
роста, растяжения, всасывания, проводящая. Строение корня
На продольном разрезе различают следующие зоны корня (участки корня растения):
- Зона роста с корневым чехликом;
- зона растяжения и начала дифференцировки клеток;
- зона всасывания;
- проводящая зона.
Зоны корня
Рисунок: зоны корня растенияРоста
Зона роста (зона деления) корня занимает кончик длиной 2-3 мм. Это зона активно делящихся клеток, меристема корня. Все ткани корня возникают из этой образовательной ткани.
Зона роста покрыта корневым чехликом, который защищает ее от повреждений и облегчает продвижение корня в почве. Клетки чехлика обладают повышенным тургором. По мере углубления корня в почве они стираются, наружный слой их слущивается, а изнутри нарастают новые клетки за счет меристемы корня.
Растяжения
В зоне растяжения клетки сильно увеличиваются в продольном направлении и становятся цилиндрическими. В них появляются большие вакуоли. Совокупный рост клеток этой зоны создает силу, благодаря которой корень углубляется в почву.
Эта зона также невелика и занимает несколько миллиметров. В верхней ее части клетки начинают специализироваться, превращаясь окончательно в зоне всасывания в сосуды, трахеиды и другие виды клеток корня.
Всасывания
Зона всасывания корня имеет длину от нескольких миллиметров до нескольких сантиметров. Поверхность ее защищена покровной тканью — кожицей с корневыми волосками. Под кожицей находится кора корня, окружающая его центральную часть с проводящей системой.
Корневые волоски и корневой чехлик развиты у наземных растений; у водных и растений-паразитов они отсутствуют. На поперечном срезе через всасывающую зону видно первичное строение корня.
Проводящая
Проводящая зона — вся остальная часть корня, начиная от всасывающей и до стебля растения. Этот участок имеет более плотную покровную ткань, утолщен, число сосудов и ситовидных трубок увеличено за счет деятельности камбия.
Этот участок имеет более плотную покровную ткань, утолщен, число сосудов и ситовидных трубок увеличено за счет деятельности камбия.
Зона проведения корня — посредник между всасывающей зоной и надземной частью растения.
Сводная таблица строения и функций зон корня
| Название зоны | Особенности строения | Функции |
|---|---|---|
| Зона деления | Мелкие живые клетки, быстро делятся | Начало всех остальных зон и тканей корня |
| Зона роста | Клетки растут и увеличиваются в размерах | Обеспечивает основной рост корня |
| Зона всасывания | Наружный слой, представлен клетками с корневыми волосками | Обеспечивает всасывание воды с растворенными в ней полезными веществами |
| Зона проведения | Хорошо развиты проводящие ткани | Транспортная |
Внутреннее строение корня растения
Наружная покровная ткань корня — кожица — отличается от кожицы стебля и листа наличием корневых волосков, отсутствием устьиц и кутикулы, легкой проницаемостью для воды, поглощающей способностью.
Клетки кожицы располагаются в один слой. Многие из них имеют корневые волоски — удлиненные цилиндрические выросты внешней стенки клеток кожицы, длиной от 0,15мм до 1см при поперечнике в сотые доли миллиметра. Ядро клетки переходит в корневой волосок и располагается обычно на самом его конце.
В цитоплазме корневого волоска кроме ядра содержатся вакуоли с клеточным соком, бесцветные пластиды. Поверхность волосков покрыта слизистым веществом, склеивающим их с частичками почвы.
Корневые волоски недолговечны. Формируются они за 30-40 часов, живут 10-20 дней, затем отмирают. На смену их на молодом участке корня образуются новые, а участок с отмершими волосками становится проводящей зоной. Количество корневых волосков на 1мм2
 Благодаря их наличию всасывающая поверхность корня увеличивается в десятки раз.
Благодаря их наличию всасывающая поверхность корня увеличивается в десятки раз.Кора корня, прилегающая к кожице изнутри, состоит из расположенных в несколько рядов клеток основной ткани. Клетки коры имеют разные размеры. Непосредственно под кожей они крупные, а в слоях, лежащих глубже, — более мелкие.
Самый внутренний слой коры (энтодерма), облегающий центральную часть корня (центральный цилиндр) с проводящей системой, состоит из одного ряда плотно уложенных клеток. Наружные их стенки (со стороны коры) тонкие, а боковые и внутренние — утолщены и непроницаемы для воды и газов.
Между толстостенными клетками есть небольшое число тонкостенных, расположенных против сосудов центрального цилиндра. Это пропускные клетки, они проводят воду из коры корня в сосуды центрального цилиндра.
Центральный цилиндр занимает срединную часть стебля и состоит из различных тканей. Наружный его слой, примыкающий изнутри к энтодерме, состоит из тонкостенных паренхимных клеток и называется перициклом, или корнеродным слоем.
Клетки перицикла (вторичная образовательная ткань) периодически делятся и дают начало боковым корням, паренхиме корня, придаточным почкам корнеотпрысковых растений, камбию.
Далее, к центру осевого цилиндра, располагается закрытый сосудисто-волокнистый пучок, в котором чередуются участки флоэмы и ксилемы, расположенные радиально. Центр осевого цилиндра корня у большинства видов растений занят одним крупным или несколькими мелкими сосудами. У некоторых видов центр занят клетками основной ткани (паренхимы), заполняющей также промежутки между флоэмными и ксилемными участками.
Зона проведения корня представлена тканью. Корни растений. Типы корневой системы. Функции корня. Зоны корня. Видоизменение корней. Особенности строения клеток
>>Зоны (участки) корня
§ 14.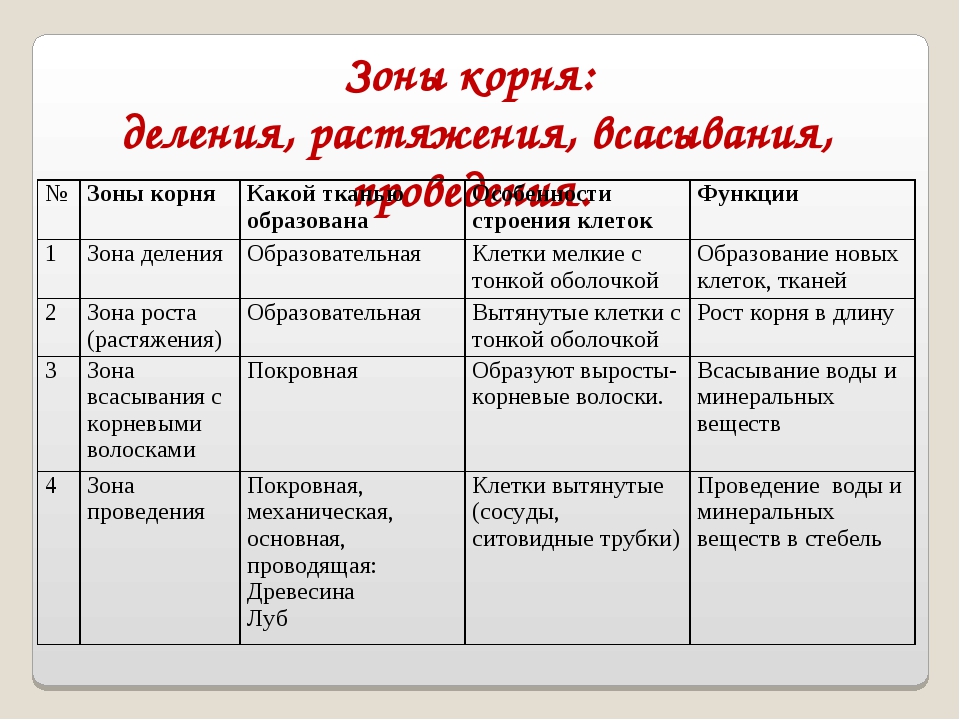
Разрежем молодой главный корень вдоль . С одной из половинок сделаем тонкий срез, приготовим из него препарат и рассмотрим под микроскопом . Клетки разных участков корня отличаются друг от друга формой и размерами 27 . Кончик корня покрыт, как наперстком, корневым чехликом. Чехлик можно заметить невооруженным глазом, рассматривая на свет кончик корня: здесь корень несколько темнее и плотнее, чем в других участках.
Корневой чехлик предохраняет верхушку корня от повреждений твердыми частицами почвы. Клетки корневого чехлика живут недолго, постепенно отмирают и слущиваются. Взамен отмерших клеток постоянно образуются новые.
Под чехликом находится участок делящихся клеток — зона деления. Этот участок образован мелкими, плотно прилегающими одна к другим живыми клетками. Клетки здесь все время делятся, число их увеличивается. Выше расположена зона роста, или растяжения; здесь клетки вытягиваются, в результате чего корень растет в длину.
Еще дальше от кончика корня поверхностные клетки образуют множество тонких и прозрачных корневых волосков. У многих растений корневые волоски можно увидеть и без микроскопа. Например, они хорошо заметны у проростков гороха. У проростков пшеницы и многих других злаков корневые волоски напоминают легкий пушок 28 .
Корневой волосок представляет собой длинный вырост наружной клетки корня.
Он одет клеточной оболочкой, под которой находятся цитоплазма , ядро , бесцветные пластиды и вакуоль с клеточным соком 27 .
Корневые волоски очень малы — длиной обычно не более 10 мм — и недолговечны. У многих растений они живут всего несколько дней, а затем отмирают. Новые волоски возникают в виде выростов более молодых поверхностных клеток, расположенных ближе к кончику корня.
Проникая между частицами почвы, корневые волоски плотно прилегают к ним и всасывают из почвы воду и другие вещества. Поэтому участок корня, находящийся выше зоны роста, на котором находятся корневые волоски, принято называть зоной всасывания.
Таким образом, зона всасывания, как и другие зоны, постоянно перемещается вслед за кончиком растущего корня и все время находится примерно на одном и том же расстоянии от него Выше зоны всасывания, то есть еще дальше от кончика корня, находится зона проведения. По клеткам этой зоны вода с растворенными минеральными веществами, поглощенная корнем, перемещается к стеблю.
Корни, особенно молодые участки, несущие корневые волоски, легко повреждаются. Корневые волоски хорошо сохраняются, когда растение пересаживают вместе с комом земли. Поэтому рассаду овощных и декоративных растений выращивают в торфоперегнойных горшочках. Рассада, выращенная в горшочках из торфа и перегноя, быстро приживается, так как корни при пересадке не страдают.
Итак, невооруженным глазом у молодого корня можно различить корневой чехлик, корневые волоски, похожие на легкий пушок, и зону проведения. Но более детально все зоны можно рассмотреть только под микроскопом.
Микроскоп позволяет увидеть сходное строение клеток одной зоны корня и отличие их от клеток других зон. Группы клеток одинакового строения и выполняющих одинаковые функции называют тканями.
Корень, как и другие органы, состоит из разных тканей: зона деления — из образовательной ткани, зона всасывания покрыта всасывающей тканью.
В состав проводящих тканей корня входят сосуды. По ним из почвы поднимается вода с растворенными в ней питательными веществами. В проводящих тканях корня находятся также клетки, по которым в корень поступают органические вещества, образовавшиеся в листьях и стеблях.
Под слоем клеток, образующих корневые волоски, расположена кора корня. Она состоит из сомкнутых округлых клеток. Оболочки клеток пропитаны пробковым веществом. Клетки коры образуют покровную ткань корня.
Прочность и упругость корня обеспечивает механическая ткань . Ее составляют вытянутые вдоль корня клетки с толстыми оболочками. Они рано теряют содержимое и заполнены воздухом.
1. Какие участки (зоны) можно различить, рассматривая молодой корень?
Какие участки (зоны) можно различить, рассматривая молодой корень?
2. Каково значение корневого чехлика?
3. Где располагается зона делящихся клеток? Чем ее клетки отличаются от клеток других зон?
4. Где располагается зона роста (растяжения) корня? Каково ее значение?
5. Что такое корневой волосок? Какое строение он имеет?
6. Почему одну из зон корня называют всасывающей?
7. Где расположена зона проведения корня? Почему ее так называют?
8. Что такое ткань?
9. Какие ткани различают в корнях растений?
> 1. Осторожно выньте из почвы проросток пшеницы и рассмотрите его. Какая зона корня покрыта приставшей почвой?
2. Прорастите семена редиса, гороха или фасоли и зерновки пшеницы. Внимательно рассмотрите корни проростков этих растений, чтобы обнаружить корневые волоски.
Корчагина В. А., Биология: Растения, бактерии, грибы, лишайники: Учеб. для 6 кл. сред. шк. — 24-е изд. — М.: Просвещение, 2003. — 256 с.:ил.
Календарно-тематическое планирование по биологии, видео по биологии онлайн , Биология в школе скачать
Содержание урока конспект урока опорный каркас презентация урока акселеративные методы интерактивные технологии Практика задачи и упражнения самопроверка практикумы, тренинги, кейсы, квесты домашние задания дискуссионные вопросы риторические вопросы от учеников Иллюстрации аудио-, видеоклипы и мультимедиа фотографии, картинки графики, таблицы, схемы юмор, анекдоты, приколы, комиксы притчи, поговорки, кроссворды, цитаты Дополнения рефераты статьи фишки для любознательных шпаргалки учебники основные и дополнительные словарь терминов прочие Совершенствование учебников и уроков исправление ошибок в учебнике обновление фрагмента в учебнике элементы новаторства на уроке замена устаревших знаний новыми Только для учителей идеальные уроки календарный план на год методические рекомендации программы обсуждения Интегрированные урокиКоторые позволяют ему осуществлять важнейшие функции в растительном организме. Внутреннее строение этого органа отличается четкой дифференциацией, благодаря которой осуществляется согласованная работа всего организма.
Внутреннее строение этого органа отличается четкой дифференциацией, благодаря которой осуществляется согласованная работа всего организма.
Что такое корень
Корнем называют осевой подземный орган растения. В зависимости от особенностей расположения различают главный, боковые и придаточные. Первый вид определить очень просто. Главный корень у растения всегда один. На нем расположены боковые. Вместе они формируют Она характерна для всех представителей класса Двудольные, включающие всем известные семейства Розоцветных, Пасленовых, Астровых, Капустных, Бобовых, и других. Прямо от побега отходят Они растут пучком. Такую корневую систему, которая называется мочковатой, имеют Однодольные растения: Злаковые, Луковые и Лилейные.
Функции корня
Главная задача подземного органа заключается в закреплении растения в почве, обеспечения его водой и растворами минеральных веществ. С помощью корня происходит поглощение из почвы соединений азота, калия, железа, магния, фосфора и других элементов. Этот процесс называется минеральным питанием. Полученные вещества растения используют для самостоятельного синтеза органических соединений.
Корень и побег осуществляют свои функции в тесной взаимосвязи. Подземный орган обеспечивает растение водой с растворами минеральных веществ. Они поступают из корня ко всем частям побега. Это восходящий ток веществ. В свою очередь, в результате фотосинтеза в листьях образуются органические вещества. Они передвигаются из побега в корень, осуществляя нисходящий ток.
Зоны корня растения
Если разрезать подземный орган вдоль его оси, можно без труда заметить зоны корня. Все они являются специализированными, с четкой взаимосвязью особенностей строения и выполняемых функций. Зоны расположены в следующей последовательности: корневой чехлик, деления, растяжения, всасывания, проведения. Уже только по названиям можно догадаться, из элементов каких тканей они состоят, и какова их роль в жизнедеятельности растительных организмов. Рассмотрим каждую из них более подробно.
Рассмотрим каждую из них более подробно.
Корневой чехлик
Чтобы проникнуть вглубь почвы, корень постоянно нарастает своей верхушкой. Эту функцию осуществляет зона деления корня, которая покрыта корневым чехликом. Он надежно защищает клетки образовательной ткани от механических повреждений, предотвращает повреждение верхушки подземного органа во время его проникновения в почву.
Корневой чехлик образован несколькими слоями живых клеток покровной ткани. Они не однородны по своему строению. Так, клетки наружного слоя постоянно разрушаются при соприкосновении с частицами почвы. Поэтому они требуют восстановления. Этот процесс происходит за счет деления клеток образовательной ткани изнутри. Корневой чехлик также играет роль своеобразного «навигатора» для подземного органа растений. Поскольку он обладает способностью воспринимать силу земного притяжения, эта зона определяет направление роста корня в глубину.
Меристема
Далее следует часть корня, объединяющая две зоны: деления и растяжения. За счет этих структур осуществляется увеличение его размеров. Поэтому ее называют зона роста корня. Какими особенностями строения обладает каждая из них?
Зона деления корня располагается за корневым чехликом. Она полностью сформирована образовательной тканью — меристемой, длина которой не превышает 3 мм. Ее клетки мелкие, плотно прилегают друг к другу, имеют тонкие стенки. Эта зона обладает уникальной способностью. При ее делении образуются клетки любых других тканей. Это очень важно для восстановления утраченных или поврежденных частей органов растительного организма.
Зона растяжения
За меристемой зона роста корня продолжается клетками другого типа. Они постоянно растут, удлиняются, приобретая фиксированную форму и размеры. Это зона растяжения. Размеры ее также незначительны: всего несколько мм. Увеличиваясь в размерах, ее клетки продвигают меристему с корневым чехликом все глубже. Зона растяжения также создана образовательной тканью. Поэтому здесь могут формироваться клетки любых типов.
Поэтому здесь могут формироваться клетки любых типов.
Зона всасывания корня
Следующая структура имеет более значительные размеры, занимая участок от 5 до 20 мм. Это зона всасывания корня. Ее основной функцией является поглощение воды с раствором питательных веществ из почвы. Данный процесс осуществляется при помощи корневых волосков, которые являются выростами клеток покровной ткани. Их длина варьирует от нескольких миллиметров до одного сантиметра. Иногда этот показатель превышает размеры самих клеток.
Корневые волоски — постоянно обновляющиеся образования. Они живут до 20 дней, после чего отмирают. Новые волоски образуются из клеток, которые располагаются вблизи зоны роста. При этом в верхней части они исчезают. Поэтому получается, что зона всасывания по мере роста корня погружается в почву все глубже.
Корневые волоски очень легко повредить. Поэтому во время пересадки растений рекомендуется переносить его вместе с почвой, в которой оно росло до этого. Структуры эти достаточно многочисленны. На 1 квадратном миллиметре образуется несколько сотен значительно увеличивает поверхность всасывания, которая в несколько сотен раз превышает площадь
Боковые корни
Зона проведения корня, или боковых корней, является самой большой. Это участок, в пределах которого подземный орган утолщается и ветвится. Здесь формируются боковые корни растения. В зоне проведения нет корневых волосков, поэтому поглощение питательных веществ из почвы не осуществляется. Зона проведения корня служит «транспортной магистралью» от зоны всасывания к наземной части растения.
Особенности внутреннего строения
Как видите, все зоны корня отличаются четкой специализацией. Это касается и внутреннего строения подземного органа. На поперечном срезе корня в зоне всасывания отчетливо видно несколько слоев. Снаружи находится покровная ткань. Она представлена одним слоем живых клеток кожицы. Именно они формируют новые корневые волоски.
Под кожицей размещается кора. Это несколько слоев основной ткани. Через них передвигаются растворы минеральных веществ из корневых волосков к элементам проводящей ткани. Внутреннюю осевую часть корня занимает центральный цилиндр. Эта структура состоит из сосудов и а также элементов механической и запасающей ткани. Вокруг центрального цилиндра находится слой клеток образовательной ткани, из которых формируются боковые корни.
Это несколько слоев основной ткани. Через них передвигаются растворы минеральных веществ из корневых волосков к элементам проводящей ткани. Внутреннюю осевую часть корня занимает центральный цилиндр. Эта структура состоит из сосудов и а также элементов механической и запасающей ткани. Вокруг центрального цилиндра находится слой клеток образовательной ткани, из которых формируются боковые корни.
Способы формирования корневой системы
Знания о строении и физиологии подземного органа растений человек издавна использует в своей хозяйственной деятельности. Так, для формирования дополнительных корней, развивающихся в поверхностном слое почвы, рекомендуется окучивать участок и подсыпать землю к основанию побегов.
Чтобы увеличить количество боковых корней применяется метод пикировки. Осуществляют его во время пересаживания рассады в открытый грунт. Для этого у проростка отщипывают кончик главного корня, в результате чего вся система становится более разветвленной. Боковые корни разрастаются, а значит, почвенное питание растений осуществляется более эффективно. Кроме того, при окучивании и пикировке их преобладающее количество развивается в верхнем слое почвы, который является более плодородным.
Итак, зоны корня представляют собой разные по особенностям строения участки осевого подземного органа растений. Все они отличаются узкой специализацией, обусловленной особенностями их строения. Различают следующие участки: корневой чехлик, деления, роста, включающие зоны растяжения и всасывания, и проведения.
Живые организмы изучает наука биология. Строение корня растения рассматривается в одном из разделов ботаники.
Корень является осевым вегетативным органом растения. Для него характерен неограниченный верхушечный рост и радиальная симметрия. Особенности строения корня зависят от многих факторов. Это эволюционное происхождение растения, его принадлежность к тому или иному классу, среда обитания. В качестве основных функций корня можно назвать укрепление растения в почве, участие в вегетативном размножении, запас и синтез органических питательных веществ. Но самая важная функция, обеспечивающая жизнедеятельность растительного организма, — почвенное питание, которое осуществляется в процессе активного всасывания из субстрата воды, содержащей растворенные минеральные соли.
Но самая важная функция, обеспечивающая жизнедеятельность растительного организма, — почвенное питание, которое осуществляется в процессе активного всасывания из субстрата воды, содержащей растворенные минеральные соли.
Типы корней
Внешнее строение корня во многом обусловлено тем, к какому типу он относится.
- Главный корень. Его образование происходит из зародышевого корешка, когда семя растения начинает прорастать.
- Придаточные корни. Они могут появляться на различных частях растения (стебель, листья).
- Боковые корни. Именно они образуют разветвления, начинаясь от ранее появившихся корней (главного или придаточных).
Виды корневых систем
Корневая система — общность всех корней, которые имеются у растения. При этом внешний вид этой совокупности у различных растений может сильно варьироваться. Причиной тому служит наличие или отсутствие, а также разная степень развития и выраженности различных типов корней.
В зависимости от этого фактора различают несколько типов корневых систем.
- Название говорит само за себя. Главный корень выступает в роли стержня. Он хорошо выражен по размеру и длине. Строение корня по данному типу характерно для Это щавель, морковь, фасоль и пр.
- Для данного типа характерны свои особенности. Внешнее строение корня, являющегося главным, ничем не отличается от такового у боковых. Он не выделяется в общей массе. Образовавшись из зародышевого корешка, он растет совсем недолго. Мочковая корневая система характерна для однодольных растений. Это хлебные злаки, чеснок, тюльпан и пр.
- Корневая система смешанного типа. Ее строение соединяет в себе особенности двух, описанных выше, типов. Главный корень хорошо развит и выделяется на общем фоне. Но при этом сильно развиты и придаточные корни. Характерна для помидора, капусты.
Историческое развитие корня
Если рассуждать с точки зрения филогенетического развития корня, то его появление произошло гораздо позже, чем образование стебля и листа. Скорее всего, толчком для этого послужил выход растений на сушу. Для того чтобы закрепиться в твердом субстрате, представителям древней флоры требовалось что-то, что может послужить опорой. В процессе эволюции сначала образовались корнеподобные подземные веточки. Позже они дали начало развитию корневой системы.
Скорее всего, толчком для этого послужил выход растений на сушу. Для того чтобы закрепиться в твердом субстрате, представителям древней флоры требовалось что-то, что может послужить опорой. В процессе эволюции сначала образовались корнеподобные подземные веточки. Позже они дали начало развитию корневой системы.
Корневой чехлик
Формирование и развитие корневой системы осуществляется в течение всей жизни растения. Строение корня растения не предусматривает наличия листьев и почек. Его рост осуществляется за счет увеличения в длину. В точке роста он покрыт корневым чехликом.
Процесс роста связан с образовательной ткани. Именно она находится под корневым чехликом, выполняющим функцию защиты нежных делящихся клеток от повреждений. Сам чехлик — это совокупность тонкостенных живых клеток, в которых постоянно происходит процесс обновления. То есть, при продвижении корня в почве застарелые клетки постепенно слущиваются, а на их месте нарастают новые. Также расположенные снаружи клетки чехлика выделяют особую слизь. Она облегчает продвижение корня в твердом почвенном субстрате.
Общеизвестно, что в зависимости от среды обитания строение растений сильно различается. Например, водные растения не имеют корневого чехлика. В процессе эволюции у них образовалось другое приспособление — водяной кармашек.
Строение корня растения: зона деления, зона роста
Клетки, появившись из со временем начинают дифференцироваться. Таким образом формируются зоны корня.
Зона деления. Она представлена клетками образовательной ткани, которые впоследствии и дают начало всем остальным типам клеток. Размер зоны — 1 мм.
Зона роста. Представлена гладким участком, длина которого составляет от 6 до 9 мм. Следует сразу за зоной деления. Для клеток характерен интенсивный рост, в ходе которого они сильно вытягиваются в длину, и постепенная дифференциация. Следует заметить, что процесс деления в данной зоне почти не осуществляется.
Зона всасывания
Этот участок корня протяженностью несколько сантиметров также часто называют зоной корневых волосков. Это название отражает особенности строения корня на данном участке. Там имеются выросты клеток кожицы, размер которых может варьироваться от 1 мм до 20 мм. Это и есть корневые волоски.
Это название отражает особенности строения корня на данном участке. Там имеются выросты клеток кожицы, размер которых может варьироваться от 1 мм до 20 мм. Это и есть корневые волоски.
Зона всасывания — это место, где осуществляется активное поглощение воды, в которой содержатся растворенные минеральные вещества. Деятельность клеток корневых волосков, в данном случае, можно сравнить с работой насосов. Этот процесс очень энергозатратный. Поэтому в клетках зоны всасывания содержится большое количество митохондрий.
Очень важно обратить внимание еще на одну особенность корневых волосков. Они способны выделять особую слизь, содержащую угольную, яблочную и лимонную кислоты. Слизь способствует растворению минеральных солей в воде. Частицы почвы благодаря слизи словно приклеиваются к корневым волоскам, облегчая процесс всасывания питательных веществ.
Строение корневого волоска
Увеличение площади зоны всасывания происходит именно за счет корневых волосков. Например, их количество у ржи достигает 14 миллиардов, образуя суммарную длину до 10 000 километров.
Внешний вид корневых волосков делает их похожими на белый пушок. Живут они недолго — от 10 до 20 дней. На формирование новых у растительного организма уходит совсем немного времени. Например, образование корневых волосков у молодых сеянцев яблони осуществляется за 30-40 часов. Тот участок, где произошло отмирание этих необычных выростов, еще в течение некоторого времени может всасывать воду, а потом его покрывает пробка, и эта способность теряется.
Если говорить о строении оболочки волоска, то, прежде всего, следует выделить ее тонкость. Эта особенность помогает волоску поглощать питательные вещества. Клетка его почти полностью занята вакуолью, окруженной тонким слоем цитоплазмы. Ядро располагается в верхней части. Пространство вблизи клетки представляет собой особый слизистый чехол, способствующий склеиванию корневых волосков с мелкими частичками почвенного субстрата. Благодаря этому гидрофильность почвы повышается.
Поперечное строение корня в зоне всасывания
Зону корневых волосков также часто называют зоной дифференциации (специализации). Это не случайно. Именно здесь на поперечном разрезе можно увидеть определенную слоистость. Она обусловлена разграничением слоев внутри корня.
Таблица «Строение корня на поперечном срезе» представлена ниже.
Следует отметить, что внутри коры тоже имеется разграничение. Ее наружный слой называется экзодерма, внутренний — эндодерма, а между ними находится основная паренхима. Именно в этом промежуточном слое происходит процесс направления растворов питательных веществ в сосуды древесины. Также, в паренхиме синтезируются некоторые жизненно важные для растения органические вещества. Таким образом, внутреннее строение корня позволяет в полном объеме оценить значимость и важность функций, которые выполняет каждый из слоев.
Зона проведения
Располагается над зоной всасывания. Самый большой по длине и наиболее прочный участок корня. Именно здесь происходит передвижение важных для жизнедеятельности растительного организма веществ. Это возможно благодаря хорошему развитию проводящих тканей в этой зоне. Внутреннее строение корня в зоне проведения обуславливает его способность транспортировать вещества в обоих направлениях. По восходящему току (вверх) идет передвижение воды с растворенными в ней минеральными соединениями. А вниз доставляются органические соединения, которые участвуют в жизнедеятельности клеток корня. Зона проведения — это место образования боковых корней.
Строение корня проростка фасоли четко иллюстрирует основные этапы процесса формирования корня растений.
Особенности строения корня растения: соотношение наземной и подземной частей
Для многих растений характерно такое развитие корневой системы, которое приводит к ее преобладанию над наземной частью. Примером может служить кочанная капуста, корень которой в глубину может вырасти на 1,5 метра. Ширина его может составлять до 1, 2 метра.
Настолько разрастается, что занимает пространство, диаметр которого может достигать 12 метров.
А у растения люцерна высота наземной части не превышает 60 см. Тогда как длина корня может составлять более 2 метров.
Все растения, обитающие в местностях с песчаными и скалистыми почвами, имеют очень длинные корни. Это обусловлено тем, что в таких почвах вода и органические вещества находятся очень глубоко. В процессе эволюции растения долго приспосабливались к таким условиям, постепенно менялось строение корня. В результате чего они стали достигать той глубины, где растительный организм может запастись необходимыми для роста и развития веществами. Так, например, корень может в глубину составлять 20 метров.
Корневые волоски у пшеницы ветвятся настолько сильно, что их суммарная длина может достигать 20 км. Однако, это не предельная величина. Неограниченный верхушечный рост корней в отсутствие сильной конкуренции с другими растениями может увеличить это значение еще в несколько раз.
Видоизменения корней
Строение корня некоторых растений может меняться, образуя так называемые видоизменения. Это своего рода приспособления растительных организмов в конкретных условиях обитания. Ниже представлено описание некоторых видоизменений.
Корневые клубни характерны для георгина, чистяка и некоторых других растений. Образуются за счет утолщения придаточных и боковых корней.
Плющ и кампсис тоже отличаются особенностями строения этих вегетативных органов. У них имеются так называемые корни-прицепки, которые позволяют им цепляться за рядом стоящие растения и другие опоры, находящиеся в их досягаемости.
Отличающиеся большой длиной и всасывающие воду, имеются у монстеры и орхидеи.
Растущие вертикально вверх дыхательные корни участвуют в выполнении функции дыхания. Имеются у ивы ломкой.
У таких овощных культур, как морковь, свекла, редис, имеются корнеплоды, которые образовались за счет разрастания главного корня, внутри которого запасаются питательные вещества.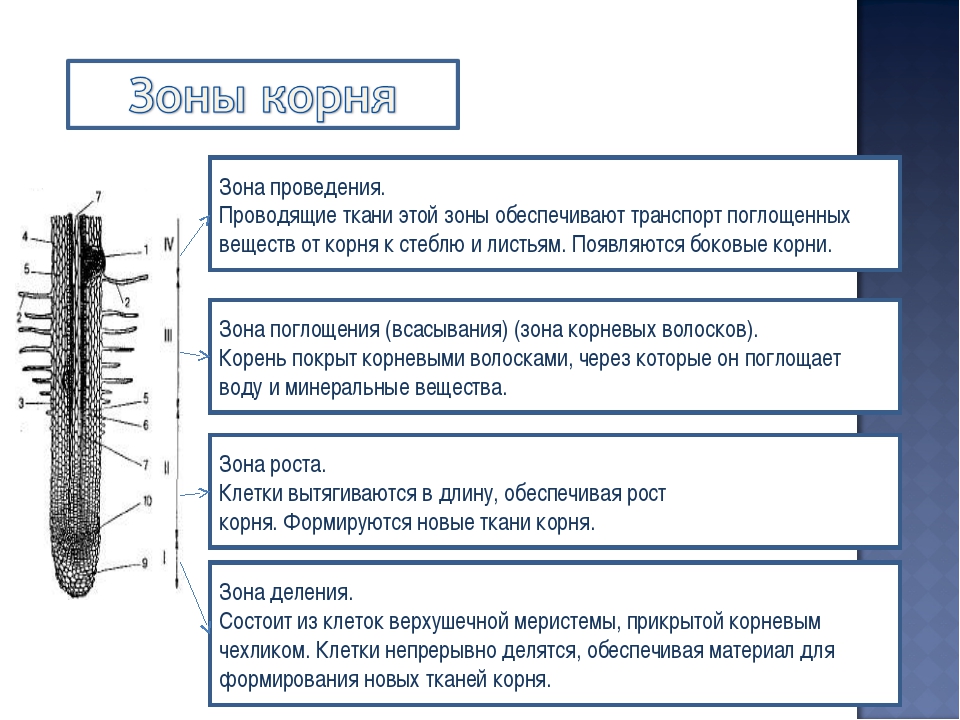
Таким образом, особенности строения корня растения, приводящие к образованию видоизменений, зависят от многих факторов. Основными являются среда обитания и эволюционное развитие.
Корневая система почти любого покрытосеменного растения состоит из множества корней. Каждый отдельно взятый корень растет своей верхушкой, или кончиком (будем считать это нижней частью корня). Благодаря этому корень может продвигаться в почве в глубь или в стороны. Это дает возможность всасывать воду и минеральные вещества там, где корни их еще не всасывали. Таким образом, чем дальше участок корня от своего кончика (и соответственно ближе к стеблю), тем он старее. В этих местах корни не растут в длину, здесь они в основном только проводят вещества в обе стороны (вверх и вниз).
Если растущую часть корня разрезать вдоль (как бы симметрично разрезать на левую и правую части), то, начиная от кончика и вверх, можно увидеть четыре зоны корня: деления, роста, всасывания, проведения. Понятно, что в зоне деления клетки делятся, в зоне роста — растут, в зоне всасывания — поглощают воду и растворенные в ней вещества, в зоне проведения — проводят воду и вещества по направлению к стеблю. То есть благодаря этим зонам корень выполняет одну из своих основных функций — обеспечивает растение водой и минеральными веществами.
Зона деления прикрыта так называемым корневым чехликом . Он прикрывает самый кончик корня и защищает зону деления от повреждений. Клетки корневого чехлика выделяют слизь.
Корень обладает так называемым положительным геотропизмом. Это означает, что он растет вниз, к центру Земли. И именно клетки корневого чехлика отвечают за эту способность.
Клетки корневого чехлика быстро слущиваются и заменяются новыми.
Под корневым чехликом и чуть выше его находится зона деления . Она состоит из клеток образовательной ткани, которые постоянно делятся. Клетки в этой зоне мелкие с тонкими стенками.
Более верхние клетки зоны деления перестают делится и начинают расти, в основном за счет вытягивания вдоль корня. Таким образом, они уже входят (или формируют) зону роста корня. Зону роста также нередко называют зоной растяжения .
Таким образом, они уже входят (или формируют) зону роста корня. Зону роста также нередко называют зоной растяжения .
Вместе, корневой чехлик и зоны деления и растяжения составляют всего несколько миллиметров. Выше находится зона всасывания , состоящая из корневых волосков. Именно в этом месте корень поглощает воду с растворенными в ней минеральными веществами.
Обычно каждый корневой волосок имеет длину не более 1 сантиметра и очень тонкий. Он представляет собой боковой вырост клетки кожицы корня (это покровная ткань). То есть один волосок — это одна клетка, причем даже не целая клетка, а ее часть. При этом корневые волоски можно увидеть у проростков многих растений невооруженным глазом. Все вместе они похожи на пушок. Под микроскопом можно увидеть, что корневой волосок является живой клеткой. В нем есть цитоплазма, ядро, лейкопласты, вакуоль, митохондрии и различные включения.
Корневые волоски живут не долго, у каждого вида по-разному. Но в среднем где-то 15 дней. Выше по корню находятся более старые волоски, и они отмирают, а снизу, сразу за зоной роста, начинают образовываться новые волоски. Таким образом, кончик корня (чехлик, зоны деления, роста, всасывания) постоянно смещается, проникая вглубь почвы.
Выше зоны всасывания (и даже немного заходя внутрь нее в центральной части корня) находится зона проведения . В центральной оси этой зоны находятся сосуды, по которым водный раствор поднимается вверх по направлению к стеблю. Сосуды относятся к проводящей ткани.
От корневых волосков до сосудов вода и минеральные вещества передвигаются по клеткам паренхимы (это основная ткань). Это передвижение осуществляется за счет так называемой сосущей силы, которую вызывает разность тургорного и осмотического давления.
В зоне проведения есть не только сосуды, но и ситовидные трубки. По ним в корень из побега поступают органические вещества. Они нужны корню для питания (от этого зависит его рост и развитие).
Корневой чехлик и зона деления
1) В каком отделе корня расположен чехлик, зачем он нужен и какими клетками представлен? Чехлик находится непосредственно на кончике корня, он играет защитную роль. Состоит он из нескольких слоев, при этом внешний слой имеет слизь, его клетки постоянно отрываются.
Состоит он из нескольких слоев, при этом внешний слой имеет слизь, его клетки постоянно отрываются.
2) Благодаря чему чехлик восстанавливает размеры? За счет зоны деления — места, где расположена образовательная ткань, которая все время делится.
3) В чем значение зоны деления помимо пополнения клеток чехлика? Она формирует все другие клетки корня.
Зона роста (растяжения)
1) Какое место имеет зона роста в корне? Расположена за зоной деления, перед зоной всасывания.
2) Клетки какой ткани входят в состав зоны роста? В чем их особенности? В нее входят частично клетки образовательной ткани, но есть и клетки, которые закончили деление и просто растут. Клетки растут благодаря поглощению влаги и формированию крупных вакуолей, поэтому этот участок корня удлиняется.
3) В чем функция зоны роста? Она толкает зону деления вперед, вглубь почвы, вместе с корневым чехликом.
4) В клетки каких тканей могут превращаться выросшие клетки зоны роста? Одна часть клеток превратится в клетки покровной ткани, вторая станет клетками ткани основной, наконец, оставшаяся часть — это клетки проводящей ткани.
5) Как на опыте доказать, что корень растет со стороны верхушки, за счет зоны деления и зоны роста? Нанесем на проросток фасоли (или конского боба) две пары черточек-меток — первую у верхушки корня, вторую его основания. Буквально спустя сутки мы увидим, что расстояние между метками увеличилось только у верхушки корня. Вывод прост — корень характеризуется именно верхушечным ростом.
Зона всасывания
1) В данной зоне расположены корневые волоски. Какое строение имеют их клетки? Оболочки клеток тонкие, слизистые, центральные вакуоли крупные. Длина волосков от 0,1 до 1,5 миллиметров, но иногда они могут достигать 8-9 мм, например, это свойственно пшенице.
2) Зачем нужны волоски? Они в ходе развития плотно сцепляются с микроскопическими комочками почвы, а содержащаяся на них слизь активно растворяет минералы в почве.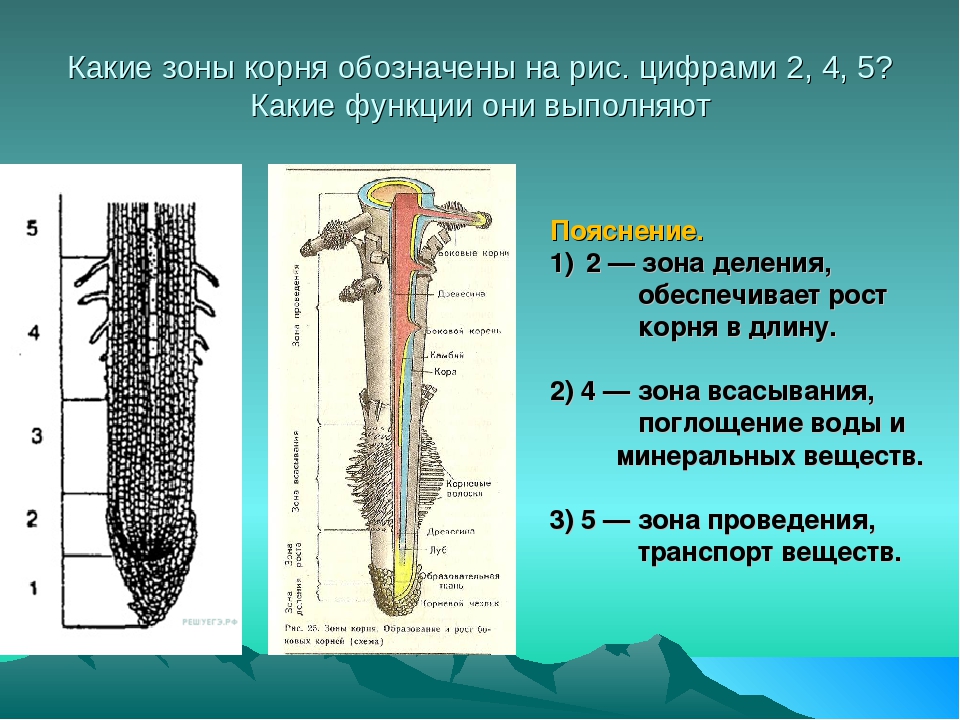 Таким образом множественные волоски увеличивают в десятки, а иногда и в сотни раз бывшую до того небольшой всасывающую поверхность корня. Впрочем, живет корневой волосок недолго, самое большее несколько дней.
Таким образом множественные волоски увеличивают в десятки, а иногда и в сотни раз бывшую до того небольшой всасывающую поверхность корня. Впрочем, живет корневой волосок недолго, самое большее несколько дней.
3) Где отмирают и где растут новые корневые волоски? Отмирают они в конце зоны всасывания, новые появляются вблизи зоны роста. Делаем вывод, что зона всасывания все время растет, проникая в новые слои почвы. Однако в целом зона всасывания сохраняет прежнюю длину.
Внутреннее строение корня в зоне всасывания
1) Наружный слой клеток, кожица (называемая ризодермой или эпиблемой), образован корневыми волосками.
2) Клетки коры корня являются следующим слоем. Они представляют собой живые, тонкостенные клетки, между которыми расположены крупные межклетники.
3) В коре можно выделить три слоя — наружный плотный (он примыкает к кожице, и в зоне проведения берет на себя функции кожицы после ее отмирания), центр коры из крупных клеток, внутренняя часть коры (представлена одним слоем смыкающихся клеток). В коре корня могут запасаться вещества, например, витамины, крахмал, белки.
4) Проводящие ткани в центре корня: центральный цилиндр, куда входят древесина и луб. Сосуды древесины центрального цилиндра — каковы они? Длинные полые трубки, без живого содержимого, с одревесневевшими стенками, по которым движется вода с содержащимися в ней необходимыми минеральными солями. Ситовидные трубки луба построены из живых клеток, для которых характерны поперечные перегородки в виде сита (отсюда и название), но без ядра. Сердцевины в корне нет. Как расположена древесина и луб в центральном цилиндре в зоне всасывания? Древесина находится в центре, ее лучи доходят до периферии центрального цилиндра. Различные виды растений имеют от трех до нескольких десятков лучей. Луб расположен между лучами древесины.
Зона проведения
1) Что происходит с клетками наружного слоя коры после того, как отмирают корневые волоски? Они также отмирают, и мертвые защищают внутренние части корня от повреждений и бактерий.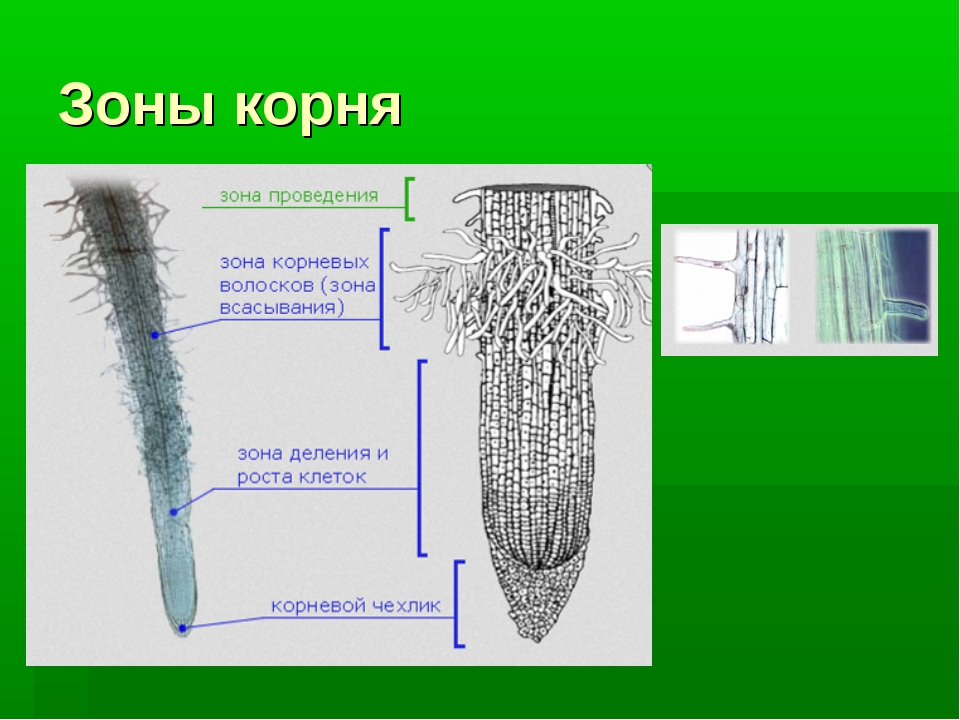 Такой участок корня уже не может всасывать, а только проводит вещества.
Такой участок корня уже не может всасывать, а только проводит вещества.
2) Растет ли зона проведения? Да. И еще на нее приходится большая часть длины долгоживущих корней.
Морфология и функции корневого окончания
В процессе роста и формирования корней древесных растений происходит изменение физиологического, анатомического и морфологического состояний всех его тканей, за исключением группы инициальных клеток, если не считать постоянного их обновления.
Указанный процесс трансформации тканей имеет хорошо выраженный цикл, состоящий из отдельных фаз, и заканчивается прекращением анатомических и морфологических изменений клеток и тканей, стабилизацией физиологических функций, составляющих данную зону корня. Указанный цикл во времени и в пространстве приурочен к определенной зоне корня, следующей за корневым чехликом. Эту зону, включая корневой чехлик, принято называть корневым окончанием.
Анатомическое и морфологическое строение корневого окончания выражено достаточно четко и тесно коррелирует с соответствующими функциями отдельных его частей и тканей. Части корневого окончания, в которых достаточно четко проявляются отдельные физиологические функции, принято называть его зонами. Все корневое окончание сопряжено с различными проявлениями роста, в связи с чем иногда указанные зоны называют зонами роста, хотя ясно выраженной границы между зонами роста не существует.
По общепринятой в ботанике схеме обычно различают 3 зоны роста корневого окончания: деления, растяжения, поглощения, или всасывания (ее еще называют зоной корневых волосков).
Корневой чехлик как бы прикрывает апикальную меристему по оси линейного роста корня. Он состоит из живых клеток, которые постепенно отлущиваются и заменяются новыми, образующимися в результате деления инициальных клеток. Долгое время корневому чехлику отводили в основном только защитную роль. В этом убеждали отлущивание внешних его клеток, обильное выделение слизи, как бы смачивающей частицы почвы и тем самым уменьшающей силу трения между ними и растущим корневым окончанием. В клетках чехлика, расположенных в его центральной (осевой) части, содержится много крахмальных зерен, которые способны перемещаться в клетке при изменении положения кончика корня в пространстве, на что соответствующим образом реагируют клетки и ткани меристемы, обеспечивая рост корня в прежнем направлении.
В клетках чехлика, расположенных в его центральной (осевой) части, содержится много крахмальных зерен, которые способны перемещаться в клетке при изменении положения кончика корня в пространстве, на что соответствующим образом реагируют клетки и ткани меристемы, обеспечивая рост корня в прежнем направлении.
Механизм управления направлением линейного роста корня является чрезвычайно сложным и далеко не изученным до конца. Корень изменяет направление роста под влиянием силы тяжести (положительный, отрицательный, нейтральный геотропизм), температуры окружающей среды (термотропизм), концентрации в почве химических веществ (хемотропизм), влажности почвы (гидротропизм). Изменение направления роста возможно только в случае реакции инициальных клеток и клеток в зоне растяжения, поскольку в последующих зонах клетки являются неподвижными по отношению к окружающим частицам почвы. Наиболее тесные и действенные связи корня с окружающей средой осуществляются в зоне всасывания, однако клетки, расположенные в этой зоне, уже не могут влиять на направление роста корневого окончания, поскольку они прекратили свой рост. Клетки корневого чехлика являются авангардом по отношению к остальным клеткам и тканям, вследствие этого они первыми могут улавливать изменения на пути ростового движения корня. Явление регулировки направления роста корневого окончания представляет одну из важнейших, еще не разгаданных тайн высших растений.
Корневой чехлик прикрывает группу инициальных, активно делящихся клеток. Это зона деления, на растущем корешке она визуально выделяется желтоватым цветом, обусловленным отсутствием более или менее заметных вакуолей внутри клеток. В зоне деления уже наблюдается первичная дифференциация вновь образовавшихся клеток. Наружный слой клеток представляет покровную ткань и называется периблемой, остальное скопление клеток — плеромой. Из клеток периблемы в дальнейшем формируется первичная кора, а из плеромы — стела. Протяженность зоны деления у разных древесных пород различна.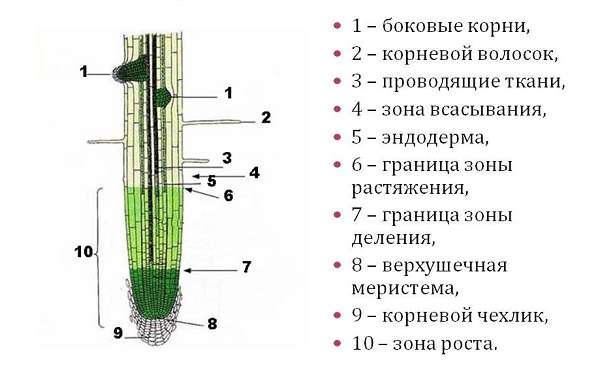 Различна она также у сосущих и ростовых корней. Меристематическая зона ростовых корней у дуба черешчатого имеет значения 250—2000 м, сосущих корней — 45—65 м. Молодые клетки, образовавшиеся в результате деления инициальных клеток и отложенные ими внутрь корневого окончания, интенсивно растут, увеличивая свои линейные размеры, растягиваясь в продольном направлении.
Различна она также у сосущих и ростовых корней. Меристематическая зона ростовых корней у дуба черешчатого имеет значения 250—2000 м, сосущих корней — 45—65 м. Молодые клетки, образовавшиеся в результате деления инициальных клеток и отложенные ими внутрь корневого окончания, интенсивно растут, увеличивая свои линейные размеры, растягиваясь в продольном направлении.
Область корневого окончания, где сосредоточено это явление, представляет зону растяжения. Визуально по сравнению с зоной деления она выделяется более светлым тоном.
В зоне растяжения происходит не только физический рост клеток. Одновременно с ростом активно протекает процесс глубокого изменения их физиологического состояния и строения, в результате чего начинается дифференциация клеток плеромы и формирование тканей, которые они образуют.
Зона растяжения сменяется (трансформируется) зоной корневых волосков, т. е. зоной всасывания. Эта зона находится на соответствующем расстоянии от инициальных клеток, однако физиологически она является прямым (неизмененным последующими превращениями) результатом их деятельности. В апексе корневого окончания самый наружный слой инициальных клеток, откладывая клетки корневого чехлика, одновременно производит клетки, которые по периферии в один слой покрывают все корневое окончание от апекса, включая зону корневых волосков. Такой покров состоит из слоя живых клеток и называется ризодермой. Ризодерма — это всасывающая ткань, клетки которой эластичны, хорошо проницаемы для воды, активно взаимодействуют с почвенным раствором. В результате этого по всей поверхности корневого окончания происходит активное всасывание воды и растворенных в ней элементов питания. Повышению интенсивности этого процесса способствует образование и функционирование корневых волосков.
Корневой волосок образуется как вырост из клеток ризодермы, представляя с ними единое целое. В процессе образования корневых волосков ядро клетки перемещается в образующийся вырост. Длина корневого волоска 0,15—2,0 мм, иногда до 3 мм, у отдельных древесных пород до 10 мм, диаметр около 10—15 мк. Потенциально каждая клетка ризодермы может образовать корневой волосок. Корневые волоски способны выделять слизь, обеспечивающую плотный контакт их с почвенными частицами. Благодаря корневым волоскам площадь всасывающей зоны может увеличиваться до 10 и более раз. Корневые волоски плотно скрепляются с почвой, поэтому их возникновение и существование становится возможным на тех участках корня, где прекращается его движение в почвенной среде. Интенсивность образования корневых волосков зависит от биологических свойств древесной породы и экологических условий, среди которых особое значение имеет влажность среды. С увеличением влажности количество корневых волосков уменьшается. Корни, произрастающие в водной среде, корневых волосков вообще не образуют. В обычных условиях на 1 см2 поверхности корня приходится в среднем от 100 до 300 (в зависимости от древесной породы) корневых волосков.
Потенциально каждая клетка ризодермы может образовать корневой волосок. Корневые волоски способны выделять слизь, обеспечивающую плотный контакт их с почвенными частицами. Благодаря корневым волоскам площадь всасывающей зоны может увеличиваться до 10 и более раз. Корневые волоски плотно скрепляются с почвой, поэтому их возникновение и существование становится возможным на тех участках корня, где прекращается его движение в почвенной среде. Интенсивность образования корневых волосков зависит от биологических свойств древесной породы и экологических условий, среди которых особое значение имеет влажность среды. С увеличением влажности количество корневых волосков уменьшается. Корни, произрастающие в водной среде, корневых волосков вообще не образуют. В обычных условиях на 1 см2 поверхности корня приходится в среднем от 100 до 300 (в зависимости от древесной породы) корневых волосков.
Например, в опытах, полученных в вегетационном домике, у саженцев акации белой насчитывалось на 1 см2 поверхности корней 520, а на корнях сосны ладанной в тех же условиях 217 корневых волосков.
Зона размещения корневых волосков на корне обычно достигает 1—2 см. Редким явлением представляется наличие корневых волосков на окончаниях корней, которые направлены вертикально в глубь почвы и достигают горизонтов с относительно стабильной повышенной влажностью (сосна, дуб и др.). При наличии микоризы корневые волоски также отсутствуют.
Продолжительность жизни корневых волосков небольшая — обычно несколько дней, иногда до 10—20 дней, отмирают они вместе с отмиранием ризодермы.
Всасывающая функция присуща всему корневому окончанию, однако максимально она проявляется только в зоне корневых волосков. Эта зона имеет решающее значение в обеспечении всего растительного организма водой и питательными элементами. Ризодерма, являясь покровной тканью корневого окончания, в функциональном физиологическом аспекте является одной из важнейших тканей организма.
Продолжительность существования ризодермы невелика — всего несколько суток и зависит от особенностей растительного организма и конкретных экологических условий. Затем ризодерма отмирает и отлущивается вместе с первичной корой корня. На участке корня после отмирания ризодермы резко снижается всасывающая активность, деятельность его ограничивается функцией проведения, транспортировки воды и питательных веществ, поступающих из зоны всасывания. Иногда эти участки корневого окончания называют зоной проведения. Распределение корневого окончания на отдельные зоны является элементом субъективным, имеющим познавательный характер и только в определенной степени с морфологической стороны характеризующим глубинные процессы развития клеток, формирования и трансформации тканей, их физиологические функции и отправления, происходящие в корневом окончании в период его роста. Не менее важным для понимания функциональной роли корня древесных растений является анализ процесса и характера дифференциации тканей, происходящего в растущем корневом окончании.
Затем ризодерма отмирает и отлущивается вместе с первичной корой корня. На участке корня после отмирания ризодермы резко снижается всасывающая активность, деятельность его ограничивается функцией проведения, транспортировки воды и питательных веществ, поступающих из зоны всасывания. Иногда эти участки корневого окончания называют зоной проведения. Распределение корневого окончания на отдельные зоны является элементом субъективным, имеющим познавательный характер и только в определенной степени с морфологической стороны характеризующим глубинные процессы развития клеток, формирования и трансформации тканей, их физиологические функции и отправления, происходящие в корневом окончании в период его роста. Не менее важным для понимания функциональной роли корня древесных растений является анализ процесса и характера дифференциации тканей, происходящего в растущем корневом окончании.
Первичная дифференциация клеток происходит сразу же в процессе их образования в результате деления инициальных клеток. От инициальных клеток, расположенных в крайней точке апикальной меристемы, образуются клетки коревого чехлика и ризодермы. Клетки, откладывающиеся при делении инициальных клеток внутрь корня, в своей совокупности представляют плерому. Однако сразу же из плеромы образуется осевой центральный цилиндр — стела. В непосредственной близости к зоне деления формируется наружный слой стелы — перицикл. Важным в биологическом аспекте является то, что клетки перицикла долго сохраняют присущую инициальным клеткам способность к новообразованиям, которая в дальнейшем проявляется при соответствующих условиях.
Клетки, находящиеся под перициклом, составляют прокамбий и быстро превращаются в проводящие ткани типа флоэмы и ксилемы. Флоэма формируется и начинает функционировать раньше ксилемы, доставляя в апекс корня пластические вещества. Несколько позднее, уже в зоне растяжения, формируются элементы ксилемы. Элементы первичной флоэмы (протофлоэма) и первичной ксилемы (протоксилема) возникают вплотную к пери цикл у.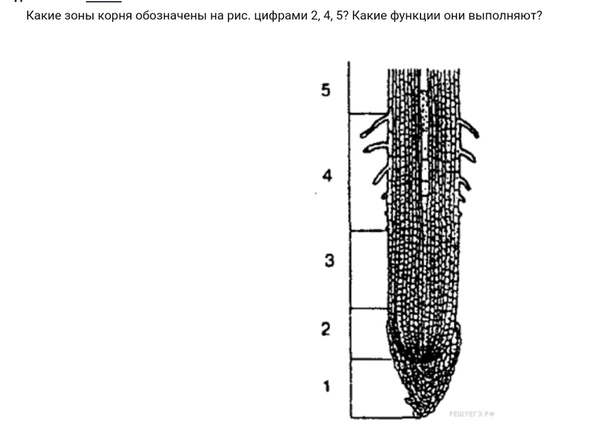 Группы их клеток первоначально чередуются друг с другом.
Группы их клеток первоначально чередуются друг с другом.
В дальнейшем развитии ксилема обгоняет флоэму, как бы вытесняя ее элементы из центральной части корня. В связи с этим в корневом окончании на соответствующем удалении от апекса расположение тканей ксилемы имеет звездчатый характер, а между лучами звезды ксилемы располагаются группы пучков флоэмы. Такое строение сохраняется недолго, оно присуще первичному строению корня.
Корневому окончанию свойственна также первичная кора. Она формируется из клеток периблемы и состоит в основном из живых паренхимных клеток с тонкими оболочками. Первичная кора покрыта слоем клеток ризодермы, снабжает их пластическими веществами и участвует в поглощении и проведении воды и питательных веществ.
Первичная кора неоднородна по структуре и функционированию ее тканей. Наружные слои ее, подстилающие ризодерму, образуют экзодерму. Экзодерма регулирует прохождение веществ от ризодермы, а после отмирания ризодермы трансформируется в защитную покровную ткань. Внутренние поверхности оболочек клеток экзодермы постепенно покрываются слоем суберина и, оставаясь живыми, теряют пропускную способность. Однако среди опробковевших (суберинизировавшихся) клеток остается часть не опробковевших, и через них в основном происходит избирательное прохождение веществ.
Внутренний слой первичной коры — эндодерма — окружает стелу. Клетки ее также постепенно покрываются суберином, однако она не становится совершенно непроницаемой благодаря наличию пропускных клеток, сохраняющих первичное строение. Схематически первичное строение корня в зоне корневых волосков представляется следующим образом: эпиблема, первичная кора, центральный цилиндр (стела).
Эпиблема (кожица) — первичная покровная ткань корня, состоящая из одного ряда сомкнутых клеток, способных образовывать корневые волоски.
Первичная кора состоит из трех достаточно хорошо различимых слоев: непосредственно под эпиблемой находится экзодерма из одного или нескольких рядов клеток, затем мезодерма, представляющая среднюю и самую большую часть коры, за мезодермой идет самый последний слой коры — эндодерма, которая непосредственно окружает центральный цилиндр.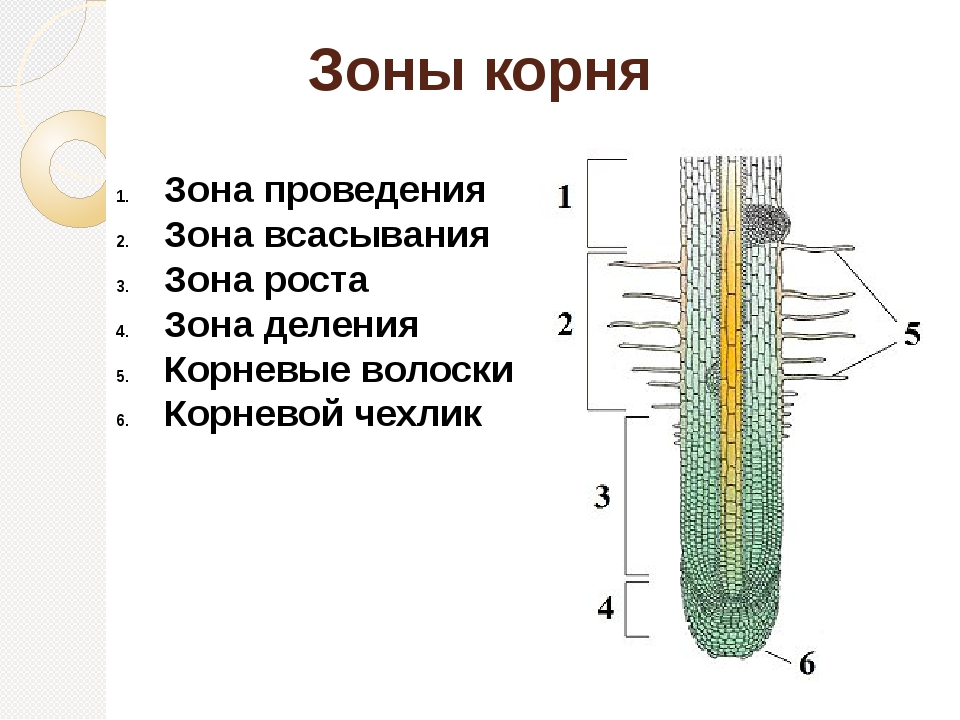
Центральный цилиндр имеет наружный слой — перицикл, примыкающий к эндодерме. Перицикл обычно представлен одним рядом клеток, однако у отдельных древесных пород он состоит из нескольких рядов. Например, в корнях ореха грецкого перицикл имеет 3—10 рядов клеток. Многослойный перицикл свойствен всем голосеменным. Центральный цилиндр включает радиальный пучок проводящей ткани, состоящей из первичных флоэмы и ксилемы, включающих элементы протофлоэмы, протоксилемы, метафлоэмы и метаксилемы.
Первичное строение корней у голосеменных и двудольных растений при развитии зародышевого корня сохраняется около 10 дней после прорастания семян и еще в фазе семядолей переходит во вторичное строение.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
Урок 14. Внутреннее строение корня. Раздел 4. Корень
Методическое пособие разработки уроков биологии 6класс
Тип урока — комбинированный
Методы: частично-поисковый, проблемного изложения, репродуктивный, объяснительно-иллюстративный.
Цель:
— осознание учащимися значимости всех обсуждаемых вопросов, умение строить свои отношения с природой и обществом на основе уважения к жизни, ко всему живому как уникальной и бесценной части биосферы;
Задачи:
Образовательные: показать множественность факторов, действующих на организмы в природе, относительность понятия «вредные и полезные факторы», многообразие жизни на планете Земля и варианты адаптаций живых существ ко всему спектру условий среды обитания.
Развивающие: развивать коммуникативные навыки, умения самостоятельно добывать знания и стимулировать свою познавательную активность; умения анализировать информацию, выделять главное в изучаемом материале.
Воспитательные:
Формирование экологической культуры на основе признания ценности жизни во всех её проявлениях и необходимости ответственного, бережного отношения к окружающей среде.
Формирование понимания ценности здорового и безопасного образа жизни
УУД
Личностные:
воспитание российской гражданской идентичности: патриотизма, любви и уважения к Отечеству, чувства гордости за свою Родину;
Формирование ответственного отношения к учению;
3) Формирование целостного мировоззрения, соответствующего современному уровню развития науки и общественной практики.
Познавательные: умение работать с различными источниками информации, преобразовывать её из одной формы в другую, сравнивать и анализировать информацию, делать выводы, готовить сообщения и презентации.
Регулятивные: умение организовать самостоятельно выполнение заданий, оценивать правильность выполнения работы, рефлексию своей деятельности.
Коммуникативные: Формирование коммуникативной компетентности в общении и сотрудничестве со сверстниками, старшими и младшими в процессе образовательной, общественно полезной, учебно-исследовательской, творческой и других видов деятельности.
Планируемые результаты
Предметные: знать — понятия «среда обитания», «экология», «экологические факторы» их влияние на живые организмы, «связи живого и неживого»;. Уметь — определять понятие «биотические факторы»; характеризовать биотические факторы, приводить примеры.
Личностные: высказывать суждения, осуществлять поиск и отбор информации; анализировать связи, сопоставлять, находить ответ на проблемный вопрос
Метапредметные:.
Умение самостоятельно планировать пути достижения целей, в том числе альтернативные, осознанно выбирать наиболее эффективные способы решения учебных и познавательных задач.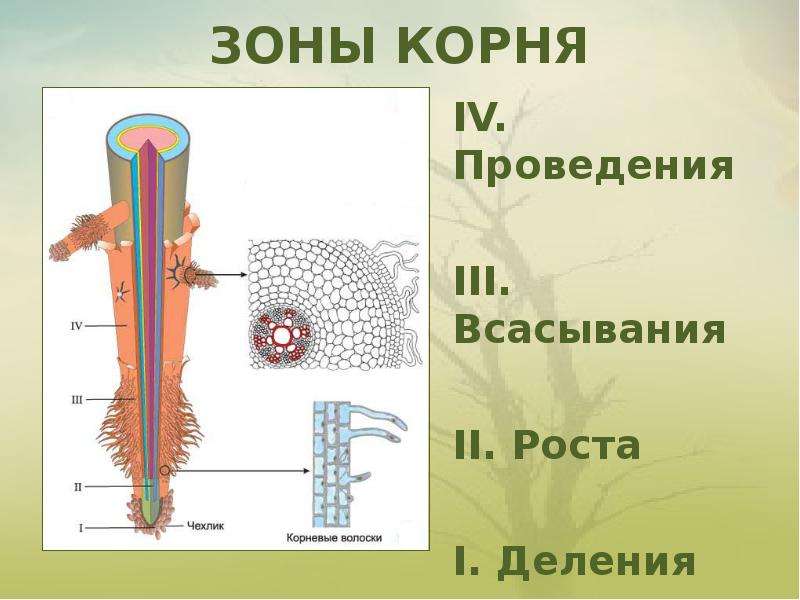
Формирование навыка смыслового чтения.
Форма организации учебной деятельности – индивидуальная, групповая
Методы обучения: наглядно-иллюстративный, объяснительно-иллюстративный, частично-поисковый, самостоятельная работа с дополнительной литературой и учебником, с ЦОР.
Приемы: анализ, синтез, умозаключение, перевод информации с одного вида в другой, обобщение.
Задачи: продолжить знакомство со строением корня, сформировать понятие о зонах корня и их функциях, раскрыть особенности строения различных зон корня на клеточном уровне; продолжить формирование понятия о тканях, познакомить с различными типами тканей на примере клеточного строения корня.
Оборудование и материалы: таблицы: «Клеточное строение корня», «Покровная ткань растений», «Образовательная ткань растений», «Проводящая ткань растений», гербарии растений, проростки гороха или пшеницы, выращенные в грубой песчаной почве (чтобы корневой чехлик был лучше заметен), лупы (на каждый стол), проростки фасоли, гороха, пшеницы, результаты опыта, доказывающего верхушечный рост корня.
Ключевые слова и понятия: корневой чехлик, зона деления, зона растяжения (зоной роста), зона всасывания (зона поглощения), корневые волоски, кора корня, зона проведения, древесина, луб, сосуды, камбий, кора, пробка.
Ход урока
Актуализация знаний
Рассказ с ошибками
(Рассказ с ошибками раздается в печатном виде каждому из учеников или один экземпляр на парту. Учащимся дается около 5 мин, чтобы прочитать его и исправить все ошибки. Затем каждый из учеников читает по одному предложению, называет и исправляет ошибки.)
Найдите и исправьте биологические ошибки в тексте.
Корень — генеративный орган растения. Все растения на земле имеют корни. Основная функция корней — это укрепление растения в почве, обеспечение его водой и минеральными веществами, а также кислородом. Корни под землей дышат кислородом, содержащимся в воздухе, находящимся между частичками почвы. Вся совокупность корней одного растения называется корневой системой. В корневой системе всех растений можно выделить главный корень. В процессе прорастания семени главный корень появляется первым. На главном корне развиваются придаточные корни, а от стебля растут боковые. Корневая система, в которой четко виден главный корень, называется стержневой. Стержневая корневая система характерна для однодольных растений. Горох, фасоль, бобы имеют стержневую корневую систему. Корневая система, в которой главный корень не отличается от прочих, называется мочковатой. Мочковатая корневая система развивается у растений, образующих корнеплоды.
Изучение нового материала
Рассказ учителя с элементами беседы
Давайте рассмотрим строение корня более подробно. Главный, боковые и придаточные корни растения имеют сходное строение. Для того чтобы разобраться с особенностями строения корня растения, рассмотрим проростки гороха или фасоли.
(Проростки и лупы желательно раздать на каждый стол.)
Рассмотрите кончик корня фасоли через лупу. Обратите внимание, что он как бы одет в маленький колпачок — корневой чехлик, который защищает верхушку корня от твердых частичек почвы, которые могут ее поранить.
Если рассмотреть срез корня под микроскопом, то хорошо видно, что корневой чехлик состоит из нескольких слоев клеток. Наружные клетки постоянно погибают и слущиваются, после чего превращаются в слизь, которая уменьшает трение.
Но почему же корневой чехлик полностью не исчезает в конце концов? Все дело в том, что под корневым чехликом расположены клетки, которые активно делятся. Эта часть корня так и называется — зона деления.
Как вы думаете, какой тканью представлена зона роста?
(Ответы учащихся.)
Она представлена образовательной тканью. Наружные клетки зоны деления постоянно обновляют корневой чехлик. Стенки их утолщаются и приобретают прочность и способность защищать нежные, молодые клетки зоны деления корня. Если растение растет на грубых, песчаных почвах, корневой чехлик становится более мощным. А у растения, обитающего на мягких, суглинистых почвах, он значительно меньше.
Внутренние клетки зоны деления как бы откладываются вверх. Они начинают активно расти. (Именно из них в дальнейшем будут строиться все остальные части корня.) Эта часть корня называется зоной растяжения, или зоной роста. Она небольшая, всего 3—9 мм в длину. Молодые клетки здесь постоянно растут, удлиняются, тем самым увеличивая длину корня. Растягиваясь, зона роста проталкивает вглубь зону деления, защищенную корневым чехликом.
Доказать, что корень растет верхушкой, очень просто.
(Демонстрируются условия опыта и его результат. Можно также дать задание ученикам, интересующимся биологией, проделать эксперимент дома, а на основе выводов подготовить доклад.)
Для этого необходимо взять проросток фасоли, бобов или гороха. При помощи туши нанести на главный корень метки на равном расстоянии друг от друга. И посмотреть на проросток на следующий день. На участке корня, расположенном ближе к основанию, расстояние между метками не изменилось, а на верхушечной части корня — увеличилось.
О чем это говорит? (Это доказывает, что корень растения растет верхушкой.)
Выше зоны растяжения расположена зона всасывания, или зона поглощения. Рассмотрите проростки фасоли, лежащие на ваших столах. Обратите внимание, что часть корня как бы покрыта легким пушком. Это корневые волоски — особые выросты наружных клеток зоны всасывания. Их длина обычно составляет 0,1—2 мм, а у некоторых растений доходит до 10 мм. Они плотно прилегают к частицам почвы и всасывают воду с растворенными в ней минеральными веществами. Кроме того, корневые волоски выделяют особую слизь, которая помогает растворять минеральную составляющую почвы для дальнейшего ее поглощения. Корневые волоски работают как насосы, всасывая питательный раствор, отсюда и название зоны. Срок жизни корневых волосков небольшой, чаще всего несколько дней, после чего они отмирают. Новые корневые волоски появляются на молодых клетках на границе зон растяжения и всасывания. Таким образом зона всасывания постоянно перемещается в почве, оставаясь ближе к концу корня.
Как вы считаете, какой в этом смысл? (В результате зона всасывания использует для питания все новые участки почвы.)
Корневые волоски очень нежные, и их легко повредить. В этом случае растение не сможет добывать из почвы минеральные вещества и воду.
Как вы думаете, что необходимо предпринимать при пересадке растений для того, чтобы не повреждать корневые волоски? (Ответы учащихся.)
Лучше всего пересаживать растения с комочком почвы, в которой они росли раньше, так как при этом не повреждаются корневые волоски.
Но какова основная роль корня, кроме укрепления растения в земле? (Минеральное питание всего растения.)
Действительно, вода и минеральные вещества от корня доставляются ко всему растению.
Но как это все происходит?
Если рассмотреть под микроскопом поперечный срез корня зоны всасывания, снаружи мы увидим один слой клеток, образующих корневые волоски. Под этим слоем расположено несколько слоев живых клеток с тонкими стенками и крупными межклетниками. Это кора корня.
Какой тканью представлена кора корня? (Ответы учащихся.)
Это покровная ткань. Середина корня практически полностью состоит из проводящей ткани, представленной двумя типами сосудов. Сосуды — длинные вытянутые полые клетки с толстыми оболочками.
Древесина — ткань, по сосудам которой вода с растворенными в ней минеральными веществами поднимается от корня растения к стеблю и листьям. Сосуды древесины представляют собой длинные полые трубки, составленные из мертвых клеток с одревесневшими стенками. Но корню необходимы органические вещества.
Откуда же они берутся? (Ответы учащихся.)
Вокруг древесины располагается еще одна проводящая ткань корня — луб. По сосудам луба органические вещества перемещаются от стебля и листьев к клеткам корня.
Участок корня, расположенный выше зоны всасывания, называется зоной проведения. Это самая длинная часть корня. Проводящая ткань здесь хорошо сформирована. Снаружи корень покрыт прочными клетками коры, под которой находится пробка — несколько слоев мертвых клеток. Живое содержимое этих клеток отмирает, оболочки утолщаются, и они становятся непроницаемы для воды и воздуха. Межклетники пробки заполнены воздухом, что является прекрасной защитой от перепадов температур, иссушения, механических повреждений. Корень в этой части не может поглощать воду и минеральные вещества, поэтому основной функцией является проведение растворенных минеральных веществ и воды.
Кроме роста в длину, корень увеличивается и в ширину. Это происходит благодаря клеткам особой образовательной ткани — камбия. Камбий — слой клеток, способных делиться и образовывать клетки других тканей. Камбий расположен между древесиной и лубом. Благодаря клеткам камбия корень постепенно утолщается за счет древесины и луба, т. е. объем древесины и луба постоянно увеличивается, а значит, возрастает проводящая способность корня.
Различные зоны корня выполняют различные функции, но все они находятся в тесном взаимодействии друг с другом, что обеспечивает нормальную жизнедеятельность всего растения.
Закрепление знаний и умений
Рассмотрите на рисунке в учебнике изображение продольного разреза молодого корня. Запишите в таблицу, какие зоны есть у корня и какие функции они выполняют.
Необходимо обратить внимание учащихся на то, что зоны корня, описанные в учебнике первыми, на рисунке располагаются ниже прочих. Поэтому заполнение таблицы следует начать со схематичного рисунка и следить за тем, чтобы текст примерно соответствовал изображению.)
Зона корня | Рисунок | Строение | Функции |
Фронтальный опрос
Ответьте на вопросы.
Что такое корневой чехлик, какова его функция?
В какой части корня расположена зона роста?
Опишите опыт, доказывающий, что корень растет верхушкой.
Как визуально отличить зону всасывания?
Приведите примеры тканей, представленных в корне растения.
Какие части корня представлены образовательной тканью?
Какова основная функция древесины и луба?
За счет чего корень растет в ширину?
Какие свойства коры и пробки помогают защитить корень от перепадов температур, иссушения, механических повреждений?
Подведение итогов урока
Творческое задание. Составить кроссворд по теме «Корень», оформить его на отдельном листе.
Задания для учеников, интересующихся биологией.
Попробовать вырастить проростки фасоли, гороха, пшеницы или другого растения в песчаной и глинистой почве и в стакане с водой и минеральной подкормкой. Через 10—14 дней аккуратно выкопать растения, промыть корневую систему и рассмотреть через лупу с 10-кратным увеличением размер корневого чехлика. Сделать вывод о том, как условия произрастания сказались на размере корневого чехлика.
Попросить у учителя биологии микроскоп и готовый микропрепарат «Митоз в кончике коня». Рассмотреть микропрепарат. Зарисовать увиденное. По каким признакам можно определить, что клетки здесь молодые и постоянно делятся?
«Внутреннее строение корня«
Корень. Клеточное строение корня
Зоны корня | Биология 6 класс #22 | Инфоурок
Ресурсы:
И.Н. Пономарёва, О.А. Корнилова, В.С. Кучменко Биология : 6 класс : учебник для учащихся общеобразовательных учреждений
Серебрякова Т.И., Еленевский А. Г., Гуленкова М. А. и др. Биология. Растения, Бактерии, Грибы, Лишайники. Пробный учебник 6—7 классов средней школы
Н.В. Преображенская Рабочая тетрадь по биологии к учебнику В В. Пасечника «Биология 6 класс. Бактерии, грибы, растения»
В.В. Пасечника. Пособие для учителей общеобразовательных учреждений Уроки биологии. 5—6 классы
Калинина А.А. Поурочные разработки по биологии 6класс
Вахрушев А.А., Родыгина О.А., Ловягин С.Н. Проверочные и контрольные работы к
учебник «Биология», 6-й класс
Биоуроки http://biouroki.ru/material/lab/2.html
Сайт YouTube: https://www.youtube.com /
Хостинг презентаций
— http://ppt4web.ru/nachalnaja-shkola/prezentacija-k-uroku-okruzhajushhego-mira-vo-klasse-chto-takoe-ehkonomika.html
Зоны молодого корня
Определение 1
Корень – это осевой орган, который обладает свойством неограниченного роста и положительным геотропизмом.
Функции корней
Данный орган растения обладает рядом жизненно важных функций, среди них:
- укрепление растения в почве и удержание наземного побега;
- минеральное питание растений;
- проведение и продвижение веществ по организму растения;
- место запасания питательных веществ;
- позволяет реализовать механизм вегетативного размножения.
Корни делятся на главные, боковые, придаточные. Главный корень развивается из зародышевого корешка и обладает верхушечной меристемой.
Боковые корни развиваются на любом другом корне образуя несколько порядков ветвления. Придаточные корни развиваются из стеблей, листьев, старых корней. Они появляются за счет развития вторичных меристем.
Зоны молодого корня и их функции
Определение 2
Зоны – это разные участки корня по длине, которые выполняют различные функции и характеризуются морфологическими особенностями. В молодом кроне обычно развиваются несколько обособленных, но взаимосвязанных между собой зон.
Зона деления представлена верхушкой корня длиной 1-2 мм. Здесь находится апикальная меристема и происходит обновление клеточных структур. Эта зона защищается корневым чехликом и образуется живыми клетками. В данной зоне также часто содержатся зерна крахмала, продуцирующие слизь.
Зона роста или растяжения имеет протяженность несколько миллиметров. В этой зоне клеточные деления отсутствуют, клетки тянуться за счет вакуолей.
Зона всасывания или корневых волосков составляет несколько сантиметров. В данном случае происходит дифференцировка и специализация клеток. Корневые волоски существуют максимально до 20 дней. При этом зона всасывания всегда удалена от кончика корня на одно и тоже расстояние с небольшими разночтениями. Отмершие клетки этой зоны довольно быстро сменяются новыми.
Что касается зоны проведения, то она находится выше зоны всасывания. Внутри этой зоны вода и минеральные соли извлекаются из почвы и передвигаются от корня к стелю и листу. В этой зоне за счет образования боковых корней происходит корневое ветвление.
Первичное строение корня формируется за счет первичной меристемы, что характерно для всех корней растения. В зоне всасывания различают три части: эпиблему, кору, осевой цилиндр. У плаунов и хвощей такое строение корня сохраняется в течение всей жизни.
Вторичное строение корня формируется за счет вторичных меристем. Процесс вторичных изменений запускается с проявления прослоек камбия между флоэмой и ксилемой. Сначала прослойки камбия разобщены, затем образуют сплошной слой. После клеточного деления радиальная симметрия характерна для первичного корневого строения.
В перицикле возникает пробковый камбий и феллоген. Наружу откладываются слои вторичной покровной ткани или пробки. Первичная кора отмирает и слущивается.
В зависимости от состава корней выделяют несколько типов корневых систем:
- мочковатая корневая система, которая образована боковыми и придаточными корнями. При этом главный корень развит слабо и очень рано прекращает свой рост. Такой тип корневой системы характерен для однодольных растений;
- стержневая корневая система характерна для двудольных растений и отличается наличием главного корня, слабым развитием других типов корней.
Строение молодого корня существенно влияет на его физиологию. Любой корень не обладает неограниченным ростом. Он растет верхушкой, на которой располагается апикальная меристема.
Поглощение корнем воды и ее передвижение к наземным органам является одной из важнейших функций корня. Такая функция возникла у растений в связи с выходом на сушу. Зоны корня приспособлены к образу жизни растения и поглощению воды, а также питательных веществ. Вода и минеральные вещества поглощаются независимо друг от друга, поскольку эти процессы основаны на различных механизмах работы клеток зон корня.
Вода поступает в растение согласно законам осмоса, благодаря корневым волоскам, а именно вакуолям, находящимся внутри них.
Минеральные вещества поступают в растения благодаря вертикальному транспорту веществ. Вертикальное перемещение воды с растворенными минеральными веществами происходит по мёртвым клеткам, не способным толкать воду к листу. Такой процесс возможен за счет наличия такого феномена как гуттация.
Определение 3
Гуттация – это процесс выделения воды у неповрежденного растения через водяные устьица в покровной ткани растения.
Гуттация является достаточно важным процессом в жизни растения.
Корни могут выполнять различные функции, образуя так называемые видоизменения корней. Например, запасающие корни выполняют функцию накопления питательных веществ. У таких корней присутствует зона запасающей паренхимы, находящейся в первичной и вторичной коре. Запасающие корни могут выглядеть как корневые клубни и корнеплоды.
Растения, которые растут на болотах и образуют корни, которые растут вверх и называются дыхательными корнями или пневматофорами. Для них хорошо развита воздухоносная паренхима, что значительно повышает уровень кислорода, поступающего в растения. Иногда воздушные корни образуют на поверхности своеобразный слой губчатой ткани – веламен.
Для того, чтобы поддерживать строение корня в любой зоне, очень часто применяют те или иные удобрения. К ним относят хлористый аммоний, кальциевую селитру, синтетическую мочевину и сульфат аммония, которые усиливают рост корней и стебля растительного организма.
Фосфорные удобрения позволяют продлить цветение и ускорить созревание плода. Калийные удобрения усиливают рост подземных органов, а именно растений и корней, луковиц, клубней. К основным органическим удобрениям относится навоз, торф, фекалии, помет, компост. Такие удобрения обладают преимуществом потому, что имеют такое качество как комплексность. В них соединены минеральные соли, органические вещества, которые благотворно влияют на развитие корня в целом и любой его зоны в частности.
«Строение и функции зон корня»
Тип – урок-путешествие.
Форма – закрепление пройденного материала и изучение нового.
Цели урока для учителя.
- Закрепить знания о строение корневой системы, изучить клеточное строение и функции корня.
- Развить логическое мышление, закрепить навыки работы с книгой.
- Развить коммуникативное общение, умение оценивать свои успехи и успехи других учащихся.
Цели урока для учащихся.
- Знать клеточное строение корня и его функции.
- Уметь определять ткани корня растения.
Оборудование.
- таблицы “Зоны корня”, таблицы с разными видами тканей,
- “Карта путешествия по корневой системе”,
- микроскопы с готовыми препаратами тканей растений,
- “Схема основных тканей растений”,
- оценочные листы на каждой парте,
- алгоритмы заполнения таблицы,
- карточками с заданиями по новому материалу,
- тесты для проверки полученных знаний,
- гербарии растений.
Рисунок 1. Карта “Путешествие по корневой системе”
Ход урока
1. Организационный момент. Приветствие
2. Сообщение цели урока
Учитель. Ребята, согласны ли вы провести урок в форме путешествия? Ведь любое путешествие – это познание нового и интересного. Мы с вами уже знаем немного о корне. Ведь знания помогают нам правильно выращивать растения. Каждому из нас приходится заниматься растениями, и каждый должен знать, что сила растения кроется в корне. Имея ещё более глубокие знания о корне, мы сможем более эффективно ухаживать за ними. Что мы ещё недостаточно хорошо знаем о корне? Какие новые сведения о корне мы можем получить на уроке?
(Эту часть можно провести в форме беседы. Учитель должен подвести детей к цели урока, чтобы цель назвали сами учащиеся).
3. Основная часть урока
Запись в тетради темы урока.
Учитель. Узнать больше о корне мы сможем, отправившись в путешествие при помощи карты “Путешествие по корневой системе (смотри рисунок 1).
На всем пути вам будут встречаться различные препятствия, вам необходимо их преодолеть и набрать как можно больше баллов, чтобы получить звание “Великого путешественника по корневой системе”. Правильно оценить свою работу и работу твоих товарищей поможет оценочный лист (смотри приложение 1).
1 этап. Путешествие по корневой системе определенного растения.
Учащиеся выполняют лабораторную работу “Стержневая и мочковатая корневая системы”. По гербарию определяют тип и особенности корневой системы, данные заносят в таблицу [1, с. 91].
Таблица №1. Стержневая и мочковатая корневая системы.
| Название растения | Тип корневой системы | Особенности строения корневой системы |
| Одуванчик лекарственный | Стержневая корневая система | Хорошо виден главный корень |
Слабые учащиеся получают гербарии знакомых растений, сильные – гербарии растений, не произрастающих в данной местности.
Выполнение – 3-5 мин. Учащиеся проверяют выполнение работы у соседа по парте и выставляют оценку при помощи оценочного листа
Учитель. Кто желает поделиться своими впечатлениями? (Ответ одного учащегося по определению типа корневой системы).
Привал.
Учитель. Для дальнейшего путешествия нам необходимо отдохнуть, сделать привал. На привале мы вспомним интересные данные о тканях, которые нам понадобятся для дальнейшего путешествия.
Учащиеся слушают сообщение ученика “Ткани растения”.
Учитель. Итак, что такое ткань? Как вы думаете, какие ткани мы не встретим в корне?
Учащиеся дают определение “ткани”, говорят, что в корне нельзя встретить фотосинтезирующую ткань.
Учитель. Молодцы! Можно смело отправляться в путь. Не забывайте оценивать свою работу.
2 этап. Путешествие внутри корня.
Учитель. Сейчас мы спустимся вниз по корню. Как вы думаете, одинаковые ли клетки в корне встречаются нам на пути? Изучим их. Для того, чтобы быстрее получить необходимую и интересную информацию, мы разделимся на группы. Каждая группа будет изучать свой участок корня, его ткани и функции. Вам также предлагается посмотреть препараты тканей растений, выяснить их строение.
- 1 группа (сильные учащиеся), изучают зоны корневого чехлика, деления и растяжения [1, с. 92-93].
- 2 группа (средние по способностям учащиеся), изучают зону всасывания [1, с. 93-94].
- 3 группа (средние по способностям учащиеся), изучают зону проведения [1, с. 94-95].
- 4 группа (слабые по способностям учащиеся), расшифровывают слова. Необходимо вставить пропущенные гласные буквы в слова данной темы при помощи учебника (смотри приложение 2).
Данные заносятся в таблицу.
Таблица №2. Клеточное строение корня
Зоны корня |
Ткани |
Особенности строения клеток |
Функции |
| 1. Корневой чехлик | Покровная | Клетки мелкие с толстой оболочкой | Защита от повреждения |
| 2. Зона деления | Образовательная | Клетки мелкие с тонкой оболочкой | Рост корня |
| 3. Зона растяжения | Образовательная | Вытянутые клетки с тонкой оболочкой | Рост корня |
| 4. Зона всасывания | Всасывающая | Корневой волосок | Всасывание растворенных веществ |
| 5. Зона проведения | Проводящая | Клетки вытянутые (сосуды, ситовидные трубки) | Перемещение веществ |
| Все зоны | Механическая | Клетки с толстой оболочкой | Опора органа |
2 и 3 группа получают алгоритм заполнения таблицы (смотри приложение 3).
Выполнение – 10 мин.
Учитель. Все справились со своим заданием. Сейчас нам необходимо собрать данные о клеточном строении корня в единую картину, также вам необходимо оценить каждого учащегося своей группы. (Выступление одного человека от группы).
Учитель. Хорошо! Прекрасно справились и с этим задание. Мы исследовали клеточное строение корня, можно возвращаться назад, но корень не хочет выпускать нас. Мы должны выполнить его условия – ответить на его вопросы.
3 этап. Привал. Закрепление изученного материала.
Учитель. Перед выполнением необходимо устроить привал, собраться с силами, мыслями. Попробуем вместе ответить на несколько вопросов. Не забывайте оценивать свою работу.
Вопросы для закрепления нового материала.
- Сколько всего зон имеет корень? Назовите их.
- Сколько разных видов тканей входит в состав корня? Назовите и покажите по таблице.
- Дайте определение ткани. Каких тканей нет в корне?
- Почему растения при пересадке болеют?
- У семени фасоли, которое проращивалось на свету, части побега вскоре стали зелеными, корень сохранил свою первоначальную окраску. Почему так произошло? Выскажите свои предположения.
Учитель. Все собрались с мыслями, силами для преодоления последнего препятствия? Хорошо! Сейчас каждый попытается ответить на вопросы корня.
4 этап. Выполнение бланкового теста по двум дифференцированным вариантам.
(Время выполнения 6 мин, смотри приложение 4).
Самопроверка теста при помощи таблицы ответов (смотри приложение 5).
4. Подведение итогов
Вот и закончилось наше путешествие. Что мы узнали?
- Корень имеет несколько зон, выполняющих несколько функций.
- Функции корня зависят от клеток корня.
Сейчас подсчитайте, пожалуйста, общее количество баллов.
У кого получилось 20 баллов? Молодцы, вам присваивается звание “Великий путешественник по корневой системе”. Вы будете нашими проводниками в следующих путешествиях по корневой системе, корню.
Теперь найдем свою оценку за урок (количество баллов разделите на четыре).
5. Домашнее задание
Учитель. Вам просто необходимо поделиться своими впечатлениями о путешествии. Свои впечатления вы можете отобразить в кроссворде, синквейне, рисунках, можно подобрать пословицы, загадки о корне.
Литература
- Пасечник В.В. Биология. 6кл. Бактерии, грибы, растения. – М.: Дрофа, 1997.
6 класс. 22. Зоны корня
Тема: Зоны корня.
Ход урока
- Организационный момент. Постановка целей урока.
Сегодня мы с вами продолжаем дальше изучать корень.
Тема нашего урока: «Зоны корня».
ПАУЗА В ВИДЕО
Слайд 2 [Вставить анимацию: тема урока.]
- Актуализация знаний.
Давайте вспомним:
Какие функции выполняет корень?
Слайд 3 [Вставить анимацию: вопрос]
ПАУЗА В ВИДЕО
Функции корня.
1.Опорная. Любое растение удерживается в почве благодаря хорошо разветвлённым корням.
2.Питающая. С помощью корней растения получают воду и минеральные соли. Какие функции выполняет корень?
3. Запасающая. В корнях многих растений откладываются про запас питательные вещества.
Слайд 4 [Вставить анимацию: функции корня.]
Показать функции корня.
Дайте определение трем типам корней
Главный корень-это…
Боковые корни — это…
Придаточные корни — это…
ПАУЗА В ВИДЕО
Слайд 5 Вставить анимацию: три определения…]
Выделяют 3 типа корней:
1. Главный — развивающийся из зародышевого корешка.
2. Боковые-отходящие от главного корня.
3. Придаточные- образовательные на нижней части стебля.
Слайд 6 [Вставить анимацию: поочередное появление трех определений]
Показать определения.
3.Изучение нового материала.
Сегодня мы проведем урок в форме путешествия. Ведь любое путешествие – это познание нового и интересного.
Сила растения кроется в корне.
Что мы ещё недостаточно хорошо знаем о корне? Какие новые сведения о корне мы можем получить на уроке?
Слайд 7 [Вставить анимацию: вопросы]
ПАУЗА В ВИДЕО
Предположительные ответы учащихся.
Когда мы изучали прорастание семян, вы, наверное, заметили, что корень проростка постоянно растет. Причем растет он своей верхушкой внизу. Такой тип роста называется верхушечным.
Слайд 7 [Вставить анимацию: изображение зон корня.]
По мере рассказа текста показывать зоны корня, соответствующие тексту.
Под лупой, если рассмотреть молодой неповрежденный корень. То видим, что он белый, ведь мы проращивали семена в банке, а не в земле.
На «хвосте» темнее остальной части корня.
Кончик корня «одет в колпачок» это корневой чехлик. Под ним спрятана зона деления корня, которая состоит из молоденьких клеток образовательной ткани. Верхушка корня покрыта корневым чехликом, предохраняющим клетки зоны деления при похождении корня сквозь почву.
Над чехликом расположен гладкий участок корня-зона роста. Именно благодаря этой своей части корень интенсивно растет в длину. Почему же рост происходит именно здесь? Так ведь в зоне деления постоянно образуются новые клетки. Они все время вытесняют вверх клетки, которые образовались раньше.
Слайд 8 [Вставить анимацию: изображение зон роста и деления корня.]
Мы поднимаемся по корню выше зоны роста. Это что еще такое? Мы запутались в каких то нитках и веревках! Куда мы попали?
Слайд 9 [Вставить анимацию: изображение корневых волосков.]
Так, ясно, это корневые волоски -тончайшие выросты клеток наружного слоя молодого корня.
Слайд 10 [Вставить анимацию: изображение зона всасывания.]
Теперь мы находимся в зоне всасывания. Корневые волоски проникают между самыми маленькими комочками почвы и плотно прилипают к ним. Эти волосы всасывают находящиеся в почве водные растворы минеральных веществ. Вот почему этот участок корня называется зоной всасывания.
Наконец-то мы добрались до этой зоны проведения. На поверхности ничего особенного не видно, вся работа идет внутри. Конечно, ведь снаружи корень в этой зоне покрыт слоем коры. Толстая кора не может поглощать воду из почвы. Здесь в корень проводят воду, полученную в зоне всасывания, к верхним надземным частям растения. Зона проведения постоянно удлиняется по мере роста корня. В старых корнях деревьев и многолетних трав основную часть длины составляет именно зона проведения.
Слайд 11 [Вставить анимацию: изображение зоны проведения.]
Давайте теперь разберем, какие ткани входят в каждую зону корня, и какие функции они там выполняют. Рассмотрим таблицу.
|
Зона корня |
Рабочая ткань |
Особенности участка корня и его клеток |
Функции |
|
Корневой й чехли |
Покровная ткань |
Живые клетки, которые постоянно обновляются. Клетки выделяют слизь, она покрывает поверхность молодого корня. |
-защищает зону деления, -облегчает продвижение корня в почве, -обеспечивает ориентацию корня в пространстве. |
|
Зона деления |
Образовательая ткань — меристема |
Клетки зоны деления тонкостенные и заполнены цитоплазмой, вакуоли отсутствуют. Зону деления можно отличить на живом корешке по желтоватой окраске, длина её около 1 мм. |
Обеспечивает рост корня. |
|
Зона роста |
Образовательная ткань |
Клетки зоны роста уже не делятся, но способны растягиваться в продольном направлении, проталкивая корневое окончание вглубь почвы. |
Обеспечивает рост корня растяжением. В пределах зоны роста происходит разделение клеток на ткани. Закладываются элементы первичной ксилемы. |
|
Зона всасывания (зона корневых волосков) |
Покровная ткань |
Длина зоны от нескольких миллиметров до нескольких сантиметров. В отличие от зоны роста участки этой зоны уже не смещаются относительно частиц почвы. Корневые волоски — это выросты клеток поверхностной ткани поглощающей зоны корня растения.Они содержат слой протоплазмы, ядро, крупную вакуоль; их тонкие, легко проницаемые для воды, оболочки плотно склеиваются с комочками почвы. Корневые волоски выделяют в почву различные вещества. Длина варьируется у разных видов растений от 0,06 до 10 мм. С увеличением влажности почвы образование замедляется; не образуются они и в очень сухой почве. По прошествии определённого времени корневой волосок отмирает. Продолжительность его жизни не превышает 10-20 дней |
Участвует в поглощении воды и минеральных веществ. Основную массу воды и питательных веществ молодые корни всасывают с помощью корневых волосков. |
|
Зона проведения |
Проводящая ткань |
В состав проводящих тканей этой зоны корня входят:
|
Проведение поглощенных веществ в надземную часть |
Слайд 12 [Вставить анимацию: таблица.]
Таблицу заполнять по мере комментирования таблицы.
4.Физкультурная минутка.
|
Раз — подняться, потянуться |
|
Слайд 13 [Вставить анимацию: физкультминутки]
Давайте еще раз, разложим все по полочкам. Какие зоны можно выделить в корне, и за какие функции отвечает каждая зона.
Корень
Зоны корня
Корневой чехлик
Зона деления Зона роста Зона всасывания Зона проведения
Рост корня, Рост корня Всасывание воды Проведение воды
развитие развитие и мин в-в и мин в-в к других зон других зон верхним органам
Слайд 14 [Вставить анимацию: схема «Зоны корня и их функции».]
Одна зона переходит в другую постепенно, резких границ между ними нет.
Основная функция корня – питание растения веществами, всасываемыми из почвы. Корень осуществляет минеральное питание растения.
Минеральное питание – это поглощение корнями растений необходимых элементов из почвы, их передвижение и усвоение растениями. Большинство питательных веществ доставляется в побеги растений в виде минеральных растворов. Проводящая система корней доводит питательные вещества ко всем тканям.
Для нормального роста и развития растениям необходимы многие элементы, но прежде всего – азот, фосфор и калий.
Азот необходим всему живому для нормального роста и развития. При недостатке азота растение отстает в развитии, листья желтеют и опадают, боковые побеги не развиваются.
Фосфор обеспечивает образование клеточной оболочки, способствует накоплению углеводов в плодах и масла в семенах.
Калий – обеспечивает развитие опорной ткани растения, нормальное развитие корней и корнеплодов и отложение питательных веществ в запас.
Слайд 14 [Вставить анимацию: подобрать изображения для определения минеральное питание.]
Для улучшения роста и развития растений в почву вносят удобрения, обогащающие почву теми или иными веществами.
Органические удобрения – вещества растительного и животного происхождения: навоз, торф, птичий помет и т.д.
Слайд 15 [Вставить анимацию: подобрать изображение для определения органические удобрения.]
Минеральные удобрения – специальные комплексы, состоящие из одного либо нескольких минеральных компонентов.
Слайд 16 [Вставить анимацию: подобрать изображение для определения минеральные удобрения.]
5.Закрепление знаний.
1.Первым при прорастании семени появляется корень:
а) главный в) придаточный
б) боковой г) главный или боковой
ПАУЗА В ВИДЕО
2.Мочковатая корневая система характерна для растении:
а) пшеницы и ржи в) лопуха и одуванчика
б) ржи и лопуха г) одуванчика и подорожника
ПАУЗА В ВИДЕО
3.Стержневая корневая система характерна для растений:
а) одуванчика и подорожника в) лопуха и одуванчика
б) подорожника и лопуха г) все ответы верны
ПАУЗА В ВИДЕО
4.Функции корня:
а) опорная в)запасающая
б)транспортная г) все ответы верны
ПАУЗА В ВИДЕО
Слайд 17 [Вставить анимацию: поочередное появление вопросов и вариантов ответов.]
Правильный ответ показывать после прочтения вопросов и вариантов ответов. Правильный ответ выделен жирным шрифтом.
Сформулируйте 3 вопроса по сегодняшней теме.
Слайд 18 [Вставить анимацию.]
8. Подведение итогов.
Спасибо вам за урок.
-До свидания!
Слайд 19 [Вставить анимацию.]
9.14: Корни — Биология LibreTexts
Теперь это серьезные корни. Но что такое корни?
Есть стержневые и волокнистые корни, первичные и вторичные корни. И они, кажется, всегда знают, куда расти. Корни — это особые органы растений. Как и почему?
Корни
У растений есть специализированные органы, которые помогают им выживать и воспроизводиться в самых разнообразных средах обитания. Основные органы большинства растений включают корни, стебли и листья.
Корни являются важными органами всех сосудистых растений.Большинство сосудистых растений имеют два типа корней: первичные корни , , растущие вниз, и вторичные корни , , ответвляющиеся в стороны. Вместе все корни растения составляют корневую систему .
Корневые системы
У растений есть два основных типа корневой системы: стержневые и волокнистые корневые системы. Оба показаны на рис. ниже.
- Системы стержневого корня имеют один толстый первичный корень, называемый стержневым корнем , с более мелкими вторичными корнями, растущими по бокам.Главный корень может проникать на глубину до 60 метров (почти 200 футов) под поверхность земли. Он может прокладывать очень глубокие источники воды и хранить много еды, чтобы помочь растению пережить засуху и другие экстремальные экологические явления. Главный корень также очень надежно закрепляет растение в земле.
- Волокнистая корневая система имеет много маленьких ветвящихся корней, называемых волокнистыми корнями , но не имеет большого первичного корня. Огромное количество нитевидных корней увеличивает площадь поверхности для поглощения воды и минералов, но волокнистые корни менее надежно закрепляют растение.
Одуванчики имеют систему стержневого корня; травы имеют мочковатую корневую систему.
Структура и функции корня
Как показано ниже на рис. , верхушка корня называется корневой крышкой . Он состоит из специализированных клеток, которые помогают регулировать первичный рост корня на кончике. Над корневой крышкой расположена первичная меристема, в которой происходит рост в длину.
Корень — сложный орган, состоящий из нескольких типов тканей.Какова функция каждого типа ткани?
Выше меристемы остальная часть корня покрыта одним слоем эпидермальных клеток. Эти клетки могут иметь корневых волосков , которые увеличивают площадь поверхности для поглощения воды и минералов из почвы. Под эпидермисом находится измельченная ткань, которая может быть заполнена накопленным крахмалом. Связки сосудистых тканей образуют центр корня. Восковые слои обеспечивают водонепроницаемость сосудистых тканей, чтобы они не протекали, что делает их более эффективными при переносе жидкостей.Вторичная меристема расположена внутри и вокруг сосудистых тканей. Вот где происходит рост толщины.
Структура корней помогает им выполнять свои основные функции. Что делают корни? У них есть три основные задачи: поглощение воды и минералов, закрепление и поддержка растений и хранение пищи.
- Поглощение воды и минералов: тонкостенные клетки эпидермиса и корневые волоски хорошо подходят для поглощения воды и растворенных минералов из почвы. Корни многих растений также имеют микоризную связь с грибами для лучшего усвоения.
- Крепление и поддержка растений: системы корней помогают закрепить растения на земле, позволяя растениям расти высокими, не опрокидываясь. Плотное покрытие может заменить эпидермис более старых корней, делая их похожими на веревку и даже более прочными. Как показано на рисунке ниже, некоторые корни имеют необычную специализацию для закрепления растений.
- Хранение пищи: у многих растений наземные ткани корней хранят пищу, производимую листьями во время фотосинтеза. Кровавый корень, показанный на Рис. ниже, хранит пищу в своих корнях в течение зимы.
Корни мангровых деревьев подобны ходулям, позволяя деревьям мангрового дерева подниматься высоко над водой. Ствол и листья находятся над водой даже во время прилива. Кровокорень использует пищу, хранившуюся зимой, для выращивания цветов ранней весной.
Рост корня
Корни имеют первичные и вторичные меристемы для увеличения длины и ширины. По мере того как корни становятся длиннее, они всегда врастают в землю. Даже если вы перевернете растение вверх дном, его корни будут стремиться вырасти вниз.Как корни «знают», в какую сторону расти? Как они могут отличить от верха? Специализированные клетки корневых крышек способны обнаруживать силу тяжести. Клетки направляют меристему на кончиках корней вниз, к центру Земли. Это обычно адаптивно для наземных растений. Вы можете объяснить почему?
По мере того, как корни становятся толще, они также не могут впитывать воду и минералы. Однако они могут быть даже лучше при транспортировке жидкостей, закреплении растений и хранении пищи (см. рис. ниже).
Вторичный рост корней сладкого картофеля дает больше места для хранения продуктов.Корни хранят сахар от фотосинтеза в виде крахмала. Какие еще крахмалистые корни едят люди?
Влияние всасывания корней на прочность на разрыв корней Chrysopogon zizanioides и его влияние на стабилизацию биосклонов
Бем В. (1979) Методы изучения корневых систем, Springer-Verlag Berlin Heidelberg. С. 126.
Google ученый
Boldrin D, Leung AK, Bengough AG (2017) Биомеханические свойства корней во время укоренения древесных многолетников.Экологическая инженерия 109: 196–206. https://doi.org/10.1016/j.ecoleng.2017.05.002
Артикул Google ученый
Болдрин Д., Леунг А.К., Bengough AG (2018) Влияние обезвоживания корней на биомеханические свойства древесных корней Ulex europaeus. Растения и почва: 1-23. https://doi.org/10.1007/s11104-018-3766-7
Google ученый
Boyer JS (1995) Измерение водного статуса растений и почв.Academic Press, Сан-Диего.
Google ученый
Couvreur V, Vanderborght J, Draye X, Javaux M (2014) Динамические аспекты доступности воды в почве для изогидрических растений: основное внимание уделяется гидравлическому сопротивлению корней. Исследование водных ресурсов 50 (11): 8891–8906. https://doi.org/10.1002/2014WR015608
Артикул Google ученый
Де Баетс С., Поесен Дж., Рубенс Б. и др.(2008) Прочность на растяжение корней и распределение корней типичных средиземноморских видов растений и их вклад в прочность почвы на сдвиг. Почва растений 305: 207–226. https://doi.org/10.1007/s11104-018-3636-3.
Артикул Google ученый
Eab KH, Likitlersuang S, Takahashi A (2015) Лабораторные и модельные исследования системы, укрепленной корнями для стабилизации откосов. Почвы и фундаменты 55 (5): 1270–1281.https://doi.org/10.1016/j.sandf.2015.09.025
Артикул Google ученый
Fan CC, Su, CF (2008). Роль корней в прочности на сдвиг укрепленных корнями почв с высоким содержанием влаги. Экологическая инженерия. 33 (2): 157–166. https://doi.org/10.1016/j.ecoleng.2008.02.013.
Google ученый
Fu JT, Hu XS, Brierley G и др.(2016) Влияние архитектурных свойств корневой системы растений на устойчивость лессовых склонов, Северо-Восточный Цинхай, Китай. Журнал горной науки 13 (5): 785–801. https://doi.org/10.1007/s11629-014-3275-y
Артикул Google ученый
Gardner WR (1960) Динамические аспекты доступности воды для растений. Почвоведение 89: 63–73.
Артикул Google ученый
Грей Д.Х., Охаши Х. (1983) Механика армирования волокном в песке.Журнал геотехнической инженерии 109 (3): 335–354. https://doi.org/10.1061/(ASCE)0733-9410(1983)109:3(335)
Артикул Google ученый
Gray DH, Sotir RB (1996) Биотехническая и почвенная биоинженерия стабилизация склонов: практическое руководство по борьбе с эрозией. Джон Вили и сыновья.
Google ученый
Grimshaw RG (1994) Трава ветивера — ее использование для стабилизации склонов и структур в тропических и полутропических условиях.В кн .: Растительность и склоны. Под редакцией Д.Х. Баркера. Институт инженеров-строителей, Лондон. С. 26–35. https://doi.org/10.1680/vasspae.20313.0003.
Google ученый
Hales TC, Miniat CF (2016) Влага почвы вызывает динамические корректировки укрепления корней, что снижает устойчивость откосов. Процессы земной поверхности и формы рельефа. https://doi.org/10.1002/esp.4039
Google ученый
Hathaway RL, Penny D (1975) Сила корней у некоторых клонов Populus и Salix.Новозеландский ботанический журнал 13 (3): 333–344.
Артикул Google ученый
Hengchaovanich D (1998) Ветиверная трава для стабилизации склонов и борьбы с эрозией, с особым вниманием к инженерным приложениям. Технический бюллетень № 1998/2. Сеть ветиверов Тихоокеанского региона. Офис Королевского совета по проектам развития, Бангкок, Таиланд.
Google ученый
Jotisankasa A, Mairaing W, Tansamrit S (2014) Пропитка и устойчивость почвенного склона ветиверной травой, подверженной осадкам, по результатам численного моделирования.В: Доклад, представленный на Ненасыщенных почвах: исследования и применение — Материалы 6-й Международной конференции по ненасыщенным почвам, UNSAT 2014, Сидней, Австралия. pp 1241–1247.
Google ученый
Jotisankasa A, Sirirattanachat T, Rattana-areekul C, et al. (2015) Технические характеристики системы Vetiver для стабилизации пологих склонов. В: Материалы 6-й Международной конференции по ветиверу (ICV-6), Дананг, Вьетнам, 5–8 мая 2015 г.Доступно по адресу http://www.vetiver.org/ICV6_PROC/ICV6_papersindex.htm, по состоянию на 14 ноября 2016 г.
Google ученый
Jotisankasa A, Taworn D (2016) Прямые испытания на сдвиг глинистого песка, армированного живым колом. Журнал геотехнических испытаний 39 (4): 608–623. https://doi.org/10.1520/GTJ20150217
Артикул Google ученый
Jotisankasa A, Sirirattanachat T (2017) Влияние корней травы на кривую удержания воды в почве и функцию проницаемости.Канадский геотехнический журнал 54 (11): 1612–1622. https://doi.org/10.1139/CGJ2016-0281
Артикул Google ученый
Kaewsaeng W (2000) Технические свойства выветренной глинистой почвы, усиленной корнем Prachuap Khiri Khan Vetiveria Nemoralis A. Camus для защиты склонов. Магистр технических наук. Технологический университет короля Монгкута Тхонбури (KMUTT), Бангкок, Таиланд.
Google ученый
Маханнопкул К. (2019) Механические свойства грунта с различной степенью насыщенности корнеармированным грунтом.Кандидат наук. Диссертация: Университет Касетсарт, Бангкок, Таиланд (в печати).
Google ученый
Ng CWW, Ni JJ, Leung AK и др. (2016) Влияние плотности посадки на рост деревьев и вызванное всасывание почвы. Géotechnique 66 (9): 711–724. https://doi.org/10.1680/jgeot.15.P.196
Артикул Google ученый
Nilaweera NS (1994) Влияние корней твердой древесины на прочность почвы на сдвиг и устойчивость откосов в Южном Таиланде.Кандидат наук. Диссертация: Азиатский технологический институт, Бангкок, Таиланд.
Google ученый
Nilaweera NS, Nutalaya P (1999) Роль корней деревьев в стабилизации склонов. Бюллетень инженерной геологии и окружающей среды 57 (4): 337–342. https://doi.org/10.1007/s100640050056.
Артикул Google ученый
Пыльца Н., Саймон А. (2005) Оценка механического воздействия прибрежной растительности на устойчивость берега ручья с использованием модели пучка волокон.Исследование водных ресурсов 41 (7): 1–11. https://doi.org/10.1029/2004WR003801
Артикул Google ученый
Preti F, Giadrossich F (2009) Укрепление корней и биоинженерная стабилизация откосов с помощью испанской метлы (Spartium junceum L.). Гидрология и науки о земных системах 13 (9): 1713–1726. https://doi.org/10.5194/hess-13-1713-2009
Артикул Google ученый
Рей Ф. (2018) Роль биоинженерных сооружений из черенков ивы в улавливании мергелых отложений: оценка трех реальных экспериментов в южных французских Альпах.Журнал горной науки 15 (2): 225–236. https://doi.org/10.1007/s11629-017-4663-x
Артикул Google ученый
Шварц М., Прети Ф., Гиадроссих Ф. и др. (2010) Количественная оценка роли растительности в устойчивости склонов: тематическое исследование в Тоскане (Италия). Экологическая инженерия 36 (3): 285–291. https://doi.org/10.1016/j.ecoleng.2009.06.014
Артикул Google ученый
Shewbridge SE, Sitar N (1989) Деформационные характеристики армированного песка при прямом сдвиге.Журнал геотехнической инженерии 115 (8): 1134–1147. https://doi.org/10.1061/(ASCE)0733-9410(1989)115:8(1134)
Артикул Google ученый
Shewbridge SE, Sitar N (1990) Модель на основе деформации для армированного песка при прямом сдвиге, Journal of Geotechnical Engineering 116 (7): 1153–1157. https://doi.org/10.1061/(ASCE)0733-9410(1990)116:7(1153)
Google ученый
Сунгворнпатансакул В., Раджани С. (2006) Прочность на сдвиг почвы, усиленной корнями ветивера.В: Материалы 6-го симпозиума по улучшению мягких грунтов и геосинтетике, Бангкок, Таиланд.
Google ученый
Tardieu F, Draye X, Javaux M (2017) Поглощение воды корнями и идеотипы корневой системы: все растение контролирует значение. Журнал зоны Вадосе 16 (9). https://doi.org/10.2136/vzj2017.05.0107
Google ученый
Teerawattanasuk C, Maneecharoen J, Bergado DT, et al.(2014) Измерения прочности корней трав ветивера и рузи. Lowland Technology International 16: 71–80. https://doi.org/10.14247/lti.16.2_71
Артикул Google ученый
Thomas RE, Pollen-Bankhead N (2010) Моделирование армирования корня с помощью модели пучка волокон и моделирования Монте-Карло. Экологическая инженерия 36 (1): 47–61. https://doi.org/10.1016/j.ecoleng.2009.09.008
Артикул Google ученый
Truong P, Van TT, Pinners E (2008) Система ветивера для стабилизации склонов, справочник инженера.Сеть Ветивер Интернэшнл.
Google ученый
Voottipruex P, Sungwornpatansakul W (2003) Технические свойства почвы, усиленной корнями ветивера. В: Материалы 12-й Азиатской региональной конференции по механике грунтов и инженерно-геологическому проектированию, Сингапур. С. 557–560.
Google ученый
Waldron LJ, Dakessian S (1981) Укрепление почвы корнями: расчет повышенного сопротивления почвы сдвигу на основе свойств корней.Почвоведение 132 (6): 427–435. https://doi.org/10.1097/00010694-198112000-00007
Артикул Google ученый
Wu TH, McKinnell III WP, Swanston DN (1979) Прочность корней деревьев и оползни на острове Принца Уэльского, Аляска. Канадский геотехнический журнал 16 (1): 19–33. https://doi.org/10.1139/t79-003
Артикул Google ученый
Yang Y, Chen L, Li N, Zhang Q (2016) Влияние влажности и диаметра корня на свойства корня при растяжении.PLOS ONE 11 (3). https://doi.org/10.1371/journal.pone.0151791
Google ученый
Чжун Р., Хе Х, Бао И и др. (2016) Оценка укрепления почвы корнями четырех преобладающих видов трав после плотин в прибрежной зоне трех ущелий водохранилища, Китай. Журнал горной науки 13 (3): 508–521. https://doi.org/10.1007/s11629-014-3397-2
Артикул Google ученый
Zhang CB, Chen LH, Jiang J (2014) Почему тонкие корни деревьев сильнее, чем более толстые: роль целлюлозы и лигнина в устойчивости склона.Геоморфология 206: 196–202. https://doi.org/10.1016/j.geomorph.2013.09.024
Артикул Google ученый
Zhang C, Zhou X, Jiang J, et al. (2019) Влияние влажности корней на тесты на растяжение корней травянистых растений. Катена 172: 140–147. https://doi.org/10.1016/j.catena.2018.08.012
Артикул Google ученый
корень | Определение, типы, морфология и функции
Корень , в ботанике, та часть сосудистого растения, которая обычно находится под землей.Его основные функции — закрепление растения, поглощение воды и растворенных минералов и их отвод к стеблю, а также хранение резервных кормов. Корень отличается от стебля, главным образом, отсутствием рубцов и почек на листьях, наличием корневой шляпки и ветвями, которые происходят из внутренней ткани, а не из почек.
Подробнее по этой теме
покрытосеменных: корневые системы
Корни закрепляют растение, впитывают воду и минералы и служат местом для хранения пищи.Два основных типа корневых систем являются первичными …
Виды корней и корневых систем
Первичный корень или корешок — это первый орган, который появляется при прорастании семян. Он прорастает вниз в почву, закрепляя саженец. У голосеменных и двудольных (покрытосеменных с двумя семенными листами) корешок становится стержневым корнем. Он растет вниз, а вторичные корни растут сбоку от него, образуя систему стержневых корней. У некоторых растений, таких как морковь и репа, стержневой корень также служит хранилищем пищи.
Два типа корневой системы: (слева) волокнистые корни травы и (справа) мясистый стержневой корень сахарной свеклы.
Британская энциклопедия, Inc.Травы и другие однодольные растения (покрытосеменные растения с одним семенным листом) имеют мочковатую корневую систему, характеризующуюся массой корней примерно одинакового диаметра. Эта сеть корней возникает не как ветви основного корня, а состоит из множества ветвящихся корней, выходящих из основания стебля.
Некоторые корни, называемые придаточными корнями, возникают не из корня, а из другого органа — обычно из стебля, иногда из листа.Они особенно многочисленны на подземных стеблях, таких как корневища, клубнелуковицы и клубни, и позволяют вегетативно размножать многие растения из стеблей или листовых черенков. Некоторые придаточные корни, известные как воздушные корни, либо проходят некоторое расстояние по воздуху, прежде чем достигают почвы, либо остаются висящими в воздухе. Некоторые из них, такие как кукуруза (кукуруза), винтовая сосна и баньян, в конечном итоге помогают поддерживать растение в почве. У многих эпифитных растений, таких как различные орхидеи и виды Tillandsia , воздушные корни являются основным средством прикрепления к непочвенным поверхностям, таким как другие растения и камни.
воздушный кореньБаньяновое дерево ( видов фикусов ) с воздушными корнями, выходящими из ветвей.
© Андрей Слиозберг / Fotolia Получите подписку Britannica Premium и получите доступ к эксклюзивному контенту. Подпишись сейчасСреди сосудистых растений существует ряд других специализированных корней. Пневматофоры, обычно встречающиеся у видов мангровых деревьев, которые растут на солончаках, представляют собой боковые корни, которые растут вверх из ила и воды и служат местом поступления кислорода для погруженной в воду первичной корневой системы.Корни некоторых паразитических растений сильно видоизменяются в гаустории, которые внедряются в сосудистую систему растения-хозяина, чтобы прокормить паразита. Узловатые корни многих членов семейства гороховых (Fabaceae) являются хозяевами симбиотических азотфиксирующих бактерий, а корни многих растений также образуют сложные ассоциации с микоризными почвенными грибами; ряд нефотосинтезирующих микогетеротрофных растений, таких как индийская трубка, питаются исключительно этими грибами.
Морфология и рост
Корни растут в длину только от концов.Самый верхушка корня покрыта корнем в форме наперстка, который защищает верхушку роста, когда она пробивается через почву. Сразу за корневой крышкой находится апикальная меристема, ткань активно делящихся клеток. Некоторые из клеток, продуцируемых апикальной меристемой, добавляются к корневому покрову, но большинство из них добавляются в область удлинения, которая лежит чуть выше меристематической области. Именно в области удлинения происходит рост длины. Выше этой зоны растяжения находится область созревания, где созревают первичные ткани корня, завершая процесс дифференцировки клеток, который фактически начинается в верхней части меристематической области.
корень и апикальная меристема побегаАпикальная меристема побега Hypericum uralum (слева) появляется на самой верхней части стебля. Сразу за апикальной меристемой находятся три области первичных меристематических тканей. Апикальная меристема корня (справа) появляется сразу за защитной крышкой корня. Сразу за апикальной меристемой отчетливо видны три первичные меристемы.
Британская энциклопедия, Inc.Основными тканями корня являются эпидермис, кора и сосудистый цилиндр, от самого внешнего до самого внутреннего.Эпидермис состоит из тонкостенных клеток и обычно имеет толщину всего в один клеточный слой. Поглощение воды и растворенных минералов происходит через эпидермис, этот процесс значительно усиливается у большинства наземных растений из-за наличия корневых волосков — тонких трубчатых расширений стенки эпидермальных клеток, которые встречаются только в области созревания. Поглощение воды происходит главным образом за счет осмоса, который происходит потому, что (1) вода присутствует в почве в более высоких концентрациях, чем в эпидермальных клетках (где она содержит соли, сахара и другие растворенные органические продукты) и (2) мембрана клетки эпидермиса проницаемы для воды, но не для многих веществ, растворенных во внутренней жидкости.Эти условия создают осмотический градиент, благодаря которому вода проникает в клетки эпидермиса. Этот поток создает силу, называемую давлением корней, которая помогает продвигать воду через корни. Давление на корни частично отвечает за подъем воды в растениях, но оно не может самостоятельно объяснять перенос воды к вершинам высоких деревьев.
корень в разрезеПоперечный разрез типичного корня, показывающий первичную ксилему и первичную флоэму, расположенные в центральном цилиндре.
Британская энциклопедия, Inc.Кора головного мозга проводит воду и растворенные минералы через корень от эпидермиса к сосудистому цилиндру, откуда они транспортируются к остальной части растения. Кора также хранит пищу, транспортируемую вниз от листьев через сосудистые ткани. Самый внутренний слой коры обычно состоит из плотно упакованного слоя клеток, называемого энтодермой, который регулирует поток материалов между корой и тканями сосудов.
Сосудистый цилиндр находится внутри энтодермы и окружен перициклом, слоем клеток, который дает начало корням ветвей.Проводящие ткани сосудистого цилиндра обычно расположены в форме звезды. Ткань ксилемы, которая несет воду и растворенные минералы, составляет ядро звезды; ткань флоэмы, несущая пищу, расположена небольшими группами между остриями звезды.
Старые корни древесных растений образуют вторичные ткани, которые приводят к увеличению обхвата. Эти вторичные ткани производятся сосудистым камбием и пробковым камбием. Первые возникают из меристематических клеток, лежащих между первичной ксилемой и флоэмой.По мере развития сосудистый камбий образует кольцо вокруг первичного сосудистого цилиндра. При делении клеток в сосудистом камбии образуется вторичная ксилема (древесина) внутри кольца и вторичная флоэма снаружи. Рост этих вторичных сосудистых тканей выталкивает перицикл наружу и разделяет кору и эпидермис. Перицикл становится пробковым камбием, производящим пробковые клетки (внешнюю кору), которые заменяют кору и эпидермис.
The Editors of Encyclopaedia Britannica Эта статья была недавно отредактирована и обновлена Мелиссой Петруццелло, помощником редактора.Узнайте больше в этих связанных статьях Britannica:
Поглощение воды корнями: последствия водного дефицита | Журнал экспериментальной ботаники
Аннотация
Переменная гидравлическая проводимость корней ( Lp r ) объясняется в терминах сложной транспортной модели. Показано, как сложная анатомическая структура корней приводит к комбинированному переносу воды и растворенных веществ. В модели важную роль играют параллельные апопластические и межклеточные (симпластные и трансцеллюлярные) пути, а также различные ткани и структуры, расположенные последовательно внутри корневого цилиндра (эпидермис, экзодерма, кора, энтодерма, стеллярная паренхима). .Обсуждается роль каспаровых тяжей и субериновых пластинок в эндо- и экзодерме корня. В зависимости от состояния развития этих апопластических барьеров общее гидравлическое сопротивление корней либо более равномерно распределяется по корневому цилиндру (молодые ненапряженные корни), либо концентрируется в определенных слоях (экзо- и энтодерма в более старых напряженных корнях). Причина изменчивости корня Lp r заключается в том, что гидравлические силы вызывают доминирующий апопластический поток воды вокруг протопластов, даже в энтодерме и экзодерме.В отсутствие транспирации поток воды является осмотическим по своей природе, что вызывает высокое сопротивление, поскольку вода проходит через множество мембран при прохождении через корневой цилиндр. Модель обеспечивает высокую способность корней впитывать воду при высокой скорости транспирации (высокие требования к воде от побегов). Напротив, гидравлическая проводимость низкая, когда транспирация отключена. В целом это приводит к нелинейной зависимости между потоком воды и силами (градиентами гидростатического и осмотического давления), которые иначе трудно объяснить.Модель учитывает особые характеристики корня, такие как высокая гидравлическая проводимость (водопроницаемость) в присутствии низкой проницаемости для ионов питательных веществ, однажды захваченных в стелу активными процессами. Низкие коэффициенты отражения корня соответствуют идее некоторых апопластных байпасов для воды внутри корневого цилиндра. Согласно модели составного транспорта, переключение из гидравлического режима в осмотический является чисто физическим. При наличии сильно пробуренных корней апопластная составляющая водного потока может быть слишком мала.В этих условиях преобладает регулирование радиального потока воды по водным каналам. Поскольку водные каналы находятся под метаболическим контролем, этот компонент представляет собой «активный» элемент регуляции. Композитный транспорт позволяет оптимизировать водный баланс побега в дополнение к хорошо известным явлениям, связанным с регулированием потока воды (газообмена) через устьица. Модель используется для объяснения реакции растений на дефицит воды и другие стрессы. Во время дефицита воды важную роль играет механизм сцепления-натяжения подъема сока в ксилеме.Обобщены результаты, подтверждающие справедливость теории когезии / растяжения. Представлены эффекты гормона стресса абсцизовой кислоты (АБК). Они показывают, что существует апопластический компонент потока ABA в корне, который вносит вклад в сигнал ABA в ксилеме. С другой стороны, (+) — цис — транс -ABA специфически влияет как на клеточный уровень (активность водного канала), так и на поток воды, управляемый градиентами осмотического давления на уровне корня, что согласуется с составным транспортом. модель.Гидравлический расход воды при наличии градиентов гидростатического давления остается неизменным. Результаты согласуются с комбинированной транспортной моделью и напоминают ранее полученные данные о высокой солености, полученные для клеточного ( Lp ) и корневого ( Lp r ) уровней. Они соответствуют известным эффектам лишения питательных веществ на корень Lp r и суточный ритм корня Lp r , недавно обнаруженный в корнях Lotus .
Введение
В прошлом было приложено много усилий, чтобы понять водный баланс растений с точки зрения регуляции транспирации, т.е.е. о том, как на устьичную проводимость будут влиять водный статус, интенсивность освещения, питание и другие факторы и как это изменится в стрессовых условиях, например, при дефиците воды (Cowan, 1977; Schulze, 1986; Kramer and Boyer, 1995; Jones , 1998). Меньше усилий было приложено для исследования входной стороны водного баланса, то есть поступления воды из почвы. Давно известно, что гидравлическая проводимость корней непостоянна (Brewig, 1937; Brouwer, 1954; Weatherley, 1982; Steudle et al ., 1987; Steudle, 1994; Steudle and Peterson, 1998; Henzler et al ., 1999). Это зависит от таких факторов, как недостаток воды и засоленность почвы, а также от потребности в воде прорастающих побегов. Другими важными факторами являются дефицит питательных веществ, аноксия, температура и тяжелые металлы (Munns and Passioura, 1984; Radin and Eidenbock, 1984; Azaizeh et al ., 1992; Birner and Steudle, 1993; Maggio and Joly, 1995; Carvajal ). et al ., 1996; Peyrano et al ., 1997; Schreiber et al. , и др. ., 1999; Henzler et al. ., 1999; Кларксон и др. ., 2000). Отсутствие текущих знаний о том, как эти факторы влияют на водоснабжение, в значительной степени связано с техническими проблемами при достаточно подробном измерении гидравлической архитектуры корней. Основные гидравлические свойства корней еще недостаточно изучены просто потому, что корни в почве намного менее доступны, чем побеги. Сравнительные измерения корневой гидравлики (разные виды, виды обработки и т. Д.) требуют измерения потоков воды, движущих сил (градиенты давления и осмотического давления между поверхностью корня и ксилемой) и площади поверхности корня. Поскольку условия выращивания меняют структуру и анатомию корня, это также необходимо охарактеризовать.
Схематично на рис. 1 показано, как переменное гидравлическое сопротивление корня влияет на водный потенциал (водное состояние) побега. Обычно паропроводы в воздушном пространстве мезофилла, устьичных порах и пограничном слое за пределами листьев представляют наивысшее гидравлическое сопротивление в континууме почва-растение-воздух (SPAC), и большая часть разницы водных потенциалов между почвой и атмосферой упадет. здесь (Нобель, 1991; Ян и Тайри, 1994).При заданной разности водных потенциалов (ψ , почва −ψ атм ) и потоке воды через континуум почва-растение-воздух, водный потенциал побега может определяться падением водного потенциала через гидравлическое сопротивление корня, которое обычно является самым высоким в жидкой части SPAC. Другими словами, чтобы улучшить водный статус побега, растение может либо увеличить устьичное сопротивление, либо уменьшить гидравлическое сопротивление корней; последнее действительно происходит (помимо регуляции транспирации).Неясно, является ли это чисто физическим регулированием или включает в себя некоторое активное регулирование гидравлических свойств корня (помимо очевидных изменений анатомии, вызванных стрессом). Как показано на рисунке, гидравлическое сопротивление ксилемы относительно невелико. Однако это может стать важным в условиях стресса, когда высокое напряжение ксилемы вызывает кавитацию и прерывает поток воды в сосудах между корнем и побегом (Tyree and Sperry, 1988).
Принято считать, что вклад активного водного транспорта в растения, т.е.е. поток воды , непосредственно связанный с метаболической реакцией (ями) , должен быть незначительным. Это связано с тем, что водопроницаемость мембран растительных клеток на несколько порядков больше, чем проницаемость растворенных веществ, что может вызвать быстрое короткое замыкание первичного активного потока воды (Steudle, 1989, 1992). Вместо того, чтобы активно впитывать воду, корни просто позволяют ей проходить через себя в ответ на градиенты водного потенциала, обычно создаваемые транспирацией. Сложная анатомическая структура корней приводит к сложной картине потока воды.При радиальном прохождении через корневой цилиндр необходимо учитывать перенос через различные ткани (эпидермис, кора, стела), а также компонент продольного потока (осевой перенос в сосудах ксилемы). Во время стресса анатомия ткани корня будет меняться в основном из-за того, что стресс (например, дефицит воды) вызывает развитие апопластических барьеров для воды и ионного потока (Стасовски и Петерсон, 1991; Талейсник и др. ., 1999). Их формирование представляет собой фундаментальную адаптивную стратегию растений для выживания в неблагоприятной окружающей среде (North and Nobel, 1995; Peyrano et al ., 1997; Циммерманн и Штейдл, 1998; Schreiber et al. ., 1999; Циммерманн и др. ., 2000). Кроме того, гидравлическая проводимость мембран клеток корня может быть снижена из-за закрытия водных каналов или аквапоринов в мембранах клеток корня. Эти транспортные белки привлекли большое внимание в последние годы (Chrispeels and Maurel, 1994; Steudle and Henzler, 1995; Maurel, 1997; Steudle and Peterson, 1998; Tyerman et al ., 1999; Kjellbom et al ., 1999). Таким образом, помимо транспорта через апопласт, есть некоторый поток воды через клеточные мембраны. В отличие от необратимых химических модификаций апопласта (полосы Каспариана, ламели суберина), интенсивность клеточного компонента может либо обратимо регулироваться путем открытия или закрытия существующих водных каналов, либо плотность или аквапорины могут быть увеличены за счет привлечения интернализованного белка. в плазматическую мембрану. Во время развития корня относительный вклад компонентов в общий поток воды может измениться, когда развиваются энтодерма и экзодерма (Frensch et al ., 1996). Дефицит воды снижает рост корней и вызывает выраженную суберизацию апопласта, а также, возможно, влияет на прохождение клеток (водные каналы). Это влияет на водный баланс, уменьшая способность корней впитывать воду.
Радиальный поток воды через корни растений вдоль апопласта с его барьерами и параллельный путь от клетки к клетке (аквапорины) лучше всего описывается сложной транспортной моделью, которая учитывает различия в движении через отдельные слои клеток, такие как экзо. — и энтодермы, а также через различные ткани (Steudle et al ., 1993, 1999; Steudle, 1994, 1997; Steudle and Frensch, 1996; Steudle and Peterson, 1998). Модель учитывает взаимодействие между параллельными потоками. Осевая составляющая потока воды может быть включена (Frensch and Steudle, 1989; Steudle and Peterson, 1998).
В этом обзоре обсуждаются некоторые основные свойства корневого переноса воды и то, как они изменятся в условиях дефицита воды. Основное внимание будет уделено относительной роли апопластных барьеров и аквапоринов, но также будут рассмотрены изменения движущих сил в соответствии с механизмом сцепления-натяжения подъема сока.Будут включены некоторые недавние результаты воздействия гормона стресса абсцизовой кислоты (АБК), а также измерения, в которых изменения химического состава апопластических барьеров были коррелированы с измерениями гидравлических свойств корней как на уровне клеток, так и на уровне корня в целом ( Freundl et al ., 1998, 2000; Hose et al ., 2000; Zimmermann and Steudle, 1998; Zimmermann et al ., 2000). Этот обзор также будет относиться к более старым данным о влиянии засоления и более поздним исследованиям стресса питательных веществ и суточной экспрессии аквапорина, которые могут быть важны в отношении дефицита воды (Azaizeh and Steudle, 1991; Azaizeh et al ., 1992; Henzler et al. ., 1999; Кларксон и др. ., 2000).
Рис. 1.
Упрощенная модель постоянного потока воды через установку. Поглощение воды корнями, транспортировка на большие расстояния в ксилеме и сопротивление устьицам обозначаются переменными гидравлическими резисторами, расположенными последовательно. Обычно сопротивление корня меньше, чем сопротивление побега, которое определяется сопротивлением паропроводов в листе и пограничного слоя вне листа.Сопротивление вдоль ксилемы относительно невелико. Следовательно, при данном потоке воды через систему гидравлическое сопротивление корня по отношению к сопротивлению устьиц будет определять водный статус (водный потенциал) побега (стрелка). Чтобы улучшить водный статус побега (ψ побег ), растение может либо улучшить водопоглощение за счет уменьшения R корня , либо избежать потерь воды за счет увеличения R побега .
Рис. 1.
Упрощенная модель постоянного потока воды через установку.Поглощение воды корнями, транспортировка на большие расстояния в ксилеме и сопротивление устьицам обозначаются переменными гидравлическими резисторами, расположенными последовательно. Обычно сопротивление корня меньше, чем сопротивление побега, которое определяется сопротивлением паропроводов в листе и пограничного слоя вне листа. Сопротивление вдоль ксилемы относительно невелико. Следовательно, при данном потоке воды через систему гидравлическое сопротивление корня по отношению к сопротивлению устьиц будет определять водный статус (водный потенциал) побега (стрелка).Чтобы улучшить водный статус побега (ψ побег ), растение может либо улучшить водопоглощение за счет уменьшения R корня , либо избежать потерь воды за счет увеличения R побега .
Роль анатомии корня: апопласт
Интерпретация корневых гидравлических данных требует достаточного знания их структуры. А именно, это относится к развитию в апопласте субериновых пластинок и каспариевых полос.Существуют большие различия между видами и условиями выращивания. За последние несколько лет был достигнут значительный прогресс в характеристике апопластических барьеров, вызванных засухой и другими факторами окружающей среды. У кукурузы поглощение воды и ионов питательных веществ может значительно снижаться во время стресса, вызванного засухой (Стасовский и Петерсон, 1991). Напряженные корни образовали суберизованную поверхность раздела между живой тканью и ризосферой, чтобы минимизировать потери воды. Оказалось, что после регидратации корневые системы восстановили свою гидравлическую проводимость (которая, однако, не была измерена) за счет роста удлинения, образуя новые корни с высокой гидравлической проводимостью.Талейсник и др. . выращивали растения в условиях нехватки воды и в контроле и сравнивали способность корней удерживать воду у растений, которые имеют экзодерму, с теми, у которых ее нет (Taleisnik et al ., 1999). Скорость потери воды в экзодермальных корнях была меньше, чем в неэкзодермальных. Более старые части корней выделяют меньше воды в сухую среду, чем молодые. Ригер и Литвин пришли к выводу на основе гидравлических измерений корневых систем (подверженных засухе и контрольных) интактных растений различных травянистых и древесных видов, что существование пробуренной экзодермы имело некоторое влияние на общий корень Lp r (Rieger and Litvin , 1999).Однако сопротивление коры оказалось равным или даже большим, чем сопротивление суберизованных слоев. Недавние измерения на вырезанных корнях молодых проростков кукурузы показали, что гидравлическая проводимость была больше в 3,6 раза (в среднем) при отсутствии экзодермы (корни, выращенные в гидропонной культуре) по сравнению с корнями, имеющими экзодерму (корни, выращенные в аэропонике). ) (Zimmermann and Steudle, 1998). В этих экспериментах возможный эффект изменения гидравлической проводимости мембран корневых клеток (водных каналов) был исключен путем измерения корневой клетки Lp (датчик клеточного давления) помимо всего корня Lp r (корневой датчик давления; Zimmermann et al ., 2000). Следовательно, было существенное влияние суберизации на общие гидравлические свойства корней.
За последние несколько лет был достигнут значительный прогресс в определении химического состава апопластических барьеров. После переваривания остальной части корня стало возможным отделить каспариновые полосы от экзо- и энтодермы, химически проанализировать эти структуры и отнести результаты к гидравлическим свойствам корня (Schreiber, 1996; Zeier and Schreiber, 1998; Schreiber и др. ., 1999; Циммерманн и др. ., 2000). Результаты показывают, что полосы Каспариана содержат как алифатический, так и ароматический суберин. Первый считается более гидрофобным и менее проницаемым для воды, второй представляет собой лигнин-подобный фенольный полимер, этерифицированный гидроксикоричной кислотой. {cw} \)
, где дополнительный верхний индекс ‘ cw означает «клеточная стенка»), которая является мерой «пассивной селективности» апопласта.{cw} \) Δπ s , также будет мало. На пути от клетки к клетке ситуация иная из-за присутствия мембран, которые демонстрируют σ s ≈1. Общий корень σ sr будет находиться между нулем и единицей, как это было обнаружено в экспериментах с датчиком корневого давления и другими методами (Miller, 1985; Steudle et al. ., 1987, 1993; Steudle and Frensch, 1989; Steudle and Peterson , 1998). Вследствие своей сложной структуры корни не ведут себя как идеальные осмометры, коэффициент отражения которых равен единице.В дополнение к низкому коэффициенту отражения корней, гидравлическая проводимость корней должна зависеть от силы (градиенты осмотического и гидростатического давления), используемой для перемещения воды через корни. Это тоже следствие композитного транспорта. При наличии разных сил два пути (апопластический и межклеточный) будут использоваться с разной интенсивностью. Оба пути будут использоваться при наличии градиента гидравлического (гидростатического давления), например, во время транспирации. Однако в присутствии осмотического градиента транспорт от клетки к клетке будет преобладать.Гидравлический компонент будет небольшим, если он вообще есть. Как следствие, гидравлическая проводимость корней должна различаться в зависимости от условий, обнаруженных экспериментально (см. Ниже).Пути для транспортировки воды, растворенных веществ и композитов
Когда вода и растворенные вещества (питательные вещества) перемещаются в корень, возникает клеточный (межклеточная) и апопластический компоненты потока (Steudle, 1989, 1992). Пассивный межклеточный компонент потока растворенных веществ должен быть небольшим из-за низкой проницаемости растворенных веществ для мембран (ионов питательных веществ).Для воды это другое дело, потому что вода движется через мембраны на несколько порядков быстрее, чем ионы. В принципе, межклеточный («протопластический») компонент должен быть разделен на симпластический и трансцеллюлярный компоненты (Рис. 2A). Первый компонент обусловлен транспортом через плазмодесматы внутри симпласта. Последнее относится к прохождению через плазмалемматы (две мембраны на клеточный слой). Вакуолярный компонент не учитывается, вклад которого должен быть небольшим (Steudle and Peterson, 1998).Экспериментально до сих пор невозможно разделить симпластический и трансклеточный компоненты. Поэтому из прагматических соображений они суммируются как межклеточный компонент. Во время радиального прохождения воды через корень могут быть комбинации путей, в которых вода может перемещаться внутри симпласта на некоторое расстояние, а затем может пересекать плазматическую мембрану и перемещаться внутри клеточной стенки. Эти комбинации могут быть включены в модели водного транспорта при условии, что делается поправка на быстрый обмен воды между путями («локальное равновесие»; Мольц и Ферриер, 1982; Вестгейт и Стейдл, 1985; Стейдл, 1989, 1992; Стейдл и Френш, 1996).Согласно этим моделям, вклад компонента от ячейки к ячейке может быть получен из измерений с помощью датчика давления в ячейке. Апопластический компонент (который на сегодняшний день недоступен для прямого измерения в неповрежденной ткани) получается из разницы между гидравлической проводимостью всего корня и межклеточного компонента (Zhu and Steudle, 1991). Когда образуются апопластные барьеры, которые прерывают поток вокруг протопластов корня в эндо- и экзодерме вдоль развивающегося корня, относительные вклады гидравлического и осмотического потока изменяются (Frensch et al ., 1996).
Композитная структура корней обеспечивает достаточно высокую и регулируемую водопроницаемость при наличии ярко выраженной способности удерживать питательные соли, перенесенные в ксилему (низкая проницаемость растворенных веществ). Этого было бы трудно достичь с помощью гомогенной мембраны или осмотического барьера. Комбинированная транспортная модель объясняет такие экспериментальные данные, как (i) низкие коэффициенты отражения корней, (ii) различия между осмотическим и гидравлическим потоком воды и изменчивость корня Lp r , и (iii) различия между корнями древесных и травянистых растений. растения.
Низкие коэффициенты отражения можно просто объяснить наличием апопластических байпасов в массивах, где полосы Каспариана еще не развиты, или даже некоторым обходным потоком воды через саму полосу Каспариана (рис. 2B). Они подтверждаются комбинированными измерениями клеток Lp и корня Lp r и экспериментами по прокалыванию (Zhu and Steudle, 1991; Azaizeh et al., ., 1992; Steudle et al. ., 1993; Peterson and Steudle, 1993).Наличие параллельных элементов с низким (апопласт) и высоким (от ячейки к ячейке) коэффициентами отражения должно приводить к циркуляционному потоку воды в корне при нулевом общем потоке воды (рис. 2B), что можно продемонстрировать с помощью компьютерного моделирования. в котором общие потоки (вода, растворенные вещества) и коэффициенты отражения были рассчитаны с использованием данных измерений корневых клеток (W Melchior и E Steudle, неопубликованные результаты). Различия между гидравлическим и осмотическим потоком воды значительны и более выражены в корнях древесных, чем травянистых растений.У древесных пород они могут достигать двух-трех порядков (Rüdinger et al ., 1994; Steudle, Meshcheryakov, 1996; Steudle, Heydt, 1997). Это соответствует теории. Переменный корень Lp r в значительной степени объясняется изменением движущей силы (см. Ниже), но обсуждались и другие причины (Fiscus, 1975; Passioura, 1988).
Рис. 2.
(A) Пути радиального потока воды через корни растений.Апопластический путь проходит вокруг протопластов. Симпластический путь опосредуется плазмодесмами, которые соединяют клеточные стенки между соседними клетками, так что образуется цитоплазматический континуум. Во время прохождения по апопласту и симпласту пересечение мембран не требуется. На трансклеточном пути необходимо пересечь две плазматические мембраны на клеточный слой. Трансклеточный путь является чем-то особенным для воды, которая имеет высокую проницаемость мембраны (гидравлическую проводимость клетки). Обычно для растворенных веществ этот компонент незначителен.Обратите внимание, что симпластические и трансцеллюлярные компоненты потока не могут быть разделены экспериментально и суммированы как межклеточный компонент потока воды (см. Текст). Из-за быстрого водообмена между протопластами и соседним апопластом, в любое время должно быть локальное равновесие потока воды между двумя компартментами. В энтодерме и экзодерме корней компонент апопластического потока может быть изменен наличием апопластических барьеров (полосы Каспариана, ламеллы суберина). Обычно считается, что они непроницаемы для воды и растворенных веществ (ионов питательных веществ).(B) Сложная составная структура корней вызывает «сложный перенос». Это связано с тем, что вдоль апопластического пути нет мембран и, следовательно, нет осмотического потока воды из-за почти нулевого коэффициента отражения от стенок. С другой стороны, через апопласт проходит значительный гидравлический поток. На пути от клетки к клетке присутствуют оба компонента потока (гидравлический и осмотический поток), но этот канал имеет относительно высокое гидравлическое сопротивление, поскольку приходится пересекать множество мембран.При отсутствии градиентов гидростатического давления (при нулевой транспирации) гидравлический поток воды через апопласт не будет. Осмотический поток будет небольшим из-за высокого сопротивления в этих условиях. В зависимости от эффективности апопластных барьеров это приводит к большим различиям между гидравлическим и осмотическим потоком, что вызывает изменчивость корня Lp r . Напряжение в ксилеме играет важную роль. При нулевом или низком расходе воды можно настроить циркуляционный поток воды внутри корня, как показано.Обратите внимание, что в отличие от (A), только одна полоса Каспарова показана на (B) для простоты. Атмосферное давление корневой среды берется в качестве эталона давления, т.е. Р м = 0.
Рис. 2.
(A) Пути радиального потока воды через корни растений. Апопластический путь проходит вокруг протопластов. Симпластический путь опосредуется плазмодесмами, которые соединяют клеточные стенки между соседними клетками, так что образуется цитоплазматический континуум.Во время прохождения по апопласту и симпласту пересечение мембран не требуется. На трансклеточном пути необходимо пересечь две плазматические мембраны на клеточный слой. Трансклеточный путь является чем-то особенным для воды, которая имеет высокую проницаемость мембраны (гидравлическую проводимость клетки). Обычно для растворенных веществ этот компонент незначителен. Обратите внимание, что симпластические и трансцеллюлярные компоненты потока не могут быть разделены экспериментально и суммированы как межклеточный компонент потока воды (см. Текст).Из-за быстрого водообмена между протопластами и соседним апопластом, в любое время должно быть локальное равновесие потока воды между двумя компартментами. В энтодерме и экзодерме корней компонент апопластического потока может быть изменен наличием апопластических барьеров (полосы Каспариана, ламеллы суберина). Обычно считается, что они непроницаемы для воды и растворенных веществ (ионов питательных веществ). (B) Сложная составная структура корней вызывает «сложный перенос». Это связано с тем, что вдоль апопластического пути нет мембран и, следовательно, нет осмотического потока воды из-за почти нулевого коэффициента отражения от стенок.С другой стороны, через апопласт проходит значительный гидравлический поток. На пути от клетки к клетке присутствуют оба компонента потока (гидравлический и осмотический поток), но этот канал имеет относительно высокое гидравлическое сопротивление, поскольку приходится пересекать множество мембран. При отсутствии градиентов гидростатического давления (при нулевой транспирации) гидравлический поток воды через апопласт не будет. Осмотический поток будет небольшим из-за высокого сопротивления в этих условиях.В зависимости от эффективности апопластных барьеров это приводит к большим различиям между гидравлическим и осмотическим потоком, что вызывает изменчивость корня Lp r . Напряжение в ксилеме играет важную роль. При нулевом или низком расходе воды можно настроить циркуляционный поток воды внутри корня, как показано. Обратите внимание, что в отличие от (A), только одна полоса Каспарова показана на (B) для простоты. Атмосферное давление корневой среды принимается за эталонное давление, т.е.е. P м = 0.
Физиологические последствия композитного транспорта
Наиболее очевидным следствием композитного транспорта является то, что подача воды корнем побега может изменяться в соответствии с потребностями побега, просто за счет «физической корректировки» корня Lp r . У растущего растения градиент гидростатического давления между почвенным раствором и ксилемой будет высоким (напряжение в ксилеме), а гидравлическое сопротивление корня — низким.Это способствует поглощению воды при увеличении спроса. Похоже, что происходит «переключение» между гидравлическим и осмотическим режимами действия корней. С другой стороны, гидравлическое сопротивление будет высоким при низких скоростях транспирации, то есть ночью или в периоды дефицита воды. В этих условиях растения могут быть защищены от чрезмерных потерь воды в почву за счет высокого гидравлического сопротивления их корней. Таким образом, модель стремится оптимизировать водный баланс, регулируя корневую гидравлику в соответствии с потребностями побега (рис.1). Скорость утечки растворенных веществ (питательных солей) сохраняется на низком уровне. Обычно коэффициенты проницаемости корней для питательных солей аналогичны коэффициентам проницаемости клеточных мембран (Steudle and Brinckmann, 1989).
Роль энтодермы и экзодермы
Измерения с вырезанными корнями молодых проростков кукурузы, выращенных в гидропонной культуре, показали, что гидравлическое сопротивление не сконцентрировано в энтодерме (что было бы общепринятым мнением). При выращивании на гидропонике у корней не образовывалась экзодерма (Peterson and Steudle, 1993; Peterson et al ., 1993; Steudle и др. ., 1993). Вместо того, чтобы концентрироваться в энтодерме, сопротивление было более равномерно распределено в живой ткани корня. Стенки сосуда, которые уже были утолщенными и содержали лигнин, составляли лишь 10–30% от общего сопротивления движению воды. Энтодерма была модифицирована каспариновыми тяжами, но субериновых ламелей не было. Таким образом, вывод о том, что энтодерма (только с полосами Каспариана) не является серьезным препятствием для движения воды, может быть сделан только для этих молодых корней, выращенных в гидропонике.Для корней, выращенных в «реальных» условиях, экстраполяция измерений на корнях, выращенных в аэропонике, и других результатов показала, что обычно это не так. Определенно, экзодерма представляет собой барьер для движения воды для корней кукурузы, выращенных в реальных условиях. Ожидается, что ламели суберина, расположенные в тангенциальной, радиальной и поперечной стенках, уменьшают гидравлическую проводимость. То же самое может произойти после утолщения обоих слоев, которые могут быть суберизованными и / или одревесневшими.Есть эксперименты, которые показывают, что это правда (North and Nobel, 1991, 1995; Melchior and Steudle, 1993).
Во время водного стресса образование субериновых пластинок в эндо- и экзодерме может привести к локализованному высокому сопротивлению потоку воды и ионов в апопласте корня. Экзодерма, таким образом, может способствовать регуляции поглощения воды корнями, как предполагают эксперименты, в которых корни выращивали в почве с дефицитом воды или в питательном растворе с высоким содержанием соли (Stasovski and Peterson, 1991; Cruz et al ., 1992; Azaizeh and Steudle, 1991; Azaizeh et al. ., 1992; Peyrano et al. ., 1997; Талейсник и др. ., 1999). Однако недавние измерения на молодых корнях лука, выращенных в вермикулите, показали, что корень Lp r увеличивался по мере развития экзодермы в константной клетке Lp (Barrowclough et al ., 2000). Эти результаты показывают, что, в отличие от кукурузы, гидравлическое сопротивление может более равномерно распределяться по корневому цилиндру у лука даже на стадиях, когда экзодерма уже развита.На сегодняшний день по техническим причинам отсутствуют количественные значения проницаемости экзо- и энтодермы интактных корней для воды. Эти подробные измерения необходимы для определения функций двух конструкций во время дефицита воды и других стрессовых ситуаций. Упомянутый выше подход Zimmermann и Steudle, хотя и ограничен в применении, может быть использован, в котором при изменении условий выращивания корни получают с экзодермисом или без него (Zimmermann and Steudle, 1998).Другой подход предполагает измерения изменений в ячейке Lp и распространения изменений водного потенциала (осмотического и гидростатического давления) по корневому цилиндру (датчик давления в ячейке). Эти эксперименты нелегко провести, но методы их проведения в принципе разработаны (Zhu and Steudle, 1991). Их следует использовать, чтобы увидеть, как изменится гидравлическое сопротивление двух слоев в ответ на дефицит воды. Суберизованные клетки экзо- и энтодермы должны иметь меньшую клетку Lp .Следовательно, скорость распространения изменений водного потенциала корневой среды через корень (измеряемая датчиком давления в клетках коры) должна быть меньше в присутствии суберизованной экзодермы.
Транспорт растворенных веществ
Вода гораздо более подвижна как на пути апопласта, так и на пути от клетки к клетке, чем растворенные вещества. Следовательно, движение воды в апопласте не может быть определено количественно, отслеживая движение апопластных красителей. Например, в экспериментах Zimmermann и Steudle внешняя среда содержала ионный апопластный индикатор PTS (3-гидрокси-5,8,10-пирен-трисульфонат; Peterson et al ., 1981; Hanson et al. ., 1985; Скиннер и Радин, 1994), чтобы проверить проницаемость корней этого ионного соединения помимо воды (Zimmermann and Steudle, 1998). СТВ проходил через корни, и, в отличие от воды, проницаемость СТВ была одинаковой, независимо от того, имел ли корень зрелую экзодерму или нет. Во время прохождения через корень PTS разбавляли до концентрации всего 0,2% от концентрации в среде. Если бы PTS были хорошим индикатором для воды, это означало бы, что апопластический компонент потока воды был только 0.2%. Это, однако, было неправдой, потому что сравнение между корнем Lp r и корневой клеткой Lp показало, что должен был быть значительный поток воды вокруг протопластов. На это также указывают измерения на других видах, помимо кукурузы (Radin and Matthews, 1989). Был сделан вывод, что, по крайней мере, для корней растений кукурузы, выращиваемых в аэропонике до возраста 3 недель, энтодерма, а не экзодерма была основным барьером для потока PTS, так что развитие экзодермы не приводило к измеримым результатам. уменьшение потока СТВ.По-видимому, заряженные растворенные вещества эффективно удерживаются в присутствии полос Каспариана.
Изображение корня, представленное до сих пор, представляет собой структуру, в которой апопласт все еще имеет некоторую проницаемость для воды, даже в экзо- и энтодерме. Этот вывод был сделан как из сравнения между корнем Lp r и клеткой Lp , так и из химического анализа апопластических барьеров (см. Выше). Это не относится к ионам питательных веществ. В молодых корнях кукурузы наблюдалось быстрое снижение корневого давления, даже когда были повреждены только несколько клеток энтодермы (Steudle et al ., 1993). Что касается PTS, диффузия ионов из стелы сильно затруднялась энтодермой. Быстрое снижение корневого давления после травмы предполагало, что через утечку имел место объемный поток раствора ксилемы, который представлял собой путь с низким сопротивлением между отделением с более высоким давлением (апопласт стелляра) и отделением с более низким давлением (среднее). Результаты подтвердили, что энтодерма действует как эффективный барьер для обратной диффузии ионов, высвобождаемых в апопласт стелы, что позволяет нарастать корневому давлению в условиях низкой транспирации.С другой стороны, гидравлическое сопротивление энтодермы довольно низкое, по крайней мере, для клеток с одними каспариевыми тяжами. Таким образом, транспортные свойства корней оптимизированы как для ионов, так и для воды. Первые собираются выборочно и надежно удерживаются в стеле, в то время как сопротивление потоку воды невелико, что обеспечивает достаточный запас воды в побег.
Являются ли водные каналы альтернативным проходом?
В побеге поток воды в значительной степени контролируется устьицами.Устьица представляют собой «активный» элемент контроля или регулирования, который регулируется состоянием воды в побегах и сигналами от корня (ABA). Кутикулу побега можно считать «водонепроницаемой» или почти такой. В корне ситуация менее ясна. Нет устьиц, а поглощение воды и питательных веществ требует прохождения через массивы непуберизованной ткани. С другой стороны, необходимо уменьшить гидравлическую проводимость, чтобы избежать значительных потерь воды в сухую почву при отключении транспирации.Похоже, что до некоторой степени сложная транспортная модель решает дилемму. Согласно модели, существует переменное сопротивление корня, которое является низким при наличии транспирации, когда напряжение (отрицательное давление) в ксилеме обеспечивает градиент водного потенциала, достаточный для впитывания воды даже из сухой почвы. При отсутствии транспирации (например, ночью) пробурение корней (при дефиците воды) обеспечивает минимизацию потерь воды в сухую почву, когда градиент водного потенциала имеет неправильную направленность.Следует отметить, что матричные силы эквивалентны гидростатическим силам (как и осмотические силы), но они физически различны. Это может вызвать различия в их роли как движущей силы. Матричные силы происходят от компонента водного потенциала, который происходит от эффектов поверхностного натяжения, а не от объемной работы, которая является источником составляющей гидростатического давления. Что касается модели, важно, как изменения в матричном потенциале почвы переводятся в изменения компонентов водного потенциала в апопласте корня (как подробно показано Steudle, 1992).Когда изменения вызывают изменение осмотического компонента, результирующий поток будет в основном переходить от клетки к клетке (высокое сопротивление). С другой стороны, когда они вызывают градиент гидростатического давления или матричных потенциалов, поток будет в значительной степени апопластическим (низкое сопротивление). При существенной нехватке воды в почве гидравлическое сопротивление корня может быть слишком высоким, чтобы обеспечить достаточное поглощение воды, даже когда градиенты водного потенциала имеют правильное направление. В этих условиях водные каналы могут действовать как клапаны, обратимо увеличивая гидравлическую проводимость и обеспечивая поглощение воды в неблагоприятных условиях.Было показано, что водные каналы могут быть активированы фосфорилированием, на которое, в свою очередь, влияют такие факторы, как водный потенциал, тургор или концентрация Ca 2+ в апопласте (Johansson et al ., 1996; Kjellbom et al ., 1999). На сегодняшний день понимание того, как эта регуляция происходит в неповрежденном корне, довольно ограничено.
Для корней Lotus japonicus было продемонстрировано, что концентрация мРНК, кодирующей предполагаемые водные каналы, демонстрирует отчетливый суточный ритм с максимумом в полдень.Это совпало с максимумом гидравлической проводимости корня, измеренной либо с осмотическим градиентом, либо с градиентом давления. Однако максимумы в корнях Lp r не коррелировали с соответствующими изменениями гидравлической проводимости клеток коры корня, как показали измерения с помощью датчика клеточного давления. Результат был интерпретирован в том, что регуляция клеточного Lp может быть ограничена энтодермальными и стеллярными клетками только как предполагают молекулярные маркеры (Schäffner, 1998).В любом случае в стеле регулирование было бы более эффективным, потому что по геометрическим причинам эта часть вносит больший вклад в общее радиальное сопротивление, чем кора и эпидермис.
Для некоторых видов, таких как фасоль ( Phaseolus coccineus ) и ячмень, было продемонстрировано, что из-за высокой мембраны Lp преобладает переход от клетки к клетке (Steudle and Jeschke, 1983; Steudle and Brinckmann , 1989). Для других эффекты различны. Было сделано несколько попыток продемонстрировать действие водных каналов (аквапоринов) в корнях, например, путем закрытия их ртутными веществами (Maggio and Joly, 1995).Однако в этих экспериментах не были полностью исключены эффекты блокирующего агента на другие переносчики и побочные эффекты. Эксперименты с пшеницей показали, что недостаток питательных веществ снижает активность водных каналов (Carvajal et al ., 1996; Clarkson et al ., 2000). Вывод был сделан из того факта, что, в отличие от контроля, ртутные вещества не влияли на гидравлическую проводимость лишенных корней. Недавно Zhang и Tyerman использовали зонд клеточного давления для измерения обратимого ингибирования водных каналов HgCl 2 в кортикальных клетках корней пшеницы (Zhang and Tyerman, 1999).Интересно, что гипоксия вызывала такие же эффекты, как и HgCl 2 , и клетки, обработанные гипоксией, не могли больше подавляться. Чжан и Тайерман не измеряли эффект закрытия водных каналов на корне Lp r , как это делали другие авторы (Azaizeh and Steudle, 1991; Azaizeh et al ., 1992). Последние авторы обнаружили, что в кукурузе засоление (100 мМ) вызывало уменьшение количества клеток Lp в 3-6 раз. Однако на корневом уровне снижение составило всего 30–60%.Результат был объяснен с точки зрения модели составного переноса значительным апопластическим байпасным потоком воды.
Для интактных междоузлий Chara на сегодняшний день имеются подробные данные о функции водных каналов (Henzler and Steudle, 1995; Schütz and Tyerman, 1997; Hertel and Steudle, 1997). Работа продемонстрировала как избирательность водных каналов, так и ее значимость для клетки Lp . Подобно клеткам коры головного мозга пшеницы (Zhang and Tyerman, 1999), закрытие водных каналов уменьшало количество клеток Lp на 75%.С другой стороны, проницаемость мембраны для небольших незаряженных растворенных веществ, таких как низкомолекулярные спирты, амиды, кетоны и т.д., не изменилась. Тщательное изучение транспортных свойств этих клеток показало, что имело место некоторое проскальзывание растворенных веществ по водным каналам (Steudle and Henzler, 1995; Hertel and Steudle, 1997; Heuzler and Steudle, 2000). Согласно результатам с Chara , водные каналы в растениях не кажутся такими избирательными, как это иногда предполагалось (Chrispeels and Maurel, 1994; Maurel, 1997; Schäffner, 1998).Отсутствуют комбинированные измерения как на корневом уровне, так и на уровне ячеек, чтобы оценить влияние блокировки канала на общий транспорт. Эти измерения должны включать клетки экзо- и энтодермы и стеларные клетки.
Механизм сцепления-натяжения обеспечивает переключение между различными видами транспорта в корнях
Нелинейная зависимость силы / потока, обнаруженная во время поглощения воды корнями растений, благоприятна для растения. Это позволяет увеличить поток воды в периоды повышенного спроса со стороны прорастающих побегов.Однако при нехватке воды гидравлическое сопротивление будет высоким, что предотвратит обратный поток в сухую почву, когда транспирация отключена. Феномен переменного корневого сопротивления трактовался по-разному. Это было интерпретировано с точки зрения изменения водопроницаемости мембран клеток корня (Brewig, 1937). Фискус разработал физическую модель, которая объяснила «кажущееся» уменьшение гидравлического сопротивления корня при высоких скоростях потока изменением движущей силы (Fiscus, 1975).Согласно «модели разбавления» Фискуса, изменение произошло из-за изменения осмотической составляющей движущей силы при постоянном корне Lp r . В условиях нехватки воды эффект должен усиливаться по мере увеличения значения осмотического давления ксилемного сока. Однако подробные измерения с помощью датчиков давления показали, что эффект разбавления не может полностью объяснить нелинейность (Zimmermann and Steudle, 1998).
Напряжения в корневой ксилеме обеспечивают основную силу, перемещающую воду через корневой цилиндр в сосуды ксилемы и вверх к побегу.Только изредка должны преобладать осмотические силы. Таким образом, механизм сцепления – натяжения играет важную роль при поглощении воды корнями, а именно в условиях водного стресса. В течение последних десяти лет Циммерманн и его коллеги ставили под сомнение правомерность теории сцепления и напряжения на основании прямых измерений давления ксилемы с помощью датчика клеточного давления (Balling and Zimmermann, 1990; Melcher et al , 1998). В ранних работах этой группы напряжение в ксилеме было довольно низким (давление менее отрицательным), и транспирация практически не влияла на него.Очень часто измеряется давление ниже атмосферного, а не давление ниже абсолютного нуля (вакуума). Значения P x , измеренные датчиком давления, были значительно выше (менее отрицательными), чем значения, обнаруженные с бомбой Шоландера (камера давления). Циммерманн и его сотрудники пришли к выводу, что теория когезии ошибочна и что барокамера неправильно измеряла водные потенциалы (давление ксилемы, P x ). Однако по мере того, как группа улучшала свои навыки измерения давления ксилемы, значения становились все более отрицательными, а напряжение доходило до 0.Было обнаружено 5–0,6 МПа, которые реагировали на транспирацию. Был сделан вывод, что нельзя полностью исключить механизм сцепления-натяжения. Скорее, он послужил бы дополнительным механизмом. Изменение отношения может указывать на то, что, в конце концов, группа встает на путь принятия теории сплоченности и напряжения. Однако это неправда, потому что старые результаты все еще цитируются и утверждают, что они верны. На них по-прежнему ссылаются и обсуждают другие, считающие, что они имеют прочную основу (Tomos and Leigh, 1999).
Недавно совместные измерения с датчиком давления и камерой давления на неповрежденных растениях кукурузы показали, что оба метода дают одинаковые значения, по крайней мере, для натяжения до 1 МПа (10 бар; Wei et al ., 1999 а , б , 2000). Это согласуется с теорией сцепления – натяжения. В отличие от ранних результатов группы Циммерманна, результаты Wei и др. . показали, что давление ксилемы сильно и быстро реагирует на изменения транспирации, что согласуется с механизмом сцепления-натяжения.Что наиболее интересно, тщательное изучение пределов датчика давления для измерения напряжений показало, что само оборудование способно выдерживать напряжения до 1,6–1,8 МПа в короткие периоды (Wei и др. . 1999 a ) . Когда кончик зонда был помещен в ксилему, верхний предел был дополнительно снижен до примерно 1 МПа. Таким образом, этот метод, как он стоит не может быть использована, чтобы подтвердить или опровергнуть существование напряженности до 10 МПа (100 бар), которые были косвенно измерена с помощью камеры высокого давления.До напряжений до 1 МПа данные согласуются с теорией сцепления – растяжения. Похоже, что ранние и настоящие утверждения группы Циммерманна в значительной степени основаны на артефактических результатах из-за ограничений используемой техники. До недавнего времени эти ограничения не подвергались тщательной проверке, хотя в литературе были предупреждения (Steudle and Heydt, 1988; Heydt and Steudle, 1991; Steudle, 1995).
Что касается поглощения воды корнями при низком водном потенциале почвы, результаты имеют решающее значение.Согласно теории сцепления-натяжения, транспирационные растения могут впитывать воду из сухой почвы с высоким корнем Lp r (как предсказывает составная транспортная модель). С другой стороны, когда транспирация отключена, root Lp r будет переключаться на низкие значения, тем самым ограничивая потерю воды в почве в этих условиях.
Транспорт ABA: корневой сигнал
Роль ABA как сигнала стресса на большом расстоянии, производимого в корне, хорошо известна.Обычно считается, что концентрация АБК в ксилеме является сигналом, который передается к побегу с потоком транспирации. Однако поглощение воды ксилемой могло бы изменить сигнал путем разбавления, если только не будет некоторого апопластического транспорта ABA, который мог бы вносить вклад в сигнал. Недавние измерения вырезанных корней подсолнечника и кукурузы показывают, что это действительно так (Freundl et al ., 1998, 2000). У подсолнечника и кукурузы, выращенных в гидропонной культуре, наблюдалось значительное сопротивление растворителю АБК, предложенной питательному раствору при низких концентрациях (0–500 нМ).Эффект компенсировал или даже чрезмерно компенсировал эффект разбавления, вызванный сопутствующим поглощением воды. Был сделан вывод, что у растений, испытывающих водный стресс, АБК, продуцируемая в корне, может апопластно транспортироваться через корень, по крайней мере, до некоторой степени. Согласно Hartung et al. ., Биосинтез ABA происходит в цитозоле всех клеток корня (Hartung et al ., 1999). Самая большая емкость обнаруживается в невакуолизированных клетках кончиков корней. Согласно модели Словика и Хартунга, АБК распределяется между апопластом и симпластом в соответствии с разницей pH между двумя компартментами (Slovik and Hartung, 1992).Более щелочной (менее кислый) pH апопласта увеличит концентрацию АБК в апопласте и, следовательно, усилит перенос растворителя АБК в побеги с потоком транспирации. Поток воды через растение будет напрямую связан с сигналом ABA, который вырабатывается в корне во время водного стресса.
В соответствии с этими выводами был тот факт, что коэффициент отражения АБК (рассчитанный из сопротивления растворителя) был близок к единице при высоком pH (все АБК в анионной форме) и низком при низком pH, когда значительное количество присутствовало в недиссоциированном виде. кислота.Когда корни кукурузы выращивались в аэропонной культуре и развивали экзодерму, как транспорт АБК, так и поток воды были снижены, что привело к концентрации АБК, аналогичной той, которая была обнаружена в отсутствие экзодермы (Freundl et al ., 2000. ). Следовательно, размер сигнала не сильно изменился. Был сделан вывод, что апопластный пассаж АБК может иметь значение во время передачи сигналов о дефиците воды от корня к побегу. Между двумя используемыми видами были большие различия в том, что апопластический компонент был намного больше у кукурузы по сравнению с подсолнечником.Различия могут быть связаны с различиями в тонкой структуре корней двух видов, а именно с ультрафильтрационными свойствами энтодермы. В энтодерме должны быть эффекты концентрационной поляризации, которые могут способствовать довольно высокому сопротивлению растворителя и низким кажущимся коэффициентам отражения АБК.
Интересно, что АБК также увеличивала гидравлическую проводимость кукурузы как на уровне отдельных клеток ( Lp, ), так и целых корней ( Lp r ), возможно, за счет воздействия на водные каналы (Hose et al ., 2000). Тесты с другими гормонами и производными АБК показали, что эффекты на Lp и Lp r специфичны для недиссоциированного (+) — цис — транс ‐ABA. Эффекты были намного сильнее на клеточном уровне, чем на корневом уровне. На корневом уровне Lp r не изменялись при наличии гидравлического потока воды, когда доминировал апопластический компонент. Это соответствует модели составного транспорта. Результаты показывают, что поток воды (транспирация), который обычно регулируется с помощью гормона стресса ABA на уровне устьиц, может взаимодействовать с сигналом ABA, исходящим от корня.С другой стороны, ABA может регулировать водопоглощение в непроходящих условиях, то есть в условиях нехватки воды. Можно предположить, что в этих условиях межклеточный компонент потока воды увеличивается за счет активации активности водных каналов.
Выводы
На основании транспортных и анатомических исследований сделан вывод, что сложная структура корней играет важную роль в регуляции водопоглощения. Сложная структура корней приводит к смешанному переносу воды и растворенных веществ.Происходит переключение между клеткой (аквапорины) и апопластическим путем, что, как считается, вызывает большую часть изменчивости гидравлической проводимости корней растений. Переключение между транспортными моделями зависит от сил, действующих в системе при наличии и отсутствии транспирации («гидравлический» и «осмотический» режимы потока воды). При высокой транспирации сила, перемещающая воду через корень, будет разницей гидростатического давления между корневой средой и ксилемой.Это позволяет использовать как апопластный, так и межклеточный пути, даже в экзо- и энтодерме. Когда транспирация отключена, остается проход от клетки к клетке, имеющий высокое гидравлическое сопротивление. Есть грубая регулировка root Lp r , которая носит чисто физический характер и не имеет аналогов в побеге. При наличии достаточной транспирации переключение на гидравлический поток воды обеспечивает доступ растения к почвенной воде при низком водном потенциале.При отсутствии транспирации корень Lp r становится мелким. Это сводит к минимуму потери воды в сухую почву. У растений, растущих при дефиците воды, рост корней замедляется, и существуют значительные апопластические барьеры, блокирующие гидравлический путь. В этих условиях корень Lp r может быть низким и регулироваться на пути от клетки к клетке путем открытия или закрытия водных каналов. Было показано, что активность водных каналов существует в корковых клетках корня, включая энтодерму.Активность водных каналов может подавляться тяжелыми металлами, гипоксией, дефицитом питательных веществ, низкой температурой и высокой соленостью. Измерения ингибирования в клетке и корне в целом Lp r согласуются с предсказаниями модели составного транспорта.
В молодых корнях кукурузы существует корреляция между химическим составом апопластных барьеров и корня Lp r . Алифатический суберин, по-видимому, действует как гидрофобный материал, который блокирует поток воды через полосы Каспариана.По сравнению с водой проницаемость корней для растворенных веществ (питательных веществ) низкая. Поглощенные ионы питательных веществ задерживаются в ксилеме и не возвращаются обратно. Существует апопластический компонент транспорта АБК в корнях, который вносит вклад в сигнал стресса в ксилеме (апопластный поток АБК). Сигнал напряжения в ксилеме зависит от интенсивности транспирации. На поток АБК влияет наличие апопластических барьеров (экзодерма). По крайней мере, до некоторой степени АБК, образующаяся в корне во время водного стресса, будет доставлена в апопласт, а затем с потоком транспирации попадет в побег.С другой стороны, АБК увеличивает водопроницаемость мембран клеток корня и осмотическую гидравлическую проводимость целых корней, как показывают измерения с помощью датчика давления клетки и корня. Эти результаты соответствуют составной транспортной модели. Напряжение питательных веществ снижает активность водных каналов и корень в целом. Lp r . В корнях Lotus суточный ритм корня Lp r коррелировал с экспрессией мРНК, кодирующей водные каналы, но не с ритмом в Lp корковых клеток.Был сделан вывод, что регуляция имеет место в энтодерме и / или стеле.
В модели композитного транспорта важную роль играет механизм сцепления-натяжения подъема сока. При высоких скоростях транспирации и напряжения в ксилеме корня предпочтительный апопластический путь для воды обеспечивает достаточное снабжение побегов водой даже в условиях нехватки воды (гидравлический режим потока воды). Был поставлен под сомнение механизм сцепления-натяжения. Прямые измерения давления ксилемы в листьях интактных транспирационных растений кукурузы теперь показывают, что механизм сцепления-натяжения действителен, по крайней мере, для диапазона напряжений до 1 МПа (10 бар).
Дальнейшая работа должна быть сосредоточена на более подробном картировании гидравлических сопротивлений в корневом цилиндре и их изменения в ответ на нехватку воды и другие нагрузки. А именно, для моделирования потока воды через корни необходимо знать гидравлическое сопротивление экзо- и энтодермы, а также стеларных клеток. Это может быть выполнено путем комбинированных измерений с помощью датчика давления в ячейке и корня, а также путем измерения распространения изменений водного потенциала (тургора) по корневому цилиндру.
Эта работа была поддержана грантом Deutsche Forschungsgemeinschaft (DFG), Schwerpunktprogramm «Apoplast».
Список литературы
Azaizeh H, Steudle E.
1991
. Влияние засоления на водный транспорт обрезанных корней кукурузы ( Zea mays L.).Физиология растений
97
,1136
–1145.Azaizeh H, Gunse B, Steudle E.
1992
. Влияние NaCl и CaCl 2 на транспорт воды через корневые клетки проростков кукурузы ( Zea mays L.).Физиология растений
99
,886
–894.Balling A, Zimmermann U.
1990
. Сравнительные измерения давления ксилемы растений Nicotiana с помощью бомбы давления и датчика давления.Planta
182
,325
–346.Barrowclough DE, Peterson CA, Steudle E.
2000
. Радиальная гидравлическая проводимость вдоль развивающихся корней лука.Журнал экспериментальной ботаники
51
,547
–557.Birner TP, Steudle E.
1993
. Влияние анаэробных условий на отношения воды и растворенных веществ, а также на активный транспорт в корнях кукурузы ( Zea mays L.).Планта
190
,474
–483.Брюиг А.
1937
. Permeabilitätsänderungen der Wurzelgewebe, die vom Spross beeinflusst werden.Zeitschrift für Botanik
31
,481
–540.Брауэр Р.
1954
. Регулирующее влияние транспирации и силы всасывания на поглощение воды и соли корнями интактных растений Vicia faba .Acta Botanica Neerlandica
3
,264
–312.Карвахал М., Кук Д.Т., Кларксон Д.Т.
1996
. Реакция растений пшеницы на недостаток питания может включать регуляцию функции водного канала.Планта
199
,372
–381.Chrispeels MJ, Maurel C.
1994
. Аквапорины: молекулярная основа облегчения движения воды через живые клетки растений.Физиология растений
105
,9
–15.Clarkson DT, Carvajal M, Henzler T, Waterhouse RN, Smyth AJ, Cooke DT, Steudle E.
2000
. Гидравлическая проводимость корней: суточная экспрессия аквапоринов и влияние стресса, связанного с питательными веществами.Журнал экспериментальной ботаники
51
,61
–70.Cowan IR.
1977
.Стоматальное поведение и окружающая среда.Успехи ботанических исследований
4
,117
–228.Cruz RT, Jordan WR, Drew MC.
1992
. Структурные изменения и связанное с ними снижение гидравлической проводимости корней Sorghum bicolor L. после воздействия дефицита воды.Физиология растений
99
,203
–212.Fiscus EL.
1975
. Взаимодействие между осмотическим потоком воды и потоком воды, вызванным давлением, в корнях растений.Физиология растений
55
,917
–922.Frensch J, Steudle E.
1989
. Осевое и радиальное гидравлическое сопротивление корням кукурузы ( Zea mays L.).Физиология растений
91
,719
–726.Frensch J, Hsiao TC, Steudle E.
1996
. Транспорт воды и растворенных веществ по развивающимся корням кукурузы.Planta
198
,348
–355.Freundl E, Steudle E, Hartung W.
1998
. Поглощение воды корнями кукурузы и подсолнечника влияет на радиальный транспорт абсцизовой кислоты и концентрацию АБК в ксилеме.Planta
207
,8
–19.Freundl E, Steudle E, Hartung W.
2000
. Апопластический транспорт абсцизовой кислоты через корни кукурузы: эффект экзодермы.Planta
210
,222
–231.Hanson PJ, Sucoff EI, Markhart AH.
1985
. Количественное определение апопластического потока через корневые системы красной сосны с использованием тринатрий-3-гидрокси-5,8,10-пирентрисульфоната.Физиология растений
77
,21
–24.Hartung W, Peuke AD, Davies WJ.
1999
. Абсцизовая кислота — гормональный сигнал стресса на расстоянии у растений при засухе и солевом стрессе. В кн .: Пессаракли М, под ред.Справочник по стрессу растений и сельскохозяйственных культур
, 2-е изд. Нью-Йорк: Марсель Деккер,731
–747.Henzler T, Steudle E.
1995
. Обратимое закрытие водных каналов в междоузлиях Chara обеспечивает доказательство сложной транспортной модели плазматической мембраны.Журнал экспериментальной ботаники
46
,199
–209.Henzler T, Streudle E.
2000
. Транспорт и метаболическое разложение перекиси водорода в Chara corallina : модельные расчеты и измерения с датчиком давления предполагают перенос H 2 O 2 через водные каналы.Журнал экспериментальной ботаники
(в печати).Henzler T, Waterhouse RN, Smyth AJ, Carvajal M, Cooke DT, Schäffner AR, Steudle E, Clarkson DT.
1999
. Суточные колебания гидравлической проводимости и корневого давления можно коррелировать с экспрессией предполагаемых аквапоринов в корнях лотоса Lotus japonicus .Planta
210
,50
–60.Hertel A, Steudle E.
1997
. Функция водных каналов в Chara : температурная зависимость потоков воды и растворенных веществ свидетельствует о переносе композитной мембраны и проскальзывании небольших органических растворенных веществ через водные каналы.Планта
202
,324
–335.Heydt H, Steudle E.
1991
. Измерение отрицательного давления в ксилеме иссеченных корней.Planta
184
,389
–396.Hose E, Steudle E, Hartung W.
2000
. Абсцизовая кислота и гидравлическая проводимость корней кукурузы: исследование клеток корня и датчика давления.Planta
(в печати).Johansson I, Larsson C, Ek B, Kjellbom P.
1996
. Основными интегральными белками плазматических мембран листьев шпината являются предполагаемые аквапорины, которые фосфорилируются в ответ на Ca 2+ и апопластный водный потенциал.Заводская ячейка
8
,1181
–1191.Джонс HG.
1998
. Стоматологический контроль фотосинтеза и транспирации.Журнал экспериментальной ботаники
49
,387
–398.Kjellbom P, Larsson C, Johansson I, Karlsson M, Johanson U.
1999
. Аквапорины и водный гомеостаз растений.Тенденции в растениеводстве
4
,308
–314.Kramer PJ, Boyer JS.
1995
.Водные отношения растений и почвы
.Орландо: Academic Press.Маджио А, Джоли RJ.
1995
. Влияние хлорида ртути на гидравлическую проводимость корневой системы томатов: данные о канально-опосредованном пути.Физиология растений
109
,331
–335.Maurel C.
1997
. Аквапорины и водопроницаемость мембран растений.Ежегодный обзор физиологии растений и молекулярной биологии растений
48
,399
–429.Melchior W, Steudle E.
1993
. Водный транспорт в корнях лука ( Allium cepa L.). Изменения осевой и радиальной гидравлической проводимости при развитии корня.Физиология растений
101
,1305
–1315.Melcher PJ, Meinzer FC, Yount DE, Goldstein G, Zimmermann U.
1998
. Сравнительные измерения давления ксилемы в транспирирующих и нетранспирационных листьях с помощью камеры давления и датчика давления ксилемы.Журнал экспериментальной ботаники
49
,1757
–1760.Миллер ДМ.
1985
. Исследования корневой функции Zea mays . III. Состав ксилемного сока при максимальном корневом давлении свидетельствует об активном переносе в ксилему и измеряет коэффициент отражения корня.Физиология растений
77
,162
–167.Molz FJ, Ferrier JM.
1982
. Математическая трактовка движения воды в клетках и тканях растений: обзор.Завод, клетки и окружающая среда
5
,191
–206.Munns R, Passioura JB.
1984
. Гидравлическое сопротивление растений. III. Влияние NaCl на ячмень и люпин.Австралийский журнал физиологии растений
11
,351
–359.Нобель PS.
1991
.Физико-химическая и экологическая физиология растений
. Сан-Диего: Academic Press Inc.,North GB, Нобелевский PS.
1991
. Изменения гидравлической проводимости и анатомии, вызванные высыханием и повторным увлажнением корней Agave desertii (Agavaceae).Американский журнал ботаники
78
,906
–915.North GB, Нобелевский PS.
1995
. Гидравлическая проводимость концентрических тканей корня Agave desertii Engelm. во влажных условиях и при высыхании.Новый фитолог
130
,47
–57.Passioura JB.
1988
. Водный транспорт внутрь и к корням.Ежегодный обзор физиологии растений и молекулярной биологии растений
39
,245
–265.Peterson CA, Emanuel ME, Humphreys GB.
1981
. Пути движения апопластных флуоресцентных индикаторов красителя через энтодерму в месте образования вторичных корней кукурузы ( Zea mays ) и бобов ( Vicia faba ).Канадский ботанический журнал
59
,618
–625.Peterson CA, Murrmann M, Steudle E.
1993
.Расположение основных барьеров для движения воды и ионов в молодых корнях Zea mays L.Planta
190
,127
–136.Peterson CA, Steudle E.
1993
. Боковая гидравлическая проводимость сосудов ранней метаксилемы в корнях Zea mays L.Планта
189
,288
–297.Peyrano G, Taleisnik E, Quiroga M, de Forchetti SM, Tigier H.
1997
. Влияние засоления на гидравлическую проводимость, содержание лигнина и активность пероксидазы в корнях томатов.Физиология и биохимия растений
35
,387
–393.Radin JW, Eidenbock MP.
1984
. Гидравлическая проводимость как фактор, ограничивающий разрастание листьев хлопчатобумажных растений с дефицитом фосфора.Физиология растений
75
,372
–377.Радин Дж. У., Мэтьюз Массачусетс.
1989
. Водные транспортные свойства клеток коркового слоя корня проростков хлопчатника с дефицитом азота и фосфора.Физиология растений
89
,264
–268.Ригер М, Литвин П.
1999
. Гидравлическая проводимость корневой системы у видов с контрастной анатомией корня.Журнал экспериментальной ботаники
50
,201
–209.Rüdinger M, Hallgren SW, Steudle E, Schulze ED.
1994
. Гидравлические и осмотические свойства корней ели.Журнал экспериментальной ботаники
45
,1413
–1425.Schäffner AR.
1998
. Функция, структура и выражение аквапоринов: в водных отношениях все еще есть сюрпризы.Планта
204
,131
–139.Шрайбер Л.
1996
. Химический состав полосок Casparian, выделенных из Clivia miniata Reg. корни: данные о лигнине.Планта
199
,596
–601.Schreiber L, Hartmann K, Skrabs M, Zeier J.
1999
. Апопластические барьеры в корнях: химический состав стенок энтодермальных и гиподермальных клеток.Журнал экспериментальной ботаники
50
,1267
–1280.Schulze ED.
1986
. Обмен углекислого газа и водяного пара в ответ на засуху в атмосфере и почве.Ежегодный обзор физиологии растений
37
,247
–274.Schütz K, Tyerman SD.
1997
. Водные каналы в Chara corallina .Журнал экспериментальной ботаники
48
,1511
–1518.Скиннер Р.Х., Радин Дж.В.
1994
. Влияние фосфорного питания на поток воды через апопластический обход корней хлопчатника.Журнал экспериментальной ботаники
45
,423
–428.Slovik S, Hartung W.
1992
. Компартментное распределение и перераспределение абсцизовой кислоты в интактных листьях.Planta
187
,37
–xx.Стасовский Э, Петерсон CA.
1991
. Влияние засухи и последующей регидратации на структуру и жизнеспособность корней проростков Zea mays .Канадский ботанический журнал
69
,1170
–1178.Steudle E.
1989
. Поток воды в растениях и его связь с другими процессами: обзор.Методы в энзимологии
174
,183
–225.Steudle E.
1992
. Биофизика воды для растений: разделение, связь с метаболическими процессами и поток воды в корнях растений. В: Somero GN, Osmond CB, Bolis CL, eds,Вода и жизнь: сравнительный анализ водных взаимоотношений на организменном, межклеточном и молекулярном уровнях
. Гейдельберг: Springer ‐ Verlag,173
–204.Steudle E.
1994
.Водный транспорт по корням.Растения и почвы
167
,79
–90.Steudle E.
1995
. Деревья под напряжением.Природа
378
,663
–664.Steudle E.
1997
. Транспорт воды через растительную ткань: роль водных каналов.Биология клетки
89
,259
–273.Steudle E, Brinckmann E.
1989
. Модель осмометра корня: вода и растворенные вещества Phaseolus coccineus .Botanica Acta
102
,85
–95.Steudle E, Frensch J.
1989
. Осмотические реакции корней кукурузы: отношения воды и растворенных веществ.Планта
177
,281
–295.Steudle E, Frensch J.
1996
. Водный транспорт в растениях: роль апопласта.Растения и почвы
187
,67
–79.Steudle E, Henzler T.
1995
. Водные каналы в растениях: меняются ли основные концепции водного транспорта?Журнал экспериментальной ботаники
46
,1067
–1076.Steudle E, Heydt H.
1988
. Искусственная осмотическая ячейка: модельная система для моделирования осмотических процессов и изучения явлений отрицательного давления в растениях.Завод, клетки и окружающая среда
11
,629
–637.Steudle E, Heydt H.
1997
. Водный транспорт через корни деревьев. В: Rennenberg H, Eschrich W, Ziegler H, eds.Деревья — вклад в современную физиологию деревьев
. Лейден, Нидерланды: Backhuys Publishers,239
–255.Steudle E, Jeschke WD.
1983
. Водный транспорт в корнях ячменя.Planta
158
,237
–248.Steudle E, Мещеряков А.Б.
1996
. Гидравлические и осмотические свойства корней дуба.Журнал экспериментальной ботаники
47
,387
–401.Steudle E, Murrmann M, Peterson CA.
1993
. Транспорт воды и растворенных веществ через корни кукурузы изменен путем прокалывания энтодермы. Еще одно свидетельство составной транспортной модели корня.Физиология растений
103
,335
–349.Steudle E, Oren R, Schulze ED.
1987
. Водный транспорт в корнях кукурузы.Физиология растений
84
,1220
–1232.Steudle E, Peterson CA.
1998
. Как вода проходит через корни.Журнал экспериментальной ботаники
49
,775
–788.Steudle E, Zimmermann HM, Henzler T.
1999
. Поглощение воды корнями растений: способы регулирования. В: Ito O, O’Toole J, Hardy B, eds,Генетическое улучшение риса для окружающей среды с ограниченными водными ресурсами
.Лос-Баньос, Манила, Филиппины: Международный научно-исследовательский институт риса (IRRI).73
–88.Taleisnik E, Peyrano G, Cordoba A, Arias C.
1999
. Водоудерживающая способность в сегментах корня, различающихся степенью развития экзодермы.Анналы ботаники
83
,19
–27.Tomos AD, Leigh RA.
1999
. Датчик давления: универсальный инструмент в физиологии растительных клеток.Ежегодный обзор физиологии растений и молекулярной биологии растений
50
,447
–472.Tyerman SD, Bohnert H, Maurel C, Steudle E, Smith JAC.
1999
. Аквапорины растений: их молекулярная биология, биофизика и значение для водных отношений растений.Журнал экспериментальной ботаники
50
, специальный выпуск ,1055
–1071.Тайри MT, Сперри Дж. С..
1988
. Действуют ли древесные растения на грани катастрофической дисфункции ксилемы, вызванной динамическим водным стрессом?Физиология растений
88
,574
–580.Weatherley PE.
1982
. Впитывает воду и проникает в корни. В: Lange OL, Nobel PS, Osmond CB, Ziegler H, eds,Энциклопедия физиологии растений
, Vol. 12B. Берлин: Springer ‐ Verlag,79
–109.Wei C, Steudle E, Tyree MT.
1999
. Подъем воды в растениях: есть ли веские основания для продолжающихся споров?Тенденции в растениеводстве
4
,372
–375.Wei C, Tyree MT, Steudle.
1999
. Прямое измерение давления ксилемы в листьях интактных растений кукурузы. Проверка теории сцепления – растяжения с учетом гидравлической архитектуры.Физиология растений
121
,1191
–1205.Wei C, Steudle E, Tyree MT.
2000
. Ответ: Подъем воды в растениях.Тенденции в растениеводстве
5
,146
–147.Westgate ME, Steudle E.
1985
. Водный транспорт в средней жилке листьев кукурузы.Физиология растений
78
,183
–191.Ян С., Тайри MT.
1994
. Гидравлическая архитектура Acer saccharum и A . rubrum : сравнение ветвей и целых деревьев и вклад листьев в гидравлическое сопротивление.Журнал экспериментальной ботаники
45
,179
–186.Zeier J, Schreiber L.
1998
. Сравнительное исследование клеточных стенок первичной и третичной энтодермальных клеток, выделенных из корней пяти видов однодольных: химический состав в зависимости от тонкой структуры.Planta
(в печати).Zhang WH, Tyerman SD.
1999
. Ингибирование водных каналов HgCl 2 в интактных клетках корней пшеницы.Физиология растений
120
,849
–858.Zhu GL, Steudle E.
1991
. Водный транспорт через корни кукурузы.Физиология растений
95
,305
–315.Zimmermann HM, Steudle E.
1998
. Апопластический транспорт через молодые корни кукурузы: эффект экзодермы.Planta
206
,7
–19.Zimmermann HM, Hartmann K, Schreiber L, Steudle E.
2000
. Химический состав апопластных транспортных барьеров в зависимости от радиальной гидравлической проводимости корней кукурузы ( Zea mays L.).Planta
210
,302
–311.© Общество экспериментальной биологии
Техасский центр исследований и распространения знаний A&M AgriLife в Сан-Анджело
Тензиометр представляет собой трубку, заполненную водой, со специальным пористым наконечником и вакуумметром. Этот инструмент измеряет всасывание воды из почвы, что аналогично процессу, который корень растения использует для получения воды из почвы.
Доступны тензиометры различной длины, как показано на рисунке слева.При установке только верхняя часть заполненной водой трубки и вакуумметр видны для показаний и технического обслуживания в полевых условиях, как показано справа.
Тензиометры относительно недорогие и многоразовые. Они просты в эксплуатации, хотя требуют некоторого обслуживания в полевых условиях. Техническое обслуживание требует добавления воды и использования ручного вакуумного насоса для удаления воздуха из трубки.
Доступны тензиометры длиной от 6 до 72 дюймов, что позволяет устанавливать их в почве на различной глубине.Один тензиометр измеряет существующие условия влажности почвы только на глубине пористого наконечника и не может контролировать условия выше или ниже этой точки. Два или более тензиометра различной длины могут быть установлены на одном участке для мониторинга состояния влажности почвы на более чем одной глубине в корневой зоне.
Всасывание грунтовой воды
Вода хранится в почве в виде пленки вокруг каждой частицы почвы и в поровых пространствах между частицами почвы. Вода, хранящаяся в поровых пространствах, удерживается поверхностным натяжением, и растение легче всего извлекает ее.Водная пленка вокруг каждой частицы почвы удерживается более сильными молекулярными силами, и растению гораздо труднее выйти из почвы.
Вода из почвы втягивается или всасывается в корень растения из-за более высокой концентрации солей в корне растения. Этот процесс извлечения воды называется «отсасыванием воды из почвы». Когда почва заполнена полем, растение должно оказывать всасывание, равное от двух до пяти фунтов на квадратный дюйм (psi), чтобы получить необходимое количество воды. По мере того, как почва становится суше, количество всасывания растением должно увеличиваться, чтобы получить достаточное количество воды для своих нужд.
Постоянная точка увядания или гибель растения происходит, когда почва высыхает до уровня, при котором растение должно оказывать всасывающее давление 220 фунтов на квадратный дюйм или более в попытке удовлетворить свои потребности в воде.
По сути, тензиометр измеряет количество всасываемой влаги из почвы, необходимое растению для извлечения воды из почвы. Чем суше почва, тем сильнее всасывание и выше показания тензиометрического вакуумметра.
Когда корни растений удаляют воду из почвы и почва становится суше, вода всасывается из пористого наконечника тензиометра в почву до тех пор, пока не будет достигнуто состояние равновесия между влажностью почвы и всасыванием тензиометра.Вода, добавленная в почву в результате дождя или орошения, обратит это действие. Более сильное всасывание, созданное ранее в тензиометре, будет втягивать воду обратно в тензиометр и понижать показания датчика.
Тензиометры измеряют отрицательное всасывание почвы до 14,7 фунтов на квадратный дюйм (100 по шкале), что является лишь частью доступной влажности почвы. Когда всасывание почвы превышает примерно 12 фунтов на квадратный дюйм (80 на манометре), водная пленка, покрывающая пористый наконечник, разрывается, что позволяет воздуху проникать в трубку. Показания манометра резко падают, затем восстанавливаются, пока воздух снова не войдет в систему.Когда это происходит, почву необходимо повторно увлажнить, а тензиометр снова заполнить, прежде чем он снова заработает должным образом.
Как показано на датчиках слева, чем выше показание, тем суше почва. Датчик, показывающий 66, указывает на то, что растение должно прикладывать давление около десяти фунтов на квадратный дюйм, чтобы извлекать воду из окружающей почвы. Датчик с показанием 9,5 указывает на то, что для извлечения воды из почвы потребуется один фунт на квадратный дюйм.
Тензиометры для различных текстур почвы
Тензиометры надежны в определенных пределах.Например, диапазон полезности зависит от текстуры почвы. Чем мельче частицы почвы (как в почвах с высоким содержанием глины), тем больше воды может удержать почва, но тем тяжелее растение должно работать, чтобы извлечь влагу из почвы. Чем крупнее частицы (как в песчаных почвах), тем меньше влаги будет удерживать почва, но больше влаги доступно для использования растениями. Следовательно, тензиометры могут измерять более широкий диапазон доступной влажности в песчаной почве, чем в почве с высоким содержанием глины.
В песчаных почвах показание шкалы 80 указывает на влажность почвы около 40 процентов от емкости поля.На песчаных почвах большинству сельскохозяйственных культур требуется дополнительная поливная вода для максимального роста растений при примерно 50 процентах емкости поля, что составляет примерно 60 на тензиометре. В песчаных почвах тензиометры могут измерять влажность почвы от 100 до 40 процентов полевой емкости, потому что относительно большие поры в этих почвах выделяют воду при более низком напряжении.
В почвах с высоким содержанием глины тензиометр будет показывать около 80, когда влажность почвы составляет около 75 процентов от полевой емкости.Глины удерживают воду при большем напряжении, чем песчаные почвы; поэтому тензиометр прерывает всасывание при более высоком уровне влажности в почвах с высоким содержанием глины. Когда датчик показывает 80, тензиометр прекращает всасывание и перестает работать. Это происходит до того, как почва высохнет до уровня, необходимого для полива большинства культур. Тензиометры могут измерять влажность почвы только в пределах от 100 до 75 процентов полевой емкости в глинистых почвах, потому что более мелкие поровые пространства удерживают воду при более высоком напряжении.
В ПОЧВАХ СО СРЕДНЕЙ текстурой диапазон измерения будет промежуточным по сравнению со значениями, показанными для песчаных и глинистых почв.
Текстура почвы также определяет скорость проникновения влаги с поверхности земли в корневую зону. Тензиометры могут помочь определить, сколько влаги действительно проникло и как быстро.
Тензиометры различных культур
Тензиометры лучше всего использовать, когда влажность почвы будет поддерживаться на уровне от 50 до 75 процентов от емкости поля, например, при выращивании культур с высокими требованиями к влажности, таких как кукуруза или овощи.
Они не измеряют диапазоны низкой влажности почвы, при которых такие культуры, как хлопок, сорго или другие мелкие зерна, могут извлекать достаточную влажность почвы.Однако при установке в начале сезона они могут указывать на глубину развития корней и извлечение воды из почвы.
Установка
Тензиометры обычно устанавливаются после укоренения урожая, и они не мешают нормальной обработке почвы в течение вегетационного периода.
Установка тензиометра может быть произведена путем ввинчивания стандартной полудюймовой трубы или другого кернового инструмента в землю на желаемую глубину, а затем удаления ее, чтобы создать отверстие точно нужного размера и глубины для тензиометра.По словам производителей, этот метод обеспечивает хороший контакт между почвой и инструментом с минимальным повреждением существующих корней и структуры почвы.
Тензиометры устанавливаются в плотно прилегающее отверстие, затем с помощью вакуумного насоса удаляется воздух из трубки. Датчик на вакуумном насосе также используется для проверки правильности работы датчика на тензиометре.
Подробные инструкции по правильной установке предоставляются производителями тензиометров.Независимо от того, установлен ли тензиометр, это следует делать с минимальным нарушением структуры почвы, типичной для данного участка. Установка в отверстие большого диаметра с последующей засыпкой не рекомендуется.
Для полива по бороздам или паводком станции можно размещать примерно на двух третях пути вниз. Однако, если пробег особенно длинный, могут потребоваться станции на каждом конце поля.
Важными моментами при выборе размещения тензиометров для спринклерного орошения являются отсутствие препятствий между дождевателем и тензиометром, а также отсутствие концентрации избыточной воды в месте установки.
Учет
Успех любой системы мониторинга влажности почвы заключается в тщательном ведении записей. Со временем ирригатор лучше поймет показания и вариации. Ниже приведен пример графика, показывающего влияние осадков, прохладной погоды и проникновения влаги.
Записи в течение вегетационного периода показывают влияние орошения и дождевых осадков на влажность почвы. Обратите внимание, что показания глубины 12 дюймов быстро реагировали на осадки и орошение в июне и июле, однако на показания глубины 36 дюймов эти события не повлияли.
Недостатки
Основным недостатком тензиометра является то, что он может работать только тогда, когда почва относительно влажная. Кроме того, показания прибора следует проводить ежедневно при высоком уровне использования воды растениями для обнаружения ложных показаний из-за попадания пузырьков воздуха в трубку. Температура ниже точки замерзания также может серьезно повредить тензиометр. Его необходимо вынуть из земли и хранить до того, как температура упадет до нуля.
Переносные тензиометры
Также доступны портативные тензиометры.Они включают в себя небольшой пористый керамический сенсорный наконечник, который сокращает время считывания. Это позволяет пользователю быстро снимать показания выборочной проверки в различных местах. Они очень чувствительны. Следовательно, в измерения легко внести ошибку. Они также требуют большего обслуживания и осторожности при обращении, чем стационарные тензиометры.
Благодарности
Эта информация была написана Майком Райзингером, USDA-NRCS, и Кеном Карвером, Район охраны подземных вод High Plains No.1
9 типов корней деревьев, растений и цветов
Вот ваше полное руководство по корням … корневым системам деревьев, растений и цветов. Мы излагаем и объясняем различные типы корней, а также функции корней (что они делают) и включаем иллюстрацию, показывающую анатомию корневой системы.
Корни — это нелистовая часть тела растения, на которой нет узлов. Это орган, который находится под поверхностью почвы.Корни также могут быть воздушными, то есть растущими над поверхностью земли, или аэрированными, что означает, что они плавают над поверхностью воды. Корни отвечают за обеспечение стеблей и листьев достаточным количеством воды и питательных веществ для их роста.
Связанные: Типы растений в горошек | Типы газовых заводов | Инструменты для обрезки растений | Типы нефритовых растений
Функции корнейКорни составляют основную часть тела растения с точки зрения объема и функций.Основные функции корней следующие:
Якорь и опораКорневая система растения обеспечивает физическую поддержку, прикрепляя тело растения к почве. Многие растения могут стоять прямо в течение сотен лет, потому что их корни глубоко уходят в почву и надежно удерживают растения на месте.
Поглощение и проводимостьКорни имеют корневые волоски, через которые корни поглощают воду и питательные вещества из почвы, которые необходимы для роста растений.Корни обладают способностью поглощать неорганические питательные вещества против градиента концентрации. После того, как вода и питательные вещества впитаются, они перемещаются вверх к стеблю и листьям.
В пустынях корни уходят глубоко в постоянные запасы воды. Считается, что пустынные районы, где растут растения, имеют запасы подземных вод. Это очень помогает в принятии решения, где копать колодец.
ТранспортировкаКорни имеют специально разработанные каналы для транспортировки поглощенных питательных веществ и воды к стеблям и листьям.Кроме того, у них также есть каналы, по которым органические продукты могут транспортироваться от надземных частей растения к корням.
ХранениеНекоторые корнеплоды, такие как морковь и сладкий картофель, служат хранилищами. В них хранятся углеводы и вода. В корнях некоторых растений, содержащихся в десертах, может храниться до 70 кг воды.
ФотосинтезФотосинтез — это процесс, с помощью которого растения готовят себе пищу.Некоторые корни способны к фотосинтезу, как, например, воздушные корни мангровых растений и эпифитных орхидей.
АэрацияУ растений, которые растут на поверхности стоячей воды, есть специальные корни, называемые пневматофорами, которые распространяют кислород из воздуха.
ДвижениеСокращающиеся корни многих луковичных растений тянут растение вниз в почву. Подземная среда более благоприятна для луковичных растений.
РазмножениеНекоторые особые корни могут воспроизводиться. Они служат средством сохранения вида. У некоторых растений, таких как зрелый агохо, вокруг ствола видны обильно растущие побеги от корней, растущих горизонтально.
Анатомия корнейКорень имеет четыре основных области :
- Корневой колпачок
- Меристематический участок
- Область удлинения
- Область созревания
Корневой колпачок присутствует на вершине корня.Это структура, похожая на наперсток. Верхушка корня нежная. Корневая шляпка защищает этот нежный конец и помогает корню глубже проникнуть в почву. Корневая шляпка выделяет слизь, которая отвечает за смазку прохода корня в почве. Эта слизь также способствует усвоению воды и питательных веществ. Корневой колпачок изнашивается, но его постоянно обновляют.
Меристематический регионТермин «меристематический» означает быстрый рост. Эта область находится чуть выше корневой крышечки.Клетки в этой области быстро растут. Они очень маленькие и очень тонкостенные. У них плотная протоплазма.
Область удлиненияКлетки в области удлинения быстро увеличиваются и удлиняются. Клетки в этой области отвечают за рост длины корня.
Область созреванияКлетки в области удлинения постепенно созревают после дифференцировки и образуют область созревания.Клетки дифференцируются, образуя различные специализированные ткани, такие как постоянная область и корневые волоски.
Корни однодольных и корни двудольныхКорни однодольных растений называются корнями однодольных, а корни двудольных растений называются корнями двудольных.
Однодольные растения имеют одну семядоль в зародыше. У однодольных корней ксилема и флоэма расположены по кругу. У двудольных растений в зародышах есть две семядоли. У двудольных корней ксилема находится в середине, а флоэма ее окружает.
Разница между корнями однодольных и корнями двудольныхРазличия между корнями однодольных и корнями двудольных следующие:
Корни однодольных дает начало только. Сердцевина хорошо развита, крупная. В корнях однодольных растений много ксилемы и флоэмы. Вторичного роста нет.
Двудольные корниПерицикл дает начало нескольким боковым корням, пробковому камбию и некоторой части сосудистого камбия.Шаг отсутствует. Количество ксилемы и флоэмы ограничено. Вторичный рост происходит в корне двудольного дерева.
Типы корнейСуществуют разные типы корней в зависимости от типа и вида растений. Основные типы:
1. Волокнистые корниВолокнистые корни встречаются у однодольных растений. Они тонкие, разветвленные, растут прямо из стебля. Эти корни имеют тенденцию расти близко к поверхности и распространяться горизонтально.Они имеют гроздевидный вид с многочисленными корнями вместе, почти одинакового размера. В мочковатой корневой системе первичный корень недолговечен. Его заменяют многочисленные корни.
Волокнистая корневая система не обеспечивает надежного закрепления растений, так как они не проникают глубоко в почву.
Характеристики:
- Волокнистые корни растут от основания стебля
- Они находятся под землей
- Они представлены в виде скоплений корней одинакового размера
- Первичный корень заменен придаточными корнями
- Не проникают глубоко в почву.
стержневые корни встречаются у большинства двудольных растений. Этот тип корня представляет собой прямое удлинение корня. В отличие от волокнистых корней, стержневые корни не являются ветвями. Вместо этого они представляют собой единственный первичный корень, который прорастает глубоко в почву. Главный корень дает начало боковым ветвям (вторичным и третичным корням), что приводит к образованию системы стержневых корней.
Ветви стержневого корня растут в акропетальной последовательности, что означает, что более длинные и старые находятся у основания, а более новые и короткие — около вершины основного корня.
Система Taproot обеспечивает очень прочное закрепление. Причина такой сильной опоры в том, что они глубоко проникают в почву.
У некоторых растений стержневой корень не зарастает слишком глубоко. Вместо этого его боковые ветви растут горизонтально по поверхности. Эти типы корней называются фидерными корнями.
Характеристики:
- Стержневые корни развиваются из корня зародыша
- Они всегда находятся под поверхностью почвы
- Они имеют один главный корень (первичный корень) с ветвями, которые растут акропетально
- Первичный корень остается на протяжении всей жизни растения.
- Они действуют как глубокие питатели, то есть проникают глубоко в почву.
Придаточные корни аналогичны волокнистым корням. Однако они могут быть подземными или воздушными (над землей). Они могут расти из любой части растения, кроме прикорневой. Обычно они растут из интермодальных узлов, стеблевых узлов и листьев. Эти корни могут быть толстыми, тонкими или модифицированными в зависимости от вида. Придаточные корни возникают в стрессовых условиях, например, при заболачивании после паводков.
Обрезки листьев и отводок у таких растений, как роза, могут привести к развитию придаточных корней.Также они развиваются при повреждении растений. Они могут увеличить шансы растения на выживание, поскольку растение размножается с помощью придаточных корней.
4. Ползучие корниПолзучие корни — это типы корней, которые не проникают глубоко в почву. Они неглубокие и широко распространены по горизонтали от основания растения. У многих деревьев ползучие корни .
5. Клубневые корниКлубневые корни — это очень толстые корни.В них хранится значительное количество пищи, чтобы прокормить все растение. Это мясистый, увеличенный и видоизмененный орган хранения. Они модифицированы из стебля.
Размножение клубневых корней осуществляется делением кроны, при котором у каждого деления кроны есть несколько почек и достаточно места для хранения, чтобы создать новое растение.
6. Водные корниВодные корни — это типы корней, которые растут в воде. Они более тонкие и хрупкие.У них есть способность пропускать кислород из атмосферы, который затем используется корнями для обмена веществ и роста. Они морфологически отличаются от почвенных корней.
7. Корни паразитовКорни паразитов — это типы корней, которые прикрепляются к другому растению и высасывают из него питательные вещества. Они не приносят никакой пользы растению-хозяину. Вместо этого они наносят серьезный ущерб, отсюда и название, корни паразита.
Факты о корняхФакты о корнях, перечисленные ниже, являются некоторыми удивительными указателями, о которых, мы уверены, вы никогда раньше не слышали:
- У большинства растений рост под землей (как корни ) столько же, сколько рост над землей.
- У сельскохозяйственных культур, выращиваемых на суше, 90% корней находятся в верхних 10 см почвы. Оставшиеся 10% корней вырастают на глубину до 2 м, чтобы достичь глубокой воды и выжить.
- В идеальных условиях корни могут расти со скоростью до 1 см в день.
- Почва прочностью менее 3 мегапаскалей подходит для роста корней.
- Корни растут случайным образом, но они растут быстрее и сильнее разветвляются на участках с благоприятными почвенными условиями.
- Рост корней идет не в сторону воды или удобрений. Вместо этого они растут вокруг этих областей, поскольку это способствует росту.
- Большая часть воды и удобрений впитывается корнями на несколько миллиметров выше кончика роста.
- Старые корни служат трубой для передачи воды и питательных веществ другим частям растения.
- В областях, где болезнь поражает кончик корня или где рост ограничен, потребление воды и питательных веществ будет ограничено.
Корни широко используются человеком. Некоторые из наиболее известных применений корней для человека:
- Многие корни используются людьми в пищу. Эти корни включают морковь, батат, картофель и редис.
- Корни являются источником некоторых важных лекарств, которые могут спасти жизнь. Растительные лекарства, такие как женьшень, ипекака, раувольфия, ашваганда, получают из корней.
- Корни содержат много клетчатки. Волокно из корней используется для изготовления метел, корзин, щеток и т. Д.
- Корни помогают предотвратить эрозию почвы путем сохранения почвы.
- Предотвращая эрозию почвы, они играют важную роль в предотвращении опустынивания.
Как упоминалось ранее, корни являются источником очень важных лекарств .Мы перечислили для вас самые известные.
1. КуркумаКорень куркумы способен лечить все, от боли в желудке до артрита, заболеваний желчного пузыря и печени, менструальных спазмов, инфекций и головных болей. Куркума в основном используется в качестве специи, но в лечебных целях корень куркумы просто феноменален.
2. ИмбирьИмбирь эффективен при диарее, тошноте и расстройстве желудка.Его также можно использовать для облегчения простуды, заложенности носа, гриппа и головных болей.
3. СолодкаСолодка издавна использовалась для лечения язвы желудка, бронхита и ангины. Согласно некоторым исследованиям, солодка также может быть полезна при лечении гепатита С.
4. АшвагандаЭтот лекарственный корень можно использовать для лечения артрита, заболеваний печени и фибромиалгии.
5. ВалерианаВалериана используется для лечения нарушений сна. Многие люди, которые пытаются избавиться от привычки употреблять снотворное, получили пользу от валерианы. Некоторые исследования показывают, что эта трава может заставить хронических бессонниц легче и быстрее засыпать.
Кто знал, что корни являются такой волшебной частью растения? Они не только сердце и легкие растения, но и приносят огромную пользу людям.
Связанные: Типы папоротниковых растений | Типы прибрежных растений | Растения и цветы, привлекающие бабочек | Типы хостов
Leave A Comment