает их и прикрепляет к их панцирю. В этих отношениях анемон получает частицы пищи от добычи, пойманной крабом, а краб защищен от хищников жалящими клетками анемона. Некоторые виды анемонов или рыб-клоунов также могут жить с морскими анемонами, потому что они вырабатывают приобретенный иммунитет к токсинам, содержащимся в нематоцистах, а также выделяют защитную слизь, которая предотвращает их ужаливание.
Структура коралловых полипов аналогична структуре анемонов, хотя отдельные полипы обычно меньше по размеру и являются частью колонии, некоторые из которых массивны и размером с небольшие постройки. Коралловые полипы питаются более мелкими планктонными организмами, включая водоросли, бактерии и личинки беспозвоночных. Некоторые антозои имеют симбиотические ассоциации с динофлагеллятными водорослями, называемыми зооксантеллами. Взаимовыгодные отношения между зооксантеллами и современными кораллами, которые предоставляют водорослям убежище, придают коралловым рифам их цвет и снабжают оба организма питательными веществами.Согласно новому исследованию международной группы ученых, эта сложная мутуалистическая ассоциация началась более 210 миллионов лет назад. То, что эти симбиотические отношения возникли во время массового распространения коралловых рифов по всему миру, предполагает, что взаимосвязь водорослей и кораллов имеет решающее значение для здоровья коралловых рифов, которые обеспечивают среду обитания примерно для одной четверти всей морской жизни. Рифам угрожает тенденция к потеплению океана, из-за которой кораллы вытесняют свои зооксантеллы и становятся белыми. Этот процесс называется обесцвечиванием кораллов.
Рифам угрожает тенденция к потеплению океана, из-за которой кораллы вытесняют свои зооксантеллы и становятся белыми. Этот процесс называется обесцвечиванием кораллов.
Anthozoans остаются полиплоидными (обратите внимание, что этот термин легко спутать с «полиплоидом») на протяжении всей своей жизни и могут воспроизводиться бесполым путем за счет образования почки или фрагментации или половым путем за счет образования гамет. Мужские или женские гаметы, продуцируемые слиянием полипа, дают начало свободно плавающей личинке планулы. Личинка поселяется на подходящем субстрате и превращается в полип на сидячей основе.
Класс Scyphozoa
Класс Scyphozoa («кубковые животные») включает все (и только) морские желе, около 200 известных видов.Медуза — важная стадия жизненного цикла, хотя в жизненном цикле большинства видов есть стадия полипа. Большинство желе имеют длину от 2 до 40 см, но самый крупный сцифозный вид, Cyanea capillata , может достигать размера двух метров в диаметре. Сцифозы имеют характерную колоколообразную морфологию ((рисунок)).
Сцифозы имеют характерную колоколообразную морфологию ((рисунок)).
Морское желе. Желе показано (а) на фотографии и (б) на диаграмме, иллюстрирующей его морфологию. (кредит а: модификация работы «Jimg944 ″ / Flickr; кредит б: модификация работы Марианы Руис Вильярреал)
В морском желе на нижней стороне животного имеется ротовое отверстие, окруженное полыми щупальцами, несущими нематоцисты.Сцифозы большую часть своего жизненного цикла живут как свободно плавающие, одинокие хищники. Рот ведет к гастроваскулярной полости, которую можно разделить на четыре взаимосвязанных мешочка, называемых дивертикулами , . У некоторых видов пищеварительная система может далее разветвляться на радиальных каналов . Подобно перегородкам у антозоязычных, разветвленные гастроваскулярные клетки выполняют две функции: увеличивают площадь поверхности для абсорбции и диффузии питательных веществ и поддерживают тело животного.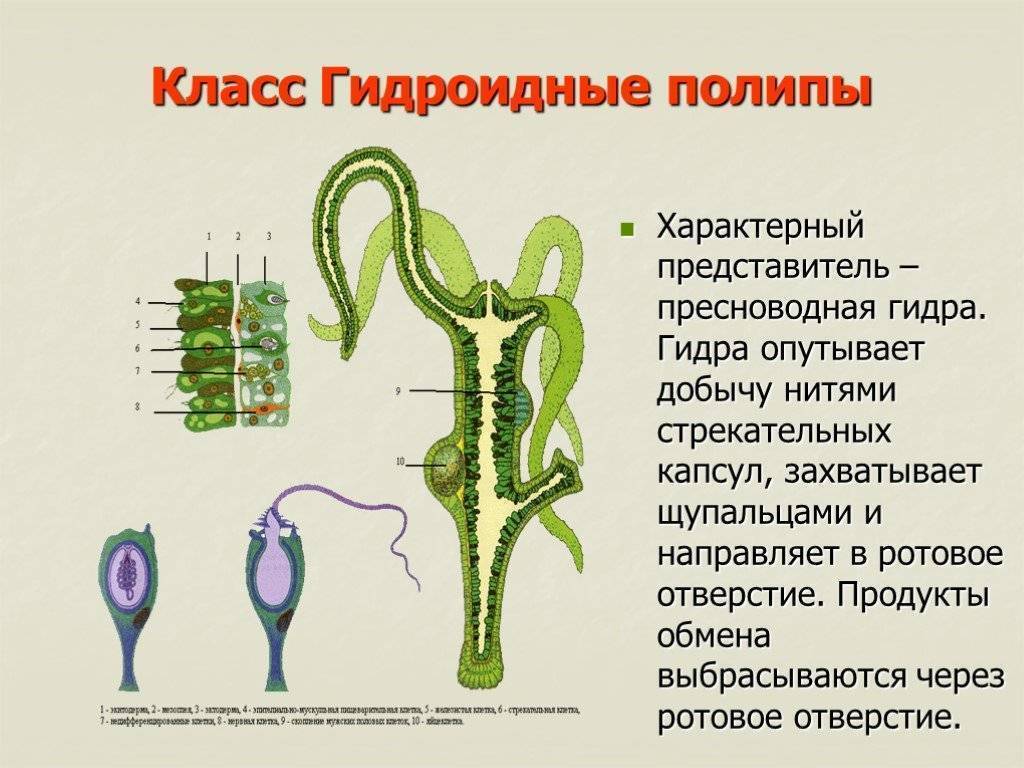
У сцифозных нервные клетки организованы в нервную сеть, которая простирается по всему телу, с нервным кольцом по краю колокола.В карманах по краю колокола могут присутствовать скопления органов чувств, называемые ропалией. У желе есть кольцо из мышц, выстилающее купол тела, которое обеспечивает сократительную силу, необходимую для плавания в воде, а также для втягивания пищи из воды во время плавания. Сцифозы имеют отдельные полы. Гонады образуются из гастродермы, а гаметы выводятся через рот. Личинки Planula образуются путем внешнего оплодотворения; они располагаются на субстрате в полиповидной форме.Эти полипы могут образовывать дополнительные полипы или сразу же начать производить бутоны медузы. У некоторых видов личинка планулы может развиваться непосредственно в медузу. Жизненный цикл ((Рисунок)) большинства сцифозов включает как половые медузоидные, так и бесполые полиповидные формы тела.
Жизненный цикл скифозов. Жизненный цикл большинства медуз состоит из двух стадий: стадии медузы и стадии полипа.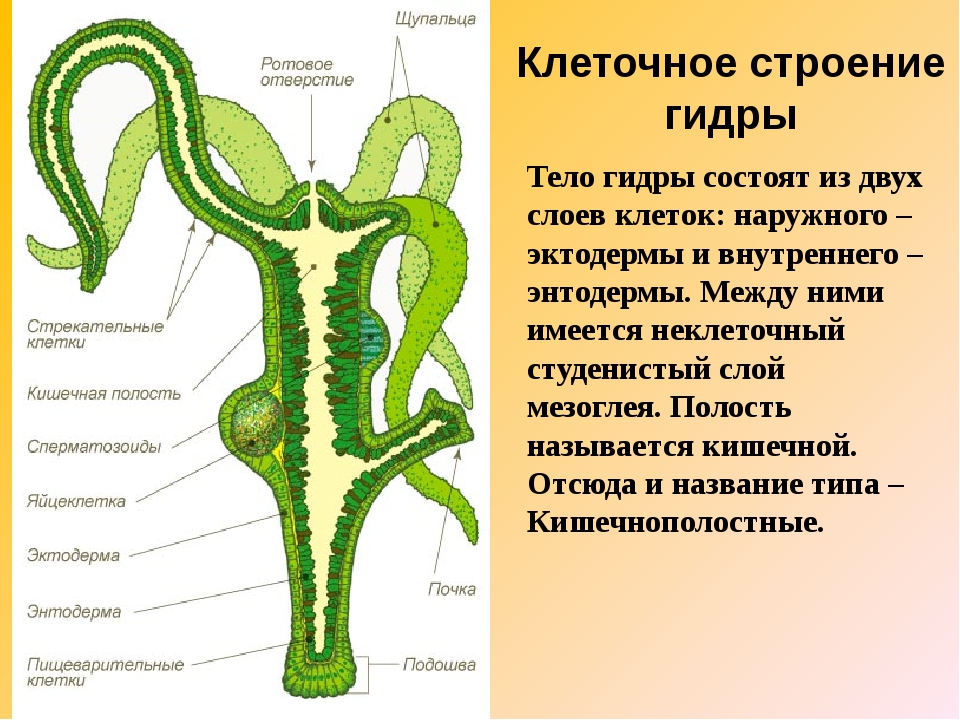 Полип размножается бесполым путем бутонизации, а медуза — половым путем. (кредит «Медуза»: модификация работы Франческо Криппы)
Полип размножается бесполым путем бутонизации, а медуза — половым путем. (кредит «Медуза»: модификация работы Франческо Криппы)
Класс Cubozoa
К этому классу относятся желе с коробчатой медузой или квадратным в поперечном сечении колокольчиком, которые в просторечии известны как «коробчатые медузы».Эти виды могут достигать размеров от 15 до 25 см, но, как правило, представители Cubozoa не такие большие, как Scyphozoa. Однако общие морфологические и анатомические характеристики кубозойных животных сходны с таковыми у сцифозов. Заметное различие между этими двумя классами — расположение щупалец. Cubozoans содержат мускулистые подушечки, называемые педалиями, по углам квадратного купола колокола, с одним или несколькими щупальцами, прикрепленными к каждой педали. В некоторых случаях пищеварительная система может распространяться на педалию.Нематоцисты могут располагаться по спирали вдоль щупалец; такое расположение помогает эффективно подчинять и ловить добычу.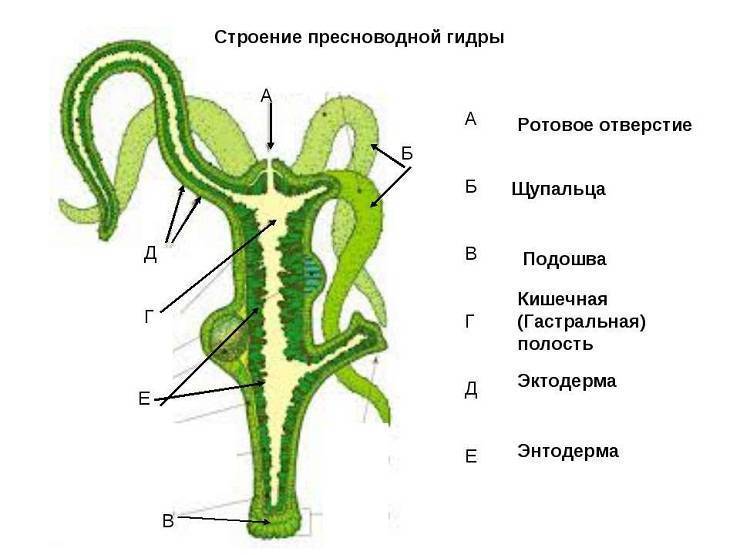 Кубозойские животные — самые ядовитые из всех книдарий ((Рисунок)).
Кубозойские животные — самые ядовитые из всех книдарий ((Рисунок)).
Эти животные необычны тем, что имеют глаза, формирующие изображение, включая роговицу, хрусталик и сетчатку. Поскольку эти структуры состоят из ряда взаимодействующих тканей, их можно назвать настоящими органами . Глаза расположены четырьмя кластерами между каждой парой педалей. Каждый кластер состоит из четырех простых глазных пятен и двух формирующих изображение глаз, ориентированных в разных направлениях.Как обрабатываются изображения, сформированные этими очень сложными глазами, остается загадкой, поскольку у кубозояновых есть обширные нервные сети, но нет отчетливого мозга. Тем не менее наличие глаз помогает кубозоям быть активными и эффективными охотниками на мелких морских животных, таких как черви, членистоногие и рыбы.
Кубозойские имеют разные полы, оплодотворение происходит внутри самки. Личинки Planula могут развиваться внутри самки или выходить наружу, в зависимости от вида. Каждая планула превращается в полип. Эти полипы могут образовывать бутоны, чтобы образовать больше полипов, чтобы создать колонию; затем каждый полип превращается в единственную медузу.
Каждая планула превращается в полип. Эти полипы могут образовывать бутоны, чтобы образовать больше полипов, чтобы создать колонию; затем каждый полип превращается в единственную медузу.
Кубозойный. (А) крошечный кубозойный желе Malo kingi имеет форму наперстка и, как все кубозойные желе, (б) имеет четыре мускулистые ножки, к которым прикрепляются щупальца. M. kingi — один из двух видов желе, которые, как известно, вызывают синдром Ируканджи, состояние, характеризующееся мучительной мышечной болью, рвотой, учащенным пульсом и психологическими симптомами. Считается, что два человека в Австралии, где чаще всего встречаются желе Ируканджи, умерли от укусов Ируканджи.(c) Знак на пляже в северной Австралии предупреждает пловцов об опасности. (кредит c: модификация работы Питера Шанкса)
Класс Hydrozoa
Hydrozoa — разнообразная группа, включающая около 3200 видов; большинство из них морские, хотя известны некоторые пресноводные виды ((Рисунок)).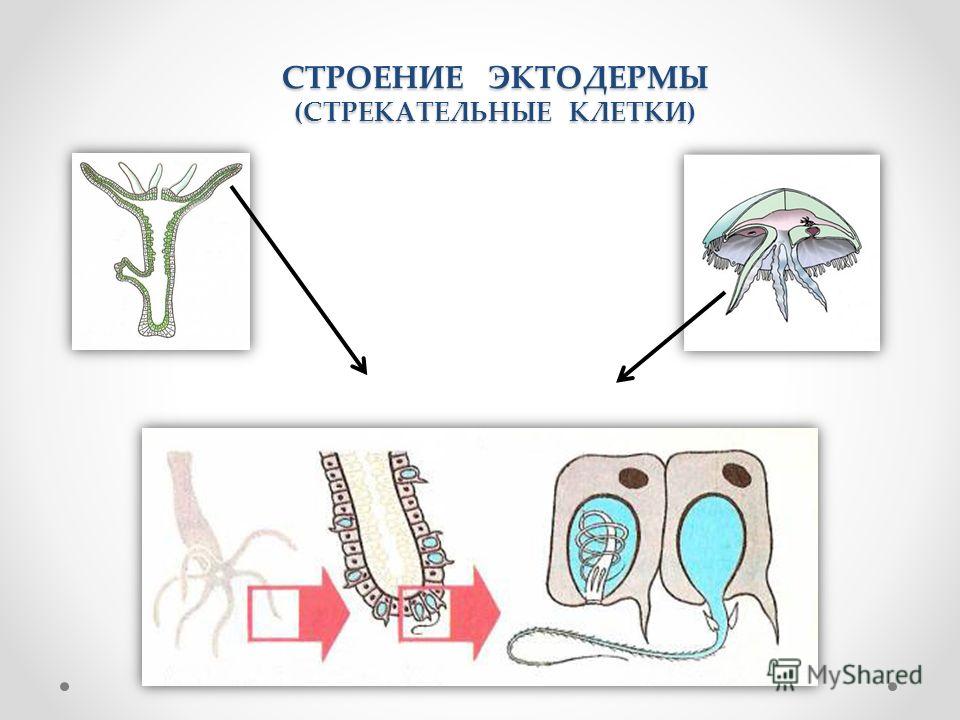 Большинство видов в своем жизненном цикле демонстрируют как полиповидные, так и медузоидные формы, хотя знакомая Гидра имеет только полиповидную форму. Медузоидная форма имеет мышечную вуаль или велум под краем колокола, поэтому ее называют гидромедузой .Напротив, медузоидная форма Scyphozoa лишена велума и называется сцифомедузой .
Большинство видов в своем жизненном цикле демонстрируют как полиповидные, так и медузоидные формы, хотя знакомая Гидра имеет только полиповидную форму. Медузоидная форма имеет мышечную вуаль или велум под краем колокола, поэтому ее называют гидромедузой .Напротив, медузоидная форма Scyphozoa лишена велума и называется сцифомедузой .
Форма полипа у этих животных часто имеет цилиндрическую морфологию с центральной желудочно-сосудистой полостью, выстланной гастродермой. Гастродерма и эпидермис имеют простой слой мезоглеи, зажатый между ними. На ротовом конце животного имеется ротовое отверстие, окруженное щупальцами. Многие гидрозоиды образуют сидячие разветвленные колонии специализированных полипов, которые имеют общую ветвящуюся гастроваскулярную полость (ценосарк), такую как обнаруженный у колониального гидроида Obelia .
Свободно плавающие колониальные виды, называемые сифонофорами, содержат как медузоидных, так и полиповидных особей, которые специализируются на питании, защите или размножении.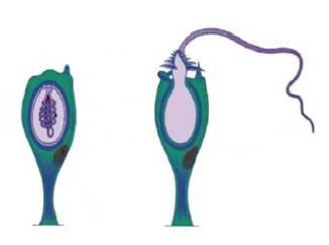 Характерный радужный поплавок португальского военного корабля ( Physalia Physalis ) создает пневматофор, с помощью которого он регулирует плавучесть, наполняя и удаляя угарный газ. На первый взгляд эти сложные суперорганизмы кажутся единым организмом; но на самом деле даже щупальца состоят из зооидов, наполненных нематоцистами.Таким образом, хотя он внешне напоминает типичную медузо-средиземноморскую медузу, P. Physalis представляет собой свободно плавающую колонию гидрозоидов ; Каждый экземпляр состоит из многих сотен организмов, каждый из которых специализируется на определенной функции, включая подвижность и плавучесть, питание, воспроизводство и защиту. Хотя они плотоядны и питаются многими морскими животными с мягким телом, у P. Physalis нет желудков, а вместо них есть специализированные полипы, называемые гастрозооидами, которые они используют для переваривания своей добычи в открытой воде.
Характерный радужный поплавок португальского военного корабля ( Physalia Physalis ) создает пневматофор, с помощью которого он регулирует плавучесть, наполняя и удаляя угарный газ. На первый взгляд эти сложные суперорганизмы кажутся единым организмом; но на самом деле даже щупальца состоят из зооидов, наполненных нематоцистами.Таким образом, хотя он внешне напоминает типичную медузо-средиземноморскую медузу, P. Physalis представляет собой свободно плавающую колонию гидрозоидов ; Каждый экземпляр состоит из многих сотен организмов, каждый из которых специализируется на определенной функции, включая подвижность и плавучесть, питание, воспроизводство и защиту. Хотя они плотоядны и питаются многими морскими животными с мягким телом, у P. Physalis нет желудков, а вместо них есть специализированные полипы, называемые гастрозооидами, которые они используют для переваривания своей добычи в открытой воде.
Physalia имеет мужские и женские колонии, которые выпускают свои гаметы в воду. Зигота развивается в отдельную особь, которая затем бесполым путем распускается, образуя новую колонию. Сифонофоры включают крупнейшие известные плавающие колонии книдарий, такие как Praya dubia , чья цепочка зоидов может достигать 50 метров (165 футов) в длину. Другими видами водорослей являются одиночные полипы ( Hydra ) или одиночные гидромедузы ( Gonionemus ). Одной определяющей характеристикой гидрозояновых является то, что их гонады происходят из эпидермальной ткани, тогда как у всех других книдарий они происходят из гастродермальной ткани.
Hydrozoans. Колония полипов Obelia (a), сифонофорные колонии Physalia (b) Physalis , известные как португальский военный, и Velella bae (c), и одиночный полип Hydra (d) имеют разные формы тела, но все принадлежат к семейству Hydrozoa. (кредит b: модификация работы NOAA; данные шкалы от Мэтта Рассела)
(кредит b: модификация работы NOAA; данные шкалы от Мэтта Рассела)
Сводка раздела
Книдарийцы представляют собой более сложный уровень организации, чем Porifera.Они обладают внешним и внутренним тканевыми слоями, между которыми находится неклеточная мезоглея. Книдарианцы обладают хорошо сформированной пищеварительной системой и осуществляют внеклеточное пищеварение в пищеварительной полости, которая проходит через большую часть животного. Рот окружен щупальцами, которые содержат большое количество книдоцитов — специализированных клеток, несущих нематоцисты, используемых для ужаливания и захвата добычи, а также отпугивания хищников. Книдарии имеют разные полы, и у многих из них есть жизненный цикл, который включает две различные морфологические формы — медузоидную и полиповидную — на разных стадиях их жизненных циклов.У видов с обеими формами медуза — это половая стадия, производящая гаметы, а полип — бесполая стадия. Виды книдарий включают отдельные или колониальные полиповидные формы, плавающие колонии или большие отдельные формы медуз (морские желе).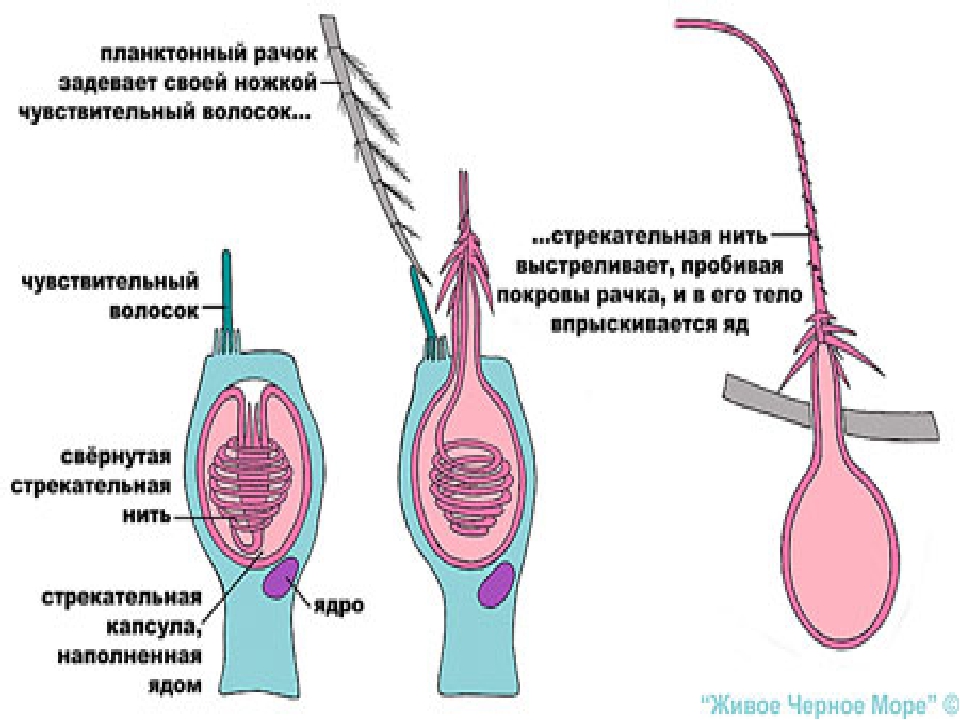
Обзорные вопросы
Книдоцитов обнаружено в _____.
- тип Porifera
- тип Nemertea
- тип Nematoda
- тип Cnidaria
Кубозоянов ________.
- полипы
- медузоидов
- полиморфов
- губки
Во время сбора образцов морской биолог находит сидящего книдария. Медузы, выросшие из него, плывут, сокращая кольцо мускулов в своих колокольчиках. К какому классу принадлежит этот экземпляр?
- Класс Hydrozoa
- Класс Cubozoa
- Класс Scyphozoa
- Класс Anthozoa
Вопросы о критическом мышлении
Объясните функцию нематоцист у книдарий.
Нематоцисты — это «жалящие клетки», предназначенные для парализации добычи. Нематоцисты содержат нейротоксин, который делает добычу неподвижной.
Сравните структурные различия между Porifera и Cnidaria.
Poriferans не обладают настоящими тканями, в то время как книдарии имеют ткани. Из-за этой разницы у poriferans нет нервной системы или мускулов для передвижения, которые есть у книдарий.
Сравните различия в половом размножении между Porifera и Cubozoans.Как разница в оплодотворении дает кубозойцам эволюционное преимущество?
Есть два основных различия между Porifera (губки) и Cubozoans (коробчатая медуза) — производство гамет и стратегия оплодотворения. Коробчатые медузы имеют разные полы, а одна губка может производить оба типа гамет. Коробчатые медузы также подвергаются внутреннему оплодотворению, в то время как губки размножаются путем внешнего оплодотворения. Внутреннее оплодотворение позволяет медузам контролировать, какие сперматозоиды используются для оплодотворения, и увеличивает вероятность встречи яйцеклеток и сперматозоидов.
Глоссарий
- Книдария
- Тип животных, диплобластных и обладающих радиальной симметрией
- книдоцит
- В Книдарии найдена специализированная стрекательная клетка
- эпидермис
- наружный слой (из эктодермы), выстилающий внешнюю часть животного
- внеклеточное пищеварение
- Пища попадает в желудочно-сосудистую полость, ферменты выделяются в полость, а клетки, выстилающие полость, поглощают питательные вещества
- гастродермис
- внутренний слой (от энтодермы), выстилающий пищеварительную полость
- желудочно-сосудистая полость
- отверстие, служащее одновременно ртом и анусом, называемое неполной пищеварительной системой
- медуза
- Свободно плавающий план тела книдарий с ртом на нижней стороне и щупальцами, свисающими с колокола
- мезоглея
- неживой гелеподобный матрикс между эктодермой и энтодермой у книдарий
- нематоциста
- гарпуноподобная органелла внутри книдоцита с заостренным снарядом и ядом для оглушения и запутывания добычи
- полип
- стеблеобразная сидячая форма жизни книдарий со ртом и щупальцами, обращенными вверх, обычно сидячая, но может скользить по поверхности
- полиморфный
- обладают множественными формами тела в течение жизненного цикла группы организмов
- сифонофор
- трубчатая конструкция, служащая для входа воды в полость мантии
Жалящие клетки Хотя книдарии сильно различаются по внешнему виду,
у них действительно есть общие характеристики, которые отделяют их от других групп. Жалящая клетка до и после выброса. Кисть — это «спусковой крючок» Это заставляет стрекательную клетку взорваться и выбросить зазубренную нить.
|
 Общей характеристикой, которая дала название этой группе, является ее жгучая
клетки (книдобласты). При прикосновении они могут выпустить колючую нить, которая
связан с ядовитым мешком.
Общей характеристикой, которая дала название этой группе, является ее жгучая
клетки (книдобласты). При прикосновении они могут выпустить колючую нить, которая
связан с ядовитым мешком. 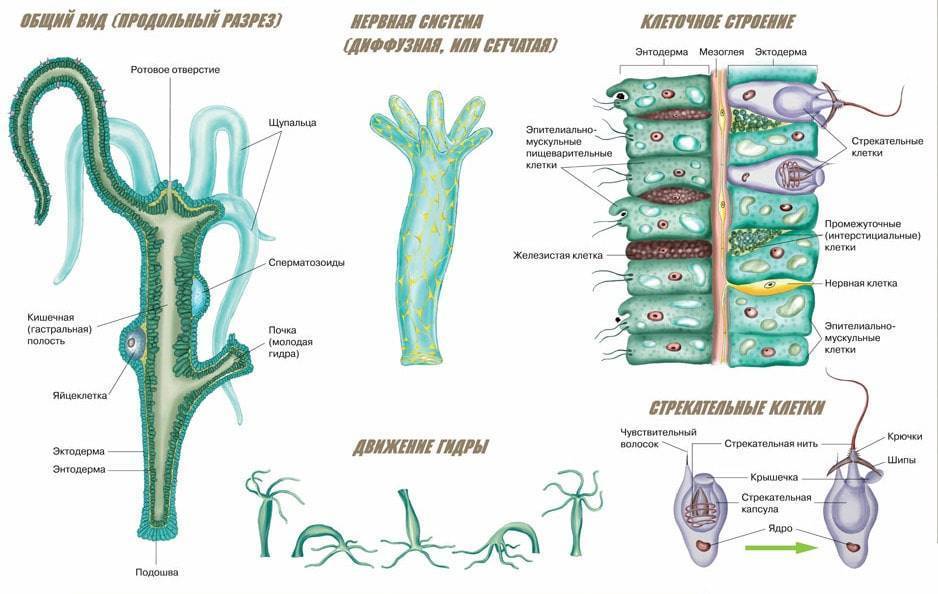 Вдоль пляжей, например, Австралия, знаки
часто публикуются предупреждения для особого типа книдариев, поле Медуза .Травмы коробчатой медузы могут быть смертельными при наличии медицинского
внимание не привлекается вовремя. В шведских водах только жалящая
медузы могут вызывать дискомфорт.
Вдоль пляжей, например, Австралия, знаки
часто публикуются предупреждения для особого типа книдариев, поле Медуза .Травмы коробчатой медузы могут быть смертельными при наличии медицинского
внимание не привлекается вовремя. В шведских водах только жалящая
медузы могут вызывать дискомфорт.Как морские слизни крадут защиту своей добычи
Не все слизни (улитки без раковин) — слизистые коричневые вредители, встречающиеся в вашем саду на заднем дворе.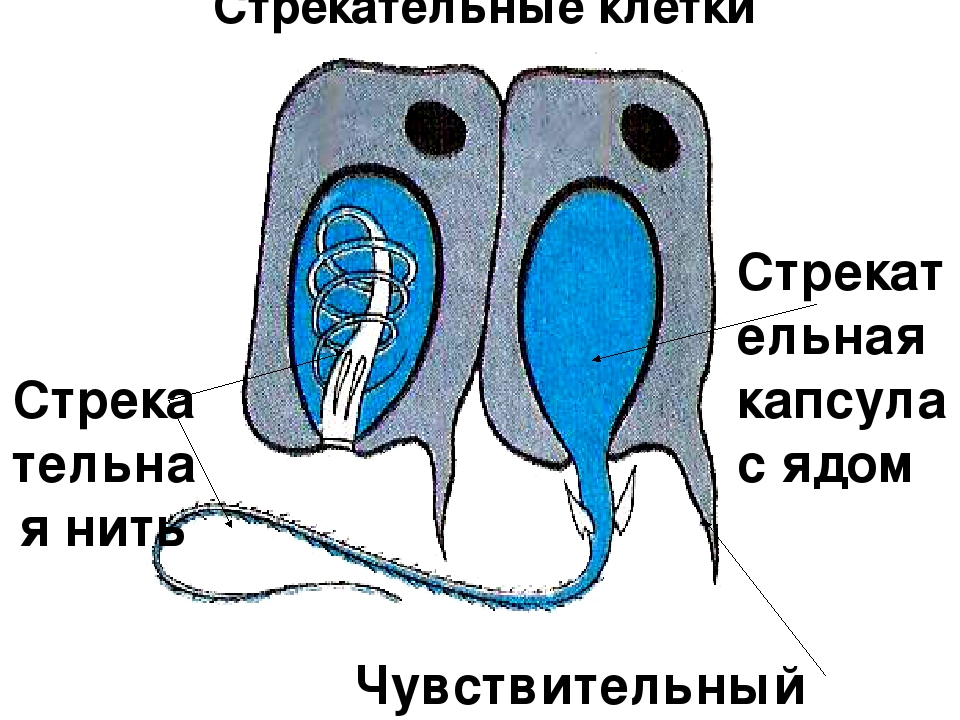 В океане они бывают самых разных цветов — некоторые соответствуют фону и их трудно заметить, но другие очень заметны. В частности, голожаберники особенно популярны среди дайверов и подводных фотографов из-за их часто ярких и красивых цветовых узоров. Однако окраска полезна не только для красивой фотографии. Яркие цвета предупреждают хищников о том, что эти голожаберники будут плохой пищей, потому что они вооружены токсинами и другими средствами защиты.
В океане они бывают самых разных цветов — некоторые соответствуют фону и их трудно заметить, но другие очень заметны. В частности, голожаберники особенно популярны среди дайверов и подводных фотографов из-за их часто ярких и красивых цветовых узоров. Однако окраска полезна не только для красивой фотографии. Яркие цвета предупреждают хищников о том, что эти голожаберники будут плохой пищей, потому что они вооружены токсинами и другими средствами защиты.
Откуда взялась эта защита? Некоторые виды в группе голожаберников, называемых эолидами (произносится как э-о-веки), крадут защиту у жалящих животных, которыми они питаются! Медузы, морские анемоны, кораллы и их родственники из типа Cnidaria захватывают пищу специальными стрекательными клетками, называемыми книдоцитами, которые выстилают их щупальца.Эти наполненные ядом клетки выделяют крошечные гарпуноподобные структуры (называемые нематоцистами), которые используются как для захвата добычи, так и для защиты от потенциальных хищников. Нематоцисты чрезвычайно эффективны в сдерживании большинства хищников книдарий, но некоторые хищники, такие как эолидные морские слизни, способны не только победить ядовитое оружие, но и использовать эту защиту в своих интересах.
Нематоцисты чрезвычайно эффективны в сдерживании большинства хищников книдарий, но некоторые хищники, такие как эолидные морские слизни, способны не только победить ядовитое оружие, но и использовать эту защиту в своих интересах.
Когда эолиды питаются этой хорошо вооруженной добычей, первое, что им нужно сделать, — это предотвратить стрельбу жалящих клеток.Похоже, они делают это, используя слизь: определенные химические вещества в слизистом следе защищают их от ужаливания. После проглатывания необожженные нематоцисты проходят через пищеварительный тракт голожаберника. Некоторые из них выводятся с остальными отходами голожаберника, но незрелые жалящие нематоцисты хранятся в мешочках, называемых книдосаками, до тех пор, пока они не созреют и не станут необходимыми.
Эолидные слизни при угрозе стреляют в украденные стрекательные клетки, а нематоцисты кусают и повреждают животных, которые пытаются съесть слизней.Как исследователь из Смитсоновского института и Университета Мэриленда, я изучаю эволюционный процесс, который позволил голожаберникам красть и хранить книдарийные нематоцисты.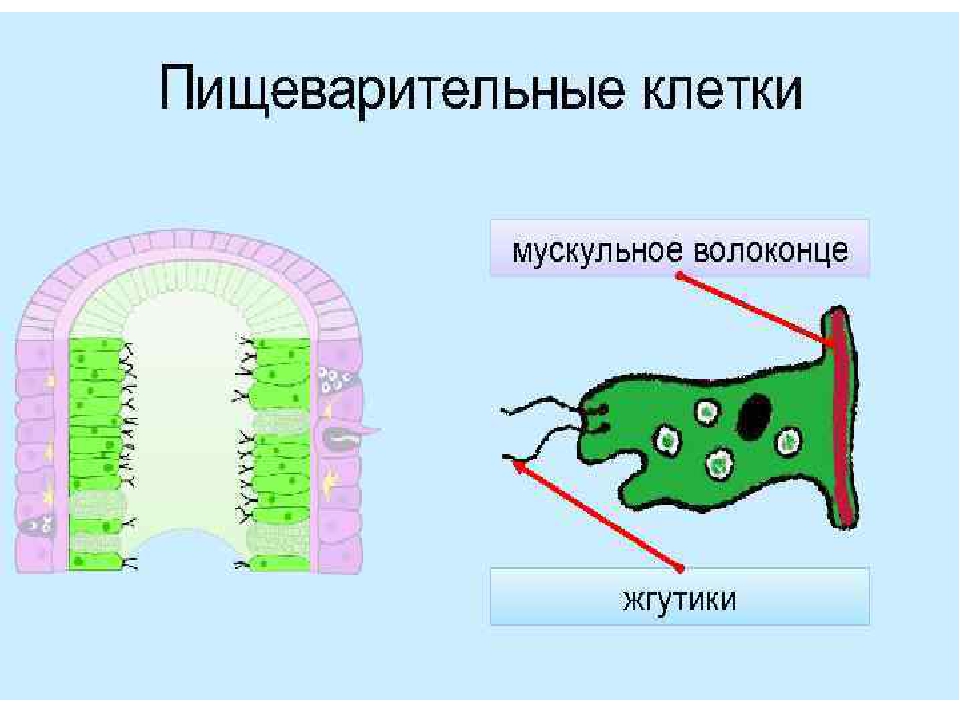 Как именно предкам этих слизней удалось использовать защиту своей жертвы для собственных нужд, до сих пор остается загадкой. Изучение истории эволюции этих разноцветных морских слизней покажет нам, как эолиды приобрели эту функцию и откуда она взялась.
Как именно предкам этих слизней удалось использовать защиту своей жертвы для собственных нужд, до сих пор остается загадкой. Изучение истории эволюции этих разноцветных морских слизней покажет нам, как эолиды приобрели эту функцию и откуда она взялась.
молекулярная карта жалящей органеллы книдарий
Анализ Hydra nematocyst 5
выявил большое количество дополнительных белков лектинового типа
в качестве компонентов капсулы (Balasubramanian et al., 2012).
Таким образом, мы предполагаем, что белок-углеводные взаимодействия
играют основную роль в сборке капсулы.
Заключение
Структурный состав книдариевой нематоцисты демонстрирует
явных сходств с внеклеточным матриксом многоклеточных животных. Миникол-
лагенов, лектинов и глизоминогликанов являются яркими примерами, а
играют важную роль в морфогенезе нематоцист. Таким образом, мы постулируем эволюционный сценарий, при котором клетки, секретирующие растворимые
токсинов, приобретают компоненты ECM для более эффективной доставки и
, вероятно, для хранения токсинов.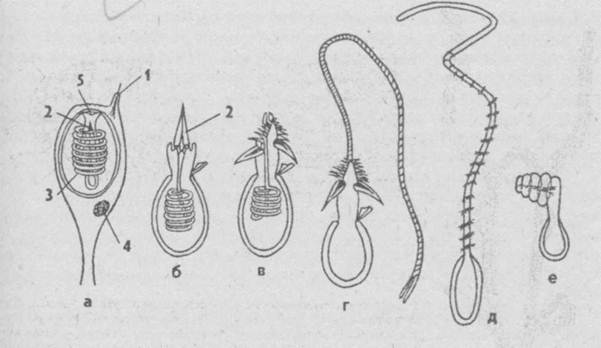 Кроме того, многие белковые токсины, такие как токсичные лектины
Кроме того, многие белковые токсины, такие как токсичные лектины
, связаны с молекулами ECM (Eble, 2010) и могут иметь
вклад в структурную часть протеома нематоцисты. Детальное сравнение
протеомов нематоцист различных видов книдарий
поможет в определении эволюционной истории этой органеллы
. Наши протеомные данные ясно указывают на консервативный набор растворимых, связанных с токсинами генов
, к которым были добавлены структурные белки
в видоспецифических вариациях (Balasubramanian et
al., в доработке).
Благодарности
Мы благодарим Бриджит Галлиот за многие полезные предложения и критическое чтение.
ing. Наша работа поддерживается грантами DFG S.Ö.
Ссылки
ADAMCZYK, P., MEIER, S., GROSS, T., HOBMAYER, B., GRZESIEK, S., BACH-
INGER, H.P., HOLSTEIN, T.W., OZBEK, S. (2008). Миниколлаген-15, новый миниколлаген
, выделенный из гидры, образует структуры канальцев в нематоцистах.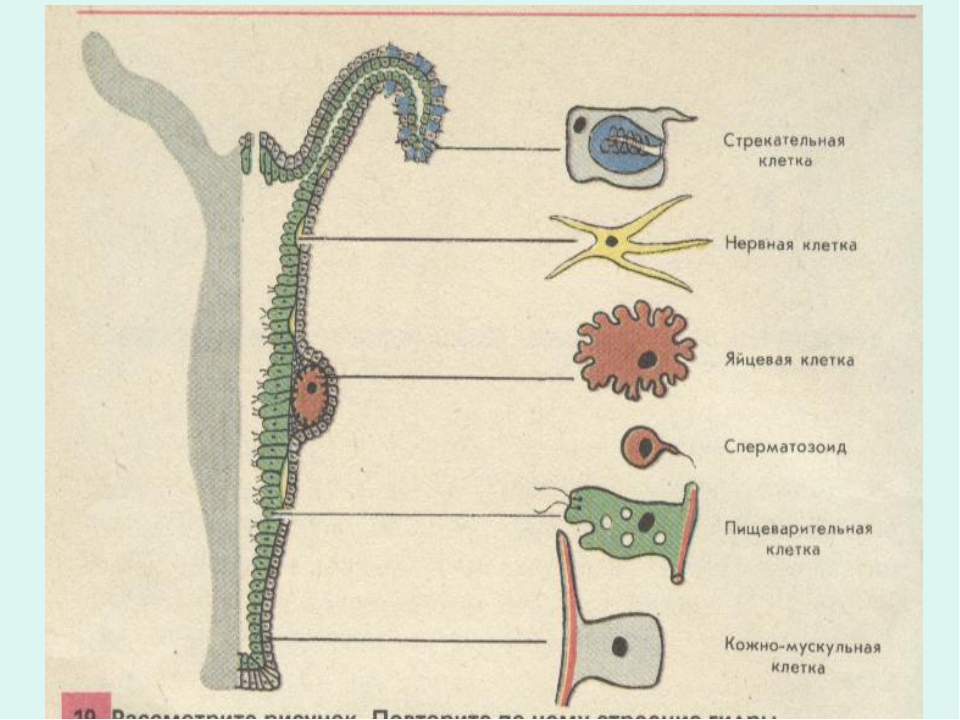 Дж. Мол
Дж. Мол
Биол 376: 1008-1020.
ADAMCZYK, P., ZENKERT, C., BALASUBRAMANIAN, PG, YAMADA, S., MURA-
KOSHI, S., SUGAHARA, K., HWANG, JS, GOJOBORI, T., HOLSTEIN, TW,
ОЗБЕК, С. (2010). Несульфатированный хондроитин стабилизирует мембранные канальцы в
органеллах книдария. J Biol Chem 285: 25613-25623.
БАЛАСУБРАМАНЯН П.Г., БЕКМАНН А., ВАРНКЕН У., ШНОЛЬЦЕР М.,
ШУЕЛЕР А., БОРНБЕРГ-БАУЭР Э., ХОЛСТЕЙН Т.В., ОЗБЕК С. (2012).
Протеом нематоцисты гидры.J Biol Chem. 287: 9672-9681. (DOI:
10.1074 / jbc.M111.328203).
BODE, H.R., DAVID, C.N. (1978). Регуляция мультипотентной стволовой клетки, интерстициальной клетки
гидры. Prog Biophys Mol Biol 33: 189-206.
БЁТТГЕР, А. и ХАССЕЛЬ, М. (2012). Hydra, модельная система для отслеживания появления границ у эвметазо. Int J Dev Biol DOI: 10.1387 / ijdb.113454ab
BOSCH, T.C., DAVID, C.N. (1991). Принятие решения в интерстициальных стволовых клетках гидры.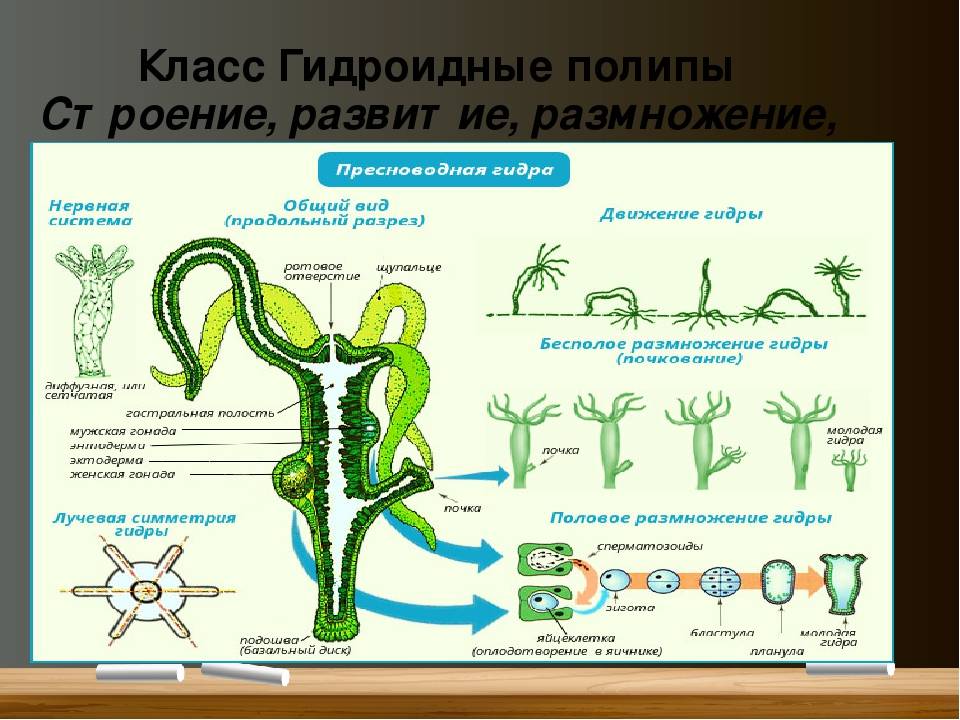
In vivo 5: 515-520.
КЭМПБЕЛЛ, Р.Д., МАРКУМ, Б.А. (1980). Миграция нематоцитов в гидре: доказательства
для руководства по контакту in vivo. J Cell Sci 41: 33-51.
CELIO, M.R., SPREAFICO, R., DE BIASI, S., VITELLARO-ZUCCARELLO, L. (1998).
Перинейрональные сети: прошлое и настоящее. Trends Neurosci 21: 510-515.
ДЭВИД, C.N. (в прессе). Интерстициальные стволовые клетки гидры: мультипотентность и принятие решения —
. Int J Dev Biol DOI: 10.1387 / ijdb.113476cd
ДЭВИД, C.N., CHALLONER, D. (1974). Распределение интерстициальных клеток и дифференцировка
нематоцист в гнездах у Hydra Аттенуата. Am Zool 14: 537-542.
ДЭВИД, C.N., GIERER, A. (1974). Кинетика клеточного цикла и развитие Hydra at-
tenuata. III. Дифференцировка нервов и нематоцитов. J Cell Sci 16: 359-75.
ДЭВИД, C.N., OZBEK, S., ADAMCZYK, P., MEIER, S., PAULY, B., CHAPMAN, J.,
HWANG, J.S., GOJOBORI, T., Холстейн, Т. (2008). Эволюция сложных структур —
Эволюция сложных структур —
структур: миниколлагены формируют книдарийную нематоцисту. Trends Genet 24: 431-438.
EBLE, J.A. (2010). Матричная биология встречается с токсинологией. Матрица Биол 29: 239-247.
ENGEL, U., OZBEK, S., ENGEL, R., PETRI, B., LOTTSPEICH, F., HOLSTEIN, T.W.
(2002). NOWA, новый белок с миниколлагеновыми Cys-богатыми доменами, участвующий в образовании нематоцист
у Hydra. J Cell Sci 115: 3923-3934.
ENGEL, U., ПЕРЦ, О., ФАУЗЕР, К., ЭНГЕЛЬ, Дж., ДЭВИД, К.Н., ХОЛСТЕЙН, Т.
(2001). Переключение дисульфидной связи во время сборки миниколлагена в матоцистах Hydra ne-
. EMBO J 20: 3063-3073.
Фаутин Д.Г. (1988). Важность нематоцист в актиниевой систематике, в:
Биология нематоцист. HESSINGER, D.A., LENHOFF, H.M., Academic Press,
, Лондон, стр. 487-500.
FAWCETT, D.W., ITO, S., SLAUTTERBACK, D. (1959). Возникновение межклеточных мостиков
в группах клеток, дифференцированных синхронно. J Biophys
J Biophys
Biochem Cytol 5: 453-460.
FUJISAWA, T., DAVID, C.N. (1981). Приверженность во время дифференцировки нематоцитов
в Hydra. J Cell Sci 48: 207-222.
FUJISAWA, T., DAVID, C.N. (1982). Обязанность во время дифференцировки стенотеле у
Hydra локализуется около границы S / G2 в терминальном клеточном цикле. Дев Биол
93: 226-230.
FUJISAWA, T., NISHIMIYA, C., SUGIYAMA, T. (1986). Дифференцировка нематоцитов в
Hydra.Curr Top Dev Biol 20: 281-290.
HAUSMANN, K., HOLSTEIN, T. (1985). Двусторонняя симметрия в комплексе книдоцил-нематоциста
пресноводной медузы Craspedacusta sowerbii Lankester (Hydrozoa,
Limnomedusae). J Ultrastruct Res 90: 89-104.
ХЕЛЛСТЕРН, С., СТЕТФЕЛЬД, Дж., ФАУЗЕР, К., ЛУСТИГ, А., ЭНГЕЛЬ, Дж., ХОЛСТЕЙН,
,Т.В., ОЗБЕК, С. (2006). Анализ структуры / функции спиналина, белка позвоночника
нематоцист гидры. FEBS J 273: 3230-3237.
HOLSTEIN, T. (1981).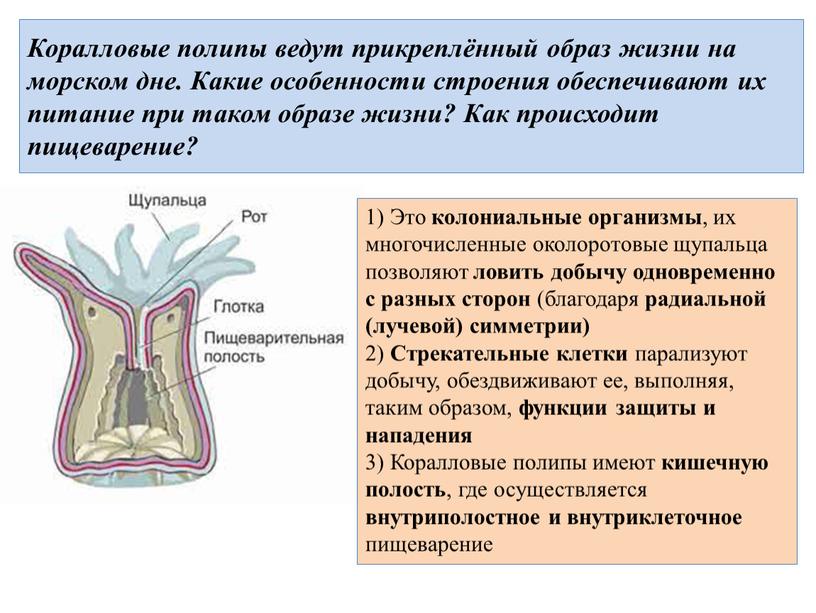 Морфогенез нематоцитов гидры и форскалии:
Морфогенез нематоцитов гидры и форскалии:
ультраструктурное исследование. J. Ultrastruct Res 75: 276-290.
Холстейн, Т., ТАРДЕНТ, П. (1984). Сверхскоростной анализ экзоцитоза:
отделяемогонематоцисты. Наука 223: 830-833.
HOLSTEIN, T.W., BENOIT, G.M., HERDER, G.V., WANNER, G., DAVID, C.N., GAUB,
H.E. (1994). Фиброзные мини-коллагены в нематоцистах гидры. Наука 265: 402-404.
HWANG, J.С., ТАКАКУ, Ю., МОМОЗЕ, Т., АДАМЧИК, П., ОЗБЕК, С., ИКЕО, К.,
ХАЛТУРИН, К., ХЕММРИХ, Г., БОШ, Т.С., ХОЛСТЕЙН, Т.В. и др., (2010).
Нематогалектин, белок нематоцисты с доменами GlyXY и галектина, демонстрирует специфический для нематоцитов альтернативный сплайсинг у гидры. Proc Natl Acad Sci
USA 107: 18539-18544.
КОЧ, А.В., ХОЛСТЕЙН, Т.В., МАЛА, К., КУРЦ, Э., ЭНГЕЛЬ, Дж., ДЭВИД, К.Н. (1998).
Спиналин, новый белок, богатый глицином и гистидином, в шипах нематоцист гидры.
J Cell Sci 111: 1545-1554.
KURZ, E.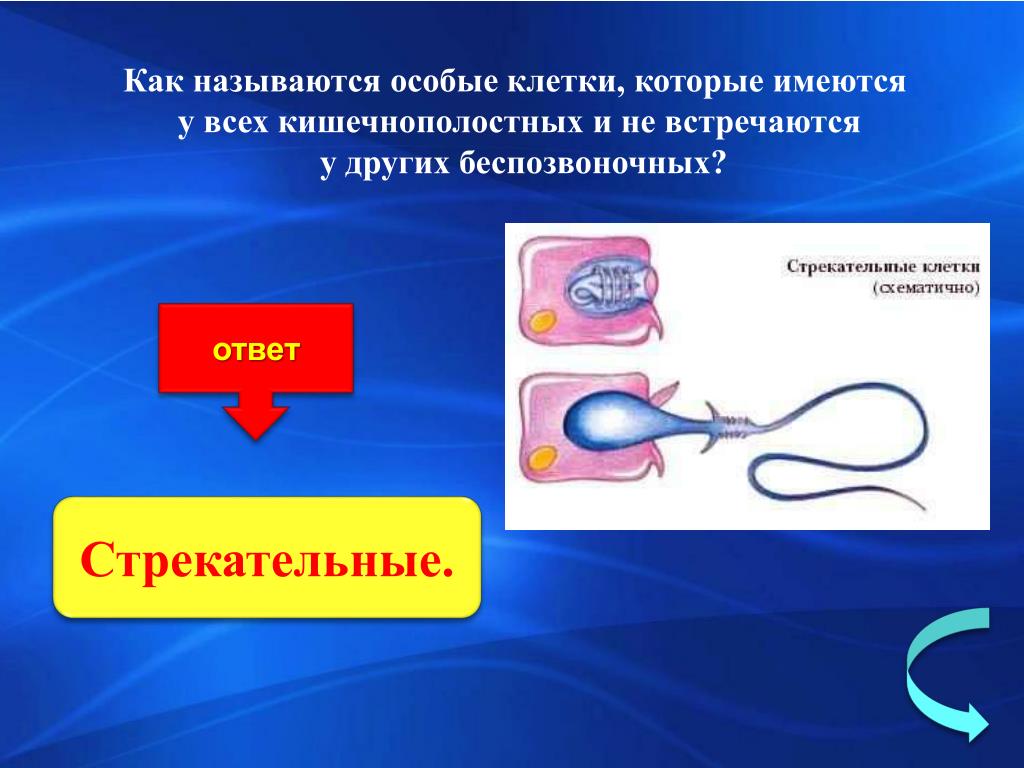 M., HOLSTEIN, T.W., PETRI, B.M., ENGEL, J., DAVID, C.N. (1991). Мини-
M., HOLSTEIN, T.W., PETRI, B.M., ENGEL, J., DAVID, C.N. (1991). Мини-
коллагенов в нематоцитах гидры. J Cell Biol 115: 1159-1169.
LENHOFF, H.M., KLINE, E.S., HURLEY, R. (1957). Богатый гидроксипролином внутриклеточный коллагеноподобный белок
нематоцист гидры. Биохим. Биофиз. Acta 26: 204-205.
LENHOFF, H.M., LENHOFF, S.G. (1988). Полипы Трембли. Scienti 9c American
258: 108-113.
MARISCAL, R.N. (1974). Экспериментальная морская биология. Academic Press, Нью-Йорк.
MEIER, S., JENSEN, P.R., DAVID, C.N., CHAPMAN, J., HOLSTEIN, T.W., GRZE-
SIEK, S., ÖZBEK, S. (2007). Непрерывная молекулярная эволюция структур белок-доменов
путем изменения одной аминокислоты. Curr Biol 17: 173-178.
NUCHTER, T., BENOIT, M., ENGEL, U., OZBEK, S., HOLSTEIN, T.W. (2006).
Наносекундная кинетика разряда нематоцист.Curr Biol 16: R316-318.
ÖZBEK, S. (2010). Книдариевая нематоциста: миниатюрный внеклеточный матрикс в пределах
секреторного пузырька.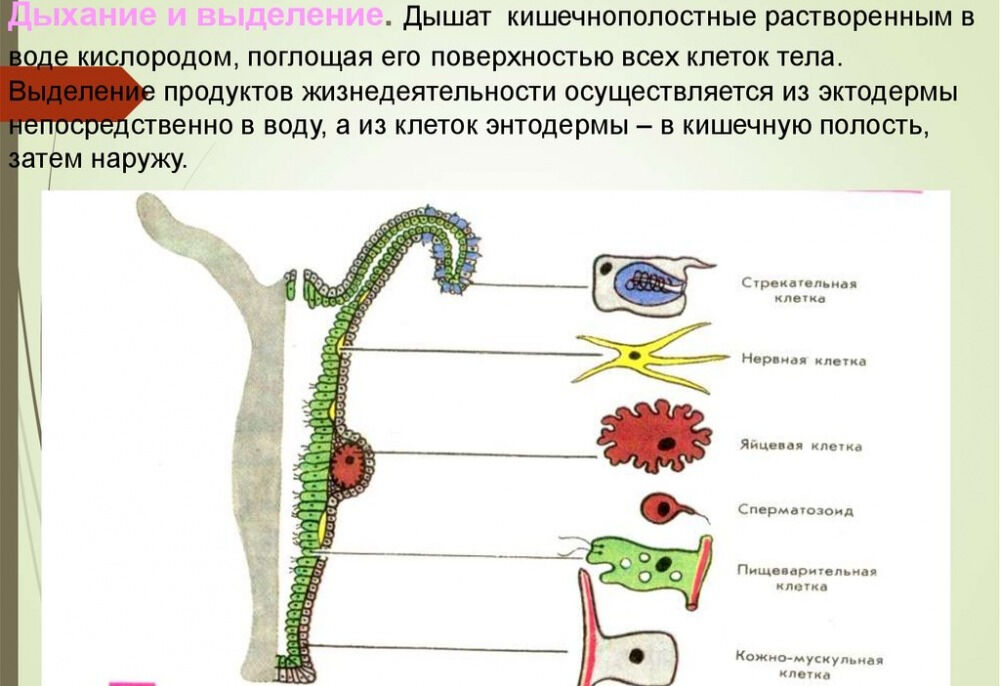
Leave A Comment