
Биология Развитие животных с превращением и без превращения
Материалы к уроку
Конспект урока
Развитие животных с превращением и без превращения
Сегодня на уроке речь пойдет о разных способах развития животных. А начнем мы урок со сказки.
«Родилась гусеница внутри яблока. Живет — поживает, вкусно кушает. Но однажды проснувшись, она обнаружила вокруг себя сгнившее яблоко. Расстроилась.
Гусенице весь мир показался гнилым, лишенным света и маленьким, как то яблоко, в котором она родилась.
— Я должна понять, зачем нужен весь этот прогнивший мир — и я в нем! Я должна вылезти из него! — закричала гусеница неизвестно кому и, собрав последние силы, в неистовом рывке проломила кожуру яблока. Какая вокруг красота. Но больше всего гусеницу очаровали прекрасные существа, порхающие среди спелых плодов и почему-то очень напоминающие ей саму себя. Это была стая красивейших бабочек. Они, то взмывали, то опускались над завороженной гусеницей. Своим волшебным танцем они рассказывали ей о том, как стать бабочкой и научиться летать вместе с ними. И это волшебство случилось».
Своим волшебным танцем они рассказывали ей о том, как стать бабочкой и научиться летать вместе с ними. И это волшебство случилось».
В каждой сказке есть истина. И, мы с вами наблюдая, за жизнью животных, видим как животные развиваются, растут, меняются. Но при этом остается между ними сходство. Котенок не так уж сильно отличается от кошки, щенок от собаки, теленок от коровы. Но в природе есть такие животные, которые при рождении, росте и развитии меняются настолько сильно, что взрослого и детёныша невозможно сравнивать друг с другом. В этом случае животное проходит через превращение, которое можно назвать вторым рождением. Например, из личинки, напоминающей червя, получается бабочка, а из головастика, похожего на рыбу развивается лягушка. Такие сложные видоизменения организма называют метаморфозами.
Метаморфозы свойственны многим животным и даже простейшим организмам. Впрочем, когда речь заходит о метаморфозах, в первую очередь говорят о насекомых. Ведь именно в этом классе происходят самые необычные превращения.
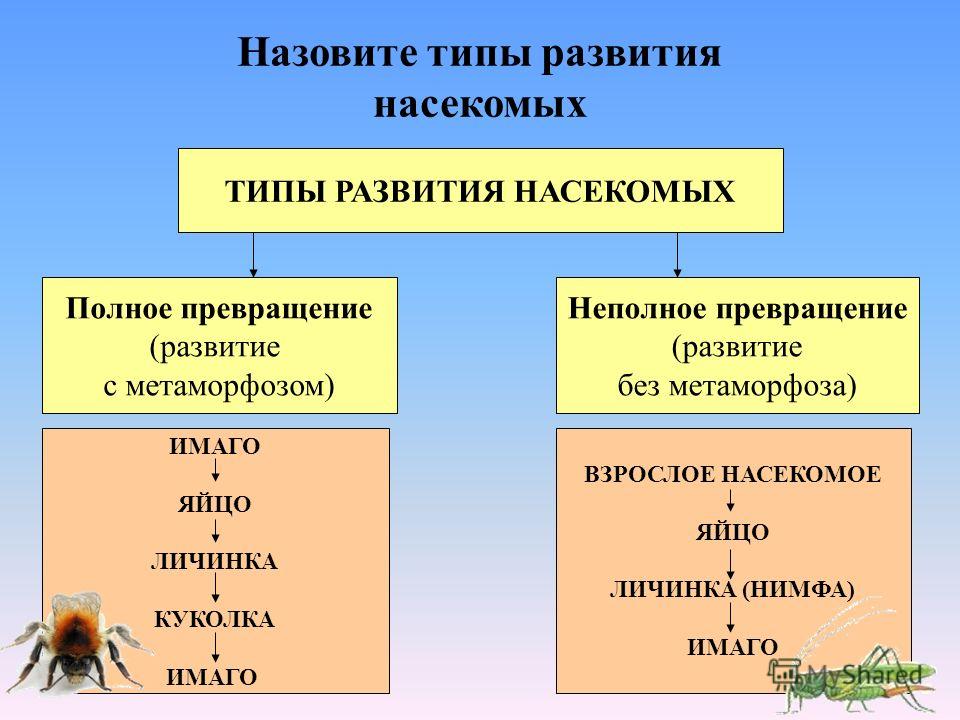
По классификации ученых метаморфозы бывают полные и неполные.
При полном метаморфозе животное проходит четыре стадии развития: яйцо, личинка, куколка и взрослая особь. Последнюю стадию развития часто называют имаго. Имаго – половозрелая стадия насекомых или взрослое насекомое.
В каждый период жизни организм насекомого выполняет своеобразный вид деятельности. В яйце происходит первоначальное развитие организма. Это развитие незаметно, но оно очень важно. Из яйца появляется личинка. Личинка постоянно ест — ее задача быстро расти и накапливать питательные вещества. Запас питательных веществ должен быть большой и его должно хватить, чтобы обеспечить жизнедеятельность куколки и даже частично взрослого насекомого.
Так, например, личинка моли живет всего три месяца и за это время успевает увеличить свой вес в четыреста раз!
Выросшая гусеница начинает готовиться к превращению в куколку.
Куколка — это стадия индивидуального развития насекомых с полным превращением, следующая за личинкой.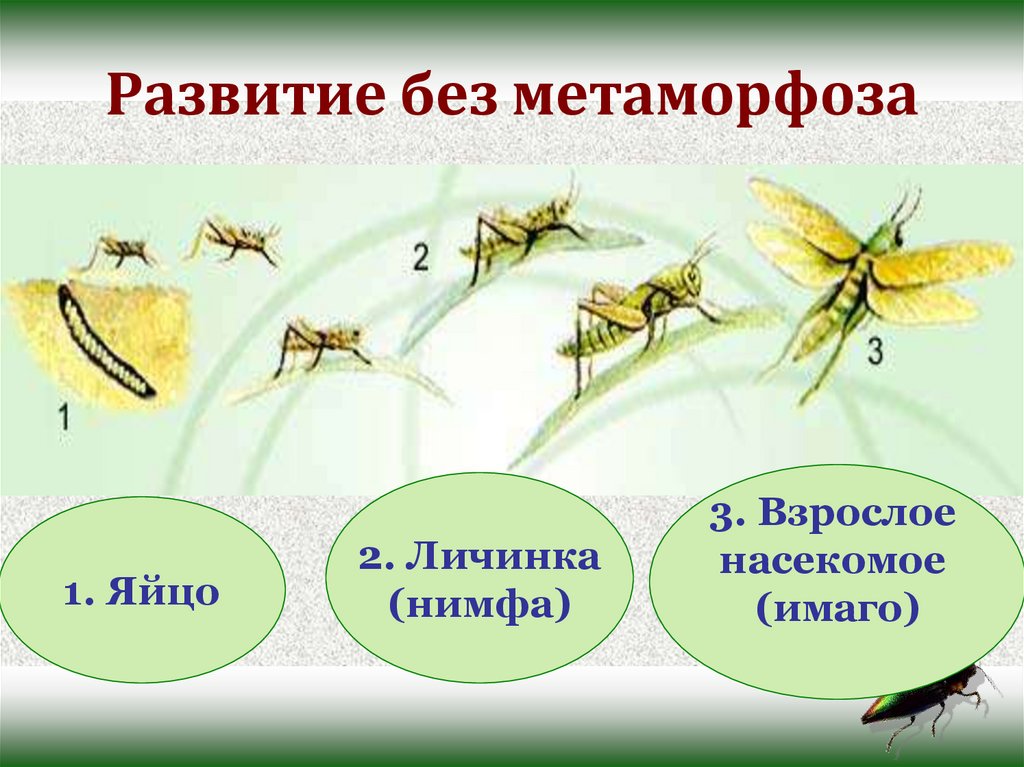 Куколка всегда прикреплена к поверхности, она неподвижна, хотя иногда и может шевелить брюшком. Она ничего не ест, однако дышит, испаряет воду и, расходует накопленные гусеницей питательные вещества. В куколке хорошо защищенное насекомое преобразуется во взрослую особь — имаго. Выйдя из куколки, насекомое покрывается хитиновым покровом, меняет характер питания и приобретает способность к полету.
Куколка всегда прикреплена к поверхности, она неподвижна, хотя иногда и может шевелить брюшком. Она ничего не ест, однако дышит, испаряет воду и, расходует накопленные гусеницей питательные вещества. В куколке хорошо защищенное насекомое преобразуется во взрослую особь — имаго. Выйдя из куколки, насекомое покрывается хитиновым покровом, меняет характер питания и приобретает способность к полету.
Основная задача взрослой особи — размножение. Чтобы ее выполнить, нужно не так уж много времени. Поэтому некоторые насекомые, выйдя из куколки, живут всего один день. Например, это можно сказать о бабочке. Взрослая бабочка почти не ест — у некоторых видов бабочек в процессе превращения из личинки в имаго исчезает рот.
В природе наряду с полным метаморфозом, существует также неполный метаморфоз. При неполном метаморфозе из яиц появляется личинка, внешне очень напоминающая взрослую особь. Но у личинки отсутствуют крылья, половая система неразвита. Личинка активно питается, развивается, растет и линяет. После нескольких линек личинка больше не растет. Личинку, которая очень похожа на взрослое насекомое называют нимфой. Стадия куколки у таких насекомых отсутствует.
После нескольких линек личинка больше не растет. Личинку, которая очень похожа на взрослое насекомое называют нимфой. Стадия куколки у таких насекомых отсутствует.
Развитие с неполным метаморфозом наблюдается у кузнечика, саранчи, стрекозы, цикады и у клопов.
Рассмотрим метаморфозы у других представителей Царства Животные.
С метаморфозом развиваются многощетинковые черви. При этом из яйца выходит планктонная личинка, способная двигаться с помощью ресничек. Тело ее шаровидное или несколько вытянутое. На верхней части расположена кучка ресничек. Это орган чувств. Затем эта шаровидная личинка растет и превращается в червя. Взрослые черви живут в илистом грунте, а личинки их свободно плавают, что позволяет виду расселяться на новых территориях.
Интересен метаморфоз у свиного цепня. У него из яйца выходит личинка с шестью крючками. Затем она превращается в финну – пузырек, с расположенными внутри головкой и шейкой, как у взрослого червя. Финна растет, и достигает величины горошины. Дальнейшее превращение во взрослого червя происходит при условии смены хозяина.
Дальнейшее превращение во взрослого червя происходит при условии смены хозяина.
У мидии из яйца тоже выходит маленькая личинка. Через некоторое время она превращается в другую личинку, которая называется парусником. Парусник плавает, питается, подрастает, оседает на грунт и превращается в молодого моллюска.
Личинки некоторых пресноводных двустворчатых моллюсков, являясь наружными паразитами рыб, вгрызаются в их кожу и питаются соками этих хозяев.
Метаморфоз свойствен и позвоночным животным. У лягушки процесс развития проходит через три стадии: яйцо, личинка, взрослая особь. Это пример неполного метаморфоза.
Лягушки откладывают в воду икру, похожую на икру рыб. Икринки разбухает и внутри видно яйцо. Яйцо многократно делится и превращается в многоклеточный зародыш.
Через две недели из икринки появляется личинка лягушки — головастик.
Головастик по внешнему виду напоминает маленькую рыбку. Дышит головастик сначала наружными жабрами, затем появляются внутренние жабры. У головастика один круг кровообращения и двухкамерное сердце, как у рыб.
У головастика один круг кровообращения и двухкамерное сердце, как у рыб.
Первые дни головастик живет за счет запасов питательных веществ икринки. Затем у него образуется рот и головастик переходит к питанию простейшими организмами. Дальнейшие изменения головастика зависят от условий среды. Чем теплее — тем быстрее развитие. Сначала у них появляются задние конечности, потом передние. Развиваются легкие — органы воздушного дыхания. Головастики начинают заглатывать воздух. Наступает время, когда хвост рассасывается, головастик превращается в лягушонка. От момента откладки икры до превращения головастика в лягушку проходит обычно около трех месяцев.
Земноводное тигровая амбистома имеет необычный метаморфоз. В мелких водоемах с высокой температурой метаморфоз у личинок тигровой амбистомы идет обязательно, и они сравнительно быстро превращаются во взрослых животных. В глубоких водоемах с низкой температурой часто остаются жить личинки, не превращаясь во взрослую особь. У некоторых подвидов амбистом метаморфоз не наступает совсем, так что для них известны только личинки — аксолотли. Часто личинки продолжают расти, у них развивается половая система, и они размножаются в личиночной стадии.
Часто личинки продолжают расти, у них развивается половая система, и они размножаются в личиночной стадии.
Аксолотль — личиночная форма тигровой амбистомы, способная размножаться, не превратившись во взрослую форму.
В процессе метаморфоза преобразуется не только внешний вид животного и строение его организма, а также условия его существования. К примеру, головастик плавает в воде, а лягушка может жить на суше. У многих низших животных, к примеру, у кишечнополостных, метаморфозы связаны со сменой поколений и служат для расселения. Полип прикреплен к поверхности, а дочерний организм — медуза способен свободно передвигаться. Причем чередуются у животных не только жизненные формы, но и половое и бесполое размножение. Такие метаморфозы называют первичными.
При метаморфозе разрушаются личиночные органы и возникают органы, присущие взрослым животным.
Метаморфоз связан с переменой образа жизни или средой обитания и имеет важное эволюционное значение.
Остались вопросы по теме? Наши репетиторы готовы помочь!
Подготовим к ЕГЭ, ОГЭ и другим экзаменам
Найдём слабые места по предмету и разберём ошибки
Повысим успеваемость по школьным предметам
Поможем подготовиться к поступлению в любой ВУЗ
Выбрать репетитораОставить заявку на подбор
Неполное превращение | Справочник по защите растений — AgroXXI
Работа с энциклопедией в полевых условиях без ИнтернетаПрофессиональный поиск по пестицидам и агрохимикатам
Подпишитесь и получайте новости где удобнее
ДЗЕН Telegram Вконтакте Дзен. новости
новости
Рекомендуем
Справочник по защите растений
Справочник по защите растений
Incomplete conversion
Статья из разделов: СловарьРазвитие насекомых
Неполное превращение – тип метаморфоза, при котором по мере своего развития насекомое проходит следующие стадии: яйцо, личинка, имаго.
Схема неполного превращения у клопаНеполное превращение и изменения морфологии насекомых
На протяжении смены стадий и между ними (при смене возрастов) насекомые с неполным превращением переживают несколько периодов линьки (экдизиса), во время которых они сбрасывают кутикулу, заменяемую новой. Это позволяет их телу постепенно увеличиваться в размерах.
При неполном метаморфозе различия между представителями разных стадий менее выражены, чем при полном. Личинка многих насекомых (например, кузнечиков) в общих чертах напоминает уменьшенную взрослую особь. Тем не менее, это правило существует не всегда. Так, у стрекоз каждая фаза жизненного цикла имеет свои особенности, благодаря которым, личинка и имаго выглядят по-разному.
Так, у стрекоз каждая фаза жизненного цикла имеет свои особенности, благодаря которым, личинка и имаго выглядят по-разному.
Схожесть между стадиями развития особенно заметна у примитивных бескрылых насекомых, у которых все визуальные изменения заключаются лишь в смене пропорций тела и увеличении его в размерах на протяжении взросления. Таких насекомых еще называют аметаболическими.
Несмотря на меньшее количество стадий развития, наличие у насекомых неполного метаморфоза не является признаком их эволюционного несовершенства; продолжительность, количество и особенности разных фаз связаны лишь с условиями существования, в которых живут насекомые. Справедливость этого утверждения можно оценить в полной мере, если вспомнить такой пример, как циклы развития тлей. На протяжении жизни эти насекомые проходят ряд специфических стадий развития, определяющихся количеством пищевых источников и их доступностью на протяжении сезона.
Гемиметаболическое развитие у поденкиХарактеристика отдельных стадий
Яйцо. Отличается теми же особенностями, что и у насекомых с полным превращением: неподвижно, может иметь большие или меньшие размеры. На протяжении данной стадии у зародыша формируются основные органы, по окончании созревания происходит выход личинки.
Отличается теми же особенностями, что и у насекомых с полным превращением: неподвижно, может иметь большие или меньшие размеры. На протяжении данной стадии у зародыша формируются основные органы, по окончании созревания происходит выход личинки.
Личинка. В молодых возрастах личинка может отличаться по внешнему облику от имаго, но по мере развития она, как правило, все больше к нему приближается. С самого начала существования личинки ее основные морфологические отличия от взрослых особей состоят лишь в отсутствии крыльев, половых органов и структур, с ними связанных.
Близость строения личинок и имаго при неполном превращении обусловлена, главным образом, тем, что различные «приобретения» появляются у таких насекомых не со сменой стадий, а постепенно, на протяжении стадии личинки, по мере смены ее возрастов. По этой причине раньше неполное превращение также называли постепенным (другие названия – частичный, прямой метаморфоз, паурометаболическое развитие). Так, у крылатых видов (например, у тараканов) зачатки крыльев появляются примерно в третьем личиночном возрасте; в отличие от них, крылья у организмов с полным метаморфозом (жуков, бабочек) становятся приобретением только последней стадии развития – имаго, а их зачатки появляются у куколок.
Неполовозрелые формы насекомых с неполным метаморфозом также называются нимфами. Некоторые нимфы живут в иной среде, нежели взрослые насекомые, например, половозрелые поденки ведут надземный образ жизни, а их личинки живут под водой. В подобных случаях личинки сильнее всего отличаются от взрослой формы: строением ротового аппарата и конечностей, формой тела, структурой органов дыхания и др. Такие нимфы также называются наядами. Вариант, при котором водноживущие наяды обладают трахеями и имеют множество других специализированных черт, отличающих их от взрослых крылатых форм с дыхальцами, называется гемиметаболическим развитием.
Имаго. На этой стадии насекомые приобретают способность к размножению, все их органы достигают максимального развития. Крылатые насекомые с неполным метаморфозом нередко отличаются архаичными чертами строения, сходством анатомии своих органов со строением аналогичных органов предковых насекомых. Яркий тому пример – жилкование крыльев у уже упомянутых стрекоз.
Близкие статьи
Имаго
Imago
Личинка
Larva
Превращение (метаморфоз) насекомых
Insect metamorphosis
Ссылки
Все статьи в разделе: Развитие насекомых
(c) Справочник AgroXXI
Интересна тема? Подпишитесь на персональные новости в ДЗЕН или Pulse или VK.Новости.
Посоветовать коллегам
Источники
Развитие животных | Определение, процесс, этапы, особенности и факты
эмбрионов разных животных
Смотреть все медиа
- Похожие темы:
- животное биологическое развитие
Просмотреть весь связанный контент →
развитие животных , процессы, которые в конечном итоге приводят к образованию нового животного, начиная с клеток, полученных от одной или нескольких родительских особей.
Общие характеристики
Размножение и развитие
У многоклеточных животных (Metazoa) размножение принимает одну из двух существенно различных форм: половую и бесполую. При бесполом размножении новая особь происходит от бластемы, группы клеток родительского тела, иногда, как у Hydra и других кишечнополостных, в виде «почки» на поверхности тела. У губок и мшанок группы клеток, из которых развиваются новые особи, формируются внутри и могут быть окружены защитными оболочками; эти тела, которые могут служить резистентными формами, способными противостоять неблагоприятным условиям среды, высвобождаются после смерти родителя. У некоторых животных родитель может разделиться пополам, как у некоторых червей, у которых отдельный червь распадается на две довольно равные части (за исключением того, что передняя половина получает рот, «мозг» и органы чувств, если они есть).
Очевидно, в таком случае нельзя сказать, кто из двух получившихся особей является родителем, а кто потомком. Некоторые офиуры (родственники морских звезд) могут размножаться, ломаясь посередине диска тела, при этом каждая из половин впоследствии отращивает свою недостающую половину и соответствующие руки.
Общей чертой всех форм бесполого размножения является то, что клетки — всегда значительное число клеток, а не только одна клетка — участвующие в образовании новой особи, не отличаются существенно от других телесных или соматических клеток. Число хромосом (тел, несущих наследственный материал) в клетках, участвующих в образовании бластемы, такое же, как и в других соматических клетках родителя, составляющих нормальную, двойную или диплоидную (2 n ), комплект.
При половом размножении новая особь образуется не соматическими клетками родительского организма, а половыми клетками или гаметами, которые существенно отличаются от соматических клеток тем, что подверглись мейозу — процессу, при котором число хромосом уменьшается до одной половина диплоидного (2 n ) числа обнаружена в соматических клетках; клетки, содержащие один набор хромосом, называются гаплоидными ( n ). Таким образом, образующиеся половые клетки получают только половину числа хромосом, присутствующих в соматической клетке. Кроме того, половые клетки обычно способны развиваться в новую особь только после того, как две из них объединились в процессе, называемом оплодотворением.
Таким образом, образующиеся половые клетки получают только половину числа хромосом, присутствующих в соматической клетке. Кроме того, половые клетки обычно способны развиваться в новую особь только после того, как две из них объединились в процессе, называемом оплодотворением.
Каждый тип размножения — бесполое и половое — имеет свои преимущества для вида. Бесполое размножение, по крайней мере в некоторых случаях, является более быстрым процессом, быстрее всего ведущим к развитию большого числа особей. Самцы и самки независимо друг от друга способны производить потомство. Большие размеры первоначальной массы живого вещества и высокая степень его организации — новая особь наследует части тела родителя, например часть пищеварительного канала, — упрощают последующее развитие и достижение этап, способный к самообеспечению легче. Новые особи, полученные в результате бесполого размножения, имеют ту же генетическую конституцию (генотип), что и их родители, и составляют то, что называется клоном.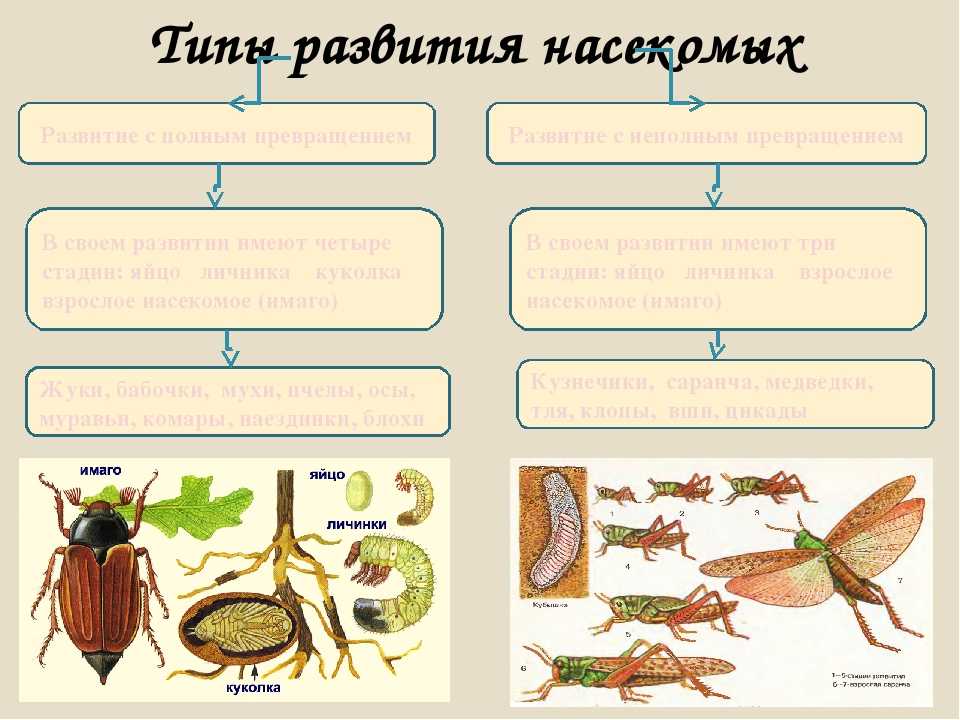
Оформите подписку Britannica Premium и получите доступ к эксклюзивному контенту.
Эволюция метаморфоза насекомых: взгляд на развитие и эндокринную систему
1. Misof B, et al. 2014. Филогеномика определяет время и характер эволюции насекомых. Наука 346, 763–767. ( 10.1126/science.1257570) [PubMed] [CrossRef] [Google Scholar]
2. Берлезе А. 1913. Intorno alle metamorphosi degli insetti. Редия 9, 121–136. [Google Scholar]
3. Иммс А.Д. 1937. Последние достижения энтомологии. Филадельфия, Пенсильвания: Блэкистон. [Google Scholar]
4. Truman JW, Riddiford LM. 1999. Происхождение метаморфоза насекомых. Природа 401, 447–452. ( 10.1038/46737) [PubMed] [CrossRef] [Google Scholar]
5. Пояркофф Э. 1914. Essai d’une théorie de la nymphe des Insectes Holométaboles. Арка Зоол. Эксп. Ген. 54, 221–265. [Google Scholar]
6. Hinton HE. 1963 год. Происхождение и функции стадии куколки. проц. Р. Энтомол. соц. Лондон сер. А 38, 77–85. [Академия Google]
7. Sehnal F, Švácha P, Zrzavy J.
1996.
Эволюция метаморфоза насекомых. В « Метаморфозе: постэмбриональное перепрограммирование экспрессии генов в клетках амфибий и насекомых» (редакторы Гилберт Л.И., Тата Дж. Р., Аткинсон Б. Г.), стр. 3–58. Сан-Диего, Калифорния: Academic Press. [Google Scholar]
В « Метаморфозе: постэмбриональное перепрограммирование экспрессии генов в клетках амфибий и насекомых» (редакторы Гилберт Л.И., Тата Дж. Р., Аткинсон Б. Г.), стр. 3–58. Сан-Диего, Калифорния: Academic Press. [Google Scholar]
8. Рейнфорд Дж.Л., Хофрейтер М., Николсон Д.Б., Мэйхью П.Дж. 2014. Филогенетическое распределение существующего богатства предполагает, что метаморфозы являются ключевой инновацией, способствующей диверсификации насекомых. ПЛОС ОДИН 9, e109085 ( 10.1371/journal.pone.0109085) [бесплатная статья ЧВК] [PubMed] [CrossRef] [Google Scholar]
9. Эрезылмаз Д.Ф. 2006. Несовершенные яйца и яйцевидные нимфы: история представлений о происхождении метаморфоза насекомых. интегр. Комп. биол. 46, 795–807. ( 10.1093/icb/icl033) [PubMed] [CrossRef] [Google Scholar]
10. Рейнольдс С. 2019. Приготовление идеального насекомого: трансформационная идея Аристотеля о полной метаморфозе насекомых. Фил. Транс. Р. Соц. Б 374, 201 ( 10.1098/rstb.2019.0074) [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
11.
12. Джиндра М. 2019. Откуда взялась куколка? Время передачи сигналов ювенильных гормонов подтверждает гомологию между стадиями гемиметаболических и голометаболических насекомых. Фил. Транс. Р. Соц. Б 374, 201
( 10.1098/rstb.2019.0064) [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
13. Truman JW, Riddiford LM. 2002. Эндокринные взгляды на эволюцию метаморфоза у насекомых. Анну. Преподобный Энтомол. 47, 467–500. ( 10.1146/annurev.ento.47.091201.145230) [PubMed] [CrossRef] [Google Scholar]
14. Ян А.С. 2001. Модульность, эволюционируемость и адаптивное излучение: сравнение геми- и голометаболических насекомых. Эвол. Дев. 3, 59–72. ( 10.1046/j.1525-142x.2001.003002059.x) [PubMed] [CrossRef] [Google Scholar]
15. Шеперд Д., Бейт С.М.
1990.
Пространственные и временные паттерны нейрогенеза у зародыша саранчи ( Schistocerca gregaria ). Разработка
108, 83–96. [Google Scholar]
Разработка
108, 83–96. [Google Scholar]
16. Трумэн Дж.В. 1996. Метаморфоз нервной системы насекомых. В « Метаморфозе: постэмбриональное перепрограммирование экспрессии генов в клетках амфибий и насекомых» (ред. Гилберт Л.И., Тата Дж. Р., Аткинсон Б. Г.), стр. 283–320. Сан-Диего, Калифорния: Academic Press. [Google Scholar]
17. Паулюс Х.Ф. 1986 год. Evolutionswege zum Larvalauge der Insekten-ein Modell für die Entstehung und die Ableitung der ozellaren Lateralaugen der Myriapoda von Fazettenaugen. Зоол. Ярб. Сист. 113, 353–371. ( 10.1111/j.1439-0469.1989.tb00345.x) [CrossRef] [Google Scholar]
18. Лю З., Фридрих М. 2004. Tribolium , гомолог стекла и эволюция глаз личинок насекомых. Дев. биол. 269, 36–54. ( 10.1016/j.ydbio.2004.01.012) [PubMed] [CrossRef] [Google Scholar]
19. Иноуэ Ю., Мито Т., Мияваки К., Мацусима К., Синмё Ю., Хэануэ Т.А., Мардон Г., Охучи Х., Нодзи С.
2002.
Корреляция паттернов экспрессии гомоторакса , таксы и без дистального отдела с проксимодистальной сегментацией зачатка ноги сверчка. мех. Дев.
113, 141–148. ( 10.1016/S0925-4773(02)00017-5) [PubMed] [CrossRef] [Google Scholar]
мех. Дев.
113, 141–148. ( 10.1016/S0925-4773(02)00017-5) [PubMed] [CrossRef] [Google Scholar]
20. Erezyilmaz DF, Riddiford LM, Truman JW. 2004. Ювенильный гормон действует при эмбриональной линьке и индуцирует нимфальную кутикулу у пряморазвивающегося сверчка. Дев. Гены Эвол. 214, 313–323. ( 10.1007/s00427-004-0408-2) [PubMed] [CrossRef] [Google Scholar]
21. Tanaka K, Truman JW. 2005. Развитие эпидермиса ног взрослых особей Manduca sexta : вклад различных популяций личиночных клеток. Дев. Гены Эвол. 215, 78–89. ( 10.1007/s00427-004-0458-5) [PubMed] [CrossRef] [Google Scholar]
22. Танака К., Трумэн Дж.В. 2007. Механизм формирования молекулярного паттерна, лежащий в основе метаморфоза грудной ноги у Manduca sexta . Дев. биол. 305, 539–550. ( 10.1016/j.ydbio.2007.02.042) [PubMed] [CrossRef] [Google Scholar]
23. Kim C-W.
1959.
Центр дифференциации, индуцирующий развитие от личинки до взрослой ноги у Pieris brassicae (Lepidoptera). Дж. Эмбриол. Эксп. Морфол.
7, 572–582. [PubMed] [Академия Google]
Дж. Эмбриол. Эксп. Морфол.
7, 572–582. [PubMed] [Академия Google]
24. Трумэн Дж.В., Хирума К., Алли Дж.П., Маквинни С.Г.Б., Чамплин Д.Т., Риддифорд Л.М. 2006. Ювенильный гормон необходим для соединения формирования имагинального диска с питанием у насекомых. Наука 312, 1385–1388. ( 10.1126/science.1123652) [PubMed] [CrossRef] [Google Scholar]
25. Thomas JB, Bastiani MJ, Bate M, Goodman CS. 1984. От кузнечика до Дрозофилы : общий план развития нейронов. Природа 310, 203–207. ( 10.1038/310203a0) [PubMed] [CrossRef] [Google Scholar]
26. Трумэн Дж.В., Болл Э.Е. 1998. Паттерны эмбрионального нейрогенеза у примитивного бескрылого насекомого, чешуйницы, Ctenolepisma longicaudata : сравнение с наблюдаемыми у летающих насекомых. Дев. Гены Эвол. 208, 357–368. ( 10.1007/s004270050192) [PubMed] [CrossRef] [Google Scholar]
27. Коэн С.М.
1993.
Развитие имагинального диска. В «Развитии Drosophila melanogaster», vol. 2 (ред. Бейт М., Мартинес-Ариас А. ), стр. 747–841. Плейнвью, Нью-Йорк: Cold Spring Harbour Press. [Академия Google]
), стр. 747–841. Плейнвью, Нью-Йорк: Cold Spring Harbour Press. [Академия Google]
28. Конопова Б., Зрзави Ю. 2005. Ультраструктура, развитие и гомология эмбриональных кутикул насекомых. Дж. Морф. 267, 339–362. ( 10.1002/jmor.10338) [PubMed] [CrossRef] [Google Scholar]
29. Kato Y, Riddiford LM. 1987. Роль 20-гидроксиэкдизона в стимуляции эпидермальных митозов во время личиночно-куколочной трансформации табачного рогатого червя, Manduca sexta . Разработка 100, 227–236. [Google Scholar]
30. Свача П. 1992. Что такое имагинальные диски, а что нет: переоценка некоторых основных понятий (Insecta, Holometabola). Дев. биол. 154, 101–117. ( 10.1016/0012-1606(92)
-I) [PubMed] [CrossRef] [Google Scholar]
31. Huet C, Lenoir-Rousseux JJ.
1976 год.
Этюд де ла миза на месте воображаемого узора Tenebrio molitor. I. Проанализируйте экспериментальный процесс восстановления в процессе морфогенеза. Дж. Эмбриол. Эксп. Превращаться.
35, 303–321. ( 10.1016/0016-6480(75)
( 10.1016/0016-6480(75)
-1) [PubMed] [CrossRef] [Google Scholar]
32. Mirth CK, Truman JW, Riddiford LM. 2009. Рецептор экдизона контролирует переключение посткритического веса на независимую от питания дифференцировку в Дрозофила крыловые имагинальные диски. Разработка 136, 2345–2353. ( 10.1242/dev.032672) [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
33. Champlin DT, Truman JW. 1998. Экдистероиды управляют двумя фазами развития глаз во время метаморфоза бабочки Manduca sexta . Разработка 125, 2009–2018 гг. [PubMed] [Google Scholar]
34. Erezyilmaz DF, Riddiford LM, Truman JW. 2006. Спецификатор куколки , широкий , направляет прогрессивный морфогенез у пряморазвивающегося насекомого. проц. Натл акад. науч. США 103, 6925–6930. ( 10.1073/pnas.0509983103) [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
35. Quennedey A, Quennedey B.
1990.
Морфогенез крыла Anlagen мучного жука Tenebrio molitor на последнем личиночном этапе. Клетка ткани
22, 721–740. (10.1016/0040-8166(90)
Клетка ткани
22, 721–740. (10.1016/0040-8166(90)
36. Clark-Hachtel CM, Linz DM, Tomoyasu Y. 2013. Информация о происхождении крыльев насекомых получена с помощью функционального анализа рудиментарных у красного мучного жука, Tribolium castaneum . проц. Натл акад. науч. США 110, 16 951–16 956. ( 10.1073/pnas.1304332110) [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
37. Cheong SPS, Huang J, Bendena WG, Tobe SS, Hui JL. 2015. Эволюция шелушения и метаморфоза у членистоногих: усиление регуляции ювенильного гормона. интегр. Комп. биол. 55, 878–890. ( 10.1093/icb/icv066) [PubMed] [CrossRef] [Google Scholar]
38. Nijhout HF, Callier V. 2014. Механизмы развития размеров тела и масштабирования тела крыльев у насекомых. Анну. Преподобный Энтомол. 60, 141–156. ( 10.1146/annurev-ento-010814-020841) [PubMed] [CrossRef] [Google Scholar]
39. Смыкал В., Даймон Т., Каюкава Т., Такаки К., Шинода Т. , Джиндра М.
2015.
Важность передачи сигналов ювенильных гормонов возникает в связи со способностью личинок насекомых к метаморфозам. Дев. биол.
390, 221–230. ( 10.1016/j.ydbio.2014.03.006) [PubMed] [CrossRef] [Google Scholar]
, Джиндра М.
2015.
Важность передачи сигналов ювенильных гормонов возникает в связи со способностью личинок насекомых к метаморфозам. Дев. биол.
390, 221–230. ( 10.1016/j.ydbio.2014.03.006) [PubMed] [CrossRef] [Google Scholar]
40. Daimon T, Uchibori M, Nakao H, Sezutsu H, Shinoda T. 2015. Нокаутные шелкопряды обнаруживают незаменимую роль ювенильных гормонов в голометаболическом жизненном цикле. проц. Натл акад. науч. США 112, E4226–E4235. ( 10.1073/pnas.1506645112) [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
41. Абулафия-Багински Н., Пенер М., Стаал Г.Б. 1984. Химическая аллатэктомия поздних эмбрионов Locusta синтетическим прекоценом и ее влияние на морфогенез личинок. J. Физиология насекомых. 30, 839–852. ( 10.1016/0022-1910(84)
-X) [CrossRef] [Google Scholar] 42. Zhou X, Truman JW, Riddiford LM.
2004.
Сверхэкспрессия wide : новый взгляд на его роль в клетках переднегрудной железы Drosophila . Дж. Эксп. биол. 207, 1151–1161. ( 10.1242/jeb.00855) [PubMed] [CrossRef] [Google Scholar]
207, 1151–1161. ( 10.1242/jeb.00855) [PubMed] [CrossRef] [Google Scholar]
43. MacWhinnie SGB, Allee JP, Nelson CA, Riddiford LM, Truman JW, Champlin DT. 2005. Роль питания в создании имагинального диска глаза и инициации метаморфоза у Manduca sexta . Дев. биол. 285, 285–297. ( 10.1016/j.ydbio.2005.06.021) [PubMed] [CrossRef] [Google Scholar]
44. Кояма Т., Сыропятова М.О., Риддифорд Л.М. 2008. Передача сигналов инсулина/ИФР регулирует изменение коммитирования в имагинальных дисках и зачатках путем подавления эффекта ювенильного гормона. Дев. биол. 324, 258–265. ( 10.1016/j.ydbio.2008.09.017) [PubMed] [CrossRef] [Google Scholar]
45. Riddiford LM. 1978 год. Вызванное экдизоном изменение клеточной приверженности эпидермиса табачного рогатого червя Manduca sexta в начале метаморфоза. Генерал Комп. Эндокрин. 34, 438–446. ( 10.1016/0016-6480(78)-8) [PubMed] [CrossRef] [Google Scholar]
46. Williams CM.
1961.
Ювенильный гормон. II. Его роль в эндокринном контроле линьки, окукливания и развития взрослых особей тутового шелкопряда Cecropia. биол. Бык.
121, 572–585. ( 10.2307/1539456) [CrossRef] [Google Scholar]
биол. Бык.
121, 572–585. ( 10.2307/1539456) [CrossRef] [Google Scholar]
47. Zhou X, Riddiford LM. 2002. Broad определяет развитие куколки и опосредует действие «статус-кво» ювенильного гормона на трансформацию куколки во взрослую особь у Drosophila и Manduca . Разработка 129, 2259–2269. [PubMed] [Google Scholar]
48. Бергот Б.Дж., Бейкер Ф.К., Серф Д.К., Джеймисон Г., Скули Д.А. 1981. Качественные и количественные аспекты титров ювенильных гормонов в развивающихся эмбрионах некоторых видов насекомых: открытие нового ЮГ-подобного вещества, выделенного из яиц Мандука секста . В биохимии ювенильных гормонов. Действие, агонизм и антагонизм (редакторы Pratt GE, Brookes GT), стр. 33–45. Амстердам, Нидерланды: Elsevier/North-Holland Biomedical Press. [Google Scholar]
49. Риддифорд Л.М., Уильямс С.М.
1967.
Влияние аналогов ювенильного гормона на эмбриональное развитие тутового шелкопряда. проц. Натл акад. науч. США
57, 595–601. ( 10.1073/pnas.57.3.595) [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
( 10.1073/pnas.57.3.595) [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
50. Lagueux M, Hetru C, Goltzene F, Kappler C, Hoffmann JA. 1979. Титр экдизона и метаболизм в связи с кутикулогенезом у эмбрионов Locusta migratoria . J. Физиология насекомых. 25, 709–723. ( 10.1016/0022-1910(79)-9) [CrossRef] [Google Scholar]
51. Темин Г., Зандер М., Руссель Дж.П. 1986 год. Физико-химические (ГХ-МС) измерения титров ювенильного гормона III во время эмбриогенеза Locusta migratoria . Междунар. Дж. Инверт. Воспр. 9, 105–112. ( 10.1080/01688170.1986.10510184) [CrossRef] [Google Scholar]
52. Имбоден Х., Ланцрайн Б., Дельбек Дж. П., Люшер М. 1978. Экдистероиды и ювенильный гормон во время эмбриогенеза у яйцеживородящих тараканов Nauphoeta cinerea . Генерал Комп. Эндокринол. 36, 628–635. ( 10.1016/0016-6480(78)
-1) [PubMed] [CrossRef] [Google Scholar]
53. Maestro O, Cruz J, Pascual N, Martin D, Belles X.
2005.
Дифференциальная экспрессия двух изоформ RXR/ультрадыхальца в течение жизненного цикла гемиметаболического насекомого Blattella germanica (Dictyoptera, Blattellidae). Мол. Клеточный эндокринол.
238, 27–37. ( 10.1016/j.mce.2005.04.004) [PubMed] [CrossRef] [Google Scholar]
Мол. Клеточный эндокринол.
238, 27–37. ( 10.1016/j.mce.2005.04.004) [PubMed] [CrossRef] [Google Scholar]
54. Маэстро Дж.Л., Паскуаль Н., Трейблмайр К., Лозано Дж., Беллес Х. 2010. Ювенильный гормон и аллатостатины в эмбрионе немецкого таракана. Биохимия насекомых. Мол. биол. 40, 660–665. ( 10.1016/j.ibmb.2010.06.006) [PubMed] [CrossRef] [Google Scholar]
55. Fernandez-Nicolas A, Belles X. 2017. Передача сигналов ювенильного гормона у гемиметаболических эмбрионов с короткой зародышевой полосой. Разработка 144, 4637–4644. ( 10.1242/dev.152827) [PubMed] [CrossRef] [Google Scholar]
56. Pecasse F, Beck Y, Ruiz C, Richards G. 2000. Krüppel-homolog , стадийно-специфический модулятор ответа предкуколочного экдизона, необходим для метаморфоза Drosophila . Дев. биол. 221, 53–67. ( 10.1006/dbio.2000.9687) [PubMed] [CrossRef] [Google Scholar]
57. Minakuchi C, Namiki T, Shinoda T.
2009. Гомолог Круппеля 1 , ранний ювенильный гормональный ответ гена ниже по течению от Метопрен-толерантный , опосредует его антиметаморфическое действие на красного мучного жука Tribolium castaneum .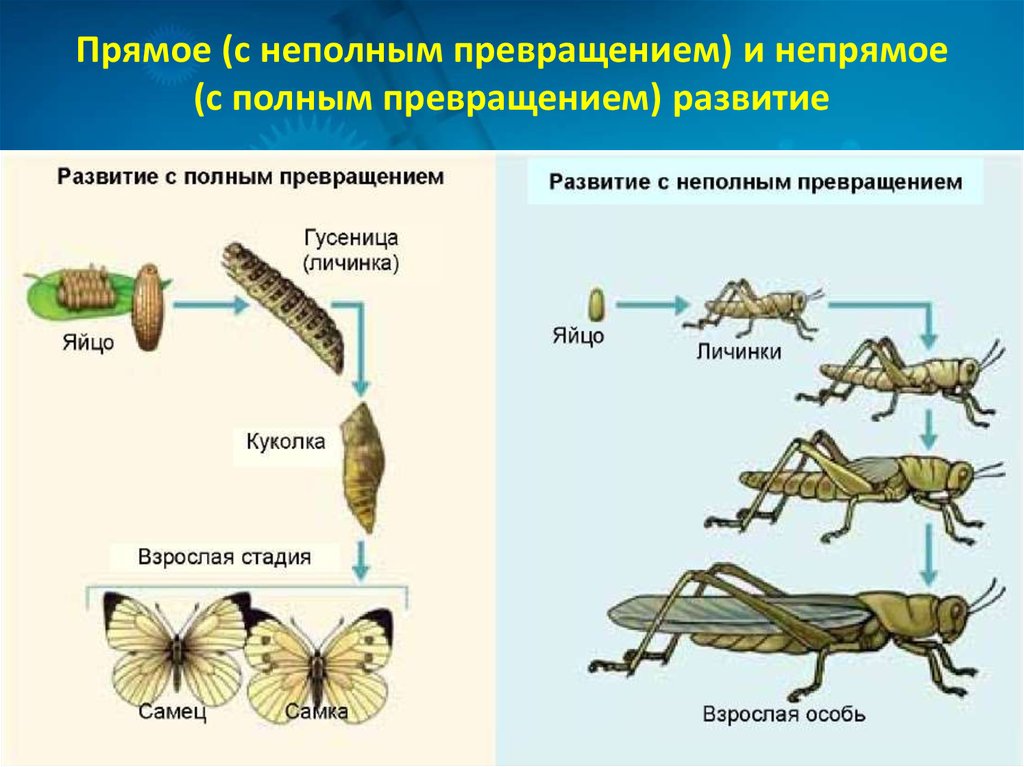 Дев. биол.
325, 341–350. ( 10.1016/j.ydbio.2008.10.016) [PubMed] [CrossRef] [Google Scholar]
Дев. биол.
325, 341–350. ( 10.1016/j.ydbio.2008.10.016) [PubMed] [CrossRef] [Google Scholar]
58. Лозано Дж., Беллес Х. 2011. Законсервированная репрессивная функция гомолога 1 Круппеля в отношении метаморфоза насекомых у гемиметаболических и голометаболических видов. науч. Респ. 1, 163 (10.1038/srep00163) [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
59. Kayukawa T, et al. 2014. Гормональная регуляция и роль в развитии гомолога Круппеля 1, репрессора метаморфоза, у тутового шелкопряда Бомбикс мори . Дев. биол. 388, 48–56. ( 10.1016/j.ydbio.2014.01.022) [PubMed] [CrossRef] [Google Scholar]
60. Minakuchi C, Namiki T, Yoshiyama M, Shinoda T. 2008. Опосредованный РНК-интерференцией нокдаун гена ювенильной гормональной кислоты O-метилтрансферазы вызывает преждевременную метаморфозу у красного мучного жука Tribolium castaneum . ФЕБС Дж. 275, 2919–2931. ( 10.1111/j.1742-4658.2008.06428.x) [PubMed] [CrossRef] [Google Scholar]
61. Ureña E, Chafino S, Manjón C, Franch-Marro X, Martin D.
2016.
Возникновение голометаболической стадии куколки требует взаимодействия между E93, гомолог Круппеля 1 и широкий комплекс. Генетика PLoS.
12, e1006020 (10.1371/journal.pgen.1006020) [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
Ureña E, Chafino S, Manjón C, Franch-Marro X, Martin D.
2016.
Возникновение голометаболической стадии куколки требует взаимодействия между E93, гомолог Круппеля 1 и широкий комплекс. Генетика PLoS.
12, e1006020 (10.1371/journal.pgen.1006020) [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
62. Bayer C, von Kalm L, Fristrom JW. 1996. Генная регуляция развития имагинального диска и слюнных желез во время метаморфоза Drosophila . В « Метаморфозе: постэмбриональное перепрограммирование экспрессии генов в клетках амфибий и насекомых» (ред. Гилберт Л.И., Тата Дж. Р., Аткинсон Б. Г.), стр. 321–361. Сан-Диего, Калифорния: Academic Press. [Академия Google]
63. Чжоу Б., Хирума К., Шинода Т., Риддифорд Л.М. 1998. Ювенильный гормон предотвращает индуцированную экдистероидами экспрессию РНК широкого комплекса в эпидермисе табачного рогатого червя, Manduca sexta . Дев. биол. 203, 233–244. ( 10.1006/dbio.1998.9059) [PubMed] [CrossRef] [Google Scholar]
64. Идзиро Т., Уракава Х., Ясукочи Ю., Такеда М., Фудзивара Ю.
2004.
Клонирование кДНК, структура генов и экспрессия генов широкого комплекса (BR-C) у тутового шелкопряда, Bombyx mori . Биохимия насекомых. Мол. биол.
34, 963–969. ( 10.1016/j.ibmb.2004.06.005) [PubMed] [CrossRef] [Google Scholar]
Идзиро Т., Уракава Х., Ясукочи Ю., Такеда М., Фудзивара Ю.
2004.
Клонирование кДНК, структура генов и экспрессия генов широкого комплекса (BR-C) у тутового шелкопряда, Bombyx mori . Биохимия насекомых. Мол. биол.
34, 963–969. ( 10.1016/j.ibmb.2004.06.005) [PubMed] [CrossRef] [Google Scholar]
65. Конопова Б., Джиндра М. 2008. Broad-Complex действует ниже по течению от Met в передаче сигналов ювенильного гормона, чтобы координировать метаморфоз примитивного голометаболана. Разработка 135, 559–568. ( 10.1242/dev.016097) [PubMed] [CrossRef] [Google Scholar]
66. Suzuki Y, Truman JW, Riddiford LM. 2008. Роль Броуда в развитии Tribolium castaneum : значение для эволюции голометаболической куколки насекомого. Разработка 135, 569–577. ( 10.1242/dev.015263) [PubMed] [CrossRef] [Google Scholar]
67. Piulachs MD, Pagone V, Belles X.
2010.
Ключевые роли гена Broad-Complex в эмбриогенезе насекомых. Биохимия насекомых. Мол. биол.
40, 468–475. ( 10.1016/j.ibmb.2010.04.006) [PubMed] [CrossRef] [Google Scholar]
( 10.1016/j.ibmb.2010.04.006) [PubMed] [CrossRef] [Google Scholar]
68. Kiss I, Beaton AH, Tardiff J, Fristrom D, Fristrom JW. 1988 год. Взаимодействия и влияние мутаций на развитие в Широкий комплекс из Drosophila melanogaster . Генетика 118, 247–259. [Бесплатная статья PMC] [PubMed] [Google Scholar]
69. Ureña E, Manjón C, Franch-Marro X, Martin D. 2014. Фактор транскрипции E93 определяет метаморфоз взрослых насекомых с гемиметаболическим и голометаболическим метаболизмом. проц. Натл акад. науч. США 111, 7024–7029. ( 10.1073/pnas.1401478111) [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
70. Kayukawa T, Jouraku A, Ito Y, Shinoda T. 2017. Молекулярный механизм, лежащий в основе опосредованной ювенильными гормонами репрессии преждевременного личиночно-куколочного метаморфоза. проц. Натл акад. науч. США 114, 1057–1062. ( 10.1073/pnas.1615423114) [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
71. Чафино С., Уренья Э., Казанова Дж., Касакуберта Э., Франч-Марро Х, Мартин Д.
2019.
Активация экспрессии гена E93 действует как триггер метаморфоза независимо от порогового размера у жука Tribolium castaneum . Сотовый представитель
27, 1039–1049. ( 10.1016/j.celrep.2019.03.094) [PubMed] [CrossRef] [Google Scholar]
Чафино С., Уренья Э., Казанова Дж., Касакуберта Э., Франч-Марро Х, Мартин Д.
2019.
Активация экспрессии гена E93 действует как триггер метаморфоза независимо от порогового размера у жука Tribolium castaneum . Сотовый представитель
27, 1039–1049. ( 10.1016/j.celrep.2019.03.094) [PubMed] [CrossRef] [Google Scholar]
72. Duncan DM, Kiefel P, Ducan I. 2017. Мутанты Drosophila 3b дефектны в митохондриальной функции и гибели личиночных клеток. G3-гены, Геномы Генет. 7 789–799. ( 10.1534/g3.116.037366) [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
73. Uyehara CM, Nystrom SL, Niederhuber MJ, Leatham-Jensen M, Ma Y, Buttitta LA, McKay DJ. 2017. Гормонозависимый контроль сроков развития посредством регуляции доступности хроматина. Гены Дев. 31, 862–875. ( 10.1101/gad.298182.117) [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
74. Конопова Б., Смыкал В., Джиндра М.
2011.
Общие и различные роли сигнальных генов ювенильных гормонов в метаморфозе голометаболических и гемиметаболических насекомых. ПЛОС ОДИН
6, e28728 ( 10.1371/journal.pone.0028728) [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
ПЛОС ОДИН
6, e28728 ( 10.1371/journal.pone.0028728) [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
75. Хуан Дж. Х., Лозано Дж., Беллес Х. 2013. Комплексные функции в постэмбриональном развитии таракана Blattella germanica проливают новый свет на эволюцию метаморфоза насекомых. Биохим. Биофиз. Акта 1830, 2178–2187. ( 10.1016/j.bbagen.2012.09.025) [PubMed] [CrossRef] [Google Scholar]
76. Erezyilmaz DF, Rynerson MR, Truman JW, Riddiford LM. 2009. Роль куколочной детерминанты wide в процессе эмбрионального развития пряморазвивающегося насекомого. Дев. Гены Эвол. 219, 535–544. ( 10.1007/s00427-009-0315-7) [Бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
77. Belles X, Santos CG. 2014. Путь MEKRE93 (толерантный к метопрену-гомолог Круппеля 1-E93) в регуляции метаморфоза насекомых и гомология стадии куколки. Биохимия насекомых. Мол. биол. 52, 60–68. ( 10.1016/j.ibmb.2014.06.009) [PubMed] [CrossRef] [Google Scholar]
78. Буаклин А., Ситтиканкаев К., Кхамнамтонг Б., Менасвета П., Клинбунга С.
2013.
Характеристика и экспрессионный анализ Ширококомплексный (Br-c) ген гигантской тигровой креветки Penaeus monodon . Комп. Биохим. Физиол. Б
164, 280–289. ( 10.1016/j.cbpb.2013.02.004) [PubMed] [CrossRef] [Google Scholar]
Буаклин А., Ситтиканкаев К., Кхамнамтонг Б., Менасвета П., Клинбунга С.
2013.
Характеристика и экспрессионный анализ Ширококомплексный (Br-c) ген гигантской тигровой креветки Penaeus monodon . Комп. Биохим. Физиол. Б
164, 280–289. ( 10.1016/j.cbpb.2013.02.004) [PubMed] [CrossRef] [Google Scholar]
79. Jiang SF, Zhang YP, Sun SM, Gong YS, Xiong YW, Qiao H, Zhang WY, Jin SB, Fu ХТ. 2015. Молекулярное клонирование, характеристика и анализ экспрессии гомолога широкого комплекса во время развития восточной речной креветки Macrobrachium nipponense . Жене. Мол. Рез. 14, 5141–5152. (10.4238/2015.May.18.4) [PubMed] [CrossRef] [Google Scholar]
80. Чжоу Б., Уильямс Д.В., Альтман Дж., Риддифорд Л.М., Трумэн Дж.В. 2009. Временные паттерны экспрессии широкой изоформы во время развития нейрональных линий у дрозофилы . Нейронный разработчик. 4, 39 (10.1186/1749-8104-4-39) [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
81. Starck JM, Ricklefs RE.
1998.
Паттерны развития: альтрициально-выростковый спектр. В книге «Птичий рост и развитие: эволюция в пределах предвыростково-выросткового спектра» (ред. Старк Дж. М., Риклефс Р. Е.), стр. 3–30. Оксфорд, Великобритания: Издательство Оксфордского университета. [Академия Google]
Starck JM, Ricklefs RE.
1998.
Паттерны развития: альтрициально-выростковый спектр. В книге «Птичий рост и развитие: эволюция в пределах предвыростково-выросткового спектра» (ред. Старк Дж. М., Риклефс Р. Е.), стр. 3–30. Оксфорд, Великобритания: Издательство Оксфордского университета. [Академия Google]
82. Минакучи С., Танака М., Миура К., Танака Т. 2011. Профиль развития и гормональная регуляция транскрипционных факторов , широкий и гомолога Круппеля 1 у гемиметаболических трипсов. Биохимия насекомых. Мол. биол. 41, 125–134. ( 10.1016/j.ibmb.2010.11.004) [PubMed] [CrossRef] [Google Scholar]
83. Vea IM, Tanaka S, Shiotsuki T, Jouraku A, Tanaka T, Minakuchi C. 2016. Дифференциальные вариации ювенильных гормонов при крайнем половом диморфизме щитовок. ПЛОС ОДИН 11, e0149459 (10.1371/journal.pone.0149459) [бесплатная статья PMC] [PubMed] [CrossRef] [Google Scholar]
84. Bernays EA.
1971.
Червеобразная личинка Schistocerca gregaria (Forskal): форма и активность (насекомые, прямокрылые).
Leave A Comment