
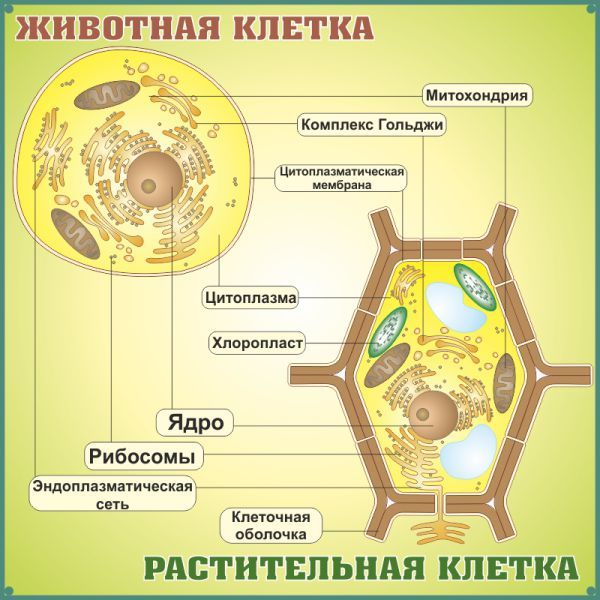
Строение животной клетки
☰
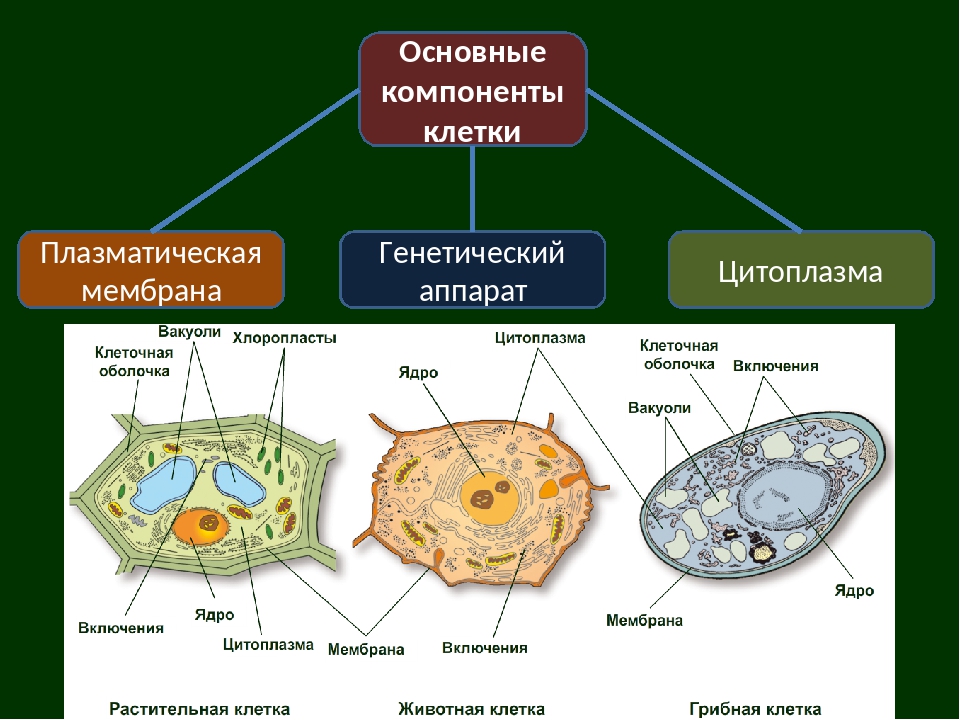
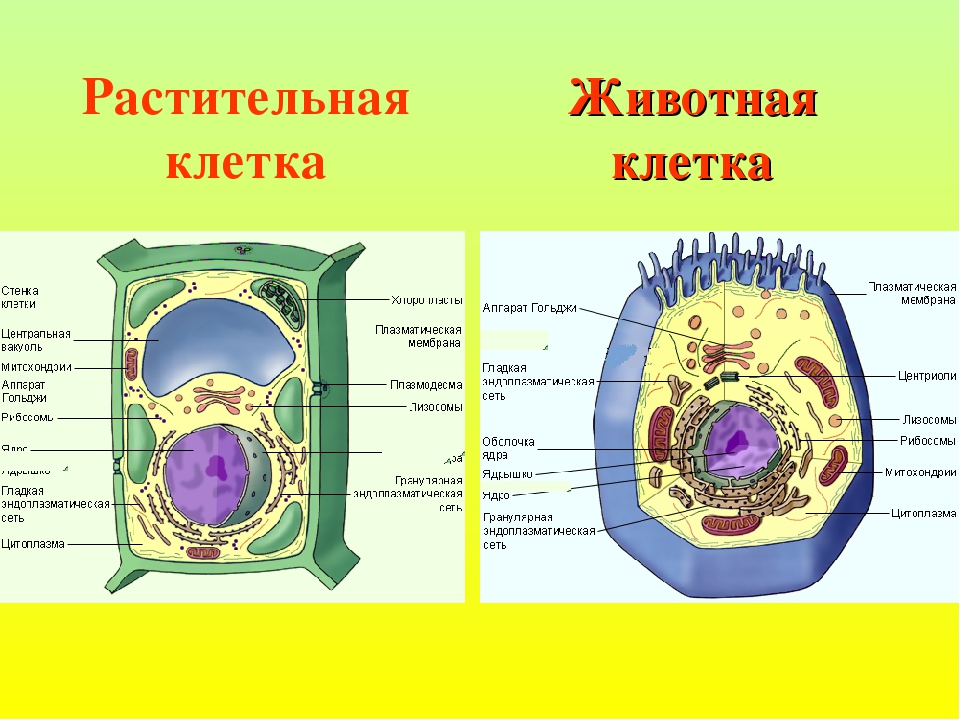
Все живые организмы имеют во многом схожее клеточное строение. Однако у клеток разных царств живого имеются свои особенности. Так клетки бактерий не имеют ядер, а у клеток растений есть жесткая целлюлозная клеточная стенка и хлоропласты. Строение животных клеток также имеет свои характерные особенности.
Чаще всего клетки животных мельче, чем клетки растений. По форме они очень разнообразны. Форма и строение животной клетки зависит от выполняемых ею функций. У сложно организованных животных тела состоят из множества тканей. Каждую ткань составляют свои клетки, имеющие характерные для них особенности строения. Но несмотря на все разнообразие, можно выделить общее в строении всех животных клеток.
От внешней среды содержимое клетки животного ограничено только клеточной мембраной. Она эластична, поэтому многие клетки имеют неправильную форму, могут незначительно изменять ее. Мембрана имеет сложное строение, в ней выделяют два слоя.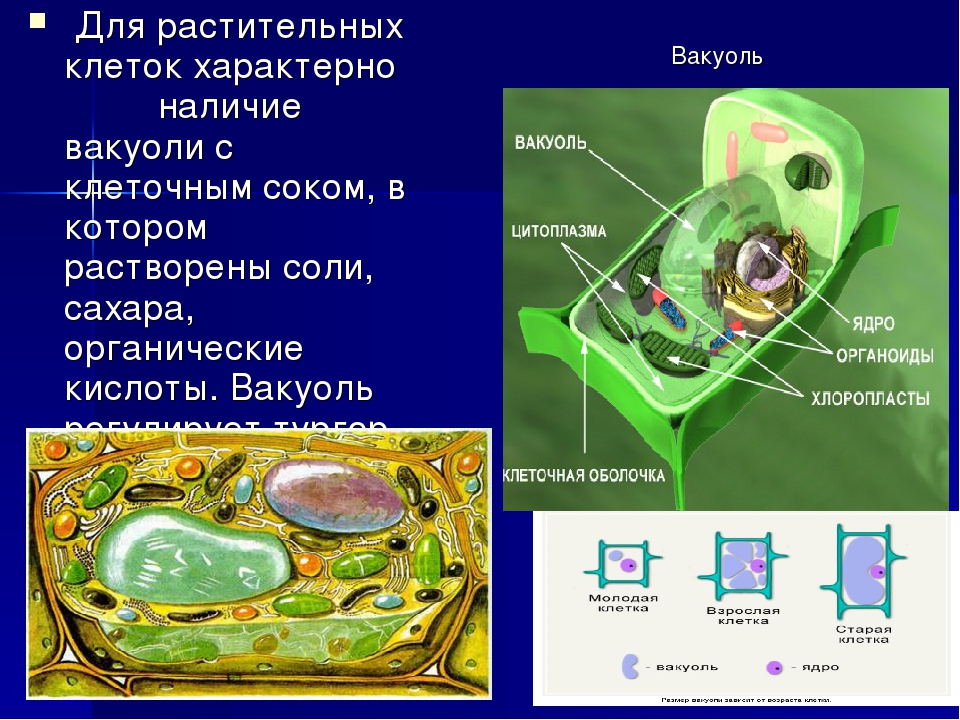
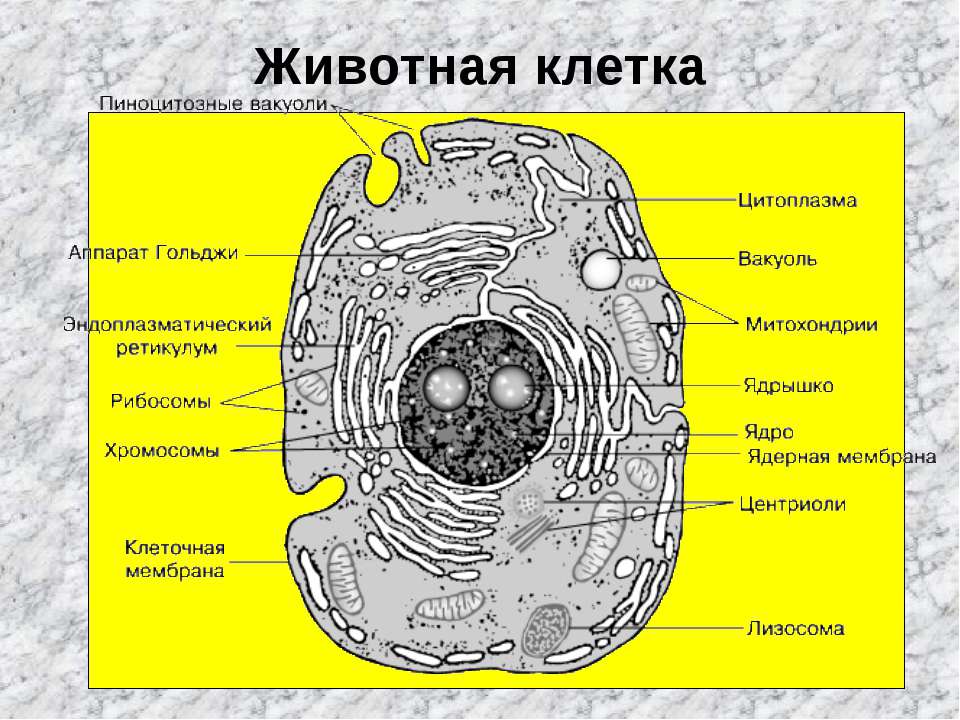
Внутри животной клетки содержится цитоплазма, ядро, органоиды, рибосомы, различные включения и др. Цитоплазма представляет собой вязкую жидкость, находящуюся в постоянном движении. Движение цитоплазмы способствует протеканию различных химических реакций в клетке, т. е. обмену веществ.
Во взрослой растительной клетке есть большая центральная вакуоль. В животной клетке такой вакуоли нет. Однако в животных клетках постоянно образуются и исчезают маленькие вакуоли
Строение животной клетки отличается от растительной еще тем, что в животной клетке достаточно большое ядро располагается обычно в центре (а у растений оно смещено из-за наличия большой центральной вакуоли). Внутри ядра содержится ядерный сок, а также находятся ядрышко и хромосомы. Хромосомы содержат наследственную информацию, которая при делении передается дочерним клеткам. Также они управляют жизнедеятельностью самих клеток.
Хромосомы содержат наследственную информацию, которая при делении передается дочерним клеткам. Также они управляют жизнедеятельностью самих клеток.
У ядра есть своя мембрана, отделяющая его содержимое от цитоплазмы. Кроме ядра в цитоплазме клетки есть другие структуры, имеющие собственные мембраны. Эти структуры называют органоидами клетки, или, по-другому, органеллами клетки. В обычной по строению животной клетке, кроме ядра, есть следующие органоиды: митохондрии, эндоплазматическая сеть (ЭПС), аппарат Гольджи, лизосомы.
Митохондрии — это энергетические станции клетки. В них образуется АТФ — органическое вещество, в последствие при расщеплении которого выделяется много энергии, обеспечивающей протекание процессов жизнедеятельности в клетке. Внутри митохондрии есть множество складок — крист.
Эндоплазматическая сеть состоит из множества каналов, по которым транспортируются синтезируемые в клетке белки, а также другие вещества. По каналам ЭПС вещества поступают в аппарат Гольджи, который в животных клетках выражен сильнее, чем в растительных. В аппарате Гольджи, который представляет собой комплекс трубочек, вещества накапливаются. Далее по мере надобности они будут использованы в клетке. Кроме того на мембране аппарата Гольджи происходит синтез жиров и углеводов для построения всех мембран клетки.
По каналам ЭПС вещества поступают в аппарат Гольджи, который в животных клетках выражен сильнее, чем в растительных. В аппарате Гольджи, который представляет собой комплекс трубочек, вещества накапливаются. Далее по мере надобности они будут использованы в клетке. Кроме того на мембране аппарата Гольджи происходит синтез жиров и углеводов для построения всех мембран клетки.
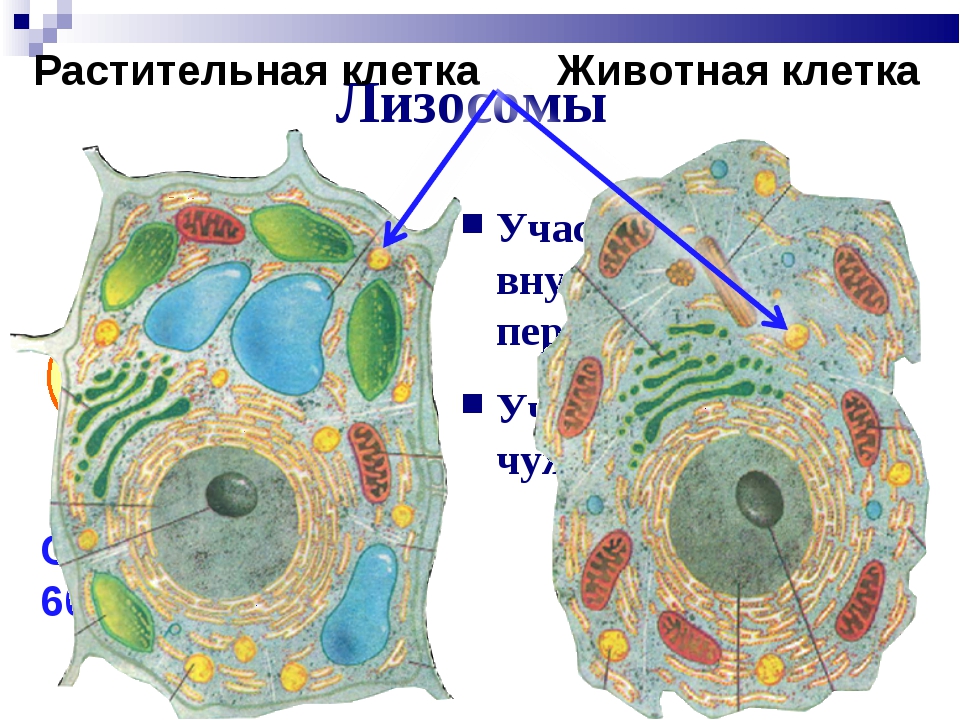
В лизосомах содержатся вещества, расщепляющие ненужные клетке и вредные для нее белки, жиры и углеводы.
Кроме органелл, окруженных мембраной, в животных клетках есть немембранные структуры: рибосомы и клеточный центр. Рибосомы есть в клетках всех организмов, а не только у животных. А вот клеточного центра у растений нет.
Рибосомы располагаются группами на эндоплазматической сети. ЭПС, покрытая рибосомами, называется шероховатой. Без рибосом ЭПС называется гладкой. На рибосомах происходит синтез белков.
Клеточный центр состоит из пары цилиндрических телец.
Клеточные включения представляют собой различные капли и зерна, состоящие из белков жиров и углеводов. Они постоянно присутствуют в цитоплазме клетки и участвуют в обмене веществ.
ЧАСТЬ 3 — КОНТРОЛЬНЫЕ РАБОТЫ — ЕГЭ 100 баллов. Биология. Самостоятельная подготовка к ЕГЭ — ЕГЭ 2018 — Произведения школьной программы
ЕГЭ 100 баллов. Биология. Самостоятельная подготовка к ЕГЭ
ВАРИАНТ 2
ЧАСТЬ 1
Ответом к заданиям 1-21 являются последовательность цифру число или слово (словосочетание).
1. Рассмотрите предложенную схему. Запишите в ответе пропущенный термин, обозначенный на схеме знаком вопроса.
Ответ:_______________.
2. Выберите три верных ответа из шести и запишите в таблицу цифры, под которыми они указаны.
Палеонтология изучает
1) сезонные миграции птиц
2) отпечатки ископаемых растений
3) взаимоотношения живых организмов
4) останки древних животных
5) влияние внешних факторов на сельскохозяйственных животных
6) следы жизнедеятельности древних организмов
3. В половой клетке пшеницы 21 хромосома. Какой набор хромосом имеют соматические клетки этого растения? В ответе запишите только количество хромосом.
В половой клетке пшеницы 21 хромосома. Какой набор хромосом имеют соматические клетки этого растения? В ответе запишите только количество хромосом.
Ответ:________________.
4. Выберите три верных ответа из шести и запишите в таблицу цифры, под которыми они указаны. Для животной клетки не характерно наличие
1) хлоропластов
2) рибосом
3) целлюлозной клеточной стенки
4) оформленного ядра
5) вакуолей с клеточным соком
6) митохондрий
5. Установите соответствие между характеристикой и типом деления клеток, к которому ее относят.
Запишите в таблицу выбранные цифры под соответствующими буквами.
6. Сколько молекул АТФ будет синтезировано в клетках амебы га подготовительном этапе энергетического обмена, в процессе гликолиза и в процессе дыхания при окислении фрагмента молекулы гликогена, состоящего из 12 остатков глюкозы?
Ответ запишите в виде последовательности цифр, в порядке их убывания.
Ответ:_______________.
7. Ниже приведен перечень характеристик изменчивости. Все они. кроме трех, используются для описания комбинативной изменчивости. Найдите три характеристики, «выпадающие» и: общего ряда, и запишите в таблицу цифры, под которыми онг указаны.
1) приводит к изменению генов
2) является ненаследственной изменчивостью
3) возникает в результате полового размножения
4) возникает в результате вегетативного размножения
5) проявляется в генотипическом разнообразии особей
6) повышает выживаемость вида в изменяющихся условиях его существования
8. Установите соответствие между зародышевыми листками, обозначенными на рисунке цифрами 1, 2, и структурами
организма, которые из них развиваются. К каждой позиции, данной в первом столбце, подберите соответствующую позицию из второго столбца.
Запишите в таблицу выбранные цифры под соответствующими буквами.
9. Выберите три верных ответа из шести и запишите в таблицу цифры, под которыми они указаны.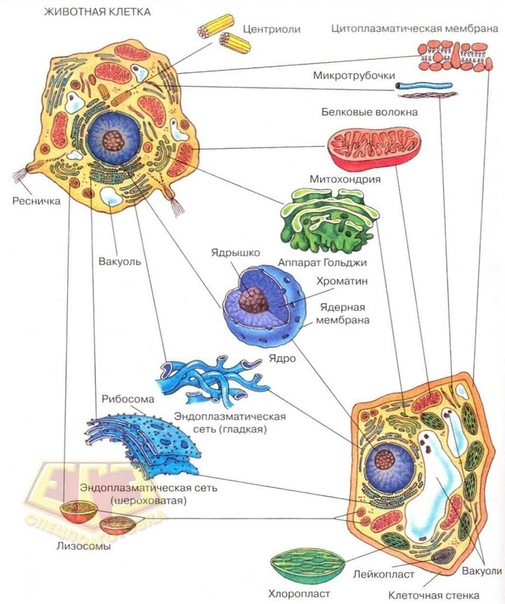
1) имеют вегетативные и генеративные органы
2) тело представляет собой таллом, или слоевище
3) имеют разнообразные ткани
4) обычно живут в водной среде
5) включают одноклеточные организмы
6) имеют цветки и плоды
10. Установите соответствие между признаком и классом животных, для которого он характерен.
Запишите в таблицу выбранные цифры под соответствующими буквами.
11. Укажите последовательность явлений и процессов, происходящих при биосинтезе белка. Запишите в таблицу соответствующую последовательность цифр.
1) образование пептидной связи
2) синтез молекулы иРНК на ДНК
3) связывание молекулы иРНК с рибосомой
4) поступление молекулы иРНК из ядра в цитоплазму
5) процесс терминации
6) взаимодействие тРНК с аминокислотой метионином, с бел- ково-синтезирующим комплексом (рибосомой и иРНК)
12. Выберите три верных ответа из шести и запишите в таблицу цифры, под которыми они указаны.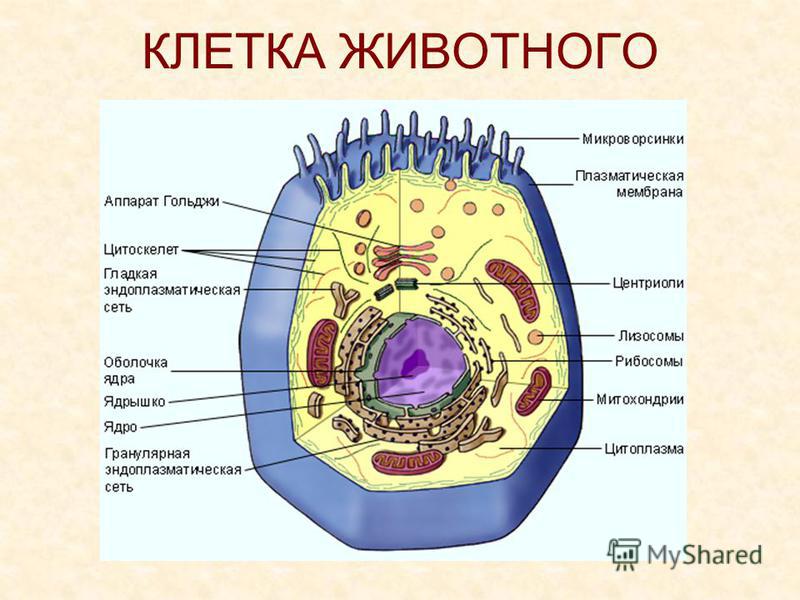 Подкожная жировая клетчатка выполняет функцию
Подкожная жировая клетчатка выполняет функцию
1) хранения генетической информации
2) запасающую
3) терморегуляторную
4) пищеварительную
5) защитную
6) выделительную
13. Установите соответствие между отделом пищеварительного
канала и процессом пищеварения, который в нем происходит.
Запишите в таблицу выбранные цифры под соответствующими буквами.
14. Укажите последовательность кровеносных сосудов, по которым проходит кровь в малом круге кровообращения. Запишите в таблицу соответствующую последовательность цифр.
1) правый желудочек
2) капилляры легких
3) легочная артерия
4) легочная вена
5) левое предсердие
15. Известно, что австралийская ехидна — яйцекладущее млекопитающее, добывающее термитов и муравьев своим длинным языком. Используя эти сведения, выберите из приведенного ниже списка три утверждения, относящиеся к описанию данных признаков этого организма. Запишите в таблицу цифры, соответствующие выбранным ответам.
Запишите в таблицу цифры, соответствующие выбранным ответам.
1) Ехидна весит до 5 кг и имеет размеры до 50 см.
2) Ехидну впервые описали в 1792 году, ошибочно причислив к муравьедам.
3) Первую ехидну-обнаружили в муравейнике, где она своим длинным липким языком, вытягивающимся на 18 см из узкой вытянутой морды, ловила муравьев.
4) Передние лапы ехидны укорочены, пальцы снабжены мощными плоскими когтями, приспособленными для разламывания стенок термитников и рытья земли.
5) Ехидна перемещает яйцо из клоаки в выводковую сумку, где имеются млечные железы без сосков, поэтому детеныши слизывают молоко с шерсти матери.
6) При опасности ехидна сворачивается в шар, пряча живот и выставляя наружу колючки.
16. Установите соответствие между признаком моллюска большого
прудовика и критерием вида, для которого он характерен.
Запишите в таблицу выбранные цифры под соответствующими буквами.
17. Выберите три верных ответа из шести и запишите в таблицу цифры, под которыми они указаны. К устойчивым экосистемам относятся:
К устойчивым экосистемам относятся:
1) вишневый сад
2) картофельное поле
3) таежный лес
4) смешанный лес
5) пшеничное поле
6) березовая роща
18. Установите соответствие между характеристикой организмов и функциональной группой, к которой они относятся.
Запишите в таблицу выбранные цифры под соответствующими буквами.
19. Установите геохронологическую последовательность возникновения групп живых организмов на Земле. Запишите в таблицу соответствующую последовательность цифр.
1) Пресмыкающиеся
2) Зеленые водоросли
3) Цветковые растения
4) Земноводные
5) Круглые черви
6) Рыбы
20. Проанализируйте таблицу «Особенности строения и функции органоидов». Заполните пустые ячейки таблицы, используя термины, приведенные в списке. Для каждой ячейки, обозначенной буквами, выберите соответствующий термин из предложенного списка.
Особенности и функции органоидов
Список терминов
1) содержит хлорофилл 5) образует кислород
2) хлоропласты 6) ЭПС
3) содержит кристы 7) клеточная стенка из целлюлозы
4) участвует в делении клетки
Запишите в таблицу цифры выбранных терминов под соответствующими буквами.
21. Изучите таблицу «Содержание белков, жиров и углеводов в продуктах питания».
Выберите утверждения, которые можно сформулировать на основании полученных результатов.
Содержание белков, жиров и углеводов в продуктах питания
Утверждения:
1) В 100 г сметаны содержится 30,2 г жиров.
2) В 100 г картофеля больше углеводов, чем в 100 г риса.
3) 100 г куриных яиц содержат больше белка, чем 100 г свинины.
4) Количество белков в горохе выше, чем в сосисках.
5) 100 г свежей капусты содержат больше углеводов, чем 100 г моркови.
Запишите в ответе номера выбранных утверждений.
Ответ:_______________.
Цитология 1-25 – правильные ответы
Кликните по вопросу, чтобы узнать ответ
1. Живые системы отличаются от неживых систем наличием —
 org/Answer»>
обмена веществ
org/Answer»>
обмена веществ
жиров
белков
раздражимости
авторегуляции
2. Термин «клетка» ввел —
 org/Answer»>
Р. Гук
org/Answer»>
Р. Гук
3. Клетку как «открытую систему» характеризуют —
обмен веществ с окружающей средой
обмен информацией с окружающей средой
уменьшение энтропии внутри системы за счет обмена энергией с окружающей средой
обмен энергией с окружающей средой
4.
 Клеточную теорию сформулировали —
Клеточную теорию сформулировали —Т. Шванн
Р. Вирхов
М. Шлейден
5. Положения клеточной теории —
всякая клетка происходит только от клетки
 org/Answer»>
клетка — основная структурная и функциональная единица живого
org/Answer»>
клетка — основная структурная и функциональная единица живого
клетки сходны по строению и свойствам
6. Для прокариотической клетки характерно наличие —
нуклеоида
мезосом
7. В прокариотической клетке отсутствуют —
 org/Answer»>
митохондрии
org/Answer»>
митохондрии
лизосомы
8. К прокариотам относят —
бактерии
цианобактерии
синезеленые водоросли
9.
 Прокариоты размножаются —
Прокариоты размножаются —бинарным равновеликим делением
цитотомией
10. Функцию дыхания в прокариотической клетке выполняют —
мезосомы
11. Органеллы, общие для про- и эукариотических клеток —
 org/Answer»>
рибосомы
org/Answer»>
рибосомы
12. Цитоплазма прокариотической клетки —
представляет единый объем
имеет цитозоль
13. В клеточной стенке прокариот присутствует —
муреин
 org/Answer»>
псевдомуреин
org/Answer»>
псевдомуреин
14. Для эукариотической клетки характерно наличие —
митохондрий
лизосом
ЭПС
15. В эукариотической клетке отсутствуют —
 org/Answer»>
мезосомы
org/Answer»>
мезосомы
16. Поверхностный аппарат животной эукариотической клетки включает —
плазматическую мембрану
гликокаликс
микротрубочки
17. В клеточной стенке эукариот могут быть обнаружены —
 org/Answer»>
целлюлоза
org/Answer»>
целлюлоза
хитин
18. Структурной основой всех мембран клетки являются —
белки
фосфолипиды
гликопротеиды
19.
 В клетку вещества не проникают в процессе —
В клетку вещества не проникают в процессе —экзоцитоза
20. Пассивный транспорт — это процессы —
осмоса
диффузии
облегченной диффузии
21.
 Функции интегральных белков плазматической мембраны —
Функции интегральных белков плазматической мембраны —осуществляют работу «натрий-калиевого» насоса
осуществляют активный транспорт
транспортируют воду и гидрофильные молекулы
22. Функции периферических белков плазматической мембраны —
являются ферментами
 org/Answer»>
обеспечивают межклеточные взаимодействия
org/Answer»>
обеспечивают межклеточные взаимодействия
обеспечивают тканевую совместимость
23. Цитоплазма эукариотической клетки —
разделена на компартменты
имеет цитоскелет
содержит микротрубочки
 org/Answer»>
имеет цитозоль
org/Answer»>
имеет цитозоль
24. Цитоскелет — это
микротрубочки
микрофиламенты
промежуточные микрофиламенты
25. Цитоскелет выполняет функцию —
 org/Answer»>
опорно-двигательной системы
org/Answer»>
опорно-двигательной системы
Клеточная мембрана
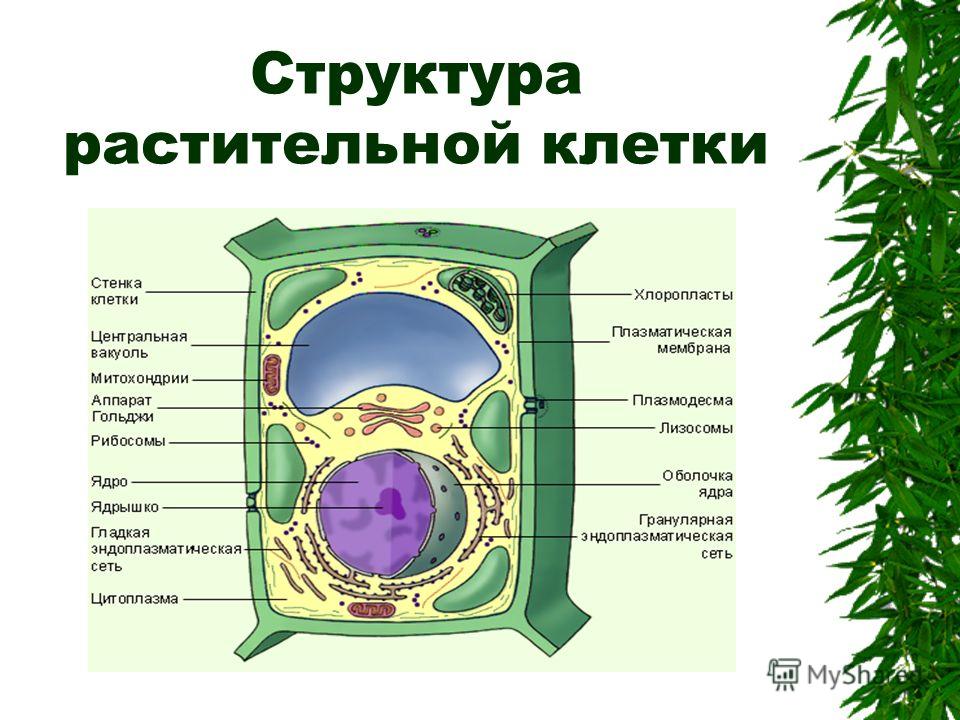
Клеточная мембрана также называется плазматической (или цитоплазматической) мембраной и плазмалеммой. Данная структура не только отделяет внутреннее содержимое клетки от внешней среды, но также входит с состав большинства клеточных органелл и ядра, в свою очередь отделяя их от гиалоплазмы (цитозоля) — вязко-жидкой части цитоплазмы. Договоримся называть цитоплазматической мембраной ту, которая отделяет содержимое клетки от внешней среды. Остальными терминами обозначать все мембраны.
Строение клеточной мембраны
В основе строения клеточной (биологической) мембраны лежит двойной слой липидов (жиров). Формирование такого слоя связано с особенностями их молекул. Липиды не растворяются в воде, а по-своему в ней конденсируются. Одна часть отдельно взятой молекулы липида представляет собой полярную головку (она притягивается водой, т. е. гидрофильна), а другая — пару длинных неполярных хвостов (эта часть молекулы отталкивается от воды, т. е. гидрофобна). Такое строение молекул заставляет их «прятать» хвосты от воды и поворачивать к воде свои полярные головки.
е. гидрофильна), а другая — пару длинных неполярных хвостов (эта часть молекулы отталкивается от воды, т. е. гидрофобна). Такое строение молекул заставляет их «прятать» хвосты от воды и поворачивать к воде свои полярные головки.
В результате образуется двойной липидный слой, в котором неполярные хвосты находятся внутри (обращены друг к другу), а полярные головки обращены наружу (к внешней среде и цитоплазме). Поверхность такой мембраны гидрофильна, а внутри она гидрофобна.
В клеточных мембранах среди липидов преобладают фосфолипиды (относятся к сложным липидам). Их головки содержат остаток фосфорной кислоты. Кроме фосфолипидов есть гликолипиды (липиды + углеводы) и холестерол (относится к стеролам). Последний придает мембране жесткость, размещаясь в ее толще между хвостами остальных липидов (холестерол полностью гидрофобный).
За счет электростатического взаимодействия, к заряженным головкам липидов присоединяются некоторые молекулы белков, которые становятся поверхностными мембранными белками. Другие белки взаимодействуют с неполярными хвостами, частично погружаются в двойной слой или пронизывают его насквозь.
Другие белки взаимодействуют с неполярными хвостами, частично погружаются в двойной слой или пронизывают его насквозь.
Таким образом, клеточная мембрана состоит из двойного слоя липидов, поверхностных (периферических), погруженных (полуинтегральных) и пронизывающих (интегральных) белков. Кроме того, некоторые белки и липиды с внешней стороны мембраны связаны с углеводными цепями.
Это жидкостно-мозаичная модель строения мембраны была выдвинута в 70-х годах XX века. До этого предполагалась бутербродная модель строения, согласно которой липидный бислой находится внутри, а с внутренней и наружной стороны мембрана покрыта сплошными слоями поверхностных белков. Однако накопление экспериментальных данных опровергло эту гипотезу.
Толщина мембран у разных клеток составляет около 8 нм. Мембраны (даже разные стороны одной) отличаются между собой по процентному соотношению различных видов липидов, белков, ферментативной активности и др. Какие-то мембраны более жидкие и более проницаемые, другие более плотные.
Разрывы клеточной мембраны легко сливаются из-за физико-химических особенностей липидного бислоя. В плоскости мембраны липиды и белки (если только они не закреплены цитоскелетом) перемещаются.
Функции клеточной мембраны
Большинство погруженных в клеточную мембрану белков выполняют ферментативную функцию (являются ферментами). Часто (особенно в мембранах органоидов клетки) ферменты располагаются в определенной последовательности так, что продукты реакции, катализируемые одним ферментом, переходят ко второму, затем третьему и т. д. Образуется конвейер, который стабилизируют поверхностные белки, т. к. не дают ферментам плавать вдоль липидного бислоя.
Клеточная мембрана выполняет отграничивающую (барьерную) от окружающей среды и в то же время транспортную функции. Можно сказать, это ее самое главное назначение. Цитоплазматическая мембрана, обладая прочностью и избирательной проницаемостью, поддерживает постоянство внутреннего состава клетки (ее гомеостаз и целостность).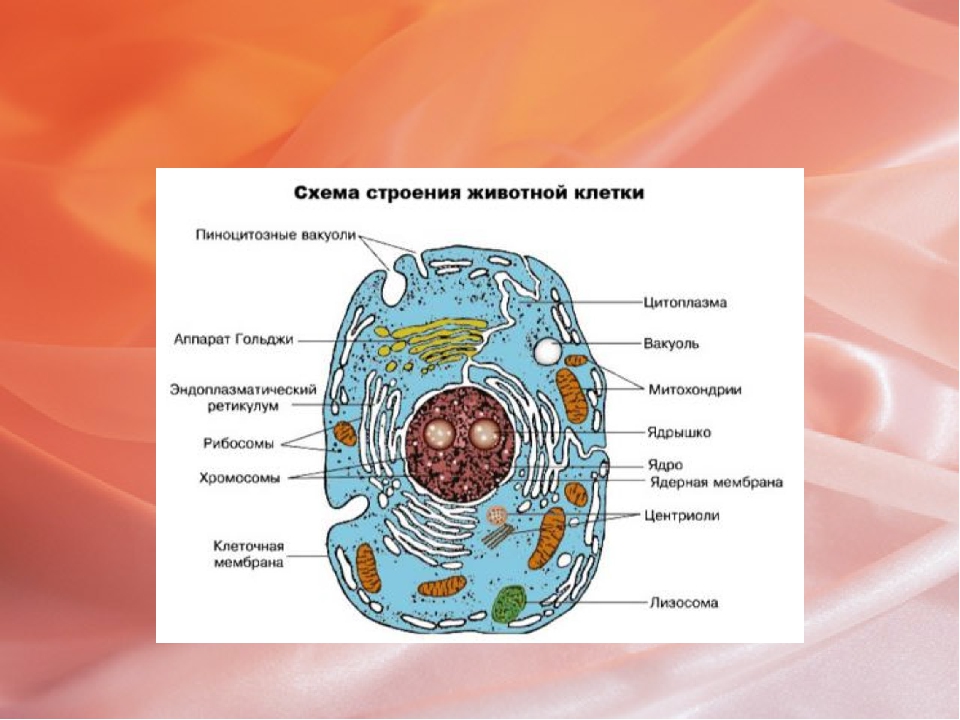
При этом транспорт веществ происходит различными способами. Транспорт по градиенту концентрации предполагает передвижение веществ из области с их большей концентрацией в область с меньшей (диффузия). Так, например, диффундируют газы (CO2, O2).
Бывает также транспорт против градиента концентрации, но с затратой энергии.
Транспорт бывает пассивным и облегченным (когда ему помогает какой-нибудь переносчик). Пассивная диффузия через клеточную мембрану возможна для жирорастворимых веществ.
Есть особые белки, делающие мембраны проницаемыми для сахаров и других водорастворимых веществ. Такие переносчики соединяются с транспортируемыми молекулами и протаскивают их через мембрану. Так переносится глюкоза внутрь эритроцитов.
Пронизывающие белки, объединяясь, могут образовывать пору для перемещения некоторых веществ через мембрану. Такие переносчики не перемещаются, а образуют в мембране канал и работают аналогично ферментам, связывая определенное вещество. Перенос осуществляется благодаря изменению конформации белка, благодаря чему в мембране образуются каналы. Пример — натрий-калиевый насос.
Перенос осуществляется благодаря изменению конформации белка, благодаря чему в мембране образуются каналы. Пример — натрий-калиевый насос.
Транспортная функция клеточной мембраны эукариот также реализуется за счет эндоцитоза (и экзоцитоза). Благодаря этим механизмам в клетку (и из нее) попадают крупные молекулы биополимеров, даже целые клетки. Эндо- и экзоцитоз характерны не для всех клеток эукариот (у прокариот его вообще нет). Так эндоцитоз наблюдается у простейших и низших беспозвоночны; у млекопитающих лейкоциты и макрофаги поглощают вредные вещества и бактерии, т. е. эндоцитоз выполняет защитную функцию для организма.
Эндоцитоз делится на фагоцитоз (цитоплазма обволакивает крупные частицы) и пиноцитоз (захват капелек жидкости с растворенными в ней веществами). Механизм этих процессов приблизительно одинаков. Поглощаемые вещества на поверхности клеток окружаются мембраной. Образуется пузырек (фагоцитарный или пиноцитарный), который затем перемещается внутрь клетки.
Экзоцитоз — это выведение цитоплазматической мембраной веществ из клетки (гормонов, полисахаридов, белков, жиров и др.). Данные вещества заключаются в мембранные пузырьки, которые подходят к клеточной мембране. Обе мембраны сливаются и содержимое оказывается за пределами клетки.
Цитоплазматическая мембрана выполняет рецепторную функцию. Для этого на ее внешней стороне располагаются структуры, способные распознавать химический или физический раздражитель. Часть пронизывающих плазмалемму белков с наружней стороны соединены с полисахаридными цепочками (образуя гликопротеиды). Это своеобразные молекулярные рецепторы, улавливающие гормоны. Когда конкретный гормон связывается со своим рецептором, то изменяет его структуру. Это в свою очередь запускает механизм клеточного ответа. При этом могут открываться каналы, и в клетку могут начать поступать определенные вещества или выводиться из нее.
Рецепторная функция клеточных мембран хорошо изучена на основе действия гормона инсулина. При связывании инсулина с его рецептором-гликопротеидом происходит активация каталитической внутриклеточной части этого белка (фермента аденилатциклазы). Фермент синтезирует из АТФ циклическую АМФ. Уже она активирует или подавляет различные ферменты клеточного метаболизма.
При связывании инсулина с его рецептором-гликопротеидом происходит активация каталитической внутриклеточной части этого белка (фермента аденилатциклазы). Фермент синтезирует из АТФ циклическую АМФ. Уже она активирует или подавляет различные ферменты клеточного метаболизма.
Рецепторная функция цитоплазматической мембраны также включает распознавание соседних однотипных клеток. Такие клетки прикрепляются друг к другу различными межклеточными контактами.
В тканях с помощью межклеточных контактов клетки могут обмениваться между собой информацией с помощью специально синтезируемых низкомолекулярных веществ. Одним из примеров подобного взаимодействия является контактное торможение, когда клетки прекращают рост, получив информацию, что свободное пространство занято.
Межклеточные контакты бывают простыми (мембраны разных клеток прилегают друг к другу), замковыми (впячивания мембраны одной клетки в другую), десмосомы (когда мембраны соединены пучками поперечных волокон, проникающих в цитоплазму). Кроме того, есть вариант межклеточных контактов за счет медиаторов (посредников) — синапсы. В них сигнал передается не только химическим, но и электрическим способом. Синапсами передаются сигналы между нервными клетками, а также от нервных к мышечным.
Кроме того, есть вариант межклеточных контактов за счет медиаторов (посредников) — синапсы. В них сигнал передается не только химическим, но и электрическим способом. Синапсами передаются сигналы между нервными клетками, а также от нервных к мышечным.
Источник:
http://biology.su/cytology/cell-membrane
Что такое клеточная мембрана
Клеточная стенка, если таковая у клетки имеется (обычно есть у растительных клеток), покрывает клеточную мембрану.
Клеточная мембрана представляет собой двойной слой (бислой) молекул класса липидов, большинство из которых представляет собой так называемые сложные липиды — фосфолипиды. Молекулы липидов имеют гидрофильную («головка») и гидрофобную («хвост») часть. При образовании мембран гидрофобные участки молекул оказываются обращены внутрь, а гидрофильные — наружу. Мембраны — структуры инвариабельные, весьма сходные у разных организмов. Некоторое исключение составляют, пожалуй, археи, у которых мембраны образованы глицерином и терпеноидными спиртами. Толщина мембраны составляет 7—8 нм.
Толщина мембраны составляет 7—8 нм.
Биологическая мембрана включает и различные белки: интегральные (пронизывающие мембрану насквозь), полуинтегральные (погруженные одним концом во внешний или внутренний липидный слой), поверхностные (расположенные на внешней или прилегающие к внутренней сторонам мембраны). Некоторые белки являются точками контакта клеточной мембраны с цитоскелетом внутри клетки, и клеточной стенкой (если она есть) снаружи. Некоторые из интегральных белков выполняют функцию ионных каналов, различных транспортеров и рецепторов.
Функции
- барьерная — обеспечивает регулируемый, избирательный, пассивный и активный обмен веществ с окружающей средой. Например, мембрана пероксисом защищает цитоплазму от опасных для клетки пероксидов. Избирательная проницаемость означает, что проницаемость мембраны для различных атомов или молекул зависит от их размеров, электрического заряда и химических свойств. Избирательная проницаемость обеспечивает отделение клетки и клеточных компартментов от окружающей среды и снабжение их необходимыми веществами.

- транспортная — через мембрану происходит транспорт веществ в клетку и из клетки. Транспорт через мембраны обеспечивает: доставку питательных веществ, удаление конечных продуктов обмена, секрецию различных веществ, создание ионных градиентов, поддержание в клетке оптимального pH и концентрации ионов, которые нужны для работы клеточных ферментов.
Частицы, по какой-либо причине неспособные пересечь фосфолипидный бислой (например, из-за гидрофильных свойств, так как мембрана внутри гидрофобна и не пропускает гидрофильные вещества, или из-за крупных размеров), но необходимые для клетки, могут проникнуть сквозь мембрану через специальные белки-переносчики (транспортеры) и белки-каналы или путем эндоцитоза.
При пассивном транспорте вещества пересекают липидный бислой без затрат энергии по градиенту концентрации путем диффузии. Вариантом этого механизма является облегчённая диффузия, при которой веществу помогает пройти через мембрану какая-либо специфическая молекула. У этой молекулы может быть канал, пропускающий вещества только одного типа.
Активный транспорт требует затрат энергии, так как происходит против градиента концентрации. На мембране существуют специальные белки-насосы, в том числе АТФаза, которая активно вкачивает в клетку ионы калия (K+) и выкачивают из неё ионы натрия (Na+). - матричная — обеспечивает определенное взаиморасположение и ориентацию мембранных белков, их оптимальное взаимодействие.
- механическая — обеспечивает автономность клетки, ее внутриклеточных структур, также соединение с другими клетками (в тканях). Большую роль в обеспечение механической функции имеют клеточные стенки, а у животных — межклеточное вещество.
- энергетическая — при фотосинтезе в хлоропластах и клеточном дыхании в митохондриях в их мембранах действуют системы переноса энергии, в которых также участвуют белки;
- рецепторная — некоторые белки, находящиеся в мембране, являются рецепторами (молекулами, при помощи которых клетка воспринимает те или иные сигналы).
Например, гормоны, циркулирующие в крови, действуют только на такие клетки-мишени, у которых есть соответствующие этим гормонам рецепторы. Нейромедиаторы (химические вещества, обеспечивающие проведение нервных импульсов) тоже связываются с особыми рецепторными белками клеток-мишеней. - ферментативная — мембранные белки нередко являются ферментами. Например, плазматические мембраны эпителиальных клеток кишечника содержат пищеварительные ферменты.
- осуществление генерации и проведения биопотенциалов.
С помощью мембраны в клетке поддерживается постоянная концентрация ионов: концентрация иона К+ внутри клетки значительно выше, чем снаружи, а концентрация Na+ значительно ниже, что очень важно, так как это обеспечивает поддержание разности потенциалов на мембране и генерацию нервного импульса. - маркировка клетки — на мембране есть антигены, действующие как маркеры — «ярлыки», позволяющие опознать клетку. Это гликопротеины (то есть белки с присоединенными к ним разветвленными олигосахаридными боковыми цепями), играющие роль «антенн». Из-за бесчисленного множества конфигурации боковых цепей возможно сделать для каждого типа клеток свой особый маркер. С помощью маркеров клетки могут распознавать другие клетки и действовать согласованно с ними, например, при формировании органов и тканей. Это же позволяет иммунной системе распознавать чужеродные антигены.

 Нейромедиаторы (химические вещества, обеспечивающие проведение нервных импульсов) тоже связываются с особыми рецепторными белками клеток-мишеней.
Нейромедиаторы (химические вещества, обеспечивающие проведение нервных импульсов) тоже связываются с особыми рецепторными белками клеток-мишеней. С помощью маркеров клетки могут распознавать другие клетки и действовать согласованно с ними, например, при формировании органов и тканей. Это же позволяет иммунной системе распознавать чужеродные антигены.
С помощью маркеров клетки могут распознавать другие клетки и действовать согласованно с ними, например, при формировании органов и тканей. Это же позволяет иммунной системе распознавать чужеродные антигены.Структура и состав биомембран
Мембраны состоят из липидов трёх классов: фосфолипиды, гликолипиды и холестерол. Фосфолипиды и гликолипиды (липиды с присоединёнными к ним углеводами) состоят из двух длинных гидрофобных углеводородных «хвостов», которые связаны с заряженной гидрофильной «головой». Холестерол придаёт мембране жёсткость, занимая свободное пространство между гидрофобными хвостами липидов и не позволяя им изгибаться. Поэтому мембраны с малым содержанием холестерола более гибкие, а с большим — более жёсткие и хрупкие. Также холестерол служит «стопором», препятствующим перемещению полярных молекул из клетки и в клетку. Важную часть мембраны составляют белки, пронизывающие её и отвечающие за разнообразные свойства мембран. Их состав и ориентация в разных мембранах различаются.
Клеточные мембраны часто асимметричны, то есть слои отличаются по составу липидов, переход отдельной молекулы из одного слоя в другой (так называемый флип-флоп) затруднён.
Мембранные органеллы
Это замкнутые одиночные или связанные друг с другом участки цитоплазмы, отделённые от гиалоплазмы мембранами. К одномембранным органеллам относятся эндоплазматическая сеть, аппарат Гольджи, лизосомы, вакуоли, пероксисомы; к двумембранным — ядро, митохондрии, пластиды. Строение мембран различных органелл отличается по составу липидов и мембранных белков.
Избирательная проницаемость
Клеточные мембраны обладают избирательной проницаемостью: через них медленно диффундируют глюкоза, аминокислоты, жирные кислоты, глицерол и ионы, причем сами мембраны в известной мере активно регулируют этот процесс — одни вещества пропускают, а другие нет. Существует четыре основных механизма для поступления веществ в клетку или вывода их из клетки наружу: диффузия, осмос, активный транспорт и экзо- или эндоцитоз.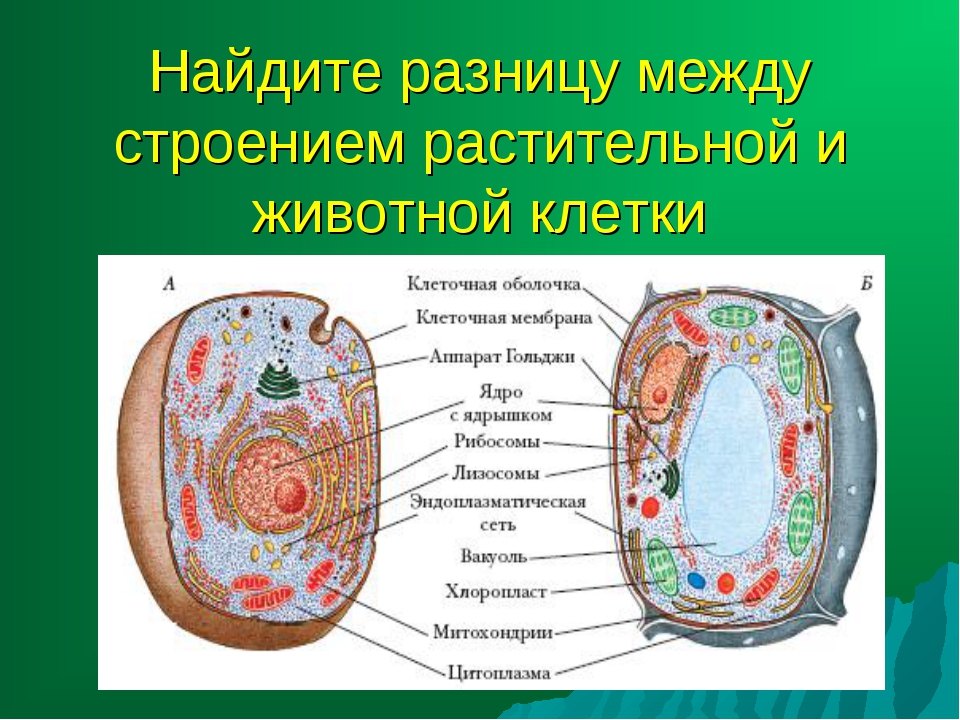 Два первых процесса носят пассивный характер, то есть не требуют затрат энергии; два последних — активные процессы, связанные с потреблением энергии.
Два первых процесса носят пассивный характер, то есть не требуют затрат энергии; два последних — активные процессы, связанные с потреблением энергии.
Избирательная проницаемость мембраны при пассивном транспорте обусловлена специальными каналами — интегральными белками. Они пронизывают мембрану насквозь, образовывая своего рода проход. Для элементов K, Na и Cl есть свои каналы. Относительно градиента концентрации молекулы этих элементов движутся в клетку и из неё. При раздражении каналы натриевых ионов раскрываются, и происходит резкое поступление в клетку ионов натрия. При этом происходит дисбаланс мембранного потенциала. После чего мембранный потенциал восстанавливается. Каналы калия всегда открыты, через них в клетку медленно попадают ионы калия.
Источник:
http://dic.academic.ru/dic.nsf/ruwiki/970083
Клеточная мембрана. Функции клеточной мембраны. Строение клеточной мембраны.
Содержание статьи
Клеточная мембрана
Клеточная мембрана или цитолемма или плазмалемма или плазматическая мембрана – эластическая молекулярная структура. Её толщина составляет от 6 до 10 нм. Рассматривая строение клеточной мембраны, можно сказать, что она состоит из белков (около 40%) и липидов (около 60%).
Её толщина составляет от 6 до 10 нм. Рассматривая строение клеточной мембраны, можно сказать, что она состоит из белков (около 40%) и липидов (около 60%).
Функции клеточной мембраны
По функциональным особенностям клеточную мембрану можно разделить на 9 выполняемых ей функций.
Функции клеточной мембраны:
1. Транспортная. Производит транспорт веществ из клетки в клетку;
2. Барьерная. Обладает избирательной проницаемостью, обеспечивает необходимый обмен веществ;
3. Рецепторная. Некоторые белки находящиеся в мембране являются рецепторами;
4. Механическая. Обеспечивает автономность клетки и её механических структур;
5. Матричная. Обеспечивает оптимальное взаимодействие и ориентацию матричных белков;
6. Энергетическая. В мембранах действуют системы переноса энергии при клеточном дыхании в митохондриях;
7. Ферментативная. Мембранные белки иногда являются ферментами. Например мембраны клеток кишечника;
8. Маркировочная. На мембране есть антигены (гликопротеины), которые позволяют опознать клетку;
9. Генерирующая. Осуществляет генерацию и проведение биопотенциалов.
Генерирующая. Осуществляет генерацию и проведение биопотенциалов.
Посмотреть как выглядит клеточная мембрана можно на примере строения животной клетки или растительной клетки.
Cтроение клеточной мембраны
На рисунке приведено строение клеточной мембраны.
К компонентам клеточной мембраны можно отнести различные белки клеточной мембраны (глобулярный, переферический, поверхностный), а также липиды клеточной мембраны (гликолипид, фосфолипид). Таже в строении клеточной мембраны присутствуют углеводы, холестерол, гликопротеин и белковая альфа спираль.
Состав клеточной мембраны
К основному составу клеточной мембраны относятся:
1. Белки – отвечающие за разнообразные свойства мембраны;
2. Липиды трёх видов (фосфолипиды, гликолипиды и холестерол) отвечающих за жёсткость мембраны.
Белки клеточной мембраны:
1. Глобулярный белок;
2. Поверхностный белок;
3. Переферический белок.
Основное назначение клеточной мембраны
Основное назначение клеточной мембраны:
1. Регулировать обмен между клеткой и средой;
Регулировать обмен между клеткой и средой;
2. Отделять содержимое любой клетки от внешней среды тем самым обеспечивая её целостность;
3. Внутриклеточные мембраны разделяют клетку на специализированные замкнутые отсеки – органеллы или компартменты, в которых поддерживаются определённые условия среды.
Структура клеточной мембраны
Структура клеточной мембраны представляют собой двумерный раствор глобулярных интегральных белков, растворенных в жидком фосфолипидном матриксе. Данная модель мембранной структуры была предложена двумя учёными Никольсоном и Сингером в 1972 году. Таким образом, основу мембран составляет бимолекулярный липидный слой, с упорядоченным расположением молекул, что вы могли видеть на этом рисунке.
Источник:
http://www.new-era.me/articles/kletochnaya-membrana-funkcii-stroenie.php
Мембрана строение функция
Мембрана строение функция
Основная структурная единица живого организма — клетка, являющаяся дифференцированным участком цитоплазмы, окружённым клеточной мембраной. Ввиду того что клетка выполняет множество важнейших функций, таких, как размножение, питание, движение, оболочка должна быть пластичной и плотной.
Ввиду того что клетка выполняет множество важнейших функций, таких, как размножение, питание, движение, оболочка должна быть пластичной и плотной.
История открытия и исследования клеточной мембраны
В 1925 году Гренделем и Гордером был поставлен успешный эксперимент по выявлению «теней|теней» эритроцитов, или пустых оболочек. Несмотря на несколько допущенных грубых ошибок, учёными было произведено открытие липидного бислоя. Их труды продолжили Даниэлли, Доусон в 1935 году, Робертсон в 1960 году. В результате многолетней работы и накопления аргументов в 1972 году Сингер и Николсон создали жидкостно-мозаичную модель строения мембраны. Дальнейшие опыты и исследования подтвердили труды учёных.
Что же представляет собой клеточная мембрана? Это слово стало использоваться более ста лет назад, в переводе с латинского оно означает «плёнка», «кожица». Так обозначают границу клетки, являющуюся естественным барьером между внутренним содержимым и внешней средой. Строение клеточной мембраны предполагает полупроницаемость, благодаря которой влага и питательные вещества и продукты распада свободно могут проходить сквозь неё. Эту оболочку можно назвать основной структурной составляющей организации клетки.
Эту оболочку можно назвать основной структурной составляющей организации клетки.
Рассмотрим основные функции клеточной мембраны
1. Разделяет внутреннее содержимое клетки и компоненты внешней среды|среды.
2. Способствует поддержанию постоянного химического состава клетки.
3. Регулирует правильный обмен веществ.
4. Обеспечивает взаимосвязь между клетками.
5. Распознает|Распознаёт сигналы.
6. Функция защиты.
Наружная клеточная мембрана, называемая также плазменной, представляет собой ультрамикроскопическую плёнку, толщина которой составляет от пяти до семи наномиллиметров. Она состоит преимущественно из белковых соединений, фосфолидов, воды|воды. Плёнка является эластичной, легко впитывает воду, а также стремительно восстанавливает свою целостность после повреждений.
Отличается универсальным строением. Эта мембрана занимает пограничное положение, участвует в процессе избирательной проницаемости, выведении продуктов распада, синтезирует их. Взаимосвязь с «соседями» и надёжная защита внутреннего содержимого от повреждения делает её важной составляющей в таком вопросе, как строение клетки. Клеточная мембрана животных организмов иногда оказывается покрытой тончайшим слоем – гликокаликсом, в состав которого входят белки|белки и полисахариды. Растительные клетки снаружи от мембраны защищены клеточной стенкой, выполняющей функции опоры и поддержания формы. Основной компонент её состава – это клетчатка (целлюлоза) – полисахарид, не растворимый в воде.
Взаимосвязь с «соседями» и надёжная защита внутреннего содержимого от повреждения делает её важной составляющей в таком вопросе, как строение клетки. Клеточная мембрана животных организмов иногда оказывается покрытой тончайшим слоем – гликокаликсом, в состав которого входят белки|белки и полисахариды. Растительные клетки снаружи от мембраны защищены клеточной стенкой, выполняющей функции опоры и поддержания формы. Основной компонент её состава – это клетчатка (целлюлоза) – полисахарид, не растворимый в воде.
Таким образом, наружная клеточная мембрана выполняет функцию восстановления, защиты и взаимодействия с другими клетками.
Строение клеточной мембраны
Толщина этой подвижной|подвижной оболочки варьируется в пределах от шести до десяти наномиллиметров. Клеточная мембрана клетки имеет особый состав, основой которого служит липидный бислой. Гидрофобные хвосты, инертные к воде, размещены с внутренней стороны|стороны, в то время как гидрофильные головки, взаимодействующие с водой, обращены наружу.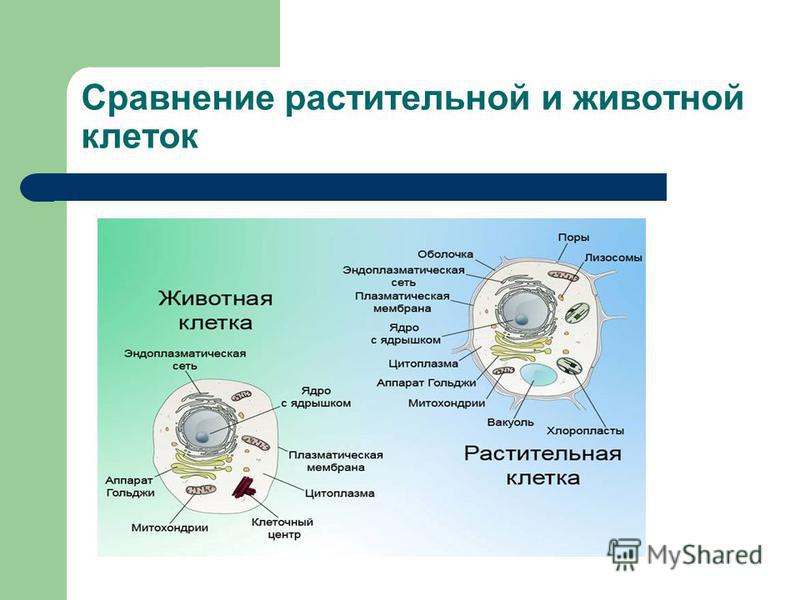 Каждый липид представляет фосфолипид, который является результатом взаимодействия таких веществ, как глицерин и сфингозин. Липидный каркас тесно окружают белки|белки, которые расположены несплошным слоем. Некоторые из них погружены в липидный слой, остальные проходят сквозь него. В результате этого образуются проницаемые для воды|воды участки. Выполняемые этими белками|белками функции различны. Некоторые из них являются ферментами, остальные — транспортными белками|белками, которые переносят различные вещества из внешней среды|среды на цитоплазму и обратно.
Каждый липид представляет фосфолипид, который является результатом взаимодействия таких веществ, как глицерин и сфингозин. Липидный каркас тесно окружают белки|белки, которые расположены несплошным слоем. Некоторые из них погружены в липидный слой, остальные проходят сквозь него. В результате этого образуются проницаемые для воды|воды участки. Выполняемые этими белками|белками функции различны. Некоторые из них являются ферментами, остальные — транспортными белками|белками, которые переносят различные вещества из внешней среды|среды на цитоплазму и обратно.
Клеточная мембрана насквозь пронизана и тесно связана интегральными белками|белками, а с переферическими связь менее прочная. Эти белки|белки выполняют важную функцию, которая заключается в поддержании структуры мембраны, получении и преобразовании сигналов из окружающей среды|среды, транспорте веществ, катализации реакций, которые происходят на мембранах.
Состав
Основу клеточной мембраны представляет бимолекулярный слой.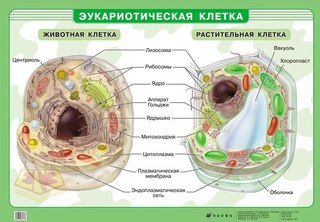 Благодаря его непрерывности клетка имеет барьерное и механическое свойства. На разных этапах жизнедеятельности данный бислой может нарушиться. Вследствие этого образуются структурные дефекты сквозных гидрофильных пор. В таком случае могут изменяться абсолютно всё|все функции такой составляющей, как клеточная мембрана. Ядро при этом может пострадать от внешних воздействий.
Благодаря его непрерывности клетка имеет барьерное и механическое свойства. На разных этапах жизнедеятельности данный бислой может нарушиться. Вследствие этого образуются структурные дефекты сквозных гидрофильных пор. В таком случае могут изменяться абсолютно всё|все функции такой составляющей, как клеточная мембрана. Ядро при этом может пострадать от внешних воздействий.
Свойства
Клеточная мембрана клетки имеет интересные особенности. Благодаря текучести эта оболочка не является жёсткой структурой, а основная часть белков и липидов, которые входят в её состав, свободно перемещается на плоскости мембраны.
В целом клеточная мембрана асимметрична, поэтому состав белковых и липидных слоёв различается. Плазматические мамбраны в животных клетках со своей наружной стороны|стороны имеют гликопротеиновый слой, который выполняет рецепторные и сигнальные функции, а также играет большую|большую роль в процессе объединения клеток в ткань. Клеточная мембрана является полярной, то есть на внешней стороне заряд положителен, а с внутренней стороны|стороны – отрицателен. Помимо всего перечисленного, оболочка клетки обладает избирательной проницательностью. Это означает, что кроме воды|воды в клетку пропускается только определённая группа молекул и ионов растворившихся веществ. Концентрация такого вещества, как натрий, в большинстве клеток значительно ниже, чем во внешней среде. Для ионов калия характерно другое соотношение: их количество в клетке намного выше, чем в окружающей среде. В связи с этим ионам натрия присуще стремление проникнуть в клеточную оболочку, а ионы калия стремятся освободиться наружу. При данных обстоятельствах мембрана активизирует особую систему, выполняющую «насосную» роль, выравнивая концентрацию веществ: ионы натрия откачиваются на поверхность клетки, а ионы калия накачиваются внутрь. Данная особенность входит в важнейшие функции клеточной мембраны.
Помимо всего перечисленного, оболочка клетки обладает избирательной проницательностью. Это означает, что кроме воды|воды в клетку пропускается только определённая группа молекул и ионов растворившихся веществ. Концентрация такого вещества, как натрий, в большинстве клеток значительно ниже, чем во внешней среде. Для ионов калия характерно другое соотношение: их количество в клетке намного выше, чем в окружающей среде. В связи с этим ионам натрия присуще стремление проникнуть в клеточную оболочку, а ионы калия стремятся освободиться наружу. При данных обстоятельствах мембрана активизирует особую систему, выполняющую «насосную» роль, выравнивая концентрацию веществ: ионы натрия откачиваются на поверхность клетки, а ионы калия накачиваются внутрь. Данная особенность входит в важнейшие функции клеточной мембраны.
Подобное стремление ионов натрия и калия переместиться внутрь с поверхности играет большую|большую роль в вопросе транспортировки сахара|сахара|сахара и аминокислот в клетку. В процессе активного удаления ионов натрия из клетки мембрана создаёт условия для новых поступлений глюкозы и аминокислот внутрь. Напротив, в процессе переноса ионов калия внутрь клетки пополняется число «транспортировщиков» продуктов распада изнутри клетки во внешнюю среду|среду.
В процессе активного удаления ионов натрия из клетки мембрана создаёт условия для новых поступлений глюкозы и аминокислот внутрь. Напротив, в процессе переноса ионов калия внутрь клетки пополняется число «транспортировщиков» продуктов распада изнутри клетки во внешнюю среду|среду.
Как происходит питание клетки через клеточную мембрану?
Многие клетки поглощают вещества посредством таких процессов, как фагоцитоз и пиноцитоз. При первом варианте гибкой наружной мембраной создаётся маленькое углубление, в котором оказывается захватываемая частица. Затем диаметр углубления становится больше, пока окружённая частица не попадёт в клеточную цитоплазму. Посредством фагоцитоза подпитываются некоторые простейшие, например амёбы, а также кровяные тельца|тельца — лейкоциты и фагоциты. Аналогичным образом клетки поглощают жидкость, которая содержит необходимые полезные вещества. Такое являние носит название пиноцитоз.
Наружная мембрана тесно соединена с эндоплазматической сетью клетки.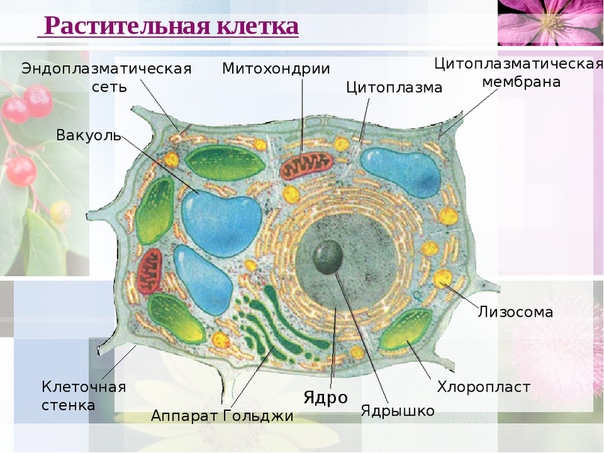
У многих типов основных составляющих ткани на поверхности мембраны расположены выступы, складки, микроворсинки. Растительные клетки снаружи этой оболочки покрыты ещё одной, толстой и отчётливо различимой в микроскоп. Клетчатка, из которой они состоят, помогает формировать опору тканям растительного происхождения, например, древесину. Клетки животных также обладают рядом внешних структур, которые находятся поверх клеточной мембраны. Они носят исключительно защитный характер, пример тому – хитин, содержащийся в покровных клетках насекомых.
Помимо клеточной, существует внутриклеточная мембрана. Её функция заключается в разделении клетки на несколько специализированных замкнутых отсеков – компартментов или органелл, где должна поддерживаться определённая среда.
Таким образом, невозможно переоценить роль такой составляющей основной единицы живого организма, как клеточная мембрана. Строение и функции предполагают значительное расширение общей площади поверхности клетки, улучшение обменных процессов.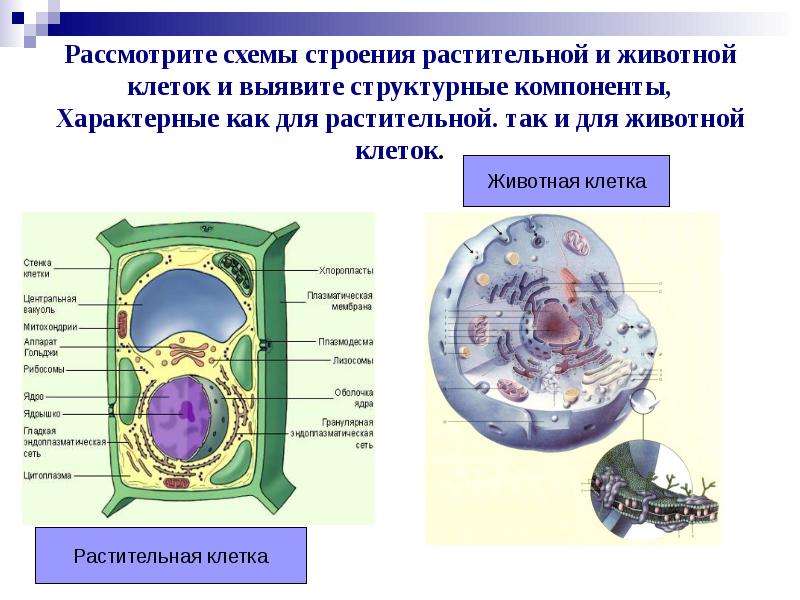 В состав этой молекулярной структуры входят белки|белки и липиды. Отделяя клетку от внешней среды|среды, мембрана обеспечивает её целостность. С её помощью межклеточные связи поддерживаются на достаточно крепком уровне, образовывая ткани. В связи с этим можно сделать вывод, что одну из важнейших ролей|ролей в клетке играет клеточная мембрана. Строение и функции, выполняемые ею, радикально отличаются в различных клетках, в зависимости от их предназначения. Посредством этих особенностей достигается разнообразие физиологической активности клеточных оболочек и их ролей|ролей в существовании клеток и тканей.
В состав этой молекулярной структуры входят белки|белки и липиды. Отделяя клетку от внешней среды|среды, мембрана обеспечивает её целостность. С её помощью межклеточные связи поддерживаются на достаточно крепком уровне, образовывая ткани. В связи с этим можно сделать вывод, что одну из важнейших ролей|ролей в клетке играет клеточная мембрана. Строение и функции, выполняемые ею, радикально отличаются в различных клетках, в зависимости от их предназначения. Посредством этих особенностей достигается разнообразие физиологической активности клеточных оболочек и их ролей|ролей в существовании клеток и тканей.
Мембрана строение функция
Природа создала множество организмов и клеток, но, несмотря на это, строение и большая|большая часть функций биологических мембран одинаковы, что позволяет рассматривать их структуру и изучать их ключевые свойства без привязанности к конкретному виду клеток.
Что такое мембрана?
Мембраны – это защитный элемент, который является неотъемлемой составляющей клетки любого живого организма.
Структурной и функциональной единицей всех живых организмов на планете является клетка. Жизнедеятельность её неразрывно связана с окружающей средой, с которой она обменивается энергией, информацией, веществом. Так, питательная энергия, необходимая для функционирования клетки, поступает извне и тратится на осуществление ею различных функций.
Структура простейшей единицы строения живого организма: мембрана клетки, ядро, органеллы, разнообразные включения. Она окружена мембраной, внутри которой располагается ядро и всё|все органеллы. Это митохондрии, лизосомы, рибосомы, аппарат Гольджи, эндоплазматический ретикулум. Каждый структурный элемент имеет свою мембрану.
Роль в жизнедеятельности клетки
Биологическая мембрана играет кульминационную роль в строении и функционировании элементарной живой системы. Только клетка, окружённая защитной оболочкой, по праву может называться организмом. Такой процесс, как обмен веществ, также осуществляется благодаря наличию мембраны.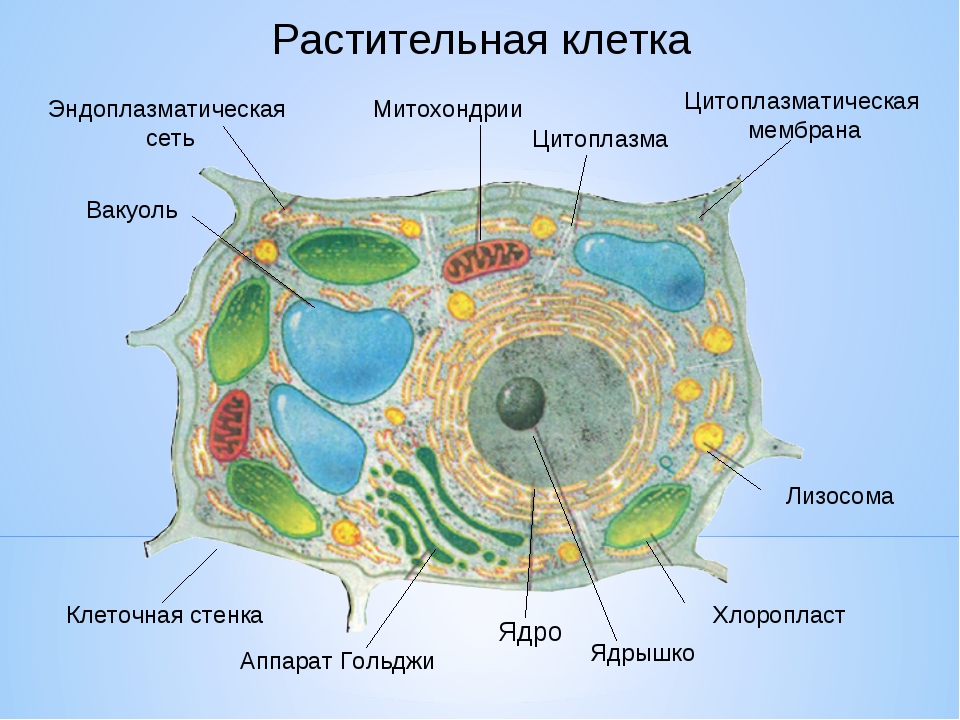 Если структурная целостность её нарушена, это приводит к изменению функционального состояния организма в целом.
Если структурная целостность её нарушена, это приводит к изменению функционального состояния организма в целом.
Клеточная мембрана и её функции
Она отделяет цитоплазму клетки от внешней среды|среды или от оболочки. Мембрана клетки обеспечивает должное выполнение специфических функций, специфику межклеточных контактов и иммунных проявлений, поддерживает трансмембранную разницу электрического потенциала. В ней имеются рецепторы, способные воспринимать химические сигналы – гормоны, медиаторы и другие биологические активные компоненты. Эти рецепторы наделяют её ещё одной способностью – изменять метаболическую активность клетки.
1. Активный перенос веществ.
2. Пассивный перенос веществ:
2.1. Диффузия простая.
2.2. Перенос через поры|поры.
2.3. Транспорт, осуществляемый за счёт диффузии переносчика вместе с мембранным веществом или посредством передачи по эстафете вещества по молекулярной цепи переносчика.
3. Перенос неэлектролитов благодаря простой и облегчённой диффузии.
4. Активный транспорт ионов.
Строение мембраны клетки
Составляющие мембраны клетки – липиды и белки|белки.
Липиды: фосфолипиды, фосфатидилэтаноламин, сфингомиелин, фосфатидилинозит и фосфатидилсерин, гликолипиды. Доля липидов составляет 40-90 %.
Белки|Белки: периферические, интегральные (гликопротеины), спектрин, актин, цитоскелет.
Основной структурный элемент – двойной слой фосфолипидных молекул.
Кровельная мембрана: определение и типология
Немного статистики. На территории Российской Федерации мембрана в качестве кровельного материала используется не так уж и давно. Удельный вес мембранных кровель из общего числа|числа мягких перекрытий крыш составляет всего 1,5 %. Более широкое распространение в России получили битумные и мастичные кровли. А вот в Западной Европе на долю мембранных кровель приходится 87 %. Разница ощутимая.
Как правило, мембрана в роли основного материала при перекрытии крыши идеально подходит для плоских кровель. Для имеющих большой уклон она подходит в меньшей степени.
Для имеющих большой уклон она подходит в меньшей степени.
Объёмы производства и реализации мембранных кровель на отечественном рынке имеют положительную тенденцию роста|роста. Почему? Причины более чем ясны:
- Срок эксплуатации составляет около 60 лет. Представьте себе, только гарантийный срок использования, который устанавливается производителем, достигает 20 лет.
- Лёгкость в монтаже. Для сравнения: установка битумной кровли занимает в 1,5 раза больше времени, нежели монтаж мембранного перекрытия.
- Простота в обслуживании и проведении ремонтных работ.
Толщина кровельных мембран может составлять 0,8-2 мм, а средний показатель веса|веса одного метра квадратного равен 1,3 кг.
Свойства кровельных мембран:
Мембрана кровельная бывает трёх типов. Главный классификационный признак – вид полимерного материала, составляющего основание полотна|полотна. Итак, кровельные мембраны бывают:
Мембрана профилированная: характеристика, функции и преимущества
Профилированные мембраны – это инновация на строительном рынке. Такая мембрана эксплуатируется в качестве гидроизоляционного материала.
Такая мембрана эксплуатируется в качестве гидроизоляционного материала.
Вещество, используемое при изготовлении, – полиэтилен. Последний бывает двух типов: полиэтилен высокого давления (ПВД) и полиэтилен низкого давления (ПНД).
Техническая характеристика мембраны из ПВД и ПНД
Источник:
http://biologyinfo.ru/page/membrana-stroenie-funkcija/
СПАДИЛО.РУ
теория по биологии цитология
Теория для подготовки к блоку «Цитология»Клеточная мембрана
Мембрана клетки = цитоплазматическая мембрана = плазматическая мембрана = плазмалемма
Образована двумя слоями фосфолипидов, которые имеют гидрофильные головки и гидрофобные хвосты. Головки расположены в сторону жидких сред: цитоплазма и внеклеточное вещество/ вещество окружающей среды, а хвосты – друг к другу. Так клеточная мембрана является достаточно плотной структурой, но в то же время пластичной и подвижной. Билипидный слой не дает содержимому клетки растекаться, а также препятствует проникновению внутрь клетки веществ, способных нанести ей вред.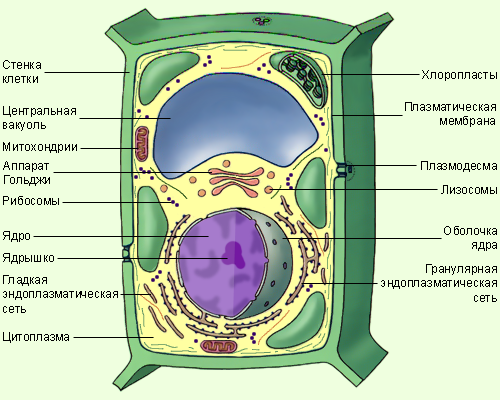
Строение клеточной мембраны
Мембрана клеток частично проницаема. Это значит, что любое вещество не может в нее проникнуть, однако и закрытой ее назвать нельзя. Так как существуют константы гомеостаза (гомеостаз – постоянство внутренней среды), определяющие содержание веществ внутри клетки, то клетка выводит ненужные ей вещества и пропускает нужные. Для этого в клетках есть разные приспособления:
Белки-рецепторы для того, чтобы узнавать молекулы веществ, приближающихся к клетке.
Белки, образующие «тоннели» в клеточной мембране для пассивного тока воды и некоторых неорганических ионов.
Клеточная мембрана помимо выборочной проницаемости обладает свойством текучести. Для захвата пищевых частиц мембрана клетки впячивается, края этого участка мембраны как бы обнимают пищу. Потом края замыкаются и пища в пищевом пузырьке, который называется фагосомой, направляется в пищеварительную вакуоль, где специальные белки-ферменты расщепят пищу. Процесс захвата клеткой твердой пищи называется фагоцитозом. Если же клетка поглощает капельку, то процесс называется пиноцитозом, а пузырек, в котором капля транспортируется в вакуоль – везикулой. Когда клетка заканчивает свои пищеварительные процессы, то ей, как и многоклеточному сложному организму, нужно вывести наружу непереваренные остатки. Тогда происходит экзоцитоз (приставка «экзо-» означает наружу), процесс обратный фагоцитозу.
Процесс захвата клеткой твердой пищи называется фагоцитозом. Если же клетка поглощает капельку, то процесс называется пиноцитозом, а пузырек, в котором капля транспортируется в вакуоль – везикулой. Когда клетка заканчивает свои пищеварительные процессы, то ей, как и многоклеточному сложному организму, нужно вывести наружу непереваренные остатки. Тогда происходит экзоцитоз (приставка «экзо-» означает наружу), процесс обратный фагоцитозу.
Мембрана клетки не представляет их себя непрерывную цепь липидов, она имеет включения в виде белков разных конфигураций. Они могут быть на поверхности мембраны, проходить сквозь нее, образовывать каналы, находиться в наружном или внутреннем слое липидов.
Во-первых, это отличительная черта эукариотических организмов. Ядро управляет процессами внутри клетки, а также хранит генетическую информацию, которая передается по наследству.
Мембрана ядра состоит из двух оболочек, пронизанных ядерными порами. Внешняя оболочка ядра шероховатая, она связана с эндоплазматической сетью клетки.
Внешняя оболочка ядра шероховатая, она связана с эндоплазматической сетью клетки.
Строение ядра. * Ядерный сок = кариоплазма.
Через поры тРНК и иРНК выходят в цитоплазму клетки. Тем временем, пока клетка не делится, в ядре располагаются деспирализованные молекулы ДНК, или же хроматин. Хроматином называются молекулы ДНК, которые связаны с белками. В профазе митоза и в профазе первого деления мейоза хроматин спирализуется, то есть наматывается на специальные гистоновые белки как проволока на карандаш. В таком виде ДНК становится компактной. В интерфазе можно увидеть огромные политенные хромосомы. Они настолько большие, что их прекрасно можно рассмотреть и в обычный световой микроскоп, однако образуются такие хромосомы далеко не во всех клетках. 1 хромосома образована 1 молекулой ДНК. Хромосомы могут быть однохроматидными и двухроматидными. Как раз-таки двухроматидными, состоящими из 2х сестринских хроматид, хромосомы становятся после процесса репликации. В центре такие хромосомы соединены особой перетяжкой – центромерой. Каждая хроматида имеет по два плеча, они могут быть разной длины, а могут быть одинаковой. На концах хроматид располагаются теломеры. Интересный факт: старением организма связано с укорачиванием теломер с течением жизни.
В центре такие хромосомы соединены особой перетяжкой – центромерой. Каждая хроматида имеет по два плеча, они могут быть разной длины, а могут быть одинаковой. На концах хроматид располагаются теломеры. Интересный факт: старением организма связано с укорачиванием теломер с течением жизни.
Строение двухроматидной хромосомы
Внутрь клетки проникают неорганические ионы, АТФ, белки и ферменты и т.д. В ядре есть жидкая составляющая, как в клетке, кариоплазма. А в кариоплазме – ядрышки, в которых происходит синтез частей рибосом. В цитоплазме формируются целые рибосомы. В одном ядре могут находиться от 1 до 7 ядрышек, образованных близкими по отношению друг к другу петлями ДНК.
Обычно в клетках располагается одно ядро, но бывают и исключения: эритроциты в ходе созревания утрачивают свое ядро, а клетки мышечной ткани – миоциты, наоборот имеют много ядер.
Источник:
http://spadilo.ru/kletochnaya-membrana-i-yadro/
§20.
 Особенности строения клеток эукариот
Особенности строения клеток эукариот
1. Как называются живые организмы, клетки которых содержат оформленное ядро?
Автотрофы, гетеротрофы, прокариоты, эукариоты.
Живые организмы, клетки которых содержат оформленное ядро, называются эукариотами.
2. В чём проявляется сходство клеток протистов, грибов, растений и животных?
● Клетки устроены по единому плану и состоят из трёх основных частей: поверхностного аппарата (включающего цитоплазматическую мембрану и надмембранный комплекс), цитоплазмы (в состав которой входят гиалоплазма, цитоскелет, включения, разнообразные мембранные и немембранные органоиды) и ядра.
● Сходным образом протекают процессы обмена веществ и энергии.
● Сходные способы деления клеток.
…и (или) другие существенные признаки.
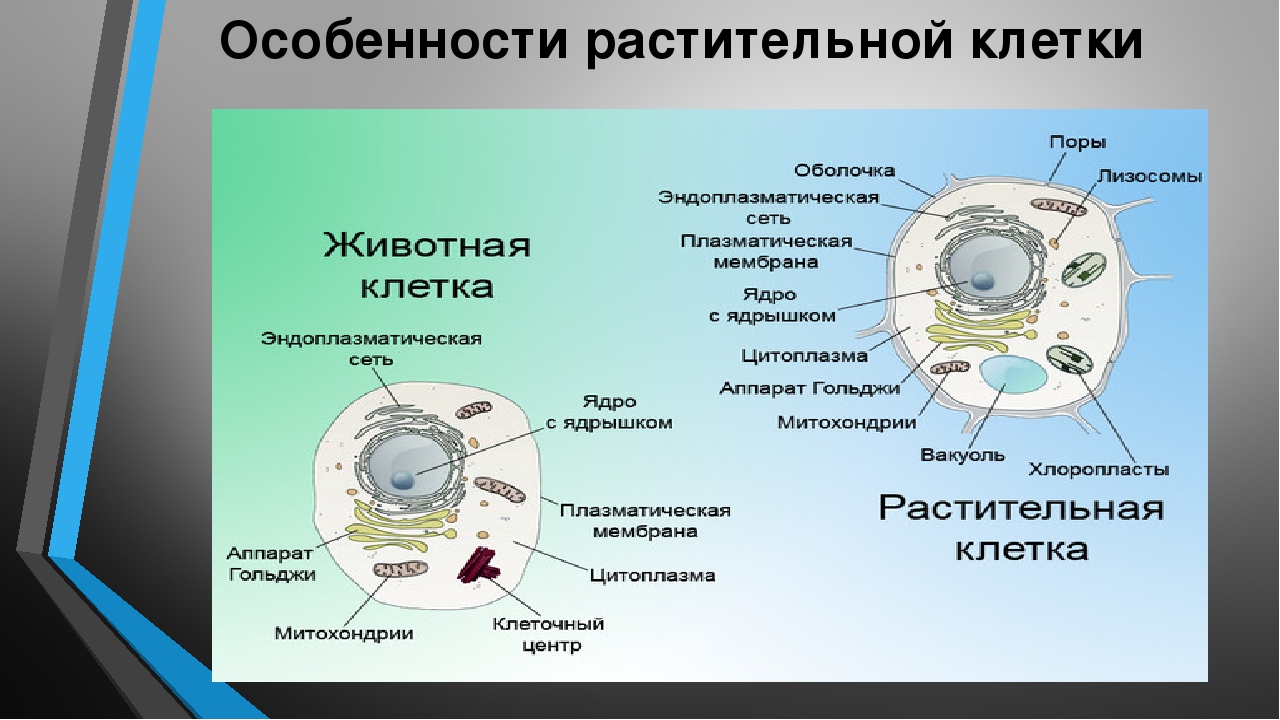
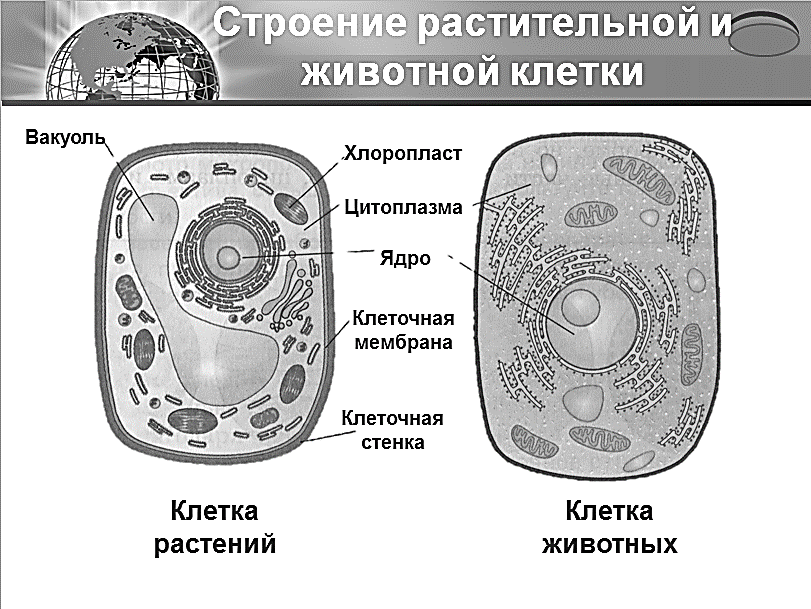
3. Чем растительная клетка отличается от животной?
● Надмембранный комплекс растительной клетки представлен жёсткой клеточной стенкой. Надмембранным комплексом животной клетки является гликокаликс.
Надмембранным комплексом животной клетки является гликокаликс.
● В отличие от животных клеток, для клеток растений характерно наличие пластид (хлоропластов, лейкопластов, хромопластов) и крупных вакуолей.
● Для клеток животных характерно наличие центриолей, которые отсутствуют в клетках большинства растений.
● Резервный полисахарид, который откладывается в клетках растений – крахмал. В клетках животных откладывается другой полисахарид – гликоген.
…и (или) другие существенные признаки.
4. Какие общие черты и какие различия можно выделить, сравнивая клетки разных групп протистов?
По типу питания выделяют три группы протистов: автотрофные, автогетеротрофные и гетеротрофные. Автотрофные и автогетеротрофные протисты называются водорослями.
Сходство:
● Все протисты являются эукариотами, следовательно, для их клеток характерно наличие плазмалеммы, ядра и цитоплазмы, включающей гиалоплазму, цитоскелет, включения, разнообразные мембранные и немембранные органоиды.
● Генетический аппарат представлен линейными молекулами ДНК, связанными с особыми ядерными белками.
● Сходные процессы обмена веществ и способы деления клеток.
Различия:
● В клетках водорослей содержатся хлоропласты (от одного до нескольких десятков) и осуществляется фотосинтез. В клетках гетеротрофных протистов пластиды отсутствуют.
● В отличие от гетеротрофных протистов, многие водоросли имеют клеточную стенку и вакуоли с клеточным соком. В клетках гетеротрофных протистов, в отличие от водорослей, есть пищеварительные вакуоли.
● Некоторые одноклеточные водоросли имеют светочувствительный глазок, а в клетках гетеротрофных протистов его нет.
● В отличие от автотрофных протистов, автогетеротрофные могут не только осуществлять фотосинтез, но и поглощать всей поверхностью тела растворённые в воде органические вещества.
● Среди водорослей есть одноклеточные, колониальные и многоклеточные формы. Гетеротрофные протисты в основном одноклеточные.
…и (или) другие существенные признаки.
5. Сравните клетки грибов, растений и животных по различным критериям. Укажите черты сходства и различия между ними.
Сходство:
● Эукариоты, их клетки покрыты плазмалеммой, содержат ядро и цитоплазму, в состав которой входят гиалоплазма, цитоскелет, включения, разнообразные мембранные и немембранные органоиды. Мембранными органоидами, наличие которых характерно для клеток всех трёх царств, являются: ЭПС, комплекс Гольджи, лизосомы и митохондрии, немембранными – рибосомы.
● Генетический аппарат представлен линейными молекулами ДНК, связанными с особыми ядерными белками.
● Сходные процессы обмена веществ и способы деления клеток.
● Являются многоклеточными (за исключением некоторых грибов).
Различия:
● Надмембранный комплекс клеток животных представлен гликокаликсом, а растений и грибов – клеточной стенкой, причём её основу у растений составляет целлюлоза, а у грибов – хитин.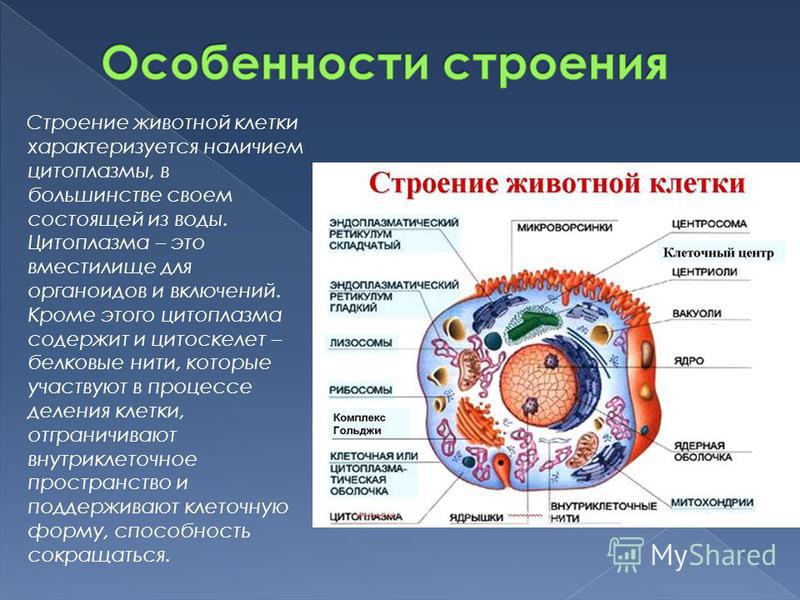
● Тип питания растений – автотрофный (в клетках имеются хлоропласты и другие пластиды, осуществляется фотосинтез), грибов и животных – гетеротрофный (пластиды отсутствуют).
● Запасной углевод клеток грибов и животных – гликоген. В растительных клетках запасается крахмал.
● В отличие от грибов и растений, для клеток животных не характерно наличие вакуолей с клеточным соком.
● Клеточный центр имеется у большинства животных клеток, но отсутствует у большинства растений и грибов.
…и (или) другие существенные признаки.
6. По содержанию белков грибы практически не уступают мясу. Почему же считается, что пищевая ценность грибов гораздо ниже, чем мяса?
Клетки грибов защищены прочной клеточной стенкой, основу которой составляют фибриллы хитина. Хитин не переваривается в организме человека и большинства животных из-за отсутствия фермента хитиназы. Поэтому белки и другие питательные вещества, содержащиеся в неповреждённых клетках грибов (сохранивших целостность хитиновой оболочки), оказываются недоступными для усвоения.
7*. Учёные предполагают, что первые (самые древние) живые организмы на Земле представляли собой наследственный материал (ДНК, РНК), который был окружен вязким раствором белков и ограничен от внешней среды мембраной. Предложите гипотезы, каким образом в процессе эволюции могло возникнуть ядро и различные органоиды, характерные для современных эукариотических клеток.
Например, аутогенная гипотеза предполагает, что эукариотическая клетка возникла путём дифференциации исходной прокариотической клетки. Вначале образовалась наружная мембрана, потом из её впячиваний сформировались отдельные структуры, образовавшие ядерную оболочку и давшие начало органоидам.
Симбиотическая гипотеза (в настоящее время её чаще называют теорией симбиогенеза или теорией эндосимбиоза) предполагает, что эукариотическая клетка возникла вследствие нескольких последовательных симбиозов.
Вначале произошло объединение крупных амёбовидных прокариотических клеток с мелкими аэробными бактериями, которые превратились в митохондрии. Затем крупные амёбовидные клетки вступили в симбиоз со спирохетоподобными бактериями (бактериями с длинными, спирально закрученными клетками), из которых сформировались центриоли и жгутики. Постепенно происходило обособление ядра.
Затем крупные амёбовидные клетки вступили в симбиоз со спирохетоподобными бактериями (бактериями с длинными, спирально закрученными клетками), из которых сформировались центриоли и жгутики. Постепенно происходило обособление ядра.
Ядерные клетки с простейшим набором органоидов могли стать предками гетеротрофных жгутиковых протистов, от которых произошли грибы и животные. Симбиоз ядерных клеток с цианобактериями, преобразовавшимися в хлоропласты, привёл к образованию одноклеточных водорослей. В дальнейшем от водорослей произошли растения.
* Задания, отмеченные звёздочкой, предполагают выдвижение учащимися различных гипотез. Поэтому при выставлении отметки учителю следует ориентироваться не только на ответ, приведённый здесь, а принимать во внимание каждую гипотезу, оценивая биологическое мышление учащихся, логику их рассуждений, оригинальность идей и т. д. После этого целесообразно ознакомить учащихся с приведённым ответом.
Дашков М.Л.
Сайт: dashkov. by
by
Вернуться к оглавлению
| < Предыдущая | Следующая > |
|---|
Тест по теме «Структура и функции органоидов клетки»
Просмотр содержимого документа
«Тест по теме «Структура и функции органоидов клетки»»
Автор-составитель учитель биологии высшей категории Страгулина Е. В.МБОУ «Школа №60» г. Ростова-на-Дону
Тест по теме «Структура и функции органоидов клетки»
Вариант 1
1. Прокариотическая клетка не имеет в своем составе:
ядра;
цитоплазмы;
ЭПС;
плазматической мембраны;
рибосом;
пластид.

2. Цитоплазма выполняет в клетке функции:
внутренней среды клетки;
дыхательного или энергетического центра;
месторасположения ядра и органоидов;
передачи наследственной информации;
связи между ядром и митохондриями;
расщепления белков, жиров, углеводов, нуклеиновых кислот.
3. Ядро клетки имеет в своем составе:
кариоплазму и хроматин;
двух мембранную оболочку;
матрикс с рибосомами и кольцевой ДНК;
ядрышко;
систему мембран, образующих цистерны и канальцы;
стопки диктиосом (телец Гольджи), переходящих в трубочки.
4. Установите соответствие между строением и функциями ЭПС и комплекса Гольджи:
Строение и функции | Органоиды клетки |
А) образует лизосомы Б) участвует в биосинтезе белка В) участвует в построении клеточной стенки Г) состоит из стопочки плоских цистерн и отделяющихся от них пузырьков Д) делит клетку на секции, где происходят противоположные реакции Е) обеспечивает транспорт веществ по трубочкам и цистернам | 1) шероховатая ЭПС 2) комплекс Гольджи |
5. Фагоцитоз – это:
Фагоцитоз – это:
захват клеткой жидкости;
захват твердых частиц;
транспорт веществ через мембрану;
ускорение биохимических реакций.
6. Какой из органоидов клетки участвует в формировании лизосом и транспорте продуктов биосинтеза?
рибосомы;
ЭПС;
комплекс Гольджи;
митохондрии.
7. Какие из перечисленных клеток содержат больше митохондрий?
клетки мякоти листа;
клетки волос млекопитающих;
клетки мозга человека;
клетки коры дерева.
8. Какой из названных организмов является прокариотическим?
амеба;
стрептококк;
инфузория;
дрожжи.
9. Роль ядрышка заключается в формировании:
хромосом;
лизосом;
рибосом;
митохондрий.

10. Какие клетки организма человека называются фагоцитами?
нейроны;
лейкоциты;
миоциты;
эритроциты.
11. Что входит в состав рибосом?
ДНК
липиды
РНК
белки
Тест по теме «Структура и функции органоидов клетки»
Вариант 2
1. Растительная клетка, в отличие от животной, имеет:
пластиды;
целлюлозную клеточную стенку;
углевод – крахмал;
хромосомы;
ядерную мембрану;
углевод – гликоген.
2. Одномембранными органоидами эукариотической клетки являются:
лизосомы;
рибосомы;
центриоли;
ЭПС;
митохондрии;
аппарат Гольджи.

3. Для животной клетки не характерно наличие:
хлоропластов;
рибосом;
целлюлозной клеточной стенки;
оформленного ядра;
вакуолей с клеточным соком;
митохондрий.
4. Установите соответствие между типом клетки и ее характеристикой.
Характеристика клетки | Тип клетки |
А) имеет крахмальные зерна Б) не имеет целлюлозной клеточной стенки В) имеет пластиды Г) содержит крупные вакуоли с клеточным соком Д) имеет клеточный центр Е) имеет целлюлозную клеточную стенку | 1) растительная клетка высших растений 2) животная клетка |
5. Кристы – это:
складки внутренней мембраны митохондрий;
складки наружной мембраны митохондрий;
межмембранные образования;
окислительные ферменты.

6. Функция гладкой ЭПС:
синтез белков;
синтез углеводов и липидов;
синтез АТФ;
синтез РНК.
7. Назовите единственное точное отличие прокариот от эукариот.
прокариоты не способны к автотрофному питанию;
прокариоты не способны к гетеротрофному питанию;
у прокариот нет митохондрий;
у прокариот нет рибосом.
8. Какое из названных веществ является запасным в клетках животных?
целлюлоза;
глюкоза;
гликоген;
белок.
9. Какой из названных функций митохондрии не обладают?
фотосинтез;
синтез белков;
способность размножаться;
синтез АТФ.
10. Какую из перечисленных функций плазматическая мембрана не выполняет?
транспорт веществ;
защиту клетку;
взаимодействие с другими клетками;
синтез белка.

11. Какая из ядерных структур принимает участие в сборке субъединиц рибосом?
ядерная оболочка
ядрышко
ядерный сок
ОТВЕТЫ К ТЕСТУ «ОРГАНОИДЫ КЛЕТКИ»
Вариант 1
1,3,6
1,3,5
1,2,4
А2,Б1,В2,Г2,Д1,Е1
б
в
в
б
в
б
в, г.
Вариант 2
1,2,3
1,4,6
1,3,5
А1,Б2,В1,Г1,Д2,Е1
а
б
в
в
а
г
б
Основные отличия строения растительных и животных клеток
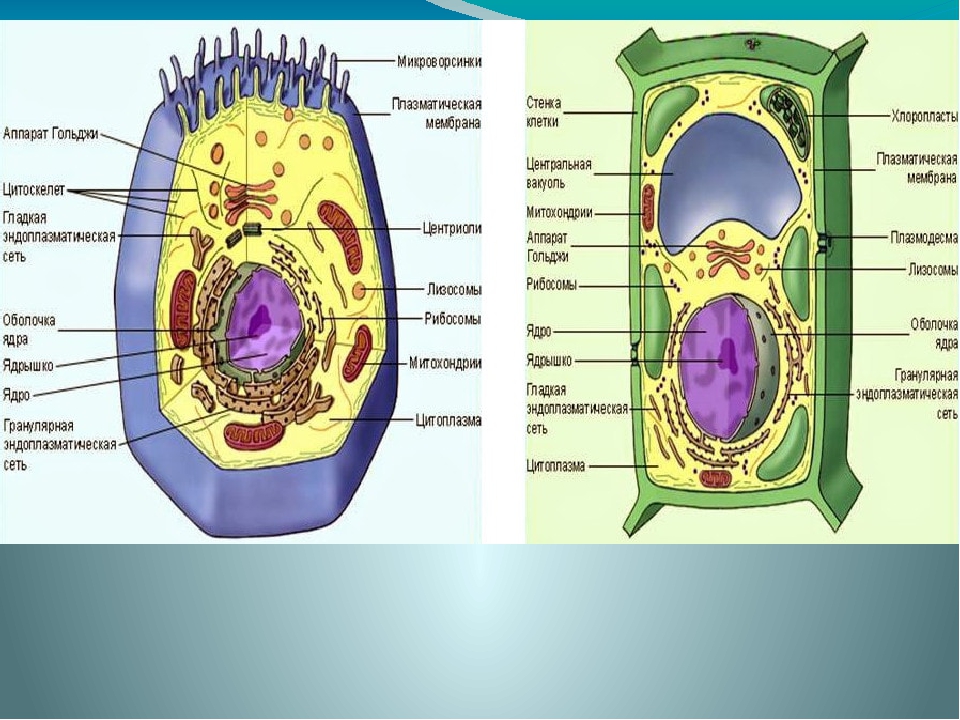
Клетки животных и растений схожи между собой, поскольку они являются эукариотическими клетками, имеющими истинное ядро, которое содержит ДНК и отделено от других клеточных структур ядерной мембраной.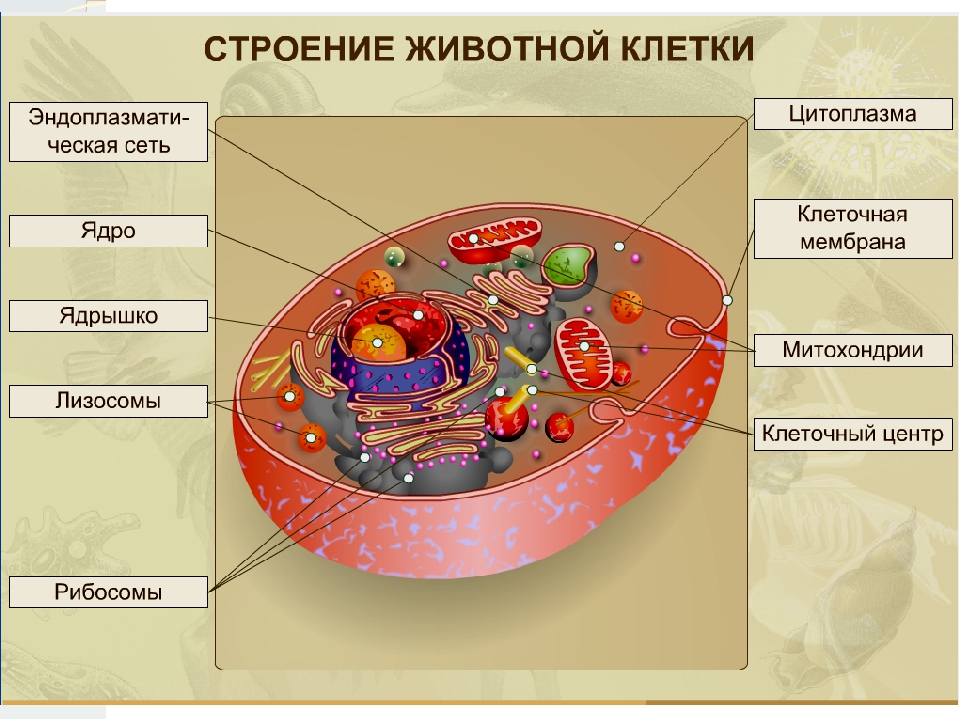 Оба типа клеток имеют сходные процессы размножения (деления), которые включают митоз и мейоз.
Оба типа клеток имеют сходные процессы размножения (деления), которые включают митоз и мейоз.
Животные и растительные клетки получают энергию, используемую ими для роста и поддержания нормального функционирования в процессе клеточного дыхания. Также, характерным для обоих типов клеток является наличие клеточных структур, известных как органеллы, которые специализированы для выполнения конкретных функций, необходимых для нормальной работы. Животные и растительные клетки объединяет наличие ядра, комплекса Гольджи, эндоплазматического ретикулума, рибосом, митохондрий, пероксисом, цитоскелета и клеточной (плазматической) мембраны. Несмотря на схожие характеристики животных и растительных клеток, они также имею множество различий, которые рассмотрены ниже.
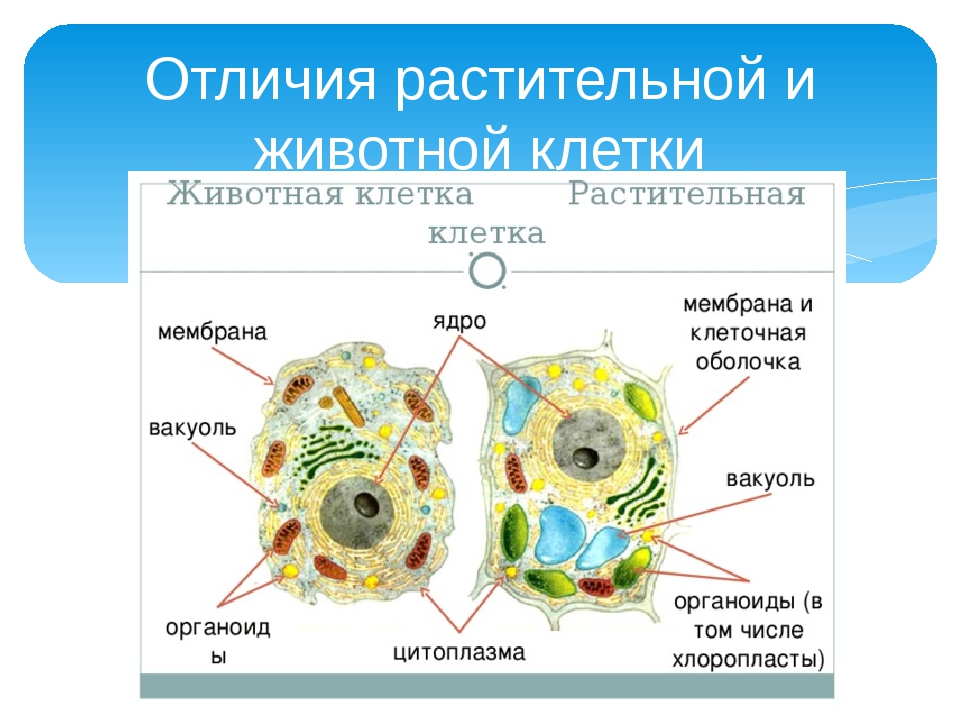
Основные различия в клетках животных и растений
Схема строения животной и растительной клеток- Размер: клетки животных, как правило, меньше, чем растительные клетки. Размер животных клеток колеблются от 10 до 30 микрометров в длину, а клеток растений — от 10 до 100 микрометров.
- Форма: клетки животных бывают разных размеров и имеют округлые или неправильные формы. Растительные клетки более схожи по размеру и обычно имеют форму прямоугольника или куба.
- Хранение энергии: животные клетки хранят энергию в виде сложного углеводного гликогена. Растительные клетки хранят энергию в виде крахмала.
- Белки: из 20 аминокислот, необходимых для синтеза белков, только 10 производятся естественным образом в клетках животных. Другие так называемые незаменимые аминокислоты получаются из пищи. Растения способны синтезировать все 20 аминокислот.
- Дифференциация: у животных только стволовые клетки способны превращаться в другие типы клеток. Большинство типов растительных клеток способны дифференцироваться.
- Рост: клетки животных увеличиваются в размерах, увеличивая число клеток. Растительные клетки в основном увеличивают размер клеток, становясь более крупными. Они растут, накапливая больше воды в центральной вакуоли.
- Клеточная стенка: у клеток животных нет клеточной стенки, но есть клеточная мембрана. Клетки растений имеют клеточную стенку, состоящую из целлюлозы, а также клеточной мембраны.
- Центриоли: клетки животных содержат эти цилиндрические структуры, которые организуют сборку микротрубочек во время деления клеток. Клетки растений обычно не содержат центриоли.
- Реснички: встречаются в клетках животных, но, как правило, отсутствуют в растительных клетках. Реснички — это микротрубочки, которые обеспечивают клеточную локомоцию.
- Цитокинез: разделение цитоплазмы при делении клеток, происходит в клетках животных, когда образуется спайная борозда, которая зажимает клеточную мембрану пополам. В цитокинезе растительных клеток образуется клеточная пластинка, разделяющая клетку.
- Гликсисомы: эти структуры не встречаются в животных клетках, но присутствуют в растительных. Гликсисомы помогают расщеплять липиды на сахара, особенно в прорастающих семенах.
- Лизосомы: клетки животных обладают лизосомами, которые содержат ферменты, переваривающие клеточные макромолекулы. Растительные клетки редко содержат лизосомы, поскольку вакуоль растения обрабатывает деградацию молекулы.
- Пластиды: в животных клетках нет пластид. Растительные клетки имеют такие пластиды, как хлоропласты, необходимые для фотосинтеза.
- Плазмодесмы: клетки животных не имеют плазмодесм. Растительные клетки содержат плазмодесмы, которые представляет собой поры между стенками, позволяющие молекулам и коммуникационным сигналам проходить между отдельными клетками растений.
- Вакуоль: животные клетки могут иметь много маленьких вакуолей. Клетки растений содержат большую центральную вакуоль, которая может составляет до 90% объема клетки.
 Они растут, накапливая больше воды в центральной вакуоли.
Они растут, накапливая больше воды в центральной вакуоли. Гликсисомы помогают расщеплять липиды на сахара, особенно в прорастающих семенах.
Гликсисомы помогают расщеплять липиды на сахара, особенно в прорастающих семенах.Читайте также: Эукариотические и прокариотические клетки: функции, строение и отличия.

Прокариотические клетки
Эукариотические клетки животных и растений также отличаются от прокариотических клеток, таких как бактерии. Прокариоты обычно являются одноклеточными организмами, тогда как животные и растительные клетки обычно многоклеточные. Эукариоты более сложны и больше, чем прокариоты. К клеткам животных и растений относятся многие органеллы, не обнаруженные в прокариотических клетках. Прокариоты не имеют истинного ядра, поскольку ДНК не содержится в мембране, а свернута в области цитоплазмы, называемой нуклеоидом. В то время как животные и растительные клетки размножаются митозом или мейозом, прокариоты чаще всего размножаются с помощью деления или дробления.
Другие эукариотические организмы
Клетки растений и животных не являются единственными типами эукариотических клеток. Протесты (например, эвглена и амеба) и грибы (например, грибы, дрожжи и плесень) — два других примера эукариотических организмов.
Не нашли, то что искали? Используйте форму поиска по сайту
Понравилась статья? Оставь комментарий и поделись с друзьями
4.
 3E: Сравнение клеток растений и животных
3E: Сравнение клеток растений и животныхХотя обе они являются эукариотическими клетками, между клетками животных и растений существуют уникальные структурные различия.
Цели обучения
- Различать структуры, обнаруженные в клетках животных и растений
Ключевые моменты
- Центросомы и лизосомы находятся в клетках животных, но не существуют в клетках растений.
- Лизосомы — это «мусоропровод» животных клеток, в то время как в растительных клетках та же функция выполняется в вакуолях.
- Растительные клетки имеют клеточную стенку, хлоропласты и другие специализированные пластиды, а также большую центральную вакуоль, которых нет в клетках животных.
- Стенка ячейки — это жесткое покрытие, которое защищает ячейку, обеспечивает структурную поддержку и придает форму ячейке.
- Хлоропласты, обнаруженные в клетках растений, содержат зеленый пигмент, называемый хлорофиллом, который улавливает световую энергию, которая управляет реакциями фотосинтеза растений.
- Центральная вакуоль играет ключевую роль в регулировании концентрации воды в растительной клетке при изменении условий окружающей среды.
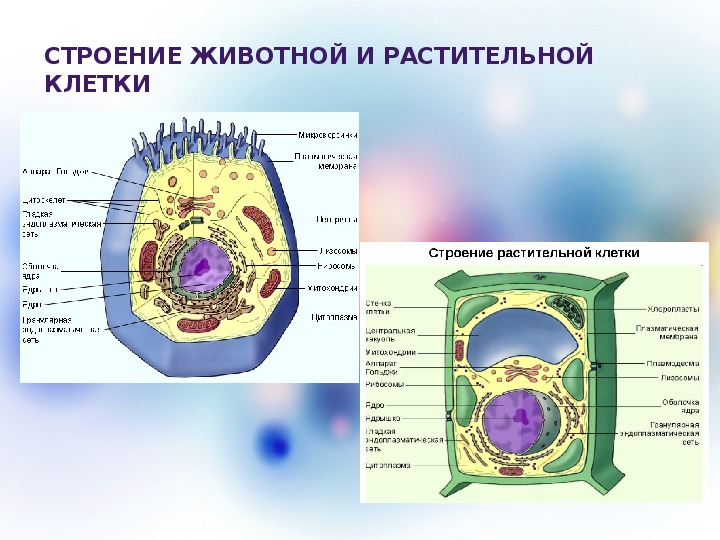
Ключевые термины
- протист : любой из эукариотических одноклеточных организмов, включая простейшие, слизистые плесени и некоторые водоросли; исторически сгруппированы в королевство Протоктисты.
- автотроф : любой организм, который может синтезировать пищу из неорганических веществ, используя тепло или свет в качестве источника энергии
- гетеротроф : организм, которому требуется внешний источник энергии в виде пищи, поскольку он не может синтезировать свою собственную
Клетки животных в сравнении с клетками растений
Каждая эукариотическая клетка имеет плазматическую мембрану, цитоплазму, ядро, рибосомы, митохондрии, пероксисомы и в некоторых случаях вакуоли; однако между клетками животных и растений существуют поразительные различия. В то время как как животные, так и растительные клетки имеют центры организации микротрубочек (MTOC), животные клетки также имеют центриоли, связанные с MTOC: комплекс, называемый центросомой. Каждая клетка животных имеет центросому и лизосомы, тогда как клетки растений не имеют. У растительных клеток есть клеточная стенка, хлоропласты и другие специализированные пластиды, а также большая центральная вакуоль, тогда как у животных клеток нет.
В то время как как животные, так и растительные клетки имеют центры организации микротрубочек (MTOC), животные клетки также имеют центриоли, связанные с MTOC: комплекс, называемый центросомой. Каждая клетка животных имеет центросому и лизосомы, тогда как клетки растений не имеют. У растительных клеток есть клеточная стенка, хлоропласты и другие специализированные пластиды, а также большая центральная вакуоль, тогда как у животных клеток нет.
Центросома
Центросома — это центр организации микротрубочек, расположенный рядом с ядрами клеток животных.Он содержит пару центриолей, две структуры, которые лежат перпендикулярно друг другу. Каждая центриоль представляет собой цилиндр из девяти троек микротрубочек. Центросома (органелла, где берут начало все микротрубочки) реплицируется до деления клетки, и центриоли, по-видимому, играют определенную роль в притяжении дублированных хромосом к противоположным концам делящейся клетки. Однако точная функция центриолей в делении клеток не ясна, потому что клетки, у которых была удалена центросома, все еще могут делиться; и клетки растений, в которых отсутствуют центросомы, способны к делению клеток.
Лизосомы
В клетках животных есть еще один набор органелл, которых нет в клетках растений: лизосомы. Лизосомы — это «мусоропровод» клетки. В растительных клетках пищеварительные процессы происходят в вакуолях.Ферменты в лизосомах способствуют расщеплению белков, полисахаридов, липидов, нуклеиновых кислот и даже изношенных органелл. Эти ферменты активны при гораздо более низком pH, чем у цитоплазмы. Следовательно, pH в лизосомах более кислый, чем pH цитоплазмы. Многие реакции, происходящие в цитоплазме, не могут происходить при низком pH, поэтому преимущество разделения эукариотической клетки на органеллы очевидно.
Стенка клетки
Стенка ячейки — это жесткое покрытие, которое защищает ячейку, обеспечивает структурную поддержку и придает форму ячейке.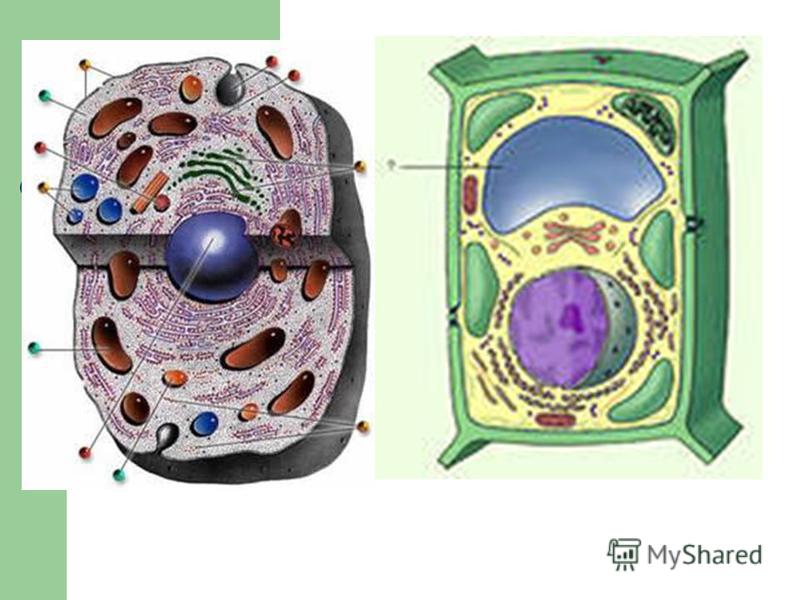 Грибковые и протистанские клетки также имеют клеточные стенки. В то время как основным компонентом стенок прокариотических клеток является пептидогликан, основной органической молекулой в стенке растительной клетки является целлюлоза, полисахарид, состоящий из единиц глюкозы. Когда вы надкусываете сырые овощи, например сельдерей, они хрустят. Это потому, что вы зубами разрываете жесткие клеточные стенки клеток сельдерея.
Грибковые и протистанские клетки также имеют клеточные стенки. В то время как основным компонентом стенок прокариотических клеток является пептидогликан, основной органической молекулой в стенке растительной клетки является целлюлоза, полисахарид, состоящий из единиц глюкозы. Когда вы надкусываете сырые овощи, например сельдерей, они хрустят. Это потому, что вы зубами разрываете жесткие клеточные стенки клеток сельдерея.
Хлоропласты
Подобно митохондриям, хлоропласты имеют собственную ДНК и рибосомы, но хлоропласты выполняют совершенно другую функцию. Хлоропласты — это органеллы растительной клетки, осуществляющие фотосинтез. Фотосинтез — это серия реакций, в которых для производства глюкозы и кислорода используются углекислый газ, вода и световая энергия.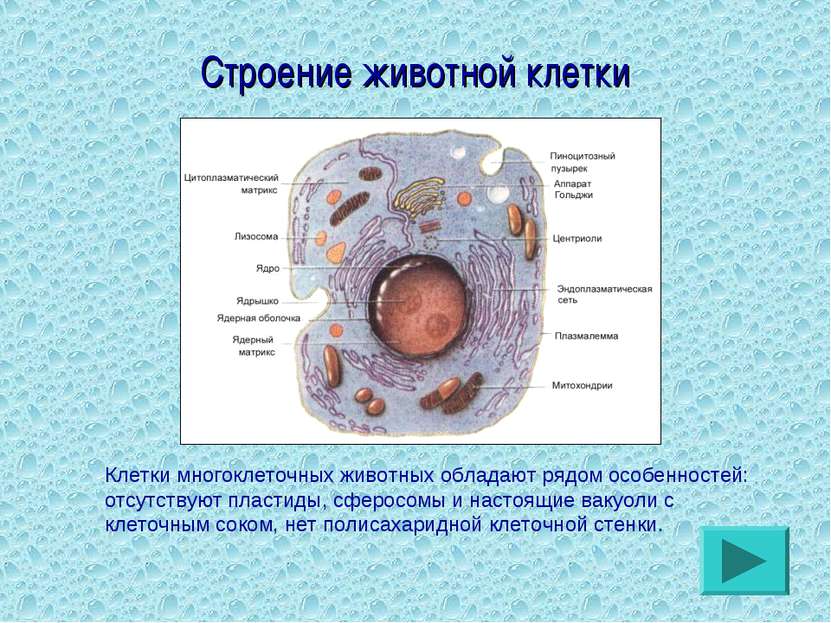 Это главное различие между растениями и животными; растения (автотрофы) могут производить себе пищу, как сахар, в то время как животные (гетеротрофы) должны принимать их пищу.
Это главное различие между растениями и животными; растения (автотрофы) могут производить себе пищу, как сахар, в то время как животные (гетеротрофы) должны принимать их пищу.
Подобно митохондриям, хлоропласты имеют внешнюю и внутреннюю мембраны, но внутри пространства, ограниченного внутренней мембраной хлоропласта, находится набор взаимосвязанных и уложенных друг в друга заполненных жидкостью мембранных мешочков, называемых тилакоидами. Каждый стек тилакоидов называется гранумом (множественное число = грана). Жидкость, заключенная во внутренней мембране, окружающей грану, называется стромой.
Рисунок \ (\ PageIndex {1} \): Структура хлоропласта : Хлоропласт имеет внешнюю мембрану, внутреннюю мембрану и мембранные структуры, называемые тилакоидами, которые сложены в грану. Пространство внутри тилакоидных мембран называется тилакоидным пространством. Реакции сбора света происходят в тилакоидных мембранах, а синтез сахара происходит в жидкости внутри внутренней мембраны, которая называется стромой.
Хлоропласты содержат зеленый пигмент под названием хлорофилл, который улавливает световую энергию, которая запускает реакции фотосинтеза.Как и в растительных клетках, у фотосинтезирующих протистов есть хлоропласты. Некоторые бактерии осуществляют фотосинтез, но их хлорофилл не относится к органеллам.
Центральная вакуоль
Центральная вакуоль играет ключевую роль в регулировании концентрации воды в клетках при изменении условий окружающей среды. Если вы на несколько дней забываете полить растение, оно увядает. Это связано с тем, что когда концентрация воды в почве становится ниже, чем концентрация воды в растении, вода выходит из центральных вакуолей и цитоплазмы.По мере того как центральная вакуоль сжимается, она оставляет клеточную стенку без поддержки. Эта потеря поддержки клеточных стенок растительных клеток приводит к увяданию растения. Центральная вакуоль также поддерживает расширение клетки. Когда центральная вакуоль содержит больше воды, клетка становится больше, не тратя много энергии на синтез новой цитоплазмы.
ЛИЦЕНЗИИ И АТРИБУЦИИ
CC ЛИЦЕНЗИОННЫЙ КОНТЕНТ, ПРЕДЫДУЩИЙ РАЗДЕЛ
- Курирование и проверка. Предоставлено : Boundless.com. Лицензия : CC BY-SA: Attribution-ShareAlike
CC ЛИЦЕНЗИОННОЕ СОДЕРЖАНИЕ, СПЕЦИАЛЬНАЯ АТРИБУЦИЯ
- Колледж OpenStax, Биология. 16 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44407/latest…ol11448/latest . Лицензия : CC BY: Attribution
- Колледж OpenStax, Эукариотические клетки.22 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m45432/latest/ . Лицензия : CC BY: Attribution
- органелла. Источник : Викисловарь. Расположен по адресу : en.wiktionary.org/wiki/organelle . Лицензия : CC BY-SA: Attribution-ShareAlike
- фотосинтез. Источник : Викисловарь. Находится по адресу : en.wiktionary.org/wiki/photosynthesis . Лицензия : CC BY-SA: Attribution-ShareAlike
- эукариотический. Источник : Викисловарь. Расположен по адресу : en.wiktionary.org/wiki/eukaryotic . Лицензия : CC BY-SA: Attribution-ShareAlike
- Колледж OpenStax, Эукариотические клетки.22 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m45432/latest/ . Лицензия : CC BY: Attribution
- Колледж OpenStax, Эукариотические клетки. 16 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44407/latest…e_04_03_04.jpg . Лицензия : CC BY: Attribution
- Колледж OpenStax, Эукариотические клетки.22 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m45432/latest/ . Лицензия : CC BY: Attribution
- Колледж OpenStax, Биология. 16 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44407/latest…ol11448/latest . Лицензия : CC BY: Attribution
- Лаура Мартин, Открытие структуры плазменной мембраны. 23 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/col10470/latest/ . Лицензия : CC BY: Attribution
- фосфолипид. Источник : Викисловарь. Расположен по адресу : en.wiktionary.org/wiki/phospholipid . Лицензия : CC BY-SA: Attribution-ShareAlike
- гипертонический. Источник : Викисловарь. Расположен по адресу : en.wiktionary.org/wiki/hypertonic . Лицензия : CC BY-SA: Attribution-ShareAlike
- гипотонический. Источник : Викисловарь. Находится по адресу : en.wiktionary.org/wiki/hypotonic . Лицензия : CC BY-SA: Attribution-ShareAlike
- Колледж OpenStax, Эукариотические клетки. 22 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m45432/latest/ . Лицензия : CC BY: Attribution
- Колледж OpenStax, Эукариотические клетки. 16 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44407/latest…e_04_03_04.jpg . Лицензия : CC BY: Attribution
- Колледж OpenStax, Эукариотические клетки.22 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m45432/latest/ . Лицензия : CC BY: Attribution
- Колледж OpenStax, Эукариотические клетки. 16 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44407/latest…e_04_03_02.jpg . Лицензия : CC BY: Attribution
- Колледж OpenStax, Клеточная мембрана.23 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m46021/latest/ . Лицензия : CC BY: Attribution
- Колледж OpenStax, Клеточная мембрана. 23 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m46021/latest/ . Лицензия : CC BY: Attribution
- Колледж OpenStax, Биология.16 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx. org/content/m44407/latest…ol11448/latest . Лицензия : CC BY: Attribution
- Колледж OpenStax, Биология. 21 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44407/latest…ol11448/latest . Лицензия : CC BY: Attribution
- гистон. Источник : Викисловарь. Расположен по адресу : http://en.wiktionary.org/wiki/histone . Лицензия : CC BY-SA: Attribution-ShareAlike
- хроматин. Источник : Викисловарь. Расположен по адресу : en.wiktionary.org/wiki/chromatin . Лицензия : CC BY-SA: Attribution-ShareAlike
- Безграничный. Предоставлено : Безграничное обучение. Адрес: : www.boundless.com//biology/de…n/nucleolus-2 . Лицензия : CC BY-SA: Attribution-ShareAlike
- Колледж OpenStax, Эукариотические клетки. 22 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m45432/latest/ . Лицензия : CC BY: Attribution
- Колледж OpenStax, Эукариотические клетки.16 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44407/latest…e_04_03_04.jpg . Лицензия : CC BY: Attribution
- Колледж OpenStax, Эукариотические клетки. 22 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m45432/latest/ . Лицензия : CC BY: Attribution
- Колледж OpenStax, Эукариотические клетки.16 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44407/latest…e_04_03_02.jpg . Лицензия : CC BY: Attribution
- Колледж OpenStax, Клеточная мембрана. 23 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m46021/latest/ . Лицензия : CC BY: Attribution
- Колледж OpenStax, Клеточная мембрана.23 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx. org/content/m46021/latest/ . Лицензия : CC BY: Attribution
- Колледж OpenStax, Биология. 23 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44407/latest…ol11448/latest . Лицензия : CC BY: Attribution
- Колледж OpenStax, Ядро и репликация ДНК.23 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m46073/latest/ . Лицензия : CC BY: Attribution
- Колледж OpenStax, Биология. 23 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44407/latest…ol11448/latest . Лицензия : CC BY: Attribution
- Колледж OpenStax, Биология.16 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44407/latest…ol11448/latest . Лицензия : CC BY: Attribution
- Колледж OpenStax, Эукариотическое происхождение. 22 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44614/latest/ . Лицензия : CC BY: Attribution
- кофактор. Источник : Викисловарь. Адрес: : en.wiktionary.org/wiki/cofactor . Лицензия : CC BY-SA: Attribution-ShareAlike
- аденозинтрифосфат. Источник : Википедия. Расположен по адресу : en.Wikipedia.org/wiki/adenosine%20triphosphate . Лицензия : CC BY-SA: Attribution-ShareAlike
- альфа-протеобактерий. Источник : Википедия. Расположен по адресу : en.Wikipedia.org/wiki/alpha-proteobacteria . Лицензия : CC BY-SA: Attribution-ShareAlike
- Митохондрия. Источник : Википедия. Расположен по адресу : en.Wikipedia.org/wiki/Mitochondrion . Лицензия : CC BY-SA: Attribution-ShareAlike
- Колледж OpenStax, Эукариотические клетки.22 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx. org/content/m45432/latest/ . Лицензия : CC BY: Attribution
- Колледж OpenStax, Эукариотические клетки. 16 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44407/latest…e_04_03_04.jpg . Лицензия : CC BY: Attribution
- Колледж OpenStax, Эукариотические клетки.22 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m45432/latest/ . Лицензия : CC BY: Attribution
- Колледж OpenStax, Эукариотические клетки. 16 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44407/latest. ..e_04_03_02.jpg . Лицензия : CC BY: Attribution
- Колледж OpenStax, Клеточная мембрана.23 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m46021/latest/ . Лицензия : CC BY: Attribution
- Колледж OpenStax, Клеточная мембрана. 23 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m46021/latest/ . Лицензия : CC BY: Attribution
- Колледж OpenStax, Биология.23 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44407/latest…ol11448/latest . Лицензия : CC BY: Attribution
- Колледж OpenStax, Ядро и репликация ДНК. 23 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m46073/latest/ . Лицензия : CC BY: Attribution
- Колледж OpenStax, Биология.23 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44407/latest…ol11448/latest . Лицензия : CC BY: Attribution
- Колледж OpenStax, Эукариотические клетки. 16 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44407/latest…e_04_03_07.jpg . Лицензия : CC BY: Attribution
- Колледж OpenStax, Биология.16 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44407/latest…ol11448/latest . Лицензия : CC BY: Attribution
- протист. Источник : Викисловарь. Расположен по адресу : http://en.wiktionary.org/wiki/protist . Лицензия : CC BY-SA: Attribution-ShareAlike
- гетеротроф. Источник : Викисловарь. Находится по адресу : en.wiktionary.org/wiki/heterotroph . Лицензия : CC BY-SA: Attribution-ShareAlike
- автотроф. Источник : Викисловарь. Находится по адресу : en.wiktionary.org/wiki/autotroph . Лицензия : CC BY-SA: Attribution-ShareAlike
- Колледж OpenStax, Эукариотические клетки. 22 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m45432/latest/ . Лицензия : CC BY: Attribution
- Колледж OpenStax, Эукариотические клетки. 16 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44407/latest…e_04_03_04.jpg . Лицензия : CC BY: Attribution
- Колледж OpenStax, Эукариотические клетки.22 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m45432/latest/ . Лицензия : CC BY: Attribution
- Колледж OpenStax, Эукариотические клетки. 16 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44407/latest…e_04_03_02.jpg . Лицензия : CC BY: Attribution
- Колледж OpenStax, Клеточная мембрана.23 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m46021/latest/ . Лицензия : CC BY: Attribution
- Колледж OpenStax, Клеточная мембрана. 23 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m46021/latest/ . Лицензия : CC BY: Attribution
- Колледж OpenStax, Биология.23 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx. org/content/m44407/latest…ol11448/latest . Лицензия : CC BY: Attribution
- Колледж OpenStax, Ядро и репликация ДНК. 23 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m46073/latest/ . Лицензия : CC BY: Attribution
- Колледж OpenStax, Биология.23 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44407/latest…ol11448/latest . Лицензия : CC BY: Attribution
- Колледж OpenStax, Эукариотические клетки. 16 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44407/latest. ..e_04_03_07.jpg . Лицензия : CC BY: Attribution
- Колледж OpenStax, Эукариотические клетки.16 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44407/latest…e_04_03_08.jpg . Лицензия : CC BY: Attribution
- Колледж OpenStax, Эукариотические клетки. 16 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44407/latest…e_04_03_10.jpg . Лицензия : CC BY: Attribution
- Колледж OpenStax, Эукариотические клетки.16 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m44407/latest. ..e_04_03_09.jpg . Лицензия : CC BY: Attribution
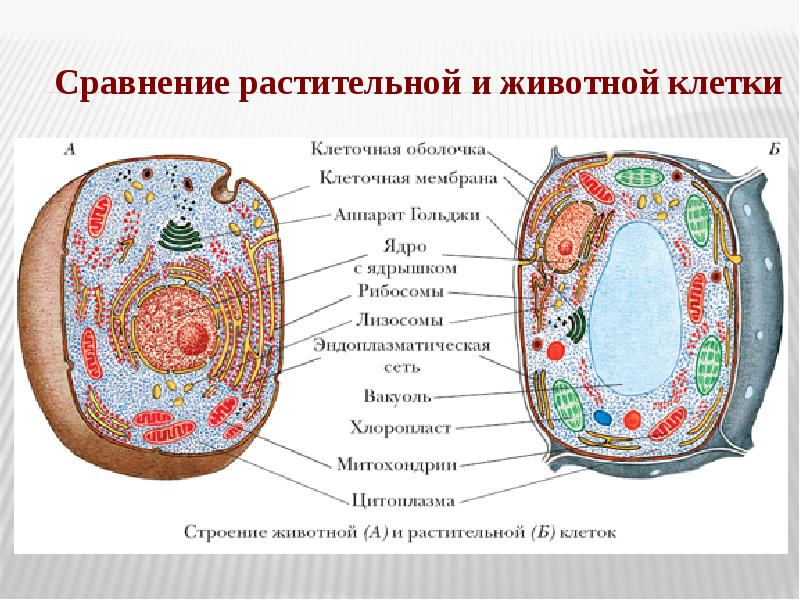 Источник : Викисловарь. Расположен по адресу : en.wiktionary.org/wiki/organelle . Лицензия : CC BY-SA: Attribution-ShareAlike
Источник : Викисловарь. Расположен по адресу : en.wiktionary.org/wiki/organelle . Лицензия : CC BY-SA: Attribution-ShareAlike 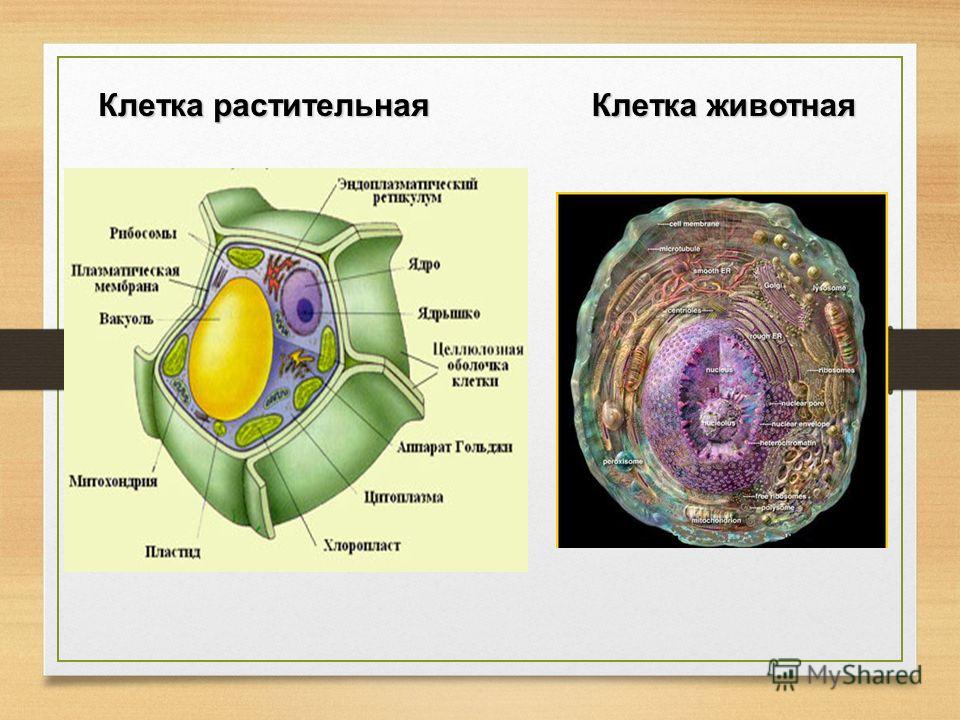 Лицензия : CC BY: Attribution
Лицензия : CC BY: Attribution  23 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/col10470/latest/ . Лицензия : CC BY: Attribution
23 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/col10470/latest/ . Лицензия : CC BY: Attribution  22 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m45432/latest/ . Лицензия : CC BY: Attribution
22 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m45432/latest/ . Лицензия : CC BY: Attribution  Расположен по адресу : http://cnx.org/content/m44407/latest…e_04_03_02.jpg . Лицензия : CC BY: Attribution
Расположен по адресу : http://cnx.org/content/m44407/latest…e_04_03_02.jpg . Лицензия : CC BY: Attribution 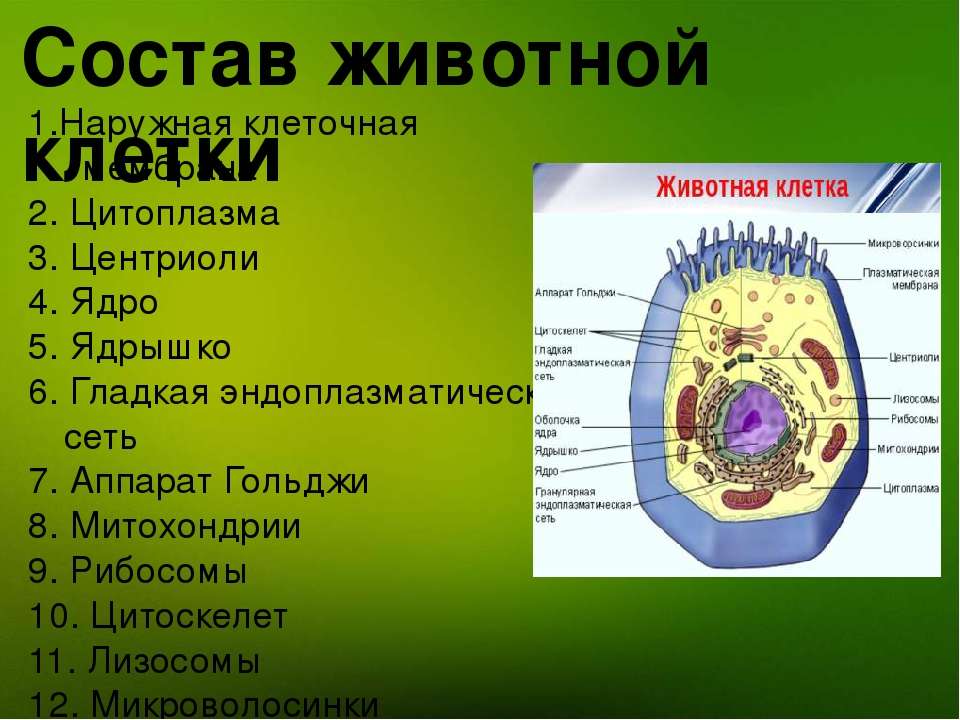 org/content/m44407/latest…ol11448/latest . Лицензия : CC BY: Attribution
org/content/m44407/latest…ol11448/latest . Лицензия : CC BY: Attribution 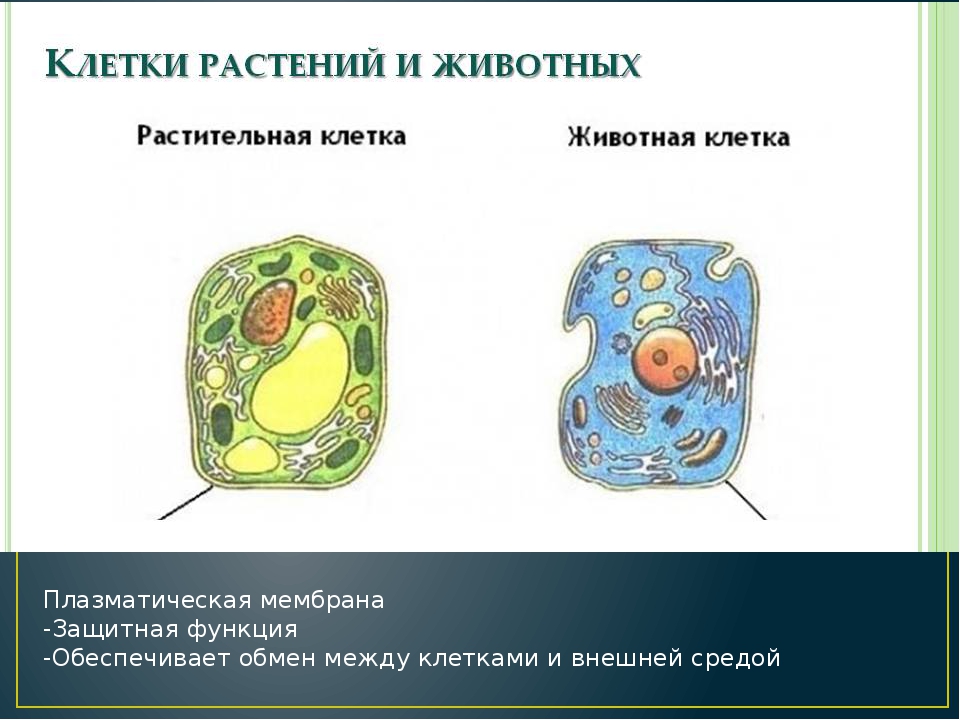 Предоставлено : Безграничное обучение. Адрес: : www.boundless.com//biology/de…n/nucleolus-2 . Лицензия : CC BY-SA: Attribution-ShareAlike
Предоставлено : Безграничное обучение. Адрес: : www.boundless.com//biology/de…n/nucleolus-2 . Лицензия : CC BY-SA: Attribution-ShareAlike  Расположен по адресу : http://cnx.org/content/m45432/latest/ . Лицензия : CC BY: Attribution
Расположен по адресу : http://cnx.org/content/m45432/latest/ . Лицензия : CC BY: Attribution  org/content/m46021/latest/ . Лицензия : CC BY: Attribution
org/content/m46021/latest/ . Лицензия : CC BY: Attribution 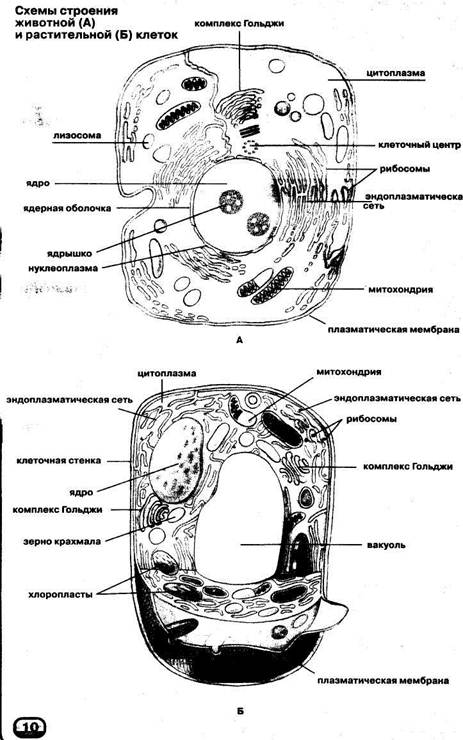 Лицензия : CC BY: Attribution
Лицензия : CC BY: Attribution 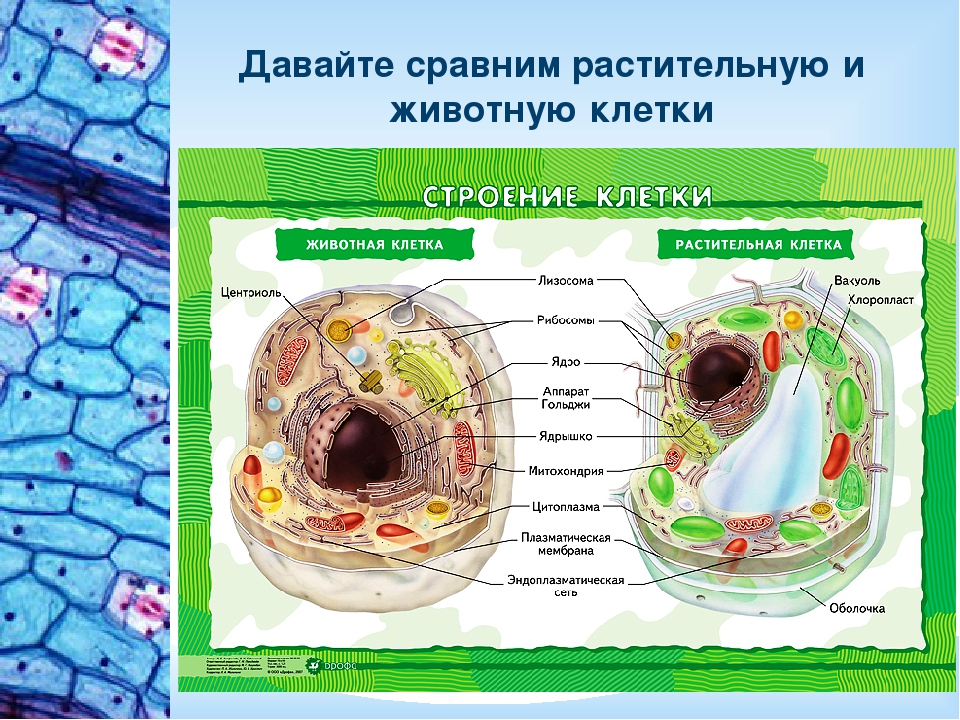 Источник : Википедия. Расположен по адресу : en.Wikipedia.org/wiki/adenosine%20triphosphate . Лицензия : CC BY-SA: Attribution-ShareAlike
Источник : Википедия. Расположен по адресу : en.Wikipedia.org/wiki/adenosine%20triphosphate . Лицензия : CC BY-SA: Attribution-ShareAlike  org/content/m45432/latest/ . Лицензия : CC BY: Attribution
org/content/m45432/latest/ . Лицензия : CC BY: Attribution  ..e_04_03_02.jpg . Лицензия : CC BY: Attribution
..e_04_03_02.jpg . Лицензия : CC BY: Attribution  23 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m46073/latest/ . Лицензия : CC BY: Attribution
23 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m46073/latest/ . Лицензия : CC BY: Attribution  Расположен по адресу : http://cnx.org/content/m44407/latest…ol11448/latest . Лицензия : CC BY: Attribution
Расположен по адресу : http://cnx.org/content/m44407/latest…ol11448/latest . Лицензия : CC BY: Attribution  22 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m45432/latest/ . Лицензия : CC BY: Attribution
22 октября 2013 г. Предоставлено : OpenStax CNX. Расположен по адресу : http://cnx.org/content/m45432/latest/ . Лицензия : CC BY: Attribution  Расположен по адресу : http://cnx.org/content/m44407/latest…e_04_03_02.jpg . Лицензия : CC BY: Attribution
Расположен по адресу : http://cnx.org/content/m44407/latest…e_04_03_02.jpg . Лицензия : CC BY: Attribution  org/content/m44407/latest…ol11448/latest . Лицензия : CC BY: Attribution
org/content/m44407/latest…ol11448/latest . Лицензия : CC BY: Attribution  ..e_04_03_07.jpg . Лицензия : CC BY: Attribution
..e_04_03_07.jpg . Лицензия : CC BY: Attribution  ..e_04_03_09.jpg . Лицензия : CC BY: Attribution
..e_04_03_09.jpg . Лицензия : CC BY: Attribution 4.7C: Сравнение клеток растений и животных
Хотя обе они являются эукариотическими клетками, между клетками животных и растений существуют уникальные структурные различия.
Цели обучения
- Различать структуры, обнаруженные в клетках животных и растений
Ключевые моменты
- Центросомы и лизосомы находятся в клетках животных, но не существуют в клетках растений.
- Лизосомы — это «мусоропровод» животных клеток, в то время как в растительных клетках та же функция выполняется в вакуолях.
- Растительные клетки имеют клеточную стенку, хлоропласты и другие специализированные пластиды, а также большую центральную вакуоль, которых нет в клетках животных.
- Стенка ячейки — это жесткое покрытие, которое защищает ячейку, обеспечивает структурную поддержку и придает форму ячейке.
- Хлоропласты, обнаруженные в клетках растений, содержат зеленый пигмент, называемый хлорофиллом, который улавливает световую энергию, которая управляет реакциями фотосинтеза растений.
- Центральная вакуоль играет ключевую роль в регулировании концентрации воды в растительной клетке при изменении условий окружающей среды.
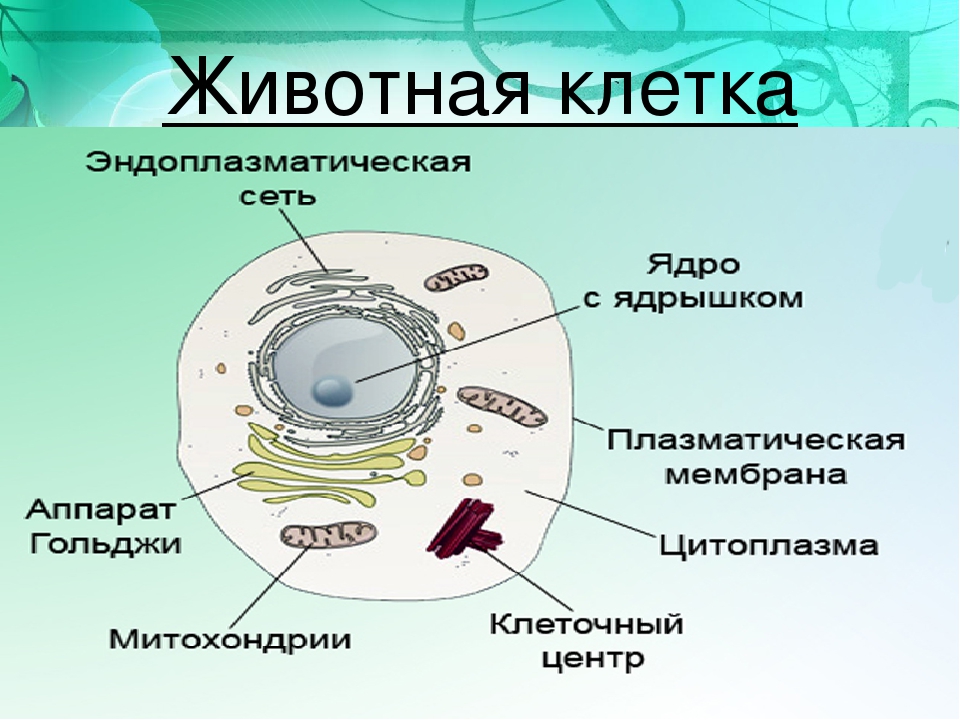
Ключевые термины
- протист : любой из эукариотических одноклеточных организмов, включая простейшие, слизистые плесени и некоторые водоросли; исторически сгруппированы в королевство Протоктисты.
- автотроф : любой организм, который может синтезировать пищу из неорганических веществ, используя тепло или свет в качестве источника энергии
- гетеротроф : организм, которому требуется внешний источник энергии в виде пищи, поскольку он не может синтезировать свою собственную
Клетки животных в сравнении с клетками растений
Каждая эукариотическая клетка имеет плазматическую мембрану, цитоплазму, ядро, рибосомы, митохондрии, пероксисомы и в некоторых случаях вакуоли; однако между клетками животных и растений существуют поразительные различия. В то время как как животные, так и растительные клетки имеют центры организации микротрубочек (MTOC), животные клетки также имеют центриоли, связанные с MTOC: комплекс, называемый центросомой. Каждая клетка животных имеет центросому и лизосомы, тогда как клетки растений не имеют. У растительных клеток есть клеточная стенка, хлоропласты и другие специализированные пластиды, а также большая центральная вакуоль, тогда как у животных клеток нет.
В то время как как животные, так и растительные клетки имеют центры организации микротрубочек (MTOC), животные клетки также имеют центриоли, связанные с MTOC: комплекс, называемый центросомой. Каждая клетка животных имеет центросому и лизосомы, тогда как клетки растений не имеют. У растительных клеток есть клеточная стенка, хлоропласты и другие специализированные пластиды, а также большая центральная вакуоль, тогда как у животных клеток нет.
Центросома
Центросома — это центр организации микротрубочек, расположенный рядом с ядрами клеток животных.Он содержит пару центриолей, две структуры, которые лежат перпендикулярно друг другу. Каждая центриоль представляет собой цилиндр из девяти троек микротрубочек. Центросома (органелла, где берут начало все микротрубочки) реплицируется до деления клетки, и центриоли, по-видимому, играют определенную роль в притяжении дублированных хромосом к противоположным концам делящейся клетки. Однако точная функция центриолей в делении клеток не ясна, потому что клетки, у которых была удалена центросома, все еще могут делиться; и клетки растений, в которых отсутствуют центросомы, способны к делению клеток.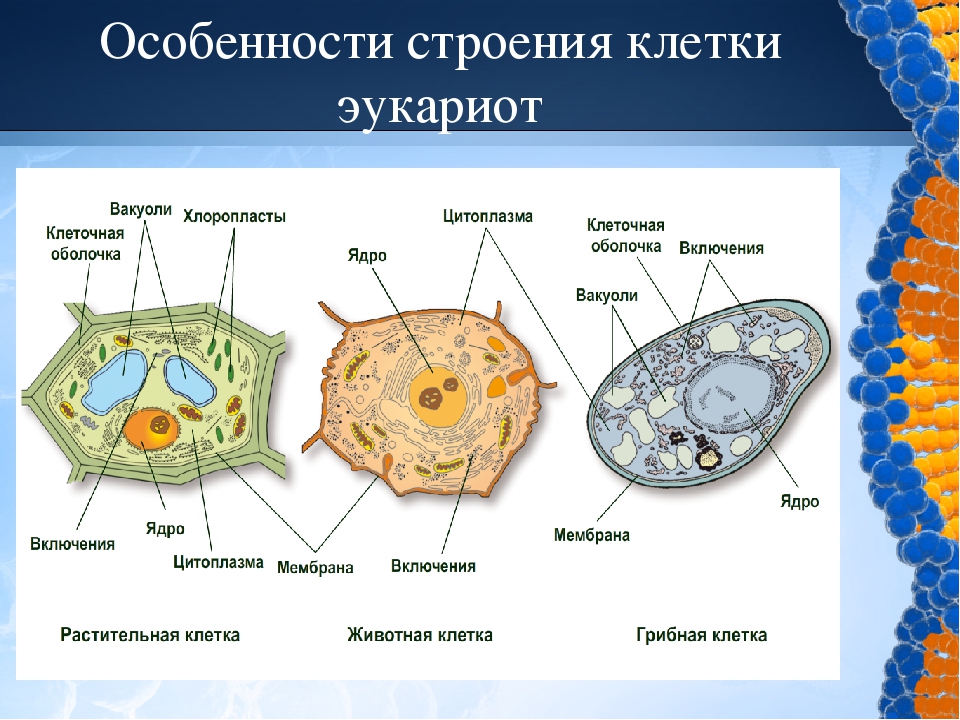
Структура центросомы : Центросома состоит из двух центриолей, которые расположены под прямым углом друг к другу. Каждая центриоль представляет собой цилиндр, состоящий из девяти троек микротрубочек. Белки нонтубулина (обозначенные зелеными линиями) удерживают вместе триплеты микротрубочек.
Лизосомы
В клетках животных есть еще один набор органелл, которых нет в клетках растений: лизосомы. Лизосомы — это «мусоропровод» клетки. В растительных клетках пищеварительные процессы происходят в вакуолях.Ферменты в лизосомах способствуют расщеплению белков, полисахаридов, липидов, нуклеиновых кислот и даже изношенных органелл. Эти ферменты активны при гораздо более низком pH, чем у цитоплазмы. Следовательно, pH в лизосомах более кислый, чем pH цитоплазмы. Многие реакции, происходящие в цитоплазме, не могут происходить при низком pH, поэтому преимущество разделения эукариотической клетки на органеллы очевидно.
Стенка клетки
Стенка ячейки — это жесткое покрытие, которое защищает ячейку, обеспечивает структурную поддержку и придает форму ячейке. Грибковые и протистанские клетки также имеют клеточные стенки. В то время как основным компонентом стенок прокариотических клеток является пептидогликан, основной органической молекулой в стенке растительной клетки является целлюлоза, полисахарид, состоящий из единиц глюкозы. Когда вы надкусываете сырые овощи, например сельдерей, они хрустят. Это потому, что вы зубами разрываете жесткие клеточные стенки клеток сельдерея.
Грибковые и протистанские клетки также имеют клеточные стенки. В то время как основным компонентом стенок прокариотических клеток является пептидогликан, основной органической молекулой в стенке растительной клетки является целлюлоза, полисахарид, состоящий из единиц глюкозы. Когда вы надкусываете сырые овощи, например сельдерей, они хрустят. Это потому, что вы зубами разрываете жесткие клеточные стенки клеток сельдерея.
Хлоропласты
Подобно митохондриям, хлоропласты имеют собственную ДНК и рибосомы, но хлоропласты выполняют совершенно другую функцию. Хлоропласты — это органеллы растительной клетки, осуществляющие фотосинтез. Фотосинтез — это серия реакций, в которых для производства глюкозы и кислорода используются углекислый газ, вода и световая энергия. Это главное различие между растениями и животными; растения (автотрофы) могут производить себе пищу, как сахар, в то время как животные (гетеротрофы) должны принимать их пищу.
Это главное различие между растениями и животными; растения (автотрофы) могут производить себе пищу, как сахар, в то время как животные (гетеротрофы) должны принимать их пищу.
Подобно митохондриям, хлоропласты имеют внешнюю и внутреннюю мембраны, но внутри пространства, ограниченного внутренней мембраной хлоропласта, находится набор взаимосвязанных и уложенных друг в друга заполненных жидкостью мембранных мешочков, называемых тилакоидами. Каждый стек тилакоидов называется гранумом (множественное число = грана). Жидкость, заключенная во внутренней мембране, окружающей грану, называется стромой.
Рисунок: Структура хлоропласта : Хлоропласт имеет внешнюю мембрану, внутреннюю мембрану и мембранные структуры, называемые тилакоидами, которые сложены в грану.Пространство внутри тилакоидных мембран называется тилакоидным пространством. Реакции сбора света происходят в тилакоидных мембранах, а синтез сахара происходит в жидкости внутри внутренней мембраны, которая называется стромой.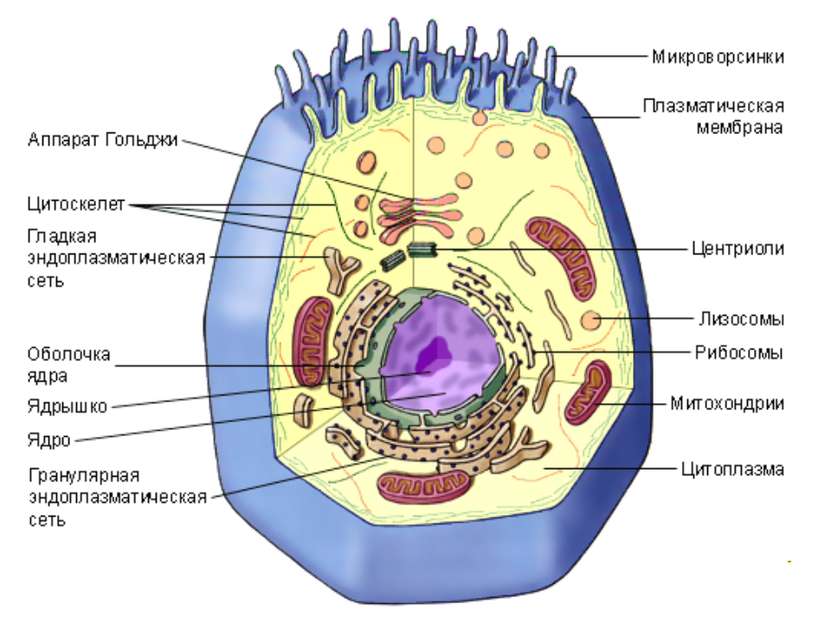
Хлоропласты содержат зеленый пигмент под названием хлорофилл, который улавливает световую энергию, которая запускает реакции фотосинтеза. Как и в растительных клетках, у фотосинтезирующих протистов есть хлоропласты. Некоторые бактерии осуществляют фотосинтез, но их хлорофилл не относится к органеллам.
Центральная вакуоль
Центральная вакуоль играет ключевую роль в регулировании концентрации воды в клетках при изменении условий окружающей среды. Если вы на несколько дней забываете полить растение, оно увядает. Это связано с тем, что когда концентрация воды в почве становится ниже, чем концентрация воды в растении, вода выходит из центральных вакуолей и цитоплазмы. По мере того как центральная вакуоль сжимается, она оставляет клеточную стенку без поддержки. Эта потеря поддержки клеточных стенок растительных клеток приводит к увяданию растения.Центральная вакуоль также поддерживает расширение клетки. Когда центральная вакуоль содержит больше воды, клетка становится больше, не тратя много энергии на синтез новой цитоплазмы.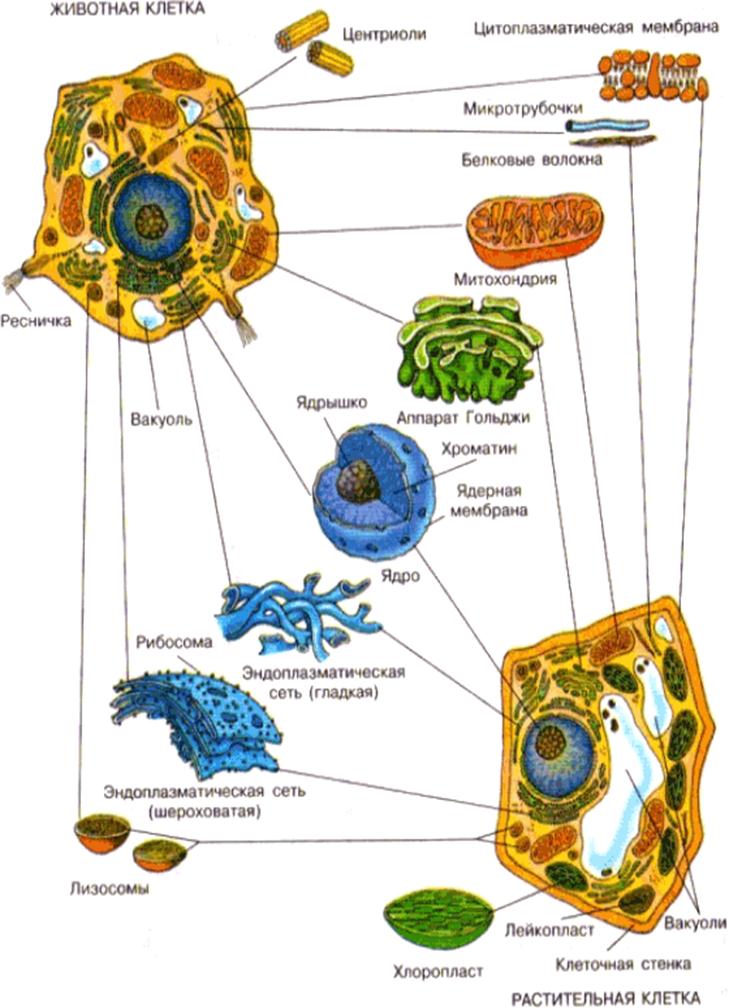
Структуры и функции растительных клеток
Клетка была впервые обнаружена в 1665 году английским ученым по имени Роберт Гук. Глядя в микроскоп, он наблюдал крошечные объекты, похожие на коробки в срезе пробки (кора дуба), и назвал эти коробки ячейками .Клетки — это основные единицы жизни, из которых состоит все живое. Эта идея лежит в основе Cell Theory .
Теория клеток
Три основных части теории клетки:
- Все живое состоит из клеток.
- Клетка — это основная единица структуры и функций всего живого.
- Клетки происходят только из других ранее существовавших клеток путем деления клеток .
В то время как некоторые организмы одноклеточные, другие состоят из множества клеток. Эти организмы называются многоклеточными (имеющими много клеток). Ячейки различаются по размеру и сложности.
Ячейки различаются по размеру и сложности.
Эукариоты — это организмы, состоящие из больших и сложных клеток, тогда как прокариоты — это организмы, состоящие из маленьких и простых клеток. Животные и растения являются примерами эукариот (имеют эукариотических клеток, ), тогда как бактерии являются примерами прокариот (имеют прокариотических клеток, ).
Структура и функции растительной клетки
Несмотря на различия в размере и сложности, все клетки в основном состоят из одних и тех же веществ, и все они выполняют схожие жизненные функции. К ним относятся рост, метаболизм и размножение путем деления клеток.
Клетки состоят из субклеточных структур, которые отвечают за различные и специфические функции. Эти структуры известны как органеллы . Некоторые из этих органелл являются общими как для животных, так и для растительных клеток.В этом разделе мы остановимся на тех частях, которые есть у растений.
- Стенка клетки: Это жесткий внешний слой клетки растения. Это делает ячейку жесткой, обеспечивая ячейку механической опорой и обеспечивая ей защиту. Клетки животных не имеют клеточных стенок.
- Клеточная мембрана: Это защитный слой, который окружает каждую клетку и отделяет ее от внешней среды.Он находится внутри клеточной стенки и состоит из комплекса липидов, (жиры) и белков, .
- Цитоплазма: Цитоплазма представляет собой густой водный раствор (на водной основе), в котором находятся органеллы. Такие вещества, как соли, питательные вещества, минералы и ферменты (молекулы, участвующие в метаболизме) растворяются в цитоплазме.
- Ядро: Ядро является «центром управления» клетки. Он содержит дезоксирибонуклеиновую кислоту (ДНК) , генетический материал, который управляет всей деятельностью клетки.Только эукариотические клетки имеют ядер (множественное число для ядер ), прокариотические клетки нет. Ядро отделено от цитоплазмы специальной мембраной под названием .
- Ядерная мембрана .
- Рибосомы: Это маленькие круглые структуры, которые производят белки. Они находятся в цитоплазме или прикрепляются к эндоплазматической сети.
- Эндоплазматический ретикулум (ER): ER — это мембранная система складчатых мешочков и туннелей.ER помогает перемещать белки внутри клетки, а также экспортировать их за пределы клетки. Существует два типа эндоплазматического ретикулума.
- Эндоплазматическая сеть шероховатая . Шероховатая эндоплазматическая сеть покрыта рибосомами.
- Гладкая эндоплазматическая сеть (без рибосом)
- Тело Гольджи: Тело Гольджи представляет собой набор покрытых мембраной мешочков, которые подготавливают белки для экспорта из клетки.
- Митохондрия (множественное число митохондрий ): это «электростанция» клетки.Он преобразует энергию, хранящуюся в пище (сахар и жир), в богатые энергией молекулы, которые клетка может использовать ( Аденозинтрифосфат — ATP для краткости).
- Лизосома: Лизосома — это пищеварительный центр клетки, который вырабатывает множество различных типов ферментов, которые способны расщеплять частицы пищи и перерабатывать изношенные компоненты клетки.
- Vacuoles: Это большие отсеки с мембраной, в которых хранятся токсичные отходы, а также полезные продукты, такие как вода.В основном они содержатся в растениях.
- Хлоропласт: Хлоропласты содержат зеленый пигмент, который улавливает солнечный свет и превращает его в сахара в процессе, называемом фотосинтезом. Сахар является источником энергии для растений и животных, которые их едят.
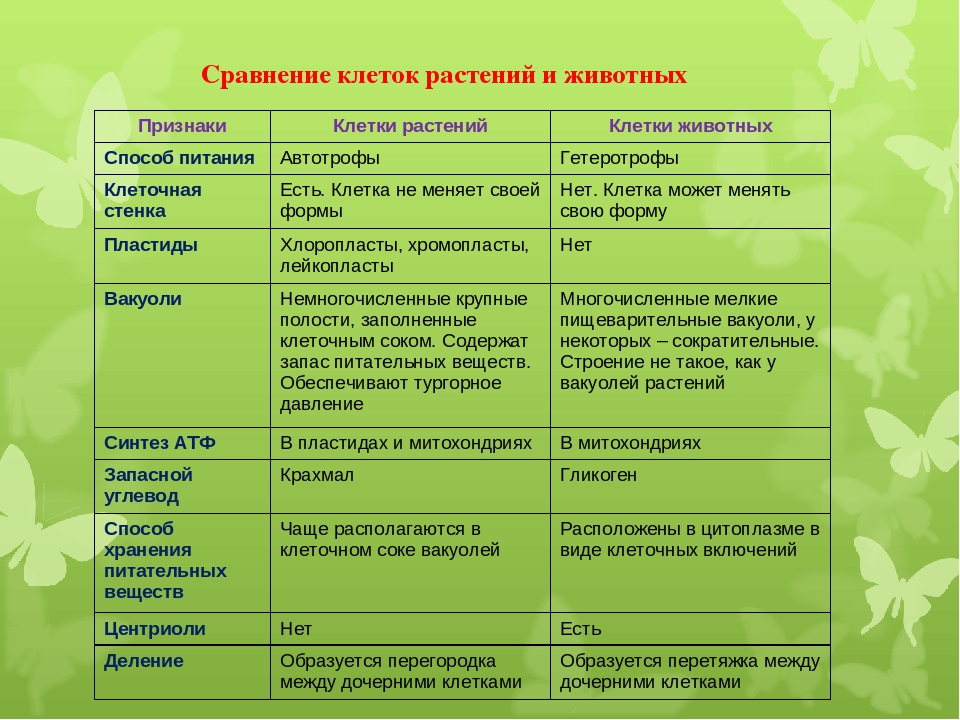

 Сахар является источником энергии для растений и животных, которые их едят.
Сахар является источником энергии для растений и животных, которые их едят.Что делает клетки растений уникальными
- Растительные клетки имеют клеточную стенку.
Растительные клетки отличаются от клеток животных во многих отношениях. Пожалуй, наиболее очевидным отличием является наличие клеточной стенки.Клеточная стенка обеспечивает растению силу и поддержку, так же как экзоскелет насекомого или паука (наш скелет находится внутри нашего тела, а не снаружи, как насекомые или пауки).
Стенка растительной клетки в основном состоит из углеводов, молекул, , целлюлозы, и лигнина , . Целлюлоза широко используется людьми для изготовления бумаги. Целлюлозу также можно превратить в целлюлозный этанол , разновидность биотоплива .Некоторые животные, такие как коровы, овцы и козы, могут переваривать целлюлозу с помощью бактерий в желудке. Люди не могут переваривать целлюлозу, которая проходит через наш организм и более известна как пищевые волокна, то есть то, что мы должны есть, чтобы наши отходы двигались должным образом! Лигнин заполняет промежутки между целлюлозой и другими молекулами в клеточной стенке. Лигнин также помогает молекулам воды перемещаться с одной стороны клеточной стенки на другую — важную функцию у растений.
Люди не могут переваривать целлюлозу, которая проходит через наш организм и более известна как пищевые волокна, то есть то, что мы должны есть, чтобы наши отходы двигались должным образом! Лигнин заполняет промежутки между целлюлозой и другими молекулами в клеточной стенке. Лигнин также помогает молекулам воды перемещаться с одной стороны клеточной стенки на другую — важную функцию у растений.
- Растительные клетки содержат вакуоли.
Большинство клеток взрослых растений имеют одну большую вакуоль, которая занимает более 30% объема клетки. В определенное время и в определенных условиях вакуоль занимает до 80% объема клетки! Помимо хранения отходов и воды, вакуоль также помогает поддерживать ячейку, потому что жидкость внутри вакуоли оказывает внешнее давление на ячейку, подобно воде внутри водяного баллона. Это называется тургорным давлением и препятствует разрушению клеток внутрь.
- Растительные клетки содержат хлоропласты.

В отличие от клеток животных, клетки растений могут использовать энергию Солнца, хранить ее в химических связях сахара, а затем использовать эту энергию. Органелла, отвечающая за это, — хлоропласт. Хлоропласты содержат хлорофилла , зеленый пигмент, придающий листьям цвет и поглощающий световую энергию. Цианобактерии , разновидность прокариотов, способных к фотосинтезу, считаются предками хлоропластов!
Хлоропласты (Источник: Kristian Peters -Fabelfroh [CC BY-SA 3.0] через Wikimedia Commons).Знаете ли вы?
Красные водоросли (многоклеточные морские водоросли) имеют хлоропласты, содержащие пигмент фикобилин, а не хлорофилл, который придает им красноватый, а не зеленый цвет.
Клетки растений и животных также имеют много общих органелл, включая ядро, клеточную мембрану (называемую плазматической мембраной , у животных), эндоплазматический ретикулум, митохондрии и цитоплазму, а также некоторые другие.
Характеристика липидов клеточной стенки из патогенной фазы Paracoccidioides brasiliensis, культивируемых в присутствии или отсутствии плазмы человека
Аннотация
Фон
Клеточная стенка гриба представляет собой сложную и динамичную внешнюю структуру.У патогенных грибов его компоненты взаимодействуют с хозяином, определяя судьбу инфекции. Настоящая работа направлена на характеристику липидов клеточной стенки из P. brasiliensis , выращенных в присутствии и в отсутствие плазмы человека. Мы сравнили результаты изолятов Pb3 и Pb18, которые представляют разные филогенетические виды, вызывающие различные паттерны экспериментального паракокцидиоидомикоза.
Методология / основные результаты
Мы сравнительно охарактеризовали фосфолипиды клеточной стенки, жирные кислоты, стерины и нейтральные гликолипиды, используя как электрораспылительную ионизацию, так и газовую хроматографию-масс-спектрометрический анализ липидов, экстрагированных органическими растворителями с последующим фракционированием в силикагеле-60. Мы обнаружили 49 видов фосфолипидов в Pb3 и 38 в Pb18, включая фосфатидилхолин, фосфатидилэтаноламин, фосфатидилсерин, фосфатидилглицерин, фосфатидилинозитол и фосфатидную кислоту. Как в Pb3, так и в Pb18 наиболее многочисленными видами были ПК и ПЭ. Среди жирных кислот C18À1 и C18À2 были наиболее распространенными видами в обоих изолятах, хотя C18À2 был более распространен в Pb18. Эффект от добавок плазмы на жирные кислоты различался в зависимости от грибкового изолята. Преобладающим видом гликолипидов был Hex-C18∶0-OH / d19∶2-Cer, хотя были обнаружены также четыре других второстепенных вида.Самым распространенным стеролом во всех образцах был брассикастерин. Отчетливые профили клеточной стенки и общего количества стеролов дрожжей предполагали, что препараты были обогащены компонентами клеточной стенки. Присутствие плазмы в культуральной среде особенно увеличивало содержание брассикастерина в клеточной стенке, а также других липидов.
Мы обнаружили 49 видов фосфолипидов в Pb3 и 38 в Pb18, включая фосфатидилхолин, фосфатидилэтаноламин, фосфатидилсерин, фосфатидилглицерин, фосфатидилинозитол и фосфатидную кислоту. Как в Pb3, так и в Pb18 наиболее многочисленными видами были ПК и ПЭ. Среди жирных кислот C18À1 и C18À2 были наиболее распространенными видами в обоих изолятах, хотя C18À2 был более распространен в Pb18. Эффект от добавок плазмы на жирные кислоты различался в зависимости от грибкового изолята. Преобладающим видом гликолипидов был Hex-C18∶0-OH / d19∶2-Cer, хотя были обнаружены также четыре других второстепенных вида.Самым распространенным стеролом во всех образцах был брассикастерин. Отчетливые профили клеточной стенки и общего количества стеролов дрожжей предполагали, что препараты были обогащены компонентами клеточной стенки. Присутствие плазмы в культуральной среде особенно увеличивало содержание брассикастерина в клеточной стенке, а также других липидов.
Выводы / Значение
Здесь представлен оригинальный сравнительный липидомный анализ клеточной стенки P. brasiliensis . Наши результаты открывают двери для понимания роли липидов клеточной стенки в биологии грибов и взаимодействия с противогрибковыми препаратами и хозяином.
brasiliensis . Наши результаты открывают двери для понимания роли липидов клеточной стенки в биологии грибов и взаимодействия с противогрибковыми препаратами и хозяином.
Образец цитирования: Longo LVG, Nakayasu ES, Gazos-Lopes F, Vallejo MC, Matsuo AL, Almeida IC, et al. (2013) Характеристика липидов клеточной стенки из патогенной фазы Paracoccidioides brasiliensis , культивируемых в присутствии или в отсутствии плазмы человека. PLoS ONE 8 (5): e63372. https://doi.org/10.1371/journal.pone.0063372
Редактор: Оскар Сарагоса, Instituto de Salud Carlos III, Испания
Поступила: 15 февраля 2013 г .; Дата принятия: 2 апреля 2013 г .; Опубликован: 17 мая 2013 г.
Авторские права: © 2013 Longo et al.Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.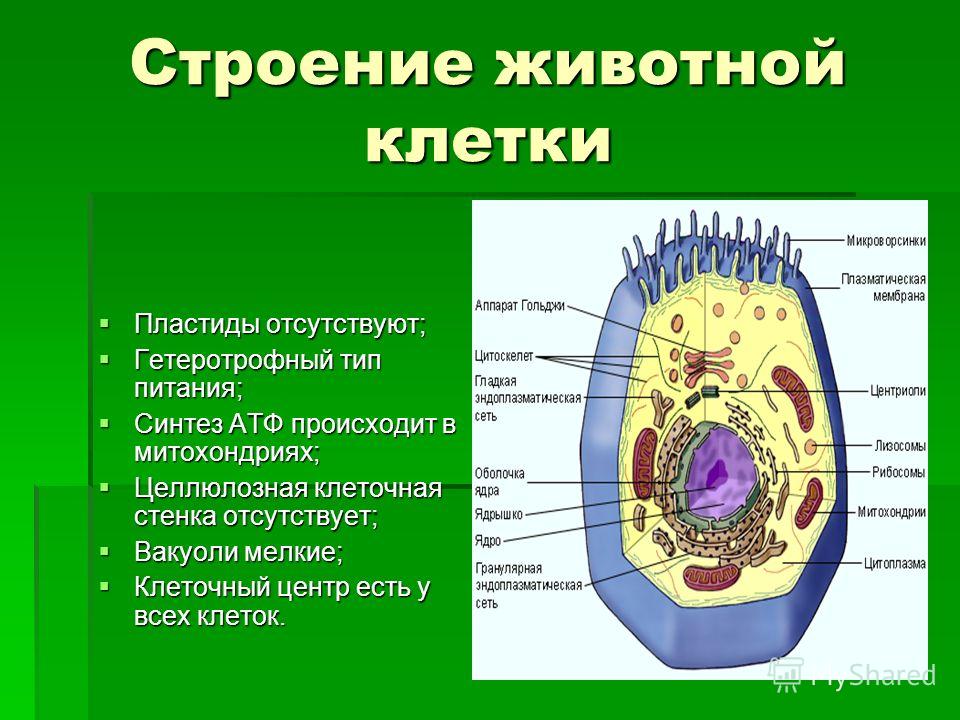
Финансирование: Финансовая поддержка была предоставлена FAPESP, CNPq и NIH (гранты № G12MD007592, 5G12RR008124-16A1, 5G12RR008124-16A1S1 и). Мы благодарны основной лаборатории анализа биомолекул BBRC / Biology / UTEP (NIH предоставляет G12MD007592, 5G12RR008124-16A1 и 5G12RR008124-16A1S1) за доступ к приборам ESI-MS и GC-MS.Финансирующие организации не играли никакой роли в дизайне исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи.
Конкурирующие интересы: Авторы заявили, что конкурирующих интересов не существует.
Введение
Клеточная стенка представляет собой сложную и динамическую структуру грибов, участвующую в таких функциях, как поддержание формы клетки, защита от механических повреждений, адаптация к морфогенезу и противостояние осмотическому давлению [1].Это основная структура, контактирующая с хозяином, поэтому она напрямую участвует во взаимоотношениях между хозяином и грибком [2].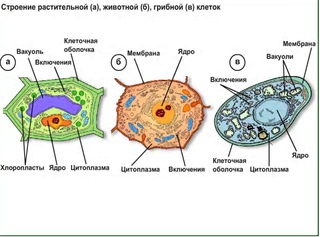 Его основные составляющие отсутствуют в клетках млекопитающих, что делает их потенциальными фармакологическими мишенями [3]. Основными составляющими клеточной стенки грибов являются структурные полисахариды, в частности глюканы и хитин, но также были описаны внешний слой маннана и несколько гликопротеинов [3].
Его основные составляющие отсутствуют в клетках млекопитающих, что делает их потенциальными фармакологическими мишенями [3]. Основными составляющими клеточной стенки грибов являются структурные полисахариды, в частности глюканы и хитин, но также были описаны внешний слой маннана и несколько гликопротеинов [3].
Диморфные грибы составляют небольшую группу патогенов, которые развиваются как дрожжи или мицелий в зависимости от температуры роста [4].В этой группе содержание липидов в клеточной стенке варьирует в зависимости от вида и / или морфологической фазы гриба, в пределах от 5,5% у патогенных дрожжей Blastomyces dermatitidis до 26% в гифах окружающей среды Sporothrix schenckii [5]. Роль липидов в клеточной стенке грибов до конца не выяснена. Наиболее охарактеризованным липидным видом клеточной стенки условно-патогенных клеток Candida albicans является фосфолипоманнан, который участвует в устойчивости грибов к агентам, нарушающим клеточную стенку, и в патогенности [6]. Глюкозилцерамид (GlcCer) уже идентифицирован в условно-патогенной клеточной стенке Cryptococcus neoformans , участвуя в почковании, росте и патогенности [7], [8].
Глюкозилцерамид (GlcCer) уже идентифицирован в условно-патогенной клеточной стенке Cryptococcus neoformans , участвуя в почковании, росте и патогенности [7], [8].
Наша модель гриба — это термодиморфный гриб Paracoccidioides brasiliensis , вызывающий паракокцидиоидомикоз (ПКМ), распространенный системный гранулематозный микоз в Латинской Америке [9]. P. brasiliensis развивается как гифы в окружающей среде и как многопучковые дрожжи в организме хозяина.Заражение начинается с вдыхания конидий окружающей среды, тогда как исход активного заболевания и тяжесть клинических форм будут зависеть от вирулентности штамма и типа иммунного ответа, который он вызывает [10]. Канетсуна с соавторами [11] изучили химический состав тщательно изолированных препаратов клеточной стенки из дрожжей и мицелия P. brasiliensis и обнаружили соотношение липидов в обеих морфологических формах от 5 до 11%. У дрожжевых мутантов липиды концентрируются во фракции клеточной стенки, растворимой в основных / кислотных растворах, что составляет до 80% ее содержания [12], [13].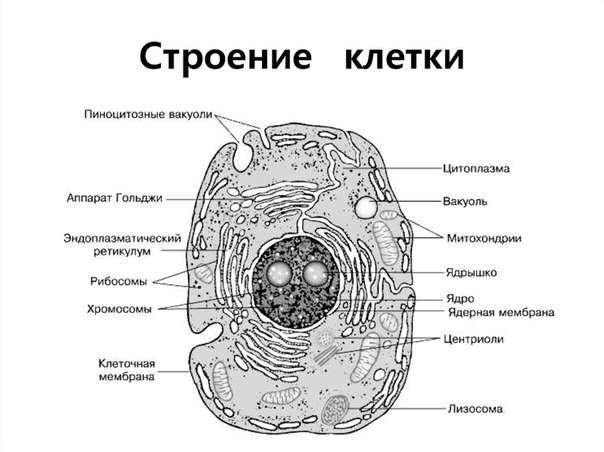 Природа липидов клеточной стенки P. brasiliensis , однако, была плохо изучена, хотя некоторые, возможно, поверхностные гликосфинголипиды [14] были охарактеризованы и показали, что они важны для роста и дифференцировки грибов [15].
Природа липидов клеточной стенки P. brasiliensis , однако, была плохо изучена, хотя некоторые, возможно, поверхностные гликосфинголипиды [14] были охарактеризованы и показали, что они важны для роста и дифференцировки грибов [15].
Целью настоящей работы было использование анализа ионизации электрораспылением (ESI-) и газовой хроматографии-масс-спектрометрии (ГХ-МС) для характеристики липидов клеточной стенки дрожжевой формы из двух филогенетически различных P. brasiliensis Pb3 и Pb18 изолирует [16].Мы также исследовали влияние плазмы человека на липидный состав клеточной стенки. Мы смогли сравнительно охарактеризовать фосфолипиды, жирные кислоты, стерины и нейтральные гликолипиды.
Материалы и методы
Реагенты
Иначе указано, все реагенты, использованные в этом исследовании, были чистыми для молекулярной биологии, ВЭЖХ или масс-спектрометрии от Sigma-Aldrich (Сент-Луис, Миссури).
P. brasiliensis и условия роста Изолирует Pb3 и Pb18 из P. brasiliensis (описанный в [17]) поддерживали при 36 ° С в твердой модифицированной среде YPD (0,5% дрожжевой экстракт, 0,5% казеиновый пептон, 1,5% глюкоза, pH 6,5). Для выделения клеточной стенки дрожжевые клетки переносили из 7-дневного твердого модифицированного YPD в 200 мл среды Ham’s F12 (Invitrogen) с добавлением 1,5% глюкозы (F12 / Glc) и с добавлением или без добавления 2% инактивированного нагреванием ( 56 ° C, один час) плазмы человека. Человеческая плазма, используемая для дополнения грибковых культур в исследовании, была предоставлена банком крови больницы Сан-Паулу (Hemocentro do Hospital São Paulo).Это было из пула образцов плазмы без идентификаторов, которые позволяют связать образцы с индивидуально идентифицируемыми живыми людьми или связанной с ними медицинской информацией. Таким образом, это исследование считается освобожденным от протокола человека в качестве субъектов, строго следуя руководящим принципам для субъектов-людей Федерального комитета по исследованиям в Сан-Паулу / больницы Сан-Паулу и Национальных институтов здравоохранения (Комитет по этике UNIFESP, номер протокола утверждения 0366/07).
brasiliensis (описанный в [17]) поддерживали при 36 ° С в твердой модифицированной среде YPD (0,5% дрожжевой экстракт, 0,5% казеиновый пептон, 1,5% глюкоза, pH 6,5). Для выделения клеточной стенки дрожжевые клетки переносили из 7-дневного твердого модифицированного YPD в 200 мл среды Ham’s F12 (Invitrogen) с добавлением 1,5% глюкозы (F12 / Glc) и с добавлением или без добавления 2% инактивированного нагреванием ( 56 ° C, один час) плазмы человека. Человеческая плазма, используемая для дополнения грибковых культур в исследовании, была предоставлена банком крови больницы Сан-Паулу (Hemocentro do Hospital São Paulo).Это было из пула образцов плазмы без идентификаторов, которые позволяют связать образцы с индивидуально идентифицируемыми живыми людьми или связанной с ними медицинской информацией. Таким образом, это исследование считается освобожденным от протокола человека в качестве субъектов, строго следуя руководящим принципам для субъектов-людей Федерального комитета по исследованиям в Сан-Паулу / больницы Сан-Паулу и Национальных институтов здравоохранения (Комитет по этике UNIFESP, номер протокола утверждения 0366/07).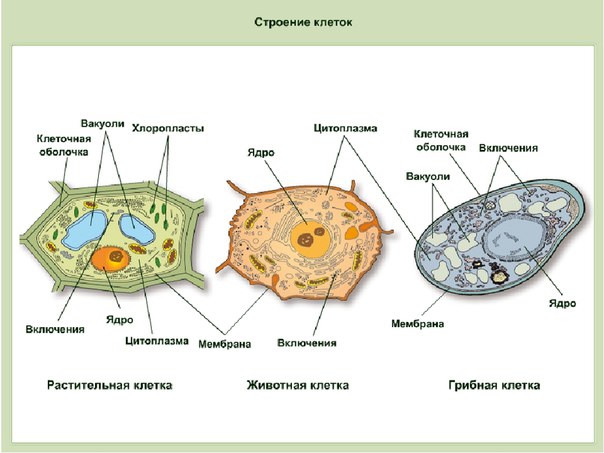 Предварительные инокуляты культивировали при встряхивании при 36 ° C в течение 4 дней, а дрожжевые клетки, собранные из четырех 200-мл колб, переносили в свежую среду (500 мл) и культивировали в течение 2 дней для очистки клеточной стенки.Клетки анализировали на жизнеспособность (> 95%) с помощью трипанового синего. Стоит отметить, что перед использованием среды с добавлением плазмы во время тестов на стерилизацию при 36 ° C наблюдали осаждение белков. Осадки дополнительно удаляли с помощью второго цикла фильтрации; Таким образом, конечная концентрация в плазме неизвестна.
Предварительные инокуляты культивировали при встряхивании при 36 ° C в течение 4 дней, а дрожжевые клетки, собранные из четырех 200-мл колб, переносили в свежую среду (500 мл) и культивировали в течение 2 дней для очистки клеточной стенки.Клетки анализировали на жизнеспособность (> 95%) с помощью трипанового синего. Стоит отметить, что перед использованием среды с добавлением плазмы во время тестов на стерилизацию при 36 ° C наблюдали осаждение белков. Осадки дополнительно удаляли с помощью второго цикла фильтрации; Таким образом, конечная концентрация в плазме неизвестна.
Очистка клеточной стенки
Дрожжи промывали 3 раза фосфатно-солевым буфером (PBS) и механически разрушали стеклянными шариками (425-600 мкм, Sigma Aldrich) в разрушителе клеток (B.Braun Biotech International GmBH, Melsungen, Germany) (6 раз по 10 мин, чередуя с 10 мин на льду). Обломки дрожжевых клеток, содержащие клеточную стенку, осаждали центрифугированием (5000 × g в течение 10 мин при 4 ° C) и промывали деионизированной водой. Клеточную стенку отделяли от цитоплазматического и мембранного содержимого тремя последовательными центрифугированиями (8000 × g в течение 45 мин при 4 ° C) в 85% сахарозе [11]. Затем фракции клеточной стенки промывали 50 раз ультрачистой деионизированной водой (Milli-Q, Millipore) и лиофилизировали.Previato et al. [18] установили этот протокол промывки клеточной стенки для Sporothrix schenckii и показали с помощью трансмиссионной электронной микроскопии, что плазматическая мембрана отсутствовала в конечных препаратах.
Клеточную стенку отделяли от цитоплазматического и мембранного содержимого тремя последовательными центрифугированиями (8000 × g в течение 45 мин при 4 ° C) в 85% сахарозе [11]. Затем фракции клеточной стенки промывали 50 раз ультрачистой деионизированной водой (Milli-Q, Millipore) и лиофилизировали.Previato et al. [18] установили этот протокол промывки клеточной стенки для Sporothrix schenckii и показали с помощью трансмиссионной электронной микроскопии, что плазматическая мембрана отсутствовала в конечных препаратах.
Экстракция и фракционирование липидов
Дрожжи и пул многочисленных препаратов клеточной стенки (100 мг) из Pb3 и Pb18 последовательно трижды экстрагировали 1 мл хлороформ: метанол (2∶1, об: об) и хлороформ: метанол: вода (2∶1∶0,8. , v: v: v) при встряхивании стеклянными шариками, как описано в Yichoy et al.[19]. Липиды из обеих экстракций смешивали и сушили в потоке N 2 . Фракционирование липидов проводили в пипетке Пастера, содержащей 500 мг сфер силикагеля-60 (60 Å, 200–400 мм, Sigma-Aldrich, Сент-Луис, Миссури). Колонку промывали последовательно 5 мл каждого метанола, ацетона и хлороформа для ВЭЖХ, и на нее загружали липиды, растворенные в 2 мл хлороформа. Стерины элюировали 5 мл хлороформа с последующим элюированием нейтральных гликолипидов 5 мл ацетона и фосфолипидов 5 мл метанола.Липидные фракции собирали и сушили в потоке N 2 . Данные по липидам получены из одной экспериментальной группы для каждого изолята, выращенного в двух параллельных условиях роста (с человеческой плазмой и без нее).
Колонку промывали последовательно 5 мл каждого метанола, ацетона и хлороформа для ВЭЖХ, и на нее загружали липиды, растворенные в 2 мл хлороформа. Стерины элюировали 5 мл хлороформа с последующим элюированием нейтральных гликолипидов 5 мл ацетона и фосфолипидов 5 мл метанола.Липидные фракции собирали и сушили в потоке N 2 . Данные по липидам получены из одной экспериментальной группы для каждого изолята, выращенного в двух параллельных условиях роста (с человеческой плазмой и без нее).
Перметилирование гликолипидов
Гликолипиды сушили в потоке N 2 и хранили в течение ночи при -20 ° C с осушающим кремнеземом, ресуспендировали в 150 мкл безводного диметилсульфоксида (ДМСО), перемешивали с несколькими миллиграммами порошкообразного безводного NaOH, смешанного с 80 мкл йодметана, и инкубируют при встряхивании 1 час при комнатной температуре.Затем добавляли деионизированную воду (2 мл) и дихлорметан (2 мл), смесь встряхивали и центрифугировали для удаления верхнего водного слоя. Органическую фазу трижды промывали 2 мл деионизированной воды и сушили в потоке N 2 .
Органическую фазу трижды промывали 2 мл деионизированной воды и сушили в потоке N 2 .
Метилирование жирных кислот
Метилирование жирных кислот проводили, как описано в [20]. Фосфолипиды инкубировали в 400 мкл 13 М гидроксида аммония (NH 4 OH): метанол (1-1, об.: Об.) В течение 1 ч при 37 ° C, сушили в потоке N 2 и дважды промывали безводным метанолом. .Образцы инкубировали в 400 мкл 0,5 М метанольной HCl (Supelco, Sigma-Aldrich) в течение 1 ч при 75 ° C и нейтрализовали 400 мкл 0,5 М NaCl. Затем образцы разделяли 1,5 мл деионизированной воды и 1,5 мл дихлорметана, и органическую фазу, содержащую жирные кислоты, дважды промывали деионизированной водой и концентрировали до 100 мкл в потоке N 2 .
ESI-MS / MS Анализ фосфолипидов и гликолипидов
Фосфолипиды и гликолипиды были проанализированы методом тандемной масс-спектрометрии с электрораспылением и ионизацией (ESI-MS / MS) на линейном масс-спектрометре с ионной ловушкой (LTQ XL, ThermoFisher Scientific, Сан-Хосе, Калифорния) в сочетании с автоматическим источником наноразмеров (Triversa NanoMate System, Адвион).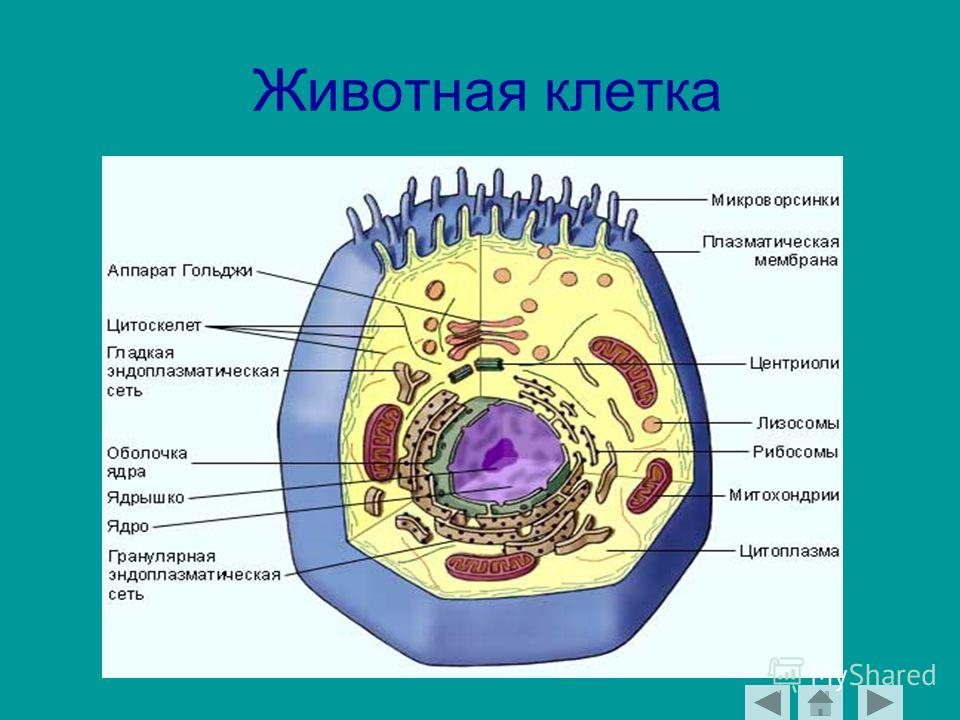 Фосфолипиды анализировали в режимах отрицательных и положительных ионов. Для анализа в режиме отрицательных ионов образцы повторно растворяли в метаноле, содержащем 0,05% муравьиной кислоты (FA) и 0,05% NH 4 OH; 2,5 мкМ фосфатидилглицерин (C12∶0 / C12∶0-PG) (Avanti Polar Lipids, Alabaster, AL) использовали в качестве внутреннего стандарта. В режиме положительных ионов образцы повторно растворяли в 10 мМ LiOH / метанол и 2,5 мкМ фосфатидилхолина (C11∶0 / C11∶0-PC) (Avanti Polar) использовали в качестве внутреннего стандарта. Спектры полного сканирования были получены в диапазоне 500–1000 m / z , а образцы подверглись картированию полных ионов (TIM) (2 a.м.у. ширина изоляции; диссоциация с импульсной добротностью (PQD) до 29% нормированной энергии столкновения; активация Q 0,7; и время активации 0,1 мс). Перметилированные гликолипиды растворяли в метаноле и анализировали, как описано для фосфолипидов, с получением спектров полного сканирования в диапазоне 500–2000 m / z и следующих настройках TIM: 700–900 диапазон m / z , 2 утра.
Фосфолипиды анализировали в режимах отрицательных и положительных ионов. Для анализа в режиме отрицательных ионов образцы повторно растворяли в метаноле, содержащем 0,05% муравьиной кислоты (FA) и 0,05% NH 4 OH; 2,5 мкМ фосфатидилглицерин (C12∶0 / C12∶0-PG) (Avanti Polar Lipids, Alabaster, AL) использовали в качестве внутреннего стандарта. В режиме положительных ионов образцы повторно растворяли в 10 мМ LiOH / метанол и 2,5 мкМ фосфатидилхолина (C11∶0 / C11∶0-PC) (Avanti Polar) использовали в качестве внутреннего стандарта. Спектры полного сканирования были получены в диапазоне 500–1000 m / z , а образцы подверглись картированию полных ионов (TIM) (2 a.м.у. ширина изоляции; диссоциация с импульсной добротностью (PQD) до 29% нормированной энергии столкновения; активация Q 0,7; и время активации 0,1 мс). Перметилированные гликолипиды растворяли в метаноле и анализировали, как описано для фосфолипидов, с получением спектров полного сканирования в диапазоне 500–2000 m / z и следующих настройках TIM: 700–900 диапазон m / z , 2 утра. ширина изоляции; PQD до 32% нормализованной энергии столкновения; активация Q 0,7; и время активации 0,1 мс. МС / МС спектры как фосфолипидов, так и гликолипидов анализировали вручную в соответствии с Pulfer and Murphy [21].
ширина изоляции; PQD до 32% нормализованной энергии столкновения; активация Q 0,7; и время активации 0,1 мс. МС / МС спектры как фосфолипидов, так и гликолипидов анализировали вручную в соответствии с Pulfer and Murphy [21].
ГХ-МС анализ стеринов
стеролов повторно растворяли в дихлорметане и анализировали методом газовой хроматографии-масс-спектрометрии (GC-MS, Trace GC Polaris Q, Thermo Fisher Scientific) с использованием колонки TR5-MS (30 м x 250 мкм x 0,25 мкм, Thermo Fisher Scientific). Температура инжектора поддерживалась на уровне 250 ° C. Температуру колонки первоначально поддерживали на уровне 170 ° C в течение 3 минут, а затем повышали со скоростью 20 ° C / мин до 280 ° C и выдерживали при этой температуре в течение 17 минут. Гелий использовался в качестве газа-носителя при расходе 1 л.2 мл / мин. Ионизацию проводили в режиме положительных ионов электронным ударом при 70 эВ и 200 ° C. Спектры были собраны в диапазоне 50–650 m / z и проанализированы путем поиска в спектральной библиотеке (библиотека NIST, доступная в Xcalibur 1.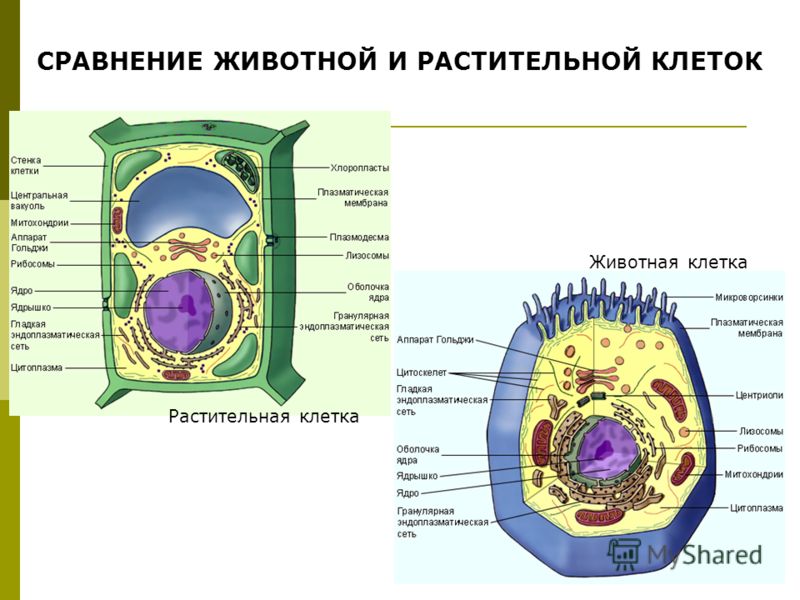 4 Srl, Thermo Fisher Scientific) и путем сравнения с внешними стандартами брассикастерина, эргостерина и ланостерина ( Avanti Polar Lipids) и холестерин (Sigma-Aldrich). Относительную количественную оценку проводили путем сравнения с внутренним стандартом стигмастерин (Avanti Polar Lipids).
4 Srl, Thermo Fisher Scientific) и путем сравнения с внешними стандартами брассикастерина, эргостерина и ланостерина ( Avanti Polar Lipids) и холестерин (Sigma-Aldrich). Относительную количественную оценку проводили путем сравнения с внутренним стандартом стигмастерин (Avanti Polar Lipids).
ГХ-МС анализ жирных кислот
Один микролитр метилированных жирных кислот в дихлорметане использовали для анализа ГХ-МС. Образцы разделяли на колонке SP-2380 (30 м × 250 мкм × 0,20 мкм, Supelco). Инжектор поддерживали при 200 ° C и использовали следующий температурный градиент: 70 ° C в течение 5 мин, приращение 4 ° C / мин до 140 ° C, 2 ° C / мин до 185 ° C и 185 ° C. ° C в течение 10 мин. В качестве газа-носителя использовали гелий со скоростью 1 мл / мин. Ионизация проводилась в режиме положительных ионов электронным ударом 70 эВ при 200 ° C.Спектры были собраны в диапазоне 30–400 m / z , и хроматограммы экстрагированных ионов были получены путем построения диагностических фрагментов-ионов при m / z 41, 43 и 55.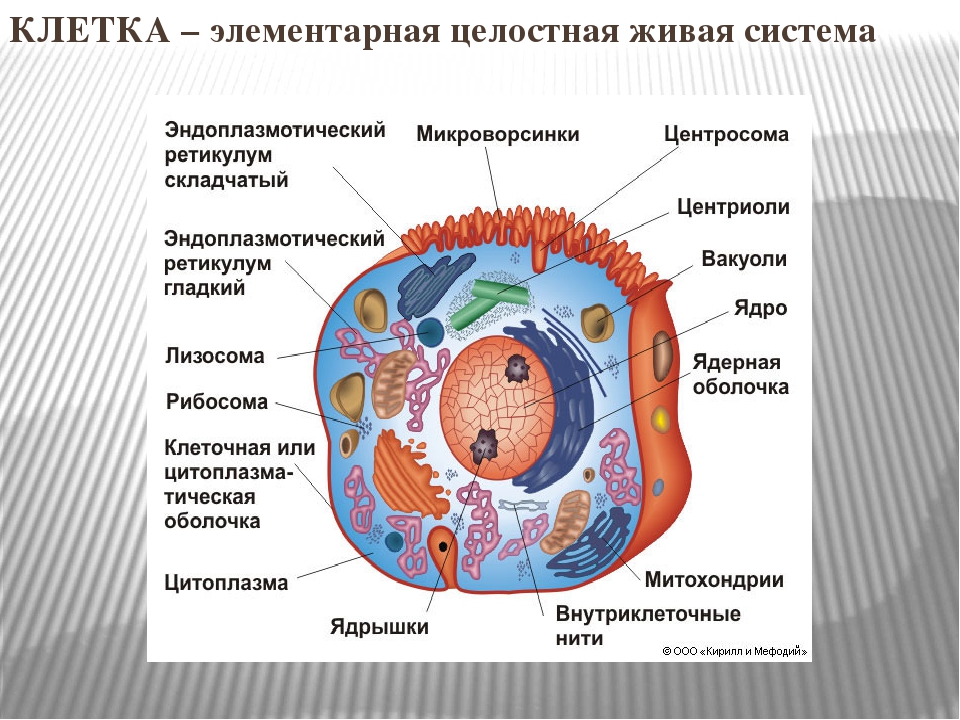 Идентификация проводилась с помощью жирных кислот. Сравнение видов с внешним стандартом смеси FAME 37 (Supelco).
Идентификация проводилась с помощью жирных кислот. Сравнение видов с внешним стандартом смеси FAME 37 (Supelco).
Результаты
P. brasiliensis липиды из тщательно выделенных препаратов клеточной стенки экстрагировали и фракционировали на колонке с диоксидом кремния-60 на фосфолипиды, нейтральные гликолипиды и стерины.Для сравнения анализировали общие липиды дрожжевых клеток. Мы сравнили результаты для Pb3 и Pb18, культивируемых в присутствии или в отсутствие плазмы человека. Эти изоляты были выбраны потому, что они представляют разные филогенетические виды [16], которые вызывают инфекцию с разными профилями у мышей B10.A [22].
ESI-MS / MS Анализ фосфолипидов клеточной стенки
Фосфолипиды из препаратов клеточной стенки Pb3 и Pb18 были ионизированы в режимах положительных (рис. 1A) и отрицательных ионов (рис. 1B) ESI-MS / MS.Спектры полного сканирования показали, что профили пиков были сходными для изолятов, выращенных в присутствии (pl) или в отсутствие плазмы, что позволяет предположить сходный состав фосфолипидов. Однако мы обнаружили различия в численности видов фосфолипидов при сравнении Pb18 и Pb3, выращенных в отсутствие плазмы, в то время как подробное изучение видов в образцах Pb18pl и Pb3pl не проводилось.
Однако мы обнаружили различия в численности видов фосфолипидов при сравнении Pb18 и Pb3, выращенных в отсутствие плазмы, в то время как подробное изучение видов в образцах Pb18pl и Pb3pl не проводилось.
Рис. 1. Спектры полного сканирования фосфолипидов клеточной стенки из Pb3 и Pb18, культивируемых в присутствии (pl) или в отсутствие плазмы.
Показан анализ ESI-MS в режимах (A) положительных и (B) отрицательных ионов. m / z , отношение массы к заряду. Указаны назначенные пики.
https://doi.org/10.1371/journal.pone.0063372.g001
Идентификация фосфолипидов проводилась путем поиска ионов диагностических фрагментов. В режиме положительных ионов были обнаружены разновидности фосфатидилхолина (PC) с аддуктом Na + или Li + или однократно протонированные (H + ) после потери нейтрали 59 Да триметиламина ((CH 3 ) 3 N или Me 3 N).Цепи жирных кислот идентифицировали по фрагментам с нейтральной потерей ([M — жирная кислота + Li + (или Na + )] + ) или по потере жирных кислот и Me 3 N ([M — жирная кислота — Me 3 + Li + (или Na + )] + ), как показано на спектре МС / МС основных частиц ПК (C18∶1 / C18∶1-PC) (рис. S1). В режиме отрицательных ионов диагностические фрагменты ионов при m / z 171, что соответствует глицерофосфату (GroP — H) — , и при m / z 153, соответствующему дегидратированному глицерофосфату (GroP — H 2 O — H) — были использованы для всех фосфолипидов, которые затем были идентифицированы с помощью аддуктов хлорида (Cl — ) или формиата (HCOO — ).Например, нейтральные потери 50 или 60 Да, связанные с N-метильной группой, связанной с Cl — [M — MeCl — H] — или формиатом [M — HCOOMe — H] — , соответственно , использовались при идентификации ПК (рис. S2). Кроме того, для идентификации каждого класса фосфолипидов использовались специфические диагностические ионы. Фосфатидилэтаноламин (PE) был идентифицирован по иону m / z 196, соответствующему дегидратированному глицерофосфоэтаноламину [GroPEtN-H 2 O-H] — (фиг.S3). Нейтральная потеря 87-Da сериновой головной группы [M — Ser — H] — и диагностического иона m / z 241, что соответствует дегидратированному фосфоинозиту [InsP — H 2 O — H] — , были использованы для идентификации видов фосфатидилсерина (PS) (рис.
S1). В режиме отрицательных ионов диагностические фрагменты ионов при m / z 171, что соответствует глицерофосфату (GroP — H) — , и при m / z 153, соответствующему дегидратированному глицерофосфату (GroP — H 2 O — H) — были использованы для всех фосфолипидов, которые затем были идентифицированы с помощью аддуктов хлорида (Cl — ) или формиата (HCOO — ).Например, нейтральные потери 50 или 60 Да, связанные с N-метильной группой, связанной с Cl — [M — MeCl — H] — или формиатом [M — HCOOMe — H] — , соответственно , использовались при идентификации ПК (рис. S2). Кроме того, для идентификации каждого класса фосфолипидов использовались специфические диагностические ионы. Фосфатидилэтаноламин (PE) был идентифицирован по иону m / z 196, соответствующему дегидратированному глицерофосфоэтаноламину [GroPEtN-H 2 O-H] — (фиг.S3). Нейтральная потеря 87-Da сериновой головной группы [M — Ser — H] — и диагностического иона m / z 241, что соответствует дегидратированному фосфоинозиту [InsP — H 2 O — H] — , были использованы для идентификации видов фосфатидилсерина (PS) (рис. S4) и фосфатидилинозитола (PI) соответственно (рис. S5). Поскольку нет никаких специфических нейтральных потерь или диагностических ионов, характерных для фосфатидилглицерина (PG) (рис. S6) и фосфатидной кислоты (PA) (рис.S7), их идентификация была достигнута с учетом молекулярной массы интактных родительских ионов. Жирные кислоты были идентифицированы по соотношению отрицательно заряженных карбоксилат-ионов m / z при m / z 255 (C16À0), 279 (C18À2) и 281 (C18À1) (рис. S6 и S7).
S4) и фосфатидилинозитола (PI) соответственно (рис. S5). Поскольку нет никаких специфических нейтральных потерь или диагностических ионов, характерных для фосфатидилглицерина (PG) (рис. S6) и фосфатидной кислоты (PA) (рис.S7), их идентификация была достигнута с учетом молекулярной массы интактных родительских ионов. Жирные кислоты были идентифицированы по соотношению отрицательно заряженных карбоксилат-ионов m / z при m / z 255 (C16À0), 279 (C18À2) и 281 (C18À1) (рис. S6 и S7).
Обнаруженные фосфолипиды показали ионов m / z между m / z 556,5 и 851,7 в режиме отрицательных ионов и между m / z 526,5 и 822,7 в режиме положительных ионов. Фосфолипиды классов PC, PE, PG, PI, PS и PA были идентифицированы как в Pb3, так и в Pb18 (таблица 1, рис.2). В целом мы идентифицировали больше видов фосфолипидов в Pb3 (63), чем в Pb18 (51), как показано в Таблице 1 и на Рис. 2. Как в Pb3, так и на рис. PA / PI и PG. Однако Pb18 показал больше видов PA (5), чем PS (4), тогда как PI (2) и PG (1) были представлены недостаточно. В обоих изолятах было лизо -C18∶1-PC и лизо -C18∶2-PC, но разновидности C18∶1-алкил / C18∶1-ацил-PE (плазмалоген или плазменилэтаноламин) были обнаружены только в Pb3 (таблица 1).
В обоих изолятах было лизо -C18∶1-PC и лизо -C18∶2-PC, но разновидности C18∶1-алкил / C18∶1-ацил-PE (плазмалоген или плазменилэтаноламин) были обнаружены только в Pb3 (таблица 1).
Рисунок 2. Количество видов фосфолипидов из каждого класса, идентифицированного в клеточной стенке Pb3 и Pb18, культивируемых в отсутствие плазмы.
Анализ ESI-MS / MS как для отрицательных, так и для положительных ионов. Подробное описание отдельных видов фосфолипидов можно найти в таблице 1.
https://doi.org/10.1371/journal.pone.0063372.g002
Оценка численности (таблица 1) была проведена путем нормализации интенсивности сигнала каждого вида фосфолипидов по внутренним стандартам C12∶0 / C12∶0-PG ( m / з 609.5) (в режиме отрицательных ионов) и C11∶0 / C11À0-PC ( m / z 600,5) (в режиме положительных ионов). В целом, количество видов между Pb3 и Pb18 было сопоставимым, но некоторые различия заслуживают внимания. Два вида ПК, C16À0 / C20À1-PC ( m / z 822,5) и C18À1 / C20À0-PC ( m / z 850,6), были в изобилии в Pb3, но не в Pb18.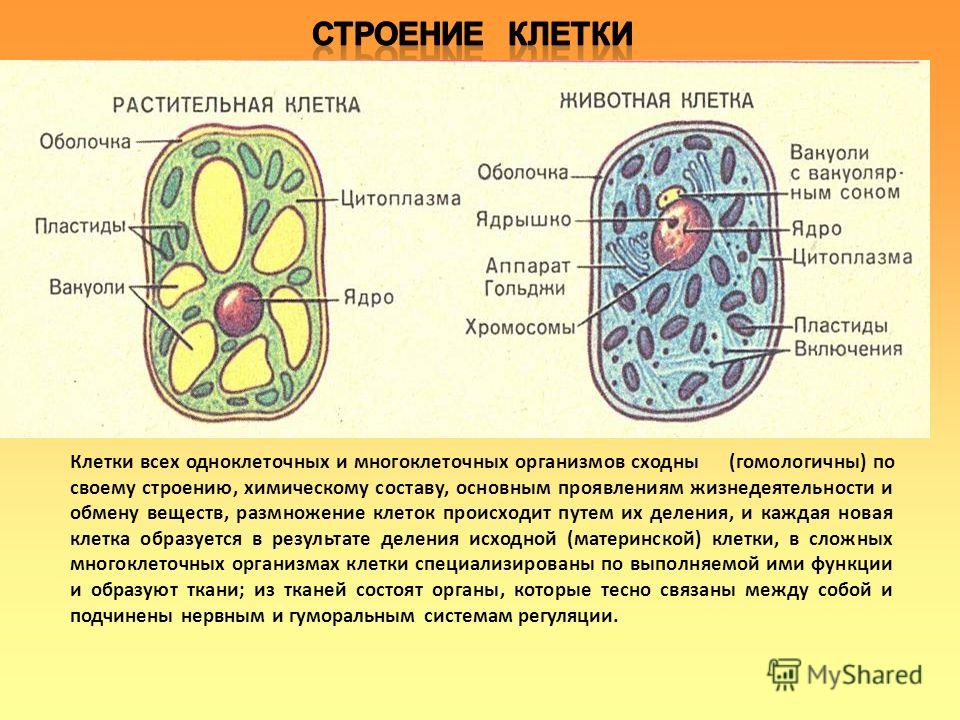 С другой стороны, C18À2 / C19À1-PC ( m / z 842,7) и C16À0 / C18À2-PA ( m / z 739,6) не были обнаружены в Pb3, но были в большом количестве в Pb18. клеточная стенка.
С другой стороны, C18À2 / C19À1-PC ( m / z 842,7) и C16À0 / C18À2-PA ( m / z 739,6) не были обнаружены в Pb3, но были в большом количестве в Pb18. клеточная стенка.
Анализ состава фосфолипидных жирных кислот клеточной стенки с помощью ГХ-МС
Жирные кислоты клеточной стенки и общие фосфолипиды дрожжевых клеток из Pb3 и Pb18, культивируемых в присутствии (pl) и в отсутствие плазмы, метилировали и анализировали с помощью ГХ-МС. Жирные кислоты были идентифицированы путем сравнения со смесью внешних стандартов, и было показано, что они имеют двойные связи в конфигурации цис . Хотя нам не удалось определить положение ненасыщенности, мы предположили, что это произошло в наиболее распространенных положениях, в частности, между C9 и C10 (Δ 9 ), в мононенасыщенных жирных кислотах или в обоих Δ 12 и Δ 15 [23].Относительное содержание жирных кислот оценивали путем нормализации к площади пика C16∶0.
Состав жирных кислот клеточной стенки был сходным во всех проанализированных образцах (рис.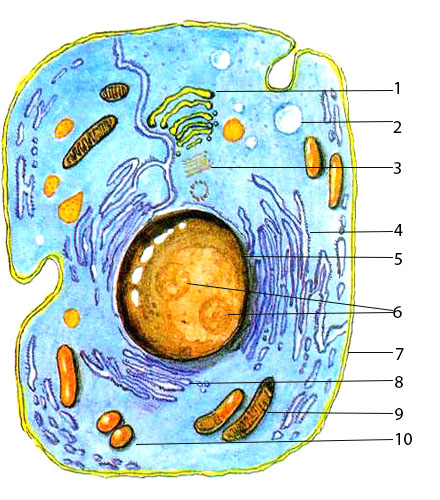 3): преобладали олеиновая кислота (C18∶1) и линолевая кислота (C18∶2), тогда как пальмитиновая кислота (C16∶0), стеариновая кислота (C18∶0). ) и пентадекановая кислота (C15∶0) были второстепенными компонентами. Однако наблюдались заметные различия в численности, особенно когда изоляты росли в присутствии плазмы. Хотя C18À1 был в равной степени представлен в обоих изолятах, C18À2 преобладал в Pb18.Более того, присутствие плазмы вызывало у Pb18 увеличение содержания обеих жирных кислот, в то время как они несколько снижались в Pb3.
3): преобладали олеиновая кислота (C18∶1) и линолевая кислота (C18∶2), тогда как пальмитиновая кислота (C16∶0), стеариновая кислота (C18∶0). ) и пентадекановая кислота (C15∶0) были второстепенными компонентами. Однако наблюдались заметные различия в численности, особенно когда изоляты росли в присутствии плазмы. Хотя C18À1 был в равной степени представлен в обоих изолятах, C18À2 преобладал в Pb18.Более того, присутствие плазмы вызывало у Pb18 увеличение содержания обеих жирных кислот, в то время как они несколько снижались в Pb3.
Рис. 3. ГХ-МС анализ фосфолипидных жирных кислот клеточной стенки из Pb3 и Pb18, культивируемых в присутствии (pl) или в отсутствие плазмы.
Содержание жирных кислот нормировано на площадь пика C16∶0.
https://doi.org/10.1371/journal.pone.0063372.g003
В совокупности анализ жирных кислот показал различия в содержании жирнокислотного состава клеточной стенки между Pb3 и Pb18, включая различное влияние плазмы в зависимости от изолята грибка. .
.
ГХ-МС анализ стеринов клеточной стенки
Фракциистеролов, элюированные из колонки с диоксидом кремния-60 с хлороформом, были проанализированы с помощью ГХ-МС, и виды были идентифицированы путем сравнения с внешними стандартами или поиска в эталонной спектральной библиотеке. Анализ относительной численности проводили путем нормализации по площади пика стигмастерола (внутреннего стандарта). Для сравнения также анализировали общие стеролы дрожжевых клеток.
Брассикастерин (эргоста-5,22-диен-3β-ол) был наиболее распространенным стеролом, идентифицированным в клеточной стенке Pb3 и Pb18, а также в общих липидах (рис.4А). В дрожжевых клетках, растущих в отсутствие плазмы, брассикастерин относительно больше содержал Pb18, чем препараты клеточной стенки Pb3. Однако в общих липидах из цельноклеточных препаратов доля Pb3 была примерно в 4 раза выше. Плазма более радикально изменила этот сценарий для брассикастерола клеточной стенки, содержание которого увеличилось как в Pb18, так и в Pb3, хотя в Pb3 он показал более выраженный эффект (в 11 раз против 2 раз в Pb18) (рис. 4A). Общий брассикастерин Pb3, напротив, снижался в присутствии плазмы, тогда как в Pb18 он несколько повышался (рис.4А).
4A). Общий брассикастерин Pb3, напротив, снижался в присутствии плазмы, тогда как в Pb18 он несколько повышался (рис.4А).
Рисунок 4. Сравнение клеточной стенки и общего количества стеринов дрожжевых клеток из Pb3 и Pb18, культивированных в присутствии (pl) или в отсутствие плазмы.
Стерины были проанализированы с помощью ГХ-МС, и их численность была нормализована по стигмастерину (внутренний стандарт). A, Сравнение количества брассикастерина, самого распространенного из обнаруженных стеролов. B — Поглощение холестерина из плазмы изолятами Pb3 и Pb18. C. Сравнение стеринов, обнаруженных в следовых количествах в образцах клеточной стенки.
https: // doi.org / 10.1371 / journal.pone.0063372.g004
Холестерин был идентифицирован только в образцах из P. brasiliensis , культивируемых в присутствии плазмы (рис. 4B), что указывает на то, что он был включен из среды во время роста грибов. В Pb3pl было включено в два раза больше холестерина, чем в клеточную стенку Pb18pl, но общий холестерин был в 4 раза выше в Pb18pl.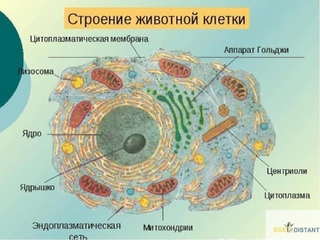
Эргостерол, эргоста-5,22-диен-3-ол ацетат, кампестерин, эргостанол, 17-альфа-ацетоксипрегненолон и 24-метилен-13-бета ланост-8-ен-3-ол также были обнаружены в небольших количествах, которые варьировалось в зависимости от изолята и условий роста (рис.4С). Интересно, что некоторые следы стеролов, по-видимому, в значительной степени индуцируются плазмой, особенно Pb3 (рис. 4C).
ESI-MS / MS Анализ гликолипидов клеточной стенки
Нейтральные гликолипиды из клеточной стенки Pb3 и Pb18 выделяли в колонке с диоксидом кремния-60, перметилировали и подвергали анализу ESI-MS / MS в режиме положительных ионов. На рисунке 5А показан один из основных видов гликолипидов, идентифицированный в спектрах полного сканирования, но четыре других были обнаружены только с помощью анализа МС / МС. Это связано с тем, что во время отбора родительских ионов в масс-спектрометре большая часть фона удаляется, что делает анализ МС / МС более чувствительным, чем сканирование полного МС.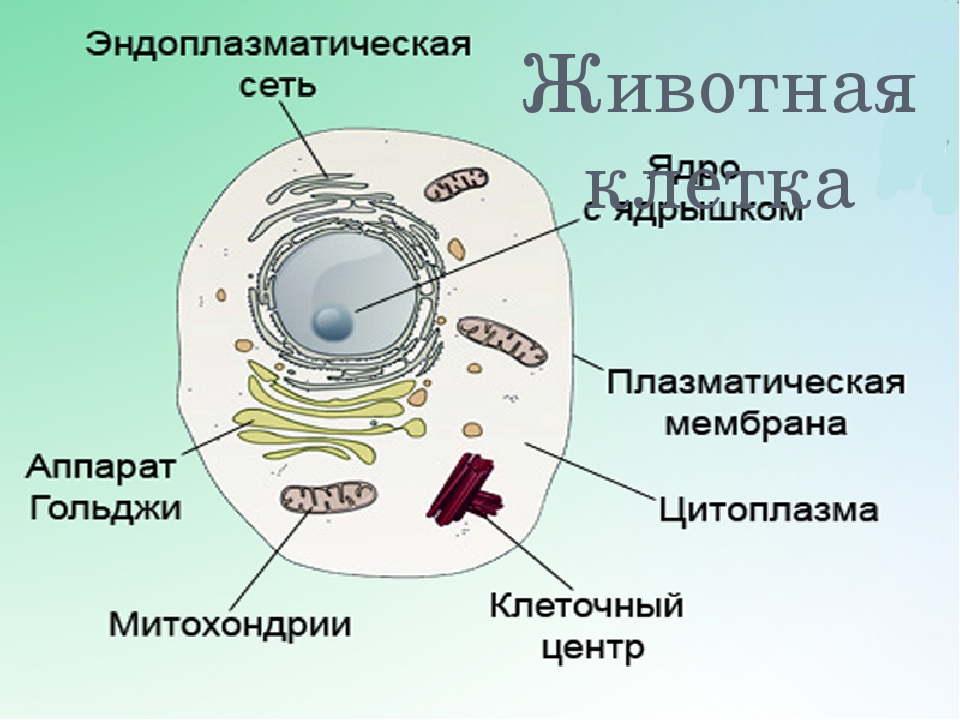 Согласно данным ручного анализа фрагментов тандемной масс-спектрометрии, наиболее распространенные виды во всех образцах были идентифицированы как Hex-C18∶0-OH / d19∶2-Cer (при m / z 876.9). Ионы диагностического фрагмента для гликолипидов, соответствующих метилированной гексозе, при m / z 227,1 [HexMe 4 — H 3 COH + Na] + и 259,2 [HexMe 4 + Na] + наблюдались. Нейтральные потери обоих фрагментов генерировали соответствующие фрагментные ионы при m / z 588.8 [M — HexMe 4 — H 3 COH + Na] + и m / z 618,8 [M — HexMe 4 + Na] + . Фрагмент перметилированного d19∶2 сфингоидного основания [d19∶2Me 2 — H 3 COH + H] + и жирная кислота C18∶0, присоединенная к части сфингоидного основания [азирин-h28∶0Me 2 — H 3 COH + H] + можно идентифицировать по фрагментным ионам m / z 290,4 и m / z 322,4 соответственно.
Согласно данным ручного анализа фрагментов тандемной масс-спектрометрии, наиболее распространенные виды во всех образцах были идентифицированы как Hex-C18∶0-OH / d19∶2-Cer (при m / z 876.9). Ионы диагностического фрагмента для гликолипидов, соответствующих метилированной гексозе, при m / z 227,1 [HexMe 4 — H 3 COH + Na] + и 259,2 [HexMe 4 + Na] + наблюдались. Нейтральные потери обоих фрагментов генерировали соответствующие фрагментные ионы при m / z 588.8 [M — HexMe 4 — H 3 COH + Na] + и m / z 618,8 [M — HexMe 4 + Na] + . Фрагмент перметилированного d19∶2 сфингоидного основания [d19∶2Me 2 — H 3 COH + H] + и жирная кислота C18∶0, присоединенная к части сфингоидного основания [азирин-h28∶0Me 2 — H 3 COH + H] + можно идентифицировать по фрагментным ионам m / z 290,4 и m / z 322,4 соответственно. Аналогичный анализ был проведен для характеристики минорных гликолипидов, идентифицированных как Hex-C16∶0-OH / d19∶2-Cer ( m / z 848.7), Hex-C18∶0-OH / d18∶2-Cer ( m / z 862,8), Hex-C18∶0-OH / d18À1-Cer ( m / z 864,6) и Hex-C18 ∶1-OH / d19∶2-Cer ( m / z 874,7) (рис. S8, S9, S10, S11).
Аналогичный анализ был проведен для характеристики минорных гликолипидов, идентифицированных как Hex-C16∶0-OH / d19∶2-Cer ( m / z 848.7), Hex-C18∶0-OH / d18∶2-Cer ( m / z 862,8), Hex-C18∶0-OH / d18À1-Cer ( m / z 864,6) и Hex-C18 ∶1-OH / d19∶2-Cer ( m / z 874,7) (рис. S8, S9, S10, S11).
Рисунок 5. ESI-MS анализ нейтральных гликолипидов клеточной стенки из Pb3 и Pb18, культивированных в присутствии (pl) или в отсутствие плазмы.
A, Спектры полного сканирования в режиме положительных ионов показывают идентифицированные гликолипиды Hex-C16∶0-OH / d19∶2-Cer ( m / z 848,7), Hex-C18∶0-OH / d18∶ 2-Cer ( м / з 862.8), Hex-C18À0-OH / d18À1-Cer ( m / z 864,9), Hex-C18À1-OH / d19À2-Cer ( m / z 874,9) и Hex- C18∶0-OH / d19∶2-Cer ( m / z 876,9). B, Тандем-МС-спектр основных идентифицированных видов гликолипидов ( m / z 876.9). m / z , отношение массы к заряду.
https://doi.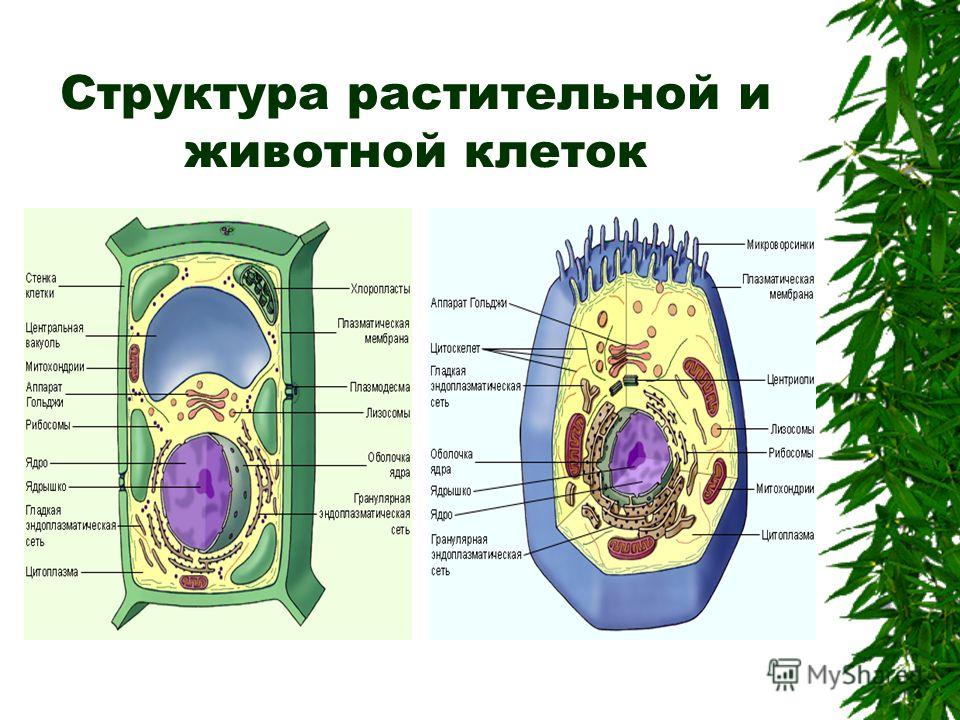 org/10.1371/journal.pone.0063372.g005
org/10.1371/journal.pone.0063372.g005
Хотя наши данные не позволили выяснить положение ненасыщенности или метильных и гидроксильных групп в основе сфингоида, Толедо и его коллеги [24] полностью охарактеризовали два вида гликосфинголипидов из дрожжей Pb18 и гиф с помощью ядерного магнитного резонанса (ЯМР) и обнаружили, что сфингоидное основание, вероятно, метилировано по C9, а ненасыщенность находится в конформации цис по C4 и C8.Следовательно, основным идентифицированным здесь нейтральным гликолипидом, вероятно, является N -2′-гидроксиоктадеканоат (4 E , 8 E ) -9-метил-4,8-сфингадиенин ( m / z 876,8).
Обсуждение
Результаты, представленные в этой работе, показывают липидный состав, включая фосфолипиды, жирные кислоты, стерины и нейтральные гликолипиды, препаратов клеточной стенки из дрожжевых клеток P. brasiliensis , сравнивая изоляты Pb3 и Pb18. Влияние плазмы на липидный состав клеточной стенки оценивали путем изучения препаратов дрожжей, культивируемых в присутствии или в отсутствие плазмы человека. Отчетливые профили клеточной стенки и общего количества стеролов дрожжей предполагали, что препараты были обогащены компонентами клеточной стенки.
Отчетливые профили клеточной стенки и общего количества стеролов дрожжей предполагали, что препараты были обогащены компонентами клеточной стенки.
Pb3 и Pb18 относятся к далеким филогенетическим группам P. brasiliensis и представляют собой виды PS2 и S1 соответственно [16]. У них были различные профили инфекции у мышей B10.A [22], BALB / C и C57BL6 (Carvalho et al., Неопубликовано), где Pb3 был менее вирулентным и вызывал больше защитных иммунных ответов, чем Pb18. Учитывая, что клеточная стенка является внешней структурой гриба и что взаимодействие между грибком и хозяином через компоненты клеточной стенки важно для установления взаимоотношений хозяин-патоген (обзор в [2]), мы полагаем, что настоящая характеристика липидов клеточной стенки может по крайней мере, частично помочь понять в будущем различия в патогенезе между этими двумя изолятами.
Значительная часть липидов клеточной стенки, вероятно, происходит из мембранных внеклеточных везикул, которые недавно были более подробно описаны в C. neoformans, Histoplasma capsulatum , Saccharomyces cerevisiae [25], [26], [27] и также у P. brasiliensis нашей группой [28]. Эти везикулы транспортируют множество компонентов во внеклеточную среду (рассмотрено в [29]) и, следовательно, им необходимо пересекать клеточную стенку. В C. neoformans они также проходят через толстую полисахаридную капсулу, как видно на изображениях просвечивающей электронной микроскопии (TEM) [25].Во время этого движения некоторые везикулы могут никогда не достичь внеклеточной среды, а просто остаться в клеточной стенке (или капсуле) и играть все еще неизвестные роли. В C. neoformans везикулы несут капсульные полисахариды, таким образом участвуя в синтезе капсул [25]. Недавно мы описали липидный состав препаратов внеклеточных везикул из Pb18 и Pb3 [30]. Сравнение данных везикул и наших настоящих результатов указывает на больше сходств, чем расхождений, следовательно, предполагая, что внеклеточные везикулы вносят вклад в значительную часть липидов клеточной стенки.
neoformans, Histoplasma capsulatum , Saccharomyces cerevisiae [25], [26], [27] и также у P. brasiliensis нашей группой [28]. Эти везикулы транспортируют множество компонентов во внеклеточную среду (рассмотрено в [29]) и, следовательно, им необходимо пересекать клеточную стенку. В C. neoformans они также проходят через толстую полисахаридную капсулу, как видно на изображениях просвечивающей электронной микроскопии (TEM) [25].Во время этого движения некоторые везикулы могут никогда не достичь внеклеточной среды, а просто остаться в клеточной стенке (или капсуле) и играть все еще неизвестные роли. В C. neoformans везикулы несут капсульные полисахариды, таким образом участвуя в синтезе капсул [25]. Недавно мы описали липидный состав препаратов внеклеточных везикул из Pb18 и Pb3 [30]. Сравнение данных везикул и наших настоящих результатов указывает на больше сходств, чем расхождений, следовательно, предполагая, что внеклеточные везикулы вносят вклад в значительную часть липидов клеточной стенки. Детали этого сравнения включены в параграфы ниже.
Детали этого сравнения включены в параграфы ниже.
В нашем анализе фосфолипиды классов PC, PE, PG, PI, PS и PA были идентифицированы в обоих изолятах, хотя Pb3 имел более высокие количества видов фосфолипидов, чем Pb18. Распределение классов фосфолипидов в целом было сходным между изолятами, где PC, за которым следовали PE, были классами фосфолипидов с наиболее многочисленными видами. Этот результат согласуется с результатами внеклеточных везикул Pb3 и Pb18 [30], а также с общим количеством фосфолипидов дрожжей и мицелия отдельных изолятов [31], [32]. Lyso -C18∶1-PC и lyso -C18∶2-PC наблюдались в небольших количествах как в Pb3, так и в Pb18 клеточной стенке, но частицы C18∶1-алкил / C18∶1-ацил-PE могли быть обнаружен только в Pb3. Во внеклеточных везикулах два эфирных фосфолипида (C16∶0-алкил / C18∶2- или C18∶1-ацил-PI) были описаны только в Pb18 [30], но не на клеточной стенке. Было высказано предположение, что эфирные фосфолипиды играют роль резервуаров для ключевых липидных медиаторов в воспалительных клетках [33], а алкилацильные разновидности фосфатидилэтаноламина в небольших количествах уже описаны в небольших количествах в клетках эритролейкемии Френда (FEL) [34], полиморфно-ядерных лейкоцитах крыс [35]. ], а также в экзосомах, происходящих из тучных клеток крыс [36].У простейшего паразита Trypanosoma cruzi гликозилфосфатидилинозитол (GPI) якоря, содержащие алкилацильный фрагмент с ненасыщенной жирной кислотой (C18∶1 или C18∶2), сильно индуцируют провоспалительные цитокины [37].
], а также в экзосомах, происходящих из тучных клеток крыс [36].У простейшего паразита Trypanosoma cruzi гликозилфосфатидилинозитол (GPI) якоря, содержащие алкилацильный фрагмент с ненасыщенной жирной кислотой (C18∶1 или C18∶2), сильно индуцируют провоспалительные цитокины [37].
В целом состав жирных кислот клеточной стенки у изолятов был сходным; однако можно выделить важные различия в численности. Подобно жирным кислотам внеклеточных везикул [30], олеиновая кислота (C18∶1) в клеточной стенке была сопоставима в Pb3 и Pb18, тогда как линолевая кислота (C18∶2) была больше в Pb18, чем в Pb3.С другой стороны, в аналогичном анализе, недавно проведенном нашей группой [30], общее количество жирных кислот, идентифицированных в дрожжевых клетках, было таким же, как и в препаратах клеточной стенки, однако с различиями в количестве: i) в дрожжах Pb3 C18∶2 был больше. чем C18∶1; ii) как C18À1, так и C18À2 были более многочисленны в Pb3, чем в Pb18, что снова указывает на то, что препараты липидов клеточной стенки были обогащены этой органеллой. Предыдущие данные о жирных кислотах из различных изолятов P. brasiliensis описывали C16∶0 [31], C18∶1 [32] или C18∶2 [38] как наиболее распространенные общие жирные кислоты в дрожжевых клетках, предполагая, что жирные кислоты состав может варьироваться в зависимости от изолята и / или условий культивирования.
Предыдущие данные о жирных кислотах из различных изолятов P. brasiliensis описывали C16∶0 [31], C18∶1 [32] или C18∶2 [38] как наиболее распространенные общие жирные кислоты в дрожжевых клетках, предполагая, что жирные кислоты состав может варьироваться в зависимости от изолята и / или условий культивирования.
Как в Pb3, так и в Pb18 клеточной стенке брассикастерин (эргоста-5,22-диен-3β-ол) был преобладающим стеролом, и его относительно больше было в Pb18. Это согласуется с тем, что мы обнаружили во внеклеточных пузырьках P. brasiliensis [30], а также в общих липидах Pb3 и Pb18 (рис. 4). Присутствие брассикастерина в дрожжах P. brasiliensis было первоначально описано San-Blas et al. [31], которые также отметили, что в фазе мицелия преобладает эргостерин. Ланостерин уже был описан как распространенный стерол у P.brasiliensis [39], хотя в нашем анализе он был обнаружен только в следовых количествах в клеточной стенке. Недавно было показано, что брассикастерин и ланостерин являются предшественниками стеролов у P. brasiliensis [40].
brasiliensis [40].
Нам удалось идентифицировать пять различных видов нейтральных гликосфинголипидов в препаратах клеточных стенок как из Pb3, так и из Pb18. В нашем анализе можно было экстрагировать только нейтральные гликолипиды, и еще предстоит определить, присутствуют ли кислые гликосфинголипиды на клеточной стенке.Наиболее распространенные Hex-C18∶0-OH / d19∶2-Cer, а также Hex-C18∶1-OH / d19∶2-Cer были идентифицированы в аналогичных относительных количествах в нашем предыдущем анализе внеклеточных везикул [30], а также в дрожжевых и гифальных клетках Pb18 [24]. Последняя работа идентифицировала глюкозу как гексозу с помощью ядерного магнитного резонанса (ЯМР). Следовательно, мы предполагаем, что идентифицированные здесь нейтральные гликолипиды являются видами глюкозилцерамида (GlcCer), которые уже были локализованы на поверхности дрожжей P. brasiliensis [14], помимо C.neoformans [7] и C. albicans [41] клеточная стенка и C. neoformans внеклеточные везикулы [25]. В C. albicans, противогрибковый растительный дефенсин RsAFP2 взаимодействует с GlcCer клеточной стенки, вызывая стресс клеточной стенки и накопление церамидов [41]. GlcCer не распознавался сыворотками пациентов с паракокцидиоидомикозом, хотя кислые гликосфинголипиды (Pb-1), содержащие концевой остаток Gal f , были описаны как иммуногенные [42]. GlcCer является ключевым элементом A.fumigatus рост [43], C. albicans вирулентность [44] и C. neoformans почкование, рост и патогенность [7], [8]. Однако в случае P. brasiliensis моноклональные антитела, направленные к GlcCer, не могли способствовать значительному ингибированию дифференцировки грибов и образования колоний [15]. Следовательно, роль GlcCer в клеточной стенке остается неизвестной.
В C. albicans, противогрибковый растительный дефенсин RsAFP2 взаимодействует с GlcCer клеточной стенки, вызывая стресс клеточной стенки и накопление церамидов [41]. GlcCer не распознавался сыворотками пациентов с паракокцидиоидомикозом, хотя кислые гликосфинголипиды (Pb-1), содержащие концевой остаток Gal f , были описаны как иммуногенные [42]. GlcCer является ключевым элементом A.fumigatus рост [43], C. albicans вирулентность [44] и C. neoformans почкование, рост и патогенность [7], [8]. Однако в случае P. brasiliensis моноклональные антитела, направленные к GlcCer, не могли способствовать значительному ингибированию дифференцировки грибов и образования колоний [15]. Следовательно, роль GlcCer в клеточной стенке остается неизвестной.
В настоящей работе мы наблюдали колебания липидного состава при культивировании P. brasiliensis в плазме человека.Как правило, многие липиды были обнаружены в пропорционально более высоких количествах при культивировании дрожжей в среде, содержащей плазму. Это также наблюдалось Chattaway и соавторами [44] в отношении липидов клеточной стенки при культивировании гиф и дрожжей C. albicans, в присутствии сыворотки. Более того, влияние человеческой плазмы на профиль транскрипции дрожжевых клеток P. lutzii (Pb01) показало усиленную регуляцию транскрипции генов, связанных с деградацией жирных кислот и ремоделированием клеточной стенки [45].Плазмосодержащая среда вызвала увеличение количества как C18À1, так и C18À2 в Pb18, тогда как она вызвала противоположный эффект в Pb3. Уже было описано, что температура культуры, возраст и состав среды могут изменять распределение жирных кислот в грибах [46], [47], [48], что подтверждает наши выводы. Наиболее драматический эффект плазмы наблюдался для брассикастерола клеточной стенки Pb3, который увеличивался в 11 раз в присутствии плазмы, тогда как общий уровень брассикастерина дрожжей снижался. Мы наблюдали, что эффект был зависимым от изолята, поскольку плазма не изменяла общий брассикастерол Pb18 дрожжевых клеток, хотя вызывал двукратное увеличение брассикастерина клеточной стенки.
Это также наблюдалось Chattaway и соавторами [44] в отношении липидов клеточной стенки при культивировании гиф и дрожжей C. albicans, в присутствии сыворотки. Более того, влияние человеческой плазмы на профиль транскрипции дрожжевых клеток P. lutzii (Pb01) показало усиленную регуляцию транскрипции генов, связанных с деградацией жирных кислот и ремоделированием клеточной стенки [45].Плазмосодержащая среда вызвала увеличение количества как C18À1, так и C18À2 в Pb18, тогда как она вызвала противоположный эффект в Pb3. Уже было описано, что температура культуры, возраст и состав среды могут изменять распределение жирных кислот в грибах [46], [47], [48], что подтверждает наши выводы. Наиболее драматический эффект плазмы наблюдался для брассикастерола клеточной стенки Pb3, который увеличивался в 11 раз в присутствии плазмы, тогда как общий уровень брассикастерина дрожжей снижался. Мы наблюдали, что эффект был зависимым от изолята, поскольку плазма не изменяла общий брассикастерол Pb18 дрожжевых клеток, хотя вызывал двукратное увеличение брассикастерина клеточной стенки. Скорости включения холестерина в клеточную стенку также различались между изолятами, они были выше у Pb3, хотя уровень общего холестерина был выше у Pb18. Возможная интерпретация этих результатов заключается в том, что дрожжевые клетки Pb18 потребляют больше холестерина по сравнению с Pb3.
Скорости включения холестерина в клеточную стенку также различались между изолятами, они были выше у Pb3, хотя уровень общего холестерина был выше у Pb18. Возможная интерпретация этих результатов заключается в том, что дрожжевые клетки Pb18 потребляют больше холестерина по сравнению с Pb3.
Вкратце, мы сравнили фосфолипиды клеточной стенки, жирные кислоты, стерины и нейтральные гликолипиды из P. brasiliensis, Pb3 и Pb18 и наблюдали влияние человеческой плазмы на состав и количество.Несмотря на некоторые расхождения, наши данные были аналогичны липидному составу, описанному нашей группой для внеклеточных везикул Pb3 и Pb18 [30]. Это подтверждает идею о том, что внеклеточные везикулы, по крайней мере, частично ответственны за состав липидов клеточной стенки. Настоящая работа существенно раскрыла природу липидной структуры клеточной стенки P. brasiliensis , открыв тем самым двери для понимания ее роли в биологии грибов, взаимодействии с противогрибковыми препаратами и с хозяином.
Дополнительная информация
Рисунок S1.
Тандем-МС-спектр C18À1 / C18À1-PC (аддукт Li + ), наиболее распространенных форм ПК, идентифицированных в режиме положительных ионов. Фрагментация была выполнена методом картирования общих ионов (TIM) с использованием PQD, а спектры были проанализированы вручную. ЧОП, фосфатидилхолин; Me 3 N, триметиламин. Указаны назначенные пики. m / z , отношение массы к заряду.
https://doi.org/10.1371/journal.pone.0063372.s001
(PPTX)
Рисунок S2.
Тандем-МС-спектр C18À1 / C18À1-PC (аддукт HCOO — ), наиболее распространенных форм ПК, идентифицированных в режиме отрицательных ионов. Фрагментация была выполнена TIM с использованием PQD, а спектры были проанализированы вручную. GroP, глицерофосфат; Я, метил. Указаны назначенные пики.
https://doi.org/10.1371/journal.pone.0063372.s002
(PPTX)
Рисунок S3.

Тандем-МС-спектр C16À0 / C18À1-PE, наиболее распространенных частиц PE, идентифицированных в режиме отрицательных ионов. Фрагментация была выполнена TIM с использованием PQD, а спектры были проанализированы вручную. GroP, глицерофосфат. Указаны назначенные пики.
https://doi.org/10.1371/journal.pone.0063372.s003
(PPTX)
Рисунок S4.
Тандем-МС-спектр C16À0 / C18À1-PS, наиболее распространенных частиц ПЭ, идентифицированных в режиме отрицательных ионов. Фрагментация была выполнена TIM с использованием PQD, а спектры были проанализированы вручную. GroP, глицерофосфат; Сер, серин.Указаны назначенные пики.
https://doi.org/10.1371/journal.pone.0063372.s004
(PPTX)
Рисунок S5.
Тандем-МС-спектр C16∶0 / C18∶2-PI, наиболее распространенных форм PI, идентифицированных в режиме отрицательных ионов. Фрагментация была выполнена TIM с использованием PQD, а спектры были проанализированы вручную.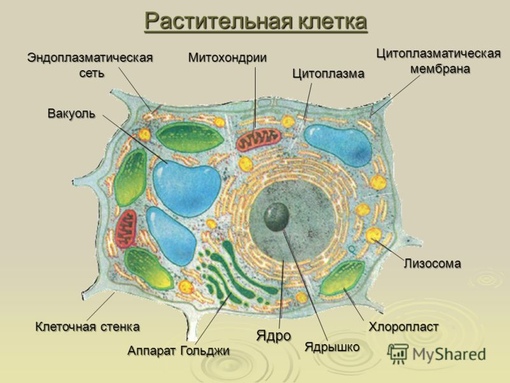 GroP, глицерофосфат; Ins, инозитол; InsP, фосфоинозитол. Указаны назначенные пики.
GroP, глицерофосфат; Ins, инозитол; InsP, фосфоинозитол. Указаны назначенные пики.
https://doi.org/10.1371/journal.pone.0063372.s005
(PPTX)
Рисунок S6.
Тандем-МС-спектр C16∶0 / C18∶1-PG, наиболее распространенных форм PG, идентифицированных в режиме отрицательных ионов. Фрагментация была выполнена TIM с использованием PQD, а спектры были проанализированы вручную. GroP, глицерофосфат. Указаны назначенные пики.
https://doi.org/10.1371/journal.pone.0063372.s006
(PPTX)
Рисунок S7.
Спектр тандемной масс-спектрометрии 16∶0 / 182-PA, наиболее распространенного вида PI-кислоты, идентифицированного в режиме отрицательных ионов. Фрагментация была выполнена путем картирования общих ионов с использованием PQD, а спектры были проанализированы вручную. GroP, глицерофосфат. Указаны назначенные пики.
https://doi.org/10.1371/journal.pone.0063372.s007
(PPTX)
Рисунок S9.

Тандем-МС-спектр гликолипида Hex-C18∶0-OH / d18∶2-Cer, идентифицированного как m / z 862,8. Фрагментацию проводили в режиме положительных ионов путем картирования общего количества ионов с использованием импульсной Q-диссоциации (PQD), а спектры анализировали вручную.Указаны назначенные пики.
https://doi.org/10.1371/journal.pone.0063372.s009
(PPTX)
Рисунок S11.
Тандем-МС-спектр гликолипида Hex-C18À1-OH / d19À2-Cer, идентифицированного при m / z 874.9. Фрагментацию проводили в режиме положительных ионов путем картирования общего количества ионов с использованием PQD, а спектры анализировали вручную. Указаны назначенные пики.
https://doi.org/10.1371/journal.pone.0063372.s011
(PPTX)
Благодарности
Мы благодарны Марсио Л. Родригесу, Леонардо Нимрихтеру и Луису Р. Травассосу за обсуждение и предложения. Благодарим Александра А. Сасаки за помощь в форматировании фигур. Мы благодарны основной лаборатории анализа биомолекул BBRC / Biology / UTEP за доступ к приборам ESI-MS и GC-MS.
Травассосу за обсуждение и предложения. Благодарим Александра А. Сасаки за помощь в форматировании фигур. Мы благодарны основной лаборатории анализа биомолекул BBRC / Biology / UTEP за доступ к приборам ESI-MS и GC-MS.
Вклад авторов
Задумал и спроектировал эксперименты: LVGL ESN MCV ALM ICA RP.Проведены эксперименты: LVGL ESN FGL. Проанализированы данные: LVGL ESN MCV FGL RP ICA. Предоставленные реагенты / материалы / инструменты анализа: RP ICA ESN FGL. Написал статью: LVGL RP ESN ICA.
Ссылки
- 1. De Groot PW, Ram AF, Klis FM (2005) Особенности и функции ковалентно связанных белков в клеточных стенках грибов. Fungal Genet Biol 42: 657–675.
- 2. Gow NA, Hube B (2012) Важность клеточной стенки Candida albicans во время комменсализма и инфекции.Curr Opin Microbiol 15: 406–412.
- 3.
Латге Дж. П. (2007) Клеточная стенка: углеводный панцирь для грибковой клетки. Mol Microbiol 66: 279–290.
- 4. Кляйн Б.С., Теббец Б. (2007) Диморфизм и вирулентность у грибов. Curr Opin Microbiol 10: 314–319.
- 5. Сан-Блас Дж., Сан-Блас Ф. (1984) Молекулярные аспекты диморфизма грибов. Crit Rev Microbiol 11: 101–127.
- 6. Mille C, Janbon G, Delplace F, Ibata-Ombetta S, Gaillardin C и др.(2004) Инактивация CaMIT1 ингибирует Candida albicans бета-маннозилирование фосфолипоманнана, снижает вирулентность и изменяет бета-маннозилирование белков клеточной стенки. J Biol Chem 279: 47952–47960.
- 7. Родригес М.Л., Травассос Л.Р., Миранда К.Р., Францен А.Дж., Розенталь С. и др. (2000) Человеческие антитела против очищенного глюкозилцерамида из Cryptococcus neoformans подавляют рост клеток и рост грибов. Заражение иммунной 68: 7049–7060.
- 8.
Rittershaus PC, Kechichian TB, Allegood JC, Merrill AH Jr, Hennig M и др.(2006) Глюкозилцерамидсинтаза является важным регулятором патогенности Cryptococcus neoformans . J Clin Invest 116: 1651–1659.
- 9. Сан-Блас Г., Нино-Вега Г., Итурриага Т. (2002) Paracoccidioides brasiliensis и паракокцидиоидомикоз: молекулярные подходы к морфогенезу, диагностике, эпидемиологии, таксономии и генетике. Med Mycol 40: 225–242.
- 10. Fortes MR, Miot HA, Kurokawa CS, Marques ME, Marques SA (2011) Иммунология паракокцидиоидомикоза.Бюстгальтер дерматол 86: 516–524.
- 11. Kanetsuna F, Carbonell LM, Moreno RE, Rodriguez J (1969) Состав клеточной стенки дрожжевых и мицелиальных форм Paracoccidioides brasiliensis . J Bacteriol 97: 1036–1041.
- 12. San-Blas F, San-Blas G, Cova LJ (1976) Морфологический мутант штамма Paracoccidioides brasiliensis IVIC Pb9. Изоляция и характеристика стен. J Gen Microbiol 93: 209–218.
- 13.
San-Blas G, San-Blas F, Ormaechea E, Serrano LE (1977) Анализ клеточной стенки требующего аденина мутанта дрожжеподобной формы Paracoccidioides brasiliensis штамм IVIC Pb9. Sabouraudia 15: 297–303.
- 14. Toledo MS, Suzuki E, Levery SB, Straus AH, Takahashi HK (2001) Характеристика моноклонального антитела MEST-2, специфичного к глюкозилцерамиду грибов и растений. Гликобиология 11: 105–112.
- 15. Толедо М.С., Таглиари Л., Сузуки Е., Сильва С.М., Страус А.Х. и др. (2010) Влияние моноклональных антител против гликосфинголипидов на рост и дифференциацию патогенных грибов. Характеристика моноклонального антитела MEST-3, направленного к Manpalpha1–> 3Manpalpha1–> 2IPC.BMC Microbiol 10: 47.
- 16. Matute DR, McEwen JG, Puccia R, Montes BA, San-Blas G, et al. (2006) Тайное видообразование и рекомбинация у гриба Paracoccidioides brasiliensis , что выявлено в генеалогии генов. Mol Biol Evol 23: 65–73.
- 17.
Morais FV, Barros TF, Fukada MK, Cisalpino PS, Puccia R (2000) Полиморфизм в гене, кодирующем иммунодоминантный антиген gp43 из патогенного гриба Paracoccidioides brasiliensis . J Clin Microbiol 38: 3960–3966.
- 18. Previato JO GP, Haskins RH, Travassos LR (1979) Растворимые и нерастворимые глюканы из различных типов клеток патогена человека Sporothrix schenckii . Экспериментальная микология. 3: 92–105.
- 19. Ичой М., Накаясу Е.С., Шпак М., Агилар С., Алей С.Б. и др. (2009) Липидомный анализ показывает, что фосфатидилглицерин и фосфатидилэтаноламин представляют собой недавно образовавшиеся фосфолипиды в раннем дивергентном простейшем, Giardia lamblia .Mol Biochem Parasitol 165: 67–78.
- 20. Мальдонадо Р.А., Куниёши Р.К., Линсс Дж. Г., Алмейда И.С. (2006) Trypanosoma cruzi олеат-десатураза: молекулярная характеристика и сравнительный анализ в других трипаносоматидах. J Parasitol 92: 1064–1074.
- 21. Pulfer M, Murphy RC (2003) Электрораспылительная масс-спектрометрия фосфолипидов. Масс-спектрометр. Ред. 22: 332–364.
- 22. Карвалью К.С., Ганико Л., Батиста В.Л., Мораис Ф.В., Маркес Э.Р. и др.(2005) Вирулентность экспрессии Paracoccidioides brasiliensis и gp43 в изолятах, несущих известный генотип PbGP43. Микробы заражают 7: 55–65.
- 23. Нельсон Д.Л. CM (2008) Принципы биохимии Ленингера. Нью-Йорк: W.H Freeman & Co., стр. 1158 стр.
- 24. Толедо М.С., Левери С.Б., Страус А.Х., Сузуки Е., Момани М. и др. (1999) Характеристика сфинголипидов микопатогенов: факторы, коррелирующие с экспрессией 2-гидрокси-жирной ацил (E) -Delta 3-ненасыщенности в цереброзидах Paracoccidioides brasiliensis и Aspergillus fumigatus .Биохимия 38: 7294–7306.
- 25. Родригес М.Л., Нимрихтер Л., Оливейра Д.Л., Фразес С., Миранда К. и др. (2007) Экспорт везикулярного полисахарида в Cryptococcus neoformans является эукариотическим решением проблемы транспорта через клеточную стенку грибов. Эукариотическая клетка 6: 48–59.
- 26. Альбукерке П.К., Накаясу Е.С., Родригес М.Л., Фразес С., Касадеваль А. и др. (2008) Везикулярный транспорт в Histoplasma capsulatum : эффективный механизм транс-клеточного переноса белков и липидов у аскомицетов.Cell Microbiol 10: 1695-1710.
- 27. Oliveira DL, Nakayasu ES, Joffe LS, Guimaraes AJ, Sobreira TJ, et al. (2010) Характеристика дрожжевых внеклеточных везикул: доказательства участия различных путей клеточного трафика в биогенезе везикул. PLoS One 5: e11113.
- 28. Вальехо М.К., Мацуо А.Л., Ганико Л., Медейрос Л.С., Миранда К. и др. (2011) Патогенный гриб Paracoccidioides brasiliensis экспортирует внеклеточные везикулы, содержащие высокоиммуногенные альфа-галактозильные эпитопы.Эукариотическая клетка 10: 343–351.
- 29. Родригес М.Л., Носанчук Д.Д., Шранк А., Вайнштейн М.Х., Касадеваль А. и др. (2011) Везикулярные транспортные системы у грибов. Future Microbiol 6: 1371–1381.
- 30. Vallejo MC, Nakayasu ES, Longo LV, Ganiko L, Lopes FG и др. (2012) Липидомный анализ внеклеточных везикул патогенной фазы Paracoccidioides brasiliensis . PLoS One 7: e39463.
- 31. Сан-Блас Дж., Урбина Дж. А., Марчан Э., Контрерас Л. М., Сораис Ф. и др.(1997) Ингибирование Paracoccidioides brasiliensis аджоеном связано с блокадой биосинтеза фосфатидилхолина. Microbiology 143 (Pt 5): 1583–1586.
- 32. Manocha MS, San-Blas G, Centeno S (1980) Липидный состав Paracoccidioides brasiliensis : возможная корреляция с вирулентностью различных штаммов. J Gen Microbiol 117: 147–154.
- 33. Chilton FH, Fonteh AN, Surette ME, Triggiani M, Winkler JD (1996) Контроль уровней арахидоната в воспалительных клетках.Biochim Biophys Acta 1299: 1–15.
- 34. Fallani A, Arcangeli A, Ruggieri S (1988) Характеристики связанных с эфиром глицерофосфолипидов в клетках эритролейкемии Френда, дифференцированных диметилсульфоксидом или гексаметиленбисацетамидом, и в неиндуцируемых клонах, обработанных индукторами. Biochem J 255: 731–735.
- 35. Рамеша CS, Пикетт WC (1987) Состав жирных кислот диацил, алкилацил и алкенилацил фосфолипидов контрольных и обедненных арахидонатом полиморфно-ядерных лейкоцитов крыс.J Lipid Res 28: 326–331.
- 36. Лауланье К., Мотта С., Хамди С., Рой С., Фовель Ф. и др. (2004) Экзосомы, происходящие из тучных и дендритных клеток, обладают специфическим липидным составом и необычной мембранной организацией. Biochem J 380: 161–171.
- 37. Алмейда И.К., Камарго М.М., Прокопио Д.О., Сильва Л.С., Мелерт А. и др. (2000) Высокоочищенные гликозилфосфатидилинозиты из Trypanosoma cruzi являются сильнодействующими провоспалительными агентами. EMBO J 19: 1476–1485.
- 38. Hahn RC, Hamdan JS (2000) Эффекты амфотерицина B и трех производных азола на липиды дрожжевых клеток Paracoccidioides brasiliensis . Противомикробные агенты Chemother 44: 1997–2000.
- 39. Перейра М., Сонг З., Сантос-Силва Л.К., Ричардс М.Х., Нгуен Т.Т. и др. (2010) Клонирование, механистический и функциональный анализ грибковой стерол-C24-метилтрансферазы, участвующей в биосинтезе брассикастерола. Biochim Biophys Acta 1801: 1163–1174.
- 40. Bertini S, Colombo AL, Takahashi HK, Straus AH (2007) Экспрессия антител, направленных к Paracoccidioides brasiliensis гликосфинголипидам во время курса лечения паракокцидиоидомикоза. Clin Vaccine Immunol 14: 150–156.
- 41. Тевиссен К., де Мелло Таварес П., Сюй Д., Бланкеншип Дж., Ванденбош Д. и др. (2012) Растительный дефенсин RsAFP2 вызывает стресс клеточной стенки, неправильную локализацию септина и накопление церамидов в Candida albicans .Mol Microbiol 84: 166–180.
- 42. Левери С.Б., Момани М., Линдси Р., Толедо М.С., Шайман Дж. А. и др. (2002) Нарушение пути биосинтеза глюкозилцерамида в Aspergillus nidulans и Aspergillus fumigatus ингибиторами UDP-Glc: церамидглюкозилтрансферазы сильно влияет на прорастание спор, клеточный цикл и рост гиф. FEBS Lett 525: 59–64.
- 43. Noble SM, French S, Kohn LA, Chen V, Johnson AD (2010) Систематический скрининг библиотеки гомозиготных делеций Candida albicans отделяет морфогенетическое переключение и патогенность.Нат Генет 42: 590–598.
- 44. Chattaway FW, Holmes MR, Barlow AJ (1968) Состав клеточной стенки мицелиальных и бластоспорных форм Candida albicans . J Gen Microbiol 51: 367–376.
- 45. Bailao AM, Shrank A, Borges CL, Parente JA, Dutra V и др. (2007) На профиль транскрипции дрожжевых клеток Paracoccidioides brasiliensis влияет плазма человека. FEMS Immunol Med Microbiol 51: 43–57.
- 46. Zarnowski R, Dobrzyn A, Ntambi JM, Woods JP (2008) Нейтральные запасающие липиды Histoplasma capsulatum : влияние возраста культуры.Curr Microbiol 56: 110–114.
- 47. Cerniglia CE, Perry JJ (1974) Влияние субстрата на жирнокислотный состав мицелиальных грибов, использующих углеводороды. J Bacteriol 118: 844–847.
- 48. Шталь П.Д., Клуг М.Дж. (1996) Характеристика и дифференциация нитчатых грибов на основе состава жирных кислот. Appl Environ Microbiol 62: 4136–4146.
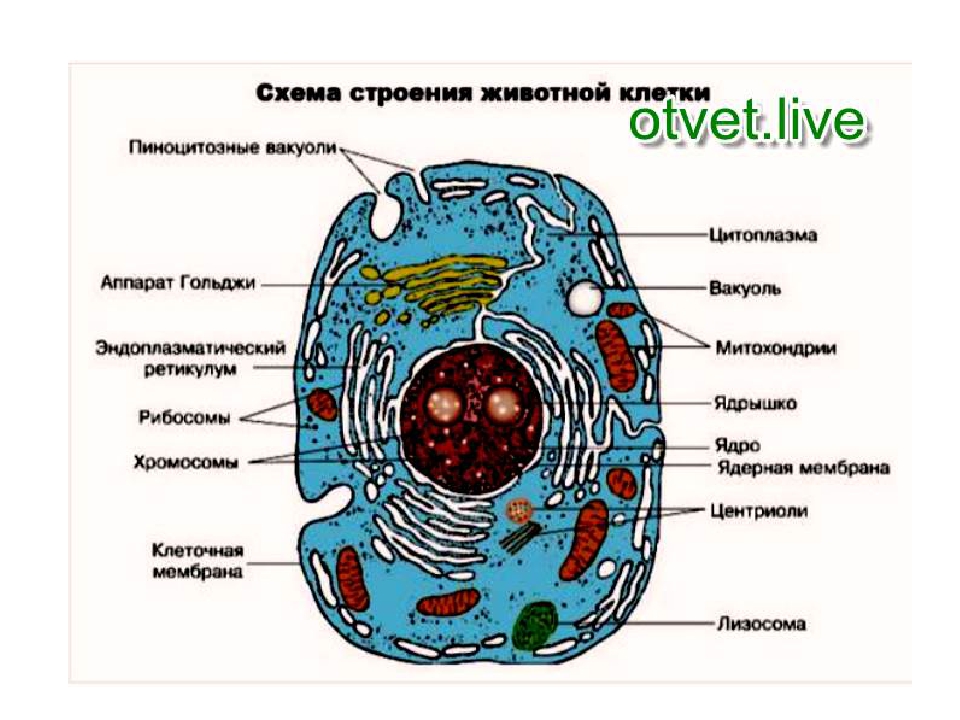
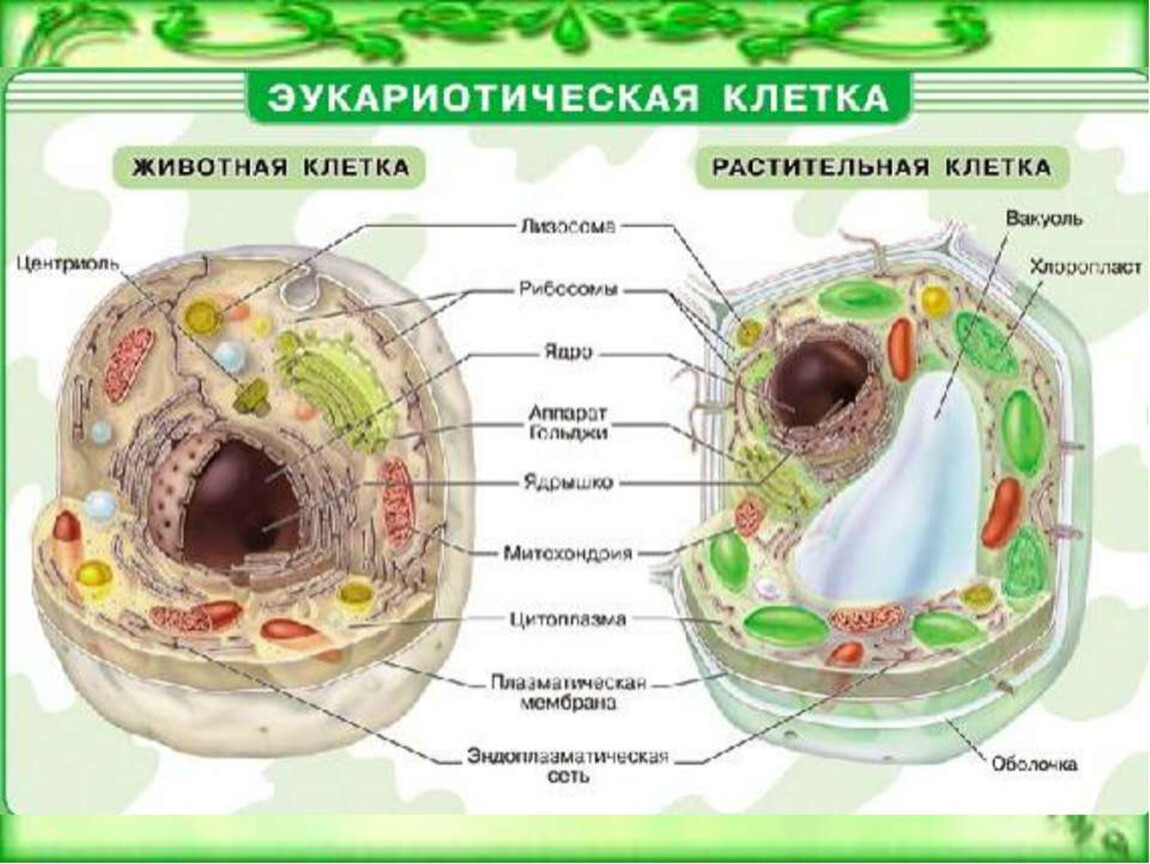 J Clin Invest 116: 1651–1659.
J Clin Invest 116: 1651–1659. Sabouraudia 15: 297–303.
Sabouraudia 15: 297–303. J Clin Microbiol 38: 3960–3966.
J Clin Microbiol 38: 3960–3966.эндоплазматической сети | Определение, функция и расположение
Эндоплазматический ретикулум (ER) , в биологии, непрерывная мембранная система, которая образует серию уплощенных мешочков в цитоплазме эукариотических клеток и выполняет множество функций, особенно важных в синтезе, складывании, модификация и транспорт белков.Все эукариотические клетки содержат эндоплазматический ретикулум (ЭР). В клетках животных ЭР обычно составляет более половины мембранного содержимого клетки. Различия в определенных физических и функциональных характеристиках различают два типа ER, известные как грубая ER и гладкая ER.
эндоплазматический ретикулумЭндоплазматический ретикулум, непрерывная мембранная система в эукариотических клетках, которая играет важную роль в биосинтезе, переработке и транспорте белков и липидов.
Encyclopædia Britannica, Inc.Популярные вопросы
Что такое эндоплазматический ретикулум?
- Эндоплазматический ретикулум (ЭР) представляет собой непрерывную мембранную систему, которая образует серию уплощенных мешочков в цитоплазме эукариотических клеток.
- Все эукариотические клетки содержат ER.
- В клетках животных ЭР обычно составляет более половины мембранного содержимого клетки.
- ER можно разделить на две функционально различные формы: гладкий эндоплазматический ретикулум (SER) и грубый эндоплазматический ретикулум (RER).
В чем разница между гладкой и шероховатой эндоплазматической сетью?
ER можно разделить на две функционально различные формы: гладкий эндоплазматический ретикулум (SER) и грубый эндоплазматический ретикулум (RER). Морфологическое различие между ними заключается в наличии синтезирующих белок частиц, называемых рибосомами, прикрепленных к внешней поверхности RER. Функции SER, сети тонких трубчатых мембранных везикул, значительно различаются от клетки к клетке, причем одной из важных ролей является синтез фосфолипидов и холестерина, которые являются основными компонентами плазмы и внутренних мембран.RER обычно представляет собой серию связанных сплюснутых мешочков. Он играет центральную роль в синтезе и экспорте белков и гликопротеинов и лучше всего изучен на секреторных клетках, специализирующихся на этих функциях. Многие секреторные клетки в организме человека включают клетки печени, секретирующие сывороточные белки (например, альбумин), эндокринные клетки, секретирующие пептидные гормоны (например, инсулин), ацинарные клетки поджелудочной железы, секретирующие пищеварительные ферменты, и хрящевые клетки, секретирующие коллаген.
Какова функция эндоплазматического ретикулума?
Эндоплазматический ретикулум (ER) выполняет важные функции, в частности, в синтезе, сворачивании, модификации и транспорте белков.Различия в определенных физических и функциональных характеристиках различают два типа ER, известные как грубая ER (RER) и гладкая ER (SER). Рибосомы на RER, которые придают RER его грубый вид, специализируются на синтезе белков, которые обладают сигнальной последовательностью, которая направляет их специально в ER для обработки. Белки, синтезируемые посредством RER, имеют определенные конечные пункты назначения, такие как клеточная мембрана, внешняя часть клетки или сам ER. SER участвует в синтезе липидов, включая холестерин и фосфолипиды, которые используются в производстве новых клеточных мембран.В клетках печени SER способствует детоксикации лекарств и вредных химикатов. Саркоплазматический ретикулум — это особый тип SER, который регулирует концентрацию ионов кальция в цитоплазме поперечно-полосатых мышечных клеток.
Когда был открыт эндоплазматический ретикулум?
ER был впервые обнаружен в конце 19 века, когда исследования окрашенных клеток показали присутствие некоторого типа обширной цитоплазматической структуры, известной тогда как гастроплазма. Электронный микроскоп сделал возможным изучение морфологии ER в 1940-х годах, когда ему было дано его нынешнее название.
Rough ER назван из-за его грубого внешнего вида, который связан с рибосомами, прикрепленными к его внешней (цитоплазматической) поверхности. Грубый ER находится непосредственно рядом с ядром клетки, и его мембрана непрерывна с внешней мембраной ядерной оболочки. Рибосомы на грубом ER специализируются на синтезе белков, которые обладают сигнальной последовательностью, которая направляет их специально в ER для обработки. (Ряд других белков в клетке, включая те, которые предназначены для ядра и митохондрий, нацелены на синтез на свободных рибосомах или на тех, которые не прикреплены к мембране ER; см. статью о рибосомах.Белки, синтезированные грубым ER, имеют определенные конечные пункты назначения. Некоторые белки, например, остаются внутри ER, тогда как другие отправляются в аппарат Гольджи, который находится рядом с ER. Белки, секретируемые аппаратом Гольджи, направляются в лизосомы или на клеточную мембрану; третьи предназначены для выделения наружу клетки. Белки, предназначенные для транспорта в аппарат Гольджи, переносятся из рибосом на грубом ER в грубый просвет ER, который служит местом сворачивания, модификации и сборки белка.
эндоплазматический ретикулум; organelle. Сканирующая электронная микрофотография ацинарной клетки поджелудочной железы, на которой видны митохондрии (синий), шероховатый эндоплазматический ретикулум (желтый; рибосомы выглядят как маленькие точки) и аппарат Гольджи (серый, в центре и внизу слева).
Пьетро М. Мотта и Томонори Нагуро / Science SourceБлизость грубого ER к ядру клетки дает ER уникальный контроль над процессингом белка. Грубый ER способен быстро посылать сигналы в ядро, когда возникают проблемы с синтезом и сворачиванием белка, и тем самым влияет на общую скорость трансляции белка.Когда неправильно свернутые или развернутые белки накапливаются в просвете ЭР, активируется сигнальный механизм, известный как ответ на развернутый белок (UPR). Ответ является адаптивным, так что активация UPR запускает снижение синтеза белка и повышение способности к сворачиванию белка ER и связанного с ER деградации белка. Если адаптивный ответ терпит неудачу, клетки направляют на апоптоз (запрограммированную гибель клеток).
Smooth ER, напротив, не связан с рибосомами, и его функции различаются.Гладкий ER участвует в синтезе липидов, включая холестерин и фосфолипиды, которые используются в производстве новых клеточных мембран. В некоторых типах клеток гладкий ER играет важную роль в синтезе стероидных гормонов из холестерина. В клетках печени он способствует детоксикации лекарств и вредных химических веществ. Саркоплазматический ретикулум — это особый тип гладкого ЭПР, который регулирует концентрацию ионов кальция в цитоплазме поперечно-полосатых мышечных клеток.
Оформите подписку Britannica Premium и получите доступ к эксклюзивному контенту. Подпишись сейчасСильно запутанная и лабиринтная структура ER привела к его описанию в 1945 году клеточными биологами Китом Портером, Альбертом Клодом и Эрнестом Фуллманом, которые создали первую электронную микрофотографию клетки. В конце 1940-х и начале 1950-х годов Портер и его коллеги Хелен П. Томпсон и Фрэнсис Каллман ввели термин эндоплазматический ретикулум для описания органелл.Позже Портер работал с американским клеточным биологом румынского происхождения Джорджем Паладе, чтобы выяснить ключевые характеристики ER.
Грубый эндоплазматический ретикулум — определение, функция и структура
Грубый эндоплазматический ретикулум Определение
Грубый эндоплазматический ретикулум (грубый ER) является частью эндомембранной системы клетки и подмножеством эндоплазматического ретикулума (ER). Эта органелла в первую очередь связана с синтезом, сворачиванием и модификацией белков, особенно тех, которые необходимо доставить к различным органеллам внутри клетки или секретировать из клетки.Грубый ER также участвует в ответе клетки на развернутые белки и играет роль в индукции апоптоза из-за его тесного взаимодействия с митохондриями.
Грубый ER характеризуется присутствием мембраносвязанных рибосом, которые придают ему характерный вид под микроскопом. Эти рибосомы выглядят как гвоздики и отличают органеллы от гладких участков ER. Некоторые белки также синтезируются цепочками рибосом, называемыми полисомами. Шероховатый ER также можно идентифицировать по его морфологии — он часто состоит из извитых, уплощенных мешкообразных структур, которые берут начало около ядра.Просвет грубого ER примыкает к перинуклеарному пространству, а мембраны грубого ER связаны с внешней ядерной мембраной.
Структура грубого эндоплазматического ретикулума
ЭПР можно морфологически разделить на две структуры — цистерны и пластинки. Грубый эндоплазматический ретикулум в основном состоит из листов — двумерного массива сплюснутых мешочков, которые проходят через цитоплазму. Помимо рибосом, эти мембраны содержат важный белковый комплекс, называемый транслоконом, который необходим для трансляции белка в грубом ER.
Структура грубого ER также тесно связана с присутствием цитоскелетных элементов, особенно микротрубочек. Когда структура микротрубочек временно нарушается, сеть ER разрушается и восстанавливается только после восстановления цитоскелета. Изменения паттерна полимеризации микротрубочек также отражаются в изменениях морфологии ER. Кроме того, когда рибосомы отделяются от слоев грубого эндоплазматического ретикулума, эти структуры могут диспергироваться и образовывать трубчатые цистерны.
Края листов ER имеют высокую кривизну, которую необходимо стабилизировать. Белки, называемые ретикулонами, и DP1 / Yop1p играют важную роль в этой стабилизации. Эти белки представляют собой интегральные мембранные белки, которые образуют олигомеры для формирования липидного бислоя. Кроме того, они также используют структурный мотив, который вставляется в одну створку мембраны и увеличивает ее кривизну. Эти два класса белков являются избыточными, поскольку избыточная экспрессия одного белка, по-видимому, компенсирует недостаток другого белка.
Функции грубого эндоплазматического ретикулума
Шероховатый эндоплазматический ретикулум играет в клетке ряд ролей, в значительной степени связанных с синтезом белка. Полипептиды синтезируются, модифицируются, складываются в их правильную трехмерную форму и сортируются по направлению к органеллам или маркируются для секреции. Он также играет важную роль в модулировании реакции клетки на стресс и в контроле качества правильного сворачивания белка. Когда количество развернутых белков увеличивается, клетки изменяют соотношение своих канальцев: листов.Это может возникать из-за большей площади, доступной в листах грубого ER для спасения развернутого белка, или может отражать потребность в отдельном протеоме грубого ER.
Протеом грубого ER отражает его специфическую роль в клетке. Он содержит ферменты, участвующие в метаболизме РНК, которые связываются с РНК и модифицируют ее. Это необходимо, поскольку органелла участвует в трансляции РНК в белок. Он также содержит белки, которые распознают различные сигнальные последовательности в растущем полипептиде и способствуют их транслокации.Ферменты и белки гликозилирования, которые действуют как молекулярные шапероны, которые обеспечивают правильную укладку синтезированных полипептидов, также являются важными белками в этой органелле. Иногда апоптоз индуцируется ER в ответ на избыток развернутого белка внутри клетки. Эта функция опосредуется митохондриями.
Синтез белков
Трансляция всех белков начинается в цитоплазме после того, как обработанный транскрипт мРНК экспортируется из ядра.Трансляция начинается со связывания рибосомы со зрелым транскриптом мРНК. Однако после образования первых нескольких аминокислот некоторые полипептиды импортируются в ER до того, как трансляция может продолжаться. Это основано на распознавании короткого отрезка аминокислот, также известного как сигнальная последовательность, многочисленными цитозольными рибонуклеопротеинами, называемыми частицами распознавания сигнала (SRP). Привязка SRP временно останавливает трансляцию и позволяет всему механизму перевода двигаться к ER.В ER возникающий полипептид проникает в органеллу через трансмембранные каналы, называемые транслоконами. Эти каналы состоят из комплекса белков, которые позволяют полипептиду проходить через гидрофобный липидный бислой мембраны ER. Канал не очень широкий, поэтому полипептид нужно вставлять в виде развернутой цепочки аминокислот. На этом этапе SRP диссоциируют от полипептида, и трансляция возобновляется. После того, как первые несколько аминокислот попадают в просвет, резидентные ферменты ER часто расщепляют сигнальную последовательность.Новые аминокислоты добавляются к растущей полипептидной цепи, поскольку рибосома остается прикрепленной к мембране ER, а возникающий белок продолжает вставляться в просвет ER. Этот процесс называется ко-трансляционным импортом в ER.
Процесс трансляции через мембраносвязанные рибосомы особенно важен для белков, которые необходимо секретировать. Таким образом, грубая ЭР заметна в клетках печени, которые секретируют сывороточный альбумин, клетках пищеварительной системы, которые секретируют ферменты, эндокринных клетках, которые синтезируют и секретируют белковые гормоны (например, инсулин), и в клетках, которые создают белки внеклеточного матрикса.Синтез белков с участием грубого ER также важен для мембраносвязанных белков, особенно таких, как G-Protein-Coupled Receptors (GPCR), которые содержат несколько гидрофобных участков и пересекают мембрану более одного раза через изгибы шпильки в своей структуре. Точная роль транслоконов и ER-резидентных белков в выполнении сложной задачи трансляции таких белков до конца не изучена.
В груди млекопитающих секреторная система, вовлекающая грубый ER, имеет решающее значение во время лактации.Отдельные слои кубовидных эпителиальных клеток участвуют в основном процессе производства молока. Ядро в этих клетках расположено ближе к базальному концу клетки, а грубый ER и аппарат Гольджи расположены близко к ядру. Белки, синтезируемые грубым ER, включают выдающийся молочный белок казеин и сывороточные белки. Эти белки упакованы в секреторные везикулы или большие мицеллы и проходят через сеть Гольджи, прежде чем слиться с плазматической мембраной, выпуская их содержимое в молочные протоки.
Сворачивание белка и контроль качества
Одним из побочных эффектов трансляции на грубый ER, когда полипептид транслоцируется в виде развернутой цепочки аминокислот, является то, что эти короткие отрезки необходимо защищать до тех пор, пока они не смогут сформировать окончательный Трехмерная структура, предотвращающая преждевременное образование агрегатов. Одним из важных механизмов обеспечения правильной укладки белка является гликозилирование возникающего полипептида с помощью ферментов, называемых олигосахарилтрансферазами.Эти ферменты являются частью транслоконного комплекса грубой мембраны ER. Гликозилирование увеличивает растворимость пептидных цепей и защищает их до тех пор, пока молекулярные шапероны не смогут связываться с ними и облегчать их складывание. Известные молекулярные шапероны грубого ER включают связывающий белок иммуноглобулина (BiP), калнексин (CNX) и кальретикулин (CRT). CNX / CRT способствует сворачиванию белков в сочетании с гликозилированием. BiP содержит субстрат-связывающую область, которая распознает гидрофобные участки в полипептиде, и АТФазный домен, который обеспечивает его сродство к этим участкам.Члены семейства белков DnaJ / Hsp40 помогают BiP в его задаче, модулируя его АТФазную активность и усиливая его взаимодействие с факторами обмена нуклеотидов. ER также содержит ферменты, которые катализируют образование дисульфидных связей и субстрат-специфических шаперонов и ферментов, необходимых для определенных белков. Он также поддерживает окислительную среду, чтобы помочь в этой задаче.
BiP, CNX / CRT и другие шапероны обогащены участками ER, которые тесно взаимодействуют с митохондриями.Этот участок ER называется MAM или митохондриально-ассоциированной мембраной. MAM становится важным центром передачи сигналов внутри клетки, который интегрирует сигналы от ER и играет роль в гомеостазе кальция, аутофагии, апоптозе и функции митохондрий.
Несмотря на эти механизмы, гарантирующие правильную укладку белков, некоторые из них необходимо удалить из системы либо из-за ошибок трансляции, либо из-за генетических мутаций, ведущих к производству дефектных белков.Это достигается за счет систем контроля качества в ER, которые «проверяют» вновь синтезированные белки. Когда полипептид не свернулся в свое нативное состояние, молекулярные шапероны снова связываются с полипептидом и делают еще одну попытку свернуть белок в его правильную форму. Когда повторные попытки терпят неудачу, неправильно свернутые белки могут быть экспортированы в цитозоль и удалены через протеасомы с использованием убиквитин-опосредованной деградации белков.
Сортировка белков
После того, как белки синтезированы и свернуты, их необходимо отправить к месту назначения.Первым шагом в этом процессе является образование пузырьков по краям шероховатой ЭПР. Эти везикулы несут груз к сети Гольджи и создаются скоординированным действием множества белков, начиная с везикулярного комплекса белков оболочки II (COPII). Фермент GTPase и фактор обмена нуклеотидов необходимы COPII для выполнения своих функций. Вместе эти белки искажают мембрану и позволяют формировать везикулу, несущую соответствующий груз. Белки, которые должны оставаться в ЭПР, перемещаются обратно через ретроградный транспорт из Гольджи с помощью пузырьков, образованных родственным белком, называемым COPI.
- Мицелла — совокупность молекул, содержащая как гидрофильные, так и гидрофобные области, диспергированные в жидкости, образующие коллоидный раствор. В водной среде мицеллы образуются с гидрофильными областями, обращенными к воде, и гидрофобными областями, изолируемыми внутрь.
- Полисома — Ассоциация между зрелым транскриптом мРНК и двумя или более рибосомами, участвующими в трансляции кодонов внутри РНК.
- Протеом — Полный набор белков, экспрессируемых в клетке, ткани, органе или организме в определенный момент времени.
- Рибонуклеопротеин — Комплекс, образованный ассоциацией рибонуклеиновой кислоты (РНК) с белками.
Тест
1. Что из утверждений относительно шероховатого эндоплазматического ретикулума верно?
A. Решающее значение для синтеза белков, которые секретируются клеткой
B. Важное значение во время лактации и производства молока
C. Шипованные рибосомами и полисомами
D. Все вышеперечисленное
Ответ на вопрос № 1
D правильный.Грубый эндоплазматический ретикулум играет важную роль в синтезе белков, предназначенных для секреции из клетки. Следовательно, он усеян рибосомами и полисомами, которые транслируют мРНК, содержащую код этих белков. Этот процесс особенно важен во время кормления грудью, поскольку молоко содержит ряд белков, которые поддерживают ребенка в первые месяцы.
2. Какой из этих молекулярных механизмов непосредственно участвует в правильном сворачивании белка в ЭР?
А. Связывание сигнальных частиц с формирующимся полипептидом
B. Транслоконы на мембране ER
C. Гликозилирование и связывание молекулярных шаперонов
D. Все вышеперечисленное
Ответ на вопрос № 2
C правильный. Из трех вариантов только гликозилирование и связывание молекулярных шаперонов играет роль в сворачивании белка внутри ER. Связывание SRP с растущей полипептидной цепью и присутствие транслоконов на мембране ER являются важными аспектами синтеза белка.Однако эти компоненты синтеза белка не участвуют напрямую в достижении полипептидом правильной трехмерной формы или нативной структуры.
3. Какой из этих белков участвует в антероградном транспорте из грубого ER в аппарат Гольджи?
A. Убиквитин и протеасома
B. Белки-шапероны CNR / CXT
C. COPII
D. Все вышеперечисленное
Ответ на вопрос № 3
C правильный .Убиквитин-опосредованная протеасомная деградация белков является важной частью механизма контроля качества внутри клетки для удаления неправильно свернутых белков. Точно так же система CNR / CXT и другие молекулярные шапероны также являются частью механизмов внутри грубого ER, чтобы гарантировать правильное формирование полипептидов. Однако по завершении этого процесса именно белки COPII участвуют в антероградном транспорте пузырьков, несущих груз к Golgi от эндоплазматического ретикулума.
Британское общество клеточной биологии
Быстрый просмотр: Центриоль, обнаруженная только в клетках животных и некоторых низших растений, состоит из коротких отрезков микротрубочек, расположенных параллельно друг другу и расположенных вокруг центральной полости в форме цилиндра.
В клетках животных центриоли расположены в центросоме и образуют ее часть, где они представляют собой парные структуры, расположенные под прямым углом друг к другу. В этом контексте они, возможно, участвуют в сборке веретена во время митоза.Центросома расположена в цитоплазме вне ядра, но часто рядом с ним.
Единственная центриоль обнаруживается также на базальном конце ресничек и жгутиков. В этом контексте оно называется «базальным телом» и связано с ростом и работой микротрубочек в ресничке или жгутике.
Чтобы просмотреть изображение центриолей в сперме дрозофилы, интерпретированное с помощью технологии CIMR GridPoint, НАЖМИТЕ ЗДЕСЬ
Центриоли представляют собой нечто вроде загадки
Центриоли присутствуют в (1) клетках животных и (2) базальной области ресничек и жгутиков у животных и низших растений (например.грамм. хламидомонады). В ресничках и жгутиках центриоли называются «базальными тельцами», но их можно рассматривать как взаимопревращаемые.
Центриоли отсутствуют в клетках высших растений.
Некоторые считают, что когда клетки животных подвергаются митозу, они получают выгоду от присутствия центриолей, которые, по-видимому, контролируют формирование волокон веретена и которые позже влияют на разделение хромосом. Однако исследования показали, что митоз может происходить в клетках животных после разрушения центриолей.Иногда кажется, что это происходит за счет аномалий в развитии веретена и последующих проблем с разделением хромосом. Недавние исследования также показывают, что эмбрионы Drosophila арестовываются очень рано, если репликация центриолей невозможна.
У высших растений митоз протекает вполне удовлетворительно, микротрубочки образуют волокна веретена, но без помощи центриолей. Таким образом, функция центриолей остается загадкой.
Структура
Центриоль состоит из коротких отрезков микротрубочек, расположенных в форме цилиндра с открытым концом, длиной около 500 нм и диаметром 200 нм.Микротрубочки, образующие стенку цилиндра, сгруппированы в девять наборов пучков по три микротрубочки в каждом.
У ресничек и жгутиков, у которых центриоли находятся в основании структуры и называются базальными тельцами, архитектура стенок и полостей немного отличается. Помимо стенок цилиндра, состоящих из девяти наборов пучков из трех микротрубочек, существуют стенки из девяти наборов из двух пучков. В обоих типах есть центральная матрица, из которой расходятся спицы, как в колесе тележки.
В клетках животных центриоли обычно располагаются парами с цилиндрическими центриолями, расположенными под прямым углом друг к другу.
Центриоли образуют вокруг себя «облако» белкового материала; это перицентриолярный материал (ПКМ). Вместе они составляют важнейшую центросому.
Функция
Центриоли функционируют как пара в большинстве клеток животных, но как единое центриоль или базальное тело в ресничках и жгутиках.
Центриоли в парах
Клетки, вступающие в митоз, имеют центросому, содержащую две пары центриолей и связанный с ними перицентриолярный материал (ПКМ).Во время профазы центросома делится на две части, и пара центриолей мигрирует к каждому концу или полюсу на внешней стороне ядерной мембраны или оболочки. В этот момент микротрубочки образуются на внешнем крае перицентриолярного материала и разрастаются радиально. Пара центриолей и ПКМ называется звездочкой. Микротрубочки от звезды на одном полюсе растут к звездочке на противоположном полюсе. Эти микротрубочки называют волокнами веретена. Некоторые из них будут прикреплены центромерами к хромосомам, выстроенным на «экваторе» делящейся клетки.Другие, хотя и не прикреплены к хроматидам / хромосомам центромерами, помогают раздвигать две части делящейся клетки.
Отдельная центриоль или базальное тело.
В основании каждой реснички или жгутика есть единственная центриоль. Эта структура и связанный с ней перицентриолярный материал создают микротрубочки в линейном направлении. Эти микротрубочки образуют большую часть внутренней части ресничек и жгутиков и в значительной степени отвечают за механические аспекты своего движения, используя белковые моторы.Центриоль у основания каждой из них также, по-видимому, в некоторой степени направляет и контролирует движение ресничек и жгутиков.
Репликация
В клетках, где центриоли присутствуют в паре, репликация происходит в течение всего клеточного цикла. В фазе G1 два центриольных цилиндра очень немного отодвигаются друг от друга. Во время фазы S новые цилиндры микротрубочек образуются рядом с двумя «материнскими» цилиндрами и под прямым углом к ним. Две пары центриолей держатся очень близко друг к другу до профазной стадии митоза.В этот момент они разделяются, и обе пары центриолей движутся по внешней поверхности ядерной оболочки к противоположным концам или «полюсам» клетки, образуя астральные полюса делящейся клетки.
Сводка
- Центриоли встречаются как спаренных цилиндрических органелл вместе с перицентриолярным материалом (ПКМ) в центросоме животной клетки.
- центриолей обнаруживаются как одиночных структур в ресничках и жгутиках в клетках животных и некоторых клетках низших растений.
- Центриоли состоят из микротрубочек.
- В клетках животных центриоли организуют перицентриолярный материал для образования микротрубочек, включая волокна митотического веретена.
- центриолей представляют собой нечто вроде загадки; они, по-видимому, влияют на результат митоза в клетках животных. Если есть удовлетворительное деление, то без них митоз все еще имеет место, но иногда с неудовлетворительным результатом. Центриоли отсутствуют в клетках высших растений, но имеет место нормальный митоз с удовлетворительными результатами.
ВАЖНОЕ УВЕДОМЛЕНИЕ
Иногда новая информация бросает вызов текущему мышлению только для того, чтобы заменяться другими идеями по мере развития знаний. Так работает наука. Британское общество клеточной биологии (BSCB) прилагает все усилия для предоставления авторитетной и актуальной информации, но не может нести ответственность за любую потерю отметок в результате использования информации на этом веб-сайте, которая может противоречить общепринятым взглядам органов, проводящих экспертизу.
Leave A Comment