
В чем биологическое значение оплодотворения у растений: особенности и описание
Размножение – это способность организмов воспроизводить себе подобных. Репродукция является одним из ключевых признаков всего живого, поэтому необходимо понимать, в чем биологическое значение оплодотворения. Этот вопрос сегодня изучен на высоком уровне начиная с основных этапов и заканчивая молекулярными и генетическими механизмами.
Что такое оплодотворение
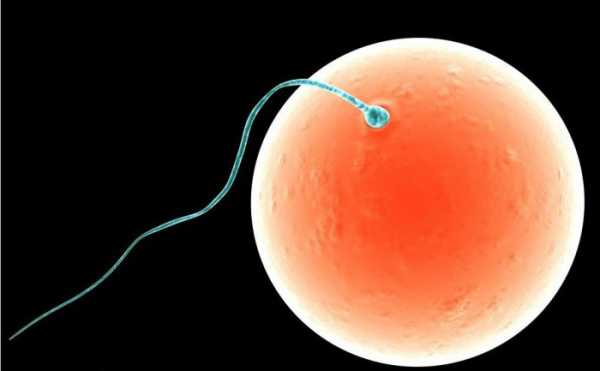
Оплодотворение – это закономерный биологический процесс слияния двух половых клеток: мужской и женской. Мужские гаметы называются сперматозоидами, а женские – яйцеклетками.
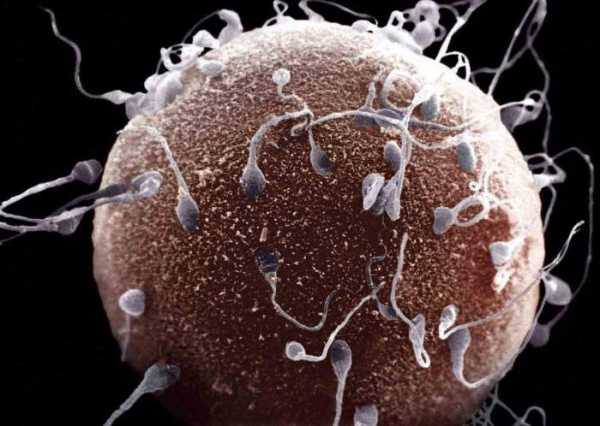
Последующим этапом после слияния половых клеток становится образование зиготы, которую можно считать новым живым организмом. Зигота начинает делиться митозом, увеличивая количество составляющих ее клеток. Из зиготы развивается зародыш.
Существует большое количество типов яйцеклеток и способов дробления. Все они зависят от таксономической принадлежности рассматриваемого живого организма, а также степени его эволюционного развития.
Каково биологическое значение оплодотворения
Размножение является основным приспособлением для продолжения рода. От репродуктивных способностей особей рассматриваемого вида зависит его будущее, поэтому у разных животных и растений есть свои способы адаптации для улучшения качества всего процесса.
Например, волки и львицы всегда защищают свое потомство от потенциальных хищников. Это увеличивает выживаемость детенышей и гарантирует в дальнейшем их приспособленность к условиям жизни. Рыбы откладывают большое количество икринок, потому что шанс внешнего оплодотворения в водной среде достаточно низок. В итоге из тысяч потенциальных мальков развиваются лишь несколько сотен.
Биологическое значение оплодотворения заключается в том, что две половые клетки от разных организмов сливаются и образуют зиготу, которая несет генетические признаки обоих родителей. Это объясняет непохожесть родственников друг на друга. И это хорошо, потому что изменение генофонда любой популяции – это эволюционный приспособительный механизм. Потомки, поколение за поколением, становятся лучше по сравнению с их родителями. В условиях постепенной смены окружающей среды (изменения климата, появление новых внешних факторов) приспособительные навыки всегда уместны.
А в чем биологическое значение оплодотворения на биохимическом уровне? Давайте рассмотрим:
- Это окончательное формирование яйцеклетки.
- Это определение пола будущего зародыша за счет соответствующих генов, принесенных мужскими гаметами.
- И, наконец, оплодотворение играет роль в восстановлении диплоидного набора хромосом, так как половые клетки по отдельности гаплоидны.
Размножение цветковых растений
Растения по сравнению с животными имеют некоторые репродуктивные особенности. Отдельного внимания требуют представители покрытосеменных, для которых характерно двойное оплодотворение (открыто русским ученым Навашиным в 1898 году).
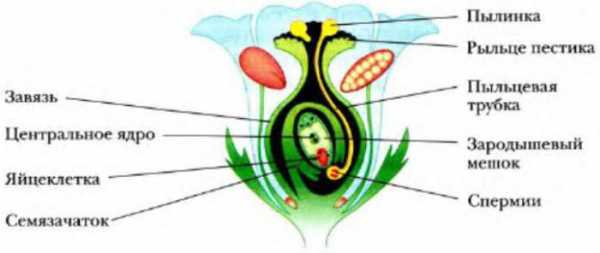
Структурами, детерминирующими половую принадлежность у цветковых растений, являются тычинки и пестики. В тычинках созревает пыльца, которая состоит из большого количество зерен. Одно зерно вмещает две клетки: вегетативную и генеративную. Пыльцевое зерно покрыто двумя оболочками, и наружная всегда имеет какие-либо выросты и углубления.
Пестик представляет собой структуру грушевидной формы, состоящую из рыльца, столбика и завязи. В завязи формируются один или несколько семязачатков, внутри которых будут созревать женские половые клетки.
При попадании пыльцевого зерна на рыльце пестика, вегетативная клетка начинает формировать пыльцевую трубу. Этот канал имеет относительно большую длину и заканчивается у микропиле семязачатка. Генеративная клетка при этом делится митозом и образует два спермия, которые по пыльцевой трубе и попадают в ткань семязачатка.
Зачем же два спермия? В чем биологическое значение оплодотворения у растений отличается от такого же процесса у животных? Дело в том, что зародышевый мешок семязачатка представлен семью клетками, среди которых есть гаплоидная женская гамета и диплоидная центральная клетка. Обе будут сливаться с пришедшими спермиями, образуя зиготу и эндосперм, соответственно.
Биологическое значение двойного оплодотворения у растений
Формирование семени – важная особенность репродукции у покрытосеменных. Для полного созревания в почве ему необходимо большое количество питательных веществ, среди которых будут различные ферменты, углеводы и другие органические/неорганические компоненты.
Эндосперм у покрытосеменных триплоидный, так как диплоидная центральная клетка зародышевого мешка слилась с гаплоидным спермием. Вот в чем биологическое значение оплодотворения у растений: тройной набор хромосом способствует высокой скорости увеличения массы ткани эндосперма. В результате семя получает много питательных веществ и запасы энергии для прорастания.
Типы семян
В зависимости от дальнейшей судьбы эндосперма, выделяют два основных вида семян:
- Семена однодольных растений. У них отчетливо виден хорошо развитый эндосперм, который занимает больший объем. Семядоля редуцирована и представлена в виде щитка. Характерен данный тип семян для всех представителей злаковых.
- Семена двудольных растений. Здесь эндосперм либо отсутствует, либо остается в виде небольших скоплений ткани на периферии. Питательную функцию у таких семян выполняют две большие семядоли. Примеры растений: горох, бобы, помидоры, огурцы, картофель.
Выводы
Конечно, называть такое оплодотворение двойным будет ошибочно, так как мы теперь знаем основные признаки и функции данного процесса. При слиянии центральной клетки со спермием не происходит формирование зиготы, а полученный генетический набор становится тройным. Все-таки семя не состоит из двух самостоятельных зародышей.
Однако биологическое значение двойного оплодотворения действительно велико. Семена при прорастании требуют большое количество органических и неорганических веществ, и данная проблема решается путем образования триплоидного эндосперма.
fb.ru
Двойное оплодотворение цветковых растений | Биология развития. Реферат, доклад, сообщение, кратко, презентация, лекция, шпаргалка, конспект, ГДЗ, тест
Двойное оплодотворение цветковых растений — это особый тип оплодотворения, который наблюдается только у цветковых (покрытосеменных) растений (рис 4). В зародышевом мешке, сформировавшемся в семязачатке завязи пестика цветка, развивается яйцеклетка. К моменту оплодотворения в зародышевом мешке помимо яйцеклетки образуется ещё двойное ядро (от слияния двух мелких клеток) и 5 других очень мелких (вспомогательных) клеток. Из проросшей на рыльце пестика пылинки по пыльцевой трубке в зародышевый мешок доставляются два спермия. Один из них сливается с яйцеклеткой, а другой спермий сливается с двойным ядром, находящимся в центральной части мешка. В итоге такого «двойного» оплодотворения из оплодотворенной яйцеклетки образуется зигота, дающая начало зародышу растения, а от слияния спермия с двойным ядром образуется особая питательная ткань, состоящая из клеток с триплоидным (3n) набором хромосом, — эндосперм, обеспечивающий питательными веществами зародыш семени.
Процесс двойного оплодотворения был открыт у цветковых растений на примере лилейных и детально изучен отечественным ботаником Сергеем Гавриловичем Навашиным в 1898 г.
Биологическое значение двойного оплодотворения у растений не совсем ясно. Однако несомненным является то, что в семенах очень быстро (опережая развитие зародыша) образуются питательные ткани с запасом высокоэнергетических питательных веществ только после оплодотворения. У цветковых растений при развитии семязачатков не тратится время на создание питательных веществ, как у голосеменных растений, поэтому развиваются они гораздо быстрее. Таким образом, благодаря двойному оплодотворению ускоряется процесс формирования и семяпочки, и яйцеклетки, и семени. Материал с сайта http://doklad-referat.ru
 |
| Рис. 4. Двойное оплодотворение цветковых растений: 1 — яйцеклетка; 2 — завязь; 3 — центральное ядро; 4 — рыльце; 5 — пылинка; 6 — пыльник; 7 — семязачаток: 8 — зародышевый мешок; 9 — пыльцевая трубка; 10 — два спермия |
Двойное оплодотворение у цветковых растений примеры
Какова функция гамет каждого организма участвующего в оплодотворении
Доклад на тему голосеменные растения кратко
Двойное оплодотворение цветковых кратко
Краткое сообщение о голосеменных растениях
Сравните оплодотворение цветковых и голосеменных растений: чем они сходны и чем отличаются?
doklad-referat.ru
Какое биологическое значение оплодотворения?
При образовании половых клеток их плоидность вдвое уменьшается, а во время оплодотворения — восстанавливается. Кроме того, перекрестное оплодотворение является одним из источников комбинативиои изменчивости: в оплодотворенной яйцеклетке в каждой паре гомологичных хромосом одна — родительская, вторая — материнская. Таким образом, половину наследственного материала будущая особь получает от отца, половину — от матери. Поэтому потомки генетически отличаются от каждого из родителей. Исключением является лишь самозаплидни организмы и представители чистых линий.
Запомните: биологическое значение оплодотворения заключается в восстановлении хромосомного набора, характерного особям определенного вида, и является одним из источников комбинативной изменчивости.
Отдельно рассмотрим биологическое значение процесса двойного оплодотворения у покрытосеменных растений. Образование триплоидные клетки, из которой впоследствии развивается эндосперм, сопровождается кратным ростом содержания ДНК в ядре. За единицу времени в триплоидные клетке синтезируется в три раза больше белковых молекул по сравнению с гаплоидной. Поэтому зародыш у покрытосеменных, имеющий больший запас питательных веществ, развивается быстрее по сравнению с голосеменными, в которых эндосперм долгое время остается гаплоидным.
Новые термины и понятия. Внешнее и внутреннее оплодотворение у животных, двойное оплодотворение у цветковых растений.
Чтобы лучше усвоить материал этого параграфа, следует вспомнить: как происходит гаметогенез? Какие источники комбинативной изменчивости? Что такое акросома? Какие особенности изогамии, гетерогамии и овогамии? Растения называют однодомными и двудомными? Известные приспособления растений к различным способам опыления? Что такое слоевище? Какое строение пильного зерна и семенного зачатке?
Оплодотворение — это процесс слияния мужской и женской половых клеток (гамет) с образованием оплодотворенной клетки (зиготы), из которой развивается новый организм.
После оплодотворения развитие нового организма начинается последовательными делениями зиготы с последующей дифференциацией клеток, тканей и органов.
Теги: конспект, образование, Оплодотворения, ответы, природа, Размножения, реферат, урокиbagazhznaniy.ru
Двойное оплодотворение у растений
Двойное оплодотворение у растений
Уникальная особенность цветковых растений — двойное оплодотворение.
В завязь покрытосеменных растений проникает два спермия, один из них сливается с яйцеклеткой, дав начало диплоидному зародышу. Другой соединяется с центральной диплоидной клеткой. Образуется триплоидная клетка, из которой возникнет эндосперм — питательный материал для развивающегося зародыша ( рис. 77 ). Этот процесс, характерный для всех покрытосеменных, открыт в конце прошлого века С.Г. Навашиным и получил название двойного оплодотворения. Значение двойного оплодотворения, по- видимому, заключается в том, что обеспечивается активное развитие питательной ткани уже после оплодотворения. Поэтому семяпочка у покрытосеменных не запасает питательных веществ впрок и, следовательно, развивается гораздо быстрее, чем у многих других растений, например у голосеменных.
У цветковых растений имеется ряд особенностей образования половых клеток и оплодотворения. Оплодотворению у них предшествует образование сильно редуцированного гаплоидного поколения — гаметофитов. После оплодотворения прорастание пыльцы цветковых растений начинается с разбухания зерна и образования пыльцевой трубки , которая прорывает спородерму в более тонком ее месте — так называемой апертуре. Кончик пыльцевой трубки выделяет специальные вещества, размягчающие ткани рыльца и столбика , в которые внедряется пыльцевая трубка. По мере роста пыльцевой трубки в нее переходят ядро вегетативной клетки и оба спермия . В огромном большинстве случаев пыльцевая трубка проникает в мегаспорангий ( нуцеллус ) через микропиле семязачатка , реже — иным образом. Проникнув в зародышевый мешок , пыльцевая трубка разрывается, и ее содержимое изливается внутрь. Один из спермиев сливается с яйцеклеткой, и образуется диплоидная зигота, дающая затем начало зародышу . Второй спермий сливается со вторичным ядром, располагающимся в центре зародышевого мешка , что приводит к образованию триплоидного ядра, развивающегося затем в триплоидный эндосперм. Весь этот процесс получил название двойного оплодотворения. Он был впервые описан в 1898 г. выдающимся русским цитологом и эмбриологом С.Г.Навашиным. Прочие клетки зародышевого мешка — антиподы и синергиды в оплодотворении не участвуют и довольно быстро разрушаются.
Как и у других высших растений , при мейозе у цветковых образуются споры . Микроспоры (мужские споры) образуются в пыльниках , из них формируются пыльцевые зерна (мужские гаметофиты). Зрелое пыльцевое зерно состоит из трех клеток — вегетативной (или клетки, пыльцевой трубки ) и двух спермиев (лишенные жгутика сперматозоиды). Мегаспоры (женские споры) формируются в семяпочках внутри завязи пестика .
Биологический смысл двойного оплодотворения весьма велик. В отличие от голосеменных , где довольно мощный гаплоидный эндосперм развивается независимо от процесса оплодотворения, у покрытосеменных триплоидный эндосперм образуется лишь в случае оплодотворения. С учетом гигантского числа поколений этим достигается существенная экономия энергетических ресурсов. Увеличение же уровня плоидности эндосперма до 3n, по-видимому, способствует более быстрому росту этой полиплоидной ткани по сравнению с диплоидными тканями спорофита .
Взаимодействие пыльцевой трубки гаметофита с тканями спорофита — сложный процесс, регулируемый химическими веществами. Так, выяснилось, что если промыть пыльцу дистиллированной водой, она теряет способность к прорастанию. Если сконцентрировать полученный раствор и обработать концентратом пыльцу, она вновь станет полноценной. После прорастания рост пыльцевой трубки контролируется тканями пестика. Например, у хлопчатника рост трубки до яйцеклетки занимает 12-18 ч, но уже через 6 ч можно установить, к какой семяпочке направляется пыльцевая трубка: в этой семяпочке начинается разрушение особой клетки — синергиды . Как растение направляет рост трубки в нужную сторону и каким образом синергида узнает о ее приближении, пока еще не известно.
Во многих случаях у цветковых растений существует «запрет» на самоопыление: спорофит «узнает» своего мужского гаметофита и не разрешает ему участвовать в оплодотворении. В некоторых случаях при этом собственная пыльца не прорастает на рыльце пестика. В большинстве же случаев рост пыльцевой трубки начинается, но затем останавливается и она не достигает яйцеклетки. Например, у первоцвета весеннего еще Ч.Дарвин обнаружил две формы цветков — длинностолбиковые (с длинным столбиком и короткими тычинками) и короткостолбиковые (столбик короткий, тычиночные нити длинные). У короткостолбиковых растений пыльца почти вдвое крупнее, а клетки сосочков рыльца мелкие. Все эти признаки контролируются группой тесно сцепленных генов.
Опыление эффективно только при переносе пыльцы с одной формы на другую. За распознавание своей пыльцы отвечают молекулы-рецепторы, представляющие собой сложные комплексы белков с углеводами. Показано, что растения дикой капусты , которые не вырабатывают в тканях рыльца молекул рецептора, могут самоопыляться. У нормальных растений рецепторы появляются на рыльце за день до открытия цветка. Если раскрыть бутон и нанести на него собственную пыльцу за два дня до распускания, то оплодотворение произойдет, а если за один день до распускания — то нет.
Интересно, что в некоторых случаях самонесовместимость пыльцы у растений определяется серией множественных аллелей одного гена, сходно с несовместимостью при пересадках тканей у животных. Эти аллели обозначаются буквой S, и число их в популяции может достигать десятков и даже сотен. Если, например, генотип производящего яйцеклетки растения — s1s2, а производящего пыльцу — s2s3, то прорастать при перекрестном опылении будут только 50% пылинок — те, что несут аллель s3. При наличии десятков аллелей большая часть пыльцы при перекрестном опылении нормально прорастает, а самоопыление полностью предотвращается.
Ссылки:
medbiol.ru
Двойное оплодотворение
Пыльцевые зерна, попав на рыльце пестика, поглощают воду, сахара и другие питательные вещества и образуют трубку. Трубка буквально прорастает в пестик до зародышевого мешка. Пыльцевая трубка проникает в зародышевый мешок, где одна мужская гамета соединяется с яйцеклеткой, образуя зиготу. После митотического деления зигота становится зародышем будущего семени. Вторая мужская гамета сливается с двумя полярными ядрами и образует эндосперм. Этот процесс известен как двойное оплодотворение.[ …]
Двойное оплодотворение представляет собой исторически сложившееся, качественно новое явление, свойственное только покрытосеменным растениям. Биологическое значение двойного оплодотворения заключается в том, что эндосперм, обогащенный двойной наследственностью, повышает жизнеспособность и приспособленность покрытосеменных, обеспечивает их преимущество перед другими растениями в современную геологическую эпоху.[ …]
Двойное оплодотворение — тип оплодотворения, свойственный покрытосеменным растениям, при котором один из спермиев сливается с яйцеклеткой с образованием диплоидной зиготы, дающей начало зародышу семени, а второй спермпй сливается с диплоидным ядром зародышевого мешка с образованием клетки, дающей начало эндосперму семенп.[ …]
После двойного оплодотворения первым обычно делится первичное ядро эндосперма (рис, 124). У пшеницы его деление начинается через 3—4 ч после опыления. В это время в цитоплазме зародышевого мешка и в первичном ядре эндосперма интенсивно накапливаются нуклеиновые кислоты (РНК и ДНК), белки, полисахариды, ферменты, витамины и другие физиологически активные вещества. С момента образования зиготы и до полного формирования зародыша эндосперм служит основным источником его питания.[ …]
Впоследствии из оплодотворенной яйцеклетки (зиготы) образуется зародыш, а из оплодотворенного ядра центральной клетки — эндосперм. Двойное оплодотворение присуще всем систематическим группам покрытосеменных растений.[ …]
Постмитотический тип двойного оплодотворения детально исследован Е. Н. Герасимовой-Навашиной у рябчика горного (Fritillaria tenella) (рис. Она же изучала промежуточный тип двойного оплодотворения у гальтонии. Премитотический процесс оплодотворения у пшеницы показан на рисунках 111, Л и 113.[ …]
Как было отмечено выше, оплодотворение у цветковых (покрытосеменных) имеет существенную отличительную особенность в виде двойного оплодотворения (С. Г. Навашин, 1896), которое сводится к тому, что в зародышевом мешке гаплоидная яйцеклетка и диплоидная центральная клетка оплодотворяются спермиями, в результате чего образуется диплоидный зародыш и триплоидная клетка, развивающаяся в клетки эндосперма (см. гл. II).[ …]
Герасимова-Навашнна Е. Н. Двойное оплодотворение покрытосеменных и некоторые теоретические аспекты//Проблемы эмбриологии.— Киев: Наукова думка, 1971.[ …]
Правильным представлением об оплодотворении у покрытосеменных мы обязаны выдающемуся русскому цитологу-эмбриологу С. Г. Навашину, открывшему в 1898 г. двойное оплодотворение у лилейных и астровых. Он впервые показал, что у цветковых растений при оплодотворении один из спермиев сливается с ядром яйцеклетки, а другой — с ядром центральной клетки или полярными ядрами (рис. 109).[ …]
У плодовых растений после завершения двойного оплодотворения из с.емяпочки формируется семя, а наружные покровы семяпочки (или интегументы) превращаются в кожуру семени. Клетки нуцеллуса используются растущим зародышем или реже превращаются в питательную ткань — перисперм. У болв-шинства плодовых и ягодных растений эндосперм и клетки нуцеллуса идут на формирование зародыша, а запасные пластические вещества локализуются в семядолях зародыша (рис. 28).[ …]
При изучении структуры спермиев в процессе двойного оплодотворения С. Г. Навашин обнаружил, что их ядра находятся в состоянии поздней телофазы. Механизм движения спермиев в пыльцевой трубке и зародышевом мешке он рассматривал как активный процесс самостоятельного передвижения.[ …]
Весь ;>тот процесс, как известно, принято намывать двойным оплодотворением. Ведь согласно общепринятому в литературе определению, оплодотворение (еипгамин) — это процесс слияния, мужской п женской половых клеток (гамет) с образованием зиготы, из которой и дальнейшем разнимается новый организм. Такое определение оплодотворения можно найти в любом учебнике биологии и в любом :нщнклопедичоском словаре (в том число в советских шщпклопедиях, в Большой и Малой). И даже в известном «Словаре русского языка» С. И. Ожегова (1973) мы читаем: «Оплодотворить. 1. Создать зародыш в ком-чем-н. слиянием мужской и женской половых клеток. 2. Послужить источником развития, совершенствования». Слияние одного из сиормпев с яйцеклеткой является, несомненно, оплодотворением, но тройное слияние по является, строго говоря, оплодотворением, так как 1) центральная клетка — не гамета и 2) в результате ¡»того слияния но образуется зигота, из которой и дальнейшем развивался бы новый организм. Очевидно, тройное слияние является оплодотворением лишь в указанном Ожеговым, втором, переносном смысле. Другими словами, в выражении «двойное оплодотворение» термин «оплодотворение» применяется в двух разных, смыслах — прямом и переносном. Тем не менее выражение «двойное онлодотиороние» настолько широко вошло в литературу, что было бы нецелесообразно его заменить (а попытки такого рода делались, в том числе известным, немецким ботаником. Достаточно, если мы будем помнить, что речь идет здесь о двух разных биологических процессах, условно объединяемых общим .названием.[ …]
Открытие в 1896 г. выдающимся русским ученым Сергеем Гавриловичем Навашииым двойного оплодотворения у покрытоременных растений положило начало новой .эре исследований в биологической науке. С. Г. Навашин является также основателем науки о ядре — кариологии.[ …]
Детально изучая поведение спермиев в зародышевом мешке, Е. Н. Герасимова-Навашина предложила различать два основных типа двойного оплодотворения: премитотический, когда объединение половых ядер происходит перед первым митозом зиготы, и постмитотический, когда объединение половых ядер наступает в начале первого митоза зиготы. Ею установлена также промежуточная форма двойного оплодотворения, характеризующаяся образованием собственной оболочки вокруг ядра спермия при медленном его погружении в ядро яйцеклетки. При этом объединение половых ядер происходит во время первого митоза зиготы (рис. 110).[ …]
Рост семенного растения начинается с прорастапия семени. Семена у покрытосеменных растений образуются из семяпочки в результате двойного оплодотворения. В пыльце сосредоточено большое количество физиологически активных веществ — фитогормонов (ауксинов), ферментов, много аминокислоты пролина. Все ето приводит к тому, что уже сразу после опыления интенсивность обмена веществ в системе пыльца — пестик резко усиливается. Пыльцевая трубка несет два слермия. В результате слияния первого спермия с яйцеклеткой образуется зигота, дающая при последующем развитии зародыш семени. Анатомически ткани зародыша целиком состоят из первичной меристемы. Оно начинает делиться и вместе с цитоплазмой образует множество клеток, составляющих эндосперм. После опыления и особенно оплодотворения завязь начинает разрастаться. Вместе с пыльцой в зародышевый мешок привносятся как сами фитогормоны (ауксины), так и ферменты, катализирующие синтез гормонов. Дыхание развивающегося после оплодотворения семени идет очень интенсивно. Семя становится центром притяжения питательных веществ. В процессе развития и роста зародыша ткань вндосперма потребляется и в моменту созревания семян может частично или полностью исчезнуть. Некоторые семена лишены андосперма, питательные вещества сосредоточены у ттнт в тканях самого зародыша (в семядолях).[ …]
У Crépis capillaris ядра спермиев при слиянии с женскими половыми ядрами находятся в периоде G прерванного митотического цикла (стадия телофазы), который они заканчивают в зиготе (рис. 112, 118). У астровых и злаковых в момент оплодотворения спермии находятся в стадии поздней телофазы митоза. На рисунке 114 показан конец двойного оплодотворения у кукурузы в момент слияния мужских ядрышек с женскими в ядрах зиготы и первичного ядра эндосперма; при этом второй сперматозоид находится в полярных ядрах; в яйцеклетке процесс слияния уже закончен, присутствует ядрышко спермия; слева виден контур второй пыльцевой трубки со спермиями.[ …]
ru-ecology.info
Двойное оплодотворение у растений :: SYL.ru
Двойное оплодотворение у растений имеет большое биологическое значение. Оно было открыто Навашиным в 1898 г. Далее рассмотрим подробнее, как происходит двойное оплодотворение у растений.
Биологическое значение
Процесс двойного оплодотворения способствует активному развитию питательной ткани. В связи с этим семяпочка не запасает вещества впрок. Это, в свою очередь, объясняет ее быстрое развитие.
Схема двойного оплодотворения
Коротко явление можно описать следующим образом. Двойное оплодотворение у покрытосеменных растений состоит в проникновении в завязь двух спермиев. Один сливается с яйцеклеткой. Это способствует началу развития диплоидного зародыша. Второй спермий соединяется с центральной клеткой. В результате формируется триплоидный элемент. Из этой клетки появляется эндосперм. Он является питательным материалом для развивающегося зародыша.
Развитие пыльцевой трубки
Двойное оплодотворение у покрытосеменных начинается после образования гаплоидного сильно редуцированного поколения. Оно представлено гаметофитами. Двойное оплодотворение цветковых растений способствует прорастанию пыльцы. Оно начинается с разбухания зерна и последующего формирования пыльцевой трубки. Она прорывает спородерму в наиболее тонком ее участке. Называется он апертура. С кончика пыльцевой трубки выделяются специфические вещества. Они размягчают ткани столбика и рыльца. За счет этого в них входит пыльцевая трубка. По мере ее развития и роста, в нее переходят оба спермия и ядро от вегетативной клетки. В подавляющем большинстве случаев проникновение пыльцевой трубки в нуцеллус (мегаспорангий) происходит посредством микропиле семязачатка. Крайне редко это осуществляется другим способом. После проникновения в зародышевый мешок происходит разрыв пыльцевой трубки. В результате все ее содержимое изливается вовнутрь. Двойное оплодотворение цветковых растений продолжается формированием диплоидной зиготы. Этому способствует первый спермий. Второй элемент соединяется с вторичным ядром, которое расположено в центральной части зародышевого мешка. Образованное триплоидное ядро впоследствии трансформируется в эндосперм.

Формирование клеток: общие сведения
Процесс двойного оплодотворения цветковых растений осуществляется особыми половыми клетками. Их формирование происходит в два этапа. Первая стадия называется спорогенез, вторая – гематогенез. В случае образования мужских клеток эти этапы именуются микроспорогенез и микрогематогенез. При образовании женских половых элементов приставка меняется на «мега» (или «макро»). Спорогенез основывается на мейозе. Это процесс формирования гаплоидных элементов. Мейозу, так же как и у представителей фауны, предшествует размножение клеток посредством митотических делений.
Образование спермиев
Первичное формирование мужских половых элементов осуществляется в особой ткани пыльника. Она называется археспориальной. В ней в результате митозов происходит формирование многочисленных эелементов – материнских клеток пыльцы. Они и вступают затем в мейоз. Вследствие двух мейотических делений образуется 4 гаплоидные микроспоры. Некоторое время они лежат рядом, формируя тетрады. После этого происходит их распад на пыльцевые зерна — отдельные микроспоры. Каждый из образованных элементов начинает покрываться двумя оболочками: внешней (экзина) и внутренней (интина). Затем начинается следующий этап – микрогаметогенез. Он, в свою очередь, состоит из двух митотических последовательных делений. После первого формируется две клетки: генеративная и вегетативная. Впоследствии первая проходит еще одно деление. В результате образуется две мужские клетки – спермии.

Макроспорогенез и мегаспорогенез
В тканях семяпочки начинает обособляться один или несколько археспориальных элементов. Они начинают усиленно расти. Вследствие такой активности они становятся значительно крупнее остальных клеток, окружающих их в семяпочке. Каждый археспориальный элемент один, два или более раз подвергается делению митозом. В некоторых случаях клетка может сразу трансформироваться в материнскую. Внутри нее происходит мейоз. В результате него формируется 4 гаплоидные клетки. Как правило, самая крупная из них начинает развиваться, превращаясь в зародышевый мешок. Три оставшиеся постепенно дегенерируют. На данном этапе макроспорогенез завершается, начинается макрогематогенез. В ходе него происходят митотические деления (у большей части покрытосеменных их три). Цитокинез не сопровождает митозы. В результате трех делений формируется зародышевый мешок с восемью ядрами. Они впоследствии обосабливаются в самостоятельные клетки. Эти элементы распределяются определенным образом по зародышевому мешку. Одна из обособленных клеток, которая, собственно, является яйцеклеткой, совместно с двумя другими – синергидами, занимает место у микропиле, в которое осуществляется проникновение спермиев. В этом процессе синергиды исполняют очень значимую роль. В них содержатся ферменты, которые способствуют растворению оболочек на пыльцевых трубках. В противоположной стороне зародышевого мешка располагаются другие три клетки. Они именуются антиподами. С помощью этих элементов происходит передача из семяпочки питательных веществ в зародышевый мешок. Оставшиеся две клетки располагаются в центральной части. Зачастую они сливаются. В результате их соединения формируется диплоидная центральная клетка. После того как произойдет двойное оплодотворение, и в завязь проникнут спермии, один из них, как выше сказано было, сольется с яйцеклеткой.
Особенности пыльцевой трубки
Двойное оплодотворение сопровождается взаимодействием ее с тканями спорофита. Оно достаточно специфично. Этот процесс регулируется активностью химических соединений. Установлено, что если пыльцу промыть в дистиллированной воде, она потеряет способность к прорастанию. Если же полученный раствор сконцентрировать, а затем ее обработать, то она снова станет полноценной. Развитие пыльцевой трубки после прорастания контролируют ткани пестика. К примеру, у хлопчатника ее рост до яйцеклетки занимает порядка 12-18-ти часов. Однако уже спустя 6 часов вполне можно определить, к какой именно семяпочке будет направляться пыльцевая трубка. Это понятно потому, что в ней начинается разрушение синергиды. В настоящее время не установлено, как растение может направить развитие трубки в нужном направлении и каким образом о приближении узнает синергида.
«Запрет» на самоопыление
Он достаточно часто наблюдается у цветковых растений. Это явление имеет свои особенности. «Запрет» на самоопыление проявляется в том, что спорофит «идентифицирует» собственного мужского гематофита и не допускает его к участию в оплодотворении. При этом в ряде случаев на рыльце пестика не происходит прорастания собственной пыльцы. Однако, как правило, рост трубки все-таки начинается, но впоследствии приостанавливается. В результате пыльца не достигает яйцеклетки и, как следствие, двойное оплодотворение не происходит. Еще Дарвиным было отмечено это явление. Так, он обнаружил у первоцвета весеннего цветки двух форм. Одни из них были длинностолбиковыми с короткими тычинками. Другие же – короткостолбиковыми. В них тычиночные нити были длинные. Короткостолбиковые растения отличаются крупной пыльцой (вдвое больше, чем у других). При этом клетки в сосочках рыльца – мелкие. Указанные признаки контролирует группа из тесно сплетенных генов.
Рецепторы
Двойное оплодотворение эффективно, когда пыльца переносится от одной формы к другой. За распознавание собственных элементов отвечают особые молекулы-рецепторы. Они представляют собой сложные соединения углеводов с белками. Установлено, что формы дикой капусты, не вырабатывающие в тканях рыльца эти молекулы-рецепторы, способны самоопыляться. Для нормальных растений характерно появление углеводно-белковых соединений за день до раскрытия цветка. Если открыть бутон и обработать его собственной пыльцой за двое суток до его распускания, то двойное оплодотворение произойдет. Если это сделать за день до открытия, то его не будет.
Аллели
Примечательно, что в ряде случаев «самонесовместимость» пыльцы в растениях устанавливается серией множественных элементов одного гена. Это явление похоже на несовместимость при пересадке ткани у животных. Такие аллели обозначают литерой S. Число в популяции этих элементов может достичь десятков или даже сотен. К примеру, если генотип растения, производящего яйцеклетки, — s1s2, а вырабатывающего пыльцу — s2s3, при перекрестном опылении прорастание будет отмечено только у 50% пылинок. Это будут те, которые несут аллель s3. Если элементов несколько десятков, то большая часть пыльцы прорастет нормально при перекрестном опылении, при этом самоопыление предотвращается полностью.

В заключение
В отличие от голосеменных, для которых характерно развитие достаточно мощного гаплоидного эндосперма вне зависимости от оплодотворения, у покрытосеменных ткань образуется только в этом единственном случае. Учитывая огромное количество поколений, таким образом достигается значительная экономия энергии. Повышение степени плоидности эндосперма, по всей видимости, способствует более скорому росту ткани в сравнении с диплоидными слоями спорофита.
www.syl.ru
СОДЕРЖАНИЕ 1. Введение 1. ВВЕДЕНИЕ Открытие в 1896 году выдающимся русским ученым Сергеем Гавриловичем Навашиным двойного оплодотворения у покрытосеменных растений положило начало новой эре исследований в биологической науке. 2. МИКРОСПОРОГЕНЕЗ И РАЗВИТИЕ МУЖСКОГО ГАМЕТОФИТА 1) МИКРОСПОРОГЕНЕЗ На ранних этапах развития цветка вслед за образованием бугорков околоцветника закладываются бугорки тычинок в виде комплекса меристематических клеток. В дальнейшем из нижней части бугорка развивается тычиночная нить, а из верхней — пыльник. Из клеток бугорка пыльника впоследствии дифференцируется группа клеток, отличающихся от остальных по структуре. Они содержат крупные ядра, ядрышки и густую цитоплазму и составляют гнездо пыльника. Обычно каждый пыльник имеет два гнезда, а каждая тычинка — два пыльника, соединенных между собой тканью — связником. Клетки, составляющие гнездо пыльника, называются первичными клетками археспория. Они интенсивно делятся тангентальными перегородками на париентальные и вторичные клетки археспория, или спорогенные клетки, которые впоследствии превращаются в материнские клетки микроспор — микроспороциты. 2) СПЕРМИОГЕНЕЗ Из первичной экзины, сформировавшейся вокруг каждой микроспоры после растворения каллозной оболочки материнской клетки, и формируются собственные оболочки микроспор — экзина (наружная) и интина (внутренняя), с момента появления которых микроспоры превращаются в пыльцевые зерна. 3. МАКРОСПОРОГЕНЕЗ И РАЗВИТИЕ ЖЕНСКОГО ГАМЕТОФИТА — ЗАРОДЫШЕВОГО МЕШКА 1) ФОРМИРОВАНИЕ СЕМЯПОЧКИ И МАКРОСПОРОГЕНЕЗ Совокупность женских генеративных органов в цветке называется гинецеем или пестиком. Полностью сформированный пестик развивается из одного или нескольких сросшихся между собой плодолистиков и имеет завязь и рыльце. Завязь может быть одно-, двух-, трех- и многогнездной. В полости завязи закладываются бугорки семяпочек, в каждой из них в дальнейшем развивается женский гаметофит — зародышевый мешок. Место заложения и прикрепления семяпочек и завязи называется семяносцем или плацентой. 2) РАЗВИТИЕ ЖЕНСКОГО ГАМЕТОФИТА — ЗАРОДЫШЕВОГО МЕШКА Существует несколько классификаций типов развития зародышевых мешков у покрытосеменных растений. В основу наиболее современных из них положены такие показатели, как число делений, начиная от макроспоры и кончая зрелым зародышевым мешком, число макроспор, участвующих в образовании зародышевых мешков, общее число ядер и клеток, входящих в состав зародышевого мешка. 4. БИОЛОГИЧЕСКОЕ ЗНАЧЕНИЕ ДВОЙНОГО ОПЛОДОТВОРЕНИЯ Правильным представлением об оплодотворении у покрытосеменных мы обязаны выдающемуся русскому цитологу-эмбриологу С.Г. Навашину, открывшему в 1898 году двойное оплодотворение у линейных и астровых. Он впервые показал, что у цветковых растений при оплодотворении один из спермиев сливается с ядром яйцеклетки, а другой — с ядром центральной клетки или полярными ядрами. 5. ЭМБРИОНАЛЬНОЕ РАЗВИТИЕ Эмбриональное развитие растений начинается после того, как оплодотворенная яйцеклетка (зигота) и слившееся со вторым спермием центральное ядро начинает делиться, При этом из зиготы развивается зародыш, а из ядра центральной клетки формируется эндосперм. Между ними с самых ранних этапов развития устанавливается тесное взаимодействие: малейшие нарушения в формировании эндосперма немедленно отражаются на развитии зародыша (главным образом на его дифференциации). При недоразвитом и аномальном эндосперме семяпочка обычно не дает полноценного семени. 1) РАЗВИТИЕ ЭНДОСПЕРМА И ПЕРИСПЕРМА У большинства покрытосеменных растений первичное ядро эндосперма, возникшее в результате слияния второго спермия с ядром центральной клетки или с двумя полярными ядрами, триплоидное. Но, в зависимости от типа развития зародышевого мешка первичное ядро эндосперма может оказаться и высокополиплоидным, если произойдет слияние спермия со многими (4-8) полярными ядрами. 2) РАЗВИТИЕ ЗАРОДЫША Оплодотворенная яйцеклетка называется зиготой, или зародышевой клеткой. Она способна развиваться в новый организм и обладает двойственной наследственностью, то есть качественно отличается как от яйцеклетки, так и от спермия. В состоянии покоя зигота находится более продолжительное время, чем первичное ядро эндосперма. Обычно она имеет грушевидную или удлиненную форму, ее ядро располагается в верхней части клетки и окружено густой цитоплазмой, в которой содержатся лейкопласты, рибосомы и др. ЗАКЛЮЧЕНИЕ Последняя треть ХIХ века ознаменовалась крупнейшими открытиями в области ботаники и цитологии: в растительной клетке были обнаружены особые внутриклеточные структуры — хромосомы, описаны способы деления ядра. Ключевые слова страницы: как, скачать, бесплатно, без, регистрации, смс, реферат, диплом, курсовая, сочинение, ЕГЭ, ГИА, ГДЗ |
referatzone.com
Leave A Comment